RESUME
La destruction par effet knock-down des bruches dans le
niébé en stock (action de choc) n'est qu'un aspect de
l'efficacité des formulations agissant comme insecticide de contact.
Cette étude avait pour objectif de vérifier la durée de la
rémanence de l'activité insecticide des huiles essentielles de
Xylopia aethiopica utilisées pures ou mélangées
avec de la poudre de fruits de la même espèce. Une
rémanence plus longue réduit en effet les risques de
réinfestation en provenance de l'extérieur ou par une nouvelle
génération après l'utilisation du produit contre les
parents. Les comparaisons ont été effectuées avec un
témoin et la survie des bruches adultes, leur fécondité et
le nombre de descendants de la première génération ont
été mesurés ainsi que la perte de poids des grains.
Les traitements testés sont le témoin
T01 (niébé seul), l'huile essentielle
diluée dans l'acétone (T02), la poudre de
Xylopia aethiopica (T03) et la poudre
mélangée à l'huile essentielle de Xylopia
aethiopica (THE). La persistance de
l'efficacité de la formulation dans le traitement THE
est meilleure vis-à-vis de la bruche du niébé pendant deux
semaines au moins. Une meilleure réduction des pontes a
été aussi observée dans le traitement
THE. La formulation huile essentielle diluée dans la
poudre de Xylopia aethiopica (THE) provoque encore 55
jours après le traitement des graines une réduction de
ponte d'environ 90%. Ce traitement a un effet résiduel plus
prolongé que pour les autres traitements. L'effet rémanent du
T03 sur la mortalité et la réduction des pontes
est plus long que le traitement T02. Le traitement
T02 provoque une mortalité semblable à celle
observée dans le traitement THE durant les six premiers
jours. Il s'agit d'un effet knock-down qui s'estompe suite à
l'évaporation de la formulation T02 (huile essentielle
diluée dans l'acétone). La rémanence de cette formulation
est donc limitée et les risques de réinfestation existent suite
à la disparition de l'effet répulsif observé les premiers
jours d'incubation.
Nos résultats indiquent qu'une protection durable et
à coût peu élevé contre Callosobruchus
maculatus est possible en utilisant les formulations à base de
fruits secs de Xylopia aethiopica. Compte tenu du fait que Xylopia
aethiopica se retrouve dans les aires de production du
niébé, l'utilisation d'un mélange de la poudre de fruits
secs et l'huile essentielle de cette espèce pour protéger les
récoltes de niébé contre les attaques
sévères de Callosobruchus maculatus peut
représenter une solution alternative supplémentaire pour les
populations locales qui traitent habituellement le niébé avec les
plantes à effet insecticide.
CHAPITRE I : INTRODUCTION
1- Introduction
Pour faire face à la demande alimentaire des
populations vivant dans les pays en développement où la
consommation de viande constitue encore un luxe, les légumineuses
à graines sont une source indispensable de protéines. Parmi ces
légumineuses, le niébé : Vigna unguiculata
L. (Walp), constitue l'un des aliments de base des populations. De par son
rôle dans la restauration de la fertilité du sol, et sa
compatibilité avec plusieurs associations culturales, cette plante est
une composante essentielle des systèmes de culture dans les zones de
savane en Afrique tropicale (Mulongoy et al., 1992). Les
graines sont riches en protéines, en carbohydrates et en vitamines
(Kholi, 1990 ; Quin, 1997). Elles contiennent plus de 25% de
protéines et sont riches en lysine et en tryptophane. Le
niébé vient ainsi équilibrer les régimes à
base de céréales et de tubercules qui sont
généralement pauvres en protéines et riches en hydrates de
carbone (Singh et Singh, 1992). Les feuilles du niébé sont
consommées comme des légumes en Afrique tandis que les
résidus constituent une bonne source de fourrage animal (Tarawali et
al., 1997).
Nonobstant son importance, les rendements du
niébé sont inférieurs dans certaines régions
à la moyenne de l'Afrique qui est de 700kg/ha (FAO, 1998). Parmi les
contraintes à la production du niébé, les insectes
ravageurs causent le plus grand problème au niébé
(Bottenberg, 1995). En effet, le niébé est attaqué par un
large spectre de ravageurs depuis la germination jusqu'à la
récolte (Jackai et Adalla, 1997). Dans la gamme très large de ces
insectes ravageurs du niébé, les Coléoptères
Bruchidae dont notamment Callosobruchus maculatus Fabricius sont parmi
les plus redoutables du fait que leurs attaques débutent au champ, pour
s'étendre ensuite à l'entrepôt où la population de
bruches peut croître rapidement. La conséquence la plus
préoccupante de ces attaques est la diminution qualitative et
quantitative de la récolte au champ ou en cours de stockage.
Plusieurs méthodes sont préconisées pour
le contrôle des ravageurs du niébé. Celles-ci incluent la
lutte biologique, la lutte chimique, la résistance de la plante
hôte et les pratiques culturales. Parmi ces méthodes, la lutte
chimique par les pesticides de synthèse est la stratégie la plus
utilisée et la plus efficace pour le contrôle des ravageurs du
niébé, puisqu'elle a montré des résultats
satisfaisants dans l'accroissement des rendements du niébé dans
maintes régions (Alghali, 1993 ; Amotabi, 1994 ; Jackai,
1995a). Malgré ces réussites, l'utilisation des insecticides de
synthèse n'est pas une stratégie recommandable pour plusieurs
raisons : leur toxicité, la perturbation de l'équilibre
biologique de l'écosystème, le coût élevé des
insecticides, la faible maîtrise des techniques d'utilisation et le
développement des souches résistantes.
Face donc à la demande croissante de promotion d'un
développement durable, et de protection de l'environnement en
l'application de l'Agenda 21 du sommet Planète Terre sur l'environnement
et le développement tenu à Rio de Jane iro en 1992 des
méthodes alternatives sont préconisées. Ainsi, suite aux
désillusions liées à l'emploi des insecticides de
synthèse, il est urgent d'identifier, d'inventorier et de recommander
des méthodes de contrôle, qui soient peu coûteuses et
efficaces, faciles à adopter pour les producteurs du Tiers Monde.
L'accent est donc de plus en plus mis sur l'exploitation des connaissances
locales à travers un recensement des composantes de la
biodiversité (Kossou et al., 2001).
Des études réalisées dans plusieurs
agrosystèmes en Afrique et particulièrement au Nord Cameroun
révèlent que les producteurs utilisent des pratiques
traditionnelles dont des extraits des plantes à effet insecticide et/ou
insectifuge pour la conservation des produits agricoles en particulier le
maïs et le niébé (Ngamo, 2004). De par leur innocuité
et leur faible toxicité pour l'homme (Isman, 2002), ces plantes à
utilités alimentaire et médicinale constituent une alternative
pour la conservation des denrées stockées.
Les huiles essentielles extraites de ces plantes aromatiques
à propriété insecticide et/ou insectifuge ont fait l'objet
d'importants travaux, notamment ceux de Delobel et Malonga (1987), Kétoh
(1998). L'évaluation de la toxicité insecticide par contact de
l'huile essentielle issue de Xylopia aethiopica Dunal, Ocimum
gratissimum L., sur Sitophilis zeamais a montré
que les huiles essentielles de O. gratissimum et X.
aethiopica sont toxiques vis-à-vis du charançon du maïs
(Kouninki, 2005). Cette même étude indique que les formulations
à 0,5% entraînent 100% de mortalité après exposition
des individus de C. maculatus pendant 24h aux grains de
niébé imprégnés de l'huile essentielle de X.
aethiopica issue des téguments. Ces résultats
révèlent donc l'effet Knock down de l'huile essentielle mais
aucune donnée sur l'efficacité à long terme et la
rémanence des produits utilisés n'est disponible.
Le présent travail a pour but d'évaluer l'effet
insecticide des formulations à base d'huile essentielle et des fruits
secs de X. aethiopica sur C. maculatus. L'étude se
propose de tester la rémanence de l'efficacité insecticide des
formulations de fruits secs de X. aethiopica sur la survie de C.
maculatus.
L'intérêt de cette étude réside
dans la recherche de formulations pouvant augmenter la rémanence des
huiles essentielles et être utilisées localement comme
biopesticide pour la protection à long terme du niébé.
CHAPITRE II : REVUE BIBLIOGRAPHIQUE
2- Revue de
littérature
2-1-
Généralités sur le niébé et Xylopia
aethiopica Dunal
2-1-1 Le niébé
(Vigna unguiculata (L.) Walp)
Le niébé est une légumineuse de la
famille des Fabaceae. Cette dicotylédone est une plante principalement
autogame, bien qu'on ait fait état d'un certain degré d'allogamie
(Rachie et Robert, 1974), favorisé par les abeilles qui assurent la
pollinisation (Zannou, 1995).
Le niébé semble trouver son point de
départ en Afrique Occidentale, et très vraisemblablement au
Nigeria où les espèces sauvages et adventices pullulent dans les
savanes et les forêts (IITA, 1982).
Sur 80 millions d'ha de superficie mondiale sous culture de
niébé, 60 millions sont emblavés en Afrique (Montimore
et al., 1997). Plus de la moitié de la production
mondiale (Pandey et Wespal, 1989) compte pour l'Afrique et est estimée
à 3,36 millions de tonnes (Duke, 1990).
Le niébé est profondément
intégré aux pratiques culturales dans les régions
tropicale et subtropicale du globe où il est généralement
associé à d'autres cultures comme le maïs, le sorgho, le mil
ou le manioc (Jackai et Daoust, 1986). Il joue un rôle capital dans les
systèmes culturaux en restaurant la fertilité des sols par la
fixation de l'azote atmosphérique. Si, à l'instar d'autres
légumineuses, le niébé a de grandes exigences en phosphore
et potassium, aucune fertilisation azotée n'est nécessaire
surtout pour les variétés qui nodulent facilement à partir
des rhizobia contenus dans le sol. Par ce processus, le niébé
cède 60 à 70 kg à l'hectare de l'azote fixé pour la
culture suivante (Rachie, 1985).
2-1-2 Xylopia aethiopica
Dunal (Annonaceae)
Plusieurs plantes ont été reconnues pour leurs
vertus insecticides et de ce fait sont utilisées depuis des
décennies dans les structures traditionnelles de stockage par les
producteurs. X. aethiopica est un arbre très répandu
en Afrique intertropicale. Il est aussi retrouvé dans les zones
forestières humides au Sénégal, au Congo Brazzaville et
dans l'île de Sao Tomé et Principe (Adjanohoun et
al., 1988).
Communément appelée le poivrier du
Guinée, cette espèce héliophile peut atteindre 15m de
hauteur, avec un diamètre de 10cm environ. Les feuilles
lancéolées sont arrondies à la base. Les fleurs sont
blanc-verdâtre et parfumées. Les fruits apocarpes sont
déhiscents à maturité. Les grains sont noirs et
entourés par un aryle jaune (Dupriez et De Leener, 1987 ;
Adjanohoun et al., 1988).
Dans certains pays, les fruits secs de X. aethiopica
(Figure 1) sont utilisés en pharmacopée traditionnelle contre
plusieurs affections dont notamment le traitement de la bronchite, de la
dysenterie, de la stérilité chez la femme et des
aménorrhées (Adjanohoun et al., 1988). Dans
l'alimentation, les fruits ou graines de X. aethiopica sont
utilisés entiers ou écrasés comme aromatisant (Dupriez et
De Leener, 1987).
Figure 1: Fruits
secs de X. aethiopica , (
www.kfunigraz.ac.at)
Les composés responsables de l'effet insecticide de
l'huile essentielle de X. aethiopica appartiennent
essentiellement aux monoterpènes (Tableau 1). Les proportions
de ces composés chimiques par groupe figurent en annexe 1.
Tableau 1:
Proportion des principaux groupes des composés chimiques
|
composés
|
%
|
|
Monoterpènes hydrocarbonés
|
83,24
|
|
Monoterpènes oxygénés
|
5,59
|
|
sesquiterpènes hydrocarbonés
|
10,58
|
2-2 Généralités sur les
coléoptères Bruchidae
Les différentes espèces de bruches se
distinguent par leur comportement, la morphologie, l'importance des dommages
causés et leurs réactions face aux méthodes de lutte. Pour
être en mesure d'apporter une réponse aux lourds
dégâts causés par ces insectes, il est absolument
indispensable de passer en revue la taxonomie, la biologie et l'écologie
de ces ravageurs dont la plupart sont d'importance économique, surtout
dans les régions chaudes du monde.
2-2-1 Taxonomie et
distribution
Les Bruchidae constituent un groupe homogène d'insectes
cléthrophages dont le développement se déroule en
général à l'intérieur d'une seule et même
graine (Delobel et Tran, 1993). La classification adoptée par Beaumont
et Cassier (1973) est la suivante :
- Règne : Animal
- Embranchement : Athropode
- Sous-Embranchement : Mandibulate ou Antennate
- Classe : Insecte
- Super-ordre :
Coléoptéroïde
- Ordre : Coleoptera
- Famille : Bruchidae
De par leur distribution, les Bruchidae constituent l'un des
groupes de ravageurs inféodés aux légumineuses à
graine les mieux connus. Elles existent sur tous les continents,
excepté l'Antarctique. Selon Credland (1990), 20 espèces se
développent aux dépens des plantes cultivées et sont
devenues des ravageurs d'importance économique. En Afrique tropicale, 16
espèces de bruches sont rencontrées sur les graines de
légumineuses alimentaires cultivées (Decelle, 1987). Ces
espèces sont représentées par au moins sept genres
(Callosobruchus, Bruchidius, Acanthoscelides,
Caryedon, Pachymerus, Zabrotes,
Spermophagus) repartis dans trois sous-familles (Decelle, 1981 ;
Taylor, 1981). Seules ces sous-familles (Bruchinae,
Amblycerinae, Pachymerinae) comprennent les espèces
nuisibles aux plantes cultivées, notamment les légumineuses
(Hoffman et al., 1962).
