à??°valuation du diagnostic par PCR directe et PCR-élisa sur les ITS des trypanosomes pathogà¨nes du bétail( Télécharger le fichier original )par Bachir SOULEY KOUATO Université Cheikh Anta Diop de Dakar - Doctorat dà¢â¬â¢état en médecine vétérinaire 2005 |

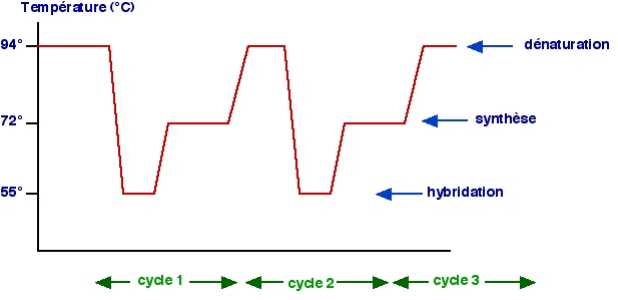
2. Méthodes2-1. Préparation des échantillons La préparation des échantillons est une étape préalable au bon déroulement de la réaction PCR, elle conditionne très souvent la sensibilité de cette réaction. 2-1-1. Echantillons d'ADN purifiés On procède d'abord à la culture des trypanosomes sur des hôtes mammifères sensibles. Sont habituellement utilisées des souris irradiées pour fragiliser leur système immunitaire et favoriser ainsi la multiplication rapide des parasites. Les prélèvements de sang s'effectuent à parasitémie très élevée. Le sang récolté est mélangé avec un tampon PSG (Phosphate Sodium Glucose), puis filtré sur une mini-colonne de DEAE-cellulose (Lahan et Godfrey, 1970). Le DEAE-cellulose ne laisse passer que les trypanosomes qui sont ensuite concentrés par centrifugation. Puis, les parasites sont lysés par congélation et traités au chelex-100. Enfin, pour confirmer l'espèce, on teste ces échantillons avec des amorces monospécifiques. 2-1-2. Traitement au chelex-100® Dans notre protocole de PCR, il n'y a pas d'extraction d'ADN à partir des échantillons de terrain. Les échantillons sont traités au chelex-100 à 5 %. Le rôle du chelex est ici de fixer les inhibiteurs de la PCR, en effet, les billes de chelex entraînent au fond du tube des éléments nuisibles à la réaction d'amplification. Le traitement consiste à triturer d'abord soigneusement les échantillons, puis à introduire dans les tubes les mêmes volumes respectifs (30 ul) de chelex-100 à 5 %. Ces tubes sont agités avec le vortex, avant d'être chauffés à l'aide d'une plaque chauffante à 56°C pendant une heure et à 94°C durant 30 minutes. Les échantillons, une fois refroidis à 4°C sont conservés à -20°C jusqu'à leur utilisation en PCR. 2-2. Principe de la PCR La PCR permet d'amplifier par une réaction enzymatique répétée, un fragment cible d'ADN balisé par un couple d'amorces, une pour chaque brin. Cette réaction repose donc sur les propriétés de l'ADN polymérase favorisant l'extension de nouveaux brins d'ADN formés. Le procédé nécessite la succession de nombreux cycles avec 3 étapes, chacune se déroulant pendant un certain temps (en général très court) et à une température bien précise. Les 3 étapes sont : Ø la dénaturation de l'ADN par ouverture de sa double hélice, l'ADN se trouve alors sous forme de simples brins (à environ 94°C) ; Ø l'hybridation des amorces sur l'ADN simple brin (de part et d'autre du fragment cible), l'une avec une séquence située en 3' sur un brin, l'autre avec une séquence également située en 3' sur l'autre brin (à environ 55°C) ; Ø l'élongation au cour de laquelle, la Taq polymérase catalyse (à environ 72°C) à partir de chaque amorce la synthèse d'un brin d'ADN (dans le sens 5'?3') en respectant la complémentarité des bases.
