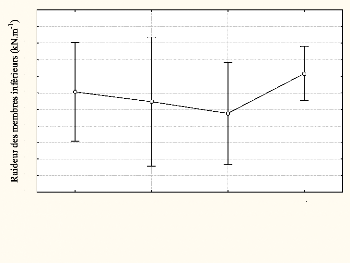
4.6. La raideur des membres inférieurs
(kleg)
La raideur des membres inférieurs a été
évaluée à 15.86 #177;2.76 kN.m-1 en moyenne et
les valeurs sont comprises entre 11.06 et 19.84 kN.m-1. Elle
évolue de 16.05 #177;2.59 kN.m-1 lors du première 100m
à 17,16 #177;1.42 kN.m-1 lors du dernier sprint. Cependant,
la différence n'est pas significative entre les 4
répétitions [F (3,21) <1]. La raideur des membres
inférieurs n'est pas affectée par la fatigue au cours du 4×1
00m.
21
20
19
18
17
16
15
14
13
12
11
10
Figure n°14 : Evolution de la raideur des membres
inférieurs au cours de la
répétition (4 x 100m)
1 2 3 4
Séries de 100m
4.7. La raideur verticale (kvert)
La raideur verticale a été évaluée
à 69.53 #177;6.35kN.m-1 en moyenne avec des valeurs
rangées entre 57.21 et 77.95 kN.m-1. Elle évolue de
72.57 #177;5.65 kN.m-1 lors du premier 100m à 69.5 #177;2.85
kN.m-1 lors du dernier sprint, mais la différence n'est pas
significative entre les 4 répétitions [F (3,21) <1].
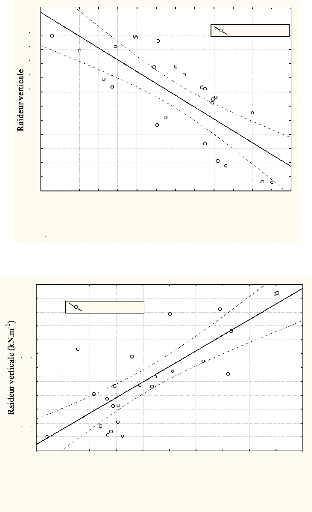
La raideur verticale varie dans le même sens que le
temps de contact [r = 0.77; p<0.05] et l'abaissement maximal des membres
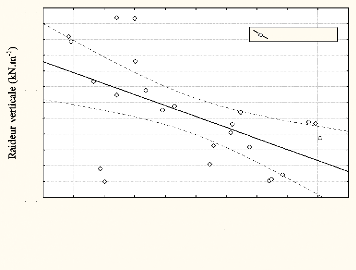
inférieurs [r = 0.8; p<0.05], au cours du 100m. Elle est inversement
corrélée [r = -0.62; p<0.05] avec la variation de la force
maximale.
82
(r = -,76)
80
95% de confiance
78
76
74
72
70
68
66
64
62
60
58
56
112 114 116 118 120 122 124 126 128 130 132 134 136 138
Figure n°15: Relation de la raideur verticale avec le temps
de contact
Temps de contact (ms)
(56)
(58)
95% de confiance
(60)
(62)
(64)
(66)
(68)
(70)
(72)
(74)
(76)
(78)
(80)
0,13 0,14 0,15 0,16 0,17 0,18 0,19 0,20 0,21 0,22 0,23
Abaissement du centre de masse (m)
Figure n°16: Relation entre l'abaissement du centre de
masse et la raideur
verticale
(56)
(58)
(60)
(62)
(64)
(66)
(68)
(70)
(72)
(74)
(76)
(78)
(80)
22 23 24 25 26 27 28 29 30 31 32
Figure n°17: Relation entre la raideur verticale et la
force maximale
aucours d'un 100m sprint
Force maximale (kN)
95% de confiance
4.8. Les concentrations sanguines en lactate ([La])
Les concentrations de sang en lactates augmente
significativement de 8.71 #177; 1.56 au premier sprint à 14.76 #177;
2.08 mmol/l à la quatrième répétition: [F (3, 21) =
35.37; p<0.05] (Voir Figure n°7). Le test post-hoc LSD de Fisher montre
une différence significative entre les 4 répétitions de
100m. Les concentrations sanguines en lactate augmentent avec la
répétition de 100m à vitesse maximale.

5. Discussion
Les valeurs moyennes du temps de contact (125 ms), du temps
d'envol (170 ms), et de l'abaissement du centre de masse (0.17 m) sont
supérieures à celles de la littérature. Par exemple, Morin
et al. (2006) ont constaté, chez des étudiants en
éducation physique et sportive, un temps de contact qui varie avec la
répétition (120.5 ms) et un temps d'envol moyen égal
à 144.5 ms qui reste inchangé. Dans les 2 études,
l'abaissement du centre de masse ne fluctue pas significativement au cours de
la répétition. La fatigue n'affecte pas l'abaissement du centre
de masse.
La différence de durée des temps de contact et
d'envol peut s'expliquer par le port de chaussure d'athlétisme chez les
sujets de notre population ce qui n'est pas le cas chez les novices qui ont
participé à l'étude de Morin et al. (2006). Le temps de
contact est plus élevé chez les experts. Selon Bishop et al.
(2006), le port de chaussures à pointes augmente la flexion dorsale de
la cheville lors de l'atterrissage. Cela peut expliquer partiellement
l'élévation des valeurs du temps de contact au sol chez les
experts sans pour autant expliquer la différence des valeurs en fonction
de l'expertise.