La grande majorité des Bruchidae nuisibles aux
légumineuses cultivées se retrouvent dans la sous-famille des
Bruchinae. Ces Bruchinae se caractérisent par un pigydium non
entièrement recouvert par les élytres, un écusson dont la
longueur ne dépasse pas la largeur ; des fémurs
postérieurs presque renflés, des yeux à échancrure
antérieure marquée (Hoffman et al., 1962). Les
espèces nuisibles de cette sous-famille appartiennent aux genres
Bruchus L., Bruchidius Schild., Acanthoscelides
Schild et Callosobruchus Pic.
Le genre Callosobruchus renferme un grand nombre
d'espèces ayant une distribution paléotropicale. Elles sont
toutes polyvoltines, et peuvent se reproduire sur les plantes, au champ et dans
les graines sèches des entrepôts. Plusieurs espèces sont
signalées sur les Fabaceae cultivées, principalement les
Phaseoleae.
2-2-2 Callosobruchus
maculatus F.
C . maculatus est une espèce très
cosmopolite qui passe pour la plus nuisible au stock du niébé
(Giga et Smith, 1983). A l'état adulte, cette bruche est
caractérisée par un dimorphisme ou des phases à
morphologie, physiologie et comportements différents. Selon ces
caractéristiques, cette espèce a été
désignée sous les noms de formes voilière ou active et de
forme normale ou non voilière (Caswell, 1961 ; Utida,
1981) :
- la forme non voilière ou normale est
caractérisée par un pigydium pigmenté chez la femelle et
peu tacheté chez le mâle. Les individus ayant cette forme ont un
muscle alaire atrophié et par conséquent, ils sont incapables de
voler (Ouédraogo, 1991). Selon les études, la
fécondité moyenne varie entre 100 oeufs par femelle (Glitho
et al., 1988) ; 60-90 oeufs (Delobel et Tran, 1993) et 91,2 oeufs par
femelle (Akpovi, 1993). Les individus sont inféodés au stock de
niébé et ont une longévité faible (6-8 jours
à 30°C) (Delobel et Tran, 1993). Les adultes de cette forme sont
toujours reproducteurs ;
- la forme voilière ou active a un pygidium clair chez
la femelle et les élytres sont tachetés chez le mâle. Les
individus de cette forme sont caractérisés par une faible
fécondité (deux oeufs en moyenne par femelle) ; leur
descendance est constituée des individus non voiliers (Glitho
et al., 1988). La longévité est d'environ un
mois à 30°C (Delobel et Tran, 1993). La teneur en eau des graines,
la température, l'humidité relative influencent l'apparition de
cette forme en élevage de masse. En effet, une forte proportion
d'adultes voiliers émergent des graines de niébé lorsque
la température est de 30,2°C, l'humidité relative 80#177;15%
et la teneur en eau 13-15% (Monge et Ouédraogo, 1990 ;
Ouédraogo et al., 1990). En outre, au cours de ses
travaux, Tchokossi (1995) a constaté que l'apparition des voiliers est
précédée par l'augmentation du nombre de larves par graine
dont l'activité entraîne l'élévation de la
température et de la teneur en eau des graines.


Figure 3: Femelle en activité de ponte
Figure 2: Femelle de bruche
(www.univ-tours.fr)
2-2-3 Développement
de Callosobruchus maculatus
i- OEuf
Après la copulation, C. maculatus
dépose ses oeufs sur les graines (Alzouma, 1981) ; les oeufs sont
de forme ovoïde et sont déposés sur le péricarpe
(Huignard et al., 1985 ; Delobel et Tran, 1993). Les oeufs sont
pondus et fixés par la femelle avec une substance gélatineuse
à la surface des graines et des gousses. La dimension des oeufs pondus
par C. maculatus varie selon le substrat et influence la survie de la
descendance. Ainsi, Fox (1994) estime que les descendants issus de gros oeufs
éclosent plus rapidement, ont un taux d'éclosion
élevé, un taux de survie larvaire élevé, se
développent plus rapidement en nymphes et produisent des adultes grands,
et plus tolérants à la pénurie alimentaire et à la
dessiccation. Kawechi (1995) affirme que les femelles de C. maculatus
peuvent anticiper la compétition larvaire future due aux conditions
de croissance en produisant de gros oeufs pour améliorer les chances de
survie de sa descendance (van Huis et de Rooy, 1998). Lors de la ponte, une
phéromone de marquage est déposée en même temps que
l'oeuf (Delobel et Tran, 1993) ; elle permet à la femelle
d'éviter les graines fortement infestées, et de réduire
ainsi la compétition inter-larvaire.
ii - Larve
L'éclosion des oeufs survient 5 à 7 jours
après la ponte dans les conditions les plus favorables. Après
éclosion, la larve qui n'a pas encore de pattes motrices, fore la paroi
externe de l'enveloppe de la gousse ou du tégument de la graine et
rentre à l'intérieur (Lenting, 2000). A la différence de
la graine où la larve peut accéder directement aux
réserves nutritives avec plus de chance de survie ; au niveau des
gousses, la jeune larve peut échouer dans le vide (espace inter-grain)
ou déboucher, au cours de son transit trophique, sur un petit grain
à faible réserve ne pouvant lui permettre d'atteindre le stade
final de développement. Dans les derniers cas, la mort de la larve est
inévitable. La larve néonée de type chrysomelien doit
creuser à travers la cloison de la gousse ou de la graine pour chercher
sa nourriture. Elle est pourvue, à cet effet, d'une plaque de chitine
sclérotinisée en forme de H. Beaucoup de larves néonates
creusent à travers l'enveloppe de la gousse et entrent dans la
première graine disponible.
Alzouma et al. (1996) affirment que des larves de
Bruchidae possèdent un équipement enzymatique leur permettant de
détruire les composés secondaires (inhibiteurs de
protéases, phytohémagglutines, acides aminés non
protéiques) présents dans les graines de leurs
légumineuses hôtes. Ces possibilités de détoxication
permettent aux larves de Bruchidae d'utiliser sans mortalité importante
et avec un très bon rendement énergétique les graines des
légumineuses riches en composés secondaires, toxiques
vis-à-vis des autres consommateurs potentiels. Néanmoins, le
tégument externe est la principale barrière qui contribue
significativement à la mortalité de la larve néonée
durant la phase de perforation (Desroche et al., 1995).
iii- La nymphe
C. maculatus se nymphose à l'intérieur
d'une seule graine après avoir construit une fenêtre au niveau du
tégument (Southgate, 1979). La durée de la nymphose varie entre 7
à 28 jours. La durée moyenne de développement est de 30
jours, mais elle varie dans des limites assez larges en fonction des
caractéristiques de la graine-hôte (Kossou et Aho, 1993).
iv- L'adulte
Au terme de la nymphose, l'adulte émerge de la graine
pour commencer un nouveau cycle après avoir traversé la
fenêtre de sortie préparée par la larve. Le pourcentage le
plus élevé d'adultes s'obtient à partir des oeufs pondus
sur les graines. Avec les pontes sur graines vertes ou mûres, la
mortalité élevée se produirait lors de la
pénétration dans la gousse (Messina, 1984). La durée de
développement oeuf-adulte est fonction des conditions
atmosphériques. Dans les conditions optimales (30-35°C ;
70-90% HR), le développement complet varie de 22 -25 jours (Rees,
1996).
2-2-4 Dégâts
et importance de Callosobruchus maculatus sur le niébé
C. maculatus exerce une pression permanente sur les
légumineuses ; cette menace est due à sa grande polyphagie
et sa faculté d'adaptation à des régions climatiques
variées (Utida, 1954). L'adulte de C. maculatus ne cause pas
directement de dégâts sur le niébé au champ ni en
stock ; les individus mâles et femelles vivent des grains de pollen
et de nectar (Alzouma, 1981 ; Messina, 1984). Ce sont les stades
juvéniles qui causent les ravages sur stock en consommant, avant le
stade pupe une partie importante des réserves nutritives des
cotylédons de la graine du niébé. L'activité de
consommation des réserves des graines par les larves provoque de la
chaleur et de l'humidité qui engendrent le développement des
champignons comme Aspergillus, Fusarium, Botrytis,
conduisant à des pertes quantitatives. En zone rurale où les
techniques de conservation des denrées agricoles sont peu
élaborées, ce ravageur entraîne des pertes
post-récoltes pouvant atteindre 100% en quelques mois (Lienard et Seck,
1994). C. maculatus entraîne non seulement une réduction
de poids sec, mais également une diminution de la qualité des
graines et de la viabilité des semences compromettant leur consommation
et leur semis (Singh et Singh, 1992 ; Odah, 1995). Selon Murdock
et al., (1997) lorsque les dégâts
excèdent un trou d'émergence par graine, la valeur marchande du
niébé s'en trouve substantiellement réduite. Tchamou
(1995) a montré qu'il existe une relation entre le nombre de trous
d'émergence des bruches sur les graines et les pertes de poids.
2-3 Ennemis naturels
associés à Callosobruchus maculatus
Le tableau 2 présente les ennemis naturels de C.
maculatus vus par van Huis (1991). La biologie des espèces majeures
d'ennemis naturels de C. maculatus et leur importance en tant qu'agent
potentiel de lutte biologique sont présentées ci-dessous.
U. lariophaga est un minuscule
hyménoptère, chalcidien qui apparaît dans les cultures
quelques jours après l'infestation des gousses par les bruches. Du point
de vue biologique, la vie de U. lariophaga se divise en deux
phases : la phase de parasite oophage au cours de laquelle la source de
nourriture est l'oeuf de l'hôte et une phase libre correspondant à
la vie imaginale durant laquelle l'adulte se nourrit de miellat de pucerons. Ce
parasitoïde est surtout attiré par les oeufs fraîchement
pondus par C. maculatus (0-2 jours après oviposition) selon van
Huis et al., (1991). L'utilité de cet insecte
réside dans le fait qu'il élimine le ravageur avant son
émergence ; ce qui fait que de U. lariophaga est un
oophage potentiel de contrôle biologique des populations de bruches
ravageurs de niébé. Le taux de parasitisme des oeufs de C.
maculatus dans les cultures de niébé varient entre
25 et 60% suivant les zones écologiques ou les conditions
expérimentales (Alzouma, 1987). Dans les greniers expérimentaux
de niébé, 33 à 60% des oeufs de C. maculatus sont
parasités (Lammers et van Huis, 1989 ; Monge et
al., 1991).
Les parasitoïdes larvophages A.
calandrae, D. basalis, E. vuilleti et E.
orientalis sont des ectoparasites solitaires. Les larves de D.
basalis, E. vuilleti et E. orientalis ne se
développent qu'aux dépens des derniers stades larvaires (larves
L4) et des nymphes de C. maculatus (Tchanssanti, 1995 ; Tchamou,
1995). Sur le plan biologique, les femelles de E. vuilleti
parasitent les hôtes portant les oeufs ou les larves de D.
basalis alors que ce dernier évite les hôtes parasités
par E. vuilleti. Ainsi, lorsque les deux espèces sont
introduites, la densité de D. basalis est faible et varie peu
tandis que celle de E. vuilleti augmente progressivement. En
présence des deux espèces, le taux d'accroissement de C.
maculatus reste élevé (van Alebeek et al.,
1993 ; Monge et al., 1995 ; Tchamou, 1995). Le taux
de parasitisme de ces espèces varie entre 7-60% (Monge et Huignard,
1991). A. calandrae pond plus d'oeufs femelles dans les
grosses larves et plus d'oeufs mâles dans les petites larves. Cette
espèce attaque de préférence les larves âgées
et les pupes. Le type de grain affecte la gamme de taille d'hôte
parasité par A. calandrae. La femelle d'A.
calandrae est capable de détecter les larves vivantes dans les
grains grâce à de nombreuses odeurs des produits de l'hôte
comme les déjections. Elle sait également détecter les
mouvements de l'hôte et les bruits de ses actions.
A. calandrae est une espèce qui a
adopté une stratégie généraliste étant
donné la grande gamme d'hôte dont il est le parasite potentiel.
Parmi les hôtes potentiels, on cite Sitophilus zeamais,
charançon du maïs; Sitophilus oryzae dans le riz, C.
maculatus dans le niébé.
