Contribution a la vaccination des volailles contre la maladie de gumboro a l'aide de vaccins inactives et vivants disponibles sur le marche de dakar( Télécharger le fichier original )par Fabrice Juliot MOUGANG EISMV / Université Cheikh Anta Diop - Doctorat en médecine vétérinaire 2008 |

Première PartieDonnées bibliographiques sur l'aviculture dans la région de Dakar ET la maladie de Gumboro. CHAPITRE 1 : GENERALITES SUR L'ELEVAGE AVICOLE DANS LA REGION DE DAKARCe chapitre est consacré à la présentation de la région de Dakar, puis à la situation actuelle de l'élevage avicole et les problèmes auxquels il est confronté. 1.1. PRESENTATION DE LA REGION DE DAKAR Pays de l'Afrique de l'Ouest, la république du Sénégal ouverte sur l'océan Atlantique est un carrefour de grandes routes maritimes et aériennes. Le Sénégal est compris entre les 12° et 16°30 latitude Nord et entre les 11°30 et 17°30 longitude Ouest (INSTITUT GEOGRAPHIQUE NATIONAL, 1977). Il couvre une superficie de 197 161 km² et est limité au Nord par la Mauritanie, à l'Est par le Mali, au Sud-est par la Guinée et au Sud par la Guinée-Bissau. La Gambie constitue une enclave tout en longueur dans le sud du Sénégal, à l'intérieur duquel elle pénètre profondément.
Figure 1 : Carte du Sénégal (source : - www.au-senegal.com/-Geographie-.htlm ) Située à l'extrême Ouest du Sénégal et du continent africain, la région de Dakar se présente comme une presqu'île de 550 km², représentant ainsi seulement 0,28% de la superficie nationale. Elle est contiguë à l'Est par la région de Thiès et entourée par l'océan Atlantique sur ses limites Nord, Ouest et Sud. La région de Dakar est divisée en quatre départements : Ø département de Dakar Ø département de Guediawaye Ø département de Pikine Ø département de Rufisque Cette situation géographique fait que cette région présente des caractéristiques climatiques particulières, notamment un microclimat de type côtier. Le relief sénégalais, presque plat est constitué de vastes plaines avec une côte basse et sablonneuse, rocheuse par endroit. Les altitudes sont partout inférieures à 150 m sauf au Sud-est dans la région de Tambacouda (JEUNE AFRIQUE, 2000). La région de Dakar est représentée par une bande côtière à dépression interdunaire humide communément appelée zone des « Niayes ». Elle s'étend de Dakar à Saint-Louis et couvre une superficie d'environ 183 km². Cette région présente un microclimat propice à l'élevage en général et à l'aviculture en particulier. Les grands traits climatiques découlent de l'influence entre de nombreux facteurs géographiques. Au Sénégal, le climat est de type sahélo-soudanien dans son ensemble. Il existe des spécificités propres à chaque région. La région de Dakar, de part sa position par rapport à la mer présente une évolution climatique différente de celles des autres régions du pays. La connaissance des vents dominants d'une région ou d'une localité est d'une importance capitale en aviculture. En effet, en plus de son impact sur la ventilation, le vent peut jouer un rôle dans le transport des agents pathogènes et des substances néfastes au confort des volailles. La région de Dakar est exposée à trois types de courants d'air aux caractéristiques thermiques, hygrométriques et directionnelles différentes. D'après JEUNE AFRIQUE, 2000, Ces dernières sont représentées par : · L'Alizé maritime : issu des archipels des Açores, c'est un vent humide et frais qui balaie les régions côtières en apportant un climat relativement doux. Il souffle du Nord vers le Nord-Ouest pendant les mois de Novembre à Mai, mais n'apporte pas de précipitations. · L'Alizé continental ou Harmattan : c'est un vent irrégulier particulièrement chaud et sec qui souffle de l'Est vers le Nord-Est pendant une période assez longue de l'année, allant du mois de mars jusqu'au début de la saison des pluies. Ce vent transporte la poussière et du sable, qui jouent un rôle dans la dissémination de certaines maladies respiratoires, surtout chez les volailles. · La Mousson : elle prend naissance au sud de l'équateur au niveau de l'anticyclone de Sainte-Hélène. c'est un vent très humide et chaud qui apporte la pluie du Sud-Ouest de juin à novembre. L'alternance de ces trois types de vents dont les déplacements sont facilités par la platitude du relief, favorise la saisonnalité du climat. La pluviométrie est caractérisée par deux types de saisons : · La saison sèche ou non pluvieuse, qui n'est sèche qu'à l'intérieur du pays ; la côte bénéficiant d'une humidité relativement élevée du fait de l'influence de la mer. · La saison pluvieuse, chaude et humide coïncide avec l'arrivée de la mousson qui envahit progressivement le pays. Les précipitations s'installent du Sud vers le Nord. Malgré sa position par rapport à la mer, la région de Dakar reçoit généralement une faible quantité d'eau. Comme le soulignent FARUQUI et coll. 2006, 450mm d'eau ont été enregistrés en 2002 ; les plus grandes quantités l'ont été au mois de septembre. La température est l'un des paramètres très importants en élevage avicole, car elle influence la prise alimentaire et constitue une source de stress chez les volailles. La région de Dakar, par sa situation est la région la plus fraîche du pays et par conséquent, la plus propice à l'aviculture (ITAVI, 1996). Elle dépasse rarement 30°C.
L'hygrométrie est la quantité d'eau ou de vapeur d'eau contenue dans l'air ambiant. Elle représente un facteur important dans l'implantation d'un élevage avicole à cause de ses effets directs ou indirects sur les oiseaux. Le degré d'hygrométrie détermine en partie la quantité d'eau consommée par les oiseaux. La région de Dakar connaît une humidité constante qui se manifeste même en saison sèche par des condensations nocturnes fréquentes. En 2007, la population du Sénégal était estimée à 12,5 millions d'habitants, soit une densité moyenne de 65 habitants au km2 (SENEGAL - MSN Encarta, 2008). Elle est inégalement repartie sur le territoire et compte une vingtaine d'ethnies. La région de Dakar abrite les 25% de cette population avec un taux de croissance de 3.69 % de 1998 à 2001.le taux d'urbanisation est de 43 % en 1999 et la densité est de 4231 habitants au km² (SENEGAL/MEF/DPS, 2001) La zone des NIAYES comprend plus de 65% de la population sénégalaise d'après les statistiques de la Direction de la Prévision et de la Statistique (DPS) cités par AHAMET (2004). Ce facteur démographique associé aux conditions climatiques favorables, fait de la région de Dakar une place de choix pour le développement de l'aviculture moderne (HABAMENSHI, 1994). Ceci s'explique par l'installation d'un grand nombre de fermes avicoles modernes dans la région de Dakar. 