| 
UNIVERSITE D'ANTANANARIVO
DEPARTEMENT DE BIOLOGIE ET ECOLOGIE VEGETALES
FACULTE DES SCIENCES Programme MEM « Modélisation pour l'Environnement à Madagascar »
Projet ANR DEDUCTION Mémoire pour l'obtention du Diplôme d'Etudes Approfondies (D.E.A.)
Biologie et Ecologie végétales
Option Ecologie Végétale DYNAMIQUE DES ECOSYSTEMES FORESTIERS
EN CONTACT AVEC LES SAVANES DANS LE
CORRIDOR FORESTIER DE FIANARANTSOA
(Sahabe-Ambohimahamasina) RANDRIANARISON A., 2008 Présenté par :
Andry RANDRIANARISON
(Maitre-ès sciences) Soutenu publiquement, le 22 Août 2009, devant les membres du jury composé de : Président du jury: Pr Charlotte RAJERIARISON Rapporteur : Dr Samuel RAZANAKA Rapporteur : Pr Vonjison RAKOTOARIMANANA Examinateur : Dr Dominique HERVE Examinateur : Dr Zo Lalaina RAZAFIARISON UNIVERSITE D'ANTANANARIVO
DEPARTEMENT DE BIOLOGIE ET ECOLOGIE VEGETALES
FACULTE DES SCIENCES 

Programme MEM « Modélisation pour l'Environnement à Madagascar »
Projet ANR DEDUCTION Mémoire pour l'obtention du Diplôme d'Etudes Approfondies (D.E.A.)
Biologie et Ecologie végétales
Option Ecologie Végétale DYNAMIQUE DES ECOSYSTEMES FORESTIERS EN
CONTACT AVEC LES SAVANES DANS LE CORRIDOR
FORESTIER DE FIANARANTSOA
(Sahabe-Ambohimahamasina) Présenté par :
Andry RANDRIANARISON
(Maitre-ès sciences) Soutenu publiquement, le 22 Août 2009,
devant les membres du jury composé de : Président du jury: Pr Charlotte RAJERIARISON Rapporteur : Dr Samuel RAZANAKA Rapporteur : Pr Vonjison RAKOTOARIMANANA Examinateur : Dr Dominique HERVE Examinateur : Dr Zo Lalaina RAZAFIARISON A Lt vv~vvoi,re ce vvov pêre «Ce ne sont pas les fils qui perpétuent la mémoire des pères, ce sont les bonnes actions et les bonnes moeurs » Alexandre III le grand. REMERCIEMENTS Ce travail a été réalisé au sein du programme MEM (Modélisation pour l'Environnement à Madagascar), une convention entre l'IRD (Institut de Recherche pour le Développement) et l'université de Fianarantsoa. Grâce à ce programme, le stage sur le terrain et la rédaction de ce mémoire se sont déroulés dans des conditions adéquates. Aussi, nous adressons nos vifs et sincères remerciements aux personnes qui nous ont apporté leurs aides et leurs conseils car sans eux, ce travail n'aurait pas pu être achevé. Nous remercions tout spécialement : · Madame Charlotte RAJERIARISON, Professeur titulaire, responsable du troisième cycle, Option Ecologie Végétale au sein du Département de Biologie et Ecologie Végétales, Faculté des sciences de nous faire l'honneur d' accepter la présidence ; vous avez fait preuve de confiance, de patience et de tolérance à notre égard ; vos qualités nous ont séduit et resteront pour nous des exemples difficiles à suivre ; nous voudrions exprimer ici la grande estime que nous avons pour vous et notre profonde et respectueuse gratitude ; · Monsieur Samuel RAZANAKA, Docteur d'Etat en Ecologie, enseignant-chercheur au sein du CNRE, qui a bien voulu défendre ce travail et qui nous a toujours montré rigueur scientifique, droiture et bienveillance. Qu'il trouve ici l'expression de notre gratitude et de notre profonde estime ; · Monsieur Vonjison RAKOTOARIMANANA, Professeur en Ecologie, titulaire d'une HDR au sein du Département de Biologie et Ecologie Végétales, pour la grande disponibilité et les accueils bienveillants que vous nous avez toujours réservé malgré vos nombreuses occupations. Pour l'honneur que vous nous faites en acceptant sans hésitation d'être le rapporteur de ce mémoire. Que ce modeste travail soit un humble témoignage de notre profonde reconnaissance et de nos remerciements les plus sincères ; · Monsieur Dominique HERVE, Docteur en agronomie, chargé de recherche à l'IRD et co-directeur du programme MEM, qui nous a toujours accueilli avec sympathie en dépit de ses multiples occupations ; vous nous avez accordé votre confiance et votre amitié et vous avez bien voulu accepter d'être l'examinateur de ce travail qui est le vôtre. Nous tenons à vous adresser nos vifs remerciements et nos plus sincères reconnaissances ;
· Monsieur Zo Lalaina RAZAFIARISON, Docteur en sociologie, option environnement et développement durable au sein du Département de Biologie et Ecologie Végétales qui nous a témoigné son aimabilité en acceptant de juger ce mémoire. En remerciement de son aimable sollicitude à notre égard, qu'il trouve ici l'expression de notre profonde gratitude et de nos très sincères reconnaissances ; · Le corps enseignant du Département de Biologie et Ecologie Végétales ainsi qu'à tous les enseignants de la Faculté des Sciences de l'Université d'Antananarivo. Qu'ils reçoivent ici l'expression de notre reconnaissance pour l'aide si précieuse et les conseils qu'ils nous ont prodigué tout au long de nos études ; · Toute l'équipe du programme MEM pour les aides et conseils ainsi que les ambiances sympathiques créées dans le laboratoire durant notre stage au sein du programme ; · La famille Jean RABELAHY de nous avoir accueilli dans leur campement, ainsi que la famille Ernest RAJOMALAHY pour leur aide et d'avoir accepté d'être notre guide lors des travaux sur terrain ; · Les autorités locales de la commune rurale d'Ambohimahamasina, Monsieur Pierre TSABO, maire de la commune, Monsieur Venance RALOHOTSY, Président du Fokontany d'Andohanimananatanana et Monsieur Samuel RALAIVAO, président du VOI lovasoa d'Andohanimananatanana, de nous avoir bien accueilli au sein de la communauté ; · La famille et les ami(e) s. Vous avez beaucoup fait pour nous ; que ce travail soit un modeste témoignage de notre reconnaissance infinie et de nos remerciements les plus sincères ; Enfin, je suis très heureux d'associer à ces remerciements, toutes les personnes qui ont contribué, de près ou de loin, à la réalisation de ce travail ainsi que tous les habitants du village de Sahabe, de Sahamaina, d'Andepo, de Vohimana et le campement d'Ambinaniranomena. GLOSSAIRE Alamainty : Forêt mature peu perturbée. Alamena : Forêt perturbée ou forêt mature de crête ou de pente. Betsileo : La dénomination générique des populations occupant la partie sud des terres centrales de Madagascar. Campement : Habitation en zone forestière. Corridor forestier : Pont biologique reliant deux massifs forestiers. Dahalo : Bandit, voleur de zébus. Dynamique : Transformation spontanée et lente, au cours de laquelle, des groupements végétaux différents se succèdent en chaque point. Ecorégion ou région écologique : Zone géographique assez large se distinguant par le caractère unique de sa morphologie, de sa géologie, de son climat, de ses sols, de ses ressources en eau, de sa faune et de sa flore. Fokontany : Division administrative territoriale dirigée par un Président du Comité Local de Sécurité (PCLS). Formation végétale : Ensemble de populations végétales de physionomie déterminée. Groupement végétal : Formations végétales appartenant au même type de forêt et au même type de sol. Kijana : Zone de pâturage de zébus. Raiamandreny : Notables, homme âgés de plus de 50 ans. Régénération : Reconstitution d'un état forestier, sans intervention de l'homme, après perturbation. Sao-draza : Evénement culturel de remerciement des ancêtres dans lequel une famille invite la communauté à fêter avec eux leurs réussites ou leurs guérisons. Savane incluse : Savane de taille réduite complètement entourée par la forêt. Savane sur versant : Savane située sur une pente, en aval de la forêt et non entourée par celleci. Station : Espace occupé par une communauté végétale, à l'intérieur duquel les conditions géomorphologiques, mésoclimatiques et biotiques (facteurs stationnels) sont globalement uniformes. Résilience : Aptitude d'un système à survivre à des altérations et à des perturbations dans sa structure et/ou son fonctionnement, et à retrouver, après la disparition de ces dernières, un état comparable à la situation initiale. Tanala ou «Ceux qui vivent dans la forêt», dénomination générique des populations occupant la région forestière située au Sud Est de l'île entre la région côtière et les Hautes Terres occupées par les Betsileo. Tevy : Nom local pour designer l'agriculture itinérante sur brûlis. Végétation climacique : Végétation qui est en équilibre avec son climat. Volambita : Rituel pour la bénédiction et protection des zébus par les ancêtres. ABBRÉVIATIONS ET ACCRONYMES ACP : Analyse en Composantes Principales. ADE-4 : Analyse des Données Ecologique : méthode Exploratoires et Euclidiennes en Sciences de l'Environnement. AFC : Analyse Factorielle de Correspondance. ANOVA : Analysis of variance (ou Analyse de variance). Classe I : Classe des individus à diamètre à hauteur de poitrine inférieur à 5 cm. Classe II : Classe des individus à diamètre à hauteur de poitrine compris entre 5 et 10 cm. Classe III : Classe des individus à diamètre à hauteur de poitrine supérieur à 10 cm. CMP tandavanala : Comité Multi-local de Planification de Fianarantsoa, association dénommée « Tandavanala ». Dhp : Diamètre à hauteur de poitrine. GCF : Gestion Contractualisée des Forêts. G 1 : Forêt à Ficus soroceoides et à Erythroxylum nitidilum G2 : Forêt à Erythroxylum nitidilum et à Amerya humbertii G3 : Forêt à Dalbergia sp. et à Polyscias ornifolia Forêt V : Forêt en contact avec les savanes sur versant. Forêt SI : Forêt en contact avec les savanes incluses. MEM : Modélisation pour l'Environnement à Madagascar. IRD : Institut de Recherche pour le Développement. VOI : Vondron'Olona Ifotony (ou COBA : COmmunauté de BAse). XL STAT : Outil d'analyse de donnée et de statistique pour le Microsoft Excel. SOMMAIRE SOMMAIRE i LISTE DES FIGURES iv LISTE DES CARTES iv LISTE DES TABLEAUX v LISTE DES PHOTOS v INTRODUCTION 1 Première partie- MILIEU D'ETUDE 3 I. LOCALISATION GÉOGRAPHIQUE 3 II. MILIEU PHYSIQUE 3 II.1. Climat 3 II.2. Hydrographie 5 II.3. Relief et pédologie 5 III. MILIEU BIOTIQUE 7 III.1. Flore et végétation 7 III.2. Faune 8 IV. MILIEU HUMAIN 8 Deuxième partie- METHODES 10 I. ETUDES PRELIMINAIRES 10 I.1. Recueil bibliographique 10 I.2. Photo interprétation 10 I.3. Prospection et choix des sites 10 II. ETUDE SUR TERRAIN 11 II.1. METHODE D'APPROCHE 11 II.2. COLLECTE DES DONNEES 13 II.2.1. Caractérisation de la végétation 13 A. Etude structurale 13 B. Etude quantitative de la végétation 15 II.2.2. Caractérisation du sol 16 II.3. ANALYSES DES DONNEES 17 II.3.1. Analyse floristique 17 A. Densité 17 B. Régénération naturelle 17 C. Dendrométrie 18 II.3.2. Analyse numérique 20 A. Test de similitude 20 B. Traitement numérique 21 II.4. CARTOGRAPHIE DE VEGETATION 23 II.5. METHODE D'ENQUETES 23 Troisième partie- RESULTATS ET INTERPRETATIONS 24 I. Caractéristiques floristiques globales 24 II. Caractéristiques édaphiques 24 III. Identification des groupements végétaux 25 III.1. Affinité floristique entre les stations 25 III.2. Analyse factorielle 26 III.3. Relation sol - végétation. 28 III.4. Structures biométriques 30 III.5. Conclusion partielle 32 IV. Description des formations forestières 32 IV.1. Forêt à Ficus soroceoides et à Erythroxylum nitidilum (G1) 32 IV.1.1. Stations et localité 32 IV.1.2. Richesse floristique 32 IV.1.3. Structure horizontale 33 IV.1.4. Structure verticale 33 IV.1.5. Régénération globale 35 IV.2. Forêt à Erythroxylum nitidilum et à Amerya humbertii G2) 35 IV.2.1. Stations et localité 35 IV.2.2. Richesse floristique 36 IV.2.3. Structure horizontale 36 IV.2.4. Structure verticale 36 IV.2.5. Régénération globale 38 IV.3. Forêt à Dalbergia sp. et à Polyscias ornifolia (G 3) 38 IV.3.1. Stations et localité 38 IV.3.2. Richesse floristique 38 IV.3.3. Structure horizontale 39 IV.3.4. Structure verticale 39 IV.3.5. Régénération globale 41 V. Synthèse des résultats sur la typologie de la végétation 41 VI. Principales menaces et pressions 43 VI.1. Mode d'élevage 45 VI.2. Exploitations forestières 48 VI.3. Culture sur brûlis ou tevy 49 VI.4. Feu de pâturage 49 VII. Evolution spatio-temporelle de la végétation 50 VII.1. Evolution des savanes incluses 50 VII.2. Principales tendances évolutives des savanes incluses 54 VII.2.1. Savane régressive 54 VII.2.2. Savane progressive 54 VII.2.3. Savane stable 56 VII.3. Dynamique des groupements forestiers en contact avec les savanes incluses 56 VII.3.1. Forêt progressive 56 VII.3.2. Forêt régressive 57 Quatrième partie- DISCUSSION et RECOMMANDATIONS 59 Discussion 59 Problèmes méthodologiques 59 Remarques sur les résultats 59 Recommandations 63 CONCLUSION GENERALE 64 BIBLIOGRAPHIE 66 ANNEXE LISTE DES FIGURES Figure 1 : Courbe ombrothermique du chef lieu de Sahabe (source World_clim_1950-2000). 5 Figure 2 : Démarche de l'étude 11 Figure 3 : Méthode d'étude de la structure verticale de la végétation (Gautier, 1994) 14 Figure 4 : Dispositif d'un placeau A 16 Figure 5 : Dendrite de similitude 26 Figure 6 : Cartes factorielles dans le plan 1-2 des 20 relevés (a) et des 138 espèces (b) 27 Figure 7 : Carte factorielle des stations 29 Figure 8 : Cercle de corrélation des propriétés physico-chimiques du sol 29 Figure 9 : Carte factorielle des 20 relevés 31 Figure 10 : Cercle de corrélation des variables biométriques 31 Figure 11 : Structure de la forêt G1 34 Figure 12 : Diagramme de recouvrement de la forêt G1 34 Figure 13 : Structure démographique de la forêt G1 35 Figure 14 : Structure de la forêt G2 37 Figure 15 : Diagramme de recouvrement de la forêt G2 37 Figure 16 : Structure démographique de la Forêt G2 38 Figure 17 : Structure de la forêt G3 40 Figure 18 : Diagramme de recouvrement de la forêt G3 40 Figure 19: Structure démographique de la forêt G3 41 Figure 20 : Schéma hypothétique de la dynamique forestière 44 LISTE DES CARTES Carte 1 : Localisation de la zone d'étude (source : FTM BD 100, 2005 ; MEM, 2009) 4 Carte 2 : Réseau hydrographique de la zone d'étude (source : FTM BD 500, 2002) 6 Carte 3 : Localisation des relevés de végétation de 2008 sur fonds de végétation 2007 (source : Atlas de la végétation de Madagascar 2007) 12
Carte 4:Le site d'étude en 1940 (extrait de la carte de la commune d'Ambohimahamasina ; source : photo aérienne 1937) 51 Carte 5 : Le site d'étude en 1957 (source : photo aérienne 1957 ; MEM, 2008) 52 Carte 6 : Evolution des limites de la lisière sur un fonds de carte 2004 (source : carte topographique 1937, photos aériennes 1957 et 1991, image Google-Earth 2004 ; MEM, 2008) 53 Carte 7: Localisation des types de savanes incluses (source : BD 100, 2005 ; MEM, 2008) 55 LISTE DES TABLEAUX Tableau I : Pourcentage de recouvrement 15 Tableau II : Richesse spécifique des stations 24 Tableau III : Matrice de similitude 25 Tableau IV : Caractères floristiques et biométriques des différentes formations 41 Tableau V : Richesse floristique et biovolume des types de forêts 42 LISTE DES PHOTOS Photo 1 : Zébus mâles castrés travaillant la terre 45 Photo 2 : Tolon'omby lors d'un Sao-draza. 46 Photo 3 : Parcage des zébus lors d'un sao-draza 46 Photo 4 : Gardiennage de zébus par les enfants 47 Photo 5 : Tevy en lisière de la forêt. 49 Photo 6 : Plantation de maïs après Tevy. 49 Photo 7 : Savane nouvellement brûlée pour le pâturage (01/2008) 57 Photo 8 : Aménagement de bas fonds en rizière 57 INTRODUCTION INTRODUCTION La forêt tropicale humide couvre 1 753 millions d'hectare soit 64% de la surface totale des forêts dans le monde (Lanley, 1992). Pendant les dernières décennies du 20ème siècle, le taux de disparition moyen annuel de forêt naturelle est estimé à 0.8% pour l'ensemble africain (FAO, 2001). Les causes directes et les plus fréquentes de la déforestation sont les activités d'aménagement et d'exploitation : feux et coupes servant aux activités productives paysannes, aux activités minières et industrielles. La déforestation conduit souvent à l'apparition de savanes. A Madagascar, les savanes couvrent 70% de l'ensemble du pays (Rakotoarimanana & Grouzis, 2008). Ces savanes sont localisées surtout sur les Hautes Terres, le Moyen Ouest et l'Ouest, caractérisées par des climats variant du type semi-humide au type sec. Néanmoins, des savanes de faibles étendues sont localisées au coeur des massifs forestiers, appelées « savanes incluses ». Ce sont ces savanes incluses qui vont faire l'objet de cette étude dans le corridor forestier de Fianarantsoa. La formation forestière de cette zone est une forêt naturelle de moyenne altitude dont le couvert boisé, très hétérogène, est constitué de mosaïque de fourrés, de forêts fragmentées linéairement par crêtes et thalwegs, de forêts continues (Serpantié et al., 2007). Ces savanes incluses se développent dans un milieu où le climat et le sol sont favorables au développement de la forêt. La présence de ces savanes incluses reste énigmatique à double titre. D'une part, ces savanes incluses se trouvent entièrement dans un milieu à climat humide, où la végétation climacique est naturellement la forêt dense humide, et d'autre part, le substrat constitué de sols de type ferrallitique, reste aussi favorable au développement de la forêt. La présence de ces savanes incluses constitue donc une situation idéale pour étudier la tendance évolutive de certains massifs forestiers. La tendance peut être progressive ou régressive. L'existence et l'évolution de ces savanes incluses rentrent dans la problématique abordée par le programme MEM. En effet, le programme MEM se propose d'étudier les dynamiques de l'occupation du sol, post-forestière à la suite de l'abattis-brûlis, de la régénération forestière et de la savanisation. La présente étude réalisée dans le cadre du programme va porter sur l'évaluation et la caractérisation de la dynamique des savanes incluses. La dynamique spatio-temporelle de la forêt, qui se traduit par des phénomènes de savanisation ou de régénération forestière, dépend-elle de l'historique de l'utilisation de chaque unité de formation végétale ? Telle est la question principale à laquelle notre travail se Introduction propose de répondre. Pour apporter des éléments de réponse à cette problématique, trois hypothèses ont été émises : - L'histoire de l'utilisation de chaque unité de formation végétale passe par des périodes de fortes et de faibles pressions exercées par l'homme. - La forêt présente une forte résilience, ainsi pendant la période de très forte pression, elle régresse graduellement, par contre dès que la pression est levée, le phénomène inverse de régénération forestière s'installe. - La résilience de la forêt est relativement faible dans les zones à sols squelettiques et dans certaines situations topographiques ou certaines expositions des pentes. Ainsi, notre étude se propose de vérifier les hypothèses énoncées précédemment à travers des relevés écologiques, combinée à une cartographie de la végétation et à une enquête auprès des paysans, se rapportant aux quatre principaux objectifs suivants : - Caractériser les différents types de groupements forestiers autour des savanes incluses. - Déterminer les différents types de contact forêts-savanes et cartographier l'évolution spatio-temporelle des limites des contacts forêts-savanes. - Décrire la dynamique des groupements forestiers en contact avec les savanes incluses. - Définir les différentes pressions exercées au niveau des forêts autour des savanes incluses. Pour répondre à ces attentes, le présent travail est subdivisé en quatre parties principales. Dans un premier temps, nous allons présenter le milieu d'étude, ensuite nous allons décrire les méthodes adoptées. Dans un troisième temps nous présenterons les résultats et les interprétations afférentes. Dans la dernière partie du travail, nous avançons une discussion qui sera suivie de quelques recommandations, puis d'une conclusion. PREMIERE PARTIE MILIEU D'ETUDE MILIEU D'ETUDE I. Localisation géographique Le corridor forestier de Fianarantsoa dénommé corridor « Fandriana-Vondrozo » se trouve à cheval entre le domaine de l'Est, et celui du centre qui longe toute la partie Est de Madagascar de façon plus ou moins continue (Humbert & Cours Darnes, 1965). Il est délimité au nord par le district de Fandriana (Région d' Amoron'i Mania) et au Sud par le district de Befotaka (Région Atsimo Atsinanana). Le corridor couvre 66 communes rurales qui se répartissent du Nord au Sud dans l'ex-province de Fianarantsoa. La zone d'étude se trouve dans la commune rurale d'Ambohimahamasina, localisée à 44 km à l'Est d'Ambalavao, soit à 101 km au Sud de Fianarantsoa. La commune rurale d'Ambohimahamasina renferme treize (13) fokontany, dont quatre (4) sont localisés à l'intérieur du corridor forestier. Ces quatre fokontany relient le pays betsileo au pays tanala par l'existence des sentiers traversant le corridor de RanomafanaAndringitra d'Ouest en Est. Ces sentiers relient des chapelets de savanes incluses. La zone d'étude (carte 1) est localisée sur le sentier reliant le fokontany d'Andohanimananatanana, chef lieu Sahabe (partie betsileo) et le fokontany de Tsianivoha (partie tanala) ; c'est le sentier le plus emprunté du corridor forestier. II. Milieu physique II.1. Climat La commune d'Ambohimahamasina est soumise à l'influence d'un climat de type tropical d'altitude, avec une alternance de deux saisons bien distinctes d'après la courbe ombrothermique (figure 1), 6 mois de saison pluvieuse (Octobre - Mars) suivis de 6 mois de saison sèche (Avril - Septembre). Pendant la saison chaude et humide qui va du mois d'Octobre à Avril, la quasitotalité de la pluviométrie environne les 90% des précipitations (1000 à 1200 mm/an) et provoque des inondations fréquentes entre le mois de novembre et janvier. Et en saison froide et humide, allant du mois de mai au mois de septembre, la précipitation arrive plutôt sous forme de crachins. Quant à la température, elle est comprise entre 15°C et 22°C et atteint son maximum en saison chaude et humide (30°C), et peut descendre jusqu'à 5°C en saison froide et humide [Web 01, 2008]. 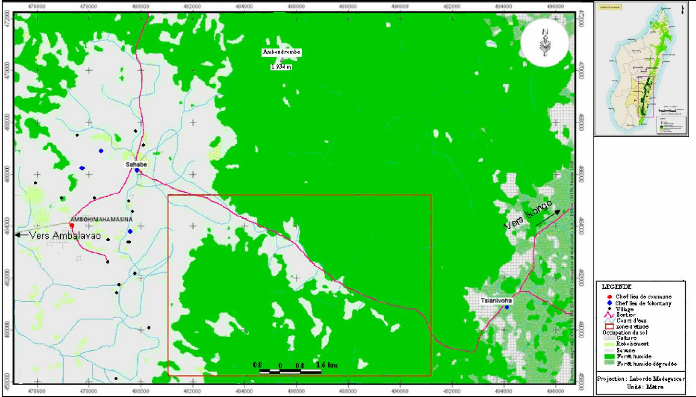
Carte 1 : Localisation de la zone d'étude (source : FTM BD 100, 2005 ; MEM, 2009) 
Temperature moyenne annuelle (C) 160 140 120 100 40 20 80 60 0 J A S O N D J F M A M J Température moyenne annuelle (°C) Mois Précipitation (mm) 350 300 250 200 50 0 150 100 Precipitation (mm) Figure 1 : Courbe ombrothermique du chef lieu de Sahabe (source World_clim_1950-2000) II.2. Hydrographie Le corridor forestier constitue un immense réservoir d'eau qui règle le régime hydrique et du climat de la région de la Haute Matsiatra. Le réseau hydrographique du site d'étude est très dense (carte 2) et plusieurs rivières prennent leur source dans la montagne d'Ambondrombe. Trois rivières, Sandragnato, Sahandranto, Matitagna, se déversent vers l'Est, tandis que la Mananatana se déverse vers l'Ouest (Rajaonarivelo, 2001). Cette rivière arrose des vallées plus ou moins étendues et aménagées constituant le bassin d'Ambalavao. II.3. Relief et pédologie Selon Boulanger & Ratoanina (1957), Chantraine (1967), Besairie (1972, 1973), le substrat géologique de la région des Betsileo appartient entièrement à un socle cristallin, plus précisément à un ensemble complexe de roches sédimentaires métamorphisées, daté de trois milliards d'années, au début du précambrien (annexe 1). 
