Caractérisation biochimique et applications potentielles des glucosidases et de la a-galactosidase du suc digestif de la larve de rhynchophorus palmarum (curculionidae)( Télécharger le fichier original )par Assoi Yapi Désiré Patrice YAPI Université ABOBO-ADJAMà Abidjan - Doctorat 2008 |

II-Glycosidases1-Réaction d'hydrolyse Les glycosidases sont des enzymes qui hydrolysent les liaisons osidiques des glycosides afin de libérer le glucide et son substituant. Elles sont divisées en deux groupes : -les á-glycosidases ; -les â-glycosidases. 1-1-Mécanismes d'action Le mécanisme d'hydrolyse enzymatique des liaisons glycosidiques le plus commun s'opère par une catalyse au cours de laquelle une paire d'acide carboxylique intervient. Dans ce cas, un acide aminé joue le rôle du donneur de proton acide/base et l'autre agit en tant que base/nucléophile. Les glycosidases peuvent alors agir avec deux modes d'actions distincts proposés par Koshland et al. (1953). Il s'agit des mécanismes d'inversion (Fig. 2) et de rétention de configuration anomérique (Fig. 3). Ces deux mécanismes sont retrouvés majoritairement chez les glycosidases. 1-1-1-Inversion de configuration anomériqueLes enzymes qui libèrent un sucre de configuration opposée, agissent avec inversion de configuration anomérique (par simple substitution). Elles utilisent un mécanisme catalytique non-covalent pour exploiter la catalyse acido-basique générale. Une molécule d'eau est activée par un acide carboxylique déprotonné (Asp ou Glu) de l'enzyme afin de générer un hydroxyle qui attaque le carbone anomérique. Selon ce mécanisme de type Sn2, il y a inversion de la configuration anomérique (Fig. 2) (Sinnott, 1990 ; Mccarter et Withers, 1994 ; Davies et Henrissat, 1995). 1-1-2-Rétention de configuration anomériqueLes enzymes agissant avec rétention de configuration anomérique, libèrent un produit dont la stéréochimie du carbone anomérique est la même que celle du substrat (après deux inversions successives). Dans ce cas, le nucléophile effectuant la première attaque sur le carbone anomérique est l'acide carboxylique déprotoné de l'enzyme (Asp ou Glu), formant un intermédiaire covalent. Par conséquent, le substituant du glucoside est relâché à cette première étape. Par la suite, un deuxième résidu acide active une molécule d'eau qui, en attaquant le carbone anomérique, libère le glucide de l'enzyme. Deux étapes de type Sn2 se suivent donc, il y a rétention de la configuration du carbone anomérique (Fig. 3) (Sinnott, 1990 ; Mccarter et Withers, 1994 ; Davies et Henrissat, 1995). 1-1-3-Quelques cas particuliersIl existe néanmoins quelques exemples de glycosidases possédant des mécanismes différents de ceux mentionnés ci-dessus. Il s'agit des cas particuliers où l'acide catalytique peut être remplacé par un phosphate inorganique. Différents groupes chimiques peuvent intervenir dans ce mécanisme et être à l'origine de la similarité de certaines glycosidases avec des enzymes présentant des activités apparemment très distinctes, comme par exemple : la maltose phosphorylase (EC 2.4.1.8, famille GH65), la sucrose phosphorylase (EC 2.4.1.7, famille GH13), ou la cellobiose phosphorylase (EC 2.4.1.20, famille GH94) (Vasella et al., 2002 ; Davies et al., 2003). Certaines glycosidases comme les myrosinases, les chitinases et les endoN-acétylglucosaminidases présentent un mécanisme moléculaire d'hydrolyse n'impliquant l'intervention que d'un seul acide aminé catalytique (Rye et Withers, 2000). D'autres impliquent des mécanismes d'oxydoréduction et/ou d'élimination (Yip et al., 2004).