1.2. L'ELEVAGE AVICOLE DANS LA REGION DE DAKAR L'élevage avicole au Sénégal contribue pour 30 % au Produit Intérieur Brut (PIB) du secteur primaire et 6 % au Produit Intérieur Brut global (SENEGAL/MAE, 2001). L'aviculture est pratiquée selon un mode traditionnel ou un mode moderne. L'aviculture traditionnelle est un type d'élevage pratiqué essentiellement en milieu rural sous un mode extensif où chaque famille paysanne possède un effectif relativement faible de poules (RAVELSON, 1990). La volaille est élevée en liberté autour des concessions et des techniques employées sont rudimentaires. Il n'y a pas de spécialisation de la production. Ce système traditionnel exploite les races locales et se caractérise par un apport minime voire nul d'intrants (aliments, médicaments) et une faible productivité : une poule locale produit en moyenne 40 à 50 oeufs par an et pèse environ 1,2 kg à 26 semaines d'âge ; un coq de même age pèse 1,4 kg (BULDGEN et coll., 1996). Ces productions sont pour l'essentiel destinées à l'autoconsommation ; les ventes sont occasionnelles. Au Sénégal les races locales sont estimées à 19.542.683 têtes. Cet effectif a progressé de 3,5 % entre 2000 et 2001 (SENEGAL/MAE, 2001). La couverture sanitaire moderne en aviculture traditionnelle est presque inexistante, mais lorsqu'une maladie apparaît, les soins se résument en l'administration des produits de la pharmacopée traditionnelle. C'est ainsi que les extraits de piment ou de feuilles et d'écorces d'Azadirachta indica dilués dans l'eau de boisson sont utilisés comme vermifuges (BULGEN et al, 1992). Secteur en expansion, l'élevage moderne est développé autour des centres urbains en raison de l'existence d'un grand marché de consommation. Il exige un niveau d'investissement et une organisation des différents intervenants. Dans ce système moderne, on distingue deux types d'élevages: élevage industriel et élevage semi industriel ou amélioré. L'élevage industriel se définit d'après LISSOT cité par KOE (2001) comme un établissement qui possède des effectifs importants, qui utilise des poussins d'un jour provenant des multiplicateurs des souches sélectionnées, qui nourrit les volailles avec des aliments complets ou des aliments supplémentés et qui pratique des mesures de lutte (prophylaxie, traitement). Ce type d'élevage utilise des équipements modernes et des techniques perfectionnées en ce qui concerne les différentes opérations. En tenant compte de cette définition, plusieurs auteurs s'accordent sur le fait qu'il existe peu d'élevages de ce type dans la région de Dakar. Toutefois, l'élevage industriel est à ses débuts avec l'exemple de la Société de Distribution du Matériel Avicole (SEDIMA). L'élevage moderne pratiqué dans la région de Dakar reste du type semi industriel (GUEYE, 1999). Il utilise des poussins d'un jour importés d'Europe ou produits au Sénégal par des couvoirs de la Société de Distribution du Matériel Avicole (SEDIMA), la Compagnie Africaine de Maraîchage d'Aviculture et d'Arboriculture Fruitière (CAMAF) et le Complexe Avicole de Mbao (CAM) entre autres. La plus grande productivité de l'élevage semi-industriel par rapport à l'élevage traditionnel justifie notre intérêt pour le secteur moderne sur lequel portera la suite de notre travail
L'effectif de l'élevage avicole moderne est passé de 4 955 651 unités à 8 568 527 unités entre 1997 et 2006. En 2004, l'élevage avicole dit semi-industriel est composé de 1 289 788 poussins ponte et de 3 994 879 poussins de chair. Ainsi 96 % des poussins retrouvés dans la filière avicole sénégalaise sont issus de la production locale, et les 4% restants proviennent de l'importation (SENEGAL.ME.CNA, 2007). L'annexe I présente les détails de ces effectifs. En 2006, il n'y a pas eu d'importations de poussins chair ou ponte. La production locale représente 8 568 527 sujets par rapport à l'année 2005 qui était de 6 752 167. Le nombre total de poussins mis en élevage a subi une hausse en valeur absolue de 1 816 360 sujets par rapport à 2005, soit 27 % en valeur relative. 1.2.1.2.2. Caractéristiques de l'aviculture moderne L'aviculture moderne utilise des races ou souches améliorées (annexe II), ces dernières reçoivent un aliment de qualité et en quantité précise. En outre, elles bénéficient d'une protection sanitaire et leurs logements sont aussi bien contrôlés. En 2001 l'effectif total de volaille (race locale et améliorée) est passé à 25.658.000 sujets contre 24.495.000 sujets en 2000 soit une hausse de 4,7 %. L'élevage traditionnel est estimé à 19.542.683 sujets et l'élevage moderne à 6.115.317 sujets en 2001 (SENEGAL.MAE, 2001). L'évolution des effectifs de volailles au Sénégal est caractérisée par une croissance progressive de la production locale (SENEGAL.ME.CNA, 2006). 1.2.1.2.3. Origine des poussins Les volailles mises en élevage en 2006 ont toute une origine sénégalaise (tableau I). La part de la production nationale de poussins nés au Sénégal (poussins nés des oeufs à couvés importés et poussins 100% sénégalais) a connu une hausse par rapport à l'année 2005, avec un taux de 100% en 2006 contre 97,4% en 2005 à cause de l'arrêt des importations de poussins d'un jour. La part des poussins 100% sénégalais a connu un hausse par rapport à l'année précédente 2005soit une valeur absolue de 366 021. Tableau I : Origine des poussins mis en élevage au Sénégal en 2006
SOURCE : SENEGAL.ME.CNA, 2007 1.2.1.2.4. Différents types de production L'aviculture moderne connaît trois types de spéculations à savoir : · la spéculation « chair » avec des élevages qui ne produisent que des poulets de chair ; · la spéculation « ponte » avec des élevages qui ne produisent que des pondeuses ; · la spéculation « mixte », qui est l'association des deux spéculations précédentes. Actuellement, l'élevage des reproducteurs de souches améliorées s'est ajouté à ces trois opérations précédentes. 1.2.1.2.4.1. Production de viande de volaille La production nationale de viande de volailles industrielles, estimée à partir des effectifs de souches améliorées de poussins « chair » mis en élevage en 2004, 2005 et 2006 et des pondeuses reformées est résumée dans l'annexe III. A ces effectifs, on applique les paramètres zootechniques qui sont : le taux de mortalité et le poids moyen à l'abattage (HABYARIMANA, 1998). La production nationale de la viande de volailles a été de 11 229 tonnes en 2006, représentant à la vente au détail un chiffre d'affaire de l'ordre de 17 milliards de francs CFA. Cette production a connu une hausse en valeur absolue de 1936 tonnes soit 23 % en valeur relative par rapport à l'année 2005. (SENEGAL/ME/CNA, 2007). 1.2.1.2.4.2. Production d'oeufs de consommation La production nationale d'oeufs de consommation a été estimée d'une part à partir de poussins « ponte » mis en place entre mars 2004 et juin 2006 (qui ont permis de déterminer un effectif moyen de 1 424 814 pondeuses en production) et d'autre part, en tenant compte de certains paramètres : · taux de mortalité : 7 % à l'entrée en ponte ; · taux de mortalité : 3 % pendant la période de ponte ; · durée d'élevage avant l'entrée en ponte : 6 mois ; · performance annuelle d'une pondeuse : 280 oeufs par poule et par an ; · prix de l'oeuf : 50 francs CFA l'unité. La production nationale d'oeufs de consommation a été de 371 millions d'unité en 2006, soit un chiffre d'affaire à la vente au détail de l'ordre de 18 milliards de francs CFA. Cette production d'oeufs a connu une croissance progressive par rapport à l'année 2005, soit une valeur absolue de 47 millions d'unités (SENEGAL.ME.CNA, 2007). Ceci s'explique par le nombre important de reconversion d'éleveurs de poulets de chair en éleveurs de poules pondeuses. Cette production d'oeufs est essentiellement assurée par l'aviculture moderne car le poids de l'aviculture traditionnelle en production d'oeufs est presque nul. Ces différents types de productions sont pratiqués dans un cadre bien organisé. Cependant, l'arrêt des importations de produits avicoles (poussins d'un jour, d'oeufs de consommation et viande de volailles) a eu un impact positif sur les importations d'oeufs à couvés. Pour l'année 2006, on a un cumul de 9 614 630 oeufs à couvés importés par la filière. Ce chiffre constitue un record et représente presque le double de l'année 2005 qui était de 4 834 550 oeufs à couvés. Ce qui confirme encore une fois de plus la demande importante et croissante en poussins d'un jour. 1.2.1.2.5. Organisation de la production La filière avicole est l'une des rares filières agroalimentaires où il existe une structure professionnelle relativement