|

MUSEUM NATIONAL D'HISTOIRE NATURELLE
Ecole Doctorale Sciences de la Nature et de l'Homme - ED
227
Année 2011 N°attribué par la
bibliothèque
| | | | | | | | | | | | |
THÈSE
Pour obtenir le grade de
DOCTEUR (PHD) DU MUSEUM NATIONAL D'HISTOIRE
NATURELLE
Spécialité : Physiologie et Biologie des Organismes
- Populations - Interactions
Présentée et soutenue publiquement
par
Issa SAWADOGO
Le 20 décembre 2011
Ressources fourragères et représentations des
éleveurs, évolution des pratiques
pastorales en contexte
d'aire protégée
Cas du terroir de Kotchari à la
périphérie de la réserve de biosphère du W au
Burkina Faso
Sous la direction de : Madame FOURNIER Anne,
Chargé de recherche, HDR
JURY
M. Jean BOUTRAIS Directeur de Recherche
Émérite en Géographie, Institut de Recherche pour
Président
le Développement (IRD).
M. Jean-François Professeur de Nutrition,
Transfert de technologie et Développement rural, Rapporteur
GRONGNET Chercheur associé à
l'INRA (Département AlimH), Agrocampus Ouest de
Rennes.
MME. Anne Chargée de Recherche en
Phytoécologie, HDR, UMR 208 PALOC, Directrice de
FOURNIER Département HNS, Institut de
Recherche pour le Développement (IRD). thèse
Dédicaces
DEDICACES
Je dédie ce travail,
A mon père, à ma mère qui m'ont
toujours accompagné par leurs prières. Vous êtes pour moi
un exemple de courage et je vous en suis très
reconnaissant.
A ma bien aimée Aguiratou et à mes
fines fleurs, Rachidatou/Laéticia, Sabrinatou et Ben Charrif. Merci pour
avoir su supporter mes multiples absences et mes sautes d'humeur. Votre amour
m'a donné l'énergie nécessaire pour aller jusqu'au bout de
cette aventure. Que Charrif me pardonne si je n'ai pas été
là quand il apprenait à apprivoiser ce monde.

I
Aux populations de Kotchari en particulier les
éleveurs. Durant cette aventure, j'ai appris à vous connaitre et
vous m'avez témoigné de votre hospitalité. Puisse ce
travail contribuer à apporter quelques pistes de solution aux multiples
contraintes auxquelles vous faites face. Un merci particulier à l'ami
Yobi pour ces heures précieuses de confidence qui me permettaient de
rectifier certaines informations collectées.
REMERCIEMENTS
Deux sentiments m'animent au moment où je
boucle ce travail : celui d'un soulagement d'avoir pu le conduire à son
terme mais aussi le souvenir qu'il est le produit d'un engagement collectif
où les uns et les autres ont apporté leurs touches de nature
diverse et d'ampleur variable. Un devoir s'impose à moi de leur en
être reconnaissant.
Avant tout je remercie le programme CORUS
(Coopération pour la Recherche Universitaire et Scientifique) phase II,
qui, en finançant le projet « Gestion de l'élevage et
des feux de brousse et conservation de la biodiversité au Burkina Faso
», a permis la mise en oeuvre de cette thèse, notamment son
volet collecte de données. Un grand merci à l'IRD qui, par la
bourse de formation continue et les commodités offertes (bureau à
Ouagadougou, Orléans et Paris), a permis que ce travail soit une
réalité et de la meilleure manière.
Je voudrais remercier Madame Anne FOURNIER, ma
directrice de thèse. Merci d'avoir accepté, alors que nous
n'étions pas sûrs de réunir les moyens nécessaires,
de m'accueillir dans votre unité (l'UR 136) qui vivait ses derniers
moments d'existence. Intellectuellement et professionnellement j'ai appris
à vos côtés. J'ai aussi appris à aborder les
questions autrement grâce à votre encadrement. Merci pour votre
dévouement, votre réactivité et votre patience
;
Monsieur Jean-Louis DEVINEAU, comment vous exprimer
mes sentiments ? J'ai abordé cette thèse la peur au ventre car je
ne maîtrisais pas les méthodes choisies comme par exemple
l'imagerie. Aujourd'hui, même si des lacunes demeurent, je dois dire que
j'ai avancé et cela grâce à votre dévouement. Que
dire du grand rôle que vous avez joué dans le pilotage de cette
thèse et dans les analyses statistiques ? Merci infiniment pour tout
;
Aux autres scientifiques et spécialistes qui
ont accepté juger ce travail, je suis sensible à l'honneur que
vous me faites. J'exprime particulièrement ma profonde gratitude
à messieurs Jean-François GRONGNET, Professeur à
Agrocampus Ouest de Rennes, Jean BOUTRAIS, Directeur de recherche à
l'IRD ainsi que Jean GANGLO, Professeur à l'Université
d'Abomey-Calavi (Bénin) pour avoir accepté promptement,
malgré leurs multiples occupations, et avec plaisir de faire partie du
jury de cette thèse. Comment ne pas se souvenir des séances de
travail au début de cette thèse avec Pr BOUTRAIS dans les locaux
de l'IRD-Orléans alors que nous cherchions les astuces
nécessaires à ce genre de travail ?
Monsieur Rainer ZAÏSS du laboratoire de
cartographie appliquée de Bondy/IRD-Iles de France (Paris), grand merci
pour avoir poursuivi ce que Mr Devineau a commencé. Je te dois de
connaitre les fondamentaux de la cartographie et des SIG. J'associe à
ces remerciements Madame Marcia De Andrade Mathieu, directrice du laboratoire
pour son accueil chaleureux;
Madame Chantal-Yvette KABORÉ-ZOUNGRANA, je
n'oublie pas que vous avez toujours été là soit pour
m'encadrer avec la rigueur scientifique qui est la votre (Mémoire
d'ingénieur et de DEA) ou alors pour nourrir la réflexion. Je
n'oublie pas aussi que, nonobstant vos fonctions actuelles très
prenantes, vous avez trouvé par moments du temps pour m'écouter
et me «passer» certaines astuces qui remobilisent ;
Monsieur Hamadé KAGONÉ, merci d'avoir
accepté accompagner ce travail en participant au comité de
thèse. Il était prévu que vous puissiez assister à
son dénouement en participant au jury mais votre emploi de temps
chargé n'a pas rendu cela possible;
Monsieur Jean-Sibiri ZOUNDI, que vous dire? Vous
m'avez «pris en main» dès mes premiers pas à la
recherche et votre voeux était que j'évolue à vos
côtés. La suite n'a pas permis cela mais vous n'avez pas
cessé, de votre position de chef LR/D, à scruter tous les
horizons qui s'ouvraient à vous pour me permettre de poursuivre ma
formation. Votre dynamisme m'a particulièrement encouragé et j'en
suis très reconnaissant surtout que même parti à l'OCDE
vous avez gardé le contact. Finalement, la vie faisant bien les choses
(le monde étant petit diront certains), vous vous retrouvez être
mon tuteur et témoin du dénouement de cette aventure à
Paris ;
Saïbou NIGNAN, mon ami et compagnon de terrain,
tu as pris une part très active dans ce travail notamment dans son volet
collecte de données botaniques. Je te dois par ailleurs d'avoir pu
améliorer mes connaissances dans ce domaine. Finalement cette
thèse a eu ce côté magique d'avoir permis que le lien entre
toi et moi se noue. Merci pour tout ce sacrifice (Bobo-Diapaga, ce n'est quand
même pas à côté !) et note que je te tiens pour
toujours et ne te lâcherai pas pour rien au monde ;
Roland GUIRÉ et Madame, ma famille de Diapaga,
permettez que je vous dise merci pour toute votre disponibilité et
l'accueil des étudiants successifs que je vous envoyais parfois sans le
minimum. Roland a été de toutes mes missions, en tout cas les
principales et a été d'un apport sans commune mesure dans la
collecte des données surtout botaniques et dans la facilitation des
contacts. Il est aussi l'auteur de certains clichés sur les races
animales. Mais il reste encore des choses à faire et je compte toujours
sur sa disponibilité en particulier dans le cadre de Biospher
Association que nous venons de créer;
Christophe MAHUZIER, l'homme disponible, je
n'oublierai pas de sitôt ta gentillesse et ta disponibilité. Mille
fois merci pour le logement à Orléans.
Mouctar ZERBO, merci infiniment pour la saisie et la
mise en forme de mes données d'enquête. Mais pouvait-il en
être autrement puisque, finalement, tu n'as fait que ton travail
d'esclave ;
Monsieur, Louis SAWADOGO, merci pour votre
disponibilité lorsque je vous ai contacté pour rapporter ce
travail même si finalement cela n'a pas été
possible.
A la grande famille SAWADOGO, merci infiniment du
soutien. Tasseré particulièrement a été là
à tous les instants de mon parcours bien avant cette thèse et je
lui dédie le produit de ces incessants voyages pendant lesquels je le
mobilisais. Je formule le voeu que tu puisses continuer à jouer pour moi
le rôle de grand frère et d'éclaireur que tu accomplis
déjà avec plein de réussite. A lui, j'associe mon grand
frère Boureima, les autres frères Alidou, Karim, Noufou et ma
soeur pleine de courage Aminata ;
Abdouramane Ousmane DIALLO, Adama BORO, André
KIÈMA, Blaise OUÉDRAOGO, Issa Boubacar CISSÉ (Hey Suka
!!), Jacques SOMDA, Karim SOULI, Ousmane DOUSSA (merci en passant pour la
relecture du présent manuscrit), Ousseini KOUDOUGOU, Nachor SORGHO,
Romaine ZANGRÉ/KONSEIGA, Salif TIEMTORÉ, Tidjeni BELOUME, Sidiki
KOULIBALI et Yacouba MAÏGA, vous êtes pour moi des

IV
exemples d'amis accomplis. Les responsabilités
de la vie nous ont éloignés les uns des autres. Néanmoins
nous ne nous oublions pas et faisons toujours l'effort de maintenir ce lien que
rien d'autre ne peut remplacer. Merci à tous de continuer dans ce sens
car finalement qu'y at-il de mieux ?
Un grand merci à tous les collègues de
l'INERA, en particulier ceux du Centre Régional de Recherches Agricoles
et Environnementales de l'Est (CRREA-Est), station de Kouaré (Fada
N'Gourma). Je voudrais particulièrement remercier le
Délégué Régional et le Chef de Programme GRN/SP
pour leur compréhension et leur soutien. Grande soeur Binta,
voilà le travail pour lequel tu n'as cessé de m'encourager
;
Toute ma reconnaissance à toute
l'administration de l'INERA en particulier, le Directeur et le chef de
département gestion des ressources naturelles et systèmes de
productions (GRN/SP) sans qui ce travail n'aurait pas été
possible ;
A mes prédécesseurs auprès de
Madame Anne FOURNIER (UR 136): Urbain YAMÉOGO, Sébastien
KIÈMA, Alexis KABORÉ et Françoise VALEA, merci pour les
sillons tracés et l'amitié que vous m'avez
témoigné. Au passage je souhaite pleine réussite à
mon compagnon de l'UMR 208 PALOC, Lassina SANOU à qui je dis par
ailleurs merci pour sa gentillesse inégalée. Je n'oublie pas ses
délicieux plats à Orléans
Tout le long de cette thèse j'ai eu l'honneur
de travailler avec un certain nombre d'étudiants (Abdoul-Wahab ZOMBRA,
Dominique OUÉDRAOGO, Kassoum OUÉDRAOGO et Etienne SODRÉ)
en formation d'ingénieur d'élevage ou de socioéconomie. Ce
fut un grand plaisir pour moi qui m'essayais à l'encadrement. Certaines
données collectées par eux ont été
exploitées dans ce rapport. Je leur dis merci et souhaite à
chacun d'eux plein succès dans ce qu'il entreprend au
quotidien.
Un grand merci à Monsieur Dominique DULIEU qui
m'a permis, grâce à son rôle de responsable scientifique
à ECOPAS, de prendre pied à la périphérie du W pour
la première fois. J'associe à lui Monsieur Philippe LHOSTE qui
m'a orienté vers l'INA P-G (aujourd'hui AgroParisTech) alors que je
cherchais une inscription en DEA et Monsieur Jean LOSSOUARN, Professeur
émérite à AgroParisTech, grâce à qui j'ai eu
cette inscription en DEA-EMTS, prémisse à cette thèse.
Alors que vous devriez participer au jury de soutenance de cette thèse,
des ennuis de santé ont rendu cette perspective impossible. Recevez mes
voeux de prompt rétablissement et merci pour votre disponibilité.
Je n'oublie pas Bernard TOUTAIN qui a suivi à distance les
péripéties de cette thèse et qui n'a pas cessé de
me présenter ses encouragements.
Loin des miens lors de mes séjours en France,
je n'étais pourtant pas seul. Je voudrais remercier toute la
communauté burkinabè à Orléans et Paris, en
particulier le couple BEOGO (Fabrice et Véronique) et enfants pour
l'assistance morale et matérielle que vous m'avez apportée.
Véro, merci pour les gâteaux. Bibi, gli ! gli ! gli.
Bien d'autres acteurs, non cités ici par omission
non voulue, ont participé, parfois activement, à cette aventure.
Qu'ils reçoivent ici mes sincères remerciements.
Un hommage posthume à KAOLO Konaté, qui
nous a quittés précocement. Il a facilité, par les moyens
octroyés et l'autorisation d'accès au parc W, la mise en route de
nos travaux.
ABRÉVIATIONS, ACRONYMES ET SIGLES
ACH Analyse ascendante hiérarchique
ACRA Association de Cooperation Rurale en Afrique et Amerique
latine
ADELE Programme d'appui au développement local à
l'Est
ADP Assemblée des Députés du Peuple
AFCM Analyse factorielle de correspondances multiples
AN Assemblée Nationale
ANOVA Analyse de variance
BDOT Base des données d'occupation des terres
BNDT Banque nationale de données terrestres
BUNASOLS Bureau national des sols
CBD Convention sur la Diversité Biologique
CC Capacité de charge
CGCT Code général des collectivités
territoriales
CGF Comté de gestion de la faune
Centre de coopération internationale en recherche
agronomique pour le CIRAD développement
CO2 Dioxyde de carbone
CORUS Coopération pour la Recherche Universitaire et
Scientifique
CRREA-Est Centre régional de recherches environnementales
et agricoles de l'Est
CSA Conseil scientifique pour l'Afrique au sud du Sahara
CSi Contribution spécifique de l'espèce i
CU Coefficient d'utilisation
CVD Conseil villageois de développement
°C Degré Celsius
DEA Diplôme d'étude approfondie
DF Disponible fourrager
DNMN Direction la météorologie nationale
dp Durée de la période
Direction provinciale de l'agriculture de l'hydraulique et des
ressources DPAHRH halieutiques
DRED-Est Direction régionale de l'économie et du
développement de l'Est
DREP Direction régionale de l'économie et de la
planification
ECOPAS Écosystèmes Protégés en
Afrique Soudano Sahélienne
EMTS Environnement: milieux, techniques,
sociétés
ETP Évapotranspiration potentielle
FAO Organisation des Nations Unies pour l'Alimentation et
l'Agriculture
FFEM Fonds français pour l'environnement mondial
FSi Fréquence de l'espèce i
Ga Groupements agrostologiques
GEF Global Environment Fund
GPS Global Positioning System

