
Année Académique 2010 - 2011
UNIVERSITE DE KINSHASA
B.P. 190/ Kinshasa XI

FACULTE DES SCIENCES
DEPARTEMENT DE
BIOLOGIE
Laboratoire de Botanique Systématique &
d'Ecologie végétale


Par
Anicet MALOM DEBENCE
Mémoire présenté et défendu en vue
de l'obtention du Grade de Licencié en Sciences Groupe : Biologie
Option : Ecologie
Directeur : Professeur Félicien Lukoki Luyeye Encadreur :
Assistant Pisco Menga
Dédicace
Gloire soit renatue à Dieu créateur, maître
ate temps et ate
circonstance, pour nous avoir protégés et
nous accompagnés jusqu'à
la réalisation ate ce
travail.
A mes parents Anicet MALOM et Rhita MAYWALA
A notre caatet
Jonas MALOM
A toute la famille LEPINA
A tous mes collègues
Et
à tous mes proches collaborateurs
Anicet MALOM DEBENCE
AVANT-PROPOS
L'élaboration d'un tel travail nous a paru au
départ relativement aisé. Mais à chaque jour qui passait,
nous nous sommes rendu compte que ce genre de travaux n'étaient pas si
facile à réaliser et, surtout pendant un temps relativement
court. Notre aventure sur le terrain a duré du 01 juillet 2010 au 07
juin 2011 pour les deux sites, soit 12 mois d'observation, dont le transport et
toute la logistique furent assurés par l'Assistant Pisco MENGA.
La présente étude est essentiellement
phénologique, à laquelle nous nous sommes efforcés
d'appliquer quelques méthodes usuelles modernes, utilisées dans
la plupart d'études réalisées en Afrique centrale. Afin
d'tre pragmatique sur le terrain, nous avions mis en place deux équipes
de travail : (1) la première basée à Bolobo était
composée de trois personnes ayant déjà prouvé leurs
compétences dans les études phénologiques en travaillant
de 2007 à 2010 aux côtés de l'Assistant Pisco MENGA. Il
s'agit de Mr Bedel IBOTO, chef d'équipe, assisté de deux
personnes locales, en la personne de Mr BESAU MUNDELE et Mr Denis MPENGE.
L'extraordinaire sens de l'orientation de l'équipe de WWF Landscape Lac
Ntumba s'est révélé un atout précieux lors des
ouvertures de nouveaux sentiers et aires d'études de certaines stations
remarquables; (2) la seconde équipe basée à Kinshasa
était constituée de deux personnes, l'Assistant Pisco MENGA et de
moi-même.
Dans le présent travail, nous avons voulu expliquer de
façon détaillée notre manière de procéder
afin que le lecteur puisse juger de la confiance qu'il peut avoir dans nos
données de base. Dans la plupart des travaux phénologiques pour
l'Afrique tropicale et équatoriale, les renseignements sur la
méthodologie sont rares et disparates. Une heureuse exception est «
Conservation en forêt pluviale - Méthodes de recherche
» de White et Edwards (2001), qui fournit beaucoup d'informations sur
la méthodologie adoptée.
Avant-propos
Cette étude permet de comprendre le rythme
phénologique de Millettia laurentii (Wengé) au cours
d'une année et les variations de la fructification avec le
diamètre.
Millettia laurentii est une essence fortement
exploitée en République Démocratique Congo, du fait des
propriétés technologiques de son bois, de sa couleur et de sa
dureté. Il s'agit en fait d'étudier la variabilité
temporelle, interindividuelle, et intersites des événements
phénologiques de Wengé, afin de juger de la
régularité phénologique (débourrement,
sénescence, durée de la saison de végétation).
Les résultats de cette étude ont des fortes
implications pour la gestion de cette espèce qui constitue actuellement
l'une des essences les plus exploitées de la RD Congo avec un volume
d'exportation équivalant à 73 800 m3 en 2008 (DGF,
2009).
Notre travail se subdivise en quatre grands chapitres : le
premier est une introduction, dans laquelle nous étalons notre
problématique, nos objectifs et les hypothèses, le
deuxième chapitre décrit les matériel et méthodes
d'étude, le troisième expose les résultats de nos
investigations, et le quatrième fait une discussion des
résultats. Une conclusion clos notre étude.
Cette étude est le fruit d'une précieuse et
généreuse collaboration de plusieurs personnes qui nous sont
chers, ainsi nous sommes reconnaissants, au terme de ce travail de leur
exprimer ici notre vive gratitude ; qu'il nous soit permis de remercier
spécialement l'Eternel Dieu, pour sa grâce divine.
Nous sommes reconnaissant aux autorités
Académiques, aux Professeurs, Chefs de travaux et Assistants de
l'Université de Kinshasa en général, et en particulier,
ceux de la Faculté des Sciences, Département de Biologie, pour
leur formation intégrale durant notre parcours universitaire.
Nous sommes très redevables envers le Professeur
Félicien LUKOKI LUYEYE, Directeur de ce travail, à qui revient
l'initiative de nous avoir dirigés et surtout en insistant sur
l'orientation qu'il faut donner à ce travail ;
Nous pensons à l'Assistant Pisco MENGA MUNKOLO pour
nous avoir encadré et qui dans l'ombre, nous a rendu des services
inestimables et inoubliables qui nous ont épargné des
sérieux faux pas ;
Nous exprimons notre gratitude au Professeur Dr BIANDA et Aux
Religieux du Centre spirituel MANRESA de Kimwenza pour nous avoir
accepté de faire les recherches durant une année dans leurs
concessions ; particulièrement aux Techniciens qui nous ont
accompagnés durant nos observations ; sans oublier notre équipe
phénologique à Bolobo, dont nous citons : Mr Bedel IBOTO, chef
d'équipe, Mr Beseau MUNDELE et Mr Denis MPENGE. Qu'ils trouvent ici
l'expression de notre gratitude ;
Nous remercions les Professeurs Roger MAVINGA du
Département des Sciences Administratifs, René BIBAKA de
l'Institut supérieur de Kikwit à Bandundu, Constantin LUBINI, du
Département de l'Environnement, et tous les Chefs de Travaux et
Assistants du Département de Biologie pour leurs contributions.
Il nous sera ingrat de ne pas relever la solidarité et
le soutient du service de climatologie de Kinshasa (METELSAT)
particulièrement à Monsieur MBALA pour nous avoir disposé
les données climatiques de Kinshasa et de Bandundu.
Nous tenons à remercier tout le comité directeur
de L'ONG Solidarité Commune « SOCO » en sigle, pour leurs
assistance et suggestions, particulièrement au président national
de cette ONG Monsieur Elysée BOKUMUANA MAPOSO, pour son soutien
financier et à qui nous sommes très reconnaissants.
Nos remerciements les plus sincères s'adressent
à la famille: MALEMA Adolphe, MALOM Anicet, LEPINA Claudine, KALAMA
Raul, MENGA Suzanne, MUNTUNTETE Van, et la famille de Docteur LEPINA Princesse
pour leur soutien moral et financier.
Nous sommes reconnaissants à tous les membres de ma
famille :mes oncles, mes frères, cousins, s~urs, neveux qui m'ont
toujours approché par leur amour fraternel dans toutes les circonstances
de ma vie ; nous citons : Jonas MALOM , Princesse MALOM, Nyota MALEMA,
Éphraïm MALEMA, Prince LEPINA, Sandrine MASOLE, Dorcas KALAMA,
Christelle YAMVWA, Noël MENGA, Didier MALEMA et tant d'autres dont les
noms ne sont pas repris ici ; qu'ils trouvent l'expression de notre franche
affection.
Il est évident que dans un réseau de relations
amicales, un cachet spécial soit adressé à tous nos
ami(e)s et connaissances : Jean Louis KOYAGIALO , Olivier MBAYO, Octavie
MOSUMBE , Tychique SINZIDI, , Trésor KASONGO, Percy PAKAY, Jean Pierre
ILENDA, Jean Pierre APANDANI, Daddi KANIKA , Carine OTUNGA, l'informaticien
Julio WELE, Lydia MANDUNDU, Ticckete MUKULU,, APISCO KEBA, Gedeon BONGO, SAM
BUTOTO, Adeline ESTE , pour leur sympathie et attachement remarquable et
à qui nous exprimons notre profonde reconnaissance.
Table des matières
DEDICACE I
AVANT-PROPOS I
REMERCIEMENTS IV
TABLE DES MATIERES VI
CHAPITRE I. INTRODUCTION 1
I.1. PROBLEMATIQUE 1
I.2. JUSTIFICATION DU CHOIX DU SUJET 2
I.3. HYPOTHESE 3
I.4. OBJECTIF 4
I.5. GENERALITES SUR LA PHENOLOGIE 4
I.5.1. Brève historique 4
I.5.2. Qu'est-ce que la phénologie ? 4
I.5.3. Les phénophases 5
I.5.4. Pourquoi observer la phénologie? 7
I.5.5. Quels facteurs influencent l'évolution
phénologique des plantes? 7
CHAPITRE II. MATERIEL ET METHODES D'ETUDE 9
II.1. MILIEU D'ETUDE 9
II.1.1. Milieu physique 9
II.1.2. Milieu biotique 12
II.2. BREVE DESCRIPTION DE L'ESPECE 14
II.3. METHODOLOGIE D'ETUDE 16
II.3.1. Installation des circuits phénologiques
16
II.3.2. Prélèvements des données
16
II.3.4. Traitement des données 18
CHAPITRE III. RESULTATS 21
III.1. PHENOLOGIE VEGETATIVE DE M. LAURENTII 21
III.2. PHENOLOGIE REPRODUCTRICE DE M. LAURENTII 22
III.2.1. Phénologie de la floraison et de la
fructification 22
24
III.2.2. Fructification et diamètre 24
CHAPITRE IV. DISCUSSION 27
4.1. LA PHENOLOGIE DE MILLETTIA LAURENTII EST LIEE A LA
SAISONNALITE DES PLUIES 27
4.2. MILLETTIA LAURENTII POSSEDE UNE FRUCTIFICATION
PRECOCE 28
CONCLUSION 30
REFERENCES BIBLIOGRAPHIQUES 31
Chapitre I. Introduction
I.1. Problématique
Le fonctionnement des écosystèmes forestiers de
la République Démocratique du Congo demeure complexe et
insuffisamment connu. L'importante diversité spécifique dont
regorgent ces écosystèmes limite la compréhension de leur
fonctionnement et in fine leur gestion durable (Blanc, 2002 ; De
Wasseige et al., 2009). Toutefois, on dénombre une grande
diversité végétale, soit un peu plus de 10.000
espèces d'Angiospermes, dont environ 3.000 sont endémiques
(ATIBT, 2006).