Tableau 2: Ennemis
naturels de Callosobruchus maculatus
|
Insectes auxiliaires
|
Références bibliographiques
|
|
TRICHOGRAMMATIDAE
Uscana lariophaga Steff
Uscana semifumipennis Girault
Uscana mukerju Mani
|
Monge et al.,(1990)
van Huis (1991)
Kapila et Agarwal (1990)
|
|
EUPELMIDAE
Eupelmus orientalis Crawf
Eupelmus vuilleti Crawf
|
van Huis (1991)
Monge et al.,(1990)
|
|
PTEROMALIDAE
Anisopteromalus calandrae How
Chaetospila elegans Westwood
Dinarmus basalis Rondani
Dinarmus vagabundus Timberleke
Lariophaga texacus Crawf
|
Heong (1981)
van Huis (1991)
Rojas-Rousse et al.,(1988)
van Huis (1991)
|
|
EURYTOMMIDAE
Eurytoma sp.
|
Prevett (1961)
|
(Source : Djidonou, 2000)
2-4 Méthodes de
contrôle de Callosobruchus maculatus
L'infestation des grains par les insectes, lorsqu'elle n'est
pas maîtrisée, entraîne de lourdes pertes. Les
méthodes de gestion des stocks généralement
utilisées sont la lutte chimique utilisant les insecticides de
synthèse ou d'origine naturelle, les méthodes traditionnelles, la
lutte biologique et la lutte intégrée.
2-4-1 La lutte chimique
L'application des pesticides de synthèse très
répandue dans les structures de stockage se fait essentiellement par
deux groupes de produits à savoir les insecticides de contact
pénétrant les tissus de l'insecte après avoir
traversé la cuticule, et les fumigènes qui agissent sur le
système respiratoire (Kossou et Aho, 1993).
Les familles d'insecticides les plus utilisées dans les
zones de culture du niébé sont les organophosphorés et
les organochlorés suivis par les pyréthrinoïdes et les
carbamates (Ngamo, 2004). Parmi les organochlorés, le lindane est
très efficace contre C. maculatus. Les composés
organophosphorés (malathion, pirimiphos méthyle) sont
caractérisés par une bonne efficacité contre les ravageurs
de stock. Les carbamates (le carbaryl) sont d'un emploi limité dans le
domaine de la protection des cultures. Les pyréthrinoïdes de
synthèse possèdent une efficacité relative contre les
coléoptères. En condition sahélienne, la
deltaméthrine à la dose 1ppm assure une protection efficace du
niébé pendant 6 à 7 mois de stockage (Hussein et
Abdel-Aal, 1982 ; Seck et al., 1991).
Parmi les fumigants, l'hydrogène phosphoré (la
phosphine : PH3) est le plus couramment utilisé dans les pays
chauds, et s'avère très efficace contre les oeufs et les larves
de C. maculatus (Singh et al., 1990). Le bromure de
méthyle (CH3Br), le phosphore d'aluminium, le tétrachlorure de
carbone, l'acide cyanydrique ont été longtemps utilisés
à titre curatif par les paysans en Afrique de l'Ouest et au Sud de
l'Inde.
Nonobstant leur efficacité, l'utilisation des
insecticides de synthèse et des fumigants n'est pas une stratégie
recommandable pour le contrôle des ravageurs. La plupart de ces produits
utilisés sont très toxiques. Selon la banque mondiale
citée par Jackai (1995b), les valeurs des Doses Létales DL50 de
plusieurs insecticides utilisés dans les exploitations sont en dessous
de la norme (200mg/kg). Ces pesticides engendrent une pollution de
l'environnement et constituent des risques pour la santé des hommes et
des animaux ; ils affectent l'équilibre biologique de
l'écosystème du niébé. De plus, les
Coléoptères des denrées ont développé des
phénomènes de résistance en raison de la
multiplicité, de la nature, de la dose et de la technique des
traitements (Delobel et Tran, 1993 ; Haubruge et Amichot, 1998 ;
Isman, 2000). Les mécanismes de résistance peuvent être
d'ordre comportemental, physiologique ou biochimique.
2-4-2 Méthodes
traditionnelles de lutte
Ces méthodes comprennent les pratiques endogènes
au milieu paysan acquises au cours de générations. Outre les
pratiques religieuses et magiques, ces méthodes comprennent les
techniques culturales, le triage de la récolte, la désinfection
des structures de stockage (GTZ, 1995 ; Lienard et Seck, 1994). En absence
d'un programme approprié de contrôle, les cultures
associées assurent dans les fermes traditionnelles moins de perte
(Kumar, 1984) en rendant l'environnement défavorable pour les ravageurs
ou en réduisant les niveaux d'infestation au champ ou alternativement
elles fournissent un environnement favorable aux ennemis naturels.
Au nombre des méthodes préventives, on peut
aussi citer l'utilisation de la température et de l'humidité,
l'addition de substances minérales, l'addition des substances
végétales et le stockage hermétique.
2-4-3 Méthodes
intégrées de lutte sans utilisation de produits de
synthèse
Les avantages offerts par les méthodes biologiques de
lutte résident dans l'absence presque totale des risques toxicologiques
et leur durabilité. Ces méthodes comprennent entre autres la
résistance de la plante hôte, la lutte biologique (utilisation des
entomophages) et la manipulation de l'environnement de l'insecte (Kossou et
Aho, 1993).
2-4-3-1 La résistance
variétale
Les études, les avancées réalisées
durant plusieurs années sur plus de 8000 variétés de
niébé ont abouti à sélectionner une
variété (Tvu 2027) qui présente une résistance
significative à C. maculatus (van Huis 1991; Lienard et
Seck, 1994 ; Kezan et al., 1994). Les facteurs que sont
une forte teneur en protéine, la texture de la graine et la paroi de la
gousse sont considérés comme étant en relation directe
avec la résistance des variétés de niébé
à l'égard de C. maculatus (Jackai et Daoust,
1986).
2-4-3-2 La lutte biologique
La lutte biologique constitue de nos jours la stratégie
sur laquelle se focalisent les attentions des institutions de recherche. Pour
la gestion des bruches de niébé, les études ont
montré que les espèces de U. lariophaga et
D. basalis respectivement parasitoïde oophage et
larvophage de C. maculatus constituent de potentiels
candidats pour une lutte biologique en Afrique de l'ouest (van Huis et
al., 1994 ; Thomas-Odjo et Gaspar, 1994 ; Monge et
al., 1995 ; Ouedraogo et al., 1996). Ainsi, un programme de
lutte biologique reste envisageable pour la protection des graines du
niébé contre C. maculatus.
2-4-3-3 Procédés
biotechniques
Ces procédés font intervenir le comportement des
ravageurs, qui vont ainsi contribuer activement à leur propre
élimination. Les méthodes de lutte employées exploitent en
l'occurrence les réactions naturelles des ravageurs face aux stimuli
émanent de l'environnement. Parmi ces procédés, on peut
citer l'utilisation d'appâts, les phéromones, les attractifs, les
répulsifs et les inhibiteurs de croissance (Gwinner et al.,
1996).
2-4-3-4 La lutte intégrée ou
« Integrated Pest Management »
La lutte intégrée suppose la
combinaison efficace et économique de plusieurs méthodes de lutte
(chimique, biologique, culturale, etc) en vue de réduire les populations
de ravageurs, pour l'obtention d'un meilleur rendement, sans trop perturber
l'environnement. Ainsi, la gestion intégrée des ravageurs est un
système écologiquement et économiquement durable dans
lequel les dommages causés par les insectes sont prévenus par
l'utilisation des facteurs naturels qui limitent la croissance des populations
des organismes, et au besoin suppléés par des mesures de
contrôle appropriées (van Lenteren et Overholt, 1994).
2-4-4 Les
phytopesticides
2-4-4-1 Généralités
Compte tenu des dangers que présente l'utilisation des
insecticides synthétiques, de nombreuses recherches sont axées
sur l'utilisation des extraits des plantes douées des
propriétés anti-insectes et dont les effets indésirables
n'ont pas été signalés chez l'homme (Isman, 2000). Il
s'agit souvent de plantes consommées dans l'alimentation humaine, par
exemple comme condiment ou épice ou des plantes à activité
médicinale. A cause du coût élevé des produits
chimiques de protection des denrées et pour pallier les pertes
post-récoltes, les producteurs utilisent des techniques alternatives
telles que : le séchage en plein air, le séchage au-dessus
d'un foyer par fumage, l'emploi des substances végétales et
minérales (terres des diatomées), l'utilisation des huiles
végétales au cours du séchage (Giles, 1964 ; Stoll,
1988 ; Bell et al., 1998 ; Stoll, 2000 ; Boeke
et al., 2004).
De nombreuses espèces végétales sont
utilisées traditionnellement pour la protection des denrées,
c'est le cas du margousier ou neem (Azadirachta indica A.
Juss.) (Meliaceae) dont les composés isolés ont
été testés (Akou Edi, 1993), des huiles d'arachide, de
coton, de soja et de maïs (Stoll, 1988 ; Stoll, 2000). De nombreuses
substances végétales se sont montrées comme des agents
répulsifs, perturbateurs du développement des ravageurs ou
insecticides, par de la plante entière ou certaines de ses parties
(Giles, 1964 ; Lambert et al., 1985 ; Bekele et Hassanali,
2001 ; Bouda et al., 2001 ; Tapondjou et al., 2002 ;
Arnason et al., 2002 ; Kim et al., 2003 ; Boeke
et al., 2004). Les familles les plus promotteuses dans la protection
des denrées au cours du stockage appartiennent aux familles des
Meliaceae, Annonaceae, Labiatae, Rutaceae, Asteracae, Canellaaceae (Jacobson,
1989).
L'utilisation des huiles essentielles pourrait aider à
réduire les effets des ravageurs des denrées stockées
à un coût acceptable vu que ces huiles peuvent être
produites artisanalement sur place. Dans les années 1970, la
possibilité d'emploi des composés naturels a été
démontrée (Szafranski et al., 1991). Les insecticides
végétaux en particulier les huiles essentielles ont fait l'objet
de nombreuses recherches dans le but de diminuer les pertes
post-récoltes : ce sont des produits biodégradables,
respectueux de l'environnement et de la faune auxiliaire.
2-4-4-2 Les huiles essentielles comme alternative de
lutte
Les huiles essentielles sont des mélanges très
complexes de monoterpénoïdes, de sesquiterponoïdes et des
phénols (Isman, 2002). Ce sont des produits obtenus à partir des
végétaux dans lesquels les caractéristiques
odoriférantes et aromatiques sont concentrées (Pruthi, 1980). Les
huiles essentielles peuvent être extraites par hydrodistillation,
distillation par vapeur d'eau et hydrodiffusion (Tuley de Silva, 1995).
Au Cameroun, les études réalisées sur les
huiles essentielles ont été pour la plupart axées sur
l'analyse de la composition chimique par chromatographie en phase gazeuse
couplée à la spectrométrie de masse (CG/SM). On peut citer
à cet effet les travaux de Amvam et al., (1991) sur
Zingiber officinale ; Jirovetz et al., (1997) sur X.
aethiopica et, Xylopia parviflora ; Ngassoun et
al., (1999) sur Plectranthus Glandulosus ; Jirovetz et
al., (2000) sur O. gratissimum et X. aethiopica.
D'autres travaux sont orientés dans l'évaluation de
l'efficacité des huiles essentielles d'O. gratissimum et X.
aethiopica soit sur C. maculatus ou S. zeamais (Kouninki
et al., 2005).
Sur le plan mondial, la production des huiles essentielles par
an est d'environ 30 tonnes, ce qui représente un chiffre d'affaires de
15 milliards de francs CFA (23 millions d'Euros) (Amvam et
al., 1991 ; Tchoumbougnang, 1997). Actuellement plus de 200
huiles essentielles font l'objet d'un commerce mondial particulièrement
actif.
Les principaux producteurs sont : les Etats-Unis, la
Chine, l'URSS, le Maroc, la Bulgarie, l'Inde, La France, l'Egypte et l'Espagne.
Le Tableau 3 donne des estimations de la production totale de quelques pays en
milliers de dollars. Ce tableau montre que la production d'huile essentielle
est absente dans bon nombre de pays producteurs de niébé. Le
développement de la production de cette huile serait non seulement
générateur de revenus et d'emplois, permettrait aussi la
fabrication des produits insecticides naturels, porteurs d'espoir pour la
protection des denrées stockées.
Tableau 3:
Estimation de la production d'huile essentielle de quelques pays en
milliers de dollars
|
Pays
|
Valeurs
|
|
USA
Chine
URSS
Maroc
Bulgarie
Inde
France
Egypte
Espagne
Algérie
Haïti
Madagascar
Côte d'Ivoire
Burkina Faso
Cuba
|
145000
110000
30000
30000
26000
25000
20000
12000
10000
8000
8000
6000
3500
500
500
|
(Source : Tchounbougnang, 1997)
2-4-4-3 Importance et originalité des huiles
essentielles des plantes aromatiques
pour la conservation des denrées
au cours du stockage
Les plantes ont été longtemps utilisées
par les paysans pour saveur des aliments ou pour protéger les produits
récoltés (Jacobson, 1989 ; Keita et al.,
2000 ; Isman, 2000). Les huiles essentielles extraites de ces plantes ont
été largement utilisées dans la lutte contre les ravageurs
de stocks (Dunkel et Sears, 1998 ; Hamraoui et Regnault-Roger, 1997 ;
Prates et al., 1998 ; Liu et Ho, 1999 ; Golop et
al., 1999 ; Tunç et al., 2000 ; Isman, 2000).
