|
Université Cheikh Anta Diop de Dakar
(UCAD)
Faculté des Sciences et Techniques (FST)
Institut
des Sciences de l'Environnement (ISE)

THESE DE DOCTORAT D'ETAT ES SCIENCES

Présentée et soutenue publiquement le 16
novembre 2009
SUJET
Potentiel et dynamique des stocks de carbone
des savanes soudaniennes et soudano-
guinéennes du Sénégal.
par
Dr. Cheikh Mbow
Jury
Président Amadou Tidiane Ba, Professeur
Titulaire, Département de Biologie
Végétale. Université Cheikh Anta Diop,
Dakar, Sénégal.
Rapporteurs Bienvenu Sambou, Maître de
Conférences, Institut des Sciences de l'Environnement. Université
Cheikh Anta Diop, Dakar, Sénégal
Amadou Tahirou Diaw, Professeur Titulaire,
Département de Géographie, Université Cheikh Anta Diop,
Dakar, Sénégal.
Membres Brice Augustin Sinsin, Professeur
Titulaire, Laboratoire d'Ecologie
Appliquée, Faculté des Sciences Agronomiques.
Université Abomey-Calavi, Cotonou, Bénin.
David Skole, Professeur, Département de
Foresterie, Michigan State University-East Lansing, USA.
Michel Verstraete, Global Environment Monitoring
(GEM) Unit, Centre Commun de Recherche-Union Européenne, Ispra (VA),
Italie.
DEDICACES
CE TRAVAIL EST DEDIE A
FEUE SOKHNA AICHA
THIAM POUR QUI L'HUMANISME
EST L'ESSENCE DE LA VIE.
A
FEU EL HADJI CHEIKH PENDA MBOW
LE SAGE PATRIARCHE, FEDERATEUR DE CONSCIENCES
ET POUR QUI LE
BIEN QU'ON FAIT POUR LES AUTRES
EST LE SENS PRINCIPAL DE LA VIE
QUE DIEU LES ACCUEILLE DANS SON AUGUSTE
PARADIS ... AMEN
Remerciement
L'Homme est le résultat d'une somme de circonstances et
de conditions d'existence, parfois aléatoires, souvent liée
à un déterminisme perçu ou abstrait qui sculpte sa
conscience et son comportement, ses succès et ses échecs. Les
relations sociales qui déterminent dans une large mesure le destin
individuel, sont souvent le fruit d'un certain hasard, ou imposé par le
groupe. Jacques Monod disait que "tout ce qui existe dans l'Univers est le
fruit du hasard et de la
nécessité". Sans chercher à
séparer les deux concepts ou verser dans une logique existentialiste,
nous considérons que, ce que l'homme réalise dans sa courte vie
dépasse souvent sa seule volonté. Il serait par conséquent
injuste de limiter l'uvre d'une personne à son mérite
exclusif.
Sous ce rapport, ce travail est alors une traduction des
dispositions et avantages que notre Créateur nous a accordés,
ainsi qu'une transcription des idées et appuis obtenus auprès de
plusieurs personnes et institutions de recherche pour lesquelles je souhaite
traduire ma profonde gratitude.
En observant les plantes et le secret qui les entoure, en
parcourant l'espace dans sa diversité, sa complexité et ses
mystères, j'ai très vite compris que la FOI ne pouvait
naître qu'en étant en contact permanent avec l'uvre immense de
DIEU. Je rends grace à Allah pour m'avoir donné les dispositions
physiques et morales qui m'ont permis de réaliser ce travail.
A travers la miséricorde du Créateur, j'ai
croisé le chemin du Professeur Amadou Tidiane Ba. Il m'a souvent
invité à la remise en question perpétuelle de
moi-même pour ne pas verser dans la facilité des
stéréotypes, des idées reçues et des dogmes. Sa
culture et son altruisme m'ont donné la chaleur nécessaire pour
couver les idées scientifiques consignées dans ce document.
Cette influence a été complétée
par l'humanisme et le paternalisme des Professeurs Bienvenu Sambou et Amadou
Tahirou Diaw, respectivement Directeur de l'Institut des Sciences de
l'Environnement et du Laboratoire d'Enseignement et de Recherche en
Géomatique de l'UCAD, pour la confiance qu'ils m'ont faite malgré
les connaissances plutôt balbutiantes que j'avais en 2004 sur la
séquestration du carbone. Ils m'ont `com-pris' et m'ont conduit avec une
douce rigueur à aimer la recherche et avoir confiance en
moi-même.
Je remercie les membres du jury : Professeur David Skole
(USA), Dr Michel Verstraete (Belgique-JRC/EU) et Professeur Brice Sinsin
(Bénin) qui ont bien voulu accepter de juger ce travail. Le Professeur
Rasmussen a été parmi les premiers à inspirer ce travail
et le Professeur Skole a été d'un apport inestimable pour la
réalisation de nombreuses tâches liées à cette
recherche. La profondeur scientifique de Michel Verstraete et ses conseils
ont
aiguisés en nous une forte curiosité dans le
domaine de la dégradation des terres au Sahel. L'esprit critique et
l'immense culture du Professeur Sinsin ont forgé en moi une estime
certaine à son égard. Le symbole que ces professeurs
représentent pour la science traduit en moi un immense honneur de les
avoir comme examinateurs. Je joins à ces remerciements une motion
particulière à Professeur Kjeld Rasmussen (Danemark) et Dr Sandra
Brown (WINROCK-Int., USA) pour leurs nombreux conseils dans
l'élaboration de ce travail.
Pour réaliser ce travail j'ai eu le soutien financier
de plusieurs institutions qu'il me plaît de remercier ici : SUN
(Sustainable Use of Natural Resources-EU), IFS (International
Foundation for Science), Start (Global Change SysTem for Analysis,
Research and Training-US) ; et DGG (Departement of Geography and
Geology-University of Copenhagen-Denmark). J'espère que ce
Mémoire sera à la hauteur des attentes et espoirs portés
à ce travail.
Pour leurs encouragements et leur assistance aussi bien
matérielle que morale, je me dois de remercier chaleureusement tous mes
collègues de l'Institut des Sciences de l'Environnement : Bienvenu
Sambou, Abou Thiam, Abdoulaye Sène, Assane Goudiaby, Cheikh Diop, Cheikh
Ibrahima Niang, François Matty et Henri Mathieu Lô. J'associe
à ces remerciements, les collègues de la FST (notamment le Pr.
Matar Mour Seck et le Pr Kandioura Noba) ainsi que tout le personnel
administratif et technique de l'ISE que sont Ousseynou Ndiaye, Doudou Samba,
Ababacar Sedikh Cissé, Linda Faye, Oula Ndiaye, Sounkarou Niassy, Oumar
Salif Gueye, ainsi qu'à Idrissa Guiro, Mamadou Diop et Fatima Niang.
J'adresse mes chaleureux remerciements à la Direction
des Eaux, Forêts, Chasse et de la Conservation des Sols (DEFCCS),
notamment son ex-Directeur Colonel Matar Cissé et les Inspecteurs des
Régions d'études que sont Babacar Faye de Tambacounda et
Souleymane Kigera de Fatick. J'associe à ces remerciements le Colonel
Bassirou Sougoufara, Maguette Kairé, Ousmane Diallo et tous les membres
du groupe de travail du projet PREFER.
Mon affiliation au LERG (Laboratoire d'Enseignement et de
Recherche en Géomatique) de l'ESP m'a permis de développer une
collaboration fructueuse avec des collègues et des chercheurs dont je
salue l'ouverture d'esprit et la sollicitude aux moments cruciaux de la
réalisation de ce travail. Que Grégoire Leclerc, Mbaye Diop,
Tidiane Sané, Alla Manga, Balla Diéye, Séraphin
Dorégo, Khady Diouf Ndir, Amadou Kâ, et à tous mes
étudiants Aminata Gueye Sarr, Awa Diouf Sylla, Idrissa Guiro, Momadou
Sow, Yacinthe Sambou, Aminata Diop, Patrick Ondo Okama, etc., trouvent à
travers ce travail un motif de satisfaction.
les nombreuses opportunités que des personnes comme
Assize Touré, Aliou Diallo, Aliou Kâ, Jacques André Dione,
Alioune Touré, Déthié Soumaré Ndiaye, et tous les
chercheurs et techniciens de ce centre ; m'ont permis de saisir.
A travers Cheikh Ndiaye Sylla et Madeleine Sarr, la DEEC
(Direction de l'Environnement et des Etablissements Classés) m'a
été d'un apport crucial pour les nombreuses formations et
ateliers (en particulier ceux du Comité National sur les Changements
ClimatiquesCOMNAC) qui m'ont permis d'être à jour sur les
questions importantes que soulève la séquestration du carbone
dans le secteur forestier.
Le Département de Géographie et Géologie
de l'Université de Copenhague a été pour moi un creuset
fertile d'échanges scientifiques qui m'a permis de tirer de larges
connaissances avec Kjeld Rasmussen, Anette Reenberg, Rasmus Fensholt, Ole
Mertz, Inge Sandholt, Thomas Theis Nielsen, Bo Elberling, Sarah Choudhary, Lars
Krogh, Mads Olander Rasmussen, Annette Norgaard, Inge Sandholt, Jens
Grundtmann, Bjarn Fog). Ma participation aux activités du GLP
(Global Land Project-IGBP/IHDP) avec Anette Reenberg, Tobias Laganke,
Lars Jørgensen et ma collaboration avec Anne Mette Lykke
(Université de Aarhus) n'ont fait que renforcer cette coopération
utile et fructueuse avec le Danemark. Le financement de mon séjour au
Danemark par le projet FIVA (International Research School of Water
Resources) a été possible grâce à son Directeur
Karsten H. Jensen et l'appui de Frederik Uldall que je remercie bien
vivement.
Ce travail ne pouvait pas se réaliser sans l'appui de
MSU (Michigan State University-USA) qui à travers le Professeur David
Skole (Directeur du GOES : Global Observatory for Ecosystem Services)
et son équipe dont Moussa Dieng, Oscar Casteneda, Jay Samek, Walter
Chomentowski, m'ont aidé pour le traitement de certaines données.
Je voudrais souligner l'apport inestimable de Dr. Sophan Chhin du
Département de Foresterie, de la même Université, sur les
analyses dendrochronologiques qui ont été difficiles à
faire ; vu la complexité du travail.
Le havre de paix que constitue ma famille à
été déterminant pendant ces années de recherche et
je veux sincèrement remercier mon épouse Salimata
Néné Thiam Mbow et mes enfants Mathiam et Aïcha pour la
douceur familiale et l'acceptation de mes longues absences pour les besoins de
cette thèse. Je souhaite que ce travail soit une véritable
satisfaction pour mon Père El Hadji Saliou Mbow, le protecteur, pour ses
prières et ses conseils ; à mes oncles Serigne, Cheikh, Falilou
et Moustapha, à mes tantes Ndéye Awa Mbow (Badiene), Khady Gueye
(Mère Khadi), Coumba Sylla, Coumba Thiam et Oumy Mbaye. Je remercie tous
mes frères et surs Mbow et Tall pour leur soutien permanent. Je souhaite
que tous mes amis, Thierno Birahim Gueye-Thier, Daha Cherif Aïdara-Che,
Ibrahima Niang-Ibou, Doudou
Diop-Dou, Sokhna Helène Diop, Gora Sarr-Farba,
Moustapha Thiam-Kamzo, Mokhtar LoMakhou, Mame Samba Mbaye-Serignebi et à
leurs épouses et époux respectifs, trouvent à travers ce
travail ; l'expression de nos sentiments de profonde reconnaissance.
Résumé
Ce travail a pour objectif de contribuer à combler le
déficit d'information sur le potentiel de séquestration de
carbone dans les écosystèmes de savane. Il a porté sur six
Forêts Classées (Wélor, Patako, Ouli, Bala-Est, Kantoora et
Mampaye) choisies pour couvrir les écosystèmes soudaniens et
soudano-guinéens du Sénégal. La démarche a
consisté en plusieurs étapes que sont : l'inventaire forestier,
la collecte de données de biomasse pour l'élaboration de
modèles allométriques, l'analyse dendrochronologique, le suivi
des formations ligneuses par des parcelles suivies depuis 1993 et cartographie
de la dynamique des formations végétales.
Sur la base des données collectées et
analysées, trois modèles allométriques utilisant des
régressions quadratique, cubique et polynomiale d'ordre 3 ont
été mis au point. Ces régressions ont permis d'une part,
d'estimer les stocks de biomasse avec un r2 > 0,9. D'autre part,
l'application des tests ANOVA (analyse de la variance) a montré une
faible différence entre la moyenne de la série
expérimentale et celles issues des modèles. Les résultats
obtenus indiquent des stocks de carbone (biomasse sur pied) très faibles
pour les formations dégradées de la Forêt Classée de
Wélor (entre 3 et 7 tC/ha) alors que les Forêts Classées de
Mampaye (20-50 tC/ha) et Kantora (40-47 tC/ha) ont les stocks les plus
importants. Des cas intermédiaires sont notés à Ouli
(12-20 tC/ha), Bala (18-28 tC/ha), et Patako (13-33 tC/ha). L'essentiel de la
biomasse se situe chez les individus de gros diamètre (> 20 cm).
Cependant, la dynamique de séquestration du carbone est portée
par les petits individus en croissance.
Le suivi de la végétation ligneuse montre que
l'essentiel des stocks de carbone du dispositif est porté par quelques
espèces, surtout celles à croissance rapide. Les espèces
qui contribuent le plus à la biomasse sont Pterocarpus erinaceus,
Bombax costatum et Terminalia macroptera. La contribution aux
stocks de carbone est fortement liée aux possibilités de
l'espèce à croître vite et à accumuler beaucoup de
biomasse. Les espèces à croissance relativement rapide sont
Detarium senegalense et Bombax costatum, suivies par
Anogeissus leiocarpus, Lannea microcarpa, Pterocarpus erinaceus,
et Terminalia macroptera.
Les analyses dendrochronologiques à partir de la
largeur des cernes fournissent des indications quant à la relation avec
des paramètres comme la pluviométrie. Mais des contraintes
d'analyse plus fine des premiers résultats subsistent encore même
s'ils confirment la faible vitesse de croissance des espèces de savane.
Les jeunes plants de savane ne se développent pas forcément plus
vite que les plantes affranchies à cause probablement des effets de
nombreux facteurs de stress comme les feux, l'aridité, la
compétition dans la croissance,
La faible productivité nette de ces
écosystèmes de savane fait que les quantités de carbone
séquestrées ne sont pas très élevées
comparées à d'autres types de formations végétales
comme les forêts tropicales, ce qui fait que le bénéfice
pour les projets MDP doit être prudemment envisagé. Les
résultats de la modélisation montrent l'effet très
destructeur de nombreuses pratiques comme les feux, les coupes de bois, les
défrichements, le surpâturage qui réduisent
significativement les stocks de carbone. Cette dégradation de
l'environnement due aux facteurs climatiques, politiques, et économiques
rend les populations très vulnérables, les obligeant à
opter pour des stratégies d'adaptation variées et qui valent plus
que les possibilités offertes par le marché actuel du carbone.
Abstract
This study is a contribution on information needs for the
potential carbon sequestration of savanna ecosystems. The research is
implemented in six Protected Forests of Senegal (Welor, Patako, Ouli, Bala-Est,
Kantoora and Mampaye), selected in various SoudanoGuinean and Guinean
ecosystems. The method is manifold including forest inventory, biomass
collection for allometric models development, dendrometry analysis, survey
plots monitored since 1993 and vegetation cover mapping.
From the data collected and analysed, 3 allometric models are
suggested based on quadratic, cubic and polynomial 3rd order
regressions. Carbon stock (above ground woody biomass) assessment based on
these models give a coefficient of determination r2 >0.9 and the
ANOVA tests shows a very low difference between mean values of experimental
data and model outputs. The application of these model on inventory data shows
low carbon stock for Welor (3-7 tC/ha), but for Mampaye (20-50 tC/ha) and
Kantora (40-47 tC/ha) the carbon stocks was better. Intermediate cases are Ouli
(12-20 tC/ha), Bala (18-28 tC/ha), and Patako (13-33 tC/ha). Most of the
biomass is trapped on individuals with DBH> 20 cm. Nevertheless, the carbon
dynamics is carried by young plants with a real growth potential.
Permanent plot data (3 plots of 1ha) show that carbon stocks
are essentially held by few species; mainly those with rapid growth. Species
with high contribution to total biomass are Pterocarpus erinaceus, Bombax
costatum and Terminalia macroptera. Hence, the specie
contribution to carbon stocks is strongly linked with its frequency, its
possibility to grow fast and its resilience to adverse stresses. The species
with rapid growth are Detarium senegalense and Bombax
costatum, followed by Anogeissus leiocarpus, Lannea microcarpa,
Pterocarpus erinaceus, and Terminalia macroptera.
The analysis of tree rings using dendrometry methods revealed
the good potential for tree dating and the possibility to link the tree ring
growth with parameters such as rainfall. The use of this method is very fuzzy
for tropical samples, but some samples gave good results that confirm the slow
growth rate of savanna species. With multiple strains in savanna ecosystems
such as fire, drought and competition, young plants do not necessarily have a
faster growth rate than mature trees as suggested by classical ecological
theories.
The low net productivity of savanna ecosystems results in
relatively low carbon sequestration compared to tropical forest stands, and
this required a measure of cautious for potential benefits from CDM projects.
The model outputs of carbon dynamics show several biomass removal factors such
as fires, wood extraction, slash and burn agriculture, overgrazing, with direct
effects on carbon stocks. This overall environmental degradation are associated
with climatic, political and economical factors that leads to high population
vulnerability that require various adaptation strategies, more vital for
population resilience to CC than the new misty carbon economy.
Par délibération, la Faculté
des Sciences et Techniques et l'Institut des Sciences de l'Environnement ont
décidé que les opinions émises dans les dissertations qui
leur sont présentées doivent être considérées
comme propres à leurs auteurs et qu'ils n'entendent leur donner aucune
approbation ni improbation.
Liste des sigles et abréviations
ACP : Analyse en Composante Principal
AMMA: Analyse Multidisciplinaire de la Mousson
Africaine
AND : Autorité Nationale
Désignée ANOVA: Analysis of Variance
AVC : Analyse des Vecteurs de Changement
BNDS: Banque Nationale de Développement
du Sénégal CASS : Carbon Accounting Simulation
Software
CC : Changement Climatique
CCNUCC : Convention Cadre des Nations Unies pour
les Changements Climatique
CBERS : Chinese, Brazilian Earth Resource
Satellite
CER: Certified Emissions Reduction
CNUED : Conférence des Nations Unies pour
l'Environnement et le Développement
COP : Conférence des Parties
CSE: Centre de Suivi Ecologique
CVD: Comité Villageois de
Développement
DBH: Diameter at Breath Hight (Diamètre
à Hauteur de Poitrine)
DEEC : Direction de l'Environnement et des
Etablissements Classés
DEFCCS: Direction des Eaux et Forêts,
Chasse et de la Conservation des Sols
ENRECA : Enhancement of Research Capacities
ESA: European Space Agency
ESP : Ecole Supérieure Polytechnique
ETM: Enhanced Thematic Mapper
FAO : Organisation des Nations Unies pour
l'Alimentation et l'Agriculture
FC: Forêt Classée
FEM : Fonds Mondial pour l'Environnement
FST : Faculté des Sciences et Techniques
GEMI : Global Environmental Monitoring Index
GES : Gaz à Effet de Serre
GIE: Groupement d'Intéret Economique
GIEC : Groupe Intergouvernemental pour l'Etude
du Climat (appelé aussi IPCC en anglais)
GIMMS : Global Inventory and Mapping System?
GOANA: Grande Offensive Agricole pour la Nourriture et
l'Abondance
GPF: Groupement de Promotion Féminine
GPS : Global Positioning System
Gt : Giga tonnes
GTZ : Gesellschaft für Technische
Zusammenarbeit (GTZ) est l'agence de coopération
technique allemande pour le développement.
IPCC : Intergovernmental Pannel for Climate
Change
ISE : Institut des Sciences de l'Environnement
ISF : International Foundation for Sience
LAI : Life Area Index
LCCS : Land Cover Classification System
LERG : Laboratoire d'Enseignement et de
recherche en Géomatique
LIDAR : Light Detection and ranging
LULUCF : Land use, Land Use Change and Forestry
MDP : Mécanisme de Développement Propre
MEA: Millenium Ecosystem Assessment
MEPN : Ministère de l'Environnement et de
la Protection de la Nature
MERIS : Medium-spectral Resolution, Imaging
Spectrometer
MODIS: Moderate Imaging System
NASA : National Aeronautics and Space
Administration NDVI : Normalized Difference Vegetation Index
(Indice de vegetation par la Difference Normalisée)
NOAA : National Oceanic and Atmospheric
Administration
NPA: Nouvelle Politique Agricole
OCB: Organisation Communautaire de Base
OMD: Objectifs du Millénaire pour le
Développement ONCAD: Office National
ONG: Organisation Non Gouvernementale
PAEFK : Projet
PAF : Plan d'Action Forestier
PAGERNA : Projet Autopromotion et Gestion des
Ressources Naturelles
PANA : Plan National pour l'Adaptation au CC
Pg : Peta grams
PGCRN : Projet de Gestion Communautaire des
Ressources Naturelles
PGIES : Projet de Gestion Intégré
des Ecosystèmes du Sénégal
PIN: Project Idea Note
PIR : Proche Infrarouge
PLD: Plan Local de Développement
PNE : productivité Nette de
l'Ecosystème
PNUE: Programme des Nations Unies pour
l'Environnement
PPB : Productivité Primaire Brute
PPG : productivité Primaire Globale
PPM : Parties par Millions
PPN : productivité Primaire Nette
PREFER : Projet de Réhabilitation des
Ecosystèmes Forestiers et Ruraux
PREVINOBA : Projet de Reboisement Villageois dan
le Nord-Est du Bassin Arachidier
PROBOVIL : Projet de Boisement Villageois
PROGEDE : Projet de Gestion des Energies
Domestiques et de Substitution
PROGERT : Projet de Gestion et Restauration des
Terres dégradées du Bassin Arachidier
PRS : Projet de Reboisement du Senegal
RADAR : Radio Detection and ranging
REC : Réduction d'Emission
Certifiée
REVA: Retour Vers l'Agriculture
SAVI : Soil Adjusted Vegetation Index
SIEF : Systèmes d'Information Ecologique
et Forestier SIG: Système D'Information
géographique
SODEVA : Société de
Développement de l'Agriculture SPOT : Satellite pour
l'Observation de la Terre
SUN : Sustainable Use of Natural Resource
TS : Température de Surface
TSAVI : Transformed Soil Adjusted Vegetation
Index UCAD : Université Cheikh Anta Diop de Dakar
UTM : Universal Tranverse Mercator
WGS : World Geodetic System
Liste des figures
Figure 1. Bilan du CO2 entre 1980
et 1990 14
Figure 2. Evolution des concentrations
de CO2 atmosphérique au niveau de 4 stations de
mesure de 1981 à 2000. 14
Figure 3. Dynamique de la
concentration du CO2 à Mauna Lao (Mesures in situ) 15
Figure 4. a) Émissions
annuelles de GES anthropiques dans le monde, 1970-2004 ; b) Parts respectives
des différents GES anthropiques dans les émissions totales de
2004, en équivalent- CO2 ; c) Contribution des différents
secteurs aux émissions totales de
GES anthropiques en 2004, en équivalent- CO2 (IPCC,
2008) 16
Figure 5. La plante et le flux du
Carbone (Waring et Running, 1998) 22
Figure 6. A) Volumes de carbone
conservés ou piégés en fonction des options
d'aménagement (Pg/an) ; B) Taux annuels moyens de conservation et de
séquestration du carbone par région géographiques (Pg/an)
(Source : Brown, 2003)
31
Figure 7. Carte des zones
climatiques et des réserves forestières du
Sénégal 49
Figure 8. Carte des densités de
population du Sénégal (base DTGC, 2004) 50
Figure 9. Carte d'occupation du sol du
Sénégal (Source JRC, GL000) 51
Figure 10. Carte des sols du
Sénégal 52
Figure 11. Localisation et typologie
des stations climatiques utilisées 53
Figure 12. Moyenne des
températures terrestres depuis la fin du 19ième Siècle
(a). Moyenne des températures terrestres et océaniques depuis la
fin du 19ième Siècle (b)
(Hadley-Centre, 2003) 54
Figure 13. Evolution des anomalies de
température (IPCC, 2008) 55
Figure 14. Ecart à la moyenne
des températures moyennes mensuelles à Kaolack, Kolda,
Tambacounda et Ziguinchor (1951-2003) 56
Figure 15. Evolution interannuelle de
la pluviométrie (1921/1922-2006) : 57
Figure 16. Dynamique spatiale des
précipitations par décennie 59
Figure 17. Les parties
concernées par la mesure des branches et des rameaux 72
Figure 18. Estimation du volume de
tronc trop volumineux 74
Figure 19. Mesures à effectuer
pour le calcul du volume d'un disque pour la densimétrie. 74
Figure 20. Tests des méthodes
allométriques utilisés dans les écosystèmes
tropicaux 77
Figure 21. Mensuration pour mesure de
volume de disque 79
Figure 22. Illustrations de quelques
étapes de la méthode 80
Figure 23. Comparaison des courbes de
la taille des diamètres et de la biomasse 82
Figure 24. Nuage de points du DBH et
de la biomasse (courbe non linéaire) 83
Figure 25. Représentation des
courbes des différentes régressions 86
Figure 26. Comparaison entre les
modèles FAO et les données de terrain (grands décalage
surtout par rapport aux gros diamètres) 87
Figure 27. Variations des mesures de biomasse par les
différents des modèles qui ont un bon
coefficient de détermination. 88
Figure 28. Comparaison des
résidus des mesures avec les estimations de la FAO 90
Figure 29. Comparaison des
résidus des mesures avec les modèles in situ. 90
Figure 30. Différence entre les
modèles quadratique et les fonctions cubiques 96
Figure 31. Schéma de la logique
de croissance des diamètres de tronc. 97
Figure 32. Biomasse estimée par
les modèles cubiques 99
Figure 33. Biomasse estimée par
les modèles Quadratiques 99
Figure 34. Biomasse estimée par
les modèles Polynomiaux 99
Figure 35. Maillage de la
Forêt Classée de Patako (mailles carrées de 250 m de
coté) 111
Figure 36. Schéma du dispositif
d'inventaire 112
Figure 37. Répartition des
effectifs par classe de diamètre 117
Figure 38. Répartition de la
biomasse en fonction des classes de diamètre pour chaque strate
|
des 6 Forêts Classées.
|
119
|
|
Figure 39. Localisation des parcelles de suivi
de la végétation
|
123
|
|
Figure 40. Délimitation des lignes de
base de la parcelle permanente
|
125
|
|
Figure 41. Délimitation des bandes de
la parcelle permanente
|
125
|
|
Figure 42. Délimitation des placettes
de la parcelle permanente
|
126
|
|
Figure 43. Détermination de la position
des individus dans les placettes
|
127
|
|
Figure 44. Mesure de DBH avec un compas
forestier
|
128
|
|
Figure 45. Abondance des
espèces des 3 placettes en 1993-1997-2003-2005
|
130-131
|
Figure 46. Croissance moyenne annuelle
des espèces les plus abondantes dans les 3 parcelles de suivi de la
végétation 133
Figure 47.
Comparaison de la vitesse de croissance des diamètres pour les trois
périodes de
suivi 134
Figure 48. Evolution de la
pluviométrie à la station de Tambacounda 134
Figure 49. Dynamique de la biomasse
totale pour la parcelle 1 135
Figure 50. Dynamique de la biomasse
totale pour la parcelle 2 136
Figure 51. Dynamique de la biomasse
totale pour la parcelle 3 136
Figure 52. Dynamique des classes de
diamètres de la parcelle 1 137
Figure 53. Dynamique des classes de
diamètres de la parcelle 2 137
Figure 54. Dynamique des classes de
diamètres de la parcelle 3 137
Figure 55. Contribution des
espèces à la biomasse totale de la parcelle 1 139
Figure 56. Contribution des
espèces à la biomasse totale de la parcelle 2 139
Figure 57. Contribution des
espèces à la biomasse totale de la parcelle 3 139
Figure 58. Courbe d'évolution
des diamètres des espèces à croissance relativement
rapide
140
Figure 59. Evolution de la biomasse
des espèces à croissance relativement rapide 141
Figure 60. Appareil Eltra SC-500 pour
l'analyse du taux de carbone et préparation des
échantillons 143
Figure 61. Schéma de
l'analyseur Eltra-500 144
Figure 62. Estimation des stocks de
carbones dans différents écosystèmes du
Sénégal. (Woomer et al., 2004) 148
Figure
63. Les 4 principales étapes de développement d'une formation
forestière ; adaptée
de Waring et Running (1998) 157
Figure 64. Structure
générale simplifiée du modèle CASS 165
Figure 65. Diagramme logique du
modèle CASS (Roxburgh S., 2004) 168
Figure 66. Exemple de configuration du
modèle CASS 169
Figure 67. Courbes de variation du
carbone dans les différents réservoirs selon des scénarios
de perturbations en cours dans les écosystèmes de savane 171
Figure 68. Intensité des feux de saison
sèche dans le Parc National du Niokolo Koba (Sud
est Sénégal) 176
Figure 69. Exploitation du bois
énergie au Sénégal Oriental 177
Figure 70. Pratiques
agroforestières dans le bassin arachidier (gauche, Faidherbia albida) et
dans le Saloum Oriental (droite, Cordyla pinnata) 178
Figure 71. Pression du bétail dans la zone de
Kaffrine (gauche) et les parcours de
transhumance au Sénégal Oriental (droite) 179
Figure 72. Elimination de l'arbre du
champ pour les cultures de rente. 179
Figure 73. Méthode de mesure de
l'épaisseur des cernes. 194
Figure 74. Photographies de quelques
échantillons de disques 197
Figure 75. Analyse de la
coévolution entre les précipitations et la croissance des cernes
199
Figure 76. Variation des écarts
normalisés de croissance des cernes et de la pluviométrie 203
Figure 77. Courbe cumulée de la croissance des
cernes de différents individu 208
Figure 78. Modèles de
croissance des espèces de savane en comparaison avec celui des
espèces tempérées. 209
Figure 79. Relation âge-biomasse
209
Figure 80. Agrandissement des
échantillons de bois des 5 espèces analysées 210
Figure 81. Etapes de transformation
des données images pour la cartographie de l'occupation du sol 220
Figure 82. Approche empirique d'extraction des
catégories nominales d'occupation et
d'utilisation des terres. Source : Prenzel, (2003) 225
Figure 83. Carte de
végétation de la Forêt classée de Mampaye 231
Figure 84. Carte de
végétation de la Forêt Classée de Bala 232
Figure 85. Carte de
végétation de la Forêt classée de Ouli 233
Figure 86. Carte de
végétation de la Forêt classée de Patako 234
Figure 87. Carte de
végétation de la Forêt classées de Kantora et Welor
(une acquisition)
235
Figure 88. Changement d'occupation
du sol et dynamique du carbone dans les Forêts
Classées de Mampaye, ouli, Balla et Patako 238
Figure 89. Carte d'occupation des sols dans le Saloum
Oriental (Kaffrine : a) localisation ; b)
carte de végétation). 239
Figure 90. NDVI annuel au
Sénégal de 2000 à 2007 243
Figure 91. Représentation
logique de l'approche de la dynamique du NDVI 245
Figure 92. Tendance des valeurs NDVI
sur le Sénégal (2000-2007) 245
Figure 93. Diagramme de l'adaptation
(Huq et Reide, 2002) 258
Figure 94. Liens entre Changements
climatiques, adaptation et atténuation (Burton et al., 2002 ; Huq et
al., 2003) 260
Figure 95. Etapes définies par IPCC
pour l'analyse de l'impact des changements climatiques
263
Figure 96. Modèle conceptuel
d'analyse de vulnérabilité proposé par le PNUE 266
Figure 97. Ecart à la moyenne
de la pluviométrie mensuelle à Kaffrine (1931-2005) 269
Figure 98. Ecart à la moyenne
de la pluviométrie mensuelle à Koungheul (1950-2005) 269
Figure 99. Evolution du bassin
arachidier depuis 1900 273
Figure 100. Evolution du prix du
maïs de 1950 à 2005, US $/t (FAO:
www.fao.org) 275
Figure 101. Evolution du prix du riz
de 1950 à 2005, US $/t (FAO:
www.fao.org) 275
Figure 102. Evolution du prix des
cultures commerciales (coton, arachide, niébé) de 1950 à
2005, US $/t (FAO :
www.fao.org ) 275
Figure 103. L'adaptation est une réponse à
plusieurs stress dont les changements
climatiques (Source, Mbow et al., 2008) 280
Figure 104. Cycle d'un projet MDP 283
Liste des tableaux
Tableau 1. Réservoirs de
carbone dans les écosystèmes de savane 27
Tableau 2. Répartition des
stocks et du flux de carbone des continents qui ont des
écosystèmes tropicaux 28
Tableau 3. Localisation des
Forêts Classées étudiées 48
Tableau 4. Régressions
allométriques proposées par Brown (1997) 76
Tableau 5. Nombre d'individus
échantillons par espèce et par Forêt Classée.
81
Tableau 6. Les modèles
calculés et leur r2 (n = 101 individus) 85
Tableau 7. Comparaison entre la
biomasse sèche (kg) est les estimations des modèles retenus 88
Tableau 8. Comparaison entre la biomasse sèche et
les estimations basées sur les modèles de
la FAO 89
Tableau 9. ANOVA pour les
différents modèles testés 95
Tableau 10. Equations
allométriques pour Combretum glutinosum 98
Tableau 11. Comparaison des
résultats des estimations entre les modèles toutes-espèces
et
les modèles spécifiques 98
Tableau 12. Caractéristiques
d'une fiche de sondage des placettes d'inventaire 111
Tableau 13. Synopsis du sondage
effectué 113
Tableau 14. Paramètres de
calcul de la biomasse 113
Tableau 15. Estimation de biomasse
avec les modèles Quadratique, Cubique et polynomiale
au niveau des Forêts Classées
étudiées 114
Tableau 16. Abondance des
espèces pour les trois parcelles 130
Tableau 17. Taux de carbone pour le
bois et l'écorce de 13 espèces dominantes des Forêts
Classées 145
Tableau 18.
Conversion des stocks de biomasse en carbone pour les différentes
strates des
Forêts Classées 146
Tableau
19. Synthèse sur quelques modèles utilisés pour la
simulation de la croissance des
végétaux. Source Adams et al., (2004),
modifiée 160
Tableau 20. Récapitulatif des
analyses dendrochronologiques 195
Tableau 21. Présentation de
quelques capteurs utilisés pour la cartographie de la
végétation
222
Tableau 22. Les scènes images
utilisées 228
Table des matières
Remerciements II
Résumé V
Abstract VI
Liste des Sigles et abréviations VIII
Liste des figures IX
Liste des tableaux XII
INTRODUCTION 1
Cadre général et justification 2
Objectifs et approche générale 7
CHAPITRE I : Le secteur forestier et les changements
climatiques 12
I.1. Le bilan global actuel du CO2 et les forêts
13
I.2. Les flux de carbone et la plante 18
1.2.1. La photosynthèse : la fixation du dioxyde de
carbone par l'arbre 18
I.2.2. La respiration autotrophe : l'émission de dioxyde
de carbone par l'arbre 20
I.3. Le bilan du carbone des forêts 23
I.4. Le rôle des forêts dans l'atténuation
des changements climatiques 25
I.5. Les réservoirs de carbone dans les forêts
27
I.6. Potentiel de séquestration de carbone des
forêts 29
1.7. La séquestration du carbone et les Mécanismes
pour un Développement Propre 32
I.8. Le MDP foresterie dans les pays en voie de
développement 33
I.9. Les questions actuelles posées par le Protocole de
Kyoto 34
I.10. L'importance du secteur forestier dans les communications
nationales (inventaires
nationaux de GES) 40
I.11. Les activités de reboisement au
Sénégal 41
I.12. La recherche sur le stock de carbone ligneux au
Sénégal 43
Conclusion 46
I.13. Contexte géographique de l'étude
48
I.13.1. Caractéristiques biogéographiques des
Forêts Classées étudiées 48
I.13.2. La variabilité climatique au
Sénégal 53
Conclusion sur le contexte géographique 60
CHAPITRE II : Modèles allométriques 64
II.1. L'importance des modèles allométriques
65
II.2. Développement et utilisation des modèles
allométriques 69
II.2.1. L'état des connaissances sur les modèles
allométriques 69
II.2.2. Généralités sur les mesures de la
masse ligneuse 70
II.2.3. Démarche adoptée pour les mesures de
terrain 72
II.2.4. Les modèles allométriques existants
75
II.2.5. Méthode retenue pour l'élaboration de
modèles allométriques 78
II.2.6. Traitement des données et élaboration du
modèle allométrique 81
II. 2.6.1. Présentation des données
collectées 81
II. 2.6.2. Régressions allométriques
développées à partir des données de terrain 83
II.2.7. Evaluation des modèles retenus 91
II.2.8. Comparaison entre les modèles `toutes
espèces' et les modèles pour une espèce :
exemple de Combretum glutinosum 97
Discussions et conclusion 100
CHAPITRE III : Biomasse et carbone des savanes soudaniennes et
soudano-guinéennes
étudiées 105
III.1. Estimation de la biomasse dans les
écosystèmes de savane 106
III.2. Cadrage conceptuel et méthode de travail 108
III.3. Méthode d'inventaire de la végétation
ligneuse dans les six Forêts Classées 110
III.4. Estimation de la biomasse des Forêts
Classées 114
III.4.1. Contribution à la biomasse par classe de
diamètre 116
III.4.1.1 Répartition des individus par classe de
diamètre 116
III.4.1.2. Répartition de la biomasse par classe de
diamètre 119
III.4.2. Dynamique du carbone par l'analyse de la croissance des
plantes 122
III.4.2.1. Méthode de suivi de la
végétation 123
III.4.2.2. Traitement des données 128
III.4.2.3. Résultats 130
III.4.2.4. Productivité des parcelles permanentes
132
III.4.2.5. Contribution des espèces à la biomasse
totale 138
III.5. Analyse du taux de carbone dans les échantillons
d'espèces ligneuses 143
III.6. Le stock de carbone des Forêts classées
étudiées 146
Conclusion 148
CHAPITRE IV : Modélisation de la dynamique du carbone
152
Introduction 153
IV.1. Les fondamentaux écologiques 156
IV.2. Etat des connaissances sur les modèles de
productivité des écosystèmes forestiers.
158
IV.3. Utilisation des modèles et la gestion durable des
forêts 162
IV.4. Modélisation des flux de carbone avec le
modèle CASS 163
IV.4.1. Description du modèle CASS 164
IV.5. Résultats de la simulation 170
IV.5.1. Analyse des résultats de la simulation 175
IV.5.2. Conditions environnementales et synthèse du
carbone 180
Conclusion 182
V.1. Méthode 191
V.2. Etat des échantillons et procédure de mesure
des cernes 193
V.3. Résultats 196
Conclusion et discussions 211
CHAPITRE VI : Dynamique de l'occupation et séquestration
du carbone 216
VI.1. Changements d'occupation du sol et implications sur la
dynamique du carbone :
l'approche cartographique 217
VI.1.1.Cartographie des changements d'occupation des sols
219
VI.1.2. Considérations pratiques pour la cartographie de
la végétation 221
VI.2. Les données et la méthode
utilisées 228
VI.2.1. Classification des images 229
VI.2.2. Résultats du traitement des images 231
VI.2.3. Suivi du NDVI à partir des données MODIS
241
Conclusion 247
CHAPITRE VII : Adaptation aux changements environnementaux
250
VII.1. La vulnérabilité à la
variabilité et au changement climatique 251
VII.2. Place de l'adaptation dans la CCNUCC 254
VII.3. Complexité et portée pratique de
l'adaptation 258
VII.4. Approches pour l'étude des impacts et de la
vulnérabilité 262
VII.5. Les outils utilisés pour le choix des
scénarios d'adaptation 266
VII.6. Méta-analyse de la vulnérabilité et
l'adaptation dans le Saloum Oriental 267
VII.6.1. Le facteur climatique 268
VII.6.2. Le facteur politique 270
VII.6.3. Le facteur économique 274
VII.6.4. Les réponses adaptives communautaires
276
VII.7. Adaptation versus atténuation, quelles
priorités pour le Sénégal ? 281
Conclusion 288
CONCLUSION GENERALE 291
ANNEXES 298
Annexe 1 : Distribution des espèces par classe de
diamètre pour les trois parcelles de suivi
de la végétation ligneuse. 299
Annexe 2 : Liste des espèces au niveau des parcelles
permanentes 307
Annexe 3 : liste des espèces inventoriées par
Forêt Classée 310
Annexe 4 : Codes VBA pour un model Simple de dynamique de
carbone 317
Annexe 5 : Source de verification des noms scientifiques des
plantes 319
Article I. INTRODUCTION GENERALE
Section 1.01 Cadre général et
justification
La problématique des changements climatiques est
devenue une préoccupation majeure de la communauté internationale
pour ses nombreuses implications sur l'avenir de l'environnement global. Les
risques liés au réchauffement planétaire
n'épargnent aucun continent et affectent tous les secteurs de la vie
économique et sociale des populations. Ces modifications du climat ou
changements climatiques désignent une variation statistiquement
significative de l'état moyen du climat ou de sa variabilité sur
de longues périodes (généralement, pendant des
décennies ou plus). Les changements climatiques peuvent être dus
à des processus internes naturels, à des forçages
externes, ou à des changements persistants d'origine anthropique qui
affectent la composition de l'atmosphère. Les impacts des changements
climatiques affectent les secteurs agricoles, forestiers,
énergétiques, les établissements humains et autres
conditions de vie des populations.
L'un des facteurs les plus dynamiques et les plus puissants de
l'environnement actuel est la modification des températures terrestres
du fait de l'effet de serre. L'effet de serre repose sur le principe selon
lequel, le rayonnement qui nous vient du soleil parvient à
pénétrer dans l'atmosphère pour être
réfléchi ou émis par la surface terrestre libérant
ainsi une chaleur qui maintient les températures moyennes globales
à 15° C au lieu de -18° C en l'absence de ce
réchauffement naturel. Ainsi, l'effet de serre à proprement
parler est un phénomène essentiel pour que la vie se
développe sur terre. Sans ces gaz les températures globales sur
terre seraient si basses que la vie y serait impossible. Ainsi, le
problème du réchauffement planétaire est une question de
seuil due à la perturbation du bilan radiatif global.
Le processus de l'effet de serre repose sur l'équilibre
entre le rayonnement issu du Soleil et la restitution de la chaleur par
l'émission terrestre. En effet, une fois absorbé par le sol, le
rayonnement se transforme en chaleur et est renvoyé sous forme de
rayonnement infrarouge à travers la restitution nocturne. Si
l'atmosphère est transparente pour une bonne partie des rayonnements
visibles, elle présente une forte capacité d'absorption du
rayonnement infrarouge du fait de la présence de certains gaz qui ont la
propriété d'absorber ces grandes longueurs d'onde ; ces gaz sont
appelés gaz à effet de serre (GES). Parmi ces gaz on note H2O,
CO2, CH4, O3, CFC, etc., qui retiennent cette chaleur restituée par la
terre et entraîne ainsi une augmentation sensible et progressive des
températures terrestres émises. Ainsi, l'augmentation de ces gaz,
notamment le CO2 et le CH4 provoquent un réchauffement global qui
affecte actuellement l'environnement.
Le Quatrième Rapport d'Evaluation (QRE) du Groupe
Intergouvernemental pour l'Etude du Climat (GIEC) souligne que 11 sur les 12
dernières années (1995-2006) figurent parmi les années les
plus chaudes depuis 1850 ; date à laquelle ont débuté les
relevés instrumentaux de la température à la surface du
globe (IPCC, 2008). Dans le Troisième Rapport d'Evaluation (TRE, 2003),
on estimait l'augmentation moyenne globale de la température de 0,6
°C (#177;0,2 °C) entre 1901 et 2000, et la valeur établie pour
1906-2005 atteint 0,74 °C (#177;0,3 °C). Les températures ont
augmenté presque partout dans le monde, quoique de manière plus
sensible aux latitudes élevées de l'hémisphère
Nord. Par ailleurs, les terres émergées se sont
réchauffées plus rapidement que les océans.
Cet état des lieux montre que l'action de l'Homme, par
une industrialisation croissante et consommatrice d'énergie, par une
exploitation démesurée des ressources naturelles, a fini de
compromettre l'environnement mondial. Cependant, les modifications du climat
sont non linéaires et discontinues (variabilité climatique) avec
néanmoins une tendance générale qui révèle
dans certaines zones semi-arides d'Afrique une hausse des températures
et une diminution des précipitations. Les manifestations des changements
climatiques sont nombreuses ; les plus manifestes étant
l'élévation du niveau des mers, les événements
climatiques extremes, la destruction de la couche d'ozone et la perturbation
des écosystèmes naturelles.
L'évaluation des impacts de ces changements climatiques
doit être basée sur une approche intégrée du fait de
leurs dimensions multisectoriel, multidisciplinaire, et multi institutionnel,
(Adejuwon et al., 2001). Il faut alors éviter les formes
d'évaluation formelle en se basant sur une série
d'hypothèses plus explicite que l'approche unilatérale
traditionnellement admise. Puisqu'il s'agit de questions complexes, il faut
tenir compte à la fois de la complexité structurelle (nombre
d'éléments en jeu) mais aussi de la complexité
auto-organisationnelle (les rétroactions et réactions du
système). Il est donc important de comprendre la nature de ce
phénomène dans sa dynamique et ses différentes dimensions
en rapport avec les modes de vie actuels fortement dépendants des
ressources naturelles. An Afrique, l'impact des changements climatiques
affectent plusieurs secteurs de productions et affectent directement les
populations dites vulnérables.
Au Sénégal, les modes de vies des populations
locales sont essentiellement basés sur le profit et les services qu'on
peut tirer des ressources naturelles. Les changements climatiques affectent par
conséquent les bases du développement durable, constituant ainsi
un facteur central dans l'appauvrissement des populations locales.
Conscient de la gravité du risque et ses implications
sur la vie des populations, la communauté des chercheurs a, à
partir des années 1980, attiré l'attention sur la
variabilité climatique observée depuis des décennies. Les
premières initiatives prises par les Nations Unies pour prendre en
compte cette menace est la mise en place d'un Groupe International d'Experts
pour l'Etude Climat (GIEC ou IPCC) en 1988. Le mandat de cette structure
était de coordonner les recherches scientifiques pour donner des preuves
des hypothèses et incertitudes liées au changement climatique.
Les résultats obtenus ont imposé l'urgence d'une décision
au niveau international pour réduire les émissions de gaz nocifs.
La conférence de Rio en 1992 a donné naissance entre autres
à la Convention Cadre des Nations Unies sur les Changements Climatiques
(CCNUCC), consolidée lors de la Troisième Conférence des
Partis (COP3) par la signature du Protocole de Kyoto en 1997.
Dans la mise en ~uvre de la CCNUCC, l'Afrique est
comptée parmi les zones où la végétation naturelle
peut contribuer à une émission de carbone par des processus de
dégradation (feux de brousse, déforestation, érosion) ; ou
de séquestration si des activités sont menées en termes de
reforestation et de reconstitution des terres dégradées. Cette
perception a entraîné une attention particulière sur les
capacités de fixation de carbone par les forêts et les savanes
tropicales d'Afrique. Des études récentes sont menées pour
documenter les capacités des sols et de la végétation des
zones de savane comme puits de carbone et leur possible contribution à
l'atténuation du changement climatique (Tiessen et al., 1998;
Fearnside, 2000; Gifford et Howden, 2001; San-Jose et
al., 2001; San Jose, 2001; Woomer et al., 2001; Manlay et
al., 2002; Thenkabail et al., 2002; Ardo et Olsson,
2003; Feral et al., 2003; Archer et al., 2004; Li et al.,
2004; Dezzeo et Chacon, 2005; Mbow, 2005).
Depuis son approbation par la Conférence des Nations
Unies sur l'Environnement et le Développement (CNUED) tenue à Rio
en 1992, la Convention n'a pu avoir un caractère opérationnel
qu'avec le Protocole de Kyoto de 1997 qui a inscrit entre autres
mécanismes de flexibilité celui appelé Mécanisme de
Développement Propre (MDP). Ce mécanisme est le seul qui
interpelle directement les pays en voie de développement. Le MDP devrait
être à la fois un instrument juridique et un cadre d'action pour
promouvoir le développement durable. Le MDP nécessite d'une part
une réduction des émissions de gaz par des alternatives
techniques non consommatrices d'énergie, d'autre part par la promotion
de la fixation de certains gaz par des puits naturels pour résorber le
surplus et ramener les taux de gaz net de l'atmosphère au minimum
acceptable. Les orientations de ce MDP au niveau des écosystèmes
de savanes consistent à promouvoir des puits substantiels de carbone par
la foresterie et la restauration des écosystèmes
dégradés (Aukland et al., 2002).
Ainsi, parmi les nombreuses initiatives prises, celle
consistant à réduire les dégagements de gaz carbonique a
retenu l'attention de plusieurs chercheurs et décideurs. C'est à
la COP7 de Marrakech en 2001 que la question de la séquestration du
carbone a été officiellement reconnue comme une option pertinente
pour atténuer le taux de carbone dans l'atmosphère. A partir des
estimations fournies par la recherche, il a été montré que
cette séquestration pourrait contribuer à une réduction de
25 % de carbone si des mécanismes efficaces étaient mis en ~uvre.
Cependant, compte tenu du fait que les pays industrialisés sont
responsables de 80 % des émissions de gaz carbonique, la voie de la
réduction par les mécanismes de flexibilité du Protocole
de Kyoto, est devenue une pierre angulaire et un enjeu de la
géopolitique environnementale actuelle. Pour les pays en voie de
développement, le principal défi réside dans le
contrôle des échanges de carbone entre le couvert
végétal et l'atmosphère. La végétation en
général est impliquée dans ce mécanisme à
deux titres (Riedacker, 2004) :
· d'une part, la photosynthèse qui conduit à
une fixation de CO2 fait de la végétation un important puits de
carbone, mais la décomposition de la matière organique en
libère.
· d'autre part, les processus de dégradation des
terres comme la déforestation et la désertification modifient le
bilan de carbone.
Il faut toutefois noter que 40 % des émissions totales
de carbone sont absorbées par la végétation terrestre
(Campagna, 1996; Ciesla, 1997; Lippke et al., 2003)
et ~50 % de la végétation forestière est constituée
de carbone (Brown, 1997). Ainsi le `puits
manquant'1 qui constituent un espoir de
réduction significative des gaz à effet de serre se trouverait au
niveau de la végétation terrestre. Cependant, dans les savanes
tropicales les quantités de carbone contenues dans le sol sont
supérieures à celles de la végétation selon
plusieurs études (Elberling et al., 2002; Ringius,
2002; Touré et al., 2002; Bartel et al., 2004;
Dumanski et al., 2004; Evrendilek et al., 2004; Wander et
Nissen., 2004; Zhao et al., 2004; Vagen et al.,
2005; Nsabimana et al., 2008). Mais du fait de l'absence de
données sur la végétation, les estimations sur leur
contenu en carbone demeurent approximatives et restent à être
complétées par des mesures quantitatives in situ.
Les travaux existants portent cependant pour la plupart sur
les stocks de carbone du sol. Ce dessein scientifique est lié au fait
que les sols tropicaux présentent des stocks de carbone
supérieurs à ceux de la biomasse végétale. Il faut
toutefois noter que l'équilibre de ces sols et le maintien durable de
leur capacité de séquestration de carbone passe par une bonne
1 Lorsqu'on fait le bilan des émissions, absorptions de
carbone par les activités humaines (combustion de carbone fossile,
déforestation et reforestation) et des absorptions par les
océans, on n'obtient pas ce qui s'accumule chaque année dans
l'atmosphère. Il manque un "puits", qui serait responsable de
l'absorption du reliquat.
conservation de l'élément protecteur majeur
qu'est la végétation subséquente et qui constitue le
début du cycle du carbone à travers la photosynthèse. Il
est important alors d'orienter la recherche sur les principaux facteurs
structurants de la dynamique écologique de ces savanes pour mieux cerner
les atouts et les contraintes pour une plus grande séquestration du
carbone atmosphérique.
Les tentatives récentes de prédiction des
changements climatiques en relation avec l'évolution du couvert
végétal fournissent des scénarios divergents. L'analyse
proposée par Sitch et al., (2008) montre une tendance à
l'augmentation des températures en Afrique de l'Ouest, mais une grande
divergence dans la prédiction des précipitations. Ces deux
paramètres centraux à l'analyse de la variabilité
climatique ont des impacts sur la dynamique de la végétation. Il
est donc important d'opérer un suivi plus détaillé et
continu des savanes en Afrique de l'Ouest en relation avec la
variabilité climatique.
Les savanes ont été étudiées
beaucoup plus au en Amérique du Sud et en Australie, mais les
caractéristiques de ces savanes sont différentes de celles de
l'Afrique de l'Ouest. Aussi est-il difficile de tirer des conclusions et de
fonder des théories solides sur l'état et la dynamique du carbone
de ces écosystèmes sans compléter le tableau avec
l'étude des savanes Africaines.
Au Sénégal, on a observé une forte
dégradation de la végétation liée à de
nombreux facteurs naturels et anthropiques. La variabilité de la
pluviométrie est un élément important dans ce processus,
mais il est essentiel de considérer les facteurs humains comme
l'exploitation du bois, la dynamique des espaces agraires, les feux de brousse,
etc. Les conséquences directes et indirectes de cette dégradation
de la végétation ligneuse sur le bilan du carbone sont peu
connues. Cependant, il est certain que la réduction de la
dégradation de la végétation ligneuse pourrait contribuer,
à travers une gestion durable des ressources naturelles, d'une part
à réduire la quantité de carbone libérée par
la décomposition de la matière organique et d'autre part à
l'augmentation de la capacité de séquestration du carbone
atmosphérique par la végétation naturelle
régénérée. L'une des recommandations de IPCC
répond de cette nécessité de mener des activités de
reboisement pour améliorer le taux de fixation de carbone dans les zones
où il possible d'en faire (IPCC, 1996; 2000).
Ainsi, la CCNUCC, à travers les MDP du Protocole de
Kyoto a, pendant ces dernières années, suscité un grand
enthousiasme auprès des pays semi arides qui y voient une
possibilité d'améliorer l'environnement à travers une
augmentation des surfaces forestières tout en bénéficiant
des crédits de carbone mis en place à travers des instruments
financiers internationaux.
Pour tirer le meilleur profit de ce marché, il faut
cependant se soumettre à une comptabilité précise des
stocks et des processus qui influencent le bilan du carbone. C'est à ce
niveau que se posent plusieurs problèmes pour l'Afrique. Les
écosystèmes de savanes ne sont pas connus sous l'angle du
carbone. Les études forestières sont plutôt conduites sous
l'angle du potentiel ligneux pour le commerce du bois, de la qualité des
sols et de la biodiversité. Il est important de mentionner qu'un grand
nombre de résultats d'analyse écologiques ou botaniques peuvent
être réutilisés dans l'analyse du carbone. Les
données d'inventaire, les processus de dégradation,
l'érosion de la biodiversité, les prélèvements
annuels, les espèces ciblées par l'exploitation sont en effet
autant de données utiles dans l'étude du carbone.
Ce travail, mené dans la moitié sud du
Sénégal, intègre des données secondaires et des
données biophysiques sur le suivi de la végétation
ligneuse qui ont permis d'estimer les stocks et la dynamique du carbone dans
les savanes soudaniennes et soudano-guinéennes du Sénégal.
Nous partons ainsi de l'exemple de six (6) Forêts Classées
représentatives de ces écosystèmes pour mener les
analyses. Il s'agit des Forêts Classées de Bala, Kantora, Mampaye,
Ouli, Patako et Wélor. Ces sites sont caractérisés par la
présence de savanes soudaniennes et soudano-guinéennes qui ont
été inventoriées dans le cadre du projet ENRECA, DAN
N° 104. Dan. 8L/203-DANIDA et les travaux de (Sambou, 2004)
livrent des informations précieuses sur leur état. Pour les
aspects humains un glissement en milieu de terroir entre Kaffrine et Koungheul
a permis d'appréhender les impacts de la dégradation
forestière sur la vulnérabilité des populations locales et
la dynamique du carbone en milieu agricole.
Section 1.02 Objectifs et approche
générale
L'objectif général de ce travail est d'estimer
les stocks et la dynamique du carbone de la végétation ligneuse
dans les savanes soudaniennes et soudano-guinéennes du
Sénégal. Les objectifs spécifiques sont les suivants :
1- mettre en place une méthode adaptée pour la
quantification de la biomasse des espèces ligneuses des savanes
soudanienne et soudano-guinéennes du Sénégal ;
2- déterminer le stock de carbone de la strate ligneuse
;
3- caractériser la dynamique temporelle du carbone
4- identifier les facteurs majeurs de dégradation de la
biomasse et les réponses apportées par les populations locales en
rapport avec les opportunités du marché du carbone.
Pour atteindre ces objectifs l'approche générale
s'articule autour des points suivants :
· élaborer des modèles
allométriques pour estimer les stocks de carbone des savanes
soudaniennes et soudano-guinéennes du Sénégal en utilisant
les données d'inventaire antérieures ;
· utiliser les meilleurs modèles
allométriques pour donner une estimation plus précise de la
biomasse ligneuse sur pied et des quantités de carbone correspondantes
;
· partir de données de suivi de la
végétation ligneuse et des modèles biogéochimiques
pour caractériser la dynamique de la biomasse et du carbone ;
· tester l'approche dendrochronologique pour retracer les
trajectoires climatiques et biologiques qui caractérisent la croissance
de la végétation dans les savanes étudiées ;
· procéder à la cartographie des changements
d'occupation des sols pour apprécier la dynamique spatiale des
ressources forestières et d'en évaluer l'impact sur le carbone
;
· synthétiser les données secondaires pour
cerner les réponses structurelles et non structurelles apportées
à la dynamique des ressources forestières dans une zone de
terroir (Kaffrine).
Les différents chapitres de ce travail s'articulent
autour de ces points et les détails méthodologiques sont
précisés dans chacune des parties du document. La
cohérence d'ensemble de l'analyse proposée est consignée
le diagramme suivant.
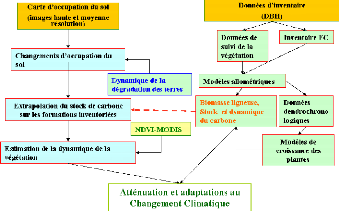
L'approche proposée ici présente deux
entrées. Une première liée aux données de terrain
sur les caractéristiques dendrologiques de la végétation
ligneuse. Les données de terrain permettent d'élaborer les
modèles allométriques, d'estimer la biomasse totale et sa
dynamique. La deuxième entrée est spatiale et permet de
caractériser l'état et la dynamique
de la végétation dans la zone d'étude. Les
réponses apportée à la dynamique de la
végétation sont analysées et comparées aux
opportunités du nouveau marché du carbone.
Cette étude s'inscrit dans la logique
d'amélioration des connaissances sur l'état et la dynamique du
carbone de la végétation ligneuse sur pied des
écosystèmes de savanes à travers l'application d'approches
biologiques et spatiales. Les résultats obtenus devraient donner au
Sénégal, et les pays qui ont des types
d'écosystèmes similaires, des arguments sur les capacités
de la végétation ligneuse à absorber le carbone
atmosphérique et permettre ainsi de développer les fondements
d'une contribution positive visant à améliorer l'état de
l'atmosphère par des activités de renouvellement du couvert
végétal tout en inscrivant de telles activités dans le
sens d'un profit durable pour les populations locales.
Bibliographie
Adejuwon, J., Azar, C., Baethgen, W., Hope, C., Moss, R.,
Leary, N., Richels, R., Ypersele, v. J.-P., 2001. Overview of Impacts,
Adaptation, and Vulnerability to Climate Change. Chapter 2, Climate Change
2001, p. 30.
Archer, S., Boutton, T.W., McMurtry, C. R., 2004. Carbon and
Nitrogen Accumulation in a Savanna Landscape: Field and Modeling Perspectives.
In: M-Shiyomi (Editor), Global Environmental Change in the Ocean and on Land.
TERRAPUB, pp. 359-373.
Ardo, J., Olsson L. 2003. Assessment of soil organic carbon in
semi-arid Sudan using GIS and the CENTURY model. Journal of Arid Environments
(54):633-651.
Aukland, L., Costa, M., P., Bass, S., Huq, S., Landell-Mills,
N., Tipper, R., Carr R., 2002. Poser les fondations d'un développement
`propre' : Préparer le secteur de l'affectation des terres. Un guide
concis du Mécanisme pour un Développement Propre. IIED,
EcoSecurities, ECCM, Londres, 39 p.
Bartel P. 2004. Soil carbon sequestration and its role in
economic development: a donor perspective. Journal of Arid Environments
(59):643-644.
Brown S., 1997. Estimating Biomass and Biomass Change of
Tropical Forests: a Primer. (FAO Forestry Paper -134). FAO - Food and
Agriculture Organization of the United Nations, Roma, 48 p.
Campagna M., 1996. Le cycle du carbone et la forêt: de
la photosynthèse aux produits forestiers. Bibliothèque Nationale
du Québec, Québec, 51 p.
Ciesla W.M., 1997. Le Changement Climatique, les Forêts
et l'Aménagement Forestier: Aspects Généraux. FAO, Rome,
126 p.
Dezzeo, N., Chacon N. 2005. Carbon and nutrients loss in
aboveground biomass a long a fire induced forest-savanna gradient in the Gran
Sabana, Southern Venezuela. Forest Ecology and Management, 209:343-352.
Dumanski J. 2004. Carbon sequestration, soil conservation, and
the Kyoto protocol: summary of implications. Climatic Change (65):255-261.
Elberling, B., Touré, A., Rasmussen K. 2002. Changes in
soil organic matter following groundnut-millet cropping at three locations in
semi-arid Senegal, West Africa. Agriculture Ecosystems & Environment
(96):37-47.
Evrendilek, F., Celik, I., Kilic S. 2004. Changes in soil
organic carbon and other physical soil properties along adjacent Mediterranean
forest, grassland, and cropland ecosystems in Turkey. Journal of Arid
Environments (59):743-752.
Fearnside P.M. 2000. Global warming and tropical land-use
change: greenhouse gas emission from biomass burning decomposition and soils in
forest conversion, shifting cultivation and secondary vegetation. Climatic
Change (46):115-148.
Feral, C.J.W., Epstein, E., H., Otter, L., Aranibar, N., J.,
Shugart, H., H., Macko, A., S., Ramontsho J. 2003. Carbon and nitrogen in the
soil-plant system along rainfall and land-use gradients in southern Africa.
Journal of Arid Environments (54):327-343.
Gifford, R.M., Howden M. 2001. Vegetation thickening in an
ecological perspective: significance to national greenhouse gas inventories.
Environmental Science & Policy, 4:59-72.
IPCC, 1996. Technologies, Policies and Measures for mitigating
Climate Change. Intergovernmental Panel on Climate Change; Technical Paper I,
94 p.
IPCC, 2000. Land use, Land Use Change and Forestry. WMO, UNEP,
377 p.
IPCC (Editor) 2008. Bilan 2007 des changements climatiques.
Contribution des Groupes de travail I, II et III au Quatrième Rapport
d'Evaluation du Groupe d'Experts Intergouvernemental sur l'évolution du
climat. Intergovernmental Panel on Climate Change, 103 p.
Li, J., Lewis, J., Rowland, J., Tappan, G., Tieszen L.L. 2004.
Evaluation of land performance in Senegal using multi-temporal NDVI and
rainfall series. Journal of Arid Environments (59):463-480.
Lippke, Garcia, B., J.P., Manrique C., 2003. The Impact of
Forests and Forest Management on Carbon Storage, Rural Technology Initiative,
College of Forest Resources, University of Washington, Seattle.
Manlay, R.J., Kairé, M., Masse, D., Chotte, J-L.,
Ciornei, G., Floret C. 2002. Carbon, nitrogen and phosphorus allocation in
agro-ecosystems of a West African savanna I. The plant component under
semi-permanent cultivation. Agriculture, Ecosystems and Environment
(88):215-232.
Mbow C., 2005. Dynamic of carbon stock in woody vegetation of
savanna ecosystems in Senegal. The Land cover change perspective. Centre for
Global Change and Earth Observations, Lansing.
Nsabimana, D., Klemedtson, L., Kaplin, B.A., Wallin G. 2008.
Soil carbon and nutrient accumulation under forest plantations in southern
Rwanda. African Journal of Environmental Science and Technology, 2
(6):142-149.
Riedacker A., 2004. Changements Climatiques et Forêts.
SILVA, RIAT, 231 p.
Ringius L. 2002. Soil carbon sequestration and the CDM:
opportunities and challenges for Africa. Climatic Change (54):471-495.
Sambou B., 2004. Evaluation de l'état, de la dynamique
et des tendances évolutives de la flore et de la
végétation ligneuse dans les domaines soudanien et
sub-guinéen au Sénégal. Doctorat D'Etat, Université
Cheikh Anta Diop de Dakar (UCAD), Dakar.
San-Jose, J.J., Montes R.A. 2001. Management effects on carbon
stocks and fluxes accross the Orinoco savannas. Forest Ecology and Management,
150:293-311.
San Jose, J.J., Montes, R.A. 2001. Management effects on
carbon stocks and fluxes accross the Orinoco savannas. Forest Ecology and
Management, 150:293-311.
Sitch, S., Huntingford, C., Gedney, N., Levy, P.E., Lomas, M.,
Piao, S.L., Betts, R., Ciais, P., Cox, P., Friedlingstein, P., Jones, C.D.,
Prentice, I.C., Woodward F.I. 2008. Evaluation of the terrestrial carbon cycle,
future plant geography and climate-carbon cycle feedbacks using five Dynamic
Global Vegetation Models (DGVMs). Global Change Biology, 14:2015-2039.
Thenkabail, P.S., Stucky, N., Griscom, B.W., Ashton, M.S.,
Diels, J., Meer, V.D., B., Enclona, a. E. 2002. Biomass estimations and carbon
Stock calculations in the oil palm plantations of African derived savannas
using IKONOS data. Center for Earth Observation (CEO), Yale University, 58p.
Tiessen, H., Feller, C., Sampaio, E.V.S.B., Garin P. 1998.
Carbon sequestration and turnover in semi-arid savannas and dry forest.
Climatic Change (40):105-117.
Touré A., 2002. Contribution à l'étude de
l'évolution des réservoirs de carbone en zone nord-
soudanienne au Sénégal., Ecole Polytechnique
Fédérale de Lausane (EPFL), 158 p. Vagen, T-G., Lal, R., Singh
B.R. 2005. Soil carbon sequestration in Sub-Saharan Africa: A
review. Land Degradation and Development (16):53-71.
Wander, M., Nissen T. 2004. Value of soil organic carbon in
agricultural lands. Mitigation and Adaptation Strategies for Global Change
(9):417-431.
Woomer, P.L., Tieszen, L.L., Tschakert, P., Parton, W.J.,
Touré A., 2001. Landscape carbon sampling and biogeochimical modelling.
A two-week skills development workshop conducted in Senegal., USGS, CSE.
Zhao, W.Z., Xiao, H.L., Liu, Z.M., Li J. 2004. Soil degradation
and restoration as affected by land use change in the semiarid Bashang area,
Northern China. Catena (In Press).
CHAPITRE I :
Le secteur forestier et
les changements climatiques
Ce chapitre est une synthèse de l'état de l'art
sur la relation entre le secteur forestier et les changements climatiques. Il
présente les informations de base sur le bilan du carbone et la
dynamique de réchauffement planétaire. Il apporte
également des précisions sur l'importance de l'arbre et des
écosystèmes forestiers dans la dynamique de séquestration
du carbone. L'analyse du cadre géographique de l'étude permet de
comprendre le contexte de l'étude et le choix des sites.
I.1. Le bilan global actuel du CO2 et les
forêts
Le dioxyde de carbone (CO2) est un des gaz à effet de
serre qui contribue naturellement au maintien de la température moyenne
de la terre à un niveau qui permet le développement de la vie.
Les températures moyennes globales de 15 °C garantissent les
processus biologiques et le maintien des équilibres
biogéochimiques. Le carbone, qui est l'une des composantes de ce gaz,
est l'objet de nombreux échanges entre les divers constituants
terrestres, atmosphériques et océaniques qui forment le cycle
global du carbone. La concentration atmosphérique de CO2, qui
était de 280 ppm (parties par million par volume) à
l'époque préindustrielle (1750-1800), est passée à
356 ppm en 1993 et actuellement, elle augmente de 1,5 ppm par an
(Schimel., 1995) pour atteindre des concentrations de 370 ppm.
Cette augmentation récente de la concentration
atmosphérique de CO2 est principalement due aux activités
humaines. En effet, pour la période 1980-1989, l'addition annuelle de
carbone au cycle planétaire par la combustion de combustibles fossiles
et la production de ciment fut en moyenne de 5,5 #177; 0,5 GtC/an (gigatonnes
de carbone/an), alors que la contribution de la déforestation
(forêt tropicale) fut de 1,6 #177; 1,0 GtC/an (Schimel, 1995).
Malgré l'importance de ces émissions (7,1 #177; 1,1 GtC/an),
l'augmentation annuelle de carbone atmosphérique n'a été
que de 3,2 #177; 0,2 GtC/an (Campagna, 1996).
La quantité de CO2 qui ne s'accumule pas dans
l'atmosphère serait absorbée par les océans et
éventuellement, déposé au fond des profonds océans,
mais aussi par les écosystèmes terrestres. Le puits manquant, ou
le sort du carbone émis qui n'est pas mesuré dans
l'atmosphère conduit encore quelques controverses (Campagna, 1996;
Waring et Running, 1998; IPCC, 2001). Le bilan global le plus récent
publié par IPCC et qui couvre la période 1980 à 1990, est
présenté par la figure 1.
Le bilan global de CO2 entre 1980 et 1990, montre un total de
7-8 Gt de carbone relâché chaque année dans
l'atmosphère à partir de la combustion des énergies
fossiles et de la déforestation. On estime que les 4-5 Gt de C sont soit
absorbés par les océans et la végétation mais le
reste, 3-4 Gt ne sont pas retrouvés par les scientifiques qui
considèrent qu'il y a un puits manquant non encore identifié
(Adams et Piovesan, 2002). Des recherches récentes soutiennent que ce
puits manquant est terrestre (Schimel, 1995), puisque les forêts ont le
pouvoir de stocker de grandes quantités de carbone.
Article I. Bilan du CO2 entre 1980 et 1990.
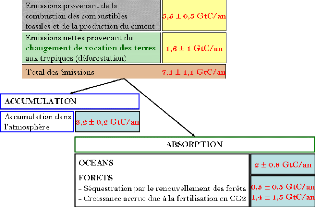
*Les écarts-types ont
été déterminés à partir d'un intervalle de
confiance de 90%
Depuis 1958, la concentration atmosphérique de CO2 fait
l'objet d'un suivi in situ à l'observatoire de Mauna Loa
à Hawaii par l'Université de Californie (USA). La figure 2
présente l'évolution de la concentration atmosphérique du
CO2 de 1981 à 2000 pour quatre stations de suivi in situ y
compris Mauna Lao. L'augmentation graduelle de la concentration
atmosphérique de CO2 suit un cycle qui comporte annuellement un maximum
et un minimum. Ce cycle annuel est attribuable à l'activité
saisonnière des écosystèmes terrestres (l'effet de la
photosynthèse notamment) de l'hémisphère Nord. Le
même processus est noté dans l'hémisphère Sud mais
cette partie du globe présente moins de terres et donc moins de
formations forestières.
Article II. Evolution des concentrations de CO2
atmosphérique au niveau de 4 stations
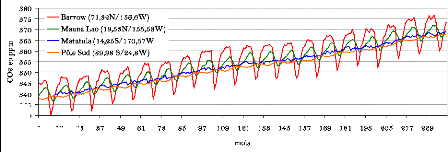
w (7 La
4/157
de mesure de 1981 à 2000.
Source : Réseau SIO-USA, 2004.
Les fortes variations saisonnières des stations de
l'Hémisphère Nord (Barrow, Mauna Lao) sont liées au cycle
phénologique de l'importante couverture de végétation de
l'hémisphère qui en sénescence perd du carbone et sa
capacité à en absorber, alors qu'en phase de croissance, les
écosystèmes forestiers absorbent une large quantité de
CO2. La surface des terres étant largement plus importante dans
l'Hémisphère Nord que dans l'Hémisphère Sud,
l'amplitude des changements dans la concentration de CO2 est plus perceptible
dans la partie Nord de la planète. Dans les deux cas, ces courbes
prouvent que la végétation peut largement contribuer au stockage
du CO2 atmosphérique.
La tendance en hausse du CO2 est aussi clairement
montrée par ces courbes. Une analyse de données de la Station de
Mauna Lao depuis 1958 montre que ces concentrations sont passées de 315
ppm à 375 ppm (figure 3a), soit une augmentation de 60 ppm au total ou
1,4 ppm/an. Il faut toutefois noter la forte variabilité
saisonnière de la concentration de CO2 du fait du cycle
phénologique des plantes (figure 3b).
Article III. Dynamique de la concentration du CO2 à Mauna
Lao (Mesures in situ).
Source: The Carbon Dioxide Research Group, University of
California, 2004.
Il est donc apparent que les forêts influencent le cycle
du carbone. Les processus d'échange de CO2 entre l'atmosphère, la
végétation et le sol passent par la photosynthèse, la
respiration autotrophe (végétale) et la respiration
hétérotrophe dans le sol (décomposition du matériel
organique qui libère du CO2) (Riedacker, 2004).
Il faut noter que la dynamique à la hausse du CO2 dans
l'atmosphère est essentiellement un fait anthropogénique qui a
été confirmé par IPCC (2008), même si quelques
études tendent à soutenir le forçage naturel comme
étant la cause essentielle des changements climatiques (cycle de
l'orbite terrestre, le volcanisme). Certains auteurs comme Ruddiman (2003)
soutiennent que le rôle de l'homme a commencé depuis plusieurs
millénaires avec les
civilisations Mésopotamiennes, Egyptiennes,
Gréco-romaines, etc., dont les activités de production agricole
ont fortement entamé de larges surfaces forestières
libérant ainsi des quantités non négligeable de carbone.
Même s'il faut garder cette perspective, les avancées
technologiques du 18ième siècle ont été le point de
départ de l'aire de fortes modifications d'origine humaine de notre
planète. L'invention de la machine à explosion utilisant les
hydrocarbures, l'avènement de nouveaux modes de communication, de
consommation, de production etc., ont été à la base
d'importantes modifications dans les concentrations de gaz à effet de
serre. On retrouve dans certains documents le terme « Anthropocène
»2 pour traduire l'importance du facteur
humain dans les modifications actuelles du climat. Compte tenu de l'importante
littérature sur la question, nous renvoyons le lecteur aux revues
spécialisées comme Sciences et Nature (
http://www.sciencesetnature.org),
Science et vie (
http://www.science-et-vie.com).
Les synthèses faites récemment par IPCC dans son
Quatrième Rapport d'Evaluation montrent l'importance relative des
activités humaines dans la libération de GES dans
l'atmosphère (figure 4).
Article IV. a) Émissions annuelles de GES anthropiques
dans le monde, 1970-2004 ; b)
Parts respectives des différents GES anthropiques dans
les émissions totales de 2004, en équivalent- CO2 ; c)
Contribution des différents secteurs aux émissions totales de GES
anthropiques en 2004, en équivalent- CO2 (IPCC, 2008).
2 L'Anthropocène est le terme créé et
utilisé par certains scientifiques, dont Paul Crutzen, prix Nobel de
chimie, pour désigner une nouvelle ère géologique, qui
aurait débuté vers 1800, avec la révolution industrielle
et où l'action de l'espèce humaine est devenue une force
géophysique agissant sur la planète. Cette ère nouvelle a
été déclenchée par le développement
industrielle avec une forte consommation d'énergie fossile qui a eu un
impact devenu majeur de l'homme sur le système Terrestre (y compris
climatique). L'anthropocène succèderait ainsi à
l'holocène qui avait débuté il y a dix mille ans.
La figure 4 a), montre que les GES ont augmenté de
près de 45% depuis 1970 (à peine 40 ans). Cette augmentation est
soutenue par la contribution significative des combustibles fossiles qui
à eux seuls ont une part de 57% des GES anthropiques alors que la
foresterie en détient 17% (figure 4 b et c). Les secteurs de
l'énergie et de l'industrie sont les plus grands contributeurs aux GES
(figure 4 c). La part relativement importante de la foresterie est liée
aux importants déboisements des forêts denses, à la
désertification et aux changements d'utilisation des terres.
L'importance du secteur forestier dans le processus de l'atténuation des
GES est fortement documentée dans le Quatrième Rapport
d'Evaluation de IPCC (IPCC, 2008). Il convient avant de décrire les
enjeux du secteur forestier dans les changements climatiques, de faire un bref
rappel sur le rôle de l'arbre dans la séquestration du carbone
atmosphérique.
I.2. Les flux de carbone et la plante
La végétation est l'élément
déterminant dans les échanges de carbones à l'interface
terreatmosphère. Le moteur de ces échanges de carbone est
assuré par la photosynthèse qui permet de fixer le gaz carbonique
et la respiration qui en libère. La base de la séquestration de
carbone par les formations forestières repose sur le bilan entre les
deux processus.
1.2.1. La photosynthèse : la fixation du dioxyde de
carbone par l'arbre
Le rôle de la photosynthèse dans la dynamique de
séquestration de carbone a été largement
étudiée par les biologistes (physiologistes) et les
écologues (Schimel, 1995; Waring et Running, 1998; Saugier, 2003). Le
rappel qu'on en fait ici est une synthèse simplifiée.
La photosynthèse, qui est activée par le
rayonnement solaire, permet à une plante de capter le CO2 de
l'atmosphère afin de synthétiser des glucides. La
photosynthèse utilise la radiation solaire visible (400 nm à 700
nm) qui représente environ 50 % de la radiation solaire globale (Waring
et Running, 1998). De cette fraction, environ 85 % de l'énergie solaire
est absorbée par les feuilles mais cette valeur peut varier
considérablement selon leur structure et l'âge des formations
végétales. Enfin, de la quantité de lumière
absorbée par la feuille, seulement 5 % sert à la
photosynthèse alors que le reste est transformé en chaleur.
L'équation suivante qui décrit la photosynthèse est :
nCO2 + 2nH2O + lumière--(CH2O)n + nO2 + nH2O
L'essentiel de la photosynthèse se fait dans le
feuillage. Les chloroplastes sont les organes contenant de la chlorophylle et
d'autres pigments qui peuvent absorber le rayonnement solaire. Cette
énergie solaire permet à la plante de dissocier
l'hydrogène (H) et l'oxygène (O2). L'hydrogène s'associe
avec le CO2 absorbé par les stomates pour générer des
composés carbonés qui seront plus tard synthétisés
en molécules plus grandes allouées à la biomasse ou
utilisées pour d'autres besoins métaboliques.
Le taux de photosynthèse varie selon les espèces
et les zones phytogéographiques, mais aussi selon le moment de la
journée et la saison. Ces variations relèvent de plusieurs
interactions entre des caractéristiques végétales comme
l'âge, la structure et l'exposition des feuilles, le développement
de la cime, le comportement des stomates, la quantité et
l'activité de Rubisco (ribulose biphosphate
carboxylase-oxygénase) et les facteurs environnementaux comme
l'intensité de la lumière, la température, la
disponibilité de l'eau, la concentration atmosphérique de CO2 et
des polluants atmosphériques et des conditions du sol (Waring et
Running, 1998). C'est grace à la photosynthèse que
l'arbre peut exercer sa fonction de puits de carbone.
Les plantes diffèrent par les types de
photosynthèse, à travers des processus physiologiques de
conversion de différents isotopes stables du CO2 en composantes
hydrocarbonées. Le dioxyde de carbone est fixé par les plantes
à travers les stomates. Il existe deux isotopes stables de CO2 : le
carbone-13 (13C) et le carbone-12 (12C) dont le rapport avec les conditions
actuelles de fortes émissions de CO2 est égal à
près de 1:84 (Waring et Running, 1998). Pendant la photosynthèse
les plantes dites C3 (les ligneux au niveau des savanes) ont tendance à
beaucoup fixer l'isotope 12C plus léger. Le rapport entre 13C/12C au
niveau de l'arbre dépend à la fois de la concentration de ces
isotopes mais aussi de la conductance stomatale des plantes qui fait varier la
fixation des isotopes de carbone selon le type de plante. La forte respiration
autotrophique des plantes C3 fait que le rendement photosynthétique est
plus faible comparé aux plantes C4 qui ont une plus grande
efficacité d'utilisation de l'eau sous l'influence d'une augmentation
des températures. Ces plantes C4 (les graminées, et autres
herbacées) peuvent aussi absorber une partie plus significative de
l'isotope 13C comparé aux plantes C3. La différence entre plantes
C3 et C4 peut être analysée alors en examinant le rapport 13C/12C.
La formule donnée par (Waring et Running, 1998) permet de calculer le
rapport 13C/12C :
ö13C(%o)= {[(13C/12C)
échantillon/13C/12C)
référence]-1}*1000
Ce rapport permet aussi de caractériser indirectement la
conductance stomatale qui induit une grande différence entre les plantes
en termes de rapport 13C/12C.
Les plantes C4 (comme les herbacées tropicales)
utilisent les processus chimiques des acides C4-dicarboxyliques pour faire la
photosynthèse; alors que les plantes C3 (les arbres) font la
photosynthèse utilisant un produit intermédiaire avec 3
molécules de carbone, (Rosenberg et al., 1983). A coté
de ces deux grands groupes on note des espèces dit CAM (Crassulacean
Acid Metabolism) ; ces plantes ont leurs stomates ouverts pendant la nuit
pour fixer du CO2 sous forme d'acide organique (plantes désertiques,
l'ananas). La photosynthèse n'est possible qu'en conditions optimales de
lumière, de température, d'humidité et de CO2. La plante
ne fait pas que la photosynthèse, elles respirent et libèrent par
conséquent du carbone.
I.2.2. La respiration autotrophe : l'émission de
dioxyde de carbone par l'arbre
Les arbres libèrent du CO2 lors de la photorespiration
et de la respiration. La photorespiration se déroule
simultanément à la photosynthèse, alors que la respiration
suit les activités métaboliques diurnes et nocturnes de
l'arbre.
- La photorespiration
La photorespiration ne s'opère que pendant le jour et
provoque la libération de CO2 dans l'atmosphère à cause
des propriétés de l'enzyme Rubisco. Cette enzyme, qui fixe le CO2
atmosphérique lors de la photosynthèse, peut également
fixer l'oxygène (O2). Lorsque l'O2 s'associe à l'enzyme au lieu
du CO2, le carbone préalablement fixé est dirigé vers le
cycle d'oxydation du carbone de la photorespiration qui mène à la
libération de CO2. La photorespiration utilise 30 à 50% du
carbone fixé lors de la photosynthèse (Waring et Running, 1998).
La photosynthèse nette est donc la différence entre la
photosynthèse et la respiration.
- La respiration
La respiration est le processus métabolique qui permet
à l'arbre de libérer du CO2 et d'utiliser l'énergie
emmagasinée dans les glucides synthétisés durant la
photosynthèse. L'équation de la respiration est :
C6H12 O2 + 6O2--)6 CO2 + 6H2O + énergie
Il y a deux types de respiration chez les arbres : la
respiration de construction et la respiration de maintenance. La respiration
est beaucoup plus intense dans les parties de l'arbre en croissance active que
dans les tissus matures et elle est très lente dans les tissus en
dormance (Campagna, 1996).
La respiration de construction utilise l'énergie des
glucides comme le glucose pour créer des structures
végétales ou d'autres composés organiques. La respiration
de construction démarre avec le début de la saison de croissance
et demeure très active tant et aussi longtemps que la plante
croît.
La respiration de maintenance sert à fournir
l'énergie nécessaire aux multiples fonctions des cellules
vivantes composant les diverses structures de l'arbre. La photorespiration, la
respiration de construction et principalement la respiration de maintenance
sont, pour les arbres, les processus métaboliques qui mènent
à une libération de CO2 dans l'atmosphère. La
respiration autotrophe s'oppose à la respiration
hétérotrophe qui est le fait des organismes vivants qui ne font
pas de la photosynthèse (les micro-organismes du sol par exemple).
- La production primaire
Le budget énergétique d'un arbre comprend 3
composantes :
- l'apport énergétique fourni par les glucides
produits par la photosynthèse ;
- les dépenses énergétiques associées
à la respiration de construction et à la respiration de
maintenance;
- les économies énergétiques
stockées dans les réserves (acides organiques, acides
aminés, glucides, amidon, lipides et protéines) et dans des
produits secondaires (tannins, pigments et régulateurs de croissance),
(Waring et Running, 1998).
Quotidiennement, la production et l'utilisation de glucides
sont influencées par de nombreux facteurs biotiques et abiotiques. La
nuit, la respiration et l'absence de photosynthèse font en sorte que
l'arbre émet du CO2. La respiration nocturne utilise environ 50 % de la
production de la photosynthèse nette et une augmentation de la
température d'environ 3 °C peut causer une augmentation
significative de la respiration de maintenance, donc une libération de
carbone. Ainsi, l'augmentation des températures globales peut
entraîner une augmentation de la respiration de maintenance et donc une
importante proportion de carbone pourrait être libérée
à cet effet. La photosynthèse débute dès
l'apparition du soleil, et augmente jusqu'au point où le CO2
absorbé par la photosynthèse équivaut au CO2
libéré par la respiration (point de compensation). Si les autres
conditions écologiques sont remplies (nutriments, eaux,
température), le taux de photosynthèse augmente en relation avec
l'intensité lumineuse jusqu'à un taux maximum (Waring et Running,
1998) qui est atteint lorsqu'un des nombreux facteurs biotiques et abiotiques,
qui influencent la photosynthèse, en limitent le fonctionnement
(facteurs limitants). Une fois le point de compensation dépassé,
l'arbre capte plus de CO2 qu'il en émet, ce qui se traduit par
l'accumulation de glucides qui serviront à la construction des
différentes parties de la plantes y compris la croissance de
l'individu.
Le carbone disponible est d'abord utilisé pour former
des bourgeons, le nouveau feuillage et les nouvelles racines. S'il reste du
carbone après cette étape, il est entreposé pour
constituer des réserves. Ce n'est qu'après que le carbone
résiduel est consacré à la croissance en diamètre
de la tige et à la synthèse de composés chimiques de
protection. La croissance du bois de tronc représente souvent moins de
20% de la production totale annuelle de matière sèche. Lorsque
les arbres produisent des organes reproducteurs, la demande en carbone est
telle qu'elle peut limiter la croissance des autres structures (Schimel, 1995).
La production, l'utilisation et l'entreposage des glucides varient
également selon les diverses périodes
d'activités végétales durant la saison de
croissance. En plus de pourvoir aux besoins usuels des arbres, les glucides
peuvent servir à soutenir des efforts ponctuels ou, à plus long
terme, elles sont utilisées pour répondre aux stress
exercés par les divers facteurs environnementaux (sécheresse,
fortes températures). Ce processus métabolique est un jeu de gain
et de perte d'énergie et le bilan final est connu sous le nom de
production primaire nette qui traduit l'accumulation finale de biomasse. La
figure 5 donne les différentes étapes de ces processus
physiologiques.
Article V. La plante et le flux du Carbone (Waring et Running,
1998)
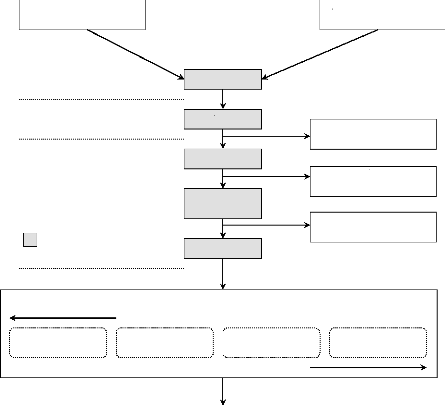
Indice foliaire
Photosynthèse
PPG
Respiration de maintenance,
jour (Rm)
Assimilation
Echanges nets
du couvert
PPN
Respiration autotrophique
Augmente avec la fertilité
Respiration de maintenance,
nuit (Rm)
Respiration de maintenance
des autres tissus
(Rm+Rs)
Système racinaire Réserves et défense
Branches et tiges Feuillage
Augmentation relative liée au manque d'eau et
nutriments
Augmentation relative liée à la RPA et la
température
Allocation du carbone
Le milieu
C-N-Eau-RPA-Vent-Temp.
Caractéristiques génétiques
Détritus
|
|
|
|
|
|
|
|
|
|
|
Respiration hétérotrophique
|
|
|
|
Respiration des
microorganismes (Rh)
|
|
|
|
|
|
|
|
|
|
|
|
|

