|
|
MALES
logie et Gestion des Monogastriques
|
|
on des performances ENT DE de reproduction ET
des
es croisé
Mars 2011 par : Sous la direction de
:
eMa

JURY
Médecine Vétérinaire
Dr Mahamadou DAOUDA, Enseignant à la FS
Année académique
2010-2011

DEDICACE

A Dieu TOUT-PUISSANT, Grand Maître
de
l'univers et de la sagesse.
Pour m'avoir donné la santé, le
courage
dans les moments de grand désespoir et
par dessus tout le
réconfort que Tu as
toujours étépour
moi.

HOMMAGES
Nous présentons nos hommages :
A notre maître de mémoire,
Marc T. KPODEKON
Professeur Agrégé de Médecine
vétérinaire, Enseignant chercheur à
l'EPAC, ex Directeur de L'Ecole Polytechnique d'Abomey-Calavi. Vous avez
accepté de diriger, avec bienveillance et disponibilité ce
travail, malgré vos multiples occupations. Votre modestie, votre esprit
de décision, votre rigueur scientifique, font de vous un grand homme qui
suscite en nous l'engagement à la recherche.
Au Professeur Issaka YOUSSAO
Maître de conférences des universités,
Enseignant chercheur à l'EPAC, Chef du Département de Production
et Santé Animales. Votre disponibilité à nous orienter
malgré vos multiples occupations s'ajoute à la grande estime et
à l'admiration que nous portons en votre personne.
Au Président de jury
Qui nous a fait l'honneur d'accepter la présidence du jury
de notre mémoire. Hommages respectueux.
Aux Honorables membres du jury
Qui nous ont fait l'honneur de participer à notre jury de
mémoire, ces hommages sont particulièrement les vôtres.
A tous les Enseignants de l'Ecole Polytechnique
d'Abomey-Calavi (EPAC) en particulier ceux du Département de
Production et Santé Animales (PSA), vous avez été les
artisans de notre formation, recevez nos hommages mérités.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
REMERCIEMENTS
A mes très chers parents
Adrien et Elisabeth GOUDJO, qui m'ont soutenu et
supporté depuis toujours, qui ont été toujours là
lorsque j'en ai besoin, sachez que je m'efforce sans cesse de me montrer digne
de vous. Longue vie à vous !
A Lucrèce AVANON
Merci pour ta générosité, ta tendresse et
ton amour indéfectible. Tu es pour moi une véritable compagne,
tes conseils et ton soutien permanent m'ont beaucoup aidé. Que Dieu
te bénisse !
A mes frères :
Léandre, Honorat et Paterne
; mes soeurs : Chantal, Ella,
Gloria, Auriane et Aurore
pour l'affection que j'ai toujours reçue de vous, ce travail est aussi
le vôtre. Je vous souhaite aussi de réussir. Que Dieu vous
garde !
A mes grands-parents
Pour m'avoir enseigné les pas à suivre dans la
vie... un grand merci.
A Monsieur Yaou A. DJAGO
Responsable-Adjoint du CECURI, vous m'avez fait
découvrir en partie la filière cunicole. Je vous remercie pour
votre écoute, pour votre gentillesse et pour tout ce que j'ai pu
apprendre avec vous.
A mon superviseur de stage, Docteur Yao
AKPO
Pour s'être montré toujours disponible et
compréhensible. Pour avoir accepté d'être mon guide et pour
m'avoir soutenu lors de mon stage au CECURI. Merci de m'avoir fait
confiance.
A Monsieur Ghislain MEDENOU
Pour votre aide et vos précieuses explications sur les
fiches d'enregistrements
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
utilisées lors des expérimentations au CECURI.
A Monsieur Boris DOMINGO
Pour vos différentes aides. Merci à vous.
A Serge DAFFA, Barnabé DESSOH et
Valdano TOSSE
Véritables amis. Nous avons réussi tous
ensemble, et surtout, nous avons réussi dans nos voies respectives. Vos
soutiens ont été l'une des clefs de ma réussite,
sachez-le.
A Jean de Kanty SENAHOUN
Des mois de stage au CECURI auraient été beaucoup
moins séduisants sans toi ! Merci pour tout.
A D. David, A. Raoul,
A. Arnaud, B. Samuel, D.
Nazaire, H. Jeannot, K. Mauris,
L. Martial, S. Ibrahim, S.
Bienvenue, T. Verkys, remarquables promotionnaires,
bonne chance à vous tous. Ce fut une aventure difficile, mais
enrichissante à tout point de vue : études, travail de groupe et
relations humaines.
Aux membres de ma famille, à mes amis, à
toutes les personnes que j'ai côtoyées et
appréciées, et qui m'ont apprécié en retour.
Merci pour tout.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
LISTE DES SIGLES ET ABREVIATIONS
ABeC : Association Béninoise des
Cuniculteurs
ACP-LP : Association Cunicole de Provenderie -La
Providence
al. : Collaborateurs en latin
CECURI : Centre Cunicole de Recherche et
d'Information
EPAC : Ecole Polytechnique d'Abomey-Calavi
FAO : Organisation des Nations Unis pour
l'Agriculture et l'Alimentation
FCFA : Franc de la Communauté Francophone
d'Afrique
?2 : HéritabilitéIA
: Insémination Artificielle
INRA : Institut National de la Recherche
Agronomique
N° : Numéro
SAGA : Station d'Amélioration
Génétique des Animaux
SAS : Statistical Analysis System
Vs : Contre
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
vii
|
UAC/EPAC/DPSA/2010 Elysée A.
GOUDJO
|
|
LISTE DES TABLEAUX
Tableau I : Taux de lapine ovulant en fonction de la couleur de
la vulve au
|
|
|
|
moment de la saillie (Boussit, 1989)
17
|
|
|
Tableau II
|
:
|
Effet de l'âge à la première saillie sur le
taux de fertilité
|
|
|
|
(Boussit, 1989)
18
|
|
|
Tableau III
|
:
|
Tailles moyennes de portées des nés totaux par mise
bas................
43
|
|
|
Tableau IV
|
:
|
Paramètres de position de la taille des portées des
nés totaux
44
|
|
|
Tableau V
|
:
|
Tailles moyennes de portées des nés vivants par
mise bas
|
45
|
|
Tableau VI
|
:
|
Paramètres de position de la taille des portées des
nés vivants...........
46
|
|
|
Tableau VII
|
:
|
Taux de
mortinatalité...........................................................
47
|
|
|
Tableau VIII
|
:
|
Tailles moyennes des portées au sevrage par mise
bas.................. .
|
47
|
|
Tableau IX
|
:
|
Paramètres de position de la taille des portées au
sevrage.................
48
|
|
|
Tableau X
|
:
|
Taux de mortalité
naissance-sevrage.......................................
|
49
|
|
Tableau XI
|
:
|
Poids moyens des portées au sevrage
|
50
|
|
Tableau XII
|
:
|
Poids moyens d'un lapereau au sevrage
|
50
|
|
Tableau XIII
|
:
|
Taux de fertilité, prolificité et intervalle entre
mises bas...................
|
51
|
|
Tableau XIV
|
:
|
Taux de gestation
|
51
|
|
Tableau XV
|
:
|
Poids moyens des
croisés.......................................................
|
54
|
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
LISTE DES FIGURES
Figure 1 : Ovaire de lapine avec deux générations
de corps jaunes après
insémination (Theau-Clément,
2005)..................................... 16
Figure 2 : Démarche générale en
sélection (Ouyed et Maignel, 2010).............. 25
Figure 3 : Plan d'accouplement des reproducteurs en
sélection 36
Figure 4 : Distribution des tailles de portées des
nés totaux (2008)................. 44
Figure 5 : Distribution des tailles de portées des
nés vivants (2008).......... 46
Figure 6 : Distribution des tailles de portées au sevrage
(2008)..................... 48
Figure 7 : Distribution des tailles de portées des
croisés nés vivants.......... 52
Figure 8 : Distribution des tailles de portées des
croisés au sevrage .......... 53
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
TABLE DES MATIERES
DEDICACE iHOMMAGES iiiREMERCIEMENTS
iv
LISTE DES SIGLES ET ABREVIATIONS viLISTE DES
TABLEAUX viiLISTE DES FIGURES viiiTABLE DES MATIERES
ix
RESUME xiv
ABSTRACT xv
INTRODUCTION 1
Première Partie : 3
1- SYNTHESE BIBLIOGRAPHIQUE 4
1.1- GENERALITES SUR LA PRODUCTION CUNICOLE 4
1.1.1- Cuniculture dans le monde 4
1.1.2- Cuniculture au Bénin 5
1.1.2.1- Difficultés d'évaluation de la production
5
1.1.2.2- Evolution de la production cunicole au Bénin 6
1.1.3- Quelques races de lapins 7
1.1.3.1- Grandes races 7
1.1.3.2- Races moyennes 8
1.1.3.3- Petites races de lapins 10
1.1.3.4.- Races de lapins nains 11
1.1.4- Aspects morphologiques 12
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.1.5- Anatomie du lapin 12
1.2- REPRODUCTION ET INSEMINATION ARTIFICIELLE CHEZ LA LAPINE
13
1.2.1- Physiologie de la reproduction 13
1.2.1.1- Développement des gonades 13
1.2.1.2- Puberté 13
1.2.1.3- Cycle oestral 13
1.2.1.4- Ovulation 14
1.2.1.5- Physiologie post-ovulatoire 14
1.2.1.5.1- Gestation 14
1.2.1.5.2- Mise bas 15
1.2.1.5.3- Pseudo-gestation 15
1.2.1.6- Comportement sexuel de la lapine 16
1.2.1.7- Facteurs de variation de la fertilité 17
1.2.1.7.1- Facteurs de variation liés à la conduite
des femelles 17
1.2.1.7.2- Facteurs liés à l'individu 18
1.2.1.7.3- Facteurs climatiques 19
1.2.1.8- Facteurs de variation de la prolificité 20
1.2.1.9- Facteurs de variation de la viabilité et du poids
au sevrage 21
1.2.2- Insémination artificielle chez la lapine :
définition et importance 21
1.2.3- Méthodes d'induction de la
réceptivité sexuelle des lapines 22
1.2.3.1- Méthodes hormonales 22
1.2.3.2- Effet du mâle 23
1.2.3.3- Séparation ponctuelle de la mère et sa
portée 23
1.3- AMELIORATION GENETIQUE 23
1.3.1- Notion de sélection 24
1.3.1.1- Objectif de sélection 24
1.3.1.2- Critères de sélection 24
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.3.1.3- Démarche générale en
sélection 25
1.3.1.4- Estimation de la valeur génétique 25
1.3.1.5- Méthodes de sélection 26
1.3.1.5.1- Sélection massale 26
1.3.1.5.2- Sélection sur ascendance 26
1.3.1.5.3- Sélection sur descendance 26
1.3.1.5.4- Sélection sur collatéraux 27
1.3.1.6- Paramètres importants en sélection 27
1.3.1.6.1- Généalogie 27
1.3.1.6.2- Héritabilité 27
1.3.1.6.3- Répétabilité 28
1.3.1.7- Progrès génétique 28
1.3.2- Croisement : apport potentiel du croisement, principaux
plan de croisement 29
1.3.2.1- Apports potentiels du croisement 29
1.3.2.1.1- Apport de gènes nouveaux 29
1.3.2.1.2- Complémentarité entre
caractéristiques 29
1.3.2.1.3- Effet d'hétérosis 30
1.3.2.1.4- Accroissement de la variabilité
génétique 30
1.3.2.2- Principaux plans de croisement 31
1.3.2.2.1- Création d'une race synthétique 31
Deuxième Partie : 33
2- METHODOLOGIE 34
2.1- Milieu d'étude 34
2.2- Animaux utilisés 34
2.2.1- Caractéristique de la souche INRA 1777 34
2.3- Balance et instruments de marquage 35
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
2.4- Répartition des animaux 36
2.4.1- Plan d'accouplement des reproducteurs en sélection
36
2.5- Critères de sélection des reproducteurs 37
2.6- Elimination des mâles et des femelles 37
2.7- Matériels d'insémination artificielle 37
2.8- Prélèvement du sperme 38
2.9- Insémination proprement dite 38
2.10- Prophylaxie 39
2.10.1- Prophylaxie sanitaire 39
2.10.2- Prophylaxie médicale 39
2.11- Mesures effectuées et données recueillies
39
2.12- Calcul des performances de reproductions 40
2.13- Analyses statistiques 40
Troisième Partie : 42
3- RESULTATS ET DISCUSSION 43
3.1- Performances de reproduction des femelles en
sélection au CECURI (2008) 43
3.1.1- Tailles moyennes de portées des nés totaux
43
3.1.2- Tailles moyennes de portées des nés vivants
45
3.1.3- Taux de mortinatalité 46
3.1.4- Tailles moyennes des portées au sevrage 47
3.1.5- Taux de mortalité naissance-sevrage 49
3.1.6- Poids moyen des portées au sevrage par mise bas
49
3.1.7- Poids moyens d'un lapereau au sevrage par mise bas 50
3.1.7- Taux de fertilité, prolificité et intervalle
entre mise bas 50
3.2- Productivité des femelles croisées avec des
mâles de souche améliorée 51
3.2.1- Taux de gestation 51
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
3.2.2- Tailles moyennes de portées des croisés
à la naissance 52
3.2.3- Tailles de portées des croisés au sevrage
53
3.2.4- Mortinatalité et mortalité naissance-sevrage
53
3.2.5- Poids moyens des croisés 54
3.3- DISCUSSION 55
3.3.1- Performances de reproduction des femelles en
sélection au CECURI (2008) 55
3.3.1.1- Tailles moyennes de portées des nés totaux
55
3.3.1.2- Tailles moyennes de portées des nés
vivants 56
3.3.1.3- Taux de mortinatalité 56
3.3.1.4- Taux de mortalité naissance sevrage 56
3.3.1.5- Tailles moyennes des portées au sevrage 57
3.3.1.6- Poids moyens des portées au sevrage 57
3.3.1.7- Fertilité et prolificité 58
3.3.2- Productivité des femelles croisées avec des
mâles de souche améliorée 59
3.3.2.1- Taux de gestation 59
3.3.2.2- Prolificité à la naissance et au sevrage
59
3.3.2.3- Poids moyens des croisés 60
CONCLUSION ET SUGGESTIONS 61
REFERENCES BIBLIOGRAPHIQUES 63
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
RESUME
Dans le cadre de l'amélioration de la
productivité numérique des lapins, un programme de
sélection a été exécuté au CECURI de Janvier
2006 à Décembre 2008. Ce programme à pour but
d'évaluer les performances de reproduction des femelles. Au total 140
femelles et 42 mâles répartis en 10 familles ainsi que 32 femelles
supplémentaires hors sélection pour égaliser les
naissances ont été utilisées. Les lapines ont
été conduites en rythme semi-intensif (saillie 10 jours
post-partum) et nourries ad libitum. La méthode de
sélection sur ascendance a été adoptée. En 2008, la
taille moyenne des portées a été de 5,53#177;2,20
nés vivants et de 3,92#177;2,10 sevrés. Les résultats ne
montrent aucun effet significatif (P > 0,05) du numéro de
portée sur la taille des portées au sevrage. La taille moyenne
des portées au sevrage en 2008 est similaire à celle
enregistrée en 2007 (3,28#177;2,37), mais inférieure à
celle observée en 2006 (4,78#177;2,18). Le poids moyen d'un lapereau
pour les deuxièmes mises bas est de 483,24#177;137,62 g et est
significativement moins élevé (P < 0,05) que celui des
troisièmes mises bas (533,53#177;155,01 g). Le taux de fertilité
a été de 65,95%. Le taux de mortinatalité et de
mortalité naissance-sevrage ont été respectivement de
2,09% et 36,61%. En Mars 2010, dans le but de créer une souche
synthétique pour améliorer la production cunicole au
Bénin, 104 femelles issues des élevages du Sud Bénin
toutes nullipares ont été inséminées. Le taux de
gestation enregistré a été de 67,31%. Les tailles de
portées (4,55#177;2,92 nés vivants, 2,71#177;2,34 sevrés)
observées sont inférieures aux normes en zones tropicales. Les
poids moyens des croisés sont de 52,20#177;16,42 g à j0,
419,66#177;136,62 g à j28 et 580,60#177;147,21 g à j35. Le taux
de mortalité naissance sevrage a été de 43,69%. La
sélection a amélioré la prolificité des lapines,
mais l'incidence des pathologies sur la viabilité des lapereaux entre la
naissance et le sevrage est encore important. Il est aussi pertinent de
rechercher des méthodes d'induction de la réceptivité
susceptibles d'améliorer non seulement la fertilité, mais aussi
la productivité globale des lapines en insémination
artificielle.
Mots-clés : lapines -- sélection -
productivité -- insémination - souche.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
ABSTRACT
In the context of improving the numeric productivity of the
rabbits, a breeding program was implemented at CECURI from January 2006 to
December 2008 in order to evaluate the reproductive performance of females. 140
females and 42 males divided into 10 families and 32 supplementary females out
selection to equalize litters. The rabbits were conducted in semi-intensive
rhythm (projecting 10 days post-partum) and fed ad libitum.
