..ÈÚÔ~Ç
4.ØÇÑÞã~Ï1Ç
k.ÑÆÇÒÌ1Ç
k.Ñ.ãÌ1Ç
République Algérienne Démocratique
et Populaire
íãáÚ1Ç
ËÍÈ1Ç j
í1~Ú1Ç
ã.áÚÊ~Ç ÉÑÇÒ
Ministère de l'Enseignement Supérieur et de
la Recherche Scientifique
|
áÍlÓ1Ç
~Æ~Ê J ÑÍÈ~Ç ã9áÚ~
17áÚ~Ç ;1äØ11Ç
|
4ÓÑÏã.Ç
|
Ecole Nationale Supérieure des Sciences de la Mer
et de l'Aménagement du Littoral

Mémoire de fin d'études en vue de
l'obtention du diplôme d'études
universitaire appliquées
(DEUA) en sciences de la mer et de
l'Aménagement du
Littoral
Thème :

Contribution à l'étude de la
croissance du Pagellus acarne (Risso, 1826) dans la baie de Bou
Ismail.
Présenté par :
Enseignante:
Berkani Abdelali Mme OUABADI. T.
Année Universitaire :
2010-2011
Sommaire
|
Introduction
|
1
|
|
Chapitre I : Généralités
|
2
|
|
1.1. Situation géographique de la Baie de Bou Ismail
|
2
|
|
1.2Cdu ilieu
|
2
|
|
. aractéristiques physiques m
|
|
|
2. Pdu de Bou Haroun
|
3
|
|
résentation port
|
|
|
3. Présentation de étudiée
|
|
|
l'espece
|
4
|
|
3.1. Présentation de la famille des Sparidés
|
4
|
|
3.2. Position systématique
|
.4
|
|
3. 3. Noms vernacuculaires
|
6
|
|
4.Répartition géographique ....
|
... .6
|
|
5. Régime alimentaire
|
.. .6
|
|
6.Reproduction
|
7
|
|
: et traitements au
|
8
|
|
Chapitre II Echantillonnage laboratoire
|
|
|
1. Méthodes d'étude
|
8
|
|
|
|
es tailles
|
8
|
|
1. 1. Fréquenc
|
|
|
|
|
1.2. Calcul des tailles moyennes
|
9
|
|
|
|
1. 3. Etude du sex ratio
|
10
|
|
2. Résultats
|
... 11
|
|
2.1. Fréquences-tailles
|
11
|
|
2.2. Les tailles moyennes
|
13
|
|
2.3. Le sex-ratio
|
...................13
|
|
Chapitre III : Etude des paramètres biologiques
...
|
14
|
|
1. Etude de l'age
|
... 14
|
|
1.1Méthode indirecte dedétermination de l'age
|
....14
|
|
Méthode de Bhattacharya (1967)
|
|
|
1.2. Résultats et discussion
|
15
|
|
2. Étude de la croissance
|
21
|
|
2.1 Croissance linéaire de Von Bertalanffy (1938)
|
21
|
|
- Estimation des parametres de croissance à l'aide du
logiciel FISAT II version (1.2.0)
|
|
|
2.1.1. Résultats
|
22
|
|
2.1.2. Discussion
|
27
|
|
2.2 Relation taille-poids
|
27
|
|
2.2.1 Méthode d'étude
|
.27
|
|
2.2.2. Résultats
|
30
|
|
2.2.3 Discussion
|
32
|
|
2.3. Croissance pondérale
|
33
|
|
2.3.1 Méthode d'étude
|
33
|
|
2.3.2. Résultats
|
33
|
|
2.3.3 Discussion
|
35
|
Introduction
Introduction
Introduction
Du fait de sa position dans le bassin
méditerranéen, avec plus de 1200 Km de côte,
l'Algérie est riche en stock plurispécifique et
spécialement en petits pélagique, ces derniers constituent un
potentiel halieutique important le long de la cote algérienne
(Klouch, Kheloui, 2010).
Parmi les ressources halieutiques dont dispose
l'Algérie, la famille des Sparidés qui présente un
potentiel économique considérable, est largement
représenté dans les captures des petits métiers et des
chalutiers (Harchouche, 1988).
Parmi les représentants de cette famille, Pagellus
acarne, connu sous le nom de (mafroune) a fait l'objet de plusieurs
recherches dans différentes régions, portant sur la biologie
(croissance, sexualité et régime alimentaire) : (Lamrini,
1983 et 1988 ; Andoloro 1982 et 1983, in Kamar,
2000) ; (Mennes, 1985 ; Stergiou et al., 1997 ;
Pajuelo & Lorenzo, 2000, in Coelho et al., 2005);
Harchouche, (1988); Benadda et Kaced, (1994) ; Brahimi, (1998) ; Rizkalla
et al., (1999); Dahamni, (2000) ; Kamar , (2000) ; Arculeo et
al., (2000) ; Coelho et al., (2005). Kheloui et
Klouch ;(2010).
Ainsi la présente étude s'articule autour de 3
chapitres :
· Le premier chapitre est consacré à la
présentation de la zone d'étude ainsi que l'espèce
étudiée ;
· Le deuxième chapitre est relatif à
l'échantillonnage et aux différents traitements au laboratoire
;
· Le troisième chapitre est réservé
à la détermination des paramètres biologiques, à
savoir l'âge, la croissance, et les mortalités
Chapitre premier
< Généralités>>
Chapitre I : Généralités
1. Présentation de la zone d'étude :
La Baie de Bou Ismail, anciennement appelée
Castiglione, est l'une des plus importante baie de la
côte algérienne car elle abrite plusieurs ports d'importance
économique notamment le port de Bou Haroun.
1.1. Situation géographique de la Baie de Bou Ismail
:
Située dans la partie centrale du littoral
algérien, à 50 km à l'Ouest d'Alger dans la wilaya de
Tipaza, entre 2°25' à l'Ouest et à 2°55' à
l'Est.
La baie est limitée par de la pointe de Cap Caxine
(Ras Acrata) à l'Est de Sidi-Fred], et par le Mont Chenoua
à l'Ouest, par la plaine de la Mitidja au Sud et la mer
méditerranéenne au Nord. (Figure 1).L'ouverture de la baie de
Bou-Ismail est d'environ 40 Km, et s'oriente du Sud-Ouest à Nord-Est.
Cette baie couvre une surface de l'ordre de 350 Km2. (Braik,
1989, in Haddouche, 2003).La baie est le
réceptacle d'oued à régime irrégulier : Mazafran,
Nador et Béni-messous. (Khaouni, 2003).
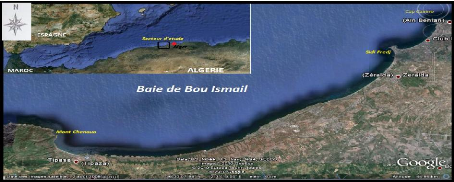
Figure 1. Situation géographique de la
baie de Bou Ismail ( Google Earth, 2009).
1.2. Caractéristiques physiques du milieu :
Le plateau continental de la baie de Bou Ismail, étant
très étroit, atteint un maximum de
13Km au niveau du mont
Chenoua, et un minimum de 3Km entre Sidi Fred] et Alger,
couvrant une
surface de 509Km, dont 70% sont consacrés au chalutage (Chavance
et
Girardin, 1986). La largeur moyenne du plateau
continental est estimée au maximum à 55Km.
La couverture sédimentaire provient de l'érosion
continentale et sous marine ainsi que de la production
biologique.
La baie de Bou Ismail est caractérisée par un
climat sub-humide, avec des étés chauds et secs et des hivers
tempérés doux et humides (Office National de la
Météorologie O.N.M, 2003).
Les mesures effectuées par Haddouche,
(2003), et Khaouni, (2003) dans la baie de Bou
Ismail, montrent que les températures des eaux superficielles sont de 22
à 24°C en juin et juillet, et de 16.4 et 18.5°C au mois de
mars et mai. Concernant la salinité, elle se situe entre 36.3 Psu
à 37.6 Psu avec une moyenne de 36.8 Psu pour le mois de mars, et entre
23.5 Psu et 37.2 Psu avec une moyenne de 35.65 psu pour le mois de mai.
2. Présentation du port de Bou Haroun :
Le port de Bou Haroun est situé à 45 Km
à l'Ouest d'Alger au centre de la baie de Bou Ismail. Orienté
Nord-est ce port ne possède aucun abri naturel. Il est classé
comme le premier port de pêche dans la région centre
E.G.P.A.P, (2006)
La zone de pêche de Bou Haroun est comprise dans la
vaste baie de Bou Ismail, en 1981, les coordonnées
géographiques relevées sur le port sont de 2° 39'50»de
longitude Est, et de 36°37' de latitude Nord. L.E.M,
(2005)
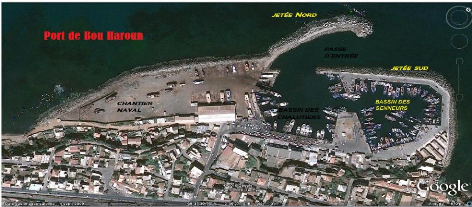
Figure 2. Situation géographique du port
de Bou Haroun (Google Earth, 2009).
Ce port présente :
- La facilité de commercialisation sur Alger.
- La population locale qui compte plusieurs familles de
pêcheurs depuis des générations. - La présence des
services de réparation.
- Son bon emplacement par rapport aux zones de pêches de
côtes algériennes.
Selon le C.N.D.P.A, (2007) la flottille au
port de Bouharoun est passée de 170 unités en 1999 à 227
unités en 2006, avec une légère diminution du nombre des
chalutiers de 2002 à 2006 suite à la saturation de ce port, mais
selon Bachouche, et al (2007) le
port de Bouharoun abrite que 22 chalutiers, 50 sardiniers et 60 petits
métiers.
3. Présentation de l'espèce
étudiée :
3.1. Présentation de la famille des Sparidés
:
La famille des sparidés est une famille de poissons
osseux (téléostéens) comprenant près de 38 genres
(Nelson, 2004).
Le nom de cette famille est dérivé du mot grec
Sparoïdes (qui veut dire poisson à
tête dorée). Communément appelés «
brèmes de mer », les sparidés se caractérisent par un
corps oblong, comprimé sur les flancs et portant des écailles
cténoïdes. Ils présentent une seule nageoire dorsale longue,
en partie épineuse, des pectorales généralement longues et
pointues, deux pelviennes thoraciques, une nageoire annale et une caudale
fourchue.
Les dents bien développées sont
différenciées en dents coniques (caniniformes), aplaties
(incisiformes) ou en pavé (molariformes) La dentition des
sparidés constitue une clé de détermination des genres et
des espèces. (Fischer et al, 1987).
3.2. Position systématique :
Grâce aux avancées de la biologie
moléculaire, les caractères retenus dans les classifications ont
atteint une diversité étonnante. En effet aux observations
morphologiques ou anatomiques classiques (la forme des écailles, la
nature du tissu osseux, l'allure des mâchoires ou le squelette)
s'ajoutent des caractères moins apparents, contenus dans la
séquence de grosses molécules biologiques (protéines, ADN,
ARN) (Léopold, 2004.in kheloui et al 2010)
La classification phylogénétique des
Ostéichtyens a été retenue pour le présent travail
selon (Lecointre et Le Guyader, 2001)
Règne : Eukaryota
Sous-Règne : Metazoa
Phylum : Chordata
Sous-Phylum : Craniata
Embranchement : Vertebrata
Super-Classe : Gnathostoma
Classe : Osteichthyes
Sous-Classe : Actinoptérygii
Super-Ordre : Teleosteei
Ordre : Perciformes
Sous-ordre : Percoidei
Famille : Sparidae
Genre : Pagellus Valenciennes, 1830
Espèce : acarne Risso, 1826.

