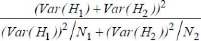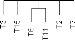Mise en place d'un suivi amphibien à Vohibola, l'une des dernières forêts humides de la côte Est de Madagascar( Télécharger le fichier original )par Ewald JAUSSAUD Université Joseph Fourier (Grenoble I ) - Master 2 pro biodiversité écologie environnement 2011 |

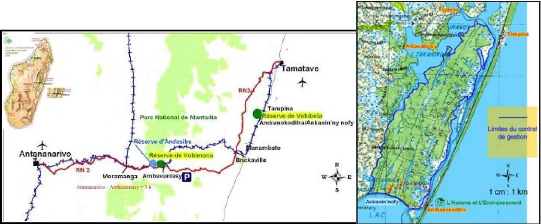
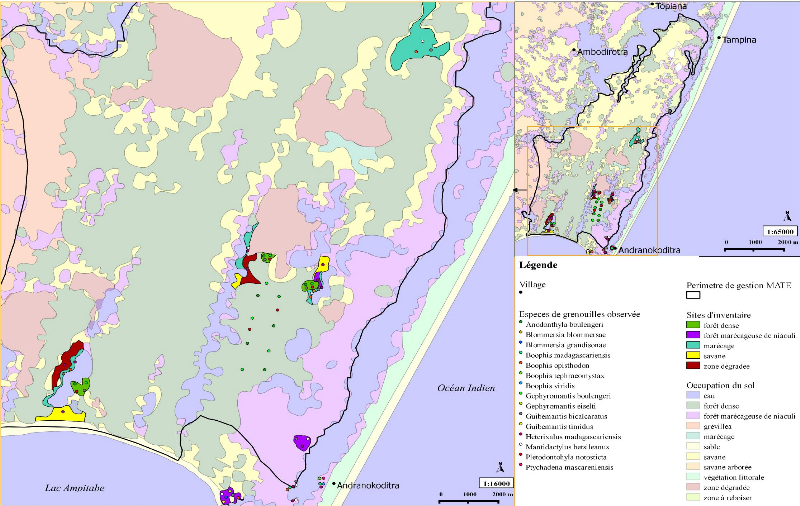
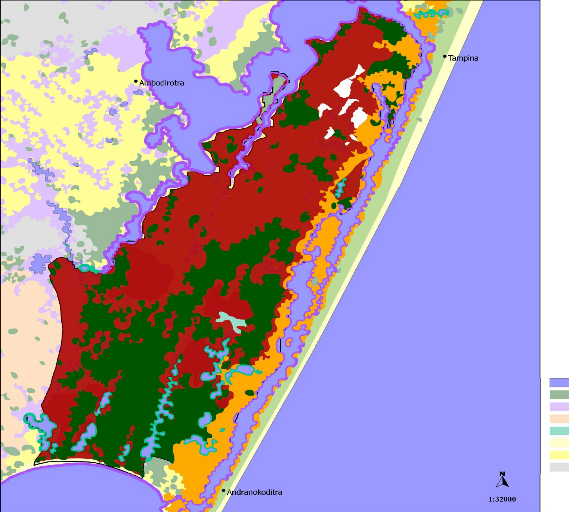
6 II Les amphibiens de la réserve de Vohibola1) La réserve expérimentale de VohibolaLa réserve de Vohibola est un territoire de plus de 2 000 ha, situé le long du canal des Pangalanes sur la côte Est de Madagascar, entre les villes de Tamatave (à 67 km au Nord) et de Brickaville (à 40 km au Sud). A un niveau local, elle est encadrée par les villages d'Andranokoditra, Tampina, Topiana et d'Ambodirotra (cf. Figure 3).
Figure 3 Localisation de la réserve de Vohibola Suite à la déforestation, qui a sévi sur la côte Est et qui a drastiquement réduit l'étendue de la forêt tropicale humide, la forêt de Vohibola constitue l'un des derniers fragments de forêt dense littorale de toute la côte Est. De ce fait, elle constitue un écosystème forestier et humide rare qu'il convient de préserver afin d'enrayer l'érosion de la biodiversité. On retrouve en effet, associée à cet écosytème, une biodiversité exceptionnelle qui se traduit par (Randrianirina & al., 2005 ; Missouri Botanical Garden, 2002) : - 7 espèces de lémuriens, qui sont endémiques de Madagascar, dont Eulemeur fulvus fulvus et Propithecus diadema respectivement « quasi-menacé » et « en danger » d'extinction selon les critères de l'IUCN (International Union for Conservation of Nature). - plus de 50 espèces d'oiseaux dont 50% sont endémiques et dont une espèce est considérée « vulnérable » tandis qu'une autre est jugée « en danger » par l'IUCN. - 19 espèces d'amphibiens, dont 1 « en danger » d'extinction et 2 commercialisées (Randrianirina & al., 2005) - 19 espèces de reptiles (Randrianirina & al., 2005), dont 10 espèces citées en Annexe 2 et 7 2 en Annexe 1 de la CITES (the Convention on International Trade in Endangered Species of Wild Fauna and Flora) qui réglemente voire interdit le commerce de ces espèces - 443 espèces de plantes endémiques de Madagascar dont 4 espèces d'arbres spécifiquement endémiques de Vohibola « en danger critique » d'extinction. Cette réserve représente donc l'un des derniers refuges possible pour de nombreuses espèces. Sa grande biodiversité fut d'ailleurs remarquée par le premier président de la république malgache qui en fit un conservatoire botanique strictement interdit à la coupe et y installa sa résidence secondaire. En 2001, le Ministère de l'Environnement, des Forêts et du Tourisme décida de la vendre pour en faire une concession forestière destinée à l'exploitation. L'Homme et l'Environnement proposa alors de la prendre en gestion pour assurer sa conservation tout en permettant aux populations locales d'en tirer profit (MATE, 2009). La forêt de Vohibola devient ainsi une réserve expérimentale (réserve naturelle protégée où sont menées conjointement des actions de conservation et des projets de développement). Cette forêt, qui fut très exploitée par le passé, si bien qu'aujourd'hui on n'y retrouve plus qu'une forêt secondaire assez jeune, est toutefois en bon état de conservation. Elle n'est pas habitée mais n'en est pas moins menacée par les pressions anthropiques des villages limitrophes (charbonnage, surexploitation des espèces à valeur économique comme l'ébène ou le bois de rose, commerce des animaux sauvages...) et par les feux qui peuvent être d'origine naturelle ou anthropique. Par ailleurs, selon les données de l'ONG, 90% de la population riveraine à la réserve vit sous le seuil d'extrême pauvreté, ce qui les rend totalement dépendants des revenus qu'ils tirent de l'exploitation de cette forêt. Bien qu'il existe des restrictions d'exploitation mises en place par l'Etat, celles-ci n'ont pas été accompagnées de solutions alternatives. Les villageois exploitaient donc "illégalement" leurs ressources naturelles immédiates (MATE, 2009). Dès lors, la forêt de Vohibola représente une zone prioritaire à gérer de manière durable afin d'allier préservation de la biodiversité et développement économique et social des villages avoisinants. C'est pourquoi MATE a poursuivi les restrictions d'exploitation de la forêt tout en proposant des alternatives économiques permettant de compenser le manque à gagner des populations locales. Elle a ainsi mis en place des projets tels que la visite guidée de la réserve et l'hébergement chez l'habitant pour ce qui concerne l'écotourisme et la production d'huile essentielle de niaouli (Melaleuca quinquennervia, plante invasive introduite le long du canal lors de sa construction afin d'en renforcer les berges) et de cannelle pour le développement économique. Côté conservation de la biodiversité, MATE a mis l'accent, depuis sa prise en gestion du site, sur le reboisement des zones dégradées, un incendie d'origine inconnue ayant ravagé une grande partie de la forêt en 2002. 8
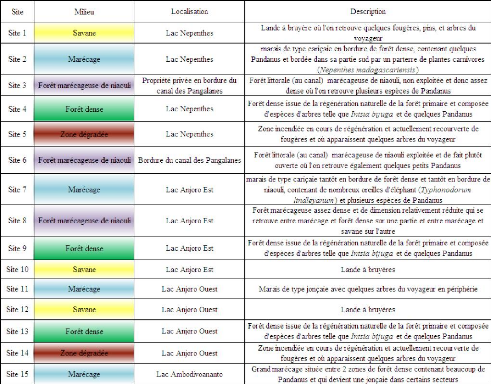
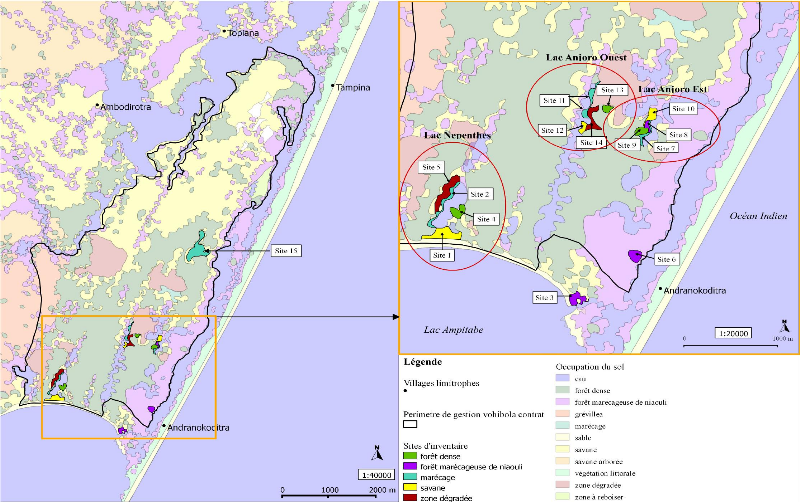
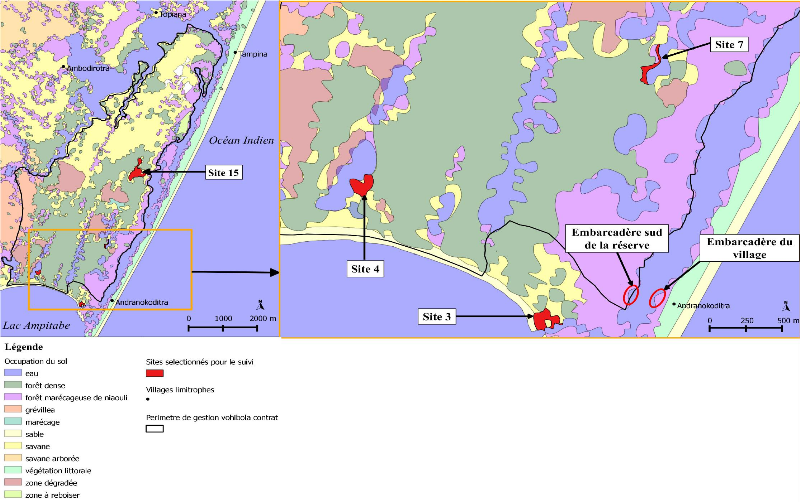
2) Les grenouilles de la réserveTous les amphibiens de Madagascar, appartiennent à l'ordre des Anoures (Glaw & Vences, 2007). De plus, il n'existe aucun crapaud, tous les amphibiens retrouvés à Madagascar, et donc à Vohibola, sont des grenouilles. Ce qui fait l'importance et l'originalité des grenouilles malgaches est le fait qu'il existe de nombreuses espèces et que celles-ci soient en grande majorité endémiques. On recense en effet 240 espèces endémiques sur un total de 242 espèces de grenouilles identifiées à ce jour. Madagascar se place ainsi au 4 ème rang mondial des pays les plus riches en espèces endémiques2. A Vohibola, sur la base des trois inventaires réalisés à ce jour (Randrianirina & al., 2005 ; Deschamps, 2007 ; Gehring & al., 2010), on retrouve potentiellement un total de 23 espèces de grenouilles dont : une espèce non endémique (Ptychadena mascareniensis), une espèce (Guibemantis bicalcaratus) citée en annexe III de la CITES et une espèce (Platypelis tetra) "en danger" d'extinction selon les critères IUCN. Toujours sur la base de ces données, et lorsque l'on s'intéresse à la systématique des amphibiens de la réserve, on constate (cf. Annexe III) que quatre des cinq familles d'amphibiens présentes à Madagascar se retrouvent à Vohibola et que les 23 espèces recensées se répartissent en 11 genres différents (soit presque la moitié de tous les genres représentés sur l'île). 2 Données IUCN, disponibles sur : http://www.iucnredlist.org/initiatives/amphibians/analysis/geographic-patterns 9 III Matériel et Méthode1) Sélection des sites d'inventaireLes sites à inventorier au sein de la réserve, dans le but de déterminer les hotspot d'amphibiens à suivre dans le futur, ont préalablement été sélectionnés par analyse cartographique. Cette étape a été réalisée à partir de la base de données cartographique de l'ONG étudiée avec le logiciel libre de cartographie SIG QuantumGIS (Qgis version 1.6.0). Cette base de données comprend notamment des informations sur le périmètre de gestion de la réserve par l'ONG et l'occupation du sol. La présélection des sites s'est faite en fonction de : - la présence d'un grand point d'eau (canal, lac, mare) dans les environs afin de maximiser le nombre d'observations, les amphibiens étant, de par leur mode de vie, intimement liés à l'eau. - la représentation des divers types de milieux (savane, zone dégradée, forêt dense composée d'espèces endémiques, forêt marécageuse de niaouli et marécage) autour de ces points d'eau afin d'inventorier l'éventail de diversité spécifique le plus large possible - l'accessibilité du site : le suivi doit pouvoir être continu dans le temps ce qui sous-entend des inventaires reproductibles. Pour cela le site doit pouvoir être accessible (accès par des chemins tracés plutôt que par traversée aléatoire de la forêt) et être retouvé aisément. Les différents types de milieux sont nommés selon la nomenclature choisie par MATE. Ainsi, on entend par savane une zone ayant subie une déforestation lors de l'incendie de 2002 et ayant été recolonisée par une espèce de bruyère appartenant au genre Philippia qui est une espèce invasive originaire d'Océanie. De même, une zone dégradée désigne également une zone incendiée mais dont la végétation est principalement représentée par une fougère (non invasive) et quelques arbres du voyageur (Ravenala madagascariensis) qui résistent aux incendies et poussent aisément en zone dégradée. Suivant les critères cités plus haut, un total de 15 sites a été prospecté. Parmi ces 15 sites (cf. Carte 1) : - on compte 4 marécages, 3 forêts denses, 3 forêts marécageuses de niaouli, 3 savanes et 2 zones dégradées - 12 sites sont regroupés autour de 3 lacs à raison de 4 sites par lac. Comme un seul de ces lacs est limitrophe d'une forêt marécageuse de niaouli, 2 sites de ce type de milieu ont été sélectionnés sur la côte orientale de la réserve qui borde le canal, c'est-à-dire là où on retrouve majoritairement le niaouli. Le 15ème et dernier site sélectionné est un grand 10 marécage choisi sur la base de sa potentielle richesse spécifique, mise en exergue par l'inventaire biologique de la réserve réalisé en 2005 (Randrianirina, 2005). - L'une des forêts marécageuses de niaouli retenue (site 3) ne se situe pas à proprement parlé dans la réserve bien qu'elle fasse partie du même territoire (cf. Carte 1). Il s'agit d'une parcelle en propriété privée. Le choix s'est porté sur ce site, car ne faisant pas partie de la zone de gestion de MATE, il n'est pas soumis à l'exploitation pour la production d'huiles essentielles. Par ailleurs, il n'est pas non plus exploité par les propriétaires de cette parcelle. Le deuxième site de ce type se trouve également en bordure de canal mais au sein de la zone de gestion et subit donc une exploitation. Ceci permet alors de distinguer deux sites d'un même habitat mais dont les propriétés d'exploitation ne sont pas les mêmes. Le troisième site correspondant à ce type de milieu se situe quant à lui à l'intérieur de la réserve et borde un lac. Il fait alors office de milieu de transition entre marécage et forêt d'une part et entre marécage et savane d'autre part. Le Tableau 1 résume l'ensemble de ces informations et reprend les caractéristiques principales de chaque site inventorié. Tableau 1 Description des sites d'inventaire