The method of selection on ancestry was adopted. In 2008, the average litter
size was 5.53#177;2.20 born alive and weaned from 3.92#177;2.10. The results
show no significant effect (P > 0.05) number worn on litter size at weaning.
The average litter size at weaning in 2008 is similar to that recorded in 2007
(3.28#177;2.37), but lower than that observed in 2006 (4.78#177;2.18). The
average weight of a rabbit for the second farrowing was 483.24#177;137.62 g and
was significantly lower (P < 0.05) than the third calving (533.53#177;155.01
g). The fertility rate was 65.95%. The rate of stillbirths and birth weaning
mortality were respectively 2.09% and 36.61%. In March 2010 in order to create
a synthetic strain to improve rabbit production in Benin, 104 females from the
farms South Benin nulliparous all were inseminated. The pregnancy rate was
recorded 67.31%. The litter sizes (4.55#177;2.92 live births, 2.71#177;2.34
pigs) are observed below the standards in the tropics. The average weight of
the Crusaders was 52.20#177;16.42 g at day 0, 419.66#177;136.62 g at day 28 and
580.60#177;147.21 g at day 35. The birth weaning mortality rate was 43.69%. The
selection has improved prolificacy of does, but the incidence of diseases on
the viability of young rabbits between birth and weaning is still important. It
is also important to seek methods of induction of receptivity could improve not
only fertility but also the overall productivity of rabbits in artificial
insemination.
Keywords: rabbits - selection - productivity -
insemination - strain.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
INTRODUCTION
Le monde entier connaît ces dernières
années des problèmes cruciaux de sous alimentation et l'Afrique
en particulier est confrontée à des difficultés
d'autosuffisance alimentaire. En effet, les efforts fournis dans le secteur de
l'élevage et celui de l'agriculture en Afrique sont encore loin de
satisfaire les besoins alimentaires des populations (FAQ, 2000). Le continent
doit encore importer en complément une part essentielle des
protéines d'origines animales qu'il consomme.
Au Bénin, la production des aliments d'origine animale
est très insuffisante par rapport à la demande des populations.
Un tel déficit se trouve comblé par les importations de produits
congelés qui ne cessent d'augmenter d'année en année. La
quête de solution durable pour remédier à ce déficit
a permis de développer l'élevage des espèces à
cycle court et prolifiques telle que le lapin. Le lapin, est un animal
homéotherme qui par divers mécanismes comportementaux et
physiologiques s'adapte aux conditions de son environnement. Cette
faculté rend certes son élevage possible dans diverses zones
climatiques mais toutefois, ne le met pas à l'abri de l'influence des
variations climatiques. Matheron et Poujardieu (1982) signalent qu'à des
températures supérieures à 30°C, le taux d'ovulation
chez la lapine est réduit de moins de 5%. De même Howarth et
al. cités par Saleil et al. (1998) ont noté chez
les lapines des pertes embryonnaires dans les mêmes conditions. Le
Bénin, n'échappe pas à ces réalités. Depuis
quelques années l'élevage de lapin évolue de façon
croissante, avec une prédominance de l'élevage semi-commercial.
Le nombre de lapines-mères est passé de 2.251 en 1998 à
17.200 en 2007 (ABeC, 2007) avec une productivité numérique au
sevrage de 20 à 25 lapereaux par femelle et par an pour les
élevages bien suivis.
L'utilisation de lapins issus d'un programme de
sélection qui tient compte des critères d'importance
économique permettra aux producteurs d'améliorer la
productivité de leurs élevages, de faire face à la demande
nationale et de tirer bénéfice du progrès
génétique réalisé. C'est ainsi que, depuis 2006 le
CECURI a mis en place un programme de sélection des reproducteurs afin
de caractériser les
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
performances de reproduction des lapins de souche locales et
de définir les meilleurs schémas de sélection pour la
production de lapins hybrides. En effet, dans les conditions tropicales la
taille moyenne des portées à la naissance admise est de 6,4
lapereaux (Djago et Kpodékon, 2000). Ce résultat comparé
à ceux des pays tempérés soit 13,0 lapereaux pour les
températures comprises entre 6 et 10°C ; 9,0 lapereaux pour les
températures supérieurs à 23°C (Lebas, 1996) est bas
et peut être amélioré. Ainsi, les travaux de
sélection entrepris au CECURI ont permis l'éradication de
certaines tares génétiques. Il en ressort que, la taille moyenne
de portées des nés totaux enregistrés est passée de
5,47 de 1995 à 1998 à 5,73#177;1,79 en 2006 et croît
à 6,04#177;2,04 en 2007 (Tanimomo, 2007 ; Hounton, 2008). De plus, Il a
été noté que la taille moyenne des portées au
sevrage est passée de 4,78 à 3,28#177;2,37 de 2006 à
2007.
Cet état des lieux des études
précédentes nous a poussés à poursuivre
l'étude des résultats des années suivantes afin de
constater l'intérêt du programme d'amélioration
génétique en cours au CECURI. La présente étude
intitulée Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche améliorée au CECURI à
pour but de faire le point des résultats de sélection en 2008 et
ceux des femelles inséminées au cours de l'année 2010.
Les objectifs spécifiques de ce travail sont
d'évaluer :
· la fertilité et la prolificité des femelles
reproductrices en sélection ;
· la viabilité et le poids au sevrage des lapereaux
;
· les performances de reproduction des femelles
inséminées.
Ce travail est subdivisé en trois grandes parties. La
première énonce des généralités et fait la
synthèse des notions de physiologie de la reproduction, de
l'insémination artificielle et de l'amélioration
génétique. La deuxième présente la
méthodologie utilisée pour réaliser l'étude et la
troisième enfin expose les résultats obtenus suivis d'une
discussion, d'une conclusion et des suggestions.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
Première Partie :

Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1- SYNTHESE BIBLIOGRAPHIQUE
1.1- GENERALITES SUR LA PRODUCTION CUNICOLE
1.1.1- Cuniculture dans le monde
Si la domestication des grandes espèces à
intérêt zootechnique (bovins, ovins, porcins) comme celle des
petites espèces (volailles) se perd dans la nuit de la
préhistoire, celle du lapin est tout encore plus récente. En
effet, originaire du Sud de l'Europe et de l'Afrique du Nord, le lapin sauvage,
Oryctolagus cuniculus, aurait été
«découvert» par les Phéniciens lors de leur prise de
contact avec l'Espagne vers l'an 1000 avant Jésus-Christ (Lebas et
al., 1996). Au temps des Romains, le lapin reste le symbole de l'Espagne.
Il semble bien que ce soit les Romains qui aient disséminé le
lapin dans l'empire comme animal gibier. Dès le 16ème
siècle, on connaît plusieurs races dont la domestication remonte
de ce fait au moyen âge. Au début du 19ème siècle,
l'élevage du lapin en clapiers se développe dans toute l'Europe
Occidentale, aussi bien en milieu rural qu'en milieu urbain (chez les ouvriers
des banlieues). Au cours des années 50, la cuniculture subit de
profondes transformations notamment dans les méthodes de production. En
même temps, apparaissent des troubles pathologiques jusqu'alors inconnus,
apparemment liés aux nouvelles méthodes «d'élevage
intensif» : entérites mucoïdes, troubles respiratoires,
troubles digestifs, etc.
La production totale du lapin dans le monde a été
estimée à 1,2 million de tonnes de carcasses en 1992 et à
1,5million en 1994 (Lebas et al. 1996).
Les principaux producteurs sont l'Italie, la Russie,
l'Ukraine, la Hongrie, la France, la Chine et l'Espagne. L'Europe assure 75% de
la production mondiale et la Chine assure la majorité de la production
restante. L'élevage du lapin est presque inexistant dans la
majorité des Pays du Proche-Orient. Des foyers d'élevage existent
dans quelques régions d'Amérique Centrale, en Asie du Sud-Est et
en Afrique. Les pays de l'Afrique Sud Sahélienne n'ont pas de tradition
cunicole en raison de l'origine assez récente de ce type
d'élevage. La viande de lapin est jusqu'à nos jours
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
encore assez peu consommée. Le lapin a fait son
apparition dans les pays du Golfe de Guinée (Côte d'Ivoire, Ghana,
Togo, Bénin et Nigeria) depuis le XIXème siècle. En effet,
il y fut introduit par les missionnaires ou les colons (Kpodékon, 1988).
A partir des années 70, les gouvernements de ces pays, afin
d'améliorer l'alimentation de leurs populations sans aggraver le
déficit du commerce extérieur, ont été
amenés à développer l'élevage du lapin :
· le Ghana en 1972 avec le projet <National Rabbit
Project» ;
· le Bénin à travers le <Centre Cunicole
de Recherches et d'Information» : CECURI en 1988 ;
· au Nigeria la Fédération des
Coopératives dispose d'un centre de diffusion des reproducteurs à
Ibadan à 100km au Nord de Lagos ;
· le Togo possède trois fermes modèles :
BENA-DEVELOPPEMENT,
BETANIA et le centre de formation de Batome (Colin et
Lebas, 1995).
1.1.2- Cuniculture au Bénin
1.1.2.1- Difficultés d'évaluation de la
production
L'une des particularités actuelles de la production
cunicole béninoise est la coexistence de deux systèmes de
production : un système traditionnel issu de petits élevages
familiaux et un système rationnel (constitué en majeurs parties
d'éleveurs membres de l'ABeC et d'autres éleveurs privés :
mikado...). La part de l'élevage traditionnel reste encore importante,
beaucoup plus que dans d'autres filières. Elle est difficile à
estimer car elle échappe aux enquêtes et aux recensements et est
très peu impliquée dans les circuits commerciaux
organisés. La production globale est donc évaluée d'une
part à partir des abattages contrôlés de l'ABeC, qui
correspondent essentiellement à la production de ses membres et d'autre
part avec les chiffres de la fabrication d'aliments qui permettent de donner
une estimation de la production traditionnelle.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.1.2.2- Evolution de la production cunicole au
Bénin
Jusqu'à la fin des années 87, il n'existait
aucune statistique fiable sur la production cunicole au Bénin. Il aura
fallu attendre les résultats d'enquêtes de Kpodékon (1988),
pour affirmer que la cuniculture est pratiquée un peu partout au
Bénin et que de nombreuses conditions favorables à son
développement sont réunies.
La cuniculture au Bénin connaît une augmentation
sans cesse croissante. En effet, en 1988 on dénombrait environ 400
élevages répartis dans tout le pays (Kpodékon, 1988) avec
en moyenne 4,1 lapines mères (Kpodekon et Coudert, 1993). Suite à
l'avènement successif de la peste porcine et de la grippe aviaire qui
ont décimé les élevages de porcs et de volailles et ont
favorisé le développement de l'élevage de lapins, on note
une forte augmentation de la production cunicole. Selon les donnés
statistiques de la FAO (2000) et de l'ABeC (2007) le nombre d'élevage
est passé de 400 en 1998 à 1350 en 2007 avec une forte
prédominance de l'élevage familiale. La production totale
annuelle de carcasses de lapins au Bénin a atteint 400 tonnes en 2005
(ABeC, 2007). Les lapins ont une origine génétique très
variée et sont le plus souvent métissés de manière
anarchique.
De plus en plus, la viande de lapin entre dans les habitudes
alimentaires des Béninois. Le marché est très variable,
avec une demande plus élevée que l'offre. La plus grosse
clientèle reste les supermarchés, les boucheries modernes, les
restaurants, les maquis (petits restaurants publics) et les hôtels, bien
qu'actuellement la demande augmente chez les particuliers qui servent cette
viande lors des anniversaires, des mariages, des cérémonies de
baptême et au cours des fêtes. Les lapins produits sont souvent
livrés, abattus s'ils sont destinés à la consommation
directe, où vivants comme reproducteurs.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.1.3- Quelques races de lapins
1.1.3.1- Grandes races
1.1.3.1.1- La Géante blanche du
Bouscat
Originaire de la France (au Bouscat, près de
Bordeaux), la race Géante blanche du Bouscat est issue du
croisement-sélection entre l'Argenté de Champagne, le
Géant des Flandres et le lapin Angora. Le mâle pèse entre 5
kg et 6 kg. C'est une race à oreilles assez épaisses, aux
extrémités arrondies et mesurant entre 17 et 18,5 cm. Sa fourrure
est bien longue (35 à 40mm), épaisse et souple avec un reflet
givré. Elle a une robe blanche. Elle présente un chanfrein assez
busqué. Son corps est robuste et assez allongé. La ligne dorsale
forme un léger arc. Les cuisses émergent quelque peu. Sa croupe
est bien développée. Le fanon est apparent chez la femelle. Les
pattes arrière sont vigoureuses. Elle a des yeux grands ouverts et de
couleur rouges. C'est une race adaptée au climat froid (ASFC, 2008).
1.1.3.1.2- La Géante des Flandres
Originaire de la Belgique, c'est une race rustique issue de
la sélection des lapins locaux. Elle pèse entre 5,5 kg et 7 kg
(3,5 kg à 4 mois). Ses oreilles sont grandes (plus de 16 cm),
épaisses, larges (entre 4 et 6 cm), portées droites et en forme
de V, unies à la base et bordées d'un liseré noir.
L'ouverture est dirigée vers l'avant. Sa fourrure est fine, lisse,
épaisse et souple. Elle peut être mate ou brillante. La couleur de
cette race varie : elle peut être gris lièvre, gris garenne, gris
fer, gris bleue, fauve, noire, bleue, blanche. Sa tête est droite, forte
et large. Son cou est imperceptible. Son corps est grand, large, long, profond
et forme un rectangle vu de dessus. Elle possède une puissante
musculature et a des épaules basses. Sa croupe est arrondie. Ses pattes
sont longues et bien musclées. C'est un lapin docile et résistant
aux stress. Elle est mise à la reproduction entre 10 et 12 mois. Elle
est très prolifique (9 lapereaux en moyenne dont 1 mort-né) et
souffre le plus souvent de l'absence de secondes incisives (ASFC, 2008).
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.1.3.1.3- Le Géant Papillon
Français
C'est une race originaire de la France et issue de la
mutation de lapins tachetés. Elle pèse 3,1 kg à 4 mois, et
présente une fourrure avec des poils recteurs discrets pour favoriser
l'expression du dessin (papillon). Elle est de couleur noire, havane ou bleue
le plus souvent. Toutes les couleurs peuvent être admises à
condition que le dessin soit bien marqué. Le dessin doit être
nettement délimité et intense. Le chanfrein est rectiligne, son
cou est imperceptible. Son corps est puissant et allongé,
uniformément développé. Ses épaules sont basses.
Ses pattes sont courtes et vigoureuses. Génétiquement, cette race
souffre du prognathisme mandibulaire (ASFC, 2008).
1.1.3.2- Races moyennes
1.1.3.2.1- La Fauve de Bourgogne
D'origine française, elle est issue d'une population
locale de lapins fauves. L'adulte pèse entre 4 et 4,5 kg. Elle a les
oreilles bien droites, légèrement en forme de V à leur
sommet, robustes, velues et serrées à la base. Elles ont une
forme arrondie à leur extrémité et une longueur comprise
entre 11,5 et 12,5 cm. Sa fourrure est dense ; les poils sont longs de 35 mm.
Elle a une couleur fauve, roux intense ou uniforme. Chez le mâle, la
tête est forte et large, ronde et non busquée (courbure
bombée) ; le cou est imperceptible et musclé. Son corps est trapu
et massif, de largeur quasi similaire en tout point (vu de dessus), plus large
au niveau du râble et des épaules. Sa poitrine est large. Sa
croupe est bien développée, musclée et remplie, sans
saillies osseuses. Ses pattes sont fortes, robustes et courtes, la largeur de
la poitrine à l'avant est bien serrée au corps à
l'arrière. Ses ongles sont de couleur noir non cornés. Elle a les
yeux bien ouverts, vifs et expressifs, jamais trop proéminents ni trop
enfoncés. Elle jouit d'une certaine vitalité. La lapine est mise
à la reproduction entre 6 et 8 mois et donne en moyenne 9 lapereaux par
portée dont 1 mort-né. Elle souffre de la maladie des muscles
blancs, du prognathisme mandibulaire et du glaucome héréditaire.
C'est une race qui est parfois croisée avec
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
les races Argentés de Champagne, Géantes Blanches
du Bouscat, Néo Zélandaises et Californiennes (ASFC, 2008).
1.1.3.2.2- La Californienne
Elle est apparue en 1922 aux Etats Unis d'Amérique
(Californie). Elle pèse entre 3,5 et 5 kg. Sa fourrure est dense et
assez lustrée. Sa robe est blanche, aux extrémités
(oreilles, pattes, queue, nez) foncées. Sa tête est forte, large,
ronde et non busquée. Son cou est imperceptible. Son corps est massif,
court, ramassé, charnu, musclé ; au râble épais, aux
épaules larges, et aux cuisses arrondies, avec une ligne dorsale
légèrement bombée. La croupe est large et bien arrondie,
sans partie saillante, à la même hauteur que les épaules.
Ses yeux sont rouges. Elle est mise à la reproduction entre 7 et 9 mois.
