Copyright (c) Vladimir DARIC
Permission is granted to copy, distribute and/or modify this
document under the terms of the GNU Free Documentation License, Version 1.2 or
any later version published by the Free Software Foundation;
with no Invariant Sections, no Front-Cover Texts, and no
Back-Cover Texts. A copy of the license is included in the section entitled
"GNU
Free Documentation License".
Stage en laboratoire :
Etude des voies nociceptives mises en
jeu
pendant la parturition, étude
par
immunomarquage de la protéine c-Fos
sur
les coupes de la moelle épinière de la
rate
Sommaire
Stage en laboratoire : Etude des voies nociceptives mises
en jeu pendant la parturition, étude par immunomarquage de la
protéine c-Fos sur les coupes de
la moelle épinière de la rate
Introduction 4
Rôle physiologique de la Douleur 4
Physiologie de la douleur 4
Les voies périphériques 4
La moelle épinière 5
Voies ascendantes 6
Les centres supérieurs 7
Parturition 7
Origine de la douleur 8
Les voies de la douleur et la distribution lombaire des neurones
nociceptifs. 9
Changements physiologiques chez la mère et chez le
foetus: pendant la gestation et pendant
l'accouchement 10
Anesthésie et analgésie, fonctionnement,
méthodes actuelles, difficultés. 12
Matériel et méthodes .15
c-fos 15
La protéine c-Fos, est-elle un bon marqueur de
l'activité nociceptive ? 16
Avantages et inconvénients de la méthode. 16
Induction du c-fos 18
Expression spinale de la protéine
c-Fos lors de la parturition chez le rat, mise en évidence par
l'immuno-marquage 19
Etude préliminaire - Expression neuronale de la
protéine c-Fos dans la moelle épinière lors de
la parturition chez la rate primipare et multipare. 19
Expression neuronale de la protéine c-Fos dans la moelle
épinière chez la rate. 19
Animaux .20
Protocole expérimental .20
Perfusion 21
Coupes frontales de moelle épinière 21
Immunohistochimie sur coupes flottantes 21
Analyses des résultats et statistiques. 22
Résultats 24
Conclusion : 24
Epilogue .26
Etude comportementale 26
Immunohistochimie de la protéine c-Fos .27
Effets de l'ocytocine en sous-cutané sur le comportement
accompagnant la parturition. 27
Etude des effets pharmacologiques de la
morphine administrée en péridurale sur la parturition...
27
Les contrôles préliminaires 27
Etude des effets
pharmacologiques de la morphine administrée en péridurale sur la
parturition...
28
Discussion et conclusion. .29
Appendice .31
Introduction
Rôle physiologique de la Douleur
Chez l'animal la douleur joue un rôle d'alarme dans
l'adaptation. Même chez l'homme, dire cela n'est pas toujours faux.
L'expérience de la douleur va conduire, normalement, vers un
comportement qui éloignera le corps de la source du mal. La sensation
douloureuse après un traumatisme peut induire certains comportements qui
favorisent le rétablissement et la guérison : le sommeil, le fait
de se nourrir et de boire ; chez l'animal le toilettage, le léchage
etc.
Physiologie de la douleur
Chez l'homme le mot « douleur » soulève une
multitude d'aspects ; sensoriel, psychologique etc. ... Nous ne nous
intéressons ici qu'à l'aspect neurologique de la douleur,
c'est-à-dire la perception du stimulus douloureux, son transport et son
intégration par le système nerveux central.
Ainsi, on définit la douleur comme la sensation ou la
perception d'effets irritants, pénibles, lancinants ou insupportables
venant d'une partie du corps, alors que la nociception constitue le processus
sensoriel à l'origine des signaux nerveux qui la déclenchent. Les
systèmes de modulation et d'intégration, qui seront
brièvement décrits par la suite, assurent le lien entre ces deux
phénomènes. Cela veut donc dire que la nociception peut exister
sans que la douleur soit perçue. De même lorsque il y a
dysfonctionnement de ces systèmes la douleur peut apparaître
spontanément, en absence de tout stimulus nociceptif.
Les premières considérations envisageaient la
douleur comme un message sensoriel de très forte intensité. Cette
idée est en partie fausse. Lorsque les circuits spécifiques de la
nociception, et les systèmes de contrôle qui modulent en
permanence le transfert de l'information aux différents niveaux du
circuit de la douleur, ont été caractérisés, on a
commencé à envisager le message nociceptif comme étant
distinct du message sensoriel classique.
Les voies périphériques
Le message nociceptif prend naissance à la
périphérie. En 1980 Perl démontre qu'il existe plusieurs
populations de fibres périphériques qui répondent à
une stimulation douloureuse. Il est généralement admis qu'il n'y
a pas de structure bien spécifique, histologiquement bien
individualisée, pouvant être qualifiée de récepteur
nociceptif. Le message nociceptif résulte de la mise en jeu de
terminaisons libres amyéliniques constituant des arborisations
plexiformes dans les tissus cutanés, musculaires, articulaires, ainsi
que dans les parois des viscères. Les fibres qui véhiculent ce
message sont à conduction lente, et en général ils
appartiennent aux deux groupes :
Fibres Aä - faiblement myélinisées
(diamètre de 1 à 5 microns). Ces fibres sont
responsables de
la perception de la douleur rapide, bien localisée ; de type
piqûre
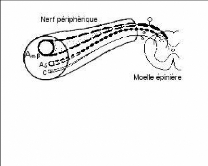
Fig 1. Fibres afférentes primaires Fibres
Aá et â - véhiculent le
message sensitif
Fibres ä et C transportent le
message nociceptif
Fibres C - non myélinisées (diamètre 0,3
à 1,5 microns), responsables de la douleur tardive, plus diffuse ; de
type brûlure
Plusieurs sous-types de ces fibres existent, mais on ne
détaillera pas leur particularités ici.
Les corps cellulaires des fibres afférentes primaires
forment les ganglions des racines dorsales ( ganglions rachidiens). Ces
neurones ont une morphologie particulière, ils n'ont pas de
prolongements dendritiques, mais seulement un axone divisé en deux
branches (neurones en T).
C'est donc au niveau du ganglion rachidien que sont
synthétisées les molécules qui transmettent le message
nociceptif au niveau de la première synapse. Pour illustrer la
diversité de neuromédiateurs existants, on peut citer :
les acides aminés excitateurs : glutamate,
aspartate
les peptides, telle la substance P, la neurokinine A, CGRP, la
galanine,
l'omatostatine, le polypeptide vaso-intestinal (VIP), la
bombesine, la neurotensine
Il est aujourd'hui banal de voir des clichés obtenus
par double marquage immunologique montrant une co-localisation de ces
molécules à l'intérieur du même neurone. Les
neuromédiateurs sont excrétés non seulement au niveau de
la moelle épinière (voie orthodromique), mais aussi au niveau des
terminaisons périphériques des fibres afférentes par le
réflexe d'axone (voie antidromique). Ainsi, on a une relation bivalente,
le stimulus nociceptif évoque un message nerveux, mais à son tour
la fibre nerveuse agit au niveau du tissu périphérique pour
éventuellement moduler le phénomène qui est à la
source du message nociceptif.
Après sa sortie du ganglion rachidien, la racine
dorsale envoie une collatérale vers le bulbe rachidien, et avant
d'entrer dans la moelle elle projette des faisceaux dans plusieurs segments
voisins, en amont et en aval de leur segment d'entré.
|
La moelle épinière
La moelle épinière a été l'objet
de nombreuses études anatomiques. Elle est formée par la
substance blanche - composée en majorité par les fibres nerveuses
- qui entoure la substance grise - constituée par les somas des
neurones. La substance grise à été subdivisée en
dix couches définies chez le chat, selon les critères
cyto-architectoniques (couches de Rexed, en 1965). La corne dorsale comprend
les couches I à VI, tandis que les couches VII à X composent la
corne ventrale (voir schéma ci-contre). Les
|
|
|
Fig 2. Couches de Rexed
Schémas représentent la subdivision de la
moelle épinière de chat, en 10 couches cellulaires distinctes (I
à X), établie par Rexed. A gauche : segment C5, au centre : le
segment T4 et à droite : le segment L7.
|
|
|
|
|
études physiologiques de la moelle confirment sans
cesse que cette division selon les critères cyto-architectoniques, a
aussi des bases fonctionnelles.
Les études anatomiques ont démontré que
les fibres afférentes primaires, C et Aä, se terminent
majoritairement dans les couches I et II. Les fibres Aä, en particulier,
projettent en général dans les couches IV-VI. La plupart des
fibres Aâ et Aä non nociceptives envoient leurs terminaisons vers
les couches III-V.
Il existe deux types de cellules qui constituent le relais
pour le message nociceptif au niveau spinal : les neurones nociceptifs
spécifiques et les neurones non spécifiques
( nommés
aussi neurones convergents ou neurones polymodaux ou encore les neurones
à large gamme réceptive - « wide dynamic range neurones
» - WDR ).
Les neurones nociceptifs spécifiques sont le
constituant majeur des couches I et II. Ces neurones sont activés
exclusivement par des fibre afférentes fines C et Aä. Pour la
stimulation cutanée, ces fibres répondent uniquement aux
stimulations mécaniques ou thermiques intenses provenant d'un champ de
récepteurs restreint. Leur réponse s'accroît avec
l'intensité de la stimulation, ce qui leur permet de coder
l'intensité du stimulus nociceptif. Néanmoins, les couches
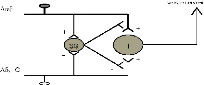
Fig 3. Gate control
Schéma de la théorie du gate control (ou
portillon) G - fibres de gros diamètre
P - fibres de petit diamètre
SG correspond a la couche de II de Rexed
T - une cellule de transmission
Ce schème permet d'expliquer comment une
stimulation sensitive permet de diminuer la sensation de douleur dans le
même zone
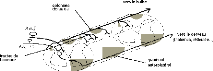
Fig 4. Voies ascendantes
:Représentation schématique de la distribution des fibres
afférentes primaires lors de leur arrivée dans la corne
postérieure et principales voies ascendantes empruntées par le
message nociceptif après relais dans la corne postérieure au
niveau des couches I, II et V.
superficielles sont formées pour une part non
négligeable par des neurones nociceptifs non spécifiques.
Les neurones nociceptifs non
spécifiques sont principalement localisés
dans les couches V-VI et autour du canal central (couche X), mais aussi, comme
il a été mentionné plus haut, dans la couche I.
Ces neurones sont activés par des
stimulations provenant de champs récepteurs assez
étendus, ils répondent à la fois à des stimulations
nociceptives et non nociceptives arrivant par des fibres Aâ, Aä et
C. Enfin, quelques neurones nociceptifs
spécifiques sont aussi présents dans
les
couches V-VI.
Il existe également des interneurones, qui communiquent
l'information, dans les deux sens, entre les couches superficielles et les
couches V-VI.
Voies ascendantes
Plusieurs observations ont très
tôt
suggéré que la douleur emprunte des voies
spécifiques dans la moelle pour atteindre les
structures supraspinales. Les patients souffrant de douleurs chroniques ont pu
être soulagés par des incisions pratiquées dans la moelle
(section du quadrant antérolatéral cf. Fig.4).