Le marché international du bois étant
très sélectif, l'exploitation du bois est alors focalisée
sur un petit nombre d'essences de grande valeur commerciale (environ 30) alors
que le pays dispose d'un potentiel important évalué à 78
essences exploitables (Malele, 2005). Renseigner l'autoécologie de
chacune de ces espèces exploitables est une première étape
essentielle à la gestion de ces écosystèmes, et plus
particulièrement dans l'établissement des plans
d'aménagement spécifique (Malaisse, 1967 ; Differt, 2001), C'est
alors que nous avions choisi d'étudier Millettia laurentii De
Wild.
Millettia laurentii constitue durant ces cinq
dernières années, l'une de deux essences les plus
exploitées en RDC. Le volume d'exportation a atteint 73.782
m3 en 2008 pour une valeur de 13.087.392,97€ (DGF, 2009).
Devant les exigences actuelles du marché international des bois
tropicaux, plusieurs études consacrées aux structures de
populations d'arbres en forJt tropicale, abordent en premier lieu les aspects
dendrométriques, phénologiques et spatiaux (Traissac, 2003;
Boyemba, 2011 ; etc.), car ils constituent l'une des bases de la connaissance
des populations et de la gestion des peuplements forestiers (Goreaud, 2000).
Les observations phénologiques sont des données
importantes pour adapter les pratiques d'exploitation à
l'écologie des espèces exploitées, notamment la
période et le diamètre d'exploitation. La plupart
d'espèces africaines exploitables produisent de nouvelles feuilles et
fleurs par pics plutôt que de manière continue au cours de
l'année (Debroux, 1998 ; Doucet, 2003 ; Kouadio, 2008 ; Yalibanda,
1999). Les connaissances relatives à la phénologie des essences
commerciales des forêts denses africaines restent encore très
parcellaires de nos jours.
Les données disponibles sont souvent dispersées
dans des sites différents, réalisées avec des
méthodes disparates ; l'effort d'échantillonnage est
généralement faible et les périodes de temps couvertes
sont discontinues (Favrichon, 1997).
Il s'avère aujourd'hui nécessaire d'enrichir les
connaissances sur la phénologie d'une espèce exploitée
comme Millettia laurentii, en particulier la phénologie de la
reproduction.
Le diamètre de fructification d'une espèce
exploitée est une information fondamentale à l'élaboration
des plans d'aménagement. Le diamètre de fructification
régulière (DFR) d'une essence est très capital pour le
maintien de semenciers potentiels devant assurer la
régénération de la ressource ligneuse dans une forêt
en cours d'exploitation. Il est l'une des conditions supplémentaires
à la détermination du diamètre minimal exploitable (DME),
car dans le cas où le DME en vigueur se révèle
inférieur au DFR, il sera nécessaire de l'augmenter (d'au moins
10 cm au DFR) pour que les conditions de fructification soient également
respectées (Durrieu de Madron & al., 2003 ; Kouadio, 2008).
Cependant, le DFR de plusieurs essences est relativement méconnu en
forêt naturelle (Durrieu de Madron & Daumerie, 2004). Pour ce qui est
connu, les valeurs varient fortement entre espèces, et parfois entre
sites pour une mrme espèce. A titre d'exemple, Doucet (2003) trouve un
DFR de 45 cm pour Lophira alata au Gabon et Yalibanda (1999) en trouve
de 35 cm au Cameroun. Sepulchre & al. (2008), lors d'une
étude au Gabon, ont déterminé le DFR de Millettia
laurentii à 45 cm, une valeur également retenue par Gillet
& al. (2008), dans un rapport issu d'une étude de 2 ans en
République du Congo.
I.2. Justification du choix du sujet
L'aménagement en forJt dense tropicale se heurte
à de nombreuses difficultés, parmi lesquelles la
méconnaissance des caractéristiques écologiques de chaque
essence (phénologie, diamètre de fructification,
tempérament, dynamique, exigences pédologiques et climatiques,
lge optimum d'exploitabilité, etc.). Ces caractéristiques
constituent l'obstacle le plus important pour la compréhension de leur
fonctionnement. En plus, les multiples interactions des espèces entre
elles et avec leur milieu font de ces forêts des
écosystèmes extrêmement complexes (Traissac, 2003).
Devant une telle complexité et du fait que le
marché international du bois est très sélectif,
l'exploitation du bois africain est alors limitée à quelques
espèces seulement, une des démarches adoptée est
l'étude de l'écologie d'espèces cibles, choisies pour un
intérTt commercial et/ou écologique, en vue de
l'établissement des plans d'aménagement spécifique. C'est
ce qui justifie le choix du Millettia laurentii dans la
présente étude, l'une des essences les plus exploiter en Afrique
centrale et particulièrement en RDC, à cause des
propriétés technologiques de son bois très
recherché sur le marché tant national qu'international.
Millettia laurentiii est une essence dont les
connaissances restent jusqu'à ce jour très parcellaires
(Algoët, 2008). Plusieurs inconnues persistent quant à son
écologie (Tchinda, 2008) : sa phénologie, son diamètre de
fructification efficace, sa vitesse de croissance, les conditions optimales de
sa régénération, les caractéristiques de son
environnement immédiat, etc. Pour un pays qui compte jusqu'à
présent sur l'extraction de cette ressource, son utilisation et sa
valorisation deviennent un enjeu majeur de développement. La
connaissance de sa phénologie et de son diamètre de
fructification permettra aux gestionnaires forestiers d'orienter de
manière objective un plan d'aménagement adapté à
l'espèce, pour que son exploitation présente et future soit
durable.
I.3. Hypothèse
Le comportement phénologique de Millettia laurentii
est étroitement liée à la saisonnalité des
précipitations (selon qu'elle est proche ou loin de l'équateur)
et à la lumière (selon qu'il est en forit dense ou en milieu
rudéral).
Quelques questions auxquelles nous devrons répondre
à travers cette étude sont notamant :
- Quels sont les différentes périodes de
phénophases de M. laurentii dans les sites
sélectionnés? La différence entre sites est-elle
significative?
- Quelles proportions d'individus produisent des fleurs et
fruits ? Durant quelles périodes de l'année et suivant quelles
quantités ? Existe-il une différence significative entre le
nombre de tiges productrices et la quantité de fleurs ou fruits produits
?
- Quel est le diamètre minimal de fructification ?
Pour répondre à ces différentes questions,
nous sommes résolus de faire 2 sentiers phénologiques dont les
descriptions sont faites dans les lignes qui suivent.
I.4.Objectif
Cette étude a pour objectif de caractériser les
phénophases de Millettia laurentii au cours d'une année
et les variations de la fructification en fonction du diamètre afin de
fournir aux gestionnaires forestiers des recommandations sur les mesures de
gestion de l'espèce.
I.5. Généralités sur la
phénologie
I.5.1. Brève historique
Les hommes s'intéressent aux rythmes saisonniers des
événements naturels depuis bien longtemps. Les plus vieux
enregistrements phénologiques connus dans le monde remontent à
l'an 812 ! Il s'agit de la date de la floraison des cerisiers de Kyoto (Japon).
Les Chinois et les Romains faisaient déjà, sans le savoir, de la
phénologie en agriculture il y a plusieurs milliers d'années, par
l'utilisation de calendriers phénologiques. On y trouve la description
de phénomènes naturels, mais on peut aussi en tirer des
règles pour le futur basées sur les expériences du
passé, celles-ci sont utiles pour prévoir le temps ou pour
estimer les récoltes. La phénologie est devenue une science
lorsqu'on a commencé à relever les dates auxquelles certains
phénomènes naturels se reproduisent chaque année (Mathieu,
2005).
La phénologie est un mot du langage scientifique,
d'origine récente, né à la fin du 19ème
siècle. Le terme a été proposé par le botaniste
belge Charles Morren en 1853. C'est à Karl Von Linné, qui fonda
en 1750 un réseau d'observations phénologiques en Suède,
que l'on doit le départ d'une véritable science de la
phénologie destinée à étudier les modifications
périodiques subies par les organismes vivants au cours des saisons
(Differt, 2001). Ce terme fut réutilisé dans les années
1950 par l'allemand Fritz Schnelle, le fondateur du grand réseau
d'observations phénologiques du service météorologique
allemand. Beaucoup de naturalistes ont prité attention à ce qui
se passait autour d'eux et enregistré des événements
phénologiques, comme la famille Marsham, une grande famille anglaise,
qui a noté quotidiennement des données phénologiques sur
des oiseaux et sur les plantes, entre 1736 et 1925 (soit 189 ans des
données !). Ces enregistrements sur un même lieu et sur une longue
période constituent des « séries
phénologique » (Mathieu, 2005). De nombreuses
séries de ce type ont existé dans la première
moitié du 20ème siècle dans la zone
tempéré de l'hémisphère Nord. Mais les observations
n'ont pas perdurées longtemps, elles avaient pour but d'utiliser la
phénologie comme marqueur des conditions météorologiques
en vue d'application pour la recherche météorologique (Fritz
Schnelle, 1955).
Malgré cet important héritage historique,
l'observation et la compréhension des phénomènes
phénologiques sont peu à peu tombés en
désuétude. Ce n'est que depuis récemment que l'on reparle
de la phénologie, dans le contexte de changement climatique.
I.5.2. 4 XV-14-ce que la phénologie ?
La vie des plantes est rythmée par des
évènements périodiques, qui se produisent chaque
année sensiblement à la mtime période, en fonction des
saisons : la plante au sortir de mauvaise saison commence par débourrer
(ouverture des bourgeons), puis les feuilles se développent, la plante
fleurit, fructifie, et meurt ou se met au repos avant la mauvaise saison
suivante. / l ANFFIMIRQUdIEFIsEpYqQIP IQtsTIst FI ETARQUESSIOOI Oa «
phénologie » et chaque étape du
développement de la plante (le débourrement, la floraison,
ODIUFtiIIFDARQ, O'VIEIIQatIRQ, IIF.) IstNNQ « stade
phénologique », Ce stade phénologique est
également appelé « phase
phénologique ou phénophase ». La
phénologie observe donc les phénomènes typiques au cours
de l'année, que l'on nomme phénophases (Mathieu, 2005).