bien organisée. Deux fédérations coexistent : l'Union Nationale des Acteurs de la Filière Avicole (UNAFA) qui représente les gros producteurs tandis que la Fédération des Acteurs de la Filière Avicole (FAFA) est le porte parole des petits éleveurs. L'aviculture moderne est un secteur organisé dans lequel interviennent divers acteurs : les sélectionneurs, les accouveurs, les éleveurs de reproducteurs, les producteurs, les provendiers et les encadreurs. Le rôle de chacun de ces acteurs est capital pour le bon fonctionnement du secteur. · Les accouveurs et éleveurs de reproducteurs Les éleveurs de reproducteurs font l'élevage des souches sélectionnées dans le but de produire des oeufs fécondés dont l'incubation donnera des poussins d'un jour destinés aux producteurs d'oeufs de consommation ou de poulets de chair. Le rôle des accouveurs se limite à l'incubation artificielle d'oeufs fécondés importés de l'étranger ou achetés auprès des éleveurs de reproducteurs locaux afin de fournir des poussins d'un jour aux producteurs. C'est le cas de la Société de Distribution du Matériel Avicole (SEDIMA), de la Compagnie Africaine de Maraîchage d'Aviculture et d'Arboriculture Fruitière (CAMAF), du Complexe Avicole de Mbao (CAM), Aviculture Sénégalaise (AVISEN), la Nouvelle Minoterie Africaine (NMA) etc. · Les producteurs Ils achètent les poussins d'un jour et les élèvent pour la production des oeufs de consommation ou de poulets de chair selon la spéculation choisie. · Les provendiers Au Sénégal, la fabrication et la vente des provendes en aviculture sont assurées par des sociétés locales telles que : la SEDIMA, le CAM, la Nouvelle Minoterie Africaine (NMA), SENTENAC etc. (SENEGAL.MA.DIREL, 1996). · Les encadreurs Ce sont des agents de structures publiques d'encadrement, les vétérinaires privés et les fournisseurs d'intrants et de poussins (HABYARIMANA, 1998). 1.2.1.2.6. Circuits de commercialisation d'oeufs et de poulets de chair Tous les produits issus de l'aviculture sont commercialisés essentiellement sur les marchés urbains pour la filière moderne, et ruraux pour la filière traditionnelle, mais également par l'intermédiaire des Bana-banas (les revendeurs informels). Les oeufs de consommation se retrouvent dans tous les circuits de distribution, du petit étal de marché aux grandes surfaces. 1.2.1.2.7. Niveaux de consommation d'oeufs et de poulets de chair au Sénégal La consommation d'oeufs peut être assimilée à la quantité d'oeufs produite par le secteur moderne puisque les importations d'oeufs de consommation sont négligeables voire inexistantes et que la production du secteur traditionnel est presque nulle (SENEGAL.ME.CNA, 2006). En 1995 la consommation d'oeuf était estimée à 19,64 oeufs par habitant au Sénégal, cette consommation est en nette augmentation depuis 1998 (KOE, 2001). La consommation de poulets de chair correspond à la quantité de poulets de chair produite par le secteur moderne et les importations de poulets congelés. En effet, en 2004, le volume des importations était de 13.700 tonnes pour une valeur de près de 13 milliards de francs CFA. Les morceaux congelés ont constitué 75 % du volume des importations. Si en 2004, la production locale de poulet de chair n'a été que de 7267 tonnes, on se rend donc compte que la majorité des consommateurs sénégalais ont privilégié les poulets congelés importés à la production locale (FRANCE.MEFI, 2005). Compte tenu du contexte actuel de la grippe aviaire, tout porte à croire qu'avec l'arrêt des importations de viande de volailles, une nette amélioration de la production locale de poulets de chair se fera sentir ; à condition que les producteurs locaux parviennent à mieux gérer les contraintes que rencontre la filière avicole au Sénégal. 1.2.2. CONTRAINTES DE L'ELEVAGE AVICOLE DANS LA REGION DE DAKAR On distingue plusieurs types de contraintes: · Les contraintes zootechniques ; · Les contraintes technico-économiques ; · Les contraintes sanitaires ; · Les contraintes Pathologiques. 1.2.2.1. CONTRAINTES ZOOTECHNIQUES L'insuffisance du niveau technique des éleveurs et l'insuffisance d'organisation des producteurs sont des facteurs qui entravent la productivité des élevages modernes. Les défaillances observées dans l'application des normes techniques d'élevage sont à l'origine de mauvaises performances. En effet, la mauvaise conception des bâtiments, les vides sanitaires mal effectués et l'absence d'hygiène souvent constatée dans les fermes ont des conséquences néfastes en élevage intensif (BIAOU, 1995). La qualité nutritive des aliments fabriqués de façon artisanale dans certaines fermes avicoles non qualifiées, la distribution irrégulière et en quantité insuffisante des aliments ainsi que la rupture prolongée des stocks d'aliments dans les fermes ne favorisent pas une production optimale de ces fermes. A ces problèmes zootechniques s'ajoutent, les contraintes technico-économiques. 1.2.2.2. CONTRAINTES TECHNICO-ECONOMIQUES L'élevage des poulets de chair comme celui des poules pondeuses n'est pas accessible à toutes les couches de la population sénégalaise. En effet, cet élevage demande des moyens financiers importants. En général, les poussins, les médicaments et 85 % du maïs destinés aux fabriques d'aliments sont des intrants importés. Les producteurs éprouvent d'énormes difficultés pour obtenir des financements nécessaires à l'achat des équipements avicoles (HABAMENSHI, 1994). La mauvaise organisation du marché et le manque de chaîne de froid pour conserver les produits invendus font que beaucoup d'aviculteurs sénégalais se limitent à des opérations ponctuelles liées à des festivités d'origines religieuses, coutumières ou familiales. (SENEGAL/MA/DIREL, 1995). En plus des contraintes technico-économiques s'ajoutent les contraintes sanitaires. 1.2.2.3. CONTRAINTES SANITAIRES Les contraintes sanitaires sont représentées par les facteurs de risque dans les poulaillers. Ces facteurs de risques sont nombreux et peuvent agir en synergie ou individuellement. Parmi ces facteurs, on peut citer : · La température C'est un facteur de stress aussi bien chez les poussins que chez les poules adultes (PARENT et coll., 1989). L'oiseau en réagissant face à l'agression thermique, s'épuise et s'expose davantage aux maladies. · L'humidité L'humidité favorise la croissance optimale des agents infectieux et infectants. Lorsqu'un poulet est soumis à un environnement à forte humidité, il devient plus réceptif aux maladies que celui qui n'est pas dans le même cadre de vie (BRUGERE-PICOUX et SAVAD, 1987). · La ventilation Le rôle de la ventilation est bien connu en aviculture car elle permet le renouvellement de l'air du poulailler. C'est d'ailleurs l'élément important qui est recherché dans l'orientation et la conception des bâtiments. Tout en évitant les grands vents, les poussières (sources d'agents pathogènes), Une bonne ventilation permet de minimiser les effets de la température et de l'humidité (IBRAHIMA, 1991). · Polluants chimiques L'ammoniac (NH3) est le polluant chimique le plus important. Il provient des oiseaux eux-mêmes ou résulte de la dégradation de la litière. Les facteurs physiques associés aux facteurs chimiques, favorisent l'apparition et l'évolution de nombreuses pathologies aviaires. 1.2.2.4. CONTRAINTES PATHOLOGIQUES (BULDGEN et coll., 1992) Les pathologies sont principalement d'origine parasitaire ou infectieuse. · Les maladies parasitaires Elles sont les plus nombreuses et sont responsables de la mortalité ou des retards de croissance dans les élevages. On retrouve entre autres :