V
GRN/SP Gestion des ressources naturelles / Systèmes de
production
GTZ Deutsche Gesellschaft für Technische Zusammenarbeit
GmbH
ha Hectare
Ha Hypothèse alternative
ha/UBT/an Hectare par unité de bétail tropical et
par an
IGB Institut géographique du Burkina
INA-PG Institut national agronomique de Paris-Grignon
IndVal Valeur indicatrice
INERA Institut de l'environnement et de recherches agricoles
IRD Institut de recherche pour le développement
ISi Indice de qualité spécifique de l'espèce
i
IUCN Union mondiale pour la nature
Jrs Jours
kg kilogramme
kgMS kilogramme de matière sèche
km kilomètre
km2 kilomètre carré
LR/D Liaison recherche/développement
m Mètre
MAB Man And Biosphere
MATD Ministère de l'administration territoriale
MECV Ministère de l'environnement et du cadre de vie
MEE Ministère de l'environnement et de l'eau
mm Millimètre
mn Minute
MRA Ministère des ressources animales
MS Matière sèche
NEC Note d'état corporel
OCDE Organisation de Coopération et de
Développement Économique
ONG Organisation non gouvernementale
PADAB II Programme danois d'appui à l'agriculture au
Burkina, phase II
PADL/T Programme d'appui au développement local de la
Tapoa
pH Potentiel hydrogène
Pi Probabilité de l'espèce i
PIB Produit intérieur brut
PICOFA Programme d'investissement communautaire en
fertilité agricole
PM Premier Ministère
PRES Présidence
Programme Concerté pour le Développement de
l'Élevage en Afrique de PROCORDEL l'Ouest
RD Recherche-développement
RGPH Recensement général de la population et de
l'habitat
SIG Système d'information géographique
SOCOMA Société Cotonnière du Gourma

VI
SOFITEX Société des Fibres Textiles

VII
tMS Tonne de matière sèche
tMS.ha-1 Tonne de matière sèche à
l'hectare
Trp Troupeau
UBT Unité bétail tropical
UBT/ha/an Unité de bétail tropical par hectare et
par an
UNESCO Organisation des Nations Unies pour l'Éducation, la
Science et la Culture
UPP Unités paysagères pastorales hors aire
protégée
UPw Unités paysagères pastorales dans l'aire
protégée
UR Unité de recherche
VNIR Visible proche infrarouge
VP Valeur pastorale
WAP Complexe W-Arly-Pendjari
WAPOK Complexe W-Arly-Pendjari-Oti-Kéran
ZOVIC Zones villageoises d'intérêt
cynégétique

VIII
SOMMAIRE
DEDICACES I
REMERCIEMENTS II
ABRÉVIATIONS, ACRONYMES ET SIGLES V
SOMMAIRE VIII
INTRODUCTION GENERALE 1
CHAPITRE I. CONTEXTE GENERAL ET PROBLEMES DE RECHERCHE
6
1.1. LE CONTEXTE DE LA RECHERCHE: PASTORALISME, AIRES
PROTÉGÉES ET ESSOR DU COTON 7
1.2. PROBLÉMATIQUE ET JUSTIFICATION DE LA RECHERCHE 11
1.3. QUESTIONNEMENT SCIENTIFIQUE ET HYPOTHÈSES DE
RECHERCHE 12
1.4. OBJECTIFS 14
1.5. DÉMARCHE CONCEPTUELLE 15
CHAPITRE II. ETAT DES CONNAISSANCES : FONCTIONNEMENT
DES
ECOSYSTEMES SAVANIENS, SOCIOLOGIE DU PASTORALISME ET
EVOLUTION DE LA POLITIQUE DE CONSERVATION 22
2.1. NATURE ET TYPES DE SAVANES 23
2.2. LES ÉCOSYSTÈMES SAVANIENS : FONCTIONNEMENT ET
PERTURBATION 24
2.3. LES INTERACTIONS ENTRE LE BÉTAIL ET LA FAUNE SAUVAGE:
QUELLE POSSIBILITÉ DE COMPROMIS ? 37
2.4. LE PASTORALISME : UN GENRE DE VIE, UN RAPPORT PARTICULIER
À L'ESPACE 38
2.5. LES GRANDES ÉTAPES DANS L'HISTOIRE DE LA CONSERVATION
40
CHAPITRE III. CARACTERES PHYSIQUES, HUMAINS ET SOCIO
-
ECONOMIQUES DE LA PROVINCE DE LA TAPOA 43
3.1. LA ZONE D'ÉTUDE ET SA LOCALISATION 44
3.2. LE MILIEU BIOPHYSIQUE 46
3.3. LE MILIEU HUMAIN 59
3.4. LES SYSTÈMES D'ÉLEVAGE 60
3.5. LES AUTRES ACTIVITÉS SOCIO-ÉCONOMIQUES 64
CHAPITRE IV. LES UNITES PAYSAGERES PASTORALES DANS LE
TERROIR ET
DANS L'AIRE PROTEGEE VOISINE : DEFINITION,
DISTRIBUTION
SPATIALE ET CARACTERISTIQUES 68
4.1. INTRODUCTION 69
4.2. MATÉRIEL ET MÉTHODES 69
4.3. RÉSULTATS ET DISCUSSION 82
4.4. CONCLUSION 107
CHAPITRE V. LES SYSTEMES D'ELEVAGE A KOTCHARI : PRATIQUES
ET
STRATEGIES D'HIER ET D'AUJOURD'HUI 108
5.1. INTRODUCTION 109
5.2. MATÉRIEL ET MÉTHODES 110
5.3. RÉSULTATS ET DISCUSSIONS 116
5.4. CONCLUSION 169
CHAPITRE VI. LE TROUPEAU AU PATURAGE :
LOGIQUES,
REPRESENTATIONS ET REALITES DU TERRAIN 171

IX
6.1. INTRODUCTION 172
6.2. MATÉRIEL ET MÉTHODES 173
6.3. RÉSULTATS ET DISCUSSIONS 178
6.4. CONCLUSION 234
CHAPITRE VII. DISCUSSION GENERALE : SYNTHESE, CONCLUSION
ET
PERSPECTIVES 236
7.1. SYNTHÈSE 237
7.2. CONCLUSION GÉNÉRALE 246
7.3. PERSPECTIVES DE RECHERCHE FUTURES 256
REFERENCES BIBLIOGRAPHIQUES 258
TABLE DES MATIERES 293
LISTE DES CARTES 308
LISTE DES ENCADRÉS 309
LISTE DES FIGURES 310
LISTE DES PHOTOGRAPHIES 313
LISTE DES TABLEAUX 314
ABSTRACT 316
RESUMÉ 317
ANNEXES 318
ANNEXE 1. ENQUÊTE SUR LES PRATIQUES ET LA
CATÉGORISATION DES PÂTURAGES 319
ANNEXE 2. GUIDE D'ENTRETIEN PERSONNES RESSOURCES 326
ANNEXE 3. ESPÈCES HERBACÉES CARACTÉRISTIQUES
DES GROUPEMENTS AGROSTOLOGIQUES 330
ANNEXE 4. LISTE FLORISTIQUE HERBACÉE 331
ANNEXE 5. LISTE FLORISTIQUE LIGNEUSE 335

INTRODUCTION GENERALE

2
L'élevage est une activité
socio-économique d'importance majeure dans le monde, en particulier en
Afrique au Sud du Sahara, il assure à la fois des fonctions
macroéconomiques et environnementales.
Sur le plan économique, les populations de l'Afrique
subsaharienne, en particulier les plus pauvres, en dépendent comme
source d'aliments et de revenus monétaires (Zoundi & Hitimana,
2008). Par ailleurs, il constitue une assurance contre les risques pour ces
populations qui tirent leurs moyens d'existence de l'agriculture. A
l'échelle de la sous-région ouest-africaine, l'élevage
contribue en moyenne pour 40% du PIB agricole (PROCORDEL,
2005) et cette valeur peut atteindre 50% si on prend en compte
la force de travail et la fumure organique (Smith et al.
1996). Numériquement, la région
sahélienne compterait jusqu'à 25 % de bovins, 33 % d'ovins et 40
% de caprins de tout l'effectif de l'Afrique subsaharienne. Au Burkina Faso,
l'élevage constitue la deuxième ressource du secteur primaire
burkinabé, représentant 27,2% de sa valeur ajoutée (MRA,
2006), il est d'ailleurs en passe de passer en première
place1. Il participe effectivement à la constitution de 25%
du PIB agricole (Wane, 2006 ; Nori, 2007), de 12% du PIB national, de 18,6% des
recettes d'exportation et se révèle être une source de
revenu pour plus de 86 % de la population active et la première forme de
capitalisation pour la quasi-totalité des ménages ruraux et
même urbains (MRA, 2005) du pays. Le cheptel, numériquement
important et diversifié (MRA, 2004) se chiffrait en 2005 entre 7,2
millions (Wane, 2006) et 7,6 millions (MRA, 2006) de têtes pour la seule
espèce bovine.
Sur le plan environnemental, une grande partie de la
région, celle formant les États sahéliens, est faite de
zones arides et subarides et l'élevage y apparaît, en particulier
dans les contrées plus au nord, comme étant la forme de
valorisation la plus efficace (Wane, 2006 ; Nori, 2007). La zone renferme, en
effet, jusqu'à 38,2% de terres arides, écologiquement fragiles,
aux écosystèmes en déséquilibre et peu favorables
au développement de la production végétale (Zoundi &
Hitimana, 2008). Dans cette bande aride, la production animale est basée
sur les ressources naturelles (fourrage herbacés et ligneux) à
distribution fortement aléatoires (dispersées dans le temps et
dans l'espace) qui sont pâturées par le bétail. Cette
valorisation qui se fait selon un mode extensif à mobilité
parfois forte lui permettant d'exploiter les complémentarités
intra-zonales ou inter-zonales, est la preuve d'une adaptation du
système au contexte environnemental. Au-delà de la
mobilité, d'autres stratégies de valorisation de ces milieux
(diversification des espèces animales, dispersion des troupeaux et
maximisation de leur taille) (Dahl, 1983 ; Wane, 2006) sont mises en oeuvre et
nourrissent la conviction des spécialistes de la question pastorale (par
exemples Benoit, 1976, 1998 & 1999 ; Toutain, 1991 ; Boutrais, 1994 ; Daget
& Godron, 1995 ; Faure, 1997 ; Touré, 1994 & 1997; Lhoste, 2004
; Wane, 2006) sur la bonne adaptation socio-économique et
écologique de l'élevage à l'environnement originel de ces
zones. Les groupes pastoraux évoluent, en effet, dans des régions
où les conditions agro-écologiques et les caractéristiques
des ressources en pâturages ne sont favorables que de manière
très variable et imprévisible du fait des conditions
pédologiques et climatiques très contraignantes (Zoundi &
Hitimana, 2008). Elles sont, en effet, «
hétérogènes, fragmentées, liées aux
régimes pluviométriques
1 Les tendances indiquent cependant que le secteur
minier en particulier l'or pourrait occuper cette place au détriment de
l'élevage et du secteur agricole, ce dernier étant jusque
là à cette place, grâce au coton.

