Chapitre 3
Tâche désincarnée : la
tâche
vectorielle abstraite
Dans ce chapitre, nous allons présenter la
première expérience pour évaluer notre modèle de la
mRF. Dans un premier temps, nous détaillerons l'expérience
réalisée, et dans un second temps nous analyserons les
résultats.
3.1 Expérience
La tàache d'esincarn'ee correspond a` une tàache
abstraite de s'election. Le principe est simple : nous donnons en entr'ee de la
mRF un vecteur de dimension 3, toutes les valeurs 'etant comprises entre 0 et
1. Ce vecteur d'entr'ee repr'esente un ensemble de valeurs issues de diverses
sources de donn'ees.
L'objectif de la mRF est de s'electionner une action. Pour
cela, nous avons d'ecid'e en nous inspirant de [Humphries et al., 2007] que
pour que la mRF choississe la bonne action, son vecteur de sortie, 'egalement
de dimension 3, doit avoir sa valeur maximum dans la même dimension que
celle contenant la valeur maximum du vecteur d'entr'ee. Par exemple, si le
vecteur d'entr'ee est (0.3, 0.5, 0.1), la mRF s'electionnera la bonne action si
et seulement si son vecteur de sortie est de la forme (a, b, c), o`u b > a
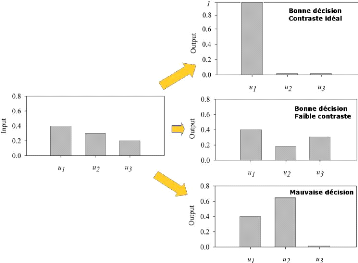
et b > c, comme le montre la figure 3.1.
Afin que l''evolution ne sur-apprenne pas un ensemble
restreint de vecteurs, ce qui empêcherait une bonne g'en'eralisation, la
mRF sera 'evalu'ee sur sa capacit'e a` s'electionner une action sur l'ensemble
de vecteurs de dimension 3 o`u nous faisons varier chaque 'el'ement d'un entre
0 et 1 d'un pas de 0.1, et en supprimant les vecteurs ayant plusieurs
composantes au maximum, afin d''eliminer les cas ambigus. Voici
contrast(X) = tu u u v
o`u k = arg max
n-1
i
xi.
?
??
(xi - xk
Xn
i=1
?
)2 ? ?
FIGURE 3.1: Ce schéma montre les deux premiers
objectifs de la tâche désincarnée : le réseau
recoit en entrée un vecteur de dimensions 3 dont les valeurs
sont entre 0 et 1, et il doit a` la fois prendre la bonne décision
(c'est-à-dire sélectionner la bonne dimension) et maximiser le
contraste.
cet ensemble de vecteurs 'ecrit formellement, ensemble qui
contient en tout 1155 { 'el'ements : }
X = (x1, x2, x3) | (x1, x2, x3) E S3 A (k = arg max
xi, -+ ((i =6 k A i E 1, 3K) -+ x > xi)
i
o`u S = {0.1 x i | i E [0, 10K}
N'eanmoins, ce premier objectif admet des solutions
inint'eressantes, comme par exemple un r'eseau de neurones qui copieraient
simplement les entr'ees en sortie. Afin d'op'erer une r'eelle s'election de
l'action, nous avons d'efini un second objectif forçant la mRF a`
maximiser le contraste entre la valeur maximum du vecteur de sortie et ses
autres valeurs. La formule g'en'erale pour calculer le contraste est la
suivante, en notant xi le ième 'el'ement du vecteur X et n sa
dimension :
/((0.6_0.5)2+(0.6_0.3)2)
contrast = 0.22. Plus le contraste est élevé,
plus l'action
2
est clairement sélectionnée.