Figure 3. Morphologie de Pagellus
acarne.( Kheloui et al., 2010)
3. 3. Noms vernacuculaires :
Harchouche, (1988) note que les noms
vernaculaires de ce poisson sont nombreux et diffèrent d'un pays
à un autre (tableau 1).
Tableau 1- Noms vernaculaires de
Pagellus acarne dans différents pays.
Pays
|
Noms vernaculaires
|
Algérie
|
Mafroune, Besoug
|
Maroc
|
Besoug, Bouka. Bogue
|
Tunisie
|
Mourjane
|
Angleterre
|
Axillary seabream
|
France
|
Pageot acarné, pageot blanc
|
Espagne
|
Aligote, Besugo
|
Italie
|
Pagello bastardo
|
|
4 .Répartition géographique :
Pagellus acarne présente une large distribution
géographique ; il se rencontre en Méditerranée, dans les
côtes Est de l'océan Atlantique du Golfe de Gascogne au
Sénégal, au niveau des Açores, Madère, des
îles Canaries et du Cap-Vert. (Parenti et al, 2004 in.
Kheloui et al, 2010).
Le pageot blanc est une espèce démersale
rencontrée sur des fonds variés, principalement sableux et
sablo-vaseux jusqu'à 500 m de profondeur mais il est plus commun entre
40 et 100 m de profondeur (Coelho et al., 2005).
5.Régime alimentaire :
Selon Fischer et coll. (1987), Pagellus
acarne fait partie des espèces omnivores à
prédominance Carnivore.
L'alimentation de Pagellus acarne consiste
principalement en crustacés (proie préférentielle),
poissons téléostéens et échinodermes, en plus des
petites quantités de céphalopodes et de polychètes Selon
les mêmes auteurs, le régime alimentaire diffère selon la
taille des individus. D'après les annélides polychètes
constituent souvent le groupe le plus représenté parmi les
proies, quelque soit l'âge du poisson et la saison. (Harchouche,
1988) in (kheloui et al., 2010)
La composition du régime alimentaire dépend de
l'abondance saisonnière et la disponibilité des proies dans le
milieu ; Rizkalla et al.,1999).in( Arkoub et al.,
2010)
6. Reproduction :
Pagellus acame est caractérisé par un
hermaphrodisme de type protandrique ;(les individus sont mâles au
début de leur vie, et deviennent femelles en vieillissant)
(Arculeo et al., 2000). Il existe également
des mâles et des femelles à développement direct.
Le cycle sexuel de P.acarne est long ; ce poisson a
une reproduction assez étalée dans le temps (Harchouche,
1988). Il présente une reproduction intermittente, de juin
à septembre en Méditerranée occidentale et de septembre
à novembre en Méditerranée orientale (FISCHER
et al., 1987). L'intensité maximale de la reproduction
se situe entre juillet et septembre pour les deux sexes (Andaloro, 1982
). En baie d'Alger elle a été déterminée
au mois de mai par Harchouche, (1988).
La taille à la première maturité sexuelle
de Pagellus acarne a été estimée à 19,85
cm au Maroc par Lamrini (1988) et à 18,3 cm en baie
d'Alger (Harchouche, 1988).
Chapitre deuxième
< Echantillonnage et traitements au
laboratoire»
Chapitre II : Echantillonnage et traitements au
laboratoire
Dans le domaine des recherches halieutiques le mode de
prélèvement des échantillons est une phase importante, car
il n'est pas facile d'obtenir un échantillon qui soit
représentatif de la population étudiée. Il constitue l'un
des problèmes les plus difficiles à résoudre pour le
scientifique dans le but d'obtenir les paramètres biologiques
(kheloui et al., 2010 in Dahamni, 2000).
Les données de Pagellus acarne
utilisées dans la présente étude proviennent des
échantillonnages effectuées durant la période
s'étalant du 11 mars au 27 avril 2011. Ces échantillons ont
été achetés au niveau du point de vente d'un
pécheur à Bologhine qui s'approvisionne au port de Bouharoun.
284 individus de Pagellus acarne ont fait l'objet de
cette étude. Ils mesurent entre 8.5 et 24 cm et sont réparties en
111 mâles, 117 femelles, et 57 indéterminés (tableau 2 en
p). Notre échantillon comporte surtout les tailles moyennes qui sont les
plus représentatives.
Les poissons ont été mesurés, ce qui
nous a permis d'établir la distribution de fréquencestailles de
l'échantillon et de calculer sa taille moyenne. Les mesures et les
pesées serviront par la suite à l'étude de la croissance
linéaire et pondérale , l'étude de l'age ; Les poissons
ont été ensuite disséqués, afin de
déterminer les sexes et d'étudier le sex-ratio .
1. Méthodes d'étude
1. 1. Fréquences-tailles
? Mensurations et pesées
Il existe de nombreuses mensurations possibles pour un
poisson. Nous avons effectué sur chaque individu 3 types de mesures (au
millimètre près) à l'aide d'une règle à
butoir appelée également ichtyomètre (figure 4). Ces
mesures sont définies comme suit :

LT, longueur totale du poisson ; comprise entre
l'extrémité de la bouche et l'extrémité de la
nageoire caudale .
Ls, longueur standard ; prise de la bouche jusqu'au niveau de
l'origine des rayons de la nageoire caudale.
LF, longueur à la fourche ; prise du bas du maxillaire
jusqu'au creux de la fourche
Seule la longueur à la fourche LF a été
utilisée pour les études ultérieures car selon
Korichi (1988), cette longueur réduit les risques d'erreurs de
mesure.
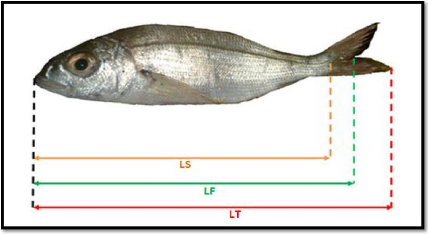
Figure 4. Différentes mensurations
effectuées sur Pagellus acarne.
Afin de déterminer le poids des individus, ceux-ci ont
été pesés au gramme près à l'aide d'une
balance de précision. Ainsi, les poids totaux (Wt), et les poids
éviscérés (We) sont relevés .
· Les mesures effectuées ont permis de calculer la
taille moyenne des males et des femelles et l'intervalle de confiance
1.2. Calcul des tailles moyennes
LF~= Ó ni.LF~
N~
LF ?= Ó ni.LF?
N?
LF : longueur à la fourche de chaque classe de taille ni
: effectif de chaque classe de taille
N : effectif total
· Calcul de l'intervalle de confiance
I.C : m #177; 1.96 * S

n
S : écart type n : effectif.
Nous avons utilisé un test de comparaison des tailles
moyennes observées et des tailles moyennes théoriques afin de
préciser le taux de signification de ces tailles moyennes.
Le test de comparaison des moyennes est basé sur
l'écart réduit calculé
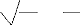
|? cal| = | ma - mb|
Sa 2 + Sb 2
na nb
et : Sa2 = Ó ni * (LF -
LF?)2
N? - 1
Sb 2 = Ó ni * (LF - LF~)2