En effet deux voies spinales principales conduisent
l'informations nociceptives vers le cerveau : les voies spino-thalamique et
spinoréticulaire. Les projections collatérales que ces voies
distribuent dans des structures bulbaires mésencéphaliques et
diencéphaliques sont très nombreuses.
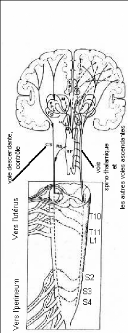
Fig 5. Voie emprunte par le message nociceptif
jusqu'aux centres supérieurs
Les centres supérieurs
Il a été observé que la stimulation
électrique de certaines zones dans le cerveau provoque chez l'animal des
comportements de fuite semblables à ceux obtenus par des stimulations
nociceptives périphériques. En revanche il est certain qu'il
n 'existe pas un centre de la douleur dans le
cerveau.
Les centres supérieurs ont pour fonction
d'intégrer le message nociceptif. C'est à ce niveau-là,
que l'on peut dire que le message nociceptif va, si toutes les conditions sont
réunies, être perçu comme de la douleur.
De nombreuses structures participent de l'intégration
du message nociceptif, dont les trois composantes principales sont :
Ø La composante sensori-discriminative qui
permet d'analyser la localisation, l'intensité et la durée de la
stimulation nociceptive ;
Ø La composante motivationnelle qui permet
d'attribuer son caractère désagréable, déplaisant,
insupportable à la sensation douloureuse ;
Ø La composante cognitive et évaluative
impliquée dans les phénomènes d'anticipation,
d'attention, de suggestion et d'expérience passée.
Le thalamus joue un rôle majeur dans l'analyse de
l'information nociceptive. De par sa partie
ventropostérolatérale, (VPL) il participe au décodage de
la composante sensoridiscriminative de la douleur, tandis que se partie plus
médiane (centralis lateralis, CL) en se connectant au cortex
moteur, participe plutôt à l'élaboration de comportements
moteurs associés à la sensation douloureuse.
La formation réticulée joue un rôle aussi
très important. Néanmoins les rôles exactes des
différentes structures qui composent la formation
réticulée sont encore très mal connus.
Parturition
Les mécanismes de la perception nociceptive sont
relativement bien connus lorsqu'il s'agit de la nociception cutanée ou
au niveau des muscles squelettiques. Les modèles de
référence, les plus étudiés, sont : l'inflammation,
et les stimulations mécanique et thermique. En ce qui concerne les
douleurs viscérales, les informations disponibles sont encore quasi
inexistantes. Ceci est d'autant plus le cas pour des phénomènes
aussi complexes que la parturition.
Le travail entrepris par l'équipe de Marie-Christine
Lombard a pour but d'apporter de nouveaux éléments sur les
mécanismes nociceptifs mis en jeu pendant la parturition. Plus
précisément, l'étude de la parturition chez le rat devrait
aboutir à l'établissement d'un modèle animal sur lequel on
pourra ensuite reproduire et étudier des situations susceptibles
d'induire une sensation douloureuse chez la femme parturiente.
Le rôle physiologique de la douleur est de donner le
signal du début du travail. Une fois ce rôle accompli on a tout
intérêt à, au moins, atténuer la douleur, car si
elle est particulièrement violente ou persistante, par les effets de
stress qu'elle déclenche, elle peut mettre en péril la
mère ou/et le foetus.
Jusqu'alors, les études n'ont
révélé aucune expression spontanée du c-fos (en
absence de stimulation nociceptive aiguë) en dehors des états
pathologiques de l'organisme ou des lésions de nerfs. La parturition est
la seule fonction physiologique qui, chez la femme, produit une douleur
viscérale intense et, chez les animaux, induit des manifestations
comportementales de douleur. La menstruation chez la femme est aussi, dans un
grand nombre de cas, ressentie comme un événement douloureux.
Le signal nociceptif responsable des douleurs
viscérales, dont les douleurs de l'accouchement font partie, a pour
origine les terminaisons nerveuses libres périphériques dans les
tissus utérins et dans les parois vaginales (Cervero F., Janig W.,
1992). Le fait que les viscéronocicepteurs périphériques
n'ont toujours pas été anatomiquement identifiés, et les
travaux étudiant les douleurs viscérales, nous amènent
à admettre qu'il s'agit probablement en grande partie de nocicepteurs
polymodaux non spécifiques, qui peuvent donc être activés
par une large gamme de stimuli (stimulations mécaniques, thermiques ou
chimiques).
Origine de la douleur
La parturition débute par l'apparition de contractions
coordonnées et rythmiques du muscle lisse utérin
(myomètre) qui succèdent aux contractions localisées et de
faible amplitude, caractéristiques de la gestation.
La parturition peut être définie comme
l'expulsion hors des voies génitales maternelles, du foetus et de ses
annexes.
La première phase correspond à
l'établissement d'une activité contractile du myomètre
impliquant des changements biochimiques du col utérin, aboutissant
à une dilatation complète de celui-ci.
La deuxième phase correspond à
l'expulsion du foetus rendue possible par la dilatation complète de
l'utérus. Les contractions utérines plus fortes expulsent alors
le foetus à travers le canal pelvien.
La troisième phase correspond à
l'expulsion des annexes foetales, facilitée par le processus des
dégénérescence placentaire et par les contractions
localisées du muscle utérin.
Pendant les phases de latence et les phases actives du
premier stade du travail, la douleur est essentiellement due aux contractions
utérines et à leur conséquences : dilatation du col
utérin et du segment inférieur de l'utérus (distension,
étirement et parfois déchirement de ces structures pendant les
contractions utérines).
Notamment :
La dilatation du col correspond à un étirement
des muscles lisses. Cette zone est très riche en fibres sensitives.
Ø Il y a une corrélation positive entre le
degré de dilatation du col et du segment inférieur et de
l'intensité de la douleur ressentie.
Ø Avant l'ouverture complète du col et
l'expulsion du liquide amniotique, le col forme une barrière ce qui
aboutit aux contractions de l'utérus dans les condition
isométriques ; La pression du liquide amniotique peut alors, en absence
de complications, atteindre 2 kPa.
Ø Lorsque le col est complètement dilaté
les parturientes ressentent une douleur en tout point similaire à celle
ressentie pendant une contraction utérine.
Pendant le second stade du travail. Les sources de la douleur
de la première phase persistent alors que de nouvelles viennent s'aj
outer. Par exemple, les tiraillements ou la compression de la paroi
péritoineale et des organes qu'elle enveloppe : les ligaments
utérins, la vessie, l'urètre, du rectum ou des ligament et
muscles de la cavité pelvienne. Cette douleur est plus intense que la
douleur ressentie au premier stade de l'accouchement avec un pic correspondant
à l'expulsion du foetus.
Les voies de la douleur et la distribution lombaire des
neurones nociceptifs
Il a longtemps été admis, et on peut encore
trouver ces données dans des publications récentes, que les nerfs
qui conduisent le message nociceptif en provenance du corps d'utérus
suivent les fibres sensorielles qui accompagnent le nerf pelvien, et se
projettent dans les segments T1 1 et T12 ; tandis que les fibres qui innervent
la partie basse de l'utérus et le col de l'utérus viennent
rejoindre la moelle au niveau S2, S3 et S4 (Gray 1973, 1980 ; Romanes 1972 ;
Williams 1985 cf Bonica). (voir fig 5. page5)
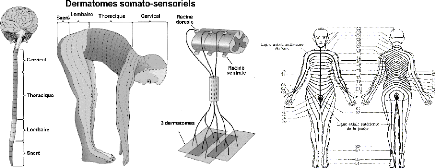
Fig 6. Dermatomes - Chaque racine sensitive regroupe des
fibres qui peuvent venir de la peau, des muscles et des viscères. Un
dermatome est le territoire cutané qui correspond à une racine
dorsale.
Selon les travaux de J.F. Bonica (Textbook of PAIN 1969, 1974,
1979) les segments mis en jeux sont le T1 0, T1 1, T1 2et L1, et tous les nerfs
en provenance de l'utérus accompagnent le nerf sympathique, alors que
les nerfs venant des structures périnéales contactent la moelle
épinière par les racines S2, S3 et S4. De plus les segments
lombaires inférieurs et les segments sacraux supérieurs envoient
quelques nerfs jusqu'aux structures pelviennes et sont ainsi impliqués
dans la nociception lors de la parturition.
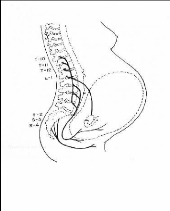
Fig 7.Schèma des principales voies nociceptives
sollicitées en cous de la parturition (selon Bonica 1994)
Comme toutes les douleurs viscérales, la douleur en
provenance de l'utérus vérifie le principe de la douleur
projetée. Ce phénomène est dû à la
ramification axonale ; il aboutit donc au fait que toutes les douleurs
viscérales sont référés au dermatome innervé
par les nerfs projetant vers le même segment lombaire que la structure
viscérale en question.
Changements physiologiques chez la mère et chez
le foetus: pendant la gestation et pendant l'accouchement
La gestation chez la femme dure en moyenne 40
semaines, les premières contractions commencent la
30éme semaine et leur intensité s'accroît
à mesure que le terme approche. Pendant la gestation, l'organisme de la
femme subit un ensemble de changements anatomiques et physiologiques qui
culminent au moment de l'accouchement. Parmi ces changements, le plus important
concerne le système respiratoire, mais on observe également des
changements neuro-endocrinologiques, cardiovasculaires,
métaboliques...
Modifications du système respiratoire
- On peut observer des modifications dans les voies respiratoires,
notamment la diminution du volume pulmonaire. On a parallèlement une
modification de la dynamique respiratoire et une augmentation du volume de la
ventilation.
Aux environs du 3ème mois de grossesse il y
a un accroissement de l'ordre de 50% du volume respiratoire du à une
augmentation spectaculaire de la ventilation alvéolaire (+70%).
L'ensemble de ces phénomènes combiné
à une diminution du volume des poumons aboutissent à une
diminution de la pression partielle du CO2 (pCO2) alvéolaire.
De surcroît, la douleur provoqué par les
contractions utérines au moment de l'acouchement, stimule fortement la
respiration. Il en résulte une augmentation nette du volume de
l'inspiration, un accroissement du taux respiratoire et par conséquent
une augmentation fulgurante de la ventilation alvéolaire. Comme
conséquence on a un écroulement de la paCO21 qui
atteint 16 à 20 mmHg et même peut descendre jusqu'au 10 à
15 mmHg. La paO22 croit légèrement pour atteindre 105
mmHg. Comme conséquence finale on a un accroissement du pH sanguin de la
mère, donc une alcalose respiratoire qui accompagne les vagues de
contractions utérines. Ces phénomènes respiratoires sont
accompagnés par une diminution du flux sanguin cérébral et
utérin et d'une augmentation de l'affinité de
l'hémoglobine pour l'O2 liée à l'augmentation du
pH3.
1 Pression partielle du CO2 artérielle et
alvéolaire
2 Pression artérielle partielle de l'O2 artérielle
et alvéolaire
3 Les variations de pH sanguin ou de pression partielle en CO2
provoquent des déplacements des courbes de dissociation de l'O2 dans le
sang. Une diminution de paCO2 ou une augmentation de pH provoque un
déplacement plus ou moins marqué de la courbe vers la gauche. Il
y a donc une diminution du relarguage de l'O2 au niveau tissulaire.