Le troisième et dernier objectif sera de respecter les
contraintes anatomiques, pour que l'évolution produise des
réseaux se rapprochant le plus possible d'une structure de réseau
de type mRF. Le score de cet objectif de plausibilitéanatomiques
correspond a` une combinaison linéaire de scores de sous-objectifs, ces
derniers représentant chacun une des contraintes anatomique suivante
(nous reprenons les notations du chapitre 1.3) :
- p doit être égal a` 80%, la formule du score est
scorep = (-1) × (0.8 - p)2 ;
- P(c) doit être égal a` 25%, la formule du score
est scorepc = (-1) × (0.25 - pc)2 ; - les synapses
des neurones de projection sont a` 45% GABAergic, la formule du score
est scoregabaergic = (-1) × (0.45 -
percentgabaergic)2 ;
- les interneurones ne peuvent pas projeter en dehors de leur
cluster, nous notons nb interneuron not within chip le nombre d'interneurones
ne respectant pas cette contrainte;
- les neurones de projection ne peuvent pas projetter dans
leur cluster, nous no-tons nb projection within chip le nombre de projection ne
respectant pas cette contrainte.
Nous n'avons pas mis de contraintes sur P(p) et P(l) car les
données anatomiques connues a` leur sujet sont assez imprécises.
Comme nous l'avons soulignédans le chapitre 2.2, les algorithmes
évolutionnistes maximisent les objectifs, d'o`u les scores
négatifs puisque se rapprocher des contraintes équivaut a` un
score se rapprochant de la valeur 0.
La formule globale pour calculer le score du troisième
objectif est :
scoreanat =
8×(scorep+scorepc+scoregabaergic)-1×nb interneuron not within chip-
0.5 × nb projection within chip
Les pondérations 8, 1 et 0.5 ont
étéchoisies arbitrairement, le seul critère étant
que chacun des scores pondérés ait une valeur très
approximativement du même ordre de grandeur, afin de maximiser les
chances que l'évolution optimise chacun d'entre eux, et non juste de
certains potentiellement aux dépens des autres.
En résumé, voici les trois objectifs que nous avons
retenus :
- Objectif 1 : nombre de bonnes décisions. Minimum : 0,
maximum : nombre de vecteurs testés;
- Objectif 2 : importance du contraste. Minimum : 0, maximum : 1
(car toutes les valeurs des vecteurs sont comprises entre 0 et 1);
- Objectif 3 : plausibilitéanatomique. Minimum : -8,
maximum : 0.
En plus de ces trois objectifs situ'es en aval, nous avons
'egalement pos'e des contraintes anatomiques en amont, c'est-à-dire au
coeur de la d'efinition des op'erateurs de mutation, afin de pousser
l''evolution a` respecter le plus possible les donn'ees anatomiques. Nous
avions d'ejàmentionn'e dans le chapitre 2.2.2 cette double application
des contraintes. Voici les contraintes que nous avons mises dans les
op'erateurs de mutation :
- impossibilit'e pour un neuron de projection de projeter dans
leur cluster;
- impossibilit'e pour un internerneurone de projeter en dehors de
leur cluster; - P(c) doit être proche 25%;
- p doit être proche 80%.
Comme nous le voyons, ces contraintes ne contiennent qu'une
partie des donn'ees anatomiques connues : l'int'erêt de les ajouter dans
les op'erateurs de mutation est que l''evolution g'enère moins
d'individus manifestement non plausibles d'un point de vue anatomique. Mettre
trop de contraintes dans les op'erateurs des mutations serait n'eanmoins
risqu'e car, outre son coit computationnel, restreindre excessivement l'espace
de recherche peut rendre plus difficile l'obtention d'une bonne solution.
Nonobstant sa simplicit'e apparente, cette tàache
d'esincarn'ee permet de valider ou d'invalider la possibilit'e pour une
structure anatomiquement proche de la mRF d'effectuer des s'elections. Nous
allons analyser les r'esultats obtenus dans la section suivante.
