INTRODUCTION
Les volailles présentent de nombreuses
particularités anatomiques et physiologiques par rapport aux
mammifères. En effet, malgré la très grande
hétérogénéité entre les différentes
espèces aviaires, l'appareil digestif des volailles reste marqué
par l'adaptation au vol, même chez les espèces qui ont perdu cette
aptitude. Cette adaptation morphologique et fonctionnelle se trouve au niveau
de la totalité des appareils et plus particulièrement l'appareil
digestif. Le tube digestif malgré les différences de
régime alimentaire est doué d'une grande capacité
d'absorption qui permet de découvrir le métabolisme basal
élevé de cette espèce.
PREMIERE PARTIE
ETUDE ANATOMIQUE
Rappel
Anatomiquement l'appareil digestif des oiseaux est
constitué par: un bec, une cavité buccale dépourvue de
dents, un gosier, un oesophage, un jabot, des estomacs sécrétoire
et musculaire, l'intestin débouchant dans le cloaque puis l'anus. Il
comprend bien sûr toutes les glandes annexes : le foie et le
pancréas. (VILLATE. D 2001; BRUGERE. H cité par BRUGERE-PICOUX.
J et SILIM. A 1992).
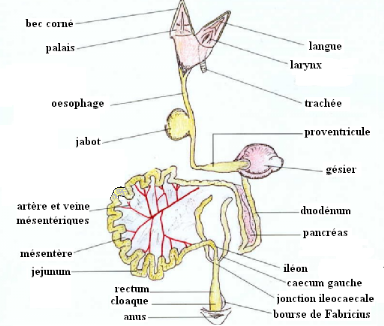
Figure 1 vue latérale du tractus digestif du
poulet après autopsie (VILLATE. D 2001).
1. REGION CRÂNIALE DU TUBE DIGESTIF
1.1 LE BEC
Le bec est utilisé avant tout pour la préhension
des aliments, il offre une grande diversité de formes dans la classe des
oiseaux qui est souvent le reflet d'une adaptation à un régime
alimentaire particulier. Le bec lamellé du Canard lui permet de filtrer
la vase. Le bec cylindrique et très long de la Bécasse lui permet
de rechercher des vers et les larves dans le sol. Les becs forts et coniques
(Poules, Dindons, Canaris, etc....) sont les moins spécialisés
mais témoignent plutôt d'un régime granivore. La forme du
bec est un des éléments importants utilisés pour la
classification scientifique ou taxonomie des oiseaux.
La partie visible du bec est une production cornée ou
rhamphothèque. Au même titre que les griffes, sa croissance est
continue. Elle doit être compensée par une usure
régulière par frottement des deux mâchoires entre elles,
sur les aliments ou sur des objets non comestibles.
Le bec est composé de deux parties : dorsalement
la maxille ou mandibule supérieure ; ventralement la mandibules ou
mandibule inférieure (ALAMARGOT. J 1982).
1.1.1 La maxille
Le squelette de la maxille est constitué principalement
de l'os prémaxillaire. Il est recouvert d'une production
cornée : la rhinothèque. La maxille est perforée de
deux narines qui sont protégées par un opercule chez la Poule et
le Pigeon, par des plumes raides chez le Canari. Chez la Perruche et les
Rapaces diurnes, les narines sont incluses dans une production charnue et
lisse : la cire.
La maxille est légèrement mobile par rapport au
crâne chez tous les oiseaux mais surtout chez les Canaris, les Perroquets
et les Perruches (ALAMARGOT. J 1982).
1.1.2 La mandibule
Le squelette de la mandibule est constitué de l'os
dentaire.
Il est recouvert de la gnathothèque,
généralement moins développée que la
rhinothèque. La mandibule est articulée avec le crâne par
l'intermédiaire de l'os carré (ALAMARGOT. J 1982).
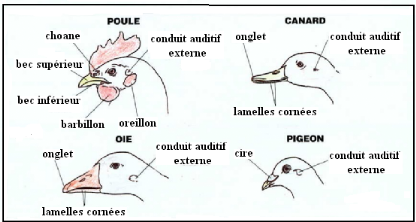
Figure 2 les différents becs des volailles
(VILLATE. D 2001).
1.2 LA CAVITE BUCCALE ET LA LANGUE
1.2.1 La cavité buccale
Elle est limitée rostralement par les bords (ou tomies)
et caudalement par le pharynx. Les limites avec le pharynx sont difficiles
à préciser anatomiquement (d'ou le nom de buccopharynx ou
d'oropharynx donné à l'ensemble bouche et pharynx).Elle ne
possède ni lèvres, ni dents.
La cavité buccale est recouverte d'un
épithélium muqueux, sauf dans sa portion rostrale ou le
revêtement est corné (rhamphothèque).
Le plafond de la cavité buccale est fendu
longitudinalement par la fissure palatine. C'est dans cette fissure que
débouchent les deux choanes (voies respiratoires) qui sont
séparées par l'os vomer.
Chez certaines espèces (Corvidés, etc....et
surtout Pélicans) le plancher de la cavité buccale est extensible
et peut servir au maintien des aliments en formant la poche gulaire. Les
oiseaux n'ont pas de voile du palais ; seul le palais dur existe. Il
possède cinq rangées de papilles filiformes chez la poule
(ALAMARGOT. J 1982).
1.2.2 La langue
Organe mobile situé sur le plancher de la cavité
buccale, la langue présente une grande variabilité de taille, de
forme et de motilité dans la classe des oiseaux. Triangulaire
(sagittée) chez la poule, elle est limitée en arrière par
des papilles filiformes cornées et possède à son apex un
pinceau de soies tactiles. Elle est recouverte d'un épithélium
corné qui lui donne une apparence dure. Elle est soutenue par l'appareil
hyoïdien (os et cartilages) et renferme l'entoglosse. Ses muscles
intrinsèques rudimentaires lui confèrent une souplesse
très réduite
(ALAMARGOT. J 1982).
1.3 LES GLANDES SALIVAIRES
Sont groupées en massifs éparpillés.
