4.3 Discussions
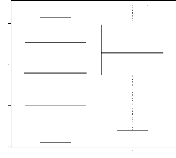
FIG. 4.21 - Corrélation entre l'apparentement
réel et l'apparentement estimé 1) dans le cas non spatial et (2)
dans le cas spatial avec 10 locus et une variance de dispersion égale
à 1
L'application pratique de notre travail a porté sur
l'étude de la diversité génétique et de la
structuration spatiale du karité dans une parcelle de jachère au
sud du Mali. Le génotype à 12 marqueurs microsatellites à
1, 30 m de hauteur de 222 arbres à karité ont été
prélevés dans une parcelle de jachère sur le site de
MPeresso. Les coordonnées géographiques de seulement certains de
ces arbres (132 arbres parmi) ont aussi été relevés. Le
nombre d'allèles par locus varie de 2 à 8 et l'indice de fixation
FIS observé varie entre --0.287 pour le locus F1 à
0.555 pour le locus G7. Le FIS global pour l'ensemble des locus est
significativement différent de 0 et ainsi la population de karité
étudiée sur le site en jachère de MPeresso est en
déséquilibre par rapport au modèle de Hardy-Weinberg. La
structuration spatiale a été étudiée aussi bien au
niveau phénotypique qu'au niveau génotypique. Au niveau
phénotypique, nous avons étudié la distribution des arbres
par classe de diamètre. Cette distribution observée
présente deux modes : le premier correspond aux jeunes arbres en
régénération et le second aux arbres déjà
présents lorsque la parcelle était encore en culture. Nous avons
effectué une analyse en composantes principales pour étudier
l'association entre les génotypes aux différents locus; nous
avons ainsi pu distinguer l'association des locus F5 et E6, d'une
|
0.80 0.85 0.70 0.75 0.80
|
|
|
1 2
|
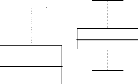
FIG. 4.22 - Corrélation entre l'apparentement
réel et l'apparentement estimé (1) dans le cas non spatial et (2)
dans le cas spatial avec 10 locus et une variance de dispersion égale
à 10

1 2
0.85 0.70 0.75
FIG. 4.23 - Corrélation entre l'apparentement
réel et l'apparentement estimé (1) dans le cas non spatial et (2)
dans le cas spatial avec 10 locus et une variance de dispersion égale
à 100
part, et celle des locus D10 et D6 d'autre part; cependant le
pourcentage d'inertie totale expliquée par les deux premiers axes
principaux est relativement faible (à peine 20%). Aussi la
réalisation du test de Mantel ne nous permet pas de conclure à
l'existence d'une association significative entre les distances
génotypiques et les distances spatiales. Cependant, il ressort du test
de Moran que la structuration spatiale du karité est
agrégée à faible et grande distances. La structuration
spatiale à grande distance pourrait être dûe aux
activités humaines. En effet, la structuration à grande distance
en jachère pourrait s'expliquer par les pratiques des populations
locales, qui entraînent une dispersion une dispersion des graines et du
pollen. Aussi, les modèles théoriques prédisent
l'existence d'une structure génétique spatiale lorsque le flux de
gènes est restreint localement et une absence de structure spatiale
lorsque le flux de gènes par les graines est extensif (Hardesty et al.,
2005). L'apparentement moyen entre les individus en fonction des classes de
distance a été déterminé selon deux méthodes
d'estimation fondées sur le calcul des moments (celle de Wang (2002) et
celle de Lynch et Ritland (1999)) et aussi selon la méthode du maximum
de vraisemblance de Milligan (2003). L'apparentent moyen décroît
significativement avec le logarithme de la distance. Mais, d'une manière
globale, l'apparentement moyen de l'échantillon
1 2
0.70 0.75 0.80 0.85
FIG. 4.24 - Corrélation entre l'apparentement
réel et l'apparentement estimé (1) dans le cas non spatial et (2)
dans le cas spatial avec 15 locus et une variance de dispersion égale
à 0.1

1 2
0.70 0.75 0.80 0.85
FIG. 4.25 - Corrélation entre l'apparentement
réel et l'apparentement estimé (1) dans le cas non spatial et (2)
dans le cas spatial avec 15 locus et une variance de dispersion égale
à 1
étudié est très faible. Nous avons aussi
noté l'existence d'une autocorrélation spatiale significative
jusqu'à une distance d'environ 15m. Cependant, le test de Mantel ne nous
permet pas de déceler une relation linéaire entre la distance
génétique et la distance géographique. Kelly et al.
(2004b) ont étudié la structuration spatiale du karité au
sud du Mali en comparant les résultats obtenus en jachère et en
forêt. Il ressort de leurs travaux que la structuration spatiale est plus
prononcée en jachère qu'en forêt. Parmi les facteurs
expliquant cela, nous pouvons relever le niveau de fructification plus
important dans la jachère dû à une moindre
compétition entre les arbres, ce qui favorise la
régénération naturelle des plants juvéniles autour
des arbres mères du fait que les graines de Vitelleria Paradoxa sont
principalement dispersées par la gravité. En forêt par
contre, le plus faible niveau de fructification qui est dû à une
compétition plus forte entre les arbres, réduit le nombre
d'arbres autour de l'arbre mère et affecte en conséquence la
structuration spatiale du Karité.
Nous avons effectué quelques simulations pour valider
le modèle spatial pour l'estimation de l'apparentement. Nous notons que
la prise en compte de l'information spatiale améliore sensiblement
l'estimation de l'apparentement génétique. Ceci est surtout
vérifié lorsque la variance de dispersion des juvéniles
autour du pied mère est faible, c'est à dire lorsque
l'hypothèse que
0.70 0.75 0.80 0.85

1 2
FIG. 4.26 - Corrélation entre l'apparentement
réel et l'apparentement estimé (1) dans le cas non spatial et (2)
dans le cas spatial avec 15 locus et une variance de dispersion égale
à 10
des individus plutôt proches spatialement sont proches
génétiquement est vérifiée.
Frequency
0 100 200 300 400 500 600
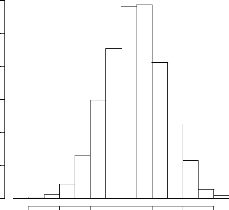
-0.007 -0.006 -0.005 -0.004 -0.003 -0.002
-0.001
IBDSpat$Bet[-c(1:100)]
FIG. 4.27 - Distribution du paramètre í
associé à la distance dans le modèle spatial pour
l'apparentement
| 

