Introduction
L'évolution de la biosphère au cours de
l'histoire de la Terre n'est pas régulière. A certaines
époques la biodiversité fût réduite de façon
importante par des extinctions de nombreux groupes, appelés :
crises biologiques.
Ces crises peuvent être classées, selon qu'elles
affectent les familles, les genres et les espèces, cela permet de
distinguer : - les crises majeures au nombre de cinq, (illustrées
par la courbe des familles d'animaux marins fig 0), - les crises
intermédiaires touchant la diversité spécifique et
générique mais aussi quelques familles ou groupes, - les crises
mineures qui se manifestent dans le milieu marin par des disparitions de genres
et d'espèces (figure 1).
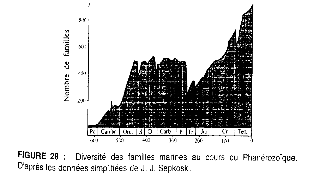
Fig 0 : Diversité des familles marines au cours du
phanérozoïque.
(D'après les données simplifiées de
J.J.Sepkoski in Lethiers 1998).
Figure 1 : Principales crises biologiques au cours du
Phanérozoïque.
(La longueur des flèches est proportionnelle à
l'intensité des extinctions, temps en MA)( D'après Sepkoski,
in F.Lethiers 1998)
L'hypothèse émise en 1984 par Raup et Sepkoski,
selon laquelle les crises biologiques ont une cyclicité, à
intéressé la communauté scientifique, puisqu`elle
suggère que les crises résultent de mécanismes, c'est
à dire d'événements récurrents. Cela explique
l'intérêt de comparer les différentes crises du point de
vue de leurs caractères paléontologiques,
sédimentologiques et géochimiques. On pourra ainsi expliquer les
mécanismes en causes.
Dans notre cas, nous serons intéressés par la
crise intermédiaire de la limite du Cénomanien-Turonien
(CT) et celle mineure du Toarcien inférieur (Ti), qui
n'échappent évidemment pas à ce principe.
Le choix de comparer, deux crises de degré
différent, peut dans un premier temps surprendre, mais il s'avère
en réalité judicieux puisque les conclusions obtenues ne
s'appliqueront pas essentiellement à un degré de crise mais
seront plus orientées vers l'ensemble des crises.
De plus, le choix de ces deux crises ne s'est pas fait de
façon aléatoire, elles possèdent un point commun
concernant un événement important puisqu'elles sont toutes les
deux caractérisées par un phénomène d'anoxie. Par
l'étude de ces deux crises géologiques de degré
différent datant respectivement de -92 millions d'années et de
-180 millions d'années, nous verrons les principes ainsi que les
méthodes utilisées, l'évolution comparée de la
biosphère puis les causes potentiellement néfastes à
l'origine des crises.
I Principes et méthodes utilisés
Deux principes éminemment simples, qui ont
été utilisés pour évaluer la santé de
l'écosystème, sont de mesurer la diversité et l'abondance
des individus. En effet, une réduction de la diversité ou une
variation de l'abondance d'individus de différents groupes en grande
expansion témoignent d'une perturbation écologique et donc
d'un stress. Et ainsi une forte diversité et une relative constance de
l'abondance soulignent de confortables conditions écologiques.
La biostratigraphie est fréquemment utilisée.
Elle permet par une analyse de la répartition des espèces dans
les strates sédimentaires, de repérer les coupures dans la pile
sédimentaire. Elle permet ainsi d'obtenir une base de datation fine. Il
est très important de posséder une bonne définition du
cadre chronologique pour pouvoir analyser les rythmes et les tendances des
changements évolutifs.
Pour ces deux crises, citons celles des ammonites (figure2)
qui a été pour cette étude la principale utilisée,
puisqu'elle présente certains avantages comme une large expansion
géographique. Cependant la géochronologie a été
réalisée en supposant une durée égale des zones
à ammonites, à cause de la petite quantité d'horizon que
l'on peut dater. Il en résulte forcement des simplifications et donc
des approximations. Cela peut aboutir, suite à des extrapolations,
à des chronologies différentes avec des périodes
temporelles variables. Mais également à des fluctuations de la
longueur temporelle de la zone d'ammonite selon la chronologie utilisée.
Ces éléments soulignent la difficulté pour obtenir la
datation qui correspond le plus à la réalité.
Figure 2 : A gauche, la chronologie et la biostratigraphie
des ammonites employées pour Ti (d'après Moses 1995).
A droite, la chronologie et la biostratigraphie des ammonites
employées pour l'intervalle autour de la limite C-T (d'après
Kauffman 1995)
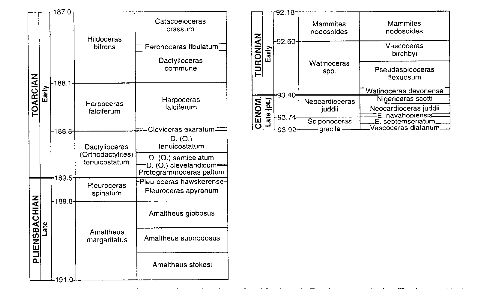
Figure 2 : A gauche, la chronologie et la
biostratigraphie des ammonites employées pour Ti (d'après Moses
1995).
A droite, la chronologie et la biostratigraphie des ammonites
employées pour l'intervalle autour de la limite C-T (d'après
Kauffman 1995).