Chaque glande possède plusieurs fins canaux excréteurs, soit une
centaine en tout. On distingue les glandes mandibulaires, palatines,
maxillaires, sublinguales, linguales, angulaires, cricoaryténoïdes,
et sphénoptérygoïdes. Les glandes salivaires sont
réduites chez certains oiseaux (Canards). La salive de la Poule
possède une amylase mais son rôle essentiel est de lubrifier et
de ramollir les aliments.
(ALAMARGOT. J 1982).

Figure 3 les glandes salivaires de la poule (VILLATE.
D 2001).
1.4 LE PHARYNX
Le pharynx est le carrefour du tube digestif et des voies
respiratoires. C'est un organe difficile à délimiter chez les
oiseaux (d'ou le nom de buccopharynx). D'un point de vue anatomique, on le
limite rostralement à la dernière rangée de papilles
filiformes du palais (après les choanes) et de la langue, et
caudalement, à l'entrée de l'oesophage, marquée
également d'une petite rangée de papilles. Revêtu d'un
épithélium muqueux simple, le pharynx est en rapport ventralement
avec la trachée par la glotte et dorsalement avec les oreilles moyennes
par une fente médiane, orifice commun aux deux trompes d'Eustache. Chez
la Perruche, cette fente est incluse dans la fissure palatine (ALAMARGOT. J
1982).
1.5 L'OESOPHAGE
L'oesophage est un organe tubuliforme musculomuqueux qui
assure le transport des aliments de la cavité buccale à
l'estomac. Il est situé dorsalement puis à droite de la
trachée dans son trajet cervical. Avant de pénétrer dans
la cavité thoracique chez certaines espèces dont la Poule et le
Pigeon, il se renfle en un réservoir, le jabot. Dans sa portion
intra-thoracique, l'oesophage redevient médian et dorsal à la
trachée. Il dévie vers la gauche après la bifurcation
bronchique (syrinx) puis passe dorsalement aux gros vaisseaux du coeur avec
lesquels il adhère quelque peu. Il se termine dorsalement au foie en
s'abouchant au proventricule.
L'oesophage est tapissé dans toute sa longueur d'une
muqueuse aux plis longitudinaux très marqués. Il possède
une musculature longitudinale interne très développée et
est très dilatable (surtout chez les Rapaces et les oiseaux piscivores)
(ALAMARGOT. J 1982).
1.6 LE JABOT
Le jabot est un élargissement de l'oesophage en forme
de réservoir situé à la base du cou, au ras de
l'entrée de la poitrine. Rudimentaire chez de nombreux oiseaux, il est
bien développé chez nos espèces domestiques (sauf chez le
Canard).Il se présente chez la Poule sous la forme d'un sac ventral
très extensible qui adhère dans sa partie ventrale à la
peau et aux muscles sous-cutanés du cou et dans sa partie caudo-dorsale
aux muscles pectoraux droits. Sa paroi, qui est très mince, a une
musculature (lisse) peu développée mais est riche en fibres
élastiques. (ALAMARGOT. J 1982).
2. REGION STOMACALE DU TUBE DIGESTIF
2.1 LE PROVENTRICULE OU VENTRICULE
SUCCENTURIE
Le proventricule est situé légèrement
à gauche dans la cavité abdominale, ventralement à
l'aorte, dorsalement au foie qui l'enveloppe partiellement .C'est un
renflement fusiforme (de 3 cm de long en moyenne chez la Poule) dont la
muqueuse est très riche en glandes à mucus. La paroi
interne ; très épaisse, est formée de lobules dont
chacun constitue une glande composée radialement à l'axe de
l'organe. Ces glandes en tube se jettent dans un canal commun à
plusieurs glandes et se déverse dans la lumière du proventricule
au sommet d'une proéminence bien marquée. La paroi du ventricule
des carnivores et des piscivores est moins épaisse et plus riche en
fibres musculaires et élastiques. Elle est alors très extensible.
Le transit des aliments ne dure que quelques minutes dans le proventricule.
(ALAMARGOT. J 1982).
2.2 LE GESIER
Le gésier est l'organe compact le plus volumineux de la
poule (6 à 8 cm de long, avec un poids d'environ 50 gr vide et 100 gr
plein). Il est situé légèrement à gauche dans la
cavité abdominale, partiellement coiffé par le foie sur son bord
crâniale. Le gésier est toujours beaucoup plus caudal qu'on ne se
l'imagine ; il est facilement palpable au travers de la paroi abdominale.
De forme sphéroïde, il est en communication crânialement avec
le proventricule et crâniomédialement avec le duodénum. Sa
cavité est sacculaire. Il est très musculeux chez les granivores
(la Poule) et chez les herbivores (l'Oie). Ses deux muscles principaux
s'unissent de chaque coté de l'organe par deux surfaces tendineuses
nacrées : les centres tendineux. Les muscles sont peu
développés chez les frugivores, les carnivores et les piscivores.
L'estomac est alors extensible. Le gésier est rattaché au sternum
et à la paroi abdominale par le ligament ventral ou
mésentère ventral, au foie par le ligament gastrohépatique
et à la paroi dorsale de l'abdomen par le mésogaster. Il partage
longitudinalement la cavité abdominale en deux compartiments ce qui lui
a valu parfois le nom « diaphragme vertical ». (ALAMARGOT.
J 1982).
3. REGION POSTÉRIEURE DU TUBE DIGESTIF
3.1 LE DUODENUM
Le duodénum est la portion de l'intestin qui fait suit
l'estomac. Il débute au pylore puis forme une grande anse qui enserre le
pancréas. Cette anse est la partie la plus ventrale de l'intestin dans
la cavité abdominale. Elle contourne caudalement le gésier et
dorsalement elle est en rapport avec les caecums. Le duodénum
reçoit deux ou trois canaux pancréatiques et deux canaux
biliaires au niveau d'une même papille. L'emplacement de cette papille
marque la fin du duodénum et le début de l'iléon. (Voir
tableau 1) (VILLATE. D 2001; ALAMARGOT. J 1982).
