
REPUBLIQUE DEMOCRATIQUE DU CONGO
UNIVERSITE DE KINSHASA

FACULTE DES SCIENCES AGRONOMIQUES
DEPARTEMENT DE GESTION DES RESSOURCES NATURELLES
Importance de la couverture au sol dans la restauration
des écosystèmes forestiers :
Cas de la Réserve de Biosphère de LUKI
au Bas-Congo
B.P.117 KINSHASA XI
?
MUDERHWA MUTABESHA Patrick
Travail de fin de cycle présenté en vue d'obtenir
le titre de gradué en Sciences agronomiques.
Option : Eaux et forêts
Directeur : KACHAKA
Encadreur : BOLALUEMBE B.
ANNEE ACADEMIQUE 2008 - 2009
Dédicace
A toute la famille MUDERHWA MUSANGANYA
Gérard et M'BUTINDI NANKAFU Félista pour tout sacrifice consenti
pour notre formation;
Nous dédions ce travail.
Etudiant Patrick MUDERHWA
REMERCIEMENTS
Rien ne pouvant s'obtenir sans quelques sacrifices dit-on, ce
travail, d'importance de la couverture au sol dans la restauration des
écosystèmes forestiers: cas de la Réserve de
Biosphère de Luki au Bas-Congo, présenté en vue d'obtenir
le titre de gradué en sciences agronomiques, option: Eaux et
Forêts est l'une de conséquences heureuses de nos sacrifices et
dont la concrétisation n'aurait pas été possible sans le
coup de pouce efficace de nombreux collaborateurs qui nous ont aidé dans
notre tâche.
Nous nous devons ainsi de remercier et de rendre grâce
à l'être suprême, DIEU, le tout puissant, source de toute
vie, force et capacités sans qui, la réalisation de ce travail
n'aurait pas été possible.
Nous témoignons notre profonde gratitude envers les
autorités de la faculté, plus particulièrement au projet
d'appui à la Formation en Gestion de Ressources Naturelles dans le
Bassin du Congo, FOGRN-BC en sigle, pour leur dévouement afin que nous
ayons une bonne formation.
Nous tenons à remercier très chaleureusement le
professeur KACHAKA SUDI, alors Doyen de la faculté des sciences
agronomiques, pour avoir dirigé ce travail et l'assistant Claude
BOLALUEMBE BOLIALE pour son encadrement très enrichissant dans la
réalisation de ce travail.
Que notre reconnaissance aie tout droit aux professeurs, alors
responsables du projet susmentionné, DAMASE KHASA, Directeur principal
MENDA KAZAYAWOKO, Point focal et à madame MELIE MONNERAT,
Conseillère sous-régionale, pour les sacrifices consentis, tant
de conseils et leur amour qu'ils n'ont cessé de nous témoigner
ainsi qu'à la coopération technique allemande GTZ, pour son
soutien qu'elle continue à nous apporter.
A nos parents MUDERHWA MUSANGANYA Gérard et M'BUTINDI
NANKAFU Félista, pour nous avoir aidé à faire nos premiers
pas dans la vie, nos chaleureux remerciements et que le tout puissant, DIEU,
les protège.
Notre gratitude s'adresse également à notre
grande soeur Béatrice MUDERHWA et toute sa famille pour tout ce qu'elles
ont enduré pour que nous arrivions à ce jour.
Enfin, nos frères et soeurs, amis et connaissances
nommément non cités qui nous ont apporté une contribution
quelqu'en soit la forme pour la réalisation de ce travail trouvent, dans
ce peu de mots, l'expression de notre reconnaissance.
Etudiant Patrick MUDERHWA
0. INTRODUCTION
Les forêts ne peuvent avoir une bonne santé que
si leur milieu est riche en nutriments du fait qu'un sol plus fertile portera
naturellement, les arbres les plus beaux, les plus hauts et les forêts
les plus denses (Plamondon, 2009).
Ebenmayer (1976) a évalué les apports annuels
de litière et leurs effets sur les accroissements des arbres dans les
forêts décidues et conifériennes de Bavière. Il a pu
démontrer qu'une grande proportion des nutriments prélevés
au sol annuellement par l'arbre, y est retournée par la litière
annuelle. Il a aussi pu établir qu'une interruption de ce cycle par
l'exploitation des litières comme fourrage pour les animaux domestiques,
pratique assez commune en Europe, récolte de débris (branches,
brindilles, bois morts) pour avoir de l'énergie, causait une
détérioration de la croissance forestière. Une loi fut
alors adoptée en Allemagne afin d'interdire cette pratique
néfaste au développement des écosystèmes
forestiers.
En outre, Plamondon (2009) signale que certaines actions
anthropiques dégradent le sol et considère qu'elles constituent
la principale cause de réduction de la fertilité des sols suite
à la perte de la matière organique et des nutriments surtout
concentrés dans la couche supérieure du sol.
Mc Kinnon et al., (1990) soulignent que ce
phénomène, de mise à nu du sol qui empêche la
forêt de se rajeunir naturellement ainsi que de continuer à
remplir ses fonctions, est accentué dans le Mayombe congolais notamment
dans la Réserve de Biosphère de Luki où
l'écosystème forestier subit une sérieuse
dégradation suite à la pression démographique.
Alors qu'une aire protégée est
appelée Réserve de Biosphère quand elle remplit les
fonctions de conservation, de développement et de support logistique
selon Maldague et al., (1997), Nsenga (2001) souligne que la
Réserve de Luki, seule Réserve de Biosphère encore
opérationnelle parmi les trois reconnues en RDC, ne remplit pas les
fonctions ci-haut citées suite non seulement à la mauvaise
politique de gestion mais surtout à la pression exercée par les
populations habitant dans la Réserve et en périphérie de
la Réserve.
Lubini (1984) et Nsenga (2001) ajoutent que les actions de
ces populations, entre autres l'agriculture itinérante sur brulis,
la récolte de bois de feu, la
carbonisation sont les principales causes qui président au
déboisement et à la déforestation.
Dans le cadre de notre travail nous
essaierons de voir si les débris végétaux, bois morts
récoltés, alors renforçant l'épaisseur de la
couverture au sol, favorise la régénération naturelle des
forêts à dominance de Terminalia superba (Limba)
Le maintien permanent de la couverture végétale
au sol serait nécessaire pour restaurer les milieux
dégradés de façon à réduire les pertes de
sols et de nutriments, à conserver la fertilité (des
forêts) et à reconstituer la composition floristique, la
structure, les fonctions et la dynamique de la végétation.
Le travail consiste en deux objectifs qui sont :
- Comparer la couverture au sol dans deux blocs
différents dominés par le Terminalia superba ;
- Identifier les espèces de sous-bois de ces
différentes parcelles.
La présente étude contribue aux connaissances
sur le rôle des écosystèmes forestiers dans le maintien de
sol et leur protection grâce à la couverture au sol qu'ils
produisent selon qu'un sol mis à nu sous couvert arborescent d'une
hauteur variant entre 7 m et 8 m génère de pertes de sols
accompagnées de celles de fertilité.
Outre l'introduction et la conclusion, le travail comporte
quatre chapitres dont le premier porte sur la revue de la littérature,
le deuxième sur la présentation de la Réserve de
Biosphère de Luki, le troisième sur les matériels et
méthodes et enfin la quatrième sur les résultats et
discussion.
CHAPITRE I. REVUE DE LA LITTERATURE
I.1. Définition de la couverture au sol
Selon Plamondon (2009), la couverture au sol est l'ensemble
de débris ligneux ou bois morts, de litières, de la
végétation basse constituée des espèces des
sous-bois excepté les espèces de la strate arborescente se
trouvant à la surface du sol.
Signalons que la couverture au sol est différente de
la couverture du sol du fait que cette dernière inclue les
espèces de la strate supérieure.
I.2. Différents types des couvertures au sol
a. Litière
La litière est un sentier pour la restitution des
éléments absorbés par la végétation au
sol.
Les additions annuelles de feuillage et des branches sont
incorporées à l'humus forestier et éventuellement une
portion s'incorpore à la matière organique du sol.
La litière est un important input puisqu'il contient
évidemment tous les éléments (Ca, K, P, Fe, N, C, Mg,
etc.) nécessaires à la croissance des arbres.La matière
organique fournit au sol permet d'entretenir une flore microscopique
très diversifiée dont l'action facilite aux arbres l'assimilation
de minéraux.