Carte 2 : Réseau hydrographique de la zone d'étude (source : FTM BD 500, 2002) Dans l'ensemble, le relief est accidenté avec des suites de collines au sommet arrondi et disséqué ainsi que des vallées plus ou moins étroites. Mais, des reliefs montagneux dont le massif sacré d'Ambondrombe culminant dans la région à une altitude de 1 936m et le sommet de Tsitondroina (1 817m), marquent aussi le paysage. A l'instar des sols des hautes terres centrales de Madagascar, les sols de la commune d'Ambohimahamasina sont de type ferralitique rouges, acides, et pauvres en humus (Rajaonarivelo, 2001). Les apports en alluvions assurent la fertilité des sols des vallées et des plaines. Dans les bas fonds, les sols sont hydromorphes. III. Milieu biotique Le corridor forestier de l'Est de Madagascar est une forêt naturelle de moyenne altitude dont le couvert boisé est très hétérogène à l'Ouest (Serpantié et al., 2007). Ce couloir forestier est formé d'une mosaïque de forêts primaires riches en espèces dont la plupart sont endémiques. Des forêts secondaires issues de défriches ainsi que des forêts artificielles issues de reboisement se rencontrent également dans ce corridor forestier. III.1. Flore et végétation Phytogéographiquement, le corridor forestier de Fianarantsoa se trouve à cheval entre le domaine de l'Est et celui du Centre. La formation du corridor forestier est une forêt dense ombrophile de moyenne altitude appartenant à la série de Weinmannia (CUNONIACEAE) et Tambourissa (MONIMIACEAE) (Humbert & Cours Darnes, 1965). Dans la commune d'Ambohimahamasina, différents types de formation végétale s'observent. Les savanes herbeuses qui sont dominées par Aristida similis en mosaïque avec les reboisements d'Eucalyptus prédominent la partie occidentale. A l'Est de cette grande savane, se trouve la forêt dense humide d'une superficie de 17 500ha (en 2001) (Rajaonarivelo, 2001) conservée à l'état primaire (végétation mature) ou secondaire formant un long corridor forestier. La flore est représentée essentiellement par les familles de CUNONIACEAE (Weinmannia sp), LAURACEAE, MONIMIACEAE, MYRTACEAE, RUBIACEAE et STERCULIACEAE (Schatz & Malcomber, 1993). III.2. Faune La forêt malgache est riche faunistiquement. Cette richesse se rencontre surtout dans les forêts humides de l'Est. Le corridor forestier Est malgache renferme des espèces rares et menacées d'oiseaux comme Philepitta souimanga, Mesitornis unicolor (OPISTHOCOMIDAE), Brachypteracias squamiger (BRACHYPTERACIDAE), Neodrepanis hypoxanthus (PHILEPITTIDAE) et Xenopirostris polleni (VANGIDAE) (Langrand, 1995). Le corridor forestier est connu surtout par ses 3 espèces de primates, Hapalemur aureus, Prolemur simus et Eulemur albocollaris, des Amphibiens tel que Mantella bernhardii, et des reptiles (Matoatoa spanrinngi), pouvant constituer l'emblème de ce corridor forestier [Web 01, 2008]. Selon la monographie de la commune rurale d'Ambohimahamasina (PCD de la commune rurale d'Ambohimahamasina, 2006), parmi les animaux communs avec le parc de Ranomafana, on peut noter dans la zone d'étude l'existence de Propithecus diadema edwardsi, Varecia variegata editorum, Eulemur rubriventer, le carnivore Cryptoprocta ferox, les reptiles tels que Furcifer campani, Furcifer minor et les amphibiens Mantella cowanii, Mantella madagascariensis. IV. Milieu humain La commune rurale d'Ambohimahamasina est majoritairement peuplée de Betsileo mais aussi de Tanala et une minorité d'Antaisaka, d'Antandroy et de Bara. La plupart du temps, les habitants ont un double résidence, un domicile dans un village et un campement dans la forêt. Ainsi, ils passent les saisons culturales en forêt pour s'assurer une conquête foncière, et la période entre les cultures au village. La commune rurale d'Ambohimahamasina, avec une population de 20 579 habitants, présente une densité de 39,3 habitants au km2 [Web 02, 2004]. La commune est la plus peuplée des communes du corridor forestier. En effet, la taille des ménages est d'environ 7 personnes mais seulement deux enfants en moyenne sont scolarisés par famille (Andriamahazo et al., 2004). Les principales activités de la population sont la riziculture et l'élevage de zébus. En effet, selon Moreau (2002), « les Betsileo sont à la fois des pasteurs et des cultivateurs (...) un Betsileo sans rizière n'est rien : on ne sait d'où il vient, ni qui sont ses ancêtres. Tout comme un Betsileo sans boeufs d'ailleurs, qui ne peut participer aux échanges sociaux ». La riziculture constitue l'activité principale de la population. La riziculture irriguée sur les bas fonds et sur les gradins aménagés depuis la base des collines et à l'intérieur même de la forêt est la plus pratiquée. D'autres cultures comme la culture de manioc, de brèdes, de maïs se pratiquent sur les tanety. La production, et particulièrement celle des rizières étant insuffisante pour l'ensemble de la population, il s'avère de plus en plus nécessaire de cultiver d'autres produits sur d'autres terrains. La période de soudure dure 3 - 6 mois. Elle commence à la fin du mois de juin (26 juin) pour les plus pauvres et le mois d'octobre pour tous (Moreau, 2002). Pour remédier à ces manques de production, les paysans choisissent d'agrandir leur parcelle cultivée en pratiquant des tevy. L'élevage de bovidés occupe une place importante dans l'économie et dans la vie socioculturelle locale. Le troupeau joue un rôle non seulement économique (capital, monnaie d'échange dans de nombreuses transactions) mais aussi social et culturel (un grand troupeau est source de prestige). DEUXIEME PARTIE METHODES METHODES I. ETUDES PRELIMINAIRES I.1. Recueil bibliographique Pour bien cerner le sujet de l'étude et mieux préparer les missions pour les relevés de données sur le terrain, des documents relatifs au thème et au milieu d'étude ont été réunis et analysés. Cette étape a permis d'obtenir les informations nécessaires sur la zone d'étude et pour la réalisation de ce présent mémoire. I.2. Photo interprétation Cette étape est indispensable pour avoir une vue d'ensemble sur la zone d'étude. Une photo-interprétation a été réalisée sur une photo aérienne de 1991 à l'échelle de 1/50 000. La photo-interprétation a été utilisée pour repérer les zones forestières et les clairières, et caractériser les unités à travers la distinction de la teinte, la structure et la texture de l'objet. La texture est définie par la forme des points élémentaires de l'objet nu. On distinguera les textures visibles à l'oeil nu des textures non visibles à une distance normale de lecture. La structure représente le mode de répartition et l'orientation de ces éléments. La teinte est la couleur des différents éléments de la photo aérienne. Pour distinguer et caractériser les unités sur les photos, des lunettes stéréoscopiques ont été utilisées pour permettre une observation en 3 dimensions. La démarche exige de placer avec une zone de recouvrement un couple de photos aériennes représentant la même zone pour avoir une interprétation en 3D du milieu. I.3. Prospection et choix des sites Les unités définies par la photo interprétation, rapportées sur une précarte, sont vérifiées sur le terrain. Une mission de vérité terrain vise surtout à localiser les sites potentiels pour les problématiques abordées dans cette étude. Le choix des emplacements des sites de relevés a été fait en tenant compte de la géomorphologie et de l'homogénéité physionomique, floristique et des substrats. A l'issue de cette mission, 2 sites ont été choisis : forêts en contact avec les savanes et forêts de référence pour déterminer le niveau de dégradation des forêts en contact par comparaison avec des forêts supposées non dégradées. Dans les forêts en contact, deux stations sont distinguées, selon les savanes : forêts en contact avec des savanes continues de versants et forêts en contact avec des savanes incluses. On suppose en effet que les premières sont plus dégradées que les secondes. Deux stations de forêts de référence selon la perception paysanne ont été distinguées : Alamainty et Alamena. Les Alamainty sont des forêts dites « naturelle » caractérisées par des arbres de gros diamètres et hautes et les Alamena sont des forêts à végétation plus dense et moins haute. Ainsi quatre stations sont retenues et cinq relevés écologiques par station sont effectués, donc 20 relevés au total (carte 3). II. ETUDE SUR TERRAIN II.1. METHODE D'APPROCHE Selon le type de sol et le degré de l'intensité des différents types de perturbations, il peut y avoir une reconstitution forestière ou un processus de savanisation. Ainsi pour comprendre cette dynamique, la démarche adoptée se subdivise en trois parties bien distinctes (figure 2). 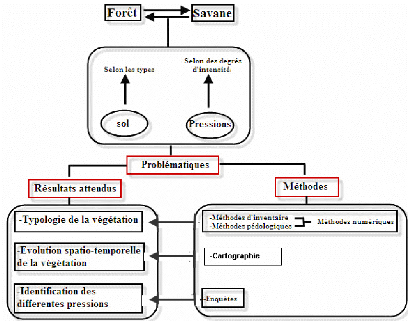
Figure 2 : Démarche de l'étude | | Méthodes | | | Carte 3 : Localisation des relevés de végétation de 2008 sur fonds de végétation 2007 (source : Atlas de la végétation de Madagascar 2007) | | | | | | | 12 | | Relevés des forêts en contact avec les savanes sur versant Relevés des forêts en contact avec les savanes incluses Relevés des forêts Alamainty Relevés des forêts Alamena | |
La première partie consiste à déterminer la typologie des différents groupements végétaux. Celle-ci consiste à utiliser des méthodes d'inventaires botaniques et pédologiques. Les résultats obtenus ont fait l'objet d'un traitement statistique. La deuxième partie traite l'évolution spatio-temporelle de la végétation. Une étude cartographique a été ainsi effectuée. Cette étude nécessite l'utilisation de la télédétection et du SIG. Des photos multi-dates ont été utilisées pour suivre l'évolution des savanes incluses et de la forêt. La troisième partie de l'étude cherche à déterminer les différentes pressions qui peuvent influencer la dynamique de la végétation. Des enquêtes ont été ainsi effectuées auprès des villageois. Cette partie traite la relation qui peut exister entre l'évolution des écosystèmes et les activités de la population. II.2. COLLECTE DES DONNEES Sur le terrain, des méthodes de transect et de placeaux ont été appliquées sur les différentes stations forestières. Ceci pour déterminer la répartition des différents individus de chaque espèce le long des pentes dans la formation. Des fosses pédologiques ont été aussi creusées pour caractériser et déterminer la nature du sol où la formation forestière se développe. II.2.1. Caractérisation de la végétation A. Etude structurale La structure de la végétation est définie comme la répartition et l'agencement des plantes les unes par rapport aux autres constituant une formation végétale (Guinochet, 1973). Elle permet d'une part, d'avoir une image réelle ou la représentation de l'ensemble de la végétation et d'autre part, de déterminer l'organisation spatiale des espèces. La structure de la végétation peut être caractérisée selon deux manières : ~ Structure horizontale La structure horizontale exprime l'agencement et la répartition des individus suivant le plan horizontal ; cette répartition se traduit par la densité des espèces dans le site d'étude, et par extension, la valeur de la surface terrière et du biovolume des individus présents. La méthode utilisée consiste à recenser les différents individus des espèces et de mesurer la hauteur maximale, la hauteur du fût, le diamètre à hauteur de poitrine (dhp). Ces recensements ont été faits dans un placeau de 900 m2. ~ Structure verticale Les strates se définissent comme étant le niveau de concentration maximale de la masse foliaire (Gounot, 1969). L'étude de la structure verticale a été effectuée suivant la méthode de Gautier (1994). Elle permet de connaître les différentes strates et de déterminer le degré de dégradation de la formation végétale à partir de la continuité de la voûte forestière. Cette méthode est basée sur l'établissement d'un diagramme de recouvrement qui correspond au mode de stratification des espèces dans la formation végétale. Le protocole de relevé consiste à tirer une corde de 50m le long de la ligne parallèle à la plus grande pente. Tous les 1 m, la hauteur du contact entre la partie vivante de la plante et le point de lecture le long de la ligne a été mesurées à l'aide d'une gaule de 7m ; au-delà de 7m, les points de contact ont été estimé visuellement (figure 3). Les données recueillies ont été ensuite transférées sur un tableur EXCEL pour visualiser le profil structural de la végétation. Le recouvrement est tiré de ce profil structural (sous forme histogramme) qui correspond au pourcentage de nombre de contacts par rapport au nombre total des points de contact. 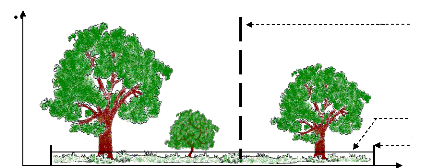
Gaule Corde Piquet Hauteur (m) Distance (m) Figure 3 : Méthode d'étude de la structure verticale de la végétation (Gautier, 1994) L'échelle de recouvrement, établie par Godron et al. (1983) a été prise comme référence ; elle correspond à l'ouverture de la strate. Plus le degré de la perturbation est élevé, plus la strate est ouverte et inversement (Tableau I). Tableau I : Pourcentage de recouvrement | Recouvrement global (%) | Type de strate | | > 90 | Strate fermée | | 75 | - 90 | Strate peu ouverte | | 50 | - 75 | Strate semi-ouverte | | 25 | - 50 | Strate ouverte | | 10 | - 25 | Strate très ouverte |
B. Etude quantitative de la végétation La méthode de placeau a été utilisée. Cette méthode permet de faire une étude quantitative de la végétation sur une surface homogène et de déterminer le type de formation végétale existant dans la zone d'étude. Sur le terrain, vingt (20) relevés ont été effectués soit cinq (5) répétitions chacun pour les quatre (4) types de forêts (les forêts en contact avec les savanes incluses, les forêts en contact avec les savanes sur versant, les forêts de référence « Alamainty » et les forêts de référence « Alamena »). Les Alamainty et les Alamena sont les deux (2) types de forêts qui existent dans la zone d'étude selon la perception paysanne. Le dispositif de relevé (figure 4) est monté de façon à ce qu'il soit parallèle à la grande pente. Le placeau a été matérialisé par des cordes et des piquets. Le placeau a été subdivisé en 3 compartiments pour faciliter le recensement des individus. Le compartiment A correspond à l'ensemble de tout le placeau soit 30m x 30m. Le compartiment B correspond au quart de A soit 15m x 15m. Le compartiment C correspond au quart de B soit 7,5m x 7,5m. Le recensement des espèces a été effectué de la manière suivante : - Toutes les espèces ayant un diamètre à hauteur de poitrine (dhp) supérieur à 10 cm ont été recensées dans tout le placeau (compartiment A). - Toutes les espèces ayant un diamètre à hauteur de poitrine (dhp) compris entre 5 et 10 cm ont été recensées dans deux (2) compartiments B. - Toutes les espèces ayant un diamètre à hauteur de poitrine (dhp) inférieur à 5 cm ont été recensées dans cinq (5) compartiments C. Les paramètres mesurés sont les suivants : l'abondance, la hauteur maximale, la hauteur du fût, le diamètre à hauteur de poitrine, le diamètre du houppier. Ils permettent de calculer les caractéristiques écologiques de chaque groupement végétal : la densité, la régénération naturelle, la surface terrière, le biovolume. 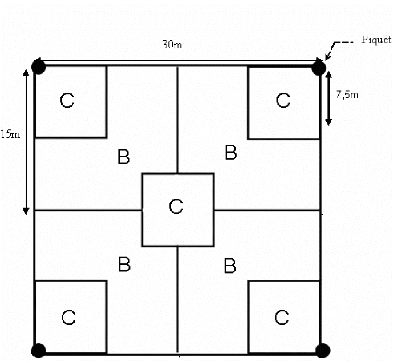
Figure 4 : Dispositif d'un placeau A II.2.2. Caractérisation du sol Les inventaires pédologiques ont été effectués en réalisant une fosse pédologique de 50x50x100cm. La fosse a été réalisée dans la zone forestière la plus représentative du milieu, en utilisant les trois critères d'homogénéités : uniformité du substrat, homogénéité physionomique de la végétation et homogénéité floristique. Sur l'ensemble de la zone d'étude, des fosses ont été creusées dans les forêts en contact avec les savanes incluses, dans les forêts en contact avec les savanes sur versant, dans les forêts Alamainty et dans les forêts Alamena. Pour chaque fosse pédologique, un sol d'environ 250g a été prélevé sur une profondeur de 0 à 20 cm. Les échantillons sont ensuite analysés au laboratoire de Radioisotope d'Antananarivo (LRI). Ceux-ci afin de caractériser la texture, le taux des éléments minéraux et organiques ainsi que l'acidité du sol. II.3. ANALYSES DES DONNEES Les paramètres floristiques et les données issues des analyses du sol en laboratoire ont fait l'objet d'analyses et de traitement. II.3.1. Analyse floristique A. Densité La densité du peuplement est le nombre total de tous les individus par rapport à la surface (Dajoz, 1975). Nb ind 