Tableau 1 la longueur et le calibre de l'anse
duodénale (VILLATE. D 2001).
|
Longueur de l'anse en cm
|
Calibre en cm
|
|
Poule
|
22-35
|
0.8-1.2
|
|
Canard
|
22-38
|
0.4-1.1
|
|
Oie
|
40-49
|
1.2-1.6
|
|
Pigeon
|
12-22
|
0.5-0.9
|
3.2 LE JÉJUNUM
Il est divisé en deux parties
- L'une proximale qui est la plus importante : tractus du
Meckel. Petit nodule, est parfois visible sur le bord concave de ses
courbures.
- L'autre distale qui s'appelle l'anse supraduodénale.
(Voir tableau 2) (VILLATE. D 2001; ALAMARGOT. J 1982).
Tableau 2 la longueur et le calibre du
jéjunum chez quelques espèces (VILLATE. D 2001).
|
Calibre en cm
|
Longueur en cm
|
|
|
0.6-1.0
|
85-120
|
Poule
|
|
0.4-0.9
|
90-140
|
Canard
|
|
1.3-1.7
|
150-185
|
Oie
|
|
0.35-0.7
|
45-72
|
Pigeon
|
3.3 L'ILÉON
Il est court et rectiligne, son diamètre et sa longueur
sont variables en fonction des espèces. (Voir le tableau 1). (VILLATE. D
2001; ALAMARGOT. J 1982).
Tableau 3 La longueur et le calibre de l'iléon
chez certaines espèces (VILLATE. D 2001).
|
Longueur en cm
|
Calibre en cm
|
|
Poule
|
13-18
|
0.7-1.0
|
|
Canard
|
10-19
|
0.4-0.8
|
|
Oie
|
20-28
|
1.0-1.5
|
|
Pigeon
|
8-13
|
0.3-0.5
|
3.4 LES CAECUMS
Un caecum se présente comme un sac qui débouche
dans le tube intestinal à la jonction de l'iléon et du rectum au
niveau d'une valvule iléocæcale. Lorsqu'ils existent, ils sont
toujours pairs, ils sont accolés à la parie terminale de
l'iléon par un méso. Ils sont en rapport ventralement avec l'anse
duodénale et dorsalement avec la portion moyenne de l'iléon. Bien
développés chez la Poule, ils sont petits chez le Canard et
l`Oie. Absents chez les perroquets, les rapaces diurnes, et les pigeons. (Voir
le tableau 4). (VILLATE. D 2001; ALAMARGOT. J 1982).
Tableau 4 la longueur et le calibre du caecum chez
quelques espèces (VILLATE. D 2001).
|
Longueur en cm
|
Calibre en cm
|
|
Poule
|
12-25
|
-
|
|
Canard
|
10-20
|
0.5-0.7
|
|
Oie
|
22-34
|
0.8-1.2
|
|
Pigeon
|
0.2-0.7
|
-
|
3.5 LE RECTUM
Le rectum fait suite à l'iléon et
débouche dans le cloaque. Le diamètre du rectum est à
peine plus grand que celui de l'iléon. A l'inverse des
mammifères, le rectum des oiseaux présente des villosités.
Le rectum réabsorbe l'eau de son contenu (fèces et urines), ces
fonctions lui ont valu parfois le nom de colorectum. (ALAMARGOT. J 1982).
3.6 LE CLOAQUE
Le cloaque est la partie terminale de l'intestin dans laquelle
débouchent les conduits urinaires et génitaux. Il est
formé de trois régions séparées par deux plis
transversaux plus ou moins nets :
3.6.1 Le coprodéum
Il est large et collecte les excréments, c'est une
dilatation terminale du rectum, la portion la plus crâniale du cloaque.
C'est dans le coprodéum que s'accumulent les fèces et les urines
avant leur émission ;
3.6.2 L'urodéum
Il est plus petit, c'est le segment moyen du cloaque. Il
reçoit les conduits génitaux et urinaires, dans sa paroi dorsale
débouchent les deux uretères. Ainsi que les deux canaux
déférents chez les mâles ou l'oviducte chez les
femelles.
3.6.3 Le proctodéum
Résulte d'une dépression de l'ectoderme
embryonnaire et s'ouvre à l'extérieur par l'anus C'est le
segment caudal du cloaque. Chez quelques espèces, il renferme
ventralement un pénis. Chez tous les jeunes oiseaux, il est relié
dorsalement à la bourse de Fabricius avec la quelle il peut communiquer
par un canal. Le cloaque s'ouvre à l'extérieure par l'orifice
cloacal : fente verticale fermée par deux lèvres
horizontales (VILLATE. D 2001; ALAMARGOT. J 1982).
4. LES GLANDES ANNEXES
4.1 LE PANCRÉAS
Le pancréas est une glande amphicrine (endocrine et
exocrine), compacte, blanchâtre ou rougeâtre, enserrée dans
l'anse duodénale. Le pancréas est issu de trois ébauches
séparées qui se constituent en deux lobes (un lobe ventral et un
lobe dorsal). Le suc pancréatique se déverse dans le
duodénum par deux ou trois canaux qui s'abouchent au même niveau
que les canaux hépatiques. (ALAMARGOT. J 1982).
4.2 LE FOIE
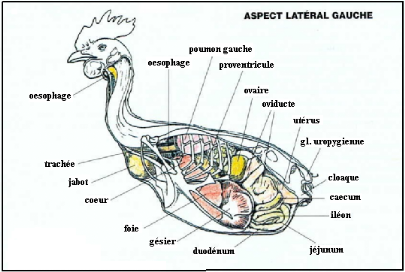
Le foie est un organe volumineux rouge sombre. C'est la glande
la plus massive de tous les viscères (33 gr environ chez la poule). Le
foie repose sur le sternum, il est séparé des parois
thorco-abdominales par les sacs aériens. Il est soutenu par quatre
ligaments (falciforme, coronaire, gastrohépatique et
hépatoduodénal). Sa face ventro-médiale porte les
impressions splénique, stomacale et intestinale. Le foie est
constitué de deux lobes réunis par un isthme transversal qui
renferme partiellement la veine cave caudale. Le lobe gauche plus petit que le
lobe droit, il est généralement marqué d'un sillon
longitudinal qui délimite le lobe accessoire du lobe gauche. Dans leur
portion crâniale, les deux lobes entourent complètement les
ventricules du coeur. Les deux lobes déversent la bile, par deux
conduits séparés. Le canal du lobe gauche (canal hépatique
gauche) s'abouche directement dans l'intestin. Le canal du lobe droit (canal
hépatique droit) se renfle d'abord en vésicule biliaire (sauf
chez le Pigeon, certains Perroquets et l`Autriche) avant de se jeter dans le
duodénum. Il porte le nom de canal cholédoque. (ALAMARGOT. J
1982).