Elle a une prolificité comprise entre 8 et 10 lapereaux à la
naissance (6 en moyenne au sevrage). Elle est adaptée aux climats
tropicaux et est souvent croisée avec le Néo zélandais
(ASFC, 2008).
1.1.3.2.3- La Néo Zélandaise
Originaire des Etats Unis d'Amérique, elle est le
résultat de la sélection des lapins colorés
américains. Elle s'acclimate très bien aux régions
tropicales. Sa robe est blanche et ses yeux sont roses. Elle pèse entre
4,5 et 5,25 kg. Ses oreilles sont très robustes aux bases,
épaisses, bien droites et assez rapprochées. Elles sont longues
de 11 à 12 cm. Sa fourrure est très dense, souple et assez
lustrée. Sa tête est pleine avec des mâchoires
développées, volumineuse, mais harmonieuse. Son corps est
puissant, développé, très massif et moyennement long avec
un dos râble, une croupe large et arrondie. Ses pattes sont assez
courtes, lourdes à l'avant et fortes à l'arrière (ASFC,
2008).
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.1.3.3- Petites races de lapins
1.1.3.3.1- Hollandais
Originaire de la Hollande, c'est une race qui possède
de petites pattes, elle est relativement animée et atteint sa
maturité à l'âge de 7 à 9 mois. Le nombre de
lapereaux par portée est compris entre 6 et 8 avec d'importantes pertes
avant le sevrage. Sa fourrure est dense, souple et courte. Elle souffre
généralement de prognathisme mandibulaire, d'ostéoporose,
de courbure de l'avant-bras, de Spinabifida (ASFC, 2008).
1.1.3.3.2- L'Argenté Anglaise
C'est une race originaire de l'Angleterre, elle pèse
entre 2,5 et 2,9 kg. Ses oreilles sont droites et mesurent entre 8,5 et 9,5 cm.
Sa fourrure est courte, dense, élastique et ses poils sont épais.
Elle possède une robe argenté (poils unis et à pointes
blanches) régulièrement répartie et des yeux
légèrement proéminents. Au sein de cette race on peut
distinguer différents individus (unicolore : argenté noir,
argenté bleu, havane. Agouti : argenté brun, argenté
crème). C'est une race à tête forte et anguleuse. Son corps
est court et trapu, rond et potelé avec une poitrine pleine, et des
épaules puissantes. Son dos est légèrement arqué,
râble et très épais. Ses pattes sont petites (ASFC,
2008).
1.1.3.3.3- Havane Français
Elle pèse entre 2,5 et 2,9 kg. Ses oreilles sont
portées droites, bien serrées et velues. Elle possède une
fourrure dense, souple et luisante avec des poils assez fins. Elle a une robe
havane soutenue (cigare foncé), très uniforme et lumineuse. Le
dessous du corps est plus mat. La tête est courte, large chez le
mâle, un peu plus fine chez la femelle. Elle possède un corps
légèrement arrondi et élégant, la poitrine et les
épaules sont pleinement formés. Ses pattes sont fines, ses yeux
sont bien ouverts et brillants de couleur brune marron foncée. Elle
donne entre 7 et 10,7 lapereaux en
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
moyenne (1 mort né). La maladie génétique
connue pour cette race est l'achondroplasie (ASFC, 2008).
1.1.3.3.4- Chinchilla
Elle provient de la France et pèse entre 2,5 et 2,9 kg
à l'âge adulte, 50 g à la naissance, 650 g à 28
jours et 1,4 kg à 77 jours. Ses oreilles mesurent entre 8,5 et 9,5 cm
elles sont très consistantes, velues et portées droites. Sa
fourrure est très dense, souple et assez longue (au moins 30 mm). Elle a
une robe grise cendrée lumineuse avec des reflets bleutés ; au
chenillé noirâtre très prononcé et ondulé sur
le dos. Le dessus de la queue est noir, le dessous du corps (ventre, menton,
queue) et le tour des yeux blancs. C'est une race docile qui produit entre 7 et
10 lapereaux en moyenne avec 1 mort né et souffre
génétiquement du tremblement paralytique, d'ataxie, de l'oeil
rouge sur patron coloré et de luxation fémorale (ASFC, 2008).
1.1.3.4.- Races de lapins nains
1.1.3.4.1- Nain Bélier
Elle est originaire de la Hollande et est le résultat
du croisement entre le nain bélier français et le nain
bélier anglais. Elle pèse entre 1,4 et 1,7 kg. Ses oreilles sont
consistantes, verticales, avec une ouverture sur les joues. Sa fourrure est
dense, souple et moyennement longue. Elle possède une très forte
tête avec le front large, le museau développé, les joues et
les mâchoires pleines. Son chanfrein est fortement busqué. Son cou
est imperceptible. C'est une race docile, assez animée et peu prolifique
3 à 7 lapereaux en moyenne 4, tous vivants (ASFC, 2008).
1.1.3.4.2- Nain Angora
C'est une race rencontré un peu partout en Europe mais
surtout en France. Elle est le résultat du croisement entre Angora et
les lapins nains. L'adulte pèse environ entre 1,2 et 1,5 kg. Ses
oreilles sont droites, consistantes et serrées. Ses poils sont longs
(sauf sur les oreilles, le museau, le nez et le front) ; les oreilles ont un
plumet
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
de poils à leur extrémité. La fourrure est
épaisse. Elle possède de petites pattes et des yeux
légèrement proéminents. Elle est mature à 6 mois
(ASFC, 2008).
1.1.4- Aspects morphologiques
Pour la majorité des races, à l'exception des
naines, l'allure générale du corps est différente selon le
sexe. Une tête large et forte, un thorax développé, des
membres relativement épais et une musculature bien
extériorisée sont généralement les
caractéristiques du mâle. Les femelles présentent toutes
proportions gardées, plus de finesse générale avec une
tête plus étroite, un corps paraissant plus allongé et une
ossature un peu plus légère. Seul l'arrière-train est plus
développé avec un bassin large (Djago et Kpodékon,
2000).
1.1.5- Anatomie du lapin
Herbivore monogastrique, le lapin est un animal dont les
dents poussent continuellement contrairement à celles des bovins par
exemple ; il les use et les affûte par des mouvements continuels des
mâchoires (Djago et Kpodékon, 2000). Chez un lapin adulte de race
moyenne (4 à 4,5 kg) ou sub-adulte, le tube digestif a une longueur
d'environ 4,5 à 5 m (Lebas et al., 1996). Il est relativement
plus développé chez le jeune lapin que chez l'adulte car le tube
digestif atteint sa taille définitive chez un lapin de 2,5 à 2,7
kg. Selon Peeters et al. (1984) et Lebas (1991), les
éléments distinctifs constituant globalement le tube digestif du
lapin sont:
· un oesophage court ;
· deux réservoirs à savoir l'estomac et le
cæcum (lieu de la fermentation des aliments par les bactéries)
dont le contenu total représente 10% du poids vif de l'animal. L'estomac
représente 40% du volume total du tube digestif ;
· L'intestin grêle (lieu de
«démontage» des aliments en leurs éléments
nutritifs de base) est la première et la plus longue partie des
intestins (3,3 m environ chez l'adulte) ;
· Le côlon est la dernière partie des
intestins et mesure environ 1,3 m de longueur chez les adultes.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.2- REPRODUCTION ET INSEMINATION ARTIFICIELLE CHEZ LA
LAPINE
1.2.1- Physiologie de la reproduction
1.2.1.1- Développement des gonades
La différenciation des gonades a lieu au 16ème
jour qui suit la fécondation. Les divisions ovogoniales commencent le
21ème jour de la vie foetale et se poursuivent jusqu'à la
naissance. Les follicules primordiaux apparaissent au 13ème jour
après la naissance, les premiers follicules à antrum apparaissent
vers 9 à 10 semaines (AERA, 1994).
1.2.1.2- Puberté
Elle correspond au moment de la vie de la lapine où
cette dernière est capable
d'ovuler et de conduire une gestation. Elle survient
généralement quand la lapine atteint les deux tiers de son poids
adulte. L'acceptation de l'accouplement survient avant l'aptitude à
ovuler (AERA, 1994). Chez les races communes, la puberté serait atteinte
entre 100 et 118 jours post-partum. Les nullipares sont rarement mises à
la reproduction avant 16 à 17 semaines.
1.2.1.3- Cycle oestral
La lapine ne présente pas de cycle oestral avec
apparition régulière de chaleurs. On parle plutôt de
période de réceptivité ou de non réceptivité
(AERA, 1994). Selon les auteurs, la durée de la période de
réceptivité est variable. D'après Brower (2006), la lapine
serait réceptive pendant 7 à 10 jours. D'autres auteurs affirment
que la réceptivité des lapines correspondrait à la
présence à la surface de l'ovaire de follicules prêts
à ovuler et durerait 5 à 6 jours (Boussit, 1989). Pour savoir si
une lapine est réceptive, on peut regarder la couleur de la vulve. En
pratique, le seul critère fiable est l'acceptation de l'accouplement.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.2.1.4- Ovulation
Elle est souvent induite par l'accouplement. Le réflexe
ovulatoire fait intervenir deux voies successives :
· la voie afférente, transmettant les stimuli du
coït, des sens et des facteurs externes au système nerveux central
;
· la voie efférente, humorale, qui induit
l'ovulation.
L'hypothalamus libère la GnRH dans le système
sanguin, qui agit au niveau de l'antéhypophyse et libère à
son tour la FSH et la LH. La FSH provoque la maturation folliculaire finale :
le follicule de De Graaf, l'ovocyte primaire termine sa première
division méiotique pour donner un follicule secondaire et un premier
globule polaire. Le pic de LH atteint son maximum 90 minutes à 2 heures
de temps après le coït. Il est responsable de la rupture des
follicules de De Graaf et de l'ovulation, 10 à 12 heures après
l'accouplement. La LH stimule également le tissu ovarien qui
sécrète alors de la progestérone. L'ocytocine,
libérée par la posthypophyse, facilite l'ovulation (Boussit,
1989).
Un nouveau pic de FSH se produit, 16 à 22 heures de
temps après le coït, entrainant la formation de nouveaux follicules
cavitaires susceptibles d'ovuler par la suite, s'il n'y a pas eu de
fécondation.
1.2.1.5- Physiologie post-ovulatoire
1.2.1.5.1- Gestation
Le corps jaune est nécessaire tout au long de la
gestation. La sécrétion hypophysaire de LH, des mécanismes
propres à l'ovaire (notamment le taux d'oestrogènes) et les
messages chimiques contrôlés par l'unité foeto-placentaire
interviennent dans le maintien du corps jaune.
La durée de la gestation varie de 30 à 33
jours. Sa fin est marquée par l'inversion du rapport des taux
d'oestrogènes et de progestérone. Le taux de progestérone
chute fortement à partir du 27ème jour (AERA, 1994).
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.2.1.5.2- Mise bas
Le taux de progestérone diminue et n'est plus
suffisant pour empêcher les contractions utérines. Les glandes
surrénaliennes foetales sécrètent des corticoïdes,
qui passent dans le sang maternel et provoquent la libération
d'ocytocine par l'hypophyse maternelle, à l'origine des contractions
utérines croissantes. Les prostaglandines PGF2á, par leur
rôle lutéolytique, diminuent encore le taux de progestérone
(Boussit, 1989).
La lapine construit un nid quelques jours avant la mise bas
avec de la paille, des copeaux, auxquels elle ajoute des poils
prélevés sur son ventre et ses cuisses quelques heures avant la
parturition. Cette dernière dure rarement plus de trente minutes.
Dès la mise bas, la lapine est de nouveau fécondable et il
n'existe pas d'anoestrus de lactation, seulement une baisse de la
réceptivité entre le 3ème et le 9ème jour
post-partum (AERA, 1994).
1.2.1.5.3- Pseudo-gestation
En l'absence de fécondation, les corps jaunes ne se
maintiennent que 15 à 19 jours et empêchent toute nouvelle
ovulation. La sécrétion de progestérone augmente jusqu'au
12ème jour et provoque des modifications de l'utérus et des
glandes mammaires identiques à celle d'une lapine gestante. Cependant,
l'absence d'unités foeto-maternelles entraine la régression de
l'endomètre dès le 16ème et le 21ème jour (Boussit,
1989).
Les pseudo-gestations sont rares en saillie naturelle,
où l'absence de gestation est le plus souvent due à une absence
d'ovulation (lapine non réceptive et absence de coït). Après
l'insémination artificielle, l'absence de gestation provient souvent
d'une absence de fécondation ou d'une mortalité embryonnaire
précoce, car l'injection de GnRH pratiquée induit presque
systématiquement l'ovulation. Cependant, Boiti et al. (1996)
ont montré que près de 20 % des lapines ont au moment de
l'insémination des concentrations plasmatiques élevées de
progestérone associées à une faible
réceptivité sexuelle et une faible fertilité. Cette
observation a été confirmée par Theau-Clément
et al. (2000). En effet, sur les 170 lapines
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
observées 24 heures après
l'insémination, 35 d'entre elles présentaient 2
générations de corps jaunes (de 11 à 33) : une
première génération de corps jaunes récents
(correspondant à l'injection de GnRH, figure 1) et une seconde
génération de corps jaunes plus anciens et proéminents.
Ces lapines étaient caractérisées par une faible
réceptivité sexuelle (22%), une faible fertilité (3%) mais
toutes avaient ovulé.
Figure 1 : Ovaire de lapine avec deux
générations de corps jaunes après insémination
(Theau-Clément, 2005)
1.2.1.6- Comportement sexuel de la lapine
En saillie naturelle, le seul critère fiable signalant
l'oestrus chez la lapine est l'acceptation de l'accouplement. Les lapines
pouvant se montrer agressives envers les mâles introduits dans leur cage,
l'accouplement doit avoir lieux dans la cage du male. Elles sont
immédiatement retirées si aucune saillie n'a lieu (Brower,
2006).
Les oestrogènes induisent le comportement d'oestrus :
en présence du mâle, la lapine s'immobilise après une
courte phase de poursuite, puis se positionne en lordose. En cas de non
réceptivité, la lapine est ramassée sur elle même et
évite le mâle, voire présence de l'agressivité.
La progestérone sécrétée par le
corps jaune après l'ovulation n'inhibe pas complètement le
comportement sexuel des lapines qui, dans certains cas, acceptent encore le
mâle en cours de gestation (AERA, 1994).
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.2.1.7- Facteurs de variation de la
fertilité
La fertilité est le nombre de femelles mettant bas sur le
nombre de femelles mises àla reproduction. Elle se trouve
influencée par plusieurs facteurs.
1.2.1.7.1- Facteurs de variation liés à la
conduite des femelles
· Réceptivité des
femelles
Le seul signe donnant une indication sur l'état
physiologique de la lapine est la couleur de la vulve : plus celle-ci est
foncée, plus la probabilité d'être en présence d'une
femelle en oestrus augmente et plus la fertilité est bonne (tableau
1).
Tableau I : Taux de lapine ovulant en fonction de la
couleur de la vulve au
moment de la saillie (Boussit, 1989)
Couleur de la vulve Taux de femelle ovulant après
saillie
Blanche 34%
Rose 41%
Rouge 63%
Violette 73%
On note qu'entre le 3ème et le 4ème jour
post-partum, la lapine n'est pas réceptive. Le rôle
majeur de la prolactine dans cette espèce inhiberait, en début de
gestation, la croissance folliculaire (Parez, 1994). Il a été
prouvé que 48 heures de séparation de la mère de sa
portée, avant le moment prévu pour une insémination,
s'accompagnait d'une diminution du niveau de prolactine, d'une augmentation de
la concentration de 17-â oestradiol et d'une augmentation du pic de LH
déclenchée par l'injection de GnRH (Theau-Clément,
2005).
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
· Age à la première
saillie
Selon Djago et Kpodékon (2000), les jeunes femelles
doivent avoir 5 mois avant d'être saillies pour la
première fois. Le tableau II montre l'effet de l'âge à la
première saillie sur la fertilité des lapines.
Tableau II : Effet de l'âge à la
première saillie sur le taux de fertilité (Boussit,
1989)
Age à la première saillie Taux de
fertilité
- de 140 Jours 85%
140 à 149 jours 86%
150 à 159 jours 72%
160 à 169 jours 78%
170 à 179 jours 80%
+ de 180 jours 79%
La compétition entre les besoins de croissance et de
production conduit à des portées plus petites et à une
production laitière moindre (Theau-Clément, 2005).
· Prolificité
Le déficit énergétique des lapines
allaitant 6 lapereaux est moindre que celui des lapines allaitant 8 lapereaux.
Ce meilleur état corporel améliore la réceptivité
et le taux de fertilité des lapines (Castellini et al.,
2003).
1.2.1.7.2- Facteurs liés à
l'individu
· Génétique
Selon AERA (1994), les femelles de petites races sont plus
précoces (3,5 à 5 mois) que les femelles de grandes races (5
à 7 mois).
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
Moce et al. (2004) ont montré que le poids du
foetus dépendait du génotype maternel, notamment de la
capacité utérine, alors que le poids de la partie foetale du
placenta était déterminé par le génotype de
l'embryon. Ainsi les foetus sont plus lourds lorsqu'ils sont portés par
des lapines ayant une grande capacité utérine. Au contraire, la
partie foetale des placentas est plus lourde pour des embryons de
génotype « faible capacité utérine ».