La figure 5 montre ainsi que le processus commence avec la
photsynthèse qui dépend de l'indice foliaire et des facteurs
environnementaux. Si on déduit de la productivité primaire
globale (PPG), la respiration de maintenance (Rm) pendant le jour on obtient
l'assimilation nette. Sur le reste on déduit la respiration de
maintenance pendant la nuit (Rm) pour obtenir le taux d'échange net du
couvert pour une journée. La productivité primaire nette (PPN)
est calculée en tenant compte des pertes liées à la
respiration autotrophique pendant la synthèse des hydrocarbones (Rs) et
celles liées à la respiration de maintenance (Rm). La PPN est
distribuée sur la base du ratio C/N qui change avec la
disponibilité des ressources en eau et des nutriments. Le renouvellement
des petites racines et des feuilles sont les principales sources
saisonnières de litière, mais toutes les composantes de la
biomasse peuvent joindre le fond de détritus. Le renouvellement annuel
des feuilles et des racines est lié avec les variations
saisonnières de l'indice foliaire et de la teneur en N. La
décomposition de la litière et la libération de CO2 par
les organismes hétérotrophiques dépend de la
qualité du substrat, de la température et de l'humidité du
milieu (conditions stationnelles). La productivité nette de
l'écosystème (PNE) est ce qui reste après avoir
déduit la respiration hétérotrophique (Rs).
Ces processus qui régissent la dynamique du carbone entre
la plante et son biotope expliquent dans une large mesure le bilan du carbone
au niveau des formations forestières.
I.3. Le bilan du carbone des forêts
Des connaissances sur les flux du carbone à travers la
végétation sont essentielles pour comprendre ce qui se passe
actuellement dans les forêts et pour prédire les
développements futurs. Le captage, la séquestration et la
libération de carbone par les forêts sont étroitement
liés au dynamisme et à la vigueur de celles-ci. Les processus les
plus importants impliquant le carbone dans les écosystèmes
terrestres sont : la photosynthèse, la respiration, la translocation,
l'allocation, l'entreposage, le renouvellement des racines fines, la
décomposition, l'influence des herbivores et la chute des feuilles, les
feux de brousse, les coupes ou autres facteurs anthropiques. La
photosynthèse, la respiration et l'allocation des glucides, entre
autres, sont indépendants et varient selon l'espèce, les
conditions stationnelles, les types d'écosystèmes et l'âge
des peuplements. Malgré les difficultés associées à
la mesure des divers processus précités, il est possible
d'estimer les flux de carbone à partir des changements annuels de la
biomasse des arbres (Ruddiman, 2003). Les concepts utilisés pour
exprimer la dynamique de la biomasse sont :
- La production primaire brute (PPB), qui est la
masse totale des composés organiques
produits par la photosynthèse ;
- La production primaire nette (PPN), qui
exprime la PPB moins la respiration
autotrophe (végétale). En d'autres termes, c'est
la masse de matière organique synthétisée par les plantes.
La PPN inclut donc toutes les augmentations de la masse des tiges, des
feuilles, des organes reproducteurs, des racines et la quantité de
tissus végétal consommé par les herbivores ou qui meurt et
devient des détritus.
- La production nette de l'écosystème
(PNE), est la PPB moins les respirations autotrophe et
hétérotrophe. La notion de production nette de
l'écosystème exclut les autres exportations de carbone de
l'écosystème comme celles provoquées par les feux,
l'exploitation forestière, l'érosion ou autres.
La connaissance de l'évolution des divers flux de carbone
dans un écosystème forestier est essentielle si on vise à
optimiser les fonctions de puits et de réservoir de carbone.
Selon certains principes de biologie végétale :
- une jeune forêt en croissance constitue un puits de
carbone ;
- une forêt mature est un réservoir de carbone en
équilibre dont les fonctions de puits et
de source de carbone sont équivalentes ;
- le remplacement d'une forêt mature par une jeune
forêt provoque une libération de CO2
vers l'atmosphère à cause de la
décomposition et du brOlage des résidus de coupe et de
production. Cette libération de CO2 pourrait être égale ou
plus importante que l'augmentation temporaire de la fixation de CO2 par la
jeune forêt, en attendant sa maturité ;
- le sol forestier est un important réservoir de carbone
du fait du cycle de la biomasse à
travers les chutes de feuilles et de brindilles.
Toutefois les différences évidentes dans
l'allocation du carbone pour les diverses espèces d'arbres sont un
élément très important à considérer si l'on
veut que les activités sylvicoles optimisent à la fois la
production forestière et la séquestration du carbone dans
l'écosystème forestier.
En somme, bien que le rôle et l'importance des
forêts dans le bilan global du carbone soient toujours l'objet
d'études, il apparaît que celles-ci séquestrent du carbone
durant leur croissance et qu'elles sont, à maturité, d'excellents
réservoirs de carbone où les échanges avec
l'atmosphère sont en équilibre. Quant au secteur forestier
(biomasse végétale vivante, sols, produits forestiers, sites
d'enfouissement sanitaires et tourbières), il serait un puits ou une
source de carbone selon la fréquence et l'ampleur des perturbations
naturelles ou anthropiques qui affectent les forêts. La forêt et le
secteur forestier pourraient constituer des
éléments efficaces pour séquestrer une
fraction plus ou moins importante du carbone émis par les
activités humaines. Le reboisement de terres agricoles
abandonnées, le remplacement de vieilles forêts par des jeunes
forêts vigoureuses et l'augmentation de la croissance des arbres par la
sylviculture sont autant d'options qui pourraient peut-être augmenter la
fonction de puits de la forêt. En définitive, c'est ce type
d'activités qui est ciblé par la CCNUCC à travers les MDP
Foresterie (Rubrique 14,
http://cdm.unfccc.int/index.html)
et dont la mise en ~uvre est détaillée par un guide de bonnes
pratiques publié par IPCC (2003).
I.4. Le rôle des forêts dans
l'atténuation des changements climatiques
On connaît depuis longtemps l'importance des
forêts pour la production de biens et de services comme les produits
ligneux, les combustibles, la conservation des ressources en sols et en eaux,
les loisirs et la diversité biologique. A présent, on
connaît aussi le rôle important des forêts dans les cycles
biogéochimiques globaux, en particulier le cycle du carbone (MEA, 2005).
Le cycle du carbone est essentiel pour son rôle de régularisation
de la concentration de gaz carbonique (CO2) - qui est un gaz à effet de
serre important - dans l'atmosphère. L'augmentation de la concentration
en CO2 dans l'atmosphère contribue au réchauffement de la
planète, et par conséquent au changement climatique. Les
principaux réservoirs de carbone sont l'atmosphère, les
combustibles fossiles, les océans, la biosphère terrestre et les
sols (voir figure 1, plus haut). Le carbone est échangé entre ces
réservoirs et l'atmosphère sous forme de gaz carbonique (CO2) par
: la combustion des combustibles fossiles, la production primaire nette et la
respiration des plantes, des sols et des autres formes de matière
organique morte, le déboisement, le boisement et le reboisement et la
repousse de la végétation après un bouleversement, et au
travers les échanges avec les océans.
Les forêts sont importantes dans le cycle global du
carbone car elles stockent de grandes quantités de carbone dans la
végétation et le sol. Elles échangent du carbone avec
l'atmosphère par la photosynthèse et la respiration et elles sont
des sources de carbone atmosphérique quant elles sont perturbées
par des causes humaines ou naturelles (feux de brousse, abattage selon de
mauvaises procédures d'exploitation, débroussaillement et brOlis
pour la conversion de la forêt à d'autres usages) et elles
deviennent des puits de carbone atmosphérique (absorption nette de CO2
de l'atmosphère) lorsqu'elles repoussent après ces
bouleversements. Les hommes peuvent, par l'aménagement des forêts,
changer la taille des réservoirs de carbone et modifier le flux de
carbone entre eux et par conséquent modifier leur rôle dans le
cycle du carbone et la façon dont ils influent sur le climat.
Les perturbations qui entraînent une libération
du carbone peuvent être d'origine naturelle ou être
provoquées par l'homme; ce sont notamment des processus qui
entraînent la conversion des forêts ou la dégradation des
terres. Parmi ces processus, on peut citer l'agriculture, le pâturage,
l'exploitation du bois, les feux de brousse, et les infestations par les
ravageurs et les maladies. Par contre, lorsqu'on reconstitue des terres
dégradées, on reconstitue la fonction de puits de carbone des
formations végétales. C'est la tendance générale
observée dans beaucoup de forêts des zones tempérées
et boréales qui ont été exploitées (abattage ou
exploitation sélective) ou déboisées dans le passé
(Brown, 2002).
Selon Brown et Gaston (1995), les forêts peuvent influer
sur le réchauffement climatique de plusieurs façons, notamment
par la production d'autres gaz à effet de serre comme l'oxyde de
carbone, l'ozone et les oxydes nitreux, et par des changements de
l'albédo, c'est-à-dire du pouvoir de réflexion de la terre
quand les forêts sont remplacées par d'autres types de couvertures
terrestres. Cependant, la principale influence des forêts réside
sur la régulation du CO2 dans l'atmosphère et, par
conséquent, sur le cycle global du carbone. Les
forêts3 couvrent environ 3,4 Gha (1 Gha =
109 ha ou 1 milliard d'hectares) (FAO, 1996). La plupart sont
situées :
- dans les zones de faible latitude (0 à 25 °N et S)
ou zone tropicale (52%),
- dans les zones de latitude élevée (50 à 75
°N et S) ou zone boréale (30%),
- dans les zones de latitude moyenne (25 à 50 °N et
S) ou zone tempérée (18%).
Il y a dans le monde quelques 1,7 Gha d'autres terres
boisées qui incluent d'autres formations comme des formations claires,
les broussailles et les jachères forestières résultant des
cultures itinérantes (FAO, 1995) qu'on ne comptabilise pas directement
dans les formations forestières au sens de la définition de la
FAO. Une partie de ces autres zones non forestières, offre des
possibilités considérables d'atténuation des
émissions de CO2 par l'aménagement forestier (par exemple, en
évitant les incendies dans la savane).
Par contre, les forêts tropicales subissent des pertes
importantes, estimées actuellement à environ 15,4 millions
d'hectares par an pour la période 1980-1990 ; mais il s'agit là
de données estimées (Brown, 2002). De 1986 à 1990, FAO
(1996) estime que 5,9 millions d'hectares par an de forêts tropicales ont
été abattues pour des besoins commerciales ou de consommation
locale. Pour la plupart, les zones déboisées sont converties en
terres agricoles, pâturages ou cultures itinérantes. Outre le
déboisement, de vastes étendues de forêts sont
exploitées à titre commercial. L'abattage diminue les stocks de
carbone sur ces
3 Les pays du Sud doivent définir la notion de
forêt dans le cadre des MDP. Trois valeurs sont requises pour
définir une forêt (critères définis par les accords
de Marrakech et par la FAO) : taux de couverture minimum de 10-30 %, surface
minimum de 0,05-1 ha, taille minimum des arbres de 2-5 m.
terres, mais les forêts peuvent se
régénérer et accumuler du carbone
(généralement davantage qu'avant l'abattage) si elles ne sont pas
gravement endommagées durant les opérations d'exploitation. Cette
reconstitution peut être plus effective si les zones forestières
font l'objet de bonnes méthodes d'aménagement et sont
protégées des forces naturelles et humaines qui pourraient nuire
à la végétation et au sol (Brown, 2002). Il faut noter
toutefois que la plupart des formations forestières tropicales subissent
plusieurs formes de dégradation insidieuses ou apparentes, diminuant
ainsi le rythme ou la capacité de séquestration du carbone.
I.5. Les réservoirs de carbone dans les
forêts
On estime que les forêts mondiales contiennent 340 Pg C
(Pg = 1015 g ou 1 gigatonne ou milliard de tonnes) dans la
végétation vivante et morte aérienne et souterraine et 618
Pg C dans le sol. A l'heure actuelle, les bilans du carbone sont incomplets
pour toutes les forêts mondiales. En outre, certaines composantes
forestières comme la teneur en carbone des détritus ligneux, des
débris végétaux et des racines sont peu connues. Cela
explique l'incertitude concernant l'estimation du réservoir de carbone
total. Les principaux réservoirs de carbone des
écosystèmes forestiers sont détaillés sur le
tableau 1.
Tableau 1. Réservoirs de carbone dans les
écosystèmes de savane
|
Réservoirs
|
Description
|
|
Biomasse vivante
|
Biomasse aérienne
|
Totalité de la biomasse aérienne vivante, y compris
les tiges, souches, branches, écorces, et feuillage.
|
|
Biomasse souterraine
|
Totalité de la biomasse de racines vivantes. Les racines
minces de moins de 2 mm de diamètre sont quelquefois exclues car souvent
il n'est pas possible de les distinguer empiriquement des matières
organiques du sol ou de la litière.
|
|
Matière organique morte
|
Bois mort
|
Totalité de la biomasse ligneuse morte qui n'est pas
contenue dans la litière. Inclut le bois au sol, les racines mortes, et
les souches de diamètre égal ou supérieur à 5 cm ou
tout autre diamètre adopté par le pays.
|
|
Litière
|
Totalité de la biomasse morte de petite taille (mois de 5
cm de diamètre), à divers stades de décomposition. Ceci
inclut la litière, les couches fumiques et humiques. Les racines
vivantes minces (inférieures au diamètre minimum adopté
pour la biomasse souterraine) sont incluses dans la litière lorsqu'il
n'est pas possible de les distinguer empiriquement de la litière.
|
|
Sols
|
Matières organiques du sol
|
Inclut le carbone organique des sols à une profondeur
donnée clairement spécifiée. Les racines vivantes minces
(inférieures au diamètre adopté pour la biomasse
souterraine) sont incluses dans les matières organiques du sol lorsqu'il
n'est pas possible de les distinguer empiriquement.
|
|
Carbone stable du sol
(Roxburgh, 2004)
|
Le carbone stable du sol est la fraction de carbone qui n'est pas
tout de suite rejetée dans l'atmosphère après le processus
de décomposition de la matière organique du sol par les
microorganismes.
|
Source : (GIEC, 2003). Complété par l'étude
de (Roxburgh, 2004)
On estime que les écosystèmes tropicaux sont une
source nette de carbone relativement importante (de 1,6#177;0,4 Pg/an en 1990).
Cette source équivaut selon Waring et Running (1998) à
près de 30 % des émissions annuelles de CO2 dues à
l'utilisation des combustibles fossiles. Contrairement aux forêts
tempérées et boréales pour lesquelles les flux de carbone
sont estimés, pour la plupart, sur la base de données provenant
d'inventaires nationaux périodiques (mesures sur le terrain), le flux de
carbone des forêts tropicales est estimé sur la base de
modèles ou de travaux basés sur des données pour le moins
lacunaires. Par conséquent, le flux net de carbone tropical pourrait
être supérieur ou inférieur aux estimations actuelles
(Riedacker, 2004).
Le tableau 2 donne une estimation des réservoirs et des
flux de carbone dans la végétation forestière (masse
vivante et morte aérienne et souterraine, y compris débris
ligneux) et dans les sols (horizon 0 et sol minéral jusqu'à 1 m
de profondeur) (Brown, 2002).
Tableau 2. Répartition des stocks et du
flux de carbone des continents qui ont des écosystèmes
tropicaux
Région Réservoirs de C (Pg) Flux de C
Végétation Sols
(Pg/an-1)
|
ZONE TROPICALE
|
|
Asie
|
41-54
|
43
|
-0,50 à -0,90
|
|
Afrique
|
52
|
63
|
-0,25 à -0,45
|
|
Amérique
|
119
|
110
|
-0,50 à -0 70
|
Le flux net estimatif de carbone des forêts mondiales
est une source de 0,9#177;0,5 Pg/an (Pg = peta grammes : 1015g),
soit environ 16 à 17% de la quantité de CO2 produite et
émise chaque année (figure 1). Ce déséquilibre est
la différence entre les puits et les sources de carbone connus et le
volume «nécessaire» pour équilibrer le bilan du
carbone. Sachant que les données primaires pour les bilans du carbone
dans les zones tempérées sont supposées être d'une
certaine précision, les chercheurs en concluent qu'une grande partie du
déséquilibre du bilan de carbone mondial doit être
cherchée dans les latitudes tropicales, pour lesquelles les estimations
proviennent de modèles qui ne font qu'estimer le bilan avec une
précision limite. Dès lors, il est évident qu'il faut,
pour résoudre cette question, effectuer des inventaires forestiers
nationaux répétés, en installant notamment des parcelles
de suivi de l'accroissement de la végétation dans les latitudes
tropicales.
I.6. Potentiel de séquestration de carbone des
forêts
Le potentiel de séquestration des forêts
dépend en grande partie des efforts de conservation qui
nécessitent des formes d'aménagement approprié permettant
un renouvellement perpétuel des écosystèmes. Les
principaux objectifs de l'aménagement forestier incluent
généralement: la production de bois industriel, la production de
bois de feu, la production de produits forestiers non ligneux, la protection
des ressources naturelles (diversité biologique, eau et sol),
l'aménagement de la faune sauvage, les loisirs, la remise en état
des terres dégradées, etc. La séquestration et la
conservation du carbone résultant de l'aménagement en vue des
objectifs susmentionnés seront un avantage supplémentaire car il
y aura une réduction de la concentration atmosphérique en CO2 et
par conséquent une atténuation du changement climatique.
L'objectif de l'aménagement aux fins de conservation
est avant tout d'éviter les émissions de carbone à travers
la conservation des réservoirs de carbone existant dans les forêts
par des méthodes comme la lutte contre le déboisement, la
protection des forêts dans des réserves, la modification des
systèmes d'exploitation et la lutte contre les autres perturbations
comme les incendies et les ravageurs. La plus grande amélioration des
pratiques de conservation du carbone sous les tropiques, pourrait provenir de
la réduction du déboisement et de la dégradation des
terres. On sait toutefois que ces pratiques sont surtout le résultat de
l'expansion de la demande en terres de culture et de pâturage ainsi que
d'exploitation de bois à des fins de subsistance et commerciales. Ces
différents besoins sont de plus en plus pressants avec les pressions
inhérentes à la croissance démographique, au
développement socio-économique et aux forces politiques. Par
conséquent, les programmes de conservation du carbone par le biais de la
réduction du déboisement doivent s'accompagner de mesures qui
accroissent la productivité et la durabilité agricoles tout en
mettant appliquant des stratégies de réduction de la
pauvreté.
Avec la mise en ~uvre des autres conventions des Nations Unies
(sur la biodiversité et sur la désertification), il y a eu
pendant ces deux dernières décennies un regain d'intéret
pour la conservation des «zones protégées». La
Convention Cadre des Nations Unies sur les Changements Climatiques compte sur
une extension des réserves forestières, une multiplication des
efforts d'aménagement, une limitation de la pression sur les formations
forestières, et une utilisation plus efficace du bois (par exemple, en
limitant les pertes après récolte lors de l'exploitation,
augmentation de l'efficacité de la combustion), pour contribuer ainsi
à fixer une part plus importante de leur carbone total.
Les approches consistant à créer des plantations
sur des terres non boisées permettent de promouvoir la
régénération naturelle ou assistée dans les
forêts non protégées (forêts communautaires), en
utilisant des techniques sylvicoles ou agroforestières. Il faut
toutefois noter que la séquestration du carbone par l'aménagement
forestier n'est qu'une solution temporaire. Il permet de produire de la
biomasse pendant un temps limité au-delà duquel peu de carbone
supplémentaire peut être accumulé. Ce processus peut
prendre plusieurs dizaines d'années, selon l'âge des formations
ciblées, la densité maximale de carbone qui peut être
obtenue, le type de forêt et les espèces choisies.
Pour réduire le carbone atmosphérique, on peut
aussi procéder à la substitution de certaines énergies non
renouvelables par la biomasse forestière. Il s'agit d'accroître
l'utilisation des forêts pour satisfaire les besoins combustibles, soit
en créant de nouvelles forêts et des plantations, soit en
augmentant la croissance et par conséquent la production potentielle de
bois. Dans le cas des forêts plantées sur des terres non
boisées pour la production de sources d'énergie comme le bois de
feu, il est possible à la fois d'augmenter la quantité de carbone
stocké au sol et, si le bois est brûlé comme combustible,
de réduire les émissions de CO2 issu des combustibles
fossiles.
Un total estimé à 700 millions d'hectares de
terre pourrait être globalement disponibles pour les programmes de
conservation et de séquestration du carbone (Brown, 2002). Cette
étendue de terre permettrait de conserver et de piéger de 60
à 87 Pg C d'ici l'an 2050. A l'échelle mondiale, le boisement et
l'agroforesterie représentent la moitié du total (38 Pg C), dont
20 % à peu près s'accumulent dans les sols, la litière des
forêts et la biomasse souterraine (Brown, 2002). Le volume de carbone qui
pourrait être conservé et piégé par ces pratiques
forestières d'ici 2050 par rapport à la situation de
référence est équivalent à environ 11 % à 15
% des émissions totales de combustible fossile pendant la même
période.
Potentiellement, les tropiques peuvent conserver et
piéger la quantité de carbone de loin la plus importante (80 %),
puis viennent la zone tempérée (17 %) et la zone boréale
(3 % seulement). Plus de la moitié du volume dans les tropiques
proviendrait de la régénération naturelle et
assistée, suivie par la protection et le `déboisement ralenti' ou
évité. Le boisement et l'agroforesterie représenteraient
moins de la moitié du volume sous les tropiques. Les taux annuels de
conservation et de séquestration du carbone avec toutes ces pratiques
augmenteraient avec le temps pour atteindre environ 2,2 Pg/an d'ici 2045
(figure 6a), l'accumulation étant plus forte dans la zone tropicale que
boréale. Le potentiel de chaque option d'aménagement est
montré aux figures 6a et 6b.
Article VI. A) Volumes de carbone conservés ou
piégés en fonction des options
d'aménagement (Pg/an) ; B) Taux annuels moyens de
conservation et de séquestration du carbone par région
géographique (Pg/an) (Source : Brown, 1996)
A B
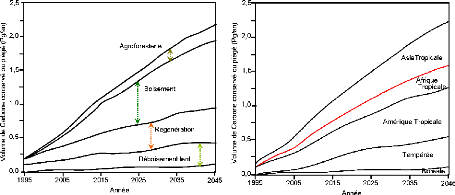
Les auteurs de ces estimations précisent que les
scénarios ci-dessus reposent sur le calcul de ce qui est physiquement
possible pour accroître le stockage et la séquestration du
carbone; ces calculs ne prennent pas en compte la faisabilité sociale et
économique de telles modifications de l'utilisation des terres et de
l'aménagement des forêts. Ils ne tiennent pas compte non plus des
effets des changements de l'accroissement de la concentration de CO2 et
d'autres polluants atmosphériques, n'intègre pas la hausse des
températures ni les modifications futures de l'utilisation des terres
dues à l'accroissement démographique. Chaque option prometteuse
d'aménagement forestier pour une atténuation des émissions
de carbone serait différente selon l'évolution du climat et de
l'atmosphère et les modifications de l'utilisation des terres. Ces
options devraient tenir compte de la demande d'une population accrue pour plus
de terre agricole et de bois ou de produits forestiers non ligneux.
Ces projections montrent que les forêts ont un
réel potentiel dans le processus de séquestration de carbone
à l'échelle globale. Il faut toutefois noter que ce potentiel
varie significativement en fonction des zones géographiques
considérées ou en fonction des activités
d'aménagement envisagées. Il apparaît d'après ces
estimations que l'agroforesterie et le boisement sont des options attractives
pour les activités MDP foresterie. Au Sénégal, la pratique
agroforestière est déjà largement répandue dans le
bassin arachidier notamment avec une association de plus en plus
pratiquée entre les cultures et des espèces comme Faidherbia
albida et Cordyla pinnata. Quant au boisement, les
activités de foresterie communautaire (bois de village), les plantations
en régie, et la mise en ~uvre de la Grande
Muraille Verte, sont autant d'activités qu'il faudrait
désormais comptabiliser parmi les initiatives de séquestration de
carbone à l'échelle nationale.
1.7. La séquestration du carbone et les
Mécanismes pour un Développement Propre
La Convention Cadre des Nations Unies sur les Changements
Climatiques fut approuvée à la Conférence des Nations
Unies sur l'Environnement et le Développement (CNUED) tenue à Rio
en 1992. La Convention Cadre des Nations Unies sur les Changements Climatiques
vise entre autres objectifs, la stabilisation des gaz à effet de serre
à un niveau qui empêcherait des changements risqués du
climat. A ce jour, 186 pays ont ratifié la Convention. Un protocole fut
esquissé à Kyoto en 1997 afin de rendre la Convention
opérationnelle. L'aspect le plus important du Protocole de Kyoto est
l'engagement juridique de 39 pays développés pour une
réduction de leurs émissions de gaz à effet de serre (GES)
de 5,2 % en moyenne par rapport aux niveaux de 1990. Ces réductions
d'émissions doivent être réalisées pendant la
période allant de 2008 à 2012: c'est ce qui a été
défini comme la `première période d'engagement'. Les pays
développés ayant des cibles de réduction des
émissions sont définis comme pays visés à l'Annexe
I, alors que ceux n'ayant pas de cibles de réduction sont
désignés comme non visés à l'Annexe I de ladite
Convention.
Le Protocole de Kyoto permet aux pays développés
de remplir leurs engagements de réduction de différentes
manières au moyen de `mécanismes flexibles'. Ceux-ci comprennent
: le marché de permis d'émissions (marché de droits
d'émissions entre pays développés), la mise en ~uvre
conjointe (transfert de droits d'émissions entre pays
développés, liés à des projets spécifiques
de réduction d'émissions), et le Mécanisme de
Développement Propre (MDP). Le MDP est le seul mécanisme de
flexibilité qui interpelle les pays en voie de développement. Il
permet aux pays développés de réaliser une partie de leurs
obligations de réduction grâce à des projets dans les pays
en voie de développement, réduisant les émissions, ou
`séquestrant' (piégeant) le CO2 présent dans
l'atmosphère.
Il faut toutefois noter que le Protocole de Kyoto en soit ne
donne que les grandes lignes directrices. En fait le Protocole n'était
qu'un accord sur les principes ; c'est bien plus tard que les significations et
les portées techniques de ces engagements ont été
clarifiées. Une rencontre charnière après Kyoto a
été la COP de Marrakech qui donna un cachet pratique à la
mise en ~uvre des décisions de Kyoto. Les COP n'ont fait que
repréciser certains choix et consolider les orientations prises.
Le Protocole de Kyoto a donc fixé les objectifs de
réduction des concentrations de CO2 de l'atmosphère. Les pays
industriels qui ont signé le Protocole, témoignent ainsi de leur
engagement, pour la période 2008-2012, à réduire leurs
émissions de gaz à effet de serre de 5,2 % sur leur niveau
d'émission de 1990. En juin 2005, 150 pays ont ratifié le
Protocole dont 34 pays industrialisés parmi les 38 qui ont eu à
le signer. Ces 34 pays contribuent pour 61,6 % des émissions des pays
industrialisés (Pearson et al., 2005).
I.B. Le MDP foresterie dans les pays en voie de
développement
Le MDP permet aux pays de l'Annexe I de mettre en ~uvre dans
les pays non Annexe I des projets de séquestration de carbone pour
atteindre des objectifs de réduction fixés par la convention. Les
Réductions d'Emissions Certifiées (REC) peuvent être
vendues aux pays Annexe I leur donnant un droit d'émission
équivalent aux crédits alloués. Ce type de projet devrait
permettre aux pays non Annexe I de promouvoir leur développement durable
sans que cela soit un investissement à perte pour les pays Annexe I.
Pour le moment, les pays en voie de développement n'ont
aucune obligation de limitation de leurs émissions de GES. Par contre,
ils ont toujours la possibilité de contribuer, de manière
volontaire, aux réductions globales des émissions, en accueillant
des projets dans le cadre du Mécanisme pour un Développement
Propre. Cette option stratégique est liée à la situation
actuelle du budget total du carbone qui présente une composante
résiduelle d'environ 3 Gt C par année (1 Gt-C (Giga tonnes de
carbone) = 1015 g) que l'on surnomme le `puits manquant'. Les
écosystèmes terrestres qui ne sont pas modifiés par les
changements d'utilisation des terres, et spécialement ceux qui se
trouvent aux latitudes moyennes de l'Hémisphère Nord, ont
été proposés comme candidats les plus probables au puits
manquant.
Le MDP a deux objectifs essentiels :
- Accorder une certaine flexibilité aux pays
développés pour la réalisation de leur
engagement à la réduction d'émissions, en
les autorisant à bénéficier de droits d'émissions
issus de projets réducteurs d'émissions ayant lieu dans les pays
en voie de développement ;
- Assister les pays en voie de développement accueillant
des projets relevant du MDP à
réaliser un développement durable.
Les bénéfices, en termes de gaz à effet de
serre, de chaque projet relevant du MDP seront
mesurés suivant des
méthodes agréées (méthodologies approuvées)
au niveau international
et seront quantifiés en unités
standard, définies en tant que `Réductions d'Emissions
Certifiées' (REC). Celles-ci sont exprimées en
tonnes d'émissions de CO2 séquestrée. Lorsque le Protocole
de Kyoto sera totalement opérationnel, ces `crédits carbone'
seront achetés et vendus au sein d'un nouveau marché de
l'environnement ; ils sont déjà en train de devenir bien
marchand.
I.9. Les questions actuelles posées par le
Protocole de Kyoto
La portée pratique de l'Utilisation des Terres, les
Changements d'Affectation des Terres et la Foresterie a subi une
évolution complexe. Le Protocole de Kyoto fait référence
à deux groupes principaux d'activités : le boisement, le
reboisement ou le déboisement (Article 3.3) d'une part, et les
activités anthropiques supplémentaires des secteurs de
l'agriculture et de l'agroforesterie (Article 3.4) d'autre part. Cependant,
l'article 12 concernant le MDP fait uniquement référence aux
`réductions d'émissions' sans mentionner aucune activité
spécifique. Ceci a conduit à diverses interprétations sur
le rôle possible de l'affectation des terres et de la foresterie au sein
du MDP.
La conférence de Bonn (Juillet 2001, COP 6), puis celle
de Marrakech (Novembre 2001, COP 7) ont amené quelques précisions
sur la réalisation des cibles de réduction des émissions.
Le boisement et le reboisement sont les seules activités relevant de
l'affectation des terres à être éligibles pour la
première période d'engagement (2008 - 2012). Une limite fut aussi
notée sur les projets d'utilisation des terres s'inscrivant dans le
cadre du MDP pour la réalisation des cibles de réduction
d'émissions par les pays visés à l'Annexe 1. Seul 1 % de
l'équivalent des émissions annuelles d'un pays
développé, pour chacune des 5 années de la période
d'engagement, peut être réalisé au moyen des puits.
Les différentes activités ciblées sont les
suivantes :
1. la création de parcelles boisées sur des
terrains communaux ;
2. le reboisement de zones marginales avec des espèces
natives : ces zones étant les berges, les escarpements, celles entre et
en marge de fragments de forêts (grâce à la plantation et
à la régénération naturelle) ;
3. les nouvelles plantations industrielles de grande
envergure ;
4. la création de plantations de biomasse pour la
production d'énergie et la substitution de combustibles fossiles ;
5. la création de petites plantations par les
propriétaires terriens ;
6. l'introduction d'arbres dans les systèmes
d'exploitation agricole existants (activités agroforestières)
;
7. la réhabilitation de zones dégradées au
moyen de plantations ou de régénération naturelle
assistée.
Rappelons que les émissions de CO2 issues des
changements d'occupation du sol représentent près de 20 % des
émissions issues des combustibles fossiles (Pearson, et al.,
2005). Les changements d'occupation du sol peuvent réduire
significativement la concentration du CO2 de l'atmosphère soit à
travers la réduction des émissions nettes, soit par la
séquestration et par le sol. Les actions comme la réduction de la
déforestation, la réduction de l'exploitation forestière,
la limitation des phénomènes d'érosion, contribuent toutes
à la minimisation des émissions de CO2.
Le Protocole de Kyoto reconnaît le rôle
joué par ces changements d'utilisation des terres sur le cycle du
carbone à travers les Mécanismes de Développement Propre
(MDP). Ce mécanisme est définit à l'article 12-2 comme
suit :
Article 12-2 : « L'objet du Mécanisme pour un
Développement "Propre" est d'aider les Parties ne figurant pas à
l'Annexe I à parvenir à un développement durable ainsi
qu'à contribuer à l'objectif ultime de la Convention, et d'aider
les Parties visées à l'Annexe I à remplir leurs
engagements chiffrés de limitation et de réduction de leurs
émissions ~ ». (CCNUCC, 1997)
- Concepts majeurs du MDP : de scénario de
référence, d'additionalité, de fuite (leakage) et de
permanence
* La ligne de base (situation de référence,
scénario de référence)
Les projets MDP sur l'afforestation et la reforestation
doivent permettre l'amélioration du stockage de carbone dans un pays
pouvant permettre une émission équivalente dans un autre pays
sans changer l'équilibre global. Il faut alors que ces projets
garantissent des « avantages réels, mesurables et durables
liés à l'atténuation des changements climatiques »
(CCNUCC., 1997). Les quantités de carbone stocké doivent
être additionnelles par comparaison au scénario de
référence qui traduit l'évolution du carbone en l'absence
du projet.
Les Accords de Marrakech définissent le scénario
de base comme « les émissions de GES quipourraient
subvenir à l'absence des activités du projet ». Ce
scénario de référence n'est accepté
qu'en utilisant une méthode approuvée par le Bureau
Exécutif de la CCNUCC avec ses exigences de transparence et de
cohérence.
* Additionalité
L'additionalité dans le Protocole de Kyoto est
exprimée en ces termes : « Réductions d'émissions
s'ajoutant à celles qui auraient lieu en l'absence de l'activité
certifiée ». (CCNUCC., 1997)
La notion d'additionalité est un élément
central des projets MDP et sa signification a été
précisée en ces termes lors de la Conférence des Parties
(COP9) tenue à Milan (Italie) en 2003 : « les projets de
reforestation et d'afforestation dans le cadre des MDP sont additionnels si la
séquestration nette de gaz à effet de serre par des puits
augmente et dépasse la somme des modifications sur les stocks de carbone
des différents segments (sol, végétation) dans la zone du
projet et qui n'aurait pas eu lieu s'il n'y avait pas de projet MDP
».
Cette définition est beaucoup plus axée sur la
composante additionalité que sur l'éligibilité des projets
MDP. La question de base demeure : quelle est la quantité de carbone
séquestrée comme résultat direct d'une transaction MDP
?
Les émissions de carbone certifiées est le bilan
des réductions ou de la séquestration des GES en retirant le
carbone préexistant de la ligne de base et les fuites. La ligne base est
un concept fondamental du MDP ; elle représente les stocks de carbone
sans intervention du projet ; on peut l'assimiler à la situation
courante (business as usual) ou scénario de
référence. L'additionalité est donc la différence
apportée par le projet en termes de réduction. Un projet est
additionnel si les réductions sont supérieures à celles de
la ligne de base. Il faut par conséquent démontrer que la ligne
de base ne permet pas une forte réduction de CO2 comparée
à l'intervention de projet de boisement reboisement. C'est pour cette
raison qu'il est plus rentable de développer un MDP foresterie dans une
zone dégradée, dont la reconstitution permet d'améliorer
la dynamique du carbone par une augmentation significative de la biomasse.
C'est en ce moment qu'un projet MDP foresterie devient éligible au
niveau du bureau exécutif. Pour prouver cette additionalité, les
méthodes approuvées sont accompagnées d'un outil
d'additionalité permettant de démontrer que les activités
menées apporte un plus par rapport à la ligne de base (IPCC,
2003). Cet outil permet d'évaluer les étapes suivantes :
- l'identification des alternatives aux activités
ciblées par le projet (les alternatives quipourraient
être le scénario de base du projet)
- une analyse de l'investissement (prouver que le projet est
rentable sur le plan
économique)
- l'analyse des barrières (les barrières
économiques, sociales, culturelles, politiques ou
administratives permettent de montrer la nature des risques
à la permanence du projet ; si ces risques sont minimes ou peuvent
être résolus, le projet est éligible) ;
- l'analyse des activités normales des populations (il
ne faut pas surtout développer des activités qui vont en contre
courant des pratiques locales, pour éviter des conflits
d'intérêt qui peuvent nuire l'avenir du projet).
Toutes ces étapes permettent de prouver la
crédibilité du projet. Pour les petits projets, on procède
uniquement à l'analyse des barrières. Ainsi
l'additionalité exprime le bilan entre l'augmentation des
émissions par des sources ou des éliminations par des puits. Les
activités additionnelles si elles permettent une fixation du carbone
atmosphérique qui est supérieure à celles qui se
produiraient en l'absence d'un projet MDP. Cette définition peut
être étendue pour inclure une additionalité dans le domaine
des finances, des investissements et de la technologie.
Dans le cadre de « l'additionalité dans le
domaine des finances », le financement de l'activité dans le
cadre de projets sera en sus du Fonds pour l'Environnement Mondial (FEM),
d'autres engagements financiers des Parties incluses dans l'Annexe I, de l'Aide
officielle au développement, et d'autres systèmes de
coopération.
Dans le cadre de « l'additionalité dans le
domaine des investissements », la valeur des Unités de
réduction des émissions/Unités de réduction
certifiée des émissions améliorera sensiblement la
viabilité financière et/ou commerciale de l'activité.
Dans le cadre de « l'additionalité dans le
domaine de la technologie », la technologie utilisée pour
l'activité sera la meilleure dont dispose la Partie hôte.
* Les fuites (leakage)
Certains projets peuvent donner de bons résultats de
séquestration de carbone dans les limites du projet. Cependant, des
changements peuvent intervenir dans les activités humaines et dans les
comportements sociaux conduisant à une réduction de la
séquestration ou à une augmentation des pertes de carbone en
dehors de la zone du projet. Ce sont ces pertes extérieures au projet
qui sont appelées fuites.
* La permanence des projets et le développement
durable
Le Protocole de Kyoto prend en compte les risques de
réémissions de carbone en relation
avec des activités
humaines (feux de brousse) ou des facteurs naturels comme la
sécheresse.
C'est ce qu'on appelle le risque de permanence qui est propre
aux projets LULUCF (Land Use, Land Use Change and Forestry). Il existe
deux types de projets : les projets temporaires et ceux dits de long terme. Le
projet doit garantir la possibilité de renouveler les stocks en cas de
projet temporaire, cinq ans après la vérification des
crédits confirmant que suffisamment de carbone a été
stocké dans la zone du projet. Cet aspect traduit les garanties que le
projet doit avoir sur les possibilités de compenser les pertes par des
activités additionnelles de séquestration de carbone ou à
travers des crédits capitalisés dans des options non-LULUFC.
L'objectif annoncé pour les pays en voie de
développement dans le cadre des MDP est de promouvoir le
développement durable. Les critères relatifs aux avantages
socio-économiques des projets MDP sont fixés par
l'Autorité Nationale Désignée (AND) qui doit voir
l'articulation avec les critères de durabilité au niveau
national. Cette approbation de l'AND fait partie du processus d'approbation des
projets MDP.
Les transactions de carbone pendant la durée du projet ne
sont pas un processus continu. Elles sont établies en fonction de la
périodicité de l'accréditation. On a deux
possibilités :
- un maximum de 7 ans renouvelable 2 fois,
- un maximum de 10 ans sans renouvellement.
Ces clauses montrent qu'on ne peut vendre que 3 fois de suite
ses stocks de carbone ; entre temps l'espace reboisé ne peut rapporter
aux populations que des services écosystémiques qui n'entravent
pas la permanence du projet. La durée totale des projets est de 20 ans
renouvelables ou 30 ans sans renouvellement selon les cas.
* Définition des activités éligibles
et cadrage conceptuel du Protocole de Kyoto
Les seuls projets éligibles pendant la première
période d'engagement (2008-2012) sont les projets d'afforestation et de
reforestation. Le niveau définition des concepts a pris beaucoup de
temps et a suscité une importante réflexion avant d'obtenir le
consensus qui a aboutit à la définition suivante des termes
boisement et reboisement.
Boisement : c'est la conversion directe par
l'homme de terres non forestières depuis 50 ans, en terres
forestières à travers la plantation, la
régénération, ou autre activités de
reconstitution.
Reboisement : c'est une conversion directe en
terres forestières des zones dégradées (mais qui ont
été occupées par des formations forestières), par
la plantation, la régénération et autres activités.
Pour la première période d'engagement, les zones
dégradées depuis Décembre 1989 sont prises en compte. Dans
la pratique, il n'y a pas de distinction entre afforestation et reforestation
dans le cadre des projets MDP.
Il faut toutefois noter que les activités comme la
gestion, la conservation ou la protection forestière et le carbone du
sol, ne sont pas éligibles pour un projet MDP lors de cette
première période d'engagement (Brown et al., 2005). Les
conditions d'éligibilité des terres sont importantes à
considérer avant de structurer un projet MDP.
* Eligibiité des terres
L'un des critères que tous les projets MDP doivent
respectés est l'absence de forêts dans la zone d'intervention des
projets entre le 31 décembre 1989 et le début du projet. Les
preuves de l'absence de forêts peuvent être fournies à
travers des images spatiales datant de 1989 ou juste avant, ou des
données officielles du gouvernement. A défaut de ces
informations, les témoignages officiels ou issus des communautés
de base devraient suffire.
* Définition de « forêts » dans le
Protocole
La signification du terme forêt est un exercice
déterminant dans le processus d'un projet MDP du fait de ses
implications dans le choix des sites des projets boisement/reboisement.
L'Autorité Nationale Désignée (AND) doit décider
pour le pays, du seuil standard de couverture du sol caractérisant une
formation forestière.
1. taux de couverture minimum entre 10 et 30 % ;
2. taille du site entre 0,05-1 ha ;
3. taille minimum des arbres entre 2-5 m.
* Les implications des critères
d'éligibiité
Il existe de nombreuses implications des seuils choisis dans
l'éligibilité des projets.
1) le taux de couverture
Un taux de couverture bas permet de considérer des
formations boisées ouvertes dans les sites potentiels des projets.
L'agroforesterie peut aussi être considérée dans les choix
de formation à faible taux de couverture, mais leur inclusion exclut
l'utilisation des formations à taux de couverture
élevé.
2) La taille du site
Le choix des sites de petite taille permet d'inclure les
petites formations forestières autour des champs de culture et des
maisons qui servent assez souvent de réserves ligneuses. Par contre, les
sites de grande taille impliquent le choix de grands espaces contigus
entraînant des avantages partagés avec la promotion de la
biodiversité, la qualité de l'eau, la réduction de
l'érosion, etc.
3) La hauteur des arbres
Les faibles hauteurs d'arbres permettent de considérer
les formations basses comme celles qui se développent sur des sols
pauvres ou en altitude. Ils induisent aussi la possibilité de
considérer les plantations commerciales comme le Coffea
arabica, C. robusta ou café, le Theobroma cacao
ou cacao, Acacia senegal ou gomme arabique, etc.
L'inverse réduirait la marge de manuvre. Il faut noter
que la hauteur des arbres s'exprime en termes de potentialités de
croissance, pas la hauteur actuelle des arbres. Ainsi, un seuil bas permet de
considérer les arbustes et non les arbres matures. Idéalement
l'Autorité National Désignée
(AND)4 devrait considérer les
écosystèmes au niveau desquels les projets ont des chances
d'atteindre leurs objectifs.
I.10. L'importance du secteur forestier dans les
communications nationales (inventaires nationaux de GES)
IPCC a retenu le principe de procéder
régulièrement à une compatibilité
détaillée des réservoirs de carbone terrestre des pays
signataires de la Convention. Lors de la COP7 à Marrakech, IPCC a mis en
place un Guide de bonnes pratiques pour la compatibilité du carbone du
secteur dit LULUCF (Utilisation, Occupation des Terres et Foresterie). Ces
directives (IPCC, 2003) donnent des indications harmonisées pour les
inventaires GES dans le secteur LULUCF à travers des méthodes
d'estimation de GES et l'utilisation d'une série de valeurs par
défaut pour convertir certaines données afin d'estimer les
différents réservoirs.
Cinq réservoirs de carbone ont été
identifiés pour les écosystèmes forestiers : la biomasse
sur pied, la biomasse racinaire, la matière organique ou débris
forestier grossier, la litière et la matière organique du sol
(IPCC, 2003). Tous ces réservoirs devraient faire l'objet d'un
inventaire à moins de démontrer que certains ne sont pas des
sources de carbone. Les inventaires nationaux de GES du Sénégal
provenant de la décomposition de la biomasse des
arbres, sont largement basés sur les valeurs par
défaut de IPCC. La collecte de données sur la biomasse des
écosystèmes forestiers pourraient contribuer largement à
améliorer ces inventaires nationaux.
Le bilan du carbone des écosystèmes forestiers
est analysé à l'heure actuelle en utilisant des modèles
écosystémiques permettant d'analyser les flux de CO2. Il faut
cependant noter que plusieurs autres études sont réalisées
en utilisant la télédétection et les données
basées sur des inventaires forestiers (Lehtonen, 2005). Même si
les modèles permettent d'éclairer les processus de base en
simulant les processus fondamentaux, leur applicabilité pose parfois
problème quand ils sont conçus pour des sites spécifiques.
De surcroît, les informations requises pour faire de simulations sont
très complexes et pas appropriées pour les savanes. La
télédétection permet quant à elle de quantifier la
productivité primaire nette avec la possibilité de couvrir de
vastes superficies de façon répétitive.
L'inconvénient de la télédétection est qu'elle ne
peut que faire des mesures indirectes du carbone et ne permet pas de
discriminer la végétation ligneuse de la végétation
herbacée. La télédétection nécessite en
plus, d'importantes données de validation qu'il n'est pas toujours
évident de collecter. Des initiatives technologiques sont entreprises
pour développer des capteurs (RADAR ou LIDAR) pour estimer la biomasse
utilisant des techniques interférométriques permettant des
estimations de volumes à partir des modèles de surface.
Les données basées sur les inventaires
forestiers permettent d'estimer les stocks de carbone pour différentes
formations. Ces méthodes sont coûteuses et lourdes, mais les
données obtenues sont relativement précises puisque directement
mesurer sur les types de végétation en question. Les
premières études sur le potentiel ligneux des formations
forestières avaient pour but d'estimer le volume de bois disponible pour
les besoins d'énergie ou de bois d'ceuvre et de service (Sambou, 2004;
Lehtonen, 2005). En plus du volume commercial ou utile pour les besoins directs
des populations, le reste de la biomasse des arbres comptent pour beaucoup dans
les stocks de carbone.
I.11. Les activités de reboisement au
Sénégal
L'historique des approches de conservation forestière
montre plusieurs séquences qui diffèrent par les objectifs de la
conservation forestière, les approches et la finalité. Il faut
noter a priori que les stratégies de reconstitution des milieux
dégradés ou de conservation ont été
influencées par les contextes démographiques, climatiques, par
les engagements internationaux, et surtout par l'apport de solutions au grand
problème de la dégradation des terres qui constitue un
réel frein à la production agricole et pastorale.
Déjà, durant la période coloniale, des
efforts importants de conservation ont été faits par la mise en
réserve de la plupart des Forêts Classées que le pays
compte actuellement. Le législateur colonial avait aussi
développé plusieurs plantations en régie avec des
boisements de Khaya senegalensis, Bombax costatum et
Tectona grandis. Les Forêts Classées de la période
coloniale ont été conservées et les plantations en
régie poursuivies pendant l'indépendance. Avec la grande
sécheresse de 1970, l'Etat du Sénégal a initié de
grands projets de reboisement en régie avec notamment la mise en place
du PRS (Projet de Reboisement du Sénégal). Jusque là les
populations locales n'étaient pas impliquées dans les efforts
structurels de reboisement. Ce n'est qu'avec l'avènement des projets de
nouvelle génération, lesquels partent du constat des
échecs antérieurs, qu'a commencé l'application de la
participation locale. L'enthousiasme et l'intérêt de l'approche
participative a engendré une littérature abondante autour de
nombreux projets des années 1980-1990 comme le PGCRN (Projet des Gestion
Communautaire des Ressources Naturelles), le PROBOVIL (Projet de Boisement
Villageois), le PAEFK (Projet d'Appui à l'Entreprenariat Forestier), le
PRECOBA (Projet de Reboisement Communautaire dans le Bassin Arachidier), le
PREVINOBA (Projet de reboisement villageois dans le Nord-ouest du bassin
arachidier), etc., qui devraient tous à travers une implication des
populations contribuer à l'atteinte des objectifs du PAFS (Plan d'Action
Forestier du Sénégal).
Les approches participatives se sont consolidées avec
l'avènement de la décentralisation (1992) qui a
transféré entre autres compétences, la gestion des
ressources naturelles non protégées aux collectivités
locales ; une nouvelle forme de participation locale apparaît à
travers la foresterie communautaire depuis 1995. Des Projets comme le PAGERNA
(Projet d'Autopromotion et de Gestion des Ressources Naturelles au Sine
Saloum), le PGIES (Projet de Gestion Intégrée des
Ecosystèmes au Sénégal), le PROGEDE (Programme de Gestion
Durable des Energies traditionnelles et de substitution) ; et des initiatives
comme celles de la coopération allemande (GTZ), avec
l'aménagement de la Forêt Classée de Dankou, sont autant
d'illustrations d'une foresterie d'un nouveau genre. L'idée est de faire
en sorte que l'aménagement et la gestion des ressources
forestières soient le fait des populations qui doivent en tirer des
bénéfices sous le contrôle du service forestier.
Jusque là, aucun des projets susmentionnés
n'évoque la question de la séquestration du carbone parmi ses
objectifs ou motivations tacites. Ce n'est qu'à la dernière phase
du PROGEDE, et la mise en place depuis 2005-2006 du PREFER (Projet de
Réhabilitation des Ecosystèmes Forestiers et Ruraux) et du
PROGERT (Projet de Gestion et de Restauration des Terres
Dégradées du Bassin Arachidier) que la question de la
séquestration du carbone
par les activités forestières a été
clairement stipulée dans les documents de travail et prise en compte
dans la mise en ~uvre des activités de projets forestiers.
Ces mutations dans l'approche sont accompagnées de
profondes réformes juridiques et institutionnelles pour faciliter la
mise en ~uvre de la foresterie rurale communautaire. Ainsi, le Code Forestier a
été révisé en 1993, la décentralisation est
devenue effective en 1996, la politique forestière (PAF) a
été revue pour mieux tenir compte des nouvelles orientations
politiques et stratégiques en matière de foresterie.
Malgré ces efforts, le déboisement est
cité parmi les facteurs structurants majeurs de la dégradation de
l'environnement au Sénégal. Les sols s'appauvrissent, les
superficies forestières sont continuellement réduites,
l'érosion et la réduction de la biodiversité sont plus que
jamais actuelles (MEPN, 1997), et ce en dépit de quelques études
qui démontrent par endroit une réhabilitation naturelle
(cicatrisation) de certains écosystèmes dans le Sahel (Bassett et
Zueli., 2000; Rasmussen et al., 2001). C'est dans ces
conditions de désertification et de déforestation que le
Sénégal s'est engagé en tant que parti signataire de la
CCNUCC à développer des projets MDP foresterie pour non seulement
séquestrer du carbone, mais aussi atténuer plusieurs effets
néfastes des changements de l'environnement et des
écosystèmes. L'état actuel des connaissances sur les
stocks de carbone des formations végétales montrent à la
fois le potentiel et les dynamiques en cours, mais révèlent
surtout les lacunes à combler pour mieux documenter les
réservoirs de carbone à l'échelle nationale.
I.12. La recherche sur le stock de carbone ligneux au
Sénégal
La question du carbone n'a été que
récemment un centre d'intérêt pour les pays
sahéliens, même si les efforts de reboisement et de reconstitution
des terres dégradées ont été un aspect central des
politiques nationales de conservation des sols et des ressources
forestières. Dans un premier temps, la pertinence du reboisement a
été mise en relation avec la production agricole et les
phénomènes d'érosion. En sus de ces aspects, le carbone
des écosystèmes est devenu une source d'enjeu économique,
écologique et social en relation avec les développements
récents relatifs aux changements climatiques (Woomer et al.,
2001; Elberling et al., 2002; Woomer, 2003; Liu et al., 2004;
Parton et al., 2004; Tschakert et al., 2004; Tschakert et
Tappan, 2004; Tschakert, 2004; Wood et al., 2004; Woomer et
al., 2004a; Woomer et al., 2004b; Mbow, 2005; Mbow et
al., 2008). La littérature montre que très peu
d'études ont été consacrées au stock de carbone
ligneux dans les écosystèmes de savane du Sénégal.
La plupart des études portent soit sur le carbone du sol (Elberling,
et al., 2002;
Tieszen et al., 2004; Tschakert et al.,
2004; Takimoto et al., 2008) soit sur une estimation de la biomasse
ligneuse basée sur des modèles écologiques (Manlay et
al., 2002; Manlay et al., 2004; Gray et al., 2005; Mbow,
2005; Lufafa et al., 2008). L'estimation du carbone ligneux requiert
une base d'information consistante à partir de données
d'inventaire qui sont peu nombreuses (Sambou, 2004). Le Sénégal
n'a pas encore réalisé l'inventaire national de ses formations
forestières même si le PROGEDE a fait un effort
considérable dans cette direction avec la mise en place d'un SIEF
(Système d'Information Ecologique et Forestier). Les inventaires
nationaux ont permis dans de nombreux pays de faire un inventaire du carbone de
la végétation ligneuse sans avoir recours à des processus
d'approximation issue des modèles. Au Sénégal, les travaux
de l'ISE dans les Forêts Classées et les Parcs Nationaux,
constituent une base utile pour estimer les stocks de carbone et leur
dynamique, (Sambou, 2004).
Pour combler le manque d'information, la plupart des analyses
des stocks de carbone a été basée sur des modèles
comme CENTURY. En utilisant le modèle CENTURY, Liu et al.
(2004) ont pu estimer une perte de 37 % du stock de carbone entre 1900 et 2000
au niveau du sol et de la végétation au Centre Sud du
Sénégal. Les résultats du modèle indiquent une
tendance à la diminution qui ne peut être corrigée qu'avec
des mesures d'aménagement et de gestion durable. Utilisant le même
modèle, Parton et al. (2004); Woomer et al. (2004a) et
Woomer et al. (2004b), ont essayé de suivre l'évolution
du carbone dans le temps en fonction des types d'utilisation des terres, et en
s'appuyant sur des données de terrain. Les résultats ont
montré une perte de 0,477 Gt de carbone de 1850 à 2000. Woomer
(2004) a essayé de relativiser ces pertes en fonction de 11 types
d'utilisation des terres et en fonction de différentes périodes.
Par contre selon ces mêmes auteurs, une amélioration des pratiques
de gestion pourrait permettre une augmentation de 0,116 Gt de C de 2000
à 2100. Dans la même lancée, Lufafa et al. (2008)
ont couplé CENTURY avec un Système d'Information
Géographique (SIG) pour générer des données de
distribution spatiale de carbone dans le vieux bassin arachidier. Ces derniers
travaillant dans les zones de jachères à Guiera
senegalensis dans la région de Thiès ont montré des
stocks de carbone variant entre 0,01 à 0,45 Mg C/ha. Ils ont conclu que
la séquestration de carbone de ces jachères dépend dans le
long terme d'un aménagement sans utilisation des feux sur les
résidus agricoles et sur la biomasse issue de la préparation des
champs ; elle dépend aussi des types de sols et des scénarios
d'aménagement. Les travaux de Lufafa et al. (2008) montrent que
si on arrive à maintenir un retour des défrichements tous les 50
ans, on pourrait augmenter les stocks de carbone de 200 à 350 % au
niveau de la végétation (sans fertilisation) et de 270 à
483 % au niveau des sols (avec un peu de fertilisation). Ainsi ces
résultats montrent qu'il existe un réel
potentiel de séquestration de carbone des terres
dégradées ; il suffit pour cela d'un aménagement
approprié.
Ces estimations sont certes utiles mais doivent être
mises en relation avec la réalité de terrain et la mise en ~uvre
de stratégies appropriées pour une amélioration du
stockage de carbone. C'est ce que Woomer et al. (2004b) ont
constaté en étudiant les stocks de carbone de la
végétation et du sol sur 5 sites le long d'un transect Nord-Sud
dans la zone sahélienne du Sénégal. Les données
collectées leur ont permis d'estimer le biovolume à partir du
taux de couverture et la hauteur moyenne du couvert. Des mesures de terrain ont
aussi permis à Touré, (2002) de procéder à des
analyses de sols, pour estimer les quantités de carbone du sol en
mettant l'accent sur les différences pédologiques et l'existence
ou non de pratiques agricoles. Cette étude a permis d'estimer les stocks
de carbone des sols dans l'ordre de 20 tG/ha pour les sols sableux et 40 tC/ha
pour les sols argileux des zones cultivées du bassin arachidier du
Saloum Oriental. Les stocks sont de 31tC/ha (sol sableux) et 64 tC/ha (sol
argileux) pour les sols non cultivés de la même zone. D'autres
études plus ouvertes sur le Sahel ont été menées
par Takimoto et al. (2008) dans des systèmes agroforestiers
(300-700 mm/an) ; ces études montrent que les stocks de carbone de la
biomasse au Sahel varient entre 0,7 et 54 Mg C/ha, alors que les stocks de
carbone du sol varient entre 28,7 et 87,3 Mg C/ha. Ces données
permettent de dire que les sols ont plus de carbone que la biomasse, mais il
faut remarquer que la dynamique de séquestration du carbone est une
chaine qui commence par la photosynthèse. Le carbone du sol
dépend largement de ce que la végétation est capable de
fixer. Le carbone du sol n'est pas éligible aux MDP, mais il est
important de bien le conserver pour améliorer les activités de
production agricole car il s'agit d'une composante essentielle de la
fertilité des terres de culture (Tieszen et al., 2004).
Pour compléter ces analyses purement physiques,
Tschakert et al. (2004); Tschakert et al., (2004) et Mbow
(2008) ont noté que malgré la dégradation des ressources
dans le Bassin Arachidier du Sénégal, les systèmes
agricoles locaux adoptent des stratégies d'adaptation avec une
diversité de méthodes de fertilisation des sols. Ces auteurs
proposent ainsi une approche d'amélioration de la séquestration
du carbone basée sur les techniques locales.
Ces études ont le mérite de dégrossir la
question du stock et de la dynamique du carbone au Sénégal.
Cependant, elles n'ont pu bénéficier de données de terrain
abondantes pour calibrer les estimations issues des modèles qu'ils ont
utilisé. Par ailleurs, la quantité de données
collectées sur le terrain reste également insuffisante pour une
meilleure estimation du stock de carbone dans les différents
écosystèmes. En effet les sites étudiés sont
presque tous
localisés dans le Nord du pays alors que l'essentiel du
potentiel végétal ligneux du Sénégal est
situé dans la moitié Sud du pays (voir carte de
végétation, figure 7).
Conclusion
Cette synthèse générale montre que le
réchauffement climatique est un fait réel dont la manifestation
directe est une augmentation des températures terrestres du fait de
l'augmentation graduelle des gaz à effet de serre (GES) de l'ordre de
1,4 ppm/an. Les impacts de ces changements climatiques sont nombreux et
affectent différents secteurs de la vie économique par la
modification des processus biophysiques qui jusque là ont connu un
certain équilibre relatif. Pour agir efficacement, la Convention Cadre
des Nations Unies sur les Changements Climatiques à travers de
nombreuses conférences des parties a pu amener la communauté
internationale à s'engager à mener des efforts de
réduction des GES émis (pays Annexe I) ou à participer aux
efforts de fixation de carbone atmosphérique à travers des
projets MDP financés par les pays pollueurs du Nord au profit des pays
en voie de développement du Sud (Pays non Annexe I). Les MDP peuvent
être de différentes formes, et on en trouve dans l'industrie, les
mines, le secteur énergétique, au niveau des déchets, etc.
Le secteur sur lequel l'Afrique a un grand intérêt est celui des
MDP foresterie qui permet de contribuer doublement aux objectifs de la
Convention, notamment sur l'atténuation et l'adaptation aux changements
climatiques.
Le secteur forestier est compté parmi les secteurs les
plus dynamiques et les plus prometteurs de l'atténuation des changements
climatiques. Il s'agit aussi d'un secteur qui mérite une attention
particulière du fait de sa forte vulnérabilité aux
modifications du climat. Les impacts des changements climatiques ont
déjà affecté plusieurs écosystèmes à
travers des modifications significatives des températures et des
précipitations. L'homme, par son lot de besoins en produits ligneux,
vient entamer les formations forestières et crée un
déséquilibre parfois irréversible dans les fonctions
naturelles du couvert végétal. Ainsi, en relation avec les
changements climatiques, les forêts sont considérées
à la fois comme facteur contribuant au problème
(dégagement de GES, feux de brousse), mais aussi et en même temps
comme un secteur à investir pour résoudre en partie le
problème des GES par la séquestration accrue des GES.
Il faut noter cependant qu'il existe une grande
variété dans les formations végétales, avec des
capacités de séquestration qui dépendent du contexte, des
impacts climatiques ou humains subis, des types d'espèces
rencontrées et des formes d'aménagement mises en ~uvre. Cette
synthèse de l'état de la question nous permet de voir clairement
que le futur de
la séquestration du carbone se situe sous les
tropiques, avec une marge de manuvre en termes d'espace disponible et des
possibilités énormes d'amélioration des modes
d'aménagement et de gestion. Aussi, pour des raisons liées au
principe du Protocole de Kyoto et à ses objectifs, les pays en voie de
développement sont-ils les réceptacles des projets MDP foresterie
pour amener les pays industrialisés à investir dans ces
projets.
Des avancées scientifiques sont notées dans le
sens d'une meilleure connaissance des stocks de carbone des savanes, mais il
reste beaucoup d'incertitudes sur les dynamiques végétales qu'on
ne peut tenter de combler qu'avec une collecte d'informations directes sur le
terrain. Ce mémoire conduit dans la moitié Sud du
Sénégal est une contribution dans ce sens. La description du
milieu biophysique de la zone d'étude permettra de mieux comprendre le
contexte géographique de l'étude.
I.13. Contexte géographique de
l'étude
I.13.1. Caractéristiques biogéographiques des
Forêts Classées étudiées
Avec une longue saison sèche qui dure 9 mois au Nord et
moins de 7 mois au Sud, le Sénégal est un pays sahélien
localisé dans une zone semi-aride caractérisée par des
précipitations faibles au Nord (300 mm) plus abondantes au Sud (>
1000 mm). Le climat du Sénégal est lié à cette
variabilité saisonnière de la pluie. De longues périodes
de sécheresse ont été enregistrées depuis 1968.
Mais depuis une décennie on constate un retour des fortes
précipitations malgré quelques années déficitaires.
L'analyse de l'évolution du climat au Sénégal est
largement abordée dans la thèse de Sagna (2006).
Le pays compte plus de 60 % de populations rurales qui
dépendent essentiellement sur les ressources naturelles pour leur
survie. Les profondes dégradations des ressources
végétales font des réserves forestières actuelles
des îlots de végétations souvent entourés par des
terres dénudées après de longues années
d'exploitation agricole. C'est au niveau de ces lambeaux de formations
végétales que cette étude a été
menée.
L'analyse des stocks de carbone et de leur dynamique a
été effectuée sur la base d'un échantillonnage
spatial et temporel. L'échantillonnage spatial a conduit à
choisir 6 Forêts Classées dans les zones soudanienne et
guinéenne du Sénégal. Les Forêts Classées
étudiées sont présentées au tableau 3.
Tableau 3. Localisation des Forêts
Classées étudiées
|
Forêt Classée
|
UTM, WGS 84/Zone 28/P
(centroïdes)
|
|
X
|
y
|
|
Wélor (zone soudano-sahélienne)
|
364501
|
1558011
|
|
Patako (zone soudanienne)
|
363759
|
1512764
|
|
Ouli (zone soudanienne)
|
610760
|
1535758
|
|
Bala (zone soudano-sahélienne)
|
714604
|
1556527
|
|
Kantora (zone soudanienne)
|
635979
|
1469743
|
|
Mampaye (zone soudano-guinéenne)
|
651556
|
1407436
|
La figure 7 montre la localisation des Forêts
classées de l'étude.
Article VII. Carte des zones climatiques et des réserves
forestières du Sénégal
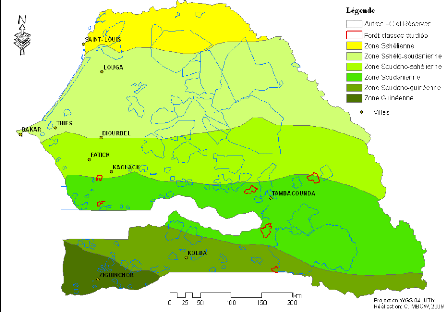
L'échantillonnage temporel a consisté à
travailler sur des données de parcelles permanentes suivies depuis 1993
dans le cadre d'un projet de coopération scientifique entre l'ISE et
l'Université de Aarhus du Danemark (Projet ENRECA, DAN N° 104. Dan.
8L/203- DANIDA).
Les Forêts Classées étudiées ont
été choisies sur la base de leur localisation dans les zones de
savane soudanienne et soudano-guinéenne du Sénégal. Aussi,
leurs localisations géographiques induisent des différences
biophysiques importantes à considérer dans le cadre des
estimations de stocks de carbone. D'abord leur disposition fait voir deux
transects : Nord-Sud et Est-Ouest. Les deux comportent des gradients du point
de vue climatique mais aussi des gradients humains sur le plan de la
démographie et des pratiques forestières. La partie Sud du pays
est la zone la plus boisée du Sénégal avec une bonne
pluviométrie et la partie Ouest est plus peuplée avec des
densités de populations plus fortes que la moyenne nationale. Ces
variations dans l'espace, les différences des conditions climatiques et
des facteurs humains conduisent à des dissemblances significatives dans
le développement des formations forestières.
Sur le plan des densités de population, on observe sur
la figure 8 que les Forêts Classées de Patako et Wélor
situées dans la partie Ouest ne sont pas très
éloignées des zones de fortes concentrations humaines. Les
Forêts Classées de la zone Est, sont situées dans des zones
de faibles densités de population (5-25 hbts/km2).
Article VIII. Carte des densités de population du
Sénégal (base DTGC, 2004)
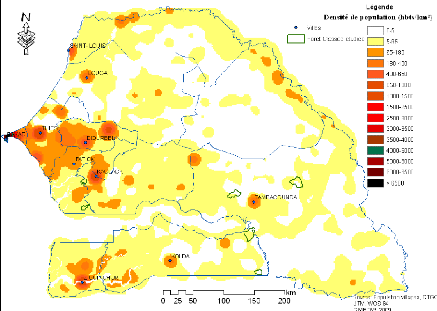
Les formations végétales de ces Forêts
Classées sont pour l'essentiel des savanes. Elles sont à
dominance arbustive pour les Forêts Classées de la zone de
transition entre la zone sahélienne et soudanienne (Bala, Ouli et Welor)
et le taux de couverture devient de plus en plus élevé avec les
forêts classées situées dans la zone soudanienne et
soudano-guinéenne (Patako, Kantora, Mampaye). Pour ces dernières,
des savanes boisées, des forêts claires et d'importantes
forêts galeries peuvent y être trouvées (figure 9).
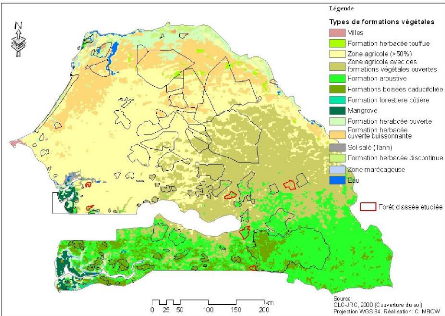
Sur le plan pédologique, les Forêts
Classées (FC) étudiées se sont développées
sur différents substrats. Les Forêts Classées de Patako et
Wélor sont localisées sur des sols ferrugineux tropicaux (sols
Dior). Dans la FC de Ouli on a une dominance de sols peu évolués
d'apports, alors qu'à Bala on a, à l'Est une dominance de
lithosols (latérite) ; à l'Ouest des sols peu
évolués d'apport. A Mampaye on observe quelques affleurements de
sols d'éboulis ou régosols issus du démantèlement
de la cuirasse latéritique, mais une bonne partie de la forêt est
concernée par les sols ferrugineux tropicaux lessivés (figure
10).
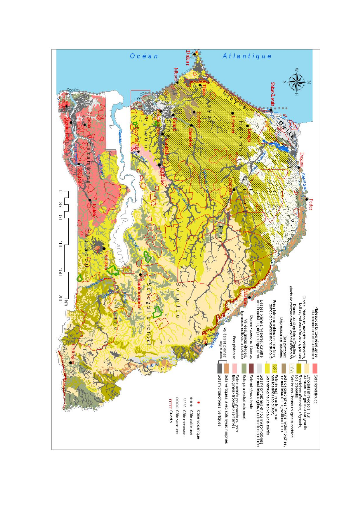
Le facteur climatique est un élément essentiel
dans la dynamique et l'état des formations ligneuses. Le
Sénégal comme tous les pays sahéliens a subi depuis
plusieurs décennies une forte variabilité climatique.
I.13.2. La variabilité climatique au
Sénégal
Pour analyser la variabilité climatique, on a
utilisé les données des stations de la moitié Sud du pays
pour couvrir ainsi la zone englobant les Forêts Classées
étudiées. La localisation et les types de ces stations sont
indiqués par la figure 11.
Article XI. Localisation et typologie des stations climatiques
utilisées.
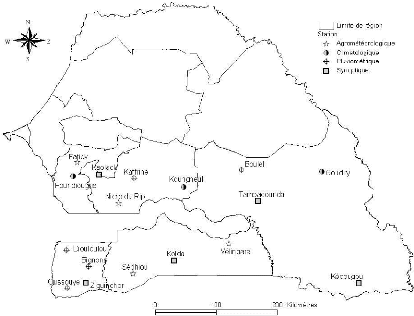
Les observations de température effectuées par
Hadley-Centre en 2003 montrent une forte augmentation des températures
globales depuis le 19ième siècle. Ces fortes variations sont
observées à la fois sur les surfaces terrestres et
océaniques (figure 12). En 2003, les températures moyennes
globales étaient supérieures de 0,7 °C à celles de
1920.
Article XII. Moyenne des températures terrestres depuis
la fin du 19ième Siècle (a). Moyenne des températures
terrestres et océaniques depuis la fin du 19ième Siècle
(b) (Hadley-Centre, 2003)
A
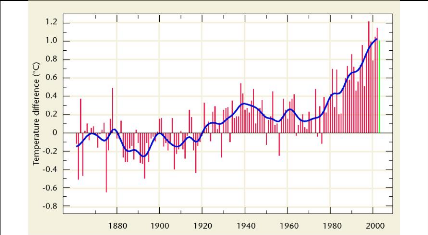
B
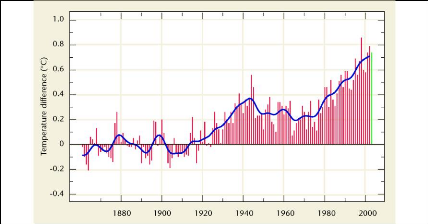
En Afrique, les variations de température obtenues
à partir des modèles montrent une augmentation de 0,8 °C
depuis 1900 si on considère les forçages naturels et anthropiques
(IPCC, 2008).
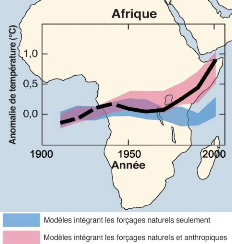
Ces variations de températures ont été
constatées au niveau des stations de la zone d'étude. Pour toutes
les stations synoptiques où des données de température
sont relevées, on y observe une anomalie positive. Ce qui est plus
frappant, c'est l'ampleur de ces anomalies, parfois assez élevée,
autour de 2 °C, soit deux fois les hausses moyennes enregistrées
sur le plan global ou Africain (figure 14). En gros, la hausse des
températures a été constatée à partir des
années 1980, ce qui correspond au point de départ des hausses
rapides observées par le Hadley-Centre (figure 12 et 13).