· Les maladies infectieuses Elles rassemblent les maladies bactériennes et virales. Ø Les maladies bactériennes et mycoplasmiques Parmi ces maladies on peut citer :
Ce sont les maladies les plus graves. Elles entraînent d'énormes dégâts car il n'existe aucun traitement contre ces maladies. On peut citer entre autres : · La maladie de Gumboro due à un Birnavirus ; · La maladie de Newcastle ou pseudo peste aviaire due à un Paramyxovirus ; · La variole aviaire due à un Poxvirus ; · Les leucoses aviaires dues à des rétrovirus ; · La bronchite infectieuse due à un Coronavirus ; · La maladie de Marek due à un Herpes virus. Bien que les maladies parasitaires soient les plus fréquentes à cause du manque d'hygiène, il faut remarquer que les maladies infectieuses (bactérienne et virale) sont les plus redoutables, puisque leurs pronostics médicaux et économiques, sont généralement catastrophiques. A l'issue de la présentation de l'aviculture au Sénégal il ressort que les contraintes pathologiques entravent sérieusement son développement. Ainsi OUMAR (1994) a montré que la maladie de Gumboro avait une prévalence de 26 % par rapport aux autres pathologies aviaires ; ensuite BAKARI (2006) a montré que cette maladie peut entraîner dans une bande de poulets de chair une perte économique de 75,81 %. Le prochain chapitre sera consacré à la présentation de la maladie de Gumboro.CHAPITRE 2 : LA MALADIE DE GUMBORO La maladie de GUMBORO est une maladie infectieuse, virulente, inoculable et contagieuse due à un virus lymphotrope de la famille des Birnaviridae dénommé IBDV (Infectious Bursal Disease Virus). Le virus attaque principalement les cellules lymphoïdes produites par la bourse de Fabricius et l'infection est suivie d'une immunodépression (VINDEVOGEL, 1992). Elle frappe tous les gallinacés et se caractérise cliniquement par des troubles digestifs (diarrhée aqueuse), des plumes ébouriffées, une mobilité réduite, de l'apathie, de l'anorexie, des tremblements et une prostration. Sur le plan anatomopathologique, elle se manifeste par une inflammation nécrosante de la bourse de Fabricius, une déshydratation très apparente au niveau des muscles, qui présentent également de nombreuses hémorragies et ecchymoses, une hypertrophie et une décoloration des reins, avec une accumulation de cristaux d'urates dans les tubules (PICOUX M., 1983). C'est la bourse de Fabricius qui présente les lésions essentielles pour le diagnostic : chez les sujets qui meurent en phase aiguë de l'infection, la bourse de Fabricius est hypertrophiée, turgescente, avec une décoloration jaune pâle. Des hémorragies intrafolliculaires peuvent être présentes et dans certains cas la bourse de Fabricius peut être totalement hémorragique et prendre l'aspect d'un caillot de sang (« cerise noire »). La maladie de Gumboro a une importance à la fois économique et médicale. Ø Sur le plan économique, elle entraîne une morbidité moyenne de 20% pouvant atteindre parfois 100% accompagnée d'une prostration sévère de la plupart des animaux durant 5 à 7 jours. La mortalité dont le taux est faible s'élève brusquement pendant 2 jours puis décline rapidement durant les 2 à 3 jours suivants. Habituellement, cette mortalité varie de 5 à 10 %, mais elle peut atteindre un pic de 60 % (VANMARCK, 1992). Les conséquences de la maladie en dehors de la mortalité se traduisent par une chute de ponte, un retard de croissance et une hétérogénéité du lot (PICAULT J. P., 1998). Ø Sur le plan médical, la maladie a un effet immunodépresseur marqué pouvant être à l'origine de certains échecs de vaccination contre la maladie de Newcastle par exemple selon STEWART et coll. (1993), rapporté par KOUZOUKENDE (2004). La maladie a été décrite pour la première fois par COSGROVE (1962) sur les jeunes volailles. Elle sévissait depuis 1957 aux USA dans l'Etat de DELAWARE plus précisément Dans la ville de Gumboro (VINDEVOGEL, 1992) A l'autopsie les poussins présentent des lésions rénales et de la bourse de Fabricius d'où la dénomination de « Néphrose aviaire » ou maladie de Gumboro. En 1962, WINTERFIELD et HITCHNER aux USA ont observé sur les poulets des lésions rénales semblables à celles décrites par COSGROVE qu'ils ont nommé « syndrome néphrite néphrose ». Puis ils ont isolé deux virus, l'un des reins et l'autre de la bourse de Fabricius de poulets atteints de cette pathologie. Ils ont démontré que le virus isolé de la bourse de Fabricius est le seul responsable des lésions induites dans cet organe. Ainsi l'appellation « maladie de Gumboro » fut dès lors réservée à l'affection virale caractérisée par la dégénérescence et la nécrose des cellules lymphoïdes de la bourse de Fabricius. Il existe deux types des souches baptisées « Holte » et « Gray » qui ont des apparentées immunologiques avec le virus de la bronchite infectieuse. Mais ils différent par le fait que celui de la bronchite entraîne des troubles respiratoires et celui de la maladie de Gumboro entraîne des lésions spécifiques au niveau de la bourse de Fabricius. En février 1975 la maladie de Gumboro a été signalée pour la première fois au Sénégal (SAGNA, 1975). La maladie de Gumboro est une maladie des gallinacés. Dans les conditions naturelles, la maladie ne s'exprime cliniquement que chez la poule. La dinde, le canard, la pintade la caille, les passereaux, l'oie et l'autruche peuvent être occasionnellement infectés mais sous une forme subclinique (VINDEVOGEL, 1992). Dans les conditions expérimentales, la poule reste sensible uniquement à l'inoculation par voie buccale. Mais la maladie de Gumboro peut être reproduite chez la poule par l'inoculation du virus par voie intraperitonéale, intracérébrale ou intraveineuse tandis que chez la souris blanche âgée de 1-14 jours elle n'est possible que par voie intracérébrale ou intraperitonéale selon BENTON et coll. (1967). 2.1.5. REPARTITION GEOGRAPHIQUE La maladie de Gumboro est une maladie cosmopolite. Des USA, elle s'est propagée dans le reste du monde, à savoir l'Europe via la grande Bretagne, l'Asie et l'Afrique où son identification a été tardive. Le virus est largement répandu à travers le monde, mais on pensait qu'il était absent dans la majorité des îles du pacifique. Cependant il a été signalé à Fidji en Polynésie française, en Nouvelle Zélande et à Vanuatu (SAVILLE, 1999). De nos jours plusieurs pays africains sont touchés par la maladie de Gumboro dont le Sénégal. 2.2.1. MORPHOLOGIE ET STRUCTURE Il a été démontré que les particules virales du virus de la maladie de Gumboro, formées par des protéines VP2 et VP3, présentent une symétrie icosaédrique de triangulation T= 13, avec un diamètre d'environ 700Å. Cette structure du virus est déterminée par cristallographie à 7Å de résolution (REY et coll., 2004). Le phénotype de virulence accrue est déterminé par la protéine majeure de capside VP2. Cette protéine constitue d'une part le moteur de la morphogenèse par ses capacités d'auto-assemblage et d'autre part un déterminant du tropisme du virus par son interaction avec des récepteurs cellulaires (COULIBALY et coll., 2003). Le génome viral est constitué d'une chaîne d'acide ribonucléique (ARN) bicaténaire et bisegmentée. 