· Stade physiologique
En élevage rationnel, les rythmes de reproduction
pratiqués soit intensifs (0 à 48 heures post-partum) ou
semi-intensif (10 à 12 jours post-partum) obligent la lapine
à assurer simultanément la gestation et la lactation.
Theau-Clément et al. (1990) de même que
Bourdillon et al. (1992), ont montré qu'en saillie naturelle,
le stade physiologique influence la fréquence d'ovulation ; elle est en
général plus faible chez les femelles saillies 0 à 48
heures post-partum. Dans le même temps ; les non allaitantes
présentent une fertilité appréciable de plus de 80%. En ce
qui concerne les femelles primipares, elles ont de sérieuses
difficultés à assurer pour la première fois
simultanément une gestation et une lactation (Chmitellin et
al., 1994). Si dans un rythme extensif d'élevage
caractérisé par la mise en reproduction des lapines non
allaitantes, la fertilité peut atteindre 95% (Theau-Clément
et al., 1990) ; le rythme semi-intensif ou intensif détermine
un état physiologique qui handicape l'expression d'une bonne performance
de reproduction chez la lapine.
1.2.1.7.3- Facteurs climatiques
La saison, généralement analysée en
fonction de la combinaison des effets d'éclairement et de
température, a été mise en évidence par Questel
(1984), qui a noté un effet significatif de ce facteur sur la
fertilité (64% en été vs 68% en automne). Dans
les conditions tropicales, l'effet de la température semble dominant,
mais on ne peut exclure un effet des variations de la durée du jour. On
observe une
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
réduction du taux de fertilité au cours de la
saison humide quand la température est élevée et
l'humidité ambiante forte (Lebas et al., 1996).
En ce qui concerne l'effet de la photopériode, les
travaux de Boussit (1989) ont montré que, le taux d'acceptation du
mâle est minimal sous 8 heures de lumière et maximal sous 16
heures.
Par rapport aux influences de la température, selon
Lebas et al. (1996), ce sont surtout les brusques variations de
température qui ont un impact négatif sur la fertilité des
lapines.
Enfin, une humidité relative trop basse (moins de 50%) se
traduit par une réduction des performances de reproduction (Lebas et
al., 1996).
1.2.1.8- Facteurs de variation de la
prolificité
La prolificité est déterminée par le
nombre de lapereaux par mise bas. Selon Lebas et al. (1996), elle
varie significativement en fonction de plusieurs
facteurs. la taille de portée
croît en moyenne de 10 à 20% entre la première et la
deuxième portée d'une lapine ; elle subit un accroissement plus
limité de la deuxième à la troisième portée
; elle reste stationnaire à la quatrième portée et peut
décroître ensuite.
Le taux d'ovulation est la première limite de la
prolificité. Il croît en moyenne avec cette dernière. Selon
Lebas et al. (1996), il serait lié à la race et à
la taille corporelle. Ainsi, une moyenne de 3,97 ovules a été
observée chez les races polonaises (race naine) et 12,88 ovules pour les
races géantes des Flandres ; les tailles des portées
correspondantes étant respectivement de 3,24 et 10,17 lapereaux.
Les facteurs environnementaux ont aussi une influence sur la
prolificité. D'après les travaux de Depres et al.
(1994), la saison de naissance a un effet significatif sur la taille de la
portée à la mise bas. Ainsi, il a été
observé un effet défavorable de la saison humide et chaude (Mai
à Novembre) sur les tailles de lapereaux à la naissance et au
sevrage (7,1 vs 8,2). Selon Kpodékon et Coudert (1993), la
taille moyenne des portées à la naissance est significativement
plus élevée (p = 0,009) au cours de la période allant de
Mai à Septembre, que pour le reste de l'année (5,9 vs
5,1 nés vivants par portée).
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
En ce qui concerne l'effet de la température, selon
Lebas et al. (1996), les réductions de prolificité en
ambiance chaude (30 ou 31°C) seraient moins imputables à la
température qu'à la réduction du poids corporel
entraînée par la baisse du niveau d'ingestion liée à
la température élevée. Par contre, il semble que la
mortalité embryonnaire augmente lorsque la température
dépasse 30 à 33°C mais à ce niveau, la part de la
réduction d'ingestion n'a pas été encore faite.
1.2.1.9- Facteurs de variation de la viabilité
et du poids au sevrage
L'un des facteurs conditionnant la viabilité des
lapereaux sous mère est la première tétée. Ainsi,
les travaux de Farougou et al. (2005) ont montré que les
lapereaux ayant accompli la première tétée
c'est-à-dire la prise du colostrum, sont plus viables que leurs
congénères n'ayant pas tétée. Par ailleurs, il est
important de noter que les portées de grandes tailles ont une influence
négative sur la prise du colostrum à la naissance.
L'expression du poids du jeune lapereau est
déterminée d'une part par son propre potentiel de croissance
appelé effet direct et d'autre part, par l'influence de sa mère,
appelée effet maternel qui se manifeste essentiellement par son aptitude
à l'allaitement et son instinct maternel (Garreau et Rochambeau,
2003).
Bolet et al. (1996), après une étude
réalisée sur l'influence du nombre de lapereaux allaités
sur la croissance des jeunes, concluent que deux effets s'opposent. D'une part,
ils constatent un effet négatif du nombre de lapereaux allaités
par femelle sur leur croissance et d'autre part un effet positif du nombre de
lapereaux nés sur la production laitière de leur mère, au
moins pendant les deux premières semaines.
1.2.2- Insémination artificielle chez la lapine :
définition et importance
L'insémination artificielle (IA) consiste à
déposer le sperme au moyen d'un instrument, au moment le plus opportun
et à l'endroit le plus approprié du tractus génital
femelle (Koutinhouin, 2010).
L'insémination artificielle de la lapine peut
être utile pour la planification de
chantier permettant l'obtention
d'animaux contemporains. Elle offre l'avantage de
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
grouper des naissances, de réduire du temps
passé pour faire les saillies, de diminuer le nombre de mâles, la
consanguinité et le risque de transmission des maladies. Elle permet
aussi la diffusion du potentiel génétique des animaux.
1.2.3- Méthodes d'induction de la
réceptivité sexuelle des lapines
1.2.3.1- Méthodes hormonales
Les traitements hormonaux ont été très
utilisés ces dernières années. Ils consistent à
administrer différents types et doses d'hormones, 2 à 3 jours
avant l'insémination.
· Pregant Mare Serum Gonadotropin (PMSG ou
eCG)
Cette molécule est une glycoprotéine de poids
moléculaire estimé entre 45 à 64 kD (Drion et
al., 1998). Elle est extraite du sérum de jument gravide. C'est une
hormone dimérique à double activité FSH et LH. PMSG est
utilisée depuis une quinzaine d'années pour induire et
synchroniser l'oestrus des lapines. Sur des lapines allaitantes au stade 11
jours post-partum, une injection de PMSG permet d'améliorer le
pourcentage de lapines réceptives au moment de l'insémination,
quelle que soit la dose (20 UI : Maertens, 1998 ; 25 UI : Theau-Clément
et Lebas, 1996). Une injection de PMSG avant l'insémination augmente
généralement la fertilité des lapines mais son
efficacité pourrait dépendre des conditions de traitement (dose,
mode d'injection, intervalle entre l'injection et l'insémination).
· La prostaglandine PGF2á
L'effet lutéolytique (régression des corps
jaunes) des prostaglandines PGF2á (naturelles ou synthétiques) a
été utilisé afin d'induire et de synchroniser les mises
bas ou pour induire la régression des corps jaunes de lapines pseudo
gestantes (McNitt, 1992). Un effet indirect de l'administration de PGF2á
au 29ème jour de gestation pour synchroniser les mises bas, est
l'augmentation de la réceptivité sexuelle et de la
fertilité (+ 16% Rodriguez et Ubilla, 1988), quand les lapines sont
inséminées à 7 jours post partum.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.2.3.2- Effet du mâle
Dans différentes situations physiologiques, la
présence du male influence l'équilibre hormonal et le
comportement des femelles des espèces ongulées : l'introduction
du mâle dans un troupeau peut être une méthode efficace de
contrôle de la reproduction. Chez la lapine, ce pourrait être une
méthode alternative aux méthodes hormonales pour induire la
réceptivité. Dans un travail préliminaire, l'introduction
d'un male parmi des femelles, 4 jours avant l'insémination n'a
modifié ni la réceptivité ni la fertilité des
femelles (Kustos et al., 2000).
1.2.3.3- Séparation ponctuelle de la
mère et sa portée
Pour un rythme de reproduction de 42 jours, dans une
situation d'allaitement libre, une séparation de 36 heures entre la
mère et sa portée est une alternative à l'utilisation
d'hormones pour induire la réceptivité des lapines et
améliorer en conséquence leur productivité (Bonanno et
al., 2005). Cette stimulation doit être appliquée juste avant
l'insémination qui est pratiquée immédiatement
après le premier allaitement suivant la séparation. Cependant,
cette courte privation maternelle déprime la croissance des jeunes.
1.3- AMELIORATION GENETIQUE
Les programmes d'amélioration génétique
des espèces d'animaux domestiques sont récurrents. Ils sont par
ailleurs très divers. Ils combinent deux grandes méthodes: le
croisement et la sélection. Le plan de
sélection va dépendre de la nature des caractères
sélectionnés, des paramètres de reproduction, des
modalités du contrôle des performances, de l'estimation des
valeurs génétiques et de la structure de la population (Ouyed et
Maignel, 2010). D'une manière générale, on peut distinguer
dans une population animale, soumise à une sélection, le noyau de
sélection où est créé le progrès
génétique et la population de production ou diffuse le
progrès génétique crée dans le noyau. Le noyau de
sélection est de taille très variable selon les plans de
sélection.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.3.1- Notion de sélection
La sélection, est définie comme étant le
moyen de modifier la composition génétique d'une population de
façon graduelle, à la faveur du renouvellement des
générations. Cette voie d'amélioration
génétique se situe à l'intérieur d'une même
population, sans intervention de reproducteurs extérieurs (Ouyed et
Maignel, 2010 ; Youssao, 2008). La mise en place d'un programme de
sélection efficace et durable nécessite plusieurs étapes
cruciales pour aboutir à des résultats concluants.
1.3.1.1- Objectif de sélection
Un objectif de sélection est un caractère ou
une combinaison de caractères que l'on veut améliorer. Ce
caractère peut ne pas être mesurable sur le candidat à la
sélection ou sur ses apparentés. Dans le cas de la production
cunicole à but expérimental, les objectifs de sélection
sont orientés vers les caractères précis (Ouyed et
Maignel, 2010 ; Youssao, 2008). Par exemple, l'amélioration de la
productivité numérique et pondérale permettrait
d'augmenter la capacité concurrentielle des producteurs de lapins.
1.3.1.2- Critères de sélection
Les critères de sélection sont des
caractères mesurables sur les candidats à la sélection ou
sur leurs apparentés. Ces mesures permettent alors de prédire la
performance ou la valeur génétique attendue sur les objectifs de
sélection, afin de classer les individus selon les objectifs
fixés (Ouyed et Maignel, 2010 ; Youssao, 2008). Dans le cas de la
production de lapins, les critères de sélection peuvent
être le nombre de lapins vivants à la naissance ou au sevrage, le
poids des lapereaux au sevrage, la vitesse de croissance pendant la
période d'engraissement, le poids des lapins à un âge fixe
(63 jours par exemple), le rendement en carcasse, etc. (Larzul et Gondret,
2005). Les programmes de sélection orientés vers des
critères à forte héritabilité devraient conduire
à des progrès génétiques plus rapides.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.3.1.3- Démarche générale en
sélection
Une fois que les objectifs et les critères de
sélection ont été bien définis, il est possible de
procéder à la sélection proprement dite. Le programme de
sélection doit être non seulement rigoureux mais aussi constant.
Le même objectif doit être poursuivi sur plusieurs
générations pour aboutir à des résultats
concluants. Le choix de la future génération de reproducteurs
à partir d'une population existante (n) constituée de candidats
à la sélection (Figure 2) comprend plusieurs étapes
à savoir : la collecte des informations, le calcul des performances,
l'estimation de la valeur génétique, le classement des individus
et enfin le choix des meilleurs reproducteurs parmi les candidats à la
sélection pour constituer la nouvelle génération (n+1)
(Ouyed et Maignel, 2010). La sélection des animaux peut être faite
en fonction des performances propres du lapin (sélection individuelle ou
sélection massale) ou sur les performances des ascendants
(sélection sur ascendance), des collatéraux (sélection sur
collatéraux) ou encore sur les performances des descendants
(sélection sur descendance ou testage). Depuis une quinzaine
d'années, de nouvelles méthodes d'évaluation et de
sélection des animaux ont vu le jour, notamment avec l'émergence
d'outils informatiques et statistiques de plus en plus puissants.
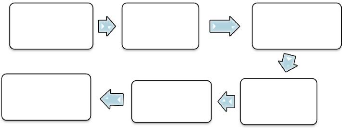
Choix
des
reproducteurs
(Génération n+1)
Candidats à la
sélection
(Génération n)
Collecte des
données
Estimation de la
valeur
génétique
Classement des
individus
Evaluation des
performances
Figure 2 : Démarche générale en
sélection (Ouyed et Maignel, 2010)
1.3.1.4- Estimation de la valeur
génétique
Cette étape est cruciale dans le processus de
sélection. La démarche de l'estimation
de la valeur
génétique fait appel à des concepts de
génétique des populations et de
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
statistiques, et bénéficie largement des
progrès récents de l'informatique. Le concept de base est que la
performance d'un animal est la résultante d'effets
génétiques et d'effets d'environnement (P = G+E) et que tout
animal transmet la moitié de ses gènes, donc de sa valeur
génétique pour un caractère donné, à ses
descendants (Ouyed et Maignel, 2010). L'évaluation
génétique, appelée couramment indexation, permet une
estimation de la valeur génétique des individus à partir
des performances mesurées sur eux-mêmes ou sur tous leurs
apparentés connus.
1.3.1.5- Méthodes de sélection
1.3.1.5.1- Sélection massale
La sélection massale consiste à choisir les
reproducteurs en fonction de leur(s) performance(s) individuelle(s). Pour
obtenir une évaluation génétique des candidats, il suffit
de contrôler des performances. Elle autorise une forte intensité
de sélection. Son principal inconvénient réside dans
l'impossibilité qu'il y a de l'appliquer à certains
caractères notamment lorsque le caractère concerné ne
s'extériorise pas chez les candidats à la sélection et
lorsque la mesure du caractère nécessite l'abattage de l'animal
(Verrier et al., 2009a).
1.3.1.5.2- Sélection sur ascendance
Elle consiste à choisir les reproducteurs en fonction
des informations dont on dispose sur leurs parents ou sur des ancêtres
plus éloignés. La mise en oeuvre de cette méthode repose
sur la disponibilité de fichier d'enregistrement rigoureux des
filiations (Verrier et al., 2009a). Cette méthode permet de
contrôler les risques de consanguinité ou d'absorption par
métissage.
1.3.1.5.3- Sélection sur descendance
La sélection sur descendance est la méthode qui
permet d'obtenir le maximum de
précision et elle est applicable pour
tous les caractères. Elle allonge notablement
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
l'intervalle de génération. Sur le plan
pratique, la mise à l'épreuve des individus de la descendance est
couteux et nécessite des infrastructures importantes et une organisation
rigoureuse (Verrier et al., 2009a). Les animaux mis à
l'épreuve doivent avoir été triés au
préalable par d'autres méthodes, plus précoces et pouvant
s'appliquer à un grand nombre de candidats.
1.3.1.5.4- Sélection sur
collatéraux
La sélection sur collatéraux consiste à
sélectionner les reproducteurs à partir de la moyenne des
performances de leurs demi- ou pleins frères-soeurs. Elle est surtout
développée dans les espèces où l'on peut disposer
de familles nombreuses : lapins, porcs,... (Verrier et al., 2009a).
Elle est en général peut précise et sa fiabilité
peut être mise en défaut en cas d'effet d'environnement commun,
phénomène relativement courant pour des pleins
frères-soeurs.
1.3.1.6- Paramètres importants en
sélection
1.3.1.6.1- Généalogie
L'évaluation génétique est basée
sur l'analyse conjointe des performances et des généalogies, et
utilise le fait que les animaux apparentés se ressemblent en termes de
performances. Les généalogies disponibles doivent être les
plus complètes et les plus précises possibles. Connaissant les
liens généalogiques entre les individus, il est facile de
calculer le taux de consanguinité.
1.3.1.6.2- Héritabilité
L'héritabilité (?2) décrit la part de
variance phénotypique d'un caractère qui est d'origine
génétique. Les valeurs de ?2 sont comprises entre 0 et 1 selon
les caractères. Plus l'héritabilité d'un caractère
est élevée, plus il se transmet d'une génération
à l'autre, et donc plus il est « facile » à
sélectionner. Dans la plupart des espèces, les caractères
de reproduction ont une héritabilité faible (<0,15), les
caractères de croissance une héritabilité moyenne (0,20
à 0,30) et les caractères de
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
composition corporelle ont une héritabilité
élevée (>0,50) (Ouyed et Maignel, 2010 ; Youssao, 2008).