Entre les périodes de contractions, on assiste à
des périodes de repos lors desquels la femme a tendance à
hypo-ventiler. Cela provoque une chute de paO2 de 10 jusqu'à -50% ( -25
à -30% en moyenne)
Effets sur le foetus :
l'alcalose respiratoire de la mère lors des
contractions induit une baisse d'échanges placentaires
d'oxygène.
l'hypoxie chez la mère qui survient pendant les phases
de relaxation entraîne une hypoxie foetale.
Effets d'analgésie :
L'analgésie partielle par emploi d'opioides ou
l'anesthésie par voie respiratoire - On observe une diminution de
l'hyperventilation lors des contractions (paCO2= 20 - 25 mmHg et
l'oxygénation augmente). Une alcalose respiratoire persiste
malgré cette diminution, et si elle s'ajoute à l'effet
déprimant des opioides pendant les périodes de repos, elle peut
provoquer de l'hypoventilation et l'hypoxie.
L'analgésie épidurale, lorsqu'elle
supprime la douleur, elle abolit également l'hyperventilation
provoquée par celle-ci, et évite l'hypoventilation pendant les
phases de repos. La paCO2 reste voisine des valeurs normales (#
28-32 mmHg) et la paO2 reste également normale (# 100
mmHg).
Modification neuro-endocrinologiques -
Plusieurs études démontrent des changements hormonaux
significatifs.
Après une stimulation nociceptive aiguë chez
l'animal on enregistre une augmentation de catécholamines (+ 20 à
40%), en particulier en ce qui concerne le taux de norepinephrine, ceci
provoque une diminution de la circulation sanguine dans l'utérus(III -
p625).
Les études chez la femme ont montré que la
douleur et l'anxiété pendant la phase active du travail peuvent
induire 300 à 600% d'augmentation du taux d'épinephrine, 200
à 400% d'augmentation du taux de norépinephrine, et 200 à
300% d'augmentation de cortisol. Une augmentation significative de
corticostéroïdes et de ACTH a également été
perçue. Le pic de ces hormones est en général
enregistré pendant ou juste après l'expulsion du nouveau
née.
L'augmentation du taux d'épinephrine est associé
aux contractions utérines pendant la phase active du travail. Il a
été démontré une corrélation positive entre
le taux d'épinephrine, et l'intensité de la douleur, donc avec
l'anxiété induite par celle-ci.
Ohno et Coll. (1986) démontrent, lors d'une
étude sur les catécholamines et les nucléotides cycliques
plasmatiques pendant le travail, une corrélation positive entre d'une
part le taux d'épinephrine et le taux cardiaque et d'autre part la
corrélation entre les taux plasmatiques de la norépinephrine et
de l'AMPc.
Effets de l'analgésie par péridurale
En supprimant ou en atténuant la douleur, la
péridurale abolit les effets de celle-ci. Pendant le travail, les taux
de toutes les hormones dont la sécrétion est stimulé par
la douleur diminuent, notamment le taux de catécholamines, de
bêta-endorphines, de ACTH et cortisol
(III - p626). Cet effet est observé uniquement chez la
femme lors du travail. Chez la femme non parturiente, le même traitement
n'induit pas de diminution de taux hormonaux.
En ce qui concerne le foetus, l'analgésie
péridurale chez la mère pendant le travail n'induit pas de
diminution de catécholamines ni de bêta-endorphines.
Changements cardiovasculaires - A partir de 6
à 8 semaines de grossesse, le volume sanguin et le volume plasmatique
augmentent. Cette hausse atteint le maximum entre la 28ème et
le 32ème semaine, pour rester constante jusqu'à la
parturition.
Au moment de l'accouchement on observe un accroissement du
débit cardiaque Pendant les périodes de contractions
utérines cette hausse est encore plus importante.
Pour une part cette augmentation est due à la perte
sanguine au niveau utérin ( 250 à 300 mL ) et à
l'amplification du retour veineux de la cavité pelvienne et des membres
inférieurs. D'autre part, la hausse de la pression systolique et
diastolique est due à l'hyperactivité sympathique
provoquée par l'anxiété, la douleur et l'effort physique
pendant le travail.
L'effort du ventricule gauche est ainsi fortement
augmenté, d'où le risque pour les parturientes ayant une atteinte
cardiovasculaire.
Effets de l'analgésie
Comme pour les autres modifications, l'épidurale, en
réduisant ou en abolissant totalement la douleur supprime les
conséquences de celle-ci. Ainsi l'hyperactivité sympathique et la
réponse neuroendocrine sont fortement diminuées et la part
d'augmentation de la fréquence cardiaque et de l'accroissement de la
pression artérielle que celle-ci provoquent sont
éliminées. Plusieurs études(III p.628)
démontrent que l'analgésie péridurale diminue le
risque de complication chez les femmes ayant une att einte cardiovasculaire.
Effets sur le métabolisme- Pendant la
grossesse le niveau du métabolisme basal augmente (#20%) ainsi que la
consommation d'O2. Pendant le travail le taux plasmique d'acides gras libres et
la concentration du lactate augmentent significativement. Cette hausse est un
effet indirect de la douleur. Laquelle, comme ca a été dit plus
haut, induit une augmentation de la sécrétion de
catécholamines ; qui ont pour l'effet de stimuler le métabolisme,
que l'on perçoit par l'accroissement du taux des acides gras circulants
et du lactate.
Anesthésie et analgésie, fonctionnement,
méthodes actuelles, difficultés
L'objectif primaire de l'anesthésie et analgésie
lors de l'accouchement est une suppression optimale de la douleur avec un
risque minimal pour la mère et l'enfant. Parmi les différentes
méthodes de l'anesthésie, on compte :
1. l'aide et préparation psychologique
- Ce groupe inclut toutes les méthodes qui n'emploient pas
d'agents pharmacologiques. Dans cette classe on trouve des techniques aussi
diverses que l'information et « éducation » de la mère
ou des
deux parents, la gymnastique prénatale ou des
techniques qui impliquent l'hypnose ou des séances de relaxation. Toutes
ces méthodes aboutissent à une diminution de
l'anxiété et de l'appréhension, ce qui permet à la
parturiente de mieux contrôler son comportement et de collaborer plus
aisément avec l'équipe d'obstétriciens. En appliquant ces
méthodes #5 à 10% de femmes ressentent peu ou pas de douleur et
l'administration de analgésiques ou anesthésiques n'est pas
requise. Chez 15 à 20% de parturientes la douleur est réduite
à un niveau modéré, et l'emploi de doses plus faibles
d'analgésiques est requise. La part restante des parturientes
n'éprouve aucune réduction de la douleur, mais
l'anxiété étant réduite, elles manifestent moins de
signes comportementaux de la douleur. Même si quelques études
montrent que les parturientes qui ont suivi ce genre de préparation ont
un travail moins long, ont moins de complications avant ou après la
parturition, moins de pertes de sang et ont des bébés « plus
heureux » que les parturientes à qui on a administré des
analgésiques ou anesthétiques, d'autres études ne montrent
aucune différence entre les parturientes qui ont été
préparées et celles qui ne l'ont pas été. On peut
estimer que la meilleure façon de faire est de combiner ces
méthodes avec l'anesthésie régionale.
2. les méthodes simples de analgésie
pharmacologique - Au début de la première phase la
douleur peut être limitée par la suggestion et administration de
sédatifs et de tranquillisants, mais pendant la phase active du travail
l'emploi d'opioïdes est souvent nécessaire. Les opioïoides
sont administrés par injection intramusculaire, ou par faibles doses en
intraveineux - parfois les injections sont faites par un dispositif
contrôlé par la parturiente. Lorsque les doses optimales sont
appliquées, il n'y a pas de dépression respiratoire significative
chez la mère mais peut apparaître une dépression
néonatale qui peut être minimisée. Cette méthode
permet de réduire la douleur modérée dans 70 à 80%
des cas, et la douleur sévère dans 35 à 60% des cas.
3. analgésie et/ou anesthésie par voie
respiratoire - L'analgésie par voie respiratoire est
très employée car elle est assez efficace et produit une relative
diminution de la douleur sans provoquer la perte de conscience et sans induire
de dépression significative chez la mère ou chez l'enfant.
L'anesthésie générale pendant
l'accouchement est encore employée malgré les gros risques
qu'elle comporte. Les raisons pour lesquelles elle est encore pratiquée
sont la rapidité relative de son induction, le contrôle facile de
la durée et de l'intensité de la sédation, et
l'élimination rapide des effets de l'anesthésie après la
fin de l'accouchement.
Compte tenu des risques l'anesthésie
générale ne devrait être appliquée que dans les cas
où aucun autre moyen de soulager la douleur n'est applicable, et dans ce
cas la présence d'un anesthésiste compétent est
impérative.
4. l'analgésie loco-régionale -
Plusieurs techniques existent :
anesthésie locale du col de l'utérus et du
segment bas de l'utérus - blocage des voies paraviscérales ;
blocage des nerfs innervant les organes génitaux
externes
La plus employée et la plus efficace est l'injection
lombaire péridurale. Nous allons commenter que cette technique,
puisqu'elle nous intéresse plus particulièrement - notre objet
d'étude étant en particulier les voies nociceptives spinales.
Avantages :
Cette technique supprime très efficacement la douleur.
En bloquant les voies nociceptives, elle abolit également les effets que
celle-ci peut provoquer dans les systèmes respiratoire, cardiovasculaire
etc. ... Lorsque les doses bien ajustées d'anesthésiques sont
appliquées, on obtient le blocage des fibres Aä et C
avec peu ou pas d'effets sur les fibres plus grosses : somatomoteurs et
tactiles. Cette méthode, bien appliquée, ne provoque pas de
dépression des fonctions de l'organisme, ni chez la mère, ni chez
le nouveau-né.
Inconvénients :
L'application de ces techniques est plus complexe, elle
demande une excellente connaissance de l'anatomie et, en général,
elle demande plus de compétences que les autres techniques. De ce fait,
elle est appliquée uniquement au sein d'un hôpital.
Complications :
Hypotension chez la mère - elle est due au
blocage des fibres vasomotrices et à la vasodilatation qui en
résulte. Cette hypotension est contrée par l'infusion d'un litre
de liquide avant d'appliquer l'anesthésie.
Réaction toxique systémique - en cas
d'injection accidentelle intraveineuse.
Anesthésie excessive ou totale - peut se
produire en cas d'injection de doses trop importantes ou en cas d'injection
accidentelle dans l'espace sous-dural de la dose d'anesthésique
prévue pour une injection épidurale. Des solutions techniques
pour résoudre ces problèmes existent.
Dans les années 50 et 60, la technique employée
aboutissait à un blocage des segments T9-10 au S5. Les fortes doses
d'anesthétiques locaux augmentaient le risque de réaction toxique
systémique, les autres risques étant également accrus.
Chez un nombre important de parturientes, après chaque injection
d'anesthétique, une baisse de l'activité utérine est
observée. Également une faiblesse ressentie au niveau des membres
inférieurs, ou même la paralysie de ceux-ci, ainsi que la
paralysie des muscles périnéaux peut survenir. Tous ces
changements ont pour effet de prolonger le second stade de la parturition et
même parfois amènent à l'utilisation des forceps.