Les évènements phénologiques se
reproduisent chaque année à la mtime pSRGuI, EP Ill OIIrMLII IT
EF\I ISITt YariIr d'uQI aQQpI O'paI, IFEITIOsAsRQt IRLIIP IQt influencés
par les conditions environnementales et en particulier par les facteurs
climatiques. &IIsANaiQsILTXILSlusieurs auteurs ont tenté de
définir la phénologie suivant leurs expériences de
terrain. Nous retenons les définitions suivantes :
- la phénologie végétale est l'étude
scientifique des variations saisonnières, de la croissance et du
développement des plantes (Fritz Schnelle, 1955) ;
- la phénologie étudie les relations entre les
phénomènes climatiques et les caractères morphologiques
externes du développement annuel des végétaux (Galoux
& al., 1967) ;
- la phénologie IlVO'palIIdII YEUDIRQs EQQuIOOII
IINpYqQIP IQts SplIRGIDIKIIOI YiIE végétale (tels que
feuillaison, floraison, fructification, jaunissement et défeuillaison),
le plus souvent annuels et liés à des variations
saisonnières de certains paramètres climatiques
(disponibilité en eau, température, etc.), (Mathieu, 2005) ;
De façon globale, la phénologie (dont l'origine
étymologique est phénoménologie) désigne, au sens
large, l'ensemble des particularités morphologiques du cycle de
développement d'un végétal, avec mention des
époques de l'année correspondantes. Au sens strict, c'est
l'étude des relations entre les phénomènes climatiques et
les caractères morphologiques externes du développement des
végétaux (Delpech et al. 1985). Par
développement, on entend toute modification qualitative dans la forme de
la plante (Durand 1967).
Le champ d'étude de la phénologie consiste
à enregistrer, dans le temps, le retour des étapes de croissance
et de développement des titres vivants et à étudier les
facteurs qui les influencent (Defila & Clot, 2000 in Differt,
2001).
I.5.3. Les phénophases
- La feuillaison : est le processus
permettant l'apparition et le développement du
feuillage, depuis le
bourgeon dormant jusqu'à la feuille adulte (COMPS & al.
1987).
- La sénescence : la fin de
la période de végétation est marquée par une
période de sénescence du feuillage, ou défoliation. Il
s'agit d'un processus débutant par l'apparition de la coloration
automnale, et aboutissant à la chute des feuilles.
- La défoliation (ou défeuillaison)
: au sens strict, est la chute des feuilles. La prise des
couleurs automnales, variant d'une espèce à l'autre, marque la
fin du cycle annuel de croissance et de fonctionnement des feuilles, (MALAISSE
1964). Pour le Wengé, par exemple, on observe le passage du vert au
jaune puis au brun, et enfin la chute des feuilles.
- La floraison : est le
processus permettant l'apparition et le développement des fleurs depuis
le bouton floral jusqu'à l'éclosion et chute de la fleur. Les
figures 1 et 2 montres des plantes en fleurs et gousses de Millettia
laurentii.

Figure 1 : Observations de feuilles et fleurs dans le site
de Kinshasa (Source : Photo Anicet MALOM)
-
- La fructification : est le
processus permettant l'apparition et le développement des fruits depuis
leur apparition jusqu'à leur chute.

Figure2 : quelques gousses de wenge dans le site de
Mont-Ngafula (Source : Photo Anicet MALOM )
I.5.4. Pourquoi observer la phénologie?
Les études phénologiques peuvent être
utilisées à différentes fins. Depuis de nombreuses
années, les observations phénologiques constituent des
données très importantes pour la compréhension du
fonctionnement des écosystèmes forestiers et en particulier pour
la croissance des arbres, car elles enrichissent la connaissance de
l'autécologie des essences (Malaisse, 1967 ; Differt, 2001). De plus,
elles constituent un outil nécessaire pour les améliorateurs
(choix des provenances selon la précocité ou la tardivité
du débourrement), (Vernier & Teiser du Cros, 1996 in
Lebourgeois & al., 2002). Enfin, elles sont depuis peu
employées afin d'étudier la réponse de la
végétation au changement climatique. En effet, à
l'échelle de temps, les données phénologiques sont
également de bons indices des effets d'un éventuel changement
climatique sur la végétation, car la température de l'air
et la disponibilité en eau ont une influence importante sur les dates de
début des phénophases (Fady, 1991).
Pour le Wengé, elles nous permettent de connaître
les variations de rythmes saisonniers (feuillaison, floraison, fructification,
jaunissement et défeuillaison) en fonction du climat et de la
région, les variations interannuelles des dates d'apparition des stades
phénologiques.
I.5.5. Quels facteurs influencent l'évolution
phénologique des plantes?
La phénologie dépend de facteurs internes et
externes. Les facteurs internes sont par exemple la prédisposition
génétique ou encore l'état de santé, les facteurs
extérieurs sont constitués des températures locales
diurnes et nocturnes, des précipitations, de la longueur des
journées (photopériodisme), mais aussi des conditions de
croissance de l'année précédente ou des conditions
météorologiques des derniers mois.
Les plantes peuvent s'adapter dans une certaine mesure
à leur environnement et à ses modifications. Cette adaptation
revêt généralement la forme d'un compromis entre les
différents besoins de la plante. Ainsi, au cours des
générations, les arbres de latitudes moyennes se sont
adaptés aux conditions climatiques locales afin, par exemple, de
minimiser le risque de dommages causés aux jeunes feuilles par le gel
tout en utilisant au maximum la période de végétation. La
capacité d'adaptation varie selon les espèces»
(Brügger, 1998 ; Defila, 1991 ; Kramer, 1996).
Ainsi, de nombreuses plantes des régions tropicales,
où il fait toujours chaud mais où existe une saison sèche,
n'entament leur croissance que lorsque l'humidité est suffisante. Dans
ces régions, le moment de la croissance est donc déterminé
par le rythme des précipitations (facteur limitatif) et non par la
température (Brügger & Vassella, 2003).
Chapitre II. Milieu d'étude,Matériel et
méthodes d'étude
II.1. Milieu d'étude
Notre étude couvre deux régions
géographiques de la République Démocratique du Congo,
Bolobo et Kinshasa. Dans les lignes qui suivent, nous parlerons
simultanément de ces deux régions qui constituent nos zones
d'étude.
II.1.1. Milieu physique
> Situation géographique
Le territoire de Bolobo se trouve dans le District des
Plateaux (issu d'une séparation de l'ancien District Léopold
II / Mai-Ndombe), dans la Province de Bandundu (en RD Congo). Il est
compris entre 16°13' et 16°32' de longitude Est et, 2°05' et
03°07' de latitude Sud. Il se trouve à 330 km de la capitale,
Kinshasa. Il est borné au Nord par le territoire de Yumbi, à
l'Ouest par le fleuve Congo qui le sépare de la République du
Congo, au Sud par le territoire de Kwamouth et à l'Est par le territoire
de Mushie (carte 1).
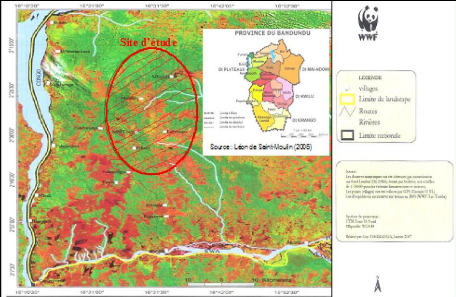
Figure 3. Province de Bandundu : localisation de la zone
d'étude à Bolobo (Source : Menga, 2007)
Par contre, la ville de Kinshasa est une agglomération
située entre 4 °18' et 4 ° 25' latitude Sud et entre 15°
19' et 15°22' longitudes Est. Elle est limitée au Nord et à
l'Est par la province de Bandundu, au Sud par la province du Bas Congo et
à l'Ouest par la République du Congo (Kikufi, 2000). Elle couvre
une superficie de 9.965 Km2 avec une densité moyenne de 441
habitants par Km2. La figure 2 donne la carte de Kinshasa, dans sa
partie urbanisée.
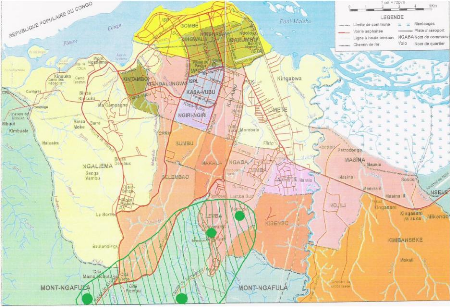
Figure 4. Province de Kinshasa : localisation de la zone
d'étude à Kinshasa (Source : Saint-Moulin & al.,
2005)
> Relief et sol
Le territoire de Bolobo est situé dans la partie Sud de
la cuvette centrale. De l'Est à l'Ouest et du Nord au Sud, le relief est
bas et monotone avec une altitude faible de 320 m en moyenne (Mpoto, 2007). Par
contre, la plaine de la ville de Kinshasa a la forme d'une banane
entourée de colline orientée dans le sens Est-Ouest. Cette plaine
s'étend sur près de 20.000 hectares avec de basses terrasses
alluviales sablonneuses situées entre 260 et 325 m d'altitude,
pénétrant sur une profondeur de près de 10 km en moyenne
(Lelo, 2008).
Les sols de Bolobo sont constitués principalement des
areno-ferrals (sols sableux à faible teneur en argile, moins de 20%).
Leur fertilité varie de moyenne à médiocre. Les meilleures
terres agricoles (sols sablo-argileux) se rencontrent dans la partie Sud
(Mpoto, 2007).
Cependant, le relief de Kinshasa est bas et monotone. Kinshasa
se trouve dans une plaine qui s'étale de la commune de Maluku à
l'Est jusqu'à l'Ouest où les pieds des collines de Ngaliema
stoppent son l'extension, avant qu'elle ne commence à affronter les
chutes de Kinsuka à Ngaliema. Les sols sont décrits comme
étant des sols à texture essentiellement sablonneuse assortie de
quelques éléments grossiers riche en fer et en aluminium. La
faible capacité de rétention en eau de ces sols leurs
confèrent une utilité marginale pour l'agriculture (Lelo,
2008).
> Hydrographie
Aucune grande rivière ne prend sa source dans le
territoire de Bolobo. Son seul avantage est celui d'rtre traversé et
baigné par les grands cours d'eau du pays en l'occurrence : le fleuve
Congo et la rivière Kasaï (pour Bolobo). On y trouve
également plusieurs petits cours d'eau.
La ville de Kinshasa est baignée par plusieurs cours
d'eau dont les plus importants sont : la N'djili ; la N'sele ; la Maï
Ndombe ; la Funa ; la Makelele ; et le fleuve Congo (Menga, 2005).
> Ecoclimat
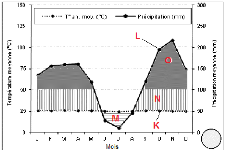
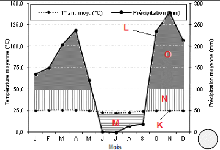
Selon les données climatiques disponibles à la
station météorologique de Binza (Metelsat) mises à notre
disposition, couvrant la période allant de 2001 à 2011, et au
regard de la figure 5, nous pouvons dire que le territoire Bolobo et la ville
de Kinshasa bénéficient respectivement d'un climat de type Aw3 et
Aw4, suivant la subdivision de Koppën.
A Bolobo, ce climat est caractérisé par une
température moyenne annuelle élevée de 25,6°C
(amplitude thermique annuelle = 1,6°C) et les précipitations
annuelles sont de l'ordre de 1.494 mm. Par contre à Kinshasa, la
température moyenne annuelle est de 24,6 °C (amplitude thermique
annuelle = 3,6 °C) et les précipitations annuelles sont de l'ordre
de 1.608 mm.