Leurs propriétés insecticides, larvicides et ovicides,
stérilisantes, antiappétentes, répulsives ont fait l'objet
de plusieurs études (Hamraoui et Regnault-Roger, 1997 ; Stoll,
2000). La sensibilité d'un insecte pour une huile essentielle
évolue en fonction de son cycle biologique. La cible d'action de ces
huiles au niveau des insectes est différente de celle des hommes, ce qui
fait qu'aucune toxicité directe sur l'Homme n'a été pour
le moment mis en évidence. En effet, les récepteurs chimiques
chez les insectes n'existent pas dans le système nerveux des
mammifères (Isman, 2000). Ainsi, il a été montré
que plusieurs composés des huiles essentielles (thymol, eugenol) sont
des neurotoxines qui interfèrent avec le neurotransmetteur octopamine
unique aux Arthropodes (Isman, 2002). A cause de leur faible persistance, les
huiles essentielles des plantes aromatiques ne présentent pas de risques
pour l'environnement. Leur activité peut varier en fonction des stades
du cycle de vie d'un insecte (Regnault-Roger, 2002). Les molécules
allélochimiques végétales appartiennent au
métabolisme secondaire : terpènes, alcaloïdes ou
glucosides, molécules qui sont facilement biodégradables par voie
enzymatique, aucun phénomène de bioamplification n'ayant
été décrit (Isman, 2002). Ces composés
allélochimiques ne développent que peu de toxicité pour
les vertébrés et sont régulièrement
consommés dans l'alimentation (Regnault-Roger, 2002).
Les composés chimiques responsables des actions sur les
ravageurs de stock sont les monoterpènes (1-8 cinéole,
l'eugenol, le camphor présents dans les huiles essentielles de O.
gratissimum et X. aethiopica) à effet insecticide sur la
survie de C. maculatus ou S. zeamais (Kouninki et
al., 2005) ou des alcaloïdes à effet inhibiteur du
développement larvaire des ravageurs comme T. castananeum
(Ngamo et al., 2001). Les recherches menées dans le but
d'étudier l'effet insecticide de certaines plantes sur de nombreux
insectes d'importance économique ont révélé des
effets destructeurs et inhibiteurs de la croissance (effet par contact, par
ingestion et par inhalation) et des effets répulsifs. Ces études
ont été réalisées sur des plantes
fréquemment utilisées localement par les paysans comme
épice, condiment, ou en médecine traditionnelle.
Les résultats des tests insecticides effectués
sur C. maculatus avec diverses formulations de plantes et quelques
formulations à base de X. aethiopica utilisées pour le
contrôle des insectes sont consignés dans le tableau 4
Tableau 4: Tests
insecticides réalisés avec des formulations de diverses
plantes
|
Références
|
Plante
|
Formulation
|
Mode d'application
|
Mode d'action
|
|
Keita et al. (1999)
|
O. gratissimum
|
Poudre
|
Mélange d'huile essentielle avec du kaolin
|
Protection du niébé contre les attaques de
C. maculatus
six mois après traitement
|
|
Keita et al. (2000)
|
Hyptis suavolens, Ocimum canum, Ocimum basilicum, Tagetes
minuta, Piper guineense
|
Poudre de kaolin extrait ethanolique
|
Fumigation
|
Effet insecticide
et pertubateur du développement de C.
maculatus
|
|
Raja et al. (2001)
|
Mentha arvensis, M. piperata, M. spicata et Cymbopogum
mordus
|
Composés volatiles de l'huile essentielle
|
Fumigation
|
Effet ovicide et insecticide sur C.
maculatus
|
|
Keita et al., (2001)
|
O. basilicum et O. gratissimum
|
Poudre
|
Mélange d'huile essentielle avec du kaolin
|
Effets fumigant et insecticide sur
les adultes, effet retardateur de la croissance de C.
maculatus
|
|
Tapondjou et al., (2002)
|
Chenopodium ambrosioides
|
Huile essentielle
|
Toxicité par contact et fumigation
|
Effets insecticide
et fumigation observés après 24 heures
sur
C. maculatus
|
|
Pascual-villabos et Balletesta-Acosta (2003)
|
Ocimum basilicum
|
Huile essentielle
|
Toxicité par contact et répulsion
|
Inhibition de l'oviposition, effet répulsif et toxique
pour les adultes de C. maculatus
|
|
Tapondjou et al. (2003)
|
Chenopodium ambrosiodes L. et Eucalyptus
saligna
|
Huile essentielle
|
Toxicité par contact
|
Effet insecticide par contact sur
C. maculatus ;
C. ambrosiodes est plus efficace que E.
saligna
|
|
Jirovetz et al. (2000)
|
X. aethiopica et O. gratissimum
|
Poudre
|
Mélange d'huile essentielle avec du kaolin
|
Effets insecticides sur S. zeamais
|
|
Ngamo et al. (2001)
|
X. aethiopica et O. gratissimum
|
Poudre
|
Mélange d'huile essentielle avec du kaolin
|
Effet insecticide
de deux huiles essentielles sur S. zeamais
|
(Source : Kouninki, 2005)
CHAPITRE III : METHODOLOGIE
3- Matériels et
méthodes
Dans plusieurs agro-systèmes en Afrique, les plantes
aromatiques sont utilisées par les producteurs pour la protection des
denrées stockées. L'utilisation de formulations (poudre ou huile
essentielle) issues de différentes parties de ces plantes a permis de
montrer l'effet knock down de ces formulations mais aucune étude n'a
permis d'apprécier l'efficacité à long terme des
formulations. Or le stockage des grains doit se faire pendant plusieurs mois,
une quantification de la rémanence est donc indispensable.
La destruction des bruches dans le niébé en
stock (action de choc) n'est qu'un aspect de l'efficacité des
formulations agissant comme insecticide de contact. Cette étude a pour
objectif de vérifier la rémanence effective en fonction du type
de formulation. Une rémanence plus élevée réduit
considérablement les risques de réinfestation en provenance de
l'extérieur ou par une nouvelle génération après
l'utilisation du produit contre les parents. Nous avons comparé
l'efficacité de l'huile essentielle de X. aethiopica seule dans
l'acétone par rapport à la poudre de fruits de X.
aethiopica seule, ou à un mélange poudre-huile. Les mesures
de l'activité biologique ont été effectuées par
rapport à des témoins. Le choix du mélange «poudre de
fruit-huile» a été guidé par la possibilité de
réaliser ainsi une formulation simple à préparer et peu
coûteuse dans les conditions locales d'utilisation.
3-1 Matériels
Elevage des bruches
L'expérimentation s'est déroulée au
Centre de Recherche sur la Biodiversité dans l'Unité d'Ecologie
et de Biogéographie de la Faculté des Sciences de
l'Université Catholique de Louvain.
L'élevage en masse de la bruche C. maculatus
et les tests menés ont été réalisés dans des
bocaux en verre (16 cm de hauteur et 9 cm de diamètre), sur les graines
de niébé V. unguiculata appelé haricot à
"oeil noirØ (Figure 4).


Figure 4: Bocaux
en verre contenant graines de niébé et bruches
Les bocaux dont les couvercles sont percés
étaient maintenus à l'obscurité dans une armoire
vitrée maintenue à une température de 28#177;2°C et
une humidité relative de 65#177;5%. La souche d'origine des bruches
provient des structures de stockage traditionnelles des producteurs du Grand-
Nord Cameroun et est entretenue dans le laboratoire depuis 2003.
Matériel
végétal
Les fruits secs de X. aethiopica ont
été achetés à Ngaoundéré au Sud du
Cameroun. La «poudre» utilisée et qui constitue le substrat
principal de la formulation est obtenue en écrasant ces fruits secs et
puis en tamisant sur une maille inférieure ou égale à 1mm.
L'huile essentielle ajoutée à cette poudre a été
obtenue par hydrodistillation pendant quatre heures à l'aide d'un
dispositif de type clevenger au laboratoire de Chimie des substances actives de
l'Université de Ngaoundéré au Cameroun. Le matériel
végétal broyé est placé dans un ballon avec de
l'eau. Le tout est porté à ébullition à l'aide
d'une plaque chauffante. La vapeur dégagée est condensée
dans une colonne de réfrigération dans laquelle circule de l'eau
en continu. Le liquide qui s'écoule de la colonne arrive dans le
séparateur où l'huile essentielle se sépare de l'eau par
décantation. Le séparateur utilisé permet de lire le
volume d'huile essentielle et de la recueillir pendant ou à la fin de la
distillation. L'huile essentielle obtenue a été
séchée sur du sulfate d'anhydride de sodium et analysée
par la chromatographie en phase liquide couplée à la
spectrométrie de masse.
3-2 Méthodes
3-2-1 Effet
rémanent des formulations de fruits secs de Xylopia aethiopica sur
la
mortalité de Callosobruchus maculatus, la ponte des
adultes
L'expérimentation comporte les traitements T01,
T02, T03 et THE contenant tous 200g de
niébé par bocal. Les traitements testés sont le
témoin T01 (niébé seul), l'huile
essentielle diluée dans l'acétone T02, la poudre
de X. aethiopica T03 et la poudre
mélangée à l'huile essentielle de X.
aethiopica THE.
Le traitement T01 ne reçoit aucun
autre ajout c'est-à-dire constitué seulement du
niébé. Dans le traitement T02, la dilution a
été réalisée à raison de 0,4ml d'huile pour
6ml d'acétone que l'on laisse s'évaporer pendant 5mn avant de
mélanger à 200g de grains. La dose de poudre
utilisée en T03 est de 1% (poids de poudre par poids de
graines) de X. aethiopica. Dans le traitement THE la
concentration d'huile essentielle ajoutée à la poudre est de 0,2%
(0,2ml d'huile essentielle pour 100g de graines) pour 2g de poudre de X.
aethiopica. Les doses d'huile essentielle ou de poudre de X.
aethiopica utilisées ont été choisies en s'appuyant
sur les résultats des travaux précédents et suite aux
essais préliminaires réalisées sur 48h.
Pour chaque traitement, un lot de 56 pots ont
été constitués correspondant à 8 pots par
traitement pour chacun des 7 jours d'infestation des graines de
niébé soit au total 224 pots pour les quatre traitements pour
toute la durée de l'essai. Tous les pots ainsi constitués ont
reçu 200g de graines de niébé chacun au jour de
lancement de l'expérimentation. Ce même jour, les pots de
tous les traitements ont été traités par
les différentes formulations : l'huile essentielle diluée
dans l'acétone pour T02, la poudre de X. aethiopica
pour T03, la poudre
mélangée à l'huile essentielle de X.
aethiopica pour THE.
Ensuite, 0, 3, 6, 9, 12, 15 et 18 jours après le
traitement des graines de niébé par les différentes
formulations, 32 pots sont retirés du lot des 224; 20 adultes
mâles et 20 adultes femelles de C. maculatus sont
introduits par pot avec 8 répétitions par traitement de graines
traitées (T02 , T03, et THE) ou non
traitées (T01). La détermination des sexes s'est
faite selon la technique de Bandara et Saxena (1995) (Figure 5).
Femelle
Mâle
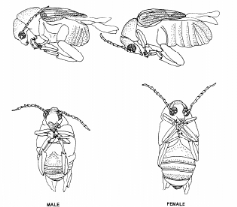
Figure 5:
Caractéristiques abdominales distinctives des mâles et
femelles
de Callosobruchus maculatus
âgés d'un jour (Bandara et Saxena, 1995)
Les différents mélanges (niébé
avec ou sans formulation) sont laissés en observation pour la mesure des
différents paramètres. Tous les trois jours après
infestation d'un lot de 32 pots, chaque pot est tamisé à l'aide
d'un tamis manuel dont les mailles ne laissent passer que les bruches. Les
bruches mortes sont recensées et le taux de mortalité est
déterminé. Etait considéré comme mort, tout
individu sans réaction des pattes et antennes après plusieurs
touches avec les pinces.
Après avoir évalué la mortalité
des bruches, 20g de graines sont prélevées par pot de graines
tamisées, par traitement avec 8 répétitions et les graines
portant les traces de ponte sont comptées sous loupe binoculaire et
pesées. Le taux de ponte a été calculé en faisant
le rapport entre le poids de graines portant les traces de ponte sur la
quantité de graines observée. Les graines sont ensuite
reversées dans chaque bocal et laissées en incubation pour
apprécier 40 jours après l'émergence de la première
génération et la perte de poids. Le Tableau 5 résume le
dispositif expérimental.
Tableau 5: Nombre de
pots par traitement par jour d'introduction des bruches
|
Jour d'introduction des bruches
|
0
|
3
|
6
|
9
|
12
|
15
|
18
|
|
Jour d'observation correspondant
|
J3
|
J6
|
J9
|
J12
|
J15
|
J18
|
J21
|
|
T01
|
8
|
8
|
8
|
8
|
8
|
8
|
8
|
|
T02
|
8
|
8
|
8
|
8
|
8
|
8
|
8
|
|
T03
|
8
|
8
|
8
|
8
|
8
|
8
|
8
|
|
THE
|
8
|
8
|
8
|
8
|
8
|
8
|
8
|
Jn : observation faite n jours après le traitement
des graines par les formulations
3-2-2 Effet des
formulations de fruits secs de Xylopia aethiopica sur l'émergence
des adultes
de Callosobruchus maculatus et la perte de poids
L'émergence de la première
génération a été observée après 40
jours d'incubation à compter de la date d'évaluation des
mortalités. Ainsi, 40 jours après observation des pots du jour
J3, les 8 pots de chaque traitement sont tamisés à l'aide d'un
tamis manuel et il a été procédé au comptage des
individus de C. maculatus émergés. Il en est de
même pour les autres jours d'observation (J6, J9, J12, J15, J18, J21).