11 Carte 1 Localisation des sites d'inventaire
12 2) InventaireLa méthodologie d'inventaire des grenouilles dans la réserve de Vohibola se base sur le protocole standardisé proposé par Arnaud LeGoff (LeGoff, 2008) et validé par l'expert herpéthologue de Conservation International à Madagascar, Mr Nirhy Rabibisoa. Ce protocole préconise notamment la recherche des amphibiens par "chasse à vue", méthode la plus employée pour ce genre d'étude (Daly & al., 2008 ; Andreone & al ., 2000, 2006 ; Vallan et al ., 2000, 2002, 2004). La méthode des pitfalls (pièges à fosse) s'est quant à elle souvent avérée inefficace car elle ne permettait pas d'observer d'èspèces différentes de celles identifiées par chasse à vue (Andreone & al., 2000, 2006), et n'a donc pas été réalisée pour cette étude. Cette méthode avait d'ailleurs été expérimentée à Vohibola lors d'un inventaire en 2007, mais n'avait permis aucune capture (Deschamps, 2007). La méthodologie de la présente étude reprend alors les principaux éléments du protocole standardisé, avec toutefois quelques modifications : - Selon cette méthodologie, par exemple, il est demandé à l'expérimentateur de relever l'hygrométrie au moment de l'inventaire. Cependant, l'ONG ne disposant pas du matériel nécessaire, ce facteur n'a pu être mesuré. - Le protocole préconise également de relever, lors de la session d'inventaire, un paramètre appelé "distance à l'eau". Ce paramètre correspond à la distance du point d'eau le plus proche à laquelle se trouve un individu observé. Techniquement, sur le terrain ce paramètre est extrêmement difficile à mesurer en raison de ce qui peut-être désigné comme "point d'eau". En effet, dans une forêt dense, un simple receuil d'eau dans une coquille vide d'escargot peut constituer un lieu de ponte pour certaines espèces. Il devient dès lors impossible de savoir à quelle distance une grenouille se situe du point d'eau le plus proche. De ce fait, le paramètre "distance à l'eau" a été jugé non pertinent et n'a pas été pris en compte lors de cette étude. - Le protocole standardisé n'envisage qu'une identification à vue (selon des caractéristiques morphologiques) alors qu'ici l'identification est réalisée à la fois de manière visuelle et acoustique. - En ce qui concerne l'analyse des résultats, un indice de biodiversité supplémentaire (indice de Hill) a été calculé, en plus des indices recommandés (indices de Shannon et Simpson), afin d'affiner l'analyse de la diversité spécifique de chaque site d'échantillonnage. 13 Période d'inventaire Les inventaires se sont déroulés sur une période de trois mois, entre le 13 février et le 15 avril, à raison de trois sessions de quinze jours. Soit une première session du 13 au 27 février, une seconde entre le 16 et le 30 mars et une troisième et dernière session du 2 au 15 avril. Méthode Les inventaires de chaque site ont toujours été effectués en binôme, c'est-à-dire avec la participation d'un guide du village, employé de MATE. L'inventaire est réalisé par transect d'une heure (effort d'échantillonnage constant) avec fouille systématique des micro-habitats (litière, arbustes, flaques d'eau...). Un transect respectant une limite de temps est préféré à un transect soumis à une contrainte de taille afin de favoriser un même effort d'échantillonnage : 100m à travers une forêt dense ne représente pas le même effort de recherche que 100m dans une zone dégradée. Les transects sont réalisés sur chaque site de jour et de nuit. Tous les sites sont donc inventoriés six fois en tout : deux fois (jour et nuit) par session, sauf pour certains sites qui n'ont pas été trouvés lors de la première session et on donc été inventoriés quatre fois. Le jour, les prospections ont lieu entre 5h30 et 7h00, au lever du soleil, afin de détecter les espèces diurnes avant que le soleil ne soit trop haut dans le ciel et donc qu'il ne fasse trop chaud (les grenouilles ayant tendance à se protéger du soleil en se réfugiant dans des troncs d'arbre ou sous les feuilles par exemple, elles deviennent alors difficiles à détecter). L'inventaire de nuit quant à lui se déroule après le coucher du soleil, entre 18h et 20h, ce qui permet d'observer et surtout d'entendre les espèces nocturnes. Pour chaque site les paramètres suivants sont renseignés lors de l'inventaire (cf. Annexe IV) : - Date et heure - Météo et température - Coordonnées GPS du transect - Description du milieu - Données de l'inventaire à proprement parler : espèce observée, nombre d'individus, stade et sexe, mode de détection, mode d'identification et n° des photos. Au début de la session d'échantillonnage, un thermomètre est placé au sol afin de déterminer la température. La température n'est alors relevée sur la fiche d'inventaire qu'après avoir renseigné les autres données (Météo, coordonnées GPS...), soit environ 5 mn. Les coordonnées géographiques sont acquises grâce à un GPS Garmin etrex modèle venture HC. Lorsque toutes ces données sont notées, le chronomètre est démarré et la prospection le long du transect débute. 14 En pratique, dès qu'un individu est repéré, une ou plusieurs photos sont prises (afin de pouvoir identifier l'individu ultérieurement) puis, si possible, l'individu est temporairement capturé et placé dans le seau prévu à cet effet. L'identification se réalise alors quand le temps de recherche est écoulé afin de bien dissocier temps de recherche et d'identification et ainsi respecter un même effort d'échantillonnage entre les sites. Plusieurs critères contribueront alors à l'identification. 3) IdentificationNe disposant pas du matériel nécessaire pour prélever puis analyser un échantillon de tissu afin d'être sûr de l'identification d'une espèce, l'identification individuelle est réalisée soit à vue, c'est-à-dire par reconnaissance de caractères morphologiques caractéristiques, soit au chant. Les individus ayant été photographiés, ceux dont l'identification posait problèmes ont pu être examinés par le professeur Nirhy Rabibisoa. Identification à vue L'identification à vue est réalisée grâce au guide "A Field Guide to the Amphibians and Reptiles of Madagascar" de Glaw & Vence (3e éd, 2007). Plusieurs paramètres concourent à l'identification des grenouilles, notamment : - la morphologie - l'habitat - les couleurs (dos, flancs, ventre, pattes...) même si ce trait de caractère est parfois trompeur - Formule palmaire et l'articulation tibio-tarsale (cf. Annexe V) - Oeil (forme de la pupille, couleur de l'iris...) - Disposition des sacs vocaux et description des glandes fémorales pour les mâles Identification au chant L'identification au chant, bien qu'elle ne permette que de détecter les individus mâles, permet un inventaire de la diversité spécifique et se révèle particulièrement importante dans une zone où la majorité des espèces ne mesure pas plus de 3 cm et se camoufle très bien dans leur environnement. De plus, les grenouilles se retrouvent parfois à plus de 2m au dessus du sol (Glaw & Vence, 2007) et sont donc non observables, d'où un intérêt supplémentaire de l'identification acoustique. Celle-ci est réalisée par enregistrement des chants (appareil photo personnel) puis comparaison avec les chants de grenouilles potentiellement présentes dans la réserve. Celles-ci sont déterminées en fonction de la répartition des espèces décrite par Glaw & Vence dans leur guide et à partir de leur présence attestée par l'inventaire biologique de la réserve réalisé en 2005 (Randrianirina & al., 2005). Les chants ont été obtenus par téléchargement depuis la base de données disponible sur le 15 site internet AmphibiaWeb. Il devient alors possible de comparer les chants enregistrés sur le terrain avec les chants de référence. 4) Analyse des donnéesRemarque : Certaines observations ont été faites en dehors des phases d'inventaire sur site. Celles-ci ont été prises en compte notamment pour déterminer la biodiversité totale de la réserve, mais, du fait qu'elles aient été obtenues en dehors de la méthodologie de transect décrite plus haut, elles n'ont pas été retenues pour le reste des analyses. Les données obtenues lors de cet inventaire permettent en premier lieu de dresser la liste des espèces que l'on retrouve dans la réserve mais aussi de calculer des indices de diversité, de mettre en évidence des différences potentielles entre sites, de comparer leur degré de similarité et d'associer les espèces aux différents habitats. Toutefois avant de traiter les données obtenues, il est nécessaire de vérifier que l'échantillonnage ait été suffisant.