Figure 4 topographie viscérale de la poule, le
coté gauche (VILLATE. D 2001).
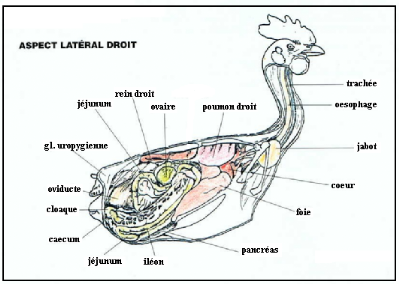
Figure 6 topographie viscérale de la poule, le
coté droit (VILLATE. D 2001).
DEUXIEME PARTIE
ETUDE PHYSIOLOGIQUE
1. PHENOMENES MOTEURS
1.1 MOTRICITE DU SEGMENT ORAL
Le profil moteur du segment oral est marqué par
l'absence de la phase de mastication.
1.1.1 Ingestion-déglutition
La préhension des aliments est assurée par le
bec, qui présente des variations morphologiques en rapport direct avec
la nature de régime alimentaire. Les particules alimentaires
ingérées ne subissent pas de modifications notables au niveau de
la bouche (absence de dents). Les simples transformations du bol alimentaire
sont liées à l'intervention des muscles hyo-branchio-lingaux et
à son humectation par la salive. La déglutition est
essentiellement un phénomène mécanique, elle est
facilitée par les mouvements de la tête. On distingue trois
phases, orale, pharyngienne, et oesophagienne. (SOUILEM. O et GOGNY. M 1994)
- La phase orale
Au cours de cette phase, la langue effectue des mouvements
rapides rostro-caudaux, de propulsion et de rétropulsion, qui durent
en moyenne 1 à 3 secondes. En même temps, on assiste à la
fermeture de la glotte et à un basculement de l'appareil hyoïde. La
progression du bol alimentaire en arrière, en direction du pharynx, est
assurée par de brefs mouvements d'extension de la tête.
- La phase pharyngienne
Elle est surtout accompagnée d'un basculement de la
langue qui recule, d'une dilatation du pharynx et d'un avancement de oesophage.
PASTEA et col. ont montrés que les bols volumineux sont poussés
immédiatement dans l'oesophage, alors que si les grains sont
administrés un par un, ils stagnent quelques temps dans le pharynx,
avant d'arriver au niveau de la jonction pharyngo-oesophagienne. La phase
pharyngienne est caractérisée par la remise en place du pharynx,
de la glotte, de l'appareil hyoïde, et de la langue (SOUILEM. O et GOGNY.
M 1994).
1.1.2 Transit oesophagien
La progression des aliments solides dans l'oesophage
résulte de la progression de salves de potentiels (durée : 6
secondes, amplitude : 350 à 400 uv à une vitesse de 0.8
à 1.2 cm/sec. Dans le cas des liquides, la progression résulte
surtout de l'effet de la pesanteur, conditionné par la position de la
tête. Le transit oesophagien est alors immédiat grâce
à l'apparition d'ondes péristaltiques rapides qui se propagent
à une vitesse de 5 à7 cm/sec. Une alimentation sous forme de
farine doit s'accumuler dans le pharynx pour atteindre un volume seuil capable
de déclencher la motricité oesophagienne. Dans ce cas
l'apparition de salves potentiels de pointe doit être
précédé en moyenne de 3 à 4 prises alimentaires.
Cette activité péristaltique beaucoup plus lente que chez les
mammifères, pourrait s'expliquer par le faible nombre de fibres
musculaires longitudinales chez la volaille (SOUILEM. O et GOGNY. M
1994).
1.1.3 Motricité du jabot
La motricité du jabot est corrélée non
seulement avec celle de l'oesophage mais aussi avec le proventricule et le
gésier. L'étude du dynamisme du jabot, par les méthodes
radioscopiques et radiographiques, fait apparaître des dilatations
brusques et des contactions partiels de pétrissage et
d'évacuation. L'analyse électromyographique (EMG) a permis de
relier la motricité oesophago-ingluviale à la prise alimentaire.
En dehors des prises, l'EMG présente de longues salves de potentiels
(durée : 6 sec) à propagation péristaltique lente,
ces salves, présentes surtout pendant la phase inter-prandiale, sont
accompagnées d'un ralentissement de la motricité du
gésier. La prise de nourriture s'accompagne d'une inhibition de
l'activité électrique du jabot, en raison de l'effet
excito-moteur sur l'oesophage et de la distension crée par l'aliment. En
outre, l'activité du jabot est corrélée à celle du
gésier. Quand le gésier est contracté, le bol alimentaire
passe dans le jabot, alors que s'il est relâché, le bol ne
pénètre pas dans le jabot. La distension du jabot est
accompagnée d'une relance de la sécrétion acide par le
proventricule. La vidange du contenu du jabot résulte de l'apparition de
10 à 20 salves de potentiels consécutives, après 1
à 3 heures de séjours des aliments stockés (SOUILEM. O et
GOGNY. M 1994).
1.2 MOTRICITE DU SEGMENT MOYEN
Elle s'intéresse à la fois le proventricule, le
gésier et l'intestin. La fréquence est évaluée
à 3 à 4 cycles par minute chez le poulet et le dindon, selon la
séquence suivante :
muscles minces du gésier ? duodénum ? muscles
épais du gésier ?proventricule.
La contraction des muscles épais et des muscles minces
du gésier est séparée dans le temps, les muscles minces se
contractent toujours avant les muscles épais, ce qui permet dans un
premier temps le passage de la partie la plus liquide du chyme dans le
duodénum. Ensuite les muscles épais, qui représentent de
véritables « mâchoires » gastriques, se
contractent pour assurer le broyage et la trituration du chyme résidant.