En pédologie, la litière est la couche
superficielle qui couvre le sol en regroupant les horizons dits
« Holorganiques ».
b. Débris végétaux
Au sens large, c'est l'ensemble de tout ce qui est du
matériel végétal rencontré au sol mais au sens
strict c'est l'ensemble de branches (se distinguant des brindilles par leurs
dimensions plus grandes), bois morts trouvés à la surface du
sol.
Ils peuvent être de dimensions variables selon les
espèces et le stade de développement et ils se retrouvent au sol
soit à cause des facteurs intrinsèques (vieillissement, ...) soit
des facteurs extrinsèques (vents violents).
Ces débris végétaux interviennent dans le
maintien de la fertilité du sol une fois décomposée par la
microflore du sol.
c. Végétation basse ou sous-bois
C'est une végétation constituant la strate
arbustive et celle herbacée. C'est-à-dire au sens large c'est
tout ce qui est en dessous de la strate arborescente.
Les composantes de la strate arbustive ont de dimensions
variables (Hauteur, DHP) dépendant des facteurs extrinsèques
(conditions du milieu) et intrinsèques (liés aux espèces
mêmes).
Cette végétation basse intervient dans le
maintien de la fertilité du sol du fait qu'elle absorbe l'impact de
gouttes de pluie au sol et favorise l'infiltration en lieu et place du
ruissellement de surface.
I.3. Cycles des éléments et nutrition des
essences forestières
Selon Anonyme (2009) la circulation des nutriments -intrants
et extrants et flux entre les différents réservoirs d'un
écosystème est un processus continu qui peut être
mesuré sur une base journalière, saisonnière, annuelle ou
même sur une révolution complète du peuplement.
On reconnait trois cycles majeurs ayant cours dans les
écosystèmes forestiers :
· Le cycle géochimique pour désigner les
intrants et les extrants en éléments nutritifs ;
· Le cycle biogéochimique pour désigner
les transferts entre le sol et la biomasse végétale ;
· Le cycle biochimique pour désigner les
transferts d'éléments à l'intérieur de l'arbre.
A. Cycles des éléments
a. Cycle géochimique des nutriments
Les intrants désignent les gains réalisés
par l'écosystème alors que les extrants identifient les pertes
qu'il subit. Les quantités de nutriments gagnées ou perdues
annuellement par un écosystème sont influencées par des
facteurs tels que les propriétés édaphiques, les
conditions climatiques, le type de végétation, les sources de
polluants naturels ou anthropiques, etc.
L'écoulement hypodermique sur les pentes peut aussi
constituer un transfert interne important. Les flux tendent vers un
équilibre dans les forêts matures lorsqu'il n'y a pas de
perturbation majeure tant humaine que naturelle.
Les principaux gains de nutriments sont attribuables aux
apports atmosphériques liquides, gazeux et particulaires (surtout N et
S), à la fixation biologique de N2, à
l'altération des minéraux (surtout P, K, Ca, Mg, Si et Al) et
à la fertilisation.
Les pertes de nutriments résultent du lessivage en
profondeur, du ruissellement de surface et de l'érosion, de
l'exploitation forestière, des incendies forestiers et de la
volatilisation. Les pertes et les gains géochimiques représentent
généralement des quantités de nutriments relativement plus
faibles que celles qui circulent à l'intérieur de
l'écosystème, entre les arbres et le sol. Les pertes peuvent
augmenter grandement à la suite d'une perturbation (voir Lamontagne
et al., 2000 pour le Boucher laurentien).
b. Cycle biochimique
Les variations saisonnières dans la composition
minérale des composantes de l'arbre sont bien connues. Il se produit une
translocation de certains nutriments des organes matures ou sénescents
vers les régions à croissance active de l'arbre et vers les
tissus de Réserve.
Ainsi, les concentrations en N, P et K sont toujours plus
élevées chez les jeunes feuilles que chez les feuilles
matures ; puis elles diminuent constamment durant la croissance active des
feuilles, demeurent constantes au cours de la période d'expansion
complète des feuilles et déclinent lors de la
sénescence.
Par contre, Ca est un élément non mobile et sa
croissance dans les tissus augmente tout au long de la saison de croissance.
Pour le feuillage, les exigences en N, P et K peuvent
être satisfaites de par la translocation de nutriments en provenance du
bois ou des branches (Côté et al., 2002). Ce processus de
transfert est toutefois beaucoup moins important pour le Ca et le Mg (Myre et
Camiré, 1994). Les réponses de croissance à long terme
fréquemment obtenues après fertilisation, spécialement
avec P et K, sont souvent expliquées par les transferts internes.
L'oubli de reconnaître ou de considérer le
transfert interne des nutriments peut conduire à des conclusions
erronées sur le fonctionnement des écosystèmes
forestiers.
En dépit de grandes variations entre différents
écosystèmes dans les quantités de nutriments en
circulation, il n'y a pas de doute que ces transferts cycliques jouent un
rôle majeur dans la productivité continue des sols forestiers.
c. Cycle biogéochimique
Les étapes successives du cycle biogéochimique
sont les suivantes : le prélèvement des nutriments du sol
par les arbres, leur rétention et leur distribution dans les arbres,
leur retour au sol par la litière, leur décomposition et leur
minéralisation.
· Le prélèvement des nutriments du sol
par les arbres
Le prélèvement annuel de nutriments
exercé par la plupart des espèces forestières est du
même ordre de grandeur que celui effectué par plusieurs cultures
agricoles ; mais comme en forêt la majeure partie de nutriments est
retournée au sol par les litières, seules de faibles
quantités sont retenues chaque année dans l'augmentation annuelle
de la biomasse.
Le prélèvement des nutriments par les arbres
forestiers est influencé par les conditions climatiques, le type et
l'âge du peuplement et le sol.
Les principales régions forestières du globe
sont soumises à des conditions climatiques différentes qui
influencent le statut nutritif de leurs peuplements respectifs.
En général, le prélèvement de Ca
est plus élevé que celui de N chez les espèces feuillues,
mais il est plus faible chez les espèces résineuses. Le
prélèvement de Mg est du même ordre de grandeur que celui
de P chez les conifères, alors qu'il en est le double chez les
feuillus.
Le stade de développement du peuplement affecte aussi
le prélèvement annuel de nutriments. Dans la phase
juvénile du développement, le taux de prélèvement
augmente annuellement. Puis à l'approche de la maturité il y a
diminution de ce taux qui peut être accompagnée d'une
réduction de la chute de litière.
Lorsque le peuplement se ferme, le prélèvement
devient relativement constant. Enfin, il diminue fortement dans la forêt
surannée.
Tableau 1 : Comparaison des espèces
arborescentes décidues et conifériennes relativement au
prélèvement, aux besoins et à la restitution en nutriments
(Kg.ha-1.an-1)
Éléments
|
Prélèvement
|
Besoins
|
Restitutions
|
|
Conifères
|
Feuillues
|
Conifères
|
Feuillues
|
Conifères
|
N
P
K
Ca
Mg
|
70
6
48
84
13
|
39
5
25
35
6
|
94
7
46
54
10
|
39
4
22
16
4
|
57
4
40
67
11
|
30
4
20
29
4
|
|
Source : Cole et Rapp, 1981.
· Rétention et la distribution des nutriments
dans les arbres
La rétention ou l'accumulation nette annuelle de
nutriments dans les arbres d'un peuplement peut être
considérée comme étant la différence entre, d'une
part le prélèvement total et, d'autre part, la quantité
retournée au sol par les exsudats et la mortalité radicalaire, la
litière aérienne et les pluviolessivats.
.
Cette rétention annuelle est ordinairement
calculée à partir des données obtenues des mesures
périodiques de la biomasse et de la composition chimique des tissus.
Etant donné que l'accumulation des nutriments suit l'accroissement de la
biomasse, il existe une phase linéaire ou exponentielle d'accumulation
durant la croissance juvénile rapide et une réduction
subséquente du taux d'accroissement à l'approche de la
maturité.
Dans le stade initial de développement d'un peuplement,
la majeure partie des nutriments est contenue dans le feuillage.
B. Nutrition des essences
forestières
B.1. Production de litière en milieu
forestier
La litière représente un des sentiers majeurs du
flux nutritionnel. Une large proportion de la production primaire nette des
écosystèmes forestiers retourne au sol annuellement. Dans le cas
de forêts matures, c'est même la presque totalité de cette
production.