Les graines ainsi tamisées sont ensuite pesées pour
apprécier la perte de poids par rapport au poids initial de 200g.
Après le pesage et le tamisage du contenu de chaque
pot, 20g de graines sont prélevés par pot et par traitement. Les
graines portant des trous d'émergence des bruches (graines
attaquées) sont séparées des graines saines. Les graines
saines et attaquées sont pesées pour avoir le poids des graines
attaquées et le poids des graines saines ; l'appréciation
des dégâts a été faite grâce aux pourcentages
d'attaque.
- Ns : nombre des graines saines,
- Na : nombre de graines
attaquées.
Le pourcentage d'attaque A est :
A%= Na/[Ns + Na]
× 100 (
www.fao.org/wairdocs)
Par ailleurs, 10 lots de graines pesant 20g chacun ont
été prélevés pour déterminer le poids moyen
d'une graine (Annexe 2).
3-3 Méthodes
d'analyse des données
Les pourcentages de mortalité, les taux de ponte et le
nombre d'individus de première génération
émergés ont été analysés par ANOVA I pour
rechercher les différences entre les différents traitements. Le
test Student-Newman-Keuls a été effectué pour classer les
résultats. Les pourcentages de mortalité, les taux de ponte, les
donnés d'émergence et les pertes de poids ont été
utilisés pour réaliser les graphes ou les diagrammes.
CHAPITRE IV : RESULTATS ET DISCUSSIONS
4- Résultats et
discussions
4-1 Résultats
La rémanence des différentes formulations a
été observée à partir de quatre
paramètres : la mortalité, le taux de ponte,
l'émergence de la première génération et les pertes
de poids. La mortalité et les pontes ont été
observées sur une période sur 21 jours à compter de la
date de traitement des graines par les formulations. Quant aux
émergences et aux pertes de poids des graines, elles ont
été mesurées sur une durée de 61 jours à
compter de la date de traitement des graines par les formulations.
4-1-1 Effet rémanent
des formulations sur la mortalité de Callosobruchus maculatus
La figure 6 montre le taux de mortalité des bruches
suivant les jours d'observation. Dans le traitement témoin
T01, les taux de mortalité varient entre 15% et 20%
pendant la durée de l'essai. Les pots infestés au jour 0, 3 et 9
présentent une mortalité de 100% pour le traitement THE
après 72h. Ce même taux de
mortalité est observé pour le traitement T02 au
jour d'infestation 0 et 3. Dans les traitements avec les formulations
(T02, T03 et THE), le taux de
mortalité décroît au fur et à mesure que la
durée d'exposition des bruches aux formulations augmente. Ainsi, la
dégradation ou l'évaporation de la matière active des
formulations entraîne une diminution de l'activité des
formulations utilisées, ce qui se traduit par une diminution du taux de
mortalité qui passe de 100% (THE et
T02) et 65,25% (T03) trois jours après
le traitement des graines à environ 21,9 %, 20,75% ; et 19,4%
respectivement pour THE, T02 et T03
après vingt-un jours. Toutefois, l'effet résiduel de la
formulation huile essentielle diluée dans la poudre de X.
aethiopica se maintient jusqu'au quinzième jour d'observation
puisqu'une mortalité de plus de 50% a été observée.
Ce qui n'est pas le cas pour les formulations des traitements T02
et T03 dont les taux de mortalité oscillent
autour de 20% déjà au douzième jour d'observation.
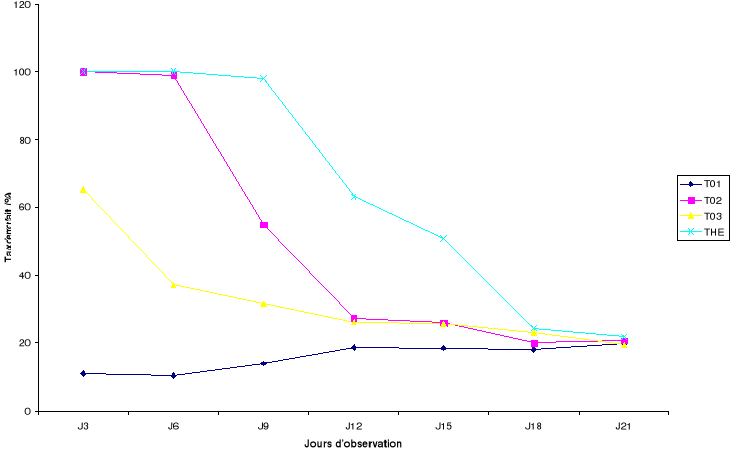
Figure 6: Evolution
des taux de mortalité en fonction de la rémanence des
formulations
Jn : observation faite n jours après le traitement
des graines par les formulations (T01 :
niébé seul, T02 : huile
essentielle diluée dans l'acétone, T03 :
poudre de X. aethiopica et THE :
huile diluée dans la poudre)
L'analyse statistique du nombre moyen de bruches morts dans
les différents traitements (Tableau 6) montre que la formulation huile
essentielle diluée dans la poudre de X. aethiopica
(THE) produit une mortalité significativement plus
élevée á =0,05 (test de Student-Newman-Keuls :
SNK) que les traitements T02 et T03 pendant
15 jours et comparativement au témoin T01, la
rémanence de la formulation THE s'étend jusqu'au
dix-huitième jour puisque à ce jour, le nombre moyen de bruches
morts dans ce traitement (9,75#177;1,83) est significativement plus
élevé que celui observé dans le témoin
(7,25#177;2,12).
Tableau 6: Nombre
moyen de bruches mort après introduction de 40 individus
|
J3
|
J6
|
J9
|
J12
|
J15
|
J18
|
J21
|
|
T01
|
4,37 #177;1,8c
|
4,25#177;2,0c
|
5,62 #177;1,9d
|
7,5#177;2,0c
|
7,37#177; 2,0c
|
7,25#177;2,1b
|
7,87#177; 1,1a
|
|
T02
|
40,0#177;0,0a
|
39,62#177;1,0a
|
22,12#177; 3,3b
|
11#177;2,2b
|
10,37#177; 1,9b
|
8#177;1,6ab
|
8,37#177; 2,1a
|
|
T03
|
26,12#177;2,9b
|
14,87#177;2,8b
|
14,87#177; 2,8c
|
10,5#177;1,6b
|
10,25 #177;1,7b
|
9,25#177;1,5ab
|
7,75#177; 1,6a
|
|
THE
|
40,0#177;0,0a
|
40,0#177;0,0a
|
40,0#177;0,0a
|
25,25#177;3,4a
|
20,37 #177;1,9a
|
9,75#177;1,8a
|
8,75 #177;1,6a
|
Les nombres suivis de la même lettre dans chaque colonne ne
sont significativement différents á=0,05 (test de SNK)
4-1-2 Effet rémanent
des formulations sur la ponte de Callosobruchus maculatus
La Figure 7 présente l'évolution des pontes pour
les différentes dates d'observation.
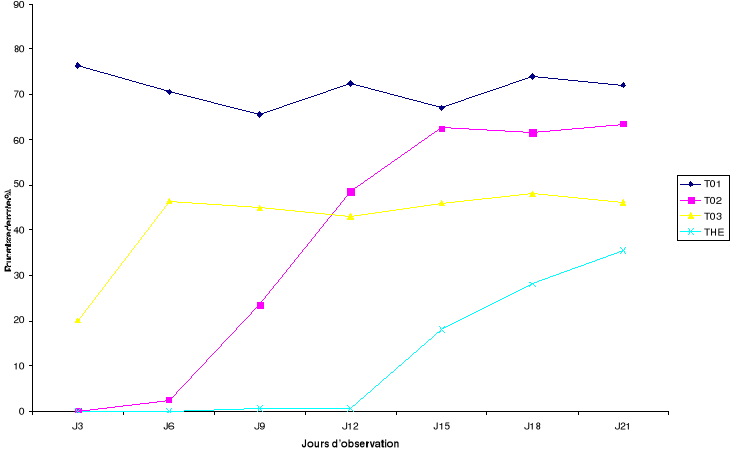
Figure 7: Evolution
des pontes en fonction de la rémanence des formulations
Jn : observation faite n jours après le traitement
des graines par les formulations (pourcentage de ponte : poids de graines
portant des traces de ponte sur poids de graines saines x 100)
Dans les pots où les graines sont infestées par
les bruches au jour 0 ; 3 ; 6 et 9 après le traitement avec la
formulation THE, l'inhibition des pontes est d'environ 100%
72h après introduction des bruches. Pour le traitement T02,
ce même taux de réduction des pontes est observé
mais seulement pour les jours d'infestation 0 et 3. Par rapport au
témoin, le traitement T03 présente une
réduction des pontes de près de 50% sur la durée de
l'essai. Le pourcentage de ponte a été plus élevé
sur les graines dans le traitement témoin que pour les
autres traitements (T02, T03 et
THE). Parmi les traitements avec formulation,
le pourcentage de ponte a été plus élevé dans le
traitement T02 après 12 jours d'incubation que
les traitements T03 et THE.
Le traitement THE apparaît comme celui présentant
le moins de graines portant les traces de pontes sur la durée de
l'essai. L'inhibition totale de la ponte des bruches par la formulation huile
essentielle mélangée à la poudre de X.
aethiopica THE a été presque totale sur
12 jours.
L'analyse statistique du poids moyen de graines portant les
traces de pontes (Tableau 7) a révélé une
différence hautement significative sur trois semaines se traduisant par
un poids de graines infestées plus élevé dans le
traitement T01 que pour les autres traitements.
Une différence significative au seuil de 5% existe entre le
poids des graines portant les traces de pontes dans le traitement THE
et les autres traitements sur la durée de
l'expérimentation. L'observation des graines avec trace de ponte sur la
durée de l'essai permet aussi de constater que la persistance d'action
de la formulation poudre de X. aethiopica seule
(T03) est meilleure au delà du 12ème
jour d'observation que pour le traitement huile essentielle seul
(T02). Il existe une différence significative entre le
poids moyen des graines portant les traces de pontes pour les traitements
T02 et T03 le 15ème
jour.
Tableau 7: Poids
moyen (g) de grain portant des traces de pontes sur 20g
|
J3
|
J6
|
J9
|
J12
|
J15
|
J18
|
J21
|
|
T01
|
14,3#177;0,6a
|
14,1#177;1,0a
|
13,1#177;1,6a
|
13,3#177;2,3a
|
13,3#177;1,2a
|
14,8#177;1,9a
|
14,0#177;0,8a
|
|
T02
|
0,0#177;0,0c
|
0,05#177;0,1c
|
4,6#177;0,9c
|
8,8#177;1,3b
|
12,5#177;0,6a
|
12,3#177;0,8b
|
12,6#177;0,9b
|
|
T03
|
4,1#177;2,1b
|
9,3#177;1,6b
|
10,0#177;1,2b
|
8,5#177;1,2b
|
9,1#177;1,9b
|
9,6#177;1,32c
|
9,2#177;0,8c
|
|
THE
|
0,0#177;0,0c
|
0,0#177;0,0c
|
0,1#177;0,3d
|
0,1#177;0,2c
|
3,7#177;0,6c
|
5,5#177;1,3d
|
7,0#177;1,0d
|
Les nombres suivis de la même lettre dans chaque colonne ne
sont significativement différents á=0,05 (test de SNK),
Jn : observation faite n jours après le traitement des graines par
les formulations
4-1-3 Effet rémanent
des formulations sur l'émergence de la première
génération
L'observation des émergences 40 jours après
chaque jour d'observation de la mortalité (Figure 8) permet
d'apprécier et de confirmer la tendance observée trois jours
après les infestations des différents supports de test par les
bruches. Le taux d'émergence pour T01 est constant et
il augmente pour les autres traitements en fonction de la perte en
activité des formulations utilisées.
Pour les traitements (T02, T03 et
THE), on note une augmentation du nombre moyen d'individus
émergés au fur et mesure de la perte en activité des
formulations. Ainsi, 55 jours après infestation des graines, le nombre
moyen de bruches émergées dans le traitement THE
est significativement moins élevé que pour les
traitements T01, T02 et T03. Les tendances
observées dans les traitements T02 et T03
se traduisent par un nombre significativement plus élevé
d'émergence pour le traitement T03 par rapport
à T02 entre le 43ème jour et le
52ème jour. Néanmoins, cette tendance a
été inversée au-delà de cette date ; ce qui se
remarque par un nombre d'individus de C. maculatus
émergés plus élevé pour le traitement T02
que pour T03.
La réduction des descendants de la première
génération est de 100% dans les pots traités (THE,
T02 et T03), 40 jours après l'observation des
mortalités. Dans les mêmes pots après 55 jours
d'incubation, la réduction des émergences est respectivement de
90%, 54% et 45% pour THE, T02 et T03 (Annexe
3). L'inhibition des émergences persiste mieux au-delà de cette
date pour la formulation huile essentielle mélangée à la
poudre de X. aethiopica (THE).
