III.2 : Variations des densités
anophèliennes
III.2.1 : Variations de la densité agressive et
du taux d'endophagie
Le tableau 2 rapporte les résultats des densités
agressives des espèces en fonction du statut des villages.
L'agressivité des femelles d'An. gambiae s.l. et d'An.
funestus a connu des variations entre les zones traitées et celle
non traitée.
Avec respectivement 2,04 Piqûre par Homme et par Nuit
(PHN) et 1 PHN, An. gambiae s.l. a été plus agressive
aussi bien à l'intérieur des habitations qu'à
l'extérieur dans le village non traité comparé à
ceux traités où l'agressivité a été
estimée à 0,87 et 0,70 PHN, respectivement à
l'intérieur et à l'extérieur.
Pour An. funestus, la même tendance a
été observée avec des densités agressives plus
faibles dans les villages, comparé à An. gambiae s.l.
Toutefois, il est important de noter que dans tous les villages suivis, An.
funestus a été plus exophage comparé à An.
gambiae s.l.
Tableau 2 : Agressivité des populations d'An.
funestus et d'An. gambiae s.l. à l'intérieur et
à l'extérieur dans les zones traitées et témoins
Capture sur homme
|
An. funestus
|
An. gambiae s.l.
|
|
Statut des zones
|
Extérieur
|
Intérieur
|
Extérieur
|
Intérieur
|
|
Zones traitées Zone non traitée
|
0,93
1,04
|
0,43
0,79
|
0,70
1,00
|
0,87
2,04
|
Les taux d'endophagie (figure 12) ont été
significativement plus élevés (p = 0,03) pour An. gambiae
s.l. dans le village témoin (41,53%) que dans ceux traités
(26,26%). Par contre, pour An. funestus aucune différence
significative n'a été observée (p = 0,67).
22
100,00
Pourcentage
75,00
50,00
25,00
T NT
0,00
An. funestus An. gambiae s.l
Statut
Figure 12 : Taux d'endophagie (en %) en fonction du statut des
villages
III.2.2 : Variations de la densité au repos
Le tableau 3 présente les densités au repos
à l'intérieur des habitations selon le statut de la zone. Les
résultats montrent que les populations d'An. gambiae s.l. ont
été 9 fois plus importantes dans la zone témoin
comparé à la zone AID. La même tendance a été
observée pour An. funestus dont les densités au repos
ont toutefois été moindre comparé à An. gambiae
s.l.
Tableau 3 : Variations de la densité au repos selon le
statut de la zone
Faune matinale résiduelle
Statut de la zone An. funestus An. gambiae s.l.
Traitées 0,60 1,04
Non traitée 2,15 9,23
III.3 : Préférences
trophiques
23
L'origine des repas sang a été
déterminée sur 62 femelles gorgées (19 An. funestus
et 43 An. gambiae s.l.), collectées au repos dans les
habitations au niveau des 3 villages. La typologie des repas montre que, de
manière globale, 38 étaient des repas mono spécifique, 23
des repas
mixtes pris sur deux hôtes différents et 1 seul
repas triple pris sur homme, boeuf et cheval. Ainsi, 87 repas ont
été identifiés.
Quelle que soit la zone d'étude
considérée, les populations d'An. gambiae s.l., et
d'An. funestus ont montré un profil plus zoophile
qu'anthropophile (figure 13). En effet, dans la zone témoin An.
gambiae s.l. s'est principalement gorgé sur cheval (65,96%), tandis
que dans celles traitées, des proportions égales de repas pris
sur cheval et boeuf (41,67%) ont été notées. Dans ces
dernières, le pourcentage de repas pris sur homme a été
faible (16,67%). Le scénario contraire est noté dans la zone
témoin où le second hôte préférentiel
était l'Homme (25,53%). Les repas sur bovins n'ont compté que
pour environ 8% du total identifié (figure 13).
En ce qui concerne An. funestus, des proportions
comparables sur l'anthropophilie ont été notées aussi bien
dans les zones traitées que dans celle témoin. Cependant, il est
important de noter que le spectre d'hôtes est élargi aux ovins
(figure 13). Dans les villages traités, les femelles de cette
espèce se sont gorgées dans les mêmes proportions (~33%)
sur homme, boeuf et cheval. Le taux d'anthrophilie a été de
26,43% pour l'ensemble des 3 villages.