Chapitre II. Matériel et méthodes
d'itude
a
b
Figures 5. Courbes ombro-thermiques de deux stations
équivalent aux deux régions d'études (a. Station
de
Bandundu-ville pour le site de Bolobo ; b.Station de N'djili pour le site
de Kinshasa), (Source : METELSAT).
Légende : (K) Courbe de
températures moyennes mensuelles (une division de l'axe = 25°C),
(L) Courbe de précipitations moyennes mensuelles (1 division de l'axe =
50 mm et 25°C = 50 mm), (M) Les lignes horizontales correspondent à
une saison relativement sèche, (N) Les lignes verticales correspondent
à une saison relativement humide, (O) Précipitations moyennes
mensuelles supérieures à 100 mm.
II.1.2. Milieu biotique >
Végétation
La végétation de Bolobo demeure jusque là
peu étudiée. Les essences suivantes sont fréquentes dans
les forJts, il s'agit de Millettia laurentii (Wenge), Milicia
exelsa (Kambala), diverses espèces d'Entandrophragma
(Entandrophragma utile, Entandrophragma angolensis, etc.), (Menga, 2007).
Selon le même auteur, on rencontre : (i) les forêts denses et
secondaires de terres fermes, caractérisées respectivement
par Strombosiopsis tetrandra et Diogoa zenkeri, ainsi que
Pentaclethra eetveldeana et Millettia laurentii ; (ii) les
forêts de sols hydromorphes, caractérisées souvent par le
groupement à Hallea stipulosa ; et (iii) les formations
herbacées des sols xéromorphes, dont la principale reste celle
caractérisée par Panicum maximum et Hymenocardia
acida.
La végétation de Kinshasa comprenait des
forêts primitives semicaducifoliées dégradées
(caractérisées par Pentaclethra macrophylla, P. eetveldeana,
Millettia laurenttii, Hymenocardia acida, Syzygium guineense, Oncoba
welwitschii), les forêts ripicoles et marécageuses
(caractérisées par Berlinia grandiflora, Paramacrolobium
coeruleum., Alstonia congoensis, Harungana madagascariensis,
etc.), des savanes (caractérisées par Loudetia demeusei,
L. simplex, Sporobolus pyramidalis, Rhynchelytrum
repens, Digitaria brazzae, Panicum maximum, etc.) et des
formations aquatiques et semiaquatiques (Compère, 1970 ; Habari,
2004).

Figure 6 : quelques individus de wenge dans le site de
Kimwenza(Source : Photo Anicet MALOM )
Avec la pression urbanistique actuelle, la forest se retrouve
secondarisée et se localise dans la région des collines de
Kimwenza et sur le plateau de Kwango. On constate le développement d'une
flore constituée des plantes introduites, dont certaines sont
cultivées et d'autres inféodées au milieu et devenues
subspontanées. (Calverton, 2007).
> Faune
La faune naturelle de la région est riche en
espèces animales diverses, adaptée chacune aux conditions
climatiques et floristiques malgré que très peu d'études
à notre disposition en font mention. D'après Menga (2007) et
Mpoto (2007), la forest est peuplée par les Pan paniscus
(Bonobos) et d'autres primates du genre Cercopithecus. On y trouve
également les Potamocherus porcerus (Potamochères),
Loxodonta africana cyclotis (Elephants de forests), Atherurus
africanus (Porc-épic), plusieurs serpents (dont les Pyton
sebae, Bitis gabonica, etc.), les Buffles de savane (Syncerus
caffer), diverses Antilopes (ex. Tragelaphus spp), ainsi que les
carnassiers tels que le lion (Panthera leo), le chacal (Canis
mesomelas), la Civette (Viverra civetta), etc. On y trouve
également plusieurs Oiseaux dont les Tisserins (Ploceus spp),
Hirondelles (Hirondo spp), Martins pécheurs (Alcedo
spp), Moineaux (Passer spp), etc. La faune ichtyologique comprend
diverses espèces dont Tilapia spp, Oreochromis niloticus,
Clarias gariepinus, Paracana obscura, Protopterus ssp, Heterobranchus
longifilis, etc.
A Kinshasa, on peut signaler l'existence de quelques d'oiseaux
(dont le plus courant sont Columba guinea (Pigeon de guinée),
Streptopelia decipiens (Tourterelle pleureuse), Alcedo cristata
(Martin pécheur huppée), Corbus albus (Corbeau
pie), Ploceus spp (Tisserins), Passer spp (Moineaux), etc.
Les rivières et ruisseaux parcourant la ville abritent une faune
ichtyologique adaptée à la pollution (par exemple,
l'Oreochromis niloticus, Tilapia zillii, Clarias gariepinus, Paracana
obscura, etc). La conservation de certaines espèces animales se
fait dans le parc de la N'SELE, le Jardin Zoologique de Kinshasa, et la
réserve de chasse de Bombo-Lumene. On peut citer Atherurus
africanus (Porc-épic), Tryonomys swinderinus (Aulacode),
Cephalophus spp. (Antilopes), Tragelaphus seriptus (Antilope
ou « Nka » en Kikongo), Cercopthercus ascanuis (Singe au nez
blanc), Viverra civetta (Civette), etc. (Ngalula, 2004 ; Menga, 2005 ;
Muleka, 2003).
II.2. Breve description de l'espice
Millettia laurentii De Wild. est une Fabaceae
de l'ordre des Fabales, caractéristique des forests denses
semi-décidues bien drainées. On le retrouve également dans
les forests secondaires matures, dans les abords des forests
marécageuses ou des forests sujettes à inondations
périodiques ainsi que dans les savanes arborées (Pauwels, 1993 ;
Latham & Konda, 2006 ; Wilks & Issembe, 2000).
Cet arbre de taille moyenne atteignant 30 (- 45) m de hauteur
présente un tronc gris pâle à aspect lisse de loin, rugueux
de près en raison de la présence des lenticelles, et souvent
couverte de lichens verts à la base. Le felt est cylindrique, souvent
légèrement courbé (rarement droit), atteignant
jusqu'à 150 cm de diamètre, et est légèrement
soutenu à la base par des piliers cylindriques, cannelées ou
légèrement empattés (Figure 7a). Les feuilles sont
composées, imparipennées, alternes, longue de 25 à 30 cm,
à (4-)6-7(-9) paires de folioles opposées plus une terminale.
L'inflorescence est en panicule terminale de 20-40 cm de long. Les fleurs sont
bisexuées, s'épanouissant en grandes grappes, de couleur
violète pâle à bleue violacée (Figure 7b). Le fruit
est une gousse déhiscente ligneuse brun-claire, linéaire, de
15-28 cm x 3-5 cm (Figure 7d). Les graines sont oblongues à
lenticulaires, aplaties, de 22- 25 mm × 18-20 mm, lisses, brun violet
(Figure 7c), (Tailfer, 1989 ; CTFT, 1952 ; Tchinda, 2008 ; Wilk & Issembe,
2000).
Chapitre II. Matériel et méthodes
d'itude

a d
b c
Figure 7. Millettia laurentii (a. fût ; b. grappe de
fleur ; c. graines matures sur une gousse ; d. gousses matures)
Cette espèce est naturellement répartie dans la
partie Sud du Bassin du Congo, en RDC et dans la zone frontière entre le
Cameroun, le Gabon et le Congo (Wilk & Issembé, 2000), mais aussi
dans le sud de la République Centrafricaine (Tchinda, 2008). Son aire de
croissance optimale est péri-guinéenne, et se situe en RDC, dans
la partie Sud-Ouest de la cuvette centrale, principalement au Nord de la
province du Bandundu et au Sud de la province de l'Equateur. Néanmoins,
on retrouve quelques tiges parsemées au Kasaï Occidental, à
Kinshasa (partie Sud du plateau de Bateke), et dans le territoire de Kasangulu
dans le Bas-Congo (Mandango & al., 1999 ; Compère, 1970 ;
Autre source : Menga, comm. pers.).
Grâce à sa couleur brun noir de chocolat au lait
et ses fines veines claires, M. laurentii a un aspect tout à
fait particulier qui fait de lui avant tout un bois
d'ébénisterie. Il est également fort convoité
pour nombre d'applications décoratives, la parqueterie lourde, le
contreplacage, l'ameublement, la menuiserie, le revêtement
extérieur, la sculpture, les panneaux, le tournage et le placage. On
l'utilise aussi comme arbre ornemental et d'alignement (CTFT, 1952 ; Latham
& Konda, 2006 ; Pauwels, 1993).
II.3. Méthodologie d'étude
II.3.1. Installation des circuits phénologiques
Pour déterminer les caractéristiques
phénologiques et le diamètre de fructification de M.
laurentii nous avions mis en place deux circuits phénologiques dans
nos deux régions d'étude, Bolobo et Kinshasa. Nous avions suivis
mensuellement dans chaque site, de façon individuelle pendant au moins
une année, la phénologie (feuilles, fleurs, et fruits) de 221
arbres numérotés et localisés géographiquement
à l'aide d'un GPS. Ce suivi couvre la période allant du 01
juillet 2010 au 07 juin 2011.
Dans la région de Bolobo, le circuit
phénologique était installé dans une mosaïque
forêt dense et forêt secondaire. Par contre à Kinshasa, il
était installé dans quelques quartiers
périphériques de la commune de Lemba (Righini, Mont-Amba,
Mbanza-Lemba, plateau de professeurs, Village Kinduku), de Mont-Ngafula
(Concession Bianda et Ecole Eucalyptus), et de Kimwenza.
Les arbres (diamètre =10 cm) ont été
sélectionnés dans les emplacements « représentatifs
» (par exemple, tous ne devaient pas être situés en
lisière de grandes éclaircies, leur houppier devaient ~tre
clairement visible d'au moins un endroit) ; ce qui fait qu'une
préférence a été portée sur les arbres
placés à une distance telle qu'ils étaient facilement
visibles depuis une piste, un chablis ou un transect.
II.3.2. Prélèvements des données
Pour effectuer nos relevés phénologiques dans
les deux régions durant une période aussi longue, nous avions
scindés les équipes de terrain en deux groupes. Le groupe de
Bolobo était dirigé par Mr Bedel Iboto (chef d'équipe) et
celui de Kinshasa par Mr Anicet Malom. Ces deux groupes travaillaient sous la
supervision de l'Assistant Pisco MENGA.