Le reflux duodéno-gastrique se produit en moyenne toutes les 15 à
20 minutes et s'accompagnant d'une simple ou double contraction brusque du
duodénum. En même temps, on note une inhibition totale de la
motricité du proventricule et du gésier, probablement par la mise
en jeu d'un mécanisme réflexe d'origine intrinsèque. Ce
mécanisme de reflux, à point de départ duodénal,
permet l'échange d'aliments entre le duodénum, le gésier
et le proventricule. Associé au transit classique, il est à
l'origine de va et vient entre ces trois compartiments. Ce reflux
duodénal peut s'interpréter comme un mécanisme
supplémentaire chez les volailles, rendu nécessaire par la
localisation paradoxale de l'estomac sécrétoire par rapport
à l'estomac mécanique, et par l'abouchement des canaux
cholédoques et
pancréatiques à l'extrémité
distale de l'anse duodénale.
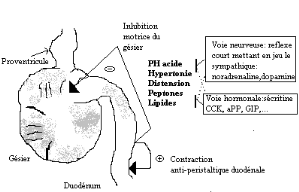 Figure 6 schéma simplifié des
mécanismes de contrôle de la vidange gastrique et du
réflexe duodéno-gastrique (SOUILEM. O et GOGNY.
M 1994).
Figure 6 schéma simplifié des
mécanismes de contrôle de la vidange gastrique et du
réflexe duodéno-gastrique (SOUILEM. O et GOGNY.
M 1994).
La distension duodénale ou l'introduction au niveau du
duodénum d'acide chlorhydrique, d'une solution hypertonique de chlorure
de sodium, d'amino-acides ou de lipides inhibent la motricité du
gésier. Cette inhibition s'installe généralement en 3
à 30 secondes et persiste pendant 2 à 30 minutes, en fonction de
la dose administrée, à l'exception des solutions à base
lipides, qui provoquent un effet inhibiteur au bout de 4 à 6 minutes, et
d'une durée de 24 à 45 minutes. Ceci laisse supposer que les
volailles sont plus sensibles aux graisses alimentaires que les
mammifères. L'effet inhibiteur exercé par une solution
hypertonique de NaCl ou des lipides est souvent accompagné de reflux
duodéno-gastriques. Le déterminisme de la motricité
gastro-duodénale est à la fois nerveux et humoral. Il existe une
phase céphalique et une phase gastrique. Le stimulus impliqué
dans la phase céphalique est représenté par la vue de
l'aliment. Le stimulus impliqué dans la phase gastrique est
l'arrivé des aliment dans le gésier. Les hormones
impliquées dans la régulation de la motricité
gastro-intestinale sont représentées par la
sécrétine, la CCK-PZ, le polypeptide pancréatique aviaire
(aPP) et le gastric inhibitory peptide (GIP). La CCK-PZ inhibent fortement la
motricité du gésier et du duodénum, mais très peu
celle de l'iléon et du cæcum. De plus elle provoque un reflux
antérograde du contenu de l'intestin. Le (aPP) est aussi à
l'origine d'une inhibition de la motricité gastrique et
duodénale. La motricité iléale est
caractérisée à la fois par la présence de
mouvements de segmentation et de péristaltisme. Le système
mésentérique est très développé à ce
niveau et se trouve contrôlé de façon classique par le
système neurovégétatif. L'activité
anti-péristaltique n'est pas l'apanage du duodénum. On la
retrouve aussi dans la portion distale de l'intestin grêle (SOUILEM. O et
GOGNY. M 1994).
1.3 MOTRICITE DU SEGMENT DISTAL
Elle s'intéresse à la fois les cæcum, le
colon et le rectum. La motricité caecale est caractérisée
par deux types de contractions :
- Des contractions majeures, puissantes, propulsives et
évacuatrices, qui peuvent se propager dans les deux sens, mais toutes
fois plus nombreuses dans le sens apex-rectum.
- Des contractions mineures, qui assurent un rôle de
mixage, et dont le rôle propulsif est très faible.
Le remplissage du caecum ne se fait pas à partir de
l'intestin grêle, mais à partir de la région recto-colique,
grâce à un anti-péristaltisme rectal permanent.
L'évacuation du contenu caecal nécessite une activation
électrique de l'ensemble des caecums, la vidange ne survient jamais
pendant la période d'obscurité, mais surtout en fin de
période d'éclairement.
La motricité recto-colique est
caractérisée par la présence d'un
anti-péristaltisme permanent vigoureux à partir du cloaque.
L'antipéristaltisme rectal est de type continu, son inhibition est
observée seulement au moment de la vidange rectale. La
défécation est liée à l'installation d'une onde
péristaltique violente qui parcourt le duodénum dans un
délai de 4 secondes environ, depuis la partie proximale du rectum
jusqu'à la région distale.
En résumé, la motricité globale du tube
digestif des volailles est caractérisée par une activité
importante et coordonnée se propageant dans les deux sens, et ce
à tous les niveaux. La durée de ce transit est estimée
à 6-10 heures. Elle varie en fonction de l'espèce, de l'age de
l'animal (plus rapide chez le jeune), du taux d'incorporation alimentaire (les
graisse augmentent la durée du transit), la température et le
stress (SOUILEM. O et GOGNY. M 1994).
2. PHENOMENES SECRETOIRES
2.1 SECRETION SALIVAIRE
La quantité de salive globale
sécrétée chez la poule est estimée à 70-30
ml par 24 heures. Elle est essentiellement de mucus,
sécrété par les glandes muqueuses, indispensable à
la lubrification de l'aliment, surtout en l'absence d'une phase de mastication.