La production de litière est contrôlée
par des facteurs climatiques et édaphiques, les premiers
contrôlant la production au niveau régional et les seconds au
niveau local.
La litière ne comprend généralement pas
que de feuilles, les données compilées par Bray et Gorham (1964)
indiquent que les autres constituants (fruits, branches, écorces, etc.)
comptent en moyenne pour 28% de la masse totale des litières
aériennes.
La contribution de la végétation de sous-bois
à la production de litière est variable selon les milieux
s'élevant de 3 à 20%.
Toutefois, pour autant que le peuplement soit fermé, la
production de litière est assez indépendante du nombre de tiges
à l'unité de surface puis la biomasse du feuillage tend à
se stabiliser après la fermeture du couvert forestier.
B.2. Restitution des nutriments au sol
La composition des litières est complexe et
évolue rapidement après la chute des feuilles et des autres
débris végétaux.
Au moment de leur chute les feuilles, qui comptent pour
#177;70% de la masse des litières épigées, apportent au
sol des constituants cellulaires (sucres, protéines, acides
nucléiques, phospholipides), des constituants structuraux des parois
cellulaires (cellulose, hémicellulose, lignine) et des constituants de
Réserve (amidon, lipides, protéines). Ces divers constituants
varient dans les proportions suivantes :
1) 60 à 70% de glucides (cellulose,
hémicellulose, amidon, oligosaccharides, monosaccharides) ;
2) 10 à 15% de protéines et substances
associées (peptides, acides aminés) ;
3) 10 à 30% de lignines, polyphénols et
tannins ;
4) #177;5% de lipides, graisses, cires, huiles et
résines.
L'analyse élémentaire des tissus foliaires
relève une teneur en cendres de l'ordre de 5 à 10% de la masse
anhydre, ce qui fait des litières forestières un important
réservoir de nutriments.
Le principal facteur de variation de la composition
élémentaire des litières est l'espèce d'origine.
La restitution de nutriments varie selon la qualité de
station, les espèces et l'âge du peuplement. Ainsi, la proportion
de nutriments prélevés retournée au sol est plus forte sur
sols riches que sur sols déficients.
La quantité de litière produite par la
végétation du sous-bois est ordinairement faible, mais sa
contribution au cycle des nutriments peut être significative étant
donné la forte concentration des nutriments et la rapidité de
décomposition de la litière de ces espèces.
B.3. Minéralisation des litières
La litière doit être décomposée et
transformée sous forme minérale (anions ou cations). Sous cette
forme, les nutriments sont alors disponibles pour le prélèvement
par les arbres.
Cette décomposition est effectuée par la faune
du sol et par la microflore (bactéries, champignons). Elle est
conditionnée par trois facteurs importants :
Ø Facteurs intrinsèques à la
litière ou à la nature même de la litière ;
Ø Facteurs extrinsèques à la
litière ou les conditions microclimatiques et chimiques de
l'environnement ;
Ø La composition des populations microbiennes et
fauniques.
1° Nature de la litière
Les facteurs intrinsèques à la litière
sont des propriétés physiques (épaisseur de la cuticule,
dureté, taille), ses propriétés nutritives (teneur en
nutriments, qualité énergétique) et la présence de
composés non nutritifs qui influencent l'attaque par les organismes du
sol (tannins, résines, composés allélopatiques) (Swift
et al., 1979). En règle générale, les
litières riches en protéines (C/N bas) et bien pourvues en
cations basiques (Ca, Mg, K) ont tendance à se minéraliser
rapidement.
Ce facteur n'est toutefois pas entièrement
indépendant des conditions du milieu. Les substances hydrosolubles et
directement fermentescibles (sucres et acides aminés) des
litières représentent une source d'énergie
immédiatement utilisable et stimulent les populations microbiennes.
Par contre, les tannins sont susceptibles de freiner l'attaque
des protéines en formant des complexes
polyphénolsprotéines alors que la lignine est un des
composés résistant le plus à la décomposition.
Plusieurs indices, tels que le rapport lignine/N (Melillo et al.,
1982), le rapport C/N (Taylor et al., 1989) ou encore les rapports
cutine/N et cutine/P (Gallardo et Merino, 1993) reflètent la
qualité de la litière au regard de sa vitesse de
décomposition (Trofymow et al., 2002).
2° Conditions microclimatiques et chimiques de
l'environnement
Les conditions ambiantes exercent une influence
décisive sur la décomposition de la litière qui sera
rapide sous des conditions de température élevée,
d'humidité et d'aération optimales et de pH du sol faiblement
acide à neutre. Les basses températures des sols des forêts
boréales expliquent en bonne partie la lenteur de décomposition
des litières.
3° Composition des populations fauniques et
microbiennes
La présence et l'activité des populations
fauniques et microbiennes sont la réalité conditionnée par
les facteurs qui précèdent. L'action de la microfaune s'exerce
à trois niveaux : il y a d'abord fragmentation des débris
végétaux conduisant à une attaque enzymatique
accélérée, tant dans le tube digestif que dans le milieu
environnant, suivie de l'incorporation de ces débris au sol
minéral à des profondeurs plus ou moins grandes, selon
l'habilité au fouissage et de la formation de complexes
organo-minéraux.
B.4. Prélèvement des ions par les arbres
Le prélèvement des ions par l'arbre est un
processus sélectif. Les éléments ne sont pas
prélevés en proportion directe de leurs concentrations relatives
dans le sol ou dans la solution du sol, non plus qu'en fonction des besoins
minimaux de l'arbre. La demande d'éléments est
contrôlée par des facteurs génétiques et varie selon
les espèces (Paré et al., 2002). Sur la base de leurs
exigences quantitatives en nutriments, les végétaux sont
classés en arbres microtrophes, mésotrophes et
mégatrophes ; les programmes de reboisement doivent tenir compte de
ces exigences différentes des espèces.
La nutrition d'un peuplement ne peut toutefois pas être
envisagée qu'en termes de besoins physiologiques des différentes
espèces.
Le bilan annuel du prélèvement ou de
l'absorption d'éléments par un peuplement, doit tenir compte
à la fois :
- De l'accumulation d'éléments dans la biomasse
végétale ;
- De la restitution d'éléments au sol par le
délavage des cimes et de l'écorce, la litière de surface,
la litière de racines et les exsudats radicalaires. Ce
prélèvement varie avec l'âge du peuplement et pour chaque
élément.
B.5. Eléments requis pour la nutrition des
arbres
Tous les végétaux ont besoin d'au moins 17
éléments pour satisfaire leurs besoins végétatifs
et reproductifs. Ces éléments sont dits essentiels, car ils ne
sont pas entièrement substituables par d'autres éléments
(Marschner, 1994). Ce sont, outre C, H et O, les éléments dits
majeurs ou « macronutriments » (N, P, K, Ca, Mg et S) et
les éléments dits mineurs ou
« micronutriments » ou
« oligoéléments » (Fe, Cu, Zn, Mn, Ni, B, Mo,
et Cl). L'appellation de ces deux derniers fait référence pour
chaque élément, à la quantité relative
nécessaire pour satisfaire les besoins de l'arbre et non à
l'importance physiologique, car chacun est essentiel au maintien de certains
processus vitaux.
Lors de l'analyse chimique des arbres, presque tous les
éléments du tableau périodique peuvent être
identifiés mais mis à part ceux cités ci-dessus, aucun n'y
joue un rôle fonctionnel défini.
Toutefois, certains éléments, tels que le Fluor
(F), le Sodium (Na), le Sélénium (Se) et l'Iode (I) sont
essentiels aux animaux. Cette particularité est importante pour les
milieux agricoles et aussi dans la perspective de la productivité
secondaire (nutrition de la faune) des écosystèmes naturels.
Tous les éléments essentiels peuvent provenir de
la minéralisation des débris organiques, phénomène
d'une importance considérable dans la très grande majorité
des écosystèmes forestiers. Ils peuvent aussi être
d'origine non organique et, sur cette base, ils sont classés en deux
groupes : d'une part ceux qui proviennent de l'air et de l'eau (C, H, O,
N) et d'autre part ceux qui proviennent de l'altération des
minéraux (Ca, K, Mg, S, Fe, Cu, Zn, Mn, Ni, B, Mo, Cl).
Cependant, dans certaines régions, les apports
atmosphériques peuvent constituer une source non négligeable de
nutriments (Ouimet et Duchesne, 2008).