Article XIV. Ecart à la moyenne des températures
moyennes mensuelles à Kaolack, Kolda, Tambacounda et Ziguinchor
(1951-2003).
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Kolda
|
|
Kaolack
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Tambacounda
|
|
Ziguinchor
|
Les précipitations ont connu aussi une grande
variabilité temporelle intra saisonnière. La figure 15 montre la
courbe des précipitations annuelles des différentes stations de
la zone d'étude, mais aussi la tendance générale et la
tendance observée depuis 1980. Ces courbes indiquent que les
précipitations ont dans l'ensemble diminué dans la partie
étudiée mais que cette diminution est entrain de
s'infléchir depuis deux décennies, avec plusieurs années
de bonnes précipitations. Les prévisions d'un Sahel plus sec par
les scénarios climatiques adoptés par IPCC (2008) ne
correspondent pas pour l'instant à ce qui est observé ces
dernières années.
Article XV. Evolution interannuelle de la pluviométrie
(1921/1922-2006)
Kaolack (1921-2006)
Kolda (1922-2006)
Tambacounda (1922-2006)
Ziguinchor (1921-2006)
Cum/a
um/an
endnce énée
Kédougou (1922-2006)
endance Deps 1980
Fatick (1921-2003)
1980
Id
0
0
i
1
1
1
né nO
Foundiougne (1921-2003)
Kaffrine (1931-2003)
oynn CAot ndance D
um/an
endnc génale
Nioro du Rip (1931-2003)
Goudiri (1951-2003)
Koungheul (1950-2003)
Boulel (1950-2003)
WWd
N
0
0
0
0
Vélingara-Kolda (1932-2003)
Oussouye (1931-2003)
Tedne D
Sédhiou (1905-2003)
L'analyse spatiale par krigeage des données de
précipitations sur l'ensemble du territoire national depuis 1961 a
permis d'analyser les variations décennales. La figure 16, illustre une
forte migration des isohyètes vers le Sud avec cependant une
poussée inverse pour la dernière période, ce qui traduit
des années de bonne précipitations.
Article XVI. Dynamique spatiale des précipitations par
décennie.
|
1961-1970
|
1971-1980
|
|
1981-1990
|
1991-2000
|
|
2000-2005
|
|
Conclusion sur le contexte
géographique
Les formations végétales étudiées
sont des savanes de différents types qui subissent les préjudices
du climat et la pression humaine. Elles sont localisées dans la zone
soudanosahélienne, soudanienne et soudano-guinéenne, zones les
plus productives du Sénégal. Les précipitations ont en
général diminué dans la partie étudiée, mais
on constate une pluviométrie positive depuis une décennie. Les
densités de populations autour de ces forêts sont faibles pour les
Forêts du Sénégal Oriental et fortes pour celles du bassin
arachidier. Cette différence ne traduit pas une relation linéaire
avec les phénomènes anthropiques de dégradation, puisque
l'exploitation forestière formelle a lieu essentiellement au
Sénégal Oriental. D'une façon générale,
toutes les Forêts Classées étudiées subissent des
formes d'exploitation qui varient par leur nature et leur ampleur. Les sols
sont de type ferrugineux tropicaux, ou des sols minéraux bruts d'apport
éolien ou hydrique avec la présence de lithosols cuirassés
dans les Forêts Classées de Kantora et Bala.
Bibliographie
Adams, J.M., Piovesan G. 2002. Uncertainties in the role of land
vegetation in the carbon cycle. Chemosphere, 49:805-819.
Bassett, T.J., Zuéli K.B. 2000. Environmental
discourses and the Ivorian Savanna. Annals of the Association of American
Geographers, 90 (1):67-95.
Brown, S., Pearson, T., Slaymaker, D., Ambagis, S., Moore, N.,
Novelo, D., Sabido W. 2005. Creating a virtual tropical forest from
three-dimensional aerial imagery to estimate carbon stocks. Ecological
Applications, 15 (3):1083-1095.
Brown, S. 1996. Rôles actuel et futur des forêts
dans le débat sur le changement climatique mondial. In Unasylva
- No. 185 - Influences forestières. - Vol. 47 - 1996/2. FAO -
Organisation des Nations Unies pour l'alimentation et l'agriculture.
Accédé le 22 juillet 2008.
Brown S. 2002. Measuring carbon in forests: current status and
future challenges. Environmental Pollution, 116:363-372.
Brown S., 2003. Measuring and Monitoring Carbon for Land-Use
Change and Forestry Projects. Personal Communication. Winrock International,
Arlington.
Brown, S., Gaston G. 1995. Use of forest inventories and
Geographic Information Systems to estimate biomass density of tropical forests:
application to tropical Africa. Environmental Monitoring and Assessment
(38):157-168.
Campagna M., 1996. Le cycle du carbone et la forêt: de la
photosynthèse aux produits forestiers. Bibliothèque Nationale du
Québec, Québec, 51 p.
CCNUCC, 1997. Protocole de Kyoto. UNFCCC, 24 p.
Elberling, B., Touré, A., Rasmussen K. 2002. Changes in
soil organic matter following groundnut-millet cropping at three locations in
semi-arid Senegal, West Africa. Agriculture Ecosystems & Environment
(96):37-47.
FAO, 1995. FAO Annuaire : produits forestiers : 1982-1993, FAO,
Food and Agriculture Organization of the United Nations, Rome (Italie),
422p.
FAO, 1996. Influences forestières, Organisation des
Nations Unies pour l'Alimentation et l'Agriculture, Rome (Italie), 79p.
GIEC, 2003. Recommandations en matière de bonnes
pratiques pour le secteur de l'utilisation des terres, changements
d'affectation des terres et foresterie, IGES, Kanagawa,
Gray K.M., 2005. Changing soil degradation trends in Senegal with
carbon sequestration payments. Monography, Montana State University,
Montana.
Hadley-Centre, 2003. Climate change. Observations and
predictions, Met Office, Hadley Center, Exeter.
IPCC, 2001. Climate change 2001: Impacts Adataptation and
Vulnerability. A report of the Working Group II. Summury for Policy Makers. 18
p. , IPCC, Geneva.
IPCC, 2003. Good Practice Guidance for Land Use, Land-Use
Change and Forestry. Penman J., Kruger D., Galbally I., Hiraishi T., Nyenzi B.,
Emmanuel S., Buendia L., Hoppaus R., Martinsen T., Meijer J., Miwa K., and
Tanabe K. (Eds). Institute for Global Environmental Strategies, Kanagawa,
Japan.
IPCC (Editor) 2008. Bilan 2007 des changements climatiques.
Contribution des Groupes de travail I, II et III au Quatrième Rapport
d'Evaluation du Groupe d'Experts Intergouvernemental sur l'évolution du
climat. Intergouvernmental Panel on Climate Change, 103 p.
Lehtonen A., 2005. Carbon stocks and flows in forest
ecosystems based on forest inventory data No. Dissertationes Forestales 11,
Finnish Forest Research Institute. Vantaa Research Center, Helsinki.
Liu, S., Kairé, M., Wood, E., Diallo, O., Tieszen L.L.
2004. Impacts of land use and climate change on carbon dynamics in
South-Central Senegal. Journal of Arid Environments (59):583-604.
Lufafa, A., Bolte, J., Wright, D., Khouma, M., Diedhiou, I.,
Dick, R.P., Kizito, F., Dossa, E., Noller J.S. 2008. Regional carbon stocks and
dynamics in native woody shrub communities of Senegal's Peanut Basin.
Agriculture, Ecosystems and Environment (In press).
Manlay, R.J., Ickowicz, A., Masse, D., Floret, C., Richard,
D., Feller C. 2004. Spatial carbon, nitrogen and phosphorus budget of a village
in the West African savanna-I. Element pools and structure of a mixed-farming
system. Agricultural Systems (79):55-81.
Manlay, R.J., Kairé, M., Masse, D., Chotte, J-L.,
Ciornei, G., Floret C. 2002. Carbon, nitrogen and phosphorus allocation in
agro-ecosystems of a West African savanna I. The plant component under
semi-permanent cultivation. Agriculture, Ecosystems and Environment
(88):215-232.
Mbow C., 2005. Dynamic of carbon stock in woody vegetation of
savanna ecosystems in Senegal. The Land cover change perspective., Center for
Global Change and Earth Observations, Lansing, 20p.
Mbow, C., Mertz, O., Diouf, A., Rasmussen, K., Reenberg A.
2008. The history of environmental change and adaptation in eastern
Saloum-Senegal. Driving forces and perceptions. Global and Planetary Change, 64
(210-221).
MEA, 2005. Rapport de synthèse de l'Évaluation des
Écosystèmes pour le Millénaire, Millenium Ecosystem
Assessment.
MEPN, 1997. Plan National d'action pour l'Environnement.
Ministère de l'Environnement et de la Protection de la Nature.
République du Sénégal, p. 158.
Parton, W., Tappan, G., Ojima, D. P., T. 2004. Ecological
impact of historical and future
land-use patterns in Senegal. Journal of Arid Environments
(59):605-623.
Pearson, T., Walker, S., Brown, S., 2005. Sourcebook for Land
Use, Land-Use Change and
Forestry Projects, Winrock International, Arlington, 64p.
Rasmussen, K., Fog, B., Madsen J.E. 2001. Desertification in
reverse? Observations from Northern Burkina Faso. Global Environmental Change
(11):271-282.
Riedacker A., 2004. Changements Climatiques et Forêts.
SILVA, RIAT, 231 p.
Saugier R.M.B. 2003. Contribution des
écosystèmes continentaux à la séquestration du
carbone. Contribution of terrestrial ecosystems to carbon sequestration.
Comptes Rendus Geosciences, 335 (6-7):577-595.
Rosenberg, N.J., Blad, B.L., Verma S.B., 1983. Microclimate.
The Biological Environment. Chapter 8-Field Photosysthesis, respiration, and
the carbon balance (pp 288-390). John Willey and Sons, New
York-Chichester-Brisbane-Toronto-Singapore, 495 p.
Roxburgh S., 2004. The CASS Terrestrial Carbon Cycle Model,
V.1.2. User Guide and Tutorial exercices., CRC for Greenhouse Accounting, GPO
Box 475. Canberra ACT 2601, Australia.
Ruddiman, W.F. 2003. The anthropogenic greenhouse era began
thousands of years ago. Climatic Change (61):261-293.
Sagna P., 2005. Dynamique du climat et son évolution
récente dans la partie ouest de l'Afrique Occidentale, Thèse de
doctorat d'État. Université Cheikh Anta Diop de Dakar, 778p.
Sambou B., 2004. Evaluation de l'état, de la dynamique
et des tendances évolutives de la flore et de la
végétation ligneuse dans les domaines soudanien et
sub-guinéen au Sénégal. Doctorat d'Etat, Université
Cheikh Anta Diop de Dakar (UCAD), Dakar.
Schimel D.S. 1995. Terrestrial Biogeochemical Cycles: Global
Estimates with Remote Sensing. Remote Sensing of Environment (51):49-56.
Takimoto, A., Nair, P.K.R., Nair V.D. 2008. Carbon stock and
sequestration potential of traditional and improved agroforestry systems in the
West African Sahel. Agriculture, Ecosystems and Environment (125):159-166.
Tieszen, L.L., Tappan, G.G, Touré A. 2004. Sequestration
of carbon in soil organic matter in Senegal: an overview. Journal of Arid
Environments (59):409-425.
Touré A., 2002. Contribution à l'étude de
l'évolution des réservoirs de carbone en zone Nord-soudanienne au
Sénégal., Ecole Polytechnique Fédérale de Lausane
(EPFL), 158 p.
Tschakert P. 2004a. The costs of soil carbon sequestration: an
economic analysis for small-scale farming systems in Senegal. Agricultural
Systems (81 ):227-253.
Tschakert, P., Khouma, M., Sène M. 2004b. Biophysical
potential for soil carbon sequestration in agricultural systems of the Old
Peanut Basin of Senegal. Journal of Arid Environments (59):511-533.
Tschakert, P., Tappan G. 2004c. The social context of carbon
sequestration: considerations from a multi-scale environmental history of the
Old Peanut Basin of Senegal. Journal of Arid Environments (59):535-564.
Waring, R.H., Running S.W., 1998. Forest Ecosystems. Analysis at
multiple scales. Academic Press, California, 370 p.
Wood, E.C, Tappan, G.G, Hadj A. 2004. Understanding the drivers
of agricultural land use
change in South-Central Senegal. Journal of Arid Environments
(59):565-582.
Woomer P.L., 2003. Field and laboratory guidelines for carbon
characteristisation in African
Drylands. printed doc, SACRED Africa.
Woomer, P.L., Tieszen, L.L., Tappan, G., Touré, A., Sall
M. 2004a. Land use change and
terrestrial carbon stocks in Senegal. Journal
of Arid Environments (59):625-642.
Woomer, P.L., Tieszen, L.L., Tschakert, P., Parton, W.J.,
Touré A., 2001. Landscape carbon sampling and biogeochimical modelling.
A two-week skills development workshop conducted in Senegal., USGS, CSE.
Woomer, P.L., Touré, A., Sall M. 2004b. Carbon stocks in
Senegal `s Sahel Transition Zone. Journal of Arid Environments (59):499-510.
CHAPITRE II :
Modèles allométriques
Ce chapitre porte sur le développement de
modèles allométriques propres aux savanes soudaniennes et
soudano-guinéennes. Le choix de cette orientation s'explique par les
limites constatées lors de la revue bibliographique et à travers
l'application des méthodes connues et couramment utilisées
jusqu'ici pour les savanes. C'est ainsi que nous présentons dans ce
chapitre la démarche utilisée et les modèles mis au point.
L'analyse s'appuie sur les données collectées dans des
Forêts Classées représentatives des savanes soudaniennes et
soudanoguinéennes du Sénégal que sont Patako,
Wélor, Ouli, Bala, Kantora et Mampaye.
II.1. L'importance des modèles
allométriques
L'allométrie d'un arbre comme ses dimensions
structurales se définit par différentes mesures et leurs
relations avec la masse ou le volume des individus (Lehtonen, 2005). Les
techniques allométriques permettent de générer ainsi la
biomasse des individus ligneux en se basant sur des mensurations de
paramètres dendrologiques sur l'individu. L'allométrie est une
vieille technique forestière et constitue les premiers types de
modèles statistiques permettant de générer des
informations quantitatives sans détruire a posteriori les individus. Le
terme générique utilisé à cet effet est
«l'approche non destructive» par opposition à celle dite
<<destructive>> qui a justement permis d'élaborer ces
modèles allométriques.
L'évaluation du potentiel ligneux des formations
forestières constitue un important domaine de recherche pour des besoins
de gestion des ressources forestières ligneuses. L'intérêt
à estimer le tonnage de bois dans une forêt donnée a
été le plus souvent connecté à des besoins
commerciaux ou de satisfaction des besoins des populations en bois. Cependant,
l'estimation des réserves de bois n'a eu à cibler que les
espèces qui ont une grande valeur marchande, énergétique
ou esthétique (Brown, 1997). Cette situation a longtemps prévalu,
depuis la période coloniale avec la réalisation de plans
d'aménagements forestiers destinés à l'exploitation des
formations forestières basée sur une bonne connaissance du
potentiel ligneux. Ainsi, l'évaluation du «volume marchand»
des forêts tropicales avait pour objectif d'estimer la partie `utile' de
l'arbre. A ce titre, les forestiers ont pu établir des relations
mathématiques simples entre des paramètres mesurés sur
l'arbre (diamètre du tronc ou sa circonférence, hauteur totale de
l'arbre, hauteurs de découpe et longueurs de billons) et le volume de
bois exploitable.
Le cubage des arbres présente un intérêt
économique permettant d'évaluer le volume commercialisable sur
une essence ou un groupe d'espèces. Il s'agit des tarifs de cubages
classiques. Les formules utilisées sont très différentes
mais utilisent toutes des mesures de tronc et de hauteur des individus.
L'équation générique utilisée est de la forme V=
kCb (k et b sont les tarifs de cubage, C est la mesure du tronc).
Des équations logarithmiques ou cubiques ont servi à
établir de telles relations entre les mesures sur l'arbre et le volume
de bois <<utile>>.
Les paramètres mesurés (lors des inventaires
forestiers) sont dès lors utilisés pour établir de telles
équations afin de calculer des volumes individuels des arbres. C'est
avec l'ajustement de ces équations aux données observées
qu'il a été possible d'établir des tables de cubage qui
expriment le volume de bois fort correspondant au volume de la tige
jusqu'à une certaine
circonférence (Rondeux, 1993). Le reste de la biomasse
de l'arbre constitué par le feuillage et les petites branches n'est pas
pris en compte. L'élaboration de tables de cubage nécessite une
approche de récolte de biomasse sur un échantillon
d'espèces de tailles différentes. Les modèles permettent
par la suite d'évaluer la phytomasse exploitable sans passer par ces
méthodes destructives.
Il apparaît alors, que ces tarifs de cubage
négligent le plus souvent la phytomasse totale des arbres et ne portent
que sur le volume de bois exploitable de certaines espèces
ciblées et n'ont été développés que pour
quelques formations végétales notamment forestières.
L'utilisation de ces équations donne ainsi une indication limitée
dans le cadre d'une étude de la biomasse ligneuse totale pour estimer le
stock total de carbone.
L'approche par tarif de cubage a une limite pour les
aménagistes dans la planification de l'exploitation forestière
pour les autres usages, comme le bois de feu par exemple qui peut être
satisfait notamment avec les petites branches. Partant de ce constat, de
nombreuses études ont été entreprises pour établir
des relations mathématiques simples entre données d'inventaire de
la végétation et biomasse totale des ligneux. Ces
équations mathématiques sont des régressions simples
(linéaires, exponentielles, logarithmiques, polynomiales), qui
permettent de dérouler une approche non destructive aboutissant à
l'estimation de la biomasse totale à partir de paramètres
mesurés sur les arbres et leur poids total. Ces relations ont un double
intérêt : d'abord, elles permettent de mieux calibrer les plans
d'aménagement en tenant compte de tout le potentiel ligneux ; ensuite
elles facilitent, dans le contexte actuel des changements climatiques,
l'estimation du stock de carbone en partant de la connaissance de la biomasse
totale. Le potentiel de stockage de carbone est central au rôle que
l'Afrique doit jouer par la réduction de la déforestation
(changements d'affectation des terres) mais aussi l'effort de reforestation qui
devient une contribution positive dans la séquestration du carbone.
Cependant, la plupart de ces essais d'estimation de la
biomasse totale des arbres ont été menés en dehors des
savanes africaines. Les chercheurs ont surtout travaillé dans des
écosystèmes tempérés ou dans les forêts
tropicales de l'Amérique du Sud (San Jose et al, 1998;
Moura-Costa et Stuart, 1999; Nelson et al., 1999; Clark et Clark,
2000; Keller et al., 2001; Brown, 2002; Fleurant et al., 2004
; Zianis et Mencuccini, 2004; Brown, 2005). La FAO au travers de la
synthèse de Brown (1997), a permis une systématisation des
méthodes existantes. Cette étude est largement utilisée
à l'heure actuelle pour estimer la biomasse des savanes africaines, avec
le risque lié aux biais non connus qu'on ne peut éviter qu'en
élaborant un modèle pour les zones étudiées.
Sur e plan pratique, l'analyse des stocks de carbone ligneux
passe nécessairement par l'estimation de la biomasse sur pied. La
biomasse ligneuse sur pied exprime la masse de matière vivante des
arbres. La masse du bois est déterminée soit par simple
pesée, soit par multiplication de son volume par sa «masse
volumique», ou masse par unité de volume exprimée en
kg/m3. La masse brute est influencée par la densité du
bois, par son degré d'humidité et éventuellement par
l'importance (épaisseur) de l'écorce. L'estimation de la masse
sèche est plus complexe, puisque la densité du bois varie en
fonction de sa teneur en eau. La densité du bois est assimilable
à sa masse à l'état humide par unité de volume
(Rondeux, 1993). Celle-ci concerne les troncs des arbres et les ensembles
constitués des branches, des feuilles et des souches. Les
méthodes d'estimation indirecte sont envisageables en ce sens qu'elles
sont non destructives et sont basées sur l'utilisation
d'équations mathématiques ou des tables exprimant la biomasse en
fonction de caractéristiques facilement mesurables comme le
diamètre à 1,3 m du sol et la hauteur totale. Les modèles
les plus utilisés se présentent généralement sous
forme de relations statistiques.
Du fait de l'intérêt grandissant dans
l'estimation des stocks de carbone des formations forestières, des
techniques spatiales (la télédétection) ont
été testées. Elles ont permis d'estimer les
éléments de structures d'une formation, souvent lourds et longs
à collecter sur le terrain. Puisque les écosystèmes de
savane sont souvent hétérogènes, des difficultés
sont notées dans les choix d'échantillonnage et la mise en
pratique des inventaires. Ainsi, l'estimation de certains paramètres
à partir des données spatiales devient très attractive
dans l'estimation de la biomasse de la végétation naturelle. Mais
les techniques proposées à ce jour sont assez complexes et
difficiles à opérationnaliser. Brown (1997); Brown et
al. (2005) proposent un système imageur multispectral en trois
dimensions, alors que plusieurs autres auteurs ont testé la technologie
LIDAR et du RADAR plus utilisée dans le domaine de l'altimétrie
topographique pour analyser la structure verticale des formations ligneuses et
d'en déduire des stocks de carbone à partir des volumes
générés sur les données en trois dimensions (3D).
Le LIDAR, le RADAR et les techniques de photogrammétrie numérique
stéréoscopiques (Télédétection 3D) ont
été proposés par Awaya et al. (2004); St-Onge
et al. (2004) et Proisy (1999), pour collecter des données sur
la hauteur et la densité des formations forestières. Avec le
développement de la Très Haute Résolution, des tests
d'estimation de la biomasse ont été réalisés
à partir des données IKONOS par Thenkabail et al. (2002)
puis par Awaya et al. (2004) pour estimer la biomasse de plantations
de palmiers à huile au Bénin et au Nigéria ou des
formations de sapin au Japon. Ces techniques sont limitées par la
lourdeur des opérations de collecte de données (complexe) et le
niveau de couverture des images qui nécessite de longs processus
techniques pour arriver à des
résultats, souvent pas directement exploitables sur la
structure de la végétation. Il faut aussinoter que les
techniques de télédétection ne permettent de distinguer,
de façon précise, les différentes espèces des
formations étudiées.
D'autres essais, purement statistiques ont été
faits, notamment avec le modèle mathématique proposé par
Fleurant et al. (2004), pour la description des caractères
morphométriques en partant des théories fractales. La
théorie fractale permet de faire une segmentation hiérarchique
des différentes parties de l'arbre (inspiré du concept de
Stralher, utilisé en hydrologie). Elle fournit alors des outils pour
l'analyse quantitative de la structure complexe des arbres. La validation de
ces types de modèles est loin de prouver qu'on puisse se passer des
données de terrain.
Les données dendrométriques issues des
inventaires de la végétation ligneuse constituent pour l'instant
la méthode la plus éprouvée pour estimer la biomasse de la
végétation ligneuse. Ces inventaires sont faits dans chaque pays
pour répondre aux besoins de la caractérisation du potentiel
forestier, de l'analyse de la biodiversité des forêts et de la
conception de plans d'aménagement pour une gestion durable des
ressources ligneuses. Les données d'inventaire sont mises à
profit pour estimer les quantités de biomasse totale des formations
dominantes. La biomasse est calculée pour chaque sujet ligneux par des
équations qui incluent le plus souvent le diamètre de l'individu
et parfois la hauteur.
II.2. Développement et utilisation des
modèles allométriques
II.2.1. L'état des connaissances sur les
modèles allométriques
Les modèles allométriques sont des relations
mathématiques entre des mesures dendrométriques des arbres
(diamètre, hauteur) et leur masse totale. L'estimation de la biomasse
est un exercice obligatoire pour déterminer les stocks de carbone dans
une formation végétale. Cette information est devenue la base de
l'évaluation et des inventaires du stock de carbone et de son
évolution dans la perspective des projets MDP (boisement, reboisement).
Ces derniers sont définis par le Protocole de Kyoto (CCNUCC, 1997) et
les accords de Marrakech (COP 7, 2001) comme un point d'ancrage de la
contribution des pays en développement (pays non Annexe I) dans l'effort
d'atténuation des changements climatiques. L'estimation de la biomasse
est une étape essentielle dans l'évaluation du stock de carbone
et les approches utilisées varient en fonction de plusieurs facteurs :
types de végétation, objectifs initiaux, méthodes
statistiques d'échantillonnage, techniques de mesures, etc.
Cependant, le modèle mathématique le plus
utilisé pour l'évaluation de la biomasse sur pied est de la forme
:
M=aDb : (a et b sont les coefficients
d'ajustement, D le diamètre à 1,3 m et M la biomasse totale),
Zianis et Mencuccini (2004).
Dans la plupart des cas, la variabilité de M est
largement expliquée par la variabilité de D. Les coefficients a
et b varient en fonction des espèces, de l'âge des arbres, des
conditions stationnelles, etc. Les techniques les plus courantes pour
déterminer les coefficients a et b sont des régressions par
moindres carrés sur des données logarithmiques
transformées de D et M issues des données de terrain. La
méthode dite destructive utilisée dans ce genre d'exercice
demande beaucoup de temps et de moyens. Le choix des sites requiert aussi une
attention particulière pour les extrapolations futures
(homogénéité et représentativité des
sites).
II.2.2. Généralités sur les mesures de
la masse ligneuse
La masse de bois est mesurée par simple pesée ou
par multiplication de son volume par sa
Mo ?
masse volumique (masse par unité de volume :
kg/m3). On parle aussi de densité pour désigner la
masse volumique. Il est relativement simple de déterminer la masse brute
de bois, mais l'estimation de la masse sèche (à l'état
anhydride) est plus complexe et requiert un séchage d'une partie de
l'échantillon à l'étuve jusqu'à atteindre une masse
constante. La masse brute est fonction de la densité, du taux
d'humidité et accessoirement de l'épaisseur de l'écorce.
Puisque la densité du bois dépend de la teneur en eau, les types
de volume de bois mesurés sont selon Rondeux (1993) :
1) le volume de bois vert ou à l'état
saturé (Vs) ;
2) le volume de bois sec à l'air ou à 12%
d'humidité (V12) ;
3) le volume de bois à l'état anhydride (Vo).
Les paramètres les plus utilisés sont la
densité à l'état anhydride et l'infra-densité.
Do= Mo/Vo (Masse volumique = Masse à l'état
anhydride/ Volume à l'état anhydride) ; D12= M12/V12 (Masse
volumique normale ou masse à l'état sec à l'air, soit 12%
d'humidité) Ds=Ms/Vs (Masse volumique à l'état vert, i.e.
bois saturé)
I=Mo/Vs (Infra-densité = Masse à l'état
anhydride/volume à l'état sec)
La masse volumique dépend des espèces. Elle est
influencée par l'essence, les variations climatiques annuelles, les
facteurs topographiques et stationnels. Il est important de retirer
l'humidité de la masse totale pour obtenir la biomasse sèche. Le
taux d'humidité est calculé de la façon suivante :
Ces fondements théoriques ont été
utilisés pour structurer une approche d'estimation de la biomasse
sèche totale des arbres.
- Les techniques expérimentales utilisées
pour l'estimation de la biomasse ligneuse
La biomasse ligneuse aérienne est la quantité,
exprimée en masse de la matière vivante des arbres. Elle concerne
les troncs et les ensembles constitués des branches, des feuilles.
Les diverses opérations à suivre pour estimer la
biomasse des arbres sont d'après Rondeux (1993) de :
1. prendre diverses mesures (diamètre à 1,3 m,
hauteur de l'individu, diamètre du houppier) ;
2. abattre l'arbre et procéder à son
débitage par catégories de diamètres ;
3. séparer les diverses composantes (tronc, branches,
ramilles, feuilles, etc.) ;
4. récolter les données relatives aux dimensions
(longueurs, diamètres, etc.)
5. peser les composantes vertes (avant séchage) ;
6. prendre des échantillons (disques sur le bois de tige
et de branche, sur les feuilles)
7. peser les échantillons verts ;
8. sécher ces échantillons à l'étuve
jusqu'à avoir un poids constant.
- Pour les troncs et grosses branches
9. peser la masse à l'état humide sur le terrain
après avoir tronçonné la tige en tenant compte des
critères de grosseur,
10. prélever et peser un échantillon sur place,
11. peser cet échantillon après séchage.
Le rapport entre la masse à l'état anhydride et
la masse l'état humide de l'échantillon est appliqué
à la masse à l'état humide totale afin d'estimer la masse
à l'état anhydride totale. La formule suivante permet de
convertir la masse humide en masse sèche.
Mat = (Ma/Mh)*Mht
Où : Mat = Masse anhydride totale ; Ma = Masse
anhydride de l'échantillon ; Mh = Masse humide de l'échantillon ;
Mht = Masse humide totale. Ma/Mh est un taux de conversion qu'il est plus
judicieux de calculer pour les différentes parties de l'arbre (tronc,
branches et feuillage).
- Pour les branches et les rameaux
Ces mesures concernent le volume ou la masse des
échantillons en faisant attention à la variation de
densité et au taux d'humidité au sein d'une même branche.
En général, la masse des branches et rameaux, concerne les deux
premiers ordres de ramification (ordre 1 et 2), figure 17.
Figure 17. Les parties concernées par la
mesure des branches et des rameaux
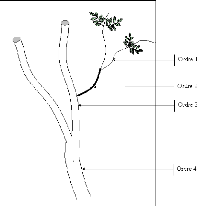
- Pour les feuilles
Il faudra peser la masse totale et faire des
prélèvements pour déterminer le taux d'humidité
afin de déduire la biomasse sèche totale. On procède
à la même démarche que pour les troncs et tiges pour
estimer la masse sèche totale.
II.2.3. Démarche adoptée pour les mesures de
terrain
- Stratification
Cette partie concerne la cartographie des Forêts
Classées ciblées pour la collecte de données de terrain.
La stratification est une représentation cartographique des
unités d'occupation du sol représentant les différentes
formations végétales présentes sur le terrain. La
télédétection et les Systèmes d'Information
Géographiques (SIG) ont permis de réaliser les cartes en
utilisant les images LANDSAT de 20015. Des
classifications et autres traitements classiques permettent de segmenter les
images en zones homogènes à partir desquelles le dispositif de
collecte a été appliqué (chapitre 3, figure 35).
5 Les données les plus récentes qu'on a pu obtenir
correspondent à cette date. La base de données LANDSAT est
maintenir d'accès libre et les travaux postérieurs devraient
être basés sur des données plus récentes.
Les inventaires antérieurs permettent de
vérifier d'une part si les espèces principales sont bien
présentes dans l'échantillon récolté et d'autre
part si la distribution des individus de ces espèces dans les
différentes classes de diamètres. Cette approche présente
des avantages sur l'alternative d'une sélection `raisonnée' des
sujets avec ses nombreux risques de biais.
Ainsi, l'ensemble des mesures s'opère sur des placettes
choisies sur le terrain, sur des sites homogènes incluant
différents types de végétation. La méthode
proposée par Rondeux (1993) et consistant à déterminer la
biomasse totale des sites, en se basant sur les arbres moyens n'a pas
été recommandée par Pearson et al. (2005) qui
préconise l'approche suivante inspirée de IPCC (2003).
- Les étapes de la collecte
En tenant compte des espèces dominantes et de la
présence d'individus de gros diamètres, les grandes étapes
de terrain sont les suivantes :
1. sélectionner l'individu (parmi les espèces
dominantes) ;
2. échantillonner près de 30 individus avec
différentes classes de diamètre, mais avec
un biais pour les gros diamètres (qui vont dominer la
biomasse) et pour les espèces dominantes de la zone (cf. inventaires
antérieurs) ;
3. mesurer le DBH (diamètre à 1,3 m) et la hauteur
des individus ; pour les individus
dont le diamètre est inférieur à 5 cm la
démarche suivante sera adoptée :
a. couper l'individu à la base,
b. séparer les tiges et les rameaux foliaires,
c. mesurer le poids de chaque composante,
d. échantillonner une partie sur les tiges et une autre
sur les feuilles 2
e. sécher l'échantillon et calculer la masse
sèche, (poids total
multiplié par taux d'humidité)
4. récolter les arbres sélectionnés
à la base du tronc ;
5. couper l'arbre en morceaux pour mesurer la masse à
l'état frais ;
6. si la découpe de gros troncs d'arbres est fastidieuse,
estimer le volume en
mesurant le diamètre sur les extrémités et
sa longueur. On évite cette alternative pour ne pas subir les biais de
mesures liés à sa mise en ~uvre.
Figure 18. Estimation du volume de tronc trop
volumineux
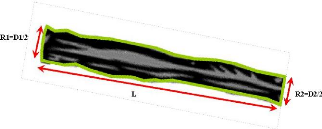
· Collecter un disque sur le bois frais, estimer le volume
(volume= H*itr2), sécher à l'étuve et mesurer
la masse sèche.
Figure 19. Mesures à effectuer pour le
calcul du volume d'un disque pour la densimétrie.
· Estimer la masse du tronc en utilisant la
densité du bois (Masse= Volume * Densité) et
ajouter les autres composantes (masse des branches, feuilles, tiges) pour
obtenir la masse totale de l'arbre.
7. calculer l'équation allométrique par les
relations entre la biomasse de l'arbre et le DBH seulement ou en le combinat
à la hauteur.
En somme l'approche allométrique est en
perpétuelle raffinement, mais le fondement
statistique est le
même ; partir des mesures de l'arbre pour en déduire sa masse. Ce
quivarie le plus souvent c'est la méthode
d'échantillonnage permettant de choisir les
individus à inclure dans le modèle en tenant
compte de la distribution spatiale, de la taille des sujets et des
espèces.
II.2.4. Les modèles allométriques
existants
Pour estimer la biomasse ligneuse sur pied, plusieurs
modèles allométriques ont été utilisés dans
la zone tropicale. Récemment, Lufafa et al., (2008), ont
développé des régressions pour estimer la biomasse d'une
espèce buissonnante (Guiera senegalensis). Les formules
utilisées par ces auteurs sont les suivantes :
· pour la biomasse sur pied à partir du
diamètre moyen de la couronne - log y = 5,12 + [0,012*uDC] (y est la
biomasse, uDC le diamètre moyen de la couronne du
sujet)6 ;
· pour la biomasse racinaire à partir du
diamètre du tronc à la base - log y = 7,52 + [0,015*uDT] (y est
la biomasse, uDT le diamètre du tronc à la base du sujet).
La difficulté avec cette méthode est l'obtention
de mesures précises du diamètre des couronnes de cette
espèce très grégaire et largement étalée sur
le sol. Le diamètre de la base des sujets est tout aussi difficile
à estimer car cette espèce est le plus souvent multicaule avec de
tout-petits troncs en grand nombre, souvent exploités pour leur bois ;
ce qui fait que les branches résiduelles ne sont que des repousses.
Pour une extrapolation des stocks de carbone, des applications
se développent de plus en plus à partir de la modélisation
spatiale, mais aussi par la recherche de corrélations entre
paramètres biophysiques mesurés sur le terrain et estimateurs de
ces paramètres avec des données spatiales. La conversion de
signatures spectrales des images Haute ou Moyenne Résolution en taux de
couverture du sol et la reconversion du taux de couverture en biomasse est une
possibilité testée par St-Onge et al. (2004); Brown
et al. (2005); Lufafa et al. (2008). Cette approche
présente des inconvénients liés aux erreurs additionnelles
issues des biais des indices utilisés : erreurs de conversion en taux de
couverture ou densité de la végétation, et erreurs
liées aux corrélations avec la biomasse. Pour minimiser ces
erreurs, il est possible de passer par une bonne caractérisation
(cartographie) des types de végétation homogène, un taux
d'échantillonnage représentatif lors des inventaires de ces types
de végétation et l'utilisation de modèles
allométriques pour convertir les données d'inventaire en
biomasse.
6 Lufafa et al., (2008) passe la diamètre
moyen de la couronne des individus de Guiera senegalensis, car cette
espèces est buissonnantes. Mais l'estimation du diamètre de la
couronne dans ce cas d'espèce présente des risques en termes de
confusion entre les individus.
Les estimations de biomasse à partir des
régressions statistiques proposées par Brown (1997)
présentent des limites au niveau de la validation ; celle-ci
étant faite sur les écosystèmes de savane de
l'Amérique du Sud, ou des forêts sèches d'Inde. Toutefois,
en relativisant les choses, on peut considérer que la mise en ~uvre de
ces équations se fonde sur des données de base collectées
dans des écosystèmes dont la répartition est fonction de
la pluviométrie (tableaux 4). Ainsi pour rester dans les conditions
climatiques proches de celles de zones considérées, l'application
de ces équations sur le Sénégal intègre les
modèles développés pour des précipitations
annuelles comprises entre 300 et 1500 mm/an.
Tableau 4. Régressions
allométriques proposées par Brown (1997)
|
Modèle allométri que
|
Zone climatique
|
Equation
|
Intervall e 0 en
cm
|
r2
ajusté
|
|
FAO 1
|
Forêts sèches Indes (pmm non indiquée)
|
Y = exp(- 1,996+2,32*ln(D))
|
5-40
|
0.89
|
|
FAO 2
|
Forêts sèche Mexique,
pmm> 900 mm
|
Y = 42.69- 12.800(D)+1.242(D2)
|
5-148
|
0.84
|
|
FAO 3
|
Forêts sèche Mexique,
pmm< 900 mm
|
Y = exp(- 2.134+2.530*ln(D))
|
Non
indiqué
|
0.97
|
|
FAO 4
|
Forêts sèche Mexique,
pmm< 900 mm
|
Y = 21.297- 6.953(D)+0.740(D2)
|
4-112
|
0.92
|
T= biomasse par sujet en kg, D = dbh en cm.
Aucune équation ne doit être utilisée pour
estimer la biomasse si le diamètre des sujets dépasse
l'intervalle indiqué. C'est ainsi que ces modèles ont
été testés sur des diamètres de moins de 80 cm
(l'essentiel des individus sont dans cette catégorie au niveau des
savanes). Cependant on note une forte divergence sur les résultats de
ces modèles allométriques quand on l'applique aux données
de savanes du Sénégal. La figure 20 montre que ces modèles
ne présentent pas les mêmes performances.
Figure 20. Tests des méthodes
allométriques utilisés dans les écosystèmes
tropicaux
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Modèles FAO (Brown, 1997)
|
Code Excel
|
|
Y = exp(-1,996+2,32*ln(D))
|
FAO1 : =EXP((-1,996)+2,32*LN(DBH))
|
|
Y = 42.69-12.800(D)+1.242(D2)
|
FAO2 : =42,69-12,8*DBH+1,24*PUISSANCE(DBH;2)
|
|
Y = exp(-2.134+2.530*ln(D))
|
FAO3 : =EXP((-2,134)+2,53*LN(DBH))
|
|
Y = 21.297-6.953(D)+0.740(D2)
|
FAO4 : =21,297-6,953*DBH+0,74*PUISSANCE(DBH;2)
|
Les tests réalisés sur ces équations
montrent une grande divergence sur la biomasse estimée à partir
de 15 cm de DBH (figure 20). Ainsi l'estimation de la biomasse des gros
diamètres constitue le principal problème de ces
régressions. Entre le modèle FAO 1 (Y = exp(- 1,996+2,32*ln(DBH))
et le modèle FAO 3 (Y = exp(-2.134+2.530*ln(DBH)), la biomasse
estimée pour les gros diamètres passe du simple au double. Ainsi,
l'utilisation de ces modèles pose le problème de surestimation ou
de sous estimation de la biomasse ce qui pourrait avoir un impact important sur
les calculs d'additionalité des projets MDP forêt.
FAO 1 (EXP((19)+23*LN(DBH))
Cependant, il n'existe pas d'approche standard pour estimer la
biomasse sur pied des zones
(P(-23)2,5(BH))
FA 0ISS
de savane basée sur des données d'inventaire du
fait du manque de données consistantes. Les méthodes
développées pour les forêts denses ou pour les savanes
d'Amérique Latine sont difficilement transposables aux formations de
savane du Sénégal qui présentent des taxons
caractéristiques (multicaules, bas branchus).
En somme, on a noté que plusieurs modèles
allométriques existent, mais il est rare d'en
trouver sur les
écosystèmes arides et subhumides d'Afrique de l'Ouest (Brown,
1997). A
6 1 6 21 26 3 36 4 46 51 56 61 6 71 76
défaut de mieux, plusieurs recherches antérieures
utilisent les modèles existants (Woomer,
DBH
2003; Woomer et al., 2004; Mbow, 2005)
développés dans des écosystèmes aux climats
similaires (mais pas identiques) à ceux des savanes
d'Afrique de l'Ouest. Les différences entre les sites d'études
entraînent des erreurs méconnues quand on applique les
modèles `exogènes'. C'est pour cette raison qu'un effort est
consenti dans chaque pays pour développer des approches adaptées
et opérationnelles. Cette volonté de développement de
méthode est sous-tendue par les problèmes méthodologiques
rencontrés lors des inventaires nationaux des gaz à effet de
serre, par le manque ou la faible fiabilité des données et par la
grande incertitude sur la précision des résultats (CGE, 2005). Il
convient dès lors de baser l'estimation de la biomasse des
écosystèmes sur des régressions allométriques
natives (endogènes), se fondant sur une collecte de données par
méthode d'abattage-pesage comme cela a été fait dans
d'autres les écosystèmes d'Amérique ou d'Europe.
Développer des équations allométriques
pour l'estimation de la biomasse est un exercice coOteux et dont la
réalisation reste complexe. Dans la perspective de l'évaluation
des stocks de carbone dans les formations forestières, les mesures
dendrométriques classiques sont de plus en plus renforcées par
des mesures de phytomasse. Dans la méthode proposée ici, il est
question d'évaluer la biomasse ligneuse sur pied. L'information sur la
biomasse sur pied des arbres et arbustes doit être recueillie sur les
parties les plus complètes possibles des individus
échantillonnés en considérant les grands et petits arbres.
La base de la collecte de biomasse pour cette thèse est
détaillée au point suivant.
II.2.5. Méthode retenue pour l'élaboration de
modèles allométriques7
Avant de procéder à la collecte de phytomasse
sur le terrain, il a été nécessaire d'analyser les
résultats de données d'inventaires antérieures sur les
Forêts Classées étudiées. Nous sommes ainsi partis
des recherches de Sambou (2004) pour identifier les espèces dominantes,
leur localisation spatiale et leur distribution dans les différentes
classes de diamètre. Il s'est agi alors de collecter les individus
au prorata de l'abondance de l'espèce et en tenant compte de
leur distribution dans les classes de diamètres. Ainsi, les
espèces les plus abondantes ont été les plus
échantillonnées et les classes de diamètres les plus
abondantes ont aussi fait l'objet de collecte de plusieurs échantillons.
Ce choix répond de l'exigence de la représentativité des
échantillons collectés. La démarche logique et
chronologique qui a été suivie est présentée comme
suit.
1. Mesurer le DBH, la hauteur, la circonférence du tronc
et du houppier avant de couper l'arbre ;
2. Couper l'arbre à la base ;
3. Faire une série de mesures :
7 Il s'agit de la méthode appliquée pour
générer nos propres modèles allométriques
· Longueur de l'arbre (de la base au sommet)
· Longueur du tronc (de la base aux premières
ramifications) ;
4. Suspendre une balance de 100 kg sur un support (en
suspension) ;
5. Répartir la biomasse de l'arbre en individualisant le
tronc, les branches et le feuillage et mesurer le poids des différentes
parties:
· Des échantillons de disques sont collectés
et pesés avec une balance électronique sur différentes
sections du tronc, sur les branches et les feuilles pour estimer après
séchage le taux d'humidité ;
· Pour les grosses branches :
o Les couper en morceaux et faire l'addition des poids
· Les troncs:
o Mesurer leur poids dans la mesure du possible,
o Couper 2 disques sur le tronc et 2 disques sur les
branches, les peser ou estimer leur volume en utilisant la formule de mesure du
volume d'un cylindre. Faire la moyenne de L1 et L2, celle de T1 et T2 (figure
21). Volume = r2ð*h
Figure 21. Mensuration pour mesure de volume de
disque
· Calculer la masse sèche de chaque partie en
utilisant le taux d'humidité mesuré après séchage
(masse humide * taux d'humidité) ;
· Ajouter le poids sec des parties pesées pour
calculer la masse totale de l'arbre.
Quelques unes de ces étapes sont illustrées
à la figure 22.
Figure 22. Illustrations de quelques
étapes de la méthode

3
5
7
1
4
2
6
8
|
Photo 1 : équipement de terrain ;
Photo 3 : mesure de circonférence (diamètre).
Photo 5 : échantillons de feuilles, branche et tronc.
Photo7 : Billes d'un tronc de Cordyla pinnata ;
|
|
Photo 2 : Balance analogique pour la mesure de biomasse ;
Photo 4 : Abattage d'un arbre
Photo 6 : pesée de la masse foliaire
Photo 8 : Séchage des échantillons à
l'étuve.
|
II.2.6. Traitement des données et élaboration
du modèle allométrique
II. 2.6.1. Présentation des données
collectées
Dans la perspective de l'élaboration du modèle
allométrique, la collecte de biomasse est basée sur des
données d'inventaire recueillies entre 2000 et 2003 (Sambou, 2004). Ces
recherches antérieures ont permis de connaître les espèces
dominantes dans chaque Forêt Classée des savanes soudaniennes et
soudano-guinéennes ciblées. Le nombre d'individus
échantillonnés par classe de diamètre pour chaque
espèce dominante a été déterminé en fonction
de l'abondance de l'espèce dans la classe de diamètre.
L'observation principale dans le choix de ces espèces est la dominance
de Combretum glutinosum dans 5 forêts étudiées.
Dans la Forêt de Wélor Acacia seyal est l'espèce
dominante. Au total, au moins trois espèces co-dominantes ont
été échantillonnées dans chaque Forêt
Classée. La liste globale des espèces ligneuses concerne 13
espèces et 101 individus ont été
échantillonnés. La distribution de ces individus dans les classes
de diamètre est indiquée sur le tableau 5.
Tableau 5. Nombre d'individus
échantillons par espèce et par Forêt Classée.
|
|
|
Forêt classée
Espèce
|
Bala
|
Kantora
|
Ouli
|
Patako
|
Mampaye
|
Wélor
|
Total
|
|
1
|
Acacia macrostachya
|
7
|
|
7
|
|
|
|
14
|
|
2
|
Acacia seyal
|
|
|
|
|
|
6
|
6
|
|
3
|
Balanites aegyptiaca
|
|
|
|
|
|
5
|
5
|
|
4
|
Bombax costatum
|
|
1
|
|
|
|
|
1
|
|
5
|
Combretum glutinosum
|
10
|
10
|
7
|
8
|
|
4
|
39
|
|
6
|
Combretum nigricans
|
|
|
|
6
|
|
|
6
|
|
7
|
Cordyla pinnata
|
|
|
2
|
4
|
|
|
6
|
|
8
|
Crossopterix febrifuga
|
|
|
|
|
3
|
|
3
|
|
9
|
Daniellia oliveri
|
|
|
|
4
|
|
|
4
|
|
10
|
Pterocarpus erinaceus
|
|
|
|
3
|
|
|
3
|
|
11
|
Sterculia setigera
|
|
|
|
|
1
|
|
1
|
|
12
|
Terminalia macroptera
|
|
8
|
|
1
|
|
|
9
|
|
13
|
Ximenia americana
|
|
|
|
|
|
4
|
4
|
|
Total
|
17
|
19
|
16
|
26
|
4
|
19
|
101
|
La biomasse humide mesurée sur le terrain est
égale à 15348,95 kg, convertie en biomasse
sèche en utilisant le facteur d'humidité calculé à
partir des échantillons des différentes parties de l'arbre
(feuilles, branches, tronc). Les échantillons de biomasse ont
été séchés à 60 °C et des pesés
réguliers ont permis d'identifier le moment de totale dessiccation quand
le poids de l'échantillon devient stable. Le temps de séchage des
troncs a souvent été plus long. Le taux

d'humidité des échantillons des
différentes parties de l'arbre est le rapport entre poids sec et poids
humide, et le facteur obtenu est multiplié par la biomasse humide. La
biomasse sèche totale de l'arbre est une somme de la biomasse
sèche du tronc, des branches et des feuilles. Dès lors, il
était possible de générer des régressions entre
biomasse sèche et DBH pour élaborer les modèles à
partir desquels on peut estimer la biomasse sèche totale.
La recherche de corrélation entre biomasse et DBH passe
par l'inspection de la distribution du nuage de points de ces deux
paramètres. Auparavant, on a diagnostiqué l'allure de la courbe
des DBH et de la biomasse. La figure 23 montre que l'augmentation du
diamètre des arbres n'avait pas la même allure graphique que celle
de la biomasse correspondante. La variation de la biomasse augmente lentement
jusqu'à un certain diamètre de tronc (~ 17 cm), pour ensuite
croître très rapidement. La variation du DBH est moins
prononcée. La variation de ces deux courbes montre qu'on n'a pas une
relation linéaire entre le DBH et la biomasse.
Figure 23. Comparaison des courbes de la taille
des diamètres et de la biomasse.
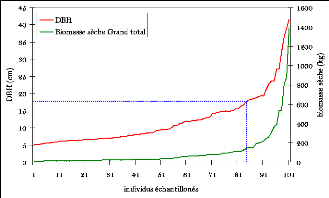
Ainsi le nuage de points de la figure 24 montre une relation non
linéaire de la forme y= bax ou y=
bxa, un modèle de croissance ou de puissance.
Figure 24. Nuage de points du DBH et de la
biomasse (courbe non linéaire)
La figure 24 montre que les plus grands risques de biais sont
situés au niveau des sujets de gros diamètre (>20cm). On
observe sur le nuage de points une plus grande dispersion des valeurs de
biomasse des gros sujets qui traduisent une certaine irrégularité
dans la relation entre celle-ci et le DBH. Ceci démontre
déjà qu'on peut obtenir une bonne prédiction de la
biomasse des sujets petits diamètres (< 20 cm) qui sont dominants
dans les écosystèmes de savane.
II. 2.6.2. Régressions allométriques
développées à partir des données de terrain
Le but des régressions est de déterminer les
valeurs paramétriques d'une fonction pour un bon ajustement de celles-ci
par rapport aux données observées (expérimentales), dans
ce cas le DBH. Les régressions sont alors des outils statistiques
permettant d'établir une relation de dépendance entre deux
variables ; une dite explicative, l'autre appelée dépendante. Le
but de la régression est triple. Elle permet de résumer la
relation existant entre une variable aléatoire dépendante `y' (la
biomasse sèche) et une variable aléatoire explicative `x' (DBH).
Le lien entre les deux variables se résume en une équation (la
régression) et quelques paramètres de précision et
d'optimisation comme le coefficient de détermination (r2), le
coefficient de corrélation (r) et les résidus du
modèle.
La détermination d'un modèle allométrique
serait un exercice simple s'il existait une relation
simple entre le DBH et
la biomasse totale. Dans de tels cas, la relation est linéaire
(y=ax+b)
DBH
et est caractérisée par une pente `a' (le pas de
la croissance) et l'ordonnée à l'origine `b' (la
valeur de
départ sur l'axe des `y'). Le calcul du modèle linéaire
sur les données collectées
offre cependant un coefficient de
détermination peu convaincant parce que la relation n'est
pas linéaire. Alors la relation entre la biomasse et le
diamètre des arbres est une fonction curviligne. C'est ainsi qu'on fait
recours à plusieurs essais de modèles mathématiques pour
ne retenir que celui (ou ceux) qui facilite grandement l'opération
d'ajustement entre les deux variables (Scherrer, 1984).
Le principe de base des régressions non
linéaires consiste à transformer les données pour arriver
à une courbe représentative de la relation entre les deux
variables (ajustement). Ainsi pour une série de données on
cherche le ou les modèles les plus adéquats pour pouvoir
prédire la valeur de la variable dépendante (biomasse) en
utilisant une fonction de la variable indépendante (DBH).
Il existe plusieurs types de régressions statistiques
(voir liste des équations plus bas). Chacune d'entre elles prédit
la variable dépendante en utilisant les paramètres p0 et p1 qui
constituent les paramètres de la régression. Le paramètre
X représente ici le DBH qui est la variable explicative, et `y' la
biomasse qui est la variable expliquée.
Liste des équations de base qui ont été
testées pour élaborer les modèles allométriques.
(1) Linéaire : y= p0 + p1 * X
(2) Logarithmique : y= p0 + p1 *ln (X)
(3) Quadratique : y= p0 + p1 *X+ p2 *X2
(4) Cubique: y= p0 + p1 *X+ p2 *X2+ p3
*X3
(5) «Compound» : y= p0 * p1X
(6) Puissance: y= p0 *X p1
(7) «Growth»: y= exp(p0 + p1 *X)
(8) Exponentiel: y= p0 * e (p1*X)
(9) Polynomial: y= p0 * X+ p1 *X1 + p2 *
X2+ p3 * X3... p0 *Xn+p
Le test de ces régressions a été
effectué avec le logiciel SPSS et le tableau 6 donne les formules, les
coefficients de détermination et la transcription de la formule sur le
logiciel Excel.
Tableau 6. Les modèles calculés et
leur r2 (n = 101 individus)
|
Modèle
|
Formule de la régression (y=)
|
R2
|
Code Excel
|
|
Exponentiel
|
7,15*e(0,151*DBH)
|
0,827
|
=6,89*EXP(0,151*DBH)
|
|
Puissance
|
0,229*DBH(2,237)
|
0,892
|
=0,229*PUISSANCE(DBH;2,237)
|
|
Growth
|
Exp(1,967+0,149*DBH)
|
0,829
|
=EXP(1,9669+0,149689*DBH)
|
|
Quadratique
|
49,84-(10,34*DBH)+
|
0,930
|
=49,84-10,34*DBH+0,89*DBH*DBH
|
|
(0,89*DBH2)
|
|
|
|
Cubique
|
-58,18+13,61DBH-
|
0,936
|
=-58,18+13,61*DBH-0,517*
|
|
0,517DBH2+0,0225DBH3
|
|
DBH*DBH+0,0225*DBH*DBH* DBH
|
|
Compound
|
7,15*1,16DBH
|
0,829
|
=7,15*PUISSANCE(1,16;DBH)
|
|
Polynomial
|
0,0225*DBH3-
|
0,936
|
=0,0225*PUISSANCE(DBH;3)-
|
|
(ordre 3)
|
0,5167*DBH2+13,613*DBH-
|
|
0,5167*PUISSANCE(DBH;2)+13,613*DBH-
|
|
58,18
|
|
58,18
|
|
Linéaire
|
23,48*DBH-178,82
|
0,784
|
= 23,48*DBH-178,82
|
|
Log10 (LN)
|
-556,76+281,17*lnDBH
|
0,547
|
=-556,76+(281,17*LN(DBH))
|
Ces régressions montrent des performances
variées. Nous observons que les coefficients de détermination
(R2) sont très différents expliquant les
différences dans l'ajustement du modèle par rapport aux
données de base. Ainsi pour une bonne approximation de la biomasse
à partir du DBH, nous retenons les régressions de R2
supérieur ou égal à 0,9. Les modèles avec un
R2 élevé conduisent à des résidus
(écarts entre le modèle et les valeurs observés)
très faibles. Les meilleurs modèles sont alors les
régressions quadratique, cubique et polynomial. Les différents
modèles testés sont montrés à la figure 25.
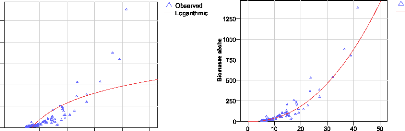
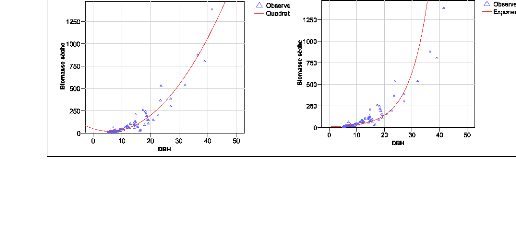
Figure 25. Représentation des courbes des
différentes régressions
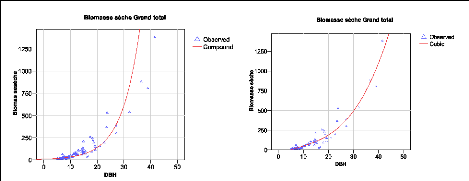
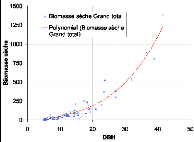
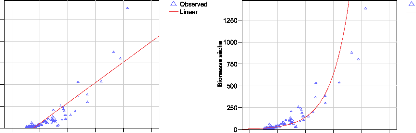

Les modèles montrant une bonne performance ont
amélioré sensiblement les résultats d'estimation de
biomasse à partir du DBH en comparaison avec ceux proposés par la
FAO. La figure 26 montre ces différences.
Figure 26. Comparaison entre les modèles
FAO et les données de terrain (grands décalage surtout par
rapport aux gros diamètres).
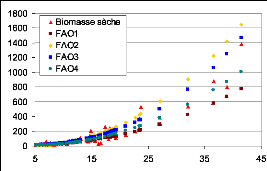
Modèles retenus (moins de décalage par rapport aux
données réelles)
DB
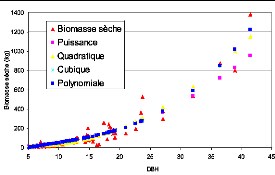
En termes de décalage (somme des résidus) entre les
estimateurs les plus performants et les mesures de biomasse, le tableau 7
montre que le polynomial donne de meilleurs résultats.
Tableau 7. Comparaison entre la biomasse
sèche (kg) et les estimations des modèles retenus.
|
Biomasse sèche
|
Puissance
|
Quadratique
|
Cubique
|
Polynomial
|
|
Total biomasse
|
10248,21 (kg mesurés)
|
9228,82
|
10194,92
|
10226,56
|
10236,12
|
|
A biomasse (résidus)
|
-1019,39
|
-53,29
|
-21,65
|
-12,09
|
|
Ordre (meilleure estimation)
|
4ème
|
3ème
|
2ème
|
1er
|
La superposition quasi-nette des modèles (figure 27)
montre que les écarts entre ces derniers sont minimes, mais certains
comme la régression polynomiale minimise mieux les écarts par
rapport aux données observées.
sa
Figure 27. Variations des mesures de biomasse
par les différents des modèles qui ont un bon coefficient de
détermination.
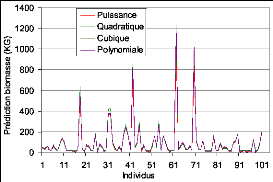
Les modèles retenus sont les plus adaptés du
fait qu'ils présentent moins de biais pour les gros diamètres et
prédisent bien les petits sujets. Dans la plupart des cas, la biomasse
des gros diamètres sont légèrement sous estimés.
Ceci pose moins de problèmes en terme de calcul d'additionalité
que dans le cas de la surestimation pour lequel on prétend comptabiliser
plus de carbone que la réalité.
Par contre, les modèles de la FAO mènent soit
à une forte sous-estimation ou à une surestimation de la biomasse
totale. Ces erreurs peuvent grandement influencer la quantification et la
caractérisation de la dynamique du carbone dans le cadre des projets
MDP. Le tableau 8 donne les résidus issus de l'application des
modèles de la FAO.
Tableau 8. Comparaison entre la biomasse
sèche (kg) et les estimations basées sur les modèles de la
FAO.
|
que Biomasse sèche
|
FAO1
|
FAO2
|
FAO3
|
FAO4
|
|
Total biomasse échantillon
|
ale
10248,21 (kg
mesurés)
|
7050,66
|
13438,96
|
11744,51
|
8424,61
|
|
? biomasse (résidus)
|
- 3197,55
|
3190,75
|
1496,3
|
-1823,6
|
|
Appréciation
|
Forte sous
estimation
|
Forte
surestimation
|
Surestimation
|
Sous
estimation
|
Les modèles proposés par Brown (1997)
présentent en général un grand décalage par 1 4 1 9
1
rapport aux grands sujets comme le montre la figure 28. Alors que
les modèles développés ndividus
présentent une différence aléatoire minime
par rapport aux données observées (figure 29).
Figure 28. Comparaison des résidus des
mesures avec les estimations de la FAO
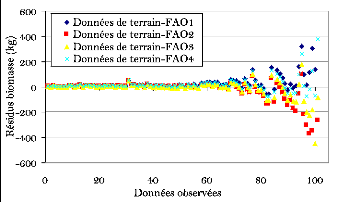
Figure 29. Comparaison des résidus des
mesures avec les modèles in situ.
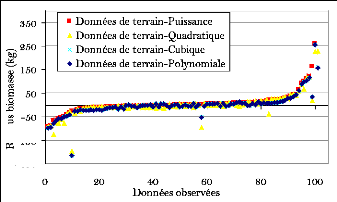
es
m
es de te
de te
L'élaboration de modèles allométriques
propres aux écosystèmes étudiés permet ainsi
Donnée de terrainQuadratique e q
d'améliorer significativement l'estimation de la biomasse
utilisant les données d'inventaire de Dées d tiPlil
la végétation. Les différences
significatives avec les modèles génériques
proposées par la 0FAO, montre qu'il est risqué d'appliquer des
modèles allométriques développés en dehors
20 40 60 80 100
des écosystèmes étudiés. Les
résultats obtenus dans ce travail peuvent permettre d'ajuster
Données observées
les estimations de carbone des projets MDP et les inventaires
de GES effectués
périodiquement pour la CCNUCC. L'absence de
régressions allométriques natives a
constitué une
source de biais pour l'estimation de la biomasse des arbres. Les nouveaux
4 0
modèles développés dans ce travail
pourraient aider à combler cette limite méthodologique.
Il faut toutefois noter que même si les modèles
obtenus permettent de mieux estimer la biomasse, il convient d'analyser leur
performance et leurs limites.
II.2.7. Evaluation des modèles retenus
L'évaluation des modèles est un exercice
délicat, dans la mesure où il n'y pas de méthode de
procédure standard. Les étapes sont nombreuses et connexes et ne
peuvent être déconnectées du processus de mise en place de
modèles forestiers (Vanclay et Skovsgaard, 1996). Un modèle doit
être évalué à travers des tests quantitatifs qui
cherchent à prouver qu'il est suffisamment bon pour prédire un
facteur donné. L'évaluation d'un modèle consiste alors
à démontrer statistiquement ou empiriquement le caractère
raisonnable des valeurs estimées à partir d'une
régression. Il existe cependant quelques procédures de base
consistant à examiner la structure du modèle pour identifier
d'éventuels dysfonctionnements. D'autres approches passent par la
collecte de données supplémentaires non biaisées
(données de référence) à des fins de comparaison
avec la prédiction du modèle en question. Ces approches ont
toutes des limites et ceci fait dire à Vanclay et Skovsgaard (1996) que
la qualité ou la performance d'un modèle ne peut être
évaluée qu'en termes relatifs et sa capacité de
prédiction est très révocable selon les cas. Toutefois,
l'évaluation d'un modèle est un processus continu tout au long de
son élaboration. Ainsi, pour évaluer un modèle il faut
vérifier l'adéquation des équations utilisées ; la
précision de ses constantes d'ajustement, la précision de la
prédiction ; et voir si le modèle satisfait aux exigences de
précision statistiques.
En outre, il faut tenir compte des considérations
qualitatives qui permettent de se prononcer sur les possibilités
d'améliorer le modèle pour les travaux futurs ou les efforts de
révision du modèle proposé.
Le souci de précision a conduit à la prise en
compte d'un certain nombre de précautions sur le choix des
échantillons (distribution spatiale, et répartition dans les
différentes classes de diamètre). Ce souci a été
maintenu depuis le début du travail par l'examen de la forme de la
courbe de la variable indépendante (DBH) et celle de la variable
dépendante (Biomasse) pour avoir une idée sur la forme que va
prendre le modèle. Des analyses complémentaires ont permis de
compléter les tests de précision.
Les tests ainsi effectués permettent de mieux
opérer le choix sur un modèle à partir de son coefficient
de détermination R2. Le coefficient de détermination,
aussi appelé coefficient de corrélation multiple, mesure la
proportion de la variation de y expliquée par la variation de x
(Scherrer, 1984). Ce coefficient est calculé de la façon suivante
:
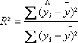
Si tous les points sont alignés, la dispersion
expliquée par le modèle est égale à la dispersion
totale et on a un modèle parfait avec un R2 égal
à 1. Ainsi, théoriquement, les meilleurs modèles sont ceux
dont le R2 est proche de l'unité. En outre, il existe
d'autres tests statistiques très éprouvés pour
compléter cette évaluation des modèles, il s'agit de
l'analyse de la variance (ANalysis Of VAriance : ANOVA). L'analyse de variance
ANOVA permet de vérifier la distribution autour de la pente de la
régression. Il s'agit en termes simples d'une comparaison des moyennes
entre la série expérimentale et les données
prédites.
- Tests statistiques par comparaison de
moyennes
Pour la comparaison entre la biomasse obtenue par mesure
directe et celle issue des différents modèles, nous avons
procédé à des tests statistiques pour évaluer la
performance de chaque estimateur de biomasse. Il existe plusieurs tests
statistiques. Les plus utilisées sont ceux qui procèdent par
comparaison de moyennes. Pour des échantillons réduits de
variances égales, on peut utiliser des tests simples comme le T-Test
qu'on peut interfacer avec les outils d'analyse de « Excel-Macros
Complémentaires ». T-Test est souvent appliqué sur de petits
échantillons de moins de 30 observations. Ce test permet d'effectuer un
T-Test de Student sur deux échantillons. Ce test, appliqué sur
deux échantillons vérifie l'égalité des moyennes de
populations de chaque échantillon. Ces tests utilisent pour ce faire des
hypothèses différentes : les variances de population sont
égales ; les variances de population ne sont pas égales ; les
deux échantillons représentent, avant et après traitement,
des observations sur les mêmes sujets.
L'application du T-Test suppose malheureusement que les deux
séries de données proviennent de distributions aux variances
identiques, ce qui n'est pas toujours garantie. On préfère
l'ANOVA au T-Test dans plusieurs situations. L'analyse de la variance est une
technique statistique permettant de comparer les moyennes de deux populations
ou plus. L'analyse de la variance n'est pas une méthode qui permet
d'étudier les différences de variances entre populations, mais
une méthode pour étudier les différences de moyenne entre
populations, pour ainsi caractériser les sources de variations sur
l'ensemble des données (Scherrer, 1984). Néanmoins, cette
méthode doit son nom au fait qu'elle utilise des mesures de variance
afin de déterminer le caractère significatif, ou non, des
différences de moyenne mesurées sur les populations (Wikipedia,
2008).
Comme le T-Test, l'ANOVA procède par un test
d'hypothèses : H0 : Ji1 = t2 (les moyennes des deux séries sont
égales)
H1 : ji1 ~ t2 (les moyennes de deux séries sont
différentes)
L'analyse de la variance est un procédé qui
permet de calculer la dispersion totale de l'ensemble des données et de
les partager en composantes de différentes sources. On a la dispersion
intergroupe appelée `factorielle' et la dispersion
intragroupe appelée `résiduelle' ou dispersion due aux
erreurs (Scherrer, 1984). Cette dernière est souvent utilisée
dans l'évaluation de la performance des régressions. Le terme
dispersion revoit alors à la somme des carrés des écarts
entre une série de valeurs et leur moyenne.
Ainsi, la dispersion totale est représentée dans
cette analyse par la somme des carrés des écarts à la
moyenne générale de l'ensemble des données recueillies
sans tenir compte de l'échantillon d'appartenance.
La dispersion à l'intérieur des groupes
(échantillons) exprime les fluctuations
d'échantillonnage,
représentées par les écarts entre les valeurs
individuelles et la moyenne de
leur propre échantillon (xij-tj). Pour
un groupe la dispersion est égale à la somme de ces
écarts élevée au carré [?
(xij-tj)2] pour tout nij, i=1 (Scherrer, 1984).
La dispersion entre groupe exprime l'écart quadratique
entre la moyenne d'un échantillon et
la moyenne générale (tj-Ji)2. La somme
des dispersions intra et intergroupe donne la dispersion totale.
L'ANOVA permet surtout de faire des tests de comparaison pour
accepter ou rejeter l'hypothèse principale (H0 : ji1 = t2) ou
l'hypothèse secondaire (H1 : Ji1 ~ t2). Le facteur F (rapport entre la
variance intergroupe et variance intragroupe) permet de comparer les deux
variances et permet de décider quelle hypothèse retenir.
1. Si la valeur de F est inférieure à la Valeur
Critique pour F (tableau 9), l'hypothèse principale est acceptée
mais on ignore le risque d'erreur (erreur de type J3 ou Type II
error).
2. Si la valeur calculée de F est supérieure ou
égale à la valeur critique pour F, l'hypothèse principale
est rejetée et le risque d'erreur présente un niveau connu
(erreur de type a ou Type I error).
Le seuil de signification utilisé dans cette analyse est
de 5%, c'est-à-dire que a = 0,05. Les résultats du test ANOVA
sont précisés au tableau 9.
Tableau 9. ANOVA pour les différents
modèles testés
|
Modèles
|
Source des
variations
|
Somme des
carrés
|
Degré de
liberté
|
Moyenne
des carrés
|
Valeur
calculée
pour F
|
Probabi-
lité
|
Valeur
critique pour
F
|
|
Puissance
|
Entre Groupes
|
5144,28
|
1
|
5144,28
|
0,163903
|
0,68
|
3,888374535
|
|
A l'intérieur
des groupes
|
6277223,97
|
200
|
31386,12
|
|
|
|
|
|
|
|
|
|
|
|
|
Growth
|
Entre Groupes
|
64055,89
|
1
|
64055,89
|
0,514684
|
0,47
|
3,888374535
|
|
A l'intérieur
des groupes
|
24891302,61
|
200
|
124456,51
|
|
|
|
|
|
|
|
|
|
|
|
|
Quadratique
|
Entre Groupes
|
14,057
|
1
|
14,05
|
0,000384
|
0,98
|
3,888374535
|
|
A l'intérieur
des groupes
|
7313779,43
|
200
|
36568,89
|
|
|
|
|
|
|
|
|
|
|
|
|
Cubique
|
Entre Groupes
|
2,32
|
1
|
2,32
|
0,000063
|
0,99
|
3,888374535
|
|
A l'intérieur
des groupes
|
7354503,26
|
200
|
36772,51
|
|
|
|
|
|
|
|
|
|
|
|
|
Exponentiel
|
Entre Groupes
|
85867,10
|
1
|
85867,10
|
0,630468
|
0,42
|
3,888374535
|
|
A l'intérieur
des groupes
|
27239139,25
|
200
|
136195,69
|
|
|
|
|
|
|
|
|
|
|
|
|
Compound
|
Entre Groupes
|
46888,04
|
1
|
46888,04
|
0,410232
|
0,52
|
3,888374535
|
|
A l'intérieur
des groupes
|
22859255,52
|
200
|
114296,27
|
|
|
|
|
|
|
|
|
|
|
|
|
Polynomial
|
Entre Groupes
|
0,72
|
1
|
0,72
|
0,000019
|
0,99
|
3,888374535
|
|
A l'intérieur
des groupes
|
7358519,95
|
200
|
36792,60
|
|
|
|
|
|
|
|
|
|
|
|
|
Logarithmique
|
Entre Groupes
|
74,67
|
1
|
74,67
|
0,002544
|
0,96
|
3,888374535
|
|
A l'intérieur
des groupes
|
5869947,57
|
200
|
29349,73
|
|
|
|
|
|
|
|
|
|
|
|
|
Linéaire
|
Entre Groupes
|
0,00056
|
1
|
0,00056
|
1,66005E
-08
|
0,99
|
3,888374535
|
|
A l'intérieur
des groupes
|
6792093,76
|
200
|
33960,47
|
|
|
|
Les résultats de l'ANOVA montrent et confirment que les
meilleurs modèles sont le Quadratique, le Cubique et le Polynomial avec
des valeurs de F très faibles, ce qui traduit la faiblesse des
écarts entre les valeurs de ces modèles et les données
observées.
Il faut ainsi noter que l'utilisation du coefficient de
détermination ne suffit pas pour estimer la précision d'un
modèle. La comparaison des moyennes, à travers des tests d'ANOVA,
entre les valeurs projetées et des données réelles permet
d'affiner le choix des modèles performants.
Toutefois, si le cubique et le polynomial sont des fonctions
cubiques et se ressemblent du point de vue des résultats, le
modèle quadratique fait une estimation différenciée en
fonction de la taille des diamètres. On constate pour les tout-petits
diamètres (5-7 cm) et les grands diamètres (> 17 cm), que le
cubique et le polynomial sous estiment légèrement la biomasse,
alors qu'ils rendent plus de biomasse pour les diamètres moyens (7-17
cm), figure 30. Ces différences entre les modèles peuvent donner
des résultats similaires sur le total de biomasse si on a une
distribution bien équilibrée des individus dans les
différentes classes de diamètres. Par contre si une
catégorie de classe de diamètre prédomine, les deux
groupes de modèles peuvent rendre des estimations relativement
différentes. Ces différences constatées peuvent être
tout de même exploitées pour fixer des limites inférieures
et supérieures d'estimation de biomasse pour chaque
écosystème. Elles peuvent aussi permettre de faire un choix
approprié d'un modèle en fonction des caractéristiques
structurelles d'une formation pour laquelle on veut estimer la biomasse
totale.
Figure 30. Différence entre les
modèles quadratique et les fonctions cubiques
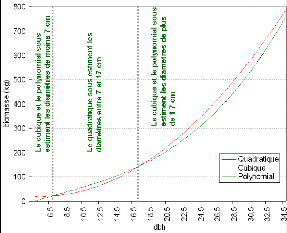
Sur le plan de la logique mathématique, le quadratique
est une fonction très simple comparée au cubique et au
polynomial, mais ces dernières expriment mieux la logique de croissance
d'un arbre. Le tronc d'un arbre augmente en volume et suit une logique cubique.
Cette logique est liée à une croissance de la surface du cylindre
du fût (2 dimensions avec Surface= ð*r2) mais aussi une
croissance verticale ce qui fait une troisième dimension expliquant la
logique cubique (figure 31).
Figure 31. Schéma de la logique de
croissance des diamètres de tronc.


On se gardera de recommander à cette étape un
modèle au détriment des autres car nous estimons que chacun
présente des avantages et des limites. Il s'agira de tenir compte de la
structure du peuplement pour opérer un choix ou procéder à
une combinaison des deux types de modèles ou même faire la moyenne
des résultats issus de ces modèles
II.2.8. Comparaison entre les modèles `toutes
espèces' et les modèles pour une espèce : exemple de
Combretum glutinosum
La série de données de biomasse collectée
sur le terrain a permis de développer des régressions
allométriques pour l'espèce Combretum glutinosum, qui
est la seule à avoir un nombre d'individus statistiquement significatif
(n=39) pour générer des régressions avec les
quantités de biomasse des différentes tailles de diamètre.
En procédant de la même façon qu'avec la base globale, les
régressions les plus précises sont encore le Cubique, le
Quadratique et le Polynomial.
Tableau 10. Equations allométriques pour
Combretum glutinosum
|
Modèle
|
Equation
|
R2
|
N
|
|
Cubique
|
y =37, 91 -
12,287*DBH+1,494*DBH2-0,0194*DBH3
|
0,908
|
39
|
|
Quadratique
|
y= -25,505+2,917*DBH+0,47*DBH2
|
0,905
|
39
|
|
Polynomial
|
y=
-0,02*DBH3+1,518*DBH2-12,467*DBH+38,681
|
0,908
|
39
|
Rappelons que le cubique et le polynomial sont deux
modèles très proches, on peut en définitive utiliser l'un
ou l'autre. Les trois modèles choisis donnent une estimation
relativement bonne de la biomasse totale (tableau 11). Les marges d'erreurs
entre les modèles génériques et les modèles
spécifiques varient entre 7,5% à 9,5% ce qui peut avoir des
implications importantes dans la caractérisation de la biomasse totale.
Si donc on veut évaluer une formation largement dominée par une
espèce, autant établir un modèle spécifique
plutôt que d'utiliser les modèles génériques plus
aptes quand il s'agit de mosaïques de formations végétales
avec plusieurs espèces.
Tableau 11. Comparaison des résultats des
estimations entre les modèles toutes-espèces et les
modèles spécifiques.
|
Biomasse mesurée
|
Cubique
Comglu total sp
|
Quadratique
Comglu total sp
|
Polynomial
Comglu total sp
|
|
3203,973
|
3165,26 2925,86
|
3158,14 2862,52
|
3204,89 2929,01
|
|
Différence en kg
|
239,4
|
295,62
|
275,88
|
|
Différence en %
|
7,56
|
9,36
|
8,6
|
Comglu= Combretum glutinosum
La différence entre les modèles et les
données réelles sont donc plus importantes pour les
modèles génériques que pour les modèles
spécifiques (tableau 11). Les modèles spécifiques sont
donc plus proches de la réalité que les modèles
intégrant toutes les espèces. Ces différences sont
importantes à considérer quand on veut estimer la biomasse d'une
formation largement dominée par une espèce. Dans ces cas de
figures les modèles intégrant une large gamme d'espèces
peuvent être inappropriés. Les travaux de Nelson et al.
(1999) montrent que l'erreur est toujours plus grande entre un modèle
intégrant une large variété d'espèces et un
modèle défini uniquement pour une espèce.
D'une façon générale les modèles
spécifiques de Combretum glutinosum introduisent les
mêmes légères différences entre polynomial et
cubique d'une part et quadratique d'autre part. Ces différences sont
liées à leur forte sensibilité à la taille de
diamètre. Les estimations rendues par les modèles n'ont jamais
dépassé les valeurs réelles de biomasse. Une
légère sous estimation est mieux qu'une surestimation dans
l'évaluation des efforts de séquestration de
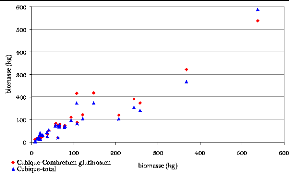
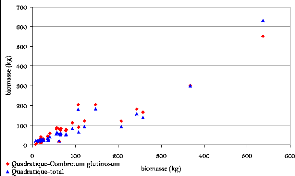
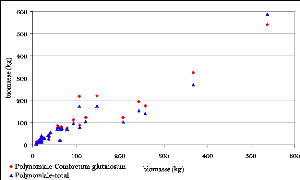
carbone. Les figures 32, 33, et 34, montrent les
différences entre les modèles spécifiques et les
modèles basées sur la totalité des espèces.
Figure 32. Biomasse estimée par les
modèles cubiques
Figure 33. Biomasse estimée par les
modèles Quadratiques
Figure 34. Biomasse estimée par les
modèles Polynomiaux
Ces figures montrent que les modèles sur toutes les
espèces sous estiment les valeurs prédites par les modèles
spécifiques. Les plus grandes sous estimations sont notées pour
les sujets de grands diamètres (> 20 cm).
II.3. Discussions et conclusion
La régression a pour but la prédiction. Elle est
une méthode puissante et utile. Mais il n'y a pas de voie royale en
statistique : toute méthode comporte des difficultés (Rouanet et
Lebaron, 2006). Ces auteurs distinguent une régression à usage
seulement prédictif et la régression à usage
explicatif. Dans la régression prédictive la liste des
variables prédictrices n'est pas limitative, et les liaisons entre ces
variables sont peu gênantes. Dans un schéma explicatif, la
variable à prédire est qualifiée de «variable
à expliquer», et les variables prédictrices sont des
«variables explicatives». Cette dernière approche est plus
délicate et se mène dans un champ de recherche de
causalité très délicat.
La problématique prédictrice est
symétrique. A partir du DBH, on peut prédire la biomasse d'un
arbre, et régresser la biomasse de l'arbre sur le DBH; mais on peut tout
aussi bien, à l'inverse, à partir de la biomasse de l'arbre,
prédire le DBH, et régresser le DBH sur la biomasse de l'arbre
(Rouanet et Lebaron, 2006). Les régressions explicatives sont elles
dissymétriques, le DBH peut servir à prédire la biomasse
mais pas l'inverse. Dans le cadre des régressions allométriques,
on se limite à des régressions prédictives.
La quantification de la biomasse passe par l'utilisation de
modèles qui définissent une relation synthétique entre une
variable dépendante et une ou plusieurs prédicateurs ou variable
indépendante. Les modèles servent à faire de la
prédiction de valeurs sur une variable non observée à
partir de mesures expérimentales réelles. Le volume, la biomasse
et le carbone peuvent être estimés en utilisant des
équations qu'on a générées à partir de
régressions statistiques. Ces régressions permettent d'approcher
la variable dépendante à partir de données
d'entrée. La qualité d'un modèle réside alors dans
son habileté à rapprocher le plus que possible les valeurs
réelles des valeurs prédites. Le modèle doit alors
reproduire les valeurs de la variable dépendante avec le moins de biais
possible (avec le moins d'écart possible).
La première forme d'analyse de la précision d'un
modèle est de comparer les valeurs estimées par le modèle
et les valeurs mesurées. Le calcul de la somme des résidus doit
se rapprocher de zéro pour qu'un modèle puisse être
considéré comme précis. Cela implique que les
résidus individuels (écarts par rapport au modèle) des
observations doivent être faibles.
Des techniques statistiques plus élaborées sont
utilisées pour évaluer la précision d'un modèle.
Parmi ces techniques:
- le t-test de l'hypothèse d'une erreur moyenne de
prédiction égale à zéro;
- les écarts absolus ;
- la distribution normale des erreurs résiduelles des
valeurs prédites ;
- les erreurs de la régression linéaire ;
- l'homogénéité des variances de l'erreur du
modèle par rapport aux données de base.
Les travaux de Vanclay et Skovesgaard (1996) donnent un
aperçu et un cadrage opérationnel pour évaluer la
qualité des modèles forestiers. La précision centrale est
qu'il faut toujours, quand on applique des modèles de volume ou de
biomasse développés pour une zone donnée ou une
espèce, procéder à la vérification de la
qualité du modèle et le tester avant son utilisation. Ces tests
peuvent nécessiter la collecte de quelques données de terrain, ou
l'utilisation de données secondaires qui pourront servir de base de
comparaison. D'après la FAO (2008), ne pas procéder par ces
tests, suppose qu'on estime implicitement que le modèle est
approprié, ce qui n'est pas toujours le cas.
Les erreurs d'un modèle proviennent principalement de
trois sources que sont les erreurs d'échantillonnage, les erreurs de
mesure et les erreurs dues au modèle. On suppose tout de même que
les mesures sur les individus sont relativement précises (limite des
instruments utilisés), mais que la méthode
d'échantillonnage et les modèles entrainent quelques erreurs.
Aucune méthode d'inventaire de la végétation ligneuse
n'est parfaite.
Dans un contexte où les données de biomasse sont
obtenues par voie de collecte sur le terrain (méthode
expérimentale), la variable indépendante qui est le DBH est le
facteur expérimental, dont on étudie les effets sur la variable
dépendante (la biomasse) à l'aide de méthodes classiques
comme l'analyse de variance. Techniquement, la régression n'est autre
que l'analyse de variance pour des facteurs expérimentaux quantitatifs
(Rouanet et Lebaron, 2006). Il faut alors que les facteurs expérimentaux
quantitatifs (DBH et biomasse) soient mesurés avec prudence et
précision sur le terrain.
En plus du coefficient de détermination R2,
des tests de signification des régressions ont été faits
pour comparer les résultats des modèles allométriques aux
quantités de biomasse mesurées. L'inférence statistique
est importante à considérer dans les régressions
allométriques. Avec les données d'observation, la seule situation
où ce cadre d'inférence est opératoire est le sondage
aléatoire. Le choix des espèces pour cette étude, est le
résultat d'un sondage au hasard dans des strates de
végétation et des grappes d'intervalle de diamètres.
Dans plusieurs autres situations, les chercheurs ont
utilisé des modèles complexes intégrant des fonctions
exponentielles ou logarithmiques. Ces fonctions ont pour but de transformer les
données brutes pour améliorer les régressions
subséquentes. Les essais que nous avons pu faire ne nous ont pas
encouragé à aller dans cette direction, parce que d'une part, on
ne connaît pas l'erreur additionnelle introduite avec la transformation
des données et d'autres part, les résultats n'ont pas
été meilleurs que ceux obtenus par des régressions
simples.
Ce chapitre a permis de mettre au point 3 modèles
allométriques performants pour les écosystèmes de savane
soudanienne et soudano-guinéenne. Ces résultats pourront servir
à ajuster les calculs de biomasse et de carbone dans ces
écosystèmes au lieu d'emprunter des modèles conçus
pour d'autres types de formations végétales. Les résultats
révèlent qu'il est possible de mieux caractériser la
biomasse s'il s'agit de modèles spécifiques qui portent sur une
espèce comparés à des estimateurs globaux. Les
modèles spécifiques sont meilleurs quand on travaille dans des
écosystèmes dominés par une seule espèce comme
c'est le cas dans certaines formations de savane avec la dominance de
Combretum glutinosum.
Bibliographie
Awaya, Y., Tsuyuki, S., Kodani, E., Takao G., 2004. Potential
of Woody Carbon Stock Estimation Using High Spatial Resolution Imagery: A Case
Study of Spruce Stands. In: M-Shiyomi (Editor), Global Environmental Change in
the Ocean and on Land. TERRAPUB, pp. 425-440.
Brown S., 1997. Estimating Biomass and Biomass Change of
Tropical Forests: a Primer. (FAO Forestry Paper -134). FAO - Food and
Agriculture Organization of the United Nations, Roma, 48 p.
Brown S. 2002. Measuring carbon in forests: current status and
future challenges. Environmental Pollution, 116:363-372.
Brown, S., Pearson, T., Slaymaker, D., Ambagis, S., Moore, N.,
Novelo, D., Sabido W. 2005. Creating a virtual tropical forest from
three-dimensional aerial imagery to estimate carbon stocks. Ecological
Applications, 15 (3):1083-1095.
Brown, S., Sathaye, J., Cannell, M., Kauppi P.E., 1996.
Management of forests for mitigation of greenhouse gas emissions. In: D.
R.T.Watson, et al. (Editor), Climate change 1995: impacts, adaptations
and mitigation of climate change: scientific analyses. Contribution du
2ième Groupe de travail au deuxième rapport
d'évaluation du GIEC. Cambridge University Press, pp. 773-798.
CCNUCC, 1997. Protocole de Kyoto. UNFCCC, p. 24 p.
CGE, 2005. Handbook on land-use change and forestry sector,
UNFCCC.
Clark, D.B., Clark D.A. 2000. Landscape-scale variation in forest
structure and biomass in a tropical rain forest. Forest Ecology and Management,
137:185-198.
Fleurant, C., Duchesne, J., Raimbault P. 2004. An allometric
model for trees. Journal of Theoritical Biology (227):137-147.
IPCC, 2003. Good Practice Guidance for Land Use, Land-Use
Change and Forestry. Penman J., Kruger D., Galbally I., Hiraishi T., Nyenzi B.,
Emmanuel S., Buendia L., Hoppaus R., Martinsen T., Meijer J., Miwa K., and
Tanabe K. (Eds). Institute for Global Environmental Strategies, Kanagawa,
Japan.
Keller, M., Palace, M., Hurtt G. 2001. Biomass estimation in
the Tapajos National Forest, Brazil. Examination of sampling and allometric
uncertainties. Forest Ecology and Management, 154:371-382.
Lehtonen A., 2005. Carbon stocks and flows in forest
ecosystems based on forest inventory data No. Dissertationes Forestales 11,
Finnish Forest Research Institute. Vantaa Research Center, Helsinki.
Lufafa, A., Bolte, J., Wright, D., Khouma, M., Diedhiou, I.,
Dick, R.P., Kizito, F., Dossa, E., Noller J.S. 2008. Regional carbon stocks and
dynamics in native woody shrub communities of Senegal's Peanut Basin.
Agriculture, Ecosystems and Environment (In press).
Mbow C., 2005. Dynamic of carbon stock in woody vegetation of
savanna ecosystems in Senegal. The Land cover change perspective. Centre for
Global Change and Earth Observations, Lansing. 25p.
Moura-Costa, P., Stuart M., 1999. Issues related to monitoring,
verification and certification of forestry-based carbon offset projects. In:
W.f.A. Conference, et al. (Editor). 18 p.
Nelson, B.W., Mesquitaa, R., Pereirab, J.L.G., S.G., A.d.S.,
Batistad, G.T., Couto L.B. 1999. Allometric regressions for improved estimate
of secondary forest biomass in the central Amazon. Forest Ecology and
Management, 117 149-167.
Pearson, T., Walker, S., Brown S., 2005. Sourcebook for Land Use,
Land-Use Change and Forestry Projects, Winrock International, Arlington. 64
p.
Proisy C., 1999. Apport des données radar à
synthèse d'ouverture pour l'étude de la dynamique des
écosystèmes forestiers, Université Paul Sabatier,
Toulouse, 103 p.
Rondeux J., 1993. La mesure des arbres et des peuplements
forestiers. Les Presses Agronomiques de Gembloux, Gembloux, 521 p.
Rouanet, H., Lebaron F., 2006. La preuve statistique: examen
critique de la régression. Séminaire "Qu'est ce que faire
preuve?" CNRS, Centre Universitaire de recherches sur l'Action Publique et
Politique, Epidémiologie et Sciences Sociales (CURAPP). 20 p.
Sambou B., 2004. Evaluation de l'état, de la dynamique
et des tendances évolutives de la flore et de la
végétation ligneuse dans les domaines soudanien et
sub-guinéen au Sénégal. Doctorat d'Etat, Université
Cheikh Anta Diop de Dakar (UCAD), Dakar. 231 p.
San Jose, J.J., Montes, R.A., Farinas, M.R. 1998. Carbon
stocks and fluxes in a temporal scaling from a savanna to a semi-deciduous
forest. Forest Ecology and Management, 105:251-262.
Scherrer B., 1984. Biostatistique. Gaëtan Morin,
Montréal, Paris, 850 p.
St-Onge, B., Jumelet, J., Cobello, M., Véga C. 2004.
Measuring individual tree height using a combination of stereophotogrammetry
and LIDAR. Canadian Journal of Forest Research (34):2122-2130.
Thenkabail, P.S., Stucky, N., Griscom, B.W., Ashton, M.S.,
Diels, J., Meer, V.D., B., Enclona, E. 2002. Biomass estimations and carbon
Stock calculations in the oil palm plantations of African derived savannas
using IKONOS data. Rapport, Center for Earth Observation (CEO), Yale
University. 58p.
Vanclay, J.K., Skovsgaard J.P. 1996. Evaluating forest growth
models. Ecological Modelling (98 ): 1-12.
Wikipedia, 2008. Wikipédia, l'encyclopédie libre
:
http://fr.wikipedia.org/wiki/Accueil.
Woomer P.L., 2003. Field and laboratory guidelines for carbon
characteristisation in African Drylands. printed doc, SACRED Africa. 20 p.
Woomer, P.L., Touré, A., Sall M. 2004. Carbon stocks in
Senegal's Sahel Transition Zone. Journal of Arid Environments (59):499-510.
Zianis, D., Mencuccini M. 2004. On simplifying allometric
analyses of forest biomass. Forest Ecology and Management, 187:311-332.
Article II. CHAPITRE III :
Article III. Biomasse et carbone des
savanes soudaniennes et soudano-
guinéennes étudiées
Ce chapitre présente les méthodes
d'évaluation existantes pour l'estimation de la biomasse des formations
végétales. Leur approfondissement nous a permis d'élaborer
une méthode de conversion des données d'inventaire forestier en
biomasse utilisant les régressions allométriques
développées dans le Chapitre 2. Les résultats montrent que
la distribution spatiale et la dynamique temporelle du carbone sont sous
l'influence de la structure des classes de diamètre et de la vitesse de
croissance des arbres. De telles analyses ont alors permis d'évaluer le
taux et le potentiel de séquestration de carbone des savanes
soudaniennes et soudano-guinéennes en partant des données
d'inventaire des Forêts Classées que sont Patako, Wélor,
Ouli, Bala, Kantora et Mampaye.
Section 3.01 III.1. Estimation de la biomasse dans
les
écosystèmes de savane
L'utilisation des modèles allométriques pour
estimer la biomasse est un exercice central dans l'estimation du carbone
ligneux des formations végétales. Les modèles
allométriques sont construites sur la base de méthodes
destructives, mais une fois que le modèle est obtenu pour un
écosystème donné on peut procéder à des
estimations non destructives de la biomasse et donc du carbone pour des
formations végétales similaires. Pour l'ensemble des
méthodes approuvées (Approaved Afforestation, Reforestation
Methods) en foresterie dans le cadre des MDP, des modèles
allométriques sont suggérés pour l'estimation de la
biomasse et de sa dynamique. Le fait d'en créer pour les
écosystèmes soudaniens et soudano-guinéens du
Sénégal devrait donner plus de précision dans la
caractérisation de la biomasse des zones d'intervention des projets des
forêts aménagées ou non. A défaut des modèles
allométriques, d'autres approches non destructives (indirectes) ont
été testées. Les données satellitaires sont de plus
en plus utilisées pour estimer la biomasse des
écosystèmes. Les techniques utilisées peuvent être
basées sur l'utilisation des données sensibles au volume de
biomasse (données RADAR par exemple) ou sur des corrélations
entre des indices spectraux comme le NDVI ou autres indices de
végétation avec la biomasse.
L'intérêt suscité par les données
satellitaires a incité l'ESA (European Spatial Agency) à
envisager un capteur RADAR sensible à la biomasse, dans le cadre des
séries de satellites appelées Earth Explorer. Dans la
seconde phase de ce programme, ESA envisage de mettre en place BIOMASS Explorer
qui doit utiliser la bande P-SAR pour le suivi des forêts à
l'échelle du globe. Cette bande permettra de développer des
modèles pour mesurer la biomasse des forêts, l'ampleur du
déboisement, mais aussi les stocks et flux de carbone (
http://sci.esa.int/,
accédé en aoflt 2008). En attendant d'avoir des données
plus précises, les estimations à partir de satellites
nécessitent un effort énorme de validation (biomasse
mesurée in situ) pour s'assurer de la qualité des
produits. Les travaux de plusieurs auteurs (Brown, 1995; Diouf et Lambin,
2000; Coops et Waring., 2001; Seaquist et al., 2003; Bronge,
2004; Fensholt, 2004; St-Onge et al., 2004; Brown et al.,
2005; Fensholt et al., 2006) ont porté sur l'estimation de la
productivité des écosystèmes ou des mesures de biomasse
basées sur l'utilisation des techniques spatiales
étalonnées avec des données de terrain. Les informations
obtenues sont très utiles par le fait qu'elles contribuent à
renseigner des zones caractérisées par une quasi-absence de
données sur la végétation, tout en offrant une perception
spatio-temporelle. Ces techniques sont cependant mises en ~uvre avec des
précisions variables et nécessitent pour la plupart des cas un
plus grand effort de validation.
Plusieurs sources de données satellites ont permis
d'obtenir des produits en rapport avec l'état de la
végétation (produits MODIS, MERIS, SPOT-VEGETATION). A l'heure
actuelle il existe peu de preuves sur l'efficacité de ces mesures pour
la quantification de la biomasse ligneuse à l'échelle locale. Il
faudrait par conséquent éviter, avec l'état actuel des
produits spatiaux, de considérer que les seules données
satellitaires peuvent permettre d'étudier les stocks de carbone et leur
dynamique. En outre, même si la télédétection peut
aider à détecter les changements de couverture du sol, les
techniques utilisées ne permettent pas pour le moment d'estimer la
biomasse en dessous de la couverture des arbres (Skutsch, 2005), ce qui
constitue une limite si les besoins d'informations portent sur la biomasse
totale.
Les chercheurs se sont longtemps consacrés à la
caractérisation de la biomasse pour estimer la production et la
productivité des écosystèmes. La biomasse totale par
unité de surface était un bon indicateur de l'état des
formations végétales. L'évaluation du capital ligneux des
forêts, est une nécessite pour tout plan d'aménagement
forestier visant une exploitation rationnelle des ressources
végétales.
La situation actuelle dressée dans le rapport sur
l'Evaluation des Ecosystèmes pour le Millénaire ou Millennium
Ecosystem Assessment (MEA, 2005), montre un tableau inquiétant de
la dynamique de dégradation accélérée des
écosystèmes terrestres pendant le dernier siècle avec
d'ailleurs une tendance à la dégradation qui se maintient.
Toujours dans le même rapport on note que `les effets néfastes
de la dégradation des services écosystémiques (la baisse
persistante de la capacité d'un écosystème à
procurer des services) sont subis de manière disproportionnée par
les pauvres, et contribuent à l'aggravation des disparités
croissantes entre les communautés et constituent parfois le facteur
principal de la pauvreté et des conflits sociaux'.
La dégradation des services d'origine
écosystémique est déjà une barrière de
taille à l'atteinte des Objectifs du Millénaire pour le
Développement (OMD) au moins pour ses objectifs 1 (réduire
l'extrême pauvreté et la faim) et 7 (assurer un environnement
durable).
Actuellement, l'enjeu relève à la fois des
besoins de développement durable comme l'a souligné le rapport
sur l'Evaluation des Ecosystèmes du Millénaire (Millenium
Ecosystem Assessement) mais avec les changements climatiques, la dimension
`puits' et `sources' de carbone place les formations forestières au
centre des facteurs de dynamiques pouvant être un problème si les
forêts sont mal gérées, mais une solution si on parvient
à reconstituer les écosystèmes dégradés. Le
premier besoin d'information pour supporter ce type d'analyse et les
décisions subséquentes, est de savoir ce dont nos forêts
disposent en termes de biomasse
pour la satisfaction durable des besoins des populations. Si
les formations forestières sont relativement bien connues depuis
plusieurs années dans la plupart des pays développés
dotés de données d'inventaires nationaux, un pays comme le
Sénégal a besoin de fournir des indications claires sur le
potentiel des formations végétales pour servir à la fois
de base de travail pour les inventaires des GES, mais aussi de
référence pour les projets MDP foresterie. De tels travaux ont
été conduits au Sénégal par Sambou (2004), le CSE
(veille environnemental portant sur le suivi de la biomasse), ou des projets de
la Direction des Eaux et Forêts, Chasse et de la Conservation des Sols
comme le PROGEDE. Les données sont collectées avec des
méthodes différentes et des objectifs variés. Le travail
de Sambou (2004), est basée sur une approche simple mais rigoureuse qui
permet une utilisation des données d'inventaire pour une estimation
appropriée de la biomasse ligneuse.
L'objectif de cette partie de ce travail est d'utiliser des
données d'inventaire de 6 Forêts Classées des domaines
soudaniens et soudano-guinéens, pour estimer les quantités de
biomasse des différentes strates (types de végétation) qui
ont été inventoriées. Ces estimations permettront de
calculer les quantités de carbone par ha et d'analyser les
différences structurelles entre les formations en question en termes de
stocks de carbone de la végétation ligneuse et leur dynamique.
Section 3.02 III.2. Cadrage conceptuel et
méthode de travail
La biomasse végétale exprime la masse totale de
matière organique des plantes exprimée en tonnes de
matières sèches par unité de surface. On peut l'estimer
pour la végétation sur pied en y incluant les herbacées et
la régénération naturelle, mais on peut aussi se limiter
à la végétation ligneuse affranchie (> 5 cm de
diamètre8). Ce travail porte essentiellement
sur la biomasse ligneuse sur pied pour les individus de plus de 5 cm de
diamètre.
La biomasse est souvent proportionnelle au volume du bois,
à la surface terrière, au diamètre du tronc ou la hauteur
des individus. La quantité de biomasse produite varie en fonction de
l'espèce et des conditions de croissance. En général on
obtient la biomasse totale de l'individu en mesurant (ou en estimant) les
différentes parties que sont le tronc, les branches et les feuilles.
Dans la plupart des études actuelles on se limite à la biomasse
sur pied, sans prendre en compte la biomasse racinaire difficile à
mesurer. Par ailleurs, ces études sont disponibles seulement pour
quelques espèces et quelques rares formations végétales,
avec une précision relativement faible (FAO, 2008).
8 Certaines plantes peuvent avoir des tailles de diamètres
inférieures à 5 cm alors qu'ils sont de vieux sujets. Les
espèces qui ont cette particularité n'apparaissent pas sur nos
données.
Dans cette étude, la biomasse racinaire n'est pas prise
en compte, car dans des mosaïques de savane il est très difficile
d'une part d'extraire toute la biomasse racinaire à cause de la
présence de sols très souvent cuirassées ; d'autre part du
fait de l'enchevêtrement des réseaux de racines appartenant
à différents individus. Il faut ajouter à ces contraintes,
les difficultés matérielles et logistiques permettant de mener de
tels travaux sur le terrain.
L'estimation des stocks de carbone dans les formations
forestières requièrent par conséquent l'existence ou la
collecte de données d'inventaire. L'estimation du taux de carbone au
niveau des écosystèmes est un exercice nécessaire pour
mieux comprendre le cycle global du carbone et ses impacts sur les gaz à
effet de serre. La Convention Cadre des Nations Unies sur les Changements
Climatiques encourage les pays à procéder à des
estimations de bilan de carbone (inventaires nationaux). Pour les pays en voie
de développement, cet exercice est confronté à de
nombreuses difficultés, liées essentiellement à l'absence
de données fiables et systématisées, et à la
rareté d'une expertise qualifiée pour procéder aux
évaluations des flux de carbone dans tous les secteurs ciblés par
la convention. Dans le domaine de la foresterie les principales contraintes
exposées sont le manque d'information sur la productivité des
forêts, le manque de données d'inventaire de la
végétation, et les données limitées sur les pertes
de biomasse (exploitation forestière, feux de brousse, etc.).
Ainsi, plusieurs équations (des régressions
statistiques) ont été développées pour certaines
espèces, ou groupe d'espèces, pour certains
écosystèmes ou biomes à travers le monde. La
problématique des modèles allométrique est largement
évoquée au Chapitre 2. Les équations qui pourraient aider
à développer ce type de recherche dans la perspective des projets
MDP sont celles qui utilisent des paramètres faciles à collecter,
qu'on retrouve dans la plupart des bases de données d'inventaire comme
le DBH et dans une moindre mesure, la hauteur des individus. Le doute
lié à l'utilisation de la hauteur des individus est qu'en
écosystème de savane, on a souvent des futs assez tortueux, ne
reflétant pas l'âge véritable du sujet. Au niveau d'une
même espèce, on peut avoir une forte variation due à la
fréquence des feux de brousse ou des coupes comme l'émondage,
à la pression sur l'espèce pour satisfaire les besoins des
populations ; mais aussi aux différences de méthodes de mesures,
le plus souvent par estimation, etc. Le paramètre le plus sûr et
plus facile à obtenir est par conséquent le DBH ou la
circonférence à 1,3 m, paramètre le plus utilisé
dans les équations allométriques courantes (Brown, 1997;
2002).
Tenant compte de ces considérations, nous avons
opté pour l'élaboration de modèles
allométriques
basés sur des régressions statistiques et celles qui ont
montré les meilleures
performances (cf. Chapitre 2) ont
été utilisées pour convertir les données
d'inventaire en
biomasse. L'inventaire de la végétation a
été réalisé entre 2001 et 2003 au niveau des six
forêts classées des domaines soudaniens et soudano-guinéens
en utilisant l'approche décrite ci-dessous.
Section 3.03 III.3. Méthode d'inventaire de la
végétation ligneuse
dans les six Forêts Classées
L'inventaire de la végétation a
été faite dans 6 Forêts Classées que sont : Bala,
Mampaye, Kantora, Ouli, Patako et Wélor. Le principal but lié
à la méthode utilisée pour cet inventaire est selon Sambou
(2004) d'être opérationnelle sur le terrain et efficiente pour des
besoins de gains de temps et de moyens financiers. L'échantillon
inventorié doit être représentatif pour des besoins
d'inférence statistique, et de permettre des traitements statistiques.
L'approche utilisée est un sondage aléatoire à deux
niveaux. Le premier niveau de sondage est un tirage aléatoire des
mailles caractéristiques ; le deuxième niveau correspond au
tirage aléatoire des placettes. La stratification des forêts
étudiées est basée sur la cartographie (utilisant des
images satellitaires LANDSAT) des zones homogènes appelées
`strates'.
Les cartes ont été quadrillées avec une
grille dont les carrés font 250 m de côté appelé
`maille' (figure 35). Les mailles sont numérotées et un premier
tirage est réalisé sur les mailles pures (qui ne chevauchent pas
entre deux strates). La liste des mailles pures constitue la base de sondage.
Des mailles retenues par un tirage aléatoire sans remise ont
été inventoriées.
L'inventaire des mailles retenues s'est fait sur la base d'un
tirage aléatoire de placettes d'inventaire à partir du centre de
la maille. Le centre de la maille est un point dont les coordonnées UTM
WGS 84 (projection de la cartographie) sont pré collectées au
laboratoire avec l'extraction des centroïdes des carrés de 250 m de
côté. Le centre de chaque maille retenue a pu être
retrouvé utilisant un GPS.
Figure 35. Maillage de la Forêt
Classée de Patako (mailles carrées de 250 m de coté).
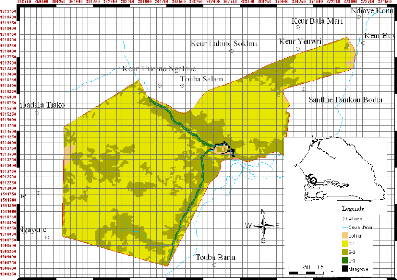
Partant du centre de la maille on tire au hasard 8 placettes
d'inventaire de 20 m de côté. Ce
tirage consiste à
extraire pour 8 directions fixes une distance tirée au hasard entre 0 et
120
m (tableau 12). Les directions sont lues sur une boussole
magnétique, et les distances tirées
sont mesurées en
utilisant un ruban métrique de 50 m de long. Le repérage de la
parcelle
passe par un jalonnage le long de l'azimut de la direction et une
mesure de la distance (quiest tirée au hasard) par rapport au
centre de la maille permet de localiser la placette sans difficulté
(figure 36).
Tableau 12. Caractéristiques d'une fiche
de sondage des placettes d'inventaire
|
Orientation en degré
|
Rose des vents
|
Distance
(da. dg)
|
|
0
|
N
|
A (da)
|
|
45
|
N-E
|
B (db)
|
|
90
|
E
|
C (dc)
|
|
135
|
S-E
|
D (dd)
|
|
180
|
S
|
E (de)
|
|
225
|
S-O
|
F (df)
|
|
270
|
O
|
G (dg)
|
|
315
|
N-O
|
H (dh)
|
Pour chaque orientation (N), on trouve la maille (A) en tirant et
en mesurant sa distance (da)
Une vue schématique de la disposition des placettes de la
maille est proposée à la figure 36.
Figure 36. Schéma du dispositif
d'inventaire
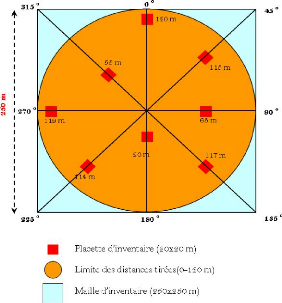
Les placettes sont de forme carrée avec comme centre la
distance tirée sur une direction donnée. Dans la placette une
série de mesures est faite sur les paramètres suivants :
1. Mesures du diamètre ou de la
circonférence des arbres (utilisant un compas forestier et un
ruban gradué en cm pour mesurer les circonférences des gros
arbres);
2. Mesures de la hauteur (dendromètre
pour les grands arbres, perche de 4 m pour les arbustes);
3. Estimations du taux de recouvrement du sol
par les houppiers des arbres et arbustes;
4. Comptages de la
régénération naturelle et du nombre de souches;
5. Observations sur les caractéristiques
du milieu, la strate herbacée, les impacts des activités
humaines, etc.
Les données d'inventaire ont été
organisées dans des fiches de saisie EXCEL pour les traitements. Les
détails de cette méthode sont donnés dans Sambou (2004).
Le tableau 13 est une synthèse des caractéristiques du sondage
réalisé.
Tableau 13. Synopsis du sondage
effectué
|
Nombre de strates
|
Nombre de mailles
|
Nombre de parcelles
|
Nombre d'individus mesurés
|
Surface inventoriée (m2)
|
|
Bala
|
3
|
23
|
184
|
2612
|
73600
|
|
Mampaye
|
3
|
23
|
280
|
4771
|
112000
|
|
Kantora
|
4
|
35
|
184
|
4279
|
73600
|
|
Ouli
|
3
|
23
|
184
|
2270
|
73600
|
|
Wélor
|
4
|
38
|
304
|
2261
|
121600
|
|
Patako
|
2
|
22
|
176
|
2214
|
70400
|
|
Total
|
19
|
164
|
1312
|
18407
|
524800
|
Afin d'aboutir à un traitement adéquat des
données pour l'estimation de la biomasse, Adansonia digitata a
été retirée de la base (diamètres très
élevés, elle intègre difficilement les
modèles)9.
Pour chaque forêt les paramètres de calcul du
tableau 14 ont été utilisés. Le but est :
- de calculer
la biomasse totale pour chaque maille des différentes strates ;
- de réaliser une agrégation des résultats
pour avoir la biomasse totale de la surface
inventoriée de chaque strate ;
- de convertir cette biomasse de la surface inventoriée en
biomasse par hectare (tableau 14).
- La biomasse par hectare est convertie en stocks de carbone
par hectare en utilisant le taux de carbone moyen des espèces dominantes
qui a été obtenu par analyse d'échantillon de bois avec
une ELTRA SC-500.
Utilisation des modèles pour estimer la biomasse
totale Tableau 14. Paramètres de calcul de la biomasse.
|
1 parcelle=20*20m = 400m2
1 maille = 8 parcelles
Surface inventoriée par maille = 8*400 = 3200
m2 Surface totale de la maille = 250*250 = 62500 m2
Surface inventoriée = 5,12 % de la surface de la maille
conversion kg?T: 1kg = 0,001 T
conversion m2?ha: 1 m2 = 0,0001 ha
Biomasse t/ha = ((biomasse maille kg/1000) / (surface maille en
m2/10000))
NB: tonnes = métrique
|
9 Il faut un modèle à part pour les espèces
à gros diamètres comme le baobab qui constituent dans le cas
d'espèce une essence qui requiert un traitement spécial.
Section 3.04 III.4. Estimation de la biomasse des
Forêts Classées
Les principaux résultats obtenus concernent : les
stocks de carbone par hectare pour chaque strate (tableau 15) des Forêts
Classées et la répartition de ces stocks de carbone par classe de
diamètre. L'utilisation des données de suivi de la
végétation ligneuse a permis d'affiner l'analyse sur la
productivité et la dynamique du carbone de ces
écosystèmes.
Tableau 15. Estimation de biomasse avec les
modèles Quadratique, Cubique et polynomial au niveau des Forêts
Classées étudiées.
Forêt classée de Bala
|
Surface Surface
Biomasse Biomasse Biomasse placettes placettes T/ ha T/ha T/ha
Quadratique Cubique Polynomial (m2) (ha) (quadratique)
(cubique) (polynomial)
|
|
S1
|
71827,27
|
110360,17
|
110413,56
|
19200
|
1,92
|
37,4
|
57,5
|
57,5
|
|
S2
|
78092,20
|
106967,62
|
107028,62
|
19200
|
1,92
|
40,7
|
55,7
|
55,7
|
|
S3
|
118474,06
|
157908,43
|
158005,56
|
35200
|
3,52
|
33,7
|
44,9
|
44,9
|
Forêt classée de Kantora
|
Surface Surface
Biomasse Biomasse Biomasse placettes placettes T/ ha T/ha T/ha
Quadratique Cubique Polynomial (m2) (ha) (quadratique)
(cubique) (polynomial)
|
|
FG
|
45851,45
|
51192,05
|
51233,31
|
6401
|
0,64
|
71,6
|
80,0
|
80,0
|
|
S1
|
212134,92
|
303608,01
|
303761,51
|
32000
|
3,2
|
66,3
|
94,9
|
94,9
|
|
S2
|
208840,70
|
244045,58
|
244187,69
|
32000
|
3,2
|
65,3
|
76,3
|
76,3
|
|
S3
|
334146,45
|
399541,13
|
399781,03
|
41600
|
4,16
|
80,3
|
96,0
|
96,1
|
Forêt classée de Mampaye
|
Surface Surface
Biomasse Biomasse Biomasse placettes placettes T/ ha T/ha T/ha
Quadratique Cubique Polynomial (m2) (ha) (quadratique)
(cubique) (polynomial)
|
|
FG
|
150978,36
|
163477,18
|
163609,58
|
16000
|
1,60
|
94,3
|
102,2
|
102,2
|
|
S1
|
64522,53
|
62383,17
|
62449,90
|
16000
|
1,60
|
40,3
|
38,99
|
39,03
|
|
S2
|
161691,75
|
165013,51
|
165170,42
|
22400
|
2,24
|
72,2
|
73,7
|
73,7
|
Forêt classée de Ouli
|
Surface Surface
Biomasse Biomasse Biomasse placettes placettes T/ ha T/ha T/ha
Quadratique Cubique Polynomial (m2) (ha) (quadratique)
(cubique) (polynomial)
|
|
S1
|
44459,99
|
61719,25
|
61749,93
|
16000
|
1,60
|
27,8
|
38,6
|
38,6
|
|
S2
|
44826,30
|
55454,10
|
55489,38
|
22400
|
2,24
|
20,0
|
24,7
|
24,8
|
|
S3
|
123459,57
|
150120,01
|
150225,65
|
35200
|
3,52
|
35,0
|
42,6
|
42,7
|
Forêt classée de Patako
|
Surface Surface
Biomasse Biomasse Biomasse placettes placettes T/ ha T/ha T/ha
Quadratique Cubique Polynomial (m2) (ha) (quadratique)
(cubique) (polynomial)
|
|
S1
|
147742,73
|
156740,02
|
156865,60
|
54400
|
5,44
|
27,1
|
28,8
|
28,8
|
|
S2
|
93434,01
|
106912,35
|
106987,94
|
16000
|
1,60
|
58,4
|
66,8
|
66,9
|
Forêt classée de Wélor
|
Surface Surface
Biomasse Biomasse Biomasse placettes placettes T/ ha T/ha T/ha
Quadratique Cubique Polynomial (m2) (ha) (quadratique)
(cubique) (polynomial)
|
|
S1
|
20830,84
|
20332,60
|
20356,39
|
25600
|
2,56
|
8,1
|
7,9
|
7,9
|
|
S2
|
12943,97
|
18904,06
|
18914,69
|
12800
|
1,28
|
10,1
|
14,8
|
14,8
|
|
S3
|
54747,36
|
61074,28
|
61128,48
|
41600
|
4,16
|
13,2
|
14,7
|
14,7
|
|
S4
|
44913,17
|
42836,22
|
42891,07
|
41600
|
4,16
|
10,8
|
10,3
|
10,3
|
FG, S1, S2, S3, S4 voir légende figure 38
Le tableau 15 montre que les sites présentant plus de
biomasse à l'hectare sont la Forêt Classée de Kantora
(dominée par des savanes boisées et des forêts claires),
suivie de celle de Mampaye (dominée par des savanes boisées). Ces
deux Forêts Classées sont toutes situées au Sud dans la
zone soudano-guinéenne. Cette importante biomasse dans ces
écosystèmes est liée à la présence de grands
sujets dont la contribution sur la biomasse totale est très
prépondérante. La Forêt Classée de Patako suit avec
la présence de savanes arborées et boisées avec de grands
sujets. Patako est suivi de près par les Forêts Classées de
Bala et de Ouli. La Forêt Classée de Wélor est la moins
dotée en biomasse (site très exploité pour le charbon de
bois).
Ces observations nous renseignent sur deux faits majeurs :
- D'une part, la quantité de biomasse n'est pas
seulement liée aux quantités d'eau précipitées
annuellement (zonalité climatique), Kantora situé plus au nord
recevant moins d'eau de pluie présente plus de biomasse par ha que
Mampaye. La fréquence des grands sujets jouent alors un rôle
significatif. A ce niveau, il faut aussi signaler l'importance des facteurs
pédologiques et la nature des terrains qui joueraient un rôle
très important sur le développement des arbres.
- D'autre part, le facteur humain, à travers la
pression sur les ressources ligneuses peut induire des différences
importantes même avec des conditions climatiques similaires. Les
Forêts Classées de Ouli et de Wélor étant
très exposées à l'exploitation forestière ont des
quantités de biomasse à l'hectare beaucoup plus faibles que la
Forêt Classée de Patako pourtant située dans la même
zone climatique.
Concernant la contribution relative des individus de petite ou
grande taille, on constate que les petits sujets peuvent donner un important
taux de couverture, mais leur impact sur la biomasse totale est moins
significatif que les grands arbres. Cependant, la présence des jeunes
sujets permet d'envisager une dynamique active de séquestration alors
que les vieux sujets ont atteint leur limite de croissance donc
présentent une faible capacité de séquestration de
carbone. En d'autres termes, les grands sujets concentrent une partie
significative du stock de carbone, mais leur contribution à la dynamique
de séquestration du
carbone est moins importante que celle des jeunes plantes. Il est
donc intéressant de voir de façon plus détaillée la
contribution des petits et grands sujets dans les stocks de biomasse.
(a) III.4.1. Contribution à la biomasse par classe
de diamètre
Pour mieux caractériser la répartition de la
biomasse en fonction de la structure des formations végétales
étudiées, il convient de voir la répartition par classe de
diamètre de la biomasse totale par strate. Ensuite, les contributions
relatives de chaque classe de diamètre ont été
calculées pour chaque strate.
(i) III.4.1.1 Répartition des individus par
classe de diamètre
La régénération naturelle (individus de
taille inférieure à 5 cm de diamètre pour les
espèces rencontrées dans cette étude) n'est pas prise en
compte dans le décompte des classes de diamètres. Le classement
des individus dans les différentes classes de diamètre montre que
l'essentiel des effectifs sont entre 5-10 cm de diamètre (figure 37). Le
nombre des sujets de gros diamètres n'est pas élevé dans
les formations de savane étudiées surtout à Ouli et
Wélor. D'autres formations comme celles de Kantora et dans une moindre
mesure la forêt de Mampaye ont une répartition de leurs classes de
diamètres plus équilibrée. Le nombre réduit de gros
sujets dans certaines Forêts Classées peut être dO à
l'importance de l'exploitation forestière et aux feux de brousse
répétitifs. Dans certaines conditions c'est la composition
floristique qui impose cette structure. Dans la Forêt Classée de
Bala par exemple la forte présence de Acacia macrostachya
(espèce qui est rarement très grande) peut influencer la
dominance des petites classes de diamètre. Le même cas est
observé lorsque les effectifs sont dominés par Combretum
glutinosum qui peut devenir grand, mais avec une forte fréquence
des feux de brousse et des conditions stationnelles d'aridité, les
grands diamètres ont du mal à se mettre en place. Ainsi, dans la
classe de diamètre 5-10, Combretum glutinosum est
l'espèce la plus représentée.
Figure 37. Répartition des effectifs par
classe de diamètre
|
Bala
|
Kantora
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Mampaye
|
Ouli
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Patako
|
|
Welor
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2- 35-4 4-4
30 2-3 40-4 4-5
50 + 45-5 50 +
35-4 3-3
|
La répartition des individus par classe de
diamètre montre qu'au-delà de 20 cm, le nombre d'individus
diminue significativement (figure 37). Cependant, ce constat ne permet pas de
se prononcer sur la contribution des différentes classes de
diamètre à la biomasse totale. La contribution absolue et
relative à la biomasse totale des strates pour les classes 5-10, 10-15,
15-20 et > 20 est calculée au niveau des 6 Forêts
Classées. La figure 38 montre que même si le nombre d'individus
est moins important dans les classes de plus de 20 cm, cette catégorie
d'individus regroupe l'essentiel de la biomasse des savanes, avec une
proportion faible d'individus. Pour la plupart des cas, les espèces dont
le diamètre est supérieur à 20 cm concentrent plus de 50%
de la biomasse totale. Dans certaines strates cette catégorie
d'espèces dispose de plus 80% de la biomasse totale. Ce constat
s'explique par la démonstration faite au chapitre 2 concernant le bond
significatif de la quantité de biomasse produite quand le
diamètre de l'arbre dépasse le seuil de 17 cm. Exemple, un
Pterocarpus erinaceus de 6 cm de diamètre fait 13,6 kg de
biomasse, celui de 11,3 cm, 63,7 kg, alors que la même espèce
à 19 cm produit 148 kg de biomasse. Cela signifie qu'il faut près
de 12 individus de 6 cm de diamètre pour arriver aux mêmes stocks
de biomasse qu'un sujet de 19 cm. Dans les cas où on a une absence ou
une faible présence de grands arbres, la biomasse est essentiellement
située dans les classes jeunes (exemple de S1 et S4-Wélor et
S3-Ouli). De telles situations traduisent cependant une certaine
vulnérabilité des formations soumises annuellement à des
feux de brousse et parfois à des activités de carbonisation
clandestines ou autorisée comme dans les Forêts Classées de
Wélor, de Ouli et de Bala.
La présence d'un nombre réduit de grands sujets,
mais au sein desquels on compte l'essentiel de la biomasse pose le
problème de l'avenir du carbone dans les formations de savane en Afrique
de l'Ouest. L'exploitation forestière vise le plus souvent de gros
sujets surtout pour le bois d'uvre et de service. Les coupes sélectives
des gros sujets réduisent radicalement les stocks de biomasse au fil des
années. Quant aux petits diamètres, leurs stocks de biomasse est
faible mais le potentiel de séquestration est plus grand que ceux des
gros sujets, s'ils sont épargnés par les feux de brousse. Les
petits sujets ont d'une part une marge de croissance importante et une vitesse
de croissance théoriquement plus grande que celle des sujets adultes.
Ces deux facteurs combinés montrent que pour favoriser la dynamique de
séquestration, il faut sans doute protéger les petits individus,
et pour maintenir les stocks de carbone, il faut porter plutôt une
attention particulière sur les gros sujets.
(ii) III.4.1.2. Répartition de la biomasse par
classe de diamètre
Les tableaux croisés dynamiques (technique statistique)
utilisés ont permis de compter les effectifs dans chaque classe de
diamètre et de cumuler leur total de biomasse. La figure 38 donne la
contribution de chaque classe de diamètre à la biomasse
totale.
Figure 38. Répartition de la biomasse en
fonction des classes de diamètre pour chaque strate des 6 Forêts
Classées.
|
Bala
|
Kantora
|
|
S1
|
FG
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
S2
|
S1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
S3
|
S2
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Qadratiq
|
|
%
|
|
|
|
|
Qu
C
|
|
|
|
|
|
|
|
|
|
Plynomi
Po
|
|
|
|
|
|
|
|
|
iqe
|
|
|
C
|
|
|
|
|
|
|
|
|
|
ly
|
|
|
|
|
|
1J%
7
|
|
|
|
|
|
|
|
|
|
|
S3
Bala (taux de couverture %)
S1 : Savane arbustive (15,7%)
S2 : Savane arbustive (18,5%)
S3 : Savane arbustive (15,7%)
Kantora (taux de couverture %) FG : Forêt
claire (75,6%)
S1 : Savane boisée (34,4%)
S2 : Savane arborée/boisée (40%)
S3 : Savane boisée/forêt claire (60,5%)
|
Mampaye
|
Ouli
|
|
FG
|
S1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
S1
|
S2
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
000
0 S2
|
S3
|
|
|
|
80
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
qu
|
|
|
|
|
|
|
|
00 6,8%
00
|
|
|
|
|
|
|
|
|
|
ia
00 ]
|
|
|
|
|
|
|
|
e % 9
0
|
|
|
|
|
|
|
|
|
|
|
[
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
%
6%
6,4% 8% 3
|
|
|
Mampaye (taux de couverture %) FG : Savane
boisée/forêt claire (68,1%)
S1 : Savane boisée (30,9%)
S2 : Savane boisée/forêt claire (51,4%)
|
Ouli (taux de couverture %)
S1 : Savane arbustive (12,5%)
S2 : Savane arbustive (11,2%)
S3 : Savane arbustive/arborée (16,4%)
|
|
Patako
|
Wélor
|
|
S1
|
S1
|
|
|
|
70000
..i, 60000 m50000
4
V, 40000
e
c7 30000 E
.o
--' 20000
10000
0
|
80000
Quadratique Cubique Polynomiale
|
64,5%
|
9000
|
|
Quadratique Cubique
|
39,0%
|
|
|
|
|
|
8000
7000
6000
m
i 5000
2
. 4000 L3
0 3000 :.°8
2000
1000
|
|
Polynomiale
|
|
36,3%
|
|
|
|
|
28,7%
|
|
95,6%
23,5%
|
|
|
|
.,rx,
|
|
|
|
|
|
|
|
4,1% '49'
|
|
17,2%
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
[5-10] (9,905) ]10-15j (9=194) j15-20] (9=69) >20
(9=127)
classe de diametre
|
0
|
C5-10] (9=262) :10-15] (n=70) ]15-20] (n=21) >so (n=i2)
c lasse de diametre
|
|
|
S2
|
S2
|
|
|
|
|
|
90000
80000
1 70000
°
60000
4 .50000
V.
7, 40000
E 30000
.o
20000
10000
0
|
Quadratique Cubique
|
744%
|
16000
14000 -,-,-.., 12000
·,,%.,
iii 10000
4
:2' 8000
e,
6000
t,
..o 4000
2000
|
|
Quadratique Cubique
|
|
78496
|
|
|
|
|
|
71,8%
|
|
|
|
|
Polynomiale
|
|
|
|
Polynomiale
|
|
|
|
|
|
67,7%
|
|
|
|
|
|
|
|
i
|
|
|
|
|
|
|
|
|
|
15,9%
|
.0,
|
|
10,5%
11,0%
|
|
|
|
|
,,,ck 9,6% 7,6% 6,7%
|
|
|
|
|
|
7,296 4096
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
[.5-10j (n=4.5.5) j10-1q (n= 16o) j15-20] (9=53) >20 (n=86)
classe de diametre
|
0
|
[5-10] (n=94) ]10-18] (n=21) ]18-20] (n=6)
>so (n=10)
classe de diamètre
|
|
|
Patako (taux de couverture %)
S1 : Savane arbustive/arborée (16%)
S2 : Savane arborée/boisée (43,6%)
|
S3
|
|
|
|
35000
,z;z 500006
c°8 28000
1
2 20000
15000
E
10000
5000
|
40000
Quadratique Cubique
Polynomiale
|
50,196
|
|
59,496
|
|
|
|
|
|
|
|
|
20,7%
|
10,3% 15,796
|
|
|
|
13,996
|
|
13,396 11,896
|
|
|
|
|
|
|
|
|
0
[5-10] (71.481) ]10-15] (9.137) ]18-20] (n=48) >20(n=82)
classe de diametre
|
|
Wélor (taux de couverture %)
S1 : Savane arbustive (8,5%)
S2 : Savane arbustive (9,5%)
S3 : Savane arbustive (12,3 %) S4: Savane arbustive (15,9%)
|
S4
|
|
|
20000
|
41,3%
|
Quadratique Cubique Polynomiale
|
|
|
18000
|
|
35,596
|
|
|
16000
"ii
|
|
|
|
|
33396
32,896
|
|
.. 14000
ct
|
|
|
|
|
|
|
|
|
|
,., 12000
il
|
|
|
|
99,0%
|
|
|
0 10000
1 8000
|
|
|
|
17,1%
|
|
|
|
|
|
13
.2 6000
.
4000
2000
|
|
|
|
|
|
|
|
|
|
|
|
|
6,11% 8,7%
|
|
|
|
|
|
|
|
|
|
|
|
0
[5-10] (n=826) ]10-15] (n=147) ]15-20] (n=26) >so (n=35)
classe de diamètre
|
La forte contribution des gros sujets aux
stocks totaux de biomasse est un fait établi par
d'autres etudes
(Clark et Clark, 2000; Joosten et al., 2004). Au niveau des savanes,
les stocks m c %
de biomasse (carbone) varient
énormément selon qu'il y ait une forte présence de gros
51 ( 7) 0] 5
individus ou pas. Dans la perspective de Kyoto (critère
d'éligibilité des terres qui sont non
clae de diamètr
forestières à la date de décembre 1990),
les formations de savane présentent un potentiel de séquestration
relativement important si les termes d'aménagement et de gestion sont
bien définis et correctement mis en ~uvre. Pour mieux asseoir la
compréhension du potentiel de ces écosystèmes, une analyse
complémentaire a été faite utilisant des données de
parcelles de suivi de la végétation10
; celles-ci ont permis de se prononcer sur la vitesse de croissance des
espèces et sur la productivité des écosystèmes de
savane.
Section 3.05 III.4.2. Dynamique du carbone par
l'analyse de la croissance des plantes
Cette analyse est basée sur des données de
parcelles de suivi de la végétation. L'idée d'utiliser ce
dispositif en même temps que les inventaires nationaux est un aspect
important dans la caractérisation du potentiel de séquestration
du carbone des écosystèmes de savanes. Une parcelle de suivi de
la végétation est un échantillon spatial
représentatif des formations végétales d'une région
et dont le suivi dans le temps permet de comprendre les caractéristiques
de la dynamique de la végétation sur le plan quantitatif et
qualitatif. Les parcelles étudiées dans le cadre de ce travail
(parcelles de 1 ha) ont fait l'objet d'un suivi depuis 1993 par un projet de
l'Institut des Sciences de l'Environnement (ISE) de l'Université Cheikh
Anta Diop de Dakar, financé par la coopération danoise, en
collaboration avec l'Institut de Botanique et d'Ecologie de l'Université
de Aarhus. C'est avec ce projet nommé ENRECA (Enhancement of
Research Capacity, DAN N° 104. Dan. 8L/203-DANIDA) que ce suivi a
été fait dans la partie sud du pays, correspondant aux
écosystèmes qui ont fait l'objet des analyses
développées dans cette étude (figure 39).
10 Les parcelles de suivi de la végétation sont
souvent appelées `parcelles permanentes'. La controverse sur le
caractère permanent de ces parcelles nous a incité à opter
pour ce terme.
Figure 39. Localisation des parcelles de suivi
de la végétation
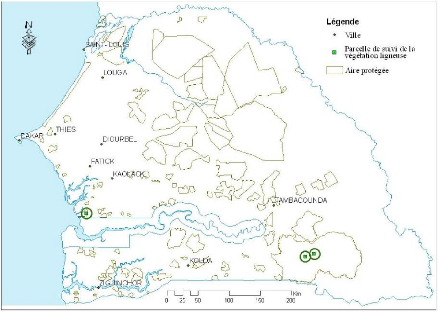
Ces parcelles de suivi de la végétation ligneuse
sont localisées dans la zone soudanienne sont représentatives des
formations végétales étudiées. Situées dans
des Parcs Nationaux, ces sites de suivi ne sont pas soumis à
l'exploitation de l'homme mais subissent les feux dits
précoces11 utilisés comme outil
d'aménagement.
(a) III.4.2.1. Méthode de suivi de la
végétation
Le suivi de la dynamique de la végétation
ligneuse est une approche éprouvée pour caractériser
l'accroissement des individus de différentes espèces. Il existe
plusieurs motivations dans l'implantation des parcelles de suivi de la
végétation. Dans certains cas, on les utilise pour
vérifier l'impact d'une contrainte comme les feux de brousse ou le
pâturage, la dynamique de la biodiversité dans une zone
donnée. Une autre dimension non directement écologique et
floristique vient s'adjoindre de nos jours à l'étude des
parcelles de suivi de la végétation. Il s'agit de la
quantification de la dynamique de la biomasse produite. Ces données de
productivité ont souvent été considérées
comme lacunaires dans les écosystèmes de savane. Avec la mise en
~uvre des MDP, il est donc capital de savoir non seulement le
11 Ces feux d'aménagements sont dits précoces mais
interviennent souvent quand la strate herbacée est très
sèche entraînant des feux violents et destructeurs.
potentiel de nos écosystèmes mais aussi la
vitesse d'accumulation de la phytomasse qui traduit concrètement le
carbone séquestré par la végétation ligneuse. La
base de données des 3 parcelles de suivi a été reprise et
a permis de faire des analyses de la dynamique de la biomasse.
- Les sites de suivi de la
végétation
Les sites de suivi ont été choisis en tenant
compte entre autres de la représentativité des
écosystèmes et de leur homogénéité. Des
sites de 1 ha ont été délimités sur des
unités topographiques homogènes avec une flore et une
végétation ne variant pas significativement dans la placette. La
dimension humaine a été aussi prise en compte dans l'emplacement
des sites, éloignés des zones directes d'intervention humaine et
sans contrainte foncière. Ces sites ne sont pas protégés
des feux de brousse et du pâturage, mais des coupes de bois ne devraient
pas s'y dérouler. Les endroits indiqués pour l'emplacement de ces
parcelles de suivi sont par conséquent le domaine forestier
protégé.
- Mise en place des parcelles de suivi de la
végétation
Au niveau de chaque site choisi, une parcelle de 1 ha (10000
m2) a été délimitée et
matérialisée par des repères discrets. L'implantation de
la parcelle de suivi de la végétation requiert 5 étapes
reprises par Sambou et al. (2007).
a) - La mise en place des lignes de base de la
parcelle
Elle consiste à mettre en place à l'aide d'une
boussole (orientation et mesure d'angle), et d'un ruban de 100 m (mesure de
distance) une première ligne droite de 100 m dite ligne de base 1 (L1)
et une deuxième ligne de 100 m dite ligne de base 2 (L2) perpendiculaire
à la première. Ces deux axes sont matérialisés par
des cordons visibles (figure 40).
Figure 40. Délimitation des lignes de
base de la parcelle permanente