2.2.2. CARACTERISTIQUES PHYSICO-CHIMIQUES ET CLASSIFICATION Le virus de la maladie de Gumboro a fait l'objet de plusieurs controverses : Ø En 1967, CHEVILLE rapporté par DIALLO (1978) décrivit des zones de regroupements du virus dans le cytoplasme de macrophages de poulets infectés. Les particules virales étaient entourées d'une trame filamenteuse. L'existence de cette trame, les caractères morphologiques de ces particules et les différentes propriétés physico-chimiques l'amenèrent à admettre que le virus de la maladie de Gumboro était un réovirus. Ø PETEK et coll., en 1967 l'assimilèrent également à un réovirus. Cette hypothèse fut réfutée par LUNGER et MADDUX qui en 1972 étudièrent au microscope électronique les transformations cellulaires survenant après infection. Ils constatèrent une altération primitive du noyau des macrophages et l'apparition d'inclusions cytoplasmiques qui sont uniquement d'origine macrophagique et non des fragments de lymphocytes phagocytés. La réplication du virus de la maladie de Gumboro, ainsi que les phénomènes morphologiques qui l'accompagnent, ressemblent à la réplication du virus Nodaruma étudié par MURPHY (1968). Ce virus Nodaruma est un picornavirus transmis par les arthropodes. En 1991, le virus de la maladie de Gumboro a été définitivement identifié et classé dans la famille des Birnaviridae. Ce virus est très stable, non enveloppé, d'un diamètre de 60 nm. Il est composé d'un double brin d'ARN entouré d'une capsule protéique et présente une attirance pour les tissus lymphoïdes notamment la bourse de Fabricius. Il détruit les lymphocytes dans tous les organes lymphoïdes provoquant une immunodépression plus ou moins sévère. Ce virus a une très grande facilité d'expansion et peut contaminer toutes les régions à forte densité avicole. Le virus de la bursite infectieuse est très résistant aux agents chimiques et physiques. Il résiste à un pH supérieur à 2 et inférieur à 12. Il présente une grande résistance à la chaleur dans le milieu extérieur. A 70° C il résiste pendant 30 minutes et à 56° C pendant 5 heures. Il présente également une grande résistance à certains agents chimiques : chloroforme, éther, acides, formol à 1% et à l'eau de javel. Il est inactivé à pH=2 (VINDEVOGEL, 1992) mais certains désinfectants sont actifs contre lui : le chloramine à 2 %, le glutaraldehyde ; le formol quant à lui n'agit qu'à une température supérieure à 20° C d'où son efficacité relative s'il est employé en hiver ou à des températures relativement basses. · Sur oeufs embryonnés La culture est faite sur oeufs embryonnés sans anticorps spécifiques ou Exempte d'Organismes Pathogènes Spécifiques (EOPS) âgés de 6 à 10 jours par inoculation intra chorio-allantoïdienne. L'embryon meurt dans 3 à 5 jours. A l'autopsie il présente :
· Sur culture cellulaire Elle est faite sur les fibroblastes du poulet, des cellules de l'embryon de dindon, de canard ou sur les lignées cellulaires des reins de lapin et de singe. Il est variable : · Dans les conditions naturelles Le virus de la maladie de Gumboro est naturellement pathogène pour les oiseaux plus précisément les gallinacés. Cette sensibilité est fonction de l'âge, d'où chez les sujets de 5 jours, il n'y a pas expression de la maladie. L'infection entraîne une immunodépression durable. Chez les sujets qui ont entre 3 et 6 semaines, la forme aiguë d'apparition brutale, est la plus observée et elle se manifeste par une diminution de l'immunité maternelle. La pathogénie est variable en fonction des souches virales. On a des souches « traditionnelles » connues depuis 1962 et qui entraînent 5 à 10 % de mortalité (BRICOUT et coll., 1974). Certains pathotypes apparus depuis 1987 entraînent un taux de mortalité de 5 à 60 % (VANMARCK, 1992). L'effet pathogène du virus dans la maladie naturelle se traduit par une hypertrophie suivie d'une atrophie de la bourse de Fabricius. · Dans les conditions expérimentales L'embryon de moins de 6 jours est moins sensible au virus que celui de 12 jours. Le passage en série sur une culture cellulaire du virus entraîne l'atténuation de son pouvoir pathogène. Le virus atténué peut être utilisé pour la production des vaccins. 2.2.4.2. Pouvoir antigénique et immunogène Le virus de la bursite infectieuse possède des antigènes qui induisent la formation des anticorps neutralisants et précipitants qu'on peut mettre en évidence par l'immunofluorescence ou par la technique ELISA. Deux sérotypes ont été identifiés: Ø Le sérotype I comprend plusieurs souches comportant les antigènes différents entre souches classiques et variantes. C'est une souche très pathogène pour le poussin et peut vaincre la protection passive des jeunes oiseaux grâce à sa différence antigénique avec la souche standard vaccinale (persistance des anticorps maternels). Ø Le sérotype II a été isolé du dindon chez lequel il ne provoque qu'une affection subclinique inapparente qui serait quand même immunosuppressive Le sérotype I est responsable de la maladie chez les poules, alors que le sérotype II se rencontre principalement chez les dindes (SAVILLE, 1999). Les deux sérotypes peuvent interférer aussi bien chez les poulets que les dindons. Par ailleurs WINTERFIELD (1969) montra que les poulets guéris de la maladie ou ayant été mis en contact avec une souche atténuée du virus, possédaient des anticorps dirigés contre les souches homologues et hétérologues. Ces travaux montrent donc l'existence de neutralisations croisées entre les différentes souches et ceci présente un grand intérêt dans la préparation des vaccins où il n'est pas nécessaire d'inclure toutes les souches connues du virus comme principe actif. Le même auteur démontre par la même occasion que l'âge où doit se faire le contact du virus avec le poussin importe beaucoup, car les poussins de trois semaines infectés développaient un taux d'anticorps neutralisants très inférieur à celui produit par les poussins de quatre semaines. Les anticorps précipitants dont l'existence a été démontrée par FARAGHER (1972) apparaissent du 2ème au 6ème jour après l'infection de la bourse de Fabricius. Il existe une variation de localisation de pouvoir pathogène : · Les souches virulentes se multiplient dans les zones inter folliculaires de la bourse de Fabricius · Les souches virulentes se multiplient dans les follicules. Le virus entre dans l'organisme par la voie orale. L'incubation de la bursite infectieuse est très brève. Le virus transite dans les lymphocytes et les macrophages intestinaux quelques heures après l'infection orale. L'envahissement hépatique précède la virémie qui assure la contamination des organes cibles dont la bourse de fabricius. Cette atteinte correspond à une « bursectomie virale » détruisant les lymphocytes B porteurs de l'immunité à médiation humorale. Il y a réaction inflammatoire de la bourse de Fabricius le 4ème jour qui suit l'infection, puis atrophie et dégénérescence en une semaine qui accompagne la nécrose des autres organes lymphoïdes. Virus Muqueuse intestinale Macrophages, lymphocytes Foie Virémie Bourse de Fabricius (multiplication du virus dans les cellules lymphoïdes B)
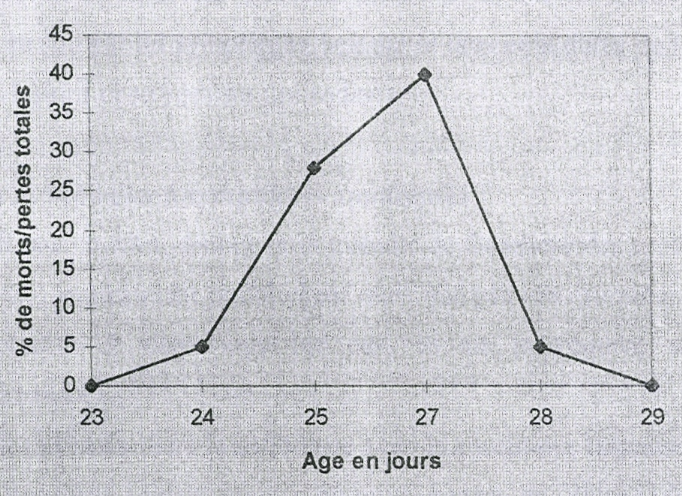