N - 1
ma : taille moyenne des femelles mb : taille moyenne
des mâles S2a : variance des femelles
S2b : variance des mâles
na et nb : nombre des femelles et des mâles
respectivement.
1. 3. Etude du sex-ratio
Les poissons ont été disséqués
pour prélever les gonades et déterminer les sexes, ce qui nous a
permis d'étudier le sex-ratio. Cette étude permet d'estimer la
proportion des individus de chaque sexe dans une population.
Le sexe ratio est le nombre de mâles par rapport au nombre
de femelles sur le nombre de femelles
Sex-ratio = M / F
F : nombre de femelles M : nombre de mâles
Afin de vérifier le résultat obtenu, on doit
utiliser un test de comparaison des pourcentages observés
et des
pourcentages théoriques en calculant l'écart réduit selon
la formule de Schwartz, (1993)
servant à
préciser le taux de signification du sex-ratio global :
|? cal | = p0 - p
p * q
n
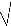
p0 : pourcentages observés
p et q : pourcentages théoriques =
0.5%
n : effectif total des mâles et des
femelles.
On compare la valeur de l'écart réduit
?cal à 1.96 au risque á=5%
Deux cas de figure peuvent se présenter :- Si
:?cal= 1.96 la différence est non significative entre le
pourcentage des mâles et celui des femelles, c'est-à-dire qu'il y
a autant de male que de femelle -Si ?cal> 1.96 la
différence est significative. ( Amrani R ., Meghouche M
A., 2010).
2. Résultats :
2.1. Fréquences-tailles
Tous les individus (284 individus) sexes confondus et de tailles
allant de 8.5 à 24 cm, sont repartis en classes de tailles avec un pas
de 0.5 cm (tableau 1, figure 2).
Tableau 2 - Fréquences-tailles des
individus de Pagellus acarne de la baie de Bou-Ismaïl,
échantillonnés durant la période de (mars-avril 2011).
Cc (cm)
|
Femelles
(N)
|
Male
(N)
|
Indéterminés
(N)
|
Sexes
confondus
(Nt)
|
8.5
|
0
|
2
|
1
|
3
|
9
|
2
|
5
|
0
|
7
|
9.5
|
0
|
4
|
5
|
9
|
10
|
3
|
5
|
2
|
10
|
10.5
|
3
|
5
|
0
|
8
|
11
|
1
|
3
|
1
|
5
|
11.5
|
1
|
1
|
1
|
3
|
12
|
2
|
3
|
2
|
7
|
12.5
|
3
|
7
|
3
|
13
|
13
|
1
|
9
|
4
|
14
|
13.5
|
5
|
10
|
3
|
18
|
14
|
5
|
1
|
4
|
9
|
14.5
|
8
|
12
|
8
|
28
|
15
|
4
|
10
|
3
|
17
|
15.5
|
2
|
4
|
2
|
8
|
16
|
1
|
0
|
1
|
2
|
16.5
|
4
|
5
|
0
|
9
|
17
|
2
|
3
|
1
|
6
|
17.5
|
3
|
3
|
2
|
8
|
18
|
2
|
0
|
0
|
2
|
18.5
|
2
|
1
|
1
|
4
|
19
|
6
|
4
|
3
|
13
|
19.5
|
2
|
1
|
0
|
3
|
20
|
2
|
0
|
0
|
2
|
20.5
|
1
|
1
|
0
|
2
|
21
|
1
|
0
|
1
|
2
|
21.5
|
2
|
0
|
0
|
2
|
22
|
13
|
0
|
1
|
14
|
22.5
|
1
|
0
|
0
|
1
|
23
|
8
|
2
|
2
|
12
|
23.5
|
15
|
5
|
4
|
24
|
24
|
12
|
5
|
2
|
19
|
Effectif total
|
117
|
111
|
57
|
284
|
|
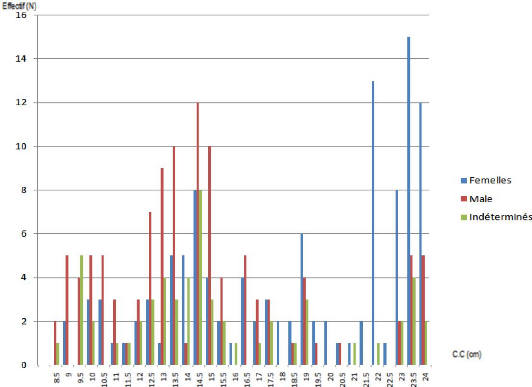
Figure 5. Distribution des fréquences de tailles
de Pagellus acarne en fonction du sexe séparé
.
Concernant les histogrammes de fréquences des mâles
et des femelles, on distingue deux phases :
· Première phase (de 8.5 à 15.5 cm) : les
mâles sont plus abondants que les femelles dans les premières
classes de tailles.
· Deuxième phase (de 16.5 à 24 cm) : les
femelles deviennent plus abondantes dans les dernières classes de
tailles.
Ces différences dans la distribution de
fréquences pourraient être liées au caractère
hermaphrodite protandrique de cette espèce (les individus sont
mâles au début de leur vie, et deviennent femelles en
vieillissant) (Arculeo et al., 2000). Concernant les
individus hermaphrodites, ceux-ci sont présents dans presque toutes les
classes de tailles.
Figure 6. Distribution des fréquences de tailles
de Pagellus acarne (sexes confondus)
On remarque dans la ( figure 6) que la courbe des individus du
mafroune (sexes confondus) présente 2 modes, le plus important se situe
à 14.5 cm et le plus petit à 22.5 cm.
On remarque également l'absence des individus de
petite taille et de grande taille, les poissons étant triés et
rangés dans des caisses selon leur calibre. Selon Harchouche
(2006), les captures des bateaux ne sont pratiquement jamais
représentatives de la population dans son ensemble étant
donné que dans la plupart des cas, ces bateaux rejettent en mer les
poissons de petites tailles.
2.2. Les tailles moyennes :
LF~ = 16.3 #177; 0.27 cm LF? = = 17.4#177; 0.27 cm
D'après ces résultats, la taille moyenne des
femelles de Pagellus acarne serait supérieure à celles
des mâles seul le teste de l'écart réduit confirmerait se
résultat. La valeur de |? cal | calculée, équivalente
à 2.54 est supérieure à celle donnée par
l'écart réduit au risque de (5%) (1,96) (t table) . La
différence est donc significative, les femelles sont plus grandes que
les mâles dans notre échantillon (2,54>1,96)
2.3. Le sex-ratio :
La valeur du sex-ratio pour le mafroune de la baie de Bou Ismail
est de :
Sex-ratio global = 0.85
La sex-ratio globale serait en faveur des femelles (Sex-ratio
global < 1). Afin de confirmer cette hypothèse, le test de
l'écart réduit est calculé et le résultat indique
une différence non significative (t calculé < t table) (0.97
< 1.96), donc il y a autant de mâles que de femelles.
Néanmoins, notons que la sélectivité des engins de
pêche et la répartition bathymétrique différente
entre les males et les femelles interviennent dans l'accessibilité de
ces derniers dans les captures.
Chapitre troisième
<< Etude des paramètres
biologiques»
Chapitre III : Etude des paramètres biologiques
1. Etude de l'âge :
L'estimation de l'âge est indispensable à
l'étude de la biologie et de la dynamique des populations de poissons.
Cette estimation forme la base pour des calculs menant à une
connaissance de la croissance, de la mortalité, du recrutement et
d'autres paramètres fondamentaux de leurs populations (Laurec
& Le Guen (1981), in Mahé et al.,
2009).
La détermination de l'age des poissons
téléostéens peut être réalisée selon
deux approches différentes :
· Les méthodes directes :
basées sur l'examen des pièces calcifiées
(écailles, otolithes, épines, vertèbres), ou sur des
techniques de marquages.
· Les méthodes indirectes :
basées sur l'étude des données de
fréquence de tailles, telles que la méthode de (Pettersen, 1891
in Sparre et Venema, 1996), de (Bhattacharya, 1967, in sparre
et Venema, 1996), de (Hasselblad, 1966 in Damir, 2008).
1.1. Méthode indirecte de détermination de
l'âge :
? Méthode de Bhattacharya (1967) :
Cette méthode a été préconisée
pour le traitement des données regroupées issus du laboratoire
l'halieutique.
Elle permet de décomposer une population en
sous-populations, en cohortes ou classes d'age. Elle se base sur une
transformation logarithmique des effectifs regroupés de taille
d'égale amplitudes `h' et de centre de classes.
On construit un graphe en portant en ordonnées pour chaque
centre de classes la quantité suivante :
Ä Log Z = Log Z(x+h) - Log Z (x)
Avec respectivement :
Z (h+x) : effectifs de la classe de longueur de centre de classe
(X+h)
Z (x) : effectif de la classe de longueur
précédente, de centre de classe (x)
On recherche les droites de pentes négatives, sur le
graphe.
Afin d'appliquer cette méthode, il est indispensable que
l'histogramme ne contienne pas des classes de taille vides, et l'intervalle h
doit être petit par rapport à chacun des écarts-types
(h/ö=2.2).
Le nombre de sous-populations correspond au nombre de droites
à pente négatives obtenues, la taille moyenne de chaque
sous-population se détermine en relevant graphiquement le point
d'intersection de ces droites avec l'axe des abscisses :
L moy = ? + (h/2)
Avec :
h : intervalle de classe
? : point d'intersection de la droite de pente négative
avec l'axe des abscisses. L moy : taille moyenne de la composante (groupe
d'age)
Les différents paramètres (moyenne ; écart
type ; variance ; effectifs) de chaque sous population sont obtenus par le
logiciel `FISAT II' (Gayanilo et al
,1997).
Les limites de confiances sont données par l'expression
suivante : P = (t* s.d.) / v N Avec respectivement : Sd :
écart-type, N : effectifs de la population, et t=1.96 pour
á=5%.
1.2. Résultats et discussion :
la méthode de Bhattacharya en utilisant la méthodes
manuelle (Microsoft Excel 2007) (tableau 3,4,figure7
) et celle avec le logiciel (FISAT II, Gayanilo et al.
2005) (tableau 8, figure 9,10)
Les données de fréquences-tailles utilisées
dans la méthode de Bhattacharya à l'aide de la méthode
manuelle (Microsoft Excel 2007) (tableau 3).
Tableau 3- Données de Fréquences
de tailles de Pagellus acarne dans la baie de Bou-Ismaïl.
|
Cc (cm)
|
Sexes confondus (Nt)
|
Ln(Nt)
|
?Ln(Nt)
|
|
8.5
|
3
|
1.0986
|
0.8473
|
|
9
|
7
|
1.9459
|
0.2513
|
|
9.5
|
9
|
2.1972
|
0.1054
|
|
10
|
10
|
2.3026
|
-0.2231
|
|
10.5
|
8
|
2.0794
|
-0.4700
|
|
11
|
5
|
1.6094
|
-0.5108
|
|
11.5
|
3
|
1.0986
|
0.8473
|
|
12
|
7
|
1.9459
|
0.6190
|
|
12.5
|
13
|
2.5649
|
0.0741
|
|
13
|
14
|
2.6391
|
0.2513
|
|
13.5
|
18
|
2.8904
|
-0.6931
|
|
14
|
9
|
2.1972
|
1.1350
|
|
14.5
|
28
|
3.3322
|
-0.4990
|
|
15
|
17
|
2.8332
|
-0.7538
|
|
15.5
|
8
|
2.0794
|
-1.3863
|
|
16
|
2
|
0.6931
|
1.5041
|
|
16.5
|
9
|
2.1972
|
-0.4055
|
|
17
|
6
|
1.7918
|
0.2877
|
|
17.5
|
8
|
2.0794
|
-1.3863
|
|
18
|
2
|
0.6931
|
0.6931
|
|
18.5
|
4
|
1.3863
|
1.1787
|
|
19
|
13
|
2.5649
|
-1.4663
|
|
19.5
|
3
|
1.0986
|
-0.4055
|
|
20
|
2
|
0.6931
|
0
|
|
20.5
|
2
|
0.6931
|
0
|
|
21
|
2
|
0.6931
|
0
|
|
21.5
|
2
|
0.6931
|
1.9459
|
|
22
|
14
|
2.6391
|
-2.6391
|
|
22.5
|
1
|
0.0000
|
2.4849
|
|
23
|
12
|
2.4849
|
0.69315
|
|
23.5
|
24
|
3.1781
|
-0.2336
|
|
24
|
19
|
2.9444
|
|
|
Effectif total
|
284
|
5.6490
|
|