d S Avec Nb ind: nombre de tous les individus S : surface de relevés B. Régénération naturelle La régénération naturelle désigne la faculté d'un écosystème à se reconstituer spontanément après destruction totale ou partielle du couvert forestier. Elle se manifeste par un ensemble de processus naturels par lesquels les espèces se reproduisent au sein d'une formation végétale (Rollet, 1979). L'étude de la régénération permet de déterminer le potentiel de régénération de la formation à travers l'évaluation du taux de régénération. La méthodologie consiste à recenser d'une part tous les individus ayant un diamètre à hauteur de poitrine (dph) supérieur à 10cm considérés comme des semenciers, et d'autres part, des individus de dhp inférieur à 10cm, interprétés comme des individus régénérés. Au delà de l'évaluation du taux de régénération, cette démarche permet également d'estimer la santé du peuplement. Ces deux caractéristiques du peuplement sont obtenues à partir des critères suivants : - La santé du peuplement peut être estimée à partir d'un graphe sous forme d'histogramme représentant les effectifs des classes de diamètre. Les classes suivantes ont été adoptées : Classe I : individus présentant un dhp inférieur à 5 cm Classe II : individus présentant un dhp compris entre 5 et 10 cm Classe III : individus présentant un dhp supérieur à 10 cm Ainsi la tendance de la santé des peuplements est définie de la manière suivante : · la courbe est en forme de J inversé, le potentiel de régénération est bon ; · elle est en forme de J, le potentiel de régénération est mauvais ; · Il y a perturbation de la régénération s'il manque une ou plusieurs classes de diamètres. - Le taux de régénération est exprimé en termes de pourcentage des individus régénérés (Nr) par rapport au nombre des individus semenciers (Ns) (Rothe, 1964). N TR = × r 100 Ns Où : TR : taux de régénération Nr : nombre des individus régénérés Ns : nombre des individus semenciers Suivant la valeur du taux de régénération, 3 cas sont possibles (Rothe, 1964) : Si TR<100% : mauvaise régénération. Si 100% <TR<1000% : bonne ou moyenne régénération. Si 1000% <TR : très bonne régénération. C. Dendrométrie · Surface terrière La surface terrière G est le recouvrement basal représenté par la surface occupée par les parties aériennes des individus d'une espèce donnée au niveau du sol, ou à hauteur de poitrine dans le cas des arbres (Parde, 1961 ; Gounot, 1969). On note Gi, la surface terrière d'un arbre et G la surface terrière d'un peuplement. La surface terrière d'un arbre (Gi) est estimée par la formule de Dawkins (1959) : ð G i d i 2 = 4 Où Gi : surface terrière en m2/ha de chaque individu i di : diamètre à hauteur de poitrine (dhp) de chaque individu i (m) La surface terrière d'un peuplement est la somme des surfaces terrières de tous les arbres qui le composent, elle dépend à la fois de la grosseur et du nombre d'arbres. Le suivi dans le temps de cette mesure donne un indice sur la productivité ligneuse d'une parcelle, via la vitesse de croissance en diamètre des arbres. La surface terrière d'un peuplement (G) est donnée par la formule : n G = ? Gi 1 Pour être plus réaliste, cet indice doit être croisé avec la hauteur des arbres. Combiné avec la hauteur, l'indice est appelé « biovolume ». ~ Biovolume Le biovolume se définit comme étant le volume de bois fourni par la végétation dans une surface donnée. Il permet d'estimer le potentiel en bois de la formation végétale. Le potentiel en bois est calculé selon la formule de Dawkins (1959) : V = 0.53 ? G i h i Avec V : biovolume de tous les individus (m3/ha) Gi : surface terrière de chaque individu i (m2/ha) hi : hauteur du fût de chaque individu (m) 0.53 : coefficient de forme Selon Roger et Rabarison (2000), le biovolume est élevé quand il est supérieur à 250m3/ha, moyen quand il est compris entre 50 et 250m3/ha, et faible, quand il est inférieur à 50m3/ha. II.3.2. Analyse numérique Les traitements numériques effectués sont de deux types, le test de similitude afin de distinguer l'affinité floristique des différentes stations et les analyses factorielles telles que l'AFC, l'ACP ainsi que l'ANOVA. Pour le deuxième type de traitement, les logiciels utilisés sont : ADE-4 et XLSTAT 7.0. L'AFC et l'ACP ont été utilisées pour déterminer la typologie et caractériser les groupements végétaux. A. Test de similitude Ce test permet de caractériser le degré de ressemblance entre les différents relevés étudiés en comparant deux à deux leurs listes floristiques. Il sert à estimer la similarité entre deux communautés végétales au moyen des pourcentages d'espèces qu'elles ont en commun. Il est déterminé à l'aide du coefficient de communauté de Sorensen Ps (Sorensen, 1948) appelé également quotient de similarité ou coefficient de similitude : Avec : · Ps : coefficient de similitude de SORENSEN (%) · a : nombre d'espèces inventoriées dans le relevé A · b : nombre d'espèces inventoriées dans le relevé B · c : nombre d'espèces communes aux deux relevés A et B. La valeur du coefficient de similitude peut varier de 0 à 100%. Elle est nulle quand il n'y a pas d'espèce commune entre les 2 communautés, alors qu'elle est de 100% quand toutes les espèces sont rencontrées à la fois dans les 2 communautés.  B. Traitement numérique a. Analyse de Variance (ANOVA) L'analyse de variance permet de tester les effets de traitement c'est-à-dire de rechercher si l'effet « traitement » est significatif ou non (Gouet & Philippeau, 1989) au seuil de probabilité 5%. En analyse de variance, un même groupement homogène est repéré par une même lettre. Cette analyse permet de classifier et de déterminer le degré de perturbation de chaque type de forêt. Avec le logiciel XLSTAT-7.0, les résultats sont obtenus avec un intervalle de confiance de 95%, c'est-à-dire que les risques d'erreur (ou probabilité p) sont de 0.05. Ainsi, · La différence est non significative (NS) si p = 0.05 ; · La différence est significative (S) si 0.01<p<0.05 ; · La différence est hautement significative (HS) si p<0.01. La richesse floristique et le biovolume ont été soumis à une analyse de variance (ANOVA) avec le test de Tukey (HSD). b. Analyse multi variée Cette méthode d'analyse permet de représenter graphiquement dans un plan les résultats obtenus et ce, soit à partir de caractères quantitatifs (variable-individus), c'est le cas de l'analyse en composantes principales (ACP), soit à partir de caractères qualitatifs (présence-absence), c'est le cas de l'analyse factorielle de correspondance (AFC). Les variables utilisées pour cette méthode sont les variables floristiques (richesse floristique, biovolume, densité) et les variables pédologiques (structure physique et chimique du sol). Pour cette analyse factorielle, nous avons utilisé le logiciel ADE-4 (Analyse des Données Ecologiques : Méthodes Exploratoires et Euclidiennes en Sciences de l'Environnement), mis au point par l'unité Biométrique de l'Université de Lyon. · Analyse factorielle de correspondance (AFC) L'Analyse Factorielle des Correspondances ou AFC est une méthode d'analyse des données lorsque les variables à étudier sont de nature qualitative ou qualitative et quantitative à la fois (Benzécri, 1981). L'AFC est basée sur la représentation sur un même plan de l'ensemble des individus et des variables. La richesse floristique de chaque relevé a été soumise à ce traitement. Ainsi, la méthode consiste : · à rassembler toutes les données recueillies à traiter sur le terrain : liste des espèces de tous les relevés ; · à établir un tableau de contingence sur le fichier EXCEL représentant les différents relevés en colonne et les espèces en lignes ; · à adopter une codification : « 1 » pour les espèces présentes et « 0 » pour les espèces absentes. Les résultats sont sous formes de graphes montrant l'ensemble des points regroupés ayant les mêmes caractéristiques appelées « nuage de forme interprétable ». L'interprétation de ces graphes est basée sur : · l'identification et signification des axes ; · la proximité des points avec les axes ; · la forme des nuages. · Analyse en composantes principales (ACP) L'Analyse en composantes principales ou ACP est une méthode d'analyse des données multidimensionnelles. Elle est utilisée si les informations obtenues sont de nature quantitative. En effet, c'est une méthode d'analyse des relations entre les caractères des peuplements végétaux et les paramètres quantitatifs (mesurables) qui influencent ces peuplements. C'est essentiellement une méthode de description et d'exploration pouvant révéler des regroupements. En d'autres termes, elle permet d'identifier les caractéristiques des groupements végétaux. La méthodologie consiste à construire un tableau de contingence sur un tableur EXCEL représentant les variables (paramètres écologiques) et les observations (relevés écologiques). Ensuite, le tableau est transféré dans le logiciel en suivant des procédures préétablies. A la fin, les graphes ainsi obtenus après traitement se présentent sous deux formes : les variables seront représentées sur un graphe appelé « cercle de corrélation » et les observations, sur une représentation plane, appelée « plan factoriel ». Les résultats ainsi obtenus sont donc représentés sur deux profils différents. Les proximités entre les variables s'interprètent en termes de corrélations ; les proximités entre les individus s'interprètent en termes de similitudes globales de valeurs observées. II.4. CARTOGRAPHIE DE VEGETATION Pour l'étude de la dynamique des savanes incluses, une carte IGN 1940 élaborée à partir d'une photo aérienne de 1937, deux photos aériennes de 1957 et de 1991, ainsi qu'une image de Google-Earth 2004, analysée en 2008, ont été utilisées. Le SIG et la télédétection ont permis d'estimer la surface des savanes incluses et leur évolution dans le temps. La qualité médiocre des photos aériennes a été contournée en augmentant les contrastes des couleurs sous le logiciel Adobe photoshop 7.0. Les photos aériennes améliorées ainsi que les images satellitales Google Earth ont été géoréférencées sur la base de données FTM (IEFN 1994 et 2000, et BD100 et BD 500, 2005) en utilisant Arcview 3.3 et son extension Image Analysis 1.1a. La méthode consistait à faire une catégorisation c'est-à-dire à identifier toutes les données sur les images. Pour cela, les images ont été catégorisées en mode binaire : forêt et non-forêt ou bois et non-bois. Une fois, la catégorisation faite, les savanes incluses prennent la forme de polygones fermés dont les surfaces sont mesurables. II.5. METHODE D'ENQUETES Pour évaluer l'impact anthropique sur l'écosystème forestier, des enquêtes ont été menées auprès des villageois, propriétaires de zébus, dans les villages ou hameaux qui se trouvent à proximité de la zone d'étude ou des villages qui ont une relation avec les formations forestières. L'enquête a été effectuée dans le Fokontany d'Andohanimananatanana. Elle portait sur les modes d'élevages et de culture dans la région, avant 1950 jusqu'à maintenant. Une fiche d'enquête a été préalablement établie (Annexe 2). L'enquête comprend quatre parties : la première partie concerne les ménages, la deuxième partie porte sur l'agriculture, la troisième partie est relative à l'élevage de zébus et la quatrième partie concerne les savanes incluses. TROISIEME PARTIE RESULTATS et INTERPRETATIONS RESULTATS ET INTERPRETATIONS Les principaux points de cette partie sont la caractérisation des formations par les caractéristiques floristiques et édaphiques, l'identification des différents groupements végétaux, et la description de ces groupements. L'évolution spatio-temporelle de la végétation ainsi que les principales menaces et pressions seront aussi abordées dans cette partie. I. Caractéristiques floristiques globales La formation dans la région est une forêt dense humide sempervirente. Sur l'ensemble des 20 relevés effectués dans les 4 stations considérées (Alamainty, Alamena, forêt en contact avec les savanes incluses, forêt en contact avec les savanes sur versant), 138 espèces ont été recensées, qui se répartissent dans 90 genres et 58 familles. Les familles les mieux représentées sont RUBIACEAE avec 12 espèces, CLUSIACEAE avec 10 espèces, EUPHORBIACEAE avec 10 espèces et ARALIACEAE avec 7 espèces. Les forêts en contact avec les savanes sur versant présentent le plus d'espèces (109) suivi par les Alamainty (94), les forêts en contact avec les savanes incluses (90). Les Alamena présentent 87 espèces (tableau II). Tableau II : Richesse spécifique des stations 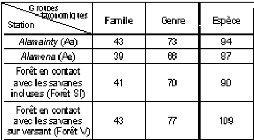
II. Caractéristiques édaphiques Les variables utilisées pour déterminer les caractéristiques du sol de chaque groupement sont la granulométrie (argile, limon, sable), le potassium échangeable, le phosphore assimilable, le rapport C/N (plus C/N est élevé, plus le taux de la matière organique dans le sol est élevé), l'aluminium échangeable, l'acidité du sol (pH). Les résultats des analyses sont présentés dans l'annexe 3. Il apparaît que la texture du sol varie suivant les stations. Il y a peu de différence entre la forêt en contact avec les savanes sur versant et la forêt en contact avec les savanes incluses. Les Alamena ont le taux de sable le plus élevé (91%). Les Alamainty ont des teneurs plus élevées en limon et plus faibles en sable (59% en moyenne). Pour les autres stations, la teneur en sable est environ 70%. Ainsi, les Alamainty, les forêts en contact avec les savanes sur versant et les forêts en contact avec les savanes incluses sont plus fertiles que les Alamena. En effet, les Alamena ont le plus faible teneur en C/N (C/N : 2) donc moins fertile et les Alamainty ont la valeur la plus élevée (C/N : 20). Les forêts en contact ont un teneur en C/N variant entre 13,6 et 17,3. III. Identification des groupements végétaux L'identification des différents groupements a été réalisée à travers des analyses numériques : le test de similitude, l'Analyse Factorielle de Correspondance (AFC) et l'Analyse en Composantes Principales (ACP). Ces analyses numériques ont été effectuées pour discriminer les types de forêt identifiés selon la perception paysanne (Alamainty, Alamena) et les types de forêts en contact. Celle-ci a été basée sur les relevés des différentes stations. Dans les Alamainty, cinq (5) relevés ont été effectués : R1, R2, R3, R4, R5 ; dans les Alamena, cinq (5) relevés ont été effectués : R6, R7, R8, R9, R10. Dans les forêts en contact avec les savanes incluses, les relevés sont R11, R12, R13, R14, R15 ; dans les forêts en contact avec les savanes sur versant, les cinq (5) relevés ont été R16, R17, R18, R19, R20. III.1. Affinité floristique entre les stations Les différents sites de relevés ont fait l'objet d'un test de similitude afin de distinguer leur degré d'affinité floristique (Tableau III). Tableau III : Matrice de similitude 
74.42 c : 76 c : 83 c : 80 Forêt SI 68.37 70.59 69.06 Alamena Forêt V Alamainty Alamena Forêt V Alamainty 77.55 74.44 Forêt SI c : 72 c : 77 c : 67 Forêt V : Forêt en contact avec les savanes sur versant Forêt SI : Forêt en contact avec les savanes incluses C=76 : Espèces communes des 2 stations (exemple) 77.55 : Taux de similarité (exemple) Les stations sont les forêts en contact avec les savanes incluses, les forêts en contact avec les savanes sur versant, les Alamainty et les Alamena. Les valeurs obtenues sont toutes supérieures à 50%, (entre 68 et 77%). Selon, le coefficient de similitude (Ps) de Sorensen, des stations présentent une ressemblance floristique lorsque le coefficient de similitude est supérieur à 50%. La végétation appartient donc à un même groupement écologique. Néanmoins, les Alamena, et les Alamainty sont les plus similaires floristiquement avec la valeur maximum du coefficient de similitude (Ps) de 77,55%. Les forêts en contact avec les savanes sur versant (forêt se trouvant en amont d'une savane non entourée par la forêt) présentent une affinité floristique avec les Alamena (74,44%) et les Alamainty (74,42%) (figure 5). 
Figure 5 : Dendrite de similitude Forêt V : Forêt en contact avec les savanes sur versant Forêt SI : Forêt en contact avec les savanes incluses. 
: Affinité floristique supérieur à 50 % : Affinité floristique supérieur a 75 % III.2. Analyse factorielle Afin de cerner les différents groupements végétaux, une analyse factorielle de correspondance sur la matrice de présence/ absence de 138 espèces dans 20 relevés a été effectuée. Les résultats sont consignés sur la figure 6. Le premier plan factoriel 1-2, explique environ 64% de la variabilité totale soit 42,07% pour l'axe 1 et 21,72% pour l'axe 2. Suivant l'axe 1, les relevés des Alamainty (G3) et des Alamena (G2) sont proches en abscisse négative. A ces relevés s'ajoutent les relevés des forêts en contact sur versant (aussi sur l'axe des abscisses négatives). Quant aux relevés des forêts en contact avec les savanes incluses, ils sont écartés en abscisse positive. Cette répartition des relevés serait liée au degré d'ouverture de la voûte forestière. L'axe 1 représente donc le degré d'ouverture de la voûte. 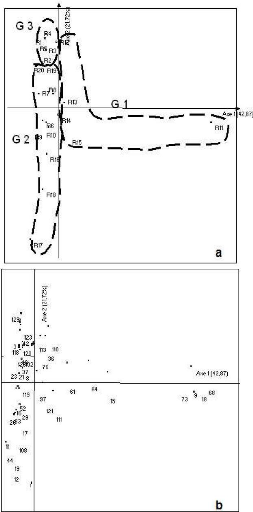
Figure 6 : Cartes factorielles dans le plan 1-2 des 20 relevés (a) et des 138 espèces (b) R1, R2, R3, R4, R5 : relevés des Alamainty ; R6, R7, R8, R9, R10 : relevés des Alamena ; R11, R12, R13, R14, R15 : relevés des forêts en contact avec les savanes incluses ; R16, R17, R18, R19, R20 : relevés des forêts en contact avec les savanes sur versant ; 1,...,129 : codage des espèces (un chiffre=une espèce). Ne sont représentées que les espèces ayant une forte contribution absolue et une bonne représentation Ainsi, trois groupements végétaux ont été identifiés : > Le groupe 1 regroupant les relevés des forêts en contact avec les savanes incluses (R11, R12, R13, R14, R15) est caractérisé par Vigna sp. (18), Peperomia sp. (9), Ficus soroceoides (73), Erythroxylum nitidilum (15), Mauloutchia sp. (64), Uapaca thouarsii (61), Pandanus sp. (111), Schefflera bojeri (121), Memecylon aff longipetalum (97), Dypsis sp. (76), Tina thouarsiana (36), Ocotea sp. (113) ; > le groupe 2 regroupant les relevés des Alamena et des forêts en contact avec les savanes sur versant (R6, R7, R8, R9, R10, R16, R17, R18, R19, R20) est caractérisé par Tambourissa hildebrandtii (3), Ephippiandra madagascariensis (4), Dypsis sp. (8), Erythroxylum nitidilum (16), Erythroxylum sp. (17), Oncostemum laurifolium (23), Bremeria sp. (26), Carissa edulis (29), Deinbolia sp. (37), Mollinaea retusa (40), Eugenia jambolana (83), Cephalostachyrum sp. (102), Schefflera vantsilana (119), Amerya humbertii (42), Psorospermum fanerana (21), Agarista salicifolia (52); > le groupe 3 regroupant les relevés des Alamainty (R1, R2, R3, R4, R5) est caractérisé par Dalbergia sp. (129), Polyscias ornifolia. (120), Ruellia cyanea (123), Polyscias sp. (118). Le groupe 3 présente un faible degré de perturbation et le groupe 2 est le plus perturbé des trois. L'axe 2 représente donc le degré de perturbation. III.3. Relation sol - végétation. Une analyse en composantes principales (ACP) est appliquée sur la matrice des quatre stations et les caractéristiques pédologiques sur une profondeur de 0 à 20 cm (figure 7 et 8). La figure 8, représente le cercle de corrélation des différentes caractéristiques physicochimiques du sol. Sur l'axe des abscisses, la teneur en potassium échangeable (K éch.), le rapport C/N, et la teneur en argile sont corrélées (abscisse positive) d'une part et l'acidité du sol et le pourcentage de sable d'autre part (abscisse négative). Le rapport C/N est le facteur le plus prépondérant de cet axe. Sur l'axe des ordonnées, la teneur en aluminium échangeable (Al ech.), en phosphore assimilable (P. Olsen) sont corrélés en ordonnées positives et s'opposent à la teneur en limon, en ordonnées négatives. L'aluminium contribue le plus à la formation de l'axe des ordonnées, ainsi, le facteur prépondérant de cet axe. 