saisonniers, divergentes en fonction du temps (variables) et
globalement caractérisées par des conditions climatiques
capricieuses (imprévisibles) » (Nori et al. 2008).
Depuis toujours, l'élevage pastoral a été
une composante majeure de l'élevage au niveau mondial. Il se pratique
sur environ 25% des terres du globe, depuis les zones arides et subarides
d'Afrique (66% des terres du continent) et la péninsule arabique aux
hautes terres d'Asie et d'Amériques latine (Nori, 2006 & 2007 ; Nori
et al. 2008). D'après la FAO (in Nori, 2007), en 2001
cet élevage fournissait 10% de la production mondiale de viande et
faisait vivre quelques 200 millions d'exploitations familiales pour près
d'un milliard de chameaux, bovins et petits ruminants. Au Burkina Faso
où l'élevage est essentiellement de type extensif (MRA, 2004),
les systèmes pastoraux (à petite et grande
transhumance)2 pratiqués principalement par les ethnies
peules, forment la majorité du cheptel (70% du cheptel bovin selon Wane
(2006)), représentant 60 % des productions de viande (MRA, 2006).
La péjoration du climat en cours depuis les
années sèches de 70 et 80 dans les zones sahéliennes et
soudaniennes de l'Afrique au Sud du Sahara a, cependant, profondément
bouleversé l'équilibre instable (Breman & De Ridder, 1991 ;
Daget & Godron, 1995) qui avait cours dans les environnements pastoraux
marquant ainsi les systèmes de vie des populations, notamment des
peuples pasteurs (Santoir 1999, Wittig et al. 2007). Si la
transhumance, système pastoral dominant dans la région,
caractérisait depuis toujours les systèmes de production de ces
sociétés, sa forme et ses manifestations se sont trouvées
ainsi profondément modifiées et tout indique que la tendance va
se poursuivre. Alors que par le passé les transhumances vers le sud de
la sous-région étaient moins importantes que celles vers le nord,
c'est plutôt l'inverse qui est observé de nos jours.
D'après Kagoné (2000), le domaine soudanien était
historiquement celui des communautés agraires sédentaires, les
éleveurs peuls de bovins zébus ne s'y aventurant que pour la
transhumance de saison sèche. De nos jours, ceux-ci investissent donc de
nouveaux territoires, notamment vers le sud dans une zone
considérée désormais comme plus clémente qui
était encore, dans un passé récent, peu utilisée
bien que relativement bien pourvue en ressources notamment fourragères
(Daget & Godron, 1995 ; Boutrais, 1990 & 1996 & 2007 ;
Kagoné, 2000 ; Convers, 2002). Les éleveurs évitaient
autrefois la partie méridionale de la région pour plusieurs
raisons : faible qualité des ressources pastorales comparées
à celles de la région strictement sahélienne (Breman et De
Ridder, 1991), risques en particulier sanitaires (présence
endémique des simulies et des glossines, vecteurs respectifs de
l'onchocercose et de la trypanosomose) (Carrière & Toutain, 1995;
Daget & Godron, 1995 ; Benoit, 1998 ; Kagoné, 2000) et risques plus
élevés de conflits avec les agriculteurs.
Face aux crises alimentaires récurrentes vécues
en zones sahéliennes, les éleveurs n'ont donc plus peur
d'affronter ces risques pourtant avérés mais qui sont en recul
dans la région nord-soudanienne (Kagoné, 2000). Bon nombre
d'entre eux s'y installent (phénomène de sédentarisation)
et, pour ceux qui ont connu une grande décapitalisation de leur cheptel,
s'adonnent plus ou moins fortement à l'agriculture (Santoir, 1998 ;
Kaboré, 2010). De nos jours, la part des agropasteurs et agro
éleveurs augmente au détriment de celle des pasteurs
2 L'élevage nomade, autre composante du
système pastoral, est marginal au Burkina, Il est cantonné
à l'extrême Nord-est du pays (Oudalan).

4
purs (Bonfiglioli, 1992 ; Seré, 1994 ; Smith et
al. 1997 ; Zoundi & Hitimana, 2008). Ces systèmes
agropastoraux sédentaires ou systèmes mixtes
agriculture-élevage qui combinent agriculture et élevage pour
l'autoconsommation et pour la vente, ont connu une croissance importante, en
raison de cette sédentarisation, de la poussée
démographique (cause d'extension des surfaces cultivées) et des
mutations socio-économiques (individualisation et pluriactivité
des exploitations, baisse des complémentarités, etc.) (Daget
Godron, 1995 ; Barrière, 1996), du transfert de propriété
du cheptel et des difficultés et conflits liés à la
transhumance (Zoundi & Hitimana, 2008) en zone soudanienne.
Tout ce processus a abouti à l'accroissement notable
des effectifs animaux dans les zones nord et sud soudaniennes des États
de la sous-région. Cela a été d'autant plus possible que,
le rôle de l'élevage dans l'économie ne cessant de prendre
de l'ampleur, nos États ont été poussés à
imaginer tant bien que mal des politiques de soutien (campagne de vaccination
à grande échelle, hydraulique pastorale, politique de
sédentarisation par le ranching, cultures fourragères, etc.)
(Touré, 1994). Si les mesures proposées n'ont pas toujours fait
la preuve de leur pertinence (Touré, 1994 & 1997), elles sont au
moins une indication de l'intérêt que les politiques portent au
secteur de l'élevage en général. Ces politiques, outre
qu'elles ne sont parfois pas adaptées ou qu'elles s'inscrivent peu dans
une perspective de long terme (Bourbouze et al. 2001), sont
généralement mises à mal du fait de
«contraintes» réelles de terrain pour l'élevage mobile
dont les principales sont une expansion démesurée du front
agricole et la mise en place d'aires protégées de mieux en mieux
surveillées (Sournia, 1987 ; Yaméogo, 2005 ; Kaboré,
2010). Or ces deux éléments sont définitivement inscrits
dans les réalités de nos pays, du moins tant qu'aucune politique
de rupture n'intervient. L'expansion de l'agriculture s'inscrit dans une
dynamique naturelle qui accompagne l'accroissement démographique tandis
que les actions de conservations relèvent d'une tendance mondiale de
préservation des écosystèmes et de la biodiversité
utile dont le taux d'extinction actuel de 10% laisse présager un
déclin très préoccupant comme le fait observer
Kièma S. (2007). Tout ceci a l'inconvénient d'exposer les
pâturages soudaniens à la surexploitation et compromet, par effet
boomerang, la survie de l'élevage. Il est donc clair que, même si
l'État exprime une politique volontariste à l'égard de
l'élevage, ce dernier est contraint de s'adapter et cela exige, de la
part des éleveurs, la mise en oeuvre de pratiques pastorales
«meilleures» (ou bonnes) sur les rares et pauvres terres communes
toujours accessibles.
Cette réalité générale
décrit parfaitement les évolutions en cours dans l'Est du
Burkina, en particulier dans la Tapoa, une province qui connaît un grand
dynamisme agricole et dont une grande partie (33% selon ACRA (2009)) est
couverte de nombreuses réserves de faunes totales et partielles. Cette
province a été et demeure toujours une destination
privilégiée pour les pasteurs sahéliens venant du nord du
pays et du Niger (Paris, 2002). Les éleveurs y sont attirés par
une disponibilité en ressources naturelles qui serait meilleure que dans
leurs zones d'origine, mais la région représente aussi une zone
de passage pour les pasteurs en transit vers le Togo et surtout le
Bénin. Toutain et al. (2001), Paris (2002) et Kaboré
(2010) indiquent en outre que la possibilité de
pénétration illégale dans le Parc du W expliquerait aussi
la préférence des pasteurs en transit ou en accueil saisonnier
pour certaines zones de la province de la Tapoa. Le terroir de Kotchari est
l'une de ces destinations privilégiées et on peut s'interroger
sur sa capacité à supporter durablement ce niveau

d'emprise. De fortes concentrations animales y sont en effet
enregistrées et la sécurité de l'élevage et des
groupes pastoraux de même que l'intégrité des ressources
naturelles, y semblent menacées dans l'avenir.
La présente recherche se propose de contribuer,
à l'échelle de l'espace d'un terroir, celui de Kotchari dans la
province de la Tapoa, à élucider les dynamiques qui y ont cours
et de proposer des interventions sur le système global qui pourraient
aider à sécuriser le système pastoral et l'atteinte des
objectifs des actions de conservation de la nature.
Le manuscrit se structure de la manière suivante, en sept
chapitres:
Dans le premier chapitre, nous décrivons la
problématique telle qu'elle se pose à la province de la Tapoa et
à notre terroir puis exposons comment nous comptons l'aborder. Ensuite,
dans le chapitre II, nous exposons les connaissances déjà
disponibles, sur lesquelles nous allons nous appuyer pour aborder les questions
d'interactions société/nature. Dans le troisième chapitre,
nous donnons un aperçu sur la zone d'étude par une description au
triple plan physique, humain et socio-économique. Dans le chapitre IV,
nous caractérisons les ressources pastorales notamment
fourragères (carte pastorale écologique) et évaluons les
risques éventuels auxquels elles s'exposent ainsi que les
opportunités et les difficultés de leur valorisation par le
bétail. Dans le chapitre V les pratiques et stratégies des
éleveurs et leur évolution sur une vingtaine d'années ont
été analysées en mettant l'accent sur trois aspects: (i)
la pression démographique, (ii) la pression animale et (iii) les
pratiques des éleveurs. Au préalable, un profil des
éleveurs présents ou fréquentant le terroir (typologie) a
été réalisé, de là les pratiques ont
été suivies pour comprendre les stratégies qui les
sous-tendent. Dans le chapitre VI, pour bien appréhender les
connaissances des éleveurs sur leurs ressources (carte pastorale
participative), leur manière de catégoriser et de classifier les
milieux qu'ils exploitent de même que les logiques en jeu, nous avons par
enquête et suivi de troupeaux au pâturage, analysé la
manière dont ils font exploiter lesdites ressources par leurs troupeaux
et les stratégies qui les sous-tendent. Cela a permis par ailleurs, de
voir comment les éleveurs mettent en oeuvre leurs connaissances et
d'identifier les contraintes qui se présentent à eux.
Enfin, dans le chapitre VII, après une brève
synthèse des principales conclusions de la recherche, nous faisons, des
propositions pour un système durable d'exploitation pastorale s'appuyant
sur les informations pertinentes fournies par la recherche avec pour
préoccupation de permettre une cohabitation viable entre les
activités pastorales et les réserves en particulier le complexe
du W.
CHAPITRE I

6
CONTEXTE GENERAL ET PROBLEMES DE
RECHERCHE

7
1.1. Le contexte de la recherche: pastoralisme, aires
protégées et essor du coton
1.1.1. La région de l'Est et l'élevage
pastoral : entre perspectives et incertitudes
L'histoire de l'élevage dans le Gurma3 est
intimement liée à celle de l'installation et de l'expansion des
peuples pasteurs peuls dans la région qui se sont faites d'Ouest en Est
(Benoit, 1998 & 1999a). Cette région que Santoir (1998)
décrit comme de « vastes, riches et vides savanes » (jusqu'au
début du 19ème siècle, la densité
humaine y était d'environ 2-3, habitants/km2), a
attisé la convoitise des Peuls qui tentaient déjà de s'y
établir dès le 15ème siècle. A cette
période déjà, les territoires avoisinant du Moogo, du
Yagha, de Torodi et du Gueladio connaissaient des charges humaines ou
pastorales élevées. Certains facteurs comme les razzias, les
attaques de fauves et les risques sanitaires (avec la peste bovine, la
trypanosomose et l'onchocercose) vont limiter cette affluence peule à
quelques zones de forte concentration humaine (Zones de Diabo, Tibga,
Comin-Yanga et autour du Gobnangou), le reste de la région situé
entre la rive droite du fleuve Niger et le Borgou au Bénin, constituant
ce que Benoit (1998) a appelé un « no man's land ». À
ce propos, l'auteur fait remarquer que « jusqu'en 1937 on ne signalait pas
une présence de bétail dans la rive droite du fleuve Niger
». Les premiers éleveurs peuls, dont les effectifs animaux sont peu
importants, vont rapidement se sédentariser4 et pratiquer
l'agriculture tandis que les autochtones gourmantchés, essentiellement
agriculteurs, continuent d'entretenir de petits effectifs de bétail peu
intégré à l'agriculture et servant essentiellement
à payer la dot et la liberté (Santoir, 1998).
Le phénomène de transhumance existait alors
déjà, mais il se limitait à de faibles déplacements
d'animaux à l'intérieur de la zone sahélienne surtout pour
chercher de l'eau. Santoir (1999) rapporte en effet, que jusqu'en janvier 1973
les transhumants sahéliens ne dépassaient guère le sud du
Yagha.
À partir de la première moitié du
19ème siècle, de nombreux facteurs vont favoriser
l'occupation intégrale de l'espace et une forte pression sur les
ressources toujours en cours de nos jours. En effet, grâce à des
recherches sur les systèmes d'élevage essentiellement
orientées vers l'amélioration génétique et des
conditions sanitaires, la lutte contre les grandes épizooties commence
à porter ses fruits (Carrière & Toutain, 1995; Benoit, 1998).
Cette situation nouvelle, ajoutée à la baisse des attitudes
guerrières, esclavagistes et génocidaires de l'époque
(Benoit, 1999a) va favoriser une croissance du cheptel local. Elle permet aussi
un début d'affluence de transhumants des zones septentrionales qui se
gardent toutefois de franchir la rivière Tapoa, bien que le parc
national du W ait fait l'objet d'une exploitation pastorale dès son
classement en 1954, (Kagoné, 2004, comm. pers.). Cette
affluence perdurera
3 Le Gurma correspond à l'origine à
un vaste territoire allant au-delà des frontières du Gourma
actuel (ou Gulmu) qui comprend les Provinces de la région administrative
de l'Est (Gourma, Gnagna, Kompienga, Komondjari, Tapoa) dont la superficie
globale est de 46256km2 (DRED-Est, 2005).
4 On signale tout de même la présence de
pasteurs aux gros effectifs dans la zone de Kantchari-Botou