Afin de déterminer les sites que l'on peut considérer comme hotspots et qui serviront donc de sites de référence pour les suivis amphibiens ultérieurs, il est nécessaire de comparer à la fois les critères qualitatifs (diversité spécifique) et quantitatifs (abondance respective des différentes espèces) de chaque site échantillonné. Pour ce faire, il existe de nombreux indices permettant d'évaluer ces critères et donc de discriminer les sites les plus pertinents pour un suivi sur le long terme. Pour chaque site ont ainsi été calculés : la richesse spécifique, l'indice de Shannon, l'indice de Simpson et l'indice de Hill. Ces indices ont été calculés à partir du nombre moyen d'individus, de chaque espèce, recensés lors des 3 sessions d'inventaire, soit : le nombre total d'individus observés divisé par le nombre de transects effectués, sauf pour les espèces trouvées uniquement de jour ou uniquement de nuit, pour lesquelles le nombre total d'individus est divisé par le nombre de transects diurnes ou nocturnes selon les cas. 16 Richesse spécifique On entend ici par richesse spécifique le nombre total d'espèces recensé sur un site. La richesse spécifique observée est un indice simple, permettant d'illustrer les caractéristiques écologiques d'un milieu. Elle correspond au nombre total d'espèces rencontrées, mais elle ne tient pas compte des abondances relatives. La richesse spécifique observée n'est qu'une approximation du nombre d'espèces réellement présentes et dépend directement de la qualité de l'échantillonnage. Indice de Shannon L'indice de Shannon (H') évalue la diversité au niveau d'un site d'échantillonnage ou d'un habitat. Cet indice est un indicateur de l'équitabilité d'un milieu, il est compris entre 0 et l'infini. Il se calcule de la manière suivante : H' = Ó [(ni/N) . ln (ni/N)] avec ni = nombre d'individus de l'espèce i et N = nombre total d'individus H' est minimal (=0) si tous les individus du peuplement appartiennent à une seule et même espèce, H' est également minimal si, dans un peuplement chaque espèce est représentée par un seul individu. L'indice est maximal quand tous les individus sont répartis d'une façon égale sur toutes les espèces (Frontier, 1983 in Grall & Hily, 2003). C'est la comparaison des indices entre sites qui renseigne sur la plus ou moins grande diversité d'un site. Il est possible de comparer les valeurs de cet indice pour un même site d'une année sur l'autre. Indice de Simpson L'indice de Simpson mesure la probabilité que deux individus sélectionnés au hasard dans la population échantillonnée, appartiennent à la même espèce. Cet indice se calcule selon la formule : D = Ó[ ni.(ni -1) / N.(N-1) ] avec ni = nombre d'individus de l'espèce i et N = nombre total d'individus D varie entre 0 et 1, 0 représentant le maximum de diversité et 1 le minimum de diversité. Cet indice est d'autant plus faible que le nombre d'espèces est grand (plus il y a d'espèce, plus la probabilité de tirer 2 individus de la même espèce devient faible). Et pour un nombre fixé d'espèces, il est d'autant plus grand que la répartition des fréquences est équitable. 17 Toutefois, cet indice donne forcément un poids plus important aux espèces abondantes. L'ajout d'espèces rares ne modifie que modérément la valeur de D (Grall & Hily, 2003). De plus cet indice ne permet pas la comparaison d'années en années d'un même site. Indice de Hill L'indice de diversité de Hill est une mesure de l'abondance proportionnelle, qui permet d'associer les indices de Shannon et Simpson. L'équation de cet indice est : Hill = (1/D) / eH' 1/D = inverse de l'indice de Simpson et eH'= exponentielle de l'indice de Shannon C'est l'indice de Hill qui semble le plus pertinent dans la mesure où il intègre les deux autres indices. Toutefois, il peut être utile d'utiliser les trois indices conjointement afin d'en extraire un maximum d'informations et de mieux comprendre la structure des communautés (Grall & Hily, 2003). Cependant dans la mesure où l'indice de Shannon est le seul qui puisse permettre la comparaison au cours du temps (nécessaire pour un suivi sur le long terme), il est celui qui nous intéresse le plus et il est donc nécessaire de le calculer. De plus, lorsqu'il n'y a qu'une espèce sur un site, les indices de Simpson et de Hill ne peuvent être calculés. De ce fait, pour la suite de l'analyse, ce seront les indices de Shannon qui seront comparés. Les indices de chaque site sont calculés avec le logiciel de statistique R (version 2.7.2). Comparaison inter-sites des indices de diversité Zones incendiées vs zones intactes On cherche ici à savoir si il existe une différence significative, en terme de diversité, entre la moyenne des valeurs des indices de Shannon des zones incendiée et celle des zone intactes. On entend par zone incendiée une zone ayant subi les ravages de l'incendie de 2002, ces sites sont représentés par les zones de savane et zones dégradées. A l'inverse, on note zone intacte les sites n'ayant pas été détruits lors de cet incendie. Afin de déterminer si il existe une différence significative, une comparaison des intervalles de confiance de la moyenne est effectuée. Puis afin d'évaluer précisément la probabilité qu'une différence (si elle existe) aussi grande puisse être due au hasard (p-value), un test de comparaison de moyenne est réalisé (test d'hypotèse). En l'occurence, comme la moyenne (ì) des indices pour la zone incendiée est égale à zéro avec un écart-type (ó) nul, on utilisera le test de comparaison d'une constante à une moyenne, dont la statistique se calcule de la manière suivante : 18 e = (ì - a) / v (s2 / n) avec s2 = [ Ó (xi - u)2 ] / (n - 1) ì = moyenne des H' des zones intactes a = constante s2 = estimation de la variance de la population n = taille de l'échantillon (nombre de H') Hypothèse nulle : H0 = Il n'y a pas de différence significative entre 0 et la moyenne des indices de Shannon des zones intactes (les différences observées sont dues uniquement aux fluctuations d'échantillonnage). Hypothèse alternative : HA = La moyenne des indices de Shannon des zones intactes est significativement différente de 0 La normalité de la distribution des données est vérifiée par le test de Shapiro-Wilk réalisé à l'aide du logiciel R. Les intervalles de confiance ont également été calculés avec ce logiciel. Zone de collecte vs zone non exploitée On cherche à savoir si la différence entre les indices de Shannon du site 3 et du site 6 est significative. Ceci revient à voir si la diversité de la forêt littorale marécageuse de niaouli exploitée est significativement différente de celle de la forêt littorale marécageuse de niaouli non exploitée. Il est possible d'évaluer une différence significative entre deux indices de Shannon grâce à un test t de student modifié (Jayaraman, 1999). En réalité, la modification porte sur l'estimation de la variance qui est adaptée à l'indice de Shannon. On a alors : t = | H'1 - H'2 | / v (Var H'1 + Var
H'2) t = statistique du test H'1 et H'2 = Indices de Shannon des sites à comparer pi = ni / N où ni : nombre d'individus de l'espèce i et N : nombre total d'individus S = nombre d'espèces observées 19 Le calcul du nombre de degrés de liberté s'effectue selon la formule suivante :
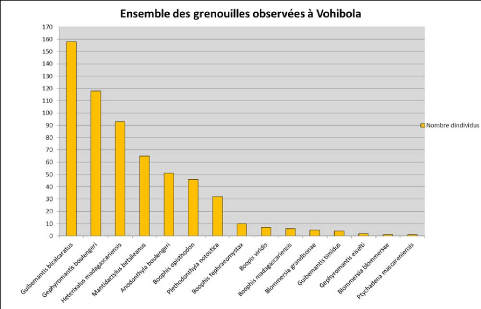
Les hypothèses du test sont : H0 = Il n'y a pas de différence significative de diversité entre les 2 sites HA = Il existe une différence significative de diversité entre les 2 sites c) Similarité des sites Un autre facteur étudié est la similarité des sites entre eux. Ce paramètre est estimé grâce à l'indice de Jaccard (Sj) qui permet de comparer entre 2 sites le nombre d'espèces communes par rapport au total des espèces recensées. Cet indice se calcule comme suit : Sj = c / (a+b-c) c = nombre d'espèces communes a = espèces du site a b = espèces du site b Cet indice varie entre 0 et 1. Il prend la valeur 0 lorsque les deux transects n'ont aucune similarité (aucune espèce en commun) et 1 lorsque la similarité est maximale (toutes les espèces sont en commun). A partir des indices de Jaccard obtenus pour chaque paire de site, il est possible de créer une matrice de distance. Cette matrice illustre la dissimilarité des sites entre eux (distance = 1 - Sj) et permet alors d'obtenir un dendrogramme regroupant les sites en fonction de leur plus ou moins grande similarité. Le calcul des indices de Jaccard, la matrice de distance ainsi que le dendrogramme sont obtenus grâce au logiciel R. 20 IV Résultats1) Nombre total d'espèces et d'individusAu total, c'est-à-dire pendant mais aussi en dehors des inventaires, 599 individus répartis en 15 espèces, ont pu être identifiés. La répartition de ces espèces au sein de la réserve est présentée sur la Carte 2. Le graphique 1 illustre quant à lui l'abondance des différentes espèces rencontrées dans la réserve de Vohibola. Graphique 1
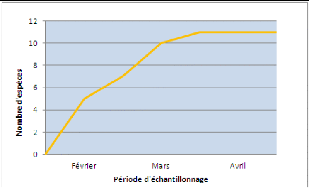
En réalité, sur les 15 espèces recensées lors de cet inventaire, 11 l'ont été sur les sites inventoriés et 4 en dehors de l'inventaire (donc hors méthodologie du transect) au niveau de l'embarcadère du village et de l'un des embarcadères de la réserve. On constate sur le graphique 1 qu'il existe de grandes disparités dans le nombre d'individus observés pour chaque espèce. Il est également intéressant de noter que l'espèce Guibemantis bicalcaratus, inscrite en Annexe III de la CITES et dont le commerce est contrôlé, est la plus représentée à Vohibola. 2) EchantillonnageLe plateau de la courbe d'accumulation totale, représentée sur le graphique 2, qui recouvre les 3 périodes d'inventaire (Février à Avril) amènent à penser que l'échantillonnage a été suffisant au
35 30 25 20 15 10 5 0 Heterixalus madagasca-riensis Guibemantis timidus Blommersia blommersae Guibemantis bicalcaratus Pletodontohyla notosticta Gephyromantis boulengeri Anodonthyla boulengeri Gephyromantis eiselti Blommersia grandisonae Mantidactylus betsileanus Boophis opisthodon cours de cette étude. En effet, à la fin du mois de Mars, l'ensemble des 11 espèces recensées avaient été identifiées. De plus, les courbes d'accumulations réalisées sur chaque site pris individuellement, semble confirmer que l'échantillonnage ait été suffisant lors de cet inventaire (cf. Annexe VI). Graphique 2 Courbe d'accumulation totale