Les paramètres relevés sur le terrain sont
consignés sur une fiche de relevés (voir tableau 1). Ces fiches
sont numérotées et comportent 2 types d'informations :
a) Les données fixes : Elles
permettent de localiser et d'identifier les arbres
concernés par le suivi phénologique. Chaque arbre était
marqué de façon permanente, sa localisation était
déterminée et son diamètre mesuré. Les principales
données fixes prisent en compte sont :
- L'habitat1: défini brièvement le milieu
dans lequel l'arbre croit.1
- 1EDICioE1F 1situe l'arbre sur le dispositif
phénologique
- N° ou code de l'arbre concerné1: 1Il
s'agit du numéro de dénombrement de l'arbre au moment de
l'installation du circuit phénologique.
- Coordonnées GPS de l'arbre1: Ces
coordonnées ont été prises 1en système
géodésique WGS 84 avec des grilles Lat/long
hddd°mm'ss.s''.1
- E1dECT1: Cette rubrique « date » renseigne
pour chaque arbre le jour (ou le mois) à laquelle les relevés ont
été faits.
- Le nom de l'observateur était inscrit sur
chaque fiche de relevé. 1
Tableau I. Exemple de fiche de prise de données
phénologiques.
Fiche n° : ......... Observateur :
........................«
Location : ......... Date:
«..............................«
Habitat : : .........«
|
FEUILLES
|
FLEURS
|
FRUITS
|
|
|
N° Placette
|
N° de l'arbre
|
Coordonnées
|
Statut
|
DBH
|
FJ
|
FM
|
FS
|
|
FRI
|
FRM
|
Remarques
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
FJ : feuilles jeunes, FM : feuilles matures, FS : feuilles
sénescentes, FRI : fruits immatures,
FRM : fruits matures
Dans cette fiche, la colonne « remarque » inclue
toutes les observations de caractéristiques pouvant présenter un
intér~t particulier ou étant en liaison avec l'une des
données relevées (la présence des dégâts dus
aux insectes, à des animaux, les branches cassées, la
défeuillaison, les lianes sur l'arbre, la maladie constatée,
chute d'arbre à proximité, indices au sol des fruits, fleurs et
semis). La quantification des indices au sol se fait de la manière
suivante : la présence peu abondante (5 indices au moins pour une
caractéristique) et la présence abondante (plus de 5 indices).
[Les données mensuelles :
1Elles sont collectées sur le terrain tous les mois, au moins
pendant une période d'au moins un an. Le principe était de
contrôler mensuellement chaque arbre, de préférence chaque
2ème semaine du mois. Elles concernaient :
- Les jeunes feuilles, les feuilles matures et les feuilles
sénescentes ; - Les fleurs;
- Les fruits immatures sur l'arbre et les fruits mErs sur
l'arbre.
La mesure de diamètre de chaque arbre a
été effectuée en suivant les conventions de Dallmeier
et al. (1992). Dans chaque site, seuls les arbres de diamètre =
10 cm dont le houppier était clairement visible d'au moins un endroit
ont été sélectionnés, numérotés, et
regroupés en 7 classes de diamètre d'une amplitude de 10 cm, en
raison d'un minimum de 30 arbres par classe de diamètre, afin d'obtenir
des données statistiquement fiables.
Le statut social de chaque arbre dans la communauté
forestière a été estimé suivant Doucet (2003) :
l'arbre est dominant, si sa cime n'est voisine d'aucune autre cime. Il est
co-dominant, si sa cime est voisine d'une ou plusieurs autres cimes sans pour
autant ~tre dominée par celle(s)-ci, et dominé, si sa cime est
dépassée par celle d'un ou plusieurs autres arbres.

Figure 8 . Anicet MALOM sur le terrain dans le site de
Kinshasa/ Mont-Ngafula (Source : Photo Anicet MALOM)
La prise de données était effectuée
toutes les deuxièmes semaines de chaque mois, et les observations
portaient sur la présence (ou l'absence) de fleurs, de fruits (immatures
et matures), et de feuilles (jeunes, matures, et sénescences), afin
d'obtenir la proportion d'arbre ayant présenté ou non une
phénophase quelconque. Les feuilles jeunes, matures et
sénescentes étaient détectées par le changement de
couleur du vert au jaune.
II.3.4. Traitement des données
Nous avions dans un premier temps examiné le
comportement phénologique de M. laurentii dans les deux sites
d'étude. Les différences entre sites ont été
testées par analyse de variance sur mesures
répétées "repeated ANOVA " à l'aide du
logiciel XLSTAT 2011.
Le type de floraison a été déterminé,
suivant la classification proposé par Hecketsweiler (1992). Selon cette
classification, on distingue trois grands types de floraison :
a) La floraison apériodique ou continue : la floraison
est continue soit parce que chaque individu a une floraison continue ou les
individus fleurissent en se relayant.
b) La floraison épisodique : on en distingue de deux
types,
- La floraison épisodique infra-annuelle : chaque
individu présente des phases multiples de floraison à n'importe
quelle époque de l'année. Aucun rythme de floraison n'est
détectable mais les individus tendent à fleurir de façon
plus ou moins synchrone à différentes époques de
l'année.
- La floraison épisodique supra-annuelle : il s'agit
des espèces dont les individus tendent à fleurir de façon
synchrone sans périodicité fixe par intervalle de plusieurs
années.
c) La floraison rythmique : on en distingue 5 types,
- La floraison rythmique infra-annuelle : la floraison se
répète après un nombre de mois déterminé
;
- La floraison rythmique annuelle : la période du
rythme de floraison est égale à une année calendaire ou
à une année climatique. Plusieurs possibilités peuvent se
présenter : tous les individus d'une espèce peuvent fleurir soit
à « date fixe », soit par groupes mais chaque groupe ne
fleurit qu'une seule fois par an, soit encore de façon asynchrone entre
eux ;
- La floraison rythmique sesquiennale : si elle se manifeste tous
les 18 mois ; - La floraison rythmique biennale : si elle a lieu tous les deux
ans ;
- La floraison rythmique triennale : si elle a lieu tous les
trois ans.
Nous avons également étudié les
variations de l'intensité de la fructification avec
le diamètre, en calculant le pourcentage d'arbres fructificatifs (i.e.
semenciers), par classe de diamètre. Les arbres ont été
regroupés en 8 classes de diamètre d'une amplitude de 10 cm,
chaque classe étant représentée au minimum par 30
individus. La détermination du DFR tient compte de la proportion
d'arbres ayant fructifié dans chaque classe de diamètre. Cette
proportion est variable selon les auteurs, ainsi Debroux (1998), Doucet (2003)
et Kouadio (2008) utilisent un seuil de 70 % alors que Durrieu de Madron &
Daumerie (2004)
NF EaNFIt NOrEEEMIAIIErFN. IEl IONTFtIFIaonN ibgalement cette
valeur de 80 % dans cette étude.
Une analyse des correspondances multiples (ACM) a
été utilisée pour mettre en évidence une
corrélation entre la floraison, le diamètre et le statut social
des arbres.
Chapitre III. Résultats
III.1. Phénologie végétative de M.
laurentii
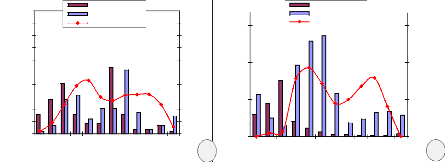
L'analyse de la phénologie foliaire de M. laurentii
dans les deux sites montre que la défeuillaison et l'apparition des
jeunes feuilles (débourrement) est étroitement liée au
régime de précipitations (Figure 9). Durant les 12 mois
d'observation, près de 55 % de tiges suivies sur le site de Bolobo ont
défeuillées, tandis que pour le site de Kinshasa très
saisonnier, on en dénote 78 %. Le pic de défeuillaison à
Bolobo s'observe à la petite et grande saison sèche, soit
respectivement en janvier et septembre, tandis qu'à Kinshasa on
l'observe uniquement lors de la grande saison sèche, soit au mois
d'aoEt.
L'ANOVA à mesure répétée
réalisée à l'aide du logiciel XLSTAT 2011 montre que les
différences entre sites sont très significatives avec risque
d'erreur de 5 % (Tableau III). Tout d'abord, les effets inter-individus qui
permettent de voir l'effet de la variable site sur la défeuillaison,
montre que le site a un impact significatif sur la défeuillaison (p <
0,05 ; Tableau III.1). Et, ensuite, les effets intra-individus qui permettent
de voir l'impact de la mesure répétée et donc du temps,
montre que le temps à un impact significatif sur la défeuillaison
(p < 0,05) mais aussi que l'interaction entre la répétition et
le site a aussi un impact significatif sur la défeuillaison (p <
0,05) (Tableau III.2).
Proportion tiges (%)
100
40
20
90
80
70
60
50
30
10
0
J A S O N D J F M A M J
Défeuillaison Débourrement Précipitation
(mm)
Mois
450
400
250
200
50
0
500
350
300
150
100
Precipitation moyenne (mm)
J A S O N D J F M A M J
a b
Mois
Proportion tiges (%)
100
40
20
90
80
70
60
50
30
10
0
Défeuillaison Débourrement Précipitation
(mm)
450
400
250
200
500
350
300
50
0
150
100
Precipitation moyenne (mm)
Figure 9. Phénologie foliaire de Millettia laurentii
(a. site de Bolobo, b. site de Kinshasa)
Le site de Bolobo a deux périodes de
débourrement qui correspondent à la petite et à la grande
saison des pluies : au mois de février et de septembre-octobre. Sur le
site de Kinshasa, elle a lieu au début de la grande saison des pluies
(octobre à décembre).
La maturation effective de feuilles commence
généralement deux mois après le débourrement. La
sénescence est maximale de février à avril sur le site de
Bolobo et de juin à septembre sur le site de Kinshasa.
Tableau II. Résultats de l'ANOVA à mesure
répétée sur la défeuillaison
III.1. Tests des effets inter-individus:
|
Source DDL Somme des Moyenne des F Pr > F
carrés carrés
|
Site 1 1,666 1,666 24,033 < 0,0001
Erreur 440 30,500 0,069
III.2. Tests des effets intra-individus :
|
Source DDL Somme des Moyenne des F Pr > F
carrés carrés
|
Répétition 11 30,549 2,777 42,025 < 0,0001
Site*Répétition 11 20,940 1,904 28,807 <
0,0001
Erreur 4840 319,844 0,066
III.2. Phénologie reproductrice de M.
laurentii
III.2.1. Phénologie de la floraison et de la
fructification
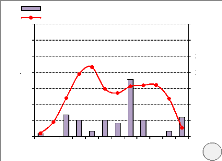
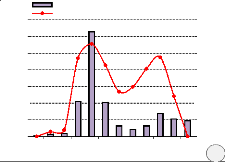
Parmi les arbres suivis, on note 80 % des tiges de
Millettia laurentii fleuries au moins une fois durant l'année
sur le site de Bolobo et 88 % sur le site de Kinshasa. Sur l'ensemble des tiges
fleuries au cours de 12 mois d'observation, environ 93 % ont fleuri une fois
l'an et 7 % deux fois l'an jà Bolobo, et 86 % une fois l'an et 14 % deux
fois l'an à Kinshasa. La floraison dure un mois en moyenne mais elle
peut atteindre jusque 3 mois chez certains individus.