I.4. Autres concepts
a. Réhabilitation
La réhabilitation des milieux dégradés
est requise sur les sites où une mauvaise gestion écologique a
contribué à détériorer les fonctions du
territoire.
La réhabilitation vise à rétablir les
fonctions de base du territoire telles les composantes du cycle hydrologique,
le recyclage des nutriments, le contrôle de l'érosion et la
productivité.
Elle s'applique donc aux écosystèmes naturels et
aux systèmes de production développés par l'homme.
La réhabilitation à l'échelle du bassin
implique un choix des espèces à utiliser, des pratiques de
conservation à favoriser, des zones à traiter et de la proportion
du paysage à rétablir.
b. Restauration
La restauration, tout comme la réhabilitation,
consiste à rétablir les processus énergétiques,
hydrologiques et biologiques du milieu, mais dans le cas de la restauration on
vise à reconstituer la composition floristique, la structure, les
fonctions et la dynamique de la végétation, ainsi que les
habitats fauniques du système originel.
c. Dégradation des terres
La dégradation est généralement
définie par le déclin temporaire ou permanent de la
capacité productive des terres (UN/FAO, 1991).
Selon une autre définition, la dégradation est
la diminution cumulative du potentiel productif des terres en termes d'usages
(cultures, pluviales ou irriguées, forêts, pâturages), de
systèmes agricoles et de ressources économiques.
La dégradation des terres est un terme global qui
réfère à la détérioration d'une ou de
plusieurs ressources du territoire tels le sol, l'eau, la
végétation, l'assise rocheuse, l'air, le climat, le relief et le
paysage (Stocking et Murnaghan, 2001).
Tableau 2 : Différentes composantes du terme
« Dégradation des terres »
Dégradation des terres
|
Dégradation du sol
|
Altération du paysage
|
Détérioration de l'eau
|
Érosion du sol
|
Épuisement des nutriments
|
Perte de biodiversité
|
Changement climatique
|
Érosion hydrique
|
Érosion éolienne
|
Réduction du couvert végétal
|
Pollution
|
Sécheresse
|
Compactage par animaux machinerie
|
Sédimentation
|
Augmentation température du sol
|
Réduction de matière organique
|
Salinisation
|
|
Source : Plamondon, 2009.
d. Fertilité du sol
Est la capacité du sol de soutenir la croissance des
arbres de manière durable, dans des conditions données de climat
et d'autres propriétés pertinentes de la terre. Elle fait partie
du concept plus large de productivité de la terre.
Les gains de fertilité par l'agroforesterie sont
basés sur l'hypothèse que les arbres recyclent plus efficacement
les nutriments et que l'apport de matières organiques améliore
les propriétés physiques du sol et les activités
biologiques.
I.5. Importance de la couverture au sol
a. Conservation des sols
Cette conservation a pour but de maintenir la
productivité des terres par le développement de techniques qui
maintiennent les pertes de sol et des nutriments ou restaurent les milieux
dégradés de façon à réduire les pertes de
sols et de nutriments à un taux acceptable appelé taux
d'érosion acceptable.
La manière la plus efficace et la plus
économique d'éviter l'érosion et la perte de
productivité du sol est de réduire l'impact des gouttes de pluie,
sinon le ruissellement à sa source.
Le maintien de la couverture au sol est la première
mesure à considérer, car elle assure une capacité
d'infiltration maximale selon le type de sol.
On doit donc conserver un couvert végétal
permettant de maintenir une couverture au sol offerte par des plantes basses,
des débris organiques ou d'un taux élevé de matière
organique incorporée dans la couche supérieure du sol.
L'érosion entraîne principalement les particules
organiques et minérales fines présentes dans la couche
supérieure du sol. Les nutriments étant principalement
concentrés dans cette couche supérieure du sol, il s'en suit une
baisse graduelle de productivité des sols.
Le couvert de la surface de sol est de loin la principale
protection du sol contre l'érosion. Le couvert arbustif et le couvert
forestier ne protègent pas le sol directement mais indirectement par la
chute de matière organique qui forme la litière protectrice
(Wiersum, 1985).
b. Infiltration et infiltrabilité des eaux de pluie
dans le sol
Selon Lassen et al., (1955), l'infiltration
représente le mouvement de l'eau près de la surface du sol. Et
c'est le processus par lequel l'eau passe à travers la surface du sol ou
l'entrée de l'eau jusqu'à une profondeur de 1,5 cm.
L'eau pénètre dans le sol par les pores, les
fissures, les orifices pratiqués par les vers ou occasionnés par
la pourriture des racines ainsi par les cavités résultants des
labours ou de la préparation du sol pour la plantation (Avila, 1980).
Par contre, l'infiltrabilité ou la capacité
d'infiltration, terme proposé par Hillel (1971) pour remplacer
l'infiltration, est le taux maximum de pénétration de l'eau dans
un sol correspondant à l'état où celui-ci se rencontre
(Hewlett, 1972). En milieu boisé sur sol grossier, cette capacité
est souvent plus élevée que les plus fortes intensités de
précipitation. Il est à noter que l'infiltrabilité d'un
sol est la valeur obtenue lorsque le taux d'infiltration devient constant
après une certaine période d'apport d'eau.
Ø Facteurs et conditions qui affectent
l'infiltration et l'infiltrabilité
La texture du sol détermine la dimension des pores, sa
conductivité hydraulique à saturation et, par conséquent,
la capacité d'infiltration.
La présence de litière ou de matière
organique incorporée dans la couche supérieure du sol
réduit le compactage par la battance de la pluie et, de ce fait,
réduit le colmatage des macrospores permettant la
pénétration rapide de l'eau dans le sol.
La rugosité de la surface favorise le stockage de l'eau
et son infiltration au détriment du ruissellement.
Lorsque la surface du sol minéral est
protégée, l'infiltrabilité est meilleure alors que le
ruissellement de surface est faible. Cet effet s'amenuise lorsque le sol est
très humide.
L'infiltrabilité augmente légèrement avec
le taux d'apport d'eau (pluie, irrigation) sur sol nu, alors qu'elle est plus
élevée et augmente plus fortement lorsque le sol est recouvert de
végétation.
La végétation par l'apport de matières
organiques, maintient la porosité de la surface du sol minéral
assurant une meilleure pénétration verticale de l'eau.
Cependant, la différence de porosité non
capillaire entre l'absence et la présence de végétation
est semblable pour chaque type de sol, ce qui démontre l'importance de
maintenir un couvert végétal permanent ou des résidus
végétaux au sol.
c. Lutte contre l'érosion hydrique en milieu
forestier
L'interception de la pluie sur les surfaces
végétales favorise la création de gouttes plus grosses par
l'amalgame des gouttelettes. Ces grosses gouttes d'eau atteignent leur vitesse
terminale de chute après une distance verticale de 7 m (Hewlett, 1982)
ou 8 m (Hoover, 1962).
Ainsi, seulement un couvert végétal bas est
efficace pour réduire l'énergie cinétique de la pluie dans
sa chute.
Une goutte tombant d'une hauteur n'a pas le temps d'atteindre
sa vitesse terminale. Lorsque la matière organique n'est pas
récoltée en forêt, l'impact de la pluie est absorbé
très efficacement par la couche de matière organique
(litière, humus), les débris ligneux et les plantes basses
(Wiersum, 1983).
Il est aussi reconnu que l'exposition du sol minéral
sous un couvert arborescent de plus de 8 m de hauteur peut causer des taux
d'érosion très élevés. Les gouttes d'eau qui
tombent du feuillage sont plus grosses que celles de la pluie et atteignent une
vitesse de chute plus élevée, ce qui augmente leur impact sur le
sol.
On note l'importance de la végétation basse, qui
absorbe l'impact des gouttes de pluie lorsque la plantation est jeune et que la
litière est en formation.
d. Régulation thermique du sol sous couvert
forestier
La couverture au sol influence grandement la quantité
de radiations solaires atteignant celui-ci.
L'effet d'une forêt dense est bien connu et même
une végétation champêtre a une influence très
notable, spécialement sur les fluctuations de température.
Les sols nus se réchauffent plus vite et se
refroidissent également plus rapidement que ceux couverts de
végétation ou de paillis.
Les températures du sol sont influencées par ce
qui les recouvre et spécialement par les résidus organiques ou
autres types de paillis placés sur la surface du sol. En plus de
diminuer l'évaporation dans l'eau de la surface du sol, les
différentes couvertures au sol tendent à amenuiser les
extrêmes de température.