jusqu'aux années 70 où, à la faveur des
grandes sécheresses (1973-74 et 1983-84)5, les flux
migratoires du cheptel sahélien vont accompagner l'occupation, devenue
plus intensive, par le cheptel local, de l'espace méridional plus au sud
désormais moins hostile. La rivière Tapoa n'est franchie par les
animaux transhumants qu'à partir de 1984 (Benoit, 1998 & 1999a).
De nos jours, la région a acquis un caractère
nettement agropastoral et grâce à une densité humaine
longtemps restée faible et à une rigoureuse politique de
conservation, elle offre un potentiel fourrager relativement intéressant
en qualité et en quantité. Ceci a fait d'elle une zone de
convergence des nombreux transhumants sahéliens (Nigériens et
Burkinabés du Nord) qui descendent de plus en plus au sud (Boutrais,
1994 ; Benoit, 1999b). Par ailleurs, l'explosion
démographique6, la diversification des activités au
sein des ménages gourmantchés qui s'intéressent de plus en
plus à l'élevage, le retour en force de la culture du coton dans
la région en 1996, sont causes aujourd'hui d'un effectif impressionnant
de bétail7 menaçant l'équilibre
écologique des parcours, les actions de conservation et, à terme,
la viabilité même du système pastoral. Selon Benoit (1998),
la pression pastorale sur la réserve de biosphère du W est un
fait majeur d'échelle internationale s'exerçant dans un contexte
de saturation générale de l'espace par le bétail.
Actuellement, deux modes d'élevage se côtoient
dans la région : le mode sédentaire et celui transhumant (Guibert
& Prudent, 2005). Le premier est celui des résidents
gourmantchés, mossi et de quelques Peuls, il est le plus important en
termes d'unité bétail tropical (UBT). Le second est celui des
pasteurs peuls qui séjournent de plus en plus longtemps sur place.
1.1.2. Les aires protégées de l'Est : un
potentiel en sursis ?
Comme on l'a déjà mentionné, un des
atouts de la région de l'Est est de disposer de plus d'espaces riches en
ressources pastorales, forestières et fauniques que d'autres
régions du pays. Autour du 14ème siècle,
à cause des attitudes hostiles qu'exprimaient entre eux les peuples de
la région, l'occupation de l'espace était contrastée. Les
espaces vides qualifiés de « no man's land » par Benoit (1998
& 1999a) vont être l'objet de grignotage au début du
20ème siècle grâce à
l'amélioration des conditions sanitaires et sécuritaires
(Santoir, 1998). La création des premières aires
protégées semble être une réaction de la puissance
coloniale face à l'occupation anarchique de l'espace, ainsi
pacifié, qui s'opérait. Il s'est finalement agi de mettre de
grandes portions de forêts hors de portée de l'occupation qui se
faisait menaçante pour elles avec le glissement progressif et
l'arrivée des populations des zones de forte concentration de l'Ouest
vers le Sud-est. Contrairement à l'Ouest du pays, l'érection des
aires de protection de la faune à l'Est, bien que motivée par le
même désir d'empêcher une grande déforestation du
pays et par des considérations esthétiques et sanitaires comme le
rapporte Kièma S. (2001 & 2007), n'a pas été
précédé de grands déguerpissements8. De
nos jours, la
5 En réalité, les années
sèches de 73-74 et 83-84 font partie de la dernière et plus
longue période sèche parmi les trois (1909-1919; 1938-1949;
1968-1985) que le pays et l'ensemble de la sous région ont connu
(Sournia, 1987)
6 A la croissance naturelle (naissances), il faut
ajouter les fortes migrations d'agriculteurs venant du plateau central du pays
à partir des années 70 et surtout 80. Entre 1970 et 1996 la
progression annuelle de la population est de 4,65% (Guibert & Prudent,
2005).
7 Les agriculteurs gourmantché capitalisent
leurs revenus provenant du coton dans l'achat de bétail.
8 Cependant, Kaboré (2010) a noté des
vagues de déguerpissements dans la zone de la réserve partielle
de faune de Pama Nord (Province du Gourma).

9
région de l'Est est la zone de plus grande
concentration d'aires protégées du pays (parcs, réserves
totales et partielles, ranchs, concessions, etc.). En effet, alors que
seulement environ 5 à 10,6 % du territoire national (Spinage &
Traoré, 1984 ; Sournia, 1987 ; Yaméogo, 2005) sont occupés
par des aires de protection de faune, elles représentent dans cette
région jusqu'à 20 à 25%9 des superficies
(Guibert & Prudent, 2005; Traoré, 2008).
Les politiques de conservation mises en oeuvre dans la
région ont suivi la tendance d'ensemble au niveau de l'Ouest africain.
Un peu partout en Afrique, on est passé successivement de l'exclusion
à la participation puis à la concertation (Aubertin, 2005),
cependant le rythme n'a pas été le même partout.
Contrairement à l'Afrique australe et orientale où
l'intérêt de l'approche de gestion participative a
été très tôt appréhendé,
l'interdiction a très longtemps prévalu en Afrique de l'Ouest
(Bayer & Ciofolo, 2004).
En Afrique de l'Ouest, pendant la période coloniale,
l'administration a mis en place des sanctuaires de faune d'où
étaient exclues les populations locales (Babin et al. 2002). Il
en a été de même dans la région de l'Est du Burkina
Faso, du parc refuge du W créé en 1926 puis des réserves
totales d'Arly (1954), de Singou (1955) et de réserves partielles comme
la Kourtiagou (1957). La mise en place de ces différentes aires a
enlevé aux populations locales le droit de regard et de jouissance de
leurs ressources en même temps que les couloirs de passage et zones de
pâture des animaux étaient occupés. Les milieux, ainsi mis
«sous cloche», n'étaient alors pas encore sous forte pression
anthropique.
A partir des indépendances, les nouveaux États
réaffirment la propriété étatique des forêts
(Babin et al. 2002). Mais après les années sèches
de 73-74 et de 83-84, ces sanctuaires sont très menacés Les
animaux transhumants franchissent pour la première fois la Tapoa, la
migration agricole est forte dans la région et les États,
manquant de moyens, n'assurent pas une surveillance adéquate des espaces
dont ils ont retiré la surveillance aux populations locales. Dans une
étude minutieuse, Kaboré (2010) explique comment
l'espace-ressource villageois et inter-villageois était régi par
les lignages (des maîtres fonciers) qui en assuraient alors la "bonne"
gestion. De fait, ces espaces qui ne sont plus de nos jours formellement sous
administration coutumière, sont laissés à eux-mêmes
et sont devenus à "accès libre" pour les braconniers et surtout
pour le bétail. De nombreux travaux dont ceux de Kièma S. (2001
& 2007), Toutain et al. (2001), Convers (2002), Paris (2002),
Riegel (2002), Boutrais (2008) et Kaboré (2010) montrent que les aires
protégées entrent depuis longtemps dans les stratégies
alternatives trouvées par les éleveurs transhumants mais aussi
sédentaires, pour faire face à la période de soudure de
saison sèche. Aussi, Sournia (1987) rapporte qu'une bonne part des
quelques 400 000 têtes de bétail transhumant ayant transité
dans la région en 1985, a pénétré dans les aires de
protection et notamment dans le parc du W. Plus récemment, un
recensement aérien a dénombré 30 000 à 50 000
têtes de bétail dans le parc du W en 1994 (IUCN, 1994). Par
ailleurs, deux recensements encore plus récents réalisés
en mai 2002 (Riegel, 2002) et avril-mai 2003 (Bouché et al.
2003) confirment cette tendance. Le recensement de mai 2002, indiquait la
présence d'environ 23 840 bovins et 1 254 petits ruminants dans le parc
W, surtout du côté Bénin. Le recensement des mois d'avril
et de mai 2003 sur l'ensemble du
9 Il est important de noter qu'en
réalité ces aires se concentrent seulement sur les territoires
des provinces les plus méridionales que sont le Gourma et surtout la
Tapoa et la Kompienga.

10
complexe WAPOK (W-Arly-Pendjari-Oti-Kéran) a permis
d'enregistrer 1 171 troupeaux de bovins totalisant 101 309 animaux dans ce
complexe WAPOK et sa périphérie proche. On note même une
tendance à la sédentarisation d'éleveurs comme c'est le
cas dans le village d'Illéla, dans la partie béninoise du W
(Kagoné, 2004). Si l'exploitation pastorale du W reste une
réalité, son ampleur serait en baisse surtout pendant la
période d'exécution du programme ECOPAS
(Écosystèmes Protégés en Afrique Soudano
Sahélienne) (2001-2008) (Fournier & Toutain, 2007) et les conditions
biologiques dans les réserves de la région de l'Est restent
relativement meilleures qu'ailleurs au Burkina, notamment dans l'Ouest comme le
rapporte Kièma S. (2001).
L'élan participatif de la conservation ne prend
réellement naissance au Burkina qu'en 1984 lors du séminaire
national sur la faune, où il est proclamé que celle-ci est
désormais l'affaire de tous (Sournia, 1987). Dans les faits, pour ce qui
est de la région de l'Est et surtout du W et des réserves
avoisinantes, il faut attendre 2001 avec la mise en place du programme ECOPAS
(Écosystèmes Protégés en Afrique Soudano
Sahélienne) pour voir un début de concrétisation de cette
volonté politique10. Ce programme, centré sur les
trois parcs nationaux contigus du W, a été conçu par les
États concernés (Bénin, Burkina, Niger) avec l'appui de
l'Union Européenne à travers le Fonds Européen de
Développement. Son objectif étant «d'arrêter et
d'inverser le processus de dégradation des ressources naturelles du parc
afin de protéger de façon durable la biodiversité, au
bénéfice des populations concernées», il a
basé son principe d'intervention sur la participation consistant en
l'implication et en la responsabilisation des acteurs locaux. Un des premiers
résultats de ce programme est le renforcement notable de la surveillance
du W, devenu, entre temps, en 1996 site du patrimoine mondial de l'UNESCO et en
2002 réserve de biosphère (Paris, 2002 ; Riegel, 2002), avec en
compensation la mise en place d'actions socio-économiques dans la zone
de transition.
Mais devant l'avancée très rapide et anarchique
du front agricole liée à la croissance naturelle de la
population, l'immigration agricole et la forte orientation des systèmes
de production au marché avec l'arrivée du coton, l'effectif
impressionnant du cheptel local et l'afflux de plus en plus massif de
bétail sahélien qui fuyait des conditions de plus en plus
insupportables, les actions du programme ECOPAS ont semblé
insignifiantes aux yeux des populations. Dans la zone de transition du W du
Niger les conflits observables sur le terrain entre les acteurs (conservateurs,
populations locales et transhumants) ont été nombreux et divers
(Paris, 2002 ; Kagoné, 2004; Sawadogo, 2004). En toile de fond se
trouvaient des différences de vue radicales sur le rôle des aires
protégées (réservoirs de biodiversité pour les uns,
vastes et riches terres ou encore stock de fourrage pour les
autres)11. Le même type de représentation de la part
des populations locales envers les aires de protection a été
observé par Kièma S. (2001), Berlin (2002) et Goungounga (2003)
dans l'Ouest du Burkina.
10 Des tentatives de gestion commune ont cependant eu
lieu dès les années 60 avec peu de succès sauf du
côté nigérien.
11 Voir aussi Kaboré (2010) pour plus de
détails sur les représentations et les revendications des
populations environnantes à l'égard des aires
protégées. Binot et al. (2006) ainsi que Harchies et
al. (2007) notent d'ailleurs qu'un peu partout en Afrique il y a une
opposition basique entre aires à vocation de protection et aires
à vocation de production.

1.1.3. Des mutations agricoles porteuses de risques pour
les équiibres socio-économique et environnemental.
A partir des périodes sèches des années
soixante dix et quatre vingt, un certain nombre de mutations tant agricoles que
pastorales ont été observées dans l'Est du Burkina Faso.
En effet, les animaux en transhumance ne s'arrêtaient plus au nord de la
rivière Tapoa. Désormais ils descendaient plus au sud
jusqu'à une limite méridionale qui se situait au Bénin et
au Togo, au-delà des frontières nationales. Par ailleurs, le
retour en force en 1996/9712 et l'explosion de la culture du coton
était patent dans la région et notamment dans la province de la
Tapoa. Les productions annuelles y sont passées de 2600 tonnes environ
en 1996 à 36513 tonnes en fin de campagne 2004/2005, puis elles ont
amorcé une baisse. Une croissance aussi rapide a eu pour corollaire des
défrichements à grande échelle qui ont
accéléré l'avancée du front agricole. De plus, les
exploitations agricoles sont devenues pluriactives et de plus en plus
orientées vers le marché. Ceci, en réduisant les
complémentarités à l'intérieur des familles ou
entre les familles, a fragilisé le tissu social. En outre, les relations
de réciprocité séculaires, qui prévalaient encore
naguère entre les communautés d'agriculteurs et d'éleveurs
(Thébaud, 1995 ; Boutrais, 1999a) ont sensiblement perdu de leur poids.
Par ailleurs, cette fragilisation du tissu social et le rapprochement plus
accentué au marché ont provoqué le morcellement des
parcelles agricoles et la compétition pour le contrôle de
l'espace-ressource (Barrière & Barrière, 1997)13
menaçant ainsi la cohésion sociale, l'intégrité des
parcours et celle des aires protégées. En même temps que
cette évolution des pratiques de transhumance et que l'essor des
cultures de rente notamment du coton, un transfert de propriété
du cheptel s'est opéré (Paris, 2002 ; Sawadogo, 2004). Toute la
région est désormais peuplée en majorité de
communautés d'agropasteurs ou d'agroéleveurs du fait de la forte
propension des agriculteurs à pratiquer l'élevage, notamment de
petits et grands ruminants. Pour leur part, de nombreux Peuls se
sédentarisent en pratiquant l'agriculture. Enfin, les mesures sanitaires
prises depuis les premières années des indépendances ont
permis d'endiguer les simulies et les glossines (Toutain et al. 2001;
Paris, 2002; Convers, 2002 ; Thébaud, 2002), vecteurs respectifs de
l'onchocercose et de la trypanosomose ; elles ont favorisé la pratique
de l'élevage.
1.2. Problématique et justification de la
recherche
L'élevage pastoral est un système qui consomme
beaucoup l'espace. Il est le mieux adapté aux conditions des
régions climatiques arides et semi-arides dans lesquelles la
pluviosité et les ressources pastorales sont sujettes à une forte
variabilité spatiale. Il est d'ailleurs maintenant reconnu et
accepté que la mobilité des animaux constitue une
nécessité écologique dans ces zones (Benoit, 1976 ;
Toutain, 2001 ; Touré, 1997). Ce système multiséculaire,
qui s'est forgé et affiné dans le temps, est très
cohérent : la mobilité très variable des animaux leur
permet d'accéder à des ressources très variées par
leur nature et leur
12 Cette culture existait déjà
traditionnellement et a même été l'objet de promotion en
1978 par le Projet de Développement Agricole Intégré
(PDAI) avec des résultats mitigés (Guibert & Prudent,
2005).
13 Barrière & Barrière (1997)
prévoient que dans un contexte de morcellement des unités de
production, la compétition pour le contrôle de l'espace-ressource
est inévitable.