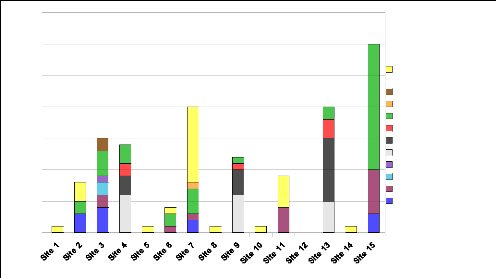
3) Abondance relative des espècesLe Graphique 3 illustre la diversité spécifique de chaque site échantillonné et la répartition du nombre des individus au sein de ces espèces. On observe que les espèces Heterixalus madagascariensis et Guibemantis bicalcaratus sont les plus représentées et que les mêmes espèces sont retrouvées sur les sites 4, 9 et 13 (forêt dense). De plus, la seule espèce retrouvée au niveau des sites 1, 5, 8, 10 et 14 est Heterixalus madagascariensis et ce, toujours dans les mêmes proportions. Ces données nous donne un premier aperçu de la similarité de certains sites (cette similarité inter-sites étant analysée de manière plus approfondie dans la partie 7). nb d'individus Graphique 3 : Abondance relative des espèces sur chaque site d'inventaire 22 Carte 2 Espèces de grenouilles retrouvées dans la réserve de Vohibola
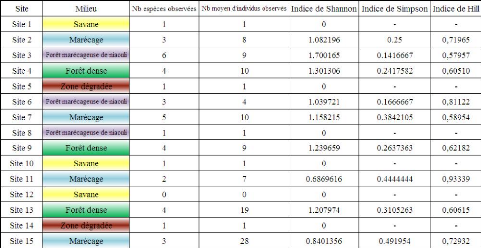
23 4) Diversité des sitesLes différentes composantes prises en compte dans l'évaluation de la diversité de chaque site échantillonné sont présentés par le Tableau 2. On constate que le site qui rassemble le plus grand nombre d'espèces est le site n° 3 tandis que le site n° 15 est celui qui contient le plus grand nombre d'individus. La comparaison des différents indices de diversité va permettre de discriminer les sites de manière plus précise et ainsi de sélectionner les plus intéressants pour un suivi amphibien. Tableau 2 Diversité des sites d'inventaire
Indices de Shannon Les zones de savanes et les zones dégradées (sites 1, 5, 10, 12 et 14) dans lesquelles une seule et même espèce a été retrouvée possèdent logiquement un indice de Shannon nul (H'=0) et s'avèrent alors être des milieux très pauvres en termes de diversité d'amphibiens. Le site le plus riche selon cet indice est la forêt marécageuse de niaouli non soumise à exploitation (site 3), viennent ensuite les zones de forêt dense (dans l'ordre croissant : sites 4, 9 et 13) puis les zones de marécage. On peut s'apercevoir que le site 7, bien que contenant plus d'espèces et autant d'individus, a une valeur de l'indice de Shannon plus faible que le site 4 (H'site7 = 1,158215 contre H'site4 = 1,301306). Ceci tient aux propriétés de l'indice de shannon qui rend compte de la répartition plus équitable des individus au sein des diverses espèces. Ainsi, le site 4 n'est pas plus diversifié que le site 7 mais ses individus se répartissent mieux entre les espèces (cf. Graphique 3). La forêt marécageuse de niaouli qui se situe dans l'intérieur de la réserve (site 8), tout comme les zones dégradées et de savane, n'est représentée que par une seule espèce présente en faible effectif. Son indice est donc nul. 24 Indices de Simpson Les indices de Simpson ne peuvent être calculés pour les sites ne contenant qu'une seule espèce, le dénominateur de l'équation étant égal à 0. L'indice pour ces sites est donc considéré comme maximal (D = 1), ce qui rend compte du minimum de diversité. Le site ayant l'indice de Simpson le plus faible est le site 3 illustrant ainsi la combinaison d'un nombre d'espèces plus important et d'une répartition équitable des individus entre les espèces. Toutefois, on constate que l'indice de Simpson du site 6 est faible et proche de celui du site 3 pourtant 2 fois plus riche en espèces, ce qui rend compte d'une répartition plus harmonieuse des individus. On remarque aussi que, excepté pour le site 3, les zones de marécage ont un indice relativement élevé (0,38 < D < 0,49) comparativement aux autres sites. Notamment, on observe que le site 4 a un indice plus faible que le site 7 qui contient pourtant plus d'espèces et autant d'individus. De même le site 4 possède un indice moins élevé que le site 13 qui regroupe plus d'individus pour autant d'espèces. Ceci illustre le poids important d'une meilleure répartition, des individus au sein des espèces, sur la diversité selon l'indice de Simpson (site 4 contre site 13) mais aussi le faible poids que cet indice apporte aux espèces rares (site 4 contre site 7). Indices de Hill La comparaison des indices de Hill permet d'avoir la meilleure vision sur la diversité d'un site. De la même manière qu'il était impossible de calculer les indices de Simpson pour les sites n'abritant qu'une seule espèce, l'indice de Hill n'est pas calculable pour les sites 1, 5, 8, 10, 12 et 14. Le Tableau 2 nous indique que le site 3 est le plus diversifié, il regroupe en effet le plus grand nombre d'espèces, un nombre d'individus moyen observé lors d'un transect assez important et montre que la répartition de ces individus est assez équitable. En se basant sur l'indice de Hill, le site 7 est le deuxième site le plus diversifié, et ce, malgré le fait que les indices de Shannon et Simpson de ce site montrent une diversité plus faible que les zones de forêt dense. Ceci reflète le fait que ce site possède à la fois plus d'espèces et autant d'individus (sauf pour le site 13), même si la répartition des individus entre les espèces présentes semble moins équitable sur ce site. Les sites de forêt dense (sites 4, 9 et 13) viennent ensuite avec un indice de Hill relativement similaire entre eux. Le site 13 bien que comptant autant d'espèces et beaucoup plus d'individus que les autres sites du même habitat, possède un indice de Hill plus grand (donc une diversité plus faible), ce qui démontre l'importance d'une répartition équitable des individus entre les espèces. De plus, le site 6, qui semblait plutôt diversifié par rapport aux autres lorsque l'on s'en remettait aux indices de Shannon et Simpson, s'avère en réalité peu diversifié selon l'indice de Hill. Cette conclusion semble logique pour un site ou l'on retrouve en moyenne seulement 4 individus 25 répartis en 3 espèces et illustre le poids trop important qui est donné à la répartition équitable des individus entre les espèces par les indices de Shannon et Simpson. Le même raisonnement s'applique pour les sites 11 et 15. ConclusionA partir de cette analyse, basée sur les indices de diversité, on peut conclure que les sites les plus diversifiés et donc les plus intéressants à suivre dans le futur sont les sites 3, 4, 7, 9 et 13. Cependant les sites 4, 9 et 13 représentant un même type d'habitat, on peut ne retenir que les sites 3, 4 et 7, le site 4 étant le plus diversifié. En effet, à eux 3 ces sites représentent l'ensemble du panel de diversité spécifique de la réserve recensé lors de cet inventaire. Toutefois ces indices ne constituent pas la panacée pour conclure sur les sites à sélectionner pour un suivi sur le long terme (voir ci-dessous : résultats complémentaires et le chapitre IV : Discussion). Ainsi d'autres éléments seront apportés afin de conclure définitivement sur ce sujet. 5) Résultats complémentairesComme souligné précédemment, 4 des 15 espèces recensées au total au cours de cet inventaire ont été retrouvées en dehors des sites sélectionnés. Il s'agit des espèces : Boophis madagascariensis, Boophis tephraeomystax, Boophis viridis et Ptychadena mascareniensis. On retrouve ces espèces au niveau de la mare et de l'embarcadère du village et au niveau de l'embarcadère qui se situe en face du village, au Sud de la réserve (cf. Carte 2). Ces espèces, et notamment B. tephraeomystax et B. viridis, sont connues pour être retrouvées dans des mares ou marécages à proximité des villages (Glaw & Vence, 2007). Ces informations n'ont pas été acquises en appliquant la méthodologie d'inventaire par transect décrite plus haut. Il s'agit d'observations ponctuelles réalisées lors des différents déplacements effectués entre le village et la réserve et donc acquises tout au long de la période d'inventaire. Ces données ne sont alors pas exploitables de la même manière que celles obtenues sur les sites d'inventaire (calcul des indices H', D et Hill) le dénombrement des individus n'étant pas réalisé dans les mêmes conditions. Elles donnent toutefois une vision de la richesse spécifique de ces zones qui reflète leur intérêt car elles regroupent un nombre important d'espèce au regard de la richesse des différents sites inventoriés mais surtout car certaines espèces n'ont été identifiées qu'à ces endroits (cf. Tableau 3). 26 Tableau 3 Espèces observées hors inventaire
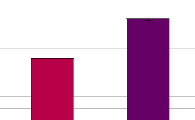
6) Comparaison inter-sites des indices de diversitéZones incendiées vs zones intactes L'indice H' de Shannon des zones ayant subies l'incendie de 2002 est constamment nul, la moyenne des indices est donc égale à 0, tout comme l'écart-type de la distribution (ì1=0; ó1=0). A l'inverse, la moyenne des indices de Shannon pour les zones intactes est différente de 0 (ì2=1.03; ó2=0.45) . La comparaison au niveau des intervalles de confiance à 99%, semble montrer que cette différence de moyenne est significative. En effet, on obtient un intervalle de confiance à 99% = [0.5619551.489303], ce qui signifie que la valeur moyenne réelle des indices de Shannon des zones intactes à 99% de chances de se situer entre ces valeurs (0.561955< ì2 <1.489303). Donc, au risque á = 1% de chances de se tromper, on peut conclure que les zones non impactées par l'incendie sont plus diversifiées que les zones ayant subies cet incendie de 2002. La distribution des indices de Shannon pour les zones intactes, suivant une loi normale (W = 0.9039, p-value = 0.2417) Ce résultat est confirmé lors du test de comparaison d'une moyenne à une constante (t=6,82 > t9,0,05=2,262 ; p<0,01). Zone de collecte vs zone non exploitée Les sites 3 et 6 représentent initialement des milieux similaires (forêt littorale marécageuse de niaouli) mais diffèrent au niveau de leur exploitation. On observe en moyenne 4 individus répartis en 3 espèces sur le site 6. On obtient alors H'site6=1.039721. 27 Pour le site 3, on observe en moyenne 9 individus par transect pour 6 espèces, ce qui donne H'site3=1.700165. Un test de Student modifié révèle (cf. Graphique 4) qu'au seuil de risque a = 10%, cette différence entre les indices de Shannon est significative (t=2,067 > t5,0,10=2,015), ce qui pourrait sous-entendre un impact de la collecte des feuilles de niaouli sur la diversité amphibienne, bien que la relation de causalité ne puissent être révélée par un test de comparaison de moyenne. Graphique 4 Comparaison des indices de Shannon en fonction de l'exploitation du site 2 1,8 1,6 1,4 1,2 1 0,8 0,6 0,4 0,2 0