Figure 7 : Carte factorielle des stations Forêt SI : forêt en contact avec les savanes incluses ; forêt V : forêt en contact avec les savanes sur versant 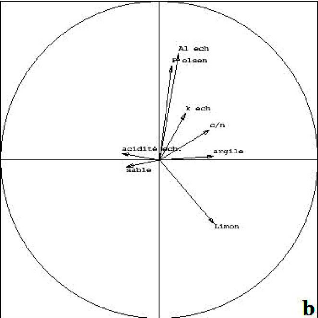
Figure 8 : Cercle de corrélation des propriétés physico-chimiques du sol K ech : Potassium échangeable, C/N : rapport en matière organique et matière minérale, P Olsen : Phosphore assimilable par la plante, Al ch. : aluminium échangeable En résumé : Le groupe 1 repose sur un sol argileux parfois sableux à moyen taux d'acidité (1,1) favorisant le développement des espèces. En effet, le taux de phosphore assimilable par les plantes est élevé (24,5 ppm) dans le sol. Le phosphore a un rôle dans la croissance des racines, l'implantation des jeunes plants, la floraison, la production et le mûrissement des fruits, la photosynthèse, la respiration et la croissance des plantes. Aussi, la richesse du sol en potassium (0,14 meq/100 g) a un effet sur la rigidité des espèces qui s'y développent. Le groupe 2 se développe sur un sol sableux et limoneux avec un faible taux de phosphore assimilable (5,1 ppm) mais un fort taux de potassium (0,14 meq/100 g) et de matière organique (14,3). Le groupe 3 est se développe sur un sol limoneux. Le sol du groupe 3 est donc favorable au développement des jeunes espèces mais le sol est très fragile. III.4. Structures biométriques En traitant les valeurs biométriques de la végétation (biovolume, densité des arbres à dhp supérieur à 10 cm, densité du peuplement, la richesse spécifique, la hauteur maximale, la régénération globale) par l'Analyse en composantes principales, les caractéristiques des différents groupements (figure 9) sont représentées sur la figure 10. Le premier plan factoriel absorbe 93% de la variabilité totale. Dans ce plan, l'axe 1 absorbe 70% de l'inertie totale. Sur le cercle de corrélation, les variables biovolume (V), densité des arbres supérieurs à 10 cm de diamètre (Nb), hauteur maximale (Hm), richesses spécifiques (Rs), sont corrélées entres elles. Le biovolume (V) et la densité des arbres supérieurs à 10 cm de diamètre avec la hauteur maximale sont les variables contribuant le plus à la formation de cet axe (abscisse négative). La variable Régénération (Rg) en abscisse positive est indépendante des autres variables. La variable densité du peuplement (D) n'intervient pas dans l'explication car elle se trouve au milieu du plan. Les relevés R15, R18, R13, R14 (figure 9) représentent une forte densité d'arbres de diamètre supérieur à 10 cm. Les relevés R1, R4, R5 sont les relevés à richesse spécifique (RS) et hauteur maximale (Hm) élevées ainsi qu'à biovolume (V) élevé. Enfin, la régénération (Rg) est élevée dans les relevés R19, R20, R9, R8, R12. 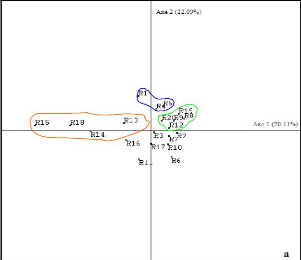
Figure 9 : Carte factorielle des 20 relevés 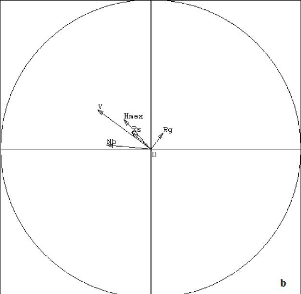
Figure 10 : Cercle de corrélation des variables biométriques V : biovolume, Hmax : hauteur maximale, Rg : potentiel de régénération, Rs : richesse spécifique, Nb : densité d'arbres supérieur à 10cm de diamètre, D : densité du peuplement III.5. Conclusion partielle Pour les 20 relevés effectués, dans le site d'étude, répartis dans les 4 stations considérées, 3 types de groupements ont été identifiés. Ces groupements sont caractérisés par leur composition floristique et la nature du substrat. Du point de vue végétation, le groupe 3 est encore considéré comme étant une formation naturelle « primaire ». La végétation renferme des espèces caractéristiques des formations humides. En outre la formation est haute (14 - 16m) avec des espèces de grands diamètres (dhp supérieur à 35 cm) et à contrefort. A l'inverse, le groupe 2 est caractérisé par des espèces arbustives héliophiles. En effet, la formation est très dense et moins haute. Celleci est causée par l'ouverture de la voûte favorisant le développement des espèces héliophiles. Le groupe 1 est une formation possédant des caractères floristiques et physionomiques se rapprochant du groupe 3. Du point de vue pédologique, le groupe 3 repose sur un sol plus riche en matière organique que le groupe 1 et le groupe 2. En outre, la texture ainsi que les éléments minéraux et organiques du sol sont favorables au développement des espèces. IV. Description des formations forestières La nomenclature des groupes a été choisie par les deux espèces les plus dominantes dans chaque groupe. Cette dominance se traduit par la contribution des espèces sur les axes lors de l'analyse factorielle. Les espèces les plus dominantes ont les valeurs de contribution les plus élevées. IV.1. Forêt à Ficus soroceoides et à Erythroxylum nitidilum (G1) IV.1.1. Stations et localité Le groupe 1 représente les forêts en contact avec les savanes incluses. La formation est localisée, loin des agglomérations et des villages, le long du sentier reliant le pays betsileo et le pays tanala. IV.1.2. Richesse floristique La formation est composée de 92 espèces réparties dans 70 genres et 41 familles. Les familles les plus représentées sont RUBIACEAE avec 10 espèces, EUPHORBIACEAE avec 10 espèces, ARALIACEAE avec 4 espèces et CLUSIACEAE avec 6 espèces. La liste des espèces est donnée dans l'annexe 4a. IV.1.3. Structure horizontale La formation est dominée par des individus de taille moyenne. La présence d'arbres de diamètre supérieure à 35cm de diamètre est rare. Le potentiel en bois est important avec un biovolume de 121m3/ha et une surface terrière de 26 m 2/ha avec un nombre d'arbres ayant un dhp supérieur à 10 cm de 1172 individus/ha. La densité du peuplement est de 21213 ind. /ha soit 2 ind. /m2. IV.1.4. Structure verticale La formation présente 3 strates, avec les émergeants. Aussi, la voûte forestière est assez fermée. Le nombre de strates et leur taux de recouvrement sont représentés dans les figures 11 et 12. La strate supérieure présente une hauteur comprise entre 10 et 15 m (figure 11) et un taux de recouvrement pouvant aller de 37 à 80% (figure 12). Les espèces les plus représentées sont : Labramia ditimena (SAPOTACEAEA), Ocotea sp. (LAURACEAE), Mauloutchia sp. (MYRISTICACEAE), Sideroxylon sp. (SAPOTACEAE), Garcinia sp. (CLUSIACEAE), Eugenia bernieri (MYRTACEAE), Ficus sp. (MORACEAE). La strate moyenne allant de 4 à 10m de hauteur ayant leur taux de recouvrement de 47 à 70%. La strate est composée par les mêmes espèces que la strate supérieure mais, les espèces suivantes sont également présentes : Dombeya lucida (STERCULIACEAE), Amerya humbertii (EUPHORBIACEAE), Mapouria parkerii (RUBIACEAE), Cyathea sp. (CYATHEACEAE), Gaertnera obovata (RUBIACEAE) et par la famille des ARALIACEAE représentée par les genres Polyscias et Schefflera. Résultats et interprétations 1 7 | 1 6 1 5 1 4 1 3 1 2 1 1 1 0 9 8 7 6 5 4 3 2 1 | |
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3 1 4 1 5 1 6 1 7 1 8 1 9 2 0 2 1 2 2 2 3 2 4 2 5 2 6 2 7 2 8 2 9 3 0 Figure 11 : Structure de la forêt G1 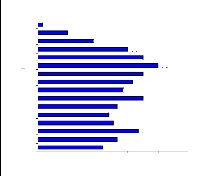
60 12_13 4_5 47 3 14_15 20 37 70 Hauteur (m) 10 11 8_9 6_7 53 57 63 70 70 80 0 20 40 60 80 100 Recouvrement (%) 2_3 0_1 50 67 53 43 Figure 12 : Diagramme de recouvrement de la forêt G1 La strate inférieure (0-4m) est composée surtout par des arbustes et des espèces herbacées tels que Cephalostachyrum sp. et Ruellia cyanea. Elle présente un taux de recouvrement compris entre 43 et 67%. Les espèces les plus fréquentes dans cette strate sont surtout Erythroxylum nitidilum (ERYTHROXYLACEAE), Drypetes madagascariensis (EUPHORBIACEAE), Oncostemum botrioïdes (MYRSINACEAE), Peperodium sp., Canthium humbertianum (RUBIACEAE). IV.1.5. Régénération globale L'allure de la figure 13 en « J » inversé indique que la formation est en bonne santé. Les individus régénérés sont plus nombreux que les individus semenciers. Ainsi, avec environ, 100 individus semenciers à l'hectare, le taux de régénération est de 474% ce quiindique une régénération moyenne de la formation. Effectifs (nb) 1200 1000 400 200 600 800 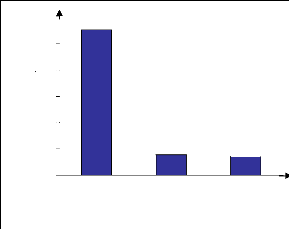
0 ss sse sse I II III Classe Classe I : 2,5<dhp<5cm Classe II : 5<dhp<10cm Classe III : dhp>10cm Figure 13 : Structure démographique de la forêt G1 IV.2. Forêt à Erythroxylum nitidilum et à Amerya humbertii G2) IV.2.1. Stations et localité Dans ce groupe sont incluses les forêts Alamena et les forêts en contact avec les savanes sur versant. Les forêts à forte densité d'arbres moyens sont localisées à l'intérieur de la forêt et les forêts en contact avec les savanes sur versant sont les plus proches des villages sur le sentier reliant le pays betsileo au pays tanala. IV.2.2. Richesse floristique L'inventaire fait état de 123 espèces réparties dans 66 genres et 39 familles. Les familles les plus représentées sont la famille des RUBIACEAE avec 10 genres et 14 espèces, EUPHORBIACEAE avec 6 genres et 14 espèces, CLUSIACEAE avec 6 genres et 8 espèces, ARALIACEAE avec 3 genres et 7 espèces. La liste floristique est donnée en annexe 4b. IV.2.3. Structure horizontale La formation est riche en individus de moyenne taille et en arbustes. De ce fait, la densité des individus est élevée avec un potentiel en bois moyen. En effet, la densité du peuplement est 27011 ind. /ha soit environ 3 ind. /m2. Respectivement, la surface terrière et le biovolume ont une valeur de 20 m2/ha et de 72 m3/ha. IV.2.4. Structure verticale La formation présente 4 strates bien distinctes représentées par les figures 14 et 15. La strate supérieure, a une hauteur comprise entre 8 à 12 m (figure 14). La strate est assez ouverte avec un taux de recouvrement allant de 43 à 70% (figure 15). Labramia ditimena (SAPOTACEAE), Weinmannia sp. (CUNONIACEAE), Uapaca thouarsii (EUPHORBIACEAE), Ocotea sp. (LAURACEAE), Anthocleista rhizophoides (LOGANIACEAE) et Syzigium onivense, Eugenia jambolana (MYRATCEAE) sont les espèces les plus représentées. La strate moyenne est entre 5 et 8m de hauteur, dont les principales espèces constitutives sont Symphonia microphilla (CLUSIACEAE), Ocotea cymosa (LAURACEAE), Cyathea sp. (CYATHEACEAE), Burasaria madagascariensis (MENISPERMACEAE), Petchia madagascariensis (APOCYNACEAE). Le taux de recouvrement environne les 57%. La strate arbustive est représentée par des espèces tels que Eliaea articulata, Calophyllum milvum, Psorospermum fanerana (CLUSIACEAE), Erythroxylum nitidilum (ERYTHROXYLACEAE), Croton glomeratus (EUPHORBIACEAE), Tina thouarsiana (SAPINDACEAE), Grewia opetala (TILIACEAE) et Oncostemum bothryoides (MYRSINACEAE). Ces espèces ont une hauteur entre 1 et 5m dont le taux de recouvrement varie de 47 à 60%. La strate herbacée est dominée par l'espèce Cephalostachyrum sp. (BAMBUSACEAE). Le taux de recouvrement, y est très élevé (environ 90%). Résultats et interprétations 
13
12 11 10 9 8 7 6 5 4 3 2 1 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 Figure 14 : Structure de la forêt G2 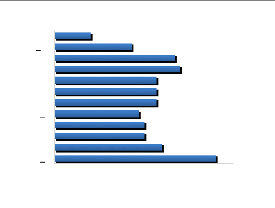
10 11 Hauteur (m) 8_9 6_7 4 5 2_3 0 1 0 20 40 60 80 100 Taux de recouvrement %) 43 67 70 57 57 57 47 50 50 60 90 20 Figure 15 : Diagramme de recouvrement de la forêt G2 IV.2.5. Régénération globale Les individus régénérés sont très nombreux et dominent la formation (figure 16). Les individus dits semenciers sont moins nombreux par rapport aux individus régénérés. Ainsi, selon la figure 16, la formation est en bonne santé avec un taux de régénération de 669%. La formation présente donc un potentiel de régénération moyen. 
Classe I : 2,5<dhp<5cm Classe II : 5<dhp<10cm Classe III : dhp>10cm 2500 2000 Effectifs (nb) 1500 1000 500 0 I II III s ss ss Classe Figure 16 : Structure démographique de la Forêt G2 IV.3. Forêt à Dalbergia sp. et à Polyscias ornifolia (G 3) IV.3.1. Stations et localité Ce sont les formations forestières composées d'arbres à gros diamètres, connues localement sous le nom d'Alamainty ou forêt naturelle. Ces forêts sont localisées au coeur du corridor loin des habitations et des sentiers. IV.3.2. Richesse floristique L'inventaire effectué fait état de 99 espèces réparties dans 73 genres et 43 familles. La liste floristique est donnée dans l'annexe 4c. Les familles les plus représentées sont les familles de RUBIACEAE avec 9 genres et 12 espèces, d'EUPHORBIACEAE avec 7 genres et 8 espèces, de CLUSIACEAE avec 5genres et 6 espèces et d'ARALIACEAE avec 2 genres et 6 espèces. IV.3.3. Structure horizontale La formation présente une rareté en espèces de moyenne taille (10cm <dhp< 35 cm) mais la présence d'arbres de diamètre supérieure à 35cm témoigne du potentiel en bois élevé. En effet, avec une densité d'arbres supérieure à 10 cm de diamètre d'environ 900 individus dans le plaçeau de 900m2, la formation présente une surface terrière de 30m2/ha et un biovolume de 169m3/ha. La densité du peuplement est de 21901 ind. /ha. IV.3.4. Structure verticale La formation présente 4 strates bien distinctes avec les émergeants (figure 17) Chaque strate présente un intervalle de taux de recouvrement représenté par la figure 18. La strate supérieure est comprise entre 10 et 16 m de hauteur (figure 17) avec un taux de recouvrement variant entre 47 à 73% (figure 18). Elle est constituée par des arbres de gros diamètre (dhp supérieur à 10 cm) des espèces suivantes : Eugenia bernieri, Syzigium emirnense (MYRTACEAE), Ocotea sp. (LAURACEAE), Albizia fastigata, Dalbergia sp. (FABACEAE). Des lianes se développent en s'agrippant sur le tronc des gros arbres afin de chercher la lumière au niveau de la canopée. Des épiphytes se développent aussi sur les troncs : des orchidées et des fougères. La strate moyenne est constituée par des arbres de moyenne taille. La hauteur des individus se situe entre 7 et 10 m. Les espèces constitutives sont Mapouria parkeri (RUBIACEAE), Macaranga alnifolia (EUPHORBIACEAE), Cyathea sp. (CYATHEACEAE), Polyscias aculeata, Polyscias sp. (ARALIACEAE). Le recouvrement de la strate varie entre 53 et 60%. La strate arbustive est dominée par la famille des RUBIACEAE telles que Psychotria sp, Canthium humbertianum, Peporidium sp., Bertiera sp., Gaertnera obovata, et quelques espèces à savoir Mollinaea retusa (SAPINDACEAE), Oncostemum laurifolium, Oncostemum falcifolium (MYRSINACEAE), Calophyllum milvum, Chrysophyllum boiviniana (CLUSIACEAE). Ces espèces ont une hauteur comprise entre 1 et 7 m dont le recouvrement est entre 43 et 53%. La strate inférieure est dominée par Ruellia cyanea (ACANTHACEAE) qui constitue un peuplement monospécifique hors les repousses des strates au-dessus. Le taux de recouvrement de cette strate est, très élevé, environ de 97%. 