12
quantité, mais dispersées dans l'espace. De nos
jours, quelle que soit l'échelle d'analyse adoptée, l'espace et
donc les ressources qui lui sont liées se raréfie et en vient
même à manquer dans certains terroirs où il devient l'objet
de compétitions rudes entre différents groupes
d'intérêts.
Nos travaux antérieurs (Sawadogo, 2004) ont
montré que les Peuls, majoritaires parmi les éleveurs dans la
région du parc W, sont considérés comme des
étrangers dans les terroirs qu'ils habitent, même lorsqu'ils sont
sédentaires. Et comme tels, le droit d'appropriation de
l'espace-ressource (Barrière & Barrière, 1997) et parfois
même d'y accéder leur est souvent dénié. Cette
constatation avait déjà été faite par d'autres
auteurs notamment Thébaud (1995) et Kaboré (2010) ailleurs au
Burkina et au Niger. On peut craindre (Ouédraogo, 2000) qu'avec la
décentralisation qui vient d'être mise en route, cette situation
n'empire, en prenant cette fois des formes légales.
On le voit donc, le problème majeur est une menace sur
la viabilité même de l'élevage pastoral et des
écosystèmes sur lesquels il repose. Pourtant la subsistance de ce
type d'élevage reste nécessaire sur un plan économique et
éthique, certains arguments écologiques soutiennent d'ailleurs
l'idée qu'à un niveau de charge raisonnable le bétail
participe à l'entretien des milieux (Boudet, 1991; Steinfeld et
al. 1997).
Au plan économique, rappelons que l'élevage
burkinabé, qui est surtout pastoral, est une activité motrice de
l'économie nationale, tant par sa contribution à la croissance du
Produit Intérieur Brut que par son rôle dans l'équilibre de
la balance commerciale (respectivement 12% et 18,6%). Il intervient aussi dans
l'amélioration des conditions de vie des populations : 86% d'entre elles
en tireraient une part non négligeable de leur revenu (MRA, 2005).
Au plan éthique, le pastoralisme et notamment la
transhumance constitue un système de vie (Daget & Godron, 1995 ;
Boutrais, 1997 ; Wane, 2006) qui est comme tout autre, respectable. Il serait
donc légitime qu'un droit soit reconnu aux peuples pasteurs (les Peuls
dans ce cas) pour qu'ils puissent continuer à le pratiquer en l'adaptant
cependant aux conditions du moment.
1.3. Questionnement scientifique et hypothèses
de recherche
Le terroir de Kotchari comme la plupart des terroirs riverains
du parc W voit son effectif en bétail s'accroître
considérablement en saison sèche du fait de la venue massive des
transhumants (Toutain et al. 2001 ; Paris, 2002 ; Sawadogo, 2004). Par
ailleurs, depuis l'arrivée de la culture cotonnière en 1996/97,
la tendance dans tous les terroirs de la zone est à l'occupation des
espaces jadis considérés comme incultes et exploitées par
le bétail local et transhumant.
Question 1 : Dans ces conditions, quel est le
niveau de saturation de l'espace et quelle place est offerte à
l'élevage notamment pastoral dans le terroir et ses environs ?
Hypothèse 1: L'afflux de
nombreux troupeaux transhumants qu'on observe chaque année sur le
terroir de Kotchari et ses environs ne s'explique pas, comme il est souvent
dit,

par la disponibilité en ressources pastorales dans
les espaces légalement accessibles (périphérie du Parc).
L'intérêt pastoral du terroir de Kotchari est plutôt
lié à l'opportunité qu'il peut offrir d'accéder
illégalement aux ressources du Parc du W
L'élevage pastoral est « un modèle constant
parmi les bergers peuls de la zone savane» (Stenning, 1959). Selon
Boutrais (1997), les pasteurs, face aux changements globaux qui
s'opèrent depuis maintenant quelques décennies, mettent en place
des stratégies d'adaptation. Ainsi par exemple, Convers (Com. pers,
2006) a relevé qu'en réponse à la surveillance accrue
des aires protégées dans la région du parc W qui
résulte de la mise en place du programme ECOPAS, les éleveurs
transhumants ont adopté trois types de trajectoires: (i) la capitulation
avec changement d'activité, (ii) l'adaptation par la mise en place de
stratégies nouvelles tendant à exclure la pâture
illégale dans les aires protégées et (iii) la
résistance. On peut penser que ce type de réaction est aussi
celui des autres catégories d'éleveurs identifiées dans la
région du terroir de Kotchari.
Question 2 : Quelles sont les décisions
que prennent les éleveurs face aux changements globaux et comment les
mettent-ils en oeuvre dans la région de Kotchari ?
Hypothèse 2: Les
systèmes d'élevage sédentaires ou mobiles subissent des
mutations perceptibles au travers des pratiques des éleveurs qui se
modifient pour s'adapter aux nouvelles conditions locales.
Question 3 : Sur quelle évaluation et
représentations du milieu s'appuient les stratégies quotidiennes
et saisonnières des éleveurs à l'échelle locale?
Hypothèse 3.1 : Dans une
localité donnée, les éleveurs évaluent et classent
les pâturages sur des critères écologiques (qualité
pastorale du moment), mais aussi en termes de risques de conflits, de risques
sanitaires, etc. L'évaluation et donc la classification d'un milieu
donné change en fonction des périodes de l'année.
Hypothèse 3.2 : Dans une
localité donnée, le choix des itinéraires par les animaux
et/ou leurs bergers repose sur cette évaluation/classification locale
qui croise une classification des milieux végétaux et une
échelle de risque. Il se fait en fonction de la distribution
spatiotemporelle et de la valeur pastorale des ressources
végétales ainsi que du niveau d'exposition aux différents
risques évoqués.
Au Burkina comme dans tous les pays de cette partie de
l'Afrique, l'élevage pastoral reste tributaire des ressources naturelles
pour la satisfaction de ses besoins alimentaires. Ces ressources naturelles,
à cause de la forte variabilité climatique saisonnière et
interannuelle, sont inégalement réparties dans l'espace et le
temps. Pour assurer la survie de leurs animaux, les éleveurs adoptent
des stratégies d'exploitation opportuniste des ressources
fourragères là où elles se trouvent, ce qui requiert une
grande mobilité dans l'espace. En effet, à certaines
périodes de l'année ou lors d'années à conditions
climatiques difficiles, les ressources se trouvent confinées dans
certaines régions ou certains espaces particuliers (bas-fonds
notamment). Ces «poches de ressources » assurent un rôle
déterminant dans la survie du bétail aux moments cruciaux de
l'année. L'accès à ces milieux, parfois qualifiés
de « ressources clefs » (Hatfield & Davies, 2006) ou de «
ressources stratégiques » ou encore de « filets de
sécurité » (Pratt & Gwynne, 1977.) est, en effet,
indispensable au fonctionnement des

14
systèmes pastoraux qui, autrement, s'effondreraient.
Ces auteurs classent les bas-fonds, les plaines d'inondations ou
marécages et les réserves sylvopastorales dans la
catégorie de ressources clefs. Dans le contexte actuel de forte pression
foncière, ces ressources sont cependant menacées de disparition
ou rendues inaccessibles pour les animaux dans de nombreuses contrées du
pays, ce qui pousse les éleveurs à la transhumance, soucieux
qu'ils sont du bien être de leurs troupeaux (Toutain et al. 2001
; Paris, 2002 ; Kagoné, 2004). Cette pression foncière sur ces
milieux particuliers, s'est accrue ces dernières années avec la
multiplication de projets de « petite irrigation villageoise ».
Grâce aux aménagements rendus possibles par les subventions, ces
projets, permettent aux populations d'occuper les abords des points d'eau
naturels ou non et d'y pratiquer une culture de saison sèche (ou de
contre saison). Ce type d'utilisation des milieux clés pour
l'élevage ne semble pas, pour le moment, toucher la région de
Kotchari, il faudrait cependant en préciser la raison : simple retard,
caractéristiques du milieu naturel ou causes culturelles ou sociales
?
Question 4 : Le terroir de Kotchari
recèle-t-il de ressources considérées comme essentielles
par les éleveurs?
Hypothèse 4. Dans le terroir
de Kotchari et ses environs, il existe des espaces comme les bas-fonds,
considérés comme étant des ressources clés pour
l'élevage pastoral.
1.4. Objectifs
1.4.1. Objectif général et
finalitéDans la présente thèse, nous
nous attelons à analyser un socio écosystème liant
végétation et bétail dans le contexte d'une
aire protégée et de sa périphérie. Les objets
d'étude sont donc à la fois le système pastoral et les
écosystèmes végétaux sur lesquels il repose.
La finalité de ce travail est de donner les bases pour
rechercher un modèle d'exploitation pastorale viable et respectant la
diversité biologique.
1.4.2. Objectifs spécifiques
De manière pratique, les objectifs assignés
à notre recherche sont de :
· Faire l'état des lieux des ressources pastorales
dans le terroir en prenant en compte l'aire protégée voisine ;
· Préciser le niveau de la pression foncière
et les contraintes qui se présentent à l'élevage;
· Identifier les types d'évolution des pratiques en
réponse à la pression foncière ainsi que les
stratégies qui les sous-tendent ;
· Appréhender les représentations des
éleveurs sur leurs ressources et la manière dont ils mobilisent
leurs savoirs techniques pour valoriser ces ressources;
· Identifier les écarts entre le discours et la
réalité pratique.

1.5. Démarche conceptuelle
1.5.1. Objet de l'étude et cadre conceptuel : le
système pastoral, un système écologique complexe et
pioté
L'objet de notre étude est le système pastoral
(ou système d'élevage mobile), considéré dans une
optique de conservation du milieu végétal. La
préoccupation majeure qui nous guide est de concilier dynamique de
développement socio-économique des communautés, notamment
pastorales, et préservation des ressources naturelles. D'un point de vue
écologique, l'activité pastorale est un facteur essentiel dans
l'évolution de l'espaceressource (Barrière, 1996 ;
Barrière & Barrière, 1997) même si, pour diverses
raisons (existence combinée d'autres facteurs, position
inférieure dans la hiérarchie de ces facteurs, accès libre
des milieux pâturés dans nos milieux) l'effet de ce facteur est
difficile à estimer (Bourliere & Hadley, 1983 ; Cole, 1986 ; Scholes
& Walker, 1993 ; César, 1994 ; Botoni, 2003 ; Kièma S., 2007
; Staver et al. 2009).
Le concept de système est un concept opératoire
ou une représentation finalisée du réel (Hubert, 1994)
susceptible d'orienter l'action destinée à transformer le
réel pour répondre à un objectif bien défini
(Landais, 1987). Cette représentation du système, avec ses
limites, ses éléments et les relations entre ces
éléments, se fait de préférence sous un angle
fonctionnel ; il relève de la stricte décision de
l'opérateur et reflète sa perception de la réalité
étudiée (Landais, 1994). L'opérateur doit donc être
conscient qu'il peut exister d'autres perceptions et donc d'autres
manières de schématiser la même réalité et
que du type de représentation choisi dépendra l'efficacité
de la démarche adoptée.
Cependant, quelle que soit la perspective personnelle d'un
opérateur qui s'intéresse à un ou des systèmes
d'élevage, sa démarche prendra obligatoirement en compte trois
pôles unanimement reconnus comme constitutifs de tels systèmes. Il
s'agit du pôle humain formé par l'éleveur ou le groupe
d'éleveurs, du pôle animal (ou troupeau) et du pôle
ressource (ou territoire) (Lhoste, 1984 ; Landais, 1987 & 1992). Il existe
plusieurs définitions des systèmes d'élevage, celle de
Lhoste (1984) est l'une des plus complètes : elle stipule que «
le système d'élevage est une combinaison des ressources, des
espèces animales et des techniques et pratiques mises en oeuvre par une
communauté ou par un éleveur, pour satisfaire ses besoins en
valorisant des ressources naturelles par des animaux ». Cette
définition adopte le point de vue de Balent & Gibon (1999) et Botoni
(2003) qui considèrent le système pastoral comme le
résultat de la gestion par l'homme des interactions
herbivore-végétation. Cette posture, qui est aussi celle que nous
adoptons, résume le système à l'éleveur ou au
groupe d'éleveurs et leurs troupeaux, vus à travers leurs
pratiques, et le territoire pastoral qui contient les ressources qu'ils
utilisent (figure I-1).
Comme on le voit, le système pastoral est un
système écologique, ouvert et piloté (Bonnet, 1990;
Landais, 1992 ; Hubert et al. 1993; Lhoste et al. 1993) dont
l'étude s'avère assez complexe. Le parti choisi ici est
d'analyser à la fois les différentes composantes du
système ainsi que leurs interrelations comme un tout. Plus
précisément, on s'est intéressé aux interactions
entre les activités pastorales et les paysages exploités.
L'approche systémique ainsi adoptée (Lhoste, 1984;
Landais 1992 & 1994 ; Daget & Godron, 1995 ; Botoni, 2003)