1.3.1.6.3- Répétabilité
La répétabilité d'un caractère
est une mesure de la corrélation entre les performances successives d'un
même animal pour un caractère répété (par
exemple la taille des portées). Cette composante est importante à
connaître en sélection car elle a un impact sur la
précision des évaluations génétiques. Ainsi, si un
caractère est très répétable, il suffira de peu de
mesures répétées pour avoir une bonne idée de la
performance moyenne de l'animal. En revanche, pour un caractère peu
répétable, il faudra accumuler beaucoup plus de données
par animal pour avoir une estimation précise de son phénotype
(Ouyed et Maignel, 2010).
1.3.1.7- Progrès génétique
Il permet de mesurer l'efficacité de la
sélection génération après
génération. Le progrès génétique est
fonction de l'intensité de sélection, de la précision de
l'évaluation génétique, de la racine carrée de la
variance génétique additive et de l'intervalle entre
générations (Lebas et al., 1996 ; De Rochambeau, 1992).
L'intensité de sélection varie en fonction de la taille de la
population et du nombre d'animaux à conserver pour la reproduction
à chaque génération. La précision de
l'évaluation génétique dépend de la
précision des mesures réalisées sur les candidats à
la sélection, mais aussi du nombre de performances accumulées
pour chaque animal et ses apparentés. L'écart type
génétique est propre à chaque caractère, cependant
il peut être diminué dans une population trop petite et/ou
consanguine. L'intervalle de génération dépend de
l'âge des parents à la naissance de leurs descendants et est
grandement influencé par la vitesse de renouvellement du troupeau
reproducteur. Ces différentes composantes sont les principaux facteurs
influençant le progrès génétique, et donc
l'efficacité d'un programme de sélection.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.3.2- Croisement : apport potentiel du croisement,
principaux plan de croisement
Le croisement consiste à l'utilisation en accouplement
des reproducteurs issus de populations différentes de la même
espèce.
· Race pure : les reproducteurs
s'unissent au sein d'une même population.
· Hybridation : les reproducteurs
appartiennent à des espèces différentes.
Appartiennent à une même
espèce des animaux dont l'accouplement est
fertile, car ils donnent naissances à un ou plusieurs jeunes viables,
eux mêmes féconds, c'est-àdire capables de produire,
à l'âge mür, des gamètes fonctionnels. Au dessous de
l'espèce, il est possible de distinguer des sous-ensembles d'animaux
présentant entre eux un certain nombre de caractéristiques
héréditaires communes, qui se distinguent des autres
sous-ensembles : on parle de races. Pour
désigner des populations fermées sélectionnées en
vue d'objectifs précis, on parle de souches
(cuniculture, aviculture) ou de lignées
(Verrier et al., 2009b).
1.3.2.1- Apports potentiels du croisement
1.3.2.1.1- Apport de gènes nouveaux
Des races différentes présentent en
général des structures différentes relativement à
plusieurs locus et peuvent différer pour leur moyenne
génétique additive pour un ou des caractères
donnés. Dans ce cas, le croisement peut permettre de progresser sur les
caractères que l'on recherche, plus que ce n'est possible par
sélection intra-race. L'amélioration de la taille de
portée dans l'espèce cunicole illustre cet intérêt
du croisement (Verrier et al., 2009b).
1.3.2.1.2- Complémentarité entre
caractéristiques
Le croisement permet de réunir, chez les mêmes
animaux, des aptitudes complémentaires, chaque race apportant des
aptitudes spécifiques. Ceci est particulièrement
intéressant quand, du fait d'antagonisme génétique, ces
aptitudes sont difficiles à sélectionner simultanément au
sein d'une seule race. Il est par
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
exemple intéressant, dans le cadre de la production de
viande, de croiser, d'une part, des lapines présentant de bonnes
aptitudes de reproduction et de bonnes aptitudes maternelles avec, d'autre
part, des lapins présentant un bon développement musculaire
(Verrier et al., 2009b). Ici, l'intérêt du croisement
peut se manifester, soit pour les caractères à
déterminisme polygénique, soit pour des caractères pour
lesquels l'existence d'un gène majeur a été
démontrée.
1.3.2.1.3- Effet d'hétérosis
Pour un caractère donné,
l'hétérosis est définie comme la supériorité
de la population croisée par rapport à la moyenne des valeurs des
deux populations parentales. La valeur de cet effet est établie sur la
base de résultats moyens. L'hétérosis varie selon le
caractère considéré (Verrier et al., 2009b).
Cette dernière permet, par exemple, de tirer partie des aptitudes
maternelles d'une race pour produire plus de lapins, et des avantages sur la
croissance d'une autre race pour que ces lapins se rendent plus rapidement au
poids du marché. L'hétérosis correspond à un
écart à l'additivité des effets des gènes : on peut
évoquer l'existence d'une dominance orientée, voire des
phénomènes d'épistasie.
1.3.2.1.4- Accroissement de la variabilité
génétique
Le croisement entre deux races ramène
systématiquement les fréquences alléliques vers des
valeurs intermédiaires (C'est évident pour les locus où
les deux races sont homozygotes chacune pour un allèle
différent). Par ce biais, le croisement permet une augmentation de la
variabilité génétique, ce qui peut permettre un
redémarrage de la sélection (Verrier et al., 2009b). De
même, le croisement brise les liens de parenté qui ont pu
s'instaurer entre les individus d'une population fermée d'effectif
limité. L'emploi du croisement dans cette perspective doit être
raisonné en fonction d'éventuels inconvénients que cette
pratique peut receler.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
1.3.2.2- Principaux plans de croisement
Les avantages du croisement correspondent aux
inconvénients de l'élevage en race pure d'où
l'intérêt d'utiliser ces deux modes d'amélioration
génétique de façon complémentaire. On peut classer
les différents types de croisement en deux principaux groupes :
· Les croisements industriels : destinés
à produire une génération terminale d'animaux
croisés tous destinés à être abattus.
Dans ce cas, on a en permanence recours à des
reproducteurs de toutes les races parentales impliquées. Ce sont : le
croisement de première génération, le croisement à
double étage.
· Les croisements destinés à
créer ou à modifier des races.
La population résultante, généralement
après un certain nombre de génération, fonctionne de
façon autonome, c'est-à-dire sans recours à des races
parentales. Ce sont : le croisement d'absorption, le croisement
d'amélioration, la création d'une race synthétique ou
croisement de métissage.
1.3.2.2.1- Création d'une race
synthétique
L'objet de ce type de croisement est de créer,
à partir de plusieurs races ou populations, une nouvelle race dite
synthétique, dans laquelle on cherche à rassembler les
qualités des diverses races parentales. En pratique, on réalise
des croisements entre les races parentales et on accouple ensuite entre eux,
à chaque génération, les produits issues du croisement en
éliminant tous les animaux non conformes à l'objectif poursuivi.
Après plusieurs générations, quand les animaux
présentent une certaine homogénéité, on peut
considérer qu'une nouvelle race est créée et entreprendre
sa sélection de façon valable du fait d'une variabilité
génétique suffisante (Verrier et al., 2009b).
L'intérêt de ce type de croisement est de permettre, de
façon plus rapide que la sélection en race pure, d'obtenir des
combinaisons favorables de caractères. Ceci est particulièrement
intéressant quand
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
l'objectif d'amélioration correspond pour plusieurs
caractères importants à une position approximativement
intermédiaire entre les deux races parentales.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
Deuxième Partie :

Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
2- METHODOLOGIE
2.1- Milieu d'étude
L'étude a été réalisée
dans les bâtiments d'élevage du Centre Cunicole de Recherche et
d'Information (CECURI). Situé sur le campus d'Abomey-Calavi à 600
m de la route inter-états Bénin-Niger, le CECURI dispose de deux
bâtiments d'élevage de 210 m2 et de 3,5 m de hauteur
avec des parois en briques et recouverts d'une toiture en tôles
ondulées sous forme de lanterneau. La ventilation est assurée par
une ouverture en longueur au sommet du toit ainsi que par des entrées
d'air suffisantes et variées en partie moyenne et en partie basse du
bâtiment. Des ouvertures sur les pignons assurent aussi l'entrée
de l'air. Les lampes électriques disposées dans les
bâtiments permettent de prolonger la lumière jusqu'à 16
heures d'éclairage par jour. De plus, la température,
l'humidité et les paramètres de ventilation, étaient
constamment contrôlées. Les données ont été
quotidiennement enregistrées. L'un des bâtiments sert à la
reproduction et l'autre à l'engraissement. Chaque bâtiment
comporte quatre batteries de cages métalliques galvanisées,
disposées en flat-deck au dessus d'une fosse d'environ 1 m de
profondeur.
2.2- Animaux utilisés
Les lapins ont été nourris ad libitum
avec des aliments commerciaux (produits par ACP-LP) répondant aux
besoins des femelles en lactation (2.500 kcal/kg d'aliment et 18% de
protéines brutes) et des lapins en engraissement (2.375 kcal/kg
d'énergie métabolisable et 16% de protéines brutes). Les
lapins reproducteurs, mâles et femelles, ont été
logés dans des cages individuelles. Le rythme de reproduction
semi-intensif a été appliqué (saillie 10 jours
post-partum) et la palpation abdominale permettait de faire le diagnostic de la
gestation. Les femelles palpées négatives ont été
accouplées une seconde fois dès leurs entrées en
chaleur.
Chaque femelle allaitait un maximum de 6 lapereaux pour la
première portée et 7
pour les portées suivantes. Les
lapereaux adoptés ont les oreilles cochées.
Des
précautions ont été prises pour adopter des
lapereaux ayant déjà tétés au moins une
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
fois. Au total, 42 mâles reproducteurs et 140 femelles
nullipares ont été utilisés lors de la sélection.
Ils sont répartis en 10 familles de 14 femelles et en 6 familles de 8
mâles ; en plus de 6 mâles et 32 femelles hors sélection.
La première génération de croisés
(F1) a été obtenue en inséminant en Mars 2010, 104
femelles de la population locale, entretenues dans les élevages du Sud
Bénin avec la semence de mâles de souche améliorée.
Les semences ont été prélevées sur des mâles
à l'élevage expérimental de la SAGA (Station
d'Amélioration Génétique des Animaux) de Toulouse,
diluée selon la technique classique, et transportée dans des
boîtes isothermes pour être mise en place 48 heures après au
CECURI. Les mâles utilisés appartenaient à 14 familles de
souche améliorée (souche INRA 1777).
2.2.1- Caractéristiques de la Souche INRA 1777
La souche INRA 1777 est une souche améliorée
qui a été crée à partir de la souche INRA 1077. Les
caractères de portées de cette dernière ont
été de 8,5 lapereaux nés totaux et 7,4 sevrés. La
souche 1077 a été longtemps sélectionnée sur la
taille de portée de même que le poids à 63 jours. Par suite
les éleveurs et les sélectionneurs ont souhaité modifier
l'objectif de sélection de cette souche pour mieux prendre en compte les
aptitudes maternelles et améliorer le poids du lapereau au sevrage. Une
réflexion combinant le bilan génétique de la souche INRA
1077, l'estimation des paramètres génétiques de nouveaux
caractères (Garreau et al., 2003) et la prédiction du
progrès génétique pour plusieurs stratégies de
sélection, ont conduit à la création de la souche
appelé 1777. Elle a été sélectionnée ensuite
pendant 3 générations sur la taille de portée et les
effets directs et maternels du poids au sevrage en appliquant une
méthode d'optimisation du progrès génétique sous
contrainte d'augmentation maximale de consanguinité fixe à 1%
(Garreau et al., 2006).
2.3- Balance et instruments de marquage
Les pesées ont été
réalisées avec une balance électronique de marque
TERAILLON
d'une portée de 10 kg. Elle a 2 g de sensibilité
entre 0 et 1 kg ; 5 g entre 1 et 4 kg et
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
10 g entre 4 et 10 kg. Des boucles d'oreilles en plastique ainsi
que des numéros de tatouage par famille ont permis d'identifier les
animaux.
2.4- Répartition des animaux
2.4.1- Plan d'accouplement des reproducteurs en
sélection
Pour limiter la consanguinité, il a été
constitué non seulement, des familles avec des animaux provenant de
différentes lignées ou progénitures du CECURI mais aussi
certaines femelles ont été accouplées avec des mâles
d'autres élevages. Au total, 10 familles de reproducteurs ont
été constituées, tout en respectant le plan d'accouplement
de la figure 3.
A
|
A
|
A
|
A
|
A
|
A
|
A
|
A
|
A
|
V
|
V
|
A
|
A
|
A
|
A
|
A
|
V
|
V
|
V
|
?supl
|
B
|
B
|
B
|
B
|
B
|
B
|
B
|
B
|
B
|
U
|
U
|
B
|
B
|
B
|
B
|
B
|
U
|
U
|
?supl
|
?supl
|
|
C
|
C
|
C
|
C
|
C
|
C
|
C
|
C
|
C
|
U
|
U
|
C
|
C
|
C
|
C
|
C
|
U
|
U
|
?supl
|
?supl
|
D
|
D
|
D
|
D
|
D
|
D
|
D
|
D
|
D
|
T
|
T
|
D
|
D
|
D
|
D
|
D
|
T
|
T
|
?supl
|
?supl
|
|
E
|
E
|
E
|
E
|
E
|
E
|
E
|
E
|
E
|
T
|
T
|
E
|
E
|
E
|
E
|
E
|
T
|
T
|
?supl
|
?supl
|
|
F
|
|
F
|
|
F
|
|
F
|
|
F
|
|
F
|
|
F
|
|
F
|
|
F
|
|
R
|
|
R
|
|
F
|
|
F
|
|
F
|
|
F
|
|
F
|
|
R
|
|
R
|
?supl
|
?supl
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
S
S
S
H
|
H
|
H
|
H
|
H
|
H
|
H
|
H
|
H
|
R
|
R
|
H
|
H
|
H
|
H
|
H
|
R
|
R
|
?supl
|
?supl
|
K
|
K
|
K
|
K
|
K
|
K
|
K
|
K
|
K
|
S
|
|
K
|
K
|
K
|
K
|
K
|
|
|
S
|
?supl
|
|
E
|
E
|
E
|
E
|
E
|
E
|
E
|
E
|
E
|
T
|
T
|
E
|
E
|
E
|
E
|
E
|
T
|
T
|
?supl
|
?supl
|
F
|
F
|
F
|
F
|
F
|
F
|
F
|
F
|
F
|
R
|
R
|
F
|
F
|
F
|
F
|
F
|
R
|
R
|
?supl
|
?supl
|
|
H
|
H
|
H
|
H
|
H
|
H
|
H
|
H
|
H
|
R
|
R
|
H
|
H
|
H
|
H
|
H
|
R
|
R
|
?supl
|
?supl
|
K
|
K
|
K
|
K
|
K
|
K
|
K
|
K
|
K
|
S
|
S
|
K
|
K
|
K
|
K
|
K
|
S
|
S
|
S
|
?supl
|
ORIGINE : FAMILLE :
|
|
|
|
|
|
|
|
|
|
|
mâle extérieur
|
|
|
|
|
Figure 3 : Plan d'accouplement des reproducteurs en
sélection.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
Ainsi, 8 rangées de cages à la maternité
et 3 rangées de cages à l'engraissement ont été
utilisées. Les lapines ont été réparties en 10
familles à savoir les familles (A, B, C, D, E) provenant du CECURI et
les familles (F, H, K, L, M) provenant d'autres élevages. Les
mâles ont été répartis en 6 familles à savoir
les familles (P, R, S) du CECURI et les familles (T, U, V) d'origine externe.
Les animaux ont été disposés dans les rangées de
cage par famille de femelle. Ainsi dans la première rangée nous
avons uniquement des femelles de la famille A et dans la deuxième,
celles de la famille B. Egalement dans chaque rangée, on retrouve des
lapins mâles dont le rôle est d'assurer les accouplements, ainsi
que des femelles supplémentaires pour égaliser les
portées. Notons qu'une femelle ne peut être accouplée par
un male n'appartenant pas à sa rangée. Cette disposition et cette
organisation ont permis d'éviter que des animaux de même
provenance soient mis en reproduction afin de réduire au maximum la
consanguinité.
2.5- Critères de sélection des
reproducteurs
La méthode de sélection adoptée est la
sélection sur ascendance. Les index de sélection des trois
premières portées des femelles détermineront le
renouvellement du troupeau. Seuls les mâles et les femelles provenant des
parents performants ont été retenus. L'objectif de la
sélection est l'accroissement du nombre de lapereaux sevrés par
cage mère. Les critères de sélection retenus sont : la
fertilité, la taille des portées à la naissance, la taille
et le poids vif corporel des lapereaux au sevrage.
2.6- Elimination des mâles et des femelles
Les femelles sont éliminées après trois
saillies négatives ainsi que les non gestantes, 136 jours après
la première saillie. A la fin des premières mises bas, les
mâles trop légers ont été éliminés.
