3. 1. 3. Morphologie des nodules
Dans la famille des légumineuses, la morphologie des
nodules et le type de nodosité développée est
déterminé par la plante hôte (Dart, 1975; Newcomb et
al., 1979; Newcomb et Tandom, 1981). Deux types majeurs de
nodosités (Figure 3) sont souvent distingués en se basant sur
l'existence ou non du méristème persistant:
(i) Nodosités à forme
indéterminée où l'activité
méristématique se maintient. De nouvelles cellules apicales sont
continuellement infectées. Cela résulte en une forme cylindrique
de la nodosité. Ces nodosités sont connues chez les
légumineuses des zones tempérées (sulla, pois, Vicia
sp., Medicago sativa L., etc...).
Revue bibliographique
(ii) Nodosités à croissance
déterminée où l'activité
méristématique cesse tôt. Les cellules infectées
engendrent d'autres cellules infectées et la nodosité en
grandissant par expansion acquiert une forme sphérique. Ce type de
nodosité existe seulement chez les légumineuses des zones
tropicales telles que le soja et le haricot (Hirsch et al., 2001).
Les deux types de nodosités partagent la même
organisation générale se basant sur l'existence d'un tissu
central entouré par plusieurs tissus périphériques. Le
tissu central contient à la fois les cellules infectées par les
rhizobiums et d'autres non. Les tissus périphériques sont
formés essentiellement par un cortex interne et un autre externe
séparés par l'endoderme nodulaire (Van de Wiel et al.,
1990).
Un troisième type intermédiaire a
été identifié chez les espèces du genre
Lupinus et Sesbania (Sesbania rostrata Brem.). Les
divisions cellulaires se font dans le cortex externe ou interne, conduisant
à la formation de nodosités déterminées ou
indéterminées (Hirsch et al., 2001).
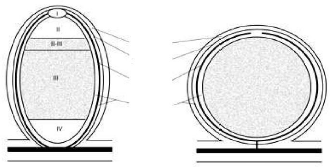
Cortex
Endoderme
Système
vasculaire
Parenchyme
A B
Figure 3: Structure des nodules de
légumineuses. A: nodule de type indéterminé. B: nodule de
type déterminé. I: zone méristématique; II: zone
d'infection; II-III: interzone II-III; IV: zone de sénescence.
(Pawlowski et Bisseling, 1996).
3. 1. 4. Génétique de la nodulation
Lors de l'étape de la reconnaissance entre les
rhizobiums et les plantes hôtes, les flavonoïdes induisent
l'activation des gènes nod et la synthèse des
lipo-chitooligosaccharides (LCOs) (Dénarié et al., 1996)
émis par la bactérie qui sont à leur tour à
l'origine de l'activation d'autres gènes végétaux ou
bactériens (Bladrgroen et Spainkc, 1998).
Revue bibliographique
Ces LCOs ont été nommés facteurs Nod et
sont responsables de changements physique et physiologique chez la plante
hôte, comme la déformation des racines, la division cellulaire et
l'organisation du primordium nodulaire. Les gènes nod sont
déterminants dans la spécificité de l'hôte
(Denarié 1992; Geurts et al., 2005; Mulder et al.,
2005; Chen et al., 2006). En effet, la longueur et la saturation
de la chaîne de l'acide gras ainsi que les différents substituants
greffés sur le squelette du LCO jouent un rôle crucial dans la
spécificité entre la bactérie et la plante (Perret et
al., 2000).
La plupart des gènes symbiotiques sont localisés
sur des méga plasmides. Les grands plasmides ne soient pas
transmissibles, mais ils sont en rapport avec d'autres plasmides très
transmissibles (Hirsch et al., 2001).
Les gènes de rhizobium essentiels pour l'infection et
la nodulation incluent les gènes nod (nod,
nol, noe), les gènes hsn (gènes de
nodulation spécifique de l'hôte) et d'autres gènes qui
codent pour l'expression et la synthèse des molécules de
structure de la surface bactérienne. Parmi ces gènes on note les
gènes ndv responsables de la synthèse des
â-glucans, les gènes exo responsables de
la synthèse des exopolysaccharides, et les gènes lps
responsables de la synthèse des lipopolysaccharides (LPS)
(Broughton et al., 2000; Spaink, 2000, 1999).
Les gènes de nodulation ne sont exprimés que par
l'action d'un activateur de transcription, le nod D. Ce dernier est
exprimé de manière constitutive (Geurts et Bisseling, 2002), il
est activé par des molécules produites par les plantes telles que
les flavonoïdes (Spaink, 1987; Zuanazzi et al., 1998).
Les gènes nod ABCIJ sont des gènes
communs très conservés chez les différentes espèces
de Rhizobium. En ce qui concerne les composants de la surface
cellulaire codés par les gènes ndv, exo et lps
se trouvent les â-glucans, exopolysaccharides et
lipopolysaccharides qui sont considérés comme d'importants
facteurs dans l'efficacité symbiotique (Breedveld et Miller, 1998). Les
â-glucans sont majoritairement des molécules du
périplasme qui permettent la croissance des bactéries sous des
conditions hypo-osmotiques (Pfeffer et al., 1994). Ils ont
également un rôle spécifique dans l'interaction symbiotique
(Breedveld et Miller, 1998). D'après Bhagwat et al. (1996), ces
molécules jouent un rôle important dans la suppression du
déclenchement du mécanisme de défense par les
phytoalexines chez l'hôte.
Revue bibliographique
| 