Figure 2 : Mécanisme d'inversion de configuration anomérique des glycosidases (D'après Davies et Henrissat, 1995)
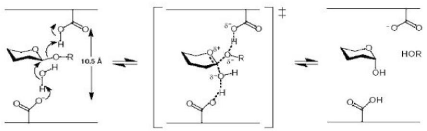
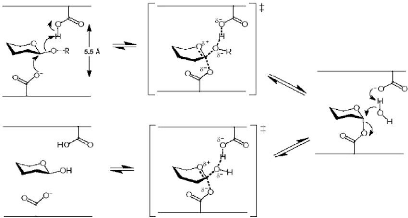
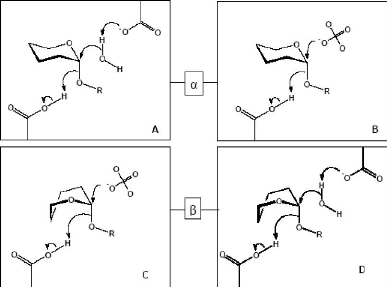
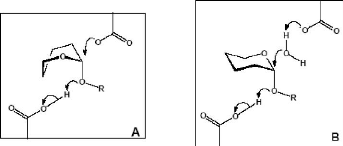
Figure 3 : Mécanisme de rétention de configuration anomérique des glycosidases (D'après Davies et Henrissat, 1995) 1-2-Relation entre les glycosidases de mécanisme d'inversion de configuration anomérique hydrolysant les liaisons alpha et beta Au niveau moléculaire, les mécanismes d'inversion et de rétention de configuration anomérique font intervenir 4 états de transition possibles de type ion oxocarbonium (Vasella et al., 2002 ; Davies et al., 2003). Parmi les â-glycosidases de mécanisme de rétention de configuration anomérique, les conformères 4H3, 2,5B et B2,5 ont été observés respectivement chez les familles GH5 et GH7 (Sulzenbacher et al., 1996 ; Varrot et al., 2003), GH11 (Sabini et al., 1999) et enfin GH26 (Ducros et al., 2002). Pour les â-glycosidases de mécanisme d'inversion de configuration anomérique, c'est la transition en conformère de type 2,5B qui est observée chez les familles GH6, GH8 et GH48 (Guerin et al., 2002 ; Varrot et al., 2003). Chez les glycosidases agissant sur les liaisons â, ces complexes de Michaelis correspondent à des distorsions de la forme cyclique nécessaire pour amener la liaison glycosidique dans une position pseudo axiale (Stam et al., 2005). Cette orientation favorise une attaque nucléophile directe (Sulzenbacher et al., 1996). Il faut noter que de telles distorsions du cycle accompagnant la formation du complexe de Michaelis sont inutiles pour les glucoamylases et leurs homologues de la famille GH15 (Stam et al., 2005). En effet, la liaison glycosidique est déjà dans une position axiale facilitant l'attaque nucléophile (Aleshin et al., 1996). En fait, le mécanisme moléculaire d'inversion des glycosidases agissant sur des liaisons á- passe par une attaque nucléophile réalisée par une molécule d'eau (Stam et al., 2005). Cette molécule d'eau, préalablement activée par un résidu basique, va attaquer le carbone anomérique du glycoside non distordu, avec, dans le même temps, rupture de la liaison glycosidique axiale (Fig. 4) (Rye et Withers, 2000 ; Zechel et Withers, 2000 ; Vasella et al., 2002). 1-3-Relation entre des alpha- et beta-glycosidases avec des mécanismes de configuration anomérique opposés Des similarités, telles que celles vues précédemment, nécessitent une distorsion du substrat dans le site actif d'une â-glycosidase, possédant un mécanisme d'inversion de configuration anomérique. Une telle distorsion place la liaison glucosidique dans une orientation axiale (Fig. 6) similaire à celle d'une liaison á dans le site actif d'une glycosidase avec un mécanisme d'inversion de configuration anomérique (Fig. 5) (Stam et al., 2005).
Figure 4 : Similarités des mécanismes chez diverses familles de glycosidases (A) Mécanisme des GHs hydrolysant des liaisons á chez les familles GH15, GH37, GH63, GH78, GH92 et GH95. (B) Mécanisme des phosphorylases de la famille GH65. (C) Mécanisme des phosphorylases de la famille GH94 hydrolysant des liaisons â. (D) Mécanisme des enzymes des familles GH8, GH9 et GH48 hydrolysant des liaisons â (D'après Stam et al., 2005).