1 7 _ 1 8 1 6 _ 1 7 1 5 _ 1 6 1 4 _ 1 5 1 3 _ 1 4 1 2 _ 1 3 1 1 _ 1 2 1 0 _ 1 1 9 _ 1 0 8 _ 9 7 _ 8 6 _ 7 5 _ 6 4 _ 5 3 _ 4 2 _ 3 1 _ 2 0 _ 1 0 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3 1 4 1 5 1 6 1 7 1 8 1 9 2 0 2 1 2 2 2 3 2 4 2 5 2 6 2 7 2 8 2 9 3 0 Figure 17 : Structure de la forêt G3 
0 1 73 73 33 47 63 53 40 40 60 60 53 43 15_16 12_13 Hauteur (m) 9_10 6_7 3_4 50 43 43 97 0 20 40 60 80 100 Recouvrement (%) Figure 18 : Diagramme de recouvrement de la forêt G3 IV.3.5. Régénération globale L'histogramme des classes de diamètres des arbres de la végétation est représenté sur la figure 19. La figure montre que les individus dites semenciers est en nombre inférieur par rapport aux espèces dites régénérées. Ce qui indique une bonne santé de la végétation. En effet, la formation présente un taux de régénération de 533% qui qualifie une régénération moyenne de la formation. 1600 1400 1200 Effectifs (nb) 1000 800 600 400 200 0 I II III s ss ss Classe Classe I : 2,5<dhp<5cm Classe II : 5<dhp<10cm Classe III : dhp>10cm Figure 19: Structure démographique de la forêt G3 V. Synthèse des résultats sur la typologie de la végétation Les données floristiques des trois (3) groupements sont consignées dans le tableau IV. Tableau IV : Caractères floristiques et biométriques des différentes formations 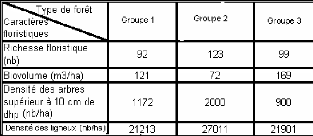
Floristiquement, les trois groupes sont riches avec un biovolume moyen mais la valeur du groupe 1 et du groupe 3 est toutefois plus élevé que celle du groupe 2, même si le groupe 2 présente la densité en gros diamètre la plus élevée. Pour déduire la classification de degré de perturbation des groupes. Les variables, biovolume et richesse floristique des stations formant les groupes, ont fait l'objet d'une analyse de variance selon le test de Tukey (HSD) qui consiste à distinguer les stations présentant une affinité. Les résultats sont consignés sur le tableau V. Tableau V : Richesse floristique et biovolume des types de forêts Modalités Richesse floristique (nb) Biovolume (m3/ha) | Stations Alamena 62 (a) 72 (a) Forêt V 61 (a) 75 (a) Alamainty (ab) 60 (a) 117 (a) Forêt SI (a) 48 (b) 121 (a) Probabilité Théorique 0.05 0.05 Calculé 0.02 (S) 0.30 (NS) |
Les moyennes suivies d'une (ou des) même (s) lettre (s) appartiennent au même groupe statistiquement homogène au seuil de probabilité de 0.05 d'après le test de Tukey HSD. S : Différence significative NS : Différence non significative Forêt V : Forêts en contact avec les savanes sur versant Forêt SI : Forêt en contact avec les savanes incluses Entre les 4 stations étudiées, le test de Tukey présente des différences significatives sur la richesse floristique avec un degré de confiance de 95%. La valeur minimale (48) a été obtenue dans les forêts en contact avec les savanes incluses et la valeur maximale autour de 60 a été obtenue dans les forêts de références et les forêts en contact sur versant (Forêt V). La valeur minimale du point de vue floristique peut indiquer la présence de prélèvement d'espèces. Les résultats du point de vue floristique ont montré que les Alamena et les Forêts en contact avec les savanes sur versant appartiennent au même groupe. La valeur du biovolume des deux forêts est dans l'intervalle des biovolumes moyens (Roger & Rabarison, 2000). L'Alamainty et les Forêts en contact avec les savanes incluses appartiennent à 2 groupes différents. Du point de vue richesse floristique, l'existence des 3 groupes est vérifiée. En ce qui concerne le biovolume, le test n'est pas significatif. Ainsi, les stations sont statistiquement homogènes. Néanmoins, la valeur biovolume entre les stations présente une large différence. Les valeurs élevées (121, 117) ont été obtenues dans les Alamainty et les forêts en contact avec les savanes incluses. Les valeurs minimales (72, 75) ont été obtenues sur les Alamena et sur les forêts en contact avec les savanes sur versant. La richesse en espèces présente 2 significations opposées. Soit, la richesse spécifique indique une forêt mature ou primaire peu perturbée soit elle peut être interprétée comme un signe de perturbation. En effet, une formation perturbée engendre l'ouverture de la voûte qui se traduit par le développement des espèces héliophiles qui pourront occuper tout l'espace de la formation. Pour discriminer l'état de la formation, le biovolume est aussi une variable non négligeable. Ainsi, une faible richesse floristique mais un biovolume élevé indique une formation mature perturbée et une richesse floristique élevée mais un biovolume faible indique une formation perturbée. Si la richesse en espèces et le biovolume sont élevés, la formation peut être considérée comme une formation peu perturbée. Les trois groupes sont aussi mis en évidence par l'existence et l'abondance de Cephalostachyrum sp.et de Ruellia cyanea dans la strate herbacée. L'abondance de Cepahalostachyrum sp. indique une formation sensible à la pénétration de la lumière ainsi, une formation ouverte et/ou perturbée. L'abondance de Ruellia cyanea, une espèce sciaphyle, indique une formation humide avec une voûte forestière assez fermée. Ainsi, le groupe 1 est une formation mature perturbée, le groupe 2 est une formation mature de crête ou de pente et le groupe 3 est une formation mature peu perturbée. VI. Principales menaces et pressions Les populations riveraines du corridor forestier sont des agriculteurs. L'élevage et la riziculture sont des pratiques complémentaires de la population locale. Parmi les activités humaines qui pourraient présenter des menaces sur la formation forestière ainsi que sur leur reconstitution, l'exploitation forestière, l'élevage de zébus, le feu de pâturage et la pratique de l'abattis-brûlis ou tevy sont les plus importantes. Ces différentes menaces influent beaucoup sur la dynamique de la forêt (figure 20). La forêt mature sous l'influence et selon l'intensité des activités humaines comme l'exploitation forestière massive et l'entrée des zébus en forêt, peut se transformer en forêt dégradée, mais peut redevenir une forêt mature si la perturbation est arrêtée. Toutefois si ces différentes pressions sont associées à la culture sur brûlis, la forêt peut disparaître laissant la place à des cultures. Après abandon cultural, la parcelle peut redevenir une forêt dégradée. Si la zone est transformée en zone de pâturage, il y a évolution vers une savane incluse. La persistance de ces savanes est favorisée par la mise à feu pour l'entretien du pâturage. 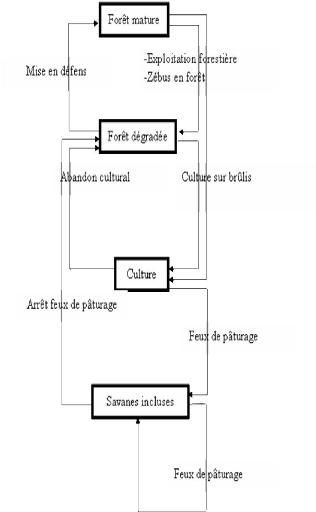
Figure 20 : Schéma hypothétique de la dynamique forestière Ainsi, les activités qui influent sur la dynamique de la végétation sont le pastoralisme à travers les modes d'élevages et le feu de pâturage, l'exploitation forestière et la culture sur brûlis ou le tevy. VI.1. Mode d'élevage Dans le Fokontany d'Andohanimananatanana, la taille moyenne des troupeaux est de 11 têtes de zébus. La taille maximale (24 têtes de zébus) est enregistrée dans les villages. Les zébus sont utilisés dans les travaux de rizières à savoir le piétinement, le labour, le hersage des mottes (photo 1), mais également pour la reproduction d'un cheptel destiné à la viande ainsi qu'à la production de fumier. Selon le besoin des paysans et la localisation des éleveurs par rapport aux forêts, le mode d'élevage est généralement de deux types : l'élevage extensif et l'élevage semi-extensif ou intensif. Le mode d'élevage dans la région dépend surtout de l'éloignement de la forêt. Aussi, la composition des troupeaux dépend du mode d'élevage pratiquée. Pour l'élevage intensif, les zébus sont surtout composés de zébus mâles castrés, avec quelques males non castrés ; Les zébus femelles sont rares. Contrairement, pour l'élevage semi extensif et extensif, le troupeau est majoritairement composé de zébus femelles avec quelques mâles non castrés. 
Photo 1 : Zébus mâles castrés travaillant la terre VI.1.1. Elevage extensif L'élevage extensif consiste à envoyer les zébus dans la forêt toute l'année. L'emplacement des zébus dans les forêts est appelé « Kijana », littéralement, zone de pâturage. Chaque famille a son propre Kijana. Ces zones de pâturage se trouvent sur une vaste étendue de forêts et de savanes incluses avec des points d'eau (rivière, lac, marais). Les zébus pâturent seulement sur les espaces ouverts pendant les périodes froides (mai - juillet) et de pluie (décembre - mars) pour avoir les premiers rayons du soleil. Les propriétaires ne les amènent dans les villages que pour le travail et pour des événements socioculturels comme le Volambita et les Sao-draza au mois de Juillet (photo 2 et 3). Lors des séjours aux villages, les troupeaux ne restent que 1 à 3 jours ; car ils risquent de tomber malade du fait du changement brusque d'alimentation. Actuellement, certains Raiamandreny pratiquent toujours l'élevage extensif. Ils habitent dans les villages, mais chargent les membres de la famille, résidant dans les campements, la responsabilité du suivi des animaux. L'élevage extensif est surtout pratiqué pour lutter contre les Dahalo. 
Photo 2 : Tolon'omby lors d'un Sao-draza. Photo 3 : Parcage des zébus lors d'un sao-draza VI.1.2. Elevage semi-extensif et intensif Avec la création des Aires Protégées à Madagascar, le mode d'élevage extensif laisse peu à peu la place aux types d'élevages semi-extensif et intensif. Le mode d'élevage est fonction de l'éloignement des villages aux forêts. Dans les villages, les zébus sont gardés dans les savanes le jour et parqués la nuit. Les zébus dans les villages sont principalement des zébus mâles castrés destinés à produire de fumier ou pour le travail aux champs. Dans les villages, le mode d'élevage est principalement de l'élevage intensif. En s'approchant de la forêt, le mode d'élevage se fait de 2 manières, soit d'une manière intensive : les zébus sont gardés dans les savanes et les abords des forêts le jour et ramenés dans les parcs la nuit, soit, d'une manière semi-extensive, où les zébus entrent en forêt du mois d'août au mois d'avril suivant. Ils sont ramenés dans les villages en lisières ou dans les campements pour 2 ou 3 jours entre le mois de septembre et octobre, pour le travail aux champs. Entre le mois de mai et le mois de juillet, le troupeau est parqué dans les parcs. Tous les membres de la famille (5 à 60 ans) sont concernés par le gardiennage de zébus (photo 4). Pour le parcage des zébus en forêts, ce sont les chefs de familles accompagnés des cousins ou des frères venant des villages qui s'en occupent. 
Photo 4 : Gardiennage de zébus par les enfants Ce mode d'élevage présente des conséquences sur la typologie des forêts et sur la dynamique des écosystèmes. VI.1.3. Conséquences de l'élevage sur les écosystèmes D'après la population riveraine, l'entrée des zébus en forêt ne présente pas de conséquence dévastatrice sur l'écosystème forestier. Au contraire l'entrée des zébus en forêt est bénéfique à cause des fumures des zébus ; ainsi, les zébus favorisent la régénération des jeunes pousses en accélérant la décomposition de la litière par piétinement. Néanmoins, les zébus en forêt mangent les jeunes pousses et les jeunes feuilles des plantes. Aussi, le piétinement des zébus rend le sol plus compact ralentissant ainsi la régénération de la forêt. De plus, la mise à feu des savanes incluses pour le fourrage des zébus en forêt ne permet pas la recolonisation de la forêt. Les savanes sont annuellement brûlées, détruisant ainsi les jeunes pousses. En élevage intensif, les fourrages fournis par les savanes sont insuffisants pour alimenter le troupeau toute l'année. Les espèces herbacées des savanes sont appréciées par les zébus pendant la période entre le mois de décembre et le mois d'avril. Le reste du temps, les zébus sont alimentés par les pailles de riz (mois septembre - octobre) et les fourrages bordant les bas fonds et rizières. Ainsi, la mise à feu des savanes pour le renouvellement des fourrages est nécessaire pour alimenter le troupeau. Toutefois, la mise à feu sur les savanes provoque d'une part sur le sol, un appauvrissement en éléments et ainsi l'incapacité de certaines espèces à se développer sur le milieu. D'autre part, elle ralentit la reconstitution forestière. VI.2. Exploitations forestières A la fin de la période coloniale, entre 1937 et 1957, des surfaces importantes de forêts ont été exploitées par les colons associés aux Merina avec une main d'oeuvre betsileo. Les espèces les plus exploitées étaient le bois d'ébène (Diospyros sp.), le palissandre (Dalbergia sp.) et le bois de rose (Dalbergia maritima). Ces espèces se font rares dans la région actuellement et ne peuvent être trouvés que dans les Alamainty (forêt dite « naturelle »). Après cette période d'exploitation massive de bois dans la région, la population riveraine fait des prélèvements dans les plantations d'Eucalyptus bordant les villages et des arbres morts pour s'approvisionner en bois de chauffe et pour la construction des maisons en bois. L'Eucalyptus est aussi utilisé pour la fabrication de charbon de bois. En forêts, les riverains prélèvent des arbustes à tige droite (CLUSIACEAE) pour la fabrication des cannes servant dans le gardiennage des zébus. Les contreforts des arbres (FABACEAE, ANACARDIACEAE) sont utilisés pour la fabrication des vannes. Les troncs des bois durs (EBENACEAE) sont utilisés pour la fabrication de manche d'angady, des mortiers et des pilons et les feuilles des Pandanus sp. (PANDANACEAE) pour la fabrication des nattes. VI.3. Culture sur brûlis ou tevy La croissance démographique oblige la population locale à migrer petit à petit vers la forêt pour la recherche de nouvelles terres cultivables ; le sol forestier étant considéré comme plus productif. De plus, la production en riz des rizières dans les bas fonds à l'extérieur de la forêt étant insuffisante, il est de plus en plus nécessaire de combiner riziculture et d'autres cultures (Photo 5 et 6) comme les maïs, des légumineuses (haricots, pois de bambara ....). Les terrains les plus exploités sont les bas fonds ou les terrains au bord de la rivière. Selon la perception locale, la présence du genre Solanum auriculatum (Seva beSOLANACEAE), Dionycha sp. (Kitonda - MELASTOMACEAE) ainsi que Cyperus sp. (Herana - CYPERACEAE) sur les abords des forêts près d'une rivière témoigne un sol de type « baiboho », indiquant une fertilité élevée du milieu. 
Photo 5 : Tevy en lisière de la forêt. Photo 6 : Plantation de maïs après Tevy. VI.4. Feu de pâturage La pratique de la mise à feu dans la région est interdite depuis 2002, date à laquelle, le GCF a été crée. La population locale n'a plus le droit d'y mettre le feu. Mais cette interdiction n'empêche en rien de trouver lors des descentes de terrain (mai 2008), des savanes nouvellement brûlées ou carrément des savanes ou des versants en train de brûler en lisière de forêt. Pour l'alimentation des troupeaux, les savanes sont annuellement parcourues par le feu avant 2003. La population locale met généralement le feu sur les savanes avec autorisation des autorités durant la période entre le 15 novembre et le 15 janvier, après la tombée des premières pluies. Néanmoins, certains pratiquent la mise à feu pendant le mois de juillet pour avoir plus de fourrage pour la période suivante de pâturage. Le feu a pour but de transformer la nécromasse sur pied en cendres, et d'accélérer la repousse de l'herbe pour l'alimentation du bétail. L'action du feu sur l'écosystème présente cependant des effets néfastes au niveau du sol et de la forêt. L'action répétée du feu sur le sol entraîne son appauvrissement en éléments organiques. Les bactéries du sol transformant les matières organiques en matières minérales ne peuvent plus y vivre. Après l'interdiction de la mise à feu par les autorités en 2003, à l'abord des hameaux, les savanes sont envahies par les Erica sp. (ERICACEAE). La qualité fourragère est ainsidiminuée. L'Erica sp. commence à coloniser le milieu après 2 ou 3 ans sans mise à feu. VII. Evolution spatio-temporelle de la végétation VII.1. Evolution des savanes incluses La carte des formations végétales (1937) ne montre pas l'existence des savanes incluses (carte 4). Toutefois, une aire de défriche a été localisée à proximité de Sahabe et le sentier a déjà existé. En 1957 (carte 5), les savanes existent dans la région. A proximité de Sahabe, les savanes sur versant occupent une surface importante. A l'Est de cette grande savane sur versant se trouvent les savanes incluses dont la surface diminue au fur et à mesure qu'on se rapproche de la falaise Tanala. La présence de ces savanes peut être mise en relation avec les activités effectuées de 1937 à 1957. Les différentes activités menée dans cette période sont : (1) l'exploitation forestière de bois précieux ; (2) exploitation minière ; (3) projet d'une route reliant le district d'Ambalavao à la côte Est. Les savanes incluses sont issues de ces diverses activités menées durant la période coloniale dans la forêt de Sahabe, entre 1937 et 1957. De 1957-2004, les savanes n'ont pas beaucoup évolué (carte 6). Toutefois, une légère augmentation des savanes incluses est notée (15%) dans la période 1957-1991. En cette période, l'élevage extensif était le mode d'élevage pratiqué par la population locale. Ainsi, la mise à feu des savanes était de mise. Par contre, dans la période 1991-2004, la reconstitution forestière est notée mais très lentement. La surface de forêt défrichée autour des savanes incluses n'est pas comblée par cette reconstitution forestière. Sur la carte 6, les limites forestières se distinguent nettement entre 1937 et 1957, alors qu'elles tendent à se confondre entre 1957 et 1991 sur un fond de carte de 2004. 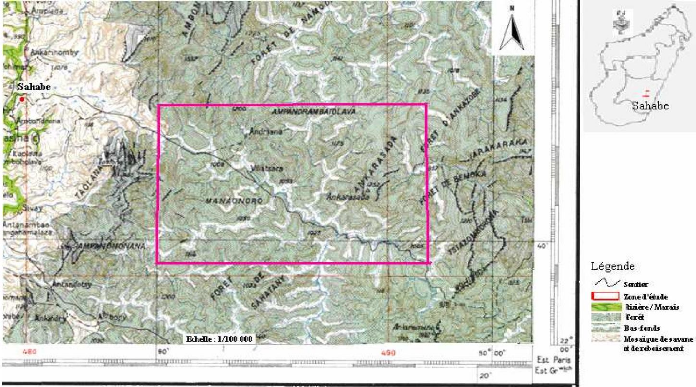
Carte 4:Le site d'étude en 1940 (extrait de la carte de la commune d'Ambohimahamasina ; source : photo aérienne 1937) 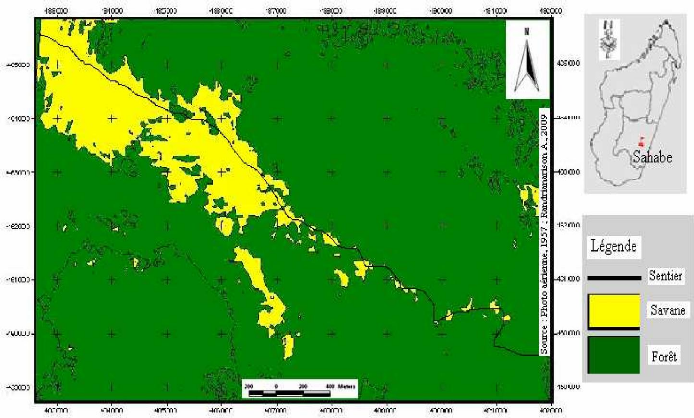
Carte 5 : Le site d'étude en 1957 (source : photo aérienne 1957 ; MEM, 2008) 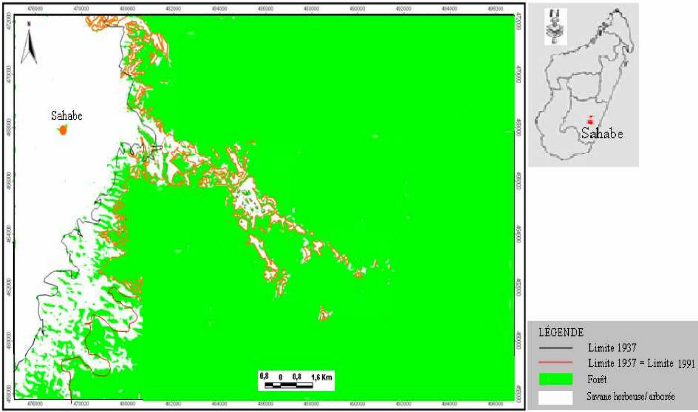
Carte 6 : Evolution des limites de la lisière sur un fonds de carte 2004 (source : carte topographique 1937, photos aériennes 1957 et 1991, image Google-Earth 2004 ; MEM, 2008) VII.2. Principales tendances évolutives des savanes incluses La typologie de la tendance évolutive des savanes incluses est basée sur l'évolution des surfaces de ces savanes. Depuis 1957, les savanes incluses n'ont pas évolué de la même manière (annexe 5) : - Quatre savanes incluses (A1, A2, A 3, A13) ont diminué de surface depuis 1957 jusqu'en 2004 ; - Deux savanes incluses (A4, campement) ont progressé en surface depuis 1957 jusqu'en 2004 ; - Cinq savanes incluses (A7, A8-A9-A10, A11, A12, A5) ont augmenté de surface depuis 1957 jusqu'en 1991 et ont diminué un peu jusqu'à 2004 ; - Une savane incluse (A6) a augmenté de surface dans le période 1957-1991 et diminué de surface dans le période 1991-2004 mais avec une surface inférieure à celle de 1957. Ces différentes unités de savanes incluses sont regroupées dans un type de tendance évolutive selon leur évolution (carte 7) : savane régressive, savane progressive et savane stable. VII.2.1. Savane régressive Les savanes incluses regroupées dans ce type de tendance sont les savanes incluses qui ont actuellement une surface inférieure à celle de 1957. Ces savanes sont surtout des savanes localisées loin des habitations humaines. Ces savanes incluses sont des savanes jadis utilisées (pâturage, déforestation, zone de culture) ; actuellement, elles se présentent soit sous forme de savoka soit avec une lisière forestière large. VII.2.2. Savane progressive Les savanes incluses regroupées dans ce type de tendance sont les savanes qui n'ont cessé d'augmenter de surface depuis 1957. Ces savanes incluses sont des unités présentant une zone marécageuse mais à dominance de savane. Les savanes incluses présentant ce type de tendance sont localisées sur des terrains hors sentiers et près d'une rivière. 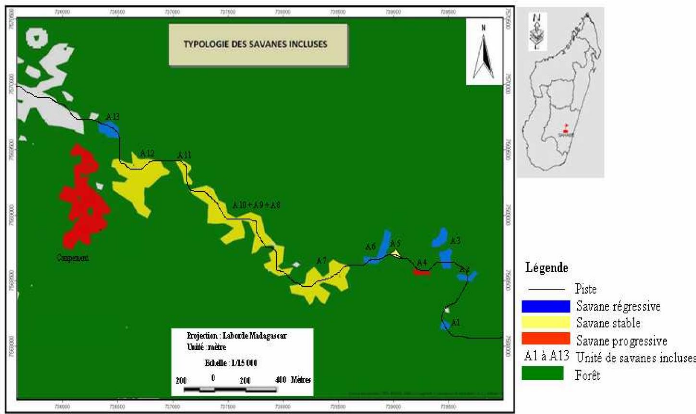
Carte 7: Localisation des types de savanes incluses (source : BD 100, 2005 ; MEM, 2008) Ces marais sont utilisés pour la riziculture. Mais avec l'insuffisance de la production, la parcelle de riz est agrandie au détriment des savanes. C'est cet aménagement qui cause l'augmentation de surface de la clairière parce que les propriétaires ont le droit de nettoyer les alentours pour se protéger contre les ravageurs. Aussi, des campements sont installés pour stocker la production. VII.2.3. Savane stable Les unités regroupées dans ce type de tendance sont des savanes qui ont connu une légère augmentation de surface après 1957 et qui ont diminué par la suite. Toutefois, la surface de ces savanes est supérieure à celle de 1957. Ces savanes incluses sont localisées le long du sentier. Ces savanes sont utilisées seulement par la population locale pour le pâturage. Ce qui fait qu'avec la maîtrise de la pratique de feu dans la région, la forêt n'est pas vraiment en danger et commence à avancer vers la savane mais avec une vitesse très lente. VII.3. Dynamique des groupements forestiers en contact avec les savanes incluses Les savanes incluses présentent donc une dynamique spatiale régressive ou progressive. Les forêts qui sont en contact avec ces savanes présentent également des évolutions régressive et progressive. Ainsi, les savanes régressives correspondent aux forêts progressives et les savanes progressives correspondent aux forêts régressives. VII.3.1. Forêt progressive Une évolution progressive de la forêt est observée sur des zones où les pressions humaines sont faibles. Ces zones correspondent à des zones utilisées comme zone de pâturage, ou des abandons culturaux. Le feu pratiqué annuellement pour l'entretien du pâturage ralentit la reconstitution forestière. En effet, sur les zones anciennement cultivées ou pâturées, les espèces herbacées sont parsemées d'espèces ligneuses. Même sur les savanes actuellement utilisées pour le pâturage (photo 7), une avancée de la forêt peut être perceptible. Celle-ci se manifeste par un phénomène d'embroussaillement, composé surtout de fougère tels que Dicranopteris sp., de Erica sp. (ERICACEAE) et des espèces ligneuses comme Aphloia theaformis (APHLOIACEAE), Haronga madagascariensis (HYPERICACEAE) et Gravesia capitata (MELASTOMATACEAE). 