CHAPITRE II. PRESENTATION DE LA RÉSERVE DE
BIOSPHERE DE LUKI
II.1. Situation géographique
La Réserve de Biosphère de Luki est
située dans la Province du Bas-Congo, à l'Ouest de la
République Démocratique du Congo, à #177;120 Km de la
côte atlantique.
D'une superficie d'environ 32 714 ha, la Réserve de
Biosphère de Luki est traversée par la quasi-totalité du
bassin hydrographique de la rivière Luki avec affluents : la Ntosi
et la Monzi, qui ensemble se jettent dans la rivière Lukunga qui est
l'un des affluents du fleuve Congo.
Elle constitue la pointe méridionale extrême du
massif forestier guinéo-congolais.
Elle est située à 13°10' et 13°15' de
longitude Est et 5°35' de latitude Sud ; l'altitude varie entre 150 m
et plus de 500 m, rarement 600 m (Lubini, 1984).
II.2. Création et historique
La Réserve de Biosphère de Luki a
été instituée par l'ordonnance coloniale du Gouverneur
Général n°05/AGRI du 12 Janvier 1937. Sa mission consistait
à l'exploitation forestière avec la sylviculture comme
préoccupation, c'est-à-dire concilier la coupe des bois d'oeuvre
avec la gestion durable des écosystèmes.
Ainsi, l'INEAC (Institut National d'Études Agronomiques
du Congo-Belge) institua une exploitation méthodique qui visait
essentiellement la connaissance de la dynamique forestière et
l'identification des normes permettant une sylviculture tropicale rentable dans
les conditions écologiques locales (Donis, 1948a).
Considérée d'abord comme une station
forestière de l'État confiée à l'INEAC dont la
gestion fut basée sur deux notions fondamentales suivantes :
- La création des parcelles expérimentales
appelées « blocs » pour besoin d'inventaires, de
relevés pédo-botaniques et d'études
forestières ;
- L'élaboration des méthodes de conversion des
futaies afin d'accroître la valeur marchande de la forêt (Kapa
et al., 1987).
Dès sa création en 1937, la station
forestière fut gérée par l'INEAC qui est devenu
actuellement l'INERA (Institut National d'Études et de Recherches
Agronomiques).
Conformément à l'ordonnance n°77-022 du 23
Février 1977, la gestion de ce domaine boisé fut retirée
de l'INERA et concédée au Ministère de l'Environnement,
Conservation de la Nature et Tourisme (MECNT).
En Mai 1979, la station boisée fut reconnue par
l'UNESCO en titre de Réserve de biosphère et faisant ainsi partie
du réseau mondial des Réserves des Biosphère.
La gestion fut donc retirée de l'INERA pour être
confiée au Comité national du programme MAB/Congo.
De 1980 à nos jours, la gestion revient au
Comité national du programme MAB (Man And Biosphere) qui est sous
tutelle du Ministère de l'Environnement, Conservation de la Nature, Eaux
et Forêts.
II.3. Cadre mésologique de la Réserve
Les aspects du cadre mésologique comprennent les
éléments climatiques (précipitations, humidité
relative, vent et nébulosité) et les éléments
édaphiques (géologie, géomorphologie, sols et
hydrographie).
II.3.1. Eco climat de Luki
Le climat de Mayombe dont fait partie celui de la
Réserve de Biosphère de Luki est sous l'influence de
l'Océan Atlantique. Il appartient au climat tropical humide du type
AW5 selon la classification de Koppen. Il est influencé par
le courant marin froid de Benguela et les Alizés du Sud-est qui
rencontrent au-dessus de la Réserve un écran de
végétation peu humide empêchant ainsi des
précipitations abondantes. Ce courant marin froid de Benguela est
responsable des petites pluies de saison sèche connues localement sous
le nom de « Masala » (Kapa et al., 1987).
L'analyse des données météorologiques de
la station de Luki de 2002 à 2005 (figure 1 et annexe 1) montre que dans
la Réserve de biosphère de Luki, il existe deux grandes saisons,
l'une pluvieuse s'étalant entre la deuxième moitié
d'Octobre et la première moitié de Mai, l'autre sèche
couvrant la période entre la seconde moitié de Mai et la
première moitié d'Octobre. La saison des pluies est
entrecoupée par une courte période de sécheresse relative,
d'ailleurs peu marquée, entre Décembre et Janvier.
La saison sèche dure quatre mois et se
caractérise par une baisse légère de température et
de fréquents brouillards matinaux ou brumes qui compensent le
déficit en eau du sol.
La température moyenne annuelle varie de 28°
à 29°C. L'humidité relative de l'air reste
élevée toute l'année ; sa moyenne annuelle est de
72,34%.
Comme on peut le constater, les précipitations sont
inférieures à la limite du seuil (1400 mm) de
développement d'une forêt ombrophile semi-sempervirente ou
semi-caducifoliée. Mais les conditions écoclimatiques de la
Réserve de Biosphère de Luki permettent le développement
de ce type de forêt qui, par rapport à son aire
géographique et sur le plan synécologique ne s'explique pas. Quoi
qu'il en soit, le phénomène de compensation par les brouillards
épais ou brumes y est pour beaucoup dans la correction du déficit
des précipitations (Toirambe, 2002).
Les vents sont très violents surtout dans les
après-midi et causent facilement la chute des arbres. Ce sont
essentiellement les Alizés du Sud-est qui sont souvent soumis à
l'action du courant marin froid de Benguela. Ce phénomène est
très fréquent pendant la saison sèche et souvent
observé sur les crêtes de collines.
II.3.2. Facteurs édaphiques
Ø Relief
La Réserve de Biosphère de Luki est
formée par une succession de collines assez jeunes à
l'échelle géologiques dont l'altitude varie entre 150 et 500 m.
Ces collines appartiennent à la chaîne de montagnes appelée
Mayombe.
L'amplitude altitudinale entre le sommet des collines et le
fond des vallées varie de 40 à 70 m et plus dans l'aire centrale.
Les pentes des collines sont parfois très raides jusqu'à 15%. Les
vallées sont très encaissées, ce qui ne permet pas
l'existence des zones marécageuses (Donis, 1948a).
Ø Géologie
Donis (1948a) cité par Lubini (1984) montre que le
soubassement de l'ensemble de la Réserve appartient au système
géologique du Mayombe qui se caractérise par quatre étages
géologiques :
- Etage de la Duizi constitué des schistes
satinés brillants ;
- Etage de Tshela constitué des roches graphiteuses et
grès feldspathiques ;
- Etage de Matadi constitué des quartzites ;
- Etage de Palabala (en amont de Luozi) constitué de
micaschistes et de gneiss satinés, de gneiss, des amphibolites et des
grès graphiteux.
Ø Hydrographie
La Réserve de Biosphère de Luki est
essentiellement située dans le bassin hydrographique de la
rivière Luki qui la traverse du Nord-est au Sud-ouest en
décrivant une grande courbe. La Luki se jette dans la rivière
Lukunga après un parcours de 69 Km.
A droite, les affluents de la Luki sont : la
rivière Ntosi ; le principal affluent avec de nombreux ruisseaux
dépendants, notamment la Vombolo, la Nkula, la Nkakala, la Kikolokolo,
la Mbondo, la Mambamba, et la Loba.
Les affluents de gauche sont : la Kikulo, la Mabakosa,
la Tadi, la Monzi, la Lukamba et la Mioka. Ces cours d'eau sont tous
caractérisés par un régime moins abondant de la
diversité halieutique (Mutambwe, 1993).
Ø Sol
Lubini (1984) distingue quatre types de sols dans la
Réserve en fonction des formations de recouvrement. Ce sont :
- Des sols rouges violacés, se développant sur
les amphibolites ;
- Des sols rouges, développés sur les
gneiss ;
- Des sols jaunes se rencontrant sur les gneiss et les
quartzites ;
- Et des sols alluvionnaires, développés sur
les alluvions récentes.
II.3.3. Eléments biotiques
Ø Végétation
La végétation de la Réserve de
Biosphère de Luki est très hétérogène et
diversifiée. Elle va des formations herbeuses aux forêts denses
humides de type semi-sempervirent en passant par des formations
édaphiques sur les sols hydro morphes. Elle regorge une diversité
spécifique qui varie entre 205 et 373 espèces par hectare, avec
une moyenne de 287 (Lubini, 1997).