2.7- Matériels d'insémination
artificielle
Les matériels utilisés pour l'insémination
artificielle sont : - des vagins artificiels ;
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
- des capotes lapin ;
- des tubes de collecte ;
- des caoutchoucs (noirs en matière synthétique)
pour la protection des semences contre les chocs thermiques ;
- un pistolet Hylajet : qui permet de contrôler la dose
introduite de façon programmée et la rapidité de
l'introduction de la semence dans les voies génitales de la femelle
(0,5ml de semence par femelle) ;
- le Receptal (GnRH : Gonadotrophine Releasing Hormone);
GONASERR (Gonadotrophine sérique, lyophilisée)
à diluer dans du sérum physiologique. Il est utilisé pour
la synchronisation des chaleurs ;
- des seringues à insuline de 1 ml ;
- des thermomètres (au-delà de 45°C) ;
- des sachets de 100 gaines bleues courbées en
plastique.
2.8- Prélèvement du sperme
- On procède dans un premier temps
à des prélèvements individuels (2
récoltes de sperme par mâle). A l'aide d'un vagin artificiel, on
masturbe le male qui, après quelque minute éjacule.
- Ensuite, on dilue la semence de chaque mâle puis on
inscrit les numéros d'identification du mâle sur chaque flacon.
2.9- Insémination proprement dite
On procède au préalable à l'injection
d'une dose de 0,5 ml de GONASERR diluée dans du sérum
physiologique à toutes les femelles 48 heures avant
l'insémination. Le contenu de chaque flacon
identifié portant le numéro du mâle est fixé au
pistolet Hylajet. Le bout courbé du pistolet est introduit dans les
voies génitales de la femelle (NB : On coupe un morceau de la gaine
bleue à partir du bout courbé d'une longueur de 10 cm environ
pour protéger le bout métallique du pistolet afin d'éviter
les cas de traumatisme vaginal). A chaque pression du pistolet, une dose de 0,5
ml de semence est introduite dans les voies génitales de la femelle.
Cette méthode
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
permet la rapidité de l'opération. Une fois le
contenu du flacon vidé, on procède à l'enregistrement du
nombre de femelles inséminées. Après insémination
chaque femelle reçoit une injection intramusculaire de 0,2 ml de
Receptal.
2.10- Prophylaxie
2.10.1- Prophylaxie sanitaire
Elle consiste à la préparation quotidienne du
pédiluve à l'entrée de chaque bâtiment. La solution
est faite de CrézylR (crézylol sodique) diluée
à 5% ou de D4+R. L'intérieur des bâtiments est
balayé tous les jours. Les abreuvoirs, les mangeoires et les cages sont
lavés mensuellement à l'aide de solution désinfectante.
2.10.2- Prophylaxie médicale
Il s'agit d'une prévention contre les maladies par
usage des principes actifs et des complexes de vitamines associés
à des oligo-éléments. Ainsi, le traitement
préventif contre la coccidiose s'effectue le plus souvent à
l'aide d'anticoccidien incorporé dans l'aliment (Cycostat
66R). Le complexe de vitamine utilisé
régulièrement est AlfacerylR.
L'OxytétracyclineR 50% est souvent utilisé pour la
prévention des infections bactériennes. L'Ivomec DR
est utilisé pour le traitement des dermatoses.
2.11- Mesures effectuées et données
recueillies
Les données de base ont été
collectées à partir des fiches individuelles des femelles ainsi
qu'a partir des fiches collectives de sélections.
- La date de l'accouplement, de la palpation, de la mise bas et
du sevrage; - le numéro du mâle;
- le numéro de la femelle;
- le résultat de la palpation;
- le numéro de la portée;
- le nombre de lapins nés vivants;
- le nombre de lapins morts nés;
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
- le nombre de lapins adoptés;
- le nombre de lapins retirés;
- le poids de la portée à la naissance (lapins
nés vivants laissés sous la mère) uniquement pour les
lapereaux des femelles inséminées;
- le nombre de lapins sevrés à 35 jours;
- le poids de la portée à 35 jours (au sevrage)
;
- le poids individuel des lapins au sevrage.
2.12- Calcul des performances de reproductions
L'ensemble des données brutes recueillies ont permis de
calculer les performances de reproduction.
Les formules utilisées pour le calcul de ces performances
sont les suivantes :
· Le taux de fertilité (%) =
(Nombre total de mises bas / nombre total de saillies) x 100.
· La prolificité à la
naissance = Nombre total des lapins nés vivants (NV) / nombre
total de mises bas.
· La prolificité au sevrage =
Nombre total de lapins sevrés (NS) / nombre total de mises bas.
· La mortinatalité (%) = (Nombre
total des lapins morts nés / nombre total lapins nés) x 100.
· La mortalité naissance - sevrage (%)
= [(total lapins nés vivants - total lapins sevrés) /
total lapins nés vivants)] x 100.
2.13- Analyses statistiques
Les analyses statistiques ont été
réalisées par le logiciel SAS (1989). Les tailles de
portées à la naissance (nés totaux et nés vivants)
et au sevrage ; les poids de portée au sevrage ont été
calculés et les moyennes ont été comparés par le
test de t. Pour les tailles de portées à la
naissance (nés totaux et nés vivants) et au sevrage ; la
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
moyenne, le mode, la médiane et les quartiles ont
été calculés par la procédure Proc
univariate du SAS (1989).
Les taux de fertilité, les taux de
mortinatalité et les taux de sevrage ont été
également calculés par numéro de mise bas et les
comparaisons ont été faites par le test bilatéral
de Z en utilisant le logiciel statistica (Stastica, 2003). Le
numéro de la portée (3 niveaux et plus : 1ère
portée, 2ème portée et 3ème portée) a
été considéré. Les données des femelles
croisées avec des mâles améliorés ont
été analysées de la même façon sans tenir
compte de la parité.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
Troisième Partie :

Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
3- RESULTATS ET DISCUSSION
3.1- Performances de reproduction des femelles en
sélection au CECURI (2008)
3.1.1- Tailles moyennes de portées des nés
totaux
Le tableau III présente les moyennes des tailles de
portées en fonction des numéros de mise bas. Pour les
premières mises bas, la taille moyenne des portées a
été de 5,06#177;2,05 lapereaux et est
significativement moins élevée (P<0,05) que celles des
deuxièmes (5,81#177;2,01 lapereaux) et des troisièmes
(6,01#177;2,25 lapereaux) mises bas. La taille moyenne des portées des
deuxièmes mises bas ne présent aucune différence
significative (P>0,05) par rapport à celle des troisièmes
mises bas.
Tableau III : Tailles moyennes de portées des
nés totaux par mise bas.
N° mise de bas
|
Moyenne
|
Déviation standard
|
1
|
5,06 a
|
2,05
|
2
|
5,81 b
|
2,01
|
3
|
6,01 b
|
2,25
|
|
Les moyennes suivies d'une lettre commune dans une
même colonne ne sont pas significativement différentes au seuil de
5%.
La figure 4 montre une distribution plus ou moins
homogène entre les faibles tailles et les fortes tailles de
portées des nés totaux. Les différents paramètres
de position de la taille des portées sont indiqués dans le
tableau IV. Les quartiles découpent la distribution des tailles de
portées en quatre parties correspondant chacune à 25% des
individus. Ces quartiles sont Q1, Q2 et Q3 avec des valeurs respectives de 4, 6
et 7. Ainsi :
· le premier quartier est compris dans l'intervalle [1, 4[
;
· le deuxième quartier est compris dans l'intervalle
[4, 6[ ;
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
· le troisième quartier est compris dans
l'intervalle [6, 7[ ;
· et le dernier quartier est compris dans l'intervalle [7,
12].
La taille moyenne de portées des nés totaux est
de 5,65#177;2,2 lapereaux ; le mode et la médiane ont la même
valeur 6. Alors la plupart des effectifs (50%) sont concentrés dans les
classes : 4, 5, et 6 ; contre 25% pour les classes : 1, 2 et 3 ; puis 25% pour
les classes de 7 lapereaux et plus.
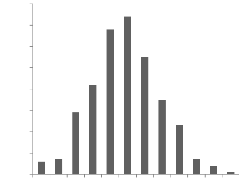
1 2 3 4 5 6 7 8 9 10 11 12
Frequences
40
80
70
60
50
30
20
10
0
Nés totaux
Figure 4 : Distribution des tailles de portées
des nés totaux (2008)
Tableau IV : Paramètres de position de la taille
des portées des nés totaux
Paramètres de position Nés totaux
Moyenne 5,65#177;2,2
Médiane 6
Mode 6
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
3.1.2- Tailles moyennes de portées des nés
vivants
Le tableau V présente les moyennes des tailles de
portées des nés vivants en fonction des numéros de mises
bas. Pour les premières mises bas, la taille moyenne des portées
a été de 4,93#177;0,19 lapereaux et est significativement plus
faible (p<0,05) que celle des deuxièmes (5,68#177;0,21 lapereaux) et
troisièmes (5,86#177;0,24 lapereaux) mises bas. La taille moyenne des
portées des deuxièmes mises bas ne diffère pas
significativement (P>0,05) de celle des troisièmes mises bas.
Tableau V : tailles moyennes de portées des
nés vivants par mise bas.
N° mise de bas
|
Moyenne
|
Déviation standard
|
1
|
4,93 a
|
2,06
|
2
|
5,68 b
|
1,91
|
3
|
5,86 b
|
2,29
|
|
Les moyennes suivies d'une lettre commune dans une
même colonne ne sont pas significativement différentes au seuil de
5%.
La figure 5 montre une distribution plus ou moins
homogène entre les faibles tailles et les fortes tailles de
portées. Les différents paramètres de position de la
taille des portées sont indiqués dans le tableau VI. Les
quartiles découpent la distribution des tailles de portées en
quatre parties correspondant chacune à 25% des individus. Ces quartiles
sont Q1, Q2 et Q3 avec des valeurs respectives : 4, 6 et 7 Ainsi :
· le premier quartier est compris dans l'intervalle [0, 4[
;
· le deuxième quartier est compris dans l'intervalle
[4, 6[ ;
· le troisième quartier est compris dans
l'intervalle [6, 7[ ;
· et le dernier quartier est compris dans l'intervalle [7,
12].
La taille moyenne des portées est de 5,53#177;2,2
lapereaux ; le mode est 6 et la médiane aussi est 6. Alors la plupart
des effectifs (50%) sont concentrés dans les classes : 4, 5 et 6 ;
contre 25% pour les classes : 0, 1, 2 et 3 ; puis 25% pour les classes de 7
lapereaux et plus.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
46
UAC/EPAC/DPSA/2010 Elysée A.
GOUDJO
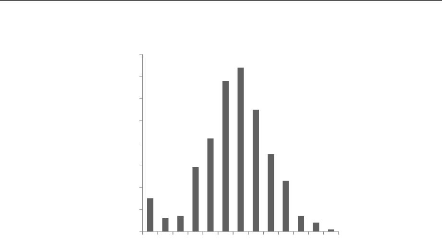
0 1 2 3 4 5 6 7 8 9 10 11 12
80
70
60
50
Frequences
40
30
20
10
0
Nés vivants
Figure 5 : Distribution des tailles de portées
des nés vivants (2008)
Tableau VI : Paramètres de position de la taille
des portées des nés vivants
Paramètres de position Nés vivants
Moyenne 5,53#177;2,2
Médiane 6
Mode 6
3.1.3- Taux de mortinatalité
Les taux de mortalité enregistrés à la
naissance par mises bas sont indiqués dans le tableau VII. Ils ont
été de 2,91%, 2,23% et 2,99% respectivement pour les
premières, deuxièmes et troisièmes mises bas. Aucune
différence significative n'a été observée entre les
valeurs enregistrées pour ce paramètre au seuil de 5%.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
Tableau VII : Taux de mortinatalité
N° de mise bas
|
Taux de mortinatalité (%)
|
1
|
2,91 a
|
2
|
2,23 a
|
3
|
2,99 a
|
|
Les moyennes suivies d'une lettre commune dans une
même colonne ne sont pas significativement différentes au seuil de
5%.
3.1.4- Tailles moyennes des portées au sevrage
La taille moyenne des portées obtenue au sevrage est
de 3,41#177;0,21 lapereaux pour les premières mises bas. Elle est de
3,63#177;0,23 lapereaux pour les deuxièmes mises bas et de 3,46#177;0,26
lapereaux pour les troisièmes mises bas. Il n'existe aucune
différence significative entre les différentes tailles de
portées au sevrage au seuil de 5%.
Tableau VIII : Tailles moyennes des portées au
sevrage par mise bas
N° mise de bas
|
Moyenne
|
Déviation standard
|
1
|
3,41 a
|
0,21
|
2
|
3,63 a
|
0,23
|
3
|
3,46 a
|
0,26
|
|
Les moyennes suivies d'une lettre commune dans une
même colonne ne sont pas significativement différentes au seuil de
5%.
La figure 6 montre une distribution bilatérale gauche
à cause de l'absence des tailles de 9, 10, 11, 12 initialement
observé chez les nés vivants ainsi qu'un pic à 0 lapereau.
Les trois quartiles découpent la distribution en quatre parties
correspondant chacune à 25% des individus. Ces quartiles sont Q1, Q2 et
Q3 avec des valeurs respectives de 3, 4 et 5. Ainsi :
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
· le premier quartier est compris dans l'intervalle [0, 3[
;
· le deuxième quartier est compris dans l'intervalle
[3, 4[ ;
· le troisième quartier est compris dans
l'intervalle [4, 5[ ;
· et le dernier quartier est compris dans l'intervalle [5,
8].
La taille moyenne des portées au sevrage a
été de 3,92#177;2,10 lapereaux, le mode et la médiane sont
respectivement de 5 et 4.
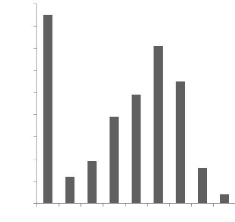
0 1 2 3 4 5 6 7 8
Frequences
90
80
40
70
60
50
30
20
10
0
sevrés
Figure 6 : Distribution des tailles de portées au
sevrage (2008) Tableau IX : Paramètres de position de la taille des
portées au sevrage
Paramètres de position Taille de portées au
sevrage
Moyenne 3,92#177;2,10
Médiane 4
Mode 5
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
3.1.5- Taux de mortalité naissance-sevrage
Les taux de mortalité entre la naissance et le sevrage
sont considérables. Ils sont présentés dans le tableau X.
Ces taux sont de 38,44%, 36,78% et 36,61% respectivement pour les
premières, deuxièmes et troisièmes mises bas. Pour ce
paramètre aucune différence significative n'a été
observée suivant les numéros de mises bas au seuil de 5%.
Tableau X : Taux de mortalité
naissance-sevrage
N° de mise bas
|
Taux de mortalité naissance-sevrage (%)
|
1
|
38,44 a
|
2
|
36,78 a
|
3
|
36,61 a
|
|
Les moyennes suivies d'une lettre commune dans une
même colonne ne sont pas significativement différentes au seuil de
5%.
3.1.6- Poids moyen des portées au sevrage par mise
bas
Les poids moyens des portées au sevrage sont
respectivement de 2203,27#177;784,51 ; 1829,11#177;867,91 et
2473,73#177;1059,30 pour les premières, deuxièmes et
troisièmes mises bas. Le poids moyen des premières mises bas est
significativement plus élevé que celui des deuxièmes mises
bas au seuil de 5%. Le poids moyen des deuxièmes mises bas est
significativement plus faible que celui des troisièmes mises bas
(P<0,05). Aucune différence significative n'a été
observée entre les poids moyens des premières et
troisièmes mises bas (P > 0,05).
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
Tableau XI : Poids moyen des portées au
sevrage
N° de mise bas
|
Moyenne
|
Déviation standard
|
1
|
2203,27 a
|
784,51
|
2
|
1829,11 b
|
867,91
|
3
|
2473,73 a
|
1059,30
|
|
Les moyennes suivies d'une lettre commune dans une
même colonne ne sont pas significativement différentes au seuil de
5%.
3.1.7- Poids moyens d'un lapereau au sevrage par mise
bas
Le tableau XII présente les poids moyens d'un lapereau
au sevrage. Les poids moyens d'un lapereau enregistrés pour les
premières et troisièmes mises bas sont respectivement de
551,66#177;147,40 g et de 533,53#177;155,01 g. Il est de 483,24#177;137,62 g
pour les deuxièmes mises bas. Il est à noter qu'il existe une
différence significative entre le poids moyen des deuxièmes mises
bas et ceux des premières et troisièmes mises bas.
Tableau XII : Poids moyens d'un lapereau au
sevrage
N° mise de bas
|
Moyenne
|
Déviation standard
|
1
|
551,66 a
|
147,40
|
2
|
483,24 b
|
137,62
|
3
|
533,53 a
|
155,01
|
|
Les moyennes suivies d'une lettre commune dans une
même colonne ne sont pas significativement différentes au seuil de
5%.
3.1.7- Taux de fertilité, prolificité et
intervalle entre mise bas
Le taux de fertilité des femelles en sélection
au CECURI au cours de l'année 2008 a
été de 65,95% et
la prolificité de 5,65 lapereaux. L'intervalle entre les
premières et
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
les deuxièmes mises bas est en moyenne de
76,16#177;24,56 jours et de 75,34#177;19 jours entre les deuxièmes et
troisièmes mises bas. Aucune différence significative n'est
observée au niveau des intervalles entre mises bas au seuil de 5%.