Photo 7 : Savane nouvellement brûlée pour le pâturage (01/2008) VII.3.2. Forêt régressive L'évolution régressive est notée sur les zones où les activités humaines sont fortes. Ces zones sont surtout des campements ou des zones nouvellement aménagées en rizières entraînant un recul de la forêt pour les besoins agricoles (photo 8). 
Photo 8 : Aménagement de bas fonds en rizière VIII. Conclusion partielle L'étude de la dynamique des écosystèmes forestiers en contact avec les savanes incluses dans le corridor forestier de Fianarantsoa nous a conduits à déterminer 3 groupements : une formation mature à Dalbergia sp. et à Polyscias ornifolia (G 3), une formation mature peu perturbée à Ficus soroceoïdes et à Erythroxylum nitidilum (G 1), et une formation perturbée à Erythroxylum nitidilum et Amerya humbertii (G2). Le groupe 1 est la formation qui entoure les savanes incluses. Toutefois, elle reste mature car la pratique de la mise à feu des savanes dans la région est bien contrôlée. L'utilisation des pare-feu est bien respectée. De plus, la période de mise à feu est seulement autorisée pendant la saison de pluie. Ainsi, la forêt est bien protégée contre le feu. Selon leurs emplacements et leurs proximités des villages ou campement, les savanes incluses ne présentent pas la même tendance évolutive. Certaines ont augmenté de surface depuis leurs apparitions jusque dans les années 1990 et d'autres ont par contre diminué. Mais quelque soit la tendance évolutive de ces savanes incluses dans les années 90, depuis, elles régressent toutes sauf celles qui sont habitées ou aménagées en rizière. Néanmoins, d'une façon générale, les savanes incluses diminuent de surface mais n'ont pas encore atteint leurs surfaces de 1957. Parallèlement, la forêt a connu ainsi une évolution suivant le mouvement de la limite forestière. Ayant un campement en forêt, les occupants commencent à aménager des rizières et à posséder des zébus. Ainsi, la pratique de la mise à feu des savanes incluses est effectuée pour l'alimentation des troupeaux. La pratique de mise à feu est la cause de la persistance des ces savanes incluses. Des tevy sont aussi effectués pour la culture des produits de contre saisons. L'entrée des zébus en forêt est une pratique qui se transmet d'une génération à une autre. Cette pratique selon la perception paysanne ne présente pas d'effet néfaste sur la forêt. Néanmoins, cette pratique modifie la physionomie et la composition floristique du milieu. Actuellement, ce mode d'élevage est de plus en plus abandonné mais laisse des traces comme l'aménagement des zones de pâturages en rizières ou en campement. QUATRIEME PARTIE DISCUSSION et RECOMMANDATIONS DISCUSSION ET RECOMMANDATIONS Cette partie portera sur les problèmes méthodologiques, les remarques sur les résultats ainsi que quelques recommandations pour améliorer la pratique agricole et pastorale. Discussion PROBLEMES METHODOLOGIQUES La matérialisation des placeaux, de dimension standard de 0.1 ha pour les forêts tropicales, a posé de nombreux problèmes au cours des différents relevés. Ceci est du au relief et à la topographie : la longueur de la pente ne dépasse pas les 50m. Or pour réaliser un placeau de 0,1ha, une dimension de 20m x 50m ou quelques fois, 10m x 100m est nécessaire. Ainsi, le dispositif du placeau effectué lors des études sur le terrain a été modifié. Un placeau de 30m x 30m a été adopté. Au moment des descentes sur le terrain, la plupart des plantes étaient à l'état végétatif, ce qui nous a posé des problèmes pour l'identification des espèces. En effet, pour la détermination des espèces, les fleurs ou les fruits sont des caractères importants pour discriminer une espèce des autres espèces ou d'un genre à l'autre. Sans ces caractères, plusieurs plantes sont restées au niveau genre. REMARQUES SUR LES RESULTATS Typologie des formations L'hétérogénéité de la formation ainsi que la présence des activités anthropiques anciennes et actuelles dans le chef lieu de Sahabe ne permet plus l'utilisation du terme forêt « primaire ». Ce terme n'est peut être plus valable pour qualifier les forêts à Madagascar. En effet, des recherches effectués à Madagascar ont montré la faible représentation de ces forêts dite « primaire » (Ranarivelo & Kotozafy, 2001 ; Moreau, 2002). Nos résultats ont corroboré ceux de Carrière et al., (2007) sur la présence de forêt mature dans le corridor. Ainsi, la formation forestière constituant le corridor de Fianarantsoa peut être qualifiée de forêt mature. Toutefois, selon Goodman & Rakotoarisoa (1998), dans la réserve spéciale du pic d'Ivohibe, un site situé plus au Sud, excepté quelques traces d'une occupation humaine ancienne, la forêt et les communautés animales ne montrent plus aucune trace récente de cette occupation humaine. C'est comme si la perturbation ancienne n'existait pas et la forêt était restée primaire. Typologie des formations selon la perception paysanne Selon la perception paysanne, la forêt dans la zone de Sahabe est de 2 types : Alamainty et Alamena. Les Alamena sont identifiées par la population locale comme des formations à forte densité d'arbres de moyen diamètre et les racines sont peu profondes. Certains arbres présentent des racines échasses pour mieux se fixer sur le substrat (sol meuble par la forte teneur en sable). La formation est aussi caractérisée par l'abondance des litières non encore décomposées. Il est à signaler aussi l'abondance d'espèces herbacées vivant en quasi-monopeuplement dans la strate herbacée ce qui les rend sensibles au feu. Quant aux Alamainty, la formation élevée et présente des individus de gros diamètre présentant des contreforts et dont la strate herbacée est dominée par la famille des Acanthacées (Ruellia cyanea). Ces 2 types de forêts renferment des espèces communes mais leur croissance est différente. Ainsi, pour la population locale, les Alamainty sont des forêts dites « naturelles » et les Alamena peuvent être des forêts dégradées, des forêts jeunes ou régénérées ou des forêts se trouvant sur un niveau topographique supérieur (crête, haut versant). Ecologiquement, les Alamena sont des formations secondaires issues d'une perturbation dont les strates tendent à se confondre. Elle est dominée par des espèces héliophiles du fait de l'ouverture de la voûte forestière favorisant le développement des espèces herbacées tels que Cephalostachyrum sp.. Les Alamainty sont des formations hautes comprenant des strates bien distinctes. La strate herbacée est dominée par des espèces sciaphiles à cause de l'humidité et de la fermeture de la voûte forestière. La forêt en contact avec les savanes sur versant et les Alamena font partie d'un même groupe. Les Alamena sont donc sensibles aux feux et peuvent rapidement se transformer en savane. Origine des savanes de la région Nombreuses sont les études réalisées pour connaître l'origine des savanes à Madagascar. D'une part, des chercheurs ont supposé que Madagascar est autrefois recouvert de forêt et affirment que la présence de ces savanes résulterait de la déforestation due à l'augmentation démographique (Perrier de la Bathie, 1921, 1936 ; Humbert, 1927 ; Gade, 1996). Ils défendent l'idée que la forêt recouvrait tout les hauts- plateaux (voire tout l'île) et que c'est l'homme avec ses actions inconsidérées qui a façonné le paysage actuel. D'autre part, des chercheurs comme Dewar & Wright (1993), Burney (1996 ; 1997), assurent grâce à des études de pollen que la végétation couvrant les Hautes Terres, il y a 2 000 ans, était plutôt de nature mixte, composée de forêts, d'arbustes et de prairies et ceci à cause de nombreux incendies pendant la quaternaire. Aussi, l'analyse de charbons montre que durant la fin du Pléistocène et le début de l'Holocène, les 3/4 des charbons proviennent de Graminées. La savane existait donc déjà pendant la quaternaire et se serait maintenue grâce à l'allongement et à l'accentuation de la période sèche à la fin de l'Holocène (1 500-1 000ans). Dans cette partie Sud de l'ex-province de Fianarantsoa, un mythe sur l'existence d'un « afotroa » circule et qui serait à l'origine des savanes (Rainihifina 1958 ; Battistini & Verin, 1966). L' « afotroa » ou un grand feu avait pris son origine dans la partie Ouest de Madagascar (région du Menabe) puis s'était répandu vers l'Est et avait brûlé certaines parties du corridor forestier. L'époque de l'existence de ce feu n'est pas connue mais la datation au C14 d'horizons tourbeux des bas fonds dans la cuvette d'Ambalavao (Portais, 1974) confirme l'existence d'un feu remontant à 600 ans environ. En effet, des prélèvements dans les sédiments lacustres montrent effectivement autour du XIIIème siècle une augmentation des feux, qui se traduit dans les sédiments par l'abondance des dépôts de charbons, la fréquence des pollens de graminées et la diminution des pollens de ligneux (Moreau, 2002). La population locale accuse ce « afotroa » d'être à l'origine des savanes et même pour certains des savanes incluses dans la région. Les savanes incluses dans la forêt de Sahabe sont apparues en 20 ans (1937-1957). Cette apparition est liée aux activités diverses effectuées du temps de la colonisation (exploitation forestière massive, exploitation minière, projet d'une route reliant le district d'Ambalavao et la côte est de l'île). La main d'oeuvre était fournie par des Betsileo de la région ou venant d'Ambalavao. Travailler pour les Merina et les colons a empêché les Betsileo de développer leur propre économie agricole. Des longues zones aménageables en lisière de forêt et en savane sont restées disponibles après la colonisation. En effet, après la colonisation, la surface des savanes incluses est restée apparemment stable (1957-2004). En effet, les savanes incluses sont utilisées seulement pour le pâturage. Mais actuellement (2008) de nouvelles trouées sont apparues. Ces trouées ne sont pas considérées comme des savanes incluses car ce sont surtout des tevy et des bas fonds nouvellement aménagés en rizières. La surface totale de ces nouvelles trouées est de 6 ha (Andrianarivo et al., 2008). Après plusieurs cycles culturaux, ces trouées peuvent devenir un jour des savanes incluses. Et comme pour les savanes incluses déjà présentes, le feu inhibe complètement la reconstitution forestière. Dynamique de l'occupation du sol Etant donné que la forêt présente déjà des difficultés à se reconstituer, les activités humaines ne font rien pour y remédier. Les pressions exercées sur la formation sont surtout le tevy, aménagement des bas fonds en rizières et le feu de pâturage laissant des vides et des creux à l'intérieur d'une vaste forêt verte. Ces ouvertures menacent réellement la continuité du corridor forestier. En effet, elles sont surtout localisées près de la falaise Tanala là où la forêt est la plus dense et la mieux préservée. La pression démographique est la cause directe de la migration des villageois vers la forêt. Cette migration s'accompagne toujours d'une recherche de terre cultivable. En outre, en forêt, les terrains et les bas fonds sont vastes, et les sols des forêts humides sont considérés comme les sols les plus riches (Vande Weghe, 2004). Ces occupations du sol sont confirmées par une analyse d'image SPOT 2004 effectuée par Serpantié et al. (2006) sur le Nord du couloir Ranomafana - Andringitra. En forêt, le sol est très fertile. Et c'est cet aspect que la population recherche en multipliant les tevy et les aménagements de bas-fonds. Mais, malgré la dégradation que ces activités provoquent sur la forêt, elles gardent encore une place primordiale dans le système de production rizicole de la population des hauts plateaux (Ravoavy & Messerli, 2001). Toutefois, dans la zone d'étude, nombreux sont les bas-fonds non encore aménagés en lisière des forêts. Mais, dans le pays betsileo, la dynamique d'occupation des bas-fonds progresse d'Ouest en Est, d'abord en savane, puis en forêt (Rakotoasimbahoaka et al., 2007). A ces pressions s'ajoute l'élevage des zébus en forêt ou l'élevage extensif. L'élevage extensif ne présente pas vraiment de menaces sur les zones de pâturage situées en forêt même. Certes, l'élevage extensif modifie la structure initiale de la végétation mais à l'échelle de la végétation, cette perturbation est faible. Selon Meyer (1991), la contribution de l'élevage dans le processus de dégradation et de déforestation est inférieure à 20% voire 11% du processus global de la déforestation. Ce processus de déforestation n'est pas alors perceptible par la population. Pour les éleveurs de zébus, ils refusent de croire que le fait de pratiquer le pâturage en forêt détruit la forêt. D'autant plus le pâturage en forêt est déjà pratiqué depuis longtemps : « cette pratique ne détruit pas la forêt, la preuve, elle est toujours là ». Ainsi, il pourrait exister un équilibre entre élevage et écosystèmes forestiers (Rakotoniaina S.N., 1994 ; Moreau, 2002 ; Ranaivoson, 2006). Recommandations Pour les Betsileo, la pratique agricole est toujours liée à la pratique pastorale. Le déséquilibre entre ces 2 pratiques présente une conséquence sur les écosystèmes. Afin qu'il y ait toujours un équilibre entre ces pratiques, quelques suggestions en vue d'améliorer les pratiques agricoles et pastorales sont proposées pour maintenir l'intégrité des écosystèmes forestiers et savanicoles : - Au cours des prospections et des séjours effectués dans la région, nous avons repéré de nombreuses zones marécageuses, sur le sentier qui n'étaient pas encore exploitées. La promotion de l'aménagement de ces bas- fonds est donc nécessaire afin de limiter les tevy en forêt et les aménagements en rizières. Mais pour l'aménagement de ces terrains, des matériels adéquats devraient leur être délivrés comme des bêches, charrues. - Les savoka occupent une place importante sur l'occupation du sol. Ainsi, la population locale devrait être incitée à exploiter ces parcelles plutôt que des parcelles en forêt. - Aussi, la promotion de l'élevage de zébus pour la production de fumier est aussi une alternative pour préserver la forêt. En effet selon Rakotoniaina (1994), l'élevage peut être considéré comme un moyen pour limiter la conquête des espaces vers la forêt. Plus une unité de production possède de boeufs, plus elle dispose de moyens de production et de fumier pour favoriser une culture intensive, d'où le détachement de cette unité de production de la forêt (Ranaivoson, 2006). Mais, afin de protéger la forêt, la mise à feu des savanes pour le pâturage de zébus devrait suivre des règles bien définies. Des suggestions sont donc à donner (Ratovonirina, 2009) : - Après quelques années (2 ou 3 ans), les Erica sp. (Anjavidy - ERICACEAE) et Vernonia sp. (Kizity - ASTERACEAE) commencent à envahir la savane. Ainsi, une rotation de mise à feu devrait être faite. En effet, si le feu passe une fois tous les 2 ou 3 ans sur une savane, la savane resterait verte tout en maintenant la qualité fourragère. - D'habitude, les agriculteurs pratiquent la mise à feu au mois de juillet. La mise à feu devrait être avancée pendant la période des pluies, avant le mois d'avril, ceci pour protéger la forêt contre un éventuel accident. - Même pendant la saison des pluies, des pare-feux devraient toujours être effectués pour limiter la puissance du feu. - Pour compléter l'alimentation des zébus, des cultures fourragères devront aussi être implantées par les agriculteurs. CONCLUSION GENERALE CONCLUSION GENERALE L'étude de la dynamique des écosystèmes forestiers en contact avec les savanes incluses dans le corridor de Fianarantsoa : Sahabe - Ambohimahamasina nous a permis de dégager que les savanes situées dans la zone d'étude sont des savanes post-forestières. Ces savanes incluses sont apparues après des exploitations massives pendant la période coloniale (avant 1947). Ces savanes ont été ensuite brûlées pour la culture et après quelques cycles de culture, la mise à feu pour l'entretien du pâturage. Le feu de pâturage est la principale cause de la persistance de ces savanes incluses. L'évolution de ces savanes dépend de l'intensité et de la fréquence des activités humaines sur le milieu. Certaines savanes incluses sont donc absorbées petit à petit par la forêt et d'autres au contraire ont augmenté de surface. Mais globalement, la surface des savanes incluses diminue laissant la place à la forêt. Néanmoins, cette diminution de surface des savanes est compensée en surface par les ouvertures en forêt, par les tevy et/ou l'aménagement des bas fonds. Du fait de ces activités, la végétation dans la région est une végétation hétérogène composée de forêt, savane, savoka, etc. La forêt est une formation mature dont le degré de perturbation varie selon sa proximité aux agglomérations. Les forêts en contact avec les savanes incluses sont considérées comme des forêts matures perturbées mais elles présentent une possibilité de régénération si toutefois l'action du feu sur les savanes, ainsi que les défrichements et les tevy, sont minimisés. Dans les zones où le feu est absent, les savanes sont hautes et la lisière est progressive. Ainsi, la dynamique de la végétation est régie par les degrés des perturbations. La forêt la plus éloignée des villages et campements est susceptible de conquérir de l'espace. En effet, la forêt des hautes terres malgaches présente une forte résilience mais ce sont les activités qui y sont effectuées qui la stoppent. Ainsi, pendant les périodes de très fortes pressions, la forêt régresse mais dès que celles-ci sont levées, elle s'installe. Le sol peut avoir aussi une influence sur la reconstitution forestière. En effet, sur les plateaux près des marais, la reconstitution forestière est significative contrairement aux forêts qui se trouvent en amont d'une savane. Conclusion générale L'étude de la lisière, combinée avec le suivi des limites et l'effet des différentes pressions évoquées, serait un atout pour aboutir à l'étude de l'évolution spatio-temporelle de la végétation. Sa physionomie, son sol, et la connaissance de sa flore et de sa superficie (ou sa longueur) pourraient nous aider dans la compréhension de la dynamique de la végétation. Aussi, cette étude peut nous donner des explications sur les différentes étapes de la dynamique selon les différents types d'évolution de la végétation. Mais pour ne pas rester sur une seule impression, des études analogues devront être effectuées sur les 8 autres sentiers traversant le corridor. Néanmoins, le présent travail permet déjà de visualiser et de comprendre les phénomènes de savanisation et de reconstitution de la forêt vis-à-vis des différentes pressions et/ou perturbations. BIBLIOGRAHIE BIBLIOGRAPHIE Andriamahazo M., Ebene Onana Y.C., Ibrahima A., Komena B.K., Razafindrandimby J., 2004 - Concilier exploitation des ressources naturelles et protection de la forêt. Cas du corridor forestier de Fianarantsoa (Madagascar). Rapport ICRA/CNRE-IRDMontpellier. Madagascar. 98 p. Andrianarivo A., Hervé D., Randrianarison A., Ratovonirina G., Razanaka S., 2008 - Dynamiques spatio-temporelles des savanes incluses en forêt : centre du corridor forestier de Fianarantsoa. In Télédétection et gestion de l'environnement : receuil des résumés, S. Randriamanga, S. Rakotoniaina, S. Rakotondraompiana, J.P. Gastellu-Etchegorry, K. Boulkroune, Y. Auda (eds). Les XIème journées scientifiques du réseau télédétection de l'AUF. Antananarivo, Madagascar du 3 au 7 novembre 2008. pp. 121-122. Disponible sur : www.reseautd.auf.org/IMG/pdf/sommaireJS2008.pdf Battistini R. & Verin P., 1966 - Les datation a Madagascar par la méthode du radio-carbone 14. Semaine Géologique. pp. 9-12. Benzécri J.P., 1981 - Histoire et préhistoire de l'analyse des données. Paris : Dunod (collection "Pratique de l'analyse des données"). Besairie H., 1972 - Géologie de Madagascar, les terrains sédimentaires. Annales géologiques de Madagascar. Fasc.XXXV. Tananarive. 463 p. Besairie H., 1973 - Précis de géologie malgache. Annales de Géologie de Madagascar. Fasc XXXVI. Tananarive. 142 p. Boulanger J. & Rantoanina M., 1957 - Carte géologique de Fort Carnot/Ambohimahamasina. Burney D.A., 1996 - Climate change and fire ecology as factors in the quaternary biogeography of Madagascar. In Biogeography de Madagascar, W.R. Lourenco (ed). ORSTOM, Paris. pp. 49-58. Burney D.A., 1997 - Theories and facts regarding Holocene environmental charge before and after human colonization. In Natural and human impact in Madagascar, S M. Goodman, BD. Patterson (eds). Smithsonian Institution Press, Washington & London. pp. 75-89. Carrière S.M., Ratsimisetra L., Roger E., 2007 - Le couloir forestier de Fianarantsoa : forêt «primaire» ou forêt des hommes. In Transitions agraires, dynamiques écologiques et consérvation : le"corridor"Ranomafana-Andingitra Madagascar, G. Serpantié, Rasolofoharinoro, S. Carrière. Actes du séminaire GEREM. Antananarivo, 9-11 novembre 2006. pp. 39-46. Chantraine J., 1967 - Géologie et prospection de la région Ambohimahasoa-FianarantsoaIfanadina. Service Géologique, Antananarivo. Notice + carte au 1/100 000, feuille O 53 et P 53. Dajoz. R., 1975 - Précis d'écologie. Ecologie fondamentale et appliquée. Gauthiers-Villars. Paris, 534 p. Dawkins A.C., 1959 - The management of natural tropical high-forest, with special reference to Uganda. Commonwealth forestry, Institute University of Oxford. England. 155p. FAO, 2001 - State of the world's forests. F A O. Rome. Gade D., 1996 - Deforestation Madagascar. Human role in changing the face of the earth. Mountain Research and Development, 16(2), pp. 101 - 116. Gautier L., 1994 - Structure et flore de la forêt sur la pente d'Andranomay. Recherche pour le développement. MRS-CIDST. pp. 14-28. Godron M., Daget P., Long G., Sauvage C., Emberger L., Le Gloch E., Poissonnet J., Wacquant J P., 1983 - Relevé méthodologique de la végétation et du milieu code et transcription sur une carte perforée. Ed. CNRS. Paris. 281 p. Goodman S.M. & Rakotoarisoa J.-A., 1998 - Un regard sur l'utilisation historique et sur la régénération des habitats naturels à Madagascar. Akon'ny Ala, 24 : 3-4. Gouet J.P. & Philippeau G., 1989 - Comment interpréter les résultats d'une analyse de variance. STAT-ITCF, Service des Etudes statistiques - Instituts techniques des céréales et des fourrages. Paris. 47 p. Gounot M., 1969 - Méthode d'étude quantitative de la végétation. Masson. Paris VI. 314 p. Guinochet M., 1973 - Phytosociologie, collection d'ecologie1. Ed, Masson et compagnie. Paris VI. 227 p. Humbert H., 1927 - La destruction d'une flore insulaire par le feu. Principaux aspect de la végétation à Madagascar. Mémoire de l'Académie Malgache, fasc. IV. 47 p.