- Délimitation des bandes de la
parcelle
Elle consiste à mettre en place des bandes de 10 m de
large perpendiculaires à la ligne de base (L1) et de 100 m de long
parallèlement à la deuxième ligne de base (L2) à
l'aide de rubans métriques et de ficelles de 100 m (figure 41).
Figure 41. Délimitation des bandes de la
parcelle de suivi
b1 b2 b10

L2
a10
a2
a1
b10
b2
b1
L1
j10
j2
j1
à la ligne de délimitation droite de la bande
(coordonnée x) et par rapport à la ligne de délimitation
séparant cette placette de la précédente
(coordonnée y). La numérotation des individus et la
détermination de leur position sont effectuées de cette
façon pour chaque placette.
Figure 43. Détermination de la position
des individus dans les placettes
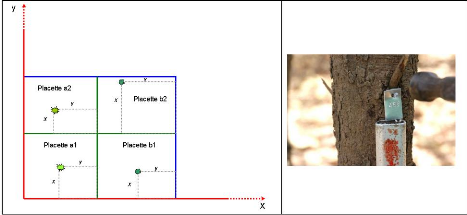
Les coordonnées géographiques des coins de la
parcelle permanente sont relevées dans la projection UTM, WGS 84.
e) - La collecte de données
Les données collectées concernent les
noms scientifiques des individus ligneux
étiquetés dans les placettes, leurs coordonnées, leur
diamètre à 1,30 m de hauteur (DBH), leur hauteur
totale, leur phénologie, leur état sanitaire, la
régénération naturelle, la composition floristique et
l'abondance des herbacées, les caractéristiques
écologiques du site et l'impact des activités humaines.
La position relative de chaque individu est estimée en
distance métrique comme indiqué à la figure 43.
La mesure du diamètre des individus de moins de 50 cm
de diamètre se fait à l'aide d'un compas forestier (figure 44).
Le diamètre des individus supérieurs à 50 cm s'obtient par
la conversion de la circonférence à 1,30 m (DBH= C/3,14).

Figure 44. Mesure de DBH avec un compas
forestier
Les données ont été collectées en
1993, 1997, 2003 et 2005. Les périodes de mesures correspondent toutes
au début de la saison sèche. Pendant cette période, le
terrain est plus accessible et la strate herbacée ainsi que la
régénération naturelle des espèces ligneuses n'ont
pas encore subi les feux de brousse.
Les données de la première année de mesure
correspondent à la situation de référence de la flore et
de la végétation de la parcelle de suivi.
(b) III.4.2.2. Traitement des données
Les données saisies sur fichier EXCEL, ont fait l'objet
d'un nettoyage des données erronées et des erreurs de saisie. Une
base de données a été compilée en mettant les
individus (enregistrements) sur des identifiants uniques. A partir de la
première année de mesure, toutes les espèces qui
apparaissent pendant les autres mesures sont dotées d'un identifiant.
Ainsi au bout du processus, toutes les espèces ont eu une clé
unique permettant de réaliser les traitements sans se tromper
d'individus. Au total, une base de 3115 enregistrements a été
établie en discriminant les individus selon leur parcelle d'appartenance
à travers une clé secondaire qui donne le numéro de la
parcelle.
Sur l'ensemble des paramètres mesurés dans le
format originel, nous nous sommes intéressés aux
paramètres que sont le nom de l'espèce et le diamètre
à 1,3 m. Le but de cette analyse est le suivi de la dynamique du
carbone, le DBH utilisé dans les modèles allométriques
étant le paramètre essentiel. Les modèles
allométriques utilisés sont ceux développés dans le
chapitre 2. Les modèles retenues sont le quadratique, le cubique et le
polynomial.
L'objectif visé en utilisant les données sur les
parcelles de suivi est de :
- suivre la productivité globale (souvent estimée)
des parcelles en tant que témoin des processus des
écosystèmes de savane similaires ;
- fournir une estimation de la croissance des espèces
(information manquante dans la littérature) ;
- connaître la contribution relative des espèces
à la production de biomasse; - approcher la dynamique de la
biodiversité et de la régénération naturelle ; -
préciser l'équilibre de la structure des formations à
travers la distribution des classes
de diamètre.
(c) III.4.2.3. Résultats
- Productivité globale des 3 parcelles
suivies
Le premier niveau d'analyse consiste à
caractériser l'abondance des espèces. Une fusion des trois
parcelles montre déjà 4 catégories d'espèces en
fonction du nombre d'individus par espèce et de l'abondance de la
régénération naturelle (figure 45). Pour l'ensemble des
parcelles de suivi, les espèces les plus abondantes sont Combretum
glutinosum, Terminalia macroptera, Cassia sieberiana, Pterocarpus erinaceus et
Hexalobus monopetalus pour une liste globale de 57 espèces. Le
nombre total d'individus des trois parcelles a augmenté de 18% entre la
première et la dernière année. Cependant, le nombre total
d'espèces n'a pas beaucoup changé depuis 1997, même si
quelques mutations ont pu être notées dans la répartition
des individus dans les classes de diamètres (tableau 16).
Tableau 16. Abondance des espèces pour
les trois parcelles
|
1993
|
1997
|
2003
|
2005
|
|
2556
|
3115
|
3116
|
3113
|
De 1993 à 1997 l'augmentation du nombre d'individus
dans les 3 parcelles cumulées est largement positive. A partir de 1997
le nombre d'individus dans l'ensemble était plus ou moins stable.
Ces informations agrégées ne donnent qu'une
vision globale de la dynamique des effectifs. Une analyse plus
détaillée par espèce pour les 4 années de suivi,
montre des situations variées. La figure 45 est une synthèse de
cette dynamique des effectifs des individus au cours du suivi.
Figure 45. Abondance des espèces des 3
placettes en 1993-1997-2003-2005 (quelques espèces très rares
dans les parcelles de suivi ne sont pas représentées)

GROUPE A :
Espèces très abondantes et bonne
régénération naturelle sur les 3 parcelles permanentes
GROUPE B :
Espèces abondantes et régénération
naturelle relativement bonne sur les 3 parcelles permanentes
GROUPE C :
Espèces en nombre faible et
régénération naturelle relativement faible sur les 3
parcelles permanentes
GROUPE D :
Espèces en nombre très faible et
régénération naturelle très rare sur les 3
parcelles permanentes
La figure 45 montre qu'en termes d'effectif, on a 4
catégories d'espèces. La première classe (A) retient
l'attention: espèces très abondantes et avec une bonne
régénération naturelle sur les 3 parcelles de suivi. Mais
l'abondance d'une espèce ne traduit pas forcément que sa
productivité est bonne à l'échelle de l'individu. Elle
traduit simplement que l'espèce se reproduit bien et imprime son
empreinte sur la physionomie générale. Pour compléter donc
l'analyse il nous a fallu analyser la productivité annuelle des
espèces dans les 3 parcelles de suivi de la végétation.
(d) III.4.2.4. Productivité des parcelles
permanentes
Croissance des espèces
Il est établi que les espèces de savane sont en
général de croissance relativement lente (Vries et al.,
1980; Breman et Dewit, 1983; Cook et Iverson, 1989; Kessler et Breman, 1991).
Les espèces de savane sont soumises à des conditions climatiques
caractérisées par une longue saison sèche et une grande
fréquence des feux de brousse. Les réponses d'adaptation à
toutes ces contraintes ralentissent la croissance des arbres. Ainsi il faut
plusieurs années à un arbre pour atteindre un gros
diamètre. C'est pour cette raison d'ailleurs que dans la plupart des
programmes de reboisement pendant les années de sécheresse, des
espèces exotiques qui se développent plus rapidement ont
été préférées aux espèces locales.
L'argumentaire est discutable, mais nous laissons ce débat à
d'autres cadres de recherche.
La vitesse de croissance des individus peut être
mesurée à travers un suivi de la croissance du diamètre
des arbres. Il faut toutefois noter que la base de données
traitée a été filtrée, et certaines données
incohérentes comme les enregistrements caractérisés par
une régression très significative de la taille du tronc ont
été retirés. Ces problèmes peuvent être
liés entre autres à des erreurs de mesure ou de transcription. Il
faut signaler que la marge de croissance entre deux périodes de mesures
est souvent égale ou légèrement supérieure à
la marge d'erreur de mesure. La croissance volumétrique s'exprime
parfois en moins d'un cm entre deux mesures ; ce qui fait qu'une erreur de
quelques mm peut entraîner une sous-estimation ou surestimation de la
taille réelle du tronc. Pour les gros sujets à croissance rapide,
le problème s'est posé avec moins d'acuité. En
reconstituant ce qui a pu l'être, les espèces qui
présentent les taux de croissance moyenne les plus significatives
constituent une liste de 24 espèces sur les 57. La croissance annuelle
exprime ici l'augmentation totale du DBH entre deux mesures et le
résultat est divisé par le nombre d'année entre deux
missions de collecte.
Il faut noter que les espèces Detarium
senegalense et Bombax costatum (espèces d'affinité
guinéenne) sont les plus productives de la série. Elles sont
suivies de près par Anogeissus leiocarpus, Lannea microcarpa,
Pterocarpus erinaceus, et Terminalia macroptera (affinité
soudanienne, figure 46).
Figure 46. Croissance moyenne annuelle des
espèces les plus abondantes dans les 3 parcelles de suivi de la
végétation.
La croissance moyenne annuelle ne rend pas compte de sa forte
variabilité temporelle. Cette variation de la croissance des arbres dans
le temps est fortement liée en partie à la quantité et la
distribution des précipitations. Entre une période sèche
et une période humide, le taux de croissance volumétrique peut
passer du simple au double. Ainsi les précipitations favorables depuis
2003 ont entraîné une bonne croissance des arbres des 3 parcelles
de suivi de la végétation. Aussi, entre 1993 et 1997 les bonnes
précipitations enregistrées expliquent une croissance assez
significative dans l'ensemble (figure 47 et 48).
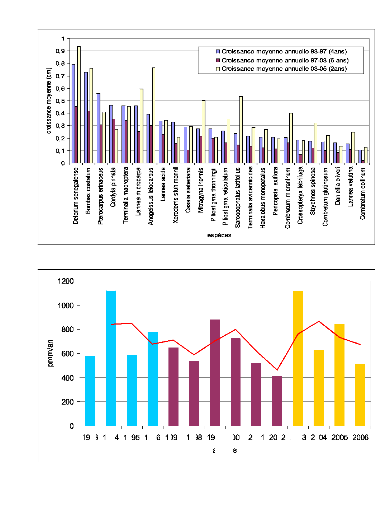
Figure 47. Comparaison de la vitesse de
croissance des diamètres pour les trois périodes de suivi.
Figure 48. Evolution de la pluviométrie
à la station de Tambacounda
Il faut toutefois noter que la production totale de biomasse
des parcelles permanentes est le
résultat du bilan entre les gains et
les pertes de biomasse. Au moment où on enregistre des
recrus
d'individus, on note aussi des disparitions d'individus du fait des coupes, des
feux de
SR
CD
CD
CD
CD
CD
CD
·
·
·
·
·
·
co
CD
CD
CAD` 1`)
0
0
0
O
\
0
0
0 .ter
brousse ou la mortalité naturelle. Aux comptes, la
croissance de biomasse dépendra de la dynamique des pertes en rapport
avec les gains. Sur les parcelles de suivi 1 et 2 (figure 49 et 50),
l'accumulation de biomasse suit la même logique que la vitesse de
croissance des individus et la vigueur de la régénération
naturelle. La forte réduction pendant la période 2007-2003 reste
liée en grande partie à la baisse des précipitations. Par
contre, la forte croissance de la biomasse au sein de la parcelle 3 est une
conjonction de l'augmentation des précipitations et d'une importante
augmentation de la régénération naturelle affranchie
(figure 51,).
La placette 2 est moins productive que les autres parce
qu'elle a moins d'individus et peu de sujets de gros diamètres. La
parcelle 3 a plus de biomasse que la parcelle 1 même si cette
dernière a plus d'individus. La différence se situe au niveau du
nombre de sujets de gros diamètre.
Figure 49. Dynamique de la biomasse totale pour
la parcelle 1
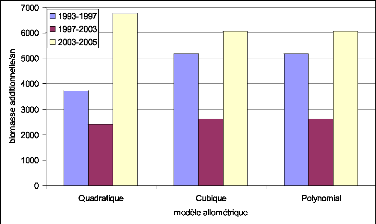
Figure 50. Dynamique de la biomasse totale pour
la parcelle 2
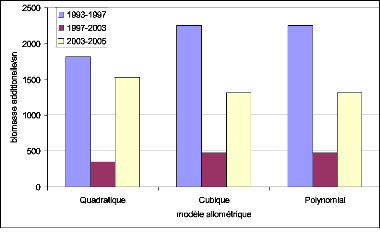
Figure 51. Dynamique de la biomasse totale pour
la parcelle 3
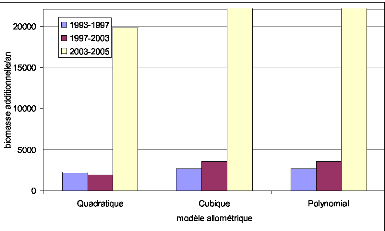
L'analyse de la dynamique des effectifs des classes de
diamètre donne une information
2003-205
additionnelle importante à considérer. La
dynamique des effectifs dans les classes de diamètre est liée
à plusieurs facteurs naturels et humains. De façon
générale, la classe 5-10 a connu une réduction des
effectifs entre 1993 et 2003, et avec le retour des pluies, et probablement la
réduction de certaines formes de pressions comme l'exploitation et les
feux de brousse, la régénération naturelle a soutenu une
augmentation significative des effectifs
Qq uqe y
dans cette classe de diamètre. Pour les effectifs de la
classe 10-15 cm les variations ont été
faibles. Une certaine
augmentation des effectifs a pu être notée dans les parcelles 1 et
3 au
niveau de la classe 15-20. Ce qui peut être noté
dans l'ensemble c'est la densification des individus de gros diamètre au
niveau des parcelles de suivi de la végétation.
Figure 52. Dynamique des classes de
diamètres de la parcelle 1.
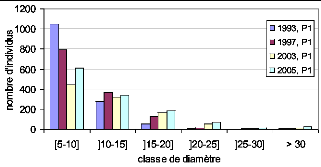
Figure 53. Dynamique des classes de
diamètres de la parcelle 2.
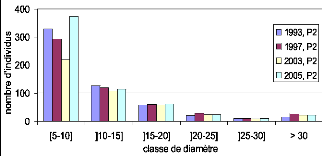
,
Figure 54. Dynamique des classes de
diamètres de la parcelle 3.
2003 P1
L'analyse de la biomasse par espèce et la contribution
relative de chacune dans les stocks
totaux de biomasse permet de mieux
caractériser le potentiel en biomasse de ces formations
végétales.
[5-10]
(e) III.4.2.5. Contribution des espèces à
la biomasse totale
La dynamique de la biomasse est dans bien des cas
portée par quelques espèces qui contribuent largement à sa
production et sa dynamique annuelle. Les espèces les plus influentes sur
la biomasse sont celles qui sont abondantes, qui
régénèrent bien et qui peuvent avoir un nombre important
de gros diamètres. Il n'y a que quelques espèces qui
répondent à ces critères. Ainsi une espèce souvent
abondante dans les parcelles étudiées comme Combretum
glutinosum n'est pas forcément celle qui contribue le plus à
la biomasse totale.
- La parcelle 1 : 55 % de la biomasse est portée par
Pterocarpus erinaceus et Terminalia macroptera ;
- La parcelle 2 : 53 % de la biomasse est portée par
Pterocarpus erinaceus, Bombax costatum et Combretum
glutinosum ;
- La parcelle 3 : 52 % de la biomasse est portée par
Pterocarpus erinaceus et Anogeissu leiocarpus.
Il apparaît alors que Pterocarpus erinaceus
est, dans ces écosystèmes soudaniens, une espèce qu'il
faut beaucoup considérer dans les stocks et la dynamique du carbone,
puisqu'elle se régénère bien et sa croissance
volumétrique est parmi les meilleurs (deuxième après
Detarium senegalense). D'autres espèces se positionnent bien
dans la production de biomasse, il s'agit de Bombax costatum et
Terminalia macroptera. Dans le cas de la parcelle 2, Combretum
glutinosum compte pour 11 % de la biomasse, alors que sa contribution est
de 6 % seulement pour la parcelle 1. Sur la parcelle 3 cette espèce fait
partie des moins productives. La contribution relative à la biomasse
totale des espèces principales est présentée sur les
figures 55, 56 et 57.
Figure 55. Contribution des espèces
à la biomasse totale de la parcelle 1.
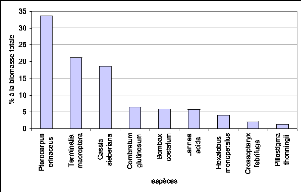
Figure 56. Contribution des espèces
à la biomasse totale de la parcelle 2.
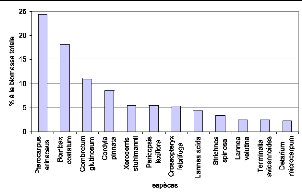

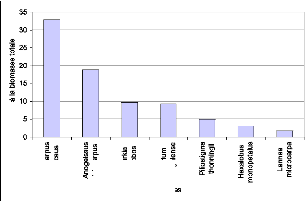
%
a
Pteroc
erina
imocE PE biglt
co co -cl Detai
CD,
2 seneaa

Figure 57. Contribution des espèces
à la biomasse totale de la parcelle 3.
- Potentiel de séquestration des espèces
à forte croissance
Partant des données sur le taux de croissance annuelle
des espèces (figure 58), il est possible de faire des projections sur la
taille d'un individu pendant une période de 30 ans. L'idée est de
chercher à préciser la valeur potentielle d'un arbre
préservé pendant cette durée sur la base de la
quantité nette de biomasse (carbone) séquestrée.
L'approche est d'appliquer le taux de croissance sur un individu de 5 cm de
diamètre et d'analyser la courbe d'évolution théorique de
son diamètre12. Il s'agit ensuite
d'appliquer une régression allométrique sur les valeurs
projetées des diamètres pour estimer la biomasse.
En prenant comme base les espèces à croissance
rapide, on obtient la courbe d'évolution des diamètres
indiquée à la figure 58.
Figure 58. Courbe d'évolution des
diamètres des espèces à croissance relativement rapide
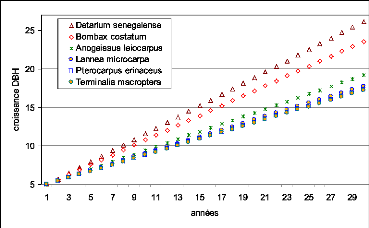
La figure 58 montre que la différence des diamètres
se creuse vers la fin de la série de 30 ans. Bomax costatum
A
L'écart de la taille des diamètres n'est pas
constant. On note que Detarium senegalense et Lannea mrocarpa
Bombax costatum ont tendance à se
démarquer du lot en devenant de grands sujets. Ces
rpu es
Tili t
aspects influent fortement sur la biomasse. En effet, en
appliquant les trois modèles allométriques retenus dans ce
travail, on arrive à des situations différentes (figure 59)
12 Cette extrapolation temporelle est indicative puisque la
vitesse de croissance de l'arbre n'est pas linéaire.
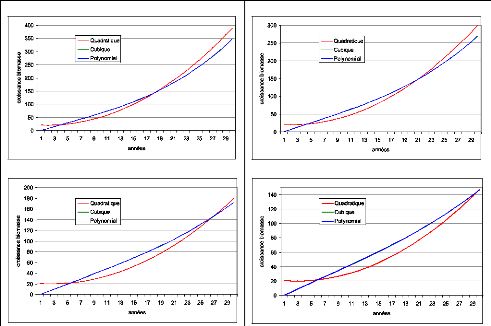
Anogeissus leiocarpus
Detarium senegalense
Bombax costatum
Lannea microcarpa
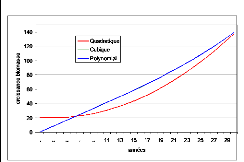
Pterocarpus erinaceus
aq uaiq
Figure 59. Evolution de la biomasse des
espèces à croissance relativement rapide
|
NB. Les modèles cubique et polynomial (ordre 3) sont si
proches qu'ils se superposent.
|
La figure 59 montre que les espèces Detarium
senegalense et Bombax costatum qui ont les plus
Quadratue Quadratique
120 C
grands taux de croissance, n'accumulent que 400 kg de biomasse au
bout de 30 ans ce qui
Polnomial Ponomal
100
correspond en moyenne à 0,4 t de carbone. Si le coût
du carbone est à son meilleur prix
u
(US$20 en moyenne, prix d'aoOt 2008) la valeur du carbone
séquestré sera de US $ 8 (4000 5 7 1 3 5 7 9 11 13 15 7 19 21 23
25 27 29
ubiq 2 5 29
lil ée nn
FCFA, pour un taux moyen de
1USD=500FCFA). A partir de cette somme (très faible du
20
reste), on aura deux possibilités ; soit d'accroître
le nombre de sujets sur de grandes surfaces,
0
5 7 9 11 13 15 17 19 2 23 25 27 9
1 5 7 11 13 15 17 19 21 23 25 27 29
ce qui permettra d'augmenter la valeur du carbone et celle des
produits forestiers non
né
ligneux (MEA, 2005) ; soit d'exploiter l'arbre pour une
valorisation plus rentable (djembé
13, meuble, bois d'énergie, etc.).
NB. La période d'accréditation de
7 ans (renouvelable une fois, c'est-à-dire 14 ans) est très
courte pour accumuler une quantité de biomasse significative. Il en est
de même pour la période d'accréditation de 10 ans non
renouvelable.
13 Le terme djembé est donné à un
instrument de musique de la famille des percussions (tam-tams) et qui se
fabrique avec des troncs d'arbres sculptés.
III.5. Analyse du taux de carbone dans les
échantillons d'espèces ligneuses
Le carbone organique total a été mesuré
en utilisant la méthode de combustion à sec à une
température de 1250°C en utilisant l'analyseur Eltra SC-500 (ELTRA
GmbH, Allemagne :
www.eltragmbh.com) avec
une précision de #177; 0.2% ; figure 60.
Figure 60. Appareil Eltra SC-500 pour l'analyse
du taux de carbone et préparation des échantillons

Eltra SC-500
Echantillons moulus de bois et
d'écorce
L'appareil Eltra SC-500 présenté à la
figure 60 est utilisé pour analyser le taux de carbone de
différents matériaux : cendre, charbon, carbonate de calcium,
calcaire, ciment, gypse, bois, plastique, etc. La méthode de la mesure
est basée sur le principe de la combustion de l'échantillon et
l'analyse des gaz émis, par leur absorption infrarouge. L'oxydation de
l'échantillon lors de la combustion à une température de
1250°C permet aux signaux infrarouge de capter la partie CO2 et SO2 lors
de la combustion. Les proportions des deux composantes sont affichées
sur l'écran de contrôle et enregistré dans l'ordinateur
connecté à l'appareil. Pour avoir une bonne combustion, de
l'oxygène pur est pompé dans la chaudière à l'aide
d'un régulateur électronique qui maintient le flux
d'oxygène à un niveau constant de 180 l/h. Un filtre à
poussière et un capteur d'humidité assurent des conditions
d'analyse à sec et sans déchets extérieurs. L'ordinateur
convertit le signal reçu en taux de carbone en utilisant le poids
d'entrée de l'échantillon.
Figure 61. Schéma de l'analyseur
Eltra-SC500
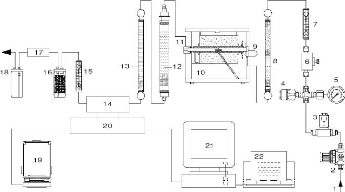
1 Entrée du gaz (O2)
2 Régulateur de pression
3 Valve pour l'oxygène
4 Changeur de pression
5 Jauge de pression
6 Ajusteur du flux
7 Affichage du débit d'oxygène
8 Purificateur de l'O2
9 Entrée chaudière
10 Chaudière
11 Sortie chaudière
12 Premier capteur d'humidité (Anhydrone)
13 Second capteur d'humidité
14 Chambre infrarouge
15 Affichage du flux de gaz mesuré
16 Allègement du volume avant pompage
17 Pompe à gaz
18 Allègement du volume après pompage
19 Balance électronique
20 Mini interface de contrôle
21 Ordinateur
22 Imprimante
Source : OPERATION MANUAL CS-500, 2004
Avec le ELTRA SC-500 une variété de
matières peut être analysée. Les méthodes de
l'analyse sont par conséquent variées. Comme le type de
combustion varie en fonction de la matière, les paramètres
d'analyse changent tout autant; il s'agit du poids de l'échantillon
expérimental, la procédure pour l'insertion de
l'échantillon dans la chaudière et la sensibilité de la
chambre infrarouge.
Les étapes pour analyser le carbone avec un Eltra SC-500
:
- Prélever 500 mg de poudre de bois et d'écorce des
espèces principales ;
- Codifier des échantillons pour les correspondances ;
- S'assurer que la température de la chaudière est
stable au moins deux heures avant introduction de l'échantillon ;
- Vérifier le capteur d'humidité, au besoin,
remplacer le capteur à magnésium ; - Vérifier que l'apport
d'oxygène a une pression de 2 à 4 bars ;
- Connecter avec l'ordinateur et la balance électronique
;
- Tarer le charriot de combustion avec la balance
électronique ;
- Disposer de 300 mg de biomasse (bois ou écorce) sur la
balance ;
- Transférer le poids à l'ordinateur ;
- Introduire le charriot contenant l'échantillon dans la
chaudière pour amorcer l'analyse ;
- Retirer le charriot de combustion immédiatement à
la fin d'une analyse.
Le carbone est mesuré au niveau de la chambre
infrarouge ; il faut éviter que des poussières
d'échantillons résiduels qui restent dans le canal de la
chaudière ne faussent les observations infrarouges.
L'analyse dure au minimum 50 secondes pour chaque passage. Il
faut noter que pour chaque échantillon deux passages ont
été faits pour plus de précision. Le résultat de
l'analyse du carbone des espèces dominantes des Forêts
Classées est indiqué au tableau 17.
Tableau 17. Taux de carbone pour le bois et
l'écorce de 13 espèces dominantes des Forêts
Classées
|
Espèce
|
Bois
|
Carbone/m1
|
Carbone/m2
|
Ecorce
|
Carbone/m1
|
Carbone/m2
|
|
Ximenia americana
|
B1a
|
51,663
|
51,757
|
E1b
|
45,799
|
45,555
|
|
Cordyla pinnata
|
B2a
|
52,163
|
52,463
|
E2b
|
47,676
|
47,812
|
|
Acacia macrostachya
|
B3a
|
50,770
|
50,670
|
E3b
|
47,133
|
47,293
|
|
Crossopterix febrifuga
|
B4a
|
53,055
|
53,520
|
E4b
|
47,373
|
48,767
|
|
Balanites aegyptiaca
|
B5a
|
50,434
|
50,463
|
E5b
|
45,113
|
44,652
|
|
Combretum glutinosum
|
B6a
|
47,216
|
46,984
|
E6b
|
46,702
|
46,594
|
|
Combretum nigricans
|
B7a
|
45,165
|
44,855
|
E7b
|
44,889
|
44,479
|
|
Bombax costatum
|
B8a
|
49,300
|
49,672
|
E8b
|
50,268
|
50,462
|
|
Sterculia setigera
|
B9a
|
49,274
|
49,564
|
E9b
|
38,523
|
38,899
|
|
Pterocarpus erinaceus
|
B10a
|
51,873
|
51,462
|
E10b
|
43,136
|
44,235
|
|
Acacia seyal
|
B11a
|
47,395
|
47,616
|
E11b
|
44,468
|
43,900
|
|
Danielia oliveri
|
B12a
|
46,560
|
46,859
|
E12b
|
48,767
|
48,320
|
|
Terminalia macroptera
|
B13a
|
52,038
|
51,965
|
E13b
|
46,556
|
46,607
|
NB. m1 et m2 correspondent respectivement à la
première et à la deuxième mesure
D'autres auteurs ont mené des analyses de taux de
carbone sur d'autres espèces. Pour Guiera senegalensis, une
espèce buissonnante des terres dégradées, Lufafa et
al. (2008) ont obtenu un taux de 0,495 (49,5%) en utilisant un LECO C
Analyzer (LECO Inc., St. Joseph, Michigan).
Ainsi le taux moyen de carbone pour les espèces de
savane est de 49,8 %. Ce taux est en cohérence avec les recommandations
de IPCC qui est de 50%. Ce contrôle rassure sur le facteur utilisé
pour la conversion de la biomasse en carbone.
Section 3.06 III.6. Le stock de carbone des
Forêts Classées étudiées
Le taux de carbone de la biomasse ligneuse utilisé
provient des analysées effectuées dans ce travail et a permis de
convertir les stocks de biomasse en carbone.
Tableau 18. Conversion des stocks de biomasse en
carbone pour les différentes strates des Forêts
Classées.
Forêt classée de Bala
T/ ha T/ha T/ha T C/ha T C/ha T C/ha
(quadratique) (cubique) (polynomial) (quadratique) (cubique)
(polynomiale)
|
S1
|
37,4
|
57,5
|
57,5
|
18,6252
|
28,635
|
28,635
|
|
S2
|
40,7
|
55,7
|
55,7
|
20,2686
|
27,7386
|
27,7386
|
|
S3
|
33,7
|
44,9
|
44,9
|
16,7826
|
22,3602
|
22,3602
|
Bala (taux de couverture %)
S1 : Savane arbustive (15,7%)
S2 : Savane arbustive (18,5%)
S3 : Savane arbustive (15,7%)
Forêt classée de
Kantora
|
T/ ha T/ha T/ha
(quadratique) (cubique) (polynomial)
|
T C/ha (quadratique)
|
T C/ha (cubique)
|
T C/ha (polynomiale)
|
|
FG 71,6
|
80,0
|
80,0
|
35,6568
|
39,84
|
39,84
|
|
S1 66,3
|
94,9
|
94,9
|
33,0174
|
47,2602
|
47,2602
|
|
S2 65,3
|
76,3
|
76,3
|
32,5194
|
37,9974
|
37,9974
|
|
S3 80,3
|
96,0
|
96,1
|
39,9894
|
47,808
|
47,8578
|
Kantora (taux de couverture %) FG : Forêt
claire (75,6%)
S1 : Savane boisée (34,4%)
S2 : Savane arborée/boisée (40%)
S3 : Savane boisée/forêt claire (60,5%)
Forêt classée de
Mampaye
T/ ha T/ha T/ha T C/ha T C/ha T C/ha
(quadratique) (cubique) (polynomial) (quadratique) (cubique)
(quadratique)
|
FG
|
94,3
|
102,2
|
102,2
|
46,9614
|
50,8956
|
50,8956
|
|
S1
|
40,3
|
38,99
|
39,03
|
20,0694
|
19,41702
|
19,43694
|
|
S2
|
72,2
|
73,7
|
73,7
|
35,9556
|
36,7026
|
36,7026
|
Mampaye (taux de couverture %) FG : Savane
boisée/forêt claire (68,1%)
S1 : Savane boisée (30,9%)
S2 : Savane boisée/forêt claire (51,4%)
Forêt classée de Ouli
T/ ha T/ha T/ha T C/ha T C/ha T C/ha
(quadratique) (cubique) (polynomial) (quadratique) (cubique)
(quadratique)
|
S1
|
27,8
|
38,6
|
38,6
|
13,8444
|
19,2228
|
19,2228
|
|
S2
|
20,0
|
24,7
|
24,8
|
9,96
|
12,3006
|
12,3504
|
|
S3
|
35,0
|
42,6
|
42,7
|
17,43
|
21,2148
|
21,2646
|
Ouli (taux de couverture %) S1 : Savane
arbustive (12,5%)
S2 : Savane arbustive (11,2%)
S3 : Savane arbustive/arborée (16,4%)
Forêt classée de Patako
T/ ha T/ha T/ha T C/ha T C/ha T C/ha
(quadratique) (cubique) (polynomial) (quadratique) (cubique)
(quadratique)
|
S1
|
27,1
|
28,8
|
28,8
|
13,4958
|
14,3424
|
14,3424
|
|
S2
|
58,4
|
66,8
|
66,9
|
29,0832
|
33,2664
|
33,3162
|
Patako (taux de couverture %)
S1 : Savane arbustive/arborée (16%)
S2 : Savane arborée/boisée (43,6%)
Forêt classée de Welor
|
T/ ha T/ha T/ha
(quadratique) (cubique) (polynomial)
|
T C/ha (quadratique)
|
T C/ha (cubique)
|
T C/ha (quadratique)
|
|
S1 8,1
|
7,9
|
7,9
|
4,0338
|
3,9342
|
3,9342
|
|
S2 10,1
|
14,8
|
14,8
|
5,0298
|
7,3704
|
7,3704
|
|
S3 13,2
|
14,7
|
14,7
|
6,5736
|
7,3206
|
7,3206
|
|
S4 10,8
|
10,3
|
10,3
|
5,3784
|
5,1294
|
5,1294
|
Welor (taux de couverture %)
S1 : Savane arbustive (8,5%)
S2 : Savane arbustive (9,5%)
S3 : Savane arbustive (12,3%) S4: Savane arbustive (15,9%)
Les résultats obtenus sur les stocks de carbone sont
moins importantes que ceux estimés par d'autres études puisque
nous n'avons pas considéré toute la biomasse sur pied. Les
travaux de Woomer et al. (2004) montrent pour la zone
soudano-guinéenne des stocks totaux de carbone de la biomasse des
forêts de l'ordre de 60 t/C/ha. Si on déduit les stocks de la
biomasse herbacée on se situe à des niveaux de stocks de l'ordre
de 40 à 50 t/C/ha pour la biomasse ligneuse sur pied. Ces
résultats sont en adéquation avec les estimations obtenues dans
les savanes boisées et forêts claires des Forêts
Classées de Mampaye, Kantora et Patako. Pour les savanes plus ouvertes
(arbustives, arborées), la même source donne des quantités
de stocks de carbone équivalentes à 30 t/C/ha, soit 20 à
25 t/C/ha pour la biomasse ligneuse sur pied. Là aussi, les
résultats que nous avons obtenus corroborent ces estimations globales.
La différence entre les résultats obtenus dans ce travaille et
ceux de travaux antérieurs est la spécificité par rapport
a des écosystèmes dont les caractérisées sont
clairement identifiées. Au moment où les autres travaux donnent
des intervalles très large nous avons abouti à des valeurs plus
précises.
Figure 62. Estimation des stocks de carbones
dans différents écosystèmes du Sénégal.
(Woomer et al., 2004). Le carré rouge concerne la zone
étudiée.
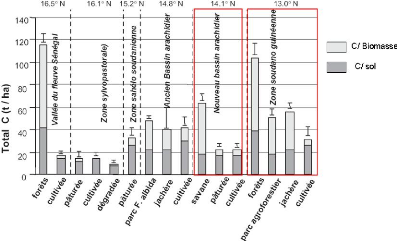
Pour ce qui est des terres avec une couverture
végétale très faible comme les jachères du bassin
arachidier, Lufafa et al. (2008) estiment des stocks de carbone
variant entre 0,035 t C/ha à 0,35 t C/ha. Dans ces
écosystèmes une importante quantité de biomasse se trouve
au niveau du système racinaire.
Section 3.07 Conclusion
L'estimation de la biomasse des individus passe le plus
souvent par l'utilisation d'équations qui estiment soit le volume soit
plus directement la biomasse. Ces équations expriment la biomasse totale
en fonction des dimensions de l'arbre, c'est-à-dire son diamètre
ou sa hauteur et plus rarement avec la surface terrière. Le besoin de
connaître la biomasse s'articule à la conception des plans
d'aménagement des forêts et l'estimation du potentiel en bois. La
satisfaction des besoins énergétiques à partir du bois
énergie requiert une connaissance des stocks de bois. La surveillance de
l'état et de la dynamique des forêts nécessite une
connaissance de la production de biomasse. Plus récemment, avec les
enjeux liés aux changements climatiques, et plus
particulièrement, le principe des MDP-forêterie, l'estimation de
la biomasse des formations forestières permet d'apprécier les
stocks de carbone et le potentiel de séquestration du carbone
atmosphérique (cf. chapitre 2).
L'observation principale qui ressort de ce chapitre est que la
structure des formations
étudiées influe très fortement
sur les stocks de carbone. Le principal élément qui
influence
les différences de stocks de carbone est le nombre
d'individus de gros diamètre. Ainsi les
différences sur le taux de couverture présente
moins d'impacts que le nombre des grands arbres.
Cette partie a permis de fournir des estimations basées
sur des données de terrain in situ qui peuvent constituer une
base de travail pour les inventaires nationaux des GES et pour les calculs de
ligne de base dans le cadre des projets MDP. Il faut toutefois noter que les
données sur les stocks de carbone des savanes sont de loin
inférieures à celles des formations denses de forêts
tropicales.
La contribution de la biomasse sur pied sur la biomasse totale
est beaucoup plus significative au niveau des formations boisées des
zones à fortes précipitations (> 800 mm/an) avec selon IPCC
(root to shoot ratio) un rapport de 0,8. Ce ratio est proche de 1 dans
les formations xériques comme celles du Ferlo par exemple. Une recherche
complémentaire devrait permettre de confirmer ces facteurs de
conversion. En termes de proportion, Lufafa et al. (2008) estiment
qu'on a près 10 fois plus de carbone dans les sols que la
végétation sur pied. Cette situation s'explique par
l'exploitation du bois des jachères, mais surtout le nettoyage des
champs avant chaque hivernage.
Les analyses montrent que le taux moyen de carbone de la
biomasse est de 49 %. L'analyse de la biomasse des savanes soudaniennes et
soudano-guinéennes ont un réel potentiel, mais les contraintes
liées aux facteurs humains et naturels font qu'une très grande
partie de la biomasse produite chaque année est perdue. Quelques
espèces à croissance rapide et qui se reproduisent bien sont le
moteur de la dynamique de carbone dans ces zones. Les projections faites
à l'échelle de projets MDP montre des avantages relativement
faibles qui pourraient décourager l'engagement dans le secteur des
projets MDP foresterie.
Concernant la dynamique du carbone, à l'absence d'autres
facteurs humains tels que les feux de brousse, la vitesse d'accumulation de la
biomasse est largement dépendante de la pluviométrie, mais aussi
de plusieurs facteurs humains de dégradation de la
végétation. Le suivi des parcelles révèle que
certaines espèces ont une croissance plus rapide (Detarium
senegalense, Bombax costatum, Anogeissus leiocarpus), mais les effectifs
d'autres espèces expliquent la contribution significative aux stocks de
carbone (Pterocarpus erinaceus, Terminalia macroptera, Combretum
glutinosum). Cependant les individus de grands diamètres (>20
cm) concentrent une large partie des stocks de carbone.
Section 3.08 Bibliographie
Breman, H. Dewit, C.T. 1983. Rangeland Productivity and
Exploitation in the Sahel. Science, 221 (4618):1341-1347.
Bronge L.B., 2004. Satellite remote sensing for estimating
leaf area index, FPAR and primary production. A literature review No. SKB
Rapport R-04-24, Swedish Nuclear Fuel and Waste Management, Stockholm, 52 p.
Brown, Pearson, S., T., Slaymaker, D., Ambagis, S., Moore, N.,
Novelo, D., Sabido W. 2005. Creating a virtual tropical forest from
three-dimensional aerial imagery to estimate carbon stocks. Ecological
Applications, 15 (3):1083-1095.
Brown S., 1997. Estimating Biomass and Biomass Change of
Tropical Forests: a Primer. (FAO Forestry Paper -134). FAO - Food and
Agriculture Organization of the United Nations, Roma, 48 p.
Brown S. 2002. Measuring carbon in forests: current status and
future challenges. Environmental Pollution, 116:363-372.
Brown, S., Gaston G. 1995. Use of forest inventories and
geographic information systems to estimate biomass density of tropical forests:
application to tropical Africa. Environmental Monitoring and Assessment
(38):157-168.
Clark, D.B., Clark D.A. 2000. Landscape-scale variation in
forest structure and biomass in a tropical rain forest. Forest Ecology and
Management, 137:185-198.
Cook, E.A. Iverson, L.R. 1989. Estimating forest productivity
with thematic mapper and biogeographical data. Remote Sensing of Environment,
28:131-141.
Coops, N.C., Waring R.H. 2001. The use of multiscale remote
sensing imagery to derive regional estimates of forest growth capacity using
3-PGS. Remote Sensing of Environment, 75:324-334.
Diouf, A., Lambin, E.F., 2000. Monitoring land-cover changes
in semi-arid regions: remote sensing data and field observations in the Ferlo,
Senegal. Journal of Arid Environment, 48:129-148.
FAO, 2008. Knowledge reference for national forest
assessment-Modelling for estimation and assessment.
http://www.fao.org/forestry/17110/en/
Roma.
Fensholt R. 2004. Earth observation of vegetation status in
the Sahelian and Sudanian West Africa: comparison of Terra MODIS and NOAA AVHRR
satellite data. International Journal of Remote Sensing, 25 (9):1641-1659.
Fensholt, R., Sandholt, I., Rasmussen, M.S., Stisen, S.,
Diouf, A. 2006. Evaluation of satellite based primary production modelling in
the semi-arid Sahel. Remote Sensing of Environment (105):173-188.
Joosten, R., Schumacher, J., Wirth, C., Schulte A. 2004.
Evaluating tree carbon predictions for beech (Fagus sylvatica L.) in
Western Germany. Forest Ecology and Management (189):87-96.
Kessler, J.J., Breman, H. 1991. The Potential of Agroforestry
to Increase Primary Production in the Sahelian and Sudanian Zones of West
Africa. Agroforestry Systems, 13 (1):41-62.
Lufafa, A., Bolte, J., Wright, D., Khouma, M., Diedhiou, I.,
Dick, R.P., Kizito, F., Dossa, E., Noller J.S. 2008. Regional carbon stocks and
dynamics in native woody shrub communities of Senegal's Peanut Basin.
Agriculture, Ecosystems and Environment (In press).
MEA, 2005. Rapport de synthèse de l'Évaluation
des Écosystèmes pour le Millénaire, Millenium Ecosystem
Assessment, 59 p.
Sambou B., 2004. Evaluation de l'état, de la dynamique
et des tendances évolutives de la flore et de la
végétation ligneuse dans les domaines soudanien et
sub-guinéen au Sénégal. Doctorat d'Etat, Université
Cheikh Anta Diop de Dakar (UCAD), Dakar.
Sambou B., Madsen, J.E., Goudiaby, A., Sonko, I., Camara,
A.A., Lykke, A.M., Mbow, C., et Bâ, A.T. 2007. Mise en place et suivi des
parcelles permanentes pour une bonne connaissance de la flore et de la
végétation des zones de savanes. Rapport, ISEENRECA. 33P.
Seaquist, J.W., Olsson, L., Ardö J. 2003. A remote
sensing-based primary production model for grassland biomes. Ecological
Modelling (169 ):131-155.
Skutsch M.M. 2005. Reducing carbon transaction costs in
community based forest management. Climat Policy (5):433-443.
St-Onge, B., Jumelet, J., Cobello, M., Véga C. 2004.
Measuring individual tree height using a combination of stereophotogrammetry
and lidar. Canadian Journal of Forest Research (34):2122-2130.
Vanclay, J.K., Skovsgaard J.P. 1996. Evaluating forest growth
models. Ecological Modelling (98 ): 1-12.
Vries, F.W.T., Krul, J.M. Keulen, H. 1980. Productivity of
Sahelian rangelands in relation to the availability of nitrogen and phosphorus
from the soil. Nitrogen cycling in West African Ecosystems:95-113.
Woomer, P.L., Tieszen, L.L., Tappan, G., Touré, A., Sall
M. 2004. Land use change and
terrestrial carbon stocks in Senegal. Journal
of Arid Environments (59):625-642.
i)
ii)
iii)
iv)
v)
vi)
vii)
viii)
ix)
x)
xi)
xii)
xiii)
xiv)
xv)
Article IV.
Article V. CHAPITRE IV :
Article VI. Modélisation de la dynamique du
carbone
|
i) Ce chapitre fait le point sur les modèles
forestiers utilisés pour analyser la dynamique des formations
végétales. Les modèles existants sont très
diversifiés (écosystémiques, déterministes,
biogéochimiques, empiriques)
et prennent difficilement en compte le facteur humain.
Cependant, parmiceux-ci, CASS (Carbone Accounting Simulation
Software) est le plus
flexible puisqu'offrant une relative transparence pour
un paramétrage des
scénarios à tester. C'est ainsi que
les résultats obtenus à partir de ce
modèle montrent
des tendances variées selon la nature et la force des
|
facteurs en jeu (coupe, feux de brousse, pâturage,
jachère, etc.) dans les savanes étudiées.
Section 6.02 Introduction
i) La dynamique du stock de carbone telle
qu'analysée dans les chapitres précédents est basée
sur des données collectées in situ ; celles-ci ont alors
permis une description des caractéristiques de distribution et de
dynamique dans le passé. Dans le présent chapitre il sera
question d'introduire la dimension temporelle par un essai de projection de ces
dynamiques dans le futur. Le devenir des formations forestières est
souvent approché par des analyses prospectives utilisant les
modèles forestiers fondés sur un certain nombre
d'hypothèses, de suppositions, de pratiques des populations, de
scénarios climatiques, etc. L'application judicieuse de ces
modèles peut donner un aperçu du devenir et du fonctionnement des
écosystèmes forestiers dans le futur.
Les premiers essais de simulation sur les forêts ont
porté sur la croissance des arbres et sont basés sur des analyses
statistiques de données d'inventaires. Ces modèles de
première génération partent de l'hypothèse que les
conditions climatiques ne changent pas radicalement dans le futur. D'autres
écologistes de la fin du 19ième siècle et du
20ième siècle se sont intéressés aux
mécanismes fonctionnels de formations forestières pour asseoir
des théories sur la succession des espèces, sur la dynamique de
la composition floristique par des approches phytosociologiques et sur les flux
de matière et d'énergie dans l'écosystème. Les
travaux de Ramade, F. en 1984 et 1989, intitulés <<
Eléments d'écologie appliquée et fondamentale »
constituent de bonnes synthèses de cet héritage
écologique. Dans la même veine, une série d'ouvrages de
base ont tenté de reconstituer les mécanismes de fonctionnement
des écosystèmes pour pouvoir en anticiper les manifestations. On
peut citer les travaux de Duvigneaud, P. sur la << synthèse
écologique » en 1980 ; l'ouvrage de Fischesser, B., portant sur
<< la vie des forêts » en 1970 ; ou l'ouvrage classique de
Guinochet, M., sur << la logique et dynamique du peuplement
végétal. Phytogéographie ; Phytosociologie » en
1973.
Ces tentatives ont été le plus souvent quasi
théoriques et moins quantitatives. Les théories de la croissance
des plantes ont introduit une notion essentielle qui est celle de climax. Cette
notion est intéressante dans la perspective des changements climatiques,
car elle traduit un état d'équilibre entre la productivité
de l'écosystème et la consommation totale d'énergie
permettant de maintenir le système. Une forêt à
l'état climacique peut donc stocker une importante quantité de
carbone, mais du point de vue de sa dynamique de séquestration, elle
peut présenter un bilan négatif. De telles formations deviennent
alors des sources plutôt que
des puits. Ainsi, avec les MDP, les notions de succession
primaires (sans perturbation) ou secondaire (avec perturbation) constituent une
base importante de l'anticipation de la dynamique des formations
forestières aménagées ou non.
La façon dont cette notion de succession a
été utilisée est cependant apparue trop simplifiée
selon certains auteurs comme Waring et al. (1998) qui
préfèrent la notion de « dynamique de la
végétation ». Ce changement de concept permet de
considérer bien plus que la dimension biodiversité et introduit
dans les projections le rôle des sols, du climat, des perturbations
humaines ou naturelles, etc. Ce changement de paradigme s'est accompagné
de mutations profondes sur les approches développées. La plupart
de ces modèles sont dénommés modèles
biogéochimiques quand ils simulent les cycles du carbone et de l'azote.
A partir de 1995, plusieurs modèles biogéochimiques sont
basés sur des approches mécanistiques, déterministes,
analytiques, empirique ou stochastiques.
Les modèles mécanistiques
désignent les modèles qui ont une structure qui représente
de façon explicite les processus physiques chimiques et biologiques.
Cette catégorie de modèle procède par une description
quantitative entre la croissance de biomasse dans les écosystèmes
et les principaux facteurs structurants. Ils sont alors théoriquement
utiles pour inférer des informations, même en dehors des zones
où initialement ils ont été élaborés.
Les modèles déterministes
donnent quant à eux une solution unique utilisant des variables
d'état de l'écosystème. Ces modèles ne simulent pas
explicitement les effets de l'incertitude des données ou leur
variabilité dans les résultats de simulation. Les variations du
résultat de ces modèles sont exclusivement dues aux modifications
dans les composantes du modèle.
Les modèles analytiques
désignent la catégorie dans laquelle les prédictions
peuvent être résolues sur la seule base de calculs
mathématiques. La plupart de ces modèles, basés sur des
calculs différentiels relativement simples, permettent de calculer le
résultat de l'interaction quantitative de plusieurs facteurs.
Certains modèles sont dits empiriques
du fait que leur structure est déterminée par les relations
observées sur des données expérimentales. Ils sont alors
souvent utilisés pour reconstituer les relations fonctionnelles entre
facteurs pouvant permettre la prévision et la description des tendances
mais uniquement au niveau des sites pour lesquels ils ont été
conçus.
La dernière catégorie de modèles qui fait
fortune en ce moment au sein de l'écologie moderne est dite
modèle stochastique. Ce sont des modèles
dynamiques qui prennent en compte la variabilité dans les
paramètres du modèle. Cette variabilité fait
référence aux différences aléatoires dues à
l'hétérogénéité des données. Cette
variabilité peut être due aux modifications des conditions
environnementales, aux variations temporelles et spatiales ou à de
simples variations aléatoires sur les mesures in situ de base.
Les résultats de ces modèles sont fonction des composantes des
modèles et de la variabilité aléatoire des
paramètres. Leur mise en application au niveau des
écosystèmes est jugé pertinente dans la mesure où
les processus écologiques des formations forestières sont assez
souvent erratiques, avec des variations spatiales et temporelles très
dynamiques.
Les théories actuelles de la modélisation des
écosystèmes considèrent les perturbations tels que les
feux de brousse, les coupes rases, les crises climatiques comme des
évènements centraux dans l'évaluation de leur dynamique.
Il faut alors bien caractériser l'effet de ces perturbations en termes
d'ampleur, de durée et de fréquence. Aussi, les réponses
naturelles des écosystèmes sont-elles prises en compte dans les
modèles (successions, résistances, compétition, etc.).
Il faut toutefois noter, même si les spécialistes
parviennent avec aisance à reconstituer l'historique des
écosystèmes du fait de l'existence de bases de données
assez complètes par endroits (longues séries d'inventaire de
végétation, données de paléoécologie, de
dendrochronologie, de sédimentologie), qu'il n'en demeure pas moins
difficile à l'heure actuelle de bien caractériser le devenir des
écosystèmes à cause de plusieurs incertitudes liées
aux trajectoires des changements climatiques et aux dynamiques
socio-économiques.
Pour mieux comprendre la logique des modèles forestiers,
il faut faire un bref rappel des fondamentaux sur la croissance des formations
forestières.
Section 6.03 IV.1. Les fondamentaux
écologiques
Pendant le processus de croissance, l'arbre modifie son milieu
et utilise les ressources disponibles pour son développement, parfois au
détriment d'autres plantes proches. La compétition entre les
arbres est un facteur important qui au fil du temps impose une certaine
structure et une composition des espèces dans un milieu donné. La
compétition est plus rude quand le milieu est monospécifique (cas
des plantations), puisque les plantes ont les mêmes besoins et
développent les mêmes stratégies ou réponses aux
différentes contraintes. Dans des milieux plus diversifiés sur le
plan de la composition floristique, les capacités d'accès aux
ressources varient en fonction des plantes du fait des différences dans
les systèmes racinaires (profondeur, distribution), et des besoins en
nutriments variables selon l'espèce (Waring et Running, 1998). Au niveau
des écosystèmes, la compétition et les processus
biophysiques qui se traduisent par une certaine structure de la
végétation influent aussi directement sur les cycles des
nutriments, du carbone et de l'eau.
Cependant, le développement d'une formation
végétale ne peut s'appréhender qu'à travers des
données historiques sur la composition et la croissance des
espèces. L'obtention de ces types de données fait l'objet de mise
en place dans différents écosystèmes, de dispositifs
adaptés comme les parcelles de suivi de la végétation
ligneuse (voir chapitre 3). Ces parcelles de suivi permettent non seulement de
suivre les phénomènes de succession et la diversité
biologique, mais constituent une bonne base pour évaluer les stocks de
carbone de la biomasse sur pied utilisant les régressions
allométriques. Mais pour arriver à reconstituer le cycle de
croissance d'une formation forestière, il faut recourir à des
modèles dynamiques qui permettent de prédire la croissance des
arbres et les modifications de sa composition pendant plusieurs dizaines
d'années.
En reprenant les étapes essentielles du
développement d'un écosystème forestier, Waring et Running
(1998), suggèrent 4 étapes suivant une dégradation totale
des arbres dans un milieu :
- début de la production de biomasse avec des
espèces pionnières (initiation, 1) - remplacement des
espèces pionnières par d'autres espèces (exclusion, 2)
- développement de la régénération
naturelle des espèces présentes (réinitiation, 3)
- maturité de la formation avec la présence
d'individus au niveau de tous les étages (4)
(grands arbres, arbres moyens,
régénération), figure 63.
Figure 63. Les 4 principales étapes de
développement d'une formation forestière ; adaptée de
Waring et Running (1998)
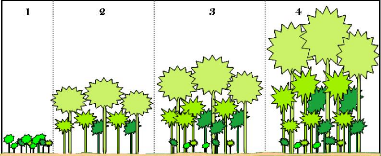
Au début, c'est-à-dire l'étape
d'initiation (1), on a la mise en place des espèces pionnières
(1). Avec la croissance des arbres et le développement de nouvelles
espèces à travers un processus d'exclusion lié à la
compétition, les espèces pionnières disparaissent (2). Les
arbres devenus grands favorisent le développement de leur propre
régénération naturelle (3). L'étape
d'équilibre est atteinte à la maturité de
l'écosystème caractérisée par la présence de
toutes les strates (4).
Ces différentes étapes correspondent à
des réponses fonctionnelles qui mènent toutes à la
production de biomasse et à son accumulation dans différents
réservoirs de l'écosystème forestier. À l'heure
actuelle, les dynamiques des savanes sont étudiées en se basant
sur la structure des classes de diamètres. Une population en bon
état est celle qui n'est pas trop perturbée avec par
conséquent une représentation des différentes classes de
diamètre (Sambou, 2004). La disparition d'une classe de diamètre
est un indicateur d'une perturbation à un moment donné. En
foresterie, on utilise un indicateur plus général appelé
la `qualité du site' (Waring et Running, 1998) ; laquelle est
estimée par un indice de site `SI', représente la hauteur des
individus dominants à un âge donné. Par exemple, SI(30)= 20
signifie que les individus dominants atteignent normalement la taille de 20 m
à l'âge de 30 ans. Si à l'âge indiqué, les
grands individus n'atteignent pas la moyenne de hauteur correspondante, le site
présente alors des signes de dégradation. Pour cet indice il faut
donc des données de suivi à long terme du genre collectées
au niveau des parcelles de suivi de la végétation ligneuse.
i) L'un des grands défis de la
modélisation est la prise en compte des réponses des
écosystèmes (rétroactions) aux perturbations naturelles ou
humaines (sécheresse, feux de brousse par exemple) et aux effets de
certaines interventions comme une fertilisation additionnelle. Ainsi,
la
simulation, dont les bases proviennent de stations
expérimentales a été une voie largement explorée en
foresterie pour reconstituer la dynamique des formations
forestières.
Section 6.04 IV.2. Etat des connaissances sur les
modèles de productivité des écosystèmes
forestiers.
Les modèles de production forestière sont l'un
des plus anciens types de modèle et remontent au
19ième siècle avec les premières tables de
correspondance qui cherchaient à prédire la production en
fonction de paramètres dendrométriques. Actuellement, la
révolution informatique permet de simuler la production des individus ou
des formations végétales.
Les indices sur le site comme la hauteur dominante ou
l'âge sont le plus souvent relevés. La plupart de ces
modèles cherchent à tenir compte de la relation complexe entre la
densité des arbres et leur croissance en diamètre et en hauteur.
Ce type de modèle est largement appliqué à
l'échelle de l'individu et certains chercheurs prennent en compte la
mortalité pour mieux affiner leur prévision.
i) Pendant les trente dernières années,
plusieurs modèles ont été développés pour
analyser différents aspects du cycle du carbone y compris la dynamique
de la végétation en relation avec les stocks de carbone.
L'estimation de la PPN (Productivité Primaire Nette) est au centre de la
plupart de ces modèles. Du fait des limites techniques et des
contraintes environnementales (facteurs et mécanismes terrestres), les
approches ont été diversifiées. Les analyses les plus
simples ont exploré les relations linéaires empiriques entre la
PPN et des paramètres telle que la température, les
précipitations utiisées dans le modèle de Miami. A l'autre
extreme, nous avons les modèles biochimiques comme TRIFFID et BIOME3
plus complexes (modèles de processus).
Ces modèles biogéochimiques ont
été testés en comparant des résultats de simulation
à des paramètres observables comme la croissance ligneuse sur une
période de temps relativement courte (~ 5 ans). Cette adéquation
entre les résultats des simulations et la croissance des arbres, suppose
une bonne évaluation de la photosynthèse, de la respiration, de
l'allocation de carbone dans les différentes parties de l'arbre et du
processus de renouvellement des tissus (Waring et Running, 1998).
ii)
Le défi lié aux modèles forestiers
réside en général sur leur capacité à bien
reproduire les perturbations des écosystèmes. Ces perturbations
étant complexes et non linéaires. Les différences dans la
plupart des modèles résident dans l'élaboration de
sous-modèles introduisant l'utilisation de feux, de coupes de bois ou
d'une longue sécheresse.
iii)
iv) Malgré leurs différences, les
résultats obtenus à partir de ces modèles sont
relativement proches sur les tendances, mais divergent sur l'ampleur des
modifications de biomasse. Il est relativement difficile de détailler
les différences entre les modèles. Un des problèmes
majeurs de la modélisation est l'harmonisation des paramètres
utiisés et la standardisation des variables tels que l'humidité,
la vitesse du vent, le type de sol, la distribution de la
végétation, etc., (Adams et al., 2004). D'autres
différences dans l'approche sont aussi notées sur la
caractérisation des types de sols et des catégories de
végétation, notamment sur la nomenclature et les critères
retenus à cet effet. Aussi, les modèles utilisent-ils un taux de
carbone global qui varie entre 280 ppmv à 360 ppmv. Ce qui a
entraîné des différences dans la prédiction qui
varient de 40 - 80 GtC par an. Beaucoup d'incertitudes accompagnent ainsi ces
résultats à cause des variations interannuelles, des
données secondaires lacunaires (sur le plan spatial et temporelle) ou
dispersées.
La comparaison des modèles ne doit pas se limiter donc
à la confrontation des résultats, mais considérer aussi
l'examen de leur structure interne pour voir les forces et les faiblesses des
sous-modèles en particulier, ceux qui permettent de
générer la PPN des écosystèmes.
Selon la structure, il existe deux grands groupes de
modèles : les modèles biogéographiques
(corrélation) et les modèles biochimiques (basés sur des
bilans de carbone). Les deux sont souvent désignés sous le terme
de modèle biogéochimique. Les modèles
biogéographiques comme BIOME et ALBIOC utilisent un quadrillage pour
renseigner les variables du climat pour chaque cellule de la grille afin de
déterminer le nombre et les types de plantes qu'on peut y rencontrer.
Les biomes et les écosystèmes sont ainsi présentés
comme des combinaisons particulières de types de plantes. Les
modèles biochimiques (exemple : TRIFFID) quant à eux,
considèrent une constante distribution des types de plantes. Les
processus biochimiques comme la photosynthèse et la respiration sont
simulés en fonction du climat local, du taux de CO2
atmosphérique, de la radiation photosynthétique active, de
l'humidité du sol et des précipitations. Les
modèles biochimiques peuvent simuler les changements de la
densité et de la productivité végétale. Le tableau
19 présente une synthèse non exhaustive des modèles
existants.
Tableau 19. Synthèse sur quelques
modèles utilisés pour la simulation de la croissance des
végétaux. Source Adams et al. (2004), modifiée
par cette étude.
|
Types de modèle
|
Description
|
Avantage
|
Limites
|
1. Modèle de Miami (Leith, 1975)
|
Empirique
Biogéographique
Basé sur les types de plante
|
~Bonne estimation de la PPN
|
Limites sur la prédiction de la PPN
Ne considère pas les changements de la
végétation
|
2. Modèle dit `n- factor'
(Post et al., 1997; King et al., 1997. )
|
Combinaison de l'approche empirique et celui dit du `n-factor'
en ajoutant un facteur de fertilisation en C sur le modèle 1 ; 30 types
fonctionnels de plantes sont considérés sur des cartes de
distribution. Allocation proportionnelle de C dans les différentes
parties de la plante (F, B, R).
|
~Bonne estimation de la PPN
Introduction d'un facteur K1 (0.6) pour pondérer la PPN
par le taux de fertilisation en carbone.
|
Ne considère pas la photosynthèse C4
La signification du facteur K1 n'est pas expliquée
|
3. Modèle dit `n- factor'
(Polglase et Wang, 1992)
|
Biogéochimique
Plusieurs types de végétation 10 biomes, le climat
et la
productivité caractéristique sont
considérés. Pondération par un facteur de croissance qui
prend en compte la fertilisation en C.
|
Prend en compte les changements de densité de la
végétation.
|
Problème sur le facteur de croissance biotique
Difficile d'utiliser ce modèle pour des endroits à
température saisonnière très variable
|
4. Modèle dit `n- factor'
(Lenton, 2000)
|
Quasi-biogéochimique
La photosynthèse= f (température et du C
atmosphérique)
La respiration = f(température et teneur en C de la
végétation).
|
~Bonne estimation de la PPN pour les climats avant
l'ère industrielle.
|
La biochimie de la plante n'est pas modélisée
explicitement. Absence de lien entre la photosynthèse et la teneur en C
de la plante. Une constante photosynthèse même si la
végétation est différente.
|
5. (Svirezhev et Bloh, 1997; 1998)
|
Quasi-biogéochimique
Similaire à 4, mais le potentiel maximum de PPN est pris
en compte en introduisant la réponse à la température, le
stress
hydrique, la fertilisation et la compétition
intraspécifique.
|
Utilisable pour une investigation
qualitative.
|
Limite sur la confusion
entre la fertilisation en CO2 et la compétition,
utilisation prudente.
|
5. Modèle TRIFFID -Cox (2001)
Simple
|
Biochimique-biogéographique (hybride). Les changements
sur la densité et la distribution de la végétation, la
photosynthèse et de la respiration. Pondération par le stress
hydrique ; prise en compte des plantes C4 ; types de
végétation considérés.
|
Relativement bon, et prend en compte la dynamique biochimique et
physiologique.
|
TRIFFID-Cox. Très complexe à utiliser, le
modèle simple est plus abordable mais pèche par la
considération d'un seul type de végétation
|
6. DEMETER
|
|
Quasi-biogéochimique- biogéographique. 16 types
fonctionnels de plantes. Le taux potentiel de photosynthèse est
calculé en modifiant la photosynthèse maximum par un facteur
linéaire qui traduit la réponse à la température.
La respiration et la photosynthèse C4 sont prises en compte.
|
???
|
Température constante, absence du facteur de fertilisation
en CO2 ni la compétition entre plantes
|
|
|
Types de modèle
|
Description
|
Avantage
|
Limites
|
7. BIOME 3 (Prentice et al.,
1992; Haxeltine et
Prentice, 1996a; b)
|
Biogéochimique-biogéographique (hybride). 7 types
de végétation. Distribution des plantes = f(limite thermique
maximum) ; prévoit les herbes dans tous les écosystèmes.
Calcul la photosynthèse f(l'éclairement, Rubisco, conductance
stomatale). La photosynthèse C4, la phénologie en fonction de la
température et du stress hydrique, la compétition
|
PPN optimisé en
fonction du LAI.
|
Modèle très complexe, les types de
végétation sont différenciés par un seul
paramètre et la dimension physiologique domine.
|
8. CENTURY 5.437 (Woomer et al.,
2001)
|
Modèle Ecosystème : COS, MOS ; Croissance des
plantes (herbacées ou ligneux) ; NPP ; C=f(température, pmm, type
de sol, utilisation des terres). Sous- modèle de croissance des plantes.
Tient compte du rapport C/N et de la décomposition de la M-O. Prend en
compte N, P et S en plus du C.
|
Très large utilisation, différents
écosystèmes, optimisé pour différents facteurs
biotiques et abiotiques. Adapté aux savanes.
|
Beaucoup de paramètres, complexe, modèle plus
adapté pour les sols et la végétation.
|
9. 3PG (CSIRO) (Landsberg et Waring, 1997)
|
Modèle Biophysique. Simulation mensuelle utilisant la
moyenne de la RPA, moyenne journalière du déficit hydrique,
températures extrêmes, précipitations
mensuelles, capacité de rétention en eau du
sol.
|
Simule la PPN, la biomasse sur pied et dans le sol.
|
Requiert des informations
pas facilement accessibles.
|
10. CASS (CRC) (Roxburgh, 2004)
|
Modèle déterministe. Simulation de carbone des
écosystèmes en tenant compte de la PPN et des facteurs de
perturbation, feux, coupe, changement occupation / utilisation des terres.
|
Flux de carbone dans l'écosystème et
libération CO2 dans l'atmosphère.
|
Facile, transparent et flexible.
|
11. PNET (Aber et al., 1996)
|
Modèle biogéochimique sur la dynamique
forestière.
|
Flux de carbone, de nitrate dans les écosystèmes
forestiers.
|
Les écosystèmes du modèle sont de type
tempéré et boréal.
|
12. DNDC (EOS, UNH)
|
Modèle déterministe.
|
Flux de carbone et d'azote en milieu agricole et sols
cultivés.
|
Ne s'applique qu'en milieu agricole.
|
13. SARRA-H (Baron et al., 1996)
|
|
Modèle empirique.
|
Simulation de la croissance des plantes et de la
productivité des cultures.
|
Ne s'applique qu'en milieu agricole.
|
|
NB. COS (carbone Organique du Sol), MOS (Matière
Organique du sol), (Productivité Primaire Nette), C/N (Rapport carbone
azote), N (Azote), P (Phosphore), S (soufre), C (carbone),
Il est difficile de dire qu'un modèle est meilleur
qu'un autre. Un modèle doit être utile en fonction des objectifs
qu'on se fixe et de la dose de subjectivité et d'imprécision dans
le processus de modélisation. Le cycle de vie d'un modèle est le
suivant : les objectifs initiaux déterminent sa structure, les objectifs
se précisent, s'étendent et la structure initiale s'y adapte,
etc. Les objectifs et la structure ont des relations hiérarchiques
puisque le système a un contexte (haut, objectifs) et des
mécanismes (bas, structure). L'essai d'évaluation des
modèles dans les paragraphes qui suivent, n'est pas une
appréciation technique, mais plutôt une analyse de leur
portée opérationnelle.
Section 6.05 IV.3. Utilisation des modèles et la
gestion durable
des forêts
Le débat actuel sur la gestion durable des ressources
forestières a beaucoup influencé l'exploration des modèles
pour simuler des options d'aménagement et des choix politiques pouvant
influencer la dynamique spatiale et temporelle des formations
végétales. Le rôle actuel et futur des ressources
forestières est devenu par conséquent, une question importante
pour sa biodiversité, son importance économique,
énergétique et son effet sur le climat. La conservation durable
des ressources forestières est très difficile à
définir du fait de la nécessité de croiser plusieurs
indicateurs parfois pas simples à quantifier. La durabilité
suppose que la productivité et l'intégrité de la
forêt soient bien conservées. Ce concept de durabilité est
conditionné dans ce cas par deux échelles d'analyse qui
interagissent : le temps (quelle période considérer ?) et
l'espace (quelle taille de forêt prendre en compte ?), (Monserud,
2003).
Avant les années 1990, la plupart des pays ont
géré leurs ressources forestières sur la base d'un
maintien de la production (biomasse totale), en mettant en second plan la
productivité (renouvellement) de la forêt. Le choix du
développement forestier durable dépend en général
de ce qu'on cherche à maintenir durablement et le plus souvent, ce choix
est orienté vers des besoins utilitaires. Ainsi, le concept de
durabilité pris dans son sens socio-économique ne peut se
concevoir sans que l'environnement (les ressources naturelles) qui en est la
base ne soit durable. Cette conception permet de prendre en compte d'autres
fonctions de la forêt pour intégrer l'état des formations,
leur intégrité, et leurs relations fonctionnelles avec les autres
éléments biophysiques du milieu, tout en jouant son rôle
économique, écologique, social et culturel pour les
générations présentes et futures. Cette définition
multidimensionnelle est très utile mais ne mène pas à une
possibilité de quantification surtout pour les petites
échelles.
Le passage de la production optimale à la production
durable a été stipulé par le rapport Brundtland sur le
développement durable en 1987 (Commission Mondiale pour l'Environnement
et le Développement) et consolidé lors du Sommet de Rio en 1992
avec l'adoption des Principes de la Forêt. Depuis, plusieurs initiatives
ont été prises comme les accords de Montréal, Kyoto,
Conventions sur la Biodiversité, sur la Désertification, etc. Un
accent particulier a été mis sur les critères et les
indicateurs pour évaluer la durabilité de la gestion
forestière à différentes échelles. Il faut
toutefois noter que, les indications de durabilité forestière
sont souvent appréciées à un niveau national alors que la
gestion de la ressource se fait au niveau de la formation, donc à une
échelle locale. Ce décalage entre
l'échelle de conception des politiques et les
échelles de prise de décision pratiques n'a pas permis une
gestion adéquate des formations forestières. Un exemple souvent
évoqué au Sénégal est la distribution des quotas
d'exploitation qui échoit souvent à des citoyens externes aux
terroirs d'exploitation (Ribot, 1998; 2002). C'est à cette
dernière échelle d'ailleurs que la plupart des modèles
forestiers ont été développés. Ainsi, le recours
aux modèles permet d'anticiper des processus et d'appuyer la prise de
décision par rapport aux prévisions les plus vraisemblables.
Ces éléments ont beaucoup influencé le
choix du modèle utilisé dans ce travail. Nous n'avons pu obtenir
que les modèles CENTURY, CASS, PNET, DNDC et SARRA-H. Les deux derniers
ont été exclus parce que ce sont des modèles de croissance
des essences agricoles. On a alors procéder à des tests des
différents autres modèles. Le modèle CENTRURY
(dernière version) simule à la fois le carbone et les flux
biogéochimiques. Les données d'entrée sont très
nombreuses et le plus souvent des valeurs par défaut sont
utilisées sans qu'on ait un contrôle sur la signification de ces
valeurs pour les écosystèmes de savane. Le modèle PNET
développé par l'Université de New Hampshire (USA), ne
dispose pas d'écosystèmes tropicaux et donc pas de formations de
savanes. De surcroit, les essais d'ajustement des types de forêts
développés dans le modèle ne permettent pas de rendre
compte de ce qui se passe dans les savanes. Seul le modèle CASS-1.2
(Carbone Accounting Software), Roxburgh (2004),
développé par le CRC (Australie) prend en compte les savanes et
les processus de perturbation comme les feux, les coupes, les transformations
d'utilisation des terres les écosystèmes tropicaux. Ce
modèle a été utilisé par conséquent pour
simuler selon les cas, quelques scénarios de modification sur les
savanes permettant de caractériser le devenir du carbone.
Section 6.06 IV.4. Modélisation des flux de
carbone avec le modèle CASS
La possibilité de modéliser les flux de carbone
dans des écosystèmes est un aspect central dans l'étude
des changements climatiques. Le modèle CASS permet de faire de la
simulation des flux de carbone dans les écosystèmes terrestres
majeurs. Les écosystèmes de savane y sont bien
représentés avec une possibilité de définir les
caractéristiques des écosystèmes particuliers ou des
variations subtiles qui se traduisent par une modification du bilan du carbone.
Le modèle prend en compte la litière, la dynamique du carbone
dans le sol, et le carbone restant des produits ligneux récoltés
(fabrication de papier, bois de chauffe, de construction, etc.). Dans ce
modèle, le carbone atmosphérique est fixé à travers
la photosynthèse et redistribué au niveau des feuilles, des
rameaux et des racines, une partie étant restituée sous forme de
respiration autotrophe. Les différentes portions du carbone
absorbé sont progressivement transférées
au sol à partir d'où elles peuvent rejoindre l'atmosphère
à travers la respiration hétérotrophe. La croissance des
plantes est fonction de la quantité de carbone qu'elles peuvent absorber
de l'atmosphère pour effectuer la photosynthèse. Ce taux
d'absorption du CO2 varie en fonction de la quantité de CO2 disponible,
des nutriments, de la température, de l'eau, de l'azote d'origine
atmosphérique, des types de formation, etc. La décomposition suit
une fonction exponentielle simple en relation avec la température et la
disponibilité des ressources en eau. L'effet des perturbations
liées aux feux de brousse et l'exploitation du bois peuvent être
pris en compte dans le modèle de la même façon que les
facteurs de modification du couvert végétal comme les changements
d'utilisation et d'occupation du sol.
(i) Il est très important aussi que les modèles
qui permettent d'établir le bilan de carbone soient basés sur des
processus physiologiques, plutôt que sur une présomption
d'homogénéité des formations végétales. En
effet, durant les prochaines décennies, les arbres seront exposés
à des changements non seulement de la concentration atmosphérique
de CO2 mais aussi peut-être à des variations extrêmes de
température ou de pluviométrie. Bien que des études
à court terme aient statué sur les effets de l'augmentation de
CO2 sur la croissance des formations forestières, il demeure que la
réponse des arbres et des forêts à des concentrations
élevées de CO2 et aux changements climatiques qui y sont
potentiellement associés reste à définir.
(b) IV.4.1. Description du modèle CASS
Le principe du modèle CASS est que les
écosystèmes terrestres peuvent être répartis en gros
dans trois réservoirs séparés : 1) le réservoir de
carbone de la végétation vivante, 2) le réservoir de
carbone de la végétation morte (litière), 3) le
réservoir de carbone du sol. Le modèle prend aussi en compte un
réservoir 4, constitué du carbone dans l'atmosphère, mais
juste pour voir son effet de fertilisation dans le processus de
photosynthèse. Des sousréservoirs peuvent être
définis dans chacun de ces grands groupes (figure 64).
Figure 64. Structure générale
simplifiée du modèle CASS