On observe sur les deux sites, une grande période de
floraison, qui coïncide globalement avec les périodes de
débourrement des feuilles et les saisons des pluies. La floraison
maximale s'observe en février pendant la petite saison de pluies pour le
site de Bolobo, et en novembre, pendant la grande saison de pluies pour le site
de Kinshasa (Figure
10). Il existe donc une grande variabilité
significative de floraison, avec risque d'erreur de 5 %, entre le site de
Bolobo un peu humide (2 mois de sécheresse et précipitations
annuelles atteignent 1800 mm) et le site de Kinshasa fortement
saisonnière (4 mois de sécheresse et une pluviosité de
l'ordre de 1600 mm/an) (Tableau IV). En effet, les effets inter-individus qui
permettent de voir l'effet de la variable site sur la floraison, montre que le
site a un impact significatif sur la floraison (p < 0,05 ; Tableau III.1).
Et, ensuite, les effets intra-individus qui permettent de voir l'impact de la
mesure répétée et donc du temps, montre que le temps
à un impact significatif sur la floraison (p < 0,05) mais aussi que
l'interaction entre la répétition et le site a aussi un impact
significatif sur la floraison (p < 0,05) (Tableau III.2).
La floraison dure un mois en moyenne mais peut atteindre
jusque 3 mois mais ce dernier cas est surtout fréquent chez les
individus qui fleurissent branche par branche.
Proportion de tiges fleuries (%)
% tiges fleuries
Courbe de précipitation_Station de Bandundu (Bolobo)
70 350
50
40
30
60
20
10
0
Juil. Août Sép. Oct. Nov. Déc. Jan.
Fév. Mar. Avr. Mai Juin
Mois
300
50
0
250
200
150
100
Precipitation moyenne (mm)
a
Proportion de tiges en fleurs (%)
40
70
60
50
30
20
10
0
Juil Août Sept Oct Nov Déc Jan Fév Mars Avr
Mai Juin
Mois
% tiges fleuries
Précipitation (mm)_Station de N'djili (Kinshasa)
350
300
250
200
50
0
150
100
b
Precipitation moyenne (mm)
Figure 10. Phénologie de la floraison de M. laurentii
(a. site de Bolobo, b. site de Kinshasa)
Tableau III. Résultats de l'ANOVA à mesure
répétée sur la floraison
IV.1. Tests des effets inter-individus :
Source DDL Somme des carrés Moyenne des carrés F
Pr > F
Site 1 4,647 4,647 72,799 < 0,0001
Erreur 440 28,088 0,064
IV.2. Tests des effets intra-individus :
Source DDL Somme des carrés Moyenne des carrés F Pr
> F
Répétition 11 34,595 3,145 40,715 < 0,0001
Site*Répétition 11 44,631 4,057 52,527 <
0,0001
Erreur 4840 373,857 0,077
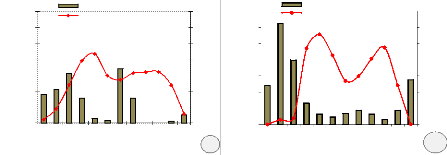
La production des fruits de Millettia laurentii suit
directement celle des fleurs, mais les fruits immatures deviennent perceptibles
à l'~il nu, environ deux mois après la floraison. Les arbres
produisent énormément de fruits immatures mais c'est seulement
près des 2/3 qui atteignent la maturité, car on observe beaucoup
des pertes prématurées de fruits immatures. La maturation des
fruits est assez longue : elle dure 6 à 8 mois, elle a eu lieu
massivement lors de la grande saison sèche. La maturité
complète avec possibilité de récolte des semences
(éclatement de gousses) coïncide à peu près avec la
période de défeuillaison maximale, soit au mois de juin pour le
site de Bolobo et d'aoIt-septembre pour le site de Kinshasa (Figure 11).
Proportion liges en gousses edatees (%)
40
70
60
50
30
20
10
0
J A S O N D J F M A M J
% tiges en gousses éclatées Courbe de
précipitation (mm)
Mois
350
300
250
200
50
0
150
100
Predpitation moyenne (mm)
a
Proportion tiges en gousses eclatees (%)
40
20
70
60
50
30
10
0
J A S O N D J F M A M J
Mois
% tiges en gousses éclatées Courbe de
précipitation (mm)
250
200
350
300
50
0
150
100
b
Precipitation moyenne (mm)
Figure 11. Eclatement de gousses de Millettia laurentii (a.
site de Bolobo, b. site de Kinshasa)
III.2.2. Fructification et diamètre
En raison de la mauvaise conformation des tiges de
Millettia laurentii évoluant à Kinshasa, dans un habitat
non naturel, nous avions considéré dans ce paragraphe, seulement
les tiges étudiées à Bolobo. Le suivi de la
phénologie des 221 individus de M. laurentii répartis
nous a également permis d'analyser les variations de la fructification
avec le diamètre. Il se révèle que des individus
fleurissent et fructifient dans toutes les classes de diamètres (Tableau
V). Le diamètre minimal pour la fructification peut donc être
considéré à partir de 10 cm. La proportion de tiges
fleuries et ayant fructifiées est néanmoins plus faible pour les
arbres de petit diamètre alors que peu d'arbres au delà de 60 cm
de diamètre ne fleurissent sans fructifier par la suite.
La floraison efficace s'effectue à partir de 30 cm de
diamètre, oil plus de 70% des individus fleurissent. A partir de 50-60
cm, la majorité des individus (environ 90%) produisent des fleurs et
donnent régulièrement des fruits.
Tableau IV. Proportion des tiges de Millettia
laurentii ayant fleurie et fructifié
|
Classes de diamètre
|
Effectifs
|
% de tiges ayant
fleurie
|
% de tiges ayant
fructifié
|
|
1 (10-20 cm)
|
30
|
30
|
26
|
|
2 (20-30 cm)
|
31
|
72
|
71
|
|
3 (30-40 cm)
|
34
|
72
|
72
|
|
4 (40-50 cm)
|
32
|
86
|
83
|
|
5 (50-60 cm)
|
30
|
91
|
91
|
|
6 (60-70 cm)
|
31
|
86
|
83
|
|
= 70 cm
|
33
|
90
|
70
|
|
|
|
|
Une Analyse des Composantes Multiples (ACM) effectuée
à l'aide du logiciel XLSTAT 2011, impliquant trois facteurs : le statut
social (Statut), la classe de diamètre (Classedhp) et la production de
fleurs (Fleurs), a montrée l'existence d'une forte corrélation
entre la floraison, le statut social et la classe de diamètre. Les
variables ont été discrétisées comme suit :
- statut : dominant = 3, co-dominant = 2, dominé = 1
- fleurs : présentes = 1, absentes = 0
- classe de diamètre 1, 2, 3, ..., 9 = Classe_dhp-1,
Classe_dhp-2, .... , Classe_dhp-9
Le tableau des valeurs test (Tableau VI) permet de savoir
quelles sont les modalités importantes pour l'interprétation
graphique du principal résultat de l'ACM : la représentation des
variables sur le plan factoriel F1/F2 (cf. Figure 12). Les valeurs-test
représentées en gras sont dites significatives au seuil á
= 0,05 et peuvent être interprétées graphiquement.
Tableau V. Valeurs test (Variables) :
|
F1 F2
|
|
Statut-1
Statut-2
Statut-3
|
6,679 -0,601 -4,944
|
1,536 -6,900 5,206
|
Classe_dhp-1
|
2,497
|
1,916
|
Classe_dhp-2
|
5,076
|
1,030
|
Classe_dhp-3
|
2,489
|
-3,313
|
Classe_dhp-4
|
-1,620
|
-3,514
|
Classe_dhp-5
|
-1,613
|
-2,448
|
Classe_dhp-6
|
-2,752
|
2,452
|
Classe_dhp-7
|
-1,767
|
3,824
|
Classe_dhp-8
|
-0,930
|
1,220
|
Classe_dhp-9
|
-1,003
|
-0,859
|
Fleurs-0
|
6,715
|
0,696
|
Fleurs-1
|
-6,715
|
-0,696
|
|
Suite à l'analyse de ce tableau, nous utiliserons les
modalités de la variable « fleur » pour interpréter
l'axe F1. Les autres variables peuvent ~tre expliquées par les deux
axes. Au vu du graphique symétrique des variables résultant de
l'ACM (Figure 12), nous pouvons tirer les conclusions suivantes :
- Millettia laurentii fleurie le plus souvent quand
elle occupe le statut social de dominant ou de co-dominant dans la
communauté forestière ;
- La production des fleurs est médiocre chez les tiges de
10-20 cm ayant le statut social de dominé ;
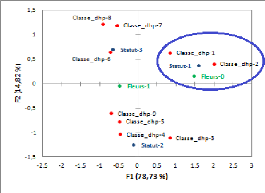
Figure 12. Représentation des variables «
floraisons », « statut social », et « classe de
diamètre » sur le plan
EFTRU7r E) Eli44X171r'58z0
4.1. La phénologie de Millettia laurentii est
liée à la saisonnalité des pluies
La phénologie foliaire et florale de Millettia
laurentii coïncide fortement avec la saisonnalité des
précipitations.
A Bolobo, la défeuillaison est maximale en Janvier,
lors de la petite saison sèche, mais aussi en septembre vers la fin de
la grande saison sèche. Par contre à Kinshasa, le pic se situe en
Septembre, vers la fin de la grande saison sèche. Ces résultats
corroborent ceux de plusieurs auteurs dont Withmore (1975) et Kouadio (2008),
qui stipulent que pour un grand nombre d'espèces tropicales la chute des
feuilles coïncide avec la saison sèche, car pendant ce temps, les
arbres sont confrontés à un stress hydrique foliaire et
racinaire, la photosynthèse est affectée, les feuilles jaunissent
puis tombent. L'impact du régime pluviométrique dans les deux
sites s'observent également dans la proportion totale des individus
ayant défeuillé durant les 12 mois d'observation. En effet, le
site de Bolobo, plus arrosé que celui de Kinshasa, présente
près de 55 % de tiges défeuillées ; alors que celui de
Kinshasa, plus sec en présente environ 78 %. Cela laisse entrevoir que
Millettia laurentii présente un comportement
semi-décidue si elle se trouve dans son aire naturelle de Bolobo, et
décidue si elle est dans un climat sec que d'habitude comme à
Kinshasa. Donc, les différences observées entre ces deux sites
résultent des conditions environnementales de chaque site. Sur le plan
individuel, les individus suivis ne défeuillent pas
nécessairement au même moment, la chute des feuilles n'est pas
simultanée pour toutes les tiges, elle se déroule parfois de
façon alternative d'un individu à l'autre ou d'une branche
à l'autre sur le mrme pied ; en plus, certains individus ne sont pas
soumis à une défeuillaison. Cela peut être lié
jà l'organisation physiologique et à la génétique
propre à chaque individu.