Il facilite ainsi le transit du bol alimentaire à travers le
bucco-pharynx et la partie proximale de l'oesophage. Chez certaines
espèces comme le moineau et l'oie, on note la présence d'une
amylase, absente chez la poule et le dindon. L'activité amylolytique
semble être corrélée avec la taille et le degré de
développement du jabot, La poule et la dinde possèdent un jabot
très développé, ce qui permet aux aliments de
séjourner un certain temps (quelques minutes jusqu'à 1 jour),
avant de gagner le proventricule, d'où la possibilité d'une
attaque de l'amidon par les amylases d'origine végétale dans un
milieu favorable. Chez le moineau et à moindre degré l'oie, le
jabot est fusiforme et ne possède pas une grande possibilité de
stockage. La présence d'une amylase salivaire pourrait constituer un
moyen de valorisation et de compensation. Le contrôle de la
sécrétion salivaire est similaire à celui des
mammifères. La prise alimentaire est le principal facteur de
déclenchement par la mise en jeu du système parasympathique
(SOUILEM. O et GOGNY. M 1994).
2.2 SECRETION INGLUVIALE
L'activité sécrétoire du jabot est
faible, voir nulle. On observe seulement une sécrétion abondante
du mucus par les glandes muqueuses de l'oesophage et de l'entrée du
jabot, favorisant l'imbibition et la macération des aliments. Il ne
semble pas y avoir de sécrétion enzymatique propre dans le jabot.
Chez le pigeon en période de reproduction, le jabot joue un rôle
important par la sécrétion d'un produit nutritif (lait de
pigeon), qui sera régurgité dans la cavité buccale des
pigeonneaux (SOUILEM. O et GOGNY. M 1994).
2.3 SECRETION GASTRIQUE
Aucune sécrétion n'a été
rapportée au niveau du gésier, mise à part la
sécrétion d'une substance protéique ressemblant à
la kératine (koïline) et formée d'un complexe
polysaccharido-protéique.
La sécrétion gastrique est assurée
uniquement par le proventricule, avec comme principale originalité la
sécrétion d'acide chlorhydrique et du pepsinogène par des
cellules spécialisées oxyntico-peptiques ou cellules principales.
Le (HCl) peut solubiliser quotidiennement 7 à 8 grammes de carbonate de
calcium, d'où le rôle non négligeable du proventricule dans
le contrôle du métabolisme calcique, surtout chez la poule
pondeuse. Le pH de la sécrétion gastrique est compris entre 1 et
2. La pepsinogène est transformée en pepsine sous l'effet du HCl
et de la pepsine elle-même. D'autres enzymes ont été
retrouvées dans le contenu gastrique, telle la lipase, très
probablement à la suite du reflux anti-péristaltique
duodénal. L'action de la sécrétion gastrique ne
débute réellement que dans le duodénum, lorsque les
aliments sont suffisamment fragmentés. Comme chez les mammifères,
on distingue trois phases de sécrétion, céphalique,
gastrique et intestinale. La prise alimentaire et l'arrivée des aliments
au niveau de l'estomac stimulent de façon importante la
sécrétion gastrique. Le (aPP) stimule à la fois la
sécrétion de H+ et de pepsine. La phase intestinale de la
sécrétion gastrique est contrôlée essentiellement
par la sécrétine et la CCK-PZ. La sécrétine stimule
à la fois la sécrétion de H+ et de pepsine, alors que la
CCK-PZ stimule la sécrétion de H+ et inhibe celle de pepsine
(SOUILEM. O et GOGNY. M 1994).
2.4 SECRETION PANCREATIQUE
Le suc pancréatique présente une couleur vert
pâle et un pH variant de 6.4 à 6.8 chez la poule, et de 7.4
à 7.8 chez le dindon. La quantité
sécrétée chez la poule est estimée à
15 à 20 ml/j. les activités enzymatiques sont faibles chez le
jeune poussin. La possibilité de digérer les glucides se
développe au cours des premiers jours (4 à 5 jours).
On distingue deux fractions, une faction aqueuse et une
fraction enzymatique. La composition de la fraction aqueuse est très
proche de celle de mammifères, avec comme principale substance, les ions
bicarbonates indispensables à l'alcalinisation du pH, et à une
activité enzymatique maximale. La fraction enzymatique comporte les
enzymes indispensables à la dégradation des lipides, des protides
et des glucides. On note la présence de ribonucléase, d'amylase,
de lipase, de chymotrypsine, de trypsine, d'élastase et de
carboxypeptidases. Il existe deux phases de sécrétion
pancréatique, une phase céphalique et une phase intestinale.
L'ingestion alimentaire stimule la sécrétion. Il existe une
corrélation entre la motricité de la partie proximale du
duodénum et la sécrétion pancréatique chez la
poule. Les contractions de la partie proximale du duodénum
favoriseraient le déplacement du chyme provenant du gésier,
à l'origine de la relance pancréatique. Les hormones
impliquées dans ce contrôle sont représentées par la
sécrétine, la CCK-PZ et le VIP. La sécrétine
augmente la sécrétion de la phase aqueuse. La CCK-PZ n'a que peu
d'effet. Le pancréas des volailles est plus sensible à l'action
du VIP que de la sécrétine. La sécrétion
enzymatique est très influencée par le régime
alimentaire ; l'ingestion régulière d'une grande
quantité de carbohydrates et de lipides augmente l'activité de
types amylase et lipase, alors qu'un régime hyperprotidique augmente
très peu l'activité de la chymotrypsine.(SOUILEM. O et GOGNY. M
1994; BRUGERE-PICOUX. J et SILIM. A 1992).
2.5 SECRETION BILIAIRE
La vésicule biliaire est absente chez quelques
espèces (autruches, pigeons). La sécrétion de la bile est
estimée à 1 ml/h chez la poule. Il s'agit d'un liquide
verdâtre, légèrement acide (pH de l'ordre 6). Les sels
biliaires sont constituées pour les deux tiers de
tauro-chéno-désoxycholate. On trouve aussi du taurocholate et des
tauro-allocholates. Ces sels sont indispensables à l'action de la lipase
pancréatique, dans la mesure où ils émulsifient les
lipides ; ils favorisent en outre nettement l'absorption intestinale de
calcium. Les pigments biliaires, tels que la bilirubine et la biliverdine sont
présents dans la bile, mais la biliverdine n'apparaît que à
l'extérieure du foie. La synthèse de la bile se développe
avec l'age ; les jeunes oiseaux digèrent male les lipides, surtout
quand ils sont à base d'acides gras saturés. Ainsi, l'addition de
sels biliaires dans l'alimentation du poussin ou du dindonneau améliore
la digestibilité des acides gras saturés, et dans moindre mesure
celle des acides gras insaturés. La sécrétion biliaire est
stimulée par l'ingestion alimentaire et par la présence de sels
biliaires dans le sang. Le contrôle hormonal fait intervenir probablement
la CCK-PZ sécrétée par l'intestin (VILLATE. D 2001;
SOUILEM. O et GOGNY. M 1994).