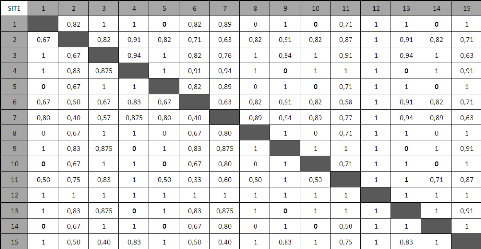
* ZC ZNC ZC = Zone collectée (site 6) et ZNC = Zone non collectée (site 3) 7) SimilaritéA partir du calcul des indices de Jaccard pour l'ensemble des sites pris deux à deux, on obtient la matrice de distance suivante (Tableau 4). Lors de l'observation de cette matrice, on remarque tout de suite que de nombreux sites sont soit fortement différents (indice de distance = 1) soit fortement similaires (indice de distance =0) entre eux. On constate notamment que tous les sites de forêts denses abritent exactement les mêmes espèces et que la seule et unique espèce retrouvée dans les savanes et zones dégradées est toujours la même. On s'aperçoit également que le site 12, où aucune espèce n'a pu être recensée, est logiquement distant de tous les autres sites qui acceuillent tous au moins une espèce de grenouille. Cette matrice des distances permet alors la construction d'un dendrogramme illustrant la répartition des groupes en fonction de leur degré de similarité (cf. Graphique 5 où T, comme transect, représente un site. Ex : T1 = site 1). 28 Tableau 4 Matrice de distance
Ce graphique confirme tout d'abord le regroupement des sites de forêt denses d'un côté et des sites de savanes et zones dégradées de l'autre. On remarque que le site 8, qui est la forêt marécageuse de niaouli situé dans l'intérieur de la réserve, est englobé dans le groupe des sites mono-spécifiques. En effet, une seule espèce y a été retrouvée et il s'agit de la même qu'en zone dégradée et qu'en savane. On remarque ensuite le regroupement des zones marécageuses (forêts de niaouli et marécages) qui partage un degré de similarité d'environ 40%. Ces données permettent donc globalement de regrouper les sites en fonction de l'habitat qu'ils représentent. Il est alors possible de déterminer les habitats spécifiques de certaines espèces au sein de la réserve de Vohibola. Graphique 5
Dendrogramme de similarité
29 8) HabitatA partir des données de l'inventaire (cf. Graphique 3) et des informations issues de l'analyse de la similarité des sites, on peut associer une diversité spécifique à un habitat donné. Plus particulièrement, on peut cibler les espèces spécifiquement retrouvé dans un type d'habitat donné au cours de l'inventaire (noms en gras sur le Tableau 5) Tableau 5 Association Habitat-Espèces
Ainsi, on peut voir que certaines espèces sont assez généralistes comme Heterixalus madagascariensis qui vit dans tous les types de milieux de la réserve (zones marécageuses, zones dégradées et de savanes) excepté la forêt dense, ou encore comme Guibemantis bicalcaratus qui se retrouve là où il y a des Pandanus, c'est-à-dire partout sauf dans les zones de savane et les zones dégradées. En effet cette espèce a systématiquement été observée dans des zones à Pandanus, qui constituent son habitat privilégié (Glaw & Vence, 2007). En revanche, il existe aussi des espèces spécialistes d'un seul habitat telles que Anodonthyla boulengeri, Gephyromantis boulengeri et Plethodontohyla notosticta qui ne vivent qu'au sein de la forêt dense. 30 Il est alors envisageable que la présence de ces 3 espèces puisse attester de la bonne santé de ce type d'écosystème et que ces espèces puissent donc être considérées en tant qu'espèces bioindicatrices. En reprenant l'ensemble des informations mises en évidence dans cette étude concernant les espèces et le type de milieu dans lesquelles elles ont été observées et en extrapolant ces données à l'ensemble de la réserve de Vohibola, il est possible d'établir une cartographie théorique des habitats pour les grenouilles (cf. Carte 3).
Carte 3 Répartition théorique des espèces de grenouilles au sein de la réserve de Vohibola Boophis madagascariensis Boophis tephraeomystax Guibemantis bicalcaratus Mantidactylus betsileanus 0 Blommersia blommersae Guibemantis bicalcaratus Mantidactylus betsileanus Anodonthyl a boulengeri Guibemantis bicalcaratus Blommersia grandisonae Gephyromantis eiselti Guibemantis timidus Mantidactylus betsileanus Hetrixalus madagascariensis Villages limitrophes Perimetre de gestion Occupation du sol eau forêt dense forêt marécageuse de niaouli grévillea marécage sable savane savane arborée M! végétation littorale zone dégradée zone à reboiser Boophis opisthodon Boophis viridis Heterixalus madagascariensis Ptychadena mascareniensis Boophis opisthodon Heterixalus madagascariensis Gephyromantis boulengeri Plethodontohyla notosticta Boophis opisthodon Guibemantis bicalcaratus Heterixalus madagascariensis 31 32 V DiscussionIdentification des individus Ne disposant pas du matériel nécessaire à une identification grâce aux tissus et n'étant pas un expert herpétologiste, il subsistera toujours un doute quant à l'identification des espèces recensées lors de cet inventaire. Cependant l'analyse selon la répartition géographique proposée dans le guide de terrain de référence (Glaw & Vence, 2007), les différents indices visuels présentés dans la partie identification de la méthodologie (cf. II) et l'association du chant à ces indices permettent de penser que les erreurs d'identification, bien que possibles, ont été marginales. Comparaison des résultats avec la littérature : quelles espèces de grenouille à Vohibola? L'étude réalisée ici a permis de mettre en évidence la présence de 15 espèces de grenouilles. Un inventaire réalisé en 2005 (Randrianirina & al., 2005) fait quant à lui état de la présence de 19 espèces d'amphibiens à Vohibola. Deux autres études, menées en 2007 (Deschamps, 2007) et en 2010 (Gehring & al, 2010), recensent quant à elle respectivement 10 et 11 espèces dans la réserve. Le Tableau 6 illustre la diversité spécifique rapportée par chacune de ces études. Il apparaît clairement de nombreuses dissimilarités dans les résultats de ces différents inventaires, par exemple l'espèce Platypelis tetra (en danger d'extinction selon l'IUCN) n'a été rapportée que lors de l'inventaire de 2005. Si l'on prend en compte toutes les espèces recencées lors de chaque inventaire, la diversité totale des amphibiens de Vohibola s'élèverait à 27 espèces. Cependant, comme le soulignent Gehring et son équipe (Gehring & al, 2010), il existe sans doute quelques erreurs d'identification lors des inventaires précédents, notamment pour les espèces : Boophis idae, Boophis mandraka, Guibemantis tornieri, Platypelis grandis et Platypelis tetra. En effet, en s'intéressant à la répartition théorique de ces espèces (Glaw & Vence, 2007 et site internet AmphibiaWeb), il semble peu probable de pouvoir les retrouver dans la réserve de Vohibola. Une des erreurs d'identification possibles concerne l'espèce Guibemantis tornieri. Lors du présent inventaire, cette grenouille n'a pas été recensée mais une espèce qui lui ressemble beaucoup du point de vue morphologique a en revanche été observée. Il s'agit de Guibemantis timidus dont la répartition géographique semble beaucoup plus en adéquation avec une localisation à Vohibola (Glaw & Vence, 2007 et site internet AmphibiaWeb). Il est donc probable qu'il y ait eu confusion entre les 2 espèces au cours des inventaires de 2005 et 2007. 33 Tableau 6 Ensemble des espèces recensées à Vohibola entre 2005 et 2011
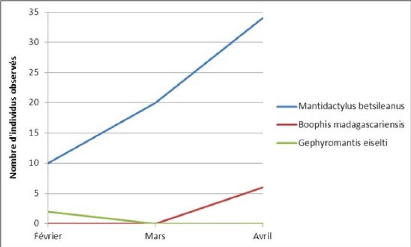
* En plus des 10 espèces du tableau Gehring identifie une sous-espèce de Gephyromantis boulengeri qui n'est pas notée ici car, faute de moyens, le niveau d'analyse de cette étude ne peut aller jusqu'à la sous-espèce et s'arrête donc à l'espèce. Etant donné que les espèces identifiées en 2010, l'ont été à partir d'analyses ADN, on part du principe qu'il existe une base de 10 espèces (et une sous-espèce) dans la réserve expérimentale de Vohibola. Si on ajoute à cela les autres espèces recensées, en retranchant à la fois les espèces mises en avant par Gehring (Gehring & al., 2010) comme étant susceptibles d'être des erreurs 34 d'identification et les espèces qui n'ont été recensées que dans une seule étude, on obtient une richesse spécifique potentielle de 14 espèces (plus une sous-espèce). On remarque également (cf. Tableau 6) que les espèces Guibemantis liber et Guibemantis pulcher n'ont été identifiées que lors de l'étude de 2010 (Gehring & al., 2010). Ceci peut s'expliquer par le fait que la distinction de ces espèces avec Guibemantis bicalcaratus est assez difficile (taille, morphologie, couleurs et habitat similaires) et pose un problème d'identification qu'une analyse ADN permet de résoudre aisément. Dans tous les cas, si les moyens le permettent, il serait intéressant, lors des futures études sur les amphibiens de Vohibola, de prélever des tissus sur les grenouilles afin de permettre l'identification sûre et définitive des espèces présentes, car à ce jour la biodiversité réelle des amphibiens de Vohibola n'est toujours pas totalement connue. Peut-être les analyses de tissus seraient-elles possibles grâce à un partenariat entre MATE et le département biologie de l'université d'Antananarivo avec laquelle plusieurs chercheurs ont déjà collaborés (Gehring & al, 2010 ; Crottini & al, 2011). Période d'inventaire Le présent inventaire a été réalisé entre février et avril 2011, ce qui ne correspond pas tout à fait à la période la plus favorable pour l'observation des amphibiens. Celle-ci se situant plutôt entre décembre et mars qui correspond à la saison chaude et pluvieuse dont les amphibiens profitent pour se reproduire. De ce fait, on peut penser que certaines espèces n'ont pas été rencontrées lors de l'inventaire car celles-ci avaient déjà accompli leur cycle de reproduction. Cette hypothèse de décalage temporel du cycle entre les espèces semble se confirmer lorsque l'on analyse l'évolution du nombre d'individus de quelques espèces au cours du temps (cf. graphique 6). On peut observer que certaines espèces voient augmenter leur effectif observé comme Mantidactylus betsileanus, que d'autres n'ont plus été observées après février (ex : Gephyromantis eiselti) et qu'à l'inverse d'autres n'ont été recensées qu'à partir du mois d'avril (ex : Boophis madagascariensis). On ne peut donc pas exclure la possibilité que d'autres espèces que celles rencontrées au cours de cette étude, aient pu déjà effectuer leur période de reproduction avant le début de l'inventaire et puissent donc être présentes sur le site de Vohibola.