Ø Flore
Les travaux de Lubini (1997) montrent que la flore de la
Réserve compte environ 1906 espèces dominées
essentiellement des plantes à fleurs (1055 angiospermes) parmi
lesquelles on note approximativement 153 monocotylédones, 902
dicotylédones.
Ø Faune
Les études de Pendje et Baya (1992) montrent que la
Réserve de Biosphère de Luki a une faune diversifiée dont
les classes des poissons, des mammifères, des oiseaux et des reptiles
sont plus exploitées. L'étude de Gata (1997) prouve à
suffisance que la faune ichtyologique qui, autrefois était riche est
aujourd'hui en péril.
II.4. Techniques sylvicoles réalisées
A sa création, l'INEAC organisa une gestion
méthodique qui était basée sur le zonage et sur les
considérations à la fois sylvicoles et agricoles de la
Réserve forestière.
Les considérations sylvo-ager aboutissent à
l'élaboration des méthodes de conversion de futaies dites
« Uniformisation par le bas » et « Uniformisation
par le haut » (Tunguni, 2001). Le travail de Donis (1948a) a
été la première étude sur la sylviculture du
Mayombe avec la référence à la Réserve
forestière de Luki.
II.4.1. Uniformisation par le Haut (UH)
Cette méthode vise la conversion de futaies d'arbres
d'âges multiples réparties par groupes ou pieds d'arbres, en une
futaie tendant vers la régularité en vue d'en améliorer
les conditions de productivité et d'exploitation (Donis et al.,
1951). Elle repose sur l'identification du potentiel d'avenir le mieux
représenté (brins, baliveaux, moyens). Ensuite, une
éclaircie est réalisée au profit de ce peuplement d'avenir
par régulation de la structure et réduction des écarts
d'âges.
Les espèces secondaires sont éliminées
lorsqu'elles sont un obstacle à la croissance des espèces
commerciales. L'éclaircie est accompagnée d'un délianage
systématique. Cette intervention entraine une modification de la
composition floristique et favoriser certaines essences.
En principe, l'uniformisation par le haut (UH) a pour
objectif de créer la situation suivante :
- Une absence de gros bois, sauf en ce qui concerne les
espèces commerciales ;
- La présence de bois moyens d'âges
divers ;
- L'uniformisation de la lumière destinée
à favoriser les régénérations existantes et
nouvelles.
Cette méthode avait concerné sept blocs (48, 49,
50, 51, 52, 53, 55) plus le bloc témoin dans la zone tampon de la
Réserve de Biosphère de Luki couvrant une superficie moyenne
estimée à 225 ha chacun.
II.4.2. Uniformisation par le Bas (UB)
L'uniformisation par le bas était
développée dans les concessions octroyées aux paysans ou
fermiers considérés comme locataires dans un système de
métayage. Cette uniformisation fut pratiquée dans la zone tampon
qui couvre 14 blocs (de 1 à 14) d'une superficie moyenne de 1631
hectares chacun. Cette méthode consistait à faire une coupe
à blanc étoc de la forêt en vue d'effectuer des plantations
de Limba (Terminalia superba), par des techniques
sylvo-agricoles. Elle fut exécutée en 1950 (Toirambe, 2001).
L'INEAC signa de 1949 à 1955 des conventions avec des
privés pour des blocs sylvo-bananiers à Monzi (Bloc 1 à
8). Ainsi furent appliquées les techniques agroforestières de
types sylvo-bananier, sylvo-cacaoyer et sylvo-caféier qui consistaient
à planter des essences forestières dans les bananeraies et les
plantations de cacao et de café. Les essences forestières
utilisées étaient selon l'importance, Terminalia superba,
Milicia excelsa, Nauclea diderrichii (Bilinga), Entandrophragma
angolense (Tiama), Entandrophragma utile (Sipo), etc. (N'landu,
1999).
CHAPITRE III. MATERIELS ET METHODES
III.1. MATERIELS
Sur le terrain, les matériels présentés
ci-dessous, ont été utilisés:
Ø Milieu expérimental
- Deux blocs à uniformisation par le bas UB1 et UB2
dans lesquels nous avions délimité les placettes ;
- Deux placettes dans lesquelles les espèces
végétales des sous - bois ont été
identifiées ;
Ø Outils de terrain
Les outils exploités pour réaliser cette
étude sont les suivants :
- GPS (Global Positioning System) : de marque GARMIN
GPSmap 60CSX, il nous a permis d'effectuer le layonnage afin d'identifier les
espèces végétales des sous - bois de nos deux
différentes placettes (le Bloc 1 et le Bloc 2) situées non
loin du village MONZI (Secteur de BUNDI, Territoire de SEKE BANZA, District de
BAS FLEUVE, Province du BAS - CONGO) ;
- Boussole et Clinomètre associés : pour
orienter le machetteur afin d'avoir une bonne direction de layonnage ;
- Machette : elle a aidé à ouvrir les
layons qui ont facilité le passage en forêt puis l'identification.
Elle a aussi permis de creuser (deux petites trappes) un profil topographique
dans chaque bloc concerné afin de comparer les épaisseurs des
horizons A0 (Litière + Humus) ;
- Latte : de 30 cm de longueur, pour mesurer
l'épaisseur des horizons A0 ;
- Stylographe : pour enregistrer les différentes
informations ou données récoltées ;
- Bloc - notes : dans lesquels les informations
étaient enregistrées.
III.2. METHODES
1° Choix du site
Le choix était fondé sur l'hypothèse
selon que dans les blocs UB où le Limba domine la
végétation, la litière au sol sera homogène et les
conditions ne seront pas très variées tandis que dans les blocs
UH où règnent l'hétérogénéité
en terme de dominance (Milicia excelsa, Pycnanthus angolensis,
Piptadeniastrum africanum, ...) les conditions de développement des
sous - bois seront très variées.
La composition de la litière au sol dans les blocs UH
est hétérogène. Or, la vitesse de sa décomposition
dépend des espèces. D'où dans ces blocs il est difficile
de déterminer les impacts de la litière sur le
développement de la végétation.
2° Les blocs concernés
A cet effet l'identification a été faite dans
deux blocs à uniformisation par le bas (UB1 et UB2) situés dans
la zone tampon de la Réserve de Biosphère de Luki, non loin du
village MONZI (Secteur de BUNDI, Territoire de SEKE BANZA, District de BAS
FLEUVE).
Ces blocs avaient subit une coupe à blanc étoc
en 1948 par les colons puis suivies d'une plantation de Limbas associés
aux cultures. Donc, ils constituent à ce jour une forêt secondaire
adulte.
3° Méthodologie
Nous avons procédé sur le terrain par des
observations directes qui nous ont permis d'avoir la situation globale de la
couverture au sol dans nos deux blocs respectifs (UB1 et UB2).
Nous avons ensuite délimité une placette de 25
ares (= 0,25 ha = 50 m x 50 m = 2500 m2)
par bloc et pour les différents types de végétations
observées dans notre zone d'étude. Nous avons
déterminé l'épaisseur moyen de l'horizon Ao puis nous
avons procédé au test de comparaison des moyennes pour
déterminer s'il existe une différence significative entre
l'épaisseur de l'horizon Ao pour les deux blocs UB1 et UB2.
Enfin, nous avons déterminé les
différentes espèces qui sont présentes dans le sous - bois
de chaque bloc retenu pour notre étude.