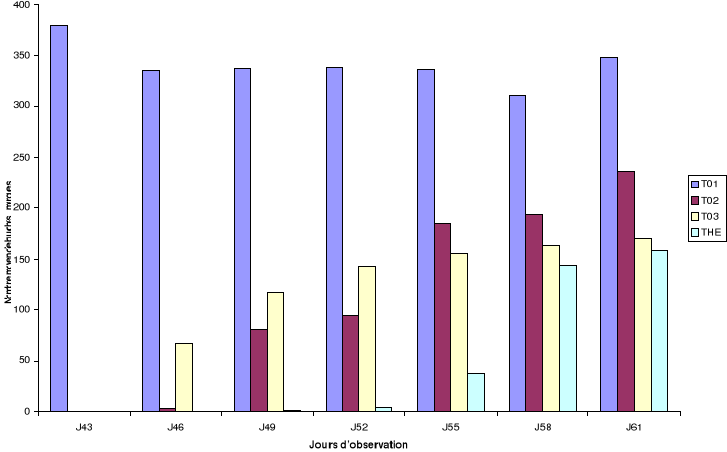
Figure 8:
Emergence des bruches en fonction de la rémanence des formulations
Jn : observation faite n jours après le traitement
des graines par les formulations
4-1-4 Effet rémanent
des formulations sur la variation du poids des graines
L'évolution du poids des graines à partir des
200g de niébé introduits dans chaque pot permet de constater que
les graines traitées présentent les plus faibles diminutions de
poids par rapport au témoin non traité (Figure 9). Les
formulations T02, T03 et THE en limitant les
attaques des bruches ont réduit les pertes post
récoltes lors de nos traitements au laboratoire. Ces résultats
sont corroborés par les pourcentages d'attaque. En effet, deux mois
après le traitement des graines par les différentes formulations,
les pourcentages d'attaque observés sont respectivement pour les
traitements 40%, 57%, 65% et 75% (Tableau 9) pour THE,
T03, T02 et T01. L'efficacité
de la formulation huile essentielle mélangée
à la poudre de X. aethiopica
THE se maintient significativement mieux comparativement
aux autres traitements jusqu'au 55ème jour après le
traitement des graines (Annexe 4). Les poids les plus bas ont été
observés dans le témoin. La diminution du poids en fonction de la
perte en activité notée pour les formulations THE, T03
et T02 serait due aux pontes plus
élevées ; ce qui se traduirait par l'utilisation des
substances de réserve du niébé par les larves.
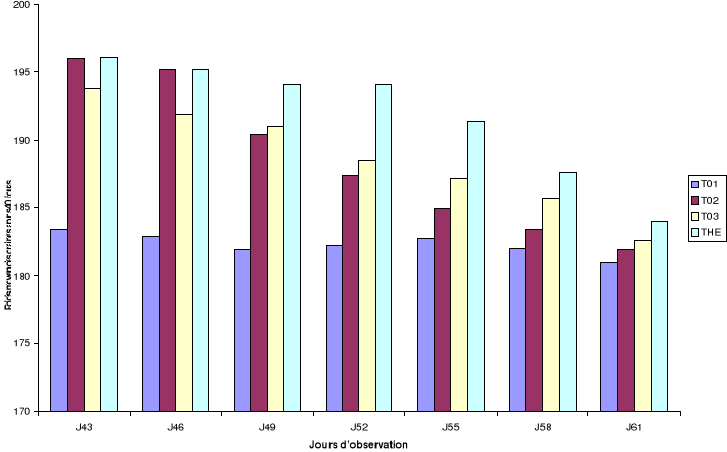
Figure 9: Poids
moyen (g) des graines sur 200g en fonction de la rémanence des
formulations
Jn : observation faite n jours après le traitement
des graines par les formulations
L'absence d'attaque des graines observée dans les
traitements THE et T02 aux jours 43 et 46
(Tableau 8) est le résultat des mortalités et des pontes
enregistrées aux jours 3 et 6. En effet, à ces jours des taux de
mortalité de 100% et une inhibition totale des pontes ont
été constatés au niveau de ces deux traitements.
Tableau 8:
Pourcentage d'attaque des graines par les bruches après 40 jours
|
J43
|
J46
|
J49
|
J52
|
J55
|
J58
|
J61
|
|
T01
|
74a
|
74a
|
71a
|
74a
|
74a
|
73a
|
75a
|
|
T02
|
0b
|
0b
|
34b
|
40b
|
53b
|
60b
|
65b
|
|
T03
|
9c
|
25c
|
41c
|
46c
|
52b
|
57c
|
57c
|
|
THE
|
0b
|
0b
|
1d
|
13d
|
16c
|
25d
|
40d
|
Les nombres suivis de la même lettre dans chaque colonne ne
sont significativement différents (á=0,05 au SNK)
Jn : observation faite n jours après le traitement
des graines par les formulations
4-2 Discussions
Les données relatives à l'efficacité des
poudres des fruits des plantes insecticides sont souvent ambiguës et quant
à leur rémanence ces données sont quasi inexistantes dans
la littérature. Les formulations testées dans notre
expérimentation ont pour intérêt d'être faciles
à mettre en oeuvre en milieu paysan et peu coûteuses.
La persistance de l'efficacité de la formulation pour
le traitement THE est meilleure vis-à-vis de la bruche
du niébé pendant deux semaines au moins ; le support de
dilution de l'huile, à savoir les micro fibres du fruit aurait pour
effet de réduire l'évaporation des matières actives,
d'augmenter la concentration en huile et prolongerait l'efficacité de la
formulation. Le traitement T02 provoque une mortalité
semblable à celle observée pour le traitement
THE durant les six premiers jours. Il s'agit d'un effet knock
down qui s'estompe suite à l'évaporation de la formulation
T02 (huile essentielle diluée dans l'acétone).
La rémanence de cette formulation est donc limitée et les risques
de réinfestation existent suite à la disparition de l'effet
répulsif observé les premiers jours d'incubation ;
d'où les nécessités de trouver des supports de
formulation. Toutefois, son action permet d'éliminer tous les individus
présents ; c'est donc une activité biologique
intéressante à l'égard de C. maculatus dont les
femelles pondent la plus grande partie des oeufs au cours des trois premiers
jours de leur état imaginal (Nwanze et al., 1975 ; Singh
et Jackai, 1985). Nonobstant un taux de mortalité plus faible
provoqué par la formulation T03 (poudre de X.
aethiopica) trois jours après le traitement comparativement aux
traitements T02 et THE, la dégradation
de cette formulation s'avère moins lente que pour le traitement
T02 et après douze jours d'incubation, le
traitement T03 présente un taux de mortalité
plus faible que T02. Les formulations testées
T03 et THE semblent donc manifester à
l'égard des adultes de C. maculatus, une toxicité
beaucoup plus par contact direct que par ingestion ou inhalation. En effet
à ce stade, les adultes ne s'alimentent pas et les boîtes
utilisées n'étaient pas étanches pour retenir les
substances volatiles notamment dans le traitement T02 (huile
essentielle diluée dans l'acétone) qui présente une
rémanence moins bonne que les traitements T03 et
THE. Les formulations à base de poudre peuvent aussi
exercer un effet mécanique sur le mouvement et la ponte des bruches
comme l'ont montré Kéita et al., 2000.
La réduction des pontes observée pour les
traitements THE et T02 pendant la
première semaine d'incubation serait le fait de la mort précoce
des adultes de C. maculatus due aux vapeurs d'huiles
essentielles et le support utilisé, comme l'ont montré Schmidt
et al. (1991), Mazibur et Gerhard (1999) en étudiant
l'effet de l'huile de Acorus calamus sur Callosobruchus
phaseoli.
L'effet résiduel prolongé observé pour la
formulation huile essentielle mélangée à la poudre de
X. aethiopica (THE) serait dû à
l'addition de l'huile essentielle à la poudre. Selon Pierrard (1986)
cité par Kéita et al., (2000), l'utilisation
d'un support de dilution de l'huile essentielle aurait pour effet de prolonger
l'efficacité de la formulation et limiterait aussi l'odeur persistante
qu'on aurait sur le graines lors de la consommation. Cette formulation aurait
une action ovicide, soit en diminuant l'adhésivité des oeufs sur
le tégument des graines ou bien en agissant sur l'embryon après
leur pénétration à travers le chorion. Une inhibition des
pontes et une action ovicide ont été observées chez la
bruche de niébé, une fois que celle-ci est traitée avec
les poudres de feuilles de Nicotiana tabacum (Solanaceae) et de O.
gratissimum (Labiatae) (Ofuya,1990), ou avec la poudre de neem (A.
indica) (Meliaceae) (Seck et al., 1991) mais il n'existe
pas de données sur la rémanence de ces formulations.
CHAPITRE V: CONCLUSION ET RECOMMANDATION
5- Conclusion et
recommandations
Les démarches suivies par la présente
étude avaient pour objet d'examiner l'efficacité et la
rémanence de trois formulations à base de fruits de X.
aethiopica en relation avec la survie des bruches adultes, leur
fécondité et le nombre des descendants de la première
génération.
Nos résultats indiquent qu'une protection durable et
à coût peu élevé contre C.
maculatus est possible en utilisant les formulations à base de
fruits secs de X. aethiopica. L'utilisation de l'huile essentielle de
X. aethiopica seule n'est pas recommandable pour une protection
à longue durée des graines de niébé. D'où la
nécessité de la mise au point d'une formulation dont le support
de dilution pourrait être la poudre des fruits secs de X.
aethiopica comme on a pu le montrer dans notre expérimentation. La
formulation huile essentielle mélangée à la poudre de
X. aethiopica serait une alternative intéressante pour la
protection des stocks du niébé puisque sa rémanence est
d'au moins deux semaines et cette formulation inhibe aussi bien la ponte mais
aussi l'émergence de la première génération. Compte
tenu du fait que X. aethiopica se retrouve dans les aires de
production du niébé, l'utilisation des fruits secs de cette
espèce pour protéger les récoltes de niébé
contre les attaques sévères de C. maculatus peut
représenter une solution alternative supplémentaire pour les
populations locales qui traitent habituellement le niébé avec les
plantes à effet insecticide.
L'effet dose de la formulation THE
pourrait être testé ultérieurement pour
prolonger l'effet résiduel de la formulation. L'effet des
différentes formulations sur les ennemis naturels de C.
maculatus doit être aussi évalué. Une étude
en milieu paysan doit être aussi envisagé en vue de tester les
mêmes paramètres dans les greniers traditionnels. Enfin, d'autres
formulations comme la poudre de kaolin et la cendre de bois enrichies de
l'huile et de la poudre de X. aethiopica sont des pistes
à explorer dans le futur. L'intérêt de la cendre est
qu'elle permet non seulement de combler les interstices mais aussi de
réduire le taux d'humidité des grains, facteur important pour une
bonne conservation des grains.
REFERENCES
BIBLIOGRAPHIES
ADJANOHOUN E. J., AHYI A. M. R., AKE ASSIL.,
BANIAKINA J., CHIBON P., CUSSET G., DOULOU V., ENZANAN A., EYME G., GOUDOTE E.,
KEITA A., MBEMBA C., MOLLET J., MOUTSAMBOTE J. M., MPATI J.& SITA P.,
1988. Contribution aux études ethnobotaniques et floristiques
en République Populaire du Congo. Collection médecine
traditionnelle et pharmacopée. ACCT. Paris. pp.21- 91.
AKOU-EDI, 1983. Effect of neem seed powder
and oil on Tribolium confusum and Sitophilus
zeamais. Proceeding second international Neem conference
Rauischholzhausen. pp. 445 -452.
AKPOVI C. S., 1993. Etude au
laboratoire de l'efficacité de Dinarmus basalis Rondani.
(hymenoptera : Pteromalidae) ectoparasite de Callosobruchus
maculatus Fabricius (Coleoptera : Bruchidae). Mémoire
d'ingénieur Agronome. Univ. Nation. du Bénin. Cotonou
Bénin ; 98p.
ALEBEEK F. A. N. van, ROJAS ROUSSE D., LEVEQUE L.,
1993. Interspecific competition between Eupelmus vuilleti and
Dinarmus basalis, two solidary ectoparasitoids of Bruchidae larvae and
pupae. Entomol. Exp. Appl. 69: pp.21- 31.
ALGHALI A.M., 1993. Intercropping as a
component in insect pest management for grain cowpea (Vigna
unguiculata L. Walp) in the Sudan Savanna of Nigeria. Insect Science
and its Application 14 (1): pp49 -54.
ALZOUMA I., 1981. Observations of the ecology
of Bruchidius atrolineatus Pic. and Callosobruchus maculatus
F. (Coleoptera: Bruchidae) in Niger. In: The Ecology of Bruchids Attacking
Legumes. Edited by Labeyrie, Sunk, The hague: pp.205-213
ALZOUMA I., 1987. Reproduction et
développement de Bruchidius atrolineatus Pic.
(Coleoptera : Bruchidae) aux dépens des cultures de Vigna
unguiculata L. Walp (Leguminosae : Papilionaceae) dans un
agrosystème sahélien au Niger. Thèse de Doctorat d'Etat,
Univ. François-Rabelais, Tours France, 162p.
ALZOUMA I., HUIGNARD J., LENGA A., 1996. Les
coléoptères Bruchidae et autres insectes ravageurs des
légumineuses alimentaires en zone tropicale. In :
Post-récolte : Principes et applications en zone
tropicale. Edité par Verstraeten C., ESTEM/AUPELF. pp. 79-103.