35 Graphique 6 Evolution du nombre d'observations de 3 espèces au cours du temps L'idéal pour le suivi des amphibiens de la réserve serait alors de commencer les futurs inventaires plus tôt, c'est-à-dire en décembre, car la période de reproduction qui correspond au pic d'activité des grenouilles malgaches est constituée par la saison des pluies qui s'étale de décembre à mars (Glaw & Vences, 2007). Toutefois, il paraît important de poursuivre les inventaires au cours du mois d'avril, tout du moins pour ce qui concerne la réserve de Vohibola, certaines espèces n'ayant pu être observées qu'à partir du mois d'avril (cf. Graphique 6). En conclusion, la période d'inventaire la plus propice pour la réalisation d'un suivi amphibien efficace à Vohibola devrait se situer entre début décembre et fin avril. Les sites de suivi L'analyse et la comparaison des résultats en termes de diversité des sites a permis de mettre en évidence la richesse et donc l'intérêt des sites n° 3, 4 et 7 pour le suivi amphibien que souhaite mettre en place MATE dans la réserve de Vohibola. En plus de ces 3 sites, on a également pu voir qu'il existait d'autres sites intéressants de par la biodiversité particulière qu'on y retrouve, il s'agit des zones d'embarcadères (et en particulier la mare) du village et de la réserve (cf. Carte 4). Certaines espèces observées ont en effet été retrouvées uniquement sur ces sites (cf. Tableau 3). De plus, l'inventaire réalisé en 2005 dans la réserve (Randrianirina & al., 2005) avait déjà mis en exergue cette zone, notamment en raison de la présence de l'espèce Boophis tephraeomystax qui, tout comme dans notre étude, n'avait été observée que sur ce site à l'époque. Toutefois, il existe une divergence assez importante en termes de composition spécifique recensée dans cette zone entre la présente étude et l'inventaire de 2005. Un examen plus approfondi de ce site serait donc particulièrement intéressant. Par ailleurs, même si ils ne constituent pas des hotspots de biodiversité, il serait intéressant de poursuivre le suivi dans les savanes et zones dégradées afin d'étudier la recolonisation du milieu par les amphibiens et peut-être corréler cette recolonisation (si recolonisation il y a) à l'évolution du milieu. Ainsi, il serait possible de suivre en parallèle l'évolution du milieu et de la biodiversité. Enfin, le site 15 mérite aussi une attention particulière car beaucoup d'espèces différentes (14 au total)3 y ont été recensées lors de l'inventaire biologique de 2005 (Randrianirina & al., 2005), et même si il y a probablement eu des erreurs d'identification (voir ci-dessus), il serait intéressant d'identifier les espèces présentes. D'autant plus que les espèces citées dans ce rapport ne ressemblent pas morphologiquement aux espèces recensées au cours du présent inventaire. Ces dernières, qui avaient d'ailleurs également été recensées en 2005, ont peu de chances d'avoir été confondues avec tant d'autres espèces. De ce fait, il est plausible que le site 15 (et la forêt qui l'entoure) recèle une biodiversité amphibienne plus importante que celle mise en évidence dans la présente étude. Ce résultat peut-être dû à la période tardive d'inventaire, ce site n'ayant pas été trouvé lors de la première session d'inventaire en février. En conclusion, 6 sites seront retenus pour le suivi amphibien (cf. Carte 4). Il est bien sûr possible et même souhaitable de rajouter des sites intéressants à cette liste, la réserve n'ayant pas pu être explorée dans sa totalité. 36 3 En 2005, l'inventaire s'est déroulé sur un site qui recouvrait à la fois forêt dense et marécage. Ici, seul le marécage a été exploré. 37 Carte 4 Sites sélectionnés pour le suivi amphibien de la réserve
38 Impact de la collecte : quelle gestion pour le niaouli? Le niaouli est une plante invasive introduite lors de la construction du canal des Pangalanes et qui aujourd'hui couvre une part importante (environ 300 ha) de la réserve. Cependant, bien que cette plante soit invasive, les zones marécageuses composées par les niaoulis constituent aujourd'hui un milieu à part entière, présent depuis près de cent ans (ce qui représente de nombreuses générations d'amphibiens). On peut alors s'interroger quant à la gestion à appliquer à ce milieu : il faut à la fois éviter l'extension du niaouli à l'intérieur de la réserve car il entraînerait une uniformisation des zones humides, et à la fois tenter de préserver un bon état des forêts de niaouli qui, comme on a pu le voir, constitue un habitat pour de nombreuses espèces de grenouilles. Bien que les résultats ne puissent être significatifs en raison d'un échantillon faible (2 sites seulement ayant été échantillonnés), on peut tout de même penser que ces résultats illustrent une tendance (à confirmer par d'autres études) et que la surexploitation pour la production d'huile essentielle représente un danger de dégradation du milieu avec un impact potentiel fort sur la faune. En effet, au niveau des zones de collecte, seules 3 espèces de grenouille peuvent être observées contre 6 espèces en zone non exploitée. De plus, à titre indicatif, dans la partie privée de la réserve (zone non exploitée), il n'est pas rare d'observer de nombreux lémuriens (Eulemur fulvus) dans la forêt de niaouli, or on a du mal à imaginer ces lémuriens sur une parcelle de ce même milieu, mais où la taille des arbres ne dépasse pas 2,5 m (contre plus de 4m pour les individus non exploités) et dont les feuilles ont quasiment toutes été prélevées. Afin de déterminer de manière plus précise l'impact de la collecte sur les zones humides à niaouli, une étude serait particulièrement intéressante à réaliser. Il serait par exemple possible d'échantillonner les sites de collecte et plusieurs zones intactes afin de comparer la diversité spécifique et le nombre d'individus que l'on retrouve sur ces sites (pour plus d'information, l'étude pourrait également être menée sur le reste de la faune retrouvée dans ce milieu). En ce qui concerne la gestion au niveau des zones de collecte, MATE a mis en place des "bonnes pratiques de collecte" censées permettre la régénération de ces zones et donc une exploitation durable du milieu. Mais si l'interdiction de coupe des troncs est respectée, la recommandation de prélever uniquement les feuilles afin d'éviter l'arrachage des branches quant à elle, n'est pas toujours suivie d'effets. En réalité, il est difficile, pour les populations concernées, d'appliquer ces "bonnes pratiques". Tout d'abord, car il n'existe pas de contrôle lors des phases de collecte et ensuite car le prix du kilo de feuilles ne le permet pas. Une étude, demandée par MATE, sur les impacts socio-économiques de la production des huiles essentielles à Vohibola (Chlémaire, 2011), estime qu'au tarif auquel sont 39 payés les collecteurs/collectrices (36 Ar4/kg), il leur est impossible de répondre à cette pratique étant donné la chute de revenus que cela occasionnerait. En effet, les branches sont arrachées afin de défolier plus rapidement car les collecteurs/collectrices ne disposent pas d'escabeau pour un effeuillage en hauteur. La défoliation sans arrachage des branches entraineraît alors une augmentation du temps nécessaire à la collecte d'un kilo de feuilles et donc une diminution des revenus. De plus, il ne faut pas oublier que ce projet est réalisé dans le cadre du commerce équitable, or, si les bonnes pratiques sont respectées et que cela induit une diminution des revenus pour les travailleurs locaux, le projet risque de ne plus répondre aux normes du commerce équitable (Chlémaire, 2011). En effet, MATE est tenue de payer ses travailleurs au salaire minimum légal (environ 90 000 Ar / mois) et se base sur les paramètres suivants pour la rémunération : - Un mois correspond à 25 jours de travail - Les collecteurs ramassent 100 kg/jour De ce fait, pour que les revenus s'alignent sur le salaire minimum malgache, le prix du kilo de feuilles est fixé à 36 Ar (36x100 = 3600 x 25 = 90 000). Or, si les collecteurs cueillent moins de 100 kg/jour (ce qui serait le cas si la collecte se faisait sans arrachage des branches) ils ne pourront plus toucher l'équivalent du salaire minimum mensuel. Cependant une augmentation du rendement en huile essentielle est possible si seules les feuilles sont collectées, ce qui permettrait à la fois une augmentation du prix au kilo et le respect des bonnes pratiques (Chlémaire, 2011). Il serait donc important de veiller au respect des bonnes pratiques pour une gestion durable du niaouli mais ceci ne peut se faire au détriment des revenus des villageois. Il faudrait alors trouver un système de rémunération plus approprié à cette activité. Isolation de la réserve de Vohibola La réserve expérimentale de Vohibola représente l'un des derniers fragments de forêt littorale humide de l'est de Madagascar, sa préservation est donc primordiale d'un point de vue environnemental. Des projets de protection sont justement mis en oeuvre. MATE entend ainsi intégrer la réserve de Vohibola au Réseau des Aires Protégées de Madagascar et inclure sa zone humide dans la convention Ramsar afin de renforcer son statut de protection, la réserve ne bénéficiant pour l'instant que d'un statut de réserve expérimentale. Mais le fait que ce type d'écosystème soit si rare implique une isolation par rapport au reste des écosystèmes semblables et en bon état de conservation. La forêt de Tampolo constitue la forêt littorale humide plus proche et se situe à plus de 150 km au nord du site de Vohibola. Cette forêt forme un habitat similaire à celle de Vohibola, et on y recense un richesse