Tableau XIII : Taux de fertilité,
prolificité et intervalle entre mises bas
Paramètres Moyennes
Taux de fertilité (%) 65,95
Prolificité 5,65 lapereaux
IMB1 (jours) 76,16#177;24,56
IMB2 (jours) 75,34#177;19
IMB1 : Intervalle entre mise bas 1 et mise bas 2
IMB2 : Intervalle entre mise bas 2 et mise bas 3
3.2- Productivité des femelles croisées
avec des mâles de souche améliorée
3.2.1- Taux de gestation
Le taux de gestation observé après
l'insémination artificielle des femelles, toutes nullipares et d'un
poids moyen de 2263,85#177;200,76 g est de 67,31% pour la première
opération. Les femelles palpées négatives après la
première insémination artificielle ont été
inséminées une seconde fois, 22 jours après la palpation
négative. Le taux de gestation enregistré à cet effet est
de 79,41% (Tableau XIV).
Tableau XIV : Taux de gestation
N° de l'insémination Nombre de femelles Taux de
gestation
1 104 67,31%
2 34 79,41%
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
3.2.2- Tailles moyennes de portées des
croisés à la naissance
La taille moyenne de portée des nés totaux est de
4,69#177;3,03 lapereaux dont 4,55#177;2,92 nés vivants.
La figure 7 montre une distribution des tailles de
portées des croisés. Les trois quartiles découpent la
distribution en quatre parties correspondant chacune à 25% des
individus. Ces quartiles sont Q1, Q2 et Q3 avec des valeurs respectives de 2, 4
et 7. Ainsi :
· le premier quartier est compris dans l'intervalle [1, 2[
;
· le deuxième quartier est compris dans l'intervalle
[2, 4[ ;
· le troisième quartier est compris dans
l'intervalle [4, 7[ ;
· et le dernier quartier est compris dans l'intervalle [7,
11].
La figure 7 montre une distribution anormale entre les
différents taille de portées des nés vivants. Le mode et
la médiane ont la même valeur 4. Alors la plupart des effectifs
(50%) sont compris dans les classes 2, 3, 4, 5 et 6 ; contre 25% pour les
classes 0, 1 et 25% pour les classes de 7 lapereaux et plus.
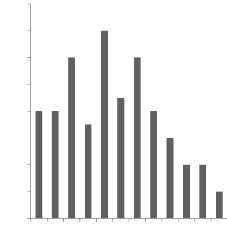
0 1 2 3 4 5 6 7 8 9 10 11
Frequences
16
14
12
10
4
8
0
6
2
Nés vivants
Figure 7 : Distribution des tailles de portées
des croisés nés vivants.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
3.2.3- Tailles de portées des croisés au
sevrage
La figure 8 montre une distribution des tailles de
portées au sevrage. Les trois quartiles découpent la distribution
en quatre parties correspondant chacune à 25% des individus. Ces
quartiles sont Q1, Q2 et Q3 avec des valeurs respectives de 0, 3 et 5. Ainsi
:
· le premier quartier regroupe 25% des tailles de
portée au sevrage de valeur 0;
· le deuxième quartier est compris dans l'intervalle
[0, 3[ ;
· le troisième quartier est compris dans
l'intervalle [3, 5[ ;
· et le dernier quartier est compris dans l'intervalle [5,
11].
La taille moyenne de lapereaux sevrés par portées
a été de 2,71#177;2,34, le mode est 0 et la médiane 3.
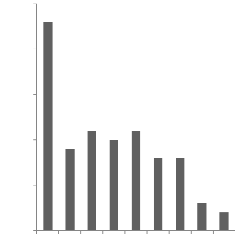
0 1 2 3 4 5 6 7 8
Frequences
25
20
15
10
5
0
sevrés
Figure 8 : Distribution des tailles de portées
des croisés au sevrage
3.2.4- Mortinatalité et mortalité
naissance-sevrage
Le taux de mortinatalité a été de 5,91%.
Entre la naissance et le sevrage, le taux de mortalité a
été de 43,69%.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
3.2.5- Poids moyens des croisés
Le tableau XV présente l'évolution du poids des
lapereaux issue du croisement entre les femelles de souche locale et des
mâles de souche améliorée. A la naissance le poids moyen de
la portée a été de 241,14#177;103,12 g. Après la
naissance, à 28 jours, le poids moyen de portée enregistré
a été de 1921,51#177;719,97 g. Les poids moyens individuels
observés à la naissance, à 28 jours puis à 35 jours
sont respectivement de 52,20#177;16,42 g, 419,66#177;136,62 g et
580,60#177;147,21 g.
Tableau XV : Poids moyens des croisés
Poids Poids
Age de la portée DS individuel DS
J0 241,14 103,12 52,20 16,42
J28 1921,51 719,97 419,66 136,62
J35 - - 580,60 147,21
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
3.3- DISCUSSION
3.3.1- Performances de reproduction des femelles en
sélection au CECURI (2008)
3.3.1.1- Tailles moyennes de portées des
nés totaux
La taille moyenne des portées enregistrée a
connu un accroissement de 12,91% entre les premières et deuxièmes
mises bas et de 3,33% entre les deuxièmes et les troisièmes mises
bas. Ces résultats sont conformes à ceux obtenus par Lebas et
al. (1996) qui ont noté que la taille de portée
s'accroît en moyenne de 10 à 20% entre la première et la
deuxième mise bas et subit un accroissement plus ou moins limité
de la deuxième à la troisième mise bas. Ceci traduit une
évolution normale des tailles de portées des lapines en
sélection.
La taille moyenne des nés totaux enregistrée est
de 5,65#177;2,21 lapereaux. Cette moyenne est supérieure à la
moyenne des tailles de portées enregistrée au CECURI de 1995
à 1998 qui est de 5,47 lapereaux. Cette observation faite serait due
à l'effet de la sélection. En effet, selon Tudela et al.
(2003), la sélection génétique a une incidence sur le
nombre de nés totaux. Ces auteurs ont obtenu 9,8 nés totaux pour
le troupeau sélectionné (souche 1077) contre 7,5 pour le troupeau
témoin (souche 9077). Par ailleurs, nous avons noté que la taille
moyenne des nés totaux enregistrée est inférieure à
celle obtenue par Hounton (2008) qui est de 6,04#177;2,04 lapereaux. Elle est
également inférieure à la moyenne de 5,73#177;1,79
lapereaux obtenue par Tanimomo (2007). Cette moyenne enregistrée en 2008
montre que les pathologies, surtout respiratoires apparues au sein du troupeau
auraient un effet négatif sur la prolificité des
lapines-mères. Mais en observant la marge existant entre la moyenne
(5,65#177;2,21 lapereaux) et les grandes tailles de portées du
quatrième quartier (7 à 12 lapereaux), nous pouvons dire que des
progrès sont encore possibles.
Toutefois les moyennes enregistrées sont largement en
dessous de celles obtenues
par Hulot et al. (1981) de même
Ouyed et al. (2007) qui sont respectivement de 9,1
et 9,32 pour la
race Californienne et la race Néo zélandaise. Ces
résultats étant
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
obtenus sur des souches européennes, des
différences significatives pourraient s'expliquer par l'influence du
type génétique maternel, notamment le taux d'ovulation sur la
prolificité.
3.3.1.2- Tailles moyennes de portées des
nés vivants
La taille moyenne des nés vivants enregistrée
est de 5,52#177;2,19 lapereaux. Parallèlement à celle des
nés totaux, cette moyenne est inférieure à la moyenne
enregistrée au CECURI par Hounton (2008) qui est de 5,84#177;2,06
lapereaux. Elle est proche de celle enregistré par Moulla et Yakhlef en
2007 sur une population locale de lapins Algériens qui est de 5,6
lapereaux. Cette moyenne montre que certains facteurs (température
élevée,...) auraient un effet négatif sur la
viabilité des lapereaux à la naissance.
3.3.1.3- Taux de mortinatalité
Le taux de mortalité est de 2,09%. Ce taux est
similaire à ceux obtenu par Tanimomo (2007) et Hounton (2008) qui sont
respectivement de 2,02% et 2,00%. Ce taux est aussi proche de ceux
enregistrés au CECURI par Kpodékon et Coudert (1993) qui ont
enregistré des taux similaires de 1991 à 1993. La
sélection n'a pas eu d'influence sur la mortinatalité. Selon
Depres et al. (1994), elle serait beaucoup plus influencée par
le stade physiologique. En effet ces auteurs ont obtenu un taux de 13,33%
lorsque les lapines sont allaitantes et un taux de 9,4% chez les lapines non
allaitantes au moment de la saillie.
3.3.1.4- Taux de mortalité naissance
sevrage
Le taux moyen de mortalité entre la naissance et le
sevrage a été de 36,61%. Ce résultat est supérieur
à celui enregistré par Akpo (2004) qui est de 21,5%. Il est au
dessus des normes (10 à 15%) indiquées dans les pays tropicaux
(Djago et Kpodèkon, 2000). Ce fort taux de mortalité
naissance-sevrage serait dû à l'apparition des troubles digestifs
dans l'élevage mais également à des maladies comme les
colibacilloses, les coccidioses et l'Entéropathie Epizootique du
Lapin.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
3.3.1.5- Tailles moyennes des portées au
sevrage
Les résultats obtenus ne montrent aucun effet
significatif du numéro de portée sur la taille moyenne au
sevrage. La taille moyenne des portées au sevrage est de 3,41#177;2,37
lapereaux. Ce résultat est proche de celui obtenu par Hounton (2008) qui
est de 3,28#177;2,37 et est inférieur à celui enregistré
par Tanimomo (2007) qui est de 4,78#177;2,18 lapereaux. Cette taille est
également inférieure à celle enregistrée au CECURI
de 1995 à 1998 qui est de 4,61 lapereaux et de 2003 à 2005 qui
est de 4,3 lapereaux. Cette moyenne rapportée à celle des
nés vivants est faible. Cette baisse de la taille de portée au
sevrage obtenue est liée aux forts taux de mortalité
naissance-sevrage. De bons résultats peuvent donc être obtenus si
l'environnement de l'élevage s'améliore, de même que si les
pathologies et les troubles nutritionnels qui entravent la vie des animaux sont
maitrisés.
3.3.1.6- Poids moyens des portées au
sevrage
Le poids moyen des portées au sevrage est de
2473,73#177;1059,3 g. Ce poids est inférieur à celui
observé par Hounton (2008) au CECURI qui est de 2830,773 g. Cette
diminution du poids pourrait s'expliquer par les abandons de portées
dues à la mort de certaines lapines-mères. Par contre, Gallal et
Khalil (1994) ont enregistré chez la race Califormienne un poids de la
portée au sevrage de 3390 g. Ce résultat étant obtenu sur
une race moyenne, des différences significatives pourraient s'expliquer
par l'influence du type génétique maternel, notamment la
capacité utérine sur le poids des foetus. Selon Moce et al.
(2004), les foetus sont plus lourds lorsqu'ils sont portés par des
lapines ayant une grande capacité utérine.
Le poids moyen d'un lapereau au sevrage est de
533,53#177;155,01 g ; ce poids est inférieur à celui
observé par Hounton (2008) qui est de 669,56 g. Il se retrouve dans la
même fourchette que ceux obtenus par Tanimomo (2007) et Akpo (2004), qui
sont respectivement de : 548#177;33,97 g et 508,97#177;19,99 g. Ce
résultat est supérieur au poids moyen obtenu par Farougou et
al. (2005) qui lors de l'étude de l'indice de la
tétée initiale sur la viabilité des lapereaux ont obtenu
un poids moyen de l'ordre de 481,3 g. Cette variation pourrait s'expliquer par
l'amélioration de l'alimentation
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
et l'effet de la sélection des femelles sur le poids
au sevrage des lapereaux. Ces résultats sont semblables à ceux de
Garreau et Rochambeau (2003), qui ont noté l'effet maternel sur le
déterminisme du poids individuel au sevrage avec une
héritabilité de l'ordre de 0,07 pour la souche INRA1077.
3.3.1.7- Fertilité et prolificité
Le taux de fertilité obtenu est de 65,95%. Il est
supérieur à celui observé par Hounton (2008) qui indique
un taux de 52,11%. Par contre, Tanimomo (2007) a observé un taux plus
élevé qui est de 69,50% au CECURI au cours de l'année
2006. Cette variation importante du taux de fertilité de 2006 à
2008 serait due à des perturbations enregistrées sur
l'état sanitaire du troupeau de sélection.
La prolificité est de 5,68 lapereaux. Elle est
inférieure à celle observée par Hounton (2008) au CECURI
qui est de 6,04. Néanmoins, des voies d'amélioration sont encore
possible. En effet, les conditions du milieu et les pathologies ont
influencé à un moment donné la productivité des
lapines.
La sélection n'a donc pas eu un effet remarquable.
Alors la poursuite de la sélection et l'amélioration des
conditions d'élevage permettraient donc une évolution à
long terme de la fertilité des lapines au CECURI. Garcia et al.
(2004) ont suggéré la possibilité d'une
sélection pour améliorer la fertilité mais avec une
espérance de progrès génétique faible. Les
héritabilités estimées sont de 0,06 pour la composante
femelle et de 0,01 pour la composante mâle.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
3.3.2- Productivité des femelles croisées
avec des mâles de souche améliorée
3.3.2.1- Taux de gestation
Le taux de gestation obtenu à la première
insémination est de 67,31%. Ce taux est plus faible que celui
rapporté par Perrier et al. (1998) sur les nullipares, qui sont
généralement très réceptives avec une
fertilité supérieure à 70%. Cette différence entre
le taux de gestation observé et celui prévu théoriquement
pourrait être lié à une faible
réceptivité des lapines malgré l'injection de
GONASERR. Toutefois, Rodriguez et Ubilla (1988),
Theau-Clément et al. (1990), Theau-Clément et Poujardieu
(1994), ont signalé que la moindre fertilité des lapines
non-réceptives au moment de l'insémination est partiellement due
à des défauts d'ovulation associée à des
défauts de gestation non liés à l'ovulation (27,1 vs
11,7 %,).
3.3.2.2- Prolificité à la naissance et
au sevrage
Les résultats montrent qu'il existe une forte
hétérogénéité entre les caractères de
la descendance. La taille des nés totaux (4,69#177;3,03 lapereaux) est
différent de celle rapporté par Lebas (1994) qui est de 6
lapereaux nés vivants par portée en tropicale. De même ces
résultats sont faibles par rapport à zone ceux obtenus par Gacem
et Bolet (2005) sur les lapines d'une population locale Algérienne
utilisée en F0 lors du croisement avec les mâles de souche
INRA2066 pour la création d'une souche synthétique qui sont de :
6,63#177;2,66 lapereaux nés totaux pour 5,95#177;2,82 nés vivants
et 4,63#177;3,19 lapereaux sevrés. Ces observations seraient dues
à un défaut de réceptivité car les lapines
réceptives ont une prolificité plus élevée que les
lapines non-réceptives au moment de l'insémination
(Theau-Clément, 1996 : + 2 lapereaux ; Rodriguez de Lara et Fallas, 1999
: + 1 lapereau). Ces auteurs affirment que, la liaison entre un niveau
élevé de réceptivité et un niveau
élevé de prolificité se retrouve quel que soit le rythme
de reproduction. Le fort taux de mortalité enregistré entre la
naissance et le sevrage (43,69%) expliquerait l'écart entre la taille de
nés vivants et des sevrés et serait due aux contraintes
d'adaptation des la souche et à l'apparition de staphylococcie des
lapereaux au nid.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
3.3.2.3- Poids moyens des croisés
Le poids moyen des lapereaux à la naissance est de
52,20 g il est de 419,66 g à 28 jours et de 580,60 g à 35 jours
d'âge. Le poids moyen à la naissance est proche de celui obtenu
par Gacem et Bolet (2005) qui est de 52,4#177;20,1 g.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
CONCLUSION ET SUGGESTIONS
Cette étude a porté sur l'amélioration
génétique des lapins de souche locale par la sélection et
par le croisement avec des mâles de souche améliorée. Elle
nous a permis d'évaluer les performances de reproduction des femelles en
sélection et des femelles inséminées.
En ce qui concerne les performances de reproduction des lapines
en sélection, nous avons noté que :
· Les tailles de portées à la naissance
sont inférieures à celles des années 2007 et 2008 mais des
progrès sont encore envisageables. De même, l'évolution des
tailles de portées au sevrage sont moins meilleures par rapport aux
résultats des années antérieures à cause du fort
taux de mortalité naissancesevrage enregistré.
· Le taux de fertilité a sensiblement baissé
par rapport à ceux enregistrés les années
antérieures alors que celui de mortinatalité a très peu
varié.
· Le poids moyen des portées au sevrage des
deuxièmes mises bas est moins élevé que ceux des
premières et troisièmes mises bas. Toutefois, le poids moyen d'un
lapereau au sevrage ne diffère pas des résultats obtenus les
années précédentes. Néanmoins des progrès
sont encore possible vue l'amélioration de l'alimentation.
· Les intervalles entre mises bas n'ont pas connu une
amélioration par rapport aux résultats des années
précédentes. Ils ont été d'environs 76 jours.