Une amélioration est obtenue lorsque l'analgésie
est limitée aux segments T10-T1 1 pendant la première phase, et
étendue aux segments sacraux pendant la seconde phase du travail. Cette
technique n'est actuellement utilisée que chez les femmes
présentant une malformation foetale ou une complication qui demande un
type spécifique de l'analgésie, pouvant être obtenue
uniquement par cette technique.
Pour encore améliorer ces techniques, une connaissance
plus approfondie des mécanismes nociceptifs mis en jeu est
indispensable. La majeure partie des résultats obtenus
aujourd'hui provient des études cliniques.
L'établissement d'un modèle animal pourrait permettre
l'application de méthodes expérimentales nouvelles. Ceci signifie
que l'on s'attaque à une tache immense et inhabituellement abondante en
inconnu.
Matériel et méthodes
L'expérience à laquelle j'ai assisté lors
de mon stage consistait à comparer la quantité de
l'immunomarquage c-Fos chez les rates primipares et multipares. La
signification de ce marquage et le rôle de la protéine c-fos
seront discutés en premier. Ensuite, l'expérience en
elle-même sera décrite, et les résultats
commentés.
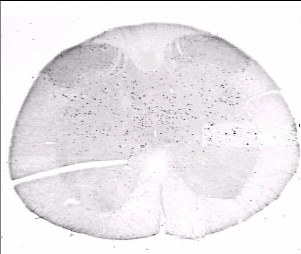
Fig 8. Marquage c-Fos sur une coupe du segment
L6
c-fos
Le gène c-fos fait partie
de la famille des gènes
d'expression précoce immédiate
(immediate-early genes, IEG), c'est un proto-
oncogène ubiquitaire. Le
produit de ce gène
est la
protéine c-Fos. Sa
surexpression dans le système nerveux peut être
induite par une stimulation pharmacologique, électrique ou
périphérique (thermique, mécanique ou chimique). C-
Fos est une phosphoprotéine qui est
généralement employée comme marqueur fonctionnel
d'activité des neurones spinaux et cérébraux
répondant, par exemple, à une stimulation nociceptive, ou pour
localiser les neurones activés par une stimulation donnée.
Le gène est activé très rapidement. Le
pic de l'ARN-m est mesuré 30 à 40 min après l'induction de
la transcription du gène et le pic de la protéine c-Fos
est mesuré 1 à 2h après celle-ci.
La protéine c-Fos se lie, immédiatement
après avoir été synthétisée, avec la
protéine c-Jun et le dimère se fixe sur le site
AP-1 (activator-protein-1) dans l'ADN pour induire la transcription des
gènes à expression tardive ( late response
genes)(III et VIII). D'autres combinaisons sont possibles :
homodimère Fos-Fos etc. Cette diversité aboutit à une
extrême finesse et variabilité de régulations.
Nombre d'études ayant pour objet l'expression spinale
du c-fos s'accordent sur l'existence d'une relation entre la
transmission de la nociception et l'expression spinale du c-fos.
Notamment, l'image qui nous est donnée par immunomarquage du c-Fos
de l'activité neuronale, s'accorde en général assez bien
avec les résultats établis par les études
électrophysiologiques ou par les traceurs anatomiques (marquage dans les
couches I, II, V et IV).
La protéine c-Fos, est-elle un bon marqueur de
l'activité nociceptive ?
La plupart des manipulations qui affectent des réflexes
nociceptifs et autres
comportements corrélés avec la douleur modifient
aussi l'expression du c-fos. Par exemple :
l'expression du c-fos est réduite par
l'administration d'analgésiques (morphine, indomethacine, ketoprofene).
De même, l'administration de substances qui interagissent avec la
transmission du signal au niveau spinal (noradrénaline, les antagonistes
du glutamate pour le récepteur à N-methyl-D-aspartate - NMDA)
ou de substances qui inhibent la NO-synthase, est capable de
réduire la synthèse de la protéine c-Fos.
l'expression du c-fos est inhibée par : la
stimulation électrique de la medulla rostro-
ventromédiane, l'activation de systèmes
endogènes d'inhibition de la nociception par
l'éléctroacupuncture, l'hypertension ou le stress
induit par l'exposition au danger.
Selon la nature de la stimulation nociceptive (chimique,
thermique ou mécanique), on observe une
hétérogénéité dans la distribution du
marquage, mais on n'est pas en mesure de dire si ces variations sont dues
à la nature (aspect qualitatif) des stimuli ou à la
différence des intensités relatives (aspect quantitatif) des
stimuli.
Cependant, une relation moins évidente existe entre les
signes comportementaux et l'expression du c-fos.
Presley[ cf. V] remarque que les rats auxquels on a
administré une forte dose de morphine, après l'injection d'une
substance irritante dans la patte (formalin test), ne
présentent pas de comportement caractéristique de la sensation de
douleur alors qu'un marquage significatif du c-Fos reste présent dans
les couches superficielles de la moelle épinière.
De même, l'administration de la cocaïne supprime
rapidement toute réaction de l'animal à la stimulation
douloureuse et n'a pas d'effet significatif sur l'expression spinale du
c-fos.
Harris(V) observe également une relation de
proportionnalité entre l'expression spinale du c-fos et la
concentration de la substance irritante injecté, alors qu'aucune
différence comportementale n'est observée pour les
différentes concentrations (formol dilué à 1.5% et
à 5%). Ceci peut être dû à la différence de
sensibilités des deux méthodes. On peut imaginer plusieurs autres
explications, qui ne seront pas développées ici, mais qui par
leur nombre rendent bien compte de la complexité du
phénomène qu'est la douleur.
Au vu de ces remarques, on peut déduire que
l'expression du c-fos dans une cellule ne signifie pas automatiquement
qu'il s'agit de nociception. Une immunoréactivité c-Fos peut
aussi refléter des processus qui ne contribuent pas directement à
la transmission nociceptive.
Avantages et inconvénients de la
méthode
Avantages :
L'immunomarquage du c-Fos est extrêmement précis.
On est en mesure de localiser précisément les cellules
marquées. De plus, l'analyse quantitative peut être faite
simplement par le comptage de noyaux marqués.
L'emploi de cette méthode permet de travailler sur des
animaux éveillés car l'usage d'anesthésiques n'est pas
indispensable comme lorsqu'on utilise l'éléctrophysiologie.
Le couplage de l'immunomarquage du c-Fos avec d'autres
techniques de marquage ou de traçage permet de caractériser les
neurones activés. De cette façon, il a été
démontré qu'une part importante de neurones exprimant le
c-fos, en réponse à une stimulation nociceptive,
expriment aussi c-Jun B et le gène de préprodynorphine.
Il a également été démontré que les neurones
immunoréactifs pour c-Fos envoient des projections vers les structures
supraspinales.
Inconvénients :
La stimulation capable d'induire l'expression du c-fos
doit être très forte et assez longue.
Comme il a été mentionné plus haut, il
n'y a pas de rapport direct entre les signes comportementaux de la douleur et
l'expression du c-fos. On est en mesure de supprimer les signes
comportementaux par administration d'analgésiques, alors qu'un marquage
c-Fos non négligeable persiste.
De plus, tous les neurones, lorsqu'il sont activés
n'expriment pas c-fos. Ceci nous oblige à faire très
attention lors de l'interprétation des résultats. L'absence de
marquage ne correspond pas forcément à une absence
d'activité. De même, la présence d'un immunomarquage c-Fos
n'est pas le signe direct de l'activité nociceptive.
Il est touj ours difficile de déterminer quel est
l'événement exact responsable de l'induction du c-fos.
Par exemple, un marquage c-Fos peut être induit par le stress
provoqué lui-même par la douleur. Il peut correspondre
également à l'activité locomotrice en réponse
à la stimulation nociceptive, ou même, être induit par
l'interruption du sommeil4.
Les protocoles expérimentaux doivent, donc, être
construits avec soin et comporter de nombreux groupes témoins afin
d'éviter une mauvaise interprétation des résultats.
Pour réellement comprendre ce qui se produit, il est
surtout indispensable de comprendre le rôle et le mode d'induction du
c-fos.
4 Le rat étant un animal nocturne, toutes les
expériences effectuées le jour perturbent son cycle de
sommeil.
Induction du c-fos
Selon les études menées sur les cultures
cellulaires, il existe deux voies d'induction du cfos. L'une met en
jeu l'activation de inositol-phosphate-protéine-kinase-C (SRE -
serum response element), et l'autre une augmentation de la concentration
intracellulaire du Ca2+.
Il a été établi que les protéines
provoquant l'induction du c-fos sont activées par une
transformation post transcriptionelle (phosphorylation), ce qui peut expliquer
la rapidité de l'induction du c-fos.
Une expérience (menée par HUGHES ET DRAGUNOW(cf.
V), 1995) établit qu'une inhibition de
la synthèse protéique aboutit à une
induction fulgurante du gène c-fos. Cela fait penser que les
protéines synthétisées de novo sont
nécessaires pour inhiber la transcription du c-fos. Et c'est
justement le produit du gène c-fos, la protéine c-Fos,
qui inhibe sa propre transcription ( trans-répression). Ce
mécanisme de trans-répression ne concerne que la voie SRE.
L'existence de régions non traduites, riches en AT dans
l'ARNm du c-fos se traduit par une instabilité de la
molécule, donc par une demi-vie courte. De cette façon, la
régulation est facilitée et peut être d'autant plus facile
et rapide.
Une fois synthétisée, la protéine c-Fos
se couple avec la protéine c-Jun à l'aide de séquences
« leucine zipper » pour former un hétérodimère
qui se lie au site activateur AP- 1 sur l'ADN, et active la transcription des
gènes cibles.
Plus globalement, c-Fos est impliquée dans une cascade
de signaux de transduction - provoquée par des événements
extracellulaires - qui est responsable de changements intracellulaires à
long terme.
Le rôle de la cascade de transcription dont c-Fos fait
partie, pourrait être de modifier la réponse nociceptive en
induisant des changements dans les voies nociceptives spinales qui aboutissent
à une augmentation de la sensibilité à la stimulation
nociceptive (hyperalgie), ou à une réponse de type nociceptif
à une stimulation non nociceptive (allodinie) (Zimmermann &
Herdegen, 1994). Ces conclusions proviennent des expériences montrant
une surexpression spinale du c-fos après les manipulations
provoquant une hyperalgie ou une allodinie5, et de celles qui
montrent que cette expression du c-fos est réduite après
traitement par les antagonistes des récepteurs NMDA ainsi que par les
inhibiteurs de la NO-synthase6.
Une autre façon dont Fos peut agir implique un
processus contraire : l'inhibition de la transmission nociceptive dans la
moelle épinière. Les événements provoquant une
hyperalgie ou une allodinie induisent l'expression de la dynorphine, un peptide
opioïde ayant des effets analgésiques connus, dans la partie
dorsale de la moelle7. Le double marquage montre la co-localisation
de la dynorphine et de la c-Fos pour plus de 80% de neurones marqués
positivement au c-Fos, après une stimulation nociceptive inflammatoire.