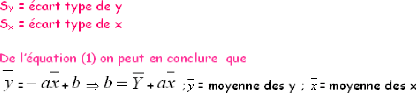
Les études de ces différents droits de
régression sont récapitulées dans le
(tableau4)
Tableau 4 - Clé âge longueur et droits de
régressions méthode manuelle (Microsoft Excel 2007) de
Pagellus acarne de la baie de Bou Ismail.
|
S /P
|
Equation de droits de régressions
|
Groupes d'ages
|
longueurs moyennes
|
|
|
(ans)
|
(Cm)
|
|
|
|
Sp1
|
Y=-0.55X+0.000107
|
I.
|
10.17
|
|
Sp2
|
Y=-0.78X+8.65
|
II.
|
12.32
|
|
Sp 3
|
|
III.
|
14.35
|
|
Sp4
|
Y=-1.66X+24.18
|
IV.
|
16.19
|
|
Sp5
|
|
V.
|
18.91
|
|
Sp6
|
|
VI.
|
19.7
|
|
Sp7
|
|
VII.
|
21.1
|
|
Sp8
|
|
VIII.
|
22.9
|
|
Sp9
|
Y=-2,76X+62,443
|
IX.
|
23.8
|
Remarque : on n'a pas besoin de déterminé
les équations des droites de régressions lorsque l'on a deux
points successifs dans le négatif, on trace directement la droite.
La figure 7 obtenue à l'aide de la
méthode manuelle (Microsoft Excel 2007) donne 9 groupes
d'age avec les tailles allant de : (10.17Cm -23.8Cm) est
illustrée ci-dessous.
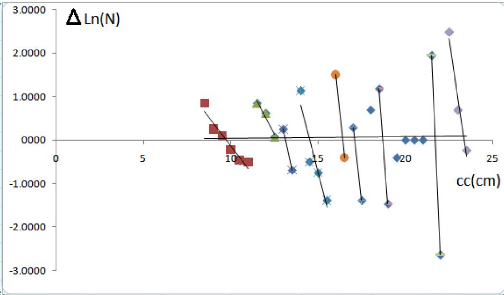
Figure 7 - Décomposition de la distribution de
fréquences de tailles de Pagellus acarne sexes confondus par la
méthode de Bhattacharya(1967)
A partir de la distribution des fréquences de taille le
logiciel (FISAT II, Gayanilo et al. 2005) a
été utilisé pour obtenir la clé âge longueur
consignée les résultats sont représenté dans
(tableau4) et dans (figures 8 et 9) qui montrent 7 groupe
d'ages allant de (10,17Cm-23,8Cm)
Tableau 5 - Clé âge longueur de Pagellus
acarne de la baie de Bou Ismail.
|
Groupes d'ages
(ans)
|
longueurs
moyennes (Cm)
|
s.d
|
Population
(effectifs Ni)
|
S .I
|
|
I
|
10.17 #177; 0.15
|
1.03
|
45.00
|
n.a
|
|
II
|
13.32 #177; 0.13
|
0.86
|
34.00
|
2.56
|
|
III
|
15.35 #177;0.16
|
1.35
|
64.00
|
3.35
|
|
IV
|
17.19 #177;0.17
|
1.25
|
17.00
|
2.65
|
|
V
|
19.31#177;0.22
|
0.96
|
20.00
|
2.62
|
|
VI
|
21.6#177; 0.5
|
1.4
|
17.00
|
1.63
|
|
VII
|
23.8#177; 0.12
|
1.1
|
55.00
|
1.71
|
S.d : écart type, S.I : indice de séparation
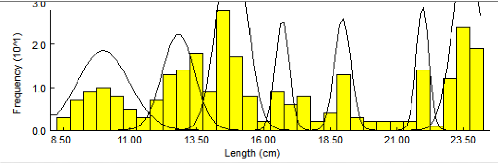
Figure 8. Décomposition de la
distribution de fréquences de tailles de Pagellus acarne sexes
confondus par la méthode de Bhattacharya(1967) (FISAT II 1.2.0,
Gayanilo et al2005).
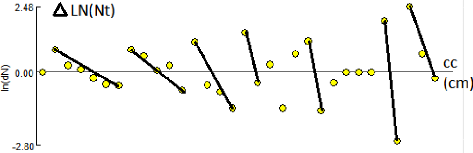
Figure 9 - Décomposition de la
distribution de fréquences de tailles de Pagellus acarne sexes
confondus par la méthode de Bhattacharya(1967), (FISAT II 1.2.0,
Gayanilo et al2005).
Discussion :
La décomposition en cohortes de la méthode de
Bhattacharya(FISAT II 1.2.0, Gayanilo et al2005),
nous donne 7 groupes d'age, et celle faite par la main
(Microsoft Excel 2007) donne 9 groupe d'ages
Le logiciel (FISAT II 1.2.0, Gayanilo et
al2005) a fait fusionner les classes dont
l'effectif total et réduit
On remarque d'après les résultats de la clé
âge-longueur que les taux de croissance ne diminuent pas en fonction de
l'age, cela pourrait s'expliquer par l'absence des petits et des grands
individus de notre échantillonnage qui est beaucoup plus
représenté par les tailles moyennes.
Selon Sparre et Venema (1996), les
données d'entrée destinées à l'analyse de
Bhattacharya sont fréquemment biaisées en raison de la
sélectivité des engins de pêche et du recrutement,
c'est-à-dire que les petits poissons sont sous-représentés
dans les échantillons de fréquence, soit qu'ils
s'échappent par les mailles des filets soit qu'ils n'aient pas encore
migré des zones d'alevinage vers les zones de péche
Les tailles moyennes par groupe d'age de Pagellus acarne
selon la littérature et dans différentes régions sont
regroupées dans le (tableau 6).
Tableau 6 - Clés âge-longueur de
Pagellus acarne selon la littérature.
|
Groupes d'âge (an)
|
|
Auteurs
|
Région
|
Méthode
d'observatio
n
|
0
|
I
|
II
|
III
|
IV
|
V
|
VI
|
VII
|
VIII
|
IX
|
|
Lamrini
(1988) (Lf)
|
Tanger
|
-
|
10.04
|
13.28
|
17.1
|
19.57
|
22.29
|
25.73
|
26.97
|
29.
27
|
30.5
|
31.5
|
|
Harchouche
(1988) (Lf)
|
baie d'Alger
|
Scalimètrie
|
-
|
11
|
13.94
|
15.39
|
17.61
|
20.69
|
23 87
|
25.
6
|
27.5
|
-
|
|
Benadda et al
(1994)
|
baie de
Bou-Ismaïl
|
Otolithométri
e
|
8.75
|
12.39
|
14.52
|
17.93
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Dahamni
(2000)
|
baie de
Bou-Ismaïl
|
Otolithométri e
|
-
|
9.83
|
12.29
|
13.76
|
15.86
|
17.83
|
19
|
20.
5
|
-
|
-
|
|
.( Kheloui et al., 2010)
|
baie de
Bou-Ismaïl
|
Bhattacharya (1967)
(FISAT II
1.2.0, Gayanilo et
al2005).
|
-
|
-
|
15.35
|
16.56
|
17.41
|
18.73
|
20.56
|
-
|
-
|
-
|
|
Présent
travail (2011)
|
baie de
Bou-Ismaïl
|
Bhattacharya (1967)
(FISAT II
1.2.0, Gayanilo et
al2005).
|
-
|
10.17
|
13.32
|
15.36
|
17.19
|
19.31
|
21.6
|
23.8
|
|
-
|
Discussion :
On remarque des différences notables entre les
résultats des tailles moyennes obtenus par chaque auteur, autant pour la
détermination du nombre de groupes d'age que pour la
détermination de la taille moyenne leur correspondant. Ceci pourrait
s'expliquer par l'emploi de méthodes différentes dans
l'estimation de l'âge, les caractéristiques chimiques de l'eau de
mer qui diffèrent d'une région à l'autre, ainsi
qu'à la qualité de l'échantillonnage ainsi que le nombre.
(Knudsen, 1962 ; Bernt et al., 1965 ; Chen
et al., 1999, in Dahamni,
2000).
Nos résultats sont le fruit de notre propres
échantillonnages (284 individus) contrairement aux études
précédente qui on compilée d'autres résultats afin
d'avoir des résultats plus précise
> On utilise les résultats de la méthode de
Bhattacharya(FISAT II 1.2.0, Gayanilo et
al2005). 7 groupes d'age pour la suite de notre
travaille.
> Le nombre de groupe d'age obtenue par taille moyenne est a
prendre avec réserve vue le nombre faible d'effectif par taille.
2. Étude de la croissance :
L'estimation des paramètres de croissance est un
élément essentiel pour l'application des modèles
halieutiques afin de déterminer l'état d'un stock
donné.
2.1 Croissance linéaire de Von Bertalanffy (1938)
:
Parmi les modèles qui décrivent la croissance
des populations, le modèle de VON BERTALANFFY (1938), est le
modèle le plus utilisé dans les études de croissance en
biologie des pêches, car il est très facile à l'emploi, il
nécessite la connaissance de trois paramètre L8, K, t0. Il
présente l'avantage de pouvoir être facilement incorporé
dans des méthodes d'évaluation des stocks et
particulièrement celui de BEVERTON ET HOLT (1957). (BA Ibrahima,
1988).
Ce modèle est basé sur une hypothèse
selon laquelle la vitesse instantanée de croissance est le