Figure 5 : Similarités dans l'orientation des liaisons dans des enzymes de mécanismes opposés (A) Similarité entre l'étape de glycosylation des enzymes appartenant au clan GH-A et (B) le mécanisme à simple déplacement des â-amylases (D'après Stam et al., 2005). 1-4-Classifications des glycosidases 1-4-1-Classification traditionnelle Dans la nomenclature de l'Union Internationale de Biochimie et de Biologie Moléculaire (IUB-MB) basée sur le type de réaction catalysée et la spécificité de substrat, les glycosidases possèdent un numéro du type EC 3.2.1.x. Les trois premiers chiffres indiquent qu'elles hydrolysent des liaisons O-glycosidiques, le dernier (x) est variable et dépend du substrat transformé (Placier, 1999). Cette classification systématique permet de nommer précisément la spécificité de substrat d'une enzyme et possède en outre, l'avantage de pouvoir classer rapidement une enzyme. Cependant, ce système ne tient pas compte des similarités de séquence protéique et donc de la structure tridimensionnelle. De plus, cette classification n'est pas systématique quant au mécanisme d'action des enzymes. Elle paraît mal adaptée aux glycosidases qui peuvent agir sur différents substrats et ne reflète pas les aspects structuraux des enzymes (Fourage, 2000). C'est notamment le cas de la â-glucosidase BglA, de Pyrococcus furiosus DSM 3638. Elle est annotée avec le numéro EC 3.2.1.21, on pourrait penser qu'elle n'a que cette activité, or elle montre des activités â-galactosidasique (EC 3.2.1.23) et â-xylosidasique (EC 3.2.1.37) (Kengen et al., 1993). Ou encore, il peut se trouver que deux numéros EC différents décrivent la même activité. C'est le cas des numéros EC 3.2.1.10 (oligo-1,6-glucosidase) et EC 3.2.1.70 (glucan 1,6-á-glucosidase). Sur le site de l'Enzyme Commission, la définition de ces deux activités est quasiment similaire. Ce qui est encore plus préoccupant, est le fait que plusieurs activités, qui sont pourtant connues depuis plusieurs années, n'ont aucun numéro EC associé, ou alors un numéro incomplet du type 3.2.1.-, où la spécificité n'est pas indiquée (Green et Karp, 2005). Ce fait révèle un manque de réactivité de l'Enzyme Commission face à la découverte de nouvelles activités. Devant cette situation, une autre classification a été proposée. Il s'agit de la classification structurale. 1-4-2-Classification structurale 1-4-2-1-Principe et classification Les polysaccharides sont la principale forme de stockage de glucides, dont l'oxydation permet de récupérer de l'énergie. Ils entrent aussi dans la composition d'éléments de structures de la plupart des organismes : le peptidoglycane des bactéries, la carapace chitineuse des insectes, la paroi cellulaire des plantes, etc (Biely, 1985 ; Subramaniyan et Prema, 2000). Une hydrolyse sélective des liaisons glucosidiques est donc nécessaire pour récupérer l'énergie via différentes voies oxydatives et libérer des monomères pour construire des parois cellulaires. Les enzymes qui modifient, créent ou lysent les liaisons glucosidiques des polysaccharides sont désignées sous le terme « Carbohydrate-Active Enzymes, ou CAZymes ». La diversité des carbohydrates entraîne donc une grande diversité chez les enzymes (glycosidases ou glycosides hydrolases, GH) dégradant les glucides par hydrolyse. Ces enzymes sont retrouvées chez des organismes provenant de tous les domaines : virus, bactéries, archéobactéries et eucaryotes, et sont impliquées dans de nombreuses voies métaboliques. Ces enzymes ont pendant leur évolution subi des mutations qui leur ont permis de se perfectionner et d'accroître leur spécificité de substrat et ainsi donner un avantage aux organismes qui les portent. De ce fait, les glycosidases couvrent un large panel de fonctions biologiques. Certaines enzymes des GHs présentent même des variations subtiles du mécanisme catalytique conduisant à des activités de type transglycosylase ou lyase, ce qui illustre l'extrême souplesse de leur reconnaissance de substrat et activité enzymatique (Stam, 2006). Les glycosidases sont les plus étudiées. Avec l'accumulation de séquences de ces enzymes, une nouvelle classification était incontournable car la diversité de propriétés et le comportement de certaines enzymes étaient inexplicables