Humbert H. & Cours Darne G., 1965 - Carte international du tapis végétal et des conditions écologiques. 3 coupures au 1/1000 000 de Madagascar. Travaux de la section scientifique et technique de l'institut Français de Pondichéry (hors série). Langrand O., 1995 - Guide des oiseaux de Madagascar. Delachaux et Niestlé, Paris. 415 p. Lanley J.P., 1992 - Dernière données sur les forêts dans le monde. Flamboyant, 23-24: 26-31. Meyer N., 1991 - The hamburger connection: how Central America's forest become North America's hamburgers. Ambio. 10(1); pp. 3-8. Moreau S., 2002 - Les gens de la lisière. La forêt, l'arbre et la construction d'une civilisation paysanne Sud-Betsileo, Madagascar. Thèse de Doctorat. Ecole doctorale Milieu, Culture et Société du passé et du présent. Paris. 667 p. Parde J., 1961 - Dendrométrie. Ed. de l'école nationale des eaux et forêts. Nancy. pp.120-349. PCD de la commune rurale d'Ambohimahamasina, 2006 - Monographie de la commune. 47 p. (Document) Portais M., 1974 - Le bassin d'Ambalavao : influence urbaine et évolution des campagnes (Sud Betsileo Madagascar). ORSTOM. Paris. 162 p. Perrier de la bathie H., 1921 - La végétation malgache. Annales de l'institut Botanicogéologique Colonial de Marseille, Sér. 3(9). pp. 12-26. Perrier de la bathie H., 1936 - Biogéographie des plantes de Madagascar. In Atlas de Madagascar. Société d'éditions Géographiques maritimes et coloniales. Paris. pp. 116-128. Rainihifina, 1958 - Lovantsaina Betsileo, Fianarantsoa. Imprimerie catholique, vol 1. 240 pp. Rajaonarivelo R.G., 2001 - Les problèmes observées entre le VOI, les organismes d'appui et d'encadrement, le service des eaux et forêts, dans le mise en oeuvre du transfert de gestion des ressources naturelles : l'exemple su site d'Ambohimahamasina. Communication du VOI Lovasoa Ambohimahamasina. 13 p. (document)
Rakotoarimanana V. & Grouzis M., 2008 - Effet à court terme du feu et du pâturage sur la qualité fourragère d'une savane à Heteropogon contortus du Sud-Ouest de Madagascar. Revue Elev. Med.Vet. Pays trop. 2008, 61(2). pp. 81-88. Disponible sur : // remvt.cirad.fr/cd/derniers-num/2008/EMUT08-081-088.pdf. Rakotoasimbahoaka C.R., Serpantié G., Toillier A., Treuil J.P., Ramamonjisoa A.B.O., Hervé D., 2007 - Un automate cellulaire pour modéliser l'aménagement des bas-fonds en fonction du peuplement. In Transitions agraires, dynamiques écologiques et conservation : le"corridor"Ranomafana-Andingitra Madagascar, G. Serpantié, Rasolofoharinoro, S. Carrière. Actes du séminaire GEREM. Antananarivo, 9-11 novembre 2006. pp. 69-81. Rakotoniaina S.N., 1994 - Contribution à l'étude des impacts des troupeaux bovins sur l'écosystème forestier du Parc National de Ranomafana. Mémoire d'Ingéniorat. Département Eau et Forêt, ESSA. Université d'Antananarivo. 63 p. Ranarivelo H.S. & Kotozafy A., 2001 - Chapitre 4. Etude de la structure et la composition floristique de la végétation dans les forêts de basse altitude (<1 000m) du couloir forestier entre le Parc National (PN) de Ranomafana et le PN d'Andringitra. Recherche pour le Développement, 17 : 47-91. Ranaivoson N.R.E., 2006 - Dynamique du système d'élevage bovin dans une zone péri- forestière de Madagascar. Cas de la commune d'Androy (Corridor forestier Ranomafana-Andringitra). Mémoire de DEA en Agro-management. ESSA, Université d'Antananarivo/Programme GEREM. 66 p. Ratovonirina G., 2009 - Dynamique des savanes incluses dans le corridor de Fianarantsoa : zone Sahabe - Ambohimahamasina. Mémoire de DEA, option Ecologie Végétale. Faculté des Sciences, Département de Biologie et Ecologie végétales, Université d'Antananarivo/Programme MEM. 65 p. Ravoavy L.N. & Messerli P., 2001 - La culture sans brûlis, une possibilité d'alternative au tavy ? Résultats des essais effectués dans la région de Beforona-Cote Est. In Sociétépaysannes, transitions agraires et dynamiques écologiques dans le sud-ouest de Madagascar, S. Razanaka, M. Grouzis, P. Milleville, B. Moizo, C. Aubry. Actes de l'atelier CNRE-IRD, 8-10 novembre 1999. pp. 293-304. Roger E. & Rabarison H., 2000 - Contexte biologique de la conservation des forêts à Madagascar. In Etude sur la politique de conservation des ressources forestières à Madagascar. 53 p. Rollet G., 1979 - Application de diverses méthodes d'analyse de données à des inventaires forestiers tropicales. Oecol plant, 14(3). pp. 319-344. Rothe P.L., 1964 - Régénération naturelle en forêt tropicale : le Dipterocarpus dyeri (vau) sur le versant cambodgien du golfe de Siam. Bois et Forêt des Tropiques, 8. pp. 386- 397. Schatz G.E. & Malcomber S.T., 1993 - Floristic composition of one hectare plot in Ranomafana National Park. In Biodiversity in Ranomafana National Park. Madagascar, P. Wright (eds). State University of New York. Press Albany (In press). Serpantié G., Toillier A., Rakotoasimbahoaka C., Hervé D., Treuil J.P., 2006 - Eléments pour la modélisation de la déforestation dans la région du corridor de Fianarantsoa (Madagascar). Colloque « Modélisation à l'interface entre natures et sociétés », NSS-IRD-CIRAD, 7-9/12/05, Montpellier (résumé et poster). Serpantié G., Toillier A., Carrière S., Razanaka S., 2007 - Déforestation à Fianarantsoa au 20ème siècle : un "corridor"en sursis. In Transitions agraires, dynamiques écologiques et conservation : le"corridor"Ranomafana-Andingitra Madagascar, G. Serpantié, Rasolofoharinoro, S. Carrière. Actes du séminaire GEREM. Antananarivo, 9-11 novembre 2006. pp. 47-56. Sorensen T., 1948 - A method of establishing groups of equal amplitude in plant sociology based on similarity of species content. 183 p. Vande Weghe J.P., 2004 - Forêt d'Afrique centrale : la nature et l'homme. Edition lannoo SA. Belgique. 357 p. WEBOGRAPHIE [Web 01], CMP Tandavanala, 2008 - Caractéristiques du corridor de Fianarantsoa. Disponible sur : http://www.cmpfianar.mg/corridorcaracteristique.html Consulté en décembre 2008 [Web 02], PRD Haute Matsiatra, 2004. Disponible sur : www.madagascar-diplomatie.ch/Politique/Région/haute Consulté en décembre 2008
Annexe 1 : Géologie de Madagascar (Besairie, 1972). 
Annexe 2 : Fiche d'enquête agropastorale CONCERNANT LES PERSONNES ENQUETEES 1. Nom et prénoms : 2. Age : 3. Sexe : 4. Domicile principal et secondaire (temporaire): 5. Nombre de personnes dans la famille : hommes, femmes, enfants, homme en age de travailler 6. Origine : Betsileo, Tanala ou autres 7. Date d'installation au village 8. Responsabilité dans la famille : chef de famille ? Bouvier ? 9. la personne enquêtée est- elle propriétaire du troupeau ou non CONCERNANT L'AGRICULTURE 1. Riziculture : nombre de parcelles - localisation ?types de rizières : bas-fonds/talweg/tourbe altitude 2. Pratiquez-vous le repiquage en ligne ?cultures de contre saison ?si oui, les quels ? 3. Relation culture - élevage : élevage contemplatif ? Ou zébus utilisés à quoi ? 4. Nombre de parcelles sur le rebord des rizières + cultures 2007_2008 5. Nombre de parcelles sur pentes + cultures 2007_2008 CONCERNANT LES ZEBUS I. TROUPEAUX 1. Consultation du « bokinomby » chez le chef fokontany afin de cibler les personnes à enquêter, d'établir l'évolution de l'effectif des zébus dans le temps et qu'on pourra comparer avec l'évolution de la surface des savanes entre 1957, 1991, 2008 2. Nombre de zébu : mâle, femelle, jeune (et les propriétaires s'ils sont différents des personnes enquêtées) 3. Ventes, achat ou prêt d'animaux 4. Evolution des nombres de zébus depuis 50 ans (augmente, stable, diminue). Raisons ? 5. Comment ils ont constitué le troupeau actuel de zébus ? (Héritage, achat, prêt, naissance...) 6. Date de constitution du troupeau initial à leur nom ? II. ALIMENTATION DU TROUPEAU 1. Elevage : · Période d'entrée en forêt · Raisons : · Quelle partie du troupeau part en forêt ? quelle partie reste ? · Période de sortie de forêt · Raisons : · Parcours des zébus dans la forêt. Jusqu'où ? Parcours connus ? parcours appropriées · Parcours des zébus dans les savanes ?parcours appropriées ? · Point d'abreuvement en forêt ? · Ressources fourragères ? (paille de riz, jachères, farihy (marais), tamboho (savane), kilanja, savanes incluses, affouragement) a. Période de l'année ? mois ressources | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | |
2. Apport élevage de bovin dans le revenu familial : · Riz, · Autres cultures, · Rhum, · Vente de travail, · Zébu, · Autres... Qui s'en occupe (gardiennage, traite, piétinage)? · Père, · Enfant (âge), · Vieux (âge), · Salarié (paiement en nature ou en argent) · L'entrée des zébus dans la forêt est- elle autorisée ou non ? Depuis quelle date ? Oui : Date Non : Date : · La mise à feu des savanes incluses est elle autorisée ou non .depuis quelle date ? Oui : Date Non : Date : Perception paysanne sur les impacts du pâturage ? CONCERNANT LES SAVANES 1) Historiques des savanes · Est-ce les savanes existaient déjà dès son jeune âge (pour les personnes enquêtées, plus de 60 ans) · Existe-t-il des mythes ou légendes racontant les origines de savanes et des forêts du corridor ? · Le chemin qui traverse le corridor existe-t-il déjà avant 1957 ? Avait-il autant ou plus d'importance que maintenant pour les étrangers, les Betsileo, les Tanala? · Jusqu'où y avaient des maisons et des hotely le long du chemin ? · Il y a 50 ans, le feu était il interdit ou non ? · D'où proviennent ces appellations des savanes ? (Andasy, Andasivazaha, Viliantsara, Ampanenitra,...) ? · Existe-t-elle une avancée ou un recul de savane d'après les enquêtées ? quelle est la cause ? · Qui sont les premiers propriétaires, il y avait 50 ans ? Généalogie 2) Les propriétaires · Vivent-ils à Sahabe? Ambohimahamasina, ailleurs ? · Ont-elles changé de propriétaires depuis 50 ans ? Comment ? 3) Le feu · Quels sont les buts de la mise à feu des savanes ? · Qui met le feu aux savanes : propriétaire, passager, gardien de zébus ? Accidentelle ou volontaire ? · Y a-t-il des dates de la mise à feu ?décision collective ou individuelle · Meilleur période de mise à feu pour la production d'herbe, pour la protection de la forêt ? · La fréquence du feu est-elle liée à la taille du troupeau · Quelle production fourragère si on met le feu ?si on ne met pas le feu ? Annexe 3 : Résultats de l'analyse chimique et granulométrique du sol
| stations | argile (%) | Limon (%) | sable (%) | | Forêt V | 23 | 21 | 56 | | Forêt V | 26 | 11 | 62 | | Forêt V | 11 | 8 | 81 | | Forêt S I | 7 | 12 | 81 | | Forêt S I | 12 | 8 | 79 | | Forêt S I | 22 | 13 | 65 | | Forêt S I | 23 | 13 | 65 | | Alamainty | 17 | 17 | 66 | | Alamainty | 15 | 32 | 53 | | Alamena | 5 | 5 | 91 | | stations | acidité
ech. | Al ech (%) | P olsen
(ppm) | k ech
(mecli100g) | c/n | | Forêt V | 1.1 | 0.7 | 4.4 | 0.12 | 8 | | Forêt V | 1.3 | 2.3 | 5.8 | 0.14 | 14.3 | | Forêt V | 1.8 | 2.9 | 18 | 0.2 | 14.3 | | Forêt S I | 1.7 | 0.6 | 3.6 | 0.05 | 0 | | Forêt S I | 0.8 | 1.8 | 9.6 | 0.07 | 14.5 | | Forêt S I | 0.5 | 3.1 | 8.6 | 0.15 | 17.3 | | Forêt S I | 1.1 | 3.8 | 24.6 | 0.14 | 13.6 | | Alamainty | 1.1 | 0.2 | 3.2 | 0.08 | 20 | | Alamainty | 0.3 | 0 | 4.7 | 0.04 | 9 | | Alamena | 1 | 0.4 | 3.4 | 0.03 | 2 |
Annexe 4 : Liste des espèces | Annexe 4a : Liste floristique du groupe 1 | | FAMILLE | Nom scientifique | Nom vernaculaire | | APHLOIACEAE | Aphloia theaformis Bennet | Fandramanana | | APOCYNACEAE | Carissa edulis Vahl | Fatsy | | Mascarenhasia arborescens D.C. | Herotra | | Petchia madagascariensis | | | AQUIFOLIACEAE | Ilex mitis (L) Radlk | Hazondrano | | ARALIACEAE | Polyscias aculeata | Vatsila | | Polyscias ornifolia | Vatsila be ravy | | Schefflera vantsilana (Baker) Bernardii | Vatsila | | Schefflera bojeri (Seeman) Bernardii | Vatsila | | ARECACEAE | Dypsis sp1. | | | Dypsis sp2. | Fanikara | | Dypsis sp3. | Rapaka | | ASTERACEAE | Brachylaena ramiflora | Mananitra | | BAMBUSACEAE | Cephalostachyrum sp. | Tsimbolo | | Nastus sp. | Voloando | | BIGNONIACEAE | Colea cf lutescens H.P | Voalava | | CELASTRACEAE | Polycardia libera O. | Miholy | | CLUSIACEAE | Calophyllum milvum RF Stevens | Vitano | | Garcinia aphanophlebia Bak. | Kimbaletaka | | Garcinia verrucosa Jum & H, Perr | Kimbamena | | Mammea madagascariensis | Kimbafotsy | | Symphonia clusoides Baker | Kimba be ravy | | Symphonia microphilla Benth & Hook | Kimba mandinindravy | | CUNONIACEAE | Weinmannia rutembergii Eng | Lalombary | | Weinmannia rhodoxylon Tul | Lalo be ravy | | Weinmannia bojeriana | Lalomaka | | CYATHEACEAE | Cyathea sp. | Lohanapa | | CYPERACEAE | Scleria arbotiva | Vendra | | EBENACEAE | Dyospiros gracilipes Hiern | Sily mainty | | ERYTHROXYLACEAE | Erythroxylum sp1. | | | Erythroxylum sp2. | | | Erythroxylum ferrugenum Cav | Malambovony | | Erythroxylum nitidilum | Menahihy | | EUPHORBIACEAE | Amyrea | humbertii | | Hazonovy | | Croton | tiligium | | | | Croton | mongue | | Sily fotsy | | Drypetes | madagascariensis (Lam) Humbert Leandri | et | Lomaka | | Drypetes | perrieri Leandri | | Lomaka ravin-tsakay | | Drypetes | madagascariensis (Lam) Humbert Leandri | et | Mariravy | | Euphorbia | tetraptera Baker | | Ramanjavona | | Macaranga | alnifolia Baker | | Mokaranana | | Macaranga | gralata Mc Pherson | | Tarambito | | Uapaca | thouarsii | | Mera | | FABACEAE | Vigna | sp. | | | | Dalbergia | sp. | | Voamboana | | LAURACEAE | Ravensara | sp. | | Molaliambo | | Cryptocaria | sp. | | Vagna | | Ocotea | sp. | | Varongy | | LILIACEAE | Dracaena | reflexa Lam | | Tavola be ravy | | Dracaena | sp. | | Tavola kely ravy | | Dianella | ensifolia | | Vazahanakampo | | LOGANIACEAE | Nuxia | sp. | | Lambinana gaga | | MALPHIGIACEAE | Acridocarpus | sp. | | Sily | | MALVACEAE | Dombeya | cf lucida Baillon | | Somanga | | MELASTOMATACEAE | Memecylon | aff longipetalum H.Perr | | Tomenja | | MONIMIACEAE | Ephippiandra | madagascariensis (Dangay) Lorence | | Ambora mavo | | Tambourissa | hildebrandtii Perkins | | Ambora | | Tambourissa | religiosa | | Kidehodeho | | MORACEAE | Ficus | soroceoides | | Kivozy | | Ficus | reflexa Thumb | | Nonoka | | MYRISTICACEAE | Mauloutchia | sp. | | | | MYRSINACEAE | Oncostemum | laurifolium Mez | | Farampandoha | | Oncostemum | bothryoides Baker | | Kitohitohy | | MYRTACEAE | Eugenia | bernieri Baill ex Drake | | Rotra | | Eugenia | jambolana Lam | | Rotra | | Syzigium | emirnense Baker | | Rotre mandinidravy | | OLACACEAE | Olax | sp. | | Afoatsinanana | | PANDANACEAE | Pandanus | sp 1 . | | Vakoa mainty | | Pandanus | sp2. | | Vakoa tsirika | | PIPERACEAE | Peperomia sp. | Benjakinoly | | RHAMNACEAE | Bathiorhamnus louvelii | | | RUBIACEAE | Bertiera sp. | | | Bremeria sp. | Fatoralahy | | Canthium humbertianum Cavaco | Fatsikahitra | | Captosperana chevatum | Tsivoka | | Gaertnera obovata (Baker) var obovata | Tsivoka kely ravy | | Mapouria parkeri | Korofokorana | | Peporidium sp. | Fatsikala | | Psychotria sp. | Voamasondrenikely | | Psychotria madagascariensis | Voamasondrenikely mandinidravy | | Tricalysia cryptocalix Baker | Tavaza | | RUTACEAE | Vepris fitoravina | Ampody | | SAPINDACEAE | Deinbolia pervillei (Blume) Radlk | Hazombato | | Mollinaea retusa Radlk | Hazombato | | Tina thouarsiana (Cambess) Capuron | Hazombato | | Tinopsis dasycarpa | Lanary | | SAPOTACEAE | Labramia ditimena H. Perr | Tarantalahy | | Sideroxylon betsimisarakum Lecomte | Nato | | SCROPHULARIACEA E | Halleria tetragona Baker | Kitoto | | STERCULIACEAE | Dombeya lucida | Hazombahy | | TILIACEAE | Grewia opetala | Mainty hoditra | | VACCINACEAE | Vaccinium emirnense Hook | Aramontsy |