en amphibiens 4 L'ariary (Ar) est la monnaie malgache : 1€ équivaut à environ 2 800 Ar (en 2011) 40 comparable (16 espèces) mais avec quelques différences lorsque l'on s'intéresse aux espèces retrouvées : 9 espèces sont communes aux deux sites (Randrianirina & al., 2005). Ceci signifie alors peut-être, qu'en cas de connexion de ces deux fragments d'un même écosystème, une biodiversité d'amphibien encore plus importante pourrait être retrouvée à Vohibola mais aussi à Tampolo. Cependant établir une connexion écologique entre ces deux fragments sur une distance supérieure à 150 km paraît difficile, d'autant que la politique environnementale malgache s'axe principalement sur la création d'aires protégées pour répondre aux recommandations de l'IUCN. Pour l'instant il vaut donc mieux se concentrer sur la bonne gestion de ces zones avant de penser à la création de corridors écologiques. Conclusion : recommandationsPour parvenir à assurer un suivi fiable, le guide du village employé de MATE, Nabé, a été formé à la méthode d'inventaire et à l'identification précise des espèces d'amphibiens. Un porte-vue réalisé spécialement pour le suivi des amphibiens de Vohibola lui a été remis sur le terrain, et un autre exemplaire a été remis au bureau de MATE à Antananarivo. Ce document contient : - des fiches d'aide à l'identification des espèces (clé de détermination) classées par type de milieu; - la liste des espèces retrouvées et de celles que l'on peut potentiellement retrouver; - une fiche d'inventaire reprenant les paramètres à noter et une fiche contenant les indices à receuillir pour une identification ultérieure d'un individu non capturé; - Un CD contenant les chants des amphibiens présents et potentiellement présents accompagné d'une liste de référence associant une espèce à un numéro de piste sur le CD. Ces documents forment une première base de travail, mais avec un peu de matériel supplémentaire il serait facile de rendre le suivi beaucoup plus efficace. En effet, sur le terrain il n'y a pas d'appareil photo en état de marche et à la disposition du guide. De ce fait, l'identification devra obligatoirement se faire sur place ce qui réduit la précision des données, étant donné que les individus observés ne sont pas toujours capturés. De plus, l'achat d'un dictaphone pour le suivi s'avérerait particulièrement pertinent pour permettre d'enregistrer les chants dont on a démontrer l'intérêt pour réaliser une étude. Mais, en plus du matériel nécessaire, cette étude à également permis de mettre en avant des éléments de réflexion dont la prise en compte pourrait permettre un suivi des amphibiens d'autant plus fiable. Le Tableau 7 présente l'ensemble des mesures préconisées pour un suivi efficace sur le long terme des amphibiens de Vohibola. 41 Tableau 7 Préconisations pour le suivi amphibien à Vohibola
Valorisation et développement durable MATE tente au travers de son intervention de concilier développement économique et social et protection de l'environnement, ce qui correspond à la notion de développement durable. MATE soutient en effet que les populations doivent pouvoir tirer profit de leurs ressources naturelles locales mais de manière raisonnée afin de permettre la préservation du milieu dont elles tirent les ressources. Ceci apparaît alors comme une action totalement légitime car l'Homme, aussi bien que toute autre espèce, doit pouvoir tirer parti des ressources de son environnement immédiat. Toutefois, plusieurs problématiques, sur lesquelles une réflexion est initiée ici, peuvent alors émerger : Tout d'abord, comme nous avons pu le voir plus haut, il existe un risque de surexploitation du milieu pour le niaouli. Il est donc primordial de mettre en place à la fois un système de surveillance permettant une exploitation réellement raisonnée de cette ressource et un système de rémunération des collecteurs plus approprié au respect des bonnes pratiques de collectes. De plus, la mise en place d'un suivi scientifique comme conseillé suite aux résultats de cette étude sur les amphibiens, serait un outil pertinent à la gestion du niaouli. 42 De plus, le projet huile essentielle, qui est le programme phare de MATE, s'inscrit dans le cadre du commerce international. En effet, on comprend facilement que les huiles essentielles et les produits cosmétiques dérivés de ces huiles ne constituent pas les besoins premiers d'une population dont environ 70%5 de la population vivait encore sous le seuil de pauvreté national en 2005. Quel problème y a-t-il au commerce international de marchandises si cela permet l'amélioration de la qualité de vie de la population? Dans le fond aucun, car l'objectif semble tout à fait louable. Il est juste surprenant d'observer qu'un organisme qui se pose en protecteur de l'environnement à une échelle locale, oublie ainsi l'impact de son activité, en terme de bilan carbone notamment, sur le reste de l'environnement à l'échelle internationale. Ne résout-on pas un problème localement en l'aggravant à l'échelle globale? Ceci, en particulier lorsque de nombreuses études scientifiques démontrent la diminution significative de l'impact de l'Homme sur l'environnement dans le cas d'échanges à une échelle locale contre la logique de commerce international actuelle (IPCC, 2001). Ensuite, le concept de valorisation sous-entend que la biodiversité est un bien qui nous appartient et qui doit nous être utile. En extrapolant : si la technologie permettait de subvenir à tous nos besoins aurait-on pour autant le droit de détruire l'environnement? De plus, il est probable que les villageois qui travaillent dans les projets de MATE ne voient dans l'environnement qu'une source de revenu. Ils n'auraient donc théoriquement aucun problème à faire une activité destructrice de l'environnement si cela leur rapportait un revenu plus élevé. Cette politique est-elle réellement durable, c'est-à-dire viable sur le long terme? Ne risque-t-on pas en associant une valeur économique à certaines ressources naturelles, de créer un nouveau marché nécessairement soumis à la loi de l'offre et de la demande, qui peut, tout en conduisant à de réels progrès sociaux (développement d'écoles ou d'infrastructures sanitaires par exemple), provoquer une surexploitation de l'environnement que l'on cherche à protéger. Car que faire si les huiles essentielles bio et équitables deviennent les produits "à la mode"? Limitera-t-on alors la production afin de protéger l'environnement devant la pression du capital économique et des avancées sociales sous-jacentes que cela représente? Rien n'est moins sûr, surtout face à une population ayant tiré les bénéfices de cette activité et dont les besoins auront évolués. En poursuivant cette logique, on peut également se demander : comment empêcher l'augmentation de la pression qu'exerce l'Homme sur l'environnement lorsque le développement économique est synonyme de croissance, donc d'augmentation de la production, du niveau de vie, de la taille des populations et nécessairement de la consommation qui les accompagne? Ceci amène finalement, à un aspect essentiel qui n'est pas, du moins pas encore, totalement traité par MATE dans sa gestion de la réserve : la sensibilisation à l'environnement. C'est-à-dire, 5 source : http://donnees.banquemondiale.org/pays/madagascar 43 permettre une réflexion aux villageaois et surtout pour les plus jeunes sur des questions comme : qu'est-ce que l'environnement? Pourquoi le protéger? Quelles sont les actions qui le dégrade? Nous appartient-il ou en faisons nous partie au même titre que n'importe quelle espèce? Avons nous un droit sur le reste de la vie? Cet aspect éducatif semble primordial si l'on souhaite un changement de notre relation à la nature qui est peut-être l'enjeu majeur de la sauvegarde de l'environnement dans le futur. Il existe actuellement au sein de l'ONG une "mission éducation". Cependant celle-ci n'existe que depuis deux ans et s'intéresse principalement à la scolarisation des enfants et au matériel scolaire. Ceci est bien sûr une base fondamentale, car il faut tout d'abord permettre aux enfants d'avoir accès à l'éducation. Mais il serait également possible d'envisager des projets d'éducation à l'environnement à moindre coût. En ce sens, la mise en place d'une ou plusieurs journées de "classe verte" dans la réserve pourrait répondre à cet objectif. Tout le monde s'accordera sur le fait que, face aux priorités qu'impliquent la situation de pauvreté dans laquelle se trouve une grande partie de la population malgache (nourriture quotidienne, logement, chauffage...), la préservation de la biodiversité est inévatiblement et logiquement reléguée au second plan. Pour autant, il ne faut pas oublier l'importance de la composante éducative qui s'avère nécessaire, d'une part dans le cadre d'un changement des comportements vis à vis de l'environnement, et d'autre part dans la compréhension et l'acceptation des actions de l'ONG (fondamentale pour la conduite d'un projet à long terme dont le but est l'autonomisation des populations). De plus, une exploitation durable de l'environnement est un gage de ressources sur le long terme. De ce fait, il est possible d'en montrer l'intêret stratégique aux populations locales. Toutefois, MATE à travers son action de valorisation des ressources naturelles s'inscrit totalement dans le cadre de la Politique Nationale de l'Environnement et de la Stratégie Nationale de Gestion Durable de la Biodiversité mise en place par l'Etat malgache. On peut citer, à titre d'exemple, le Madagascar Action Plan (MAP) élaboré en 2006 et appliqué pour le quinquennat 2007-2012. Ce nouveau plan d'action met l'accent sur le développement rural et à cet égard, la valorisation des plantes aromatiques à travers l'exportation des HE fait partie des activités prioritaires identifiées par le MAP. La stratégie de préservation de la biodiversité qui prédomine actuellement dans le monde, est la valorisation des ressources naturelles, censée allier création de revenus et protection de l'environnement. Peut-être faut-il effectivement que la biodiversité apporte un gain direct à l'Homme avant qu'il ne puisse penser à préserver l'environnement pour ce qu'il est, mais peut-être aussi que cette vision inculque l'idée, que la nature, si elle ne présente pas d'intérêt, devient inutile et donc quelque chose qu'il n'est pas nécessaire de conserver. 