Figure 15 : Les étapes de la PCR et cycles thermiques 2-3. Protocole de PCR classique 2-3-1. Réalisation du mélange réactionnel Le mélange des différents réactifs de la PCR est appelé « master mix » ou mélange réactionnel, à ce mélange est ajouté l'échantillon à tester. Le volume des différents constituants le mélange réactionnel dépend du nombre de réactions à effectuer. En PCR classique, un volume final de 10 ul de mélange réactionnel est reparti dans des tubes eppendorf de 0,2 ml, on y ajoute 1ul de l'échantillon à amplifier. La concentration et le volume individuel des différents réactifs sont inscrits dans le tableau de l'annexe 5. Le volume total d'un réactif donné est obtenu en multipliant son volume individuel par le nombre de réactions. Pour palier l'incertitude sur le pipetage, on prépare une quantité de mix légèrement supérieure au nombre de réactions. Il est utile de rappeler que la Taq polymérase est ajoutée en dernière position (juste avant de distribuer le mélange réactionnel), en outre, une fois le mélange réalisé, les tubes sont brièvement agités avec le vortex. 2-3-1. Programme d'amplification L'amplification se fait l'aide d'une machine appelée « Thermocycleur » programmable à différentes températures. Le protocole de la PCR classique utilise le thermocycleur MJ Research Inc, PTC 200TM (qui peut contenir les tubes eppendorf de 0,2 ml). Le programme d'amplification initialement dénommé « programme Diag », conformément aux étapes de la PCR est composé : d'une étape de préchauffage à 94°C pendant 30 secondes ; suivie d'un cycle composé d'une étape de dénaturation à 94°C pendant 30 secondes, une phase d'hybridation à 55°C pendant un minute et une phase d'élongation à 72°C pendant une minute. Ce cycle est répété 30 fois et se termine par une élongation finale à 72°C pendant 10 minutes et une phase de réfrigération à 4°C. Dans l'objectif de réduire les hybridations non spécifiques, une optimisation de ce programme a été réalisée sur la base de la température d'hybridation des amorces qui sont spécifiques d'espèce. En faisant la moyenne de la température de fusion des 2 amorces, des programmes d'amplification propres à chaque espèce ont été conçus pour les principaux trypanosomes pathogènes du bétail fréquemment rencontrés. Il s'agit des programmes « NRP» pour T. brucei, « TV » pour T. vivax, et « TCS » pour T. congolense type savane (Tableau I). Tableau I : Programmes d'amplification de la PCR monospécifique
2-3-1. Electrophorèse et visualisation des produits PCR Les produits PCR obtenus, sont migrés sur un gel d'agarose 1,5 %. On pèse 1,875 g d'agarose qui est dissout dans 125 ml de tampon TBE (Tris Borate-EDTA), puis chauffé pendant 2 minutes dans un four micro-onde. On laisse refroidir et on y ajoute 2,5 ul (10 mg / ml) de BET (Bromure d'Ethidium). Le mélange est coulé pour polymériser dans une cuve sur laquelle sont placés des peignes, dont les empreintes sur le gel constitueront les puits de dépôt des échantillons. Un ul de tampon de charge2(*) (Loading buffer) est mélangé à 10 ul de produits PCR et déposé dans un puits du gel. De part et d'autre du gel, on dépose 3ul de marqueur de poids SL 1003(*) (Step Ladder 100, promega®) dont la taille des fragments d'ADN croit de100 à 100 bases et va jusqu'à 2000 bases. La migration se fait dans une cuve remplie de TBE (on y rajoute après le dépôt du BET), à 120 V pendant une heure. La visualisation des produits PCR se fait sous lampe UV reliée à un dispositif photographique en liaison avec un ordinateur permettant de sauvegarder l'image sur disquette. 2-4. Protocole de PCR-ITS avec les amorces Tryp 4R et Tryp 4S 2-4-1. Réalisation du mélange réactionnel Comme pour la PCR classique, le volume final des différents réactifs du master mix dépend du nombre de réactions à effectuer. Par contre, dans le protocole de PCR-ITS, un volume de 11 ul de mélange réactionnel est reparti dans des tubes de 0,5 ml. Le master mix ajouté à un volume de 3 ul d'échantillon, constitue le mélange réactionnel prêt pour l'amplification, soit donc un volume final de 14 ul. En plus, à la différence de la PCR monospécifique, ici on met dans le mélange réactionnel, le DMSO à 5% du volume total, le contrôle positif est constitué de 1 ul d'ADN associé au master mix. Afin d'optimiser les résultats de l'amplification, certains paramètres ont été variés. Ainsi, une concentration de 0,3 uM pour chacune des 2 amorces et une concentration de 0,4 UI / 10 ul pour la Taq polymérase ont donné les meilleurs résultats (Annexe 6). 