Hypertrophie Atrophie Figure 2 : Pathogénie de la maladie de Gumboro La bourse de Fabricius est un organe lymphoïde primaire, impair et médian rencontré uniquement chez les oiseaux. Cet organe présente également certaines propriétés des organes lymphoïdes secondaires. Elle est située dorsalement au cloaque. Sa cavité est recouverte longitudinalement par un épithélium plissé, formant environ 15 bourrelets primaires et 7 secondaires. Le développement de la bourse de Fabricius commence à partir du 4ème jour de la vie embryonnaire et atteint son maximum à l'âge de 4 semaines. La bourse de Fabricius régresse entre la 10ème et la 23ème semaine, ce qui consiste à un épuisement lymphoïde physiologique qui s'achève vers l'âge de la maturation sexuelle. Au cours de la vie embryonnaire, les cellules souches des lymphocytes vont migrer du foie et du jaune d'oeuf vers le thymus et la bourse de Fabricius. C'est ainsi que les lymphocytes T issus du Thymus seront responsables de l'immunité à médiation cellulaire, et les lymphocytes B issus de la bourse de Fabricius seront responsables de l'immunité humorale grâce aux immunoglobulines qu'ils fabriquent (SALIM et REKIK, 1992). Une fois passés dans la lymphe et le sang, les lymphocytes seront chargés de bloquer une éventuelle intrusion des agents porteurs des antigènes correspondants. La présence de la bourse de Fabricius est surtout nécessaire pendant les deux premières semaines, ce qui ne signifie pas que l'oiseau puisse s'en passer sans risque, de la 2ème à la 10ème semaine (SCALA et coll., 1988). C'est dans cet organe lymphoïde que le virus attaque les lymphocytes B et s'y multiplie avec un effet cytolytique entraînant les réactions inflammatoires qui se traduisent par une hypertrophie de la bourse de Fabricius. A la suite de la destruction des lymphocytes B, il se produit une dépression immunitaire. Cette dépression nuit à la protection contre les maladies bactériennes telles que les colibacilloses et les salmonelloses (WYETH, 1976). 2.3.2. CONSEQUENCES PHYSIOPATHOLOGIQUES Les conséquences immédiates de cette affection sont une immunosuppression quasi immédiate entraînant de graves échecs aux vaccinations diverses (Newcastle, bronchite infectieuse, Marek). La disparition de certaines barrières immunitaires entraînera l'éclosion d'affections parasitaires, virales et bactériennes variées. Diverses hypothèses sont émises quant à l'origine des lésions et symptômes des formes graves : · Coagulation intra vasculaire disséminée ou CIVD (il y a libération de thromboplastine à partir de la bourse de Fabricius lésée) ; · La maladie à immuns complexes, avec vascularite qui provoquerait les lésions hémorragiques et en partie l'atteinte rénale. Les autres conséquences physiopathologiques sont entre autre des diarrhées entraînant des déshydratations aggravées par l'absence d'abreuvement. Ce qui a pour conséquence l'accumulation de cristaux d'urate dans les reins et les uretères laissant présager un pronostic médical sombre ; L'infection précoce chez les poussins de 5 jours entraîne une immunodépression subclinique tandis qu'a partir de la 3ème semaine on a la forme clinique aiguë. 2.4.1. EPIDEMIOLOGIE DESCRIPTIVE La maladie de Gumboro affecte naturellement les poulets mais aussi les dindons, les cailles, les passereaux et les canards. Les zones les plus affectées sont les zones où se concentre un grand nombre de volailles. Les mortalités enregistrées évoluent selon une courbe de mortalité en cloche pathognomonique de la maladie de Gumboro ou courbe de PARKHUST (figure 3). A Dakar, la maladie de Gumboro évolue généralement sous une forme enzootique. Cependant, il y a des périodes particulières comme l'hivernage où nous assistons à des épizooties.
Figure 3 : Courbe caractéristique de mortalité de la forme aiguë de la maladie de Gumboro selon PARKHUST cité par ABDEL-AZIZ (2007). 2.4.2. EPIDEMIOLOGIE ANALYTIQUE Ø Espèce La maladie se rencontre surtout chez le genre Gallus, le canard et le dindon développent des formes subcliniques inapparentes. Les sources sont les animaux malades ou morts, les fientes, l'eau, les litières et les aliments contaminés. Ø L'âge La maladie de Gumboro se nomme souvent « la maladie aux deux visages » car durant les deux premières décades de vie, l'infection précoce provoque une immunosuppression sévère. De 3 à 6 semaines c'est l'age de la plus grande sensibilité au virus. L'immunité naturelle est fonction de l'état immunitaire des reproducteurs. Ø L'individu Les variations de sensibilité individuelles sont liées aux résidus d'immunité passive (persistance des anticorps maternels). Ø Le milieu Le stress et les mauvaises conditions d'hygiène sont les facteurs favorisants l'apparition et la persistance de la maladie de Gumboro chez les animaux sensibles au virus. Ø Transmission Les animaux se contaminent soit par contact direct avec les malades soit par l'intermédiaire des vecteurs passifs contaminés par les fientes. L'excrétion virale persiste deux semaines après la contamination et tous les animaux peuvent être porteurs. La voie de contamination est soit orale soit respiratoire. Il n'y a pas de transmission par l'oeuf. 2.4.3. EPIDEMIOLOGIE SYNTHETIQUE L'introduction du virus dans le milieu se fait par le biais des échanges commerciaux des volailles. L'existence de nombreux vecteurs passifs (eau contaminée, litière contaminée etc.) et des animaux réservoirs du virus (canards, dindons) font que la maladie évolue durant toute l'année. 2.5. DIAGNOSTIC DE LA MALADIE DE GUMBORO 2.5.1. DIAGNOSTIC SUR LE TERRAIN 2.5.1.1. ELEMENTS CLINIQUES ET EPIDEMIOLOGIQUES Un troupeau affecté présentera une morbidité très élevée sur les poulets de 3 à 6 semaines accompagnée d'une prostration sévère de la plupart des animaux durant 5 à 7 jours. La mortalité s'élève brusquement pendant 2 jours puis décline rapidement durant les 2 à 3 jours suivants. Habituellement, la mortalité varie de 5 à 10 %, mais elle peut atteindre 30 à 40 %. Les principaux symptômes sont une diarrhée aqueuse pouvant contenir des caillots de sang (BRUGERE-PICOUX, 1974 ; ROSENBERGER, 1989 ; VINDEVOGEL, 1992), des plumes ébouriffées, une mobilité réduite, de l'anorexie, des tremblements et une prostration. 2.5.1.2. ELEMENTS NECROPSIQUES On peut reconnaître la maladie de Gumboro lors de l'ouverture du cadavre de poulet suspect. Les lésions observables à l'examen nécropsique incluent une déshydratation très apparente au niveau des muscles, qui présentent également de nombreuses hémorragies et ecchymoses, une hypertrophie et une décoloration des reins, avec une accumulation de cristaux d'urates dans les tubules. C'est la bourse de Fabricius qui présente les lésions essentielles pour le diagnostic : chez les sujets qui meurent en phase aiguë de l'infection, la bourse de Fabricius est hypertrophiée, turgescente, avec une décoloration jaune pâle (HANSON, 1967). Des hémorragies intrafolliculaires ou des substances caséeuses sur les feuillets peuvent être présentes (ROSENBERGER, 1989 ; VINDEVOGEL, 1992) et dans certains cas la bourse de Fabricius peut être totalement hémorragique et prendre l'aspect d'un caillot de sang «cerise noire ». Un oedème péribursal de couleur jaune paille est présent chez de nombreux sujets. Il faut cependant être prudent pour écarter les affections qui peuvent ressembler à la maladie de Gumboro ; d'où l'intérêt de prendre en compte les éléments différentiels. 2.5.1.3. DIAGNOSTIC DIFFERENTIEL Certaines maladies peuvent prêter à confusion avec la maladie de Gumboro soit par les syndromes, soit par le taux et la durée de la mortalité, soit par des lésions observées sur des cadavres. Il faut en effet distinguer la maladie de Gumboro des symptômes toxiques qui certes, apparaissent brutalement mais peuvent entraîner jusqu'à 100 % de mortalité et dans tous les cas, ils ne provoquent pas de lésions caractéristiques. Aussi il faut faire la différence avec la coccidiose qui est responsable de la diarrhée et de mortalité brutale mais n'entraîne jamais de lésions de bourse de Fabricius. L'une des affections à ne pas confondre avec la maladie de Gumboro est la maladie de Newcastle. Elle provoque des lésions hémorragiques et affecte tous les animaux quelque soit leur âge et persiste plus longtemps dans l'élevage avec des mortalités élevées (jusqu'à 100%). Elle provoque en outre, des signes nerveux et respiratoires qui n'apparaissent pas dans la maladie de Gumboro. Les hémorragies au niveau du proventricule sont situées sur les papilles sous forme de taches. Du fait de l'atteinte rénale, il faut aussi écarter le syndrome néphrite néphrose qui s'accompagne de signes respiratoires et qui n'entraîne aucune lésion de la bourse de Fabricius. On peut écarter aussi la lipoïdose hépatorénale, qui du fait de la faible mortalité qu'elle entraîne sur des sujets de 3 semaines et les lésions rénales peuvent être confondues avec la maladie de Gumboro. Mais, là encore il n'y a pas de lésions de la bourse de Fabricius. Cependant, il y a des affections comme l'avitaminose A, la leucose lymphoïde et la maladie de Marek qui peuvent entraîner des lésions de la bourse de Fabricius mais à l'histologie on s'aperçoit que dans l'avitaminose A, il s'agit d'une métaplasie épithéliale et dans la maladie de Marek et les leucoses, il s'agit des processus tumoraux. Si le doute persiste encore malgré toutes les investigations, on fait appel au diagnostic de laboratoire. 