16
repose sur l'enquête pastorale telle que la
conçoivent Daget & Faure (2003) et la cartographie, elle prend en
compte l'organisation et le fonctionnement du système dans sa
globalité. Il s'agissait pour nous d'explorer tous les pôles du
système en acquérant des données sociologiques
(étude des acteurs, de leurs représentations et de leurs
pratiques) et naturalistes (étude des ressources naturelles en
particulier végétales) qui sont ensuite croisées. Compte
tenu de l'objet de notre étude et de nos objectifs, nous avons
privilégié les pôles humains (les pratiques) et celui des
ressources en insistant sur les interactions entre eux et sur les effets de ces
interrelations sur leurs dynamiques respectives.
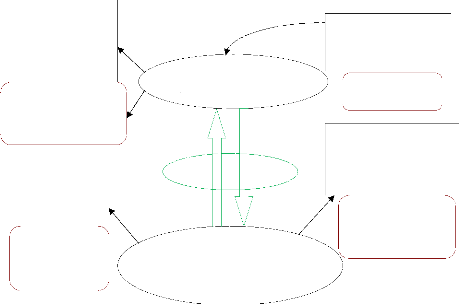
Indicateurs de gestion du milieu: -
Régime des feux - Organisation des parcelles
agricoles
Indicateurs relatifs à la
végétation
- Recouvrement
- Richesse floristique - Diversité floristique -
Valeur pastorale
Méthodes
- Abondance-dominance - Interception linéaire
- Bibliographie
Méthodes - Enquêtes
- Cartographie des milieux
Pratiques pastorales
(Interactions herbivore-végétation : animaux
d'élevage, et relations avec la faune sauvage)
Pâturages (fourrage,
points d'eau, cures salées)
Interactions
Climat, sols, politiques nationale et régionale en
matière de développement rural et de conservation
Indicateurs de pratiques - Utilisation de
l'espace
- Comportement alimentaire - Stratégies des pasteurs
- Cartographie/SIG
Méthodes
- Suivis de troupeaux - Enquêtes informelles -
Enquêtes formelles
Méthodes Bibliographie
Figure I-1 : Schéma du modèle conceptuel
étudié (adapté de Botoni, 2003)
1.5.2. Définition des termes et concepts
Tout le long de la thèse nous aurons recours à
un certain nombre de termes et concepts dont la définition peut varier
selon les auteurs et les écoles de pensée. Il convient donc de
préciser l'acception qui est la nôtre.
1.5.2.1. Les aires de conservation
Une aire protégée est un espace
naturel identifié, circonscrit et géré comme tel. L'IUCN
(1994), définit une aire protégée comme "une portion
de terre, de milieu aquatique ou de milieu marin, géographiquement
délimitée, vouée spécialement à la
protection et au maintien de la diversité biologique, aux ressources
naturelles et culturelles associées; pour

ces fins, cet espace géographique doit être
légalement désigné, réglementé et
administré par des moyens efficaces, juridiques ou autres". Les
formes de gestion tout comme l'intérêt scientifique, biologique ou
patrimonial peuvent y être divers, et les aires protégées
se déclinent ainsi en forêts classées, aires de protection
faunique, parcs nationaux, réserves de biosphère, réserves
naturelles intégrales, réserves de faune totales ou partielles,
sanctuaires, ranches, refuges locaux, zones villageoises d'intérêt
cynégétique (MEE, 1997; MECV, 2004).
Un parc national est une aire protégée,
administrée principalement dans le but de préserver les
écosystèmes et aux fins de récréation (MEE,
1997).
Une réserve est une zone
protégée en raison de son intérêt écologique
et où les activités humaines sont, en principe,
réglementées. Dans une « réserve intégrale
» ou « naturelle », ou encore « biologique », toute
activité humaine est interdite, y compris le ramassage.
Une réserve de biosphère est une aire
déclarée comme bien du patrimoine mondial en raison de ses
spécificités biologiques, écologiques, culturelles ou
historiques (MEE, 1997). Elle peut être terrestre, côtière
ou marine et est protégée au niveau international dans le cadre
du programme MAB (Man And Biosphere) de l'UNESCO, en vue d'une conservation de
la diversité biologique, associée à un
développement économique, social et culturel durable ainsi
qu'à des activités scientifiques. Le programme MAB affirme
l'importance d'asseoir les stratégies de conservation de la nature sur
la connaissance d'une part, mais aussi sur le développement
économique et social des populations, dans le respect des cultures
locales (Génot & Barbault, 2004). Les réserves de
biosphère sont placées sous la juridiction souveraine de
l'état où elles sont situées (Da Lage &
Metailié, 2000). Elle comprend une aire centrale, qui
bénéficie d'une protection intégrale, d'une zone
tampon, où peuvent être menées des
expérimentations de recherche, et enfin d'une zone
périphérique ou de transition soumise à
l'exploitation agricole.
Une réserve naturelle intégrale est une
aire protégée administrée principalement aux fins
d'étude scientifique.
1.5.2.2. Notions de biodiversité, de
conservation et d'écologie des parcours
L'écosystème est un système
constitué par l'ensemble des êtres végétaux et
animaux vivant dans un milieu physique donné en interaction
étroite avec ce dernier (Ozenda, 1982). Des relations complexes
(synergie, complémentarité, concurrence, etc.) lient ces
êtres vivants entre eux et à leur environnement physique. Rares
sont aujourd'hui les écosystèmes totalement naturels. La plupart
de écosystèmes ont déjà été plus ou
moins profondément artificialisés (et fragilisés) par les
interventions anthropiques.
La diversité biologique, ou son
équivalent plus récent (Veuille 2006 ; Barbault, 2008), la
Biodiversité, évoqué pour la première fois par
Édouard Wilson en 1985 et médiatisé lors du sommet de la
terre de Rio de 1992 (Barbault, 2006), est un concept global, indiquant la
propriété des systèmes vivants à être
différents (Betsch et al. 2003). Selon l'échelle
à laquelle on se place (Le Guyader, 2006) on parle de diversité
génétique, de diversité spécifique (ou des
organisations) ou de diversité écologique (ou diversité
des écosystèmes) (Norse et al. 1986).

18
Le concept biodiversité est assez complexe et selon
Barbault (1993), il fait appel à trois approches possibles : une
approche écologique qui se préoccupe de ses rôle et place
dans le fonctionnement des écosystèmes ; une approche
éthique où la biodiversité est considérée
comme un patrimoine naturel devant, à ce titre, être transmise aux
descendants; une approche économique, qui perçoit la
biodiversité comme une réserve de ressources potentielles
à exploiter. Cette dernière approche attribue à la
biodiversité quatre types de valeurs (Heywood, 1997 ; Barbault 1995 ;
Betsch et al. 2003): les valeurs d'usage (consommation
directe, production, recréation), les valeurs
écologiques (régulation des eaux, des climats, des invasions
biologiques, lutte contre l'érosion, etc.), les valeurs
d'option (matériaux nouveaux, usages pharmaceutiques, etc.) et les
valeurs d'existence (consentement à payer).
Signalons que l'un des résultats majeurs du sommet de
Rio a été la mise en place de la Convention sur la
Diversité Biologique (CBD) qui est entrée en vigueur le 29
décembre 1993. La CBD considère que la préservation de la
biodiversité à toutes les échelles ainsi que l'utilisation
durable des ressources naturelles sont des préoccupations communes de
l'humanité et qu'elles font partie intégrante du
développement durable.
La notion de conservation a connu une
évolution dans son acception (Génot & Barbault, 2004;
Aubertin et al. 2008). Signifiant à l'origine une protection
intégrale des espaces délimités (sanctuarisation) avec
exclusion des humains, on lui donne aujourd'hui une vision
intégrée; c'est-à-dire, une préservation des
ressources naturelles qui inclut leur exploitation humaine.
Un indicateur est une valeur calculée
à partir de paramètres donnant des indications sur ou
décrivant l'état d'un phénomène, de l'environnement
ou d'une zone géographique (OCDE, 1993).
Un indicateur biologique ou bio-indicateur,
est un organisme ou ensemble d'organismes qui, par référence
à des variables biochimiques, cytologiques, physiologiques,
éthologiques ou écologiques, permet, de façon pratique et
sûre, de caractériser l'état d'un écosystème
ou d'un éco complexe et de mettre en évidence leurs modifications
(Blandin, 1986).
L'espace pastoral est l'entité territoriale
sur laquelle évoluent les systèmes d'élevage pastoraux ou
systèmes d'élevage mobile. Il comprend l'ensemble des terres
(terres naturelles ou modifiées consacrées à
l'élevage, terres cultivées et accessibles après les
récoltes, terres réservées aux cultures
fourragères, plantations, parcours forestiers, etc.) parcourues par les
animaux en vue d'y prélever leur nourriture (César, 1994 ;
D'Amico et al. 1995 ; Botoni, 2003).
Milieu en équilibre / déséquilibre :
certains écologues du pastoralisme (Breman & De Ridder, 1991 ; Daget
& Godron, 1995 ; Scoones, 1999) distinguent les « milieux
équilibrés » d'une part et les « milieux en
déséquilibre» ou en « équilibre
instable » d'autre part. Les premiers se rencontreraient dans les
pays et régions suffisamment humides où les régimes
pluviométriques sont stables. Des mécanismes classiques de
rétroaction (impact négatif sur la végétation
lorsque le cheptel devient important et dépasse une «
capacité de charge » avec risque de dégradation à
long terme) y ont cours : le contrôle des charges animales donnera alors
des résultats. Les écosystèmes en
déséquilibre se rencontrent au contraire en milieu

semi-aride et surtout aride, où le facteur limitant est
l'eau (la pluviométrie) plutôt que la fertilité des sols
(Breman et De Ridder, 1991) et l'impact du contrôle des charges devient
aléatoire.
Le terroir, désigne un ensemble spatial
agronomiquement homogène caractérisé par une même
structure et une même dynamique écologique, ainsi que par un
même type d'aménagement agricole (Bonnet, 1990). Dans une
problématique de gestion de l'espace, le terroir villageois est souvent
assimilé à « finage » (Vielzeuf, 1986 in
Bonnet, 1990). Dans cette thèse nous assimilons le terroir au finage
entendu comme « un espace dont une communauté agricole
définie par les liens de résidence, tire l'essentiel de sa
subsistance ; autrement dit la portion de sol environnant où se
localisent les champs et où paît le bétail, dans la mesure
où celui-ci se trouve associé d'une façon ou d'une autre
à la culture » (Sauter, 1962 in Bonnet, 1990).
1.5.2.3. Concepts de pastoralisme et de pratiques
pastorales
La mobilité est un phénomène historique,
caractéristique des peuples pasteurs qui en usent pour gérer
l'imprévu et les risques (Scoones, 1995 ; Nori, 2006 & 2007 ; Nori
et al. 2008) dans les milieux arides et subarides. Le terme
pastoralisme se réfère par ailleurs, selon Wane (2006), aux modes
de conduite des troupeaux sur pâturage naturel, et donc, aux
systèmes où l'élevage est pratiqué de
manière extensive avec peu ou pas de complémentation et sans
pratique de cultures fourragères.
Un élevage pastoral ou élevage
mobile est, selon Brunschwig et al. (2001), un système
d'exploitation basé sur l'utilisation de superficies composées en
majorité de parcours non récoltables et dont l'utilisation est
assurée uniquement par le pâturage des animaux. Pour Asiedu et
al. (2009), si plus de 90 % de la matière sèche
consommée par le bétail provient des pâturages cela est
suffisant pour qualifier le système de pastoral. Adoptant une approche
économique, Swift (1988) considère qu'un système de
production pastorale est un système dans lequel au moins 50 % du revenu
brut des ménages proviennent de l'élevage ou d'activités
qui lui sont liées ou dans lequel plus de 15 % de la consommation
d'énergie alimentaire des ménages se composent de lait ou de
produits laitiers produits au sein des ménages. A contrario,
selon le même auteur, un élevage agropastoral est un
élevage dans lequel, le revenu brut des ménages est
généré à plus de 50 % par l'agriculture ou dans
lequel entre 10 à 50 % de ce revenu proviennent de l'élevage.
Dans le système agropastoral, le cheptel est fortement dépendant
du fourrage cultivé (Nori, 2007).
Si l'on se réfère à l'amplitude des
déplacements14 et à leur fréquence, on peut
distinguer l'élevage sédentaire (ou sur parcours villageois),
l'élevage transhumant (petite et grande transhumance) et
l'élevage nomade.
L'élevage sédentaire est le type
d'élevage extensif qui implique le moins de mobilité. La
mobilité, réduite, est généralement interne
à l'espace des terroirs villageois ou des villages les plus proches et
il n y a pas de déplacements cycliques (Nori, 2007).
14 Nous faisons référence aux
déplacements "habituels" qui doivent être distingués des
déplacements d'urgence imposés par les crises (sécheresse,
épidémie, conflit).