L'ordre d'apparition des deux protéines suggère que l'expression
de la préprodynorphine
5 (Hunt, Pini & Evan 1987 ; Ma. Q. P. & Woolf, 1996 ;
Menetrey, Gannon, Levine &Basbaum, 1989 ; Noguchi, Kowalski, Traub,
Solodkin, Iadarola & Ruda, 1991 ; Presley, Menetrey, Levine & Basbaum,
1990 ; Williams, Evan & Hunt, 1990)
6 (Chapman, Honoré, Buritova & Besson, 1995 ;
Kenl,Gogas, Lichtblau, Pollock, Mayers, Basbaum & Wilcox, 1991 ; Roche,
Cook, Wilcox & Kajander, 1996)
7 (Iadarola, Douglass, Civelli & Naranjo, 1988 ;
Iadarola,Brady, Draisei, & Dubner, 1988 ; Millan.M.J., Millan.M.H.,
Pilcher, Czlonkowski, Herz & Colpaert, 1985, 1986 ; Ruda, Iadila, Cohen
& Young, 1988 ; Weiche, Millan.M.J. Holt, Nohr & Herz, 1989)
pourrait être régulée par la c-Fos. Les
arguments directs confirmant cette théorie ont été
apportés par Hunter et al. (1995) qui a constaté que les rats
chez qui on a bloqué spécifiquement la synthèse de la
protéine c-Fos par injection d'oligo-désoxynucleotides antisens
ARNm du c-fos, lorsqu'ils ont reçu une stimulation nociceptive
(par injection de formol dilué dans la patte), ne synthétisent
plus d'ARNm de la préprodynorphine. On observe également une
diminution du seuil de la réponse à la douleur lors du test du
formol.
Fos apparaît donc comme étant impliquée
dans un mécanisme d'inhibition de la transmission nociceptive lors d'une
stimulation intense ou prolongée.
Expression spin ale de la protéine c-Fos lors de la
parturition chez le rat, mise en évidence par l'immuno-marquage
Etude préliminaire - Expression neuronale de la
protéine c-Fos dans la moelle épinière lors de la
parturition chez la rate primipare et multipare
L'expérience qui a précédé celle
à laquelle j'ai assisté avait pour but d'étudier
l'expression de la protéine c-Fos au niveau de la moelle
épinière chez la rate primipare et multipare. Voici un bref
résume de cette expérience. Les mêmes techniques ont
été utilisées dans l'experience qui l'ont suivi. Les
techniques seront donc détaillées dans la description de
celle-ci.
A J-2 1 de gestation, les rates reçoivent oralement une
dose de 5mg/kg de mifepristone ( RU-486 ) ce qui provoque une «
synchronisation » des naissances. Chez les primipares, 3 délais
après l'expulsion du premier petit ont été
étudiés : 1, 2 et 4h. Chez les multipares le délais de 2h
a été choisi, car les études préalables ont
montré que ce délai correspond à une forte l'expression de
c-Fos.
Au délai choisi, la rate est anesthésiée
et perfusée. Les segments T12 à S2 sont prélevés et
préparés selon les méthodes classiques pour la coupe au
microtome. Les coupes flottantes de 40 um sont traitées par la
méthode immunohistochimique indirecte (Avidine/Biotine). Les neurones
immunopositifs (F+) sont ensuite comptés dans 6 à 10 coupes, non
sériées, par segment.
Résultats : Chez les primipares, des neurones F+
sont présents bilatéralement dans tous les segments
étudiés. Ils prédominent en L5-S1
où le SPN8 présente un forte densité
de neurones F+. La distribution laminaire varie le long de la moelle
épinière. L'administration du RU-486 n'a pas d'effet significatif
sur l'expression de la protéine c-Fos. Les neurones F+ sont
présents dans toutes les couches. Ils sont les plus nombreux dans les
couches ventrales et leur distribution est la suivante :
Couches ventrales > couches V -VI > III - IV > X >
I - II
Quelques moto-neurones du VM sont F+ en L4-L5. Le nombre total
de neurones F+ n'est pas significativement différent entre 1h et 2h,
mais diminue significativement à 4h.
Expression neuronale de la protéine c-Fos dans la moelle
épinière chez la rate
8 noyau parasympathique spinal
Animaux
Les expériences ont étés
réalisées sur des rats femelles adultes de souche SpragueDawley
de 270g (témoins non gestantes), 370g (primipares) et 450g (multipares,
4 à 5 parturitions). Les rates gestantes ont été
accouplées chez le fournisseur (Charles River). Dans le laboratoire, les
animaux ont été gardés dans une pièce isolée
dédiée exclusivement aux femelles. Les animaux ont à leur
disposition de la nourriture et de l'eau en permanence, et sont maintenus une
animalerie thermostatée et dans laquelle la période
d'éclairement est de 1 2h.
Protocole expérimental
Quatre groupes expérimentaux (n=4 à 6) ont
été étudiés :
Les deux premiers groupes sont formées pas les rates
gestantes :
1. Rates primipares gestantes traités au RU-486 ( per
os - 5mg/kg dans une solution de carboxyl-méthyl-cellulose, 1ml/kg). Ce
traitement permet de synchroniser les naissances, elles ne sont pas
provoquées car elles ont eu lieu à terme. L'expérience est
alors envisageable car ainsi les mises bas sont regroupées dans une
période de quelques heures9. Trois sous-groupes ont
été formés pour étudier 4 périodes avant ou
après la mise bas:
· trois heures avant la mise bas (estimé)
· immédiatement après l'expulsion du premier
petit
· 2 heures après l'expulsion du premier petit
· 4 heures après l'expulsion du premier petit
trois groupes témoins existent également :
2. les rates témoins gestantes perfusées avant la
mise bas
3. et 4. deux lots de rates non gestantes de même
âge : traitées au RU-486 et traitées uniquement avec le
véhicule dans les conditions identiques.
J-2 1
Dans la matinée, on administre aux rates, par voie
orale, le RU-486, afin de synchroniser les naissances. Un groupe témoin
a été constitué afin de démontrer que cette
opération ne provoque pas de marquage c-Fos (groupe « gestantes
traitées véhicule uniquement »).
J-22
Les deux groupes témoins ; gestantes et non gestantes
(traitées ou non par le RU-486) sont perfusées. Pour les animaux
gestants, on compte le nombre d'embryons par branche de l'utérus.
9 L'expérience a exigé un travail de plus de 1 9h
dan une seule journée, pour cette raison la synchronisation des
naissances est indispensable
Perfusion
Les rates sont sacrifiés et anesthésiées
par une injection intra-péritonéale de penthiobarbital sodique
(55 mg/kg, Sanofi), puis perfusées via l'aorte avec 300 ml de tampon
phosphate salin (PBS, tampon phosphate 0,1M, pH=7,4, chlorure de sodium 0,9%)
afin d'éliminer le sang et 500 ml de paraformaldéhyde (Fluka)
à 4% dans du PBS afin de fixer les tissus.
La moelle épinière et le cerveau sont ensuite
prélevés et post-fixés 4 heures dans le même
fixateur, puis cryoprotégés 24 heures à 4°C dans une
solution de saccharose à 30% (D (+) sucrose, Fluka) dans du tampon
phosphate (PB) (0,1 M, pH=7,4).
Coupes frontales de moelle épinière
Les segments lombaires T12 à S2 sont isolés et
nettoyés de leurs racines dorsales et ventrales, puis coupés
à l'aide d'un microtome à congélation en coupes frontales
sériées de 40 um recueillies dans du PB.
Immunohistochimie sur coupes flottantes
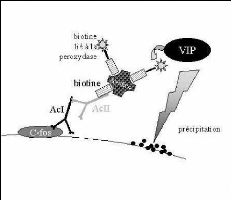
Fig 9: Schéma de la réaction
immunohistochimique - ABC (avidine/biotine)
Les coupes sont incubées pendant 60 minutes dans du
sérum normal de chèvre à 3% (NGST, Gibco BRL) dilué
dans du PBS contenant 0,3% de Triton X-100 (Prolabo). Cette étape permet
de bloquer les sites antigéniques non spécifiques susceptibles
d'être reconnus par l'anticorps secondaire (obtenu chez la
chèvre). Le Triton X-100 est un détergent qui perméabilise
les membranes et permet ainsi une meilleure pénétration des
différents réactifs.
Puis, les coupes sont incubées avec l'anticorps
primaire (dilué dans du NGST 1%) pendant 20 heures à 4°C.
Le lendemain, après 2 rinçages dans du
NGST 1%, les coupes sont incubées avec l'anticorps
secondaire biotinylé (anticorps anti-lapin obtenu chez la chèvre,
Laboratoires Vector) dilué dans du NGST 1%, pendant 1 heure, sous
agitation, à température ambiante.
Puis, après 2 rinçages au NGST 1%, le complexe
ABC-HRP (kit Vectastain, laboratoires Vector), dilué dans du PB 0,1M,
est ajouté et laissé incuber 1 heure sous agitation à
température ambiante.
La dernière étape (réaction enzymatique)
se fait par ajout de VIP.
La réaction est stoppée par plusieurs
rinçages successifs dans du PB 0, 1M.
Les coupes sont ensuite montées sur des lames
gélatinées, puis séchées à l'étuve
à 37°C pendant au moins 30 minutes.
Une déshydratation est effectuée dans des bains
successifs d'éthanol à 75%, 90 % et absolu.
Après un dernier bain dans du toluène, les lamelles
sont montées sur les lames avec de l'Eukitt (Baume d'inclusion pour
techniques histologiques, O.Kindler GmbH & Co.).
Analyses des résultats et statistiques
Les coupes sont regardées au microscope optique,
objectif 10. Les noyaux marqués sont dessinés grâce
à une chambre claire, sans tenir compte de l'intensité du
marquage. Ils sont ensuite comptés à l'intérieur de six
zones délimitées arbitrairement :
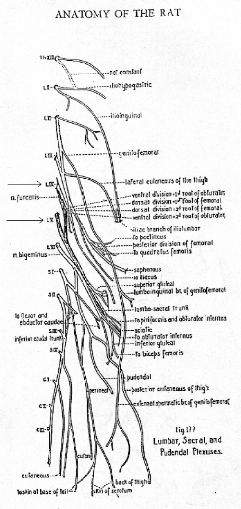
Fig10 : Schéma du système nerveux
périphérique du rat. Les nombreuses ramifications expliquent
comment une stimulation nociceptive localisé peut engendrer du marquage
Fos dans de nombreux segments.
> zone 1 : les couches superficielles (couche I et couche
II)
> zone2 : les couches III et IV
> zone3 : les couches V et VI
> zone4 : la partie ventrale (couches VII,VIII et corne
ventrale)
> zone5 : la couche X
> zone6 : le noyau parasympathique spinal (SPN)
Le comptage des neurones c-Fos est effectué sur 5
coupes par segment, non consécutives et représentatives du
marquage. Cette analyse est réalisée sur les différents
groupes. Si les groupes sont homogènes, on réalise une ANOVA
suivie d'un test de Fisher (PLSD, logiciel Statview 4.0).
Résu ltats
Ø Le RU-486 n'a pas d'effet significatif sur l'expression
spinale de la protéine c-Fos dans les segments T12 à S2 quelque
soit le délai.