De manière spécifique, les villages traités
ont présenté un taux d'anthropophilie faible de 22,2%,
comparé à celui du village non traité (27,5%). Cette
différence n'est cependant pas significative (p-value=0,87).
24
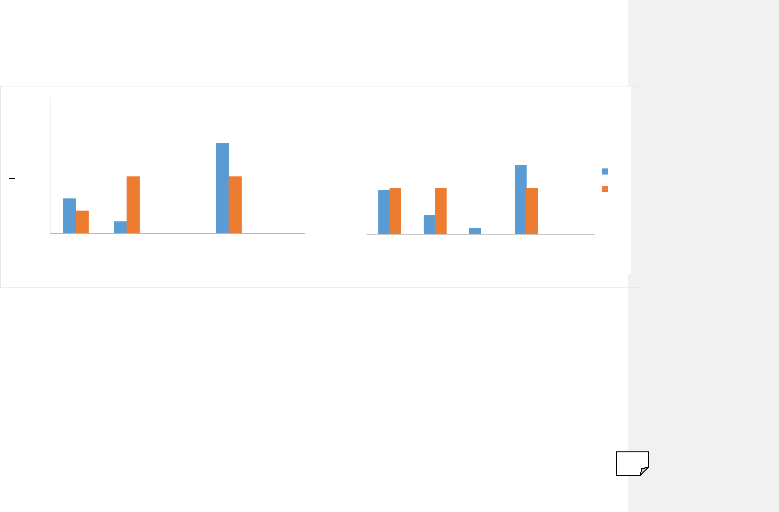
Figure 13 : Spectres d'hôtes des populations d'An.
gambiae s.l. et d'An. funestus en fonction du statut de
traitement des zones d'étude
25
An. funestus
homme boeuf mouton cheval poule
Hôtes
NT T
homme boeuf mouton cheval poule
Hôtes
100
An. gambiae s.l.
Proportion (%)
75
50
25
0
III.4 : Taux de parturité
Au total 113 femelles à jeun capturées sur
appât humain ont été disséquées pour la
détermination de l'âge physiologique. Les résultats sont
présentés dans le tableau 4. L'analyse montre que la proportion
de femelles pares pour An. gambiae s.l. et An. funestus varie
entre zones traitées (55% et 66,67% respectivement) et zone
témoin (52,27% et 35,71% respectivement). Cependant, les
différences observées entre les zones n'ont pas été
significatives. Il en est de même pour la parturité entre les
populations d'An. gambiae s.l. et d'An. funestus.
Tableau 4 : Taux de parturité selon les zones
|
An. gambiae s.l.
|
An. funestus
|
|
|
Statut
|
Total disséqué
|
Parturité (n)
|
Total disséqué
|
Parturité (n)
|
P
|
|
Zones traitées
|
20
|
55,00% (11)
|
21
|
66,67% (14)
|
0,65
|
|
Zone non traitée
|
44
|
52,27% (23)
|
28
|
35,71% (10)
|
0,25
|
p 0,94 0,06
(n) = effectif testé ; p = Valeur de p
III.5 : Indice Circumsporozoïtique :
ICS
Au total, 120 femelles dont 99 dans les deux villages
traités et 21 dans celui non traité (témoin) ont
été testées pour la recherche d'infection à
Plasmodium falciparum par la méthode d'ELISA CSP. Aucune
infection à Plasmodium falciparum n'a été
détectée.
III.6 : Sensibilité des populations d'An.
gambiae s.l. aux insecticides
Parallèlement à l'étude de dynamique des
populations de vecteurs, celle de sensibilité des populations d'An.
gambiae s.l. vis-à-vis des Carbamates (Bendiocarb) et des
Organophosphorés (Pyrimiphos-méthyl, Malathion et
Fénitrothion) a été également évaluée
dans le district de Nioro.
Au total, 443 spécimens ont été
exposés aux 4 molécules d'insecticides appartenant à deux
familles susmentionnées. Les résultats, enregistrés dans
le tableau 5, montrent que les populations d'An. gambiae s.l. ont
été totalement sensibles aux Organophosphorés et aux
Carbamates.
26
L'identification spécifique d'un
sous-échantillon de la population testée, par PCR, a
révélé la présence d'An. arabiensis
(67,6%) et d'An. gambiae (32,84%).