2.6 SECRETION INTESTINALE
La sécrétion de base est estimée à
1.1ml/h pour un poulet de 2.5 à 3.5 kg. La source de la
sécrétion est représentée seulement par les glandes
de Lieberkühn. Le suc intestinal est jaune pâle et renferme du
mucus, des électrolytes et des enzymes. Les extraits de la muqueuse
intestinale sont capables de digérer les glucides, les lipides et les
protides. On y trouve amylase, di-saccharidases, peptidases, lipases, maltases,
sucrases, isomaltases, entérokinases, etc.... La lactase est absente
d'où l'intolérance au lactose. Le lait et ses sous-produits
provoquent e la diarrhée et des troubles intestinaux. La plus grande
activité di-saccharidase a été retrouvée au niveau
de la partie proximale de l'iléon. Les activités maltase et
sucrase sont présentes dés la naissance, ce qui donne la
possibilités au poussin de digérer les sucres dés les
premiers jours (4 à 5 jours).la digestion des lipides est
médiocre à la naissance, et seuls les lipides insaturés
sont utilisés. La valorisation des lipides saturés ne devient
possible qu'à l'age de 4 à 8 semaines. (SOUILEM. O et GOGNY. M
1994).
3. DIGESTION MICROBIENNE
L'activité microbienne a été
signalée essentiellement au niveau du jabot et surtout des caecums.
Cette flore bactérienne (surtout de bactéries gram
négatives), favorise le renouvellement rapide de
l'épithélium intestinal. La flore bactérienne du jabot est
surtout constituée de lactobacilles (Lactobacillus acidophilus), qui
contribuent à la baisse relative du pH local, par la
sécrétion d'acide lactique, d'acides organiques et d'acides gras
volatils. Les oses issus de la dégradation bactérienne des
glucides sont absorbés en très faible quantité et sont
souvent utilisés comme source d'énergie par les bactéries.
Les cæcums constituent un milieu favorable pour la multiplication
bactérienne : le milieu est anaérobie, très liquide,
stagnant partiellement (l'évacuation ne se fait que toutes les 6
à 8 heures en moyenne), et le pH est de l'ordre de 6.5 à 7.5.
L'attaque des glucides se traduit par une activité amylolytique
très faible, ainsi qu'une production de gaz (CO2, méthane) et
d'acides gras volatils. Concernant la digestion de la cellulose, les poulets
normaux sont capables d'utiliser 17% de la cellulose de la ration. Les caecums
jouent un rôle dans la digestion des protéines et dans la
récupération de l'azote non protéique, la digestion des
protéines est mieux valorisée chez les sujets normaux que chez
ceux qui ont subit l'ablation des cæcums. La flore caecale
anaérobie est capable, de décomposer l'acide urique (principal
composé azoté excrété par les reins), issu de
l'urine véhiculée par le reflux antipéristaltique.
L'ammoniac produit est incorporé dans la synthèse d'acides
aminés, qui sont utilisés par les bactéries et
généralement peu absorbés par la paroi des caecums. Comme
chez les mammifères, la flore digestive des oiseaux, déprime
l'utilisation des lipides, en réduisant le rôle des sels
biliaires. Enfin, la flore bactérienne au niveau des caecums est capable
de synthétiser les vitamines hydrosolubles, surtout du groupe B, telle
la vitamine B12 (SOUILEM. O et GOGNY. M 1994; BRUGERE-PICOUX. J et SILIM. A
1992).
4. ABSORPTION DES NUTRIMENTS
4.1 EAU ET ELECTROLYTES
L'eau est absorbée selon un mécanisme passif qui
dépend de la pression osmotique. Chez les oiseaux, les pressions
osmotiques enregistrées dans le tube digestif, peuvent dépasser
deux fois la pression osmotique du sang. Dans ces conditions, le flux de l'eau
devrait se faire dans le sens d'une excrétion depuis les cellules vers
la lumière intestinale (diffusion simple). Dans le cas du Na, la
concentration intracellulaire est inférieure à la concentration
luminale, son absorption s'effectue en descendant le gradient
électrochimique sans nécessiter d'énergie. Le transport du
K est essentiellement passif. Le transport actif fait intervenir une K-ATPase
située sur la membrane apicale. L'absorption intestinale du Ca
dépend de nombreux facteurs liés à la composition du
régime alimentaire et au stade physiologique de l'animal. Lorsque
l'aliment apporte du Ca en quantité suffisante par rapport au besoin, le
mécanisme d'absorption peut être assimilé à une
simple diffusion dépendant de la différence du potentiel
électrochimique. Dans le cas d'une déficience, le
mécanisme de transport est actif, dépendant à la fois de
la parathormone et de la vitamine D. chez la poule pondeuse, l'absorption du Ca
est augmentée en même temps que la quantité de CaBP
(Calcium Binding Protein) intestinale et plasmatique. L'influence de
l'état physiologique est surtout remarquable aux heures de formation de
l'oeuf ; le pourcentage de Ca absorbé par rapport à
l'ingéré varie pour le jéjunum supérieure de 45
à 17 p.100 selon que l'oeuf est en formation où non. Le fer
absorbé est stocké sous forme de ferritine. Il est
transporté à l'intérieure des cellules par la
transferrine. Son passage dans le milieu sanguin est régulé en
fonction des besoins de l'organisme (LARBIER. M et LECLERCQ. B 1992).
4.2 MONOSACCHARIDES
Dans le cas du glucose et de galactose, le système est
une combinaison ternaire : monosaccharide-transporteur-sodium.