Ø Les segments où l'expression du c-Fos est
maximale sont L5 et L6. Ces segments correspondent aux afférences
principales du nerf pelvien (voir fig.13).
Ø La distribution au cours du temps du marquage est la
suivante :
témoins < -3h < 0 h < 1h < 2h
> 4h
Ø Dans tous les segments la distribution du marquage par
couche est la suivante : Couches ventrales > couches V -VI > X > II-IV
<I-II Conclusion :
La parturition induit l'expression du c-fos dans les
régions de projection des afférences nociceptives. La
distribution du marquage laisse supposer que la composante nociceptive est
surtout mediée par le nerf pelvien en provenance du corps de
l'utérus et du col de l'utérus.
Compte tenu du temps nécessaire pour l'induction et la
synthèse de la protéine C-Fos, le marquage à 1h et
à 2h est probablement dû aux stimuli qui ont lieu avant et pendant
l'expulsion du premier petit.
Ces premiers résultats suggéreraient que lors de
la parturition chez le rat les segments spinaux qui reçoivent des
stimulations nociceptives sont ceux qui reçoivent les afférences
de l'utérus, du segment inférieur de l'utérus, de la
vessie et du rectum.
Il n'est pas démontré qu'il s'agit forcement des
afférences nociceptives. Mais la présence des neurones
immuno-positifs dans les couches I-II, V-VI, X et du SPN10 le
suggèrent fortement (cf. Fig. 12).
10 noyau parasympathique spinal
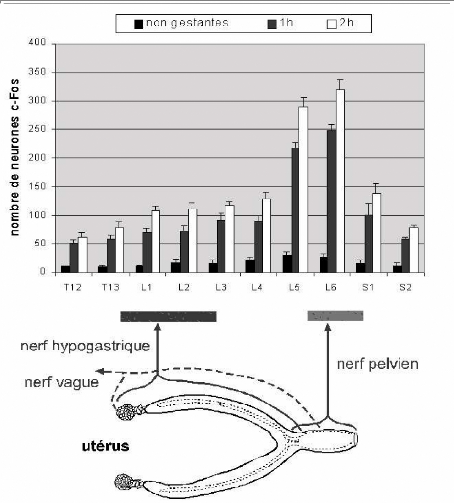
Fig. 13 : Graphique supérieur
: Distribution du nombre moyen par coupe de 40 um des neurones
exprimant la protéine c-Fos dans la moelle épinière chez
des rates non gestantes et des rates parturientes à 1 heureet 2 heures
aprèla naissane du premier petit.
Fig.12 : Répartition des
neurones c-fos en fonction des
Schéma inférieur : projection
des afférences sensitives issues de l'utérus.
Il est à noter le pic de l'expression de la
protéine c-Fos dans les segments L5 et L6, qui correspondent aux
afférances du nerf pelvien.
L'expression c-fos ets maximale à 2h, elle est
également très forte à 1h. Compte tenu du délais
d'expression de la protéine, on peut admettre que celle-ci est induite
par les contractions utérines qui précédent l'expulsion du
premier petit.
Epilogue
Les travaux décrits ont été
effectués en 1999. Ils font partie d'une recherche plus globale. En
fait, certaines données de la litt érature suggèrent que
dans la période de périparturition, l'intégration du
message afférent nociceptif est modifiée. En effet, des
études comportementales ont montré qu'en fin de gestation, les
seuils de réponse à des stimulations nociceptives étaient
élevées chez la rate (Gintzler, 1980), la truie (Jarvis et al,
1997) et également chez la femme (Iwasaki et al., 1991; Cruz et al.,
1996). Il semble que ces modifications soient en partie liées au
système opioïde endogène. Sachant qu'un des sites d'action
majeur des opioïdes est la moelle épinière (rappelons que
les couches superficielles de la moelle épinière sont
considérées comme le site de projection des afférences
nociceptives et que la présence de nombreux récepteurs
opioïdes à ce niveau a été démontrée),
il est possible que cette plasticité d'intégration du message
nociceptif ait lieu dès ce niveau. De manière surprenante, aucune
étude anatomofonctionnelle n'a été réalisée
au niveau du premier relais de la transmission des informations nociceptives,
que constitue la moelle épinière. Ce projet de recherche
s'insère donc dans une étude globale s'intéressant
à ce que nous pourrions appeler la mise en place d'une plasticité
physiologique au niveau de la moelle épinière.
Je vais succinctement décrire les travaux récents,
qui même s'ils ont été effectués après mon
stage permettent de mieux comprendre la finalité des travaux auxquels
j'ai pu assister.
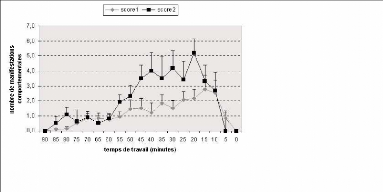
Figure 14 : Représentation du nombre de
manifestation de score 1 et de score 2 en fonction du temps. Le temps 0
correspond à l'arrivé du premier petit.
Etude comportementale
Les rates non gestantes et préparturientes ont
été observées et filmées afin de définir les
comportements caractéristiques. Deux types de comportement ont
été observés chez les rates préparturientes alors
qu'ils ne l'étaient pas pour les rates non gestantes. Il s'agit de
l'étirement d'une des deux pattes postérieures (score 1) et de
l'aplatissement du bassin accompagné de l'étirement des pattes
postérieures (score 2). Soulignons, que ces deux manifestations
comportementales sont des postures qui ont été décrites
dans des modèles de douleur viscérale, tels que le modèle
induit par injection intrapéritonéale d'acide acétique
(Schmauss et Yaksh, 1984), par implantation de calculs artificiels dans
l'uretère (Giamberardino et al., 1994), et après inflammation des
cornes utérines (Wesselmann et al., 1998).
Pour définir les effets de différents agents
pharmacologiques sur le déroulement de la parturition, « le
temps de travail » a été arbitrairement défini
comme le temps entre le premier mouvement stéréotypé t
l'arrivée du premier petit.
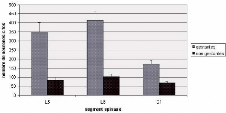
Fig 15: Distribution des neurones c-Fos en fonction des
segments spinaux L5, L6 et S1 chez des rates gestantes et non
gestantes.
Immunohistochimie de la protéine c-Fos
Cette étude comportementale a été
menée en parallèle avec une étude de l'expression spinale
du c-fos. En effet après avoir été
observées toutes les rates ont été sacrifiées,
perfusées, et les segments L5, L6 et S1 ont été
prélevés (comme il a précédemment été
démontré, c'est dans ces segments que l'on enregistre un pic
d'expression de la protéine c-Fos).
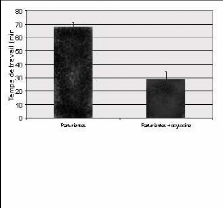
Fig. 16: Temps de travail moyen mesuré chez des
rates parturientes et chez des rates parturientes ayant reçu de
l'ocytocine en souscutané.
Effets de l'ocytocine en sous-cutané sur le
comportement accompagnant la parturition
L'ocytocine (10 ug / kg) a été injectée
en sous-cutané et les résultats ont été
comparés avec une injection de sérum physiologique. Les scores 1
et 2, ainsi que le score total ont été comparés pour les
deux groupes, et il a été démontré que l'ocytocine
ne modifie pas le nombre total des manifestations
comportementales ni leur proportion
respective. En revanche
une réduction
significative du temps du travail a
été
observée après l'injection de
l'ocytocine.
Etude des effets pharmacologiques de la morphine
administrée en péridurale sur la parturition
Les contrôles préliminaires
La morphine est administrée par voie péridurale.
Ainsi le modèle étudié est proche du modèle
clinique correspondant à un accouchement provoqué et sous
péridurale. Cette technique d'injection est la moins traumatisante
possible : elle permet en effet d'éviter des lésions de la moelle
et/ou des fibres afférentes primaires. Les difficultés de cette
approche résident dans le fait qu'elle a été très
peu utilisée chez le rat, seules trois publications font état de
la possibilité d'utiliser cette technique chez le rongeur (en
particulier Durant et Yaksh 1986) ; en conséquence il n'y a pas eu de
travail de référence portant sur les effets/doses de substances
telles que les opioïdes. Le choix des doses a donc été
difficile car les travaux qui ont porté sur les effets spinaux
d'analgésique ont utilisé une approche intrathécale. La
dose à administrer par voie péridurale chez les rates a
été choisie en se basant sur les nombreuses données
obtenues chez le rat par approche intrathécale et sur les données
fournies par la clinique.
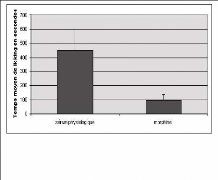
Fig.17: Représentation du temps moyen de «
licking » chez des rats ayant reçu du sérum physiologique ou
de la morphine en péridurale.
Afin de vérifier que la pose du microcathéter
n'engendre pas de gêne et que la morphine administré par ce moyen
là a l'effet souhaité, une batterie de tests a été
effectuée. De la morphine (à la dose 50 ug / 10ul) ou du
sérum physiologique ont été injectés par voie
péridurale, et suivis 15 minutes après par une injection
intraplantaire de 50ul de formol dilué à 5 % dans une patte
postérieure. Les animaux ont été observés pendant
60 minutes après l'injection de formol dilué.
Le comportement qui a été observé et
quantifié était le léchage de la patte enflammé
(licking).
Il a été constaté que l'injection de la
morphine en péridurale diminue très significativement le temps de
licking et également, une heure après l'injection de morphine, il
a été vérifié grâce à une table
à 45°, que la pose du cathéter et la morphine en
péridurale n'entraînaient pas de dysfonction motrice apparente.
Etude des effets pharmacologiques de la morphine
administrée en péridurale sur la parturition
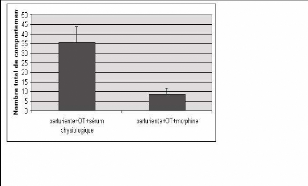
Fig. 18 : Représentation du nombre total de
comportement observés chez des rates ayant reçu soit du
sérum physiologique ou de la morphine par voie
péridurale
Les rates gestantes, dès l'apparition du premier
mouvement stéréotypé sont injectées par 10 ul de
sérum physiologique ou 10 ul
contenant 30 ug de morphine. A cette injection est
associée simultanément
une deuxième injection d'ocytocine en
sous-cutané à la dose 10 ug / kg.
Les scores 1 et 2 sont comptabilisés et l'on mesure
également le temps de travail. Il s'est avéré que le temps
de travail ne diffère pas significativement entre les deux groupes
d'animaux. Cependant dans le groupe ayant reçu de la morphine en
péridurale en plus de l'ocytocine en sous-cutané, le nombre des
comportements stéréotypés a significativement
diminué.
Discussion et conclusion
Il est a présent clair que la parturition provoque une
forte activité (que l'on visualise à l'aide du marquage de la
protéine c-Fos) dans les segments L5, L6 et S1 qui correspondent au site
de projection des afférences du tractus génital chez la rate.
Cette augmentation bilatérale concerne l'ensemble des couches de la
moelle épinière, y compris les couches superficielles (I, II),
les couches V-VI, la couche X, et le noyau parasympathique spinal (SPN). Ces
résultats suggèrent fortement qu'une partie de l'information
arrivant à la moelle pendant la parturition est de nature
nociceptive.