Tableau 5 : Sensibilité d'An. gambiae s.l. aux
Carbamates et aux Organophosphorés dans le district de Nioro
Carbamates Organophosphorés
Bendiocarb Pyrimiphos-méthyl Malathion
Fénitrothion
|
100***
|
100***
|
100***
|
100***
|
|
(114)
|
(109)
|
(116)
|
(104)
|
() = effectifs testés ; *** = Susceptible
27

III.7 : Discussion
Au cours de cette étude, 5 des 20 espèces
déjà décrites au Sénégal (Diagne et
al., 1994) ont été collectées. Il s'agit d'An.
gambiae s.l., An. funestus, An. pharoensis, An.
ziemanni et An. squamosus. Parmi celles-ci, An. gambiae
s.l., et An. funestus, ont été les plus abondantes
quelle que soit la méthode de collecte ou la zone
considérée.
Pour An. gambiae s.l, sa prédominance pourrait
s'expliquer par la présence de ses gîtes
préférentiels, constitués par des collections d'eau
temporaires peu profondes et ensoleillées liées aux pluies
(Gillies et De Meillon, 1968). Quant à An. funestus,
l'abondance notée dans les différents villages suivis,
principalement à Ndramé ndimbe, pourrait être liée
à la présence de cours d'eau permanents à
végétation dressée, favorable au développement de
ses larves. Cette même observation a été rapportée
par Gueye (1969) au cours de ses travaux dans le bassin du Saloum.
Le comportement exophage, moins endophile et zoophile
observé au cours de cette étude chez les populations d'An.
funestus dans les villages traités, pourrait trouver son
explication par l'effet des aspersions intra-domiciliaires, combiné
à l'utilisation des moustiquaires imprégnées
d'insecticides, rendant difficile l'accès à l'homme. En effet,
l'utilisation des insecticides à effet rémanent conduit à
suspecter une hétérogénéité au sein des
populations d'An. funestus. D'autant plus que des études
réalisées au Burkina Faso mettaient en évidence
l'existence de deux unités taxonomiques à capacité
vectorielle et à indice d'anthropophilie différents (Boccolini et
al., 1994); alors qu'à Madagascar des études simi1aires
soulignaient des variations liées à l'anthropophilie (Boccolini
et al., 1992). Au Mali et au Burkina Faso des observations comparables
faisaient état d'un important polymorphisme (Boccolini et al.,
1998). D'autre part, la cohabitation des populations humaines avec les animaux
domestiques pourrait offrir une large gamme d'hôtes alternatifs,
d'où la déviation trophique notée à travers les
repas pris fréquemment sur cheval, boeuf et dans une moindre mesure sur
mouton.
Commentaire [EN1]:
A l'image d'An. funestus, le même comportement
a été également observée chez les populations
d'An. gambiae s.l. Ceci pourrait laisser penser que les espèces
du complexe sont en majorité représentées par An.
arabiensis, dont le comportement exophage, moins endophile et zoophile est
bien connu (Gillies et De Meillon, 1968 ; Niang et al., 2016). Ceci
lui permet de se soustraire aux interventions de LAV et a pour
conséquence un changement de la composition spécifique de la
population d'An. gambiae s.l. comme suggéré dans
d'autres contrées (Derua et al., 2012 ; Kitau et al.,
2012).

Au cours de cette étude, aucune infection à
Plasmodium falciparum n'a été détectée sur
les spécimens testés dans les différents sites. Ceci
pourrait être mis en relation avec les comportements de piqûre et
de repos. En effet, le faible contact homme-vecteur ne permettrait pas aux
espèces de s'infecter.
Commentaire [EN2]:
Nos travaux ont rapporté des taux de parturité
faibles à modérés et similaires entre les deux
espèces vectrices retrouvées dans les deux zones. Pour les deux
taxa, ces taux pourraient être expliqués par l'AID, dont
l'objectif premier est de réduire la longévité des
populations vectrices (Najera et Zaim, 2002).
L'étude a également révélé
une sensibilité totale des populations d'An. gambiae s.l aux
Carbamates et aux Organophosphorés, à l'instar d'autres zones du
pays (Faye et al., 2011). Toutefois, une étude récente
dans le Sénégal Oriental a confirmé la résistance
des populations d'An. gambiae s.l. au Fénitrothion (Niang
et al., 2016).
| 