CHAPITRE IV. RESULTATS ET DISCUSSION
A. Identification des espèces des sous - bois
des placettes des blocs UB1 et UB2
Après observation des placettes
installées dans les deux blocs UB1 et UB2, nous avons identifié
les espèces suivantes reprises dans le tableau ci - dessous.
a. Bloc UB1
N°
|
Famille
|
Espèces (noms scientifiques)
|
1
|
Anacardiaceae
|
Pseudospondias microcarpa
Memecylon spp
|
2
|
Annonaceae
|
Monodora myristica
|
3
|
Apocynaceae
|
Funtumia elastica
|
4
|
Bignoniaceae
|
Markhamia sessilis
|
5
|
Burseraceae
|
Dacryodes buettneri
|
6
|
Combretaceae
|
Pteleopsis hylodendron
|
7
|
Euphorbiaceae
|
Croton spp
Macaranga spinosa
|
8
|
Fabaceae
|
Milletia drastica
Hylodendron gabunense
Piptadeniastrum africanum
|
9
|
Flacourtiaceae
|
Oncoba welwitschii
|
10
|
Irvingiaceae
|
Irvingia grandifolia
|
11
|
Meliaceae
|
Trichilia spp
Carapa procera
Lovoa trichiloïdes
Entandrophragma angolense
|
12
|
Moraceae
|
Trilepisium madagascariensis
Musanga cecropioïdes
Myrianthus arboreus
Antiaris welwitschii
|
13
|
Myristicaceae
|
Pycnanthus angolensis
Staudtia kamerunensis
|
14
|
Rutaceae
|
Zanthoxylum kivuensis
|
15
|
Sapindaceae
|
Blighia spp
|
16
|
Sterculiaceae
|
Sterculia tragacantha
Cola brunelli
|
17
|
Ulmaceae
|
Holoptelea grandis
Celtis soyauxii
|
|
b. Bloc UB2
|
N°
|
Famille
|
Espèces (noms scientifiques)
|
|
1
|
Anacardiaceae
|
Lannea welwitschii
Pseudospondias macrocarpa
|
|
2
|
Annonaceae
|
Monodora myristica
|
|
3
|
Apocynaceae
|
Alstonia bonnei
Funtumia elastica
|
|
4
|
Burseraceae
|
Dacryodes buettneri
|
|
5
|
Euphorbiaceae
|
Ricinodendron heudelottii
Croton spp
Macaranga spinosa
|
|
6
|
Fabaceae
|
Milletia versicolor
Milletia drastica
Amphimas spp
Albizzia ferruginea
Piptadeniastrum africanum
|
|
7
|
Meliaceae
|
Entandrophragma angolense
Entandrophragma candollei
Carapa procera
|
|
8
|
Moraceae
|
Trilepisium madagascariensis
Musanga cecropioïdes
Myrianthus arboreus
Milicia excelsa
|
|
9
|
Rutaceae
|
Zanthoxylum kivuensis
|
|
10
|
Sapindaceae
|
Blighia spp
|
|
11
|
Sterculiaceae
|
Sterculia tragacantha
|
|
12
|
Ulmaceae
|
Holoptelea grandis
|
Lors de l'identification, nous avons constaté que dans
le bloc UB2 le passage était facile que dans le bloc UB1. Ce qui nous a
conduits à introduire la notion de densité (nombre d'arbres par
unité de surface) qui est fonction de la richesse d'un sol en
éléments nutritifs.
Les sous - bois du bloc UB1 étaient denses par rapport
à ceux de UB2. Ainsi c'est sur base de nos observations directes que
nous étions à mesure de conclure que la couverture au sol
(débris ligneux, litières, ...) était importante dans le
bloc UB1 que dans le bloc UB2.
B. Epaisseurs des horizons A0 dans les
blocs UB1 et UB2
Ø Comparaison des épaisseurs moyens des
horizons A0 dans les blocs UB1 et UB2
En ce qui concerne l'épaisseur de l'horizon
A0 du sol (donc la première couche du profil), les trappes
(petits dispositifs de forme carrée) que nous avions creusées
dans différents endroits de ces placettes nous ont permis d'obtenir les
résultats ci - après et présentés dans le tableau
suivant.
Tableau 3 : Epaisseur des horizons A0 dans
les trappes
|
Epaisseur couche (Cm)
|
|
N°
|
UB1
|
UB2
|
|
1
2
3
4
5
6
|
6,8
8,6
7,8
5,9
9,6
8,1
|
5,7
7,0
4,5
4,4
5,7
8,1
|
|
7,8
|
5,9
|
|
46,8
|
35,4
|
|
373,62
|
219,2
|
Test d'hypothèse d'égalité de deux
moyennes par l'ANOVA



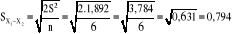

 Avec
Avec . On rejette l'hypothèse nulle.
. On rejette l'hypothèse nulle.
Le test d'hypothèse ci-haut fait, de comparaison de
deux moyennes, nous montre qu'il y a une différence significative entre
l'épaisseur moyen de l'horizon Ao pour les deux blocs UB1 et UB2 du fait
que l'hypothèse nulle (Ho) d'égalité entre ces moyennes
est rejetée. Donc, les conditions de développement, des
espèces forestières, sont différentes dans nos deux blocs
respectifs.
Selon que l'épaisseur de l'horizon A0
constituée de la matière organique décomposée nous
permet d'apprécier la richesse d'un sol en termes
d'éléments nutritifs, ces résultats nous montrent que le
sol du bloc UB1 où l'épaisseur moyen de l'horizon
A0 est 7,8 cm est plus fertile que celui du bloc UB2 où
l'épaisseur moyen de l'horizon A0 est de 5,9
cm.
C. Les débris ligneux
Concernant les débris ligneux au sol, signalons qu'ils
étaient importants dans le bloc UB1 que dans le bloc UB2. A cet effet,
nous pouvons dire qu'ils étaient récoltés pour servir
comme bois de feu par la population locale selon que le bois constitue la
principale source d'énergie des villages autour et dans la
Réserve de Biosphère de Luki.
CONCLUSION ET SUGGESTION
Ce travail portant sur l'importance de la couverture au sol
dans la restauration des écosystèmes forestiers : Cas de la
Réserve de Biosphère de Luki a consisté à comparer
la couverture au sol dans deux blocs différents dominés par le
Terminalia superba (Limba) ainsi qu'à identifier les
espèces de sous-bois de ces blocs.
En effet, nous avons eu à comparer l'épaisseur
moyen de l'horizon Ao, du profil topographique, de la litière en
décomposition de 6 trappes installées dans chaque placette et
dans chaque bloc, ainsi que les débris ligneux associés à
la litière non décomposée au sol alors que nous avions
identifier 30 espèces groupées en 17 familles dans la placette
du bloc UB1 et 25 espèces groupées en 12 familles dans celle du
bloc UB2.
Il ressort de cette étude que la placette du bloc UB1
présente un épaisseur moyen de l'horizon Ao relativement
supérieur à celui de la placette du bloc UB2. En effet,
l'épaisseur moyen de l'horizon Ao de la placette du bloc UB1 est de 7,8
cm alors que celui de la placette du bloc UB2 est de 5,9 cm. La couverture
au sol dans le bloc UB1 est caractérisée par une quantité
importante de débris ligneux contrairement à celle
observée dans le bloc UB2 ce qui a favorisé une bonne
régénération naturelle des espèces de sous-bois du
premier bloc.
En outre, les résultats de notre étude
corroborent ceux d'Ebenmayer (1976) qui confirme que le
développement des essences forestières est lié au maintien
de la couverture au sol.
Il sied de mentionner que toute initiative de mise en
défens des savanes anthropiques comme celle réalisée par
le World Wide Found for nature, WWF en sigle, dans la Réserve de
Biosphère de Luki au Bas-Congo depuis 2006, favorise le rôle de la
couverture au sol dans la restauration des écosystèmes
forestiers.
Tout compte fait, nous pouvons suggérer à
tous les gestionnaires des aires protégées en
général et de la Réserve de Biosphère de Luki en
particulier ainsi que des forêts d'avoir une vision très
particulière à la couverture au sol car elle contribue à
la bonne croissance des arbres de plus de 8 m de haut tout en maintenant la
fertilité du sol. Nous proposons enfin que les séances de
sensibilisations soient organisées auprès des communautés
locales pour qu'elles sachent l'importance et la place de cette couverture dans
la restauration des forêts afin de les gérer durablement.
REFERENCES BIBLIOGRAPHIQUES
Anonyme, 2009. Géologie, dépôts de surface et
sols forestiers, In: Anonyme (Eds), Manuel de foresterie : Nouvelle
édition entièrement revue et augmentée, Multi
mondes ; Québec, pp 47-124.
Avila H., 1980. Cité par Plamondon A., 2009. Bassins
versants et érosion : application en forêt tropicale humide,
Notes de cours, 3ème Graduat Foresterie, Faculté de
Foresterie, de Géomatique et de Géographie, Université
Laval, Québec, 287p.
Bray J.R., et Gorham R., 1964. Litter production in forests of
the world, Adv. Ecol. Res. 2: 101-157.
Cole D.W., et Rapp M., 1981. Elemental cycling in forest
ecosystems, dans D.E. Reichle (ed.), Dynamic properties of forest ecosystems,
IBP 23, Cambridge Univ., Press, Cambridge, p 341-409.