20
La transhumance, selon Lhoste et al. (1993)
et Wane (2006), est « un système de production animale
caractérisé par des mouvements saisonniers de caractère
cyclique, d'amplitude variable. Ces mouvements saisonniers
préétablis (Nori, 2007) s'effectuent entre des zones
écologiques complémentaires, sous la garde de quelques personnes,
la plus grande partie du groupe restant sédentaire ». Selon
l'ampleur du déplacement on parle de petite transhumance15
(délocalisation temporaire et à une courte distance des animaux
pour éviter les dommages causés dans les champs pendant la saison
des pluies) ou de grande transhumance16 (Bierschenk & Le Meur,
1997 ; Convers, 2002). Cette dernière concerne surtout les bovins
(Saidou, 1986) et donne lieu à une minutieuse organisation (Toutain et
al. 2001).
Le nomadisme se réfère à une
pratique de mobilité en élevage dans laquelle les pasteurs n'ont
pas d'habitat fixe et permanent (Wane, 2006 ; Nori, 2007) : toute la famille
suit les déplacements du troupeau, parfois sur de longues distances (des
centaines de kilomètres). Ils se déplacent avec leurs troupeaux
et du fait de cette mobilité, les nomades pratiquent peu
d'activités agricoles.
Notons que la transhumance tout comme le nomadisme
relèvent du genre de vie pastoral (Benoit, 1979 ; Boutrais, 1992; Daget
& Godron, 1995).
Les pratiques sont définies comme l'ensemble
des actions mises en oeuvre dans l'utilisation du milieu (Blanc-Pamard &
Milleville, 1985) ou les façons dont l'opérateur met en oeuvre
une opération technique (Lhoste & Milleville, 1986).
Selon ces auteurs, l'approche des pratiques renvoie à
trois séries de questions : (i) l'identification des pratiques et leur
caractérisation, (ii) l'évaluation de leurs effets (impacts sur
le milieu, le bétail à travers sa dynamique et son niveau de
production) et la recherche des causes qui les motivent (les
stratégies). Guérin & Hubert (1995) épousent ce point
de vue lorsqu'ils affirment que les manières de faire des
éleveurs peuvent être caractérisées par leurs
modalités (pratiques), leur efficacité (résultats des
actions) et leur opportunité (motivations des actions), selon que l'on
s'intéresse aux aspects décisionnels, descriptifs ou
techniques.
La difficulté majeure pour comprendre le
fonctionnement des exploitations d'élevage est de mettre en
évidence le projet de l'éleveur, d'analyser sa cohérence
avec des choix stratégiques qu'il s'agit d'identifier. En analysant les
« manières de faire » des éleveurs, on en arrive
à mettre en lumière les décisions qu'ils prennent et leurs
objectifs. Landais & Deffontaines (1989) disaient si bien à ce
propos qu'« on connaît les projets par les pratiques, on
comprend les pratiques par les projets ». Dans le processus
d'identification des projets des éleveurs, il ne s'agit cependant pas de
rendre compte du processus de décision lui-même (Girard, 1995)
mais de se focaliser sur les pratiques pour expliquer la cohérence dans
laquelle s'inscrivent un certain nombre de décisions.
15 yawtooru en langue peule (Convers,
2002)
16 bartoje en langue peule (Convers,
2002)

La relation troupeau/végétation est
pilotée par un éleveur (Lhoste, 84 ; Landais 92 & 94) qui met
en oeuvre un certain nombre de pratiques, elles-mêmes fonction des
informations dont il dispose sur l'état de cette relation (Guérin
& Hubert, 1995) et de ses projets propres.
En milieu pastoral, il existe trois types de pratiques (Landais,
1994)
· Les pratiques d'élevage stricto sensu
à travers lesquelles les éleveurs interviennent directement sur
le troupeau. Elles concernent (i) les pratiques d'agrégation ou
de constitution du troupeau ou encore d'allotement qui concernent la formation
des groupes d'animaux, (ii) les pratiques de conduite du troupeau qui
regroupent toutes les opérations d'entretien (soins, reproduction,
alimentation, etc.) effectuées sur les animaux afin d'améliorer
leurs performances, (iii) les pratiques d'exploitation qui concernent
toutes les opérations de prélèvements (traite, abattage,
tonte, etc.), (iv) les pratiques de renouvellement qui ont trait aux
actions de renouvellement de la composition (réforme des animaux
âgés et des malades, sélection de jeunes d'allotement,
etc.) et (v) les pratiques de valorisation (transformation et mise en
marché) qui s'appliquent aux productions animales (fromage, charcuterie,
etc.) .
· Les pratiques fourragères regroupent
toutes les opérations agronomiques qui ont lieu sur les pâturages.
Dans le contexte soudano-sahélien, les pratiques fourragères sont
quasi-absentes, les éleveurs se contentant bien souvent d'exploiter
l'herbe naturelle avec peu ou pas d'actions agronomiques.
· Les pratiques de gestion du pâturage et des
stocks fourragers qui mettent en relation le troupeau aux parcelles
fourragères.
CHAPITRE II

22
ETAT DES CONNAISSANCES :
FONCTIONNEMENT DES ECOSYSTEMES
SAVANIENS, SOCIOLOGIE DU PASTORALISME
ET EVOLUTION DE LA POLITIQUE DE
CONSERVATION

23
2.1. Nature et types de savanes
La végétation du domaine soudanien du Burkina
Faso, domaine dont relève notre zone d'étude, est essentiellement
constituée de savanes considérées comme des
écosystèmes complexes et dynamiques. Il existe une abondante
littérature sur l'origine et la nature des formations savanicoles
(Schimper, 1935 ; Burtt-Davy, 1938 ; Trochain, 1940 ; Aubreville, 1957 ; Birot,
1965 ; Schnell, 1971 ; Boudet, 1978 ; Beani & Dessi, 1984 ; Fournier, 1991
; César, 1992 ; Breman & Kessler, 1995 ; Menaut et al.
1995, etc.). Un point de vue considère les savanes comme étant
des formations naturelles climaciques17 (Schimper, 1935 ; Birot,
1965 ; Petit, 1990 ; Ramade, 1994) pendant que certains spécialistes de
la question (par exemples Schnell, 1971 ; Dajoz, 1982) admettent que la nature
des climax tropicaux est forestière, les savanes étant alors des
formations secondaires résultant d'actions anthropiques (Dajoz, 1982 ;
Harchies et al. 2007). Une hypothèse toute récente
(Beerling & Osborne, 2006) veut que ces formations, constituées
d'herbes à photosynthèse de type C4 et de ligneux de type C3 et
qui sont apparues il y a environ 8 millions d'années, soient fortement
dépendantes de la manière dont le feu influence la microphysique
des nuages, le climat et l'écologie. D'après cette
hypothèse, le feu serait favorable à l'expansion des
herbacées et limiterait la densité des ligneux,
phénomène qui est amplifié par une atmosphère peu
chargée en dioxyde de carbone (CO2). Ils ajoutent que les
mécanismes qui président à ce processus peuvent être
perturbés (rehaussés ou abaissés) du fait d'une pression
continue sur les écosystèmes.
Selon Schnell (1971) les formations de savane avaient
été classées par Burtt-Davy (1938) parmi les formations
herbacées tropicales avant que la rencontre de Yangambi (Aubreville,
1957) ne les situe définitivement dans le grand groupe des formations
mixtes forestières et graminéennes et formations
graminéennes. Elles se localisent en Afrique en zone intertropicale
(zone soudanienne au Nord et zone zambézienne au Sud) (Dajoz, 1982 ;
Menaut et al. 1995).
Breman & Kessler (1995) définissent la savane
comme une formation de transition entre les formations forestières
fermées et les formations herbeuses ou tout simplement le désert.
Ces auteurs sont rejoints par Sankaran et al. (2005) qui notent que
« Les écosystèmes purement herbacés et purement
ligneux constituent les extrêmes d'un continuum « savane » qui
peut être défini comme une entité fonctionnelle liée
à l'équilibre herbes-arbres ». Trochain (1940) et
Ozenda (1982) abordent la savane sous l'angle physionomique et la
considèrent comme « un tapis graminéen ouvert, tout au
moins au niveau du sol, de hauteur variable, obligatoirement parsemé
d'arbustes clairsemés et parfois d'arbres isolés ».
Pour Beani et Dessi (1984), la savane est un écosystème à
climat tropical qui se caractérise par une herbe xérophile et par
une couverture irrégulière d'arbres et d'arbustes. Mais la
définition la plus précise de la savane à notre sens, est
celle de Boudet (1978) qui la qualifie de « type de
végétation dont le couvert herbacé majoritairement
à graminées pérennes, d'au moins 80 cm
17 De climax, notion contestée de nos jours
(Génot, 2006) et définie de diverses manières. Selon
Ramade (1984) le climax est « une association stable d'espèces qui
caractérisent qualitativement et quantitativement l'ultime phase de
développement d'une biocénose dans une succession » alors
que Fischesser et Dupuis-Tate (1996), dans le guide illustré de
l'écologie, le considère comme un « groupement vers lequel
tend la végétation d'un lieu dans des conditions naturelles
constantes, en l'absence d'intervention de l'homme. Sa structure dépend
étroitement des facteurs climatiques et de la nature du sol ».

de hauteur, comprend deux strates (inférieure et
supérieure) s'organisant le plus souvent en touffes continues et
brûlant régulièrement chaque année ». En
effet, lorsque la savane est régulièrement parcourue par le feu,
sa composition floristique reste stable et chaque unité de
végétation demeure identique à elle-même dans le
temps (César, 1992) à condition, bien entendu, qu'aucun autre
facteur de perturbation n'intervienne.
Lors de son colloque de Yangambi, tenu du 29 juillet au 8
août 1956 (Aubreville, 1957), le conseil scientifique pour l'Afrique au
sud du Sahara (CSA), a défini 5 types de savanes allant des forêts
claires aux savanes herbeuses. La classification s'appuie surtout sur la
hauteur et la densité (ou le recouvrement) de la strate herbacée
(tableau II-1).
Au plan de la composition floristique surtout
herbacée, les formations de savane sont faites essentiellement de
graminées (en termes de masse végétale et de nombre
d'individus) avec une forte présence de Cypéracées et de
légumineuses (Fournier, 1991).
Tableau II-1. Classification des formations de savane africaine
lors du colloque de Yangambi en 1956 (Aubreville, 1957).
Type de Formation Hauteur Recouvrement
Forêt claire
Savane boisée
Savane
arborée
Savane arbustive
Savane herbeuse
|
> 8 m > 8 m > 8 m < 8 m < 8 m
|
70 à 90 % 20 à 70 % 2 à 20 % 2 à
70 % 0 à 2 %
|
|
2.2. Les écosystèmes savaniens :
fonctionnement et perturbation
Les principaux déterminants des formations
végétales de savane sont le sol, la topographie et le climat
(Schnell, 1971 ; César, 1992). La diversité, la structure et le
fonctionnement des savanes sont affectés par la variation dans l'espace
et le temps de ces facteurs, auxquels s'ajoutent les effets de perturbations
comme le feu, la pâture animale, les prélèvements humains
et les défrichements agricoles (Bourlière & Hadley, 1983 ;
Cole, 1986 ; Scholes & Walker, 1993 ; César, 1994 ; Fournier et
al. 2001 ; Jentsch, 2001 ; Staver et al. 2009). Cependant,
lorsqu'un facteur intervient de façon régulière et
prévisible, comme la pâture animale (Gaucherand, 2005) ou les feux
de brousse (César, 1992), son statut de perturbation est discutable, il
peut être considéré comme faisant partie intégrante
du fonctionnement de l'écosystème, structurant alors les
communautés animales et végétales comme l'ont
théorisé Allen & Starr (1982). D'après ces auteurs, la
perturbation, lorsqu'elle est récurrente, finit par s'intégrer au
système qui, en retour, exerce sur lui un certain contrôle
permettant ainsi sa régulation et la stabilisation de l'ensemble. La
perturbation régulière s'incorpore ainsi au système et,
comme par exemple pour le feu en savane, devient nécessaire à son
fonctionnement (Fournier et al. 2001).

25
Schnell (1971), White & Pickett (1985), White &
Jentsch (2001) ainsi que Turner et al. (2003), définissent les
perturbations, qui peuvent être d'origine naturelle ou
anthropogénique, comme étant généralement des
événements relativement discrets ou brusques qui modifient la
structure des écosystèmes, celle des communautés
végétales ou animales et modifient le stock des ressources, la
disponibilité de substrats et l'environnement physique. D'après
Huston (1994), ce sont des processus aléatoires pouvant se traduire par
une perte soudaine de la biomasse au sein d'une communauté sur un laps
de temps significativement plus court que celui nécessaire à
l'accumulation de cette biomasse. Cette perte de biomasse peut libérer
de l'espace et donc des ressources pour de nouveaux organismes (Roxburgh et
al. 2004).
Les effets des perturbations sont complexes et variables, ils
dépendent de l'état initial du milieu, mais aussi de leur
fréquence, de leur intensité, de l'échelle et de la
période (Frost et al. 1986). Les perturbations seraient «
régulatrices de diversité biologique à l'échelle du
court-temps et génératrices de cette même diversité
à l'échelle du long temps » (Blondel, 2003).
L'évolution des écosystèmes savaniens sous l'effet de ces
facteurs peut être progressive ou régressive (Boudet, 1978), elle
aboutit à un nouvel état d'équilibre à la suite
d'un remaniement du cortège floristique initial.
2.2.1. Les feux et leurs rôles dans le
fonctionnement des écosystèmes de savane
2.2.1.1. Importance socio- économique et types
de feux
Historiquement, le feu a toujours été le
compagnon de l'homme qui en faisait déjà usage (Bruzon, 1994 ;
Trabaud, 1995; Mazoyer & Roudart, 1998) initialement pour divers usages
comme la chasse, l'essartage et l'écobuage (Beani & Dessi, 1984 ;
Hoffmann, 1985 ; Bruzon, 1994 & 1995 ; Lavorel et al. 2007). Pour
beaucoup de peuples africains, en effet, le feu est un outil essentiel pour
chasser, éclaircir les paysages, contrôler les maladies et
détruire les résidus de culture (Frost et al. 1986 ;
Lecomte, 1995 ; Lavorel et al. 2007).
La classification des feux repose sur divers critères.
Ainsi, Hoffmann (1985), César (1992) et Bruzon (1994), se focalisant sur
les périodes de leur survenue, définissent des feux
précoces, des feux tardifs (ou de saison sèche ou encore de
contre-saison). Monnier (1981), s'intéressant à la localisation
spatiale du feu dans la végétation, distingue des feux de fauche
(les plus violents), des feux rampants, des feux d'humus et des feux de cime ou
de buissons. En se plaçant sous l'angle de la valorisation des milieux,
on peut distinguer à l'instar de Bruzon (1994), des «feux
pastoraux» et des «feux agricoles».
2.2.1.2. Les feux, un facteur de régulation des
savanes
Les spécialistes du fonctionnement et de la dynamique
des écosystèmes savaniens sont, de nos jours, unanimes sur le
fait que les feux, qu'ils soient d'origine naturelle (la foudre) ou
anthropique, sont une composante à part entière des savanes
(Lamotte, 1970 ; César, 1991 & 1992 ; Dolidon, 2005). Ils
contribuent à maintenir cet écosystème (Lamotte, 1979 ;
Schnell, 1971; Monnier, 1981 ; Bruzon, 1990 ; Fournier, 1991 ; César,
1992 ; Scholes & Walker, 1993 ; Gaucherand, 2005 ; Fournier & Devineau,
2009) avec lequel ils ont de tout