2.5.2. DIAGNOSTIC DE LABORATOIRE 2.5.2.1. DIAGNOSTIC HISTOPATHOLOGIQUE L'examen histopathologique met en évidence des lésions oedémateuses, hémorragiques et nécrosantes ou l'atrophie folliculaire de la bourse de Fabricius. 2.5.2.2. DIAGNOSTIC VIROLOGIQUE Deux techniques sont couramment utilisées pour mettre en évidence le virus de la maladie de Gumboro. Il s'agit de l'immunofluorescence et la technique de l'inoculation. Elle consiste à inoculer des broyats de bourse de Fabricius suspects aux poulets sensibles (3 à 6 semaines d'âge et dépourvus d'anticorps spécifiques). Ensuite rechercher au bout de 3 jours sur les bourses de Fabricius des poulets inoculés des lésions histopathologiques caractéristiques. Et enfin au bout de 6 jours, rechercher des lésions macroscopiques sur les cadavres. En raison de la contamination fréquente de la bourse de Fabricius par d'autres virus on préfère utiliser la rate qui donne de bons résultats. Les prélèvements peuvent aussi être inoculés à la membrane chorioallantoidienne des oeufs embryonnés de 10 jours dépourvus d'anticorps spécifiques. Les embryons meurent au bout de 3 à 4 jours et les lésions observées sont des oedèmes de la tête, du cou et de l'abdomen ; des congestions et des hémorragies dans le tissu conjonctif sous cutané et une coloration verdâtre du jaune d'oeuf et du liquide allantoïdien. L'inoculation peut aussi se faire sur culture cellulaire de fibroblastes de poulet, des cellules d'embryon de dindon ou de canard. La multiplication du virus provoque au voisinage des noyaux des cellules infectées, des inclusions cytoplasmiques éosinophiles à contours irréguliers. 2.5.2.2.2. L'immunofluorescence Elle consiste à mettre en évidence les antigènes du virus au niveau des calques de la bourse de Fabricius sur lame, grâce à la réaction antigène - anticorps en utilisant des immunoglobulines antivirus Gumboro marquées par la fluorescéine. 2.5.2.3. DIAGNOSTIC SEROLOGIQUE La sérologie met en évidence le passage des antigènes viraux. En effet, des prélèvements seront effectués sur les oiseaux à des intervalles bien définis pour voir la cinétique des anticorps. Trois techniques sont utilisées : · la technique de précipitation en milieu gélosé : sensible et peu onéreuse ; · la technique de séroneutralisation : sensible mais délicate et onéreuse; · la technique ELISA (Enzyme Linked Immunosorbent Assay) : facile à mettre en oeuvre mais nécessite des Kits ELISA relativement coûteux. Elle est utilisée pour la détection des anticorps induits par l'IBDV et donne de très bons résultats. Il n'y a pas de traitement spécifique contre la maladie de Gumboro. Un traitement symptomatique peut consister en l'administration d'électrolytes dans l'eau de boisson et lutter contre les agents opportunistes (coccidies et bactéries). Elle repose sur les règles d'hygiène de base dans l'élevage aviaire : · élevage bien isolé avec des locaux bien conçus, faciles à nettoyer, à désinfecter à dératiser etc. · élevage en bande unique avec pour chaque poulailler, un ouvrier et du matériel propre ; · le nettoyage et la désinfection doivent être effectués après chaque bande suivant un ensemble de procédures strictes ; un vide sanitaire de 15 jours au minimum doit précéder l'arrivée d'une nouvelle bande. Le virus est très résistant et persiste longtemps dans le milieu extérieur. La rencontre du virus et du poussin est donc inévitable, précoce et individuelle car il n'y a pas de transmission verticale. La contagion est simultanée pour tous les poussins. Une bonne protection des poussins passe par la vaccination des parents car les anticorps maternels persistent quatre semaines si les poules sont bien vaccinées. La vaccination est actuellement la seule méthode efficace dans la prévention des maladies virales. Elle permet de renforcer les défenses immunitaires de l'individu contre un microbe, en injectant ce dernier sous une forme qui n'est plus pathogène (qui ne provoque pas la maladie) ou qui ne peut pas se répliquer. Ainsi la vaccination protège l'organisme contre le virus qui a servi à fabriquer le vaccin. Idéalement, les vaccins devraient protéger non seulement contre les manifestations cliniques et mortalité, mais encore prévenir la perte en gain de poids et l'immunosuppression associée à la maladie. Une gamme de vaccins a été décrite dans des rapports par THORNTON et PATTISON (1975) et THORNTON (1976) dans lesquels il a été montré que certains vaccins réunissaient ces qualités idéales, mais d'autres étaient encore suffisamment virulents pour provoquer la maladie qu'ils étaient destinés à prévenir. On distingue deux sortes de vaccins : · Vaccins à virus inactivés : En 1964, WINTERFIELD et HITCHNER rapportent que les vaccins inactivés sont inefficaces en matière d'immunité. Cette conception a été révisée. En effet selon BENNEJEAN (1977) des vaccins inactivés ont été expérimentés, mais les concentrations virales nécessaires à l'obtention de l'immunité sont actuellement trop élevées pour permettre une production industrielle. DESBORGES (1999) a montré que le vaccin inactivé est totalement insensible aux anticorps maternels des poussins. Ce vaccin induit une protection progressive et de longue durée. · Vaccins à virus vivants : Dans ce domaine les vaccins ont connu deux périodes : une première période où on utilisait des vaccins à virus pleinement virulents et une période où les vaccins furent atténués. Les vaccins à virus pleinement virulents sont préparés à partir de suspension de bourse de Fabricius de poulets infectés. A l'heure actuelle, ces vaccins sont abandonnés au profit des vaccins à virus atténués. CONSTANTIN (1988) a montré que les vaccins vivants atténués utilisés très précocement seront neutralisés par les anticorps d'origine maternelle chez les poussins. Pour une vaccination efficace contre la maladie de Gumboro avec les vaccins vivants, FERRE et coll., (2005) ont montré qu'il faut un taux d'anticorps d'origine maternelle compatible avec la souche vaccinale soit 350 en ELISA (kit IDEXX, dilution 1/500) pour les vaccins intermédiaires et 500 pour les vaccins à souches dites « chaudes ». · Programme de vaccination : Il faut chercher à obtenir des poussins un niveau immunitaire élevé et uniforme. Le relais immunitaire actif du poussin doit être pris au moment optimum : les poussins sont sensibles entre trois et six semaines au virus de Gumboro et la persistance des anticorps maternels peut entraver la bonne réponse vaccinale. Il faut bien choisir le protocole vaccinal. Il y a actuellement recrudescence de cas aiguës malgré la vaccination. Une vaccination trop précoce est neutralisée par les anticorps maternels ce qui permet l'installation d'un virus sauvage plus pathogène. Il faut suffisamment d'anticorps maternels pour maîtriser une éventuelle souche sauvage mais pas trop pour ne pas neutraliser le virus vaccinal. De plus le