Les manifestations comportementales retenues sont
réellement reflet de contractions utérines, comme l'ont
montré les observations comportementales. En effet, le fait que
l'ocytocine, qui est un inducteur de fortes contractions utérines,
diminue le temps de travail sans réduire significativement le nombre de
comportements stéréotypés, laisse penser que les
comportements observés chez les rates parturientes sont en relation
étroite avec les contractions utérines (Fig. 14).
Ces manifestations comportementales pourraient, constituer
soit un réflexe moteur mettant en jeux les muscles squelettiques
abdominaux et facilitant l'expulsion du foetus, soit une réaction
nociceptive de l'animal à la sensation « douloureuse »
accompagnant la contraction utérine. Pour déterminer cela, les
effets de la morphine péridurale sur le temps de travail et sur le
nombre de manifestations comportementales avaient été
évalués.
L'administration péridurale de morphine diminue
significativement le nombre de manifestations comportementales sans modifier le
temps de travail (Figure 14). La disparition des séquences
comportementales n'affectent pas le délai d'expulsion du foetus, ce qui
suggère que ces manifestations ne sont pas nécessaires au bon
déroulement de la parturition, mais correspondent plutôt à
une réaction de l'animal à la sensation nociceptive accompagnant
la contraction utérine.
D'aucuns pourront remarquer que la façon dont
l'ocytocine a été injectée n'est pas très proche de
ce qui se produit in vivo. Il a été démontres que
lors de la parturition la dilatation du col utérin, en particulier lors
de l'expulsion du foetus stimule par voie réflexe les noyaux
hypothalamiques. Cette stimulation induit une synchronisation de
l'activité de l'ensemble des neurones, donnant lieu à une
activité pulsatile de libération d'ocytocine dans le
système sanguin (Belin. V et al, 1984), en parallèle à une
augmentation des récepteurs à ocytocine du myomètre
utérin, contribue au renforcement de la fréquence des
contractions utérines : c'est le réflexe de Ferguson. L'ocytocine
entraîne donc la contraction des fibres lisses du myomètre. Ces
contractions sont d'autant plus importantes en début de parturition,
dues à l'augmentation des récepteurs à l'ocytocine du
myomètre utérin (Fuchs et al, 1991). L'injection en
sous-cutané a été choisie pour des raisons de
commodité, car une injection intra-veineuse, mimant mieux la situation
naturelle, autait été beaucoup plus lourde techniquement et
traumatisante pour l'animal, car elle demande une intervention chirurgicale
préalable.
L'intérêt de ce modèle réside dans
le fait que la majorité des accouchements en clinique actuellement sont
provoqués (par administration d'ocytocine), et sont souvent faits sous
péridurale. En effet, très souvent, l'administration des
cocktails pharmacologiques périduraux (morphinique, anesthésique
local, agoniste á2 adrénergique) aboutit à une
anesthésie plutôt qu'à une analgésie. Les patientes
ne ressentent plus les contractions utérines, et ne peuvent donc pas
coordonner leurs efforts à la contraction. Une meilleure connaissance de
la mise en jeu des systèmes de contrôles endogènes
permettra peut être de proposer des substances plus spécifiques du
contrôle des douleurs associées aux contractions et d'adapter les
doses administrées. On pourrait ainsi améliorer le confort et la
participation active des patientes dans le processus d'expulsion.
Dans ce but, l'équipe de Marie-Christine Lombard
projette d'étudier le potentiel analgésique de substances
agissants au niveau des différents types de récepteurs
opioïdes en utilisant ce modèle.
Appendice
Préparation des réactifs pour la perfusion
1. Paraformaldehyde 4%
· preparation d'une solution de paraformaldehyde è
8% (pour préparer 1 L de solution)
80g de para
1L de PB 0,1
· protocole :
chauffer la quantité voulue de PB 0.1% à
70°C tout en agitant
peser le para sous hotte, puis verser dans le PB 0,1
préalablement chauffé.
laisser le mélange reposer jusqu'à ce qu'il
devient transparent
laisser refroidir
filtrer
dilution avant l'emplois (500mL de para à 8%
filtré + 500 mL de PB 0,1)
2. PBS
· 250 mL de PB 0,4
· 750 mL H2O distilée
· 9g de NaCl
Préparé la veille de la manipulation et
stocké à température ambiante 3. Sucrose 30%
· 150g de sucrose D
· 500 mL de PB 0,1
Préparation de réactifs pour immunohistochimie
1. PB 0,4
· 62,4g NaH2PO4 dans 1L d'H2O bi-distillée
· 356g Na2HPO4 dans 5L d'H2O bi-distillée
5L de Na2HPO4 + 0,9L de NaH2PO4 sont filtrés
2. PB 0,1
· 250 mL de PB 0,4 + 750 mL d'H2O distillé
3. PBS
· 1L de PB 0,1
· 9g d'NaCl
4. PBST
· 1L de PB 0,1
· 9g d'NaCl
· 3mL de triton 100
(conservation à 4°C)
5. NGST 1%
· 1mL NG (normal goat) dans 100mL de PBST
6. NGST 3%
· 3mL de NG dans 100 mL de PBST
7. Anticorps primaire
· Aliquots de 25 uL à 1\10 préalablement
préparés et congelés
· Dilution du AC I (anticorps primaire) à 1\7500
8. Anticorps secondaire (anti rabbit)
· l'AC II lyophilisé est dilué dans 1 mL
d'H2O distillée
· on dilue 4 gouttes de ACII dans 30mL de NGST 1%
9. VIP
· Dans 10 mL de PB 0,1 on mélange
3 gouttes de réactif 2
3 gouttes de réactif 3
3 gouttes d'H2O2
entre ces différents réactifs on ajoute 3 gouttes
du réactif 1
Bibliographie
I. CHAPRON Charles, BENHAMOU Dan, BELAISCH-ALLART Jöelle,
DUBUISSON Jean-Bernard -
La Douleur en Gynécolologie,
ARNETTE BLACKWELL 1996
II. MALTIER Jean-Paul, LEGRAND Chantal, BREUILLIER Michelle -
La Parturition from
DUPOUY Jean-Paul, Hormones et grandes
fonctions, ELLIPSES 1998
III. BONICA J. John - Labour Pain from Textbook of PAIN, third
edition edited by Patrick
D. Wall, Ronald Melzack, CHURCHILL LIVINGSTONE
1994
IV. CHAPMAN, J. -M. BESSON - Pharmacological Studies of
Nociceptive Systems Using the C-Fos Immunohistochemical Technique : An
Indicator of Noxiously Activated Spinal Neurones, from Handbook of Experimental
Pharmacology, Vol 130 The Pharmacology of Pain, Springer-Verlag 1997
V. Justin A. HARRIS - Using c-fos as a Neural Marker of Pain,
Brain Research Bulletin,
Vol 45, No 1, pp 1-8, 1998
VI. S. SIMONATO, G. BREGOLA, A. DONATINI, C. BIANCHI, L.BEANI,
S. FERRI and P. ROMUALDI -
Kindled Seizure-induced c-fos and Prodynorphin mRNA Expressions
are Unrelated in Rat Brain, European Journal of Neuroscience, Vol 8,pp.
2064-2067, 1996
VII. A. J. TODD, R. C. SPIKE, A. R. BRODBELT, R. F. PRICE and S.
A. S. SHEBAB - Some
Inhibitory Neurons in Spinal Cord Develop c-fos Immunoreactivity
After Noxious Stimulation, Neuroscience, Vol 63, No 3, pp. 805-816, 1994
VIII. BURITOVA Jaroslava - Physio-pharmacologie des processus
liés à l'inflammation produite par l'administration
intra-plantaire de caragenine, étudiés par l'intermédiaire
de l'expression de la protéine c-Fos au niveau de la moelle
épinière du rat, Thèse
IX. C. THIBAULT, M. -C. LEVASSIEUR - La reproduction chez les
mammifères et l'homme, Elipses-
Inra, 19??
X. M. NAVARRO - Mémoire de DEA : Etude de la
nociception lors de la parturition chez la rate : approches comportementale,
pharmacologique et immunohistochimique de l'expression spinale de la
protéine c-Fos ; Travail réalisé sous la direction des
Drs. M.C. Lombard et J.M. Besson, 2000
GNU Free Documentation License Version 1.2 November
2002
Copyright (C) 2000,2001,2002 Free Software Foundation, Inc.
59 Temple Place, Suite 330, Boston, MA 02111-1307 USA Everyone is
permitted to copy and distribute verbatim copies of this license document, but
changing it is not allowed.
0. PREAMBLE
The purpose of this License is to make a manual, textbook, or
other functional and useful document "free" in the sense of freedom: to assure
everyone the effective freedom to copy and redistribute it, with or without
modifying it, either commercially or noncommercially. Secondarily, this License
preserves for the author and publisher a way to get credit for their work,
while not being considered responsible for modifications made by others.
This License is a kind of "copyleft", which means that
derivative
works of the document must themselves be free in the same sense.
It complements the GNU General Public License, which is a copyleft license
designed for free software.
We have designed this License in order to use it for manuals for
free software, because free software needs free documentation: a free program
should come with manuals providing the same freedoms that the software does.
But this License is not limited to software manuals; it can be used for any
textual work, regardless of subject matter or whether it is published as a
printed book. We recommend this License principally for works whose purpose is
instruction or reference.
1. APPLICABILITY AND DEFINITIONS
This License applies to any manual or other work, in any medium,
that contains a notice placed by the copyright holder saying it can be
distributed under the terms of this License. Such a notice grants a world-wide,
royalty-free license, unlimited in duration, to use that work under the
conditions stated herein. The "Document", below, refers to any such manual or
work. Any member of the public is a licensee, and is addressed as "you". You
accept the license if you copy, modify or distribute the work in a way
requiring permission under copyright law.
A "Modified Version" of the Document means any work containing
the Document or a portion of it, either copied verbatim, or with
modifications and/or translated into another language.
A "Secondary Section" is a named appendix or a front-matter
section of the Document that deals exclusively with the relationship of the
publishers or authors of the Document to the Document's overall subject (or to
related matters) and contains nothing that could fall directly within that
overall subject. (Thus, if the Document is in part a textbook of mathematics, a
Secondary Section may not explain any mathematics.) The relationship could be a
matter of historical connection with the subject or with related matters, or of
legal, commercial, philosophical, ethical or political position regarding
them.
The "Invariant Sections" are certain Secondary Sections whose
titles are designated, as being those of Invariant Sections, in the notice that
says that the Document is released under this License. If a section does not
fit the above definition of Secondary then it is not allowed to be designated
as Invariant. The Document may contain zero Invariant Sections. If the Document
does not identify any Invariant Sections then there are none.
The "Cover Texts" are certain short passages of text that are
listed,
as Front-Cover Texts or Back-Cover Texts, in the notice that
says that the Document is released under this License. A Front-Cover Text may
be at most 5 words, and a Back-Cover Text may be at most 25 words.