Côté B., Fyles W et Djalilvand H., 2002.
Increasing N and P resorption efficiency and proficiency in northern deciduous
hardwoods with decreasing foliar N and P concentrations, Ann. For. Sci:
59 : 275-281
Donis C., 1948a. Essai d'économie forestière au
Mayombe. Publications de l'INEAC, série scientifique, 37, Bruxelles,
92p.
Donis et al., 1951. Cité par Angoboy B., 2006. Etat
actuel des arboreta dans la Réserve de Biosphère de Luki au
Mayombe, memoire, inédit, Faculté des sciences agronomiques,
Université de Kinshasa.
Ebenmayer E., 1976. Die gesammte lehre derwaldstreu mit
Rucksicht auf die chemische Statik des waldbaues. J. Springer, Berlin.
Gallardo A., et Merino I., 1993, Leaf decomposition in two
Mediterranean ecosystems of southern Spain: influence of substrate quality,
Ecology. 74: 152-161.
Gata D., 1997. Étude des impacts humains, estimation
de degré de péril de la biodiversité et principe
directeurs pour une gestion durable de ressources disponibles. MAB/Congo,
MECNT, 37p.
Hewlett J., 1972. Cité par Plamondon A., 2009. Bassins
versants et érosion : application en forêt tropicale humide,
Notes de cours, 3ème Graduat Foresterie, Faculté de
Foresterie, de Géomatique et de Géographie, Université
Laval, Québec, 287p.
Hewlett J., 1982. Principles of forest hydrology, University of
Georgia, Press, Athens, GA, USA, 183p.
Hillel M., 1971. Cité par Plamondon A., 2009. Bassins
versants et érosion : application en forêt tropicale humide,
Notes de cours, 3ème Graduat Foresterie, Faculté de
Foresterie, de Géomatique et de Géographie, Université
Laval, Québec, 287p.
Hoover B., 1962. Cité par Plamondon A., 2009. Bassins
versants et érosion : application en forêt tropicale humide,
Notes de cours, 3ème Graduat Foresterie, Faculté de
Foresterie, de Géomatique et de Géographie, Université
Laval, Québec, 287p.
Kapa B., Malele M., Nkiama M. et Ritvisay M., 1987.
Aménagement de la Réserve Biosphérique de la Luki. SPIAF,
67p.
Lamontagne S., Carignan R., D'Arcy P., Prairie L. et
Paré D., 2000. Element export in runoff from eastern Canadian Boreal
shield drainage basins following forest harvesting and wildfires can. J. Fish.
Aquat. Sci. 57. (Suppl 29:118-128).
Lassen et al., 1955. Cités par Plamondon A., 2009.
Bassins versants et érosion : application en forêt tropicale
humide, Notes de cours, 3ème Graduat Foresterie,
Faculté de Foresterie, de Géomatique et de Géographie,
Université Laval, Québec, 287p.
Lubini A., 1984. La Réserve de Biosphère de
Luki, MAB/Congo, MECNT, Kinshasa, 92p.
Lubini A., 1997. La végétation de la
Réserve de Biosphère de Luki au Mayombe (Zaïre). Jard. Bot.
Nat. Bel. Opéra Botanica 10, 155p
Maldague M., Mankoto S. et Rakotomavo T., 1997. Notions
d'aménagement et de développement intégré des
forets tropicales. ERAIFT, UNESCO, MAB, PNUD, 375p.
Marschner H., 1994. Mineral nutrition's of higher plants,
2ème éd., Academic Press, London, 889p.
Mc Kinnon J., Child D. et Thorsell J., 1990.
Aménagement et gestion des aires protégées tropicales,
UICN, Gland, Suisse, 289p.
Melillo J., Aber D. et Muratore F., 1982, Nitrogen and lignin
control of hardwood leaf litter decomposition dynamics, Ecology: 63:
621-626.
Mutambwe S., 1993. Présentation de la Luki et de son
bassin versant, MAB/Zaïre, 53p.
Myré R., et
Camiré C., 1994. The
establishment of stem nutrient distribution zones of
European larch and tamarack using principal component analysis, Trees 9:
26-34
N'Landu J., 1999. Rapport du chargé des travaux
forestiers sur l'État actuel de la réserve de biosphère de
Luki. Projet MAB-Luki, 3p.
Nsenga L., 2001. Étude de la gestion des aires
protégées en République Démocratique du Congo, cas
de la Reserve de Biosphère de Luki-Mayombe; 139p.
Ouimet R., et Duchesne L., 2008. Dépôts
atmosphériques dans les forêts au Québec :
retombées actuelles en forêt et tendances au cours des 20 à
30 dernières années, Naturaliste canadien (Sous presse).
Paré D., Lamontagne S. et Prairie L., 2002. Assessing
the geochemical of balance of managed boreal forests, Ecol. Indicators 1:
293-311
Pendje G. et Baya M., 1992. La Réserve de
Biosphère de Luki (Mayombe, Zaïre) : Patrimoine floristique et
faunique en péril. UNESCO, Paris, 62p.
Plamondon A., 2009. Bassins versants et érosion :
application en forêt tropicale humide, notes de cours,
3ème Graduat Foresterie, Faculté de Foresterie, de
Géomatique et de Géographie, Université Laval,
Québec, 287p.
Stocking M. et Murnagan G., 2001. Cités par Plamondon
A., 2009. Bassins versants et érosion : application en forêt
tropicale humide, Notes de cours, 3ème Graduat Foresterie,
Faculté de Foresterie, de Géomatique et de Géographie,
Université Laval, Québec, 287p.
Swift F., Huber L. et Schull H., 1979, Decomposition in
terrestrial ecosystems, Univ. California Press, Bervelly, 384p.
Taylor G. et Bradle J., 1989, Nitrogen and Lignin content as
predictors of Litter decay rates: a microcosm test, Ecology. 70: 97-104.
Toirambe B., 2001. Valorisation des produits forestiers non
ligneux (PFNL) dans la Réserve de Biosphère de Luki Bas-Congo.
Mémoire inédit de DESS, ERAIFT, 163p.
Toirambe B., 2002. Cité par Angoboy B., 2006. Etat
actuel des arboreta dans la Réserve de Biosphère de Luki au
Mayombe, memoire, inédit, Faculté des sciences agronomiques,
Université de Kinshasa.
Trofymow S. et Delph L., 2002, Rates of litter decomposition
over 6 years in Canadian forests: influence of litter quality and climate, can.
J. For. Res. 32: 789-804.
Tunguni S., 2001. Cité par Angoboy B., 2006. Etat
actuel des arboreta dans la Réserve de Biosphère de Luki au
Mayombe, memoire, inédit, Faculté des sciences agronomiques,
Université de Kinshasa.
UN/FAO., 1991. Report of the Seventh Session of the FAO. Panel
of Experts on Forest Gene Resources. December, 1989, FAO, Rome.
Wiersum L., 1985. Cité par Plamondon A., 2009. Bassins
versants et érosion : application en forêt tropicale humide,
Notes de cours, 3ème Graduat Foresterie, Faculté de
Foresterie, de Géomatique et de Géographie, Université
Laval, Québec, 287p.
Wiersum L., 1983. Cité par Plamondon A., 2009. Bassins
versants et érosion : application en forêt tropicale humide,
Notes de cours, 3ème Graduat Foresterie, Faculté de
Foresterie, de Géomatique et de Géographie, Université
Laval, Québec, 287p.
TABLE DES MATIERES
DEDICACE 1
AVANT-PROPOS 2
INTRODUCTION 3
CHAPITRE I : REVUE DE LA LITTERATURE 5
I.1. Définition de la couverture au sol 5
I.2. Différents types des couvertures au sol 5
I.3. Cycles des éléments et nutrition des essences
forestières 6
I.4. Autres concepts 13
I.5. Importance de la couverture au sol 15
CHAPITRE II : PRESENTATION DE LA RESERVE DE BIOSPHERE DE
LUKI 18
II.1. Situation géographique 18
II.2. Création et historique 18
II. 3. Cadre mésologique de la réserve 19
II.4. Techniques sylvicoles réalisées 21
CHAPITRE III : MATERIELS ET METHODES 25
III.1. Matériels 25
III.2. Méthodes 25
CHAPITRE IV : RESULTATS ET DISCUSSION 26
CONCLUSION ET SUGGESTION 30
REFERENCES BIBLIOGRAPHIQUES 31
TABLE DES MATIERES 35