temps co-évolué (Frost et al. 1986 ;
Fournier, 1990). Ils constituent donc un élément fondamental de
fonctionnement de l'écosystème au même titre que le climat
(Naveh, 1975). Vu sous cet angle, le feu peut être
considéré comme un facteur naturel de conservation ou de
régulation en zone de savane (César, 1991 ; Hoffmann et
al. 2003), il évite l'évolution de celle-ci vers les
formations forestières (Schnell, 1971 ; Monnier, 1981 ; Fournier, 1991).
Ce rôle du feu dans l'entretien des écosystèmes de savane
est confirmé par Aubreville (in Dolidon, 2005) qui affirme que «
si les feux de savane ne sévissaient pas chaque année, la
reconstitution de la forêt se produirait instantanément sur une
grande partie de l'aire qu'elle a perdue ».
2.2.1.3. Impacts des feux sur les
écosystèmes
Les feux interviennent dans le fonctionnement global des
écosystèmes en accélérant, contrariant ou
supprimant certains de ses processus (germination et croissance des
espèces, successions végétales, etc.). Leurs effets sont
fonction de leur intensité, de leur fréquence, de la saison
à laquelle ils interviennent (Alexandre, 1989 ; Lavorel et al.
2007), mais aussi du type de végétation, de la topographie et de
la nature du sol dans le site considéré (Bruzon, 1995; Pyne et
al. 1996). Les impacts des feux sur les milieux peuvent s'analyser
à travers les changements provoqués dans la
végétation (richesse floristique, composition spécifique,
diversité spécifique, structure spatiale, types biologiques,
physionomie et phénologie) et dans les caractéristiques
physico-chimiques des sols (teneur en nutriments et en microorganismes).
2.2.1.3.1. Les feux et leurs impacts sur la
végétation
Les feux entretiennent les savanes en agissant comme des
agents sélectifs et régulateurs qui permettent d'empêcher
le remplacement de la strate herbacée par la végétation
boisée (Shantz, 1947 ; César, 1991 & 1994 ; Lavorel et
al. 2007) et, comme rappelé précédemment,
l'évolution des formations savanicoles vers les types forestiers parfois
denses (Schnell, 1971 ; Monnier, 1981 ; Fournier, 1991 ; Jacquin, 2010). Leurs
effets sont multiformes, ils diffèrent notamment selon la nature
ligneuse ou herbacée de la formation végétale
considérée et les types biologiques végétaux qui
ont développé à l'égard du feu des
stratégies assez diversifiées (Schnell, 1971; Ozenda, 1982 ;
César, 1992 ; Bruzon, 1995). Du point de vue de l'utilisation pastorale
des milieux, on note que les feux influencent le niveau de la production
primaire nette, son évolution cyclique (disponibilité
saisonnière du fourrage) ainsi que sa qualité fourragère
(appétence et qualités nutritionnelles) (Schnell, 1971).Par
ailleurs, le moment (précoce ou tardif) de survenue des feux peut
être déterminant sur l'évolution des
écosystèmes pastoraux.
En ce qui concerne les types biologiques, les
hémicryptophytes (les graminées pérennes surtout) et les
géophytes disposent respectivement soit de bourgeons basilaires
protégés dans les talles et qui rejettent après le passage
du feu (Fournier, 1991) soit de semences enfouies dans le sol et supportant
l'élévation de température consécutive au passage
des feux. L'effet dépressif observé chez les ligneux, est bien
moindre ou parfois absent chez les herbacées chez lesquelles on peut
noter, dans le temps, une stimulation du tallage et un élargissement des
touffes (Bruzon, 1995).

27
Les feux précoces ont des effets peu nocifs sur la
végétation ligneuse, ils retardent mais ne compromettent pas son
développement. Ces feux surviennent, en effet, à une
période où l'herbe est encore un peu humide (stress hydrique
faible), et se contentent de consumer la litière de feuilles et des
herbes fanées (Beani & Dessi, 1984) là où
l'accumulation de combustible de l'année précédente n'est
pas élevée. Manquant donc d'assez de combustible (César,
1994), ils sont bas, lents et leur température n'est pas très
élevée ; ils n'affectent que la couche supérieure du sol
préservant ainsi les racines et les graines, ils ne s'attaquent
pratiquement pas aux arbres. Au contraire, en éliminant une bonne partie
de la strate herbacée pendant son passage, ils réunissent les
conditions (abaissement de la compétition herbe-arbre) d'une bonne
croissance des ligneux plus tard au moment de leur reprise. De ce fait, en
favorisant le développement des ligneux, ils seraient favorables aux
savanes arborées ou boisées (César, 1994). Les feux
pastoraux, généralement précoces, qui visent, selon
Boutrais (1994), à favoriser, par des repousses, le renouvellement du
pâturage (Bruzon, 1994 ; César, 1994) relèvent, en
conséquence, de stratégies du court terme (Boutrais, 1994). Ils
conduisent, en effet, à long terme à l'embroussaillement des
formations de savane et à la perte d'une bonne partie du fourrage
herbacée.
Notons qu'en zone subhumide, si le feu est précoce -
pas assez précoce cependant pour interrompre la mise en réserves
des nutriments dans les organes souterrains (GTZ, 1979 in Hoffmann,
1985) - les repousses des graminées vivaces après le passage des
feux précoces sont plus importantes à cause de l'humidité
des sols encore élevée à cette période (Bruzon,
1995), ce qui accroît la valeur pastorale des milieux.
Cependant, les feux précoces
répétés, en favorisant le développement de la
végétation ligneuse, vont entraîner l'élimination
des hémicryptophytes, les plus recherchées par le bétail
(Daget & Godron, 1995 ; Kagoné, 2000), au profit des types
biologiques de plus grande taille (chamaephytes et nanophanerophytes) et une
réduction de la contribution des Graminées. A la longue il se
produit un embroussaillement puis une reforestation (Boutrais, 1994 ;
César, 2005). Ceci a des incidences négatives sur la valeur
pastorale et les potentialités fourragères de la savane qui
dépendent des graminées pérennes. Rippstein (1985) a
observé par exemple, suite à des études conduites dans
l'Adamaoua (Nord-Cameroun), qu'à partir d'un recouvrement ligneux de 40%
environ, la productivité et la valeur pastorale des milieux devenaient
faibles.
Les effets des feux dits « tardifs » sont
généralement plus nocifs que ceux des feux précoces
surtout sur la strate ligneuse (César, 1994 ; Bruzon, 1995). En effet,
au moment où ces feux surviennent (précisément en saison
sèche chaude), le combustible herbacé, assez sec, est abondant ce
qui permet un feu violent au moment même où les ligneux sont en
train d'émettre de nouveaux organes, notamment les feuilles et aussi,
pour certains, de fleurs et de fruits (Schmitz et al. 1996). Ces
organes, y compris les jeunes branches, sont alors brûlés,
obligeant les ligneux à une seconde et épuisante foliaison
(César, 1992). La croissance en zone de savane peut ainsi être
réduite faute de feuilles pour assurer la photosynthèse. Par
ailleurs, les rejets, les arbustes et les jeunes pousses,
particulièrement vulnérables (Beani & Dessi, 1984), sont en
grande partie détruits, ce qui empêche l'installation de nouveaux
individus ligneux (Bruzon, 1995). Étant inclus dans la strate
herbacée, ces derniers, encore fragiles, sont en effet
brûlés par les feux (Western & Maitumo, 2004 ; Bond &
Keeley ; 2005)
lors de leur passage. Les feux, en particulier le type
tardif, empêcheraient donc l'embroussaillement des milieux et
favoriseraient l'installation de savanes arbustives (César, 1994, cf.
figure II-1).
Cependant, la réponse des ligneux à l'action du
feu, même tardif, serait assez diverse selon l'espèce
considérée (Schnell, 1971) et l'âge de l'individu ligneux.
Selon Bruzon (1995) en effet, certaines espèces comme Afzelia
africana Smith ex Persoon et Nauclea latifolia Smith sont
résistantes ou «pyrotolérantes» (on les qualifie alors
de pyrophytes), elles rejettent abondamment à partir de souches munies
d'écorces épaisses, isolantes et subérifiées
protégeant les tissus vivants de leurs troncs. Elles ont, en outre, des
graines qui supportent les températures élevées. Par
ailleurs, d'après Schnell (1971), les arbres surtout les plus jeunes,
rejettent de la base et prennent un port buissonnant, lorsqu'ils sont atteints
par le feu.
Figure II-1. Rôle du feu dans l'évolution des
savanes (Source : César, 1994)
Légende :
En l'absence de feu, la végétation de savane
évolue vers la forêt dense. Le feu annuel maintient la
végétation des savanes, les feux précoces (courant
novembre) permettent l'installation de savanes arborées ou
boisées, tandis que les feux tardifs (mars) plus violents (le
combustible est devenu sec en ce moment) n'autorisent qu'une savane arbustive
claire.
2.2.1.3.2. Les feux et leurs impacts sur les
sols

Schnell (1971) puis Bruzon (1995) expliquent clairement et en
détail comment le feu influence les qualités chimiques et
biochimiques des sols. De manière générale, la couverture
du sol par la cendre induit une modification de sa température et de son
humidité relative. Par ailleurs, le sol est enrichi en
éléments assimilables comme le calcium, le magnésium, le
potassium et surtout le phosphore, de même, l'activité des
actinomycètes et des bactéries est stimulée par suite
d'une élévation du pH. Ces observations sont confirmées
par de nombreux

29
travaux conduits sous divers horizons (Senthilkumar et al.
1997 ; Jeensen et al. 2001 ; Wan et al. 2001). Les
travaux de Senthilkumar et al. (1997) apportent des précisions
sur le fait que seule la couche supérieure du sol (0-10 cm) est
affectée notamment par une augmentation de la population de la
microfaune ainsi que leurs activités enzymatiques du fait justement de
la disponibilité en matière organique.
Des feux fréquents et intenses (Bird et al.
2000 ; Parker et al. 2001 ; Mills & Fey, 2004) ou de plus en plus
tardifs (Bruzon, 1995) conduisent cependant à des effets contraires.
Mills & Fey (2004), à partir d'expérimentations menées
sur des savanes sud-africaines, observent, en effet, une destruction de la
matière organique produite par la végétation et la
litière, ce qui affecte, selon Laclan et al. (2002), la
disponibilité en nutriments pour la faune du sol. Roscoe et al.
(2000) au Brésil n'observent pas de différence dans la chimie des
sols brûlés par rapport à ceux mis en défens. Ils
constatent en particulier que les teneurs en carbone et azote ne changeaient
pas significativement, mais cela peut s'expliquer par le fait que ces auteurs
n'ont pas considéré séparément la couche (0-10 cm)
très affectée et celle (10- 20 cm) qui ne l'est pas du tout.
Au-delà de ces impacts directs, selon Bruzon (1995), le
feu en éloignant ou détruisant la faune du sol, joue sur la
structure de celui-ci. Par ailleurs, l'adsorption et l'infiltration de l'eau
diminuant par suite de la destruction de la litière et du couvert
végétal, l'évaporation se trouve accrue.
2.2.1.3.3. La végétation de savane, une
végétation adaptée au feu.
Les plantes de savane, par suite de l'élimination des
espèces sensibles, sont essentiellement des espèces
adaptées au feu auxquelles s'ajoutent des espèces qui lui sont
sensibles et qui se cantonnent sur des sites habituellement
épargnés par les flammes (anciennes termitières,
affleurements rocheux, cuirasses) (César, 1992 & 1994). Hoffmann W.
et al. (2003) montrent que les espèces ligneuses de savane ont
une plus grande tolérance au feu que celles de forêt, elles
développement une écorce plus épaisse, ont une plus grande
capacité de rejeter après le passage du feu et leurs jeunes
pousses atteignent plus vite la taille de reproduction ce qui leur donne plus
de chance d'atteindre la maturité entre deux feux consécutifs.
Cette évolution vers les formations forestières se produit
progressivement par la fermeture du milieu avec la disparition de la strate
herbacée graminéenne et la densification de la strate ligneuse
dans laquelle prend place des espèces auparavant peu compétitives
du fait du feu (Bruzon, 1995).
Le feu entretient donc le cortège floristique des
savanes - le « pyroclimax » (Schnell, 1971) - le protégeant
contre la concurrence des espèces forestières, dans cette mesure
il constitue un