A "Transparent" copy of the Document means a machine-readable
copy, represented in a format whose specification is available to the general
public, that is suitable for revising the document
straightforwardly with generic text editors or (for images
composed of pixels) generic paint programs or (for drawings) some widely
available drawing editor, and that is suitable for input to text formatters or
for automatic translation to a variety of formats suitable for input to text
formatters. A copy made in an otherwise Transparent file
format whose markup, or absence of markup, has been arranged to
thwart or discourage subsequent modification by readers is not Transparent. An
image format is not Transparent if used for any substantial amount of text. A
copy that is not "Transparent" is called "Opaque".
Examples of suitable formats for Transparent copies include plain
ASCII without markup, Texinfo input format, LaTeX input format, SGML or XML
using a publicly available DTD, and standard-conforming simple HTML, PostScript
or PDF designed for human modification. Examples of transparent image formats
include PNG, XCF and JPG. Opaque formats
include proprietary formats that can be read and edited only by
proprietary word processors, SGML or XML for which the DTD and/or processing
tools are not generally available, and the
machine-generated HTML, PostScript or PDF produced by some word
processors for output purposes only.
The "Title Page" means, for a printed book, the title page
itself,
plus such following pages as are needed to hold, legibly, the
material this License requires to appear in the title page. For works in
formats which do not have any title page as such, "Title Page"
means the text near the most prominent appearance of the work's title,
preceding the beginning of the body of the text.
A section "Entitled XYZ" means a named subunit of the Document
whose title either is precisely XYZ or contains XYZ in parentheses following
text that translates XYZ in another language. (Here XYZ stands for a specific
section name mentioned below, such as "Acknowledgements", "Dedications",
"Endorsements", or "History".) To "Preserve the Title"
of such a section when you modify the Document means that it
remains a section "Entitled XYZ" according to this definition.
The Document may include Warranty Disclaimers next to the notice
which states that this License applies to the Document. These Warranty
Disclaimers are considered to be included by reference in this License, but
only as regards disclaiming warranties: any other implication that these
Warranty Disclaimers may have is void and has no effect on the meaning of this
License.
2. VERBATIM COPYING
You may copy and distribute the Document in any medium, either
commercially or noncommercially, provided that this License, the copyright
notices, and the license notice saying this License applies
to the Document are reproduced in all copies, and that you add no
other conditions whatsoever to those of this License. You may not use technical
measures to obstruct or control the reading or further copying of the copies
you make or distribute. However, you may accept compensation in exchange for
copies. If you distribute a large enough number of copies you must also follow
the conditions in section 3.
You may also lend copies, under the same conditions stated above,
and you may publicly display copies.
If you publish printed copies (or copies in media that commonly
have printed covers) of the Document, numbering more than 100, and the
Document's license notice requires Cover Texts, you must enclose the copies in
covers that carry, clearly and legibly, all these Cover Texts: Front-Cover
Texts on the front cover, and Back-Cover Texts on the back cover. Both covers
must also clearly and legibly identify you as the publisher of these copies.
The front cover must present the full title with all words of the title equally
prominent and visible. You may add other material on the covers in addition.
Copying with changes limited to the covers, as long as they preserve the title
of the Document and satisfy these conditions, can be treated as verbatim
copying in other respects.
If the required texts for either cover are too voluminous to fit
legibly, you should put the first ones listed (as many as fit reasonably) on
the actual cover, and continue the rest onto adjacent pages.
If you publish or distribute Opaque copies of the Document
numbering more than 100, you must either include a machine-readable Transparent
copy along with each Opaque copy, or state in or with each Opaque copy a
computer-network location from which the general network-using public has
access to download using public-standard network protocols a complete
Transparent copy of the Document, free of added material. If you use the latter
option, you must take reasonably prudent steps, when you begin distribution of
Opaque copies in quantity, to ensure that this Transparent copy will remain
thus accessible at the stated location until at least one year after the last
time you distribute an Opaque copy (directly or through your agents or
retailers) of that edition to the public.
It is requested, but not required, that you contact the authors
of the Document well before redistributing any large number of copies, to give
them a chance to provide you with an updated version of the Document.
4. MODIFICATIONS
You may copy and distribute a Modified Version of the Document
under the conditions of sections 2 and 3 above, provided that you release the
Modified Version under precisely this License, with the Modified Version
filling the role of the Document, thus licensing distribution and modification
of the Modified Version to whoever possesses a copy of it. In addition, you
must do these things in the Modified Version:
A. Use in the Title Page (and on the covers, if any) a title
distinct from that of the Document, and from those of previous versions (which
should, if there were any, be listed in the History section of the Document).
You may use the same title as a previous version if the original publisher of
that version gives permission.
B. List on the Title Page, as authors, one or more persons or
entities responsible for authorship of the modifications in the Modified
Version, together with at least five of the principal authors of the Document
(all of its principal authors, if it has fewer than five), unless they release
you from this requirement.
C. State on the Title page the name of the publisher of the
Modified Version, as the publisher.
D. Preserve all the copyright notices of the Document.
E. Add an appropriate copyright notice for your modifications
adjacent to the other copyright notices.
F. Include, immediately after the copyright notices, a license
notice giving the public permission to use the Modified Version under the terms
of this License, in the form shown in the Addendum below.
G. Preserve in that license notice the full lists of Invariant
Sections and required Cover Texts given in the Document's license notice.
H. Include an unaltered copy of this License.
I. Preserve the section Entitled "History", Preserve its Title,
and add to it an item stating at least the title, year, new authors, and
publisher of the Modified Version as given on the Title Page. If there is no
section Entitled "History" in the Document, create one stating the title, year,
authors, and publisher of the Document as given on its Title Page, then add an
item describing the Modified Version as stated in the previous sentence.
J. Preserve the network location, if any, given in the Document
for public access to a Transparent copy of the Document, and likewise the
network locations given in the Document for previous versions it was based on.
These may be placed in the "History" section. You may omit a network location
for a work that was published at least four years before the Document itself,
or if the original publisher of the version it refers to gives permission.
K. For any section Entitled "Acknowledgements" or "Dedications",
Preserve the Title of the section, and preserve in the section all the
substance and tone of each of the contributor acknowledgements and/or
dedications given therein.
L. Preserve all the Invariant Sections of the Document,
unaltered in their text and in their titles. Section numbers or the equivalent
are not considered part of the section titles.
M. Delete any section Entitled "Endorsements". Such a section
may not be included in the Modified Version.
N. Do not retitle any existing section to be Entitled
"Endorsements" or to conflict in title with any Invariant Section.
O. Preserve any Warranty Disclaimers.
If the Modified Version includes new front-matter sections or
appendices that qualify as Secondary Sections and contain no
material copied from the Document, you may at your option designate some or all
of these sections as invariant. To do this, add their titles to the list of
Invariant Sections in the Modified Version's license notice. These titles must
be distinct from any other section titles.
You may add a section Entitled "Endorsements", provided it
contains nothing but endorsements of your Modified Version by various
parties--for example, statements of peer review or that the text has been
approved by an organization as the authoritative definition of a standard.
You may add a passage of up to five words as a Front-Cover Text,
and a passage of up to 25 words as a Back-Cover Text, to the end of the list of
Cover Texts in the Modified Version. Only one passage of Front-Cover Text and
one of Back-Cover Text may be added by (or through arrangements made by) any
one entity. If the Document already includes a cover text for the same cover,
previously added by you or by arrangement made by the same entity you are
acting on behalf of, you may not add another; but you may replace the old one,
on explicit permission from the previous publisher that added the old one.
The author(s) and publisher(s) of the Document do not by this
License give permission to use their names for publicity for or to assert or
imply endorsement of any Modified Version.
5. COMBINING DOCUMENTS
You may combine the Document with other documents released under
this License, under the terms defined in section 4 above for modified versions,
provided that you include in the combination all of the Invariant Sections of
all of the original documents, unmodified, and list them all as Invariant
Sections of your combined work in its license notice, and that you preserve all
their Warranty Disclaimers.
The combined work need only contain one copy of this License, and
multiple identical Invariant Sections may be replaced with a single copy. If
there are multiple Invariant Sections with the same name but different
contents, make the title of each such section unique by adding at the end of
it, in parentheses, the name of the original author or publisher of that
section if known, or else a unique number. Make the same adjustment to the
section titles in the list of
Invariant Sections in the license notice of the combined work.
In the combination, you must combine any sections Entitled
"History" in the various original documents, forming one section Entitled
"History"; likewise combine any sections Entitled "Acknowledgements", and any
sections Entitled "Dedications". You must delete all sections Entitled
"Endorsements".
6. COLLECTIONS OF DOCUMENTS
You may make a collection consisting of the Document and other
documents released under this License, and replace the individual copies of
this License in the various documents with a single copy that is included in
the collection, provided that you follow the rules of this License for verbatim
copying of each of the documents in all other respects.
You may extract a single document from such a collection, and
distribute it individually under this License, provided you insert a copy of
this License into the extracted document, and follow this License in all other
respects regarding verbatim copying of that document.
7. AGGREGATION WITH INDEPENDENT WORKS
A compilation of the Document or its derivatives with other
separate and independent documents or works, in or on a volume of a storage or
distribution medium, is called an "aggregate" if the copyright resulting from
the compilation is not used to limit the legal rights of the compilation's
users beyond what the individual works permit. When the Document is included in
an aggregate, this License does not apply to the other works in the aggregate
which are not themselves derivative works of the Document.
If the Cover Text requirement of section 3 is applicable to these
copies of the Document, then if the Document is less than one half of the
entire aggregate, the Document's Cover Texts may be placed on covers that
bracket the Document within the aggregate, or the electronic equivalent of
covers if the Document is in electronic form. Otherwise they must appear on
printed covers that bracket the whole aggregate.
8. TRANSLATION
Translation is considered a kind of modification, so you may
distribute translations of the Document under the terms of
section 4. Replacing Invariant Sections with translations requires special
permission from their copyright holders, but you may include translations of
some or all Invariant Sections in addition to the original versions of these
Invariant Sections. You may include a translation of this License, and all the
license notices in the
Document, and any Warranty Disclaimers, provided that you also
include the original English version of this License and the original versions
of those notices and disclaimers. In case of a disagreement between the
translation and the original version of this License or a notice or disclaimer,
the original version will prevail.
If a section in the Document is Entitled "Acknowledgements",
"Dedications", or "History", the requirement (section 4) to Preserve its Title
(section 1) will typically require changing the actual title.
9. TERMINATION
You may not copy, modify, sublicense, or distribute the Document
except as expressly provided for under this License. Any other attempt to copy,
modify, sublicense or distribute the Document is void, and will automatically
terminate your rights under this License. However, parties who have received
copies, or rights, from you under this License will not have their licenses
terminated so long as such parties remain in full compliance.
10. FUTURE REVISIONS OF THIS LICENSE
The Free Software Foundation may publish new, revised versions
of the GNU Free Documentation License from time to time. Such new
versions will be similar in spirit to the present version, but may differ in
detail to address new problems or concerns. See
http://www.gnu.org/copyleft/.
Each version of the License is given a distinguishing version
number. If the Document specifies that a particular numbered version of this
License "or any later version" applies to it, you have the option of following
the terms and conditions either of that specified version or of any later
version that has been published (not as a draft) by the Free Software
Foundation. If the Document does not specify a version number of this License,
you may choose any version ever published (not as a draft) by the Free Software
Foundation.