Les résultats sur les performances de reproduction des
femelles inséminées nous ont permis de constater que :
· La réceptivité des femelles semble
être faible malgré l'injection de GONASERR et serait
à l'origine du taux de gestation obtenue après la première
insémination artificielle ;
· Parallèlement au niveau de
réceptivité faible, on observe aussi un faible niveau de
prolificité ;
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
· Le taux de mortalité est élevé entre
la naissance et le sevrage en raison de l'apparition de la staphylococcie des
lapereaux au nid ;
· En ce qui concerne la croissance des lapereaux, les
résultats montrent que l'utilisation de mâles de souche 1777
permet une amélioration de la vitesse de croissance des lapereaux.
Au regard des résultats obtenus dans cette
étude, la sélection sur la taille des portées à la
naissance et la viabilité des lapereaux au sevrage pourrait au bout de
quelques années améliorer la productivité numérique
des lapines ainsi que le poids de leurs progénitures. De même il
est important de rechercher des méthodes d'induction de la
réceptivité susceptibles d'améliorer non seulement la
fertilité, mais aussi la productivité globale des lapines en
insémination artificielle.
Nous suggérons :
· Que les travaux de sélection soient poursuivis
sur plusieurs générations en adoptant des pratiques
d'élevage plus appropriées en vue d'évaluer le
progrès génétique ;
· Qu'il est important de compléter les travaux
par d'autres essais pour évaluer les performances de croissance et de
rendement de carcasse des lapins à l'abattage ;
· De poursuivre la démarche de création de
lignée synthétique car dans le contexte actuel de la production
cunicole au Bénin, elle présente des avantages, notamment pour la
création et l'approvisionnement des élevages familiaux.
· Avant d'envisager l'utilisation de cette nouvelle
souche, il conviendra de poursuivre son évaluation et de réaliser
une comparaison rigoureuse avec la souche locale afin de vérifier
notamment si cette souche peut s'adapter aux conditions climatiques locales.
.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
REFERENCES BIBLIOGRAPHIQUES
ABeC, 2007. Répertoire actualisé
des éleveurs de lapin. Cotonou, ABeC. 26 p.
AERA, 1994. La reproduction chez le lapin.
Association pour l'Etude de la Reproduction Animale. Maison Alfort : 4-11.
Akpo B., 2004. Etude de quelques performances
zootechniques de vingt familles de lapins au CECURI. Mémoire de fin de
cycle : Diplôme d'ingénieur des travaux. UAC, EPAC, Abomey-calavi,
Bénin, 34-48.
ASFC, 2008. Répertoire
actualisé des races de lapins. Association Scientifique Française
de Cuniculture. Cuniculture Magazine, Vol, (33), 56-66. [En ligne]
accès internet
http://www.noslapins.com,
consulté le 18 Novembre 2010
Boiti C., Canali C., Monaci M., Stradioli G., Verini
Supplizzi A., Vacca C., Castellini C., FacchinA E., 1996. Effect of
postpartum progesterone levels on receptivity, ovarian response, embryo quality
and development in rabbits. 6th World Rabbit Congress, 9-12 July, 1996,
Toulouse, France, Vol 2, 45-50.
Bolet G., 1998. Problèmes liés
à l'accroissement de la productivité chez la lapine. INRA
Prod Anim, Juin 1998, 11, 235-238.
Bolet G., Garreau H., Hurtaud J., Saleil G.,
Esparbié J., Falieres J., TheauClément M., Bodin L., 1996.
Canalising selection on within litter variablility of birth weight in
rabbits: Responses to selection and characteristcs of the uterus of the does.
In Proc.: 9th World Rabbit Congress, 10-13 June, 2008. Verona, Italy,
51-55.
Bonanno A., Mazza F., Di Grigoli A., Tornambè
G., 2005. Both 48-hour doe-
litter separation and 2-day controlled
suckling improved fertility of 11-day lactating
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
does similarly to 20 IU of PMSG. Cost Action 848. Joint
Scientific Meeting: Management and housing of rabbit does: reproductive
efficiency and welfare interactions. June 23-25, 2005, Palermo,
Italy.
Bourdillon A., Chmettellin F., Jarrin D., Parez V.,
Rouillere H., 1992. Effect of a PMSG treatment on breedingresults of
artificially inseminated rabbits. 5th World Rabbit Congress,
July 25-30 1992, Corvallis, USA, Vol A, 530-537.
Boussit F., 1989. Reproduction et
insémination artificielle en cuniculture. Rambouillet : Association
française de cuniculture ; 1989 : 46-82.
Brower M., 2006. Prationer's guide to pocket pet
and rabbit theriogenology., 66 :618-623
Castellini C., Dal Bosco A., Mugnai C., 2003.
Comparison of different reproduction protocols for rabbit does: effect
of litter size and mating interval. Livestock Production Science,
83: 131-139.
Chmitellin F., Rouillere R., Bureau J., 1994.
Performances de reproduction des femelles en insémination
artificielle en post partum. 5èmes Journées de la Recherche
Cunicole, 12-13 Décembre, 1990, Paris, France. Tome I,
Communications N° 4.
Colin M., Lebas F., 1995. Le lapin dans le
monde. Paris : Edition Association Française de Cuniculture. 287 p.
De Rochambeau H., 1992. Les bases de la
génétique quantitative : Le progrès
génétique et sa réalisation dans les expériences de
sélection. INRA Prod. Anim., 1992, hors série
«Éléments de génétique quantitative et
application aux populations animales», 83-86.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
De Rochambeau H., 1998. La femelle parentale
issue des souches expérimentales de l'INRA. Evolutions
génétiques et perspectives. 7èmes Journées de
la Recherche Cunicole, 13-14 mai 1998, Lyon, France. ITAVI. 3-14.
Depres E., Theau-Clément M., Lorvelec O., 1994.
Productivité des lapines élevées en Guadeloupe:
influence du type génétique, de l'allongement de la durée
d'éclairement de la saison et du stade physiologique. In: Proc.
5èmes Journées de la Recherche Cunicole en France, Tome
I, Communications N°6 et 7.
Djago Y., Kpodékon M., 2000. Le guide
pratique de l'éleveur de lapins en Afrique de l'Ouest. Cotonou :
Impression 2000. 106 p.
Drion P.V., Remy B., Houtain J.Y., Mc Namara M., Baril
G., Heyman Y., Cognie Y., Theau-Clément M., Leboeuf B., Ectors F.,
Segers K., Beckers J.F., 1998. Utilisation
répétée des gonadotropines exogènes dans le
contrôle de la reproduction : justifications, relations
structure-activité biologique, effets secondaires potentiels. Une
synthèse. Annales de Médecine Vétérinaire,
142, 373- 396.
FAO, 2000. Production et santé animales
n° 19 - Collection FAO: ISBN 92-5- 203441-2 - ISSN 0253-3731.
Farougou S., Kpodékon M., Koutinhouin B., Braih
O.H.D., Djago Y., 2005. Incidence de la tétée initiale
sur la viabilité des lapereaux. Revue Africaine de Santé et
de Production Animales, EISMV de Dakar, 3 (3-4)
218-221.
Gacem M., Bolet G., 2005. Création
d'une lignée issue du croisement entre une population locale et une
souche améliorée pour développer la production cunicole en
Algérie. 11èmes Journées de la Recherche Cunicole,
29-30 Novembre 2005, Paris, France, ITAVI. 15-18.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
Gallal E., Khalil M.H., 1994. Developpement of
rabbit industry in Egypt. In Rabbit production in hot climates. Option
méditerranéenne, Vol. 8, 43-46.
Garcia M., Sanchez J., Rafael O., Ramon J., Piles M.,
2004. Variabilité et relation phénotypique de plusieurs
caractéristiques de production et de qualité du sperme chez le
lapin. 10p.
Garreau H., De Rochambeau H., 2003. La
sélection des qualités maternelles pour la croissance des
lapereaux. 10èmes Journées de la Recherche Cunicole, 10-20
Novembre 2003, Paris, France, ITAVI. 64-69.
Garreau H., Duzert R., Tudela F., Baillot C., Ruesche
J., Grauby G., Lillelarroucau C., De Rochambeau H., 2006. Gestion et
sélection de la souche INRA 1777 : Résultats de trois
générations de sélection. 11èmes
Journées de la Recherche Cunicole, 29-30 Novembre 2005, Paris,
France, ITAVI. 19-22.
Hounton G., 2008. Evaluation des performances
de reproduction des lapines en sélection au CECURI au cours de
l'année 2007. Mémoire de fin de cycle : Diplôme
d'ingénieur des travaux. UAC, EPAC, Abomey-calavi, Bénin, 65
p.
Hulot F., Matheron G., 1981. Effets du
génotype, de l'âge et de la saison sur les composantes de
reproduction chez la lapine. Annales de Génétique et de
Sélection Animale, 13,131-150.
Kpodékon M., 1988. Le point sur
l'élevage du lapin en République du Bénin. Perspectives
d'avenir. Cuni-Science, 4 (2), 15-26.
Kpodékon M., Coudert P., 1993. Impact
d'un centre cunicole de recherche et d'information sur la recherche et le
développement de la cuniculture au Bénin. World Rabbit
Science (1993), I (1), 25-30.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
Koutinhouin B., 2010. Cours de biotechnologie de
la reproduction des monogastriques. UAC, EPAC, Abomey-calavi, Bénin, 60
p.
Kustos K., Eiben C.S., Szendro Z.S.,
Theau-Clément M., Godor J.Z.S., 2000. Effect on reproductive
traits of male presence among rabbit does before artificial insemination
(Preliminary results). 7th World rabbit Congress, 4-7 July 2000,
Valencia Spain, 161-166.
Larzul C., Gondret F., 2005. Aspects
génétiques de la croissance et de la qualitéde la viande
chez le lapin. INRA. Prod. Anim. 2005, 18 (2).
119-129.
Larzul C., Gondret F., Combes S., De Rochambeau H.,
2005. Divergent selection on 63-day body weight in the rabbit:
response on growth, carcass and muscle traits. Genet. Sel. Evol.
37 (2005) 105-122.
Lebas F., 1991. Alimentation pratique des lapins
en engraissement. Cuniculture Magazine. 6 :
273-281.
Lebas F., Coudert P., de Rochambeau H., Thibault R.,
1996. Le lapin: élevage et pathologie. Collection FAO:
Production et santé animales, N°19, FAO, Rome, 40- 120.
Maertens l., 1998. Effect of flushing,
mother-litter separation and PMSG on the fertility of lactating does and the
performance of their litter. World Rabbit Science,
Vol.6 (1), 185-190.
Matheron G., 1980: Etude comparative à
priori de deux rythmes de reproduction chez le lapin. 11èmes
Congrès Mondial de Cuniculture, Barcelone, Avril 1980.
McNitt J.I., 1992. Endocrinological approaches
for commercial rabbit production. J. Appl. Rabbit Res.,
15, 364-397.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
Moce M., Climent A., Blasco A., 2004. The
effet of divergent selection for uterine capacity on fetal land placenta
development at term in rabbits: maternal and embryonic genetic effects. J.
Anim. Sci.; 82 : 1046-1052.
Moulla F., Yakhalef., 2007. Evaluation des
performances de reproduction d'une population locale de lapins en
Algérie. 12èmes Journées de Recherche Cunicole, 27-28
Novembre 2007, Le Mans, France. 45-48.
Ouyed A., Lebas F., Lefrançois M., Rivest J.,
2007. Performances de reproduction des lapines de races pures
(Néo-Zélandais Blanc, Californien et Géant Blanc du
Bouscat) et des croisés, en élevage assaini au Québec.
12èmes Journées de la Recherche Cunicole, 27-28 Novembre,
2007, Le Mans, France, INRA-ITAVI.
Ouyed A., 2009. Évaluation du rendement
en carcasse, en muscle et du poids des différentes parties des lapins de
lignées pures et hybrides. Rapport final. 84 p.
Ouyed A., Maignel L., 2010. Mise en place d'un
programme d'évaluation génétique pour l'espèce
cunicole. Rapport final. 44 p.
Parez V., 1994. Reproduction chez la lapine.
Bull G.T.V. 43-46.
Perrier G., Theau-Clément M., Poujardeu B.,
Delhomme G., 1998. Essai de conservation de la semence de lapin
pendant 72 heures. 7èmes Journées de la Recherche Cunicole,
13-14 Mai, 1998, Lyon, France, 237-240.
Perrier G., Theau-Clément M., Jouanno M.,
Drouet J.P., 2000. Reduction of the GnRH dose and inseminated rabbit
doe reproductive performance. 7th World Rabbit Congress, 4-7 July, 2000,
Valencia, Espagne. Vol A, 225-230.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
Peeters J.E., Charlier G.J., 1984. Le complexe
entérite du lapin de chair en élevage rationnel.
Cuni-Sciences, 2 (3): 14.
Questel G., 1984. Contribution à
l'étude de la fertilité chez le lapin domestique. Mémoire
de fin de formation, INRA Paris-Grignon, France, 65 p
Rodriguez J.M., Ubilla E., 1988. Effect of
sexual receptivity on ovulation response in rabbit does induced with GnRH.
4th Congress of World Rabbit Science Association, 10-14 October,
1988, Budapest, Hungary, Tome II, 504-508.
Rodriguez De Lara R., Fallas L.M., 1999.
Environmental factors and physiological factors influencing kindling
rates and litter size at birth in artificially inseminated doe rabbits.
World Rabbit Science, 7 (4), 191-196.
Saleil G., Goby J.P., Rochon J.J., Richard F., Bochec
V., 1998. Influence des conditions climatiques sur la production du
lapin élevé en plein air. 7èmes Journées de la
Recherche Cunicole,12-13 Décembre, 1998, Paris, France.
Tome I, Communications N°6
Sellier P., 1992. La gestion des populations: la
diversité des plans d'amélioration génétique.
INRA Prod. Anim. hors serie Génétique Quantitative,
229-235.
Tanimomo E., 2007. Evaluation des
performances de reproduction des lapines en sélection au CECURI au cours
de l'année 2006. Mémoire de fin de cycle : Diplôme
d'ingénieur des travaux. UAC, EPAC, Abomey-calavi, Bénin, 58
p.
Theau-Clément M., Bolet G., Roustant A., Mercier
P., 1990. Comparaison de
différents modes d'induction de
l'ovulation chez les lapines multipares en relation
avec leur stade
physiologique et la réceptivité au moment de la mise à
la
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
reproduction. 5èmes Journées de la Recherche
Cunicole,12-13 Décembre, 1990, Paris, France. Tome I,
Communications N°6.
Theau-Clément M., Lebas F., 1994. Etude
de l'efficacité de la ciclogonine (PMSG) pour induire la
réceptivité chez la lapine. Cuniculture,
115, 21 (1), 5-11.
Theau-Clement M., Poujardieu B., 1994.
Influence du mode de reproduction, de la réceptivité et
du stade physiologique sur les composantes de la taille de portée des
lapines. 6èmes Journées de la Recherche Cunicole, 6-7
Décembre, 1994, La Rochelle, France, Vol 1, 187-194.
Theau-Clément M., Lebas F., 1996.
Effect of a systematic PMSG treatment 48 hours before artificial
insemination on the productive performance of rabbit does. World Rabbit
Science 4 (2), 47-56.
Theau-Clément M., Boiti C., Mercier P.,
Falieres A J., 2000. Description of the ovarian status and fertilising
ability of primiparous rabbit does at different lactation stage,
Proceedings of the 7th World Rabbit Congress Valencia Spain, Vol A,
259- 266.
Theau-Clément M., 2001. Etude de
quelques facteurs de contrôle de l'interaction entre la lactation et la
reproduction chez la lapine conduite en insémination artificielle.
Thèse, doctorat, Institut National Polytechnique, Toulouse, 103 p.
Theau-Clément M., 2005. Reproduction et
physiologie de la reproduction au 8ème Congrès Mondial de
Cuniculture. Cuniculture Magazine ; 32 : 38-44.
Tudela F., Hurtaud J., Garreau H., Rochambeau H., 2003.
Comparaison des
performances zootechniques de femelles parentales
issues d'une souche témoin et
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
d'une souche sélectionnée pour la production
numérique. Résumé. 10èmes Journées de la
Recherche Cunicole,. VOL 30, 46-51
Verrier E., Rognon X., Leroy G., Heams T., 2009a.
La sélection : paramètres du
progrès
génétique, comparaison des méthodes disponibles,
AgroParisTech, 71-96.
[En ligne] accès internet :
http://www.inapg.fr/dsa/uvf,
consulté le 13 Octobre 2010.
Verrier E., Rognon X., Leroy G., Heams T., 2009b.
Le croisement: apports potentiels du croisement, principaux plans de
croisement, AgroParisTech, 97-126. [En ligne] accès internet
http://www.inapg.fr/dsa/uvf,
consulté le 13 Octobre 2010.
Youssao A.K.I., 2008. Cours
d'Amélioration génétique des productions animales. UAC,
EPAC, Abomey-calavi, Bénin, 82 p.
Evaluation des performances de reproduction des
lapines en sélection et des femelles croisées avec des
mâles de souche INRA 1777 au CECURI.
| 