résultat de deux processus physiologiques opposés : l'anabolisme
proportionnel à la surface des organismes et le catabolisme
proportionnel au volume de leur corps (poids).
L t= L8 (1- e- k (t- t0)
)
L t : Longueur du poisson à l'instant t.
L8 : Longueur asymptotique théorique qui serait
atteinte par le poisson s'il croit indéfiniment,
à ne pas confondre avec la taille maximale atteinte par le
poisson.
K : Coefficient de croissance ou vitesse de croissance,
caractérisant la vitesse avec laquelle l'espèce croit vers sa
taille asymptotique (L8).
to : Age théorique auquel la longueur du
poisson est nulle (ans).
- Estimation des paramètres de croissance à
l'aide du logiciel FISAT II version (1.2.0) :
Recommandé par la FAO, le logiciel FISAT II est
basé sur l'équation de Von Bertalanffy (1938), il permet de
fournit initialement des évaluations des différents
paramètres, puis il utilise des algorithmes itératifs.
Les couples de données âge-longueur obtenus par la
méthode de Bhattacharya, sont introduits dans le programmes FISAT II
version (1.2.0) (Gayanilo et al,
2005).
2.1.1. Résultats :
La taille asymptotique L8 obtenue par logiciel FISAT II
L8= 24.01 Cm
La taille asymptotique L8 obtenue par la méthode de
pauly (L8= Lmax /0.95)
L8= 25.26 Cm
La taille asymptotique L8 obtenue par la moyenne des deux (FISAT
II et pauly) L8= 24.63 Cm
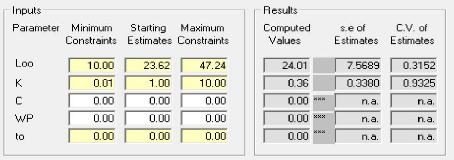
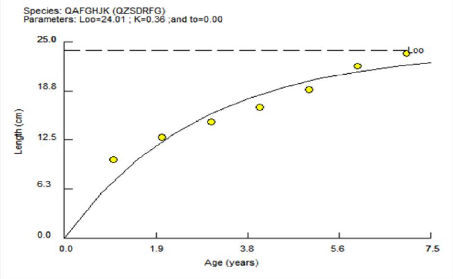
Tableau 7 - Paramètres de croissance de
l'équation de Von Bertalanffy obtenus par le logiciel FISAT II pour
Pagellus acarne de la baie de Bou Ismail.
- Le taux de croissance : K= 0.36 / an.
- La taille asymptotique : L8= 24.01Cm.
- Age théorique auquel la longueur du poisson est nulle
(ans) : to=0 e- k (t- t0) )
L t= L (1-
La détermination du L8 graphiquement par le logiciel
FISAT II version (1.2.0) est représenté par la
(figure 9)
Figure 10- Courbe de croissance linéaire
de l'équation de Von Bertalanffy obtenus par le logiciel FISAT II pour
Pagellus acarne de la baie de Bou Ismail.
Tableau 8 - Paramètres de croissance de
l'équation de Von Bertalanffy obtenus par par le logiciel FISAT II pour
Pagellus acarne de la baie de Bou Ismail.
METHODES
|
PARAMETRES
|
Pauly
|
|
|
FISAT II
version (1.2.0)
|
|
la moyenne des deux
méthodes (FISAT II
et
pauly)
|
|
L8 (Cm)
|
25.26
|
|
|
24.01
|
|
|
24.63
|
|
|
K (/an)
|
|
|
|
0.36
|
|
|
|
|
|
t0 (an)
|
|
|
|
0
|
|
|
|
|
|
Equati de
Von Bertalon anffy
(1938)
|
L t=25.26 (1-6
|
0.36
|
( t))
|
L
t=24.01
(1-e
|
-0(
|
t ))
|
L t=24.63 (1- e-0.36
|
( t ))
|
Les longueurs moyennes observées et théoriques aux
différents groupes d'ages, obtenus avec les paramètres de
croissance
Tableau 9- Les longueurs moyennes
observées et théorique obtenue par Pagéllus
acarné
|
Groupes d'âges
|
Longueurs théoriques
|
Longueurs observés
|
|
1
|
10.17
|
7.44623209
|
|
2
|
12.32
|
12.6412919
|
|
3
|
14.35
|
16.2657622
|
|
4
|
16.19
|
18.7944693
|
|
5
|
18.91
|
20.5586884
|
|
6
|
19.7
|
21.7895423
|
|
7
|
21.1
|
22.6482799
|
|
8
|
22.9
|
23.2474008
|
|
9
|
23.8
|
23.6653933
|
Le tableau 10 nous a permis de tracer une courbe de croissance
linéaire représentant les tailles théoriques et les
tailles observés de pagellus acarné (figure 14)
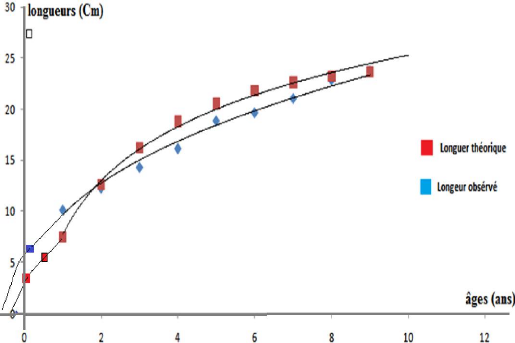
Figure 11- Courbe théorique et
obsevée de croissance linéaire de l'équation de Von
Bertalanffy pour Pagellus acarne de la baie de Bou
Ismail.
Paramètres de la croissance linéaire de
Pagellus acarne selon la littérature et dans différentes
régions sont regroupées dans le (tableau 10).
Tableau 10- Paramètres de la croissance
linéaire de Pagellus acarne selon différents auteurs et
différentes régions.
|
Auteur
|
Région
|
L8 (cm)
|
K (an-1)
|
to (ans)
|
Méthode
|
|
Andaloro (1982)
|
Détroit de
|
29.78*
|
0.3203
|
-0.26
|
Wetherall et al(1986)
|
|
in Dahamini (2000)
|
Messine
|
26.23**
|
0.4187
|
-0.22
|
|
|
Harchouche (1988)
|
Baie
|
37.07
|
0.1455
|
|
|
|
|
|
|
|
-
|
|
d'Alger
|
25 05
|
0.1917
|
-
|
|
|
|
32.80
|
0.164
|
|
|
|
Lamrini in
|
Tanger
|
44.07
|
0.14
|
|
|
|
Harchouche (1988)
|
|
|
|
-
|
|
|
Derdiche et al
|
Béni-Saf
|
24.47
|
0.30
|
|
|
|
(1990)
|
|
|
|
|
|
|
Habib et al (1990)
|
Baie d'Alger
|
38.48
|
0.198
|
-
|
Wetherall et al(1986)
|
|
Belkessam et Issolah (1990)
|
Baie Bou- lsmail
|
27.65
|
0.27
|
|
Wetherall et al(1986)
|
|
Benadda et Kaced (1994)
|
Baie Bou- lsmail
|
22.78
|
0.35
|
0.5
|
Wetherall et al (1986)
|
|
|
52.16
|
0.08
|
-2.23
|
Ford-Walford (1946)
|
|
Dahamni (2000)
|
Baie Bou-
|
32.6
|
0.105
|
-2.4
|
Ford-Walford (1946) Abranson -
|
|
lsmail
|
35.86
|
0.088
|
-2.67
|
Tomlinson(1971)
|
|
Erzini et al (2001)
|
Portugal
|
32.07
|
-0.2
|
-2.6
|
|
|
Coelho et al (2005)
|
Sud du
|
32.3
|
0.18
|
-2.56
|
|
|
Portugal
|
|
|
|
|
|
Kheloui et al., (2010)
|
Baie de Bou Ismail
|
27.55
|
0.29
|
0
|
FISAT II
|
|
Présente étude
|
Baie de Bou
|
24.01
|
0.36
|
0
|
FISAT II
|
|
(2011)
|
Ismail
|
|
|
|
|
2.1.2. Discussion :
La taille asymptotique (L8) obtenue par la méthode de
Pauly (1985) et celle obtenus par la méthode du logiciel FISAT II
(1.2.0) est rapprochée.
Les longueurs moyennes observées et théoriques aux
différents groupes d'ages, obtenus avec les paramètres de
croissance sont similaires, cela nous indique qu'il y a un bon ajustement
entres les tailles théoriques et les tailles observées issus du
FISAT II (1.2.0).
Selon la littérature (tableau 10), la valeur de la
longueur asymptotique estimée par la présente étude (L8=
24.63 Cm), est proche de celle obtenue par.( Kheloui et al.,
2010) et Benadda et Kaced (1994) (dans la baie de Bou
Ismail), En revanche les résultats obtenus par
Lamrini (1988) au Maroc, et Coelho et
al (2005) au Portugal, sont surestimées par rapport
ànos valeurs, cela peut s'expliquer par la diversité
des méthodes utilisées pour l'évaluation des
paramètres de croissance, la sensibilité du
modèle de Von Bertalanffy aux clés âge-longueur
utilisées, ainsi qu'à la qualité de
l'échantillonnage.
Selon Sparre et Venema (1996), la
différence des paramètres de croissance peut exister Ventre les
stocks de la même espèce ; ainsi des cohortes successives peuvent
se développer différemment selon les conditions du milieu.
2.2 Relation taille-poids :
2.2.1 Méthode d'étude :
La relation taille-poids lie le poids d'un poisson à sa
longueur, elle permet de déterminer le poids des individus dont on
connait la taille ou inversement (Dubuit, 1975). Cette
relation est