Quant à l'apparition de feuilles (débourrement),
les pics s'observent au début des deux saisons des pluies (en
février et octobre) pour le site de Bolobo, et lors de la grande saison
de pluies pour le site de Kinshasa. Ces périodes s'apparentent donc
à l'amélioration des conditions climatiques pour le
fonctionnement des arbres.
La floraison de Millettia laurentii s'effectue de
façon synchrone avec l'apparition des nouvelles feuilles, comme chez
d'autres espèces caractéristiques des forts semi-décidues
d'Afrique centrale, à l'instar des Entandrophragma spp
jà M'baïki en Afrique centrale (Medjibe et Hall, 2001). Les
précipitations semblent être le principal facteur stimulant la
floraison chez l'espèce malgré que des floraisons occasionnelles
peuvent néanmoins ftre observés tout au long de l'année.
Ce qui rend le type de floraison difficilement identifiable avec exactitude ;
ce type de floraison varie donc en fonction de l'échelle d'observation
comme suggéré par Newstrom et al. (1994).
A l'échelle du peuplement (sites), la floraison
paraît continue (Hecketsweiler, 1992) pour les deux sites, car les
individus fleurissent en se relayant, on retrouve des floraisons occasionnelles
de faibles amplitudes tout au long de l'année. Le pic de floraison
s'observe en février, lors de la petite saison de pluies sur le site de
Bolobo, et un peu plus tard, en novembre sur le site de Kinshasa. Ce constat
vient en appui de celui de Martineau (1929) cité par Capon (1947), qui a
observé également chez certaines espèces de la RD Congo,
des floraisons au début de la petite et de la grande saison des pluies.
A l'échelle de l'individu, la floraison présente un rythme annuel
(sensu Hecketsweiler, 1992), car la majorité d'individus
fleurissent par groupes mais chaque groupe ne fleurit qu'une seule fois par an
(environ 93 % pour Bolobo et 86 % pour Kinshasa).
La fructification suit directement la floraison, les jeunes
fruits deviennent perceptible à l'~il nu un à deux mois
après la floraison. La proportion d'arbres portant les fruits matures
reste assez faible jà cause d'une importante chute
prématurée des fruits immatures.
4.2. Millettia laurentii possède une
fructification précoce
Que ça soit dans le milieu forestier de Bolobo ou
urbain de Kinshasa, les individus de Millettia laurentii suivis, dans
toutes les classes de diamètre possible, fleurissent et fructifient.
Cependant, cette floraison dépend en grande partie du statut social de
l'individu dans la communauté forestière comme observé
à Bolobo, un milieu fermé. Les individus qui sont souvent
dominés (cas des individus appartenant à la classe de
diamètre 1 et 2) ont des problèmes de floraison, car il y a peu
de rayonnement solaire à leur porté. Par contre, dans un milieu
ouvert comme Kinshasa, cette variable (i.e statut social) n'as pas
d'incidence sur la floraison des individus car le milieu est ouvert, toutes les
tiges bénéficient donc d'un
éclairement très important, et la différence
des classes de diamètre sur la floraison ne se fait pas sentir.
Le diamètre minimal pour la fructification (DMF) de
Millettia laurentii est donc de 10-20 cm. Comparativement aux
résultats de Sepulchre et al. (2008), qui retiennent la valeur
de DMF de 20 cm au Gabon, nous pouvons dire que Millettia laurentii
est une espèce à fructification précoce. Cette
précocité de floraison garantie le maintien de l'espèce
dans la forest et sa capacité à se perpétuer. En effet,
l'espèce possède un taux de mortalité juvénile
très élevée, et pour compenser cette mortalité et
sa pérennisation dans le milieu forestier, cette précocité
de fructification est d'une nécessité extr~me. Donc, sa
stratégie de cicatricielle durable des milieux forestiers et sa
capacité à produire rapidement des semences fertiles, lui
confère l'aptitude à la colonisation des trouées
forestières (Menga, communication personnelle).
Le diamètre de fructification régulière
(DFR) est de 50-60 cm. Ce diamètre est légèrement
supérieur à celui de 45 cm trouvé au Gabon par Sepulchre
et al. (2008) et au Congo par Gillet et al. (2008), et valide
le diamètre minimal pour l'exploitabilité (DME) de
l'espèce, fixé par l'administration forestière du pays,
qui est de 60 cm. En effet, il est généralement admis que le DME
d'une essence commerciale doit ~tre supérieur d'au moins 10 cm au DFR
afin de laisser le temps aux arbres semenciers de fructifier pendant au moins
quelques décennies avant d'1tre exploités (Durrieu de Madron et
Daumerie, 2004), or ce qui est le cas avec Millettia laurentii, donc
ce qui atteste que le DME actuel n'a pas d'influence négative sur la
reproduction de l'espèce.
Conclusion
A l'issue de cette étude sur le comportement
phénologique des populations de Millettia laurentii à
Bolobo et à Kinshasa, nous pouvons dire qu'il existe une
légère variabilité intraspécifique d'une part, et
inter-sites d'autre part dans l'apparition des caractères
phénologique de Millettia laurentii. Cette variabilité
offre un large choix de sélection des périodes d'exploitation et
de diamètre d'exploitation aux gestionnaires forestiers.
Cette étude a permis de montrer que la
phénologie des populations de Millettia laurentii
répondait principalement au gradient climatique (la
pluviométrie). D'autres paramètres non étudiés
pourraient certainement intervenir pour la réguler sur le plan
individuel (la physiologie et le génotype propres de l'individu, etc.).
Les résultats de cette étude ont des implications pour la
gestion de cette espèce qui constitue actuellement l'une
des essences les plus exploitées de la RDC. Le diamètre de
fructification régulière de Millettia laurentii
étant de 50 cm, les arbres ne peuvent ~tre exploités
qu'à partir d'un diamètre minimal de 60 cm, afin de garantir le
maintien d'un bon nombre de semenciers et pouvoir leur laisser le temps
nécessaire de fructifier au moins pendant quelques décennies
avant d'itre exploitée. Il serait aussi préférable que les
arbres exploitables ne soient pas abattus pendant leur période de
reproduction (floraison, fructification) mais plutôt après la
période d'éclatement de gousses qui coïncide avec la
défeuillaison. C'est pendant cette période de
défeuillaison que peut se faire la récolte des graines. Des
études similaires sont recommandées pour les autres essences
telles que Entanshophragma utile, Entanshophragma
angolensis,Hallea stipulusa,Milicia excelsa qui sont aussi
particulièrement exploitées, souvent de manière
artisanale, dans les zones de faible densité.
Ces études permettraient de donner des instructions
utiles dans l'exploitation des écosystèmes contenant ces essences
et qui sont dégradés, de façon anarchique.
La protection de semenciers est la seule voie d'emprcher la
disparition totale de ces essences dans les écosystèmes fragiles
exposés à l'exploitation pour des besoins agricoles ou
d'énergie bois (charbon de bois, bois de chauffe).
Références bibliographiques
Algoët B. (2008). Phenologie et regeneration du Wenge
(Millettia laurentii) en RDC. Inedit, Mém. de fin
d'étude, AgroParisTech, 95p. + Annexes.
ATIBT (2006). Synthèse sur les forêts
equatoriales-Volet faune sauvage. Ed. ATIBT, 50p. Blanc P. (2002). Être
plante à l'ombre des forits tropicales. Nathan, Paris, France, 428 p
Boyemba B.F. (2011). Ecologie de Pericopsis elata
(Harms) Van Meeuwen (Fabaceae), arbre de forêt tropicale
africaine à repartition agregee. Thèse de doctorat, ULB,
Bruxelles, 181 p.
BRÜGGER R. & VASSELLA A. (2003) : Les plantes au
cours des saisons - Guide pour observation phénologique.
Geographica Bernensia, Bern, 56p.
Calverton, (2007) : enquête demographique et de sante en
republique democratique du Congo (EDS, RDC), Ministère du plan avec la
collaboration du Ministère de la sante publique, 2eme ed. DDE Maryland,
Kinshasa, RDC 482p.
Capon M. (1947). Observations sur la phénologie des
essences de la forest de Yangambi. Comptes rendus de la semaine agricole
de Yangambi (du 26 fevrier au 5 mars 1947). Public. INEAC,
2ème partie, Hors serie, Communication n°66, pp
849-861.
Compère P. (1970). Carte des sols et de la
végétation du Congo, du Rwanda et du Burundi, n°25
Bas-Congo : Note explicative de la carte de la végétation. INEAC,
Bruxelles, pp 27-28.
Comps B., Letouzey J., Savoie J.M. (1987) : Phénologie
du couvert arborescent dans une chesnaie-hestraie d'Aquitaine. Annales des
Sciences forestières, vol. 44, pp. 153-170.
CTFT (Centre Technique Forestier Tropical) (1952). Wenge. Revue
Bois et Forêts des Tropiques 25: 329-332.
Dallmeier F., Kabel M. et Rice R. (1992). Methods for
long-term biodiversity inventory plots in protected tropical forest. In
: Dallmeier F. (ed.). Long-term monitoring of biological diversity in
tropical forest areas: methods for establishment and inventory of permanent
plots. MAB Digest 11. UNESCO, Paris, 11-46.
De Wasseige C., Devers D., de Marcken P., Eba'a Atyi R., Nasi
R. et Mayaux P. (2009). Les forests du Bassin du Congo : Etat des
forests 2008. Office des publications de l'Union européenne,
425p.
Debroux L. (1998).
/11Ppg1fIPIULaLsLIRUtsLtERSIRIPsLIRnapLEITLlEL1fUOIRnLaIsLSRSulations
agaaL- Exemple du moabi (Baillonella toxisperma Pierre) dans la forest du Dja,
Cameroun. Thèse de doctorat, FUSAGX, 283p.
Delpech R., Dume G. & Galmiche P. (1985)
: Typologie des stations forestières - Vocabulaire.
Ministère de l'Agriculture, Direction des Forests et Institut pour le
Developpement Forestier, 243p.
DGF (2009) : Rapport annuel d'activités - Exercice
2008. Kinshasa, 25p.
Differt D. (2001). Phenologie des espèces arborees -
Synthèse bibliographique et Analyse des donnees du Reseau national de
Suivi à long terme des Écosystèmes forestiers (RENECOFOR).
Rapport scientifique. Nancy : UMR ENGREF-INRA - LERFOB, 97 p. + annexes.
Doucet J-L. (2003). l'Alliance délicate de la
gestion et de la biodiversité dans les forits du Gabon.