Une des solutions utilisée est basée sur une
analyse des taux d'accumulation de roche à court terme et sur des
évaluations sur la durée des interruptions. Ces faits supposent
des approximations, tels que les taux de concrétion qui doivent
êtres connus etc.....
Cette méthode est employée par Harries et Little
pour ces deux crises. On supposera que les zones d'ammonites ont des
durées égales.
Une des premières difficultés consiste à
situer très précisément les limites de la crise.
Celle-ci a été effectuée sur des
compilations d'un champ de données globales familiales et
génériques.
Pour étudier les crises, il a été
judicieux d'établir différents intervalles :
- Intervalle d'extinction lorsque le taux
d'extinction est supérieur au taux d'apparition.
- Intervalle de survie lorsque le taux d'extinction
est environ égal au taux d'apparition.
- Intervalle de reconquête quand le taux
d'extinction est inférieur au taux d'apparition.
Dans un souci de clarté les taxons ont
été subdivisés en différentes
catégories :
- Espèces éteintes.
- Espèces survivantes à
l'intérieur de laquelle on distingue les nouvelles espèces
provenant de lignées survivantes et les nouvelles espèces
provenant de lignées ayant nouvellement évolué.
L'étude de paramètres géochimiques
permet, tel que la mesure du delta C13 lorsqu'elle présente des
variations, de montrer des perturbations dans l'environnement. Le fait d'avoir
utilisé le même matériel biologique et de regarder les
modifications relatives qu'il subit pour ces deux crises permet de
réaliser une comparaison rigoureuse. Cependant étant donné
que celles-ci ont souvent une différence de durée, ce sont les
types de changement très représentatifs de
l'événement d'extinction pour des critères,
telle que la diversité, qui ont été observés.
II Données sur la Biosphère lors de la
crise du Toarcien inférieur et celle du Cénomanien-Turonien
Tous les principes exposés précédemment
s'appliquent donc entre la crise Ti et la crise CT (qui possède une
limite difficile à définir mais qui semble se situer à la
limite CT).
Les données qui ont été choisies par ces
scientifiques viennent d'un nombre de sections qui représente des
éléments plus solides que sur simplement une section, où
des changements locaux ne peuvent pas être différenciés des
événements plus répandus. Notons que les données de
CT sont encore plus significatives puisqu'elles viennent d'un nombre de
sections plus important. Cependant il serait intéressant de produire des
barres d'erreurs significatives sur les données mais cela est impossible
puisque celles-ci ont été collectées, compilées et
comparées entre de nombreuses localités différentes,
d'où la somme d'erreurs inhérente en un point varie
considérablement.
Pour étudier ces deux crises qui n'ont affecté
que le milieu marin, un matériel commun a été
utilisé :
- Les ammonites.
- Les bivalves benthiques.
- Les bivalves semis-enfouis.
- Les bivalves suspensivores.
- Les gastéropodes.
- Des taxons rares tels que les scaphopodes, brachiopodes
inarticulés des serpumides.
(Les données fournies pour la crise Ti et CT viennent de
Paleogeography, Paleoclimatology, Paleoecology 154 (1999) P 39-66) Fig 3 et
4.

Figure 3 : Stratigraphie concernant le Ti.
(Paleogeography, Paleoclimatology, Paleoecology 154 (1999))
A) Données sur la crise Toarcien
inférieur.
La Localisation des sections étudiée se trouve
dans le Nord-ouest de l'Europe en deux endroits, les sections
côtières du bassin de Cliveland, du Nord du Yorkshire
(Angleterre) et dans le sud du bassin allemand Baden wür hemberg
(figure 5)
Figure 5: Localités du Nord Ouest de l'Europe
utilisées pour examiner l'extinction du ET, les données
proviennent de deux localités en Allemagne et en Angleterre. (Little 96)
(Paleogeography, Paleoclimatology, Paleoecology 154 (1999) P 43)
L'intensité de l'extinction a touché 8% des
familles, 26% des genres et d'après des estimations 33 à 53% des
espèces.
La figure 3 montre une disparition de 90% de tous les taxons.
La sous-division de l'intensité d'extinction dans les groupes, nous
montre un impact dévastateur de celle-ci au moins sur le biotope de
l'Europe du Nord.
Certains groupes ont été particulièrement
touchés citons : les ammonites, les bélemmites, les bivalves
se nourrissant de fouisseurs en suspensions ou déposés,
gastéropodes et brachiopodes. Seul certains d'entre eux possèdent
des survivants au niveau des espèces : il s'agit des bivalves
benthiques, des crinoïdes et des taxons rares avec respectivement 78%, 50%
et 0% d'extinction. Cela montre un impact important malgré le nombre de
nouvelles espèces dérivant des lignées survivantes ou des
réfugiées. Quand tous les bivalves benthiques sont inclus,
la diversité de ce groupe diminue seulement de 24%, beaucoup moins que
pour les autres familles benthiques.
Les céphalopodes augmentent leurs dominances relatives,
alors que la grande majorité des autres espèces subissent des
réductions. Les données montrent un changement dans la
distribution des taxons qui correspond à l'horizon des disparitions.