Wt = a Lt
b
donnée par l'expression :
Wt : Poids du poisson
Lt : Longueur du poisson
a : Constante
b : Coefficient d'allométrie (coefficient de croissance
relative entre le poids et la longueur).
Par une transformation logarithmique, l'équation
linéarisée se présente sous la forme : Ln Wt = b Ln LT +
Ln a
Pour étudier la croissance relative de Pagellus
acarne de la baie de Bou Ismail, 12 couples de valeurs (We, LF) on
été retenus. L'ajustement linéaire se fait par la
méthode des moindres carrés.
Selon la valeur de b, trois cas peuvent se présenter :
b est égal à 3, la croissance est dite
isométrique ; les deux variables We et LF ont le même taux de
croissance(K), le poids croit en même temps que la longueur du
poisson.
b est inférieur à 3, l'allométrie est
minorante ; le poids croit relativement moins vite que la longueur,
b est supérieur à 3, l'allométrie est
majorante ; le poids croit plus vite que la taille de l'individu.
Un test de conformité doit-être fait afin de
confirmer la valeur du paramètre b :

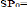



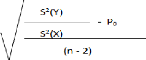
On compare le tcal avec le ttable :
P= 3 pente théorique
P0 : pente observée
S2(Y) : variance de (y= Ln Wt)
S2(X) : variance de (x= Ln Lt)
n-2 : degré de liberté (d.d.l = n-2,
á=5%)
-Si t cal < t table : la différence est non
significative. -Si t cal > t table : la différence est
significative.
Le (tableau-11) récapitule la
distribution des poids totaux par classe de taille regroupé a partir de
la série des données (échantillonnages entre la
période du : du 22 mars au 10 avril 2011), nous a permet
de déterminer les paramètres de la croissance relative
Tableau- 11- Répartition des poids totaux
par classe de taille( Pagellus acarne de la baie de Bou Ismail).
|
CC (cm)
|
Wt (g)
|
|
7.88
|
10.13
|
|
9.22
|
20.09
|
|
9.58
|
29.13
|
|
9.88
|
31.65
|
|
10.53
|
29.14
|
|
11.08
|
33.11
|
|
11.18
|
34.12
|
|
11.98
|
42.14
|
|
12.59
|
30.00
|
|
13.07
|
39.00
|
|
13.56
|
46.25
|
|
14.06
|
53.00
|
|
14.57
|
40.00
|
|
15.09
|
61.92
|
|
15.62
|
66.93
|
|
16.08
|
71.93
|
|
16.38
|
76.93
|
|
17.10
|
81.94
|
|
17.54
|
86.94
|
|
18.18
|
91.94
|
|
18.53
|
96.95
|
|
19.14
|
101.95
|
|
19.78
|
110.95
|
|
19.89
|
119.95
|
|
20.48
|
129.27
|
|
20.88
|
138.60
|
|
21.78
|
147.92
|
|
22.28
|
157.25
|
|
22.38
|
166.57
|
|
22.48
|
175.89
|
|
23.06
|
185.22
|
|
23.39
|
194.54
|
Chapitre III Etude des paramètres biologiques
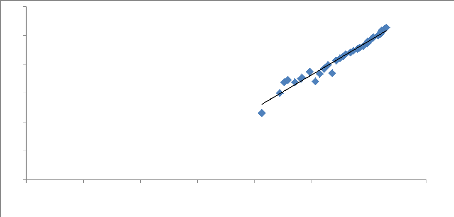
y = 2.344x - 2.2254
R2 = 0.9634
LN(Wg)
1
LN(Lt)
0
0 0.5 1 1.5 2 2.5 3 3.5
6
5
4
3
2
Figure 11. Courbe linéaire de la relation
taille poids.
2.2.2. Résultats :
La valeur du coefficient de corrélation (R) est
très proche de 1, ce qui traduit une forte corrélation positive
entre les deux variables (Wévi, Lt)
Les paramètres a et b et r ont été
calculés, les résultats obtenus sont exprimés dans le
(tableau12et 13).
Ln Wt = b Ln LT + Ln a
Ln a= -- 2.22 a=e -0.22 a=0.10
b=2.34 (b < 3)
L'allométrie est minorante ; le poids croit relativement
moins vite que la longueur á confirmé par le test de
conformité : On compare le tcal avec le ttable (1.24<1 ,96)
t cal >
t table : la différence non significative donc le
Pagellus acarne a une isométrie entre le poids
éviscéré (We), et la longueur á la fourche (LF). et
non pas une allométrie minorate.
Tableau 12 -- calcule Un test de
conformité tcal (á=5%)
|
p-p0
|
S2(Y) : variance
de (y= Ln Wt
|
S2(X) : variance
de (x= Ln Lt
|
S2(Y) /
S2(X)
|
(S2(Y) /
S2(X)
)-
p0
|
N-2
|
sp0
|
tcal (á= 5%)
|
|
1.3020
|
0.66
|
0.54597141
|
0.095732
66
|
0.17534373
|
282
|
0.53
|
1.24
|

We =0.11 * LF 2.34
Les paramètres (a, b) ainsi que le type
d'allométrie de la relation taille-poids de Pagellus acarne
sont données dans le (tableau - 13).
Tableau 13 - Paramètres de la relation
taille poids pour Pagellus acarne de la baie de Bou Ismail.
|
Paramètres
|
A
|
B
|
R
|
t cal
(á=5%)
|
t table
|
type d'allométrie
|
|
Valeurs
|
0.11
|
2.34
|
0 .96433
|
1.24
|
1 ,96
|
Isométrie
|
Le type d'allométrie obtenu par le test tcal=1.24
< à 1.96, différence non significative donc le
Pagellus acarne a une isométrie entre le poids
éviscéré (We), et la longueur à la fourche (LF). et
non pas une allométrie minorate
Relation taille-poids de la Pagellus
acarné
Wt (g)
250.00

|
200.00
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
150.00
|
|
|
|
|
|
|
|
|
|
|
100.00
|
|
|
|
|
|
|
|
|
|
|
50.00
|
|
|
|
|
|
|
Lt ( cm)
|
|
0.00
|
|
|
|
|
|
|
|
|
|
|
|
0.00 5.00 10.00 15.00 20.00 25.00
Figure 15- Relation taille-poids de.
Pagellus acarne de la baie de Bou Ismail.
Tableau 14- Relation taille-poids de
Pagellus acarne selon différents auteurs et différentes
régions.
|
Auteurs
|
Régions
|
Relation taille-poids
|
|
Lamrini
(1983)
|
Sud-Marocain
|
W= 0.02LF2.98
|
|
Lazar, et al.,
(1985)
|
Méditerranée marocaine
|
W= 0.0364LF2.825
|
|
Lamrini
(1988)
|
Agadir Casablanca Larache
Tanger
|
2.96
W= 0.026LF
W= 0.014LF3.22 W= 0.008LF3.3
W= 0.023LF2.97
|
|
Benadda et Kaced
(1994)
|
Baie de Bou-lsmail
|
W=0.055LF3.39
|
|
Dahamni
(2000)
|
Baie de Bou-lsmail
|
W=0.06LF3.37
|
|
Santos et
al
(2000)
|
Sud du Portugal
|
W = 0.0085LT3.153
|
|
(kheloui et al., 2010)
|
Baie de Bou Ismail
|
We = 0.015 * LF 3
|
|
Présente étude
|
Baie de Bou Ismail
|
We =0.11 * LF 2.34
|
2.2.3 Discussion :
La relation taille poids obtenue indique une isométrie
entre la taille et le poids, d'après le test t Le coefficient
d'allométrie b est significativement égal à 3, nous
pouvons dire alors que cette relation est de type isométrique
c'est-à-dire le poids croit en même temps que la longueur.
La comparaison de la relation taille poids des différentes
régions et des différents auteurs (tableau 14) indique que le
coefficient d'allométrie b de Pagellus acarne est
généralement au voisinage de 3 et que la croissance de ce dernier
suit une isométrie.
Cependant, le taux d'allométrie chez une espèce ne
présente pas un caractère universel absolu de ce fait les
donnée de littérature (tableau 13) ne peuvent pas être
comparés afin de conclure à des différences réelles
entre population éloignées. Mouneimme (1981,
in Korichi, 1988)
Notons que des variations de la taille et du poids du poisson
peuvent se manifester au cours d'un changement des facteurs du milieu.
(Kundsen, 1962)
2.3. Croissance pondérale :
2.3.1 Méthode d'étude :
La combinaison de l'équation de croissance linéaire
de VON BERTALANFFY (1938) avec la relation taille-poids permet d'obtenir
l'équation de la croissance pondérale
L'expression de la croissance pondérale s'écrit de
la manière suivante :