stock d'anticorps maternels se dilue plus ou moins vite en fonction de la vitesse de croissance du poussin. Une souche lourde diluera beaucoup plus vite son immunité transmise qu'une souche label à croissance lente. Il existe donc un moment optimum de vaccination difficile à déterminer Les poussins venant des reproducteurs vaccinés héritent d'un taux d'anticorps élevé, destinés à les protéger pendant les 3 premières semaines de vie. Cette protection théorique est, malheureusement souvent prise à défaut lorsque la pression virale est élevée ou lorsqu'on est en face de souches sauvages très virulentes. La sensibilité du poussin au virus de la maladie de Gumboro et le risque d'infection précoce nécessite une vaccination au cours des premiers jours de la vie (VINDEVOGEL et coll., 1976). Tel est le cas des oiseaux dépourvus d'anticorps maternels : ils doivent alors être vaccinés à la naissance avec une souche atténuée qui n'entraîne ni lésions de la bourse de Fabricius, ni un effet immunodépresseur. Dans le cas contraire, la vaccination doit être différée, car les souches vaccinales atténuées sont neutralisées par les anticorps maternels ou doit être pratiquée avec des souches plus agressives, qui confèrent une bonne protection en présence d'anticorps maternels, sans qu'apparaissent les lésions de la bourse liées au manque d'innocuité relatif de ces souches. L'étude expérimentale de la dynamique des anticorps maternels révèle que ces derniers disparaissent en une dizaine de jours dans le cas des anticorps précipitants et en 20 à 30 jours dans le cas des anticorps de type neutralisant. En effet, dans la pratique il existe des grandes variations du taux des anticorps maternels chez les poussins car ces derniers proviennent de troupeaux de reproducteurs différents. Dans la région de Dakar, l'observation de la cinétique des anticorps a révélé que 52,6% des poussins produits à Dakar avaient un seuil de protection bas à partir de la 3ème semaine (CARDINALE et coll., 1998). Il se peut donc, en retardant la vaccination avec des souches atténuées, que la contamination intervienne à une période où l'immunité active n'a pas pris le relais de l'immunité passive, ou en utilisant des souches plus agressives. Il se peut que les poussins ne possèdent pas un taux d'anticorps maternels suffisants pour empêcher le développement des lésions de la bourse (VINDEVOGEL et coll., 1976). Dans ces conditions, il serait souhaitable d'immuniser les reproducteurs dont la descendance est soumise à des risques possibles d'infection. Une deuxième vaccination est obligatoire entre le 25ème et le 28ème jour de la vie. Faute de quoi les sujets porteurs d'anticorps à la naissance mais qui ne sont plus protégés entre le 25ème et le 30ème jour peuvent payer un lourd tribut au virus (signes cliniques et dépression immunitaire) Malgré les mesures sanitaires et médicales préconisées pour lutter contre la maladie de Gumboro, on observe sur le terrain de plus en plus des échecs de la vaccination. C'est dans le but de mieux comprendre et expliquer les raisons de l'échec de la vaccination contre la maladie de Gumboro, que nous avons entrepris une étude comparative des taux d'anticorps et de lymphocytes B produites par la bourse de Fabricius et responsables de l'immunité dans la maladie de Gumboro en conditions expérimentales. Ceci fera l'objet de la deuxième partie de notre travail. DEUXIEME PARTIE : ETUDE EXPERIMENTALE CHAPITRE 1 : MATERIEL ET METHODES Notre travail s'est déroulé de septembre 2007 à juin 2008, soit une durée de dix mois. Lors de cette expérimentation, les bandes de poussins étaient conduites dans les bâtiments situés dans l'enceinte de l'Ecole Inter Etats des Sciences et de Médecine Vétérinaires de Dakar (E.I.S.M.V.).
Figure 4: Carte de Dakar (source : www.senegal-online.com) Le matériel animal est représenté par des coquelets de souche Isabrown et des poulets de chair de souche Cobb500. La bande de poulets de chair comportait 520 poussins et celle de coquelets 160 poussins. Les poussins d'un jour provenaient du couvoir de la Sénégalaise de Distribution de Matériel Avicole (SEDIMA) pour les coquelets et les poulets de chair. Les poussins ont été nourris avec de l'aliment de SEDIMA pour le stade démarrage et l'aliment industriel de la Nouvelle Minoterie Africaine (NMA) pour les stades croissance et finition aussi bien des poulets de chair et des coquelets. Avant l'arrivée des poussins, nous avons effectué un vide sanitaire dans le bâtiment d'élevage. Le local a été nettoyé avec de l'eau et des détergents, rincé et ensuite désinfecté. Pendant 15 jours la salle a été close afin de permettre la destruction des germes de surface. Elle a été ouverte 24h avant l'arrivée des poussins. La litière, composée de copeaux de bois a été installée ainsi que quelques cartons pour servir de cloisons pour délimiter une aire de démarrage d'environ 2 m2. Pour assurer le chauffage des oiseaux, nous avons utilisé des ampoules de 100 watts disposées à environ 1 m du sol. Les poulets de chair et les coquelets ont été nourris avec deux types d'aliments : l'aliment SEDIMA pour le stade de démarrage et l'aliment NMA pour les stades de croissance et finition.
Nous avons utilisé dans notre élevage, les différents vaccins disponibles sur le marché sénégalais (tableau II).
1.2.3. MATERIEL DE PRISE DE SANG Pour le prélèvement de sang, nous avons utilisé des seringues, des aiguilles, des tubes à essai, des portes tubes, une glacière contenant des carboglaces pour la conservation. Des marqueurs ont été utilisés pour identifier les échantillons. 1.3.1. PROTOCOLES DE VACCINATION 1.3.1.1. Bandes de poulets de chairDans notre élevage expérimental, nous avons utilisé pour les deux bandes de poulets de chair trois types de vaccins : v le vaccin inactivé adjuvé qui a été utilisé par voie sous cutanée ; v les vaccins vivants lyophilisés souche intermédiaire et souche chaude qui ont été administrés par voie occulo-nasale (trempage du bec). Les différents protocoles de vaccination utilisés dans les bandes sont fixés par nous même (tableaux III et IV). Le protocole de vaccination de la bande NO1 de poulets de chair est récapitulé dans le tableau III. Tableau III : Protocole de vaccination de la bande NO1 de poulets de chair
J9= 9ème jour de la vie ; J25= 25ème jour de la vie ; S/C = sous cutanée Les poussins du lot1 étaient vaccinés à J9 avec le vaccin inactivé adjuvé (L191644) en injection par la voie sous cutanée. Les poussins du Lot2 quant à eux ont été vaccinés deux fois à J9 et J25 respectivement avec un vaccin inactivé adjuvé (L191644) et un vaccin vivant lyophilisé à souche intermédiaire (2305R2S1B) Le protocole de vaccination de la bande NO2 de poulets de chair est récapitulé dans le tableau IV. Tableau IV : Protocole de vaccination de la bande NO2 de poulets de chair
J18= 18ème jour de la vie Les poussins du lot1 étaient vaccinés à J18 avec le vaccin vivant à souche chaude (2305R2S1B) par la voie occulo-nasale. Nous avons utilisé différents vaccins et différentes modalités de vaccination à des dates différentes. Les poussins ont été regroupés en lots identifiables avant d'être vaccinés. Un marquage à la peinture a permis d'identifier les oiseaux de différents lots. Certains poussins ont été marqués sur la tête et d'autres sur les ailes avec des peintures de différentes couleurs. Le marquage est refait régulièrement lorsqu'il commence à s'effacer. Pour éviter la contamination par le virus vaccinal (vaccin vivant atténué), les lots témoins ont été élevés dans des bâtiments différents des lots vaccinés. |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||