Annexe 4 b | Annexe 4b : Liste floristique du groupe 2 | | FAMILLE | Nom scientifique | Nom vernaculaire | | ANNONACEAE | Xylopia | buxifolia HBN | Ramiavo | | APHLOIACEAE | Aphloia | theaformis Bennet | Fandramanana | | APOCYNACEAE | Carissa | edulis Vahl | Fatsy | | Petchia | madagascariensis | | | AQUIFOLIACEAE | Ilex | mitis (L) Radlk | Hazondrano | | ARALIACEAE | Polyscias | aculeata | Vatsila | | Polyscias | repanda | Vatsila | | Polyscias | ornifolia | Vatsila | | Polyscias | sp1. | Vatsila | | sp2. | Vatsila | | | Schefflera | vantsilana (Baker) Bernardii | Vatsila | | Schefflera | bojeri (Seeman) Bernardii | Vatsila | | ARECACEAE | Dypsis | sp1. | | | sp2. | Fanikara | | sp3. | Rapaka | | | ASTERACEAE | Brachylaena | ramiflora | Mananitra | | Helichrysium | cordifolium | | | Vernonia | garnieriana Klatt | Kandalahy | | Vernonia | delapsa (Baker) | Maranitsiatoraka | | BAMBUSACEAE | Cephalostachyrum | sp. | Tsimbolo | | Nastus | sp. | Voloando | | BIGNONIACEAE | Colea | cf lutescens H.P | Voalava | | CELASTRACEAE | Mystroxylon | aethiopicum | Fanazava | | Polycardia | libera O. | Miholy | | CLUSIACEAE | Calophyllum | milvum RF Stevens | Vitano | | Eliaea | articulata Cambees | Ketoke | | Garcinia | sp. | Kimbaletaka | | Garcinia | verrucosa Jum & H, Perr | Kimbamena | | Mammea | madagascariensis | Kimbafotsy | | Mammea | bongo | Kimbaletaka | | Psorospermum | fanerana Baker | Fanera | | Symphonia | microphilla Benth & Hook | Kimbamandinidravy | | CUNONIACEAE | Weinmannia | rutembergii Eng | Lalombary | | Weinmannia | rhodoxylon Tul | Lalo be ravy | | CYATHEACEAE | Cyathea | sp. | Lohanapa | | CYPERACEAE | Scleria | arbotiva | Vendra | | EBENACEAE | Dyospiros | gracilipes Hiern | Sily mainty | | ERICACEAE | Agarista | salicifolia Hooker | Kavodia | | ERYTHROXYLACEAE | Erythroxylum | | sp. | | | | Erythroxylum | | ferrugeneum Cav | | Malambovony | | Erythroxylum | | nitidilum Bak | | Menahihy | | EUPHORBIACEAE | Amyrea | | humbertii | | Hazonovy | | Croton | | mongue | | Sily fotsy | | Croton | | glomeratus Aug D.C | | | | Drypetes | | madagascariensis | | | | Drypetes | | Madagascariensis (Lam) Humbert et Leandri | | Lomaka | | Drypetes | | perrieri Leandri | | Lomaka ravintsakay | | Drypetes | | madagascariensis (Lam) Humbert Leandri | et | Mariravy | | Euphorbia | | tetraptera Baker | | Ramanjavona | | Macaranga | | alnifolia Baker | | Mokaranana | | Macaranga | | gralata Mc Pherson | | Tarambito | | Uapaca | | thouarsii | | Mera | | FABACEAE | Albizia | | fastigiata (E Meyer) Oliver | | Volomborona | | Dalbergia | | sp. | | Voamboana | | HYPERICACEAE | Harongana | | madagascariensis | | Harongana | | ICACINACEAE | Cassinopsis | | madagascariensis | | Hazomafaitra | | Cassinopsis | | madagascariensis H. Br | | Tsivoka be ravy | | LAURACEAE | Cryptocaria | | sp. | | Vagna | | Ocotea | | cymosa (Nees) Palacky | | | | Ocotea | | sp. | | | | Ocotea | | sp. | | Varongy | | Ravensara | | sp. | | Molaliambo | | LILIACEAE | Dianella | | ensifolia | | Vazahanakampo | | Dracaena | | reflexa Lam | | Tavola be ravy | | Dracaena | | sp. | | Tavola k ravy | | LOGANIACEAE | Anthocleista | | amplexicaulis Baker | | Ndendemilahy | | Anthocleista | | rhizophoboides Baker | | Ndendemilahy | | Anthocleista | | madagascariensis | | Ndendemy | | Nuxia | | capitata | | Lambinana | | MALVACEAE | Dombeya | | cf lucida Baillon | | Somanga | | MELASTOMATACEAE | Medinilla | | ibytiensis | | Kitonda | | Memecylon | | aff longipetalum H.Perr | | Tomenja | | MELIACEAE | Malleastrum | | sp. | | | | Malleastrum | | madagascariensis | | | | MENISPERMACEAE | Burasaria | | madagascariensis A.D.C. | | Masoposahina | | MONIMIACEAE | Ephippiandra | | madagascariensis (Dangay) Lorente | | Ambora mavo | | Tambourissa | | hildebrandtii Perkins | | Ambora | | Tambourissa | | religiosa | | Kidehodeho | | MORACEAE | Ampalis | sp. | Ampaly | | Ficus | soroceoides | Kivozy | | MYRSINACEAE | Oncostemum | laurifolium Mez | Farampandoha | | Oncostemum | falcifolium Mez | Farampandoha lahy | | Oncostemum | sp. | Farampandoha mavo | | Oncostemum | bothryoides Baker | Kitohitohy | | Oncostemum | cf reticulatum Mez | Kitohitohy | | Oncostemum | sp. | Kitohitohy | | MYRTACEAE | Eugenia | bernieri Baill ex Drake | Rotra | | Eugenia | jambolana Lam | Rotra be ravy | | Eugenia | jambolana Lam | Rotra | | Syzigium | emirnense Baker | Rotra mandinidravy | | OLACACEAE | Olax | sp. | Afoatsinanana | | OLEACEAE | Norhonia | cf leandriana H Perr | Tsilaitra | | Norhonia | sp. | Tsilaitra | | PANDANACEAE | Pandanus | sp. | Vakoa | | Pandanus | sp. | Vakoa mainty | | Pandanus | sp. | Vakoa tsirika | | PROTEACEAE | Dilobeia | thouarsii Roem et Schult | Hazovao | | RUBIACEAE | Bertiera | sp. | | | Bremeria | sp. | Fatoralahy | | Canthium | humbertianum Cavaco | Fatsikahitra | | Captosperana | chevatum | Tsivoka | | Gaertnera | obovata subsp sphaerocarpa | | | Gaertnera | obovata Baker | | | Mapouria | parkeri | Korofokorana | | Mapouria | angustifolia | Korofokorana
mandinidravy | | Peporidium | sp. | Fatsikala | | Psychotria | sp. | Voamasondrenikely | | Psychotria | madagascariensis | Voamasondrenikely mandinidravy | | Pyrostria | major | Lomaka mavo | | Tricalysia | cryptocalix Baker | Tavaza | | RUTACEAE | Vepris | fitoravina | Ampody | | Vepris | ampody H.Perr | | | SAPINDACEAE | Deinbolia | perveillei | Hazombato | | Mollinaea | retusa Radlk | Hazombato | | Tina | thouarsiana (Cambess) Capuron | Hazombato | | Tinopsis | sp. | Hazombato | | Tinopsis | dasycarpa | Lanary | | SAPOTACEAE | Labramia | ditimena H. Perr | Tarantalahy |
Annexe 4c | SCROPHULARIACEAE | Halleria | tetragona Baker | Kitoto | | | STRELIZIACEAE | Ravenala | madagascariensis | Ravenala | | TILIACEAE | Grewia | opetala | Mainty hoditra | | VACCINACEAE | Vaccinium | emirnense Hook | Aramontsy | | Annexe 4c : Liste floristique du groupe 3 | | Famille | Nom scientifique | Nom vernaculaire | | ACANTHACEAE | Ruellia cyanea | Velatra | | ANNONACEAE | Xylopia sp. | Ramiavo | | Xylopia buxifolia Baill | | | APHLOIACEAE | Aphloia theaformis Bennet | Fandramanana | | APOCYNACEAE | Mascarenhasia arborescens D.C. | Herotra | | Petchia madagascariensis | | | AQUIFOLIACEAE | Ilex mitis (L) Radlk | Hazondrano | | ARALIACEAE | Polyscias aculeata | Vatsila | | Polyscias repanda | Vatsila | | Polyscias ornifolia | Vatsila | | Polyscias sp. | Vatsila | | Polyscias sp. | Vatsila | | Schefflera vantsilana (Baker) Bernardii | Vatsila | | ARECACEAE | Dypsis sp1. | | | Dypsis sp2. | Fanikara | | Dypsis sp3. | Rapaka | | ASTERACEAE | Brachylaena ramiflora | Mananitra | | Vernonia delapsa (Baker) | Maranitsiatoraka | | BAMBUSACEAE | Cephalostachyrum sp. | Tsimbolo | | Nastus sp. | Voloando | | BIGNONIACEAE | Colea cf lutescens H.P | Voalava | | CELASTRACEAE | Polycardia libera O. | Miholy | | CLUSIACEAE | Calophyllum milvum RF Stevens | Vitano | | Chrysophyllum boiviniana Perr | Rehiaky | | Garcinia aphanophlebia Bak | Kimbaletaka | | Garcinia verrucosa Jum & H, Perr | Kimbamena | | Mammea madagascariensis | Kimbafotsy | | Symphonia microphilla Benth & Hook | Kimba mandinidravy | | CUNONIACEAE | Weinmannia rutembergii Eng | Lalombary | | Weinmannia rhodoxylon Tul | Lalo be ravy | | CYATHEACEAE | Cyathea sp. | Lohanapa | | CYPERACEAE | Scleria arbotiva | Vendra | | EBENACEAE | Dyospiros gracilipes Hiern | Sily mainty | | ERYTHROXYLACEAE | Erythroxylum sp. | Malambovony | | Erythroxylum nitidilum Bak | Menahihy | | EUPHORBIACEAE | Amyrea humbertii | Hazonovy | | Croton mongue | Sily fotsy | | Drypetes madagascariensis (Lam) Humbert et Leandri | Lomaka / mariravy | | Drypetes perrieri Leandri | Lomaka ravin-tsakay | | Euphorbia tetraptera Baker | Ramanjavona | | Macaranga alnifolia Baker | Mokaranana | | Macaranga gralata Mc Pherson | Tarambito | | Phyllantus castirum Willem | Hazontana | | FABACEAE | Dalbergia sp. | Voamboana | | Albizia fastigiata (E Meyer) Oliver | Volomborona | | ICACINACEAE | Cassinopsis madagascariensis | Hazomafaitra | | LAURACEAE | Cryptocaria sp. | Vagna | | Ocotea cymosa (Nees) Palacky | | | Ocotea sp1. | Varongy | | Ocotea sp2. | Varongy laka | | Ravensara sp. | Molaliambo | | LILIACEAE | Dracaena reflexa Lam | Tavola be ravy | | Dracaena sp. | Tavola | | Dianella ensifolia | Vazahanakampo | | LOGANIACEAE | Anthocleista rhizophoboides Baker | Ndendemilahy | | MALVACEAE | Grewia cuneifolia | Afopotsy | | Dombeya cf lucida Baillon | Somanga | | MELASTOMATACEAE | Memecylon aff longipetalum H.Perr | Tomenja | | MENISPERMACEAE | Burasaria madagascariensis A.D.C. | Masoposahina | | MONIMIACEAE | Ephippiandra madagascariensis (Dangay) Lorence | Ambora mavo | | Tambourissa hildebrandtii Perkins | Ambora | | Tambourissa religiosa | Kidehodeho | | MORACEAE | Ampalis sp. | Ampaly | | Ficus soroceoides | Kivozy | | MYRSINACEAE | Oncostemum laurifolium Mez | Farampandoha | | Oncostemum falcifolium Mez | Farampandoha lahy | | Oncostemum bothryoides Baker | Kitohitohy | | MYRTACEAE | Eugenia bernieri Baill ex Drake | Rotra | | Syzigium emirnense Baker | Rotra mandinidravy | | OLACACEAE | Olax sp. | Afoatsinanana | | OLEACEAE | Norhonia cf leandriana H Perr | Tsilaitra | | PANDANACEAE | Pandanus sp 1 . | Vakoa karababika | | Pandanus sp2. | Vakoa mainty | | PROTEACEAE | Dilobeia thouarsii Roem et Schult | Hazovao | | RHAMNACEAE | Bathiorhamnus louvelii | | | RUBIACEAE | Bertiera sp. | | | Canthium humbertianum Cavaco | Fatsikahitra | | Canthium sp. | Vandrika | | Captosperana chevatum | Tsivoka | | Gaertnera obovata subsp sphaerocarpa | | | Gaertnera obovata (Baker) var obovata | Tsivoka mandinidravy | | Mapouria parkeri | Korofokorana | | Peporidium sp. | Fatsikala | | Psychotria sp. | Voamasondrenikely | | Psychotria madagascariensis | Voamasondrenikely madinidravy | | Pyrostria major | Lomaka mavo | | Tricalysia cryptocalix Baker | Tavaza | | RUTACEAE | Vepris fitoravina | Ampody | | Vepris ampody H.Perr | | | SAPINDACEAE | Deinbolia pervellei (Blume) Radlk | Hazombato | | Mollinaea retusa Radlk | Hazombato | | Tinopsis dasycarpa | Lanary | | SAPOTACEAE | Labramia ditimena H. Perr | Taratalahy | | Sideroxylon. betsimisarakum (Lecomte) | Nato | | STERCULIACEAE | Dombeya sp. | Hafopotsy | | Tina thouarsiana (Cambess) Caparon | Hazombato | | STRELIZIACEAE | Ravenala madagascariensis | Ravenala | | TILIACEAE | Grewia opetala | Mainty hoditra | | indet 4 | Soanata |
Annexe 5 Annexe 5 : Superficie (ha) et évolutions (%) des savanes incluses inventoriées
| savanes incluses | 1937 | 1957 | 1991 | 2006 | rapport entre
1957 et
1991 (%) | rapport entre 1991 et 2006 (%) | | A1 | 0,00 | 0,36 | 0,23 | 0,10 | -36,11% | -56,52% | | A2 | 0,00 | 3,38 | 1,76 | 0,57 | -47,93% | -67,61% | | A3 | 0,00 | 0,30 | 1,50 | 1,40 | +400% | -6,67 | | A4 (aménagée en riziere) | 0,00 | 0,25 | 0,23 | 0,40 | -8% | +73,91% | | A5 | 0,00 | 0,20 | 0,48 | 0,45 | +140% | -6,25% | | A6 | 0,00 | 2,20 | 3,00 | 1,82 | +36,36% | -39,33% | | A7 | 0,00 | 5,86 | 8,16 | 7,26 | +39,25% | -11,03% | | A8+A9+A10 | 0,00 | 8,49 | 14,02 | 11,02 | +65,14% | -21,40% | | A11 | 0,00 | 0,71 | 1,19 | 1,09 | +67,61% | -8,40% | | A12 | 0,00 | 13,66 | 12,84 | 9,93 | -6% | -22,66% | | A13 | 0,00 | 2,57 | 2,11 | 1,42 | -17,90% | -32,70 | | campement | 0,00 | 11,67 | 12,95 | 14,71 | +10,97% | +13,59 |
Auteur : Andry RANDRIANARISON Sujet : « DYNAMIQUES DES ECOSYSTEMES FORESTIERS EN CONTACT AVEC LES SAVANES DANS LE CORRIDOR FORESTIER DE FIANARANTSOA » A Madagascar, le paysage est formé surtout par les écosystèmes forestiers et savanicoles avec une prédominance des écosystèmes savanicoles ; ces savanes existent surtout sur le moyen ouest, ouest et sud-ouest mais des savanes exceptionnelles sont localisées au coeur même d'un massif forestier. Ce présent travail, effectué dans le corridor de Ranomafana-Andringitra (Fianarantsoa -Madagascar), a pour but d'apporter des éléments d'explications à la présence et à l'évolution de ces savanes incluses. Pour atteindre cet objectif, on a eu recours à la méthode de Gautier, de placeau, et on a creusé des fosses pédologiques, pour caractériser la structure verticale et horizontale de la formation forestière ainsi que les caractères du sol. Le traitement statistique (AFC, ACP, ANOVA) de ces paramètres permet de typer les différentes formations forestières. Des études cartographiques sur la base d'une carte topographique de 1940, des photos aériennes de 1957 et de 1991, une image Google-Earth 2004, combinées avec les enquêtes, ont été effectuées pour dater et suivre l'évolution des savanes incluses. Ces démarches ont montré que les savanes incluses sont apparues dans le période 1937-1957. Depuis leur apparition, ces savanes ont évolué selon trois types de tendance (type progressif, stable, régressif). L'apparition et l'évolution de ces savanes incluses sont conditionnées par les activités humaines (exploitation massive, tevy, feu de pâturage) sur la végétation forestière et herbeuse. Ainsi, on a pu dégager que la forêt n'est plus primaire mais secondaire, et on a pu déterminer trois types de groupements forestiers : forêt à Dalbergia sp.et à Polyscia ornifolia, forêt à Erythroxylum nitidilum et à Amerya humbertii, forêt à Ficus soroceoides et à Erythroxylum nitidilum. La forêt à Ficus soroceoides et à Erythroxylum nitidilum est le type de forêt qui est en contact avec les savanes incluses. Selon l'intensité des pressions humaines pesant sur les savanes, la forêt en contact connaît des évolutions régressive ou progressive. Pour conclure, l'homme est responsable de l'avancement ou le recul de la forêt, dont le maintien reste précaire du fait de ses activités. Mots clés : Corridor forestier de Fianarantsoa, forêt, savanes incluses, dynamique, pâturage, formation végétale, Sahabe, Ambohimahamasina, Madagascar. Encadreurs : Pr Vonjison RAKOTOARIMANANA
Dr Samuel RAZANAKA Author: Andry RANDRIANARISON Subject: «DYNAMIC OF THE FOREST ECOSYSTEMS IN CONTACT WITH THE SAVANNAS IN THE FOREST CORRIDOR OF FIANARANTSOA» To Madagascar, the landscape is especially formed by the forest ecosystems and savanicoles with a predominance of the ecosystems savanicoles; these savannas especially exist on the west, west and southwesterly means but some exceptional savannas are localized at the very heart of a forest massif. This present work, done in the corridor of RanomafanaAndringitra (Fianarantsoa - Madagascar), has goal to bring elements of explanations to the presence and to the evolution of these included savannas. To reach this objective, one had resort to the method of Gautier, of placeau and one did pedological pits to characterize the vertical and horizontal structure of the forest formation as well as the characters of soil and in order to extract the parameters of it for the statistical treatment (AFC, ACP, and ANOVA) to mark the different forest formations. Of the cartographic studies on a topographic card of 1940, of the aerial photos of 1957 and 1991, and an image Google-Earths 2004, combined with the investigations, have been done to date and to follow the evolution of the included savannas. These adopted steps showed that the included savannas appeared in the period 1937-1957. Since their apparitions, these savannas evolved according to three types of tendencies (progressive, steady, regressive type). The apparition and the evolution of these included savannas are conditioned by the human activities (massive exploitation, tevy, fire of grazing) on the forest and grassy vegetation. Thus, one was able to clear that the forest is not more primary but secondary and one could determine three types of forest groupings: forest with Dalbergia sp. and Polyscia ornifolia, forest with Erythroxylum nitidilum and Amerya humbertii, forest with Ficus soroceoides and Erythroxylum nitidilum. The forest with Ficus soroceoides and Erythroxylum nitidilum is the type of forest that is in contact with the included savannas. According to the intensity of the human pressures weighing on the savannas, the forest in contact knows some evolutions regressive or progressive. To conclude, the man is responsible for the advancement or the receding of the forest, the forest maintenance remained precarious because of his activities. Key words: Forest corridor of Fianarantsoa, forest, included savannas, dynamic, grazing, vegetation, Sahabe, Ambohimahamasina, Madagascar. Advisors: Pr Vonjison RAKOTOARIMANANA Dr Samuel RAZANAKA |