W t= W8 (1- e- k (t- t0) ) b avec W8= a
L8b (Ricker, 1975)
Avec respectivement :
Wt : Poids à l'instant
t
W8 : poids asymptotique correspondant à la
longueur asymptotique L8 (W8 est obtenu en
remplaçant le L de la relation taille poids par le L8 obtenu
pour notre échantillonnage la moyenne des deux méthodes
(FISAT II et pauly)
b : Constante déterminée par la relation
taille-poids
k et to : paramètres de l'équation de VON
BERTALANFFY(1938)
W8 : Poids asymptotique correspondant à L8
(Djemmali, 2005)
K, t0 sont les paramètres de croissance linéaire,
tandis que a et b sont les paramètres de la relation taille-poids.
2.3.2. Résultats :
Le poids asymptotique W8 est estimé en remplaçant
le L de la relation taille poids par le L8 = 24.63Cm (la moyenne des
deux méthodes (FISAT II et pauly)).
Le poids asymptotique résultant est de :
W8 = 198.34g
L'équation de la croissance pondérale pour
Pagellus acarne (sexes confondus) s'écrit :
W t= 198.34 (1- e- 0.36 (t ) ) 3.

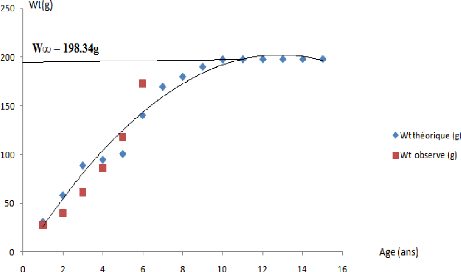

Figure 12- Courbe de croissance pondérale
de Pagellus acarne de la baie de Bou Ismail.
Tableau15- les poids asymptotiques obtenus pour
Pagellus acarne dans différentes régions et pour
différents auteurs (Dahamni, 2000).
|
Auteurs
|
Régions
|
L8 (cm)
|
W8(g)
|
taille extrême d'échantillon
(cm)
|
|
Andaloro
(1982)
|
Sicile
|
26.23 M 29.78 F
|
212.82
314.47
|
8-28 cm
|
|
Lamrini
(1988)
|
Tanger
|
44.07
|
1757.25
|
8-32 cm
|
|
Benadda et
Kaced
(1994)
|
Baie de Bou Ismail
|
22.79
|
225.81
|
6-22.5 cm
|
|
Dahamni (2000)
|
Baie de Bou Ismail
|
35.86
|
971.8
|
8.5-25.5 cm
|
|
(kheloui et al.,
2010)
|
Baie de Bou Ismail
|
27.55
|
313.65
|
6.5-25.5 cm
|
|
Présente étude
(2011)
|
Baie de Bou Ismail
|
24.63
|
|
8.5-24cm
|
2.3.3 Discussion :
On remarque que les points observés se rapprochent de la
courbe théorique, le groupe d'age I n'a pas été
déterminé par manque d'effectifs des petits individus. En ce qui
concerne les autres groupes d'age un bon ajustement des points est
observé, à noter qu'une amélioration des effectifs des
grands et des petits individus serait appréciable pour bien distinguer
les écarts des poids observés et des poids théoriques.
D'après la littérature nos résultats se
rapprochent de ceux obtenus par(kheloui et al., 2010) alors
qu'on retrouve une divergence avec les autres auteurs.
Cette divergence est probablement du à la
fiabilité des échantillonnages utilisés, de la
diversification des zones étudiées, ainsi que de la variation des
longueurs et du poids asymptotiques obtenus.
Conclusion générale
Conclusion générale :
Au terme de ce présent travail, ayant eu pour objectif, la
contribution à l'étude de la croissance de Pagellus
acarne (Risso, 1826) de la baie de Bou-Ismail, plusieurs
remarques ont été retenues.
Les valeurs des tailles moyennes calculées sont de 17.4
#177; 0.27 cm pour les femelles et de 16.3 #177; 0.27 cm pour les mâles.
Il est à remarquer que la taille moyenne des femelles est
supérieure à celle des mâles confirmé par le teste
de l'écart réduit La valeur de |? cal | calculée,
équivalente à 2.54 est supérieure à celle
donnée par l'écart réduit au risque de (5%) (1,96) (t
table). La différence est donc significative, les femelles sont plus
grandes que les mâles dans notre échantillon (2,54>1,96)
Chez les individus de Pagellus acarne, le sex-ratio
global = 0.80, il est supérieure à 1, ce qui indique que les
femelles restent plus abondantes que les mâles, le test de l'écart
réduit est calculé et le résultat indique une
différence non significative (t calculé < t table) (0.97 <
1.96), donc il y a autant de mâles que de femelles.
La méthode de battachaya donne une clé âge
- longueur obtenue par le logiciel FISAT II version 1.2.0, à
été retenue pour la présente étude. Cette
clé âge longueur est caractérisée par 7 groupes
d'ges, avec des longeures moyenne allant de .8cm à 24 cm.
Le nombre de groupe d'ge obtenue par taille moyenne est a prendre
avec réserve vue le nombre faible d'effectifs par taille.
L'étude de la croissance à été
réalisée, les résultats retenus pour les paramètres
de croissance sont ceux obtenus par le logiciel FISAT II version 1.2.0.
La longueur asymptotique (L8) de Pagellus
acarne, fournie par l'équation de la croissance linéaire de
Von Bertalanffy, est de 24.63 cm. Le paramètre de courbure (K), qui
détermine la rapidité du poisson d'approcher la longueur
asymptotique, est de 0.36 /an.
La relation taille poids a permet de conclure à une
isométrie de croissance chez Pagellus acarne avec un poids
asymptotique de 198.34
Enfin, certaines réserves sont à prendre en
considération en raison de la coute période
d'échantillonnage.
Des études plus poussés sur une longue
période permettaient d'obtenir des résultats plus proches de la
réalité.
Références bibliographiques
Références bibliographiques
Andalaro F., 1982. Résumé des
paramètres biologiques sur P.acarne de la mer
tyrrhénienne méridionale et de la mer ionienne septentrionale.
Inst. Zool. Univ. de Messine .Italie : 89-92.
Arculeo M., Bruslé-Sicard A., Potoschi., S Riggio.
2000. Investigations on gonadal maturation in Pagellus acarne
(Piscies, Sparidae) in the Strait of Messina (Sicily). Ital. J. Zool., 67 (4):
333-337.
BA Ibrahima S., 1988. Biologie et dynamique des
populations d'anchois engraulis encrasicolus des côtes
mauritaniennes. Thèse de doctorat en biologie. Université de
Bretagne occidentale. 132 p
Bachouche S ., Nacer L ., 2007.
Aménagement des ports de pêche : Bouharoun et Khemisti.
Mémoire d'ingénieur. I.S.M.A.L.
Belkessam D., Issolah F., 1990. La
pêche chalutière au port de Bou Haroun : étude statistique
des débarquements et aperçu sur quelques paramètres
biologiques et dynamiques d'espèces cibles. Mémoire
d'ingénieur. I.S.M.A.L. 96 p.
Benadda S., Kaced N ., 1994 . Contribution
à l'étude du niveau d'exploitation d'un sparidé
Pagellus acarne (Risso, 1826) dans la baie de Bou-ismail.
Mémoire d'ingénieur en halieutique I.S.M.A.L 67p
Bernt I., Dybern., 1965. The life cycle of Curva
intestinalis (L) f.typica in relation to the environment of temperature
O.I.K.O.S ., 16 (1-2)
Bertalanffy L. V., 1938. A quantitative theory
of organic growth. (Inquiries on growth laws II). Hum. Biol. 10 : 181-213.
Bhattacharya C.G., 1967. A simple method of
resolution of a distribution into gaussian components. Biometrics, 23 (1):
115-135
Brahimi H., 1998. Contribution à
l'étude de la fécondité d'un poisson de la famille des
sparidés : Pagellus acarne (Risso, 1826). Mémoire de
diplôme d'études supérieures U.S.T.H.B Alger 56 p.
Braik D.J., 1989. Etude de la dynamique
sédimentaire devant Bou-Ismail-sédimentologie
Morphologie-Problèmes d'érosion du littoral-Aménagement.
Thèse Magistère en Géologie. U.S.T.H.B, 174 p.
C.N.D.P.A (Centre National du Développement de la
Pêche et de l'Aquaculture)., 2007. Monographie de la wilaya de
Tipaza. 28 p.
Chavance P ., Girardin M., 1986. Niveaux
d'exploitation en 1982 et potentialités régionales de la
pêcherie chalutière algérienne. Application d'un
modèle de production composite. FAO Rapp. Sur les pêches, 347, pp.
113-134.
Chen Y., Harold., Harvey H., 1999. Spatial
structuring of length -at age of the benthivorous white sucker (Catostomus
commersoni) in relation to environmental variables Aquat. Living
.Ressoure, . 12(6):
Coelho R., Bentos C., Correira J.M.S.,
Gonçalves P., Monteiro J., Ribeiro P.G., Lino., K Erzini.,
2005. Age, growth and reproduction of the axillary seabream, Pagellus
acarne (Risso, 1826) from the south coast of Portugal. An International Journal