par rapport à la seule référence EC. Ces raisons ont amené Bernard Henrissat, à partir de 1989, à chercher une autre alternative pour classer ces enzymes selon des propriétés structurales qui leur sont propres. Il a donc créé une classification, basée sur la similarité de séquences en acides aminés, à partir d'analyses de clusters hydrophobes, ou `hydrophobic cluster analysis', HCA (Gaboriaud et al., 1987) de séquences protéiques de cellulases (Henrissat et al., 1989). Elle est basée sur la détection de segments structuraux constituant le coeur hydrophobe des protéines globulaires (Fig. 6). Grâce à cette approche, des similarités dans le repliement tridimensionnel peuvent être détectées entre des protéines possédant des identités de séquences très basses (< 20 %). Les séquences ayant une forte similarité entre elles ont été classées dans les mêmes familles. A l'origine, seules 6 familles de cellulases ont été créées et les informations étaient enregistrées sous un simple format tabulaire. En 1991, un effort a été fourni pour approfondir ce début de classification (Henrissat, 1991), car plusieurs des familles obtenues par similarité de séquence, présentaient des enzymes ayant des activités sur d'autres substrats que la cellulose (Stam, 2006). Cette classification diffère de la classification de l'IUB-MB par le fait que des enzymes de spécificité différente peuvent appartenir à la même famille. Elle permet de prendre en considération la structure tridimensionnelle ainsi que le mécanisme moléculaire d'action des glycosidases. Elles sont ainsi regroupées en familles (notées GH) en fonction des similarités dans leur séquence d'acides aminés. Certaines glycosidases sont multifonctionnelles. Elles contiennent des domaines catalytiques qui appartiennent à différentes familles des glycosidases. Puisqu'il existe une relation directe entre similarités de séquence en acides aminés et similarités de repliement (Chothia et Lesk, 1986), il peut être admis que les membres d'une même famille possèdent des repliements similaires. Cette situation permet de prédire les structures tridimensionnelles générales des sites actifs des membres de chaque famille, si cette information est connue pour un ou plusieurs de ses représentants (Chothia et Lesk, 1986). Le mécanisme d'action d'une enzyme étant dicté par la structure du site actif et par la position des différents groupes fonctionnels, le mécanisme et la stéréochimie de la réaction seront conservés dans une famille (exception faite de la famille GH4). Toutes les glycosidases agissent généralement par un mécanisme qui implique deux résidus importants. Chez ces enzymes, les acides aminés catalytiques sont le plus souvent des aspartates et/ou des glutamates (Tableaux I et II) (Davies et al., 1995). Cet effort a abouti à la classification de 291 séquences en acides aminés en 35 familles de GHs différentes (Henrissat, 1991), dont plusieurs ont été déjà polyspécifiques. Cette courte liste de familles a été, par la suite, mise à jour plusieurs fois (Henrissat et Bairoch, 1993 ; Henrissat et Bairoch, 1996). A l'heure actuelle, 112 familles de glycosidases différentes sont répertoriées et peuvent être consultées sur internet ( http://afmb.cnrs-rs.fr/CAZY/org.html). Par ailleurs, beaucoup de ces familles, présentant des similarités dans le repliement tridimensionnel des enzymes qu'elles contiennent, ont été regroupées pour former des clans ou super-familles (Henrissat et al., 1995). On dénombre actuellement 14 clans (Clan GH : Clan des GlycosylHydrolases) appelés A, B, C, D, E, F, G, H, I, J, K, L, M et N (Tableau I ). Quatre familles d'enzymes ont été supprimées, ce sont les familles 21, 40, 41 et 60. Certaines familles d'enzymes n'ont pas été rangées en clans ou dans des clans (Tableau II). Des enzymes dont les structures ont été identifiées n'ont pas été à leur tour classées en familles. Réactualisée en permanence (Henrissat et Bairoch, 1993 ; Henrissat et al., 1996), la classification CAZy (Carbohydrate-Active enZymes) est maintenant étendue à d'autres classes d'enzymes actives sur les sucres qui sont les glycosyltransférases (91 familles), les polysaccharides lyases (18 familles), leurs modules non-catalytiques et les carbohydrates estérases (15 familles).