L'amplitude de la foulée, qui est égale à
2.08 m en moyenne, semble normale par rapport aux résultats de la
littérature. Mero et al. (1992) l'ont moyenné entre 2 et 2.6m. En
comparant nos résultats à cette étude, nous remarquons que
la fréquence de la foulée (3.9 Hz) était inférieure
à 5Hz, sachant que cette valeur correspondait à une valeur
enregistrée chez des sprinters experts. Les fréquences de la
foulée de notre population ont diminué de 9 % entre la
première et la dernière répétition. Ces
résultats ressemblent à ceux de Morin et al. (2006) chez une
population de novices, i.e. une diminution de 8.03 % dans des conditions
identiques à celle de notre protocole expérimental.
L'entraînement ne permet pas de diminuer l'effet de la fatigue sur la
fréquence de la foulée.
Les valeurs de la force maximale du premier sprint (2.76 kN)
sont supérieures à celles de Mero et al. (1992) chez des experts
en condition de course à vitesse maximale (1.76 kN). Ces chercheurs ont
démontré que la force maximale augmentait dans le même sens
que la vitesse de course (9.96 m.s-1). La différence des
valeurs de la force
maximale peut donc être expliquée par les
différences de performance entre les deux populations.
Les valeurs moyennes de la force maximale (2.68 kN) sont
similaires aux résultats de Morin et al. (2006) chez des novices et dans
un protocole identique à celui de notre expérimentation. A notre
connaissance, aucune étude n'a démontré, à ce jour,
que la répétition de 100m à vitesse maximale affecte la
force des membres inférieurs.
Le temps au 100m a été calculé à
12.43 s, c'est à dire que nos sujets ont couru en moyenne à une
vitesse égale à 8.06 m.s-1 sur l'ensemble des 4
répétitions. La performance du premier 100m (12.43 s) est
nettement inférieure au record du monde du 100m qui est égale
à 9.69 s. La performance de notre population est éloignée
de celle énoncée au début (11.54 s). Cela peut être
expliqué par la différence de motivation entre une
compétition officielle et une expérimentation scientifique. Le
protocole (4 x 100m) est compris par les athlètes comme étant un
protocole de fatigue. Cela n'éloigne pas l'hypothèse que les
sprinters répartissent leurs énergies, et ne courent pas chaque
sprint avec la vitesse maximale possible, comme l'indique notre consigne
orale.
La vitesse de course a diminué de 8.18 m.s-1
au premier 100m à 7.89 m.s-1 au quatrième sprint,
c'est-à-dire de 8.36 %. Morin et al. (2006) ont constaté une
diminution de 11.6 % dans des conditions similaires chez des novices, avec une
vitesse moyenne qui diminue de 8.10 m.s-1 au premier sprint à
7.10 m.s-1 à la quatrième répétition. La
baisse de la performance avec la fatigue due à la
répétition est plus élevée chez les novices (3.5
%). L'expertise permet de diminuer les effets de la fatigue sur la performance
au 100m.
La raideur des membres inférieurs a
évolué de 16.05 #177;2.59 kN.m-1 lors de la
première répétition de 100m à 17,16 #177;1.42
kN.m-1 au cours de la dernière répétition. Elle
diminue de 19.5 kN.m-1 à 18.3 kN.m-1 chez des
novices dans des conditions similaires [23]. La fatigue n'affecte la raideur
des membres inférieurs, ni chez experts, ni chez les novices. Est-ce une
stratégie du système nerveux central pour contrôler les
différents paramètres mécaniques ? La raideur des membres
inférieurs est-elle une
variable macroscopique commandée par le SNC pour conserver
et optimiser les paramètres de la foulée au cours de la course
?
La raideur verticale diminue de 72.57 à 69.5
kN.m-1 entre le premier et le dernier sprint. Ces valeurs sont
supérieures à celles des novices [23] dont la raideur verticale
diminue significativement de 93.9 à 74.9 kN.m-1. Morin et al.
(2006) ont constaté que la diminution de la raideur verticale est la
conséquence de la diminution de la vitesse. Cela n'est pas le cas chez
les experts dont la raideur verticale ne varie pas avec la
répétition. La fatigue n'affecte pas la raideur verticale chez
les sprinters confirmés.
Au cours du 100m, la raideur verticale varie dans le
même sens que le temps de contact et l'abaissement maximal des membres
inférieurs. La raideur verticale permet de soutenir le temps de contact
et l'abaissement du centre de masse au cours d'un 100m sprint. Cela permet au
système musculotendineux de restituer de l'énergie
élastique au cours d'un temps court, ce qui explique
l'insensibilité de l'abaissement du centre de masse à la
décélération de fin de course. La raideur verticale permet
d'optimiser la foulée lors d'un 100m en réduisant le temps de
contact et la fluctuation verticale du centre de masse.
Les taux de lactate enregistrés sont similaires
à ceux de Bret et al. (2001) qui ont démontré une
corrélation entre la performance (11.7s) et les concentrations de
lactate sanguin (12.5 mmol.l-1) lors de compétition
officielle de 100m, ce qui n'est pas le cas pour Hautier et al. (1994) qui ont
constaté des taux de lactates inférieurs aux valeurs
enregistrées par Bret et al. (2001) (8.5 mmol.l-1) et
indépendants des performances au 100m (valeurs de lactates
enregistrées lors d'une compétition officielle). De même,
Nummela et al. (1992) ont démontré que les concentrations de sang
en lactates n'étaient corrélées qu'avec la performance
finale au 400m et qu'elles étaient indépendantes de celles des
distances inférieures, i.e. le 300, le 200 et le 100m. Les
filières énergétiques sollicitées lors d'une
série de 4 répétitions de 100m (anaérobie
alactique) ne sont pas les mêmes que celles sollicitées lors d'une
épreuve 400m (anaérobie lactique), donc, il n'est pas possible de
comparer nos résultats à ceux de Nummela et al. (1992).
| 