3.2 Résultats
Tout d'abord, voici les paramètres que nous utiliserons
tout au long des exp'eriences, sauf indication contraire :
- taille de la population : 500 individus;
- nombre de g'en'erations : 500;
- nombre de clusters : 4;
- nombre de neurones initial par cluster (en plus des neurones
d'entr'ee et de sortie) : tirage al'etoire uniforme entre 3 et 10, les clusters
pouvant avoir un nombre de neurones diff'erents au sein de la même
mRF;
- probabilit'e d'ajouter/supprimer un neurone/une connexion
(inter ou intra-cluster) : 0.05;
- probabilit'e de modifier le poids d'une connexion (inter ou
intra-cluster) : 0.1; - nombre d'it'erations pour propager une entr'ee dans le
r'eseau : 100.
Les 'evolutions calcul'ees nous donnent des individus
pr'esentant au bout de 300 g'en'e-
rations plus de 95% de bonnes d'ecisions,
voire dans certains cas 100%, respectant
presque parfaitement les
contraintes anatomiques et dont les vecteurs de sortie ont
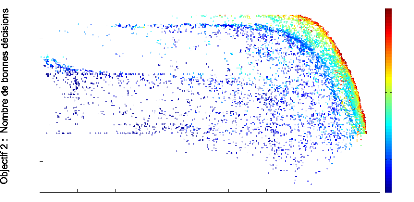
un contraste supérieur a` 0.6. Le graphique 3.2 montre
un front de Pareto 2D obtenu typiquement lorsque l'on représente les
scores des objectifs de bonnes décisions et de contaste obtenus par
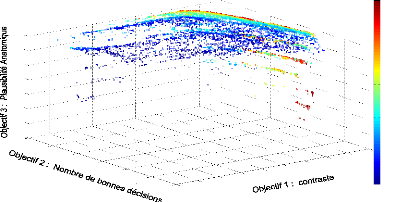
chaque individu. Le graphique 3.3 montre un front de Pareto 3D montrant les
scores des 3 objectifs.
1200
Numéro de génération
500
1000
400
800
300
600
200
100
0
400
200
0
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9
Objectif 1 : contraste
FIGURE 3.2: Front de Pareto de dimension 2 repr'esentant les
scores des objectifs de contaste en abscisse et de bonnes d'ecisions en
ordonn'ee obtenus par chaque individu de chaque g'en'eration au cours de
l''evolution.
Prenons les 5 meilleurs individus présentant 1155 de
bonnes décisions et regardons la moyenne de leurs résultats :
- score de nombre de bonnes décisions : 1155 (ce qui
correspond au maximum théorique);
- score de contraste : 0.68761 (le maximum théorique est
1);
- score de plausibilitéanatomique : -1.20792e-15 (le
maximum théorique est 0).
Le tableau suivant résuméles statistiques
anatomiques moyennes de ces 5 individus.
Numéro de génération
0
0
-1
-2
-3
-4
-5
-6
-7
1200
1000
800
0.7
0.8
600
400
200
0.2
0.3
0.4 0.5
0.6
0
0
0.1
500
400
300
200
100
FIGURE 3.3: Front de Pareto de dimension 3 repr'esentant les
scores des objectifs de bonnes d'ecisions, de contaste ainsi que de
plausibilit'e anatomique obtenus par chaque individu de chaque g'en'eration au
cours de l''evolution.
|
Statistiques sur
les 5 meilleurs
individus
|
Contrainte
dans op'erateurs
de mutation
|
Contrainte
dans objectif
|
Données
anatomiques
connues
|
|
nombre total
de neurones
|
57
|
Aucune
|
Aucune
|
NA
|
|
nombre total
de connexions
|
117
|
Aucune
|
Aucune
|
NA
|
|
p
|
80%
|
80%
|
80%
|
80%
|
|
percentgabaergic
|
45%
|
Aucune
|
45%
|
45%
|
|
P(c)
|
25%
|
25%
|
25%
|
25%
|
|
P(p)
|
8.73626%
|
Aucune
|
Aucune
|
< 10%
|
|
P(l)
|
8.61552%
|
Aucune
|
Aucune
|
< 10%
|
|
Nombre de
interneurones
projettant en
dehors de
leur
cluster
|
0
|
0
|
0
|
0
|
|
Nombre de
neurones de
projection
projettant
dans
leur cluster
|
0
|
0
|
0
|
0
|
Ce résultat montre qu'en respectant les données
anatomiques connues de la mRF, cette dernière peut effectuer une
tàache de sélection de l'action. Pour comparaison, [Humphries et
al., 2007] dont cette tàache est inspirée obtient environ 75% de
bonnes décisions, sans se pencher sur le contraste, et considère
que cela suffit pour montrer la possibilitéd'une sélection de
l'action.