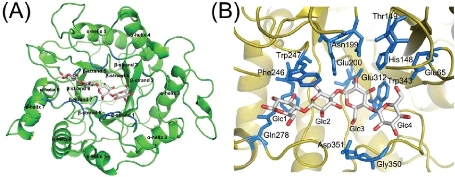
39 Figure 6 : Structure tridimensionnelle de l'endocellulase (famille 5 des glycosidases) de Thermus caldophilus (A) et de son site actif (B) liant le cellotétraose (D'après Kim et al., 2006) Tableau I : Mécanismes réactionnels et acides aminés impliqués dans la catalyse enzymatique des familles des glycosidases rangées en clans a = le premier groupement cité est le composé nucléophile/base, le second est celui du donneur de proton
41 Tableau II : Mécanismes réactionnels et acides aminés impliqués dans la catalyse enzymatique des familles des glycosidases non rangées en clans Familles de glycosidase Mécanisme Acides aminés impliqués dans la réactionnel catalyse enzymatique Nucléophile/Base Donneur de proton 3 rétention Asp Glu 4 rétention non applicable non connu 6, 45, 74 inversion Asp Asp 9, 25, 75 inversion Asp Glu 19, 64, 71, 78, 81 inversion non connu non connu 14 inversion Glu Glu 109 rétention non applicable non connu 29 rétention Asp non connu 44 rétention Glu Glu 112 inversion non connu Asp 47, 67 inversion non connu Glu
57 rétention Glu non connu 66 rétention Asp Asp 84 rétention oxygène du carbonyl Asp 90 inversion Glu et Asp Asp 91 inversion Glu non connu 94 inversion phosphate Asp 95 inversion Asn activé par Asp Glu 108 non connu non connu Glu 42 1-4-2-2-Division des familles des glycosides hydrolases en sous familles Les familles de protéines définies dans CAZy permettent d'avoir un certain pouvoir prédictif : le même repliement, le même mécanisme d'action, et les mêmes résidus catalytiques sont conservés à l'intérieur d'une même famille. Or depuis quelques années, l'équipe CAZy participe à des projets de séquençages de génomes, intervenant au niveau de l'identification et de l'annotation fonctionnelle des gènes de CAZymes (Stam, 2006). Les centres de génomique sont conscients de l'accumulation d'erreurs et des limitations de la méthode « best BLAST hit ». Ils se tournent donc vers des équipes spécialisées capables d'apporter la meilleure annotation possible. Etant accessible en ligne depuis 8 ans, CAZy est devenue une des principales références en ce qui concerne les CAZymes. Bien que l'assignement d'une enzyme à une famille particulière permet de prédire plusieurs caractéristiques structurales et mécanistiques, il n'était pas toujours possible de fournir une prédiction précise sur la fonction de cette enzyme (Stam, 2006). Pour répondre à ce problème et améliorer les procédures d'annotation de séquences génomiques, Stam et al. (2006) ont fourni un effort de division des familles de GHs en sousfamilles suivant l'idée communément acceptée que des enzymes possédant des séquences très similaires doivent partager des propriétés biochimiques très proches (Fig. 7). La famille GH 13, qui regroupe beaucoup de glycosidases a été subdivisée en 40 sous-familles (Tableau III) (Stam et al., 2006). Des á-amylases (EC 3.2.1.1) appartenant à cette même famille ont été rangées dans 13 sous familles qui sont GH13_1, GH13_2, GH13_5, GH13_6, GH13_7, GH13_15, GH13_19, GH13_24, GH13_27, GH13_28, GH13_32, GH13_36 et GH13_39. Quant aux á-glucosidases (EC 3.2.1.20), elles se retrouvent dans des sous familles différentes de celles des á-amylases. Il s'agit des sous-familles GH13_17, GH13_21, GH13_30 et GH13_40 (Tableau III). Tableau IIIa : Division de la famille GH13 en sous familles
D'après Stam, 2006. 44 Tableau IIIb : Division de la famille GH13 en sous familles (suite et fin)
D'après Stam, 2006.
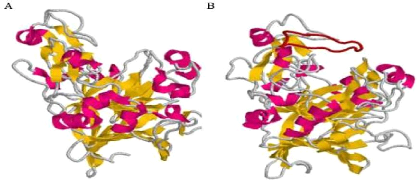
Figure 7 : Comparaison entre la structure d'une endoglucanase et celle d'une cellobiohydrolase de la famille GH7. Structure d'une cellobiohydrolase (A), et d'une endoglucanase (B) de la famille GH7, représentative des sous-familles GH7_2 et GH7_1, respectivement (D'après Stam, 2006). Les hélices á sont en violet, les feuillets â en jaune. La boucle supplémentaire présente chez les cellobiohydrolases est en rouge. Figure réalisée avec Rasmol (Sayle et Milner-White, 1995) à partir des structures PDB 1DYM et 7CEL 5 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||