2-4-1. Programme d'amplification Pour l'amplification des ITS, c'est le thermocycleur MJ Research Inc, PTC 500TM pouvant contenir les tubes de 0,5 ml qui est utilisé. Le programme d'amplification auparavant mis en oeuvre est le programme T50. Ce dernier comporte une étape de préchauffage à 94°C pendant 2 minutes, un cycle composé d'une étape de dénaturation à 94°C durant 20 secondes, d'une étape d'hybridation des amorces à 50°C pendant 45 secondes, suivie d'une étape d'extension se déroulant à 72°C pendant 30 secondes. On effectue 38 cycles au bout desquels il y a une élongation finale à 72°C pendant 2 minutes et enfin une phase de réfrigération à 4°C. Dans le but d'améliorer la sensibilité, mais aussi et surtout la spécificité de la réaction PCR avec les amorces Tryp 4R et Tryp 4S, l'optimisation du programme d'amplification a conduit à une variation en gradient de la température d'hybridation. Ainsi le programme intitulé T4RS a été finalement retenu et appliqué aux amorces Tryp 4R et Tryp 4S. Ce programme est composé d'abord d'une étape de préchauffage à 94°C pendant 2 minutes et de 40 cycles. Un cycle d'amplification comprend une étape de dénaturation à 94°C pendant 30 secondes, suivie d'une étape d'hybridation (à température variable) qui dure 45 secondes, puis une étape d'élongation à 72°C pendant 30 secondes. Les températures d'hybridation vont de 42°C à 50°C et croissent de 2 à 2°C, le nombre de cycles augmente avec l'augmentation de la température d'hybridation. On a pour 42°C et 44°C 2 cycles d'amplification, 6 cycles pour 46°C, 8 cycles avec une température d'hybridation de 48°C et enfin 22 cycles pour 50°C. A la fin des 40 cycles on a une étape d'élongation finale (à 72°C pendant 2 minutes) précédent une phase de réfrigération à 4°C. 2-4-1. Electrophorèse, visualisation et interprétation des produits PCR Les produits PCR obtenus à la suite de l'amplification avec les amorces Tryp 4R et Tryp 4S sont migrés sur un gel d'agarose selon un processus analogue à celui de la PCR classique. Il existe néanmoins quelques différences, le gel d'agarose est dans ce cas de 2%. On pèse donc 2,5 g d'agarose dissout comme pour la PCR monospécifique dans 125 ml de tampon TBE. La migration par électrophorèse, se fait aussi à 120V, mais avec une durée un peu plus longue qui est en moyenne d'une heure 30 minutes. La visualisation des produits d'amplification sur gel d'agarose est identique à tous les protocoles de PCR. L'interprétation des résultats est rendue possible par le marqueur de poids SL 100 déposé aux deux extrémités du gel. Les tailles des produits PCR avec les amorces Tryp 4R et Tryp 4S des principaux trypanosomes pathogènes du bétail sont au préalable connues (voir annexe 3). En effet, le SL 100 permet non seulement de connaître et ou de confirmer la taille des produits PCR, mais aussi d'identifier les témoins positifs (ADN des différentes espèces). Le résultat positif d'un échantillon donné se traduit par l'apparition d'une bande ou d'un signal au même niveau que le témoin positif d'une espèce donnée. On aura donc selon les cas une ou plusieurs bandes apparaissant à différents niveaux. 2-5. PCR ELISA 2-5-1. Principe et méthodes La PCR-ELISA est une méthode d'amplification d'une séquence d'ADN cible qui repose sur les mêmes principes de la PCR, mais utilisant pour sa révélation l'hybridation d'une sonde d'ADN couplée à une réaction enzymatique. Le principe général du marquage (ici non radioactif) se fonde sur la fixation d'une molécule marqueur sur l'ADN sonde selon des modalités proches de celles utilisées pour le marquage immuno-enzymatique (Kodjo et al., 1993). Deux méthodes sont habituellement utilisées pour les analyses de PCR-ELISA. Ø Dans la première méthode, les produits d'amplification obtenus avec des amorces marquées à la biotine, sont directement fixés au fond de la microplaque préalablement sensibilisée à la streptavidine. Puis les produits PCR sont hybridés par une sonde spécifique marquée à la digoxigénine ou à la fluorescéine. La présence de cette sonde est ensuite détectée par des anticorps anti-digoxigénine ou anti-fluorescéine, conjugués à l'enzyme d'un chromogène telle que la peroxydase ou la phosphatase alcaline. Après l'ajout du substrat (peroxyde pour la peroxydase et NADPH pour la phosphatase alcaline), il apparaît une coloration caractéristique pour les échantillons positifs. Ø La deuxième méthode utilisée dans la présente étude, consiste à hybrider les produits PCR marqués à la digoxigénine avec une sonde d'ADN spécifique (à l'ITS d'une espèce de trypanosome) marquée à la biotine et se trouvant fixée au fond d'une microplaque sensibilisée à la streptavidine. La révélation des produits PCR se fait à l'aide d'anticorps anti-digoxigénine conjugués à la peroxydase, après l'ajout du substrat (ABTS® ou TMB peroxydase+ H2O2). Cette méthode fait intervenir quatre étapes principales : 1). La première étape consiste au marquage des produits PCR avec la digoxigénine pendant la réaction d'amplification.