L'énergie est nécessaire pour permettre le passage
basolatéral du Na. Selon le modèle de Crane, lorsque la
concentration en Na est suffisante, le glucose se fixe sur le transporteur. Le
complexe (G-T-Na) passe sur la face cytosolique de la membrane
entérocytaire. Dans le milieu intracellulaire, la concentration de Na+
est faible, celui-ci se détache du transporteur et du même coup,
l'affinité diminue pour le glucose qui se libère à son
tour.
Le transporteur retrouve sa structure initiale et son
activité pour transférer de nouvelles molécules de
glucose. La teneur de Na+ dans le cytosol est maintenue faible grâce
à la Na - K - ATPase qui expulse activement Na en faisant entrer K sur
les faces basolatérales de l'entérocyte.
Pour les oses autres que le glucose et le galactose,
l'absorption intestinale s'effectue selon le mécanisme de simple
diffusion ou de diffusion facilitée. Ainsi pour le fructose, il s'agit
d'une diffusion facilitée indépendante de Na et l'énergie
(LARBIER. M et LECLERCQ. B 1992).
4.3 ACIDES AMINES
La vitesse d'absorption des acides aminés dépend
de la nature de ces derniers. Les acides aminés neutres sont
transportés selon un mécanisme Na dépendant. Pour les
acides aminés basiques l'absorption intestinale s'effectue selon le
mécanisme de Na-dépendant, beaucoup moins actif que celui des
acides aminés neutres. Dans le cas des acides aminés
dicarboxyliques, le flux procède par un mécanisme actif mais
partiellement Na-dépendant. La vitesse d'absorption des acides
aminés comme celle des glucides simples, dépend de nombreux
facteurs liés à l'état nutritionnel des animaux et
à la composition du régime alimentaire. D'une façon
générale, elle est fortement diminuée chez les poulets
nourris avec un aliment hypoénergétique. Elle est en revanche
augmentée sous l'influence d'une subdéficience en acides
aminés (LARBIER. M et LECLERCQ. B 1992).
4.4 LIPIDES
Les lipides sont capturés par les entérocytes
selon une simple diffusion. Dans le cas du cholestérol, il doit
être sous fore polaire pour être absorbé par une
protéine spécifique. La ré-estérification est
réalisée grâce à la
cholestérol-estérase et la
cholestérol-acetyl-transférase. Les phospholipides alimentaires
sont hydrolysés par la phospholipase pancréatique puis
absorbés sous forme de lysophospholipides (LARBIER. M et LECLERCQ. B
1992).
4.5 VITAMINES
La vitamine A est consommée sous forme de ß
carotènes ou d'ester, qui est hydrolysé par une estérase
pancréatique. L'absorption est passive, insensible à l'anoxie,
augmentée en présences de sels biliaires. A l'intérieur de
l'entérocyte, le carotène est scindé puis
estérifié avant d'être incorporé dans les
lipoprotéines transporteuses de lipides. Le mécanisme de
transport de la vitamine D est comparable à celui de la vitamine A. A
l'intérieur de l'entérocyte on a une hydroxylation avant le
passage dans le sang porte. L'hydrolyse luminale de l'ester
précède l'absorption de la vitamine E qui sera par la suite
transportée comme les nutriments lipidiques. La vitamine K1 est
transportée activement par un système
énergie-dépendant mais Na-indépendant. Les vitamines K2 et
K3 sont absorbées passivement par les lipoprotéines sans subir de
transformation intra-entérocytaire. La vitamine B1 nécessite un
transport spécifique et du Na. Pour la B12 le mécanisme
d'absorption est passif lorsque celle-ci se trouve en forte concentration dans
la lumière intestinale. Pour les concentrations physiologiques, la
vitamine se combine à une protéine d'origine gastrique (F1 ou
facteur intrinsèque) qui la protège contre l'action de la flore
bactérienne. Dans le jéjunum et surtout l'iléon, la
vitamine se lie à un récepteur membranaire qui assure son
transport, le mécanisme est énergie-indépendant. Le
transfert au sang est réalisé grâce à deux
transporteurs, les transcobalamines 1 et 2. La vitamine pp est
transférée activement. Il est en même pour l'acide
pantothénique dont le système de transport est dépendant
à la fois du Na et de l'énergie. La B2 est d'abord
hydrolysée. Les molécules de riboflavine obtenues sont
absorbées activement selon un mécanisme Na dépendant
stimulé par les sels biliaires. Enfin les folates sont
hydrolysées avant d'être absorbé en présence d'un
transporteur : la Folate Binding Protein (FBP) selon un mécanisme
actif, saturable, spécifique et énergie-dépendant
(LARBIER. M et LECLERCQ. B 1992).
CONCLUSION
Le tube digestif des oiseaux
présente des particularités fondamentales par rapport aux
mammifères. Les adaptations fonctionnelles, sont en parfaite
corrélation avec l'anatomie du tube digestif. La valorisation importante
de l'aliment ingéré par les volailles témoigne de la
grande efficacité de la digestion et des mécanismes d'absorption
malgré la présence d'un tube digestif court et d'un transit
intestinal rapide.
LA BIBLIOGRAPHIE
1. ALAMARGOT. J 1982.
- L'appareil digestif et ses annexes, pages 15-32.
- In : Manuel d'anatomie et d'autopsie aviaires.
- Edition : Le point vétérinaire.
2. BRUGERE-PICOUX. J et SILIM. A 1992.
- Particularités de la physiologie des oiseaux, pages
15-24.
- In : Manuel de pathologie aviaire.
- Edition : chaire de pathologie médicale du
bétail et des animaux de basse-cour, école nationale
vétérinaire D'Alfort.
3. LARBIER. M et LECLERCQ. B 1992.
- Absorption des nutriments, pages 38-47.
- In : Nutrition et alimentation des volailles.
- Edition : INRA.
4. SOUILEM. O et GOGNY. M 1994.
- Particularités de la physiologie digestive des
volailles, pages 525-537.
- Revue de la médecine vétérinaire,
juillet 1994, 145.
5. VILLATE. D 2001.
- L'appareil digestif, pages 27-38.
- In : Les maladies des volailles.
- Edition : INRA.