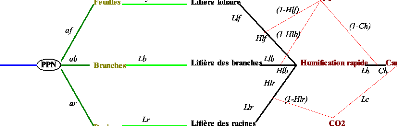
La principale entrée du modèle est le carbone
atmosphérique fixé par les plantes (PPN, gC/m2/an) et
les principales sorties sont le stock de carbone des différents blocs et
sous-blocs et le CO2 libéré dans l'atmosphère. Les
paramètres (af, ab, ar) sont les portions de carbone réparties
entre les feuilles, les branches et les racines ; af + ab + ar = 1. Le taux
à travers lequel le carbone de la végétation vivante est
transféré à la litière est défini par la
durée moyenne d'existence du carbone au niveau des feuilles, des
branches et des racines (Lf, Lb, Lr). Pour exemple, si Lf est de 5 ans, alors
1/5ième du carbone total des feuilles est perdu pour la
litière foliaire pour le même espace de temps.
Une fois que le carbone se trouve dans la litière, il
peut se produire trois choses : le carbone
Végétation Litière Sol
peut y rester jusqu'à l'année suivante, il peut
se retrouver dans l'humus du sol, la litière peut se décomposer
pour libérer le carbone dans l'atmosphère. L'humus peut subir une
décomposition et libérer le carbone ou le convertir en carbone
stable du sol.
Lf CO2
Flls Litièe fiaie
Les bases mathématiques du modèle combinent 8
équations différentielles simultanées et
af (-Hb)
Hlf
chaque équation décrit une des huit dynamiques
des 8 blocs de carbone. Ces équations
b Lb Llb
N nc p
traduisent la vitesse à laquelle le carbone des
différents blocs change dans le temps. Pour
Hlb Lh Ch
calculer le stock de carbone, ces équations sont
intégrées par le modèle utilisant l'algorithme
1
de 4ième ordre de Runge Kutte avec un
contrôle au pas de simulation.
a Llr
- Au niveau de la végétation
vivante
dC feuille, branche, racine/dt : traduit la vitesse
à laquelle le carbone passe des feuilles, branches, racines à la
litière.
- Au niveau de la litière
C_litière-feuille/L_feuille : traduit le taux
de transformation du carbone de la litière vers le
bloc humus. Le
second terme
`h_litière-feuille*C_litière-feuille/L_litière-feuille'
est la quantité
C C C C C
de carbone de la litière foliaire
décomposée, c'est-à-dire la portion qui retourne dans
? h ? h C ? (1 C
re felle
? lire branche
? litière racine
? hums hums )
L L L L L
l'atmosphère. Il en est de même pour les branches et
les racines.
- Au niveau du sol
Les trois premiers termes de l'équation traduisent les
gains en carbone à partir de trois
b
sources de litière. Le quatrième terme est une
conversion du carbone de l'humus du sol et le dernier exprime la
décomposition du carbone de l'humus du sol.
Le carbone fixé au sol s'exprime comme suit :
Quand une simulation est faite sur une zone avec une faible
végétation qui croît dans le temps, la PPN augmente
à un taux de 1,05 et la courbe prend une forme sigmoïdale (forme de
S) rendue par la formule suivante :
Le terme a contrôle la vitesse de l'augmentation de la
PPN.
Dans ce modèle on peut inclure des modifications de
variables climatiques comme la température, le CO2 atmosphérique,
l'humidité du sol, la fertilisation du sol, le rythme de la
décomposition de la matière organique, etc. Le poids des blocs
est fonction du taux de fixation et de libération du carbone
Le principe de modélisation de la dynamique du carbone
est basé sur le bilan entre la quantité de carbone qui entre dans
le système par le processus de la photosynthèse et celle qui en
sort par la consommation des herbivores, les pertes de feuilles, etc.
Ces équations de base permettent de comptabiliser les
flux de carbone entre les trois réservoirs terrestres que sont la
biomasse vivante, la litière et le carbone du sol. A partir de la
synthèse des plantes qui fixent le CO2, la respiration autotrophe et
hétérotrophe est déduite des quantités de carbone
fixées ; ensuite le modèle prend en compte les perturbations de
différente nature : feux de brousse, coupe, changement d'utilisation des
terres, changement d'occupation des sols, etc. Pour chacun de ces processus, la
modification des stocks et des flux de carbone dépend de la durée
du phénomène et de son ampleur. Les principales composantes du
modèle sont montrées à la figure 65.
Figure 65. Diagramme logique du modèle
CASS (Roxburgh, 2004)
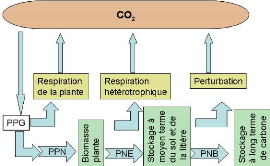
PPG (Productivité Primaire Globale) ; PPN
(Productivité Primaire Nette) ; PNE (Productivité Nette de
l'Ecosystème) ; PNB (Productivité Nette du Biome).
Le modèle CASS permet alors d'observer les grands
processus qui interagissent pour caractériser les trajectoires du
carbone dans les écosystèmes terrestres. Aussi, dans un souci de
transparence et de simplicité, ce modèle accorde une grande
importance à la description simple des processus écologiques.
Cependant ses deux principales limites sont :
- qu'en conditions réelles, les cycles de carbone de l'eau
et des nutriments évoluent en
parallèle et interagissent ; les autres cycles agissant
comme des facteurs extérieurs
(facteurs de modification des scénarios `environmental
modifiers') ;
- et que la croissance y suit le schéma de la
`productivité maximale' spécifiée et modifiée en
fonction des facteurs écologiques.
La prise en compte des différents paramètres est
indiquée à la figure 66, qui détaille l'architecture du
modèle codifié en VBA-Excel (voir reprise des codes à
l'Annexe 4).
Figure 66. Exemple de configuration du
modèle CASS
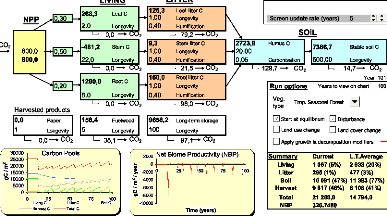

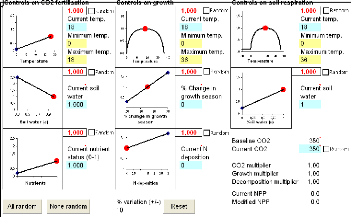

Perturbation Changement d'occupation du sol
00 CO2
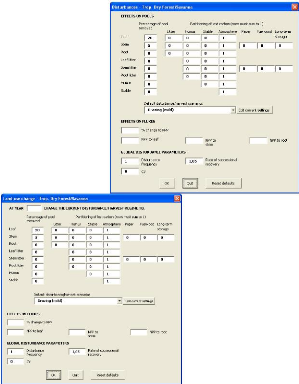
Section 6.07
IV.5. Résultats de la simulation
Pour structurer les paramètres de base, on a
combiné les données de la littérature et les
données collectées dans le cadre de ce travail. Les stocks moyens
de carbone de la ligne de référence au niveau des sols et de la
litière ont été ajustés en se basant sur les
travaux de Elberling et al. (2002); Touré (2002); Batjes
(2003); Manlay et al. (2004); Tieszen et al. (2004); Gray
(2005); Farage et al. (2007); Nsabimana et al. (2008). Pour
la biomasse sur pied, les données de terrain et les résultats de
Brown (1997); Moura-Costa et Stuart (1999); Woomer et al. (2004);
Barbosa et Fearnside (2005); Lufafa et al. (2008) ont servi de base
pour renseigner le modèle.
En se basant sur les valeurs par défaut du
modèle on a accepté la répartition de la biomasse pour la
savane boisée (30% feuilles, 50% bois ; 20% racines) ; pour la savane
arborée ou arbustive (25% feuilles, 45% bois : 30% racines), pour les
formations herbacées (60% feuilles, 10% ligneux ; 30% racines) ; pour
l'agriculture (50% feuilles ; 10% ligneux ; 40% racines). Ces valeurs peuvent
être mieux ajustées, mais on s'est basé pour la
définition du ratio de la biomasse racinaire sur les valeurs IPCC
concernant le « Root/Shoot ratio » (rapport biomasse
racinaire/ biomasse sur pied) (IPCC, 2003).
Les données sur les températures maximum et
minimum, l'humidité des sites, la concentration de CO2 dans l'air (370
ppm) ont été ajustées.
La donnée de base du modèle est l'assimilation
du CO2 atmosphérique. Ce paramètre est rendu dans le
modèle par la PPN qui en constitue la porte d'entrée. Les valeurs
de PPN ont été obtenues par analyse de données
satellitaires (MODIS). Les PPN annuelles de 2000 à 2006 ont
été acquises à partir du serveur
http://daac.ornl.gov/NPP/npp
home.htm; et les moyennes de PPN de différentes formations ont
été appliquées. Le format MOD17A3.A2000.
Sahel_subset_npp_1km a été téléchargé et le
Sénégal échantillonné. Les valeurs moyennes de PPN
sont calculées en utilisant des polygones représentatifs des
formations ciblées.
Les différentes simulations ont concerné (voir
figure 67) des savanes soumises aux feux de brousse (simulation 1 et 2) ; des
savanes exploitées pour le bois (simulations 2 et 4) ; des terres de
culture mises en jachère (simulation 5) ; des zones boisée
aménagées dont le bois est utilisé pour les besoins
d'énergie (simulation 6) ; des savanes pâturées (simulation
7 et 8) ; et des formations de savanes boisées/forêts claires
transformées en terres agricoles. Ces choix de scénarios sont
inspirés des cas fréquents de situations qu'on rencontre dans
les

écosystèmes étudiés ; Il est possible
de diversifier les cas et combiner les facteurs mais l'interprétation
des résultats devient plus délicate à faire.
Figure 67. Courbes de variation du carbone
dans les différents réservoirs selon des scénarios de
perturbations en cours dans les écosystèmes de savane
(durée de la simulation = 100 ans)
|
Résultat graphique (100 ans)
|
Paramètres clés
(g/C/m2)
|
|
Simulation 1
|
Savane arborée brûlée
PPN/an : 650 C/Feuille : 100 C/Bois : 3500 C/Racine : 1700
C/Litière feuille : 215
C/Litière du bois : 525
Humus : 5000
C stable du sol : 6500
Collecte industrielle : 0
Collecte bois énergie : 400
C long terme : 4000 Perturbation : Feux de
brousse de moyenne intensité tous les 3
|
|
|
|
|
|
C/Racines mortes : 130
|
|
|
|
|
|
ans
|
|
Simulation 2 (a)
|
Savane boisée /forêt claire
brûlée
(PPN/an 800)
C/Feuille : 250
C/Bois : 4000
C/Litière feuille : 400
C/Litière du bois : 600
C/Racines mortes : 175
Humus : 7000
C stable du sol : 8500
Collecte industrielle : 0
Collecte bois énergie: 600
C long terme : 16000 Perturbation : feux de
brousse de forte intensité tous les 3 ans
|
|
|
|
|
|
|
|
|
|
C/Racine : 1900
|
|
|
|
|
|
|
Résultat graphique (100 ans)
|
Paramètres clés
(g/C/m2)
|
|
Simulation 2 (b)
|
Même scénario que 2a, mais Perturbation :
feux de brousse de faible intensité tous les 3 ans
2a simule les feux tardifs, alors que 2b les feux
précoces
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Simulation 3
|
Savane aménagée exploitée
PPN : 700
C/Feuille : 180 C/Bois : 7780 C/Racine : 1499 C/Litière
feuille : 213
C/Litière du bois : 501
C/Racines mortes : 150
Humus : 9571
C stable du sol : 3500
Collecte industrielle : 0
Collecte bois énergie: 196
C long terme : 5016 Perturbation : exploitation
de moyenne intensité tous les 15
ans
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Simulation 4
|
Savane boisée, forêt claire
exploitée
PPN : 800
C/Feuille : 250
C/Bois : 4000
C/Racine : 1900
C/Litière feuille : 400
C/Litière du bois : 600
C/Racines mortes : 175
Humus : 7000
C stable du sol : 8500
Collecte industrielle : 0
Collecte bois énergie: 600 C long terme : 16000
Perturbation : Exploitation intensive de bois
tous les 10 ans
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Cb l
carbon
|
|
|
Résultat graphique (100 ans)
|
Paramètres clés
(g/C/m2)
|
|
Simulation 5
|
Terre de Culture mise en jachère
PPN : 400
C/Feuille : 400
C/Bois : 120
C/Racine : 350
C/Litière feuille : 250
C/Litière du bois : 115
C/Racines mortes : 50
Humus : 1200
C stable du sol : 1500
Collecte industrielle : 0
Collecte bois énergie: 0
C long terme : 1600 Perturbation : terre
cultivée mise en jachère, devient savane après 10 ans
d'exploitation
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Simulation 6
|
Terre dégradée protégée puis
exploitée pour le bois et régulièrement
brûlée
PPN : 800
C/Feuille : 250
C/Bois : 4000
C/Racine : 1900
C/Litière feuille : 400
C/Litière du bois : 600
C/Racines mortes : 175
Humus : 7000
C stable du sol : 8500
Collecte industrielle : 0
Collecte bois énergie: 600
C long terme : 16000 Perturbation : Exploitation
de bois pendant les 25 premières années (tous les 7 ans), puis
exploitation faible combinée à des feux de faible
intensité tous les 7 ans.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Résultat graphique (100 ans)
|
Paramètres clés
(g/C/m2)
|
|
Simulation 7
|
Savane pâturée intensivement PPN :
650
C/Feuille : 150
C/Bois : 3500
C/Racine : 1750
C/Litière feuille : 220 C/Litière du bois : 525
C/Racines mortes : 130 Humus : 5000
C stable du sol : 6500
Collecte industrielle : 0 Collecte bois énergie: 400 C
long terme : 8000 Perturbation : Année 25 introduction
d'importants pâturages.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Simulation 8
|
Savane herbeuse-arbustive faiblement
pâturée, sans feu, sans coupe
PPN: 400
C/Feuille : 300
C/Bois : 60
C/Racine : 900
C/Litière feuille : 400 C/Litière du bois : 30
C/Racines mortes : 20 Humus : 2500
C stable du sol : 3000
Collecte bois énergie: 0
C long terme : 5000 Perturbation :
pâturage faible à partir de l'année 25
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Collecte industrielle : 0
|
|
|
|
Simulation 9
|
Savane boisée, forêt claire
transformée en terres agricoles
PPN : 800
C/Feuille : 250
C/Bois : 4000
C/Racine : 1900
C/Litière feuille : 400 C/Litière du bois : 600
C/Racines mortes : 175 Humus : 7000
C stable du sol : 8500 Collecte industrielle : 0 Collecte bois
énergie: 600 C long terme : 16000
Perturbation : terre défrichée
pour mise en culture à partir de l'année 25.
|
|
|
Car
Ca
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
(a) IV.5.1. Analyse des résultats de la
simulation
Les deux premières simulations (1 et 2) permettent
d'analyser l'impact des feux de brousse au niveau des savanes.
Les feux de brousse constituent un phénomène récurrent,
quasiannuel et qui affectent les savanes chaque saison sèche. Les feux
affectent ces écosystèmes directement par la consommation de
biomasse et indirectement par la modification des processus écologiques
comme le cycle de l'eau, les cycles biogéochimiques, les échanges
d'énergie et de matière entre les composantes de
l'écosystème et surtout par la décomposition de la
matière organique entraînant une forte libération de
carbone, etc. Les travaux antérieurs (Kuhlbusch et Krutzen., 1995;
Fearnside, 2000; Mbow et al., 2000; Mbow et al., 2003;
Nielsen et al., 2003; Saarnak et al., 2003; Zhang et
al., 2003; Mbow et al., 2004; Barbosa et Fearnside, 2005) ont
montré le rôle important joué par les feux dans les
savanes. Il faut toutefois noter que, l'impact du feu dépend de
plusieurs facteurs : intensité du feu, types de formations
végétales affectés et leur état de dessiccation ;
conditions météorologiques et topographiques, etc.
La simulation 1 montre un feu d'intensité moyenne sur
une savane arborée mais qui survient tous les trois ans. Pendant les
premières années, les stocks de carbone diminuent assez
rapidement, pour ensuite rester stable pour le reste du temps. On remarque la
variation parallèle du carbone de la litière et de la biomasse
après chaque feu ; alors que le carbone du sol est moins sensible
à ces variations événementielles dues aux feux.
Pour la simulation 2 (a, b), on a essayé de voir
l'impact d'un brOlis sévère et de faible intensité sur des
formations de savane boisée (plus dense) sur un pas de temps de 3 ans.
Il apparaît que la chute des stocks de carbone est spectaculaire pour les
feux intenses, y compris pour le carbone du sol. L'intensité du feu fait
varier significativement les stocks de carbone de tous les réservoirs.
Ainsi à défaut d'empêcher les feux, il faut aller dans le
sens d'utiliser des feux de moindre intensité, qu'on désigne sous
le vocable `feux précoces'. Il s'agit de feux contrôlés
dans des conditions d'humidité et de charge de biomasse bien
précises. Les feux de forte intensité désigne ici les feux
tardifs en milieu de saison sèche (figure 68).
Figure 68. Intensité des feux de saison
sèche dans le Parc National du Niokolo Koba (Sud est
Sénégal)

Les simulations 3 et 4 concernent les coupes (exploitations)
de bois dans les formations de savane. L'exploitation
forestière est un des facteurs les plus puissants et les plus
manifestes de la dégradation des savanes. Les besoins en bois, pour
satisfaire la demande d'énergie, le marché du bois de service et
du bois d'uvre, constituent une rubrique importante que ne peuvent supporter
les formations forestières. La frénésie de l'exploitation
dépasse assez souvent la capacité des écosystèmes.
Au Sénégal, les besoins d'énergies sont largement
satisfaits à partir des formations forestières. La
dégradation de ces dernières a suscité plusieurs
stratégies comme le reboisement, les bois de village, les plantations en
régies, les plantations privées, la mise en défens, etc.
Ces initiatives prises par le service forestier n'ont pas empêché
un développement rapide des défrichements et des coupes
(déforestation). La déforestation est ciblée par le
protocole de Kyoto comme une importante source de carbone, mais aussi une
solution à l'atténuation du CO2 atmosphérique si des
mesures adéquates sont prises. Plusieurs études portant sur les
changements d'occupation du sol (Grepperud, 1996; Giese et al., 2003;
Lambin et al., 2003; Reenberg et al., 2003; Tappan et
al., 2004) montrent des tendances de dégradation dont l'impact sur
les stocks de carbone est sans équivoque (figure 69).
La simulation 3 montre une formation de savane bien
aménagée, dont l'exploitation est mise en adéquation avec
les stocks de bois disponibles. Les stocks de carbone de la biomasse sur pied,
de même que ceux du sol sont légèrement affectés.
Par contre, la simulation 4 montre une situation de surexploitation. Pendant
l'intervalle sans exploitation, la végétation a tendance à
repartir, mais le retour de l'exploitation ramène les quantités
de carbone à un niveau très bas. Il apparaît ainsi, que la
mise en adéquation entre l'exploitation et le potentiel ligneux est une
voie salutaire pour préserver non seulement l'équilibre des
formations en question (Ravindranath et al., 2001; Riedacker, 2004;
Sambou, 2004) ; mais
aussi pour améliorer la contribution de ces
écosystèmes à l'atténuation du taux de carbone
atmosphérique.
Figure 69. Exploitation du bois énergie
au Sénégal Oriental

La troisième catégorie de simulation porte sur
les terres dégradées. Les scénarios
retenus ici, portent sur une perspective de reconstitution ou de
réhabilitation. Les terres dégradées peuvent prendre
plusieurs formes. Nous en retenons deux: une jachère qui devient savane,
ou une terre dégradée mise en défens mais soumise aux feux
et à l'exploitation de bois. On constate que les terres
dégradées ont un potentiel de reconstitution assez rapide, mais
la vitesse de reconstitution du carbone du sol est plus lente que celui de la
biomasse (simulation 5). Le carbone du sol semble se reconstituer moins vite et
peut faire 3 fois moins le carbone de la biomasse selon la simulation 5. Une
jachère en reconstitution diffère ainsi des savanes par le taux
de carbone du sol qui y est moins important ; les rapports sont inversés
en savane (carbone du sol plus important que le carbone de la biomasse,
simulations 1-2-3-4). C'est pour cette raison que dans la plupart des
jachères, le cycle de retour s'est raccourci, à certains endroits
du Sénégal, à cause de cette impression de reconstitution
du milieu liée au couvert végétal. Les études sur
le carbone des jachères confirment cet important potentiel de stockage
de carbone (Hashimotio et al., 2000; Tschakert et al., 2004;
Wood et al., 2004; Vagen et al., 2005). Certains auteurs
(Lufafa, et al., 2008; Takimoto et al., 2008) soulignent la
présence importante des arbres en zone de jachères ; lesquels
peuvent initier le retour des cycles d'échanges entre la biomasse et le
sol. C'est pour cette raison d'ailleurs que les pratiques
agro-forestières sont vivement encouragées pour les terres
caractérisées par une perte significative de fertilité
(figure 70). Les arbres font remonter les nutriments non accessibles aux
plantes cultivées et entretiennent une fertilisation organique par les
chutes et la décomposition de feuilles (Mbow et al., 2008).
Figure 70. Pratiques agroforestières dans
le bassin arachidier (gauche, Faidherbia albida) et dans le Saloum
Oriental (droite, Cordyla pinnata)

La simulation 6, par contre est une savane qui dans un premier
temps est exploitée pour son bois tous les 7 ans pendant les 25
premières années. L'atténuation des
prélèvements même avec une présence de feux de
moindre intensité permet par la suite de conserver l'essentiel du
carbone des différents réservoirs. Cette simulation montre que
malgré le passage des feux et la collecte de bois de chauffe
(d'intensité moindre), la productivité de
l'écosystème est relativement bonne. La simulation 6 montre qu'un
écosystème qui n'est pas continuellement exploité
même avec un passage régulier des feux de moindre intensité
peut jouer un rôle important dans le stockage de carbone.
Les simulations 7 et 8 traduisent l'influence du
pâturage au niveau des stocks de carbone (figure 71). La
simulation 7 montre une importante activité pastorale après 25
années dans une savane arborée. Le carbone de la biomasse est
fortement réduit et se stabilise rapidement si la charge animale n'a pas
augmenté entre temps. Le carbone du sol est aussi réduit mais pas
avec la même ampleur que celui de la biomasse. Si la pression du
bétail est constante, ce type de formation peut se maintenir en
l'absence d'autres facteurs de dégradation. Par contre, une formation
herbacée faiblement pâturée dans des conditions de bonne
pluviométrie et sans intervention d'autres facteurs de
dégradation, peut mener à une reconstitution des stocks de
carbone (Hary et al., 1996). Ainsi la question du pâturage se
pose en termes de charge ou de pression du bétail.
Figure 71. Pression du bétail dans la
zone de Kaffrine (gauche, photo, A. Reenberg) et les parcours de transhumance
au Sénégal Oriental (droite)

La dernière simulation (simulation 9) est une
reconstitution d'un défrichement agricole dans une
savane boisée (figure 72). Ces types de situations sont souvent
rencontrés dans la zone d'étude avec l'avancée du front
cotonnier et du front arachidier. Ces spéculations nécessitent
une coupe rase des arbres au niveau des champs. Des pratiques de brûlis,
de coupes ou de mortalités provoquées des arbres finissent au
bout de quelques années par dénuder des formations
végétales relativement denses.
Figure 72. Elimination de l'arbre du champ pour
les cultures de rente.

Ces différentes simulations montrent les importantes
quantités de carbone des sols de
savane. Il faut remarquer cependant
que les processus qui régissent le recyclage du carbone
de ces
écosystèmes sont fortement influencés par les processus
qui se déroulent au niveau de
la végétation (Archer et al., 2004). Le
carbone du sol est le résultat des différents échanges de
matières végétales entre les plantes et le sol. La
végétation devient ainsi le moteur de la dynamique du carbone
dans les différents réservoirs de ces écosystèmes.
Si la végétation est bien conservée, le carbone de
l'écosystème se reconstitue assez vite ; par contre la
dégradation de la végétation nuit, directement aux autres
réservoirs dans des délais plus ou moins courts. C'est une des
raisons pour lesquelles le Protocole de Kyoto n'a pas rendu éligible le
carbone du sol, mais juste le carbone de la végétation qui est
très vulnérable aux modifications d'utilisation des terres.
Ces simulations permettent ainsi d'apprécier la
dynamique de la végétation et surtout le test de
scénarios. Lesquels, peuvent être complexes (plusieurs
combinaisons possibles) et requièrent beaucoup d'informations in
situ pour bien ajuster les simulations. Parallèlement, IPCC a, dans
le cadre des méthodes approuvées et du guide des bonnes pratiques
sur la foresterie, développé des méthodes de calcul de la
dynamique du carbone des écosystèmes pour la comptabilité
du carbone des projets MDP.
Ces méthodes approuvées sont à l'heure
actuelle (septembre 2008) au nombre de 10 avec une méthode
consolidée :
http://cdm.unfccc.int/methodlogies/ARmethodologies/approved
ar.html. Ces méthodes d'élaboration introduisent la notion
d'additionalité et leurs méthodes de calcul. Un des aspects
importants de ces méthodes approuvées est la prise en compte des
fuites. Les fuites sont non seulement les prélèvements sur la
biomasse mais aussi les dégagements de GES liés aux
activités du projet. Le facteur climatique peut renforcer ou contrer ces
processus humains. Dans le modèle CASS les facteurs écologiques
sont pris en compte à travers le module `Environmental
Multipliers' que sont essentiellement la température,
l'humidité, la fertilisation et la saisonnalité.
(b) IV.5.2. Conditions environnementales et
synthèse du carbone
Dans le cadre des changements climatiques, des modifications
sont entrevues au niveau du cycle de l'eau, de la température, et des
concentrations de CO2. Ces éléments affectent la
photosynthèse et donc le processus de stockage de carbone des
plantes.
Le Sénégal fait partie des zones arides et
semi-arides caractérisées par une forte variabilité des
précipitations. Les pluies annuelles ont fortement diminué
pendant les quarante dernières années. Un léger mieux est
noté au cours des 10 dernières années avec une
augmentation des pluies de forte intensité pendant le mois d'aoflt.
La réduction des précipitations entraîne
un stress hydrique qui réduit l'efficacité de la
photosynthèse. La
réduction de l'humidité du sol influe
directement sur l'ouverture des stomates des plantes qui, en réduisant
leur transpiration atténue la fixation du carbone et sa diffusion dans
la plante (Rosenberg et al., 1983). L'eau constitue alors un facteur
limitant dans la production de biomasse.
Les températures globales ont aussi
augmenté. Au Sénégal, les données des stations
synoptiques montrent une hausse de près 1° C depuis 1980 (chapitre
I). La photosynthèse n'est pas significativement affectée par les
variations de températures dans les marges normales d'adaptation de la
plante. Mais avec de fortes variations de température, la respiration de
la plante est très affectée. Les arbres tropicaux (plantes C3)
sont plus sensibles aux variations de températures que les plantes C4
(herbacées) qui peuvent assimiler plus vite d'ailleurs quand les
températures augmentent. Avec le réchauffement de la terre, un
stress thermique plus important devrait être noté sur les plantes
C3 (Rosenberg et al., 1983). Aussi, l'accroissement des
températures peut augmenter la respiration au niveau des sols et des
racines, et favoriser une décomposition rapide de la litière. Il
s'en suivra naturellement de plus grandes émissions de CO2 dans
l'atmosphère.
Les concentrations de CO2
atmosphérique sont en constante augmentation. Dans le chapitre
I nous avons montré que le CO2 augmente en moyenne de 1,4 ppm/an.
Théoriquement, en augmentant la concentration de CO2
atmosphérique, la fixation du carbone par les plantes augmente ; mais
faudrait-il que les autres conditions de température, de lumière
et d'humidité soient réunies. Des essais de Rosenberg et al.
(1983), montrent une augmentation linéaire de la
photosynthèse avec l'augmentation du CO2 atmosphérique entre 220
et 400 ppm. Les plantes C3 (arbres tropicaux) supportent plus l'augmentation de
CO2 que les plantes C4 (les Poaceae). Les plantes C3 augmentent plus
leur photosynthèse que les plantes C4 dans le cas d'une augmentation de
la concentration de CO2 atmosphérique. Ces différences ont
été notées dans des serres dont l'air ambiant a
été enrichi en CO2.
D'autres facteurs comme le vent et des turbulences ont
été évoqués comme pouvant influencer la
productivité des plantes. Le vent a essentiellement un effet
mécanique à travers les distorsions sur la physionomie des
formations. Il faut aussi noter que la température, l'humidité ou
le taux de carbone du sol influent sur la respiration des plantes
(dégagement de CO2). La respiration dépend de la
température qui influence le processus d'oxydation de l'humus,
l'activité métabolique de la flore et de la faune, et enfin la
respiration des racines. L'humidité du sol, quant à elle, est un
facteur déterminant dans le processus de décomposition de la
litière. Ainsi, les modifications de ces paramètres dans le cadre
des changements climatiques entraînent d'importantes mutations dans les
échanges biogéochimiques entre formations végétales
et atmosphère.
Section 6.08 Conclusion
La modélisation est un exercice complexe et
risqué. La diversité des modèles et les divergences
d'approches montrent que le sujet reste largement ouvert. Aussi, faut-il dans
le choix d'un modèle, considérer la maîtrise des
paramètres d'entrée et la possibilité de produire des
résultats vraisemblables et conformes à la réalité
de terrain. A l'heure actuelle, la grande majorité des modèles
sont des `boîtes noires', difficiles à pénétrer et
d'appropriation très contraignante. Le modèle CASS utilisé
dans cette partie du travail, a l'avantage d'être transparent et dispose
d'un paramétrage adapté pour les savanes et ses processus.
Les résultats obtenus montrent qu'au niveau des
écosystèmes de savanes d'importantes modifications peuvent
subvenir soit à cause de facteurs naturels ou humains. En l'absence de
perturbations humaines, les formations végétales
dégradées ont tendance à se reconstituer. Le but de
l'aménagement forestier est justement d'équilibrer les
prélèvements avec la capacité des
écosystèmes à se reconstituer. Les facteurs humains
influent sur les stocks de biomasse différemment selon la nature de la
perturbation et de son amplitude. Nous avons testé les
conséquences de plusieurs activités humaines (feu, collecte de
bois, pâturage, abandon/jachère, plantation etc.). Il
apparaît selon les résultats des simulations que les
écosystèmes de savane sont très sensibles (réaction
rapide après le passage d'un feu ou des coupes à blanc) et les
délais de reconstitution peuvent parfois prendre beaucoup de temps (au
moins 25 ans). Le suivi dans la modélisation des autres
réservoirs de carbone (litière et carbone du sol) montre que les
modifications de la végétation influent directement sur ces
réservoirs, même si les modifications sont plus lentes que celles
de la biomasse sur pied. Ainsi, tout effort de conservation des formations
ligneuses a un effet d'entraînement sur l'amélioration des stocks
de carbone des autres réservoirs.
Toutefois, pour introduire les variations entre les
différentes formations de savane, dont le concept est unifié dans
le modèle, nous avons procédé à la modification des
paramètres de base (PPN et allocation feuilles, branches, racines). Les
simulations proposées dans cette partie sont par conséquent des
scénarios vraisemblables selon les activités en cours dans les
savanes. Cependant, les dynamiques futures du climat, les changements de
précipitations, de température, de concentration de CO2 etc.,
peuvent affecter significativement la fixation du CO2 atmosphérique et
les processus de décomposition des plantes entraînant une
libération de GES. Ces nouvelles conditions écologiques sont en
prendre en compte dans le paramétrage du modèle.
Bibliographie
Adams, A., White, A., Lenton T.M. 2004. An analysis of some
diverse approaches to modelling terrestrial net primary productivity.
Ecological Modelling (177):353-391.
Aber, J.D., Reich, P.B., Goulden M.l. 1996. Extrapolating leaf
CO2 exchange to the canopy: a generalized model of forest photosynthesis
validated by eddy correlation. Oecologia (106): 257-265
Archer, S., Boutton, T.W., McMurtry, C. R., 2004. Carbon and
Nitrogen Accumulation in a Savanna Landscape: Field and Modeling Perspectives.
In: M-Shiyomi (Editor), Global Environmental Change in the Ocean and on Land.
TERRAPUB, pp. 359-373.
Barbosa, R.I., Fearnside P.M. 2005. Above-ground biomass and
the fate of carbon after burning in the savannas of Roraima, Brazilian
Amazonia. Forest Ecology and Management, In Press, 22 p.
Baron, C., Perez, P., Maraux F., 1996. SARRA: Système
d'Analyse Régionale des Risques Agroclimatiques. Module SARRABIL: bilan
hydrique à la parcelle. Analyses comparatives des bilans annuels ou
pluriannuels. Analyses des risques climatiques. Guide d'utilisation, CIRAD-CA,
Montpellier, 32 p.
Batjes N.H. 2003. Estimation of soil carbon gains upon
improved management within croplands and grasslands of Africa Environment,
Development and Sustainability (6): 133-143,.
Brown S., 1997. Estimating Biomass and Biomass Change of
Tropical Forests: a Primer. (FAO Forestry Paper -134). FAO - Food and
Agriculture Organization of the United Nations, Roma, 48 p.
Elberling, B., Touré, A., Rasmussen K. 2002. Changes in
soil organic matter following groundnut-millet cropping at three locations in
semi-arid Senegal, West Africa. Agriculture Ecosystems & Environment (96):
37-47.
Farage, P.K., Ardo, J., Olsson, L., Rienzi, E.A., Ball, A.S.,
Pretty J.N. 2007. The potential for soil carbon sequestration in three tropical
dryland farming systems of Africa and Latin America: A modelling approach. Soil
and Tillage Research ( 94):457-472.
Fearnside P.M. 2000. Global warming and tropical land-use
change: greenhouse gas emission from biomass burning decomposition and soils in
forest conversion, shifting cultivation and secondary vegetation. Climatic
Change (46):115-148.
Giese, L.A.B., Aust, W.M., Kolka, R.K., Trettin C.C. 2003.
Biomass and carbon pools of
disturbed riparian forests. Forest Ecology and Management,
180:493-508.
Gray K.M., 2005. Changing soil degradation trends in Senegal with
carbon sequestration
payments. Monograph, Montana State University, Montana.
Grepperud S. 1996. Population Pressure and Land Degradation: The
Case of Ethiopia. Journal of Environmental Economics and Management
(30):18-33.
Hary, I., Schwartz, H-J., Pielert, V.H.C., Mosler C. 1996. Land
degradation in African pastoral systems and the destocking controversy.
Ecological Modelling (86):227-233.
Hashimotio, T., Kojima, K., Tange, T., Sasaki S. 2000. Changes
in carbon storage in fallow forests in the tropical lowlands of Borneo. Forest
Ecology and Management, 126:331-337.
Haxeltine, A., Prentice I.C. 1996a. BIOME3: an equilibrium
terrestrial biosphere model based on ecophysiological constraints, resource
availability and competition among plant functional types. Global
Biogeochemical Cycles, 10 (4):693-709.
Haxeltine, A., Prentice I.C. 1996b. A general model for the light
use efficiency of primary production. . Functional Ecology, 10:551-561.
IPCC, 2003. Good Practice Guidance for Land Use, Land-Use Change
and Forestry. Penman J., Kruger D., Galbally I., Hiraishi T., Nyenzi B.,
Emmanuel S., Buendia L.,
Hoppaus R., Martinsen T., Meijer J., Miwa K., and Tanabe K.
(Eds). Institute for Global Environmental Strategies, Kanagawa, Japan.
King, A.W., Post, W.M., Wullshleger S.D. 1997. . The potential
response of terrestrial carbon storage to change in climate and atmospheric
CO2. Climate Change, 35:199- 227.
Kuhlbusch, T.A.J., Crutzen P.J. 1995. Towards a global
estimate of black carbon in residues of vegetation fires representing a sink of
atmospheric CO2 and a source of CO2. Global Biogeochemical Cycles, 9
(4):491-501.
Lambin, E., F., Geist, H.J., Lepers E. 2003. Dynamics of land
use and land cover change in
tropical regions. Annual Reviews of
Environmental Resources (28):205-241.
Landsberg, J.J., Waring R.H. 1997. A generalised model of
forest productivity using simplified concepts of radiation use efficiency,
carbon balance and partitioning Forest Ecology Management, 95:209-228.
Leith H., 1975. Modeling the primary productivity of the world.
In: Leith, et al. (Editor), Primary Productivity of the Biosphere. .
Springer-Verlag, New York.
Lenton T.M. 2000. Land and ocean carbon cycle feedback effects on
global warming in a simple Earth system model. Tellus, 52B:1159-1188.
Lufafa, A., Bolte, J., Wright, D., Khouma, M., Diedhiou, I.,
Dick, R.P., Kizito, F., Dossa, E., Noller J.S. 2008. Regional carbon stocks and
dynamics in native woody shrub communities of Senegal's Peanut Basin.
Agriculture, Ecosystems and Environment (In press).
Manlay, R.J., Ickowicz, A., Masse, D., Floret, C., Richard,
D., Feller C. 2004. Spatial carbon, nitrogen and phosphorus budget of a village
in the West African savanna-I. Element pools and structure of a mixed-farming
system. Agricultural Systems (79):55-81.
Mbow, C., Goita, K., Bénié G.B. 2004. Spectral
indices and fire behavior simulation for fire risk assessment in savanna
ecosystems. Remote Sensing of Environment (91):1-13.
Mbow, C., Nielsen, T.T., Rasmussen K. 2000. Savanna Fires in
East-Central Senegal: Distribution Patterns, Resource Management and
Perceptions. Human Ecology, 28 (4):561-583.
Mbow, C., Sambou, B., Ba, A.T., Goudiaby A. 2003. Vegetation
and fire readiness in main morphological units of Niokolo Koba National Park
(Southeast Senegal). Danish Journal of Geography, 103 (1):55-62.
Mbow, C., Mertz, O., Diouf, A., Rasmussen, K., Reenberg, A.,
2008. The history of environmental change and adaptation in eastern
Saloum-Senegal - driving forces and perceptions. Journal of Arid Environment,
64: 210-221.
Monserud R.A. 2003. Evaluating forest models in a sustainable
forest management context. FBMIS, 1:22-47.
Moura-Costa, P., Stuart M., 1999. Issues related to monitoring,
verification and certification
of forestry-based carbon offset projects. In:
W.F.A. Conference, et al. (Editor).
Nielsen, T.T., Rasmussen, K., Mbow, C., Touré A. 2003. The
fire regime of Senegal and its determinants. Danish Journal of Geography, 103
(1):43-53.
Nsabimana, D., Klemedtson, L., Kaplin, B.A., Wallin G. 2008.
Soil carbon and nutrient accumulation under forest plantations in southern
Rwanda. African Journal of Environmental Science and Technology, 2
(6):142-149.
Polglase, P.J, Wang Y.P. 1992. Potential CO2-enhanced Carbon
Storage by the Terrestrial Biosphere Australian Journal of Botany,
40:641-656.
Post, W.M., King, A.W., Wullshleger S.D. 1997. Historical
variations in terrestrial biospheric carbon storage. Global Biogeochemical
Cycles, 11 (1 ):99-109.
Prentice, C., Cramer, W., Harrison, S.P., Leemans, R.,
Monserud, R.A., Solomon A.M. 1992. A global biome model based on plant
physiology and dominance, soil properties and climate Journal of Biogeography,
19:117-134.
Ravindranath, N.H., Sudha, P, Rao S. 2001. Forestry for
sustainable biomass production and carbon sequestration in India. Mitigation
and Adaptation Strategies for Global Change (6):233-256.
Reenberg, A., Mbow, C., Diallo B., 2003. Temporal and spatial
diversity of Sudano-Sahelian landscapes: Land use and land cover dynamics in
forest reserves and their margins. In: D.A. Wardell, et al. (Editor),
Negotiated frontiers in Sudano-Sahelian landscapes Implication for natural
resource management strategies. Sahel-Sudan Environmental Research Initiative
(SEREIN), Denmark, pp. 137-161.
Ribot J., 1998. Decentralization and participation in Sahelian
Forestry: Legal instruments of central political-administrative control,
Harvard Center for Population and Development Studies. Cambridge, 47 p.
Ribot J., 2002. African Decentralization. Local Actors, Powers
and Accountability, United Nations Research Institute for Social Development.
Democracy, Governance and Human Rights Paper Number 8, Geneva.
Riedacker A., 2004. Changements Climatiques et Forêts.
SILVA, RIAT, 231 p.
Rosenberg, N.J., Blad, B.L., Verma S.B., 1983. Microclimate.
The Biological Environment. Chapter 8-Field Photosynthesis, respiration, and
the carbon balance (pp 288-390). John Willey and Sons, New
York-Chichester-Brisbane-Toronto-Singapore, 495 p.
Roxburgh S., 2004. The CASS Terrestrial Carbon Cycle Model,
V.1.2. User Guide and Tutorial exercises, CRC for Greenhouse Accounting, GPO
Box 475. Canberra ACT 2601, Australia.
Saarnak, C., Nielsen, T.T., Mbow C. 2003. A local-scale study
of the trace gas emissions from vegetation burning around the village of Dalun,
Ghana, with respect to seasonal vegetation changes and burning practices.
Climate Change (56):321-338.
Sambou B., 2004. Evaluation de l'état, de la dynamique
et des tendances évolutives de la flore et de la
végétation ligneuse dans les domaines soudanien et
sub-guinéen au Sénégal. Doctorat d'Etat, Université
Cheikh Anta Diop de Dakar (UCAD), Dakar, 231 p.
Svirezhev, Y.M., Bloh, v. W. 1997. Climate, vegetation and global
carbon cycle: the simplest zero-dimensional model. Ecological Modeling,
101:79-95.
Svirezhev, Y.M., Bloh, v. W. 1998. A zero-dimensional
climate-vegetation model containing global carbon and hydrological cycle.
Ecological Modeling, 106:119-127.
Takimoto, A., Nair, P.K.R., Nair V.D. 2008. Carbon stock and
sequestration potential of traditional and improved agroforestry systems in the
West African Sahel. Agriculture, Ecosystems and Environment (125):159-166.
Tappan, G.G., Sall, M., Wood, E.C., Cushing M. 2004. Ecoregions
and land cover trends in Senegal. Journal of Arid Environments (59):427-462.
Tieszen, L.L., Tappan, G.G, Touré A. 2004. Sequestration
of carbon in soil organic matter in Senegal: an overview. Journal of Arid
Environments (59):409-425.
Touré A., 2002. Contribution à l'étude de
l'évolution des réservoirs de carbone en zone nordsoudanienne au
Sénégal., Ecole Polytechnique Fédérale de Lausane
(EPFL), 158 p.
Tschakert, P., Khouma, M., Sène M. 2004. Biophysical
potential for soil carbon sequestration in agricultural systems of the Old
Peanut Basin of Senegal. Journal of Arid Environments (59):511-533.
Vagen, T-G., Lal, R., Singh B.R. 2005. Soil carbon sequestration
in Sub-Saharan Africa: A Review. Land Degradation and Development
(16):53-71.
Waring, R.H., Running S.W., 1998. Forest Ecosystems. Analysis at
multiple scales. Academic Press, California, 370 p.
Wood, E.C, Tappan, G.G, Hadj A. 2004. Understanding the drivers
of agricultural land use
change in south-central Senegal. Journal of Arid
Environments (59):565-582.
Woomer, P.L., Tieszen, L.L., Tschakert, P., Parton, W.J.,
Touré A., 2001. Landscape carbon sampling and biogeochimical modeling. A
two-week skills development workshop conducted in Senegal., USGS, CSE.
Woomer, P.L., Touré, A., Sall M. 2004. Carbon stocks in
Senegal's Sahel Transition Zone. Journal of Arid Environments (59):499-510.
Zhang, Y.H., Wooster, M.J., Tutubalina, O., Perry G.L.W. 2003.
Monthly burned area and forest fire carbon emission estimates for the Russian
Federation from SPOT VGT. Remote Sensing of Environment (87):1-15.
Article VII.
Article VIII. CHAPITRE V :
Article IX. Analyse dendrochronologique des
principales espèces
Pour parvenir à une analyse fine sur la dynamique des
arbres, la méthode dendrométrique a été
testée avec succès dans les écosystèmes
tempérés. Le débat sur son efficacité en milieu
tropical reste ouvert même s'il existe de réels potentiels. Sous
ce rapport, l'application de cette méthode montre les
possibilités offertes pour d'une part, caractériser la dynamique
du carbone à travers la datation des espèces de savane et d'autre
part permettre de décrire la variabilité climatique à
partir des cernes des arbres. Sur la base de ces informations, des
modèles de croissance des espèces principales (Cordyla
pinnata, Ptercocarpus erinaceus, Combretum glutinosum, Terminalia macroptera,
Acacia macrostachya) sont établis en rapport avec les conditions
écologiques des savanes soudaniennes et soudano-guinéennes. Outre
ces résultats, on peut signaler le poids des interactions et contraintes
écologiques dans ces écosystèmes, ce qui explique le
caractère non linéaire et lent de la croissance des arbres sur
les sites étudiés.
Section 9.01 Introduction
L'une des questions centrales de la recherche sur les
formations de savane s'articule autour de la dynamique, de la
productivité ou du rythme de croissance des espèces
végétales. Cette question peut être liée à la
discussion sur la gestion durable des ressources forestières pour une
reconstitution des formations dégradées et pour la
séquestration du carbone dans l'esprit de Kyoto. L'analyse de la
dynamique de la végétation est développée dans la
plupart des études sur la base de la répartition des classes de
diamètre, de l'évolution de la composition floristique ou par
l'utilisation des modèles écologiques. Ces analyses sont le plus
souvent orientées sur la structure de la végétation. A
l'échelle des individus, il est possible de suivre la croissance des
arbres en se basant sur l'accroissement du diamètre du tronc. Cette
option est adoptée dans le traitement des parcelles de suivi de la
végétation ligneuse. Cependant, vu la faible fréquence des
mesures, il est difficile de déterminer une relation claire entre la
dynamique du volume de bois et la variation pluviométrique par
exemple.
La zone soudanienne et soudano-guinéenne connaît
d'importantes variations climatiques et écologiques dues aux
fluctuations de la pluviométrie depuis les années 1970. Le
déplacement des isohyètes est une manifestation claire de la
réduction généralisée des pluies (Sagna, 2006).
Pour appréhender l'impact de cette crise climatique, la recherche s'est
intéressée aux impacts sur la biodiversité, sur la
productivité des écosystèmes et les impacts
socioéconomiques. Cette variabilité climatique se manifeste aussi
par des années excédentaires avec une pluviométrie qui
dépasse la normale. Ces extrêmes ont aussi des impacts sur la
croissance des arbres et se manifeste quantitativement et qualitativement sur
le développement des plantes.
Pour suivre l'impact de cette variabilité climatique
sur la dynamique des écosystèmes, plusieurs méthodes
indirectes d»analyse de l'historique des formations
végétales (étude des pollens, analyse des sédiments
du sol) ont été proposées mais les essais n'ont pas
toujours donné des résultats satisfaisants (Worbes, 2002). Le
besoin de mieux comprendre la dynamique de croissance des arbres et l'effet de
la variabilité pluviométrique sur la végétation
suscite de nos jours un intérêt renouvelé de la
dendrochronologie. La dendrochronologie (Du grec : dendron = l'arbre,
kronos = le temps, logos = l'étude) est une
méthode de datation, précise à 1 an voire à 6 mois
près, qui permet de déterminer la période durant laquelle
un arbre a vécu et de préciser l'année et la saison de son
abattage. Grâce à elle, on peut dater l'âge des individus,
la vitesse et les conditions de leur croissance. Ces paramètres
permettent de reconstituer la variabilité du climat, notamment les
précipitations, pendant la durée de vie de la plante. Des
études on d'ailleurs pu compléter certaines séries
climatiques lacunaires utilisant les résultats d'analyse
dendrochronologique (Tarhule et
Hughes, 2002). Elle apporte, en plus d'une datation, de
nombreux renseignements dans les domaines climatiques et écologiques
à partir de la structure du bois. L'analyse du cambium a amené
certains auteurs comme Worbes (2002), à rappeler l'importance des
phytohormones des bourgeons, de l'anatomie du bois à travers les
propriétés du xylème (arrangement des vaisseaux, la
structure du parenchyme et des fibres du bois).
Le principe de base repose sur le fait que la croissance des
végétaux ligneux à un rythme annuel est soumise à
l'alternance des saisons. En effet, les arbres produisent un cerne de
croissance pendant chaque saison des pluies grâce aux assises du cambium
situé sous l'écorce. La largeur des cernes varie essentiellement
en fonction des quantités d'eau de pluie et de leur répartition.
A l'intérieur de chaque cerne, on peut distinguer la période de
saison sèche (poreux et clair) et celle de la saison des pluies (dense
et foncé). L'épaisseur des cernes dépend de la
pluviométrie. Pendant les années pluvieuses, le cerne est plus
large, alors que durant les années sèches, le cerne est plus
étroit. Pour une même espèce, dans une même zone
climatique, les cernes de croissance évoluent en théorie de la
même manière. La différence entre espèces de la
même zone réside dans la vitesse de croissance de celles-ci. On
peut alors, après avoir mesuré l'épaisseur des cernes sous
loupe binoculaire établir une courbe de référence
normalisée pour une région climatique donnée.
L'épaisseur ou la densité des cernes est donc,
d'une part, un indicateur de l'histoire du climat passé et, d'autre
part, un point de repère pour dater une période de croissance. On
peut mesurer les cernes et en tirer des graphiques qui représentent
l'évolution de la croissance d'un arbre. Les années sont
représentées sur la ligne horizontale de gauche à droite
(abscisse), l'épaisseur des cernes figure sur la ligne verticale
(ordonnée).
Ces cernes sont facilement visualisables sur les souches sous
la forme d'anneaux concentriques. En comptant les cernes et en les analysant,
on peut établir, pour chaque espèce d'arbre et par région,
un catalogue de référence permettant de reconstituer la
variabilité climatique. Le comptage des cernes d'arbre est
théoriquement assez simple
La dendrochronologie apporte, en plus d'une datation, de
nombreux renseignements dans les domaines climatiques et écologiques
à partir de la structure du bois. L'analyse du cambium a amené
certains auteurs comme Worbes (2002), à rappeler l'importance des
phytohormones des bourgeons, de l'anatomie du bois à travers les
propriétés du xylème (arrangement des vaisseaux, la
structure du parenchyme et des fibres du bois).
Le comptage des cernes d'arbre est théoriquement assez
simple. Cependant, la formation des cernes se fait grace aux assises du cambium
situé sous l'écorce. A l'intérieur de chaque cerne, on
peut distinguer la période de saison sèche (poreux et clair) et
celle de la saison des pluies (dense et foncé). L'épaisseur des
cernes dépend de la pluviométrie. Pendant les années
pluvieuses, le cerne est plus large, alors que durant les années
sèches, le cerne est plus étroit. Pour une même
espèce, dans une même zone climatique, les cernes de croissance
évoluent en théorie de la même manière. La
différence entre espèces de la même zone réside dans
la vitesse de croissance de celles-ci. On peut alors, après avoir
mesuré l'épaisseur des cernes sous loupe binoculaire
établir une courbe de référence normalisée pour une
région climatique donnée.
L'épaisseur ou la densité des cernes est donc,
d'une part, un indicateur de l'histoire du climat passé et, d'autre
part, un point de repère pour dater une période de croissance. On
peut mesurer les cernes et en tirer des graphiques qui représentent
l'évolution de la croissance d'un arbre. Les années sont
représentées sur la ligne horizontale de gauche à droite
(abscisse), l'épaisseur des cernes figure sur la ligne verticale
(ordonnée).
L'analyse des cernes est largement utilisée en
foresterie, botanique, climatologie et en sciences de la terre. Les
différentes applications connues montrent que la dendrochronologie est
très utile en climatologie, en écologie forestière et en
gestion des ressources forestières (Stahle, 1999). La dimension
changement climatique et potentiel de séquestration du carbone est une
nouvelle possibilité d'application insuffisamment explorée. En
effet, certains chercheurs sont souvent découragés par les
difficultés liées à leur mesure du fait de la forte
variabilité climatique qui affecte la formation des cernes des
espèces tropicales (Worbes, 1995). La question posée est de
savoir si les cernes visibles sont annuelles ou pas ? Est-ce que le comptage et
les mesures sur les cernes permettent de reconstituer la variabilité
climatique ? Quelles sont les espèces dominantes qui ont un réel
potentiel de datation avec la dendrochonologie ?
Les études en Afrique sont rares et portent sur
quelques sites et sur quelques espèces. Les références sur
les espèces tropicales sont éparses même s'il a
été prouvé la présence de cernes clairs sur
certaines espèces, notamment celles qui ont un bois relativement dur.
Beaucoup d'études ont montré que la mesure des cernes des
espèces tropicales est un exercice difficile, même si la
synthèse établie par Stahle (1999) prouve qu'il existe de grandes
possibilités non exploitées pour ces espèces.
C'est pour cette raison et pour consolider les
résultats obtenus au chapitre 3 et 4 sur la dynamique des
espèces, que nous proposons une analyse plus fine de la dynamique de
croissance des espèces en se basant sur la croissance des cernes.
Ce chapitre porte sur l'analyse de 5 espèces dominantes
des savanes inventoriées que sont : Combretum glutinosum,
Pterocarpus erinaceus, Acacia macrostachya, Terminalia macroptera et Cordyla
pinnata. Ces espèces ont été les plus
représentatives en termes de biomasse dans les Forêts
Classées étudiées. Le but visé est de
compléter les échelles d'analyse qui portent sur les groupements
(données inventaires) ; le suivi de groupes d'espèces
(données des parcelles permanentes), et le niveau spécifique
(suivi de la croissance des individus de différentes espèces).
Section 9.02 V.1. Méthode
Le net découpage saisonnier des zones soudaniennes et
soudano-guinéennes étudiées dans ce travail nous donne un
certain espoir sur l'utilisation de la dendrochronologie pour l'analyse de
l'impact de la variabilité climatique sur les arbres. Cependant, la
saisonnalité climatique ne garantit pas forcément une perception
nette des cernes. Les difficultés sont inhérentes à
l'espèce et à la variabilité intra-saisonnière des
pluies.
Les méthodes dendrochronologiques sont nombreuses et
peuvent être classées selon qu'elle soit destructive (collecte
d'échantillons de bois impliquant la coupe de l'arbre) ou non
destructive (carottage ou méthode électrique). Le plus souvent,
l'analyse du bois (technologie du bois) implique l'utilisation des sections du
tronc d'arbre pour analyser les propriétés du bois
(densité, cernes, structure, couleur). Cependant, certaines techniques
modernes cherchent à analyser la dynamique de croissance du cambium en
mesurant sa résistance électrique. Les différences de
résistance traduisent une variation de la teneur en hydrocarbone due aux
phases de dormance et de croissance. Le Shigomètre est un type
d'instrument utilisé à cet effet (Worbes, 1995) mais
l'interprétation des résultats issus de cette technique n'est pas
aisée.
Les méthodes les plus éprouvées pour
l'analyse des cernes des arbres sont basées sur les disques de tronc
prélevés après abattage de l'arbre (méthode
destructive). L'utilisation des dendromètres permet une mesure continue
des diamètres de cernes et donc de l'activité cambiale qu'on peut
alors comparer avec les données climatiques.
l'échantillon et en faisant ressortir le caractère
effectif de la croissance du bois plutôt que de se limiter à
l'enregistrement d'une variable non appréciée.
Pour être plus précis dans la
caractérisation de la dynamique de croissance d'un arbre, les
échantillons de bois font l'objet d'une analyse anatomique qui consiste
à déterminer la structure du bois pour mieux définir les
lignes de discontinuité. L'analyse de la densité des
différents cernes est aussi une pratique courante. Cette technique
utilise la technologie des rayons X pour identifier la délimitation des
cernes du fait de la variation de densité (ce type d'analyse fera suite
à ce travail). Ainsi, la variation de densité présente une
forte relation avec la saisonnalité climatique. La densitométrie
aux rayons X n'a pas donné de bons résultats sur les
espèces tropicales du fait de la structure complexe du bois des
angiospermes et des nombreuses variations intra-annuelles (Worbes, 2002).
D'autres techniques utilisent la variation des isotopes stables du bois comme
le C13, mais cette méthode est coûteuse et les résultats
parfois difficilement comparables à la variabilité climatique.
Pour comparer la largeur des cernes avec la variabilité
climatique, ces derniers doivent être transformés en indice de
variation (standardisation) pour des besoins de comparaison. Les
précipitations souvent utilisées pour les comparaisons en milieu
tropical sont des cumuls mensuels. Les séries temporelles des
précipitations mensuelles sont comparées à celles des
cernes d'arbres pour mettre en évidence l'influence des
précipitations sur la croissance des arbres.
Il est aussi important de mentionner l'importance de
procéder aux techniques de datation croisée. Cette technique
consiste à mettre en correspondance la largeur des cernes d'un individu
avec des caractéristiques structurelles des arbres d'autres
espèces dans la zone étudiée pour identifier
l'année exacte de la formation des cernes. La datation croisée
des cernes est une méthode de vérification et de consolidation de
l'analyse dendrochronologique et dépend ainsi de leur reproduction
adéquate. Elle est basée essentiellement sur le principe selon
lequel l'effet des facteurs climatiques affecte différemment les cernes
selon les espèces. Certaines espèces ont naturellement des cernes
étroits, d'autres n'en ont pas pour la même saison, au moment
où certaines espèces ont des cernes très distincts. On
utilise alors les espèces qui ont des cernes nets de cette saison pour
combler les lacunes sur les autres espèces.
L'approche utilisée dans cette étude a
consisté à réutiliser les disques d'arbre qui ont servi
à estimer le taux d'humidité dans le cadre de
l'élaboration des modèles allométriques. Les
échantillons ont été polis en utilisant du papier de
menuiserie pour mieux exposer les cernes (Stahle, 1999 ; Tarhule et Hughes,
2002).
Section 9.03 V.2. Etat des échantillons et
procédure de mesure
des cernes
Les échantillons analysés sont ceux des
espèces principales qui ont servi à la mise en place des
modèles allométriques (cf. Chapitre 2). Les études
classiques utilisent une stratégie qui consiste à
sélectionner les espèces reconnues pour le caractère
distinct des cernes (Stahle, 1999 ; Stahle et al., 1999). Cette option
n'était pas possible pour ce travail, puisqu'il n'a pas
été recensé de travaux sur les espèces dominantes
du Sénégal.
Une comparaison avec les cernes distincts observés sur
les arbres des régions tempérées (comme le Pin Rouge) a
permis de constater un grand flou sur les échantillons de savanes
analysés dans ce travail. Tahrule et Hughes (2002) montrent qu'en savane
Africaine les espèces comme Combretum glutinosum, Acacia
macrostachya et Pterocarpus erinaceus sont difficilement
utilisables pour ce type d'analyse. Il s'agit pourtant d'espèces
très fréquentes. Ces auteurs soutiennent qu'on pourrait se
rabattre sur des espèces comme Cordyla pinnata qui
présente une netteté relative de leurs cernes.
Cependant, certains échantillons pourraient être
mesurés en utilisant les algorithmes d'analyse d'image (exemple:
CDendro:
http://www.cybis.se/forfun/dendro/index.htm).
Ces techniques passent par des filtres, la détection des bordures qui
sont couramment dans le domaine de télédétection. On a
préféré toutefois utiliser une loupe binoculaire pour
détecter les cernes (iVelmex :
http://www.velmex.com/manual
tree ring sys.html). A chaque fois que cela a été possible,
on a mesuré la largeur des cernes dans deux directions opposées
en allant de l'extérieur vers le centre. Dans certains cas cette double
mensuration n'a pas été possible.
Les mesures ont été très difficiles avec
les échantillons de Cordyla pinnata et Combretum
glutinosum. Les cernes sont le plus souvent discontinus ou disparaissent
en se fusionnant avec d'autres cernes ou en se biseautant. Dans de telles
situations, l'âge des échantillons et la largeur des cernes
deviennent difficiles à déterminer. Dans certains cas, il
était simplement impossible de procéder à l'analyse des
cernes du fait d'une lecture très difficile de ces dernières.
Pour analyser les cernes les échantillons ont
été polis (papier-sable) pour mieux mettre en exergue la
structure du bois. Pour chaque disque, deux itérations ont
été faites quand c'est possible dans deux directions
perpendiculaires (figure 73).
Figure 73. Méthode de mesure de
l'épaisseur des cernes.
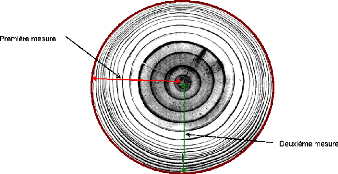
L'option pour la mesure des cernes dans deux directions a
été choisie pour minimiser les biais liés à la
variabilité de la largeur des cernes dans le même
échantillon. La moyenne des deux mesures a été faite pour
chaque échantillon. Pour certains échantillons on s'est
contenté d'une seule mesure. Au total 48 échantillons ont
été analysés sur les 67 soit 70% des échantillons
préparés pour cette analyse. Certains de ces échantillons
très jeunes n'ont pas été inclus dans l'analyse
croisée avec la pluviométrie. Le tableau 20 fait le point sur les
traitements faits des échantillons.
Tableau 20. Récapitulatif des analyses
dendrochronologiques
Echantillons datables
|
Espèce
|
année
|
collecte
|
âge
|
observation
|
|
ACAM01
|
1983
|
2007
|
25
|
|
|
ACAM04
|
1989
|
2007
|
19
|
|
|
ACAM04
|
2002
|
2007
|
6
|
Tj
|
|
ACAM06
|
1996
|
2007
|
12
|
Tj
|
|
ACAM08
|
2000
|
2007
|
8
|
Tj
|
|
ACAM09
|
2000
|
2007
|
8
|
Tj
|
|
ACAM10
|
2001
|
2007
|
7
|
Tj
|
|
ACAM11
|
2000
|
2007
|
8
|
Tj
|
|
ACAM12
|
1997
|
2007
|
11
|
Tj
|
|
COGL6PA
|
1986
|
2007
|
22
|
|
|
COGL7PA
|
1988
|
2007
|
20
|
|
|
COGL8PA
|
1987
|
2007
|
21
|
|
|
COGL9PA
|
1987
|
2007
|
21
|
|
|
COGL12PA
|
1973
|
2007
|
35
|
|
|
COGL14OA
|
1990
|
2007
|
18
|
|
|
COGL15OA
|
1991
|
2007
|
17
|
|
|
COGL16OA
|
1984
|
2007
|
24
|
|
|
COGL17OA
|
1982
|
2007
|
26
|
|
|
COGL18OA
|
1988
|
2007
|
20
|
|
|
COGL31KA
|
1947
|
2007
|
61
|
|
|
COGL33KA
|
1976
|
2007
|
32
|
|
|
COGL35KA
|
1984
|
2007
|
24
|
|
|
COPI2PA
|
1967
|
2007
|
41
|
|
|
COPI3PA
|
1988
|
2007
|
20
|
|
|
PTER1P
|
1989
|
2007
|
19
|
|
|
PTER2P
|
1989
|
2007
|
19
|
|
|
PTER3P
|
1980
|
2007
|
28
|
|
|
TEMA1P
|
1993
|
2007
|
15
|
|
|
TEMA2K
|
1987
|
2007
|
21
|
|
|
TEMA3K
|
1999
|
2007
|
9
|
Tj
|
|
TEMA4K
|
1987
|
2007
|
21
|
|
|
TEMA5K
|
1979
|
2007
|
29
|
|
|
TEMA6K
|
1996
|
2007
|
12
|
|
|
TEMA8K
|
2001
|
2007
|
7
|
Tj
|
|
TEMA9K
|
1999
|
2007
|
9
|
Tj
|
Echantillons non datables
|
espèce
|
année
|
collecte
|
âge
|
observation
|
|
ACAM02
|
|
2007
|
|
Ts
|
|
COGL1WA
|
|
2007
|
|
Ts
|
|
COGL3WA
|
|
2007
|
|
RBND
|
|
COGL4WA
|
|
2007
|
|
RBND
|
|
COGL5PA
|
|
2007
|
|
Ts
|
|
COGL23BA
|
|
2007
|
|
RBND
|
|
COGL27BA
|
|
2007
|
|
TS
|
|
COGL30KA
|
|
2007
|
|
TS
|
|
COPI1PA
|
|
2007
|
|
TS
|
|
COPI4PA
|
|
2007
|
|
TS
|
|
COPI4PA
|
|
2007
|
|
TS
|
|
COPI5OA
|
|
2007
|
|
TS
|
|
COPI6OA
|
|
2007
|
|
TS
|
ACAM : Acacia macrostachya COGL
: Combretum glutinosum PTER :
Pterocarpus erinaceus TEMA : Terminalia
macroptera COPI : Cordyla pinnata
Tj : Trop jeune
Ts : Cernes très biseautés RBND : Cernes pas
distincts
Les données sur la croissance des cernes ont
été comparées à la dynamique des
précipitations. Les courbes cumulées de la croissance des cernes
ont permis d'analyser les modèles de croissance des espèces de
savane.
Section 9.04 V.3. Résultats
Les cernes des espèces tropicales ne sont pas
suffisamment distincts pour être nettement identifiables sur une loupe
binoculaire. Les images de la figure 74 montrent la nature floue de la
structure des cernes et nous édifient sur les difficultés
subséquentes pour les analyser. La photographie de Pinus
resinosa, espèces tempérée, (figure 74, Photo n°
6) a été introduite pour avoir un élément de
comparaison avec les espèces des écosystèmes
tempérées qui présentent des cernes bien distincts.
La théorie classique de la croissance des arbres n'est
pas très apparente sur l'évolution des cernes des plantes de la
végétation naturelle des savanes. Il est soutenu que les arbres
croissent plus rapidement à leur jeune age et la croissance ralentit
quand l'arbre vieillit. On observe bien sur l'échantillon 6 que cette
théorie s'applique bien aux espèces tempérées.
Mais
l'observation des échantillons de savane ne nous permet
pas de faire cette déduction sigénéralement
acceptée.
Les hypothèses qui peuvent être
évoquées sont relatives aux facteurs de stress soumis à la
plantule et qui limitent sa croissance rapide (compétition avec les
herbacés, sols squelettiques peu profonds, feux de brousse, fortes
températures). Ces facteurs réduisent la croissance et parfois
éliminent certains cernes. En définitive, la rapidité de
la croissance n'est pas exclusivement liée au cumul annuel de la
pluviométrie. La distribution spatiale et temporelle des pluies est
aussi un élément déterminant.

Figure 74. Photographies de
quelques échantillons de disques
5
3
1
6
2
4
1 : Terminalia macroptera 2 : Pterocarpus
erinaceus
3 : Cordyla pinnata 4 : Combretum glutinosum
5 : Acacia macrostachya 6 : Pinus
resinosa (espèce tempérée,
référence)
Les résultats obtenus se démarquent de ce que
les dendrochronologues ont observé dans les écosystèmes
tempérés, en l'occurrence la bonne correspondance entre la
pluviométrie annuelle et la largeur des cernes. En milieu tropical la
nature des espèces, les différents facteurs stationnels et les
contraintes exercées sur la plantes se traduisent par une
évolution quasi erratique des cernes. Cette absence de
régularité dans la croissance des arbres rend difficile la mise
en relation entre la croissance des cernes et les facteurs écologiques,
de même que la pluviométrie, décrite comme le facteur
majeur de développement des plantes. La figure 75 montre plusieurs cas
de figures :
- grand décalage entre la pluviométrie et la
croissance des cernes (ACAM01 et ACAM04) ;
- faible décalage entre la pluviométrie et la
croissance des cernes (TEMA4) ;
- bonne correspondance entre la croissance des cernes et la
pluviométrie (COGL31, COGEL12, COPI2).
Ce déphasage entre les fortes précipitations
annuelles et la faible croissance de cernes est plus caractéristiques
chez les jeunes sujets qui ont un système racinaire peu profond et qui
sont plus exposés à la compétition. La reprise de
croissance de ces jeunes sujets après le feu est aussi toujours plus
lente du fait d'une faible efficacité dans l'utilisation de
l'humidité du sol. Cette logique s'applique aussi pour certains sujets
adultes.


Figure 75. Analyse de la coévolution
entre les précipitations et la croissance des cernes
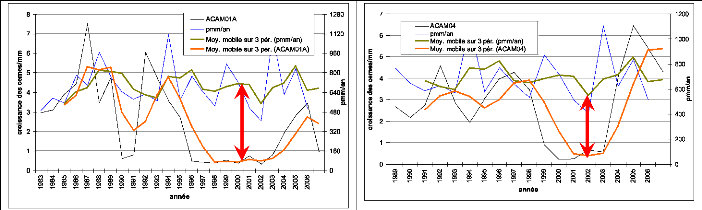
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5 4,5 4 3,5 9
3
8000
000
,
|
|
moileu
|
|
100
900
600
900
800
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
C C 6 3
60
pmm/a
|


|
12
|
pmm/an
|
900
|
|
|
|
|
|
10 9 8
E 7
E in
o 6
E
8
a 5
m
-0
. 4
a
co
3
2
1
|
|
pmm/an
PTER2P
PTER3P
Moy. mobile sur 3 per. ( PTER2P) Moy. mobile sur 3 per. (
PTER3P) Moy. mobile sur 3 per. (pmm/an)
|
|
1000
900 800 700 600
500 -E'
E o.
400
300
200
100
|
|
10
E
E 8
Ft
i
6
:
'0
c
·
co
e 4
2
2
|
PIER3PA
Moy. mobile sur 3 per. (pmm/an) Moy. mobile sur 3 per.
(PTER3PA)
|
|
750
600
.!!
450 E
E
0.
300
150
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 0
|
|
?o) 8 0) Si 0) 8 8 8 8 2 0) 8 8 8 8 8 8
CO 8
CD 0) 0) 0) 0) 0) 0) 0) 0) 0) 0) 0 0 0 0 0 0 0
0
NNNNNNNN
années
|
|
0 s
8 `,,,' g 8 2 8 `al 1 8 8 8 8 8 CO8
0) 0) 0) 0) 0) 0) 0) 0) 0) 0) 0 0 0 0
N N N N
année
|
|
|
TEMA5K
|
|
|
|
|
9
8
7
o
0
E 6 8
m5 8 15
4
i
.g 3
1
|
|
1500 1350 1200 1050
900
c
i
75E
E
600o.
450
300
150
|
|
6
E E if / PT
Mo3
a 0 3 Mo
8
c
Q9n 3 2 mo
8 mo
1
0
|
pmm/an
Moy. mobile sur 3 per. (TEMA5K) Moy. mobile sur 3 per.
(pmm/an)
|
|
1500
1000
c
-,°
750E
0. 500 0
0
|
|
|
5
1250
|
|
|
|
|
|
|
|
|
|
2
|
TEMA4K
pmm/an
Moy. mobile sur 3 per. (TEMA4K)
Moy. mobile sur 3 per. (pmm/an)
o 0
|
|
|
|
|
|
8 8 0 8 CT) cal) Ca
0) '8 8 0) 0) 8 E ccJ 8 8
'é,' 8 8
0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, 0, c, c, c, c, c, c,
c, c,
N NNNNNNN
année
|
|
P-.) 8 g 8 2 g 2 8 F 2
0) 8 0) `01 ca a s (8 18 8 0) 8 8
8 8 300 8 8 F 90
a, a, a, a, a, a, a,a, a, a, a, a, a, a,
a, a, a, a, a, a, on ono ono
NNNNNNNN
50 200 75
année
PTERa, :
3 Pterocarpus erinaceus
|
|
0
ACAM : Acacia macrostachya
2 TEMA4K
pmm/an
COGL: Combretum glutinosum
m
0 n H
o
r 4 Pz'
cz(-"' .4, ,t
4 2 .4
Cr. c., cz
'1 t..> . c%
co A Q
4
4 A Z
ON q ct,
z a N. r, cz .., ,
Q
|
COPI: Cordyla pinnata
TEMA: Terminalia macroptera
|
Les vitesses de croissance sont très variables selon
les espèces et leurs conditions stationnelles de croissance. Des
espèces comme Pterocarpus erinaceus
échantillonnées dans la Forêt Classée de Patako
peuvent avoir des maximums de croissance annuelle de 10-12 mm/an, alors que
leurs minimums peuvent être très faibles jusqu'à atteindre
moins de 1 mm/an14.
Cordyla pinnata présente parfois une
croissance assez rapide avec des maximums qui tournent autour de 6-7,5 mm/an et
des minimums faibles de moins de 1 mm/an.
Terminalia macroptera croît en conditions
favorables entre 5-7,7 mm/an avec des minima inférieurs à 1
mm/an.
Acacia macrostachya, se développe avec des
maximums de 6,5-7 mm/an et des minimums inférieurs à 1 mm/an
Pour Combretum glutinosum les maximums de croissance
des individus sont entre 4,5-5,5 mm/an et les minimums sont inférieurs
à 1mm/an.
Les variations moyennes de croissance du DBH
présentées dans le chapitre 3 confirment cette croissance
relativement lente comparée aux espèces équatoriales qui
peuvent avoir une croissance soutenue de plus de 8-9 mm/an (Worbes et
al., 2003).
On constate sur la plupart des échantillons une sorte
de décalage entre la croissance des cernes et la pluviométrie.
Les fortes pluies se traduisent par une relative forte croissance mais avec un
déphasage variable d'au moins une ou deux années. Ce constat fait
penser que la plante ne tire pas immédiatement profit des fortes
précipitations.
La difficulté de faire la relation entre pluie et
croissance des arbres apparaît déjà avec les courbes de
coévolution (figure 76). Il est important tout de même d'analyser
les anomalies à travers des valeurs standardisées de croissance
par la méthode des écarts moyens
Z= xi-moyenne/écart type
La figure 76 montre quelques paradoxes entre les cumuls
pluviométriques et la croissance des cernes. Sur quelques années
la pluviométrie a été positive et la croissance des cernes
négative ou inversement.
14 Dans les parcelles de suivi de la végétation,
avec le suivi du DBH, Pterocarpus erinaceus présente une
moyenne de croissance annuelle de 4,2 mm/an, Cordyla pinnata de 3,8
mm/an ; Terminalia macroptera de 4,1 mm/an ; Combretum
glutinosum de 1,7 mm/an ; (voir chapitre 3).