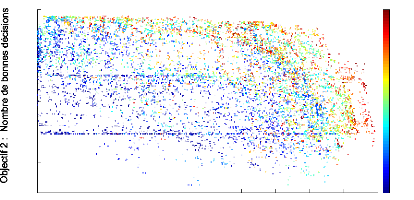
Nous avons également fait tourner le programme en
enlevant les contraintes situées dans les opérateurs de mutation.
Comme le montre le graphique 3.4, cela a pour effet d'augmenter le nombre
d'individus ayant une faible plausibilitéanatomique, car nous voyons que
le front 2D représentant les scores des objectifs de bonnes
décisions et de contaste obtenus par chaque individu est beaucoup moins
nette que celui obtenu dans l'expérience initiale dont nous venons de
présenter les résultats. Cette observation correspond a`
l'intuition que nous avions. Néanmoins l'évolution
présente toujours des individus, certes plus rares que dans
l'expérience initiale, presque parfaitement plausibles anatomiquement,
de contraste supérieur a` 0.5 et dont le taux de bonnes décisions
est supérieur a` 95%.
1
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9
Numéro de génération
1200 400
1000
300
800
200
600
400
200
100
0
Objectif 1 : contraste
FIGURE 3.4: Front de Pareto 2D sans contraintes anatomiques
lors des mutations. Le front represente les scores des objectifs de bonnes
decisions et de contaste obtenus par chaque individu.
Une autre variante que nous avons essayée est d'enlever
les contraintes des opérateurs
des mutations et d'éliminer en
plus l'objectif de plausibilitéanatomique. Autrement
dit, nous avons
cherchéa` voir comment évoluerait un réseau sans
contrainte anato-
mique vis-à-vis de la tàache de
sélection.
Les résultats montrent que d'une part les
réseaux obtenus ont des performances similaires a` celles obtenues avec
les réseaux contraints, voire légèrement meilleures (cf
graphique 3.5) si nous considèrons le contraste, et que d'autre part en
faisant des analyses statistiques sur la structure de ces réseaux, ils
ne tendent pas vers des structures de type mRF. Cela signifie que les
données anatomiques connues sur la mRF ne représentent ni un
avantage (car il existe d'autres structures de réseau tout aussi
performantes) ni un inconvénient pour la sélection.
1000
300
200
0
1
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9
Numéro de génération
1200
800
200
600
100
400
Objectif 1 : contraste
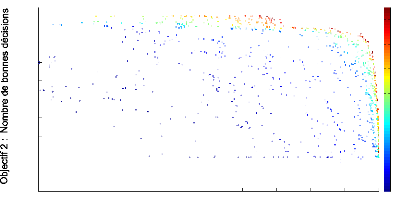
FIGURE 3.5: Front de Pareto 2D de l''evolution sans aucune
contrainte anatomique (ni dans les mutations, ni dans un objectif). Le front
repr'esente les scores des objectifs de contaste en abscisse et de bonnes
d'ecisions en ordonn'ee obtenus par chaque individu de chaque g'en'eration au
cours de l''evolution.
Cette première série d'expériences
basées sur une tàache désincarnée a
montréla
capacitécomputationnelle de la mRF a` effectuer une
tàache de sélection. Notre modèle
est plus performant que celui de Humphries dans cette
tàache et nous y avons ajoutécomme contrainte
supplémentaire le contraste. Nous allons a` présent effectuer
une
seconde série d'expériences pour analyser les
performances de la mRF sur une tàache incarnée de simulation
robotique.
| 