AMOTABI C.I., 1994. Field evaluation of some
insecticides for the control of insect pests of cowpea (Vigna
unguiculata L. Walp) in the Sudan Savanna of Nigeria. Jour. Pest
Management, 40 : pp.13-17.
AMVAM ZOLLO P. H., MENUT C., NGO BAHAGBEGE A. &
ATOGO B.M., 1991. Etude des constituants volatiles du
gingembre (Zingiber officinale Roscoe) du Cameroun. Biociences
Proceeding, 2: pp.280-282.
ARNASON J.T., DURST T., & PHILOGENE B.J.R.,
2002. Prospection d'insecticides phytochimiques des plantes
tempérées et tropicales communes ou rares In :
Biopestides d'origine végétales, ed Tec & doc.
Londres-Paris-New York. pp. 40-75.
BANDARA K. A. N. P., SAXENA R. C., 1995. A
technique for handling and sexing Callosobruchus maculatus (F.) adults
(Coleoptera: Bruchidae). J. stored Prod. Res. Vol.31, No.I, pp.
97-100.
BEAUMONT A., CASSIER P., 1973. Biologie
animale : des protozoaires aux métazoaires
épithélioneuriens. Tome2. Dumod, Université ;
918p.
BEKELE J. & HASSANALI A., 2001. Blend
effects in the toxicity of the essential oils constituent of Ocimum
kilimandssharicum and Ocimum kenyense (Labiateae) on two post
harvest insect pest. Phytochemistry 57: pp. 385-391.
BELL A., MUCK O., MUTULU P. & SCHNEIDER H.,
1998. La protection intégrée des denrées
stockées est une affaire rentable !. GTZ. Eschborn . 37p.
BOEKE S.J., BAUMGARD I. R., VAN HUIS A., DICKE M.
& KOSSOU D.K., 2004. Toxicity and repellence of African plants
traditionally used for the protection of stored cowpea against
Callosobruchus maculatus. Journal of stored products
Reasearch 40: pp. 423-438.
BOTTENBERG H., 1995. Farmers perception of
crop pest control practices I rainfed cowpea cropping systems in Kano,
Nigeria. Inter. J. of Pest Management 41(4): pp.195-200.
BOUDA H., TAPONDJOU L.A., FONTEM D. A., & GUMEDZOE
M.Y.D., 2001. Effect of essential oil from leaves of
Ageratum conyzoïdes, Lantana camara and Chromolaena adorata
on the mortality of Sitophilus zeamais (Coleoptera,
Curculionnidae). Journal of stored products Research. 37: pp.
103-109.
CASWELL G.H., 1961. The infestation of cowpea
in the Western regions of Nigeria. Trop. Sci.3: pp. 154-159.
CREDLAND P.F., 1990. Biotype variations and
host plant change in bruchids: cause and effects in the evolution of bruchids
pests. Bruchids and legumes: Economics Ecology and coevolution. FUJI K. (ed.);
pp. 271- 287.
DECELLE J., 1981. Bruchidae related to grain
legumes in the perspectives. Biologie survey of Canada Monograph Service
N°1: 439p.
DECELLE J., 1987. Les
coléoptères nuisibles aux légumineuses alimentaires
cultivées dans la région afrotropicale. Colloque sur les
légumineuses alimentaires. Univ. de Niamey, 1985. AUPELF ; pp.
188-200.
DELOBEL A., MALONGA P., 1987. Insecticidal
properties of six plant materiels against Caryedon serratus (OI.)
(Coleoptera: Bruchidae). Journal of Stored Products Research 23: pp.
173-176.
DELOBEL A., TRAN M., 1993. Les
coléoptères des denrées alimentaires entreposées
dans les régions chaudes. ORSTOM/CTA. Faune tropicale 32. Paris.
424p.
DESROCHES P. E., EL SHAZLY, MANDON N., DUC G.,
HUIGNARD J., 1995. Development of Callosobruchus chinensis L.
and C. maculatus F. (Coleoptera: Bruchidae) in seeds of Vicia
faba L. differing in their tannin, vicine and convicine contents. J.
stored Prod. Res. 31: pp. 83-89.
DJIDONOU, 2000. Contribution au recensement
des composantes de la biodiversité pour une lutte intégrée
contre les bruches du niébé (Vigna ungunculata L.
Walp) : cas de la zone de transition, Plateau-Vallée de
l'Ouémé au Bénin. Mémoire d'Ingénieur
Agronome FSA/UNB 128p.
DUKE J.A., 1990. Introduction of food
legumes. In Singh S.R.(ed.) Insect pests of food legumes. Chichester,
John Wiley et sons. pp. 1-42.
DUNKEL. FV. & SEARS LJ., 1998. Fumigant
Properties of physical preparation from mountain big sagebrush, Artemisia
tridentate Nut spp Vaseyena (Rydb) Beetle for stored grains insects.
Journal of stored products Research. 34(4): pp. 307-321.
DUPRIEZ H. & DE LEENER P., 1987. Jardins
et vergers d'Afrique. Apica, Enda, CTA, Wageningen. Collection Terres et vies.
380p.
F.A.O., 1998. La situation mondiale de
l'alimentation et de l'agriculture. 371p.
FOX C.W., 1994. The influence of egg size on
offspring performance in the seed beetle, Callosobruchus maculatus.
Oikos 71: pp. 321-325.
GIGA D. P., SMITH R.H., 1983. Comparative
life history studies of four Callosobruchus species infesting cowpea
with special reference of Callosobruchus rhodesianus Pic (Coleoptera:
Bruchidae). J. stored prod. Res. 19: pp. 189- 198.
GILES P. H., 1964. The storage of cereals by
farmers in Northems Nigeria. Trop. Agric. Trinidad, 41(3): 197-212.
GLITHO I.A., NUTO Y., ATTOH A., SAMBENA B., KOUNNOU
K., 1988. Ecologie et biologie de la reproduction des Bruchidae
parasites des légumineuses alimentaires cultivées au Togo et au
Bénin. Rapport ABN (Biosciences), Lomé ; 81p.
GOLOP P., MOSS C., DALES M., FIDGEN A. EVAN J.,
GUDRUPS I., 1999. The use of spices and medicinals as bioactive
protectants for grains. FAO Agricultural Services Bulletin, 137.
GTZ, 1995. Le stockage et la conservation du
niébé. Fiche technique, 11p.
GWINNER J., HARNISH R., MUCH O., 1996. Manuel
sur la manutention et la conservation des grains après récolte.
GTZ. Gmbh, 368p.
HAMRAOUI A. REGNAULT-ROGER C., 1997.
Comparaison des activités insecticides des monoterpènes sur deux
espèces d'insectes ravageurs des cultures Ceratitis capitata et
Rhopalosiphum padi. Acta Bot. Gallica, 144 : pp. 413-417.
HAUBRUGE E., AMICHOT M., 1998. Les
mécanismes responsables de la résistance aux insecticides chez
les insectes et acariens. Biotechnol. Agron. Soc. Enviro.2 (3) :
pp . 161-174.
HOFFMANN A., LABEYRIE V., BALACHOWSKY A.S.,
1962. Les Bruchidae. In Entomologie appliquée à
l'agriculture. A.S.BALACHOWSKY Ed. Masson public 1 : pp.
434-494.
HUIGNARD J., LEROI B., ALZOUMA I., GERMAIN J.F..,
1985. Oviposition and development of Bruchidae atrolineatus
Pic and Callosobruchus maculatus F. (Coleoptera: Bruchidae) in
Vigna unguiculalata L. Walp cultures in Niger. Insect Sci.
Applic. Vol.6, pp. 691-699.
HUIS A. van, 1991. Biological methods of
bruchids control in thevtropics. A review Insect Sci. Appl. Vol.12,
N°1/2/3; pp.87-102.
HUIS A. Van, de ROOY M., 1998. The effect of
leguminous plant species on Callosobruchus maculatus (Coleoptera:
Bruchidae) and its egg parasitoid Uscana lariophaga (Hymenoptera:
Trichogrammatidae). Bull. Ent. Research 88: pp. 93-99.
HUIS A. van, SCHUTTE C., COOLS M.H., FAMGET P., van
der HOER K., PIQUET S.P., 1994. The role of semiochemicals in host
location by Uscana lariophaga, egg parasitoid of Callosobruchus
maculatus. Proceeding 6th International working conference on
stored product protection. April 17-23, 1994. Camberra, Australia, vol.2,
pp.1158-1164.
HUSSEIN M. H., ABDEL-AAL Y.A., 1982. Toxicity
of counpounds against the cowpea seed beetle Callosobruchus maculatus
F. (Coleoptera: Bruchidae). Int. Pest Control 24 pp. 12-16.
IITA, 1982. Le niébé: manuel de
formation, Ibadan (Coll. Series de manuel 11); 127p.
ISMAN M. B., 2000. Plant essential oils for
pest and disease management. Crop protection. 19: pp. 603-608.
ISMAN M. B., 2002. Problèmes et
perspectives de commercialisation des insecticides d'origine botanique
In : Biopesticides d'origine végétales. pp 300-311.
JACKAI L. E. N., 1995a. Legume Pod borer
Maruca vitrata (Fab.) and its principal host plant, Vigna
unguiculata L. Walp. Use of selective insecticide sprays as aid in the
identification of useful levels of resistance. Crop Protection, 14:
(4): pp. 299-306.
JACKAI L.E.N., 1995b. Safe use of insecticide
in agriculture. IITA research guide 15, 40p.
JACKAI L.E.N., ADALLA C.B., 1997. Pest
management of borers of cowpea and beans. Insect Science and its
application, 16: pp. 237-250.
JACKAI L.E.N., DAOUST R.A., 1986. Insect pest
of cowpea. Ann. Review of Ent. 31: pp. 95-119.
JACOBSON M., 1989. Botanical pesticides:
past, present and future. Arnason J.T., Philogene B. J. R., & Morand P.,
eds. Insecticides of plant origin. ACS Symposium Series, 384, pp.
1-10. Washington DC USA.
JIROVETZ L., BUCCHBAUER G. & NGASSOUM M. B.,
1997. Investigation of the essential oils from the dried fruits of
Xylopia aethiopica (West African «Peppertree») and
Xylopia parviflora from Cameroun. Ernäbrung-Nutrition, 21: pp.
324-325.
JIROVETZ L., BUCHBAUER G., NGSSOUM M.B., NGAMO T. L.
& ADJOUDJI O., 2000. Isecticidal activity and chemical composition
of essential oils of Ocimum gratissimum and Xylopia
aethiopica from Cameroun. In: ISEO 31st international symposium
on essential oils. Hamburg/Germany. September 10-13-2000.
KAWECKI T.J. 1995. Adaptative plasticity of
egg size in response to competition in the cowpea weevil, Callosobruchus
maculatus (Coleoptera: Bruchidae). Oecologia 102: pp. 81-85.
KEITA M.S., VINCENT CHARLES., SCHMIT J. P., RAMMASWANY
S., BELANGER A., 2000. Effect of various essential oils on
Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). Journal of
stored products Research. 36: pp. 355-364.
KETOH K. G., 1998. Utilisation des huiles
essentielles de quelques plantes aromatiques du Togo comme biopesticides dans
la gestion des stades de développement de Callosobruchus
maculatus (Coleoptera : Bruchidae). Thèse unique,
Université du Bénin, Lomé, Togo, 136p.
KEZAN Z., HUESING J.E., SHADE R. E., MURDOCK L.L.,
1994. Cowpea trysin inhibitor and resistance to cowpea weevil
(Coleoptera: Bruchidae) in cowpea variety «Tvu 2027». Environ.
Entomol. 23(4): pp. 987-991.
KHOLI K.S., 1990, «Kohinoor» a
nutritinos fodder cowpea for animals. Indian Farming, 39: pp. 15-29.
KIM S.I., ROH J.Y., KIM D.Y., LEE H.S., AHN Y.J.,
2003. Insecticidal activities of aromatic plant extracts and essential
oils against Sitophilus oryzae and Callosobruchus chinensis.
Journal of Stored Products Reasearch. 39: pp. 293-303.
KOSSOU D.K., AHO N., 1993. Stockage et
conservation des grains alimentaires tropicaux: Principes et Pratiques. Les
éditions du Flamboyant, 125p.
KOSSOU K. D., GBEHOUNOU G., AHANCHEDE A., AHOHUENDO
B., YACOUBA B., van HUIS A., 2001. Endogenous cowpea production and
protection practies in Bénin. Insect. Sci. Applic. (21)
2 : pp. 30-30.
KOUNINKI H., HAUBRUGE E., NOUDJOU F. E., LOGNAY G,
MALAISE F., NGASSOUM M. B., GOUDOUM A., MAPONGMETSEM P. M., NGAMO I. S., HANCE
T., 2005. Potentiel uses of essential oils from Cameroon applied as
fumigant or contact insecticides against Sitophilus zeamais Motsch.
(Coleoptera: Curculionidae). Commun Agric Appl Biol Sci. 70(40): pp.
787-792.
KOUNINKI H., 2005. Etude de la
toxicité des huiles essentielles de quelques plantes traditionnellement
utilisées au Nord du Cameroun contre les Bruchidae et les Curculionidae.
Mémoire de fin de DEA, 82p.
KUMAR R., 1984. Insect pest control. Edward
Arnold Ltd, New York ; 298p.
LAMBERT J.D.H., GALE J., ARNASON J.T., PHILOGENE
B.J.R., 1985. Bruchid control with traditionally used insecticidal
plants Hyptis spicigera and Cassia nigricans. Insect Sci.