Figure 76. Variation des écarts
normalisés de croissance des cernes et de la pluviométrie
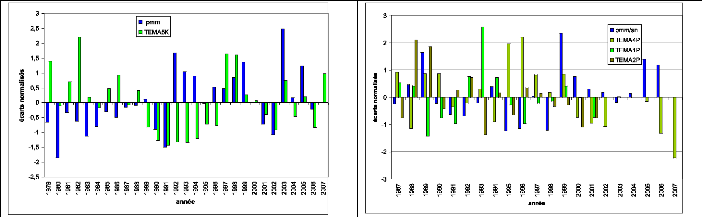
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2,
PT
pmm/
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1,
ecarts a la . C
N J
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
r:,
1967
1969 1971 1973 1975 1977 1979 1981 1983 1985
as
1987
·
1989
1991 1993 1995 1997 1999 2001 2003 2005 2007
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
PTER : Pterocarpus erinaceus ACAM : Acacia
macrostachya COGL: Combretum glutinosum COPI: Cordyla
pinnata
TEMA: Terminalia macroptera
|
|
3
0
1
(karts normalises
..Is . E, p -, n
3 0 -, 0 0 0 -
· 0 N3 C
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
L'absence de correspondance dans la variation des
précipitations et la croissance des cernes n'est pas un
phénomène particulier à une espèce donnée.
Elle est observée dans la plus part des échantillons à des
moments donnés de leur croissance.
La relation entre la précipitation et la croissance des
cernes dépend de plusieurs facteurs qui influencent directement ou non
la vitesse de croissance des arbres. Parmi ceux-ci on peut retenir les points
qui suivent.
- La distribution spatiale et temporelle des
précipitations
Dans la zone sahélienne on observe une forte
variabilité climatique (chapitre 1) qui se manifeste par une variation
interannuelle des cumuls pluviométriques. Les saisons pluvieuses sont
entrecoupées par des épisodes secs qui freinent la croissance des
arbres. A l'intérieur de chaque saison on peut observer de longues
pauses pluviométriques qui ralentissent les quantités d'eau
absorbées par les plantes. Ces deux facteurs sont soulignés dans
plusieurs travaux (Denett et al., 1985 ; Malo et Nicholson, 1990 ;
Berger et al., 1996 ; Dahlberg, 2000) qui montrent la relation entre
la variation pluviométrique et la production végétale. Il
faut toutefois noter que si les températures constituent un facteur
central à la phénologie des plantes en milieu
tempéré, dans le domaine tropical par contre la quantité
et la distribution temporelle de l'eau de pluie est le facteur essentiel, les
températures n'étant pas un facteur limitant.
- L'influence des feux de brousse
Les savanes africaines sont connues pour les nombreux feux de
brousse qui affectent directement la composition et la dynamique de la
végétation. De nombreuses études sur l'écologie des
feux dans les savanes prouvent l'effet direct de la chaleur sur la croissance
des arbres (Ahlgren et Ahlgren, 1960 ; Janzen, 1967 ; Afoloyan, 1978 ;
Brookman-Amissah et al., 1980 ; Trabaud, 1987; Sabiiti et
al., 1988; Ehrlich et al., 1997; Arthur et al., 1998).
De nombreux individus échantillonnés ont subi des blessures dues
aux feux de brousse qui créent un stress réduisant la croissance
normale de la plante.
- Les conditions écologiques
Les conditions stationnelles sont souvent
évoquées dans la caractérisation du spectre biologique, la
structure et les facteurs de développement des plantes (Arrhenius, 1921;
Barnes et al., 1951; Adams, 1967; Hills et al., 1968; 1979;
Akpo, 1997; Dahlberg, 2000). Les différences dans la production de
biomasse des plantes dépendent de l'efficacité physiologique des
plantes mais aussi des conditions écologiques qui expliquent que pour
une même espèce on observe des rythmes de croissance très
variés selon les conditions du milieu.
Sur plusieurs échantillons de la même
espèce, on note des rythmes de croissance très différents
ne permettant pas d'ailleurs d'avoir une bonne relation entre l'âge et la
biomasse totale. Les conditions écologiques agissent notamment sur la
disponibilité de l'eau et des sels minéraux, la profondeur
racinaire, la compétition intra spécifique, etc.
La vitesse de croissance des arbres dépend d'une
conjonction de circonstances qui fait jouer le type de sol, la distribution et
les quantités de pluies, la composition spécifique et la
concurrence entre espèces, les facteurs de stress comme la
température et les feux de brousse. En milieu tropical, la force de
chacun de ces facteurs peut favoriser ou non la dynamique de croissance de la
plante. Il est par conséquent très rare d'avoir une dynamique
régulière de croissance. Les facteurs tels que le feu et les
fortes températures sont si sévères que la plante
croît de façon plus lente comparée à certaines
espèces du milieu tempéré.
Tous ces facteurs font que la théorie écologique
qui consiste à dire que les plantes jeunes croient plus rapidement que
les vieux sujets est à considérer avec prudence pour les
espèces tropicales. Ce constat pourrait être lié aux
hypothèses suivantes :
- la jeune plantule non affranchie est plus sensible au stress
lié au feu, à la température et à l'aridité
et croît très lentement au début selon les conditions du
milieu ; le démarrage peut donc être très lent (figure
77, Combretum glutinosum) ;
- la plante affranchie est plus résistante aux feux et
dispose d'un enracinement plus profond qui lui permet d'être plus
résilient à l'aridité ; sa croissance peut être plus
rapide que celle de la jeune plantule (figure 77, Terminalia
macroptera) ;
- c'est à la dernière phase de sa croissance que
l'effet de la sénescence se fait sentir sur la plante avec un
ralentissement de la croissance.
La figure 77, qui est une courbe cumulée de la
croissance de quelques individus, montre des cas très différents
les uns des autres, rendant difficile une généralisation sur le
rythme de croissance des plantes dans les savanes. Il est aussi très
difficile d'admettre que les jeunes plantes ont toujours une croissance
très rapide vu le nombre de facteurs comme les feux, la
compétition, le type de sol et l'enracinement, qui entrent en jeu.

Figure 77. Courbe cumulée de la
croissance des cernes de différents individus.
Pterocarpus erinaceus (PTER3PA)
Combretum glutinosum (COGL31KA) Combretum
glutinosum (COGL12KA)
Cordyla pinnata (COPI12)
Acacia macrostachya (ACMA01)
Terminalia macroptera (TEMA5K)
Ces 6 exemples nous ont permis de déterminer deux
grandes catégories de croissance. Une
croissance normale qui
correspond aux théories écologiques et qui montre que les
plantes
0
jeunes sont plus rapides en croissance (modèle de
croissance classique ; exemple COGL12) ; 0 une autre qui correspond à la
réalité de la plupart des espèces de savanes avec une
croissance
3
lente au début, rapide après affranchissement et
qui se ralentit à la sénescence. Cette réalité
0
5
t-r
CD
P,
r3
1-1-
-
C2
.1
CD
-4
00
montrée à la figure 78.
0 se voit sur PTER3PA (Figure 77). Une
représentation de ces deux modèles de croissance est
Dans ces différents cas on observe une forte
irrégularité de la croissance des arbres due à
différents facteurs écologiques et climatiques.
Figure 78. Modèles de croissance des
espèces de savane en comparaison avec celui des espèces
tempérées.
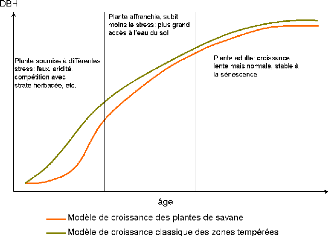
L'influence de tous ces facteurs fait qu'il est difficile
d'avoir une relation claire entre l'âge des individus et la biomasse
correspondante. Le coefficient de détermination très faible
(R2=0,47) de la figure 79 confirme la faiblesse de cette relation
âge-biomasse.
Figure 79. Relation âge-biomasse
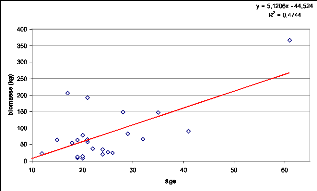
Ces résultats sont conformes à ceux de Worbes
et al. (2003) sur des échantillons du Cameroun qui ont
montré que l'âge des arbres est peu corrélé avec le
diamètre du tronc et la hauteur des individus.
- Analyse des tissus du bois

Figure 80. Agrandissement des
échantillons de bois des 5 espèces analysées
Acacia macrostachya
Combretum glutinosum
Cordyla pinnata
Pterocarpus erinaceus
Localisation des discontinuités sur les
échantillons considérés. Grossissement : 20 x ??
Terminalia macroptera
L'analyse de la structure du bois des échantillons
montre que le potentiel dendrochronologique existe pour certaines
espèces de savane. Le comptage des cernes posent cependant des
difficultés liées à la suppression de certains cernes et
le caractère flou des discontinuités. La figure 80
représente des agrandissements de certains échantillons de
bois.
L'observation des discontinuités par photographies
(agrandies ~20X) montre la difficulté structurelle d'identifier les
discontinuités entre cernes successifs. Les espèces
présentées ici ont un certain potentiel de datation mais le
détail des cernes ne garantissent pas toujours une analyse
systématique de la croissance des arbres. Sur un lot de base de 67
individus seule une trentaine a pu être analysée.
Section 9.05 Conclusion et discussions
Ce chapitre montre à la fois la complexité de
l'analyse dendrochronologique des échantillons tropicaux, mais aussi le
potentiel de cette méthode pour la datation de certaines espèces
et le suivi de la croissance des arbres. La mise en relation entre la
croissance des cernes et la pluviométrie montre qu'il n'est pas
aisé d'établir une relation significative. La raison est
liée à l'importance d'autres facteurs comme les feux de brousse,
l'ampleur des sécheresses, la compétition entre espèces,
la nature du sol et l'enracinement des arbres. Le caractère erratique de
la contribution de ces différents facteurs ne facilite pas le
développement de modèles ou la structuration de théories
robustes sur la croissance des espèces de savanes. Le rythme de
croissance des individus échantillonnés montre le plus souvent
une contradiction avec certaines théories écologiques qui
admettent que les jeunes plantes croissent plus vite que les adultes. Dans les
écosystèmes de savane, la croissance des arbres dépend de
la combinaison de plusieurs facteurs, dont certains sont favorables et les
autres sont des contraintes au développement des plantes. Ainsi, chaque
essence réagissant différemment aux nombreux facteurs qui
déterminent la croissance, il est donc nécessaire de les traiter
séparément et avec beaucoup de précautions.
Il y a des cas d'extrême difficulté de lecture
due à la densité de succession des cernes qui rend incertaine la
limite de chacun d'entre eux. Les travaux de Worbes (1995) soulignent que les
cernes annuels sont plus nets quand on a une année pluviométrique
monomodale et courte de 2-3 mois. Dans les écosystèmes où
les précipitations annuelles sont bimodales on a tendance à avoir
deux cernes (cas de l'Afrique de l'Est et de pays comme la Côte
d'Ivoire). Dans les zones équatoriales où la distinction des
saisons n'est pas nette, les cernes observés ne peuvent pas être
connectés à la variabilité du climat (Worbes, 2002). Dans
certains cas, la formation des cernes est liée à l'occurrence de
la crue qui, du fait de l'immersion permanente entraîne des conditions
d'anoxie qui réduisent l'activité racinaire, ce qui
entraîne une dormance du cambium (Worbes, 1995) du fait du déficit
d'eau. Les analyses de Daniellia oliveri par Tarhule et Hughes (2002)
montrent une présence de doubles cernes qui alternent avec les
séquences de cernes nets. Certaines difficultés liées aux
blessures sur les troncs ont été signalées par Worbes
(1995) et Tarhule et Hughes (2002). Avec l'impact de l'exploitation de l'homme
(bois, écorce, feux de brousse), certains cernes sont très flous,
ou incomplets.
Pour certaines espèces comme Isoberlina doka,
les cernes développés en période de sécheresse sont
très proches pour être discriminables. La sécheresse peut
entraîner aussi des déviations ou fusions de cernes qui
s'opèrent de façon irrégulière.
Les études en Afrique de l'Ouest de Tarhule et Hughes
(2002) révèlent que la potentialité d'utiliser les cernes
pour l'analyse de la croissance des arbres donne des performances variables
selon les espèces. Ces auteurs ont ainsi identifié trois
catégories d'espèces. Celles qui permettent d'avoir de bons
résultats comme Cassia sieberiana, Cordyla pinnata, Daniellia
oliveri, Isoberlina doka, Tamarindus indica, Acacia seyal et Gmelina
arborea ; et celles qui donnent des résultats passables comme
Afzelia africana, Burkea africana, Detarium microcarpum, Acacia
polyacantha, Boscia senegalensis, Prosopis africana, Pericopsis
laxiflorus.
Selon cette étude, il existe une liste d'espèces
qui ne permettent pas d'analyser de façon adéquate les cernes des
arbres. Dans ce groupe on note Lannea acida, L. microcarpa, L. velutina,
Sclerocarya birrea, Bombax costatum, Bauhinia rufesencs, Anogeissus leiocarpus,
Combretum glutinosum, C. micranthum, C. nigricans, C. paniculatum, Terminalia
laxiflora, T. macroptera, Diospyros mespiliformis, Hymenocardia acida,
Strychnos spinosa, Khaya senegalensis, Faidherbia albida, Acacia macrostachya,
Dichrostachys cinerea, Parkia biglobosa, Entada africana, Ficus sp.,
Pterocaprus erinaceus, Crossopteris febrifuga, Mitragyna inermis, Gardenia sp.,
Vitellaria paradoxa, Sterculia setigera, Vitex doniana et Balanites
aegytiaca.
Nous notons cependant, à travers les analyses faites
dans ce travail que quelques unes des espèces jugées comme peu
utiles pour l'analyse dendrochronologique, ont un réel potentiel pour la
datation et l'analyse de la croissance des cernes : il s'agit de Combretum
glutinosum, Terminalia macroptera, Acacia macrostachya et Pterocaprus
erinaceus. La différence avec les autres auteurs peut être
liée à la nature des sites étudiés et les
différences dans le déroulement des saisons
pluviométriques. En effet dans de nombreuses zones tropicales on observe
deux saisons des pluies qui font que les cernes annuelles sont difficilement
décelables ; ce n'est pas le cas des zones soudaniennes et
soudano-guinéennes.
Une autre source de différence sur les résultats
s'articule autour de la méthode d'analyse. Tarhule et Hughes (2002) ont
utilisé des échantillons par carottage qui présentent
moins de détails que les disques prélevés sur des troncs.
Mais il est évident que la plupart des espèces de savanes
citées dans leur travail présente des niveaux de
difficultés variés, pour l'analyse dendrochronologique. Selon
Tarhule et Hughes (2002), les Caesalpiniaceae donnent souvent de bons
résultats.
Cependant, Worbes, (2002) a donné un tableau
détaillé qui évalue la discrimination des cernes en
fonction de la position de ces derniers par rapport au centre du tronc. Il en a
conclu que certaines espèces comme Pterocarpus erinaceus, Swartzia
sp., Annona sp, ont des cernes nets, alors que d'autres comme
Spondias mombin, Ceiba pentandra et Cordia sp, ne permettent
aucune analyse dendrochonologique avec les technologies actuelles. Entre ces
deux extrêmes on a des espèces qui sont datables malgré
quelques difficultés.
L'une des difficultés de la dendrochronologie tropicale
réside selon Worbes (2002) sur l'asynchronie phénologique des
espèces. Certaines espèces perdent les feuilles de façon
prématurée et d'autres tardivement. La présence de
feuilles sur l'arbre est un indicateur de la dynamique de la
photosynthèse qui est la base de l'assimilation et de la production de
biomasse. Les travaux de Chhin et Wang (2008) montrent aussi que pour les
écosystèmes boréaux la croissance des cernes dépend
de l'activité de production foliaire qui s'initie une année et se
développe l'autre année. Donc une année bien pluvieuse se
traduit par une forte croissance des cernes, mais une année après
(décalage temporel). Les analyses que nous avons effectuées,
montrent l'absence de corrélation entre les précipitations
annuelles ou mensuelles avec la croissance des cernes. Un déplacement
des données de cernes d'une année n'aide pas pour autant. Donc
l'hypothèse du décalage peut être valable mais ne
correspond pas systématiquement à une année ou un à
nombre d'année précis. Chhin et Wang (2008) ont essayé de
corréler les précipitations à la superficie des bandes de
cernes, mais dans le cas des savanes, l'irrégularité des
tracés des cernes rend cette option très délicate à
opérer.
Dans les analyses, on a aussi testé la relation entre
la croissance des cernes et les précipitations du mois d'aoflt. Le
résultat n'est pas différent de celui avec les
précipitations annuelles. La raison est que les précipitations
annuelles sont très largement dépendantes des
précipitations du mois d'aoflt. La température n'étant pas
un facteur limitant sous les tropiques, cette donnée n'a pas
été testée dans cette analyse.
Pour obtenir une bonne interprétation des cernes dans
les écosystèmes de savane, il faudra faire une analyse mixte de
plusieurs espèces, à plusieurs ages pour atténuer
l'anastomose des cernes due aux facteurs évoqués.
Bibliographie
Adams, M.E. 1967. A study of the ecology of Acacia
mellifera, A. seyal, and Balanites
aegyptiaca in relation to land-clearing. Journal of
Applied Ecology, 4 (kt):221-237.
Afolayan, T.A. 1978. The effect of fires on
the vegetation in Kainji Lake National Park,
Nigeria. OIKOS., 31.
Ahlgren, I.F. Ahlgren, C.E. 1960. Ecological effects of forest
fires. The Botanical Review, 26:483-533.
Akpo, L. 1997. Phenological interactions between tree and
understory herbaceous vegetation of a sahelian semi-arid savanna. Plant
Ecology, 131:241-248.
Arrhenius, O. 1921. Species and area. Journal of Ecology, 9:-.
Arthur, M.A., Paratley, R.D. Blankenship, B.A. 1998. Single
and repeated fires affect survival and regeneration of woody and herbaceous
species in an oak-pine forest. Journal of the Torrey Botanical Society,
125:225-236.
Barnes, H. Stanbury, F.A. 1951. A statistical study of plant
distribution during the colonisation and early development of vegetation on
China clay residues. Journal of Ecology, 39:171-181.
Berger, A., Grouzis, M. Fournier, C. 1996. The water status of
six woody species coexisting in the Sahel (Ferlo, Senegal). Journal of Tropical
Ecology, 12:607-627.
Brookman-Amissah, J., Hall, J.B., Swaine, M.D. Attakorah, J.Y.
1980. A re-assesment of a fire protection experiment in North-Eastern Ghana
savanna. Journal of Applied ecology, 17:85-99.
Chhin, S., Wang, G.G. 2008. Climatic response of Picea
glauca seedlings in a forest-prairie ecotone of western Canada. Annals of
Forest Sciences. 65 :207, 8p.
Dahlberg, A.C. 2000. Vegetation diversity and change in
relation to land use, soil and rainfall - a case study from North-East
District, Botswana. Journal of Arid Environments, 44 (1):19-40.
Dennett, M.D., Elston, J. Rodgers, J.A. 1985. A reappraisal of
rainfall trends in the Sahel. J.of Climatology, 5:353-361.
Ehrlich, D., Lambin, E.F. Malingreau, J.P. 1997. Biomass
burning and broad-scale land-
cover changes in Western Africa. Remote Sensing of Environment,
61 (2):201-209.
Hills, T.L. Randall, R.E., 1968. The ecology of the
forest/savanna boundary, Montreal, 128
p.
Janzen, D.H. 1967. Fire, vegetation structure, and the ant x
Acacia interaction in Central America. Ecology, 48 (kt):26-35.
Malo, A.R. Nicholson, S.E. 1990. A study of rainfall and
vegetation dynamics in the African Sahel using normalized difference vegetation
index. Journal of Arid Environment, 19:1-24.
Sabiiti, E.N. Wein, R.W. 1988. Fire behaviour and the invasion of
Acacia sieberiana into savanna grassland openings. African Journal of
Ecology., 26:301-313.
Sagna P., 2005. Dynamique du climat et son évolution
récente dans la partie ouest de l'Afrique Occidentale, Thèse de
Doctorat d'État. Université Cheikh Anta Diop de Dakar, 778 p.
Stahle D.W. 1999a. Useful strategies for the development of
tropical tree-ring chronologies. IAWA Journal, 20 (3):249-253.
Stahle, D.W., Mushove, P.T., Cleaveland, M.K., Roig, F.,
Haynes G.A. 1999b. Management implications of annual growth rings in
Pterocarpus angolensis from Zimbabwe. Forest Ecology and Management
(124):217-229.
Tarhule, A., Hughes M. 2002. Tree-ring research in semi-arid West
Africa: need and potential. Tree-ring Research, 58 (1/2):31-46.
Trabaud, L., 1987. Fire and survival traits of plants. In: L.
Trabaud (Editor). The role of fire in ecological systems. SPB Academic
Publishing, The Hague, pp. 67-89.
Worbes M. 1995. How to measure growth dynamics in tropical trees.
A review. IAWA Journal, 16 (4):337-351.
Worbes M. 2002. One hundred years of tree-ring research in the
tropics-a brief history and an outlook to future challenges. Dendrochronologia,
20 (1/2):217-231.
Worbes, M., Staschel, R., Roloff, A., Junk W.J. 2003. Tree
rings analysis reveals age structure, dynamics and wood production of a natural
forest stand in Cameroon. Forest Ecology and Management (173):105-123.
Article X. CHAPITRE VI
Article XI. Dynamique de l'occupation du sol et
séquestration du carbone
Ce chapitre centré sur l'approche spatiale, permet de
caractériser la dynamique de la végétation. Il fait
d'abord le point sur les avantages et limites de l'analyse spatiale à
partir des données satellitaires. Ceci nous a alors amené
à considérer deux échelles (locale et nationale) utilisant
des images LANDSAT (1990-2000) et MODIS (2000-2007). Les échelles
temporelles et spatiales prises en compte montrent une forte variabilité
de l'occupation du sol et de sa dynamique. A cet égard, les aires
protégées présentent des caractéristiques
différentes de celles des terroirs où le rôle de
l'agriculture est prépondérant dans la dynamique de la biomasse
et donc du carbone.
Section 11.01 VI.1. Changements d'occupation du sol et
implications sur la dynamique du carbone : l'approche cartographique
La dégradation des terres, plus particulièrement
le déboisement conduit sans équivoque à une
réduction significative des stocks de carbone. La réduction de la
productivité des terres conduit à la diminution du potentiel de
séquestration de carbone par la végétation naturelle. Les
conséquences environnementales de la déforestation sont
très profondes. Elles incluent les modifications spatiales et
temporelles des terres de culture, le déboisement, l'expansion des
pâturages, etc. Ces modifications sont le résultat des effets
climatiques et humains. Le facteur humain est très complexe ; il est
lié à la rareté des ressources qui entraîne une
pression sur ce qui reste, aux dynamiques de marché qui changent les
opportunités, aux interventions politiques qui parfois sont peu
appropriées, la réduction des capacités d'adaptation des
populations due à une forte vulnérabilité
socio-économique, et les mutations sociales et organisationnelles. Les
modifications des services et biens tirés des écosystèmes
influent à leur tour sur les facteurs de dynamiques de l'occupation du
sol. Ainsi certaines conséquences deviennent des causes de
dégradation (Lambin et al., 2001; Lambin et al., 2003;
Verstraete et al., 2008; Verstraete et al., 2009). La
question des changements d'occupation des sols est transversale à
plusieurs aspects du développement rural, basé essentiellement
sur les ressources naturelles. La croissance économique et les
conditions de vie des populations sont très dépendantes des
ressources naturelles.
L'interdépendance entre l'utilisation des ressources
naturelles et le développement durable n'est nulle part plus
évidente que dans les pays en voie de développement où la
survie de la grande majorité de la population dépend de
l'agriculture, des ressources forestières et des ressources animales
(Abdelgalil et Cohen., 2001). La diversité des pratiques et la
complexité sociale des besoins locaux rend difficile l'articulation
politique à toutes les situations (Chokor et Odemerho., 1994).
Actuellement, la relation étroite entre les changements d'occupation des
sols et ses conséquences économiques et sociales est largement
étudiée (Grepperud, 1996; Jones, 1996 ; Barbier, 2000 ; Sankhayan
et Hofstad, 2001 ; Muchena et al., 2004). Cependant, un
intérêt grandissant est noté sur le rôle de la
dégradation des terres et les changements climatiques.
Dans le cadre des changements climatiques, les changements
d'occupation des terres sont considérés comme une source
importante de dégagement de CO2 au niveau des pays tropicaux. Les
défrichements agricoles et l'exploitation forestière
éliminent un grand nombre d'arbre dont la biomasse libère
beaucoup de carbone en se décomposant. En plus, la dégradation
des formations végétales réduit le potentiel de
séquestration de carbone. Pour
ces deux raisons, il est important d'évaluer la dynamique
spatiale et temporelle des ressources forestières pour en
connaître les implications sur le bilan terrestre du carbone.
L'étude des changements d'occupation des sols et
d'utilisation des terres est un exercice relativement difficile à mener
directement sur le terrain, pour des raisons méthodologiques et des
implications financières coûteuses. La
télédétection permet d'acquérir un grand nombre
d'informations sur de vastes surfaces et de façon continue en utilisant
une variété de capteurs satellitaires et une diversité
d'approches (Benz et al., 2004).
Les approches méthodologiques, fortement liées
aux types de données, connaissent aujourd'hui une évolution qui
reflète la diversité et la finesse de la résolution des
images utilisées (photographies aériennes et images
satellitaires) ; ce qui contribue à la réalisation de nombreux
projets cartographiques à des échelles variées. En effet,
les outils de télédétection et de Systèmes
d'Information Géographique (SIG) ont donné une impulsion
remarquable au dessein de la représentation et de l'analyse des
attributs spatiaux plus connus sous l'appellation `occupation du sol'.
Les outils et les méthodes mises en ~uvre -
facilités par l'intégration informatique - tentent d'apporter une
réponse à la problématique de cartographie
harmonisée de la végétation qui réside dans la
conciliation de trois aspects : l'étendue de la zone couverte, le niveau
de détail souhaité et la désignation des classes
identifiées. Ces paramètres sont inhérents aux objectifs
de la recherche, à la finalité manifestée par
l'utilisateur ou l'aménagiste ainsi qu'aux types de données
utilisées.
L'aménagement de l'espace est basé sur deux
concepts clés que sont le temps et l'espace. La
télédétection prend en compte ces deux aspects. Ces deux
concepts déterminent les notions d'échelle spatiale et
temporelle. L'échelle spatiale traduit à la fois l'espace couvert
et la finesse de l'information acquise, alors que l'échelle temporelle
renvoie à la régularité d'acquisition de cette
information. L'analyse des changements de l'occupation des sols est
fondamentalement basée sur cette relation temps-espace.
Les résultats de l'analyse spatiale essentiellement
quantitatifs, sont utilisés pour répondre à plusieurs
objectifs de planification, d'aménagement et de gestion des ressources
naturelles. Les analyses structurales et temporelles menées par
télédétection permettent de décrire les changements
de l'occupation des sols. Alors que l'utilisation des terres fait
référence aux interactions entre les communautés humaines
et leur milieu (Prenzel, 2003). En d'autres termes, l'occupation des sols
traduit la structure spatiale de l'espace géographique et l'utilisation
des terres fait référence à la fonction de cet espace.
Ainsi, une unité d'occupation
des sols peut avoir donc différents types
d'utilisations et plusieurs types d'occupation des sols peuvent être
utilisés pour le même dessein socio-économique. Dans cette
partie du mémoire nous travaillons essentiellement sur l'occupation du
sol en utilisant des techniques de traitement d'images satellitaires.
(a) VI.1.1.Cartographie des changements
d'occupation des sols
La végétation terrestre est souvent le premier
objet de surface rencontré par l'énergie solaire. Ainsi pour la
plupart des types de surface, les images satellitaires, les photos
aériennes ou les informations obtenues par vidéographie,
permettent d'enregistrer les caractéristiques de la
végétation. La télédétection a
été ainsi exploitée pour étudier plusieurs
thèmes liés à la végétation : les
plantations, l'infestation de parasites au niveau des zones de culture,
l'estimation de la production des formations boisées, la cartographie
des types de végétation, etc., (Cambell, 1996).
La composante végétale est très complexe
et change dans le temps ; ses propriétés spectrales
étudiées à différentes échelles (feuilles,
plantes ou couvert végétal), variant fortement selon le milieu,
la saison et les phases de croissance. Ces fortes variations des
propriétés de la végétation expliquent la
complexité des procédures d'analyse des données
satellitaires pour la caractérisation des différents paysages
à la surface terrestre (Flasse et Verstraete, 1993). Cette
difficulté rend évident le besoin d'une convergence dans les
démarches cartographiques et la désignation des classes de
végétation.
L'essentiel des défis de la cartographie tourne autour du
concept `d'harmonisation' des méthodes. Les principaux
points qui requièrent cet effort de convergence sont les suivants :
- l'harmonisation des procédures de classification,
- l'harmonisation des légendes (nomenclature),
- l'harmonisation des méthodes de collecte de
données sur le terrain,
- l'harmonisation des types de projection et des
paramètres de cartographie.
Ces exigences sont utiles pour arriver à une
comparabilité des cartes réalisées dans les
différents pays africains.
L'utilisation de la télédétection et des
données géospatiales pour la cartographie de
la
végétation et l'occupation des terres est devenue une
activité courante des institutions
intéressées par la
gestion des ressources végétales (Achard et al., 1996 ;
Achard et al., 1996).
L'imagerie satellitaire joue un rôle incontournable dans
le processus de caractérisation et d'aménagement des
écosystèmes. L'acquisition et l'analyse des données
spatiales requièrent cependant un minimum d'investissement temporel et
financier lié à l'acquisition et au traitement des images et des
données de terrain. Le cartographe est souvent confronté à
deux exigences : le choix des images adaptées au type de cartographie et
la conception de la démarche liée à la nature des
données.
Le spectre optique (Visible et PIR : 400-3000 nm) est le plus
utilisé en télédétection pour les études de
l'occupation du sol. Les processus d'absorption, de réflexion, de
diffusion et d'émission qui se passent dans cet intervalle du spectre
électromagnétique ont servi à la caractérisation de
la surface terrestre. Les quantités spectrales mesurées pour un
objet donné dépendent des propriétés de cet objet.
Par conséquent, les propriétés d'un objet de surface
peuvent être inférées à partir de ses
propriétés spectrales.
Prenzel (2003) suggère une technique d'inversion simple
pour passer des mesures de réflectance aux propriétés des
objets de surface en utilisant les fonctions suivantes:
radiation mesurée = f(propriété de
l'objet) où propriété de l'objet = f'(radiation
mesurée).
L'analyse des images satellitaires doit prendre en compte le
niveau de l'information à travers lequel les données sont
présentées pour lier les quantités spectrales avec une
réalité thématique. Les différents niveaux de
traitement des données permettent de simplifier, de
généraliser la réalité complexe pour que
l'interprétation soit plus simple. La figure 81, illustre le lien entre
les différents ordres de traitement pour arriver à un
résultat proche de la variable thématique en question
(l'occupation du sol).
Figure 81. Etapes de transformation des
données images pour la cartographie de l'occupation du sol.
Données brutes
Réflectance
Occupation du sol
NDVI, TS
LAI, Biomasse
+ d'informations thématiques
Un autre aspect de la complexité de la
télédétection réside dans le cheminement qu'on
peut
adopter pour arriver à des résultats. On peut noter deux
principales approches : l'approche
déterministe et celle empirique.
La première permet de faire des analyses quantitatives
précises alors que la seconde basée sur des lois
de probabilité permet d'approcher la réalité avec un
certain niveau de confiance. Les approches déterministes
requièrent plus de validation alors que les modèles empiriques
sont simples à développer et demandent moins d'effort de
validation. L'utilisateur doit ainsi trouver un compromis entre ces deux
extremes pour l'étude des changements de l'occupation du sol. Les
analyses déterministes comprennent : l'estimation de la chlorophylle,
l'indice foliaire, le contenu en nitrates et la productivité primaire
nette (quantités biophysiques). Les analyses empiriques couvrent : la
classification des groupes spectraux, les réseaux de neurones, la
segmentation, etc. Le résultat de ces modes de traitement est nominal.
On peut développer des méthodes mixtes qui essaient de tirer les
avantages entre les deux approches à travers des corrélations
linéaires ou non. La méthode adoptée dans le cadre de ce
travail est une approche empirique basée sur la classification des
images satellitaires.
(b) VI.1.2. Considérations pratiques pour la
cartographie de la végétation
La cartographie thématique est un exercice
délicat puisqu'il nécessite un choix judicieux des données
de base. Toute image est utile pour un type de cartographie
particulière. En fonction de la finalité du travail et de
l'étendue de la zone, le cartographe doit opérer un choix
judicieux des images. Quatre principales étapes sont importantes
à considérer :
- Identification des besoins (cartographie à grande ou
à petite échelle) ;
- Recherche des données adaptées au type de
cartographie (le type d'image détermine les possibilités de faire
une carte détaillée ou non) ;
- Identification de la meilleure période pour
l'acquisition des images (dans les zones de savane la fin de la saison des
pluies avant le début des feux de brousse, est souvent indiquée
pour cette acquisition) ;
- Elaboration de la méthode de cartographie adaptée
aux objectifs (celle-ci est relative à l'objectif du projet).
Le choix d'un type d'image doit être un compromis entre
le niveau de détail souhaité, l'étendue de la zone
d'étude et le coOt des données. Le tableau 21 donne quelques
détails sur le potentiel de certaines sources de données pour la
cartographie de l'occupation du sol.
Tableau 21. Présentation de quelques
capteurs utilisés pour la cartographie de la
végétation.
|
Capteurs
|
Taille du pixel
|
Echelle de cartographie de la
végétation
|
Méthodes utilisées (non
exhaustives)
|
|
Photographie aérienne
|
Dépend de l'échelle (1 m-15 m)
|
Plan / Régionale (1/10.000 - 1/250.000)
|
Interprétation visuelle, vectorisation
|
|
IKONOS, QuickBird, GeoEye
|
<1 m
|
Locale (1/10.000 - 1/25.000)
|
Classification, Numérisation directe Décompte
d'arbres, taux
de couverture
|
|
SPOT-HRV, CEBERS
|
20 m (XS), 10 m (P)
|
Locale (1/25.000 - 1/50.000)
|
Classification, ACP, Numérisation directe,
|
|
SPOT-5
|
2,5 m
|
Locale (1/10.000 - 1/25.000)
|
Classification, Numérisation directe
|
|
LANDSAT- TM-ETM
|
30 m (XS), 15 m (P)
|
Locale (1/50.000 - 1/200.000)
|
Classification, ACP, Numérisation directe,
|
|
MODIS (Terra/Aqua)
|
500 m ®-250 m (PIR)
|
Locale / régionale (1/250.000 - 1/500.000)
|
Indices de végétation, Classification,
|
|
MERIS
|
250 m
|
Locale / régionale (1/250.00 - 1/500.000)
|
Indices de végétation, Classification,
|
|
SPOT- VEGETATION
|
1km
|
Régionale et globale (à partir de 1/500.000)
|
Classification
|
|
NOAA-AVHRR
|
1 km
|
Régionale et globale (à partir de 1/500.000)
|
Indices de végétation, Classification, Ts
|
|
SPOT VEGETATION
|
1 km
|
Régionale et globale (à partir de 1/500.000)
|
Indices de végétation, Classification
|
|
ERS (RADAR)
|
25 m-30m
|
Locale et régionale (1/100.000 - 1/200.000)
|
Ratio, texture
|
Du fait de la diversité des données et des
résultats cartographiques, le Projet AFRICOVER (initié par la
FAO) a entamé un processus d'harmonisation des procédures de
classification des formations végétales de l'Afrique tropicale.
Cette initiative lancée en 1994 à Addis Abéba, a pour
tâche de proposer une classification standardisée qui pourrait
servir à la cartographie de l'occupation du sol en Afrique (AFRICOVER,
1997). Deux parmi les objectifs majeurs du projet nous semblent
particulièrement intéressants pour cette analyse, il s'agit :
- du développement de modèles de classification
à partir d'images satellites et de photographies aériennes ;
- de la conformité des types de classification aux
légendes actuellement en vigueur en Afrique.
Ce dernier point pose le problème de la `normalisation'
terminologique des types de végétation en rapport avec les
légendes existantes. Par ailleurs, dans la démarche
méthodologique proposée par AFRICOVER (1997), à travers le
`Land Cover Classification System' ou LCCS', peu de précisions
sont données sur les critères de seuillage des taux de couverture
pour séparer les formations dites fermées, ouvertes et
clairsemées. Il en de même sur l'association entre familles
végétales et types biologiques (situant au même niveau
arbres et arbustes, Graminae et Euphorbiacea). L'autre
difficulté réside sur l'utilisation du critère de la
taille des feuilles comme élément discriminant surtout dans les
milieux de savane caractérisées par une relative
hétérogénéité foliaire. Les principes de
classification de la couverture végétale ont été
développés au milieu des années 1980 par la FAO. De facto,
l'établissement de la nomenclature de couverture de terre de
GlobCover-2005 a suivi les principes du système FAO-LCCS (le
système de classification de la couverture de terre, qui est un ensemble
de règles et un logiciel pour créer une nomenclature). Cette
approche prête plus d'attention à la densité et la
physionomie de la végétation. Ces approches ont été
encouragées pour pallier les limites des décisions prises en 1954
à Yangambi (Congo) par des botanistes de l'Afrique tropicale (voir
Sambou, 2004).
Les approches sont très nombreuses, mais quelque soit
la technique ou la méthode utilisée, la cartographie de la
végétation est largement dépendante des types de
données source et des objectifs fixés. Par conséquent, le
type de classification doit répondre dans une large mesure aux
caractéristiques des images utilisées et au niveau de
précision souhaité. Un choix judicieux des images devient alors
un préalable dans tout processus de représentation
cartographique.
Article XII.
Le choix des données et le traitement des
images
Une bonne utilisation de la télédétection
pour la cartographie des changements de l'occupation des sols et de
l'utilisation des terres suppose un choix judicieux des données et des
méthodes (Rogan et Chen., 2004). Les contraintes liées
à l'étude des changements de surface sont de deux ordres : les
limites techniques et les contraintes environnementales.
- Les contraintes techniques
Les données satellitaires sont décrites à
travers leurs résolutions spatiales, spectrales, temporelles et
radiométriques. Les contraintes technologiques apparaissent dans la
façon dont ces résolutions se traduisent sur les données.
D'une façon générale, la résolution spatiale est
inversement corrélée à la couverture spatiale. Cette
contrainte technique limite la
portée des études sur la dynamique de l'espace.
De la même manière, une faible couverture spatiale se traduit par
une répétitivité moins importante (Rogan et
Chen., 2004). Il existe aussi des contraintes d'ordre techniques
liées à des facteurs externes difficiles à
contrôler. Ces contraintes sont selon Collins et Woodcock (1994; 1996) :
i) les variations des conditions atmosphériques, ii) les
différences d'illumination, iii) les différences de
calibration.
- Les contraintes méthodologiques
Les algorithmes de traitement des images sont essentiellement
basés sur les valeurs individuelles des pixels et ne prennent que
rarement en compte l'information contextuelle des segments spectraux qui
représentent les objets de l'image (Benz et al., 2004).
L'analyse du signal est très dépendante de l'incertitude
associée aux bruits systématiques de l'image. Aussi, les
différentes étapes du traitement des images introduisent des
artefacts et des ambiguïtés qui peuvent aggraver
l'imprécision de la donnée finale. Il faut signaler par ailleurs
que les objets géographiques n'ont pas de limites linéaires
nettes. Les analyses actuelles (exception faite de la théorie des
ensembles flous) ne prennent pas en compte les classes composites, les zones de
transition, les mosaïques qui sont plus difficiles à segmenter.
Dans certaines conditions de savane, il faut une bonne dose de
généralisation pour rendre les cartes lisibles.
- Les contraintes environnementales
L'occupation du sol est un facteur très dynamique dans
le temps. L'évolution des types de couverture du sol incluent : les
perturbations (nuages, feux de brousse, maladies, exploitation de certaines
ressources naturelles), les cycles naturelles des écosystèmes
(succession des saisons, variabilité interannuelle). Pour suivre ces
changements, le choix de la période d'acquisition des images est aussi
crucial que le choix des capteurs et les algorithmes de détections
utilisés. Le choix de la période d'acquisition des images
satellitaires doit être à peu près la même chose pour
les différentes dates d'analyse. Pour déterminer cette
période, il faut une relative bonne connaissance des
caractéristiques environnementales de la zone d'étude.
- Prétraitement des images pour la
détection des changements
Le prétraitement des images satellitaires a pour but de
minimiser les erreurs liées à l'acquisition des données
images, les bruits systématiques associés au capteur, la
correction géométrique des images et le masquage des fragments de
la scène contaminés par une couverture nuageuse. Ces
différents aspects du prétraitement sont regroupés en deux
catégories : la correction géométrique et la calibration
radiométrique. Une bonne rectification géométrique est
essentielle pour l'analyse des changements d'occupation du sol ; toute erreur
majeure dans la correction géométrique peut entraîner des
biais importants
dans l'analyse, surtout au niveau des zones de contact de
différents types d'occupation du sol. Aussi, les caractéristiques
radiométriques des images de différentes dates doivent être
intercalibrées pour pouvoir comparer les grandeurs spectrales qui
garantissent que les résultats de changements obtenus traduisent une
réalité thématique factuelle sur le terrain plutôt
que des artefacts liés aux capteurs, à l'état de
l'atmosphère ou à la topographie locale. Il faut par
conséquent une compatibilité radiométrique et
géométrique entre les images. Le modèle
générale d'analyse des images satellitaires pour extraire les
catégories de l'occupation des sols est schématisé dans la
figure 82.
Figure 82. Approche empirique d'extraction des
catégories nominales d'occupation et d'utilisation des terres. Source :
Prenzel, (2003).
Prétraitement :
corrections géométriques, et calibration
radiométrique
CN, traités
année Y

CN, année X
CN, année Y
Validation avec
des
données
indépendantes



Classification des
changements
Classification thématique
des
changements
Intégration dans un système
de
planification

SIG
CN, traités
année X
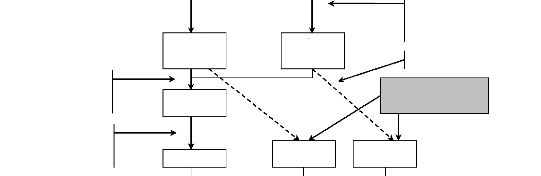
Identification de changement
:
différence, rapport, analyse de
vecteur de
changement
Analyse booléenne : Masquage des zones
sans changements
CN améliorés
de X - Y
Amélioration
de contraste
année X
Changement
Classification
année X
Amélioration
de contraste
année Y
Classification
année Y
Calibration et validation
avec des
données
indépendantes
Améliorations spectrales :
Amélioration de contraste
Ratio de bandes, Filtres spectraux, ACP, Indices spectraux
Classification thématique
- Détection de changements d'occupation du
sol
Le besoin de supports d'interprétation simplifiés
de la distribution des types de végétation a
conduit à
de nombreuses approches méthodologiques utilisant des outils et
techniques
variés. A cet égard, l'étude des
caractères spatio-temporels des formations végétales par
le
biais de la représentation cartographique, a permis
quelques progrès dans la gestion des ressources naturelles. Plusieurs
approches ont été utilisées pour la détection de
changement de l'occupation du sol. Les plus fréquemment citées
sont les suivantes.
- La technique de soustraction d'images. Les images
de changements sont faciles à interpréter puisque leurs
histogrammes présentent une distribution normale. Les pixels qui n'ont
pas changé sont au milieu de l'histogramme alors que les pixels qui ont
muté sont soit à droite soit à gauche du centre en
fonction de leur brillance (Rogan et Chen, 2004).
- Les analyses en composante principales. C'est une
transformation orthogonale basée sur une matrice de corrélation
(standardisée) ou de variance covariance (non standardisée). Les
travaux basés sur cette méthode ont montré que le choix du
type de matrice peut influencer les résultats statistiques des
composantes principales. Les ACP sont uniquement basées sur les
propriétés statistiques des images, ce qui rend difficile leur
utilisation pour des périodes d'acquisition différentes.
- La Transformation multitemporelle des indices
Tasseled-Cap (Kauth et Thomas). C'est aussi une transformation orthogonale
multibandes proposée par Collin et Woodcock (1994; 1996) pour
générer différentes images qui présentent une
stabilité ou non pour le brightness (brillance), le
greeness (verdure) et le wetness (humidité). Cette
approche a l'avantage de ne pas dépendre des caractéristiques
statistiques des scènes de la série temporelle
utilisée.
- L'analyse des vecteurs de changements (AVC). Cette
méthode permet de générer deux types d'informations :
l'importance et la direction du changement. La classification d'image qui suit
le calcul des vecteurs permet de spatialiser les zones qui ont connu une
dynamique spatiale. Cette méthode est cependant limitée par le
masquage de la variance statistique des `classes de changements' par les
`classes stables'.
- La classification d'image, segmentation d'image et les
systèmes intelligents. Cette méthode concerne les
procédures utilisées avant et après la classification
d'image. Pour toute classification supervisée ou non supervisée,
des données de validation doivent avoir été
collectées pour une bonne validation des résultats. Ces
méthodes de classifications sont les plus utilisées pour la
cartographie des changements de surface. Les méthodes basées sur
les arbres de décision ou les réseaux de neurones ont
montré d'énormes avancées par rapport aux méthodes
classiques puisque ces méthodes ne sont pas tributaires des
hypothèses statistiques paramétriques. Elles sont ainsi
adaptées à l'analyse de données multimodales,
bruitées ou lacunaires.
- L'Analyse texturale consiste à regrouper les
unités de paysage selon leur homogénéité spatiale.
Dans les formations de savane caractérisées par la variation des
densités d'arbres et de taux de couverture du sol, cette approche
pourrait être très utile.
- La théorie des ensembles flous. Les
classifications basées sur cette théorie sont souvent
appliquées au niveau d'espaces géographiques dont le passage
d'une unité à l'autre fait apparaître une forme de
transition qui ne permet pas de définir une frontière nette.
- Le Multitemporal spectral mixture analysis. Cette
technique est plutôt utilisée quand il s'agit de séries
temporelles souvent de basse résolution (NOAA-GIMMS, Pathfinder).
- Les Déductions statistiques. Cette approche
est relativement simple et consiste à comparer les superficies des types
de couverture du sol de deux cartes d'occupation du sol.
Section 12.01 VI.2. Les données et la
méthode utilisées
L'approche ici mise en ~uvre s'appuie sur les connaissances
tirées des applications de la télédétection
à l'étude de la végétation. Sur cette base et
à partir des traitements des données LANDSAT et du contrôle
de terrain, nous proposons une procédure de cartographie de la
végétation à l'échelle locale dans les
écosystèmes de savane.
Le choix des données LANDSAT dans le cadre de ce travail
s'explique alors et dépend des quatre exigences ci-dessous :
- la nécessité d'avoir des informations les plus
récentes possibles ;
- la possibilité de couvrir de façon
instantanée de vastes superficies ;
- l'exigence d'avoir des supports images permettant une
cartographie détaillée ;
- et, la possibilité de répéter les prises
de vue dans une perspective de suivi de la dynamique spatiale de la
végétation.
Les données images de niveau 1G ont été
reçues sous un format GeoTIFF (Geographic Tagged Image File
Format). Ce format est caractérisé par l'association
d'informations géodésiques et cartographiques qui ont fait
qu'aucune rectification géométrique n'a été
nécessaire. Chacune des bandes est délivrée en niveaux de
gris non compressés avec une dynamique de 8 bits. Le tableau 22 donne
des indications sur les données acquises.
Section 12.02
Tableau 22. Les scènes images
utilisées
|
Forêts
|
Path-row
|
Dates
|
|
Année 1
|
Année 2
|
|
Bala
|
203-50
|
1989
|
2000
|
|
Kantora
|
203-51
|
1989
|
2000
|
|
Mampaye
|
203-51
|
1989
|
2000
|
|
Ouli
|
203-50
|
1990
|
2000
|
|
Patako
|
205-50
|
1989
|
1999
|
|
Wélor
|
205-50
|
1989
|
1999
|
|
Zone Kaffrine (terroir agricole)
|
204-50
|
1989
|
1999
|
Les conditions d'acquisition n'ont pas toujours permis d'avoir
des intervalles chronologiques de dix ans avec des images de qualité.
Ainsi la cartographie de l'occupation des sols pour les deux acquisitions
souhaitées n'a pas pu se faire pour Wélor et Kantora. Les images
ont été acquises entre novembre et décembre de chaque
année. La considération de la zone de terroir est basée
sur l'hypothèse selon laquelle, les tendances de l'occupation du sol en
zones non protégées est plus sévère que celles des
zones classée. En outre la dynamique de la
végétation en zone de terroir est un facteur
important dans l'évaluation de la vulnérabilité des
populations aux modifications de l'environnement.
Section 12.03
(a) VI.2.1. Classification des images
Nous avons procédé à l'extraction des
fenêtres spatiales correspondant aux Forêts Classées
étudiées. Une composition colorée 7-4-2 rehaussée a
permis par analyse visuelle de décider du nombre de classes
représentatives sur chaque image. Pour harmoniser la procédure,
une classification non supervisée (ISODATA) a été faite
sur chaque fenêtre d'image.
La définition de critères de classifications
permet d'éviter des divergences liées à la
subjectivité (arbitrage) et au mode de traitement automatique. La
classification réalisée part d'un certain nombre de principes que
sont :
- L'exhaustivité qui renvoie à la
nécessité d'individualiser toutes les classes qui apparaissent
sur les images ;
- L'exclusivité mutuelle qui permet
d'éviter qu'un type de végétation ne soit
représenté dans différentes classes de la zone
d'étude ;
- La flexibilité qui permet de pouvoir modifier
la nomenclature ou faire des fusions de
classe si les données annexes et le terrain
présentent des réalités différentes ;
- L'objectivité qui permet de se concentrer sur
les caractéristiques de la végétation et éviter
l'interprétation subjective et une typologie relative ;
- La représentativité, pour que les
classes identifiées soient des entités spatiales
significatives sur le terrain ;
La classification non supervisée par ISODATA a
été utilisée pour éviter les divergences
liées aux besoins de cohérence (conformité) dans le choix
de zones d'entraînement pour deux dates successives. ISODATA utilise un
algorithme qui effectue d'abord un calcul des moyennes de classes et ensuite de
façon itérative, les pixels de l'image sont rangés dans
les classes en utilisant le critère de la distance (spectrale) minimale.
Toutes les bandes du visible, moyen infrarouge et proche infrarouge ont
été utilisées pour la classification. Le nombre
d'itérations est de `n' bandes + 1, ce qui fait 7 itérations. Ce
nombre d'itérations (fixé de façon empirique) permet une
bonne convergence du résultat de traitement. Les itérations
permettent de recalculer les moyennes et d'affiner la classification des pixels
en fonction des nouvelles moyennes calculées. Des seuillages sur la
valeur des pixels ont permis de mieux séparer les classes et d'en
éliminer ceux ne correspondant pas à une réalité
thématique
précise. Ainsi, les pixels sont tous rangés dans
les classes qui ont les moyennes spectrales les plus proches. Le processus est
répété jusqu'à ce que tous les pixels soient
rangés dans des classes et ceci au bout des 7 itérations.
Les résultats de la classification sont soumis à
un filtre majoritaire pour simplifier la cartographie. Le filtre majoritaire
permet de supprimer les pixels isolés dans les grandes classes. La
fenêtre mobile (kernel) utilisée est 7x7 pixels et la classe
dominante dans cette fenêtre est appliquée à tous les
pixels qui s'y trouvent. Ainsi, la classe dominante est `nettoyée' des
pixels isolés qui altèrent l'homogénéité de
la classe.
L'étape suivante est une superposition du
résultat de la classification avec la composition colorée et une
interprétation visuelle interactive permet de donner une signification
thématique à chaque classe. L'outil de vectorisation automatique
a permis de convertir la classification en format vectoriel.
Le résultat est transféré dans un SIG qui a
permis de regrouper les classes qui ont la même signification
thématique et de calculer les surfaces de chaque classe.
L'interprétation des images est basée sur une
connaissance du terrain et des caractéristiques spectrales des objets
dans les différentes bandes de longueur d'onde. Les données de
terrain constituent une référence importante dans
l'interprétation et la classification des images. Les observations de
terrain ont permis de vérifier le lien entre les classes de
végétation au sol et les classes spectrales sur les images. La
vérification sur le terrain a été faite en fin de saison
des pluies, ce qui correspond aux dates d'acquisition des scènes.
(b) VI.2.2. Résultats du traitement des
images Article XIII.
Article XIV. Dans l'intervalle d'une décennie,
l'analyse des cartes d'occupation du sol des Forêts Classées
étudiées indique plusieurs cas de figure. Les figures 83-87
fournissent le détail des thèmes classifiés, avec une
bonne acquisition pour Mampaye, Ouli, Bala et Patako et des perturbations
liées aux feux de brousse pour Kantora et Wélor sur les
données de 1988. Ceci explique la médiocre qualité de ces
dernières images pour cette application et par conséquent
l'impasse sur leur classification et interprétation.
Figure 83. Carte de végétation de la
Forêt classée de Mampaye
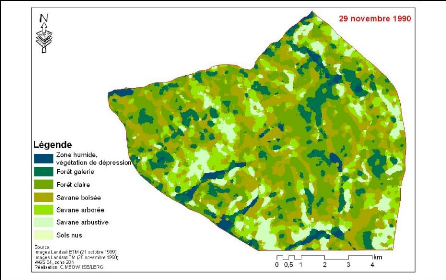
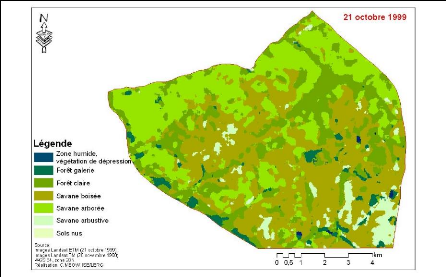
Figure 84. Carte de végétation de
la Forêt Classée de Bala
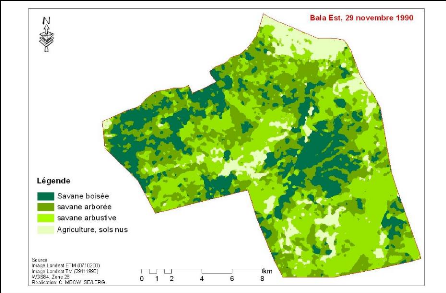
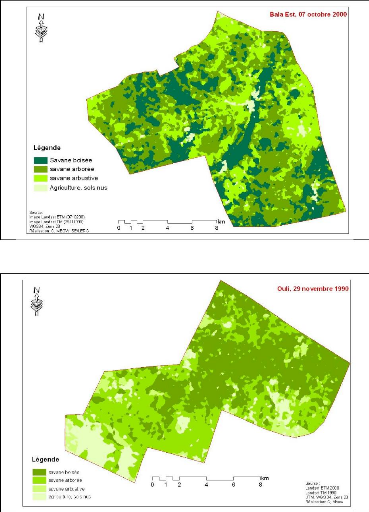
Figure 85. Carte de végétation de
la Forêt classée de Ouli
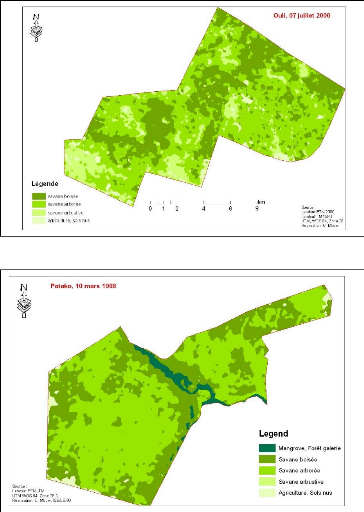
Figure 86. Carte de végétation de
la Forêt classée de Patako
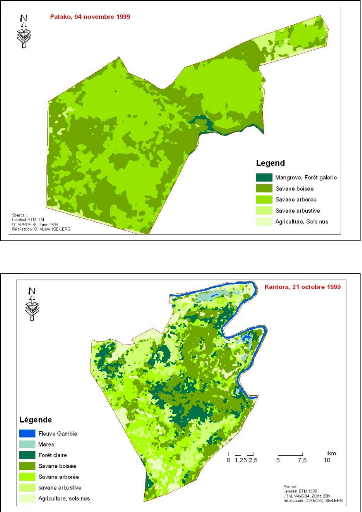
Figure 87. Carte de végétation de
la Forêt classées de Kantora et Wélor (Une acquisition)
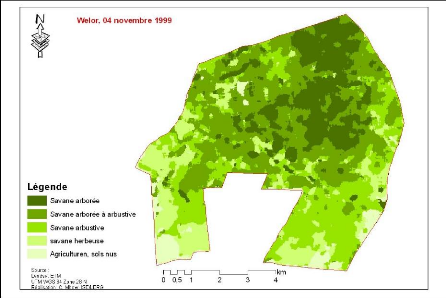
Les cartes produites à intervalle de dix ans indiquent
une absence de dynamique linéaire des changements d'occupation du sol.
Cependant, il existe des modifications persistantes des formations denses comme
les formations de vallées qui se transforment de plus en plus en des
formations ouvertes (cas de la Forêt Classée de Mampaye et de
Patako). Des savanes arborées ou boisées semblent se reconstituer
par endroits dans la Forêt classée de Bala. Une forte
fragmentation de la végétation est notée sur le site de
Ouli et dans une moindre mesure à Bala. Ces différents cas de
figures remettent en question des théories de dégradation
absolues, et parfois confortent les thèses selon lesquelles, la
dégradation de la végétation est un mythe qu'il faut
prendre parfois de façon relative (Bassett et Zuéli, 2000;
Rasmussen et al., 2001 ; Rasmussen et al., 2006). Des
études récentes basées sur des séries temporelles
de données NOAA-GIMMS/PATHFINDER ou même les composites NDVI de
MODIS soutiennent que le Sahel est entrain de reverdir. Il faut
considérer à ce niveau que les analyses sont à petite
échelle et les intervalles d'acquisition de données partent le
plus souvent de périodes sèches vers des périodes humides.
Ces facteurs peuvent influencer le résultat, notamment la pente de la
tendance observée. Il est difficile à l'état actuel des
connaissances de trancher cette question. Il faut par conséquent prendre
chaque situation à part et analyser les tendances en fonction du milieu
et des interventions en cours.
Au chapitre 3, les données in situ des
parcelles de suivi de la végétation ligneuse
introduisent
l'importance du facteur pluviométrique qui peut
radicalement modifier un couvert végétal au
bout de quelques
années. La nature du substrat et les facteurs humains affectent aussi
l'état
et la dynamique du couvert végétal. Sur les
sites étudiés, il est très difficile de discriminer les
facteurs naturels des facteurs humains (résultats des simulations). Il
faut rappeler qu'on a plusieurs agressions sur les formations
forestières que sont : les feux de brousse, les coupes, les
défrichements agricoles, etc.
Toutefois, l'implication des changements d'occupation du sol
sur le bilan du carbone dépendra à la fois des modifications de
la densité des peuplements mais aussi de la conservation des jeunes
plantes en croissance qui ont un grand potentiel de fixation de CO2. Si par
exemple des formations végétales matures sont exploitées,
il faudra mettre en place des mécanismes de reconstitution des milieux
dégradés pour compenser et même inverser le bilan de
carbone. Certaines dynamiques positives observées à Bala par
exemple peuvent êtres liées à une réduction de
l'exploitation forestière en un moment donnée. Inversement, la
persistance de la dégradation à Ouli s'explique en grande partie
par une forte pression sur les ressources forestières, du fait des
activités de carbonisation. Les variations de superficies de la figure
88, montrent que pour Mampaye et Ouli, on a en générale une
tendance vers des formations plus ouvertes, alors que pour Patako et Bala, les
variations sont plus erratiques. Ces observations peuvent largement
contrastées avec la réalité des zones de terroirs qui
montrent une forte dynamique de dégradation des formations
végétales naturelles.
Figure 88. Changement d'occupation du sol et
dynamique du carbone dans les Forêts Classées de Mampaye, Ouli,
Balla et Patako.
Pour avoir une meilleure perception des changements
d'occupation du sol dans la zone d'étude, nous avons choisi la zone de
Kaffrine (Saloum Oriental) où on a une superposition de plusieurs
activités humaines : agriculture, élevage, activités
sylvicoles, etc. le fait que la zone soit considérée pendant
longtemps comme un creuset de la production arachidière et un espace
privilégié d'exploitation agricole, permet d'entrevoir et de
mieux comprendre ce que
12000
cette activités peut générer comme
modifications au plan de la dynamique spatiale, (figure
10000
S
NI,
89).80
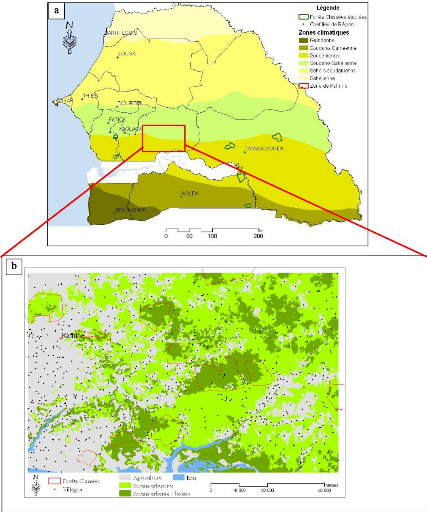
Année:1989
Figure 89. Carte d'occupation des sols dans le
Saloum Oriental (Kaffrine : a) localisation ; b) carte de
végétation).
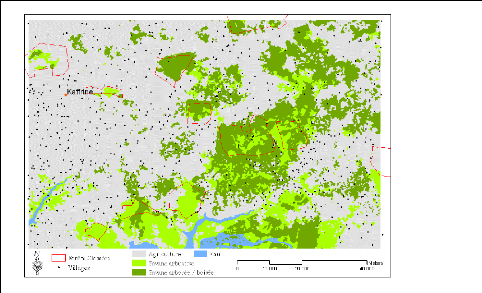
Année:1999
Les terres agricoles ou dénudées de cette zone
ont augmenté de 58 % en 13 ans (de 281 ha, elles sont passées
à 486 ha), au moment où les formations de savane ont
été réduites de 56% (passant de 473 ha à 264 ha
pour la même période). Les estimations de biomasse du chapitre 3
montrent que les savanes arbustives/arborées ont des charges de biomasse
de 35 t C/ha (#177; 5 t) en moyenne. La réduction des savanes de 209 ha
entraîne une perte totale de carbone estimée à 7315 tC soit
562 tC par an sur l'ensemble de la zone cartographiée.
De tels résultats ont sous-tendu la forte conviction
d'une dégradation des terres des écosystèmes semi-arides
et constituent un facteur important de libération du carbone dans
l'atmosphère (IPCC, 2000). Ainsi, les projets MDP visent à
reconstituer ces milieux dégradées qui sont le plus souvent
d'anciennes terres de culture ou des zones de surpâturages.
Dans le Saloum Oriental, la monoculture de l'arachide et les
techniques agricoles utilisées
expliquent en bonne partie le recul de
formations forestières. Des études sur le carbone du
sol montrent déjà de fortes réductions
(Feller, 1977; Elberling et al., 2002; Touré, 2002 ),
quisont une conséquence directe de la dégradation de
la végétation ligneuse et d'un épuisement
progressif des terres. Le chapitre 7 expose les implications
de cette forte dégradation de l'environnement sur la
vulnérabilité des populations de plus en plus exposées aux
fortes variations du climat.
Les études de cas, à des échelles locales
montrent des résultats variables selon les milieux, les conditions
climatiques et humaines. Pour mieux caractériser la dynamique de
l'espace, nous proposons une analyse à l'échelle nationale des
dynamiques d'indices de végétation.
(a) VI.2.3. Suivi du NDVI à partir des
données MODIS
Les indices de végétation
(IV)15 sont les premières sources
d'information pour un suivi opérationnel de la couverture du sol. Ils
correspondent à des mesures radiométriques sur la
variabilité spatiale et temporelle de l'activité
photosynthétique de la végétation, laquelle est
reliée à des variables biophysiques de la couverture du sol et
plus particulièrement de la couverture végétale. De
nombreux indices de végétation ont été
proposés depuis une vingtaine d'années et sont basés,
presque tous, sur les propriétés spectrales de la
végétation verte dans le PIR et le Rouge. Ils reposent
généralement sur la grande différence radiométrique
entre le PIR (760-900 nm), où la réflectance est liée
à la structure cellulaire des feuilles, et le Rouge (630-690 nm)
où la chlorophylle entraîne une forte absorption du rayonnement
incident.
Ces indices permettent de mesurer la vigueur de la
végétation et sont plus sensibles à la détection de
la biomasse que les bandes spectrales prises individuellement. Chaque indice
spectral possède un potentiel discriminatoire qui permet de l'utiliser
dans des situations variées, mais il n'existe pas encore d'indice
universel pouvant s'adapter à tous les types de végétation
et à toutes les conditions écologiques.
Il existe un grand nombre d'indices de
végétation (PVI, SAVI, TSAVI, MSAVI, GEMI, NDVI, etc.), et chacun
essaie d'optimiser la caractérisation des états de surface en
évitant le maximum de biais, comme l'effet du sol, de
l'atmosphère, etc. Les IV ne sont pas des mesures directes de la
biomasse ou de la production primaire, mais sont corrélées
à la couverture foliaire et à la quantité de biomasse.
Pour l'estimation de l'état de la végétation, ils doivent
être capables de différencier les caractéristiques de la
végétation de celles du sol. Ceci n'est possible que si les
réflectances des deux objets de surface sont uniques et si l'indice de
végétation choisi est sensible à ces différences de
réflectance. L'indice de végétation le plus utilisé
à l'heure actuelle est le NDVI, différence normalisé entre
le PIR et R (PIRR/PIR+R).
15 De nouvelles recherches préfèrent la radiation
photosynthétiquement active (PAR) à la place du NDVI
Pour analyser la dynamique de la végétation
à l'échelle nationale, nous avons utilisé les
données MODIS-250m (NDVI). Ce produit a pu être
téléchargé sur le site du Global Land Cover Facility
(
http://www.landcover.org/data/ndvi).
Les bandes 1 et 2 du capteur MODIS (Moderate Resolution Imaging
Spectroradiometer) du satellite Terra de la NASA ont été
utilisées pour produire ces données. Celles-ci sont obtenues sous
forme de composites de 16 jours en format Geotiff. Les 20 composites de 16
jours (entre les jours juliens 49-353) ont été
intégrées pour générer 8 composites annuelles, de
2000 à 2007 (figure 90).
Les variations de la production végétale
semblent à cette échelle très liée aux variations
de la pluviométrie. Les années très pluvieuses de 2005
montrent une importante productivité dans la partie sud du pays. Il
apparaît donc que la variation du NDVI varie de façon zonale en
relation avec l'importance des pluies. Une telle situation montrée par
la figure 91, ne permet pas de mesurer la dynamique temporelle et spatiale de
la végétation en termes de direction et d'ampleur de
changements.
Figure 90. NDVI annuel au Sénégal
de 2000 à 2007.
2006 2007
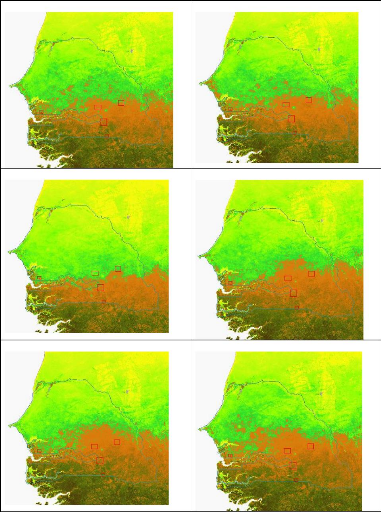
2000 2001
2002 2003
2004 2005
-1 0 + 1
100 km % Sites
étudiés

Nous avons ainsi procéder à une analyse simple,
consistant à considérer les droites de tendance des NDVI des
images composites pendant les 8 années d'acquisition de données.
La logique suivie est de segmenter l'image en fonction de l'importance des
modifications de valeurs de NDVI. Chaque pixel a une valeur de NDVI sur les 8
ans de suivi. Si les valeurs de NDVI diminuent de façon
générale on a une courbe de tendance descendante, et si elles
augmentent on a une courbe de tendance ascendante. Chaque courbe de tendance
est caractérisée par sa pente et son coefficient de
détermination (R2). L'approche développée est
de créer un produit cartographique qui montre les valeurs de
coefficients de détermination (R2) sur l'image. La figure 91
décrit la logique de l'approche.
Figure 91. Représentation logique de
l'approche de la dynamique du NDVI
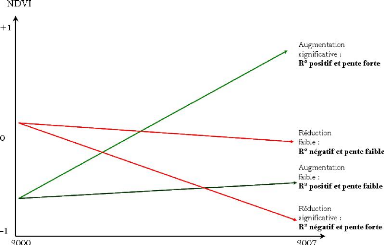
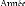
La figure 92 est une spatialisation de ce concept
d'évaluation des changements d'état de surface.
Figure 92. Tendance des valeurs NDVI sur le
Sénégal (2000-2007)