Avant la crise, nous pouvons constater que la distribution de la
diversité entre les différents taxons est équitablement
répartie. Suivant les extinctions, la domination déplace surtout
les ammonites et les bivalves fouisseurs. Cela tend à montrer une
modification de la structure écologique.
La distribution des taxons présente un changement entre
« l'intervalle de fond » de la crise et les intervalles de
repopulations fig6.
La diversité cumulative est relativement constante
entre avant et après les intervalles d'extinctions, dans un champ de 59
à 63 taxons (fig 6A). A certains points, cela semble être le
reflet de la durée plus longue de l'intervalle de repopulation et celle
plus courte de l'intervalle d'extinction. La distribution de la
diversité entre l'intervalle de fond et les intervalles d'extinctions
montre que la plupart des taxons ont disparu lors de cette dernière (fg
6B). Les céphalopodes se diversifient passant de 14 espèces
pendant « l'intervalle de fond » à 43 taxons durant
le début de la repopulation.
Cette tendance est minorée dans les deux groupes de
céphalopodes qui augmentent de 10 à 24 taxons (les ammonites) et
de 4 à 17 taxons (pour les bélemmites). Les autres groupes, la
plupart benthiques voient leurs diversités atteindre un taux
supérieur à 50% fig 6A.
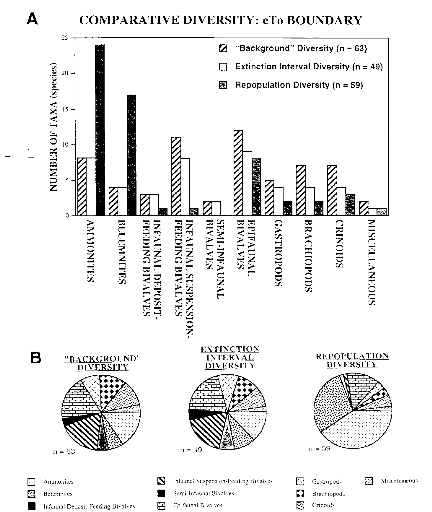
Figure 6: A) Diversité comparée de quelques taxons
pour la crise CT
B) Représentation de la structure
des taxons pour CT
(Paleogeography,
Paleoclimatology, Paleoecology 154 (1999) P 54)
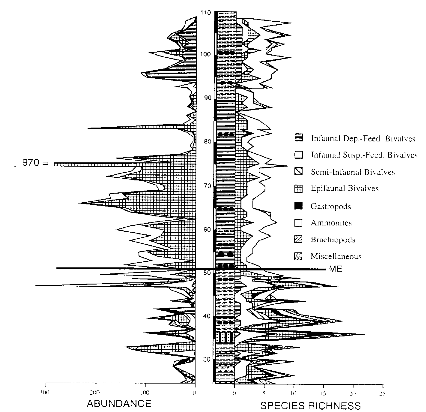
L'intervalle de pré-extinction de la faune est
caractérisé par une diversité relativement
élevée, avec des valeurs moyennes de 8 espèces (fig7). De
plus la diversité est distribuée équitablement entre les
différents groupes des taxons.
Figure 7 : Abondances et tendances de la diversité
pour ET pour la section du Saltwick Nab. (Paleogeography, Paleoclimatology,
Paleoecology 154 (1999) P 57)
L'intervalle de survie, qui est
marqué par une diminution de la diversité, a des valeurs moyennes
chutant à 4.2 et une domination des bivalves benthiques et des
ammonites.
L'intervalle de reconquête voit la diversité
augmenter jusqu'à 5.5 (grâce notamment à la
réapparition des brachiopodes et des bivalves se nourrissant des
benthiques déposés).
Pour l'intervalle de repopulation la répartition se
fait ainsi : 43% pour les espèces nouvelles provenant de
lignées ayant nouvellement évolué, 41% pour les
espèces nouvelles provenant de lignées ayant survécu et
16% de survivants, (figure 8).
Les céphalopodes, les ammonites et les bellemmites
représentent 93% des espèces nouvelles provenant de
lignées ayant nouvellement évolué. Le taux d'extinction de
certains de ces groupes pendant l'intervalle de fond et l'intervalle
d'extinction, suggère une continuation de leur modèle
évolutif. Ils possèdent des évolutions différentes
des autres groupes, au moins à l'intérieur de l'Europe de
l'Ouest. Les organismes pélagiques (tels que les ammonites) sont
florissants dans ces habitats. Ces lieux sont peu ou pas affectés par
les mécanismes créant l'extinction dans le domaine benthique.
Les survivants, comme les bivalves benthiques, ont un taux
d'évolution plus bas que les ammonites. Les autres groupes, tels que les
bivalves fouisseurs, gastéropodes, brachiopodes contiennent uniquement
des espèces nouvelles provenant de lignées ayant survécu.