2). Dans la deuxième étape, les produits PCR sont dénaturés et hybridés avec une sonde spécifique marquée à la biotine. La sonde fixe spécifiquement une séquence interne de l'ADN cible.
3). A la 3ème étape, l'ensemble sonde spécifique marquée à la biotine hybridée avec la séquence cible d'ADN, est immobilisé dans une microplaque sensibilisée à la streptavidine. 4). Enfin à la quatrième étape, les produits PCR marqués à la digoxigénine, hybridés et immobilisés dans la microplaque, sont détectés grâce à des anticorps anti-digoxigénine conjugués à la peroxydase (Anti-DIG-POD). La révélation se fait par l'ajout du substrat (ABTS® ou TMB peroxydase+H2O2). La lecture des microplaques se fait à 450 nm au spectrophotomètre. vert Digoxigénine (DIG-DUTP) Sonde marquée à la biotine Anticorps anti-digoxigénine (Anti-DIG-POD) Microplaque sensibilisée à la streptavidine Figure 16 : Symboles utilisés pour expliquer le principe de la PCR-ELISA2-5-2. Protocole de PCR-ELISA La détection des trypanosomes dans les échantillons biologiques (ADN et échantillons de terrain) par PCR-ELISA s'effectue suivant une méthode qui comporte les étapes suivantes : la réalisation du mélange réactionnel, l'amplification de ce mélange, et enfin la révélation par ELISA des produits PCR hybridés avec les sondes. Afin de vérifier la taille attendue des produits PCR, la révélation par ELISA est précédée d'une détection des produits d'amplification sur gel électrophorétique. Cette étape est facultative dans le protocole de la PCR ELISA, elle n'est utilisée que pendant la phase expérimentale. 2-5-2-1. Réalisation du mélange réactionnel A la différence de la PCR directe, le mélange réactionnel de la PCR-ELISA contient des dNTP marqués avec de la digoxigénine sous forme de 2'-desoyuridine-5'triphosphate (DIG-dUTP). Les dTTP et les DIG-dUTP sont mélangés dans des proportions 19 / 1, avec 1,9 mM pour les dTTP et 0,1 mM pour les DIG-dUTP. Les dATP, dCTP et dGTP gardent la même molarité (0,2 mM) comme dans les autres protocoles de PCR. 2-5-2-2. Programme d'amplification Les tubes contenant le mélange (master mix et échantillon) sont agités brièvement avec le vortex, puis placés dans le thermocycleur (MJ Research Inc, PTC 500™). L'amplification se fait selon le programme T4RS décrit plus haut (cf. programme d'amplification pour la PCR-ITS). 2-5-2-3. Détection des produits PCR sur gel d'agarose
C'est une étape qui n'est pas indispensable dans le protocole de la PCR ELISA, mais elle est nécessaire, car elle permet de vérifier la taille attendue des produits d'amplification, elle permet en plus de détecter les éventuelles contaminations susceptibles d'interférer avec la révélation ELISA. L'électrophorèse est réalisée avec un gel d'agarose à 2 % dans les mêmes conditions décrites pour la PCR directe. 2-5-2-4. Révélation des produits d'amplification par ELISA La détection des produits PCR marqués à la digoxigénine et hybridés avec les sondes se déroule suivant un protocole analogue à celui de la technique ELISA. Pour chaque échantillon à tester, on prévoit deux puits de la microplaque Maxisorp® sensibilisée à la streptavidine. Les étapes de cette opération sont décrites au niveau de l'annexe 8.