Sites étudiés


100 km %
-1 0 +1
L'interprétation d'un tel produit permet de voir que la
dynamique de la végétation n'est pas aussi linéaire que ne
le montrent les cartes annuelles prises individuellement. On observe en effet
plusieurs zones de fortes réductions de l'activité
chlorophyllienne (zone de Bignona, Ferlo, Bassin arachidier) ; mais on observe
une augmentation significative de la verdure dans la partie nord du
Sénégal Oriental. Ces variations du NDVI sont relatives. Un
regain de l'activité chlorophyllienne peut se traduire par une forte
production de biomasse ou par une modification de la composition
spécifique des formations végétales. Pour les Forêts
Classées étudiées, le cas de Patako montre une
réduction significative des valeurs de NDVI, alors que les autres
Forêts Classées semblent maintenir une bonne activité de
production. La Forêt Classée de Bala laisse apparaître une
forte augmentation sans doute liée à la réduction des
activités d'exploitation forestière dans cette zone.
L'analyse du NDVI montre une perspective plus globale de la
dynamique de la végétation et permet de mieux apprécier de
façon qualitative le potentiel de séquestration de carbone et les
tendances observées pendant une période de temps donnée.
Ce type de produit pourrait constituer une base relativement utile pour la
détermination des sites d'intervention en termes de reconstitution des
milieux dégradés.
Section 14.02 Conclusion
La télédétection constitue un important
volet pour une caractérisation de l'état et de la dynamique des
formations végétales. Les modifications de la couverture
végétale influent fortement sur le bilan du carbone et
nécessitent qu'on puisse caractériser la dynamique de l'espace
géographique afin de mieux définir les zones en fonction de leur
état de dégradation ou de conservation. Ces informations sont par
ailleurs très utiles dans l'élaboration et la mise en ~uvre de la
gestion et l'aménagement des formations et des groupements
végétaux. Les apports les plus remarquables concernent :
- les capacités de spatialisation et de
généralisation des résultats acquis sur des surfaces
limitées ;
- la souplesse de traitements des résultats grace à
l'outil informatique ;
- le complément de la mémoire collective,
apporté par les données diachroniques, quipermettent
en plus une évaluation de certains phénomènes.
Il faut cependant garder à l'esprit que les cartes de
végétation ne donnent qu'une information spatiale
générale. Elles ne peuvent pas par exemple remplacer les
données de terrain, elles ne peuvent que les compléter. Comme
tout outil, la télédétection doit être
utilisée judicieusement et se méfier des résultats
esthétiques obtenus sous forme graphique. La qualité de
l'information cartographique que l'on peut tirer des images satellitaires
dépend de la pertinence des connaissances acquises avant ou après
le traitement des images. Un spécialiste de la
télédétection (appliquée à l'étude de
la végétation) doit chercher à acquérir un capital
d'expérience et une aptitude à convertir l'information «
abstraite >> des images sur la base d'une connaissance sOre de la
réalité « concrète >> ; ce qui établit
un `dialogue image-terrain'. Les efforts qui sont entrain d'être
menés tant sur le plan d'une meilleure connaissance des
phénomènes biologiques que sur celui de l'amélioration de
la sensibilité spectrale des capteurs, permettent d'envisager un
affinement dans la précision des analyses à plus ou moins long
terme.
Les dynamiques observées dans ce chapitre montrent une
variation différenciée en fonction des milieux et des
interventions humaines. L'implication de l'augmentation du NDVI dans la
dynamique du carbone est parfois délicate à induire. Le plus
souvent, une augmentation des valeurs de NDVI traduit une activité
photosynthétique plus importante résultant d'une
séquestration de carbone. Cependant, la dynamique peut être
positive (R2> 0), alors la pente (taux de croissance) est
très faible. C'est ce qu'on a pu observer par exemple dans la partie
nord du Sénégal et le Sud de la Mauritanie.
Bibliographie
Abdelgalil, E.A., Cohen S.I. 2001. Policy modelling of the
trade-off between agricultural development and land degradation--the Sudan
case. Journal of Policy Modeling (23):847-874.
Achard, F., Ansseau, C., Beaubien, J., Defourny, P., Jaton,
A., Lemieux, G., Royer, A., Thomson K., 1996. La
télédétection des forêts. In: AUPELF, et
al. (Editor), Précis de Télédétection Volume
II. AUF, Québec, pp. 227-265.
AFRICOVER, 1997. Classification de l'occupation du sol, FAO,
Roma.
Barbier E.B. 2000. The economic linkages between rural poverty
and land degradation:
some evidence from Africa. Agriculture, Ecosystems and
Environment (82):355-370. Bassett, T.J., Zuéli K.B. 2000. Environmental
discourses and the Ivorian Savanna. Annals of
the Association of American Geographers, 90 (1):67-95.
Benz, U.C., Hofmann, P., Willhauck, G., Lingenfelder, I.,
Heynen M. 2004. Multi-resolution, object-oriented fuzzy analysis of remote
sensing data for GIS-ready information. Journal of Photogrammetry & Remote
Sensing (58):239-258.
Cambell J.B., 1996. Introduction to Remote Sensing. Taylor and
Francis, Londres, 622 p. Chokor, B.A., Odemerho F.O. 1994. Land degradation
assessment by small-scale traditional
African farmers' implications for sustainable conservation
management. Geoforum,
25 (2):145-154.
Collins, J.B., Woodcock C.E. 1994. Change detection using the
Gramm-Schmidt Transformation applied to mapping forest mortality. Remote
Sensing of Environment (50):267-279.
Collins, J.B., Woodcock C.E. 1996. An assessment of several
linear change detection techniques for mapping forest mortality using
multitemporal LANDSAT-TM data. Remote Sensing of Environment (56):66-77.
Elberling, B., Touré, A., Rasmussen K. 2002. Changes in
soil organic matter following groundnut-millet cropping at three locations in
semi-arid Senegal, West Africa. Agriculture Ecosystems & Environment
(96):37-47.
Feller Christian. 1977. Évolution des sols de
défriche récente dans la région des Terres Neuves
(Sénégal Oriental). Cahiers ORSTOM, Série Biologie, XII
(N° 3):199-211.
Flasse, P.F., Verstraete, M. , 1993. Monitoring the
environment with vegetation indices: comparison of NDVI and GEMI using AVHRR
data over Africa, Joint Research Centre, Ispra.
Grepperud S. 1996. Population Pressure and Land Degradation:
The Case of Ethiopia. Journal of Environmental Economics and Management
(30):18-33.
IPCC, 2000. Land use, Land Use Change and Forestry. WMO, UNEP,
377 p.
Jones S. 1996. Discourses on Land Degradation in the Uluguru
Mountains, Tanzania: Evolution and Influences. Journal of Rural Studies, 12
(2):187-199.
Lambin, E.F., Turner, B.L., Geist, H.J., Agbola, S.B.,
Angelsen, A., Bruce, J.W., Coomes, O.T., Dirzo, R., Fischer, G., Folke, C.,
George, Homewood, P.S., Imbernon, J., Leemans, R., Lin, X., Moran, E.F.,
Mortimore, M., Ramakrishna, P.S., Richards, J.F., Skanes, H., Steffent, W.,
Stone, G.D., Svedin, U., Veldkamp, T.A., Vogel, C., Xu J. 2001. The causes of
land-use and land-cover change: moving beyond the myths. Global Environmental
Change, 11:261-269.
Lambin, E., F., Geist, H.J., Lepers E. 2003. Dynamics of land
use and land cover change in
tropical regions. Annual Reviews of Environmental Resources
(28):205-241.
Mayaux, P., Grandi, D., G., Malingreau J.P. 2000. Central African
Forest Cover Revisited: a
multisatellite analysis. Remote Sensing of Environment
(71):183-196.
Muchena, F.N., Onduru, D.D., Gachini, G.N., Jager, D. A. 2004.
Turning the tides of soil degradation in Africa: capturing the reality and
exploring opportunities. Land Use Policy (In Press).
Prenzel B. 2003. Remote sensing-based quantification of
land-cover and land-use change for planning. Progress in Planning
(61):281-299.
Rasmussen, K., Fog, B., Madsen J.E. 2001. Desertification in
reverse? Observations from Northern Burkina Faso. Global Environmental Change
(11):271-282.
Rasmussen, K., Nielsen, T.T., Mbow, C., Wardell A.D. 2006.
Land degradation in the Sahel: An apparent scientific contradiction.
Proceedings of the 17th Danish Sahel Workshop, 6-7 November 2006:37-43.
Rogan, J., Chen D.M. 2004. Remote sensing technology for
mapping and monitoring land-cover and land-use change. Progress in Planning
(61):301-325.
Sankhayan, P.L., Hofstad O. 2001. A village-level economic
model of land clearing, grazing, and wood harvesting for sub-Saharan Africa:
with a case study in Southern Senegal. Ecological Economics (38):423-440.
Touré A., 2002. Contribution à l'étude de
l'évolution des réservoirs de carbone en zone nordsoudanienne au
Sénégal., Ecole Polytechnique Fédérale de Lausanne
(EPFL), 158 p.
Verstraete, M.M., Brink, A.B., Scholes, J.B., Beniston, M.,
Smith M.S. 2008. Climate change and desertification: Where do we stand, where
should we go? Global and Planetary Change, 64:105-110.
Verstraete, M.M., Scholes, R.J., Smith M.S. 2009. Climate and
desertification: looking at an old problem through new lenses. Frontiers in
Ecology and the Environment, doi:10.1890/080119:9p.
Article XV. CHAPITRE VII :
Article XVI. Adaptation
aux changements
environnementaux
|
Après avoir discuté les concepts clés que
sont l'impact, la vulnérabilité et l'adaptation, ce
chapitre
fait le point sur les paramètres importants des changements
environnementaux quiinfluent sur la vie des populations. Avec la
dynamique des ressources forestières comme
entrée, nous avons essayé de démontrer
que le climat est un facteur de vulnérabilité au même titre
que les facteurs économiques, démographiques et politiques. La
combinaison de ces différents éléments est
nécessaire pour mieux saisir les stratégies et options
d'adaptation dans les zones de savane étudiées. Les
résultats obtenus montrent que les formes d'adaptation portent sur des
stratégies au champ (fertilisation, technique de culture,
diversification, etc.) ou des réponses en dehors du champ (migration,
commerce, dynamique organisationnelle, formation, etc.). Ces résultats
devraient conduire à une documentation plus poussée sur la
question de l'adaptation, de ses enjeux et de ses avantages pour les
populations vulnérables par rapport aux bénéfices
générés par les projets se séquestration du
carbone.
|
Section 16.01 VII.1. La vulnérabilité
à la variabilité et au changement climatique
Les chapitres précédents montrent une
dégradation des écosystèmes et une réduction
significative de la biomasse, donc des stocks de carbone des zones de terroir
et dans une moindre mesure des Forêts Classées
étudiées. Ils ont aussi montré que même si les
savanes présentent un important potentiel de carbone, les conditions
actuelles du marché ne permettent pas au secteur forestier de
résoudre les questions de développement durable des populations
locales dont la survie dépend essentiellement d'une agriculture
productive. Le MDP dans le domaine de la foresterie laisse encore de nombreuses
questions sans véritables réponses. Il s'agit notamment des
avantages directs que peuvent tirer les populations locales très
pauvres. Ce constat s'explique entre autres par le fait que ce sont les Etats
qui se sont engagés et non les populations, détentrices de la
décision finale au quotidien. De cette situation découle
également d'autres interrogations particulièrement sur le
foncier, la répartition des revenus des transactions du crédit du
carbone et le degré d'engagement des populations concernées.
Il faut toutefois noter que les procédures pour
accéder à l'argent du carbone sont très complexes et
longues, et s'opposent à l'urgence de la survie des populations locales.
Ces questions de base font qu'il est important d'orienter la recherche sur
l'adaptabilité des populations aux changements climatiques. L'adaptation
est un ensemble de réponses directes ou non aux impacts observées
ou implicites des changements climatiques qui permettent la résilience
des communautés de base très vulnérables. Dans cette
partie de la recherche, nous explorons les facteurs structurants de la
vulnérabilité des populations locales et les options d'adaptation
mises en ~uvre, par exemple lors des périodes de stress liées aux
sécheresses récurrentes des années 1970-1992 et à
la dégradation de l'environnement. Pour procéder à cette
analyse, nous considérons toujours le cas du Saloum Oriental,
caractérisé par une profonde dégradation des ressources
naturelles, forestières en particulier.
L'étude de la vulnérabilité et de
l'adaptation nécessite un cadrage conceptuel, non seulement pour
comprendre les termes mais aussi pour mieux évaluer les approches et
méthodes développées afin d'analyser en profondeur les
composantes essentielles de la question et des stratégies actuelles en
matière de changement climatique.
Un bon éclairage a été fait dans de
nombreux documents et présentations scientifiques. Nous
en tentons
ici une synthèse tirée pour l'essentiel des travaux de
Adejuwon et al. (2001);
Burton et al. (2001); Olmos
(2001); Huq et Reid (2002); Adger et al. (2003); Huq et al.
(2003);
Janssen et Ostrom (2006); Winograd (2006). A cet égard,
la littérature montre une pluralité de significations selon les
institutions et les disciplines. Nous mettrons l'accent sur la
compréhension faite des termes vulnérabilité et adaptation
des populations rurales d'Afrique semi aride face aux changements
climatiques.
Le terme vulnérabilité a été
beaucoup utilisé dans le domaine des risques et des catastrophes
naturels, où il signifie: « le degré selon lequel une
unité exposée est soumise à une perturbation ou à
une contrainte, et traduit du coup la capacité (ou
l'impossibilité) de l'unité à risque d'en faire face, ou
de s'adapter de manière fondamentale (en devenant un nouveau
système ou en disparaissant) » (Kasperson et al.,
2000). Dans le cas des catastrophes le terme est utilisé dans le
sens de «degré de perte résultant d'un
phénomène néfaste » (glossaire UNOCHA
-Coordination des affaires humanitaires des Nations Unies).
Sur le plan socio-économique, la littérature sur la
pauvreté et le développement mettent l'accent sur les conditions
sociales, économiques et politiques actuelles (Winograd, 2006).
IPCC recommande une définition de la
vulnérabilité presque exclusivement reliée aux changements
climatiques (Adejuwon et al., 2001; IPCC, 2001; 2003):
«degré selon lequel un système est susceptible, ou se
révèle incapable, de faire face aux effets néfastes des
changements climatiques, notamment à la variabilité du climat et
aux conditions climatiques extrêmes. La vulnérabilité est
fonction de la nature, de l'importance et du taux de variation climatique
auxquels un système se trouve exposé; de sa sensibilité,
et de sa capacité d'adaptation».
Dans ses différentes composantes, il apparaît que
la vulnérabilité est une notion plurielle qui est fonction d'un
côté du ou des risques, des dangers et de l'exposition au stress ;
de l'autre, des options et réponses d'adaptation. Les
déterminants de la vulnérabilité peuvent être
biophysiques, dans ce cas on s'intéresse aux processus
écologiques de la vulnérabilité, l'exposition et la
susceptibilité à des processus de changements environnementaux.
Ces déterminants peuvent être d'ordre socio-économique,
dans ce cas on se focalise sur les déterminants politiques,
socioéconomiques, culturels et institutionnels de la
vulnérabilité
Les systèmes ont la propriété de
réagir au stimulus pour atténuer leurs conséquences
négatives. Ce type de réponse correspond au concept
d'adaptation dont la mise en ~uvre dépend des
capacités du système lui-même à s'adapter. Le terme
adaptation est perçu comme l'ensemble des stratégies d'ajustement
d'un système pour atténuer les impacts des changements
climatiques, de tirer parti de la nouvelle situation ou de se résigner
aux conséquences. Par contre, le terme
vulnérabiité traduit la
susceptibilité du système aux impacts négatifs. Plus un
système est doté de capacités d'adaptation, moins grande
sont
ressentis les impacts des changements environnementaux. Les
capacités d'adaptation peuvent être celles mobilisées par
le système et/ou celles mises en ~uvre par les sociétés
humaines qui vivent dans ce système. On parlera alors d'adaptation
autonome (réaction normale et naturelle du système), ou
d'adaptation planifiée (réaction d'anticipation liée au
projet des hommes). A partir de ces deux situations Robledo et Forner (2005)
déduisent deux types de vulnérabilités :
- Vulnérabilité = impacts potentiels -
capacités d'adaptation du système
- Vulnérabilité = impacts - capacités
d'adaptation autonome - adaptation planifiée
L'adaptation peut être considérée alors
comme l'ajustement d'un système en réponse avec un stimulus ou
ses effets (Burton et al., 2002). Selon IPCC (2001), les
systèmes qui s'adaptent aux changements climatiques sont soit naturels
(écosystèmes forestiers) ou humains, et les formes d'adaptation
cherchent à modérer les impacts ou exploiter les
opportunités liées aux changements.
Le terme « impact » traduit de façon simple,
exprime l'ensemble des conséquences négatives (ou positives selon
la perception) liées aux changements climatiques. Du fait de la grande
variabilité de situations géographiques, environnementales et
socio-économiques, un facteur comme les changements climatiques peut
avoir des conséquences négatives sur une zone et entraîner
des effets positifs ailleurs. L'exemple peut être pris sur l'augmentation
de la pluviométrie des zones inondables qui peut causer d'énormes
revers alors que pour les sociétés dont l'agriculture
dépend de la pluie, ces hausses de pluies sont dans certaines
proportions un facteur bénéfique (Robledo et Forner,
2005). Les précipitations de 2005, 2008 et 2009 au
Sénégal and dans la sous-région constituent des exemples
récents de cette situation. L'importance de l'impact des changements
climatiques dépend de quelques facteurs que sont son ampleur
(l'importance du changement), sa probabilité d'occurrence (ainsi que la
probabilité de son intensité), son taux et sa durée
(rapidité des modifications et la durée de celles-ci), mais aussi
de la réactivité du système, de sa tolérance et de
ses capacités d'adaptation.
On parle souvent de résilience, et ce
terme traduit le résultat de l'adaptation puisqu'elle traduit la
capacité d'absorption du problème par le système. La
résilience traduit les capacités existantes qui permettent
d'absorber les crises sans effort supplémentaire, alors que l'adaptation
traduit de nouvelles options, attitudes et pratiques permettant de faire face
à des stress et à des perturbations externes qui peuvent
être d'ordre politique, social, économique ou environnemental
(Adger et Brooks, 2002; Adger et al., 2003).
En termes pratiques donc l'adaptation est une réponse
à la vulnérabilité à travers un changement de
procédures, de pratiques et d'options de développement visant
à limiter ou effacer les dommages potentiels ou à tirer
bénéfice des opportunités créées par la
variabilité et les changements climatiques (Winograd, 2006). Ces
réponses dépendent de ce qu'on appelle la (ou les)
capacité (s) d'adaptation qui exprime les moyens dont dispose une
communauté, un individu à s'adapter aux effets et aux impacts du
changement climatique (y compris la variabilité climatique).
L'adaptation dépend essentiellement des ressources économiques,
sociales, techniques et humaines d'une société. Il faut toutefois
noter que l'adaptation exprime aussi les profits, les avantages ou
opportunités qu'on peut tirer de nouvelles conditions climatiques. Dans
ce cadrage, les systèmes les plus vulnérables sont
évidemment ceux qui sont sensibles aux infimes modifications du
climat.
Cette notion d'adaptation est souvent opposée à
celle de l'atténuation qui exprime les
mécanismes ou actions visant à réduire l'exposition et la
susceptibilité à des perturbations ou des stress. Il s'agit
essentiellement des interventions humaines visant à réduire les
émissions de gaz à effet de serre en provenance de
différentes sources ou à renforcer leur absorption par des
puits.
On peut alors voir l'adaptation comme un aspect de
l'atténuation en Afrique car la promotion du développement
durable permet d'investir des stratégies visant un environnement durable
à faible émission de carbone
Section 16.02 VII.2. Place de l'adaptation dans la
CCNUCC
L'adaptation est une importante dimension de la Convention sur
les Changements Climatiques, car même si chaque pays respectait les
engagements de Kyoto en terme d'émission de GES, les teneurs de ces gaz
dans l'atmosphère vont prévaloir pendant encore plusieurs
années. Il est important alors de combiner l'adaptation à
l'atténuation pour promouvoir une réponse
équilibrée à la variabilité climatique. Sur ce
plan, il faut souligner que l'ampleur des impacts des changements climatiques
dépendra d'une part de la vigueur de la dynamique de modification du
climat et d'autre part du dynamisme et de l'adéquation des
réponses apportées à ces changements. Il s'agira d'un jeu
de force.
Le futur de l'adaptation dépendra aussi de la
capacité à anticiper sur les modifications du climat. Les
résultats des modèles sophistiqués repris par IPCC dans
son Quatrième Rapport d'Evaluation, montrent parfois de fortes
différences (divergences) dans les scénarios sur le long terme
(horizon 2100), dépendant de différents niveaux de stabilisation
possibles pour
les gaz et l'évolution de l'équilibre climatique
global. La dissimilitude des projections réside aussi sur les
scénarios retenus sur le plan atmosphérique, économique,
infrastructurel, social et démographique.
Le futur de l'adaptation est alors une question de coOts pour
les solutions et réponses apportées aux changements climatiques.
Plusieurs alternatives envisagées à l'heure actuelle, en termes
de consommation énergétique, de modification des systèmes
de culture, d'amélioration de la productivité, etc.,
nécessitent un investissement de départ que des populations
vulnérables ne peuvent pas envisager dans le cadre de l'auto adaptation.
Dès lors, il sera nécessaire aux Etats de structurer une forme de
gouvernance qui inscrit l'adaptation à tous les niveaux politiques pour
mieux amortir les impacts des changements climatiques. Toutefois, il faut
veiller à ce que Adejuwon et al. (2001) appellent la mal
adaptation qui peut générer encore plus de coOts que l'absence
d'adaptation. Les stratégies d'adaptation qu'on met en place doivent
être minutieusement examinées pour s'assurer de leur
adéquation avec la réalité, le contexte, et les moyens
disponibles pour les réaliser.
La plus grande contribution de l'Afrique et du
Sénégal à l'effort d'atténuation passera par un
grand engagement pour l'adaptation. Une bonne adaptation entraîne des
effets collatéraux qui atténuent la déforestation, la
dégradation des terres, ciblées comme facteur de
libération de GES. Les efforts d'atténuation en Afrique
requièrent, vu le contexte économique, le choix d'options
à moindre coOt sur le plan économique et social, ce qui ne ferait
que renforcer la vulnérabilité si les préoccupations des
populations ne sont pas prises en compte. Ainsi, il est important de souligner
que l'engagement sur l'atténuation est une option qui ne doit pas
masquer l'adaptation qui est une nécessité.
Par conséquent, les premières réponses
aux changements du climat sont articulées sur des actions
internationales à travers la CCNUCC qui s'est plus appesantie sur
l'atténuation même si l'adaptation est mentionnée dans la
convention en son Article 4-e. Toutes les parties doivent
«Préparer, en coopération, l'adaptation à
l'impact des changements climatiques et conçoivent et mettent au point
des plans appropriés et intégrés pour la gestion des zones
côtières, pour les ressources en eau et l'agriculture, et pour la
protection et la remise en état des zones frappées par la
sécheresse et la désertification, notamment en Afrique,.
.».
Dans le Protocole de Kyoto, les pays industrialisés (sauf
l'Australie, les USA et la Russie) (Partis Annexe I de la Convention) ont
accepté de réduire leurs émissions de GES à un
niveau de 5 % au-dessous des niveaux de 1990 et ceci entre 2008 et 2012
(première période d'engagement). Le Protocole a aussi
autorisé les partis à développer des activités de
puisage
de GES à travers des stratégies de modification
des formes d'utilisation des terres et de la foresterie (déforestation
évitée, séquestration de carbone). Les quantités de
GES, notamment de CO2 séquestré peuvent faire l'objet de
transactions commerciales entre pays du Sud et pays du Nord. Les deux autres
alternatives sont la «mise en ~uvre conjointe" entre pays
développés et les «mécanismes de développement
propre» (MDP) qui encouragent la réduction des émissions
à travers des projets entre pays en développement plus
vulnérables et pays développés. Sur le plan du suivi, le
Protocole de Kyoto a établi les inventaires nationaux (Communications
Nationales) pour tous les partis afin de bien évaluer les efforts
consentis dans le sens de la réduction des GES et mieux orienter la mise
en ~uvre de la convention (CCNUCC, 1992). Le but de toutes ces alternatives est
de réduire les GES, en s'appuyant sur le bras scientifique de la
Convention connu sous le nom IPCC (GIEC en Français) qui est
chargé de faire continuellement le point scientifique sur la question
des changements climatiques afin d'éclairer la décision
politique. IPCC a à son compte 4 rapports (1990, 1995, 2001 et 2007).
Ces rapports ont été consolidés par des rapports
complémentaires et des rapports spéciaux pour préciser
certaines questions spécifiques. Ces rapports prennent en compte le
maximum de facteurs possibles pour mieux comprendre les causes et les
conséquences des changements climatiques. Les analyses couvrent une
large gamme de thèmes depuis la physique de l'atmosphère avec les
forçages radiatifs, les impacts des changements climatiques, les
techniques d'atténuation et les propositions politiques pour
répondre à ces nombreux défis. A côté des
rapports généraux, IPCC a développé une
série de rapports régionaux qui mettent en exergue les
spécificités en fonction des continents et des
sous-régions.
La Convention est soutenue dans sa mise en ~uvre par des
institutions internationales comme la Banque Mondiale, le Fonds Mondial pour
l'Environnement (FEM), le Programme des Nations Unies pour l'Environnement
(PNUE), mais aussi plusieurs institutions régionales.
Au niveau des pays, la Convention encourage la participation
à la mobilisation des informations scientifiques permettant à
IPCC de structurer ses rapports scientifiques (communications nationales). Le
rôle des Etats signataires est surtout de faire en sorte que les
décisions prises soient appliquées afin de prouver sa
contribution effective dans un
cadrage basé sur le principe : « à chacun
selon ses besoins, à chacun selon ses capacités ».
Iis'agit de l'engagement commun et différencié. Les
stratégies nationales sont laissées à la
libre appréciation des Etats. Dans ce contexte, les
énergies renouvelables sont citées parmi
les solutions les
plus explorées dans ce contexte. En Afrique, le développement des
cultures
de bioénergie est entrain de prendre une dimension
débordante. Cette activité qui pourrait
être lucrative ne devrait pas être un piège
qui ralentirait l'investissement sur l'adaptation qui elle couvre une dimension
socio-économique essentielle pour la survie des populations locales.
L'adaptation intègre de façon cohérente la question du
développement durable de populations très vulnérables et
dépendantes des ressources naturelles pour leur survie.
La question du développement durable est en soit
antérieure à la CCNUCC. Elle a été lancée
depuis 1987 par la Commission Mondiale sur l'Environnement (Commission
Brundtland) qui a défini le développement durable comme un
«développement permettant de satisfaire les besoins actuels
sans compromettre la capacité des générations futures de
satisfaire leur propres besoins». Son but est la mise en place d'un
système de gestion de l'environnement stable dans la production de
ressources suffisantes et perpétuelles pour répondre
continuellement aux besoins de la société. La question principale
du concept est de savoir jusqu'où cette notion de besoins des
populations doit aller (Pyramide de Maslow). Faut-il la limiter aux besoins de
nourriture, d'habillement, de logement, de santé ou devrait-il inclure
les notions de confort, de commodité, de qualité de vie, etc. Le
consensus est loin d'être atteint sur ce point (Adejuwon et al.,
2001).
Ces questions de développement durable se posent dans un
contexte particulier caractérisé par une situation où:
- les impacts physiques des changements climatiques sont
supposés avoir de larges répercussions sur les systèmes de
production ;
- les pays africains sont caractérisés par une
forte dépendance d'une agriculture peu performante pour l'essentiel
pluviale ;
- la pauvreté parfois chronique offre peu d'espoir
immédiat d'atténuation ;
- les capacités économiques et techniques sont
encore loin d'atteindre un niveau appréciable.
Dans ce contexte, le Sénégal comme les autres
pays du Sahel est très sensible aux modifications de la
pluviométrie qui sont des manifestions de la variabilité
climatique. Cette sensibilité est liée au faible niveau de
développement économique, à la dépendance aux
ressources naturelles, ainsi qu'à l'occurrence de
phénomènes extrêmes qui interviennent sur des espaces
temporels très courts (Thomas et al., 2005). Des modifications
brusques et profondes comme la sécheresse, des pluies excessives, des
modifications saisonnières du climat (événements
extrêmes), sont autant de facteurs qui peuvent exposer les populations
rurales à de nouvelles épreuves, posant de vrais défis
politiques et stratégiques aux Etats. Les capacités d'une
société à contenir les phénomènes,
dépendent des avantages qu'on peut en tirer ou des inconvénients
qu'on doit éviter. Les situations sont variables, certains
groupes peuvent assez rapidement trouver leur compte dans les
changements qui s'opèrent, alors que d'autres sont plus
vulnérables. Ces différences posent le problème de la
relativité de la vulnérabilité des sociétés
humaines aux impacts des changements climatiques en relation avec les
capacités d'adaptation.
Section 16.03 VII.3. Complexité et
portée pratique de l'adaptation
L'adaptation est une notion très complexe. Elle
relève d'interactions complexes entre les facteurs de
vulnérabilité (facteurs humains et physiques) et les
éléments de réponses (économiques, techniques,
sociale, politiques) structurées par les populations.
Plusieurs modèles conceptuels ont été
proposés pour caractériser la vulnérabilité et
l'adaptation. Le modèle développé par Huq et Reide (2002)
montre des interrelations complexes entre l'adaptation et les facteurs qui
l'influencent.
Figure 93. Diagramme de l'adaptation (Huq et
Reide, 2002)
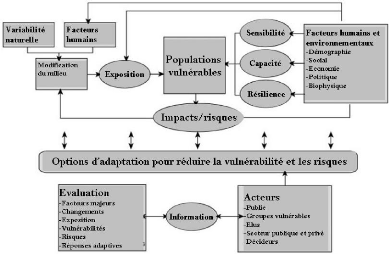
D'autres modèles simplifiés ont
été proposés par Adejuwon et al. (2001);
Burton et al. (2001); Olmos (2001); Adger et Brooks (2003); Thomas
et al. (2005) ; Burton et al. (2002) ; Winograd (2006) mais ils
ont tous en commun une vision transversale qui fait intervenir des facteurs
humains et physiques montrant essentiellement que l'adaptation est un facteur
dynamique qui a commencé bien avant et qui est en mutation
perpétuelle dans le temps et dans l'espace. C'est grace aux ajustements
des pratiques des populations aux nouvelles conditions
environnementales que les populations du Sahel ont
été résilientes aux différentes modifications du
climat. Ces mutations sont elles-mêmes fonction de la dynamique des
facteurs structurants qui sous-tendent la vulnérabilité (facteurs
climatiques, économiques, politiques, environnementaux, ..) et des
capacités des systèmes d'apporter des réponses
adéquates.
Dans la CCNUCC, il existe deux formes d'adaptation qui
impliquent deux types de stratégies. Le premier consiste à
prévenir les interférences graves des activités humaines
sur l'atmosphère avec la concentration de gaz à effet de serre,
il s'agit là de l'atténuation. Le second
consiste à minimiser la vulnérabilité des
sociétés humaines et des écosystèmes aux
changements climatiques par les processus d'adaptation. Dans
les deux cas, il est nécessaire de mesurer l'impact des changements
climatiques pour dimensionner l'effort sur la forme d'adaptation choisie. Les
enjeux pour des pays comme le Sénégal se situent au niveau de la
réduction de la vulnérabilité. Selon Burton et al
(2002), il existe un dilemme entre l'effort d'atténuation et celui
d'adaptation. Selon cet auteur, plus l'impact est important plus l'effort
d'atténuation l'est; mais plus l'effort d'adaptation est grand pour la
réduction de la vulnérabilité, moins il sera urgent de
réduire les émissions de GES. Il faut équilibrer entre les
efforts d'atténuation et ceux d'adaptation en utilisant une approche
appropriée.
Dans la même lancée, Burton et al.
(2002) et Winograd (2006) ont essayé de proposer un canevas
permettant d'harmoniser l'approche d'évaluation de la
vulnérabilité en simplifiant la prise en compte des relations
entre les différents facteurs. La figure 94 insiste sur la mise en
relation dynamique entre les impacts et la vulnérabilité pour
mieux appréhender les formes d'adaptation, plus particulièrement
celle dite autonome et qui relève des populations affectées.
Figure 94. Liens entre Changements Climatiques,
Adaptation et Atténuation (Burton et al., 2002 ; Huq et
al., 2003).
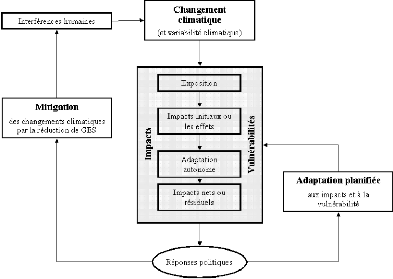
Dans la plupart des cas, le climat n'affecte les
sociétés humaines qu'indirectement par la modification des
conditions et du milieu de vie. Pour les sociétés humaines
dépendantes des ressources naturelles pour leur survie, les effets des
changements climatiques sont à la fois directs et indirects puisque ces
sociétés ne se donnent pas de marges de manuvre pour
d'éventuelles modifications des conditions climatiques.
L'adaptation peut prendre plusieurs formes. Selon Adejuwon
et al (2001), les adaptations communautaires sont invariablement des
réponses réactives aux stress issus des changements climatiques
sans interventions des services publics. Parallèlement on peut avoir une
série de mesures structurelles à travers la mise en ~uvre de
stratégies nationales visant à réduire au
Sénégal la vulnérabilité des populations dans
plusieurs domaines :
- agricole (exemple GOANA, REVA),
- forestier (projets pour satisfaire les besoins en produits
ligneux)
- stress hydrique (bassins de rétention, Projet Sectoriel
Eau, amélioration des systèmes d'irrigation) ;
- niveau de revenu des populations (microcrédits,
subventions, diversification des activités de production) ;
- social (dynamique organisationnelle, éducation,
santé) ;
- démocratique (bonne gouvernance, démocratie
interne), etc.
Les formes d'adaptation communautaires sont les plus
difficiles à réussir du fait de la pauvreté chronique de
certaines communautés. Que l'adaptation soit une initiative directe des
populations ou initiée par l'Etat, elle est toujours un processus
complexe pas exclusivement et nécessairement lié aux changements
climatiques. D'autres déterminants peuvent influencer la
vulnérabilité des populations en plus des changements
climatiques. On peut citer les plus apparents que sont les politiques de
développement mal adaptées (ajustement structurel) et la
dynamique des prix de produits de base sur le marché international (prix
de l'arachide, prix du pétrole, prix du cacao) (Adger et Brooks, 2002).
La figure 103 donne une idée simplifiée du concept d'adaptation
en Afrique (Mbow et al., 2008). Les formes d'adaptation
planifiées peuvent être réactifs (pour les chocs et les
stress) ou anticipatif pour contenir des impacts prévus des changements
climatiques.
Un aspect important de la vulnérabilité et de
l'adaptation est sa dimension historique. L'analyse dans le temps est non
seulement un regard sur les formes d'adaptation du passé, mais aussi une
analyse de la façon dont un groupe social projette de construire son
avenir et d'en mesurer l'adéquation par rapport aux défis
annoncés par la tendance actuelle. Le recours au passé peut se
faire grâce aux profils historiques utilisés par les sociologues,
mais aussi à la revue de la littérature et des archives qui
capitalisent des réponses que chaque communauté a pu
développer face à des problèmes environnementaux.
L'ouverture vers le futur est moins évidente et n'est basée
à l'heure actuelle que sur un développement de scénarios
ou de modèles conceptuels du genre « si. alors » sans beaucoup
de certitude vu le caractère dynamique et quelque peu aléatoire
de l'adaptation (plus on s'ajuste au système, plus on le maîtrise
et moins on devient vulnérable). Sur ce plan, certains
spécialistes proposent l'approche par analogie qui veut qu'on cherche
des conditions similaires des manifestations des changements climatiques dans
le passé ou au niveau d'autres groupes sociaux contemporains et
identifier les stratégies (les bonnes) ayant permis de survivre aux
conditions difficiles liées au climat (Adger et Brooks, 2002).
Là-dessus, la dimension culturelle (réceptivité des
technologies colportées, barrières culturelles), pourrait
constituer un problème dont il faut tenir compte.
Il existe une grande variété dans les formes
d'adaptation, mais dans bien des situations où la
vulnérabilité est très avancée (état de
pauvreté extrême), les solutions sont difficiles à
envisager dans des formes d'adaptation autonome. Dans des conditions de
pauvreté, les formes d'adaptation qui accrochent plus les populations
sont celles où le retour à l'investissement se fait dans un
très court terme, du fait de l'urgente nécessité à
trouver une
solution de survie. L'auto adaptation peut être
entravée par certaines barrières notamment financières
(microcrédit), institutionnelles/juridique (police forestière,
décentralisation) ; ou le manque d'expertise pour le
développement de l'innovation locale (capacités techniques).
C'est pour ces raisons, en grande partie, que l'adaptation des populations
locales porte sur des solutions simples à bénéfice
immédiats. Parfois, cette préférence pour les solutions
à action rapide dévie les populations locales des perspectives du
long terme. Et quand le court terme l'emporte, on peut engager des alternatives
qui mettent en péril les ressources dans le long terme (« advienne
que pourra »). C'est ainsi qu'on a souvent vu des populations s'adonner
à la collecte de ressources forestières notamment le bois de
valeur et les fruits. Les bénéfices qu'on peut en tirer incitent
à une exploitation aussi démesurée que non
planifiée et dans un temps assez court. Cette situation entraînant
alors un épuisement rapide de la ressource. La
vulnérabilité qui a été atténuée en
apparence pendant un court terme devient encore plus cruciale parce que la
ressource sur laquelle on s'est basée est vite dilapidée. C'est
pour cette raison que les plans d'aménagement et de gestion sont
essentiels à la gestion des ressources naturelles.
Chaque groupe social a eu à faire face dans le
passé à des défis environnementaux qui ont
nécessité des formes d'adaptation variables en nature et en
engagement communautaire. Il s'agit donc d'une construction sociale
influencée par des dynamiques politiques et institutionnelles (Adger
et al., 2003).
Section 16.04 VII.4. Approches pour l'étude des
impacts et de la vulnérabilité
Les méthodes pour évaluer les impacts des
changements climatiques et la vulnérabilité des systèmes
nécessitent une approche interdisciplinaire. Pour faire simple, certains
auteurs proposent une superposition d'un scénario climatique à un
scénario social.
Cf.
http://unfccc.int/program/mis/meth/view.html.
Les méthodes développées pour l'analyse des
impacts de l'adaptation et de la vulnérabilité suivent les lignes
directrices de IPCC (2001). Trois grandes approches sont proposées :
- l'approche impact : est la plus simple et est basée sur
des relations cause-effet. Elle
permet d'identifier les changements de variables climatiques
particulières et leur
impact potentiel sur les éléments du système
affecté ;
- l'approche interaction : en plus de l'identification des
impacts climatiques, cette méthode prend en considération les
processus qui accentuent ces impacts, entraînant ainsi une prise en
compte de la vulnérabilité sur d'autres facteurs ayant des
impacts négatifs sur le système ;
- l'approche intégrée : les
éléments étudiés doivent être
intégrés dans un système qui lui-même est
intégré de sorte que toutes les interactions possibles entre les
systèmes et leurs éléments affectés par les
changements climatiques puissent être analysées.
Figure 95. Etapes définies par IPCC pour
l'analyse de l'impact des changements climatiques.

Burton et al. (2002) développent des
idées qui sous-tendent bien évidemment ces étapes. Par
exemple, si on veut évaluer les impacts futurs des changements
climatiques, on ne se limite pas nécessairement à des
scénarios climatiques, mais on doit aussi considérer
l'état du système affecté en incluant les scénarios
socio-économiques. Les situations économiques actuelles et
futures et la vulnérabilité qui en découle sont plus
déterminantes sur les impacts nets que les changements climatiques
projetés. Les étapes 6 et 7 supposent que les réponses
adaptatives sont connues ce qui n'est toujours pas le cas.
Il est important de noter que le point 4 de cette approche
donne plus d'importance aux changements futurs et ignore les impacts et les
problèmes de vulnérabilité actuels. En outre cette
étape 4 requiert la sélection de scénarios climatiques ou
des projections issues des Modèles Climatiques Globaux. Ces informations
combinées à des données biophysiques et
socio-économiques ont permis le développement d'une importante
littérature synthétisée dans le rapport de IPCC (2001).
Mais dans ce rapport, les impacts biophysiques sont plus apparents et plus
convaincants que les dimensions socio-économiques. Les plus gros efforts
au sein des pays ont été consentis sur l'utilisation des
scénarios climatiques et les impacts généraux ou de
premier ordre, alors que peu d'attention a été portée
jusque là aux adaptations.
La méthode standard proposée par IPCC
présente des limites pour une analyse approfondie de la
vulnérabilité et des stratégies d'adaptation. Ces
contraintes sont liées à l'utilisation des scénarios
climatiques. Pour les étapes 6 et 7, on est souvent confronté
à une longue liste d'impacts potentiels des changements probables du
climat sur la socio-économie. Il existe deux niveaux d'incertitudes :
celui lié aux scénarios climatiques, et celui liés aux
futures conditions socio-économiques ; ces incertitudes se propagent sur
les analyses de la vulnérabilité et des impacts. Il devient
dès lors difficile de prédire ce qui s'adapte à quoi et
les analyses d'adaptation et de vulnérabilité deviennent
spéculatives quand elles sont basées sur des prédictions
climatiques et socio-économiques incertaines. Cette situation a
réduit l'ardeur des décideurs politiques et des scientifiques,
confrontés à des défis plus urgents et plus
prégnants comme la productivité économique et le
développement durable.
Il faut ajouter à cela que les Modèles
Climatiques Globaux sont le plus souvent à des échelles
grossières qui ne correspondent pas aux échelles
opérationnelles d'analyse de la vulnérabilité. A ce titre,
des recherches sont entreprises pour arriver à des détails plus
fins (downscaling) permettant d'obtenir des informations plus
fouillées au niveau local. Il faut toutefois noter que ces analyses
fines sont souvent assujetties à une certaine imprécision qui
impose la prudence dans l'utilisation des résultats (Burton et
al., 2002).
Au Sénégal, les stratégies d'adaptation
sont plus associées aux variabilités climatiques et aux
phénomènes externes à la variabilité climatique.
Une analyse adéquate de l'adaptation nécessite une prise en
compte de la combinaison de plusieurs variables et la durée des
séquences climatiques. Il faut ajouter que la plupart des
évaluations d'impacts des changements climatiques ne prennent pas en
compte la large panoplie d'adaptations possibles. Par exemple, les
modèles de productivité agricole (qui se focalisent sur
l'irrigation, la fertilisation, la croissance des plantes...) ne prennent pas
en compte les stratégies non agricoles comme les facteurs du
marché, les droits de propriété terrienne, etc.
Les analyses des impacts par la méthode standard posent
aussi le problème de prise en compte du contexte politique et des
acteurs en jeu surtout ceux qui sont les plus exposés.
Une meilleure analyse des impacts, de la
vulnérabilité et des formes d'adaptation nécessite une
amélioration significative en partant des études de la
première génération développée selon la
méthode IPCC. Dans cette logique, il faut surtout éviter
d'adopter une logique prescriptive. Le principe du diagnostic va être
fondé sur un cadre conceptuel commun flexible permettant d'organiser la
recherche sur l'adaptation et la vulnérabilité. Les questions de
base que l'on peut se poser sont du genre : quelle est la nature de la
vulnérabilité ?
Comment va - t - on les mesurer ? Est - il possible de
créer une référence commune pour la
vulnérabilité ? Quel est le meilleur moyen de réduire la
vulnérabilité ? Qu'est ce qu'il faut considérer pour la
réduction de la vulnérabilité ? Quelles sont les
responsabilités de ceux qui sont considérés comme les
couches les plus vulnérables et celles qui doivent apporter leur
assistance pour la réduction de la vulnérabilité ?
De façon détaillée on cherchera dans le
cadre d'une étude de vulnérabilité à
répondre aux questions non exhaustives indiquées ci-dessous :
- Q uelle a été l'expérience récente
de la zone en termes de variabilité climatique et l'occurrence
d'extrêmes ?
- Q uels ont été les impacts économiques et
comment sont-ils répartis dans l'espace et dans les groupes sociaux ?
- Q uels sont les autres impacts non économiques (impacts
sociaux, environnementaux) connus ?
- Y a-t-il des tendances sur la variabilité climatique et
les événements extrêmes, si oui, à quoi pourra-t-on
les attribuer ?
- Y a-t-il des tendances sur les dommages engendrés, si
oui comment pourrait-on les expliquer ?
- Quelles ont été les options politiques et les
mesures prises pour réduire la vulnérabilité et quel est
le bilan de ces choix ?
- Jusqu'où vont les pratiques d'adaptation et quelles sont
les barrières, les obstacles, et les facteurs favorisants l'adaptation
?
- Comment les politiques publiques influencent-elles les impacts
et les formes d'adaptation ?
- Comment les politiques sur les risques climatiques
influent-elles sur la politique
économique et le développement durable ou les
autres stratégies en place ?
- Quel lien y a t-il entre les politiques sur les risques
climatiques et celles sur la
qualité de l'air, l'ozone, et autres questions
atmosphériques ?
- Quel lien y a t-il entre les politiques sur les risques
climatiques et la gestion des ressources naturelles et celle de l'environnement
?
- Quel est le niveau d'implication des acteurs dans le processus
de mise en place des politiques et comment cette implication peut être
améliorée ?
Le PNUE propose aussi une méthode relativement
similaire à celle de IPCC mais qui est plus précise s'il s'agit
d'étudier des aspects thématiques particuliers comme les impacts
sur les ressources forestières.
Figure 96. Modèle conceptuel d'analyse
de vulnérabilité proposé par le PNUE,
http://130.37.129.100/english/o
o/instituten/ivm/research/climatechange/handbook.htm
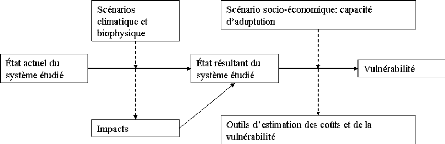
Dans l'analyse des impacts des changements climatiques, il est
important de considérer plusieurs types de scénarios. Il s'agit
en fait de superposer plusieurs scénarios et de chercher à
établir les interactions entre différentes variables physiques,
économiques, sociales, climatiques, etc.
Section 16.05 VII.5. Les outils utilisés pour
le choix des scénarios d'adaptation
L'opinion des experts est un outil
très utilisé pour opérer des choix de scénarios et
les analyses des relations entre ces scénarios et les
éléments du système affecté, tout en permettant de
poser des hypothèses pour anticiper le déroulement des
événements.
Les modèles : en plus des
modèles climatiques, dits Modèles de Circulation Globale, on a
plusieurs autres modèles de type biophysique, bioclimatique,
socio-économique et environnemental en utilisant les résultats
des modèles climatiques. Le modèle FORET est un exemple
permettant de suivre la dynamique des forêts en fonction de la
modification de différentes variables climatiques.
Les outils pour estimer le coût : ce
sont des outils monétaires pour analyser l'impact économique des
changements climatiques et les formes d'adaptation possibles. Les coOts
associés à des secteurs non marchands comme la
biodiversité sont cependant difficiles à appréhender.
L'analyse par contingence est une nouvelle approche pour mettre en exergue la
valeur des impacts à travers le `consentement à payer' pour un
service écosystémique.
Les SIG sont utilisés pour structurer
et intégrer des informations spatiales de type physique ou humain et
opérer des projections en utilisant les outils d'analyse
géostatistiques. Ils permettent de visualiser, de superposer et
d'examiner les stratégies d'adaptation.
Les bases de données capitalisent les
informations historiques sur différents aspects liés aux
changements climatiques, écologiques, économiques, sociaux, etc.
Ces bases peuvent indiquer des tendances qu'on peut utiliser pour anticiper
l'avenir.
En se fondant sur les études antérieures, il est
possible de considérer les informations contenues dans la
littérature et qui constituent des éléments de
vulnérabilité et d'adaptation en un endroit donné. Nous
partons du constat que les populations locales se sont toujours adaptées
à différents facteurs de stress et une méta-analyse de la
littérature grise permet de retracer les facteurs de stress et leur
évolution ainsi que les choix d'adaptation adoptés localement.
Dans les développements qui suivent nous appliquons cet exercice
à la zone du Saloum Oriental, de Kaffrine à Koungheul.
Section 16.06 VII.6. Méta-analyse de la
vulnérabilité et l'adaptation dans le Saloum Oriental
Le Saloum Oriental est caractérisé par une
profonde dégradation des ressources forestières en zone de
terroir (voir chapitre 6, carte d'occupation des sols). Ces modifications
rapides sont liées à des facteurs humains et naturels internes ou
exogènes. Cette zone est située au cur du Bassin
Arachidier16 qui est un espace de production
agricole dominé par la culture de l'arachide, en combinaison avec des
cultures vivrières.
L'utilisation des données historiques est une approche
bien appropriée pour analyser les tendances et l'ampleur des changements
ainsi que les interactions complexes entre l'homme et son environnement. L'un
des éléments les plus étudiés pour expliquer les
changements de l'environnement reste la dynamique des précipitations
annuelles qui affectent directement la production végétale, les
ressources forestières, pédologiques et pastorales, ainsi que les
modes de vie des populations locales. La variabilité climatique n'est
cependant pas le seul facteur de stress auquel les populations doivent trouver
des réponses. Il existe des facteurs économiques et politiques
qui influent sur les choix locaux d'utilisations de l'espace et des ressources
naturelles (Mortimore et Adams, 2001). Dans cette analyse de données et
d'informations acquises depuis les années 1950, nous avons essayé
de comprendre les facteurs principaux de la dynamique de l'environnement et
comment des réponses
16 Nom donnée à la zone où prédomine
la culture de l'arachide au Sénégal.
structurelles et autonomes ont aidé à absorber
nombre d'impacts liés à la dégradation de
l'environnement.
(a) VII.6.1. Le facteur climatique
La pluviométrie est un des premiers facteurs
évoqué par les populations comme cause de la dégradation
des ressources naturelles. En effet, la forte dépendance des populations
aux ressources naturelles les rend très vulnérables à une
diminution des pluies qui entraîne une réduction rapide des
services des écosystèmes. Au Sénégal, plusieurs
études ont montré une tendance générale de
réduction des précipitations même si on note un
léger mieux pendant les 10 dernières années. On a
enregistré par le passé une succession de séquences
sèches et humides sur les enregistrements pluviométriques du
Sahel (anomalies négatives), (L'Hôte et al., 2002) :
- 1910-1916 : période sèche de 7 ans
- 1950-1967 : période humide de 18 ans
- 1970-1993 : période sèche de 23 ans
Les bonnes pluviométries de 1994, 1999, 2003, 2005 et
2008, laissent penser à un retour d'une séquence humide du Sahel.
Cependant les modèles retenus par IPCC dans son quatrième rapport
d'évaluation projettent des climats plutôt arides pour le Sahel en
rapport avec les changements climatiques (IPCC, 2008).
L'analyse des écarts à la moyenne
pluviométrique sur deux stations de la zone d'étude de cas montre
une forte récession pluviométrique avec des signes de retour
d'une séquence humide depuis au moins 2003 (figures 97 et 98).
Figure 97. Ecart à la moyenne de la
pluviométrie mensuelle à Kaffrine (1931-2005)
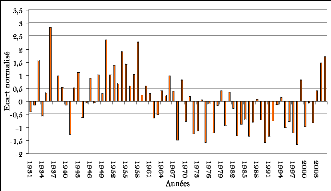
Figure 98. Ecart à la moyenne de la
pluviométrie mensuelle à Koungheul (1950-2005)
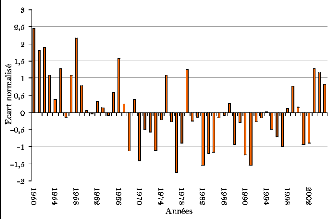
_
La longue sécheresse qui a commencé dans les
années 1970 a eu d'énormes impacts sur les ressources
forestières. Ces impacts sont de deux ordres. D'une part, on note une
forte diminution de la productivité végétale avec
notamment une forte mortalité des individus et une faible
régénération naturelle. D'autre part, les populations
compensent les déficits de production liés à la faible
pluviométrie par une pression additionnelle sur les ressources
forestière. Jusque là utilisées pour satisfaire des
besoins de base, les forêts deviennent une source de revenus et aiguise
ainsi les appétits mercantiles qui entraînent une
dégradation
Années
.
rapide. Aussi, est-il important de noter que la
réduction de la productivité agricole a poussé la plupart
des agriculteurs à étendre l'espace agricole en défrichant
plus de forêts (agriculture itinérante).
Le climat est par conséquent un facteur de dynamique
important dans les changements environnementaux. Il interagit avec d'autres
facteurs comme le facteur politique.
(b) VII.6.2. Le facteur politique
Depuis la période coloniale, les options politiques en
matière d'agriculture et de gestion des ressources naturelles ont connu
des évolutions pour répondre à plusieurs défis de
développement économique et social. La politique agricole du
Sénégal s'est longtemps articulée sur les performances de
la production arachidière. L'approche productiviste qui a fait du
Sénégal le premier pays producteur d'arachide a été
le soubassement de plusieurs options politiques qui vont de l'approche
socialiste, avec une forte implication de l'Etat qui a mis en place des
coopératives agricoles, à une libéralisation totale du
secteur en passant par la Nouvelle Politique Agricole.
Selon Gaye (2000), l'impact des politiques sur la production
agricole chez le paysan peut être étudié sur trois grandes
périodes.
- l'époque coloniale allant jusqu'en 1960,
- l'époque du Programme Agricole (PA) de 1960-80,
- la période des réformes structurelles avec
comme éléments marquants la Nouvelle Politique Agricole (NPA)
initiée en 1984-85, suivie du Programme d'Ajustement du Secteur Agricole
(PASA) à partir de 1995.
Pour compléter ce tableau, on note depuis les
années 2000 une forte approche programme (pour le maïs (Zea
mays), le bissap (Hibuscus sabdarifa), le manioc (Manihot
esculenta), le sésame (Sesamum indicum), etc.), avec
notamment l'acceptation du privé dans les transactions des produits
agricoles.
La stratégie des autorités coloniales a
été centrée sur la production de l'arachide (Arachis
hypogaea) pour l'exportation et l'importation de riz pour la consommation
(Mbodj, 1992, Gaye, 2000). L'arachide constituait l'élément
principal des revenus étatiques et du PIB. L'intervention publique
s'exerçait à travers les coopératives. Malgré leurs
multiples tares, elles sont devenues l'instrument principal de l'Etat
indépendant pour l'exécution des politiques agricoles.
La politique arachidière du Sénégal a
aussi consisté à un déplacement massif de
populations
pour occuper les `terres neuves' qu'il faut défricher en
peu de temps et exploiter pour la
production agricole. Le Saloum Oriental a
été ainsi le site par excellence de destination des
migrants de l'ethnie Sérère ou des agriculteurs de
la confrérie Mouride (Rocheteau, 1975 ; Garenne et Lombard, 1988).
La décision politique de coloniser les terres neuves
est consignée dans le deuxième Plan Quadriennal de
Développement Economique et social (1965-1969) qui a fait état du
besoin de déplacer une fraction de la population du bassin arachidier
dont la forte densité de population limiterait le développement,
et de la nécessité d'ouvrir à l'agriculture les
territoires inexploités du Saloum Oriental et du Sénégal
Oriental (Maymard, 1974). Il faut toutefois noter que le déplacement de
populations dans cette zone a débuté dans les années 1940
dans le secteur de Boullèle notamment, au gré de la mise en place
de la ligne de chemin de fer (Diouf, 2006). Deux options soutiennent cette
politique de colonisation :
- un mouvement migratoire ouolof, dit «spontané"
mais en réalité impulsé par des chefs
religieux de la confrérie Mouride;
- un mouvement migratoire organisé en faveur des
Sérères, populations connues selon le rapport pour leur bonne
qualité d'agriculteurs.
A l'époque plus de 200000 migrants devraient être
déplacés en 15 ans, mais le plan n'a pas été clair
sur l'organisation de cette migration. Ainsi, il y a eu beaucoup d'anarchie
dans l'installation des villages et très rapidement on a constaté
une forte dégradation des ressources naturelles forestières dans
la zone ainsi que les ressources pédologiques (Feller, 1977) et ceci
malgré un encadrement rapproché par les services techniques de
l'agriculture. L'Etat a auparavant réformé la législation
foncière (Loi 64-46 de 1964 portant Code Foncier) pour permettre aux
colons d'utiliser les `terres vacantes sans maître', qui sont
essentiellement dans les zones boisées, qu'il fallait donc
défricher. La progression n'était en définitive
limitée que par la présence de sols gravillonnaires ou
cuirassés, peu profonds ou l'existence de Forêts Classées ;
ces dernières seront sollicitées plus tard sous forme
d'empiètement ou de contrats de culture. Ainsi, l'arrivée de
migrants combinée à la croissance démographique naturelle
a entraîné une forte demande en terres de cultures qu'on ne
pouvait satisfaire qu'à travers de vastes opérations de
défrichement des formations forestières (voir chapitre 6,
Changement occupation du sol dans le Saloum Oriental).
L'intervention très poussée de l'Etat portait
aussi sur le contrôle du système de commercialisation, la
détermination des prix des principaux produits agricoles, l'octroi de
crédit pour les facteurs de production, et la réalisation des
infrastructures de base. Pour bien asseoir sa politique, l'Etat a eu pendant
cette période à instituer des structures d'encadrement et de
vulgarisation comme l'Office National de Coopération et d'Assistance au
Développement (ONCAD) et la Société de
Développement et de Vulgarisation Agricole
(SODEVA). Ces structures étaient des gouffres à
sous et les déficits enregistrés ont impacté tout le
système (Casswell, 1983).
Par une combinaison de la réalité du prix au
producteur sur le marché international (prix faible) et les charges pour
le paysan en termes de remboursement des prêts d'intrants, ce dernier se
retrouvait avec un revenu trop faible ne lui permettant pas d'investir.
Le fardeau des dettes accumulées par le système
coopératif, l'évolution défavorable des cours mondiaux de
l'arachide, ainsi que d'autres facteurs comme la hausse du prix du
pétrole, ont conduit l'Etat à entreprendre des réformes
structurelles. Ainsi, la Nouvelle Politique Agricole (NPA) centrée sur
l'autosuffisance alimentaire s'est fondée sur le principe de
désengagement de la puissance publique (Gray, 2002 ; Kaling, 2003). La
réduction des superficies cultivées en arachide et le
déclin du niveau d'équipement des exploitations agricoles sont
étroitement liés à la NPA caractérisée par
des restrictions en matière de crédit. Ces éléments
ont entraîné une forte vulnérabilité des populations
au moment déjà, où les prémisses de la
sécheresse des années 1970, commençaient à faire
ses effets sur la production agricole.
Au lieu de favoriser la durabilité de la production
agricole, la forte orientation mercantile de cette politique a plutôt
joué sur l'état d'esprit des populations locales tournée
vers le souci du gain. Les terres se sont alors rapidement
dégradées à cause d'un système de production qui
exclut l'arbre au champ. En plus le système d'un labour superficiel
(matériel agricole rudimentaire) réduit la fertilité des
sols en addition à l'exportation de la totalité de la plante
(gousse et foins), ce qui ne permet pas de compenser la baisse de
matière organique.
D'autres facteurs exogènes viennent renforcer cette
crise. On peut citer les crises du pétrole (1970-1972, 2005-2008) et
l'ajustement structurel qui ont fortement contribué à
l'accentuation de la pauvreté et de la vulnérabilité des
populations locales. En cherchant à répondre à ces crises,
les principales réponses sont souvent orientées vers l'extension
des terres agricoles pour répondre à la fois à la demande
de l'exportation et aux besoins de subsistance. La figure 99 montre l'extension
du bassin arachidier depuis 1900 (adapté du rapport Portères de
1952).
Figure 99. Evolution du bassin arachidier depuis
1900
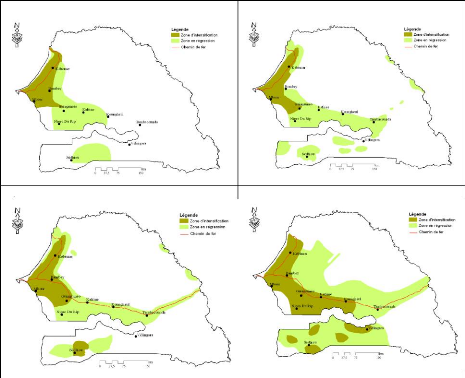
1900
1925
1910
1937
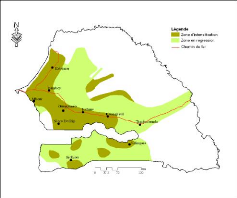
Depuis 1960
|
Ces cartes sont reprises de Mbow et al. (2008) et ont
adaptées des cartes Protères (1952).
|
Cette extension agricole s'est faite au détriment des
ressources forestières, de plus en plus confinées dans les
espaces de conservation comme les Forêts Classées et les Parcs
Nationaux.
La libéralisation totale du secteur pour des
agriculteurs non préparés à affronter la
réalité du marché a simplement déstructuré
la production arachidière. Récemment, l'Etat a initié des
programmes de production intensive du manioc, du maïs, du sésame,
du bissap qui apparaissent beaucoup plus comme des réactions
émotives qu'une base de politique agricole durable. Avec l'exposition
croissante aux avaries du climat et fluctuations imprévisibles des
produits du marché mondial, des initiatives ont été prises
comme le Plan REVA (Retour vers l'Agriculture) et la Grande Offensive Agricole
pour la Nourriture et l'Abondance (GOANA). Ces plans cherchent encore un
financement malgré les gros efforts de l'Etat d'impulser le processus
dans des délais très courts.
Au niveau local, on sent le développement de nouvelles
variétés comme le niébé et le sorgho fourragers, le
melon d'eau, qui parfois sont en association avec les cultures de mil permet
d'éviter l'envahissement des cultures par les adventices, tout en
améliorant les rendements par la fixation d'azote. Ces nouvelles
pratiques agricoles permettent d'assurer à la fois la production de
fourrage et de céréales.
Il apparaît ainsi que les populations rurales subissent
de façon prégnante les conséquences des options
politiques, facteurs importants de stress du fait de ses implications dans
l'accentuation de la pauvreté rurale. Ces facteurs politiques agissent
en même temps que des facteurs économiques.
(c) VII.6.3. Le facteur économique
Les facteurs économiques sont essentiellement
liés au prix des matières premières sur le marché
international. Le prix des céréales, du riz et du maïs
notamment a connu une réduction constante depuis 1950, au moment
où le prix des cultures commerciales augmentait légèrement
(figure 100-102). Il faut noter que si à un certain moment il
était rentable de s'adonner à une culture de rente pour acheter
des céréales importées et avoir toujours une marge de
bénéfice, cette réalité n'est plus vraie depuis de
nombreuses années du fait des faibles différences voire du
dépassement du prix des céréales sur les prix des cultures
de rente. Par exemple en 2004, le prix de l'arachide sur le marché
international était de 185 $/t, alors que celui du riz était de
217 $/t ; en 2008 le prix du riz avoisinait les 1000 $/t entraînant la
crise des prix dont les pays sahéliens comme le Sénégal
(grand importateur) ont profondément souffert.
Figure 100. Evolution du prix du maïs de
1950 à 2005, US $/t (FAO:
www.fao.org)
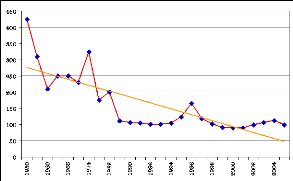
Figure 101. Evolution du prix du riz de 1950
à 2005, US $/t (FAO:
www.fao.org)
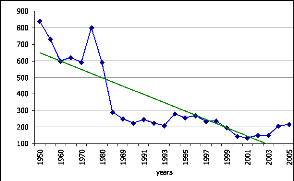


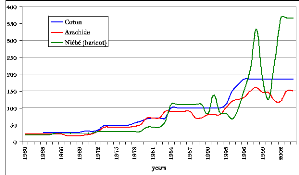

Figure 102. Evolution du prix des cultures
commerciales (coton, arachide, niébé) de 1950 à 2005, US
$/t (FAO :
www.fao.org )
La crise énergétique qui absorbe beaucoup de
ressources de l'Etat et la forte pression démographique accentuent cette
crise économique qui expose les populations à une pauvreté
accrue et donc à une forte vulnérabilité
socio-économique.
Pendant la période de gloire de l'arachide, il existait
un système bancaire d'investissement du secteur agricole. La Banque
Nationale de Développement du Sénégal (BNDS), assurait les
crédits nécessaires pour le fonctionnement du système.
Cette banque a fait faillite à l'image du secteur et les
possibilités de crédit des populations étaient devenues
fortement réduites. Le déficit du crédit, surtout du
microcrédit a inhibé pendant de longues années les
possibilités des populations agricoles du bassin arachidier qui
n'arrivent plus à sécuriser leur système de production
(Dièye et Guèye, 2003). Cette situation nouvelle rompt avec les
périodes de l'Etat providence pendant lequel l'encadrement agricole
était accompagné de plusieurs possibilités de
crédits sous forme d'intrants et d'équipements agricoles. La
crise était devenue tellement profonde en milieu rural que l'Etat en
2008 a mobilisé des fonds d'aide sociale (10 milliards de F CFA-XOF)
destinés à apporter de la nourriture de soudure à des
populations sérieusement affectées par une profonde crise
agricole.
(d) VII.6.4. Les réponses adaptatives
communautaires
Les formes d'adaptation sont nombreuses et variées. Il
est difficile, partant du cas du Saloum Oriental, de lier les réponses
adaptatives aux seuls changements climatiques. Les facteurs économiques
et politiques sont tout aussi déterminants dans le processus
d'appauvrissement des populations locales. Pour simplifier l'analyse, nous
présentons les formes d'adaptation selon une typologie qui
dépendra des options et des formes d'adaptation, mais aussi selon les
échelles de temps et d'espace de l'adaptation sur la zone
étudiée.
Les populations peuvent développer des
stratégies d'amélioration de la production agricole (adaptation
au champ) ou explorer d'autres activités permettant d'assurer leur
survie (adaptation hors champ). Les formes d'adaptations au champ sont
très variables.
* Les agriculteurs développent des
stratégies de reconstitution de la fertiité de terre à
travers les options suivantes (adaptation réactive et préventive)
:
- la fertilisation organique : utilisation de quelques
résidus agricoles, des bouses de
vache, en lieu et place des fertilisants minéraux qui sont
chers et peu accessibles ;
- la promotion de l'agroforesterie qui consiste à
promouvoir un nombre d'arbres plus
important dans le champ et autour du champ (haie vive) pour
d'une part atténuer
l'érosion, et d'autre part faire jouer à l'arbre
(surtout Faidherbia albida et Cordyla
pinnata) le rôle de pompe de nutriments profonds
que les racines des plantes cultivées ne peuvent atteindre ;
- les jachères de courte durée pratiquées
de plus en plus sont une réponse à la saturation foncière.
Il s'agit donc d'une réponse au manque de terre. Cependant, quand les
jachères sont courtes, elles ne permettent de reconstituer la
fertilité du sol. Dans de tel cas on parle de mal-adaptation.
* Les changements de techniques et d'options culturales
(adaptation préventive et planifiée) :
- alternance de cultures (alternance de plantes fixatrices
d'azote comme l'arachide à des plantes vivrières comme le mil) ;
cette approche réduit les pertes rapides de fertilité des sols et
est combinée à un choix de semences à cycle court ;
- diversification agricole, par l'introduction et la promotion
de nouveaux cultivars comme le bissap, le melon d'eau, le sésame, qui
peuvent générer des revenus et compenser les faibles performances
de la culture arachidière. Les cultures introduites sont adaptées
aux conditions de faible pluviométrie. Il faut aussi noter que dans la
zone, la culture vivrière du mil a pris une grande importance pour des
raisons de sécurité alimentaire des populations locales, mais
aussi parce que le mil est devenu un important `vivrier marchand' des centres
urbains en pleine expansion (en particulier Touba, Kaolack et Dakar) ;
- utilisation des bas fonds, moins inondés et l'abandon de
certains champs de plateaux aux sols pauvres ;
- promotion de la culture de contre saison, avec notamment le
maraîchage comme activité émergente, promue par des projets
de développement et des ONG ; le maraîchage occupe ainsi les
populations pendant la période sèche longue de 8 mois, ce qui
leur permet d'avoir des revenus additionnels non négligeables, tout en
améliorant leur niveau de vie ;
- le développement de l'élevage intensif
(stabulation).
A côté de ces stratégies au champ,
d'autres initiatives adaptives sont développées dans d'autres
secteurs de la vie économique et sociale afin d'atténuer les
impacts des contraintes liées à la variabilité climatique
et aux facteurs politico-économiques.
Les dynamiques récemment observées dans le
Saloum Oriental montrent une amélioration de la dynamique
organisationnelle en termes de solidarité de groupe. Les populations
mettent en place, avec l'appui des projets de développement ou des ONG
des comités locaux appelés Organisations Communautaires de Base
(OCB) qui peuvent être des Comités
Villageois de Développement (CVD), des Groupements de
Promotion Féminines (GPF), des Groupements d'Intérêt
Economiques (GIE), des Associations Sportives et Culturelles (ASC), des
organisations d'orientation socio-religieuses ou Dahiras, etc. Cette dynamique
organisationnelle est réconfortée par le contexte de
décentralisation qui donne plus de responsabilités aux
Collectivités Locales, responsables de la gestion des ressources
naturelles entre autres compétences transférées. La mise
en place de Plans Locaux de Développement (PLD) montre un souci de mise
en cohérence de la politique locale avec une identification des acteurs
et des interventions prioritaires susceptibles de promouvoir le
développement à la base. Cependant, les PLD écrits en
français ne sont pas accessibles à la majorité des
populations locales. Dans de nombreux cas, les PLD représentent plus une
liste de souhaits, de projets à faire par l'Etat, ou des bailleurs de
fonds extérieurs ou les ONG, qu'une stratégie cohérente
permettant d'identifier les problèmes de gestion durable des ressources
naturelles. Il faut retenir toutefois que les réseaux sociaux et les
organisations sociales constituent la première ligne de défense
des ménages pour faire face aux chocs et met en avant les femmes qui du
fait de certaines pesanteurs sociales se déplacent moins facilement que
les hommes.
Ce contexte nouveau est confronté cependant à
des barrières liées au financement des besoins prioritaires,
à la connaissance des textes de la décentralisation et aux
questions de démocratie interne (Ribot, 1998 ; Ellison, 2004). Il est
reconnu que la diversification des revenus en milieu rural et la mise en place
de services financiers desservant les populations rurales sont les domaines de
politiques et de dépenses publiques accusant des lacunes importantes. En
outre, la présence encore apparente de l'Etat dans la politique
d'exploitation forestière limiterait les capacités de
contrôle de la dégradation des ressources forestières par
les populations locales.
Une autre dynamique positive est l'engagement plus
significatif tourné vers l'alphabétisation et l'éducation
de base. La zone du Saloum Oriental comme pour plusieurs zones du
Sénégal tire profit du programme structurel de l'Etat pour
améliorer l'éducation de base et le niveau d'instruction des
populations. Les capacités d'innovation et l'utilisation efficiente des
ressources naturelles requièrent un bon renforcement des
capacités techniques locales. L'engouement et l'intéret
porté à l'éducation s'explique par une volonté de
plus en plus marquée de recherche d'un emploi salarié
sécurisé plutôt que la dépendance à une
production agricole contingente. Les interventions diverses et variées
des structures de développement fortifient cette volonté de
renforcement des capacités locales dans le cadre des approches
participatives qui privilégient les besoins des populations locales.
Par ailleurs, des actions de terrain menées pour
réduire l'occurrence des feux de brousse, améliorer la couverture
végétale par des activités de reboisement ou de mise en
défens sont autant de réponses à la dégradation de
l'environnement. A ce niveau, beaucoup d'efforts de communication sont
menés à travers les radios rurales ou communautaires, les
assemblées villageoises, etc. Ces actions sont parallèlement
conduites avec une valorisation commerciale de certains produits forestiers
ligneux et non ligneux (Mertz et al., 2009). Cette dernière
option est encore marquée par des tares qu'il faut corriger pour que le
souci du profit immédiat, favorisé par les opportunités
des marchés (louma ou marchés hebdomadaires), ne prenne le dessus
sur la conservation durable des ressources forestières.
L'une des formes d'adaptation non agricole les plus sensibles
en termes social et économique est la migration des populations. Une des
premières réponses à la dégradation des ressources
naturelles, base de survie des populations, est d'aller faire fortune ailleurs
(Mertz et al., 2009). Les études sur les migrations rurales
montrent qu'il s'agit d'une question complexe. De façon
générale, les ruraux se déplacent par étapes,
d'abord en direction des villes secondaires (Touba, Kaolack), vers les villes
côtières comme celles de la zone des Niayes, Saint-Louis et Mbour
(pour la pêche et le maraîchage) ; ensuite une bonne partie arrive
à Dakar pour se déployer dans le secteur dit `informel'. Puisque
l'agriculture n'est plus `compétitive' comparée au recyclage des
ordures, au petit commerce urbain, au travail de gardiennage, à
l'agriculture périurbaine, les populations rurales sont attirées
par les grandes villes et étoffent les grands effectifs de population,
surtout de celle de Dakar. La chaine de migration explique le regroupement des
familles au niveau des quartiers périphériques dans des
conditions écologiques et économiques très
précaires (Mbow et al., 2008).
Quelque soit les réponses apportées à la
dégradation de l'environnement, on peut faire une analyse temporelle ou
spatiale fine permettant de mieux caractériser les dynamiques en cours.
En effet, l'adaptation peut concerner des réponses individuelles sur de
petites surfaces (le lopin de terre, la vie du ménage), ou collectives
(reboisement, projets de groupes au sein des OCB, initiatives des CR). Ces
réponses peuvent aussi être menées comme des
réactions immédiates à un problème ponctuel (vendre
du bois pour inscrire son enfant à l'école, produire du charbon
pour payer les frais sanitaires d'un malade), ou se projeter dans le long terme
par l'implantation de vergers, ou la mise en défens forestier. Parfois,
le court terme peut constituer une contrainte sur le long terme ; par exemple
une forte exploitation de bois de Vèn (Pterocarpus erinaceus)
pour gérer des besoins immédiats peut nuire à la
conservation durable des ressources naturelles si l'ampleur de cette forme
d'exploitation est grande et si elle est non planifiée.
Chacun des éléments de réponse
développés dans le cas du Saloum Oriental constitue une
simplification et un rappel des grands facteurs qui jouent dans la
vulnérabilité et l'adaptation. L'idée est de comprendre
l'interaction des mécanismes fonctionnels qui régissent la
dynamique de l'environnement et les réponses dynamiques des populations
par rapport à sa dégradation. La figure 103 est une
synthèse des idées développées ici. Elle permet de
mieux comprendre comment se pose la question de l'adaptation dans le Saloum
Oriental.
Figure 103. L'adaptation est une réponse
à plusieurs stress dont les changements climatiques. Source : Mbow
et al. (2008)
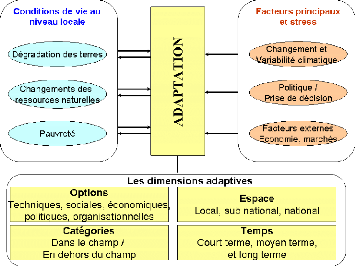
Dans la mesure où l'adaptation a été
posée comme une priorité dans le cadre des changements
climatiques pour l'Afrique, nous proposons une analyse croisée avec les
efforts d'atténuation véhiculés dans les pays en voie de
développement par la mise en ~uvre de projets MDP.
Section 16.07 VII.7. Adaptation versus
atténuation, quelles priorités pour le Sénégal
?
Le MDP est un des mécanismes de marché flexible
du Protocole de Kyoto qui permet aux pays Annexes I d'engager des projets de
réduction de GES dans les pays en voie de développement pour
pouvoir utiliser les crédits d'émission
générés (CER) pour atteindre leur objectifs de
réduction. Après la première année de la
première période d'engagement, il convient de questionner les
intentions du MDP et les enjeux pour l'Afrique. Selon plusieurs documents
officiels, l'objectif du MDP est double ; il permet d'une part d'aider les pays
industrialisés à atteindre leurs objectifs de réduction
des GES émis tout en permettant un transfert de technologies et un appui
économique au pays en voie de développement.
La première interprétation qu'on peut faire de
cette allégation est que le privilège du développement
industriel est réservé aux pays du Nord, et que les pays du Sud
doivent se contenter de nettoyer l'atmosphère.
La deuxième interprétation réside dans le
fondement éthique du MDP. Bien avant que les changements climatiques
aient été considérés comme défi majeur de
l'humanité, la question du développement de l'Afrique a
été posée à travers une amélioration du
système agro-sylvo pastoral. Les échos et engagements sur ce
problème d'amélioration de ces systèmes de production
n'ont pas été, à l'époque, aussi favorablement
accueillis qu'ils ne le sont aujourd'hui, parce que simplement la question est
devenue à la fois un problème éthique et une solution
potentielle à la réduction des GES. En d'autres termes, le
principe de la responsabilité commune et différenciée
engage, pour la recherche de solution, toute la communauté, mais les
bénéfices réels au bout du compte sont mal répartis
au sein de cette communauté.
En outre, le Sénégal, dans le souci de tirer le
meilleur parti de cette nouvelle économie verte a eu l'idée de
développer le secteur des biocarburants. Cette option pourrait
contrarier l'engagement pour la production agricole dans le but d'arriver
à une production alimentaire suffisante pour les besoins du pays. A cet
égard, les programmes comme la GOANA et le REVA, semblent plus
cohérents au contexte actuel du pays parce qu'orientés vers la
production agricole pour l'autosuffisance alimentaire. Dès lors que la
sécurité alimentaire est une préoccupation majeure dans le
contexte de pauvreté rurale marquée, l'esprit de promouvoir les
biocarburants devient peu attractif. Ainsi, au lieu d'orienter les grandes
ambitions agricoles vers le développement de Jatropha curcas,
il serait mieux de mettre
l'accent sur les cultures vivrières ou la maîtrise
de l'eau pour atténuer la crise du milieu agricole.
Exemple du prix du carbone
La valeur de la tonne de carbone sur le marché
international tourne dans le meilleur des cas autour de 20 US $ (prix d'aoflt
2008). Il faut d'abord noter que l'effort à consentir pour capturer 1
tonne de carbone correspond à un effort correspondant à la
production de 2 tonnes de biomasse. L'utilisation de cette biomasse pour la
satisfaction des besoins énergétiques donne une plus grande plus
value aux populations que sa vente sur le marché du carbone. Dès
lors, il faut penser aux caractères compétitifs de ces projets
MDP qui ne doivent pas être réduits à leur simple et
triviale dimension financière. Vu sous l'angle de cette tyrannie,
l'esprit des MDP - qui est d'abord une responsabilité sociale et un
devoir à l'endroit de l'environnement et non pas une loi aveugle du
marché- risque d'être altéré par le jeu de la
spéculation des cartels et autres lois ultra capitalistes.
Le développement de projets MDP est un processus
très complexe. Il sera difficilement à la portée des
populations locales pourtant essentiellement visées par cette mesure. Le
cycle d'un projet MDP nécessite beaucoup de temps qui dépasse
largement les délais de réponses requises par les populations
pour parer à l'urgence des questions de pauvreté. La figure 104
schématise le cycle d'un projet MDP.
Figure 104. Cycle d'un projet MDP
(synthèse des étapes)
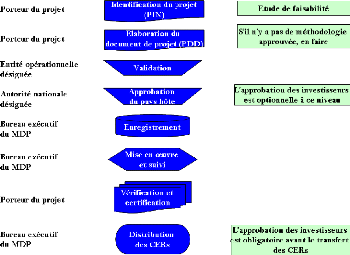
Les détails à considérer dans la
structuration de projets MDP sont donnés dans les développements
qui suivent.
Le Project Idea Note (PIN)
C'est l'idée de projet proposé par un porteur
sur la base d'une étude de faisabilité pour évaluer les
possibilités de développer un projet complet. Cette étude
de faisabilité prend en compte le potentiel de réduction de GES,
les coOts associés, l'additionalité et la possibilité que
le projet soit approuvé par le pays hôte, le Bureau
Exécutif du CDM et l'investisseur si déjà
identifié. En somme, le PIN donne une idée générale
d'un projet, mais en fournit au moins les informations indicatives sur
l'additionalité et le financement.
Le Project Design Document (PDD)
Il s'agit du projet détaillé avec un
scénario de base, une méthode de suivi, un calendrier et les
périodes d'accréditation, les impacts environnementaux et
sociaux. C'est ce document ainsi ficelé qui sera soumis aux
étapes administratives et de validation entre l'AND, le Bureau
Exécutif et l'Entité Opérationnelle avant sa mise en
~uvre. En gros un PDD comporte 5 grands points :
A. La description générale du projet
B. Le scénario de base et la méthode de suivi
C. La durée du projet et la période
d'accréditation choisie
D. Les impacts environnementaux et sociaux
E. L'avis ou les commentaires des partis prenants En plus il
faut produire 4 annexes :
Annexe 1. Les contacts de l'instituions (s) qui soumet le projet
Annexe 2. Les informations sur le financement public
Annexe 3. Les informations détaillées sur le
scénario de base Annexe 4. Le plan de suivi
Les méthodes approuvées
Tout projet MDP doit se référer à une
méthode approuvée pour la description de son scénario de
base, de la méthode de suivi, de l'additionalité etc. Au total,
10 méthodes sont approuvées par la Convention (AM-AR1-10, avec
une méthode consolidée ACM-1, à la date d'Aoflt 2008). En
développant un PDD, il faut choisir la méthode approuvée
qui du point des activités sont les plus proches de ce qu'on a choisi de
développer comme projet. La méthode approuvée est une
standardisation des caractéristiques des scénarios de base, du
calcul de l'additionalité et des réductions d'émission. La
méthode de suivi y est aussi harmonisée à travers les
paramètres et modes de suivi des activités.
La validation par l'Entité Opérationnelle
Désignée (EOD)
Il s'agit d'une évaluation indépendante du projet
qui vérifie si les conditions du MDP sont respectées au moins au
niveau des aspects suivants :
- le PDD répond-il aux exigences du MDP ?
- la méthodologie choisie est-elle applicable et
correctement utilisée ?
- le projet est-il additionnel ?
- les documents d'approbation de l'autorité nationale
désignée sont-ils obtenus ?
C'est après ces vérifications que le projet est
mis sur la page web de la convention dans la rubrique validation, pour des
commentaires ouverts pendant un mois avant validation finale. Pendant ce
processus le porteur du projet est régulièrement invité
à effectuer des clarifications et corrections en fonction des remarques
issues des évaluateurs. Cette validation peut être une relecture
du document de projet, mais elle peut comporter une visite de site. Le document
ne peut être enregistré par le Bureau Exécutif
qu'après avoir franchi ces étapes.
Mise en oeuvre du projet
La mise en ~uvre du projet doit être basée sur un
respect strict de la méthode de travail.
Une comptabilité
correcte du carbone séquestré est requise avec
l'élaboration régulière de
rapports de suivi qui doivent être transmis à
l'Entité Opérationnelle Désignée, laquelle doit
être différente de celle qui a évalué le projet. A
cette étape, la vérification consiste à voir si les
émissions certifiées déclarées dans le document de
projet sont atteintes lors de la mise en ~uvre. L'entité
opérationnelle qui fait l'évaluation de la mise en ~uvre envoie
les résultats du projet pour que des Certificats d'Emissions
Réduits (CERs) lui soient délivrés par le Bureau
Exécutif. Ces certificats d'émissions réduits sont vendus
sur le marché du carbone.
Les concepts du CDM foresterie
Les pays en voie de développement doivent
définir la notion de forêt avant de pouvoir mettre en ~uvre tout
projet de boisement ou reboisement dans le cadre du Protocole de Kyoto. Pour ce
faire, des critères nationaux doivent être définis mais en
conformité avec ceux définis par les Accords de Marrakech sur la
surface minimale, le taux de couverture et la hauteur des arbres. Dans ce
cadre, les critères pour définir une forêt sont les
suivantes :
- surface minimale entre 0,05-1,0 ha,
- taux de couverture des arbres 10-30 %,
- auteur des arbres 2-5 m à maturité.
Dans la plupart des pays, ces critères de
définition ne sont pas encore fixés. L'une des difficultés
majeures liées au choix de ces critères est son influence
à la fois sur les terres éligibles, sur la faisabilité des
projets, mais aussi sur la cohérence avec les objectifs du pays (Neeff
et al., 2006). Si on choisit des critères qui maximisent les
terres éligibles et la faisabilité, on peut introduire une
contrainte sur les objectifs de développement du pays, par exemple sur
ses activités de développement agricoles. Un choix optimal est
donc très difficile à faire.
Il faut noter que l'application de ces critères au
Sénégal convertit en forêt la plupart des formations de
savane, les jachères de longue durée, quelques vergers, etc. Ces
critères ont été un ajustement de la première
définition donnée par la FAO (surface minimale 1 ha, hauteur
minimum des arbres 7m, et taux de couverture > 20 %). L'abaissement des
seuils a entraîné une augmentation rapide et consistante (300
millions d'ha) des surfaces forestières du globe (Neeff et al.,
2006). Ces définitions ne précisent pas la différence
entre forêts naturelles et forêts artificielles.
Les activités éligibles sont le boisement
(afforestation) et le reboisement (reforestation). Le
boisement fait référence aux
conversions directes en zones forestières par activités humaines,
des terres qui n'étaient pas des forêts depuis au moins 50 ans,
par une plantation d'arbres, des semis direct, ou par promotion de la
régénération naturelle. Le
reboisement
est une conversion directe par l'homme de terres non
forestières (après déboisement), en terres
forestières par des plantations, des semis directs, ou par la promotion
de la régénération naturelle. Pour la première
période d'engagement, les terres éligibles au reboisement sont
celles qui n'ont pas été des forêts depuis le 31
décembre 1989.
Les contraintes d'un projet MDP foresterie
La rédaction de projets MDP bancables dans le domaine
de la foresterie est un exercice ardu et complexe. Il y a plusieurs
difficultés liées à l'élaboration de ces types de
projets. La première difficulté est leur manque de
flexibilité. On est même tenté de dire que les
mécanismes de flexibilités ne sont pas aussi flexibles qu'on le
dit et qu'on peut l'entendre. A ce sujet et pour qu'un projet puisse être
retenu, il faut tenir compte de quelques contraintes présentées
ci-dessus.
- Le projet doit satisfaire aux exigences et rigueur d'une
méthode élaborée ailleurs dans d'autres
écosystèmes notamment ; il existe à l'heure actuelle
près de 6 AR (Afforestation, Reforestation Methods) `applicables' au
niveau des écosystèmes tropicales, mais aucune n'a
été proposée pour répondre aux exigences des
savanes de l'Afrique de l'Ouest.
- Une bonne partie des données requises pour calculer
l'additionalité et les quantités de carbone certifié, ne
sont pas disponibles pour le moment (la productivité des
écosystèmes, la vitesse de croissance des espèces ; les
fuites liées à la satisfaction des besoins locaux, etc.).
- La difficulté pour caractériser la ligne de
référence (absence de données et de documentation).
- Les approches sont très techniques et il existe peu de
spécialistes de la question dans les pays d'Afrique.
- Les nombreuses étapes d'approbation avant la
finalisation d'un projet MDP : par exemple il faut élaborer un PIN,
soumettre le PIN à l'AND, qui l'examine et le soumet au Comité
National sur les changements climatiques. Si l'idée est acceptée,
un budget est sollicité, après rédaction, une
re-soumission est faite auprès de l'AND, qui le transmet au Bureau
Exécutif pour avis des experts. Si le projet est accepté parce
techniquement valable, il est finalement déposé auprès
d'un bailleur de fonds.
- L'approche est trop structurelle pour des populations
locales qui se retrouvent mieux dans la foresterie communautaire que dans des
systèmes qui rappellent étrangement la logique des plantations en
régie.
La complexité du cycle des projets MDP rend difficile
l'accès aux fonds alloués à cette activité et
explique la part congrue de l'Afrique dans les transactions de carbones sur le
marché international. Cependant le potentiel est là et ne
s'articule pas nécessairement aux activités éligibles de
la phase 2008-2012. Une étude menée par Skurtsch (2005) montre un
grand potentiel de la foresterie communautaire dans les pays en
développement pour les CDM. L'argumentaire de cet auteur est basé
sur les avantages financiers d'une telle approche compte tenu des coOts minimes
liés à leur mise en ~uvre comparé aux grands projets de
boisement/reboisement. Il suffit dans ce cas de bien former les populations
pour les travaux techniques de suivi et d'évaluation.
Même si les plantations en régie sur de grands
espaces peuvent jouer un rôle actif dans le puisage du carbone, il faut
remarquer qu'elles sont d'un faible apport pour l'atténuation directe de
la vulnérabilité des populations si ces dernières ne
peuvent y tirer profit dans le cadre d'une valorisation durable. Le plus
souvent les plantations en régie ont des fortunes différentes au
Sénégal. Elles sont en général sous l'apanage de la
DEFCCS sans grande intervention des populations. Par contre,
l'aménagement forestier communautaire déjà en cours mais
non éligible d'après Skurtsch (2005), peut pourtant donner des
résultats satisfaisants en termes de conservation des
écosystèmes et de développement local. Ces
résultats n'ont pas été articulés dès le
départ avec les changements climatiques, mais il faut noter que la
reconstitution des formations végétales à travers ces
projets contribue significativement à l'atténuation des GES mais
aussi et surtout à la réduction de la fragilité des
écosystèmes et à l'augmentation du potentiel de
séquestration de carbone au fil des années. Plusieurs organismes
et ONG appuient déjà avec succès de telles entreprises
à travers des microcrédits et une impulsion d'une dynamique
organisationnelle permettant aux populations locales de mieux s'organiser et de
promouvoir une démocratie interne au sein des structures
d'aménagement et de gestion. La foresterie communautaire s'articule bien
avec les réformes administratives et de gouvernance des deux
dernières décennies et portant sur la décentralisation. La
gestion des ressources naturelles est une compétence
transférée entre autres et donne droits de contrôle et de
gestion durable de ces ressources aux communautés de base. Si donc dans
le cadre des MDP on ne s'inscrit pas dans la tendance actuelle en
matière de GRN, on peut passer largement à côté des
objectifs initiaux.
Section 16.08 Conclusion
Dans ce chapitre nous avons voulu montrer les enjeux de
l'adaptation au Sénégal. Comme on peut s'en rendre compte, le
concept est complexe du fait des différents facteurs qui interviennent
à des échelles de temps et d'espaces, et qui entraînent des
impacts d'ampleur variée affectant en même temps les modes et le
niveau de vie des populations rurales. La situation de pauvreté
constatée dans le cas étudié apparaît en grande
partie comme une conséquence de plusieurs facteurs internes et externes
d'origine physique ou naturelle. La dégradation de l'environnement est
en effet liée à plusieurs facteurs biophysiques (dont le climat),
économiques et politiques. Les réponses apportées par les
populations sont variées et s'articulent essentiellement autour de
l'amélioration de la production agricole et forestière, mais
aussi autour de plusieurs stratégies génératrices de
revenus pour faire face aux défis de la pauvreté. On peut voir
à travers ces réponses adaptives ce qu'on peut qualifier de
`pratiques sans regret' qu'on aurait apporté avec ou pas de changement
climatique. Ces réponses visent une large panoplie de facteurs
influençant directement ou non la pauvreté rurale. Ainsi, compte
tenu de la forte vulnérabilité des populations pauvres, nous
avons questionné, en prenant l'exemple du Saloum Oriental,
l'opportunité des projets MDP qui constituent une option par rapport
à l'adaptation qui elle est une nécessité.
L'argumentaire développé dans ce chapitre
suggère un focus sur l'adaptation (agriculture, gestion des ressources
forestières pour la satisfaction des besoins des populations). Avec
l'aridification prévue du climat, il faut concentrer les efforts sur la
maîtrise de l'eau et la promotion de l'agriculture durable et d'un
élevage intensif (l'élevage extensif va beaucoup souffrir du
manque de fourrage si l'aridification se confirme). Ces options permettent de
répondre aux besoins des populations qui pourront en définitive
avoir un important impact sur l'atténuation. Si on réduit la
pauvreté et la dépendance aux ressources naturelles on pourra
atténuer du coup la dégradation des ressources
forestières.
Sur le plan des coOts de mise en ~uvre, la foresterie
communautaire est plus avantageuse, du fait de la facilité de
mobilisation de la main d'uvre, de la proximité et de la bonne
surveillance des zones d'intervention, de la réduction des processus de
dégradation (feux de brousse, coupe, empiètements). En plus des
avantages économiques, sociaux et écologiques, la foresterie
communautaire pourra si elle est retenue pour la deuxième période
d'engagement (2012-2017) donner plus d'enthousiasme et d'engagement aux
populations rurales.
Bibliographie
Adejuwon, J., Azar, C., Baethgen, W., Hope, C., Moss, R.,
Leary, N., Richels, R., Ypersele, v. J.-P., 2001. Overview of Impacts,
Adaptation, and Vulnerability to Climate Change. Chapiter 2, Climate Change
2001. IPCC, 30 p.
Adger, W.N., Brooks N., 2002. Does global environmental change
cause vulnerability to disaster?, CSERGE and Tyndall Centre for Climate Change
Research, University of East Anglia, Norwich.
Adger, W.N., Huq, S., Brown, K., Conwaya, D., Hulmea M. 2003.
Adaptation to climate change in the developing world. Progress in Development
Studies, 3 (3):179-195.
Burton, I., Challenger, B., Huq, S., Klein, R.J.T., Yohe G.,
2001. Adaptation to Climate Change in the Context of Sustainable Development
and Equity. Chapiter 18, Climate Change 2001. IPCC, 36 p.
Burton, I., Huq, S., Lim, B., Pilifosova, O., Schipper E.L.
2002. From Impacts Assessment to
Adaptation Priorities: the Shaping of Adaptation Policy. Climate
Policy (2):145-159. Casswell N., 1983. Autopsie de I'ONCAD : La politique
arachidière au Sénégal, 1966-1 980.
Leiden, 35 p.
CCNUCC, 1992. Convention Cadre des Nations Unies sur les
Changements Climatiques. In: N. Unies (Editor). Nations Unies, p. 31.
CCNUCC, 1997. Protocole de Kyoto. UNFCCC, 24 p.
Dièye, P.N, Guèye N. 2003. Les systèmes
agriculture-élevage au Sénégal : importance,
caractéristiques et contraintes. Improving Crop-Livestock Systems in
West and Central Africa, 26 p.
Diouf, A. 2006. Dynamique de l'occupation du sol et logiques
de gestion des ressources naturelles dans la Forêt Classée de
Kassas (Kaolack). Mémoire de DEA. Institut des Sciences de
l'Environnement, Faculté des Sciences et Techniques, Université
de Dakar; 119 p.
Ellison K., 2004. An Overview of Decentralization: Causes, Trends
and Challenges, ARD, Inc., USAID, 19p, Washington.
Feller Christian. 1977. Évolution des sols de
défriche récente dans la région des Terres
Neuves (Sénégal Oriental). Cahiers ORSTOM,
Série Biologie, XII (N° 3):199-211. Garenne, M., Lombard J., 1988.
Migrations, changements sociaux et développement.
Journées démographiques de l'ORSTOM, Paris, p.
27.
Gaye M. 2000. Région de Diourbel: politiques Nationales
affectant l'investissement chez les petits exploitants. Drylands Research,
Working Paper 12:58.
Gray J.K., 2002. The Groundnut Market in Senegal: Examination of
Price and Policy Changes. PhD, Virginia Polytechnic Institute and State
University.
Huq, S., Rahman, A., Konate, M., Sokona, Y., Reid H., 2003.
Main stream adaptation to climate change in Least Developed Countries (LDCS),
IIED, Climate Change Programme, London, 42 p.
Huq, S., Reid H., 2002. Report of Adaptation Day at COP 8, IIED,
Delhi, India.
IPCC, 2001. Climate change 2001: Impacts Adataptation and
vulnerability. A report of the Working Group II. Summury for Policy Makers. 18
p., IPCC, Geneva.
IPCC, 2003. Definitions and methodological options to
inventory emissions from direct human-induced degradation of forests and
vegetation of other types, IGES, Kanagawa.
IPCC (Editor) 2008. Bilan 2007 des changements climatiques.
Contribution des Groupes de travail I, II et III au quatrième Rapport
d'Evaluation du Groupe d'Experts Intergouvernemental sur l'Evolution du climat.
Intergouvernmental Panel on Climate Change, 103 p.
Janssen, M.A., Ostrom E. 2006. Resilience, vulnerability, and
adaptation: A cross-cutting theme of the International Human Dimensions
Programme on Global Environmental Change. Global Environmental Change,
Editorial (16):237-239.
Kaling C., 2003. La modernisation de l'agriculture dans la
région du Sine Saloum de 1960 à 1966. DEA, Université
Cheikh Anta Diop, Dakar.
L'Hôte, Y., Mahé, G., Somé, B., Triboulet,
J.P. . 2002. Analysis of a Sahelian annual rainfall index from 1896 to 2000 ;
the drought continues. Hydrological Sciences Journal, 47 (4).
Maymard J., 1974. Projet de colonisation des terres neuves. Le
point de la situation au 20 mars 1974. ORSTOM, Dakar, p. 8.
Mbodj Mohamed, 1992. La crise trentenaire de l'économie
arachidière. In: M.-C.D. (éd.) (Editor), Sénégal.
Trajectoires d'un État. CODESRIA, Dakar, pp. 95-135.
Mbow, C., Diop, A., Diaw, A.T., Niang C.I. 2008. Urban sprawl
development and flooding at Yeumbeul suburb (Dakar-Senegal). African Journal of
Environmental Science and Technology, 2 (4):075-088.
Mbow, C., Mertz, O., Diouf, A., Rasmussen, K., Reenberg A. 2008.
The history of environmental change and adaptation in eastern Saloum-Senegal.
Driving forces and perceptions. Global and Planetary Change, 64: 210-221.
Mertz, O., Mbow, C., Reenberg, A., Diouf A. 2009. Farmers'
perceptions of climate change
and agricultural adaptation strategies in rural Sahel.
Environmental Management ( 43):804-816.
Mortimore, M.J., Adams W.M. 2001. Farmer adaptation, change and
`crisis' in the Sahel. Global Environmental Change (11):49-57.
Neeff, T., Luepke, v., H., Schoene D., 2006. Choosing a forest
definition for the Clean Development Mechanism, FAO, Roma, 18 p.
Ogunseitan O.A. 2008. Clean Carbon Communism. African Journal of
Environmental Science and Technology., 2 (5):Editorial.
Olmos S., 2001. Vulnerability and Adaptation to Climate Change:
Concepts, Issues, Assessment Methods, Climate Change Knowledge Network.
Ribot J., 1998. Decentralization and participation in Sahelian
Forestry: Legal instruments of central political-administrative control.,
Harvard Center for Population and Development Studies. 47 p, Cambridge.
Ribot J., 2002. African Decentralization. Local Actors, Powers
and Accountability, United Nations Research Institute for Social Development.
Democracy, Governance and Human Rights Paper Number 8, Geneva.
Robledo, C., Forner C., 2005. Adapatation of forest ecosystems
and the forest sector to climate change, FAO-Forest and Climate Change Working
Paper 2.
Rocheteau G. 1975. Pionniers Mourides au
Sénégal: Colonisation des Terres Neuves et transformations d'une
économie paysanne. Cahiers ORSTOM, Série Sciences Humaines, XII
(1):19-53.
Skutsch M.M. 2005. Reducing carbon transaction costs in community
based forest management. Climat Policy (5):433-443.
Thomas, D., Osbahr, H., Twyman, C., Adger, N., Hewitson B.,
2005. Adaptations to climate change amongst natural resource-dependant
societies in the developing world: across the Southern African climate gradient
No. Technical Report 35, Tyndall Centre for Climate Change Research, London.
Winograd M., 2006. Concepts, cadres et méthodologies pour
évaluer la vulnérabilité et les stratégies
d'adaptation. C3D, ENDA, p. 32 p.
Article XVII. CONCLUSION GENERALE
Conclusion générale
Les changements climatiques constituent un des plus grands
défis du 21ième siècle qui a fini de mobiliser
la communauté internationale à travers une dynamique
institutionnelle et décisionnelle pour des solutions collectives
appropriées. Les causes des changements climatiques sont d'origine
humaine. C'est donc au niveau des activités humaines que l'essentiel des
solutions sont à rechercher. A travers les nombreux rapports
scientifiques, les dégagements de CO2 sont considérés
comme principal gaz à effet de serre ciblé par la CCNUCC à
travers les engagements de Kyoto. Pour les pays développés, les
principales initiatives ciblent les secteurs de l'énergie, l'industrie,
le transport, le bâtiment, et les changements de comportement individuel
et collectif. Un nouveau courant environnementaliste prend forme et s'affirme
de plus en plus à partir de l'indice de consommation de carbone
(carbon footprint) pour chaque activité et pour chaque
individu. En plus de ces aspects, l'Afrique est sollicitée pour une
contribution volontaire aux efforts de réduction, à travers
notamment le secteur forestier qui est une source de 17% sur les
dégagements de GES.
Le secteur forestier est un domaine d'intervention
relativement ouvert qui prend en compte plusieurs possibilités
contribuant toutes à la reconstitution des écosystèmes
dégradées. Le Protocole de Kyoto a sélectionné un
certain nombre d'activités éligibles au titre des MDP qui sont le
boisement et le reboisement. L'ouverture des activités éligibles
sur d'autres secteurs comme la mise en défens, le carbone du sol,
l'agroforesterie, etc., est mise en perspective dans les négociations de
la Convention et devraient être approuvés avec la COP de
Copenhague de Décembre 2009. Pour l'instant (Juillet 2009) toutes les
autres interventions qui ne consistent pas à planter directement des
arbres à travers des projets de protection forestière, de
foresterie communautaire, de régénération assistée,
de garnissage, etc., ne sont pas comptées parmi les activités de
MDP foresterie pendant la première période d'engagement
2008-2012. Cette limitation réduit les possibilités offertes par
les techniques forestières pour donner une pleine mesure à la
mise en ~uvre de la convention en Afrique.
Au Sénégal, la réponse à cet appel
aux MDP butte sur un déficit informationnel sur les
écosystèmes forestiers. Les données nécessaires
pour caractériser l'état actuel des stocks de carbone des types
de végétation ne sont connues que de façon partielle et
par des estimations indirectes (modèles). Les estimations faites
à ce jour utilisent pour la plupart des outils et approches
développés pour d'autres types de milieux. Un grand nombre
d'études sur la biomasse s'est focalisé sur les tarifs de cubage,
qui sont des matrices de conversion de mesures sur les arbres en bois fort
commercial. Cette approche ne vise pas la totalité de la
biomasse des arbres et reste donc peu utile pour les besoins
d'évaluation de la biomasse totale des arbres dans les
écosystèmes de savanes. Aussi, il était important
d'asseoir une base de travail appropriée par l'élaboration de
modèles allométriques fondés sur des mesures in
situ. Pour cela, 6 Forêts Classées situées dans les
zones soudaniennes et soudano-guinéennes, ont servi de support ; il
s'agit de Bala, Kantora, Mampaye, Ouli, Patako et Wélor. Nous proposons
3 modèles allométriques mettant en relation le DBH et la biomasse
sèche des arbres. Le DBH est un paramètre collecté pour
tout inventaire de la végétation. Ainsi, pour que les
modèles suggérés soient applicables à une bonne
partie des savanes d'Afrique de l'Ouest, nous avons utilisé ce
paramètre DBH pour les régressions allométriques. La
méthode retenue, évite de développer des outils trop
complexes du genre transformations logarithmiques ou exponentielles sur les
données brutes avant élaboration des modèles. Nous avons
donc voulu, dans ce choix méthodologique, éliminer toute source
d'erreur supplémentaire sur les données. Ainsi, les
modèles obtenus (régression quadratique, cubique et polynomiale
d'ordre 3) ont permis d'estimer les stocks de biomasse avec un r2
> 0,9 et les tests ANOVA (analyse de la variance) ont montré une
faible différence entre la moyenne de la série
expérimentale et celles issues des modèles.
Les approches d'estimation de biomasse proposées par la
FAO, comparées aux données expérimentales ont
montré de grandes différences de résultats. A cet
égard, les résidus sont très grands et entraînent
une surestimation ou une estimation de la biomasse selon les cas.
Les 3 modèles retenus après une série de
tests ont été utilisés pour estimer les stocks de carbone
des Forêts Classées. Les résultats obtenus montrent des
stocks de carbone très faibles pour les formations
dégradées de la Forêt Classée de Wélor (entre
3 et 7 tC/ha) alors que les Forêts Classées de Mampaye (20-50
tC/ha) et Kantora (40-47 tC/ha) ont les stocks les plus importants. Des cas
intermédiaires ont été notés au niveau de Ouli
(12-20 tC/ha), Bala (18-28 tC/ha), et Patako (13-33 tC/ha). L'essentiel de la
biomasse se situe au niveau des individus de gros diamètre (> 20 cm).
Cependant, la dynamique de séquestration du carbone est portée
par les petits individus en croissance. Ces résultats montrent que pour
réduire les émissions de carbone issue des savanes, il faut
atténuer la dégradation des gros sujets, mais pour promouvoir une
contribution des savanes à la séquestration du carbone, il faut
des logiques de préservation qui protègent les petits sujets.
Ainsi, le rôle des savanes dans le cadre de l'atténuation des
émissions de CO2 s'articule à des efforts d'aménagement et
de gestion qui permettent de maintenir un équilibre de la structure de
la végétation dans le sens d'une bonne représentation des
différentes classes de diamètre. Les stocks de carbone du sol, y
compris la biomasse racinaire, ceux de la litière ou de la biomasse
herbacée et des plantules
ne sont pas pris en compte. Ces résultats concernent donc
uniquement la biomasse ligneuse sur pied des individus de taille
supérieure ou égale à 5 cm de diamètre.
La caractérisation des stocks de carbone est une chose
importante et utile pour définir les lignes de base ou situation de
référence. Il est tout aussi essentiel de déterminer la
dynamique des formations ligneuses en termes d'accumulation de biomasse. La
caractérisation de la dynamique est basée sur l'analyse de
données de parcelles de suivi de la végétation ligneuse (3
parcelles de 1 ha chacune) suivies depuis 1993 dans la zone d'étude.
Cette analyse a permis de caractériser la production globale de chaque
parcelle mais aussi d'obtenir des informations sur la croissance des
espèces prises individuellement. Elle a également conduit
à saisir la dynamique de stockage du carbone qui, en l'absence de coupe,
est largement expliquée par la variation pluviométrique. De cette
dynamique, nous comprenons par ailleurs que l'essentiel des stocks de carbone
des parcelles de suivi est le fait de quelques espèces, surtout celles
à croissance rapide et qui peuvent atteindre de gros diamètres.
Les espèces les plus abondantes des 3 parcelles de suivi sont
Combretum glutinosum, Terminalia macroptera, Cassia sieberiana, Pterocarpus
erinaceus et Hexalobus monopetalus ; mais les espèces qui
contribuent le plus à la biomasse sont Pterocarpus erinaceus ;
Bombax costatum et Terminalia macroptera. Ainsi, la contribution
aux stocks de carbone n'est pas simplement une question d'effectifs, elle est
aussi liée aux possibilités de l'espèce à
croître vite et accumuler ainsi beaucoup de biomasse au fil du temps.
D'ailleurs, les espèces qui croient vite dans ces
écosystèmes sont Detarium senegalense et Bombax
costatum (espèces d'affinité guinéenne) ; elles sont
suivies de près par Anogeissus leiocarpus, Lannea microcarpa,
Pterocarpus erinaceus, et Terminalia macroptera. Les modifications de la
biodiversité n'ont pas été très grandes pendant la
période de suivi, mais les quantités de biomasse ont connu une
première phase de régression entre 1997 et 2003 du fait
probablement des faibles quantités de pluies puis on observe une
dynamique positive à partir de 2003 du fait certainement de la bonne
pluviométrie.
Pour mieux caractériser le rythme de croissance, des
analyses dendrochronologiques ont été faites. Pendant longtemps
cette méthode a été jugée très probante pour
analyser le développement des plantes mais aussi la relation entre la
vitesse de croissance des arbres et des facteurs environnementaux comme les
précipitations, la température, les phases de crue et
décrue des fleuves pour les espèces de galerie. De nombreuses
difficultés subsistent cependant pour les espèces tropicales qui
contrairement aux espèces des milieux tempérées
présentent des cernes pas toujours faciles à discriminer. Le
potentiel pour la datation des individus et la mise en relation avec des
paramètres comme la pluviométrie existe mais demande beaucoup
d'efforts pour que les résultats soient exploitables. Les
résultats obtenus
confirment la faible vitesse de croissance des espèces
de savane. On a noté pour la plupart des cas une réponse
irrégulière par rapport au signal pluviométrique ; l'effet
des facteurs de stress comme les feux, l'aridité, la compétition
dans la croissance. Ces facteurs font que la courbe de croissance des
espèces de savane étudiées est `atypique' par rapport
à ce que nous enseignent les théories écologiques. A cause
des nombreux facteurs de stress, les jeunes plants de savane ne croît pas
forcément plus vite que les plantes affranchies.
L'analyse de ces données montrent que les stocks de
carbone peuvent être assez importants quand la végétation
est bien conservée (Forêt Classée de Kantora et Mampaye),
lorsque notamment il existe dans de telles formations des espèces qui
ont la capacité de stocker de façon relativement rapide de la
biomasse. Mais si on projette la croissance des espèces les plus
performantes, on se rend compte qu'au bout de 30 ans la valeur marchande du
carbone stocké ne représente pas des montants importants (< 20
USD) devant les avantages qu'on peut en tirer avec l'utilisation des arbres
pour du bois énergie ou de service, activités plus rentables en
terme de revenus. Nous avons ainsi discuté de la question de la
`compétitivité' des MDP face aux autres alternatives
d'utilisation du bois en termes d'avantages comparatifs. Si par
conséquent le prix du carbone n'évolue pas dans le sens de
l'incitation aux MDP, les populations peuvent toujours préserver les
ressources forestières jusqu'à une certaine période pour
les exploiter et en tirer un revenu optimal. Ce risque pose le problème
du principe de permanence qui est un des piliers des MDP foresterie.
Dans la même lancée, des modèles de
croissance forestière ont été testés pour donner un
aperçu sur le futur en se basant sur un certain nombre de
scénarios réalistes. Ces scénarios prennent leur logique
sur les pratiques et facteurs de dynamique en cours comme les feux de brousse,
la jachère, le reboisement, l'exploitation forestière, le
pastoralisme et les défrichements agricoles. Il apparaît selon les
résultats de la simulation que les facteurs les plus
sévères de dégradation de la biomasse sont l'exploitation
forestière et les défrichements. Par contre, la
végétation naturelle se reconstitue assez rapidement en l'absence
de prélèvement. Le pâturage et les feux de savane
réduisent les stocks de carbone mais avec une ampleur plutôt
moindre. Les simulations ont aussi confirmé que les stocks de carbone du
sol sont plus importants mais leur dynamique est dictée par des
processus qui se déroulent au niveau de la végétation sur
pied. On constate néanmoins que dans les phases de reconstitution de la
végétation (reboisement, jachère de longue durée),
les stocks de carbone de la biomasse peuvent avant la phase d'équilibre
dépasser ceux du sol. Le carbone du sol se dégrade et se
reconstitue donc plus lentement que celui de la biomasse végétale
sur pied.
La perspective spatiale basée sur les changements
d'occupation des sols montre que les plus importantes dynamiques se
déroulent en milieu de terroir. D'une façon
générale les formations végétales connaissent une
certaine régression. Cette régression est parfois moins
perceptible au niveau des Forêts Classées, où par endroits
la végétation s'est reconstituée au moment où
d'autres faciès (notamment les formations denses de vallée ou les
savanes boisées) ont connu des reculs significatifs. Par contre, dans
les zones de terroir comme la zone Kaffrine-Koungheul, les observations
montrent une avancée soutenue des terres de culture au détriment
de la végétation naturelle. Des pertes engendrées peuvent
aller jusqu'à 4tC/ha/an.
La Haute Résolution (LANDSAT-TM et ETM) utilisée
pour cette analyse ne permet pas de voir les dynamiques plus régionales.
Ainsi, les données NDVI de MODIS à une résolution de 250 m
sur l'ensemble du Sénégal ont permis de suivre les modifications
dans la productivité végétale de 2000 à 2007 (8
ans). Les résultats montrent des endroits avec une dynamique positive
(quelques parties de Bala) et d'autres avec une forte régression
(Wélor, Ouli). A ce niveau, les facteurs climatiques interagissent avec
les facteurs humains dans les processus en question. Le principal enseignement
qu'on en tire est que la dynamique de la végétation n'est pas
linéaire, et présente une grande variabilité spatiale ;
elle procède par des allers-retours selon la force des facteurs en jeu
(climat et actions anthropiques).
Dans les zones de terroir, la dégradation notée
est un élément important à considérer dans
l'analyse de la vulnérabilité des populations locales face aux
changements environnementaux. En reprenant le cas du Saloum Oriental
(Kaffrine-Koungheul), l'analyse de la vulnérabilité et de
l'adaptation des populations locales est basée sur la recherche des
facteurs structurants majeurs qui semblent aller au-delà des impacts de
la variabilité climatique et inclut les facteurs politiques et
économiques à différentes échelles spatiales et
temporelles.
La variabilité climatique, les politiques agricoles et
forestières peu favorables aux populations rurales, des conditions de
marché fortement dominées par l'esprit capitaliste n'ont pas
favorisé une économie rurale stable. La pauvreté qui en
résulte a suscité de nombreuses réponses adaptives qui
consistent en gros à améliorer le système agricole (avec
des succès et échecs) mais aussi à trouver d'autres
sources de revenus comme le commerce, l'immigration, les changements
organisationnels, etc. L'étude de la vulnérabilité
nécessite des schémas analytiques complexes, de même que la
caractérisation des réponses apportées. On a montré
que certaines réponses étaient des sources de
vulnérabilité et qu'il pouvait y avoir antinomie entre les
réponses immédiates pour absorber des problèmes ponctuels
et les
exigences d'un développement durable inscrites dans le
long terme. L'exemple de l'exploitation forestière vers Koupentoum en
constitue une illustration claire. Ainsi, en voulant trouver des revenus
immédiats lors de la sécheresse de 1970 et 1980, plusieurs
paysans se sont reconvertis en exploitants forestiers. Au bout du compte le
fondement de leur survie a été dilapidé parce que les
activités n'ont pas été planifiées. Compte tenu de
la profonde pauvreté du milieu rural nous avons questionné
l'opportunité des MDP en tant que mesure d'atténuation, face
à l'adaptation qui pose un problème de survie immédiat et
futur pour les populations locales pauvres. Nous soutenons qu'il faut mettre
l'accent sur l'adaptation qui prend en compte d'ailleurs la bonne conservation
et la gestion durable des ressources forestières, non pas seulement pour
son carbone, mais surtout pour les services que les ressources
forestières peuvent apporter à l'équilibre
socio-économique des populations rurales. La priorité des pays
sous-développés comme le Sénégal étant
l'atténuation de la pauvreté rurale qui est une
nécessité et non une option que constituent les MDP.
ANNEXES
Annexe
Section 17.01 Annexe 1. Distribution des
espèces par classe de diamètre pour les trois parcelles de suivi
de la végétation ligneuse.
Parcelle de suivi 1
1993, P1
|
[5-10]
|
]10-15]
|
]15-20]
|
]20-25]
|
]25-30]
|
> 30
|
Total individus
|
|
Acacia macrostachya
|
3
|
0
|
0
|
0
|
0
|
0
|
3
|
|
Acacia sieberiana
|
1
|
0
|
0
|
1
|
0
|
0
|
2
|
|
Allophylus africanus
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Bombax costatum
|
13
|
7
|
4
|
1
|
2
|
0
|
27
|
|
Borassus aethiopum
|
0
|
0
|
0
|
1
|
|
1
|
2
|
|
Cassia sieberiana
|
234
|
65
|
18
|
2
|
0
|
0
|
319
|
|
Combretum glutinosum
|
163
|
19
|
0
|
0
|
0
|
0
|
182
|
|
Crossopteryx febrifuga
|
47
|
8
|
0
|
0
|
0
|
0
|
55
|
|
Dichrostachys cinerea
|
4
|
0
|
0
|
0
|
0
|
0
|
4
|
|
Grewia bicolor
|
2
|
0
|
0
|
0
|
0
|
0
|
2
|
|
Grewia flavescens
|
3
|
0
|
0
|
0
|
0
|
0
|
3
|
|
Guiera senegalensis
|
2
|
0
|
0
|
0
|
0
|
0
|
2
|
|
Hexalobus monopetalus
|
119
|
9
|
0
|
0
|
0
|
0
|
128
|
|
Hymenocardia acida
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Lannea acida
|
58
|
8
|
4
|
1
|
0
|
0
|
71
|
|
Lannea velutina
|
5
|
0
|
0
|
0
|
0
|
0
|
5
|
|
Lonchocarpus laxiflorus
|
3
|
1
|
0
|
0
|
0
|
0
|
4
|
|
Mitragyna inermis
|
24
|
1
|
0
|
0
|
0
|
0
|
25
|
|
Oncoba spinosa
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Parkia biglobosa
|
1
|
1
|
0
|
0
|
0
|
0
|
2
|
|
Piliostigma reticulatum
|
15
|
1
|
0
|
0
|
0
|
0
|
16
|
|
Piliostigma thonningii
|
23
|
6
|
1
|
0
|
0
|
0
|
30
|
|
Pterocarpus erinaceus
|
119
|
31
|
8
|
0
|
0
|
3
|
161
|
|
Sarcocephalus latifolius
|
3
|
0
|
0
|
0
|
0
|
0
|
3
|
|
Spondias mombin
|
0
|
2
|
1
|
0
|
0
|
0
|
3
|
|
Tamarindus indica
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Terminalia avicennioides
|
1
|
2
|
0
|
0
|
0
|
0
|
3
|
|
Terminalia laxiflora
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Terminalia macroptera
|
195
|
116
|
20
|
|
|
|
331
|
|
Xeroderris stuhlmannii
|
0
|
1
|
0
|
0
|
0
|
0
|
1
|
|
Ziziphus mucronata
|
7
|
0
|
0
|
0
|
0
|
0
|
7
|
|
Total
|
1050
|
278
|
56
|
6
|
2
|
4
|
1396
|
1997, P1
|
[5-10]
|
]10-15]
|
]15-20]
|
]20-25]
|
]25-30]
|
> 30
|
Total individus
|
|
Acacia macrostachya
|
4
|
0
|
0
|
0
|
0
|
0
|
4
|
|
Acacia sieberiana
|
1
|
0
|
0
|
0
|
1
|
0
|
2
|
|
Allophylus africanus
|
2
|
0
|
0
|
0
|
0
|
0
|
2
|
|
Bombax costatum
|
7
|
9
|
4
|
1
|
4
|
2
|
27
|
|
Borassus aethiopum
|
0
|
0
|
0
|
0
|
0
|
1
|
1
|
|
Cassia sieberiana
|
184
|
96
|
34
|
3
|
0
|
1
|
318
|
|
Combretum glutinosum
|
125
|
26
|
0
|
0
|
0
|
0
|
151
|
|
Crossopteryx febrifuga
|
29
|
5
|
1
|
0
|
0
|
0
|
35
|
|
Dichrostachys cinerea
|
5
|
0
|
0
|
0
|
0
|
0
|
5
|
|
Grewia bicolor
|
2
|
0
|
0
|
0
|
0
|
0
|
2
|
|
Grewia flavescens
|
6
|
0
|
0
|
0
|
0
|
0
|
6
|
|
Hexalobus monopetalus
|
108
|
20
|
2
|
0
|
0
|
0
|
130
|
|
Lannea acida
|
46
|
15
|
7
|
0
|
1
|
0
|
69
|
|
Lannea velutina
|
4
|
1
|
0
|
0
|
0
|
0
|
5
|
|
Lonchocarpus laxiflorus
|
3
|
1
|
0
|
0
|
0
|
0
|
4
|
|
Mitragyna inermis
|
31
|
2
|
0
|
0
|
0
|
0
|
33
|
|
Oncoba spinosa
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Parkia biglobosa
|
0
|
2
|
0
|
0
|
0
|
0
|
2
|
|
Piliostigma reticulatum
|
13
|
4
|
0
|
0
|
0
|
0
|
17
|
|
Piliostigma thonningii
|
17
|
8
|
0
|
0
|
0
|
0
|
25
|
|
Pterocarpus erinaceus
|
82
|
50
|
23
|
6
|
0
|
3
|
164
|
|
Sarcocephalus latifolius
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Spondias mombin
|
0
|
1
|
1
|
1
|
0
|
0
|
3
|
|
Tamarindus indica
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Terminalia avicennioides
|
0
|
1
|
0
|
0
|
0
|
0
|
1
|
|
Terminalia laxiflora
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Terminalia macroptera
|
117
|
122
|
54
|
2
|
0
|
0
|
295
|
|
Xeroderris stuhlmannii
|
0
|
1
|
0
|
0
|
0
|
0
|
1
|
|
Ziziphus mucronata
|
7
|
0
|
0
|
0
|
0
|
0
|
7
|
|
Total
|
797
|
364
|
126
|
13
|
6
|
7
|
1313
|
2003, P1
|
[5-10]
|
]10-15]
|
]15-20]
|
]20-25]
|
]25-30]
|
> 30
|
Total individus
|
|
Acacia macrostachya
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Acacia sieberiana
|
1
|
0
|
0
|
0
|
1
|
0
|
2
|
|
Allophylus cobbe
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Anogeissus leiocarpus
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Bombax costatum
|
1
|
6
|
3
|
3
|
1
|
6
|
20
|
|
Borassus aethiopum
|
0
|
0
|
0
|
0
|
0
|
1
|
1
|
|
Capparis decidua
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Cassia sieberiana
|
88
|
95
|
35
|
4
|
1
|
1
|
224
|
|
Combretum glutinosum
|
18
|
12
|
0
|
0
|
0
|
0
|
30
|
|
Crossopteryx febrifuga
|
21
|
4
|
0
|
1
|
0
|
0
|
26
|
|
Dichrostachys cinerea
|
2
|
0
|
0
|
0
|
0
|
0
|
2
|
|
Grewia flavescens
|
7
|
0
|
0
|
0
|
0
|
0
|
7
|
|
Hexalobus monopetalus
|
100
|
24
|
2
|
0
|
0
|
0
|
126
|
|
Lannea acida
|
33
|
16
|
9
|
1
|
1
|
0
|
60
|
|
Lannea microcarpa
|
0
|
1
|
0
|
0
|
0
|
0
|
1
|
|
Lannea velutina
|
3
|
0
|
0
|
0
|
0
|
0
|
3
|
|
Lonchocarpus laxiflorus
|
3
|
0
|
1
|
0
|
0
|
0
|
4
|
|
Mitragyna inermis
|
27
|
7
|
1
|
0
|
0
|
0
|
35
|
|
Oncoba spinosa
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Parkia biglobosa
|
0
|
2
|
0
|
0
|
0
|
0
|
2
|
|
Piliostigma reticulatum
|
13
|
5
|
0
|
0
|
0
|
0
|
18
|
|
Piliostigma thonningii
|
15
|
7
|
2
|
0
|
0
|
3
|
27
|
|
Pterocarpus erinaceus
|
51
|
54
|
32
|
18
|
4
|
0
|
159
|
|
Sarcocephalus latifolius
|
1
|
1
|
0
|
0
|
0
|
0
|
2
|
|
Spondias mombin
|
0
|
0
|
1
|
1
|
1
|
0
|
3
|
|
Stereospermum kunthianum
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Tamarindus indica
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Terminalia avicennioides
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Terminalia macroptera
|
47
|
84
|
79
|
23
|
0
|
|
0
|
233
|
|
Xeroderriis sthuhlmannii
|
0
|
1
|
0
|
0
|
0
|
|
0
|
1
|
|
Ziziphus mucronata
|
6
|
0
|
0
|
0
|
0
|
|
0
|
6
|
|
Total
|
441
|
319
|
165
|
51
|
9
|
|
11
|
996
|
|
2005, P1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Total
|
|
[5-10]
|
]10-15]
|
]15-20]
|
]20-25]
|
]25-30]
|
>
|
30
|
individus
|
|
Acacia sieberiana
|
1
|
0
|
0
|
0
|
1
|
0
|
2
|
|
Allophylus cobbe
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Anogeissus leiocarpus
|
2
|
0
|
0
|
0
|
0
|
0
|
2
|
|
Bombax costatum
|
2
|
6
|
3
|
3
|
2
|
12
|
28
|
|
Borassus aethiopum
|
0
|
0
|
0
|
0
|
0
|
2
|
2
|
|
Capparis decidua
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Cassia sieberiana
|
96
|
108
|
38
|
6
|
1
|
2
|
251
|
|
Combretum glutinosum
|
27
|
11
|
1
|
0
|
0
|
0
|
39
|
|
Crossopteryx febrifuga
|
41
|
5
|
0
|
0
|
0
|
0
|
46
|
|
Dichrostachys cinerea
|
2
|
0
|
0
|
0
|
0
|
0
|
2
|
|
Grewia flavescens
|
10
|
0
|
0
|
0
|
0
|
0
|
10
|
|
Hexalobus monopetalus
|
144
|
29
|
3
|
0
|
0
|
0
|
176
|
|
Hymenocardia acida
|
2
|
0
|
0
|
0
|
0
|
0
|
2
|
|
Lannea acida
|
39
|
18
|
11
|
2
|
1
|
0
|
71
|
|
Lannea microcarpa
|
1
|
0
|
1
|
0
|
0
|
0
|
2
|
|
Lannea velutina
|
2
|
1
|
0
|
0
|
0
|
0
|
3
|
|
Lonchocarpus laxiflorus
|
4
|
0
|
1
|
0
|
0
|
0
|
5
|
|
Mitragyna inermis
|
51
|
12
|
1
|
0
|
0
|
0
|
64
|
|
Oncoba spinosa
|
3
|
0
|
0
|
0
|
0
|
0
|
3
|
|
Parkia biglobosa
|
0
|
2
|
0
|
0
|
0
|
0
|
2
|
|
Piliostigma reticulatum
|
16
|
8
|
0
|
0
|
0
|
0
|
24
|
|
Piliostigma thonningii
|
35
|
10
|
2
|
0
|
0
|
0
|
47
|
|
Pterocarpus erinaceus
|
74
|
60
|
31
|
21
|
8
|
8
|
202
|
|
Sarcocephalus latifolius
|
2
|
0
|
1
|
0
|
0
|
0
|
3
|
|
Spondias mombin
|
0
|
0
|
0
|
2
|
1
|
0
|
3
|
|
Stereospermum kunthianum
|
2
|
0
|
0
|
0
|
0
|
0
|
2
|
|
Tamarindus indica
|
3
|
0
|
0
|
0
|
0
|
0
|
3
|
|
Terminalia avicennioides
|
0
|
1
|
0
|
0
|
0
|
0
|
1
|
|
Terminalia macroptera
|
45
|
69
|
90
|
35
|
1
|
0
|
240
|
|
Xeroderriis sthuhlmannii
|
0
|
0
|
1
|
0
|
0
|
0
|
1
|
|
Ziziphus mucronata
|
6
|
0
|
0
|
0
|
0
|
0
|
6
|
|
Total
|
612
|
340
|
184
|
69
|
15
|
24
|
1244
|
Parcelle de suivi 2
1993, P2
|
[5-10]
|
]10-15]
|
]15-20]
|
]20-25]
|
]25-30]
|
> 30
|
Total individus
|
|
Bombax costatum
|
0
|
0
|
2
|
4
|
2
|
7
|
15
|
|
Combretum crotonoides
|
8
|
0
|
0
|
0
|
0
|
0
|
8
|
|
Combretum glutinosum
|
137
|
36
|
9
|
0
|
0
|
0
|
182
|
|
Cordyla pinnata
|
5
|
5
|
3
|
0
|
2
|
1
|
16
|
|
Crossopteryx febrifuga
|
37
|
14
|
4
|
1
|
0
|
0
|
56
|
|
Daniellia oliveri
|
11
|
11
|
3
|
0
|
0
|
0
|
25
|
|
Detarium microcarpum
|
8
|
11
|
2
|
0
|
0
|
0
|
21
|
|
Grewia flavescens
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Hexalobus monopetalus
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Lannea acida
|
5
|
5
|
5
|
1
|
2
|
0
|
18
|
|
Lannea microcarpa
|
1
|
0
|
2
|
1
|
0
|
0
|
4
|
|
Lannea velutina
|
12
|
3
|
4
|
2
|
0
|
0
|
21
|
|
Lonchocarpus laxiflorus
|
2
|
0
|
0
|
0
|
0
|
0
|
2
|
|
Maytenus senegalensis
|
3
|
0
|
0
|
0
|
0
|
0
|
3
|
|
Pericopsis laxiflora
|
4
|
14
|
8
|
5
|
0
|
0
|
31
|
|
Pterocarpus erinaceus
|
4
|
1
|
6
|
3
|
5
|
8
|
27
|
|
Strychnos spinosa
|
38
|
9
|
1
|
0
|
0
|
0
|
48
|
|
Terminalia avicennioides
|
36
|
7
|
1
|
0
|
0
|
0
|
44
|
|
Trichilia emetica
|
4
|
2
|
0
|
0
|
0
|
0
|
6
|
|
Vitex madiensis
|
5
|
2
|
0
|
0
|
0
|
0
|
7
|
|
Xeroderris stuhlmannii
|
8
|
7
|
8
|
2
|
1
|
0
|
26
|
|
Total
|
330
|
127
|
58
|
19
|
12
|
16
|
562
|
1997, P2
|
[5-10]
|
]10-15]
|
]15-20] ]20-25]
|
]25-30]
|
> 30
|
Total individus
|
|
Bombax costatum
|
0
|
0
|
2 3
|
2
|
10
|
17
|
|
Combretum crotonoides
|
7
|
40
|
11 0
|
0
|
0
|
58
|
|
Combretum glutinosum
|
110
|
0
|
0 0
|
0
|
0
|
110
|
|
Cordyla pinnata
|
5
|
3
|
4 2
|
1
|
2
|
17
|
|
Crossopteryx febrifuga
|
31
|
15
|
9 1
|
0
|
0
|
56
|
|
Daniellia oliveri
|
8
|
5
|
4 0
|
0
|
0
|
17
|
|
Detarium microcarpum
|
8
|
10
|
3 0
|
0
|
0
|
21
|
|
Grewia flavescens
|
2
|
0
|
0 0
|
0
|
0
|
2
|
|
Hexalobus monopetalus
|
1
|
0
|
0 0
|
0
|
0
|
1
|
|
Lannea acida
|
6
|
3
|
4 2
|
3
|
1
|
19
|
|
Lannea microcarpa
|
2
|
0
|
1 0
|
1
|
2
|
6
|
|
Lannea velutina
|
10
|
4
|
2 3
|
0
|
0
|
19
|
|
Lonchocarpus laxiflorus
|
1
|
0
|
0 0
|
0
|
0
|
1
|
|
Maytenus senegalensis
|
3
|
0
|
0 0
|
0
|
0
|
3
|
|
Pericopsis laxiflora
|
3
|
14
|
7 6
|
0
|
0
|
30
|
|
Pterocarpus erinaceus
|
3
|
1
|
2 7
|
3
|
11
|
27
|
|
Strychnos spinosa
|
38
|
10
|
1 0
|
0
|
0
|
49
|
|
Terminalia avicennioides
|
41
|
5
|
1 1
|
0
|
0
|
48
|
|
Terminalia macroptera
|
1
|
0
|
0 0
|
0
|
0
|
1
|
|
Trichilia emetica
|
3
|
2
|
0 0
|
0
|
0
|
5
|
|
Vitex madiensis
|
3
|
3
|
0 0
|
0
|
0
|
6
|
|
Xeroderris stuhlmannii
|
7
|
4
|
10 3
|
1
|
0
|
25
|
|
Total
|
293
|
119
|
61 28
|
11
|
26
|
538
|
2003, P2
|
|
|
|
|
|
|
|
Total
|
|
[5-10]
|
]10-15]
|
]15-20]
|
]20-25]
|
]25-30]
|
>
|
30
|
individus
|
|
Bombax costatum
|
0
|
0
|
1
|
3
|
2
|
|
7
|
13
|
|
Combretum crotonoides
|
26
|
4
|
0
|
0
|
0
|
|
0
|
30
|
|
Combretum glutinosum
|
66
|
36
|
12
|
0
|
0
|
|
0
|
114
|
|
Cordyla pinnnata
|
3
|
2
|
5
|
3
|
0
|
|
3
|
16
|
|
Crossopteryx febrifuga
|
25
|
11
|
8
|
1
|
0
|
|
0
|
45
|
|
Daniellia oliveri
|
4
|
4
|
3
|
0
|
0
|
|
0
|
11
|
|
Detarium microcarpum
|
2
|
6
|
5
|
0
|
0
|
|
0
|
13
|
|
Grewia flavescens
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Hexalobus monopetalus
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Lannea acida
|
3
|
6
|
3
|
1
|
3
|
|
0
|
16
|
|
Lannea microcarpa
|
1
|
1
|
2
|
1
|
1
|
|
0
|
6
|
|
Lannea velutina
|
4
|
2
|
2
|
2
|
0
|
|
0
|
10
|
|
Lonchocarpus laxiflorus
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Maytenus senegalensis
|
3
|
0
|
0
|
0
|
0
|
|
0
|
3
|
|
Pericopsis laxiflora
|
4
|
6
|
8
|
2
|
1
|
|
0
|
21
|
|
Pterocarpus erinaceus
|
1
|
2
|
1
|
6
|
3
|
|
12
|
25
|
|
Strychnos spinosa
|
29
|
14
|
1
|
0
|
0
|
|
0
|
44
|
|
Terminalia avicennioides
|
34
|
5
|
1
|
0
|
0
|
|
0
|
40
|
|
Terminalia macroptera
|
1
|
1
|
0
|
0
|
0
|
|
0
|
2
|
|
Trichilia emetica
|
3
|
2
|
0
|
0
|
0
|
|
0
|
5
|
|
Vitex madiensis
|
2
|
2
|
0
|
0
|
0
|
|
0
|
4
|
|
Xeroderris stuhlmannii
|
5
|
5
|
6
|
5
|
0
|
|
1
|
22
|
|
Total
|
219
|
109
|
58
|
24
|
10
|
|
23
|
443
|
2005, P2
|
[5-10]
|
]10-15]
|
]15-20]
|
]20-25]
|
]25-30]
|
> 30
|
Total individus
|
|
Annona senegalensis
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Bombax costatum
|
2
|
0
|
1
|
2
|
3
|
7
|
15
|
|
Combretum crotonoides
|
45
|
5
|
0
|
0
|
0
|
0
|
50
|
|
Combretum glutinosum
|
103
|
42
|
12
|
0
|
0
|
0
|
157
|
|
Cordyla pinnnata
|
9
|
1
|
6
|
3
|
0
|
3
|
22
|
|
Crossopteryx febrifuga
|
34
|
12
|
8
|
2
|
0
|
0
|
56
|
|
Daniellia oliveri
|
4
|
4
|
3
|
0
|
0
|
0
|
11
|
|
Detarium microcarpum
|
7
|
5
|
6
|
0
|
0
|
0
|
18
|
|
Grewia flavescens
|
1
|
0
|
0
|
0
|
0
|
0
|
1
|
|
Hexalobus monopetalus
|
3
|
0
|
0
|
0
|
0
|
0
|
3
|
|
Lannea acida
|
6
|
6
|
3
|
1
|
3
|
0
|
19
|
|
Lannea microcarpa
|
1
|
1
|
2
|
1
|
1
|
0
|
6
|
|
Lannea velutina
|
7
|
2
|
2
|
2
|
0
|
0
|
13
|
|
Lonchocarpus laxiflorus
|
1
|
1
|
0
|
0
|
0
|
0
|
2
|
|
Maytenus senegalensis
|
4
|
0
|
0
|
0
|
0
|
0
|
4
|
|
Pericopsis laxiflora
|
5
|
5
|
8
|
2
|
1
|
0
|
21
|
|
Pterocarpus erinaceus
|
1
|
1
|
2
|
5
|
4
|
12
|
25
|
|
Strychnos spinosa
|
64
|
15
|
2
|
0
|
0
|
0
|
81
|
|
Terminalia avicennioides
|
48
|
5
|
1
|
0
|
0
|
0
|
54
|
|
Terminalia macroptera
|
0
|
1
|
1
|
0
|
0
|
0
|
2
|
|
Trichilia emetica
|
6
|
2
|
0
|
0
|
0
|
0
|
8
|
|
Vitex madiensis
|
10
|
2
|
0
|
0
|
0
|
0
|
12
|
|
Xeroderris stuhlmannii
|
11
|
6
|
5
|
6
|
0
|
1
|
29
|
|
Total
|
373
|
116
|
62
|
24
|
12
|
23
|
610
|
|
1993, P3
|
[5-10]
|
Parcelle de suivi 3
]10-15] ]15-20] ]20-25]
|
]25-30]
|
> 30
|
Total individus
|
|
Allophylus africanus
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Anogeissus leiocarpus
|
3
|
3
|
0
|
4
|
2
|
|
10
|
22
|
|
Anthostema senegalense
|
2
|
1
|
0
|
1
|
0
|
|
0
|
4
|
|
Bombax costatum
|
3
|
0
|
0
|
0
|
0
|
|
1
|
4
|
|
Cassia sieberiana
|
0
|
1
|
1
|
0
|
0
|
|
0
|
2
|
|
Cissus populnea
|
4
|
0
|
0
|
0
|
0
|
|
0
|
4
|
|
Combretum collinum
|
9
|
9
|
1
|
0
|
0
|
|
0
|
19
|
|
Combretum glutinosum
|
5
|
0
|
0
|
0
|
0
|
|
0
|
5
|
|
Combretum micranthum
|
11
|
1
|
0
|
0
|
0
|
|
0
|
12
|
|
Combretum nigricans
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Combretum tomentosum
|
1
|
0
|
1
|
0
|
0
|
|
0
|
2
|
|
Cordyla pinnnata
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Crossopteryx febrifuga
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Detarium senegalense
|
7
|
0
|
0
|
2
|
0
|
|
3
|
12
|
|
Diospyros mespiliformis
|
2
|
2
|
0
|
1
|
0
|
|
2
|
7
|
|
Erythrophleum suaveolens
|
0
|
0
|
0
|
0
|
0
|
|
1
|
1
|
|
Feretia apodanthera
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Gardenia erubescens
|
2
|
0
|
0
|
0
|
0
|
|
0
|
2
|
|
Grewia bicolor
|
1
|
1
|
0
|
0
|
0
|
|
0
|
2
|
|
Hexalobus monopetalus
|
53
|
38
|
7
|
0
|
1
|
|
1
|
100
|
|
Lannea microcarpa
|
2
|
1
|
0
|
2
|
1
|
|
3
|
9
|
|
Lonchocarpus laxiflorus
|
0
|
0
|
1
|
0
|
0
|
|
0
|
1
|
|
Parkia biglobosa
|
0
|
0
|
0
|
0
|
0
|
|
5
|
5
|
|
Piliostigma thonningii
|
7
|
4
|
15
|
11
|
4
|
|
7
|
48
|
|
Pterocarpus erinaceus
|
25
|
26
|
4
|
12
|
5
|
|
27
|
99
|
|
Saba senegalensis
|
4
|
3
|
0
|
0
|
0
|
|
1
|
8
|
|
Sarcocephalus latifolius
|
18
|
17
|
2
|
2
|
1
|
|
0
|
40
|
|
Tamarindus indica
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Trichilia emetica
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Terminalia macroptera
|
0
|
0
|
0
|
0
|
0
|
|
1
|
1
|
|
Vitex madiensis
|
4
|
0
|
0
|
0
|
0
|
|
2
|
6
|
|
Total
|
164
|
107
|
32
|
35
|
14
|
|
64
|
416
|
|
1997, P3
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Total
|
|
[5-10]
|
]10-15]
|
]15-20]
|
]20-25]
|
]25-30]
|
>
|
30
|
individus
|
|
Allophylus africanus
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Anogeissus leiocarpus
|
4
|
2
|
1
|
2
|
3
|
|
10
|
22
|
|
Anthostema senegalense
|
0
|
3
|
0
|
0
|
1
|
|
0
|
4
|
|
Bombax costatum
|
4
|
0
|
0
|
0
|
0
|
|
1
|
5
|
|
Cassia sieberiana
|
0
|
1
|
1
|
0
|
0
|
|
0
|
2
|
|
Cissus populnea
|
5
|
0
|
0
|
0
|
0
|
|
0
|
5
|
|
Combretum collinum
|
12
|
9
|
3
|
0
|
0
|
|
0
|
24
|
|
Combretum glutinosum
|
3
|
0
|
0
|
0
|
0
|
|
0
|
3
|
|
Combretum micranthum
|
27
|
1
|
0
|
0
|
0
|
|
0
|
28
|
|
Combretum nigricans
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Combretum tomentosum
|
3
|
1
|
0
|
0
|
0
|
|
0
|
4
|
|
Cordyla pinnnata
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Crossopteryx febrifuga
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Detarium senegalense
|
8
|
3
|
0
|
0
|
1
|
|
5
|
17
|
|
Diospyros mespiliformis
|
2
|
0
|
2
|
0
|
0
|
|
1
|
5
|
|
Erythrophleum suaveolens
|
0
|
0
|
0
|
0
|
0
|
|
1
|
1
|
|
Feretia apodanthera
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Gardenia erubescens
|
3
|
0
|
0
|
0
|
0
|
|
0
|
3
|
|
Grewia bicolor
|
2
|
1
|
0
|
0
|
0
|
|
0
|
3
|
|
Hexalobus monopetalus
|
53
|
46
|
7
|
1
|
0
|
|
1
|
108
|
|
Lannea microcarpa
|
2
|
0
|
0
|
0
|
3
|
|
3
|
8
|
|
Lonchocarpus laxiflorus
|
0
|
0
|
1
|
0
|
0
|
|
0
|
1
|
|
Parkia biglobosa
|
0
|
0
|
0
|
0
|
0
|
|
5
|
5
|
|
Piliostigma thonningii
|
8
|
1
|
9
|
14
|
4
|
|
9
|
45
|
|
Pterocarpus erinaceus
|
26
|
19
|
8
|
9
|
5
|
|
30
|
97
|
|
Saba senegalensis
|
11
|
4
|
0
|
0
|
0
|
|
1
|
16
|
|
Sarcocephalus latifolius
|
17
|
15
|
3
|
2
|
2
|
|
0
|
39
|
|
Tamarindus indica
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Trichilia emetica
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Terminalia macroptera
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Vitex madiensis
|
2
|
0
|
0
|
0
|
0
|
|
0
|
2
|
|
Total
|
197
|
106
|
35
|
28
|
19
|
|
67
|
452
|
|
2003, P3
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Total
|
|
[5-10]
|
]10-15]
|
]15-20] ]20-25]
|
|
]25-30]
|
>
|
30
|
individus
|
|
Allophyllus cobe
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Anogeissus leiocarpus
|
3
|
1
|
1
|
0
|
4
|
|
10
|
19
|
|
Anthostema senegalense
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Bombax costatum
|
2
|
0
|
0
|
0
|
0
|
|
1
|
3
|
|
Cassia sieberiana
|
0
|
1
|
1
|
0
|
0
|
|
0
|
2
|
|
Cissus populnea
|
0
|
1
|
0
|
0
|
0
|
|
0
|
1
|
|
Combretum collinum
|
3
|
5
|
3
|
0
|
0
|
|
0
|
11
|
|
Combretum glutinosum
|
1
|
3
|
0
|
0
|
0
|
|
0
|
4
|
|
Combretum micranthum
|
30
|
2
|
0
|
0
|
0
|
|
0
|
32
|
|
Combretum nigricans
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Combretum tomentosum
|
0
|
1
|
0
|
0
|
0
|
|
0
|
1
|
|
Cordyla pinnnata
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Crossopteryx febrifuga
|
0
|
1
|
0
|
0
|
0
|
|
0
|
1
|
|
Detarium senegalense
|
5
|
1
|
4
|
0
|
2
|
|
7
|
19
|
|
Diopyros mespiliformis
|
1
|
1
|
2
|
0
|
0
|
|
0
|
4
|
|
Erythrophleum suaveolens
|
0
|
0
|
0
|
0
|
0
|
|
1
|
1
|
|
Feretia apodanthera
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Gardenia ternifolia / triacantha
|
2
|
0
|
0
|
0
|
0
|
|
0
|
2
|
|
Grewia flavescens
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Hexalobus monopetalus
|
39
|
48
|
8
|
3
|
0
|
|
1
|
99
|
|
Lannea microcarpa
|
0
|
0
|
0
|
0
|
3
|
|
2
|
5
|
|
Lonchocarpus laxiflorus
|
0
|
0
|
1
|
0
|
0
|
|
0
|
1
|
|
Parkia biglobosa
|
0
|
0
|
0
|
0
|
0
|
|
4
|
4
|
|
Piliostigma thonningii
|
1
|
2
|
4
|
12
|
3
|
|
3
|
25
|
|
Pterocarpus erinaceus
|
18
|
17
|
10
|
5
|
7
|
|
29
|
86
|
|
Saba senegalensis
|
13
|
4
|
2
|
0
|
0
|
|
1
|
20
|
|
Sarcocephalus latifolius
|
8
|
14
|
1
|
3
|
2
|
|
0
|
28
|
|
Tamarindus indica
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Trichilia emetica
|
0
|
1
|
0
|
0
|
0
|
|
0
|
1
|
|
Terminalia macroptera
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Vitex madiensis
|
1
|
1
|
0
|
0
|
0
|
|
0
|
2
|
|
Total
|
129
|
104
|
37
|
23
|
21
|
|
59
|
373
|
|
2005, P3
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Total
|
|
[5-10]
|
]10-15]
|
]15-20]
|
]20-25]
|
]25-30]
|
>
|
30
|
individus
|
|
Allophyllus cobe
|
6
|
0
|
0
|
0
|
0
|
|
0
|
6
|
|
Anogeissus leiocarpus
|
5
|
2
|
1
|
0
|
3
|
|
11
|
22
|
|
Anthostema senegalense
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Bombax costatum
|
3
|
1
|
0
|
0
|
0
|
|
1
|
5
|
|
Cassia sieberiana
|
0
|
1
|
1
|
0
|
0
|
|
0
|
2
|
|
Cissus populnea
|
0
|
1
|
0
|
0
|
0
|
|
1
|
2
|
|
Combretum collinum
|
4
|
6
|
3
|
0
|
0
|
|
0
|
13
|
|
Combretum glutinosum
|
2
|
3
|
0
|
0
|
0
|
|
0
|
5
|
|
Combretum micranthum
|
40
|
3
|
0
|
0
|
0
|
|
0
|
43
|
|
Combretum nigricans
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Combretum tomentosum
|
6
|
1
|
0
|
0
|
0
|
|
0
|
7
|
|
Cordyla pinnnata
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Crossopteryx febrifuga
|
0
|
1
|
0
|
0
|
0
|
|
0
|
1
|
|
Detarium senegalense
|
7
|
0
|
3
|
2
|
0
|
|
10
|
22
|
|
Diopyros mespiliformis
|
4
|
1
|
2
|
0
|
0
|
|
1
|
8
|
|
Erythrophleum suaveolens
|
0
|
0
|
0
|
0
|
0
|
|
1
|
1
|
|
Feretia apodanthera
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Gardenia ternifolia / triacantha
|
2
|
1
|
0
|
0
|
0
|
|
0
|
3
|
|
Grewia flavescens
|
7
|
0
|
0
|
0
|
0
|
|
0
|
7
|
|
Hexalobus monopetalus
|
47
|
46
|
12
|
3
|
0
|
|
1
|
109
|
|
Lannea microcarpa
|
0
|
0
|
0
|
0
|
3
|
|
2
|
5
|
|
Lonchocarpus laxiflorus
|
0
|
0
|
1
|
0
|
0
|
|
0
|
1
|
|
Parkia biglobosa
|
0
|
0
|
0
|
0
|
0
|
|
4
|
4
|
|
Piliostigma thonningii
|
2
|
0
|
4
|
13
|
1
|
|
5
|
25
|
|
Pterocarpus erinaceus
|
21
|
18
|
11
|
4
|
7
|
|
30
|
91
|
|
Saba senegalensis
|
23
|
4
|
3
|
0
|
0
|
|
1
|
31
|
|
Sarcocephalus latifolius
|
15
|
14
|
2
|
2
|
3
|
|
0
|
36
|
|
Tamarindus indica
|
1
|
0
|
0
|
0
|
0
|
|
0
|
1
|
|
Trichilia emetica
|
0
|
1
|
0
|
0
|
0
|
|
0
|
1
|
|
Terminalia macroptera
|
0
|
0
|
0
|
0
|
0
|
|
0
|
0
|
|
Vitex madiensis
|
2
|
0
|
0
|
0
|
0
|
|
1
|
3
|
|
Total
|
199
|
104
|
43
|
24
|
17
|
|
69
|
456
|
|
Section
17.02 Annexe 2. Liste des espèces au niveau
des parcelles de suivi
|
|
|
Liste des espèces (1993)
|
Liste des espèces Nombre (1997)
|
Nombre Liste des espèces (2003)
|
Nombre Liste des espèces (2005)
|
Nombre
|
|
Régénérartion
|
163 -
|
815 -
|
1096 -
|
814
|
|
1
|
Acacia macrostachya
|
3 Acacia macrostachya
|
4 Acacia macrostachya
|
3 Acacia macrostachya
|
1
|
|
2
|
Acacia sieberiana
|
2 Acacia sieberiana
|
2 Acacia sieberiana
|
2 Acacia sieberiana
|
2
|
|
3
|
Allophylus africanus
|
2 Allophylus africanus
|
3 Allophylus africanus
|
1 Allophyllus cobe
|
7
|
|
4
|
|
|
|
Annona senegalensis
|
1
|
|
5
|
Anogeissus leiocarpus
|
22 Anogeissus leiocarpus
|
22 Anogeissus leiocarpus
|
20 Anogeissus leiocarpus
|
24
|
|
6
|
Anthostema senegalense
|
4 Anthostema senegalense
|
4 Anthostema senegalense
|
6
|
|
|
7
|
Bombax costatum
|
47 Bombax costatum
|
47 Bombax costatum
|
41 Bombax costatum
|
43
|
|
8
|
Borassus aethiopum
|
1 Borassus aethiopum
|
1 Borassus aethiopum
|
1 Borassus aethiopum
|
1
|
|
9
|
|
|
|
Capparis decidua
|
1
|
|
10
|
Cassia sieberiana
|
321 Cassia sieberiana
|
320 Cassia sieberiana
|
254 Cassia sieberiana
|
249
|
|
11
|
Cissus populnea
|
5 Cissus populnea
|
5 Cissus populnea
|
1 Cissus populnea
|
2
|
|
12
|
Combretum collinum
|
21 Combretum collinum
|
24 Combretum collinum
|
18 Combretum collinum
|
13
|
|
13
|
Combretum crotonoides
|
8 Combretum crotonoides
|
7 Combretum crotonoides
|
4 Combretum crotonoides
|
50
|
|
14
|
Combretum glutinosum
|
369 Combretum glutinosum
|
315 Combretum glutinosum
|
229 Combretum glutinosum
|
202
|
|
15
|
Combretum micranthum
|
14 Combretum micranthum
|
28 Combretum micranthum
|
34 Combretum micranthum
|
43
|
|
16
|
Combretum nigricans
|
1 Combretum nigricans
|
1 Combretum nigricans
|
1 Combretum nigricans
|
1
|
|
17
|
Combretum tomentosum
|
2 Combretum tomentosum
|
4 Combretum tomentosum
|
1 Combretum tomentosum
|
7
|
|
18
|
Cordyla pinnata
|
17 Cordyla pinnata
|
17 Cordyla pinnata
|
16 Cordyla pinnnata
|
23
|
|
19
|
Crossopteryx febrifuga
|
111 Crossopteryx febrifuga
|
92 Crossopteryx febrifuga
|
83 Crossopteryx febrifuga
|
102
|
|
20
|
Daniellia oliveri
|
25 Daniellia oliveri
|
17 Daniellia oliveri
|
13 Daniellia oliveri
|
11
|
|
21
|
Detarium microcarpum
|
21 Detarium microcarpum
|
21 Detarium microcarpum
|
19 Detarium microcarpum
|
18
|
|
22
|
Detarium senegalense
|
12 Detarium senegalense
|
17 Detarium senegalense
|
20 Detarium senegalense
|
22
|
|
23
|
Dichrostachys cinerea
|
4 Dichrostachys cinerea
|
5 Dichrostachys cinerea
|
4 Dichrostachys cinerea
|
2
|
|
24
|
Diospyros mespiliformis
|
7 Diospyros mespiliformis
|
5 Diospyros mespiliformis
|
5 Diopyros mespiliformis
|
8
|
|
25
|
Erythrophleum suaveolens
|
1 Erythrophleum suaveolens
|
1 Erythrophleum suaveolens
|
1 Erythrophleum suaveolens
|
1
|
|
Liste des espèces (1993)
|
Nombre
|
Liste des espèces (1997)
|
Nombre Liste des espèces (2003)
|
Nombre
|
Liste des espèces (2005)
|
Nombre
|
|
26
|
Feretia apodanthera
|
1
|
Feretia apodanthera
|
1
|
|
-
|
|
|
|
27
|
Gardenia erubescens
|
2
|
Gardenia erubescens
|
3
|
Gardenia erubescens
|
3
|
Gardenia ternifolia / triacantha
|
3
|
|
28
|
Grewia bicolor
|
5
|
Grewia bicolor
|
5
|
Grewia bicolor
|
1
|
|
|
|
29
|
Grewia flavescens
|
4
|
Grewia flavescens
|
8
|
Grewia flavescens
|
8
|
Grewia flavescens
|
19
|
|
30
|
Guiera senegalensis
|
2
|
|
|
|
|
|
|
|
31
|
Hexalobus monopetalus
|
234
|
Hexalobus monopetalus
|
239
|
Hexalobus monopetalus
|
232
|
Hexalobus monopetalus
|
288
|
|
32
|
Hymenocardia acida
|
1
|
|
-
|
|
|
Hymenocardia acida
|
2
|
|
33
|
Lannea acida
|
89
|
Lannea acida
|
87
|
Lannea acida
|
84
|
Lannea acida
|
89
|
|
34
|
Lannea microcarpa
|
13
|
Lannea microcarpa
|
13
|
Lannea microcarpa
|
12
|
Lannea microcarpa
|
13
|
|
35
|
Lannea velutina
|
26
|
Lannea velutina
|
24
|
Lannea velutina
|
17
|
Lannea velutina
|
16
|
|
36
|
Lonchocarpus laxiflorus
|
7
|
Lonchocarpus laxiflorus
|
6
|
Lonchocarpus laxiflorus
|
6
|
Lonchocarpus laxiflorus
|
8
|
|
37
|
Maytenus senegalensis
|
3
|
Maytenus senegalensis
|
3
|
Maytenus senegalensis
|
3
|
Maytenus senegalensis
|
4
|
|
38
|
Mitragyna inermis
|
25
|
Mitragyna inermis
|
33
|
Mitragyna inermis
|
35
|
Mitragyna inermis
|
63
|
|
39
|
Oncoba spinosa
|
1
|
Oncoba spinosa
|
1
|
Oncoba spinosa
|
1
|
Oncoba spinosa
|
3
|
|
40
|
Parkia biglobosa
|
7
|
Parkia biglobosa
|
7
|
Parkia biglobosa
|
6
|
Parkia biglobosa
|
6
|
|
41
|
Pericopsis laxiflora
|
31
|
Pericopsis laxiflora
|
30
|
Pericopsis laxiflora
|
26
|
Pericopsis laxiflora
|
22
|
|
42
|
Piliostigma reticulatum
|
16
|
Piliostigma reticulatum
|
17
|
Piliostigma reticulatum
|
19
|
Piliostigma reticulatum
|
23
|
|
43
|
Piliostigma thonningii
|
79
|
Piliostigma thonningii
|
71
|
Piliostigma thonningii
|
66
|
Piliostigma thonningii
|
73
|
|
44
|
Pterocarpus erinaceus
|
290
|
Pterocarpus erinaceus
|
288
|
Pterocarpus erinaceus
|
275
|
Pterocarpus erinaceus
|
313
|
|
45
|
Saba senegalensis
|
9
|
Saba senegalensis
|
16
|
Saba senegalensis
|
20
|
Saba senegalensis
|
32
|
|
46
|
Sarcocephalus latifolius
|
43
|
Sarcocephalus latifolius
|
40
|
Sarcocephalus latifolius
|
36
|
Sarcocephalus latifolius
|
39
|
|
47
|
Spondias mombin
|
3
|
Spondias mombin
|
3
|
Spondias mombin
|
3
|
Spondias mombin
|
3
|
|
48
|
|
|
|
|
|
|
Stereospermum kunthianum
|
2
|
|
49 Strychnos spinosa
|
48
|
Strychnos spinosa
|
49
|
Strychnos spinosa
|
46
|
Strychnos spinosa
|
81
|
|
50 Tamarindus indica
|
1
|
Tamarindus indica
|
1
|
Tamarindus indica
|
1
|
Tamarindus indica
|
4
|
|
51
|
Terminalia avicennioides
|
47
|
Terminalia avicennioides
|
49
|
Terminalia avicennioides
|
47
|
Terminalia avicennioides
|
55
|
|
52
|
Terminalia laxiflora
|
1
|
Terminalia laxiflora
|
1
|
Terminalia laxiflora
|
1
|
Terminalia macroptera
|
244
|
|
53
|
Terminalia macroptera
|
332
|
Terminalia macroptera
|
296
|
Terminalia macroptera
|
253
|
Trichilia emetica
|
9
|
|
Liste des espèces (1993)
|
Liste des espèces Nombre (1997)
|
Nombre Liste des espèces (2003)
|
Nombre Liste des espèces (2005)
|
Nombre
|
|
54 Trichilia emetica
|
6 Trichilia emetica
|
6 Trichilia emetica
|
6 Vitex madiensis
|
15
|
|
55 Vitex madiensis
|
13 Vitex madiensis
|
8 Vitex madiensis
|
5 Xeroderriis sthuhlmannii
|
1
|
|
56 Xeroderris stuhlmannii
|
27 Xeroderris stuhlmannii
|
26 Xeroderris stuhlmannii
|
25 Xeroderris stuhlmannii
|
29
|
|
57 Ziziphus mucronata
|
7 Ziziphus mucronata
|
7 Ziziphus mucronata
|
6 Ziziphus mucronata
|
6
|
|
Total
|
2556 Total
|
3115 Total
|
3116 Total
|
3113
|
|
Section 17.03 Annexe 3 : liste des espèces
inventoriées par Forêt
Classée
Forêt Classée de Bala
Affinités
|
|
No
|
Espèces
|
Familles
|
phytogéographiques
|
|
1
|
Acacia ataxacantha
|
Mimosaceae
|
soudanienne
|
|
2
|
Acacia macrostachya
|
Mimosaceae
|
soudanienne
|
|
3
|
Acacia sp
|
Mimosaceae
|
soudanienne
|
|
4
|
Adansonia digitata
|
Bombacaceae
|
soudanienne
|
|
5
|
Anogeissus leiocarpus
|
Combretaceae
|
soudanienne
|
|
6
|
Baissea multiflora
|
Apocynaceae
|
soudanienne
|
|
7
|
Balanites aegyptiaca
|
Balanitaceae
|
sahélo-soudanienne
|
|
8
|
Bombax costatum
|
Bombacaceae
|
soudanienne
|
|
9
|
Boscia angustifolia
|
Capparidaceae
|
soudanienne
|
|
10
|
Boscia salicifolia
|
Capparidaceae
|
soudanienne
|
|
11
|
Boscia senegalensis
|
Capparidaceae
|
soudanienne
|
|
12
|
Boscia sp
|
Capparidaceae
|
soudanienne
|
|
13
|
Combretum crotonoides
|
Combretaceae
|
soudanienne
|
|
14
|
Combretum glutinosum
|
Combretaceae
|
soudanienne
|
|
15
|
Combretum lecardii
|
Combretaceae
|
soudanienne
|
|
16
|
Combretum micranthum
|
Combretaceae
|
soudanienne
|
|
17
|
Combretum nigricans
|
Combretaceae
|
soudanienne
|
|
18
|
Combretum paniculatum
|
Combretaceae
|
soudanienne
|
|
19
|
Combretum sp
|
Combretaceae
|
soudanienne
|
|
20
|
Comiphora africana
|
Burseraceae
|
soudanienne
|
|
21
|
Cordyla pinnata
|
Caesalpiniaceae
|
soudanienne
|
|
22
|
Crossopteryx febrifuga
|
Rubiaceae
|
soudanienne
|
|
23
|
Dicrostachys cinerea
|
Mimosaceae
|
soudanienne
|
|
24
|
Dombea quinqueseta
|
Sterculiaceae
|
soudano-guineenne
|
|
25
|
Feretia apodanthera
|
Rubiaceae
|
soudanienne
|
|
26
|
Gardenia sp
|
Rubiaceae
|
soudanienne
|
|
27
|
Grewia bicolor
|
Tiliaceae
|
sahélo-soudanienne
|
|
28
|
Grewia flavescens
|
Tiliaceae
|
sahélo-soudanienne
|
|
29
|
Guiera senegalensis
|
Combretaceae
|
soudanienne
|
|
30
|
Hymenocardia acida
|
Euphorbiaceae
|
soudanienne
|
|
31
|
Lannea acida
|
Anacardiaceae
|
soudanienne
|
|
32
|
Loeseneriella africana
|
Hippocrateaceae
|
soudanienne
|
|
33
|
Lonchocarpus laxiflorus
|
Fabaceae
|
soudano-guineenne
|
|
34
|
Maerua angolensis
|
Capparidaceae
|
soudanienne
|
|
35
|
Maytenus senegalensis
|
Celastraceae
|
sahélo-soudanienne
|
|
36
|
Ormocarpum bibracteatum
|
Fabaceae
|
soudano-guineenne
|
|
37
|
Pericopsis laxiflorus
|
Fabaceae
|
soudano-guineenne
|
|
38
|
Piliostigma reticulatum
|
Caesalpiniaceae
|
soudanienne
|
|
39
|
Pterocarpus erinaceus
|
Fabaceae
|
soudanienne
|
|
40
|
Pterocarpus lucens
|
Fabaceae
|
soudanienne
|
|
41
|
Securidaca longepedunculata
|
Polygalaceae
|
soudanienne
|
|
42
|
Securinega virosa
|
Euphorbiaceae
|
soudanienne
|
|
43
|
Sterculia setigera
|
Sterculiaceae
|
soudanienne
|
|
44
|
Stereospermum kunthianum
|
Bignoniaceae
|
soudanienne
|
|
45
|
Strychnos spinosa
|
Loganiaceae
|
soudanienne
|
|
46
|
Terminalia avicennioides
|
Combretaceae
|
soudanienne
|
|
47
|
Terminalia macroptera
|
Combretaceae
|
soudanienne
|
|
48
|
Xeroderrys stuhlmannii
|
Fabaceae
|
soudano-guineenne
|
|
49 Ziziphus mauritiana
50 Ziziphus mucronata
Forêt Classée de Kantora
|
Rhamnaceae
Rhamnaceae
|
sahélo-soudanienne soudanienne
Affinités
|
|
No
|
Espèces
|
Familles
|
phytogéographiques
|
|
1
|
Acacia macrostachya
|
Mimosaceae
|
soudanienne
|
|
2
|
Acacia sieberiana
|
Mimosaceae
|
soudano-guinéenne
|
|
3
|
Albizia ferruginea
|
Mimosaceae
|
soudanienne
|
|
4
|
Annona senegalensis
|
Annonaceae
|
soudano-guinéenne
|
|
5
|
Baissea multiflora
|
Apocynaceae
|
soudanienne
|
|
6
|
Bombax costatum
|
Bombacaceae
|
soudanienne
|
|
7
|
Borassus aethiopum
|
Arecaceae
|
soudanienne
|
|
8
|
Boscia angustifolia
|
Capparidaceae
|
soudanienne
|
|
9
|
Burkea africana
|
Caesalpiniaceae
|
soudanienne
|
|
10
|
Canthium sp
|
Rubiaceae
|
soudanienne
|
|
11
|
Cassia sieberiana
|
Caesalpiniaceae
|
soudano-guinéenne
|
|
12
|
Cissus populnea
|
Vitaceae
|
soudano-guinéenne
|
|
13
|
Combretum crotonoides
|
Combretaceae
|
soudanienne
|
|
14
|
Combretum glutinosum
|
Combretaceae
|
soudanienne
|
|
15
|
Combretum micranthum
|
Combretaceae
|
soudanienne
|
|
16
|
Combretum molle
|
Combretaceae
|
soudanienne
|
|
17
|
Combretum nigricans
|
Combretaceae
|
soudanienne
|
|
18
|
Combretum tomentosum
|
Combretaceae
|
soudanienne
|
|
19
|
Cordyla pinnata
|
Caesalpiniaceae
|
soudanienne
|
|
20
|
Crossopteryx febrifuga
|
Rubiaceae
|
soudanienne
|
|
21
|
Daniellia oliveri
|
Caesalpiniaceae
|
soudanienne
|
|
22
|
Detarium microcarpum
|
Caesalpiniaceae
|
soudanienne
|
|
23
|
Dicrostachys cinerea
|
Mimosaceae
|
soudanienne
|
|
24
|
Dombeya quinqueseta
|
Sterculiaceae
|
soudano-guineenne
|
|
25
|
Entada africana
|
Mimosaceae
|
soudanienne
|
|
26
|
Erythrophleum africanum
|
Caesalpiniaceae
|
soudano-guineenne
|
|
27
|
Feretia apodanthera
|
Rubiaceae
|
soudanienne
|
|
28
|
Ficus glumosa
|
Moraceae
|
soudanienne
|
|
29
|
Gardenia sp
|
Rubiaceae
|
soudanienne
|
|
30
|
Grewia bicolor
|
Tiliaceae
|
sahélo-soudanienne
|
|
31
|
Grewia flavescens
|
Tiliaceae
|
sahélo-soudanienne
|
|
32
|
Guiera senegalensis
|
Combretaceae
|
soudanienne
|
|
33
|
Hannoa undulata
|
Simaroubaceae
|
soudanienne
|
|
34
|
Hexalobus monopetalus
|
Annonaceae
|
soudanienne
|
|
35
|
Hymenocardia acida
|
Euphorbiaceae
|
soudanienne
|
|
36
|
Khaya senegalensis
|
Meliaceae
|
guinéenne
|
|
37
|
Lannea acida
|
Anacardiaceae
|
soudanienne
|
|
38
|
Lannea microcarpa
|
Anacardiaceae
|
soudano-guineenne
|
|
39
|
Lannea velutina
|
Anacardiaceae
|
soudano-guineenne
|
|
40
|
Lonchocarpus laxiflorus
|
Fabaceae
|
soudano-guineenne
|
|
41
|
Maerua angolensis
|
Capparidaceae
|
soudanienne
|
|
42
|
Maytenus senegalensis
|
Celastraceae
|
sahélo-soudanienne
|
|
43
|
Mitragyna inermis
|
Rubiaceae
|
soudanienne
|
|
44
|
Ozoroa insignis
|
Anacardiaceae
|
soudanienne
|
|
45
|
Parkia biglobosa
|
Mimosaceae
|
soudano-guineenne
|
|
46
|
Pavetta cinereifolia
|
Rubiaceae
|
soudano-guineenne
|
|
47
|
Pericopsis laxiflora
|
Fabaceae
|
soudano-guineenne
|
|
48
|
Piliostigma reticulatum
|
Caesalpiniaceae
|
soudanienne
|
|
49
|
Prosopis africana
|
Mimosaceae
|
Mimosaceae
|
|
50
|
Pterocarpus erinaceus
|
Fabaceae
|
soudanienne
|
|
51
|
Sarcocephalus latifolius
|
Rubiaceae
|
soudano-guineenne
|
|
52
|
Sclerocarya birrea
|
Anacardiaceae
|
soudanienne
|
|
53
|
Securidaca longepedunculata
|
Polygalaceae
|
soudanienne
|
|
54
|
Securinega virosa
|
Euphorbiaceae
|
soudanienne
|
|
55
|
Stereospermum kunthianum
|
Bignoniaceae
|
soudanienne
|
|
56
|
Sterculia setigera
|
Sterculiaceae
|
soudanienne
|
|
57
|
Strophantus sarmentosus
|
Apocynaceae
|
soudanienne
|
|
58
|
Strychnos spinosa
|
Loganiaceae
|
soudanienne
|
|
59
|
Terminalia avicennioides
|
Combretaceae
|
soudanienne
|
|
60
|
Terminalia macroptera
|
Combretaceae
|
soudanienne
|
|
61
|
Trichilia emetica
|
Meliaceae
|
soudano-guineenne
|
|
62
|
Vitex madiensis
|
Verbenaceae
|
soudanienne
|
|
63
|
Xeroderrys stuhlmannii
|
Fabaceae
|
soudano-guineenne
|
|
64
|
Ximenia americana
|
Olacaceae
|
soudanienne
|
|
65
|
Ziziphus mucronata
|
Rhamnaceae
|
soudanienne
|
Forêt Classée de Mampaye
|
|
|
Affinités
|
|
No
|
Espèces
|
Familles
|
phytogéographiques
|
|
1
|
Acacia sieberiana
|
Mimosaceae
|
soudano-guinéenne
|
|
2
|
Allophylus africanus
|
Sapindaceae
|
guinéenne
|
|
3
|
Andira inermis
|
Fabaceae
|
guinéenne
|
|
4
|
Annona senegalensis
|
Annonaceae
|
soudano-guinéenne
|
|
5
|
Anogeissus leiocarpus
|
Combretaceae
|
soudanienne
|
|
6
|
Baissea multiflora
|
Apocynaceae
|
soudanienne
|
|
7
|
Bombax costatum
|
Bombacaceae
|
soudanienne
|
|
8
|
Borassus aethiopum
|
Arecaceae
|
soudanienne
|
|
9
|
Bridelia micrantha
|
Euphorbiaceae
|
soudano-guinéenne
|
|
10
|
Canthium sp
|
Rubiaceae
|
soudanienne
|
|
11
|
Cassia sieberiana
|
Caesalpiniaceae
|
soudanienne
|
|
12
|
Cissus populnea
|
Vitaceae
|
soudanienne
|
|
13
|
Combretum crotonoides
|
Combretaceae
|
soudano-guinéenne
|
|
14
|
Combretum glutinosum
|
Combretaceae
|
soudano-guinéenne
|
|
15
|
Combretum lecardii
|
Combretaceae
|
soudanienne
|
|
16
|
Combretum micranthum
|
Combretaceae
|
soudanienne
|
|
17
|
Combretum nigricans
|
Combretaceae
|
soudanienne
|
|
18
|
Cordyla pinnata
|
Caesalpiniaceae
|
soudanienne
|
|
19
|
Crossopteryx febrifuga
|
Rubiaceae
|
soudanienne
|
|
20
|
Daniellia oliveri
|
Caesalpiniaceae
|
soudanienne
|
|
21
|
Detarium microcarpum
|
Caesalpiniaceae
|
soudanienne
|
|
22
|
Dicrostachys cinerea
|
Mimosaceae
|
soudanienne
|
|
23
|
Diospyros mespiliformis
|
Ebenaceae
|
soudanienne
|
|
24
|
Entada africana
|
Mimosaceae
|
soudanienne
|
|
25
|
Erythrophleum africanum
|
Caesalpiniaceae
|
soudano-guineenne
|
|
26
|
Erythrophleum suaveolens
|
Caesalpiniaceae
|
guinéenne
|
|
27
|
Feretia apodanthera
|
Rubiaceae
|
soudanienne
|
|
28
|
Ficus glumosa
|
Moraceae
|
soudano-guineenne
|
|
29
|
Gardenia sp
|
Rubiaceae
|
soudanienne
|
|
30
|
Grewia bicolor
|
Tiliaceae
|
soudanienne
|
|
31
|
Grewia flavescens
|
Tiliaceae
|
soudanienne
|
|
32
|
Guiera senegalensis
|
Combretaceae
|
sahélo-soudanienne
|
|
33
|
Hannoa undulata
|
Simaroubaceae
|
sahélo-soudanienne
|
|
34
|
Hexalobus monopetalus
|
Annonaceae
|
soudanienne
|
|
35
|
Holarrhena floribunda
|
Apocynaceae
|
guinéenne
|
|
36
|
Hymenocardia acida
|
Euphorbiaceae
|
soudanienne
|
|
37
|
Khaya senegalensis
|
Meliaceae
|
soudanienne
|
|
38
|
Lannea acida
|
Anacardiaceae
|
soudanienne
|
|
39
|
Lannea velutina
|
Anacardiaceae
|
soudanienne
|
|
40
|
Lonchocarpus laxiflorus
|
Fabaceae
|
soudano-guineenne
|
|
41
|
Lophira lanceolata
|
|
soudano-guineenne
|
|
42
|
Maerua angolensis
|
Capparidaceae
|
soudano-guineenne
|
|
43
|
Maytenus senegalensis
|
Celastraceae
|
soudano-guineenne
|
|
44
|
Mitragyna inermis
|
Rubiaceae
|
soudanienne
|
|
45
|
Oncoba sp
|
Flacourtiaceae
|
soudano-guineenne
|
|
46
|
Ormocarpum bibracteatum
|
Fabaceae
|
soudano-guineenne
|
|
47
|
Ozoroa insignis
|
Anacardiaceae
|
sahélo-soudanienne
|
|
48
|
Parkia biglobosa
|
Mimosaceae
|
soudanienne
|
|
49
|
Parinari curatellifolia
|
Rosaceae
|
soudano-guineenne
|
|
50
|
Pericopsis laxiflora
|
Fabaceae
|
soudano-guineenne
|
|
51
|
Piliostigma reticulatum
|
Caesalpiniaceae
|
soudanienne
|
|
52
|
Piliostigma thonningii
|
Caesalpiniaceae
|
soudano-guineenne
|
|
53
|
Prosopis africana
|
Mimosaceae
|
soudano-guineenne
|
|
54
|
Pterocarpus erinaceus
|
Fabaceae
|
soudanienne
|
|
55
|
Pteleopsis suberosa
|
Combretaceae
|
soudano-guineenne
|
|
56
|
Saba senegalensis
|
Apocynaceae
|
soudano-guineenne
|
|
57
|
Sarcocephalus latifolius
|
Rubiaceae
|
soudano-guineenne
|
|
58
|
Securinega virosa
|
Euphorbiaceae
|
soudanienne
|
|
59
|
Stereospermum kunthianum
|
Bignoniaceae
|
soudanienne
|
|
60
|
Sterculia setigera
|
Sterculiaceae
|
soudanienne
|
|
61
|
Strophantus sarmentosus
|
Apocynaceae
|
soudanienne
|
|
62
|
Strychnos spinosa
|
Loganiaceae
|
soudanienne
|
|
63
|
Terminalia avicennioides
|
Combretaceae
|
soudanienne
|
|
64
|
Terminalia laxiflora
|
Combretaceae
|
soudano-guineenne
|
|
65
|
Terminalia macroptera
|
Combretaceae
|
soudanienne
|
|
66
|
Trichilia emetica
|
Meliaceae
|
soudanienne
|
|
67
|
Tricalysia okelensis
|
Rubiaceae
|
soudano-guineenne
|
|
68
|
Vernonia colorata
|
Asteraceae
|
soudano-guineenne
|
|
69
|
Vitex madiensis
|
Verbenaceae
|
soudanienne
|
|
70
|
Xeroderrys stuhlmannii
|
Fabaceae
|
soudano-guineenne
|
|
71
|
Ximenia americana
|
Olacaceae
|
soudanienne
|
|
72
|
Ziziphus mucronata
|
Rhamnaceae
|
soudano-guineenne
|
Forêt Classée de Ouli
|
No
|
Espèces
|
Familles
|
Affinités phytogéographiques
|
|
1
|
Acacia macrostachya
|
Mimosaceae
|
soudanienne
|
|
2
|
Acacia seyal
|
Mimosaceae
|
soudanienne
|
|
3
|
Anogeissus leiocarpus
|
Combretaceae
|
soudanienne
|
|
4
|
Azadirachta indica
|
Meliaceae
|
introduite
|
|
5
|
Baissea multiflora
|
Apocynaceae
|
soudanienne
|
|
6
|
Balanites aegyptiaca
|
Balanitaceae
|
sahélo-soudanienne
|
|
7
|
Bombax costatum
|
Bombacaceae
|
soudanienne
|
|
8
|
Boscia angustifolia
|
Capparidaceae
|
soudanienne
|
|
9
|
Boscia salicifolia
|
Capparidaceae
|
soudanienne
|
|
10
|
Boscia sp
|
Capparidaceae
|
soudanienne
|
|
11
|
Combretum collinum
|
Combretaceae
|
soudanienne
|
|
12
|
Combretum crotonoides
|
Combretaceae
|
soudanienne
|
|
13
|
Combretum glutinosum
|
Combretaceae
|
soudanienne
|
|
14
|
Combretum micranthum
|
Combretaceae
|
soudanienne
|
|
15
|
Combretum molle
|
Combretaceae
|
soudanienne
|
|
16
|
Combretum nigricans
|
Combretaceae
|
soudanienne
|
|
17
|
Cordyla pinnata
|
Caesalpiniaceae
|
soudanienne
|
|
18
|
Crossopteryx febrifuga
|
Rubiaceae
|
soudanienne
|
|
19
|
Dicrostachys cinerea
|
Mimosaceae
|
soudanienne
|
|
20
|
Entada africana
|
Mimosaceae
|
soudanienne
|
|
21
|
Feretia apodanthera
|
Rubiaceae
|
soudanienne
|
|
22
|
Gardenia sp
|
Rubiaceae
|
soudanienne
|
|
23
|
Grewia bicolor
|
Tiliaceae
|
sahélo-soudanienne
|
|
24
|
Grewia flavescens
|
Tiliaceae
|
sahélo-soudanienne
|
|
25
|
Guiera senegalensis
|
Combretaceae
|
soudanienne
|
|
26
|
Hannoa undulata
|
Simaroubaceae
|
soudanienne
|
|
27
|
Hexalobus monopetalus
|
Annonaceae
|
soudanienne
|
|
28
|
Lannea acida
|
Anacardiaceae
|
soudanienne
|
|
29
|
Maerua angolensis
|
Capparidaceae
|
soudanienne
|
|
30
|
Maytenus senegalensis
|
Celastraceae
|
sahélo-soudanienne
|
|
31
|
Ormocarpum bibracteatum
|
Fabaceae
|
soudano-guineenne
|
|
32
|
Piliostigma reticulatum
|
Caesalpiniaceae
|
soudanienne
|
|
33
|
Piliostigma thonningii
|
Caesalpiniaceae
|
soudano-guineenne
|
|
34
|
Pterocarpus erinaceus
|
Fabaceae
|
soudanienne
|
|
35
|
Sclerocarya birrea
|
Anacardiaceae
|
soudanienne
|
|
36
|
Securidaca longepedunculata
|
Polygalaceae
|
soudanienne
|
|
37
|
Securinega virosa
|
Euphorbiaceae
|
soudanienne
|
|
38
|
Stereospermum kunthianum
|
Bignoniaceae
|
soudanienne
|
|
39
|
Sterculia setigera
|
Sterculiaceae
|
soudanienne
|
|
40
|
Strychnos spinosa
|
Loganiaceae
|
soudanienne
|
|
41
|
Tamarindus indica
|
Caesalpiniaceae
|
soudanienne
|
|
42
|
Terminalia avicennioides
|
Combretaceae
|
soudanienne
|
|
43
|
Terminalia macroptera
|
Combretaceae
|
soudanienne
|
|
44
|
Xeroderrys stuhlmannii
|
Fabaceae
|
soudano-guineenne
|
|
45
|
Ziziphus mauritiana
|
Rhamnaceae
|
sahélo-soudanienne
|
|
46
|
Ziziphus mucronata
|
Rhamnaceae
|
soudanienne
|
Forêt Classée de Patako
|
|
|
Affinités
|
|
No
|
Espèces
|
Familles
|
phytogéographiques
|
|
1
|
Acacia macrostachya
|
Mimosaceae
|
soudanienne
|
|
2
|
Acacia sieberiana
|
Mimosaceae
|
soudano-guinéenne
|
|
3
|
Afzelia africana
|
Caesalpiniaceae
|
soudano-guinéenne
|
|
4
|
Albizia zygia
|
Mimosaceae
|
soudano-guinéenne
|
|
5
|
Allphylus africanus
|
Sapindaceae
|
guinéenne
|
|
6
|
Annona senegalensis
|
Annonaceae
|
soudano-guinéenne
|
|
7
|
Anogeissus leiocarpus
|
Combretaceae
|
soudanienne
|
|
8
|
Antiaris toxicaria
|
Moraceae
|
guinéenne
|
|
9
|
Azadirachta indica
|
Meliaceae
|
introduite
|
|
10
|
Baissea multiflora
|
Apocynaceae
|
soudanienne
|
|
11
|
Balanites aegyptiaca
|
Zygophyllaceae
|
sahélo-soudanienne
|
|
12
|
Bombax costatum
|
Bombacaceae
|
soudanienne
|
|
13
|
Bridelia micrantha
|
Euphorbiaceae
|
soudano-guinéenne
|
|
14
|
Cassia sieberiana
|
Caesalpiniaceae
|
soudanienne
|
|
15
|
Cissus populnea
|
Vitaceae
|
soudanienne
|
|
16
|
Cola cordifolia
|
Sterculiaceae
|
soudano-guinéenne
|
|
17
|
Combretum glutinosum
|
Combretaceae
|
soudano-guinéenne
|
|
18
|
Combretum lecardii
|
Combretaceae
|
soudanienne
|
|
19
|
Combretum micranthum
|
Combretaceae
|
soudanienne
|
|
20
|
Combretum nigricans
|
Combretaceae
|
soudanienne
|
|
21
|
Combretum tomentosum
|
Combretaceae
|
soudano-guinéenne
|
|
22
|
Cordyla pinnata
|
Caesalpiniaceae
|
soudanienne
|
|
23
|
Crossopteryx febrifuga
|
Rubiaceae
|
soudanienne
|
|
24
|
Daniellia oliveri
|
Caesalpiniaceae
|
soudanienne
|
|
25
|
Detarium microcarpum
|
Caesalpiniaceae
|
soudanienne
|
|
26
|
Dicrostachys cinerea
|
Mimosaceae
|
soudanienne
|
|
27
|
Dombeya quinqueseta
|
Sterculiaceae
|
soudano-guinéenne
|
|
28
|
Elaeis guineensis
|
Arecaceae
|
guinéenne
|
|
29
|
Entada africana
|
Mimosaceae
|
soudanienne
|
|
30
|
Feretia apodanthera
|
Rubiaceae
|
soudanienne
|
|
31
|
Ficus sur
|
Moraceae
|
soudano-guineenne
|
|
32
|
Gardenia erubescens
|
Rubiaceae
|
soudano-guineenne
|
|
33
|
Gardenia sp
|
Rubiaceae
|
soudano-guineenne
|
|
34
|
Guiera senegalensis
|
Combretaceae
|
sahélo-soudanienne
|
|
35
|
Hannoa undulata
|
Simaroubaceae
|
sahélo-soudanienne
|
|
36
|
Hexalobus monopetalus
|
Annonaceae
|
soudanienne
|
|
37
|
Hymenocardia acida
|
Euphorbiaceae
|
soudanienne
|
|
38
|
Khaya senegalensis
|
Meliaceae
|
soudanienne
|
|
39
|
Lannea acida
|
Anacardiaceae
|
soudanienne
|
|
40
|
Landolphia heudelottii
|
Apocynaceae
|
guinéenne
|
|
41
|
Lannea velutina
|
Anacardiaceae
|
soudanienne
|
|
42
|
Lepisanthes senegalensis
|
Sapindaceae
|
guinéenne
|
|
43
|
Lonchocarpus laxiflorus
|
Fabaceae
|
soudano-guineenne
|
|
44
|
Maerua angolensis
|
Capparidaceae
|
soudano-guineenne
|
|
45
|
Malacantha alnifolia
|
Sapotaceae
|
guinéenne
|
|
46
|
Maytenus senegalensis
|
Celastraceae
|
soudano-guineenne
|
|
47
|
Neocarya macrophylla
|
Rosaceae
|
soudano-guineenne
|
|
48
|
Newbouldia laevis
|
Bignoniaceae
|
soudano-guineenne
|
|
49
|
Ozoroa insignis
|
Anacardiaceae
|
soudanienne
|
|
50
|
Parkia biglobosa
|
Mimosaceae
|
soudano-guineenne
|
|
51
|
Pericopsis laxiflora
|
Fabaceae
|
sahélo-soudanienne
|
|
52
|
Piliostigma reticulatum
|
Caesalpiniaceae
|
soudanienne
|
|
53
|
Piliostigma thonningii
|
Caesalpiniaceae
|
soudano-guineenne
|
|
54
|
Prosopis africana
|
Mimosaceae
|
soudano-guineenne
|
|
55
|
Pseudospondias microcarpa
|
Anacardiaceae
|
guinéenne
|
|
56
|
Pterocarpus erinaceus
|
Fabaceae
|
soudanienne
|
|
57
|
Saba senegalensis
|
Apocynaceae
|
soudano-guineenne
|
|
58
|
Sclerocarya birrea
|
Anacardiaceae
|
soudanienne
|
|
59
|
Securidaca longepedunculata
|
Polygalaceae
|
soudanienne
|
|
60
|
Securinega virosa
|
Euphorbiaceae
|
soudano-guineenne
|
|
61
|
Spondias mombin
|
Anacardiaceae
|
soudano-guineenne
|
|
62
|
Stereospermum kunthianum
|
Bignoniaceae
|
soudanienne
|
|
63
|
Sterculia setigera
|
Sterculiaceae
|
soudano-guineenne
|
|
64
|
Strychnos spinosa
|
Loganiaceae
|
soudanienne
|
|
65
|
Terminalia avicennioides
|
Combretaceae
|
soudanienne
|
|
66
|
Terminalia laxiflora
|
Combretaceae
|
soudanienne
|
|
67
|
Terminalia macroptera
|
Combretaceae
|
soudanienne
|
|
68
|
Trichilia emetica
|
Meliaceae
|
soudanienne
|
|
69
|
Tricalysia okelensis
|
Rubiaceae
|
soudano-guineenne
|
|
70
|
Vitex madiensis
|
Verbenaceae
|
soudanienne
|
|
71
|
Xeroderrys stuhlmannii
|
Fabaceae
|
soudano-guineenne
|
|
72
|
Ximenia americana
|
Olacaceae
|
soudano-guineenne
|
|
73
|
Ziziphus mauritiana
|
Rhamnaceae
|
soudanienne
|
Forêt Classée de Wélor
|
|
|
Affinités
|
|
No
|
Espèces
|
Familles
|
phytogéographiques
|
|
1
|
Acacia ataxacantha
|
Mimosaceae
|
soudanienne
|
|
2
|
Acacia macrostachya
|
Mimosaceae
|
soudanienne
|
|
3
|
Acacia senegal
|
Mimosaceae
|
soudanienne
|
|
4
|
Acacia seyal
|
Mimosaceae
|
soudanienne
|
|
5
|
Acacia sieberiana
|
Caesalpiniaceae
|
soudano-guinéenne
|
|
6
|
Adansonia digitata
|
Bombacaceae
|
soudanienne
|
|
7
|
Anogeissus leiocarpus
|
Combretaceae
|
soudanienne
|
|
8
|
Azadirachta indica
|
Meliaceae
|
introduite
|
|
9
|
Balanites aegyptiaca
|
Balanitaceae
|
sahélo-soudanienne
|
|
10
|
Bombax costatum
|
Bombacaceae
|
soudanienne
|
|
11
|
Borassus aethiopum
|
Arecaceae
|
soudanienne
|
|
12
|
Boscia salicifolia
|
Capparidaceae
|
soudanienne
|
|
13
|
Boscia sp
|
Capparidaceae
|
soudanienne
|
|
14
|
Cadaba farinosa
|
Capparidaceae
|
soudanienne
|
|
15
|
Calotropis procera
|
Asclepiadaceae
|
sahélo-soudanienne
|
|
16
|
Capparis tomentosa
|
Capparidaceae
|
soudanienne
|
|
17
|
Combretum aculeatum
|
Combretaceae
|
soudanienne
|
|
18
|
Commiphora africana
|
Burseraceae
|
soudanienne
|
|
19
|
Combretum glutinosum
|
Combretaceae
|
soudanienne
|
|
20
|
Combretum micranthum
|
Combretaceae
|
soudanienne
|
|
21
|
Combretum nigricans
|
Combretaceae
|
soudanienne
|
|
22
|
Combretum sp
|
Combretaceae
|
soudanienne
|
|
23
|
Cordyla pinnata
|
Caesalpiniaceae
|
soudanienne
|
|
24
|
Crateva andansonii
|
Capparidaceae
|
soudanienne
|
|
25
|
Dicrostachys cinerea
|
Mimosaceae
|
soudanienne
|
|
26
|
Diospyros mespiliformis
|
Ebenaceae
|
soudanienne
|
|
27
|
Feretia apodanthera
|
Rubiaceae
|
soudanienne
|
|
28
|
Gardenia sp
|
Rubiaceae
|
soudanienne
|
|
29
|
Grewia bicolor
|
Tiliaceae
|
sahélo-soudanienne
|
|
30
|
Grewia flavescens
|
Tiliaceae
|
sahélo-soudanienne
|
|
31
|
Guiera senegalensis
|
Combretaceae
|
soudanienne
|
|
32
|
Hexalobus monopetalus
|
Annonaceae
|
soudanienne
|
|
33
|
Lannea acida
|
Anacardiaceae
|
soudanienne
|
|
34
|
Loeseneriella africana
|
Hippocrateaceae
|
soudanienne
|
|
35
|
Maerua angolensis
|
Capparidaceae
|
soudanienne
|
|
36
|
Maytenus senegalensis
|
Celastraceae
|
sahélo-soudanienne
|
|
37
|
Mitragyna inermis
|
Rubiaceae
|
soudanien
|
|
38
|
Neocarya macrophylla
|
Rosaceae
|
soudano-guinéenne
|
|
39
|
Piliostigma reticulatum
|
Caesalpiniaceae
|
soudanienne
|
|
40
|
Sclerocarya birrea
|
Anacardiaceae
|
soudanienne
|
|
41
|
Securidaca longepedunculata
|
Polygalaceae
|
soudanienne
|
|
42
|
Securinega virosa
|
Euphorbiaceae
|
soudanienne
|
|
43
|
Stereospermum kunthianum
|
Bignoniaceae
|
soudanienne
|
|
44
|
Sterculia setigera
|
Sterculiaceae
|
soudanienne
|
|
45
|
Strophantus sarmentosus
|
Apocynaceae
|
soudanienne
|
|
46
|
Tamarindus indica
|
Caesalpiniaceae
|
soudanienne
|
|
47
|
Ximenia americana
|
Olacaceae
|
soudanienne
|
48 Ziziphus mauritiana Rhamnaceae
sahélo-soudanienne
Section 17.04 Annexe 4 : Codes VBA pour un model
Simple de dynamique de carbone
Dim PPN, LVivant, LLitiere, LSol, hf As Double Dim iVivant,
iLitiere, iSol As Double
Dim CVivant, CLitiere, CSOl, CTotal As Integer Dim Annee, NbAnnee
As Integer
Sub Main()
initialisation
For Annee = 1 To NbAnnee
CVivant = CVivant + (PPN - CVivant / LVivant)
CLitiere = CLitiere + (CVivant / LVivant - hf * CLitiere /
LLitiere - (1 - hf) * CLitiere / LLitiere)
CSOl = CSOl + (hf * CLitiere / LLitiere - CSOl / LSol)
printresults Annee, CVivant, CLitiere, CSOl
Next Annee
End Sub
Sub initialisation()
'charger le nombre d'annees pour executer le modèle a
partir de la feuille excel' NbAnnee = Range("b1")
'Charger la valeur du PPN a partir de la feuille Excel' PPN =
Range("b5")
'Charger le reste des parametres a partir de la feuile excel'
LVivant = Range("b6") LLiriere = Range("b7") LSol =
Range("b8")
hf = Range("b9")
'Charger le stock de carboen initial'
'et le stock de carbone actuel a partir des valeurs suivantes'
iVivant = Range("b13") iLitiere = Range("b14") iSol =
Range("b15") CVivant = iVivant
CLitiere = iLitiere
CSOl = iSol
'Creer un grand rectangle dans la feuille de calcul entre les
cellule e1 a h5004. 'pour entrer 2000 annees de simulations'
Set Rangeto = Worksheets("Feuil1").Range("e1:h5004")
's'assurer que les cellules sont vides'
Rangeto.Cells = Empty
'Renseigner les entetes ainsi que le stock de carbone initial sur
la feuille de calcul'
Rangeto.Cells(1, 1) = "Annee" Rangeto.Cells(1, 2) = "CVivant"
Rangeto.Cells(1, 3) = "CLitiere" Rangeto.Cells(1, 4) = "CSol"
Rangeto.Cells(2, 1) = "0" Rangeto.Cells(2, 2) = iVivant
Rangeto.Cells(2, 3) = iLitiere Rangeto.Cells(2, 4) = iSol End Sub
Sub printresults(Annee, CVivant, CLitiere, CSOl) End Sub

Configuration de la feuille EXCEL
Section 17.05 Annexe 5. Source de verification des
noms scientifiques des plantes
Section 17.06
Conservatoire et Jardins Botanique de Genève :
http://www.villege.ch/musinfo/bd/cjb/africa/index.php?langue=an
Base de données en ligne :
Moteur de recherche dans la base de données.
NB. Cette sources incluse toutes les informations disponibles
dans :
Jean-Pierre Lebrun et Adélaide L. Stork, 1992 :
Enumération des plantes à fleurs d'Afrique Tropicale. 4
volumes.
La base de données ne nécessite pa un mot de passe
: l'accès est libre
| 