of Marine Sciences. Thalassa, 2005, 21(1) :79-84.
Dahamni H., 2000. Contribution à
l'étude de l'exploitation de Pagellus acarne de la baie de Bou
Ismail. Mémoire d'ingénieur en Halieutique. I.S.M.A.L. 82 p.
Damir N., 2008. Contribution à
l'étude de l'écologie et de la croissance de Raja asterias
(Delaroche, 1809) Mémoire d'ingénieur d'état
ISMAL
Derdiche o., Stambouli A., Djabali F., Djekrir F., 1990.
Détermination d'une unité d'effort de pêche pour
les flottilles chalutières et sardinières de Beni-Saf. Pelagos
1995.Num spécial.
Djemmali I., 2005. Evaluation de la biomasse
piscicole dans les plans d'eau douce tunisiens : Approches analytique et
acoustique. Thèse de doctorat en sciences agronomique. Institut
National Agronomique Tunisie. 218p
Dubuit M. H., 1975. Etude de la relation
taille-poids chez Raja naevus (Rajidae) coefficient de condition.
J.Cons .Int .Explor. Mer.36(2) :166-169
E.G.P.A.P (Entreprise de Gestion des Ports et des abris
de Pêches et de Plaisance), 2006. Guide des ports de pêche
Algériens. Edition spéciale MINAPPECH, 67p.
Erzini K., Bentes L., Lino P. G., Ribeiro J., Coelho
R., Monteiro P., Correia C., Gonçalves J. M.S., 2001. Age and
growth of seven sparidea species of the south coast of Portugal. Univ of the
Algarve, 8000 Faro. Portugal.
Fischer W., Bauchot M.L., Schneider M., 1987.
Fiches d'identification des espèces pour les besoins de la
pêche. Méditerranée et mer Noire. Ed. F.A.O. Rome,
2 : 761-1530.
Gayanilo F.C. Jr, & Pauly D., 1997.
FAO-ICLARM Stock Assessment Tools: Reference Manual. FAO Computerized
Information Series/Fisheries. No. 8. Rome, 262 p.
Gayanilo F.C. Jr., Sparre P. & Pauly D., 2002.
- The FAO-ICLARM Fish Stock Assessment Tools (FISAT II, user's guide)
2000-2004 FAO, Rome, Italie. Version 1.1.2. 173p.
Gayanilo F.C., Sparre P., Pauly D., 2005.
FAO-ICLARM Outils d'évaluation des stocks II (FISAT II).
Version révisée. Guide d'utilisation. FAO Série
informatique. Péche. No. 8, Version révisée. Rome,
FAO.190p.
Habib A., Koudil M., 1990. La péche au
chalut et à senne au port d'Alger : étude statistique des mises
à quai et détermination du niveau d'exploitation de quelques
espèces cibles. Mémoire d'ingénieur.I.S.M.A.L.
145p
Haddouche I., 2003. Etude de la pollution en
baie de Bou-Ismail par les sels nutritifs et les métaux lourds en
juillet 2002. Mémoire d'ingénieur I.S.M.A.L.
Harchouche K ., 2006. Contribution à la
systématique du genre Spicara ; écologie, biologie
et exploitation de Spicara maena (Poisson,
Téléostéen) des côtes algériennes.
Thèse de doctorat en océanographie. U.S.T.H.B.,
Alger: 210p.
Harchouche K., 1988. Contribution à
l'étude de la biologie et l'écologie d'une espèce de
sparidae dans la baie d'Alger : « Le pageot blanc », P.
acarne (RISSO, 1826). Thèse de magistère en
océanographie. U.S.T.H.B., Alger: 170 p.
Kamar A., 2000. Contribution à la
systématique du genre Spicara et à l'estimation des
paramètres de croissance et d'exploitation de Pagellus acarne
(Risso, 1826) et Spicara maena (Linné, 1756).
Mémoire d'ingénieur d'état en aquaculture U.S.T.H.B.
75 P
.
Khaouni L .,2003. Etude des variations
spatio-temporelle des sels nutritifs et de l'oxygène dissous en baie de
Bou Ismail. Mémoire d'ingénieur. I.S.M.A.L.
Klouch ., Kheloui., 2010 Contribution à
l'exploitation de Pagellus acarne (Risso, 1826) de la baie de Bou-Ismail. .
Mémoire d'ingénieur. E.N.S.S.M.A.L.
Kinacigül H.T., Akyol O., Metün G., Saygi
H., 1999. A Systematic Study on the Otolith Characters of Sparidae
(Pisces) in the Bay of Izmir (Aegean Sea). Ege University,
Fisheries Faculty, 35100, Bornova, Izmir - TURKEY: 357-364.
Knudsen B., 1962. Growth and reproduction of
house Mice at three different temperature Olkos . vol 13.fasc 1 :1-14.
Korichi H.S, 1988. Contributions à
l'étude des deux espèces de saurels Trachurus trachurus
(Linné, 1758), et Trachurus méditerraneus (Steindachner,
1868) et de la dynamique de Trachurus trachurus dans la baie de Bou
Ismail (Algérie). Thèse de magister en halieutique,
I.S.M.A.L. 260 p
L.E.M (Laboratoire d'Etude Maritime), 2005.
Dragage et aménagement du port de Bouharoun. 20 p.
Lahmari L., Selama A., 2008. Impacte de la
variation de la mortalité naturelle (M) par âge sur l'estimation
de la biomasse exploitable de la sardinelle (Sardinella aurite,
Valenciennes(1847)) de la région centre de la côte
Algerienne. Mémoire d'ingenieur . I.S.M.A.L. 260 p.
Lamrini A., 1988. Les sparidés de la
côte atlantique marocaine, Reproduction, croissance et exploitation de
cinq espèces Thèse.doc.en sciences naturelles. Univ .sciences
techniques .Brest. 382 p
Lamrini A., 1983. Les Sparidae du plateau
continental saharien du sud marocain, reproduction, croissance et alimentation
de Pagellus acarne et de Pagellus coupei , Doct. De
3ème cycle, Univ. Bretagne Occidentale, 291p.
Laurec A. & Le Guen J.C., 1981. Dynamique
des populations marines exploitées, Rapports scientifiques et
techniques, Tome I, Concepts et modèles. Rapp. Scient. et tech.,
Éditions CNEXO, n° 45 : 118 p.
Lazar N., Benbouchaib K., Zouiri M., Idelhaj A., 1985.
Resultats de la campagne exploratoire du plateau continental de la
Mediterranee marocaine. FAO Fish. Rep., 347: 85-98 p.
Lecointre G., Le Guyader H., 2002. La
classification phylogénétique du vivant 2e édition. Paris:
Editions Belin.
Léopold M., 2004.- Guide des poissons de
mer de Guyane. Ed. Ifremer, 216 p.
Mennes F., 1985. Multispecies assessment of fish
stocks off the Western Sahara region with emphasis on the family Sparidae.Naga,
3: 5-10. n°96/040, 174p.
Nelson, J.S. 1994. Fishes of the world.
3ème édition. John Wiley and Sons, New York. 600 p.
O.N.M (Office National de la Météorologie),
2003. Centre climatologique national BP 153 DAR EL BEIDA 16100 ALGER.
ALGERIE.
Pajuelo J.G. & Lorenzo J.M., 2000.
Reproduction, age, growth and mortality of axillary seabream, Pagellus acarne
(Sparidae), from the Canarian archipelago. J. Appl. Ichthyol. 16: 41-47.
Parent P., Poly W.J., 2004 . Sparus
axilaris Perez 1820, a senior synonym of the axillary sea bream
Pagellus acarne(Risso 1827) invalidated by 'reversal of precedence'
Cybium 28(2):171-173.
Pauly D., 1980. On the interrelation ships
between natural mortality, growth parameters and mean environmental temperature
in 175 fish stocks. J. Cons. C.I.E.M., 39 (2) : 175-192.
Pauly D., 1983. Some simple methods for the
assessment of tropical fish stocks. Fao Fish. Tech. Pap. 234, Rome, 52p.
Pauly D., 1984. Length converted catch curves: a
powerfull tool for fisheries research in the tropics (part II), fishbyte, 2
(1): 9-17.
Santos M. N., Monteiro C., Erzini K., 2000.
Aspects of the biology and gillnet selectivity of the axillary
seabream (Pagellus acarne) and common Pandora (Pagellus erythrinus)
from the Algarve (South Portugal). Universidade do Algarve, Campus de Gambelas,
8000, Faro, Portugal.
Schwartz D., 1983. - Méthodes
statistiques à l'usage des médecins et des biologistes. Ed.
Flammarion; Coll. Stat. Biol. en Médecine : 199p.
Schwartz D., 1992. - Méthodes
statistiques à l'usage des médecins et des biologistes. Ed.
Flammarion; Coll. Stat. Biol. en Médecine : 306p.
Sparre P., & Venema S. C., 1996.
Introduction à l'évaluation des stocks de poissons
tropicaux. Prem. part. Manuel. FAO Doc. Tech. Pêches, Rome : 306
(1) : 401p.
Stergiou K.I., Christou E.D., Georgopoulous, D.,
Zenetos, A., Souvermezoglou, C. (1997). The Hellenic seas: physics,
chemistry, biology and fisheries. In: Ansell, A.D., Gibson, R.N. & Barnes,
M. Eds., Oceanography and marine biology: an annual review, UCL Press, pp.
415-538.