Dans un premier temps, on vérifie la capacité d'hybridation des sondes en testant 3 sondes d'une espèce donnée avec l'ADN de cette même espèce. Puis cette hybridation est effectuée avec un ensemble d'échantillons constitués d'ADN, de buffy coat et d'organes de mouches testés positifs à l'espèce correspondante aux 3 sondes. On réalise deux fois ces tests à l'issue desquels, les sondes présentant les meilleurs résultats sont choisies pour la suite des travaux. Les résultats doivent se traduire par une bonne sensibilité, c'est-à-dire le maximum d'échantillons révélés positifs, mais également des densités optiques relativement élevées à la lecture au spectrophotomètre. A l'issue de ces premiers essais les sondes STV2, STCS1 et STCS2, STCF1 et STB2 ont été retenues pour respectivement T. vivax, T. congolense type savane, T. congolense type forêt et T. brucei. 2-5-4. Evaluation des sondes Avant l'utilisation des sondes retenues candidates pour le test ELISA, il convient de vérifier une fois encore leur efficacité. Il s'agit de procéder au contrôle de la spécificité et de la sensibilité des sondes sur des ADN purifiés et sur des échantillons de terrain. On réalise une PCR plurispécifique (utilisant le couple d'amorces Tryp 4R et Tryp 4S) amplifiant au total 10 échantillons d'ADN purifiés des principales espèces de trypanosomes pathogènes (4 ADN pour T. vivax, 4 pour T. congolense type savane, 1 pour T. congolense type forêt et 1 pour T. brucei). Les produits d'amplification sont hybridés avec toutes les sondes choisies et révélés par la technique ELISA. Ce test est ensuite effectué sur des prélèvements sanguins de bovins et des organes de mouches testés positifs (pour chacune des espèces de trypanosomes) à la PCR utilisant les amorces Tryp 4R et Tryp 4S. Le signal à la révélation ELISA ne doit être positif que lorsque la sonde correspond à l'ADN de l'espèce pour laquelle elle a été dessinée ou à l'échantillon de terrain infecté par cette espèce. La sensibilité de la sonde est appréciée en fonction du nombre d'échantillons détectés positifs par la technique ELISA et aussi en fonction de la valeur de la densité optique. 2-5-5. Optimisation de la technique Dans le but d'améliorer la spécificité, mais aussi la sensibilité du test ELISA, certains paramètres, dont entre autre la température d'hybridation et le volume des produits d'amplification ont été variés. La température optimale d'hybridation varie en fonction de la température de fusion (Tm), qui elle-même varie en fonction de la sonde. La température d'hybridation optimale doit être inférieure à au moins 5 à 10°C de la Tm. En dessous de ces valeurs, on observe fréquemment des hybridations non spécifiques qui sont à l'origine des faux positifs à la révélation ELISA. A l'étape d'hybridation du protocole de révélation des produits PCR, nous avons appliqué des températures allant de 37 à 63°C en fonction de l'espèce de trypanosome à détecter. Tableau II : Températures d'hybridation des sondes utilisées pour la PCR ELISA
NB : Les meilleurs résultats ont été obtenus avec une température de 60°C retenue pour toutes les sondes. Par ailleurs, le volume des produits PCR s'est avéré comme un paramètre déterminant la sensibilité de la technique de révélation ELISA. Ainsi, pour T. vivax le volume des amplifiats est passé de 5 à 7 ul, pour les autres taxons de trypanosomes le volume reste égal à 5 ul. Enfin, la fixation de la sonde marquée à la biotine (avec le tampon d'hybridation porté à une température de 60°C) dans la plaque avant d'y rajouter le mélange produit PCR / NaOH a considérablement amélioré les résultats du test.
* 2 Le tampon de charge ou tampon de dépôt est constitué de glycérol et d'un colorant (Xylène et bromophénol). Le glycérol entraîne l'échantillon au fond du puits et le colorant permet de visualiser le front de migration pendant l'électrophorèse. * 3 Le SL 100 permet de connaître la taille des produits obtenus pendant la lecture du gel. |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||