44 BIBLIOGRAPHIERapports non publiés : - Chlémaire Nadia, 2011. Etude d'impact socio-économique de la production d'huiles essentielles à Vohibola - Deschamps Chloé, 2007. Inventaire amphibiens et étude sur les amphibiens pandanicoles de la réserve expérimentale de Vohibola. - Gehring P-S. & Ratsoavina F., 2009. A rapid assessment of the amphibians and reptiles of the Vohibola forest, eastern Madagascar - Le Goff Arnaud, 2008. Protocole standardisé de suivi des amphibiens. - MATE, 2009. Plan de gestion de la réserve expérimentale de Vohibola 2009. - Missouri Botanical Garden, 2002. Inventaire botanique de la forêt de Vohibola. - Randrianirina J.E, 2005. Inventaire biologique de la forêt littorale de Vohibola, Madagascar. Articles / Publications : - Andreone F., Randrianirina J.E. & Aprea G., 2000. Species diversity of Amphibia, Reptilia and Lipotyphla (Mammalia) at Ambolokopatrika, a rainforest between the Anjanaharibe-Sud and Marojejy massifs, NE Madagascar. Biodiversity and Conservation 9: 1587-1622. - Andreone F., Cadle J., Cox N., Glaw F., Nussbaum R., Raxworthy C., Stuart S., Vallan D. & Vences M., 2005. Species Review of Amphibian Extinction Risks in Madagascar: Conclusions from the Global Amphibian Assessment. Conservation Biology : 1790-1802 - Andreone F., Mercurio V. & Mattioli F., 2006. Between environmental degradation and international pet trade: conservation strategies for the threatened amphibians of Madagascar. Natura - Soc. it. Sci. nat. Museo civ. Stor. nat. Milano, 95 (2): 81-96. 45 - Andreone F., Bungard M. & Freeman K., 2007. Amphibiens menacés de Madagascar. Museo regionale di scienze naturali. Réalisé dans le cadre du projet "A Conservation Strategy for the Amphibians of Madagascar" et de l'Amphibian Specialist Group / IUCN. - Andreone F. & Randriamahazo H., 2008. Sahonagasy Action Plan : Programme de Conservation pour les Amphibiens de Madagascar. Museo regionale di scienze naturali. Réalisé dans le cadre du projet A Conservation Strategy for the Amphibians of Madagascar et de l'Amphibian Specialist Group / IUCN. - Crottini A., Glaw F., Casiraghi M., Jenkins R. K. B., Mercurio V., Randrianantoandro C., Randrianirina J. E., & Andreone F., 2011. A new Gephyromantis (Phylacomantis) frog species from the pinnacle karst of Bemaraha, western Madagascar. ZooKeys 81: 51- 71. doi: 10.3897/zookeys.81.1111 - Daly, J. W. , Garraffo H.M., Spande T.F., Giddings L., Saporito R.A., Vieites R. D. & Vences M., 2008. Individual and Geographic Variation of Skin Alkaloids in Three Species of Madagascar Poison Frogs (Mantella). J Chem. Ecol. 34: 252-279. - Gehring P-S, Ratsoavina F. M. & Vences M., 2010. Filling the gaps - amphibian and reptile records from lowland rainforests in eastern Madagascar. Salamandra 46 : 214-234, ISSN 0036-3375 - IPCC, 2001 Climate Change 2001: The Scientific Basis. Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change [Houghton, J.T.,Y. Ding, D.J. Griggs, M. Noguer, P.J. van der Linden, X. Dai, K. Maskell, and C.A. Johnson (eds.)]. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, 881pp. - Jayaraman K., 1999. Manuel de statistique pour la recherche forestière. Rapport pour l'Organisation des Nations Unies pour l' Alimentation et l'Agriculture, la coopération Hollandaise et la Commission Européenne. 238 pp. - Vallan D., 2000. Influence of forest fragmentation on amphibian diversity in the nature reserve of Ambohitantely, highland Madagascar. Biological Conservation 96 : 31-43. - Vallan, D., 2002. Effects of anthropogenic environmental changes on amphibian diversity in the rainforests of eastern Madagascar. Journal of Tropical Ecology 18 :725-742. - Vallan, D., F. Andreone, V. Raherisoa, & R. Dolch., 2004. Does selective wood exploitation affect amphibian diversity? The case of An'Ala, a tropical rainforest in eastern Madagascar. Oryx 38 : 410-417. 46 - Vieites D., Wollenberg K., Andreone F., Köhler J., Glaw F. & Vences M., 2009. Vast underestimation of Madagascar's biodiversity evidenced by an integrative amphibian inventory. PNAS 106 : 8267-8272. Ouvrages : - Glaw F. & Vences M., 2007. A field guide to the amphibians and reptiles of Madagascar. 3e édition. Cologne, Vences & Glaw Verlag, 496 pages. Sites Web : AmphibiaWeb : http://amphibiaweb.org/search/index.html IUCN : http://www.iucnredlist.org/ I Annexe I1) Poids de chacune des menaces qui s'exercent sur les amphibiens
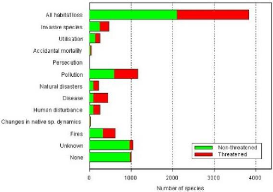
(Source : http://www.iucnredlist.org/initiatives/amphibians/analysis/major-threats) 2) Classements des pays selon leur richesse en espèces d'amphibiens
(Source : http://www.iucnredlist.org/initiatives/amphibians/analysis/geographic-patterns) I Annexe I (bis)3) Classement des pays en fonction du nombre d'espèces endémiques d'amphibiens
(Source : http://www.iucnredlist.org/initiatives/amphibians/analysis/geographic-patterns) Annexe IIOrganigramme de l'ONG l'Homme et l'Environnement
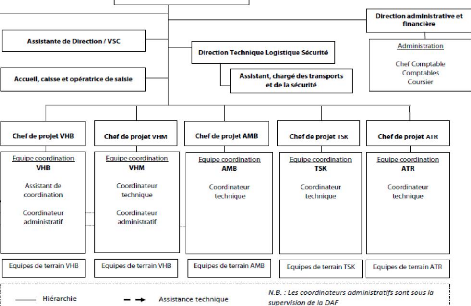
Direction executive Antennes de représentation
l'étranger Programme International d'échanges DGA chargee du suivi Evaluation SIG r Carlo
Hiérarchie -- --4. Assistance technique supervision de la DAf N_8..- Les coordnuteurs odmirrrstratrfs sont sores !a Equ ipes de terrain VHR Equips de terrain VHR Eq u ipes de terrain AMR Equipes de terrain TSK Eq u ipes de terrain ATR Chef de projet VHM Chef de projet TSK Chef de projet ATR Chef de projet VHR Chef de projet AhM16 Equipe coordination VHB Assistant de coordination Coordinateur administratif Equipe coordination VHM Coordinateur technique Coordinateur administratif Fitt line rnnrfination AMH Coordinateur technique FNline rnordinatiort TSK Coc rdi nateu r technique Eauine coordination ATR Coordinateur technique II POLEACTION ENVIRONNEMENTALE
Programme Eco-tourisme Appui â la production et commercialisation Annexe IIISystématique des amphibiens de Vohibola Famille Genre Espèces Discoglossidae Hyperoliidae Hoplobatrachus Heterixalus Aglyptodactylus Heterixalus madagascariensis Heterixalus punctatus
Mantellidae Boehmantis Guibemantis Boophis idae Gephyromantis boulengeri Gephyromantis eiselti Guibemantis bicalcaratus
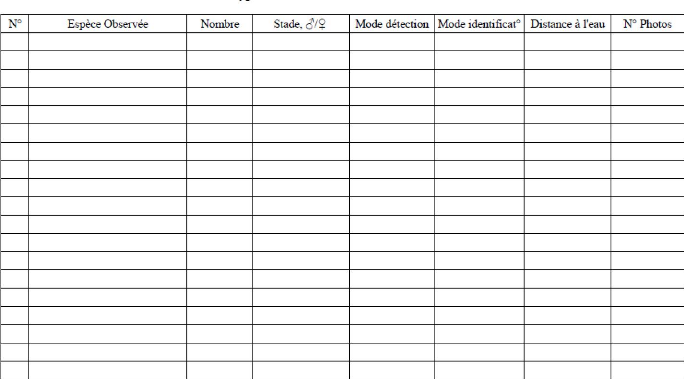
Microhylidae Ptychadenidae Anodonthyla Platypelis Stumpffia Ptychadena Anodonthyla boulengeri III Platypelis grandis Platypelis tuberifera Pletodontohyla notosticta Stumpffia sp. Ptychadena mascareniensis IV Annexe IVFiche d'inventaire terrain Denomination Site : (:oord- GPS Description site :
Espice Obser -ee Toirxlre Stadeo/- Niode detection Mode iclentiticat' Distance ; l'eau Date/Heure : N1etëo T° H -zro : V Annexe VMoyens d'idendification des grenouilles
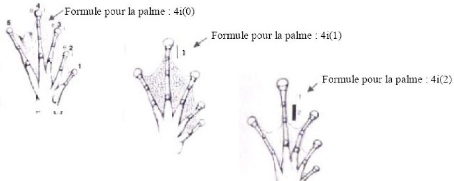
VI Annexe VI
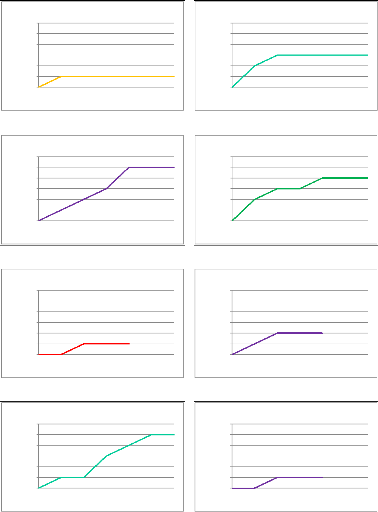
6 5 4 3 2 1 0 Nombre d'espèces 6 5 4 3 2 1 0 Nombre d'espèces 0 60 120 180 240 300 360 6 5 4 3 2 1 0 Nombre d'espèces 0 60 120 180 240 300 360 Courbe d'accumulation : site 1 Nombre d'espèces 0 60 120 180 240 300 360 Durée d'échantillonnage (minutes) 6 5 4 3 2 1 0 Courbe d'accumulation : site 2 Durée d'échantillonnage (minutes) Courbe d'accumulation : site 3 0 60 120 180 240 300 360 Durée d'échantillonnage (minutes) Courbe d'accumulation : site 4 Durée d'échantillonnage (minutes) 6 5 4 3 2 1 0 Nombre d'espèces 0 60 120 180 240 300 360 Courbe d'accumulation : site 5 Nombre d'espèces 0 60 120 180 240 300 360 Durée d'échantillonnage (minutes) 6 5 4 3 2 1 0 Courbe d'accumulation : site 6 Durée d'échantillonnage (minutes) 6 5 4 3 2 1 0 Nombre d'espèces Courbe d'accumulation : site 7 0 60 120 180 240 300 360 Durée d'échantillonnage (minutes) Courbe d'accumulation : site 8 Nombre d'espèces 0 60 120 180 240 300 360 Durée d'échantillonnage (minutes) 6 5 4 3 2 1 0 Courbe d'accumulation de chaque site échantillonné Annexe VI (bis)
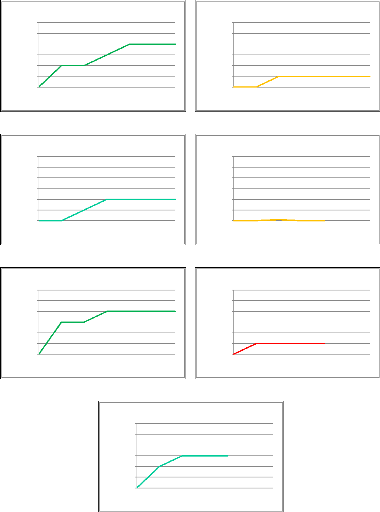
6 5 4 3 2 1 0 Nombre d'espèces 0 60 120 180 240 300 360 6 5 4 3 2 1 0 Nombre d'espèces 0 60 120 180 240 300 360 Courbe d'accumulation : site 9 Durée d'échantillonnage (minutes) Courbe d'accumulation : site 10 Nombre d'espèces 0 60 120 180 240 300 360 Durée d'échantillonnage (minutes) 6 5 4 3 2 1 0 6 5 4 3 2 1 0 Nombre d'espèces Courbe d'accumulation : site 11 0 60 120 180 240 300 360 Durée d'échantillonnage (minutes) Courbe d'accumulation : site 12 Nombre d'espèces 0 60 120 180 240 300 360 Durée d'échantillonnage (minutes) 6 5 4 3 2 1 0 6 5 4 3 2 1 0 Nombre d'espèces Courbe d'accumulation : site 13 0 60 120 180 240 300 360 Durée d'échantillonnage (minutes) Courbe d'accumulation : site 14 Nombre d'espèces 0 60 120 180 240 300 360 Durée d'échantillonnage (minutes) 6 5 4 3 2 1 0 Courbe d'accumulation : site 15 Durée d'échantillonnage (minutes) VI
| 
Changeons ce systeme injuste, Soyez votre propre syndic 
"Ceux qui vivent sont ceux qui luttent" | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||