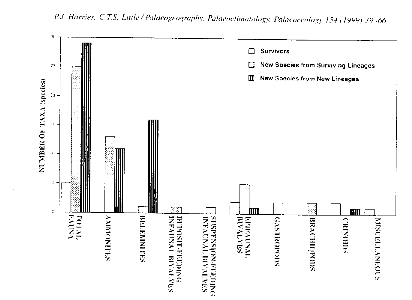
Figure 8 : Histogramme montrant la distribution de la
repopulation des taxons pour ET pour les espèces éteintes, les
espèces survivantes, les nouvelles espèces provenant de
lignées ayant survécu et les nouvelles espèces qui ont
évolué. (Paleogeography, Paleoclimatology, Paleoecology 154
(1999) P 59)
L'événement du Ti a vu également un
grand changement chez les radiolaires, en effet, les formes
caractéristiques du Trias terminal et du Jurassique
« basal » sont remplacées par des taxons
caractéristiques du Jurassique supérieur et du
Crétacé inférieur. Notons également une
modification dans le plancton du Ti (Rie S.Hori, 1996)
B) Données sur la crise
Cénomanien-Turonien
Figure 9: Localités du Bassin intérieur de
l'Ouest en
Amérique du Nord, utilisées pour examiner
l'événement d'extinction CT.( Paleogeography, Paleoclimatology,
Paleoecology 154 (1999) P 43).
La Localisation des sections étudiées se trouve
dans le Nord de l'Amérique dans le bassin intérieur Ouest, citons
Pueblo Collorado, figure 9.
L'intensité de l'extinction a touché 8% des
familles, 26% des genres et d'après une estimation 33 à 53% des
espèces.
Cette extinction atteint 79% des espèces macro
invertébrées à l'intérieur du bassin
intérieur de l'ouest en Amérique du Nord (WIB) (figure 4).
L'extinction au niveau des espèces est de 93% pour les
ammonites, 75% pour les bivalves inocéramides, 69% pour les autres
bivalves benthiques, 90% pour les bivalves semi-fouisseurs, 68% pour les
bivalves fouisseurs, 84% pour les gastéropodes, 73% pour les taxons
miscelleux.
Comme pour la crise Ti, les céphalopodes augmentent
leurs dominances relatives, figure 10, alors que la grande majorité des
autres groupes subit des réductions.
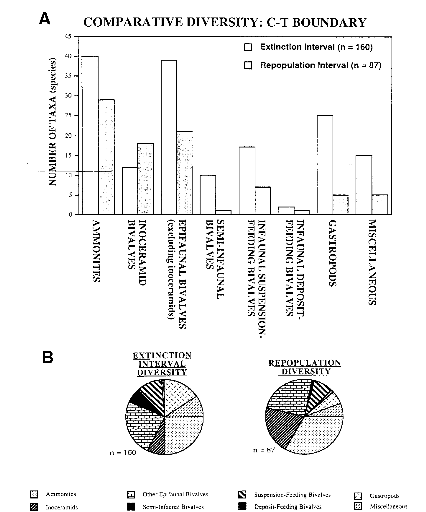
Figure 10:A) Diversité comparée de quelques
taxons pour la crise CT
B) Représentation de la structure des
taxons pour CT
(Paleogeography, Paleoclimatology, Paleoecology 154 (1999) P
55)
Pour l'intervalle de repopulation la répartition se
fait ainsi : 39% concernent les espèces nouvelles provenant de
lignées ayant survécu, 36% sont des survivants, 23% des
espèces nouvelles provenant de lignées ayant nouvellement
évolué (la majorité sont des céphalopodes avec les
ammonites) et 2% des ancêtres de la crise (qui servent de réserve
pour les premières radiations qui sont limitées aux
inocéramides), la figure 11 montre cette répartition.
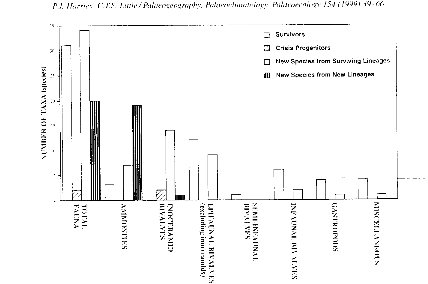
Figure 11 : Histogramme montrant la distribution de la
repopulation des taxons pour CT pour les espèces éteintes, les
espèces survivantes, les nouvelles espèces provenant de
lignées ayant survécu et les nouvelles espèces qui ont
évolué. (Paleogeography, Paleoclimatology, Paleoecology 154
(1999) P 60)
Certains bivalves, comme les rudistes furent
particulièrement affectés. En moins d'un million d'années,
10 genres de rudistes sur 17 disparaissent, 8 s'effacent pour
« réapparaître » que deux millions
d'années plus tard. Cette extinction frappe massivement les rudistes
à aragonite dominante, tels que les caprotinidaes et les
caprinidaes, tandis que les rudistes à calcite dominant tels que
les Radiolitidaes ne sont pas affectés. Notons également
que les foraminiféres benthiques sont également touchés
par cette crise puisque trois espèces sur 21 passent cette limite
(figure 12).
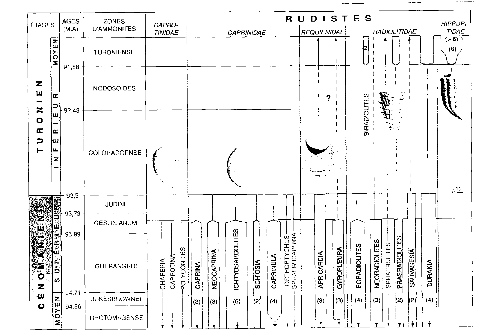
Figure 12 : La crise des rudistes lors de la limite CT dure
2 millions d'années. La communauté s'est épanouie de
nouveau au Turonien moyen, mais après un renouvellement important des
familles qui la composait.(le nombre d'espèces est entre
parenthèses) « Pour la science no 28 P90 »
Lors de cette crise, des organismes de taille bien plus
volumineux furent affectés tels que les ichthyosaures. Ce
groupe de reptiles retourné à la vie aquatique marine disparu
lors de cette crise. Les ichthyosaures qui sont apparus au Trias
inférieur, se nourrissaient entre autre de bélemmites en rejetant
le rostre. Ils furent probablement victimes de leurs trop grandes adaptations
morphologiques, physiologiques alors que d'autres reptiles aquatiques moins
spécialisés tels que les Mosasaures continuaient au-delà
de cette crise. (figure 13)
Figure 13: Reconstitution d'un ichthysaure
Evolution de la biosphére et événements
géologiques P298, F.Lethiers
L'extinction du CT a le même modèle que celle du
Ti, en effet, le début « énergétique »
de l'extinction est assez élevé (4.4 espèces pour la
première moitié de l'intervalle d'extinction) puis
équitablement distribué figure 14.
Figure 14 :Abondances et tendances de la diversité
pour CT. (Paleogeography, Paleoclimatology, Paleoecology 154 (1999) P 58)
Figure 9 : Abondance et tendance de la
diversité pour CT
Paleogeography, Paleoclimatology, Paleoecology 154 (1999) P
58)
Figure 9 : Abondance et tendances de la diversité
pour CT pour la zone du Pueblo. Paleogeography, Paleoclimatology, Paleoecology
154 (1999) P 58)
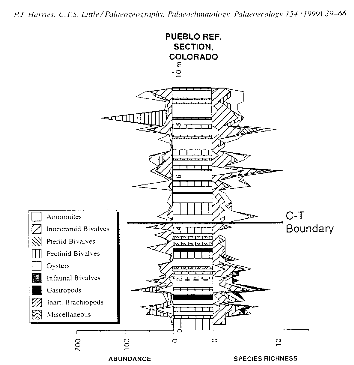
Tant que cet intervalle d'extinction progresse, la
diversité diminue à une moyenne de 2.3 espèces ; la
faune est dominée par les bivalves benthiques, spécialement les
inocéramides. Dans l'intervalle d'extinction, la diversité
augmente peu jusqu'à 2,4 espèces ; les bivalves
épifaunes continuent à dominer avec les inocéramides et
les huîtres. Quelques huîtres, telles que Pseudoperna
bentorensis étaient dépendantes des valves
inocéramides qu'ils utilisaient comme substrat.
Notons l'extinction et la radiation observées chez les
radiolaires pour le CT (Erbacher et Thurow, 1996)
C) Bilan
Notons que les données de la crise Ti ne concernent
qu`une partie de l'intervalle de reconquête. Cela entraîne des
conclusions limitées. Cependant des tendances peuvent être
soulignées. En effet, les données montrent une augmentation de la
diversité et également un changement concernant la dominance.
Ce changement montre une amélioration des conditions
avec notamment une grande source de nourriture riche en éléments
organiques à l'origine des dépôts noirs argileux.
L'intervalle de reconquête du pré-CT indique une
hausse de la diversité et de l'abondance sauf chez les bivalves
benthiques. Cela s'explique probablement par le fait que les bivalves
benthiques représentent une portion très faible de la faune
pendant cet intervalle de fond.
Les pulsations suivant directement les frontières des
extinctions pourraient simplement représenter:
- Des événements évolutifs similaires
à d'autres radiations de l'intervalle de fond à
l'intérieur du groupe
- Une hausse dans les niches des colonnes d'eau
supérieures associée avec des pics de transgressions.
Une analyse des morphotypes d'ammonites du CT suggère
que les formes habitant les plus hautes zones atteintes par les colonnes d'eau
ne sont pas affectées par l'extinction de masse.
En outre, les données sur les céphalopodes
montrent que certains taxons ne sont pas sensibles aux même pressions
sélectives et évolutives que les organismes vivants dans le
domaine benthique. Cette différence s'exprime dans les taux rapides
d'évolution subit par les céphalopodes alors que certains groupes
benthiques ont une extinction marquée. Pour Ti, l'extinction principale
d'ammonites précède l'extinction en masse. Cette évolution
rapide fut favorisée par l'événement d'extinction,
créant une place vacante dans l'écosystème dans laquelle
les céphalopodes se seraient développés. En effet,
lorsqu'un taxon s'éteint, il libère une niche écologique.
Celle-ci est alors utilisée par un autre taxon, ce qui lui permet de se
« développer ».
Néanmoins aucune disparition ne semble affecter les
compétiteurs potentiels à l`intérieur du domaine
pélagique.
La réponse biotique pour les taxons marins de ces deux
événements est équivalente en général.
Les extinctions ont des valeurs très proches. La
diversité est initialement élevée et distribuée
équitablement.
Les tendances à long terme de la diversité
montrent des traits similaires pour les événements du Ti et du CT
Notons que la diversité pendant la repopulation du CT
est seulement légèrement supérieure à la
moitié de celle durant l'intervalle d'extinction, cela entraîne
que les changements dans la dominance sont relatifs plus qu'absolus.
Toutes ces données (fig4) qui peuvent paraître un
peu rébarbatives permettent de mettre en évidence quatre
principales tendances qui sont valables pour ces deux crises :
- 1er tendance : La diversité
ne varie presque pas lors de l'extinction et les intervalles de
repopulation sont relégués à « l'intervalle de
fond ». Cela tend à un grand renouvellement sauf s'il y a des
survivants. Ce model est le plus souvent réalisé dans les groupes
à renouvellements rapides, tels que les ammonites et les
bélemmites pour Ti et les inocèramides du CT fig 3 et 4.
- 2ème tendance : Elle se
caractérise par une extinction de la majorité des taxons avant la
« frontière », suivi par l'explosion
évolutive très tôt dans la repopulation. Cela est
illustré par les bivalves benthiques et les gastéropodes pour les
deux extinctions ou les brachiopodes pour Ti.
- 3ème tendance : Les groupes ont
quelques survivants mais aussi un retard évolutif avant l'apparition de
nouveaux taxons, citons les crinoïdes de Ti et les bivalves fouisseurs du
CT.
- 4éme tendance : Elle répond au
concept « espèces lazares », cela traduit la
disparition pour une période d'un groupe, qui trouvant refuge dans un
lieu, réapparaît lors de la repopulation. Cela est valable pour
les gastéropodes du Ti.
Ces deux crises présentent un intervalle de survie
possédant une diversité et une abondance basses, reflétant
la perturbation écologique de l'écosystème. Mais
également un intervalle de reconquête étendu fig 3 et 4.
Toute la repopulation peut être
considérée comme un événement important de
diversification. Il existe plusieurs formes de réponses à
l'événement : citons celle des ammonites ou des belemmnites
du Ti et celle des inocéramides du CT, qui évoluent et rayonnent
rapidement, cela suggère que les extinctions de masse
représentent, pour certains groupes, un élément de
radiation.
La réponse biotique suggère que les conditions
rapidement améliorées permettent une expansion de la
diversité et de l'abondance.
Harrie et Little ont étudié ces crises dans des
sédiments qui ont été déposés dans des mers
épicontinentales, tandis que Philip montre que le milieu récifal,
lieu de vie des rudistes, a été également affecté
par la crise CT.
L'ensemble des modèles des crises Ti et CT sont
identiques à des échelles variées. De même, les
intervalles d'extinction, de survie et de reconquête possèdent la
même ressemblance. Dans les deux cas, les organismes planctoniques et
nectiques habitant la partie supérieure de la colonne d'eau sont
protégés des événements d'extinctions. Ces groupes
ont aussi maintenu leur évolution.
Les bivalves benthiques ne furent pas affectés et
devinrent dans les deux cas les organismes dominant dans les dernières
portions de l'intervalle d'extinction et dans l'intervalle de survie.
III Evénements et preuves intervenants lors de
ces crises.
Ces deux crises Ti et CT présentent respectivement
des intervalles d'extinction de 0.6 et 0.42 millions d'années, cela
tend à montrer qu'il n'y a pas un seul événement
catastrophique commandant l'extinction.
Cela laisse supposer deux hypothèses :
- Soit un événement qui a agi sur un grand laps
de temps.
- Soit une conjonction de plusieurs causes.
Plusieurs événements se sont passés
à ces époques qui ont pu provoquer ces crises, cependant il est
très difficile de pouvoir définir la principale.
Ces deux événements arrivent juste avant et
pendant un grand pic eustatique avec une élévation du niveau
marin (transgression). Cela a entraîné lors de ces intervalles une
augmentation des routes épicontinentales marines. L'extension des mers
épicontinentales a continuellement varié au cours des temps
géologiques. Ces phénomènes affectent évidemment
les différents organismes, selon l'effet surface-biodiversité. Il
existe une relation entre la surface occupée et le nombre de taxons pour
le benthos marin, figure 15, cette augmentation de surface des mers
épicontinentales est favorable à l'augmentation de la
diversité.
Figure 15 : Illustration de l'effet
surface-biodiversité,
( évolution de la biosphère et
événements géologiques, F.Lethiers P127)
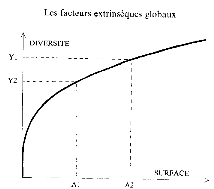
L'ensemble de ces deux crises est associé à une
anoxie très répandue et un dépôt de schiste argileux
noir.
Ces deux crises possèdent une lithologie très
proche dans le bassin environnemental :
- Ti possède le schiste argileux et des
concrétions carbonatées,
- CT des schistes argileux ou marnes et des
concrétions carbonatées.
Ces deux intervalles correspondent à un climat chaud
mais constant. De nombreux scientifiques pensent qu'il fut engendré par
un effet de serre. Ces conditions ont amené une circulation
océanique difficile. De plus la formation d'une eau chaude salée
dans la partie inférieure de l'eau met en évidence une
stratification de la mer d'où un potentiel pour l'anoxie. En effet, la
teneur en O2 dépend de la salinité et de la température
qui, lorsqu'elles augmentent, provoquent une baisse de la teneur en O2 comme
l'illustre la figure 16.

Figure 16 : Dissolution de O2 dans l'eau en fonction de
la salinité et de la température (valeurs de saturation à
l'équilibre, sous Patm normale dans l'air saturé en vapeur d'eau.
(Ivanoff 1972) Evolution de la biosphére et événements
géologiques, F.Lethiers P158
Cela affecte nombre de taxons puisque l'oxygène dissous
dans l'eau est nécessaire à la respiration d'organisme marin
même si certains, comme les bivalves benthiques, tolèrent une
carence en oxygène. La similarité dans la réponse biotique
des deux crises suggère que les événements
favorisés par l'anoxie possèdent une structure
« prévisible ».
Certaines données géochimiques, comme le delta
du Carbone13 montre qu'il existe un lien solide entre le modèle de
repopulation et les conditions environnementales. Le delta 13C positif
enregistré dans plusieurs aires peut être comparé. Ces
courbes (figure 17) s'avèrent remarquablement similaires, bien que
quelque peu compensées, et suggèrent que les perturbations
environnementales étaient au moins régionales. Le champ de
digression va de 0.38% à 0.25% selon le lieu de mesure pour Ti. Pour CT,
les digressions du delta 13C mesurées dans le bassin intérieur de
l'Ouest en Amérique du Nord sont de 0.3% en moyennes. La tendance est
similaire pour les deux crises.
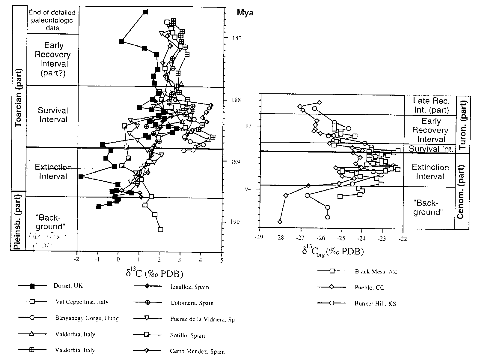
Figure 17 : Points montrant la tendance du delta C13 pour Et
et CT .Les données dans l'ancien d'après Jenkyns et Clayton
(19986-1997), Jimenez (1996) et pour le supérieur Pratt 1985.Dans les
deux cas les données ont été placées à
l'intérieur de l'encadrement temporel de la figure (celles des
ammonites).
( Paleogeography, Paleoclimatology, Paleoecology 154 (1999) P
63)
Les digressions positives du delta 13C, toutes les deux
caractéristiques des extinctions Ti et CT qui ont été
reliées à des événements anoxiques,
suggèrent que l'amplitude et la durée de l'anoxie et son stress
environnemental associé, étaient au moins un acteur significatif
dans la détermination de la réponse biotique.
Les digressions du delta 13C peuvent être
utilisées en tant qu'outil de prédiction puisque lorsqu'elles
reviennent vers des niveaux de l'intervalle de fond, elles suggèrent une
amélioration environnementale et donc que la reconquête est
initiée. Il existe deux hypothèses pour expliquer les
digressions : certains pensent qu'elles sont dues à l'augmentation
de la productivité, d'autres penchent pour la diminution du recyclage
des matières organiques, mais quelles que soient les causes de ces
digressions elles représentent un piégeage du carbone organique.
Cette préservation à l'intérieur des sédiments est
dépendante et peut-être la cause de cette anoxie.
Notons que le principal intervalle de perturbation
géochimique pour les deux événements est temporairement
similaire (0.8 et 0.65 millions d'années pour respectivement Ti et CT).
La fin de la digression du delta 13C correspond à la
fin de l'intervalle de survie, cela indique que les perturbations de
l'environnement enregistrées par le delta 13C affectent également
les biotopes. Cela semble pouvoir également s'appliquer à la
crise Crétacé Tertiaire puisque la perturbation majeure
a une très courte durée et l'intervalle de survie est de ce fait
inexistant.
Pour les données du CT l'intervalle d'extinction
initié à la première digression majeure est absente pour
Ti, cela suggère qu'il se produit une traîné à
l'arrière de l'intervalle d'extinction. Ce point est soutenu par
l'association des plus frappantes étapes d'extinction avec la partie
supérieure de l'intervalle d'extinction (figure 17).
Il existe à la limite CT (il y a 95 millions
d'années) la présence d'un taux d'iridium en Amérique du
Nord et du Sud, qui suggère l'hypothèse d'un
astroproblème. Celui-ci serait arrivé en Amérique du Nord,
dans l'Alberta à Steen River. Cet astroproblème par sa taille
relativement modeste (cratère de 25 km de diamètre) a pu jouer un
rôle dans la crise CT, mais on ne peut pas lui imputer la crise à
lui seul, en effet plusieurs astroproblémes de dimension
supérieure ne sont pas rattachés à des extinctions
significatives, pour illustrer cela citons l'impact majeur du cratère de
Montagnais, en Atlantique Nord il y a 51 millions d'années, qui ne
correspond à aucune extinction importante malgré une dimension de
45 km de diamètre (Lethiers 1997 P102). De plus, un cratère de 25
km « apparaît » tous les 1 million d'années
environ (d'après Raup (1992)).
A la limite CT il existe également de fortes anomalies
des rapports isotopiques de l'oxygène (18O/16O) et du carbone (13C/12C)
ainsi que du manganèse, mais l'avis des géochimistes diverge sur
leur interprétation. Ainsi M Kuyper pense qu'il s'est produit une baisse
de température liée à une chute en CO2
atmosphérique, d'où une baisse de l'effet de serre due à
une séquestration d'une grande quantité de matière
organique marine et continentale dans les parties profondes des océans.
Il s'appuie sur le fort enrichissement des couches en matière organique
dans ces milieux à cette époque, résultant, semble t-il
d'une stagnation des eaux profondes (Pour la science no28 P91).
Ces conclusions tirées par ce scientifique sont
diamétralement opposées à celles proposées
précédemment, cela souligne la difficulté
d'interprétation des faits.
On peut donc légitimement penser que ces crises
résultent d'une conjonction de plusieurs événements. Si
certains semblent plus importants, comme l'anoxie, ils ne peuvent pas expliquer
les crises à eux seuls.
Cependant notons l'absence de données sur l'influence
de ces crises sur la flore et sur l'aspect écologique. Cela peut
être du à trois raisons : soit une absence de données,
cela semble peu plausible; soit un choix délibéré de ne
pas en parler; soit qu'aucun changement ne s'y produit. Ces données si
elles existent auraient pu, peut être, nous apporter des
éléments qui auraient permis de trancher entre les
hypothèses contraires des scientifiques.
Quoi qu'il en soit, on peut penser que l'extinction de
certains organismes ont du influencer le développement de certains types
de flore. De plus, la disparition d'une seule catégorie
d'éléments floraux peut entraîner un
déséquilibre écologique et entrer dans une conjonction de
causes qui peut-être à l'origine d'une cascade de disparitions.
Cela nous démontre bien l'importance qu'ils revêtent.
Les similarités dans les réponses biotiques de
ces événements remettent en question les hypothèses tel
que l'événement « imprécis ». Cette
hypothèse relègue une bonne partie de l'histoire de la vie au
rôle de chance.
Néanmoins, la similarité dans les modèles
biotiques ente Ti et CT implique que si les mécanismes d'extinctions
sont similaires, la réponse biotique est prévisible.
Les événements de natures différentes
font que les lignées ne sont pas atteintes de la même façon
et avec la même intensité : cela dégage la notion de
sélectivité des crises.
Il peut être difficile de prédire les
espèces exactes qui survivraient mais pas les modèles
basés sur les groupes taxoniques. Les événements
anoxiques, très fréquents dans les crises, dévastent les
éléments fouisseurs, limitent les taxons benthiques,
spécialement gastéropodes et brachiopodes articulés. Ils
permettent la prolifération des taxons des parties supérieures
des colonnes d'eau aussi bien qu'à des taxons benthiques
spécifiques.
Conclusion
Ces crises qui semblent revenir cycliquement sont liées
le plus souvent à une sommation de plusieurs causes similaires.
Cependant les crises sont de natures différentes parce que tous les
événements n'interviennent pas à chaque fois et qu'ils se
combinent de manières différentes. Elles permettent aux
espèces survivantes et apparaissantes de se développer en
utilisant les niches écologiques laissées vacantes par les
espèces qui ont disparu. Cela permet parfois une redistribution des
espèces dominantes dans la compétition perpétuelle
qu'elles se livrent, en généralisant, nous pouvons citer les
mammifères qui grâce à la crise Crétacé
Tertiaire ont pu se développer. Les crises permettent donc à
certaines espèces bridées de prendre une place conséquente
dans la biosphère.
Bibliographie
Erbacher, Thurow, Influence of anoxic events on the evolution of
mid-Cretaceous radiolaria in the North Atlantic and Western Tethys, Marine
Micropaleontology
Harries et Little (1999) The early Toacian and the
Cenomanian-Turonian mass extinctions, Paleogeography, paleoclimatology,
paleoecology 154 P39- 66 Harries et Little
Lethiers (1998) Evolution de la biosphère et
événements géologiques, « Gordon and Breach
science Publishers »
Philip (07/2000) Une extinction dans les mers tropicales de
l'ère secondaire, Pour la Science, no 28, p89-91
Rie S.Hori (1996), The Toarcian radiolarian event in bedded
cherts from southwestern Japan, Marine Micropaleontology
Résumé
La biosphère fut au cours de son histoire plusieurs
fois bouleversée par de des extinctions plus ou moins importants. Ces
disparitions de taxons permettent, la plupart du temps, une redistribution de
la diversité. Certains groupes peuvent se développer, en
utilisant les niches écologiques ainsi libérées.
Il y a 92 millions d'années et 180 millions
d'années, des événements sont intervenus modifiant de
manière importante la biosphère marine. Cela a permis de
dégager respectivement deux crises ; la crise intermédiaire
du Cénomanien- Turonien et celle mineure du
Toarcien inférieur.
Le choix de ces deux crises s'est réalisé
à causes d'événements semblables, telle que l'anoxie.
Le fait de posséder des traits communs permet d'effectuer des
comparaisons.
Après avoir énoncé quelques principes
d'analyses et de méthodes utilisées, nous pouvons émettre
certaines « règles » qui semblent s'appliquer, c'est
à dire, se généraliser à l'ensemble des crises.
Cela permet également de dégager certaines tendances. Ces crises
géologiques, qui sont des événements cycliques, sont
liées à une sommation de causes.
Même si certaines sont plus importantes que qu'autres,
il est rare qu`une seule supporte la responsabilité de la crise.
Mots clés :
Crise, Cénomanien-Turonien, Toarcien inférieur,
extinction, principe.
| 