Applic. 6(2): pp. 167-170.
LAMMERS P.M., van HUIS A., 1989. Uscana
lariophaga Steffan (Hym: Trichogrammidae), egg parasitoid of the stored
insect pests Callosobruchus maculatus F. and Bruchidius
atrolineatus Pic. (Coleoptera: Bruchidae) population studies in the field
and in the stored in Niger. Proc. Conf. Int. Pest Management 3: pp.
1013-1022.
LENTERAN J.C. van, OVERHOLT W.A., 1994.
Ecology and integrated pest management. Insect sci. Appli. Vol. 15,
N°6: pp.557-582.
LENTING M.I., 2000. Preferences and
performances of Callosobruchus maculatus F. (Coleoptera: Bruchidae) on
pods of various plant species towards naturals vegetation manipulation on
cowpea production in Benin. Thesis of Msc n°003. Univ. Wageningen, 31p.
LIENARD V., SECK D., 1994. Revue des
méthodes de lutte contre Callosobruchus maculatus F.
(Coleoptera : Bruchidae), ravageur des graines de niébé
(Vigna unguiculata L. Walp) en Afrique Tropicale. Insect Sci.
Appli. Vol. 15, N°3 : pp.301-311.
LIU Z. L., HO H. S., 1999. Bioactivity of
the essential oil extracted from Evodia rutae carpa Hook F et
Thomas against the grains storage insects Sitophilus zeamais Motsch
and Tribolium castaneum (Herbst). Journal of stored products
Research. 35 (4): pp. 317-328.
MESSINA J.F., 1984. Influence of cowpea pod
maturity on the oviposition choices and larval survival of a bruchid beetle
Callosobruchus maculatus. Ent. Esp. Appli. Vol 35. pp.
241-248.
MAZIBUR H.S., GERHARD J., 1999. Journal
Stored Prod. Res.35 : p.285.
MONGE J.P., DUPONT P., IDI A., HUIGNARD J.,
1995. The consequence of interspeific competition between Dinarmus
basalis Rond and Eupelmus vuilleti Crw (Hymenoptera: Eupelmidae)
on the development of their host populations. Acta Oecologia 16(1): pp.
19-30.
MONGE J.P., OUEDRAOGO A.P., 1990.
Déterminisme du polymorphisme imaginal chez Callosobruchus
maculatus F. (Coleoptera : Bruchidae). Dourdan, France. Ed. INRA,
1990. Les colloques de l'INRA N°52.
MONGE J.P., OUEDRAOGO A.P., HUIGNARD J.,
1991. Development of two Bruchid species : Bruchidius
atrolineatus (Pic) and Callosobruchus maculatus (F.) and their
larval parasitoïdes during storage of cowpea seeds Vigna
unguiculata (Walp) in West Africa. In F. Fleurat Lessard et P. Ducon
(eds), Proc. 5th, Int. Working Conf. Stored Product Prot. Bordeaux, Sept. 9-14,
1990, vol.1: pp. 151-160.
MORTIMORE M.J., SINGH B.B., BLADE S.F., 1997.
Cowpea in traditional cropping systems. Advances in cowpea research; IITA.
JIRCAS. pp.99-112.
MULONGOY M., GUEYE K., SPENCER D.S.C., 1992.
Biological nitrogen fixation and sustainability of tropical agriculture, IITA,
Ibadan, Nigeria. 69p.
MURDOCK L.L., SHADE R.E., KITCH L.W., NTOURKAM G.,
LOWENBERG-DEBOER J., HUESING J.E., MOAR W., CHAMBLIS O.L., ENDONDO L.,
WOLFSON J.L., 1997. Post-harvest storage of cowpea
research. Copublication of IITA and JIRCAS: pp. 303-312.
NGAMO L.S.T., NGASSOUM M.B., JIROVETZ L., ADJOUDJI O.,
NUKENINE E.C. MOUKALA O.E., 2001. Protection of stored maize against
Sitophilus zeamais (Most) by the use of essential oils of species from
Cameroun. Med. Fac. Landbouww. Univ. Gent, 66/2a., pp. 473-478.
NGAMO L.S.T., 2004. Rapport technique
(Septembre 2003-Février 2004), Projet interuniversitaire ciblé
(PIC), 34p.
NGASSOUM M.B., JIROVETZ L., FLEISCHHACKER W.,
BUCHBAUER G., YONKEU S., 1999. Investigation of aromatic plants from
Cameroon : GC/MS analysis of essential oils of Plectranthuus
glandulosus (Lamiacea). 30TH International Symposium on
Essential oils (30TH ISEO). Leipzig (Germany), September 5-8: pp.
1999.
NWANZE K.F., HORBER E., PITTS W., 1975.
Evidence for ovipositional preference of Callosobruchus maculatus for
four varieties of cowpea. Environn. Ent.4: pp. 409-412.
ODAH B.K., 1995. Impact du
développement de Callosobruchus maculatus F. (Coleoptera :
Bruchidae) sur les graines de trois variétés de Vigna
unguiculata L. Walp. Thèse de D.E.A., Univ. du Bénin,
Lomé-Togo; 67p.
OFUYA T.I., 1990. Oviposition deterrence and
ovicidal proporties of some plants powders against Callosobruchus
maculatus in stored cowpea (Vigna unguiculata) seeds. J. Pest
Manag. 43 : pp. 89-94.
OUEDRAOGO P.A., 1991. Le déterminisme
du polymorphisme imaginal chez Callosobruchus maculatus F.
(Coleoptera : Bruchidae), son importance sur la biologie de cette bruche.
Thèse Doctorat. Univ. Tours (France); 197p.
OUEDRAOGO P.A., MONGE J.P., HUIGNARD J.,
1990. Importance of temperature and seed water content on the
induction of imaginal polymorphism in Callosobruchus maculatus F.
(Coleoptera: Bruchidae). Ent. Exp. Appl. 59: p.66.
OUEDRAOGO P.A., SOU S., SANON A., MONGE J.P., HUIGNARD
J., TRAN B., CREDLANG P.F., 1996. Influence of temperature and
temperature and humidity on population of Callosobruchus maculatus
(Coleoptera: Bruchidae) and its parasitoid Dinarmus basalis
(Pteromalidae) in two climatic zones of Burkina Faso. Bull. Ent.
Research 86, pp.695-702.
PANDEY R.K., WESTPHAL E., 1989. Vigna
unguiculata L. Walp. In: L.J.G.V.D. Masson and Sadikin Someantmadji
(eds.), PROSEA, N°1, Pulses. Pudoc, Wageningen, pp. 77-81.
PRATES H.T., SANTOS J.P., WAQUIL J.M., OLIVEIRA AB.
& FOSTER J.E., 1998. Insecticidal activity of monoterpenes against
Rhyzopertha dominica (F) and Tribolium castaneum (Herbst).
Journal of stored products research. 34(4): pp. 243-249.
PREVETT P.F., 1961. Field infestation of
cowpea (Vigna unguiculata) pod by beetles of the families Bruchidae
and Curculionidae in northern Nigeria. Bull. Ent. Research. Vol.52
pp.635-645.
PRUTHI J.S., 1980. Spices and condiments:
chemistry, microbiology and technology. Adv. Food Res. Suppl. 4,
Academic Press, New-York (USA). 449p.
QUIN F.M., 1997. Introduction. Advances in
cowpea research. Sayce Publishing, Devon, UK. pp.9-15.
RACHIE K.O., 1985 Introduction. Singh S. R.
et Rachie K.O.(eds) cowpea research production and utilization. John wiley and
sons, Chichester, London. 6p.
RACHIE K.O., ROBERT L.M., 1974. Grain legumes
of lowland tropics. Advances in Agronomy 26: pp. 44-50.
REES P.D., 1996. Coleoptera. In: integrated
Management of insect in stored Products. pp.1-39.
REGNAULT-ROGER C. 2002. De nouveaux
phytoinsecticides pour le troisième millénaire ? In :
Biopesticides d'origine végétales, ed Tec & doc.
Londres-Paris-New York . pp. 19-39.
SECK D., SIDIBE B., HAUBRUGE E., HEMPTIME J.L., GASPAR
H., 1991b. La protection chimique des stocks de niébé et
de maïs contre les insectes au Sénégal. Meded Fac
Landbouwet. Rijikuniv. Gent. 56/3b : pp.1225-1233.
SCHMIDT G.H., RISHA E.M., NAHAL A.K.M., 1991.
Reduction of progeny of some stored-products Coleoptera by vapours of
Acorus calamus oil. J. Stored Prod. Res. 38: pp. 129-138.
SINGH B.B., SINGH S.R., 1992.
Sélection du niébé résistant aux bruches. La
recherche à l'IITA, N°5 Septembre 1992. pp. 1-5.
SINGH S.R., JACKAI L.E.N., 1985. Insect pests
of cowpea in Africa : their life cycle, economic importance, potential for
control. Cowpea, research, production and utilization. John Wiley and Sons,
Chichester, London pp. 217-231.
SINGH S.R., JACKAI L.E.N., dos SANTOS J.H.R., ADALLA
CB., 1990. Insect pests cowpeas. Insects pests of tropical foods
legumes, Chichester, John Wiley and sons, pp.43-90.
SOUTHGATE B.J., 1979. Biology of the
Bruchidae. Ann. Rev. Entomology 24, pp. 449-473.
STOLL G., 1988. Protection naturelle des
végétaux en zones tropicales. CTA/AGRECOL. République
Fédérale d'Allemagne. 180p.
STOLL G., 2000. Natural crop protection in
the tropics. CTA/AGRECOL. 376 p.
SZAFRANSKI E., BLOZDZ B., DROZDZ B., 1991.
Activité biologique des extraits de quelques plantes des environs de
Kisangani (Zaïre). Belg. Journal bot. 124(1): pp. 60-70.
TAPONJOU L., ADLER C., BOUDA H., & FONTEM D. A.,
2002. Efficacy of powder and essential oil from Chenopodium
ambrosioides leaves as post-harvest grain protectants against six-stored
product beetles. Journal of Stored Products Research, 38: pp.
395-40.
TARAWALI S.A., SINGH B.B., PETERS M., BLADE S.F.,
1997. Cowpea haulms as fooder; In Advances in cowpea research. Sayce
Publishing, Devon, UK. pp.313-325.
TAYLOR T.A., 1981. Distribution, Ecology and
importance of Bruchids attacking grain legumes. Ed. by V. Laberyrie; Junk, The
Hague pp.199-203.
TCHAMOU K.S., 1995. Impact des
hyménoptères parasitoïdes: Dinarmus basilis R. et
Eupelmus vuilleti Craw ; sur les populations de
Callosobruchus maculatus F. dans un système de stockage de
niébé : Vigna unguiculata L. Walp. Mémoire
de D.E.A. de Biologie de Développement. Univ. du Bénin.
Lomé-Togo ; 54p.
TCHASSANTI M.T.A., 1995. Dynamique des
populations de trois Coléoptères Bruchidae et de leurs
parasitoïdes dans les cultures de niébé : Vigna
unguiculata Walp. Mémoire de D.E.A. de Biologie de
Développement. Univ. Lomé-togo ; 45p.
TCHOKOSSI E.Y., 1995. Conditions d'induction
de la quiescence reproductrice chez Callosobruchus maculatus F. la
bruche du niébé (Vigna unguiculata Walp) dans un
système de stockage en zone Guinéenne. Mémoire de D.E.A.
de Biologie de Développement ; Univ. du Bénin,
Lomé-Togo ; 44p.
TCHOUMBOUGNANG F., 1997. Contribution
à la détermination des teneurs, des caractéristiques
chimiques et des activités antifongiques des huiles essentielles de
quelques plantes aromatiques, condimentaires et médecinales du Cameroun.
Thèse de Doctorat 3ème Cycle en Biochimie.
Faculté des Sciences, Université de Yahoundé I (Cameroun),
270p.
THOMAS-ODJO A., GASPAR A., 1994. Utilisation
de Dinarmus basalis Rond. (Hymenoptera : Pteromalidae) pour le
contrôle de Callosobruchus maculatus Fab. (Coleoptera :
Bruchidae). Mended. Fac. Landbouw. Univ. Gent. 59/2a: pp. 451-456.
TULEY DE SYLVA K., 1995. United Nations
Industrial development Organisation, Vienna. 232p.
TUNÇ I., BERGER B.M., ERLER F. & DAGLI F.,
2000. Ovicidal activity of essential oils from five plants against two
stored product insects. Journal of stored product Research. 36: pp.
161-168.
UTIDA S., 1954. Phase dimorphism observed in
the laboratory population of the cowpea weevil Callosobruchus
quadrimaculatus. Oyo-Dobuts-Zasshi, 18: pp. 161-168.
UTIDA S., 1981. Polymorphism and phase
dimorphism in Callosobruchus maculatus; In: The ecology of
Bruchids attacking legumes. Ed. by Labeyrie, Junk, The Hague,
pp.143-47.
ZANNOU T.E., 1995. Observations des pontes
des bruches et de leurs hyménoptères parasitoïdes oophages
dans un système de culture de niébé : Vigna
unguiculata L. Walp. au Sud Bénin. Mémoire de D.E.A. de
Biologie de Développement ; Univ. du Bénin ;
Lomé-Togo. 39p.
|
|