Thèse de doctorat, Faculte Universitaire des Sciences Agronomiques
Gembloux, 323 p + Annexes.
Durand R. (1967) : Action de la
température et du rayonnement sur la croissance. Annales de
Physiologie Végétale, 9 : 5-27.
Durieu de Madron L. & Daumerie A. (2004). Fruiting
diameter in different species in the natural forests of the Central African
Republic. Bois et Forests des Tropiques, 281 (3) : 87-95.
Durrieu de Madron L., Favrichon V., Dupuy B., Bar Hein A.,
Maitre H.F. (1998a). Croissance et productivité en forêt dense
humide : bilan des expérimentations dans le dispositif de Mopri -
Côte d'Ivoire (1978 - 1992). CIRAD-Forest, Montpellier. Serie
FORAFRI, document 3, 72 p.
Fdy B. (1991) : Variabilité du débourrement
végétatif du Sapin de Céphalonie (Abies
cephalonica
Loudon) en plantation. Annales des Sciences
forestières, vol. 48, 1991, pp. 73-85
Favrichon V. (1997). Réaction de peuplements
forestiers tropicaux à des interventions sylvicole. Bois et Forest
de tropiques, 254 (4) : 1-24.
Galoux, A. ; Schnock, G. ; Grulois, 0. (1967) : La Variabilite
phenologique et les conditions climatiques. -Bulletin de la Societe royale de
Botanique de Belgique, vol. 100, pp. 309-314.
Gillet J.F., Ngalouo B. et Missamba-Lola A. (2008).
Rapport d'analyse - Volet dynamique forestière. Projet CIB-FFEM
- "Suivi du programme dynamique forestière - agroforesterie -
inventaires faunes". 99 p.
*RTRX3) I(TEli) $ SSRIMIIIIW\TRIMERNEOMMSDKOBVRUWAP Splptli
l'p\MITIA la modelisation des peuplements complexes. Thèse de doctorat
en Sciences forestières, ENGREF, 362 p.
Habari J.P. (2004) : Etude ecologique et phytosociologique de la
vegetation de la vallee de la Lukaya/ Kinshasa. Mem. DEA, Depart. Biol. Fac.
Sciences/ UNIKIN. 30 p + Annexes.
Hecketsweiler P. (1992). Phénologie et
saisonnalité en foret gabonaise, l'exemple de quelques espèces
ligneuses. Thèse de doctorat, Université de Montpellier II,
266p.
Kikufi, B. (200) : Etude de la végétation et de la
flore du Bassin de la rivière Funa à Kinshasa. Mém.
Licence, Dép. Biol., Fac. Sciences/ UNIKIN, Inédit, 85 p.
Kouadio L. (2008). Mesures sylvicoles en vue
d'améliorer la gestion des populations d'essences forestières
commerciales de l'Est du Cameroun. Thèse, FUSAGx, Gembloux, 253p. +
annexes.
Kramer K. (1997): Phenology and growth of European
trees in relation to climate change. Phenology, Seasonal Climates
12 : 39-50.
Latham P. et Konda K.M. (2006). Quelques plantes utiles de la
Province du Bas-Congo, RD Congo ; 2ème éd. Paul Latham
et Konda ku Mbuta, Bruxelles, p. 204
Lebourgeois F., Differ J., Granier A., Breda N. & Ulrich
E. (2002) : Premières observations phénologiques des
peuplements du réseau national de suivi à long
terme des écosystèmes forestiers (renecofor). Rev. For.
Fr. LIV - 5-2002, pp407-418.
Lebrun J.-P. et Stork A. (2008). Tropical african Flowering
plants. Ecology and distribution. vol.4 Fabaceae
(Desmodium-Zornia). Conservatoire et Jardin botaniques de la Ville de
Genève. Hors série, 291 pp.
Lelo (2008). Ville & environnement de Kinshasa. Ed.
l'harmattan 2008, paris 281p
http : // www.librerie
harmanttan.com.
Malaisse F. (1967). Contribution à l'étude
des hetraies d'Europe occidentale - Note 6 : aperçu climatologique et
phénologique relatif aux hetraies situées sur l'axe Ardennes
belges. Provence. 14ème Congrès IUFRO, Munich,
section 21, pp. 325-334.
Malaisse, F. (1964) : Contribution à l'étude des
hêtraies d'Europe occidentale. Note 6 : aperçu climatologique et
phénologique relatif aux hêtraies situées sur l'axe
Ardennes belges Provence. 14e Congrès IUFRO, Munich, 1967, II, section
21, 1967, pp. 325-334.
Malele S. (2005). Présentation du secteur forestier de la
RDC. Exposé compagnonnage industriel Belgique-RDC-France, Kinshasa, le
13 juin 2005, 41p.
MALOM (2009). Etude floristique et ethnobotanique des plantes du
Quartier Livulu dans la commune de Lemba. Tfc Unikin Fac. Sciences dep.
Biologie 62p.
Mandango M.A, Idrissa A., Kayumba L. et Malulu N. (1999).
Etude écologique et phytosociologique de la forêt à
Pentaclethra macrophylla au sud de Kinshasa (RDC). Doc. Phytos, N.S,
vol. XIX, Camerino, 349-
Mathieu G. (2005). Phénologie et changement
climatique. Centre de Recherches sur les écosystèmes
d'altitude (CREA), Programme phénoclim, déc. 2005, 5p.
Medjibe V. et Hall J. (2001). Seed dispersal and its
implications for sylviculture of African Mahogany (Entandrophragma
spp.) in undisturbed forest in the Central African Republic. Forest Ecology and
Management, 5810 : 1-9.
Menga P. (2005). Etude phytosociologique de vegetation
Forestière de « symphonies naturelles » a Kinshasa/Ngaliema.
Mem. UNIKIN Fac. Sciences dep. Biologie. Opt. Ecologie 47p.
Menga P. (2007). Contribution à l'étude
floristique et phytosociologique de la végétation de la region de
Bolobo-Mushie (Bandundu/RDC). Mem. DEA, Laboratoire de Botanique
Systématique et d'Ecologie Végétale, Université de
Kinshasa, 79 p.
Mpoto I. (2007). Carte Postale du District de Maï-Ndombe, RD
du Congo /Bandundu. BEDRI, Kinshasa, 6p. «
http://fr.wikipedia.org/wiki/Mai-Ndombe
%28district%29 ».
Muleka (2003) : Inventaire des Vertebres terrestres de la forest
des Symphonies Naturelles à Kinshasa/ Ngaliema. Mem. Lic., Dep. Biol.
ISP/Gombe Inedit, 43 p.
Newberry D.M., Chuyong G.B. et Zimmermann L. (2006). Mast
fruiting of large ectomycorrhizal African rain forest trees - importance of dry
season intensity, and the resource-limitation hypothesis. New
phytologist, pp. 561-579.
Newstrom L.E., Frankie G.W. et Baker H.G. (1994). A new
classification for plant phenology based on flowering patterns in lowland
tropical rain forest trees at La Selva, Costa Rica. Biotropica
26, 141-159.
Ngalula M ; (2004) : Caracteristiques physico-chimiques et
ichthyologiques des ruisseaux de « Symphonies Naturelles » à
Kinshasa/ Ngaliema. Mem. Lic., Dep. Biol., ISP/ Gombe, Inedit, 23 p.
Nshimba H. (2008). Etude floristique, ecologique et
phytosociologique des forests de l'île Mbiye à Kisangani, RD
Congo. Thèse de doctorat, ULB, 272 p.
Pauwels L. (1993). Nzayilu N'ti - guide des arbres et arbustes
de la region de Kinshasa Brazzaville. Scripta Botanica Belgica. Volume 4.
Jardin botanique national de Belgique, Meise, Belgium. 495 p.
Saint moulin (2005) : Atlas de l'organisation administrative de
la République démocratique du Congo, centre d'études pour
l'action sociale CEPAS-Kinshasa,235p
Schnelle, F. (1955): Plante - Phenologie. Akademische
Verlagsgesellschaft Geest & Porting K.-G., Leipzig.
Sepulchre F., Dainou K. et Doucet J.L. (2008). Etude de la
vulnérabilité de 18 essences ligneuses commerciales d'Afrique
centrale reprise sur la liste rouge IUCN. Gembloux, Nature plus - FUSAGx,
Laboratoire de Foresterie des Régions tropicales et subtropicales. 50
p.
7 IXD-rEz MEEM EZ EUIRTITtNC-QN-11'S IIIIX- EF-Qtral-I
IK-QtifTEDARQ SIDEIX-II-s SIiQcISIXx arbres. Tome 2. CTA, Wageningen, Pays Bas.
pp. 465#177;1271.
Tchinda A.T. (2008). Millettia laurentii De Wild.
[Internet] Fiche de Protabase. Louppe, D., Oteng-Amoako, A.A. & Brink, M.
(Editeurs). PROTA (Plant Resources of Tropical Africa /E5 HMMFKIFHM EYpJplilHM
ECHEerpirtnH EtrFstIFalHil E : alHntuHa E3 CA M EBaM.E <
http://database.prota.org/recherche.htm>.
Traissac S. (2003). Dynamique spatiale de Vouacapoua americana
(Aublet), arbre de forest à répartition agrégée.
Université Claude Bernard - Lyon 1, 216 p. (Thèse).
Van Schaik CPo (1986). Phenological changes in a Sumatran rain
forest. J. Trap. Ecal. 2:327- 47
9 al-Q 71 -1 ) aXr- 7 7 (19M). S Ibr-III-sTERTITWE-Qs-NI'S IBTX-
&-QtrIl-I ltiGE M5 ( , Agence de Coopération Culturelle et
Technique, Paris, France. 565 p.
White L. et Edwards A. (2001). Conservation en forest
pluviale africaine - Méthodes de recherché. Ed, WCS, New
York, 456 p.
Withmore T.C. (1975) : Tropical Rain Forest of the Far East.
Clarendon Press, Oxford, 282p. Whitmore T.C. (1975). Tropical Rain
Forest of the Far East. Clarendon Press, Oxford, 282p.
Wilks C. et Issembe Y. (2000). Les arbres de la Guinée
Equatoriale: Guide pratique GIC-QtifTEDARQ: rtigIRQ cRQt1Q-Qtal-1
13ERj-IA&8 5 ( ) ,TBata, Guinée Equatoriale. 546 p. + photos.
Yalibanda Y. (1999). Phénologie en forest dense de
Ngotto (R.C.A.) - Bilan de trois années tiIRETHITtiRn.
Séminaire FORAFRI de Libreville (12-16 octobre 1998). In : Nasi
R., Amsallem I. & Drouineau S. (eds). La gestion des forests denses
africaines agjRgati'hgiE CActes du séminaire FORAFRI de Libreville,
12-16 octobre 1998. Cédérom, CIRAD-forest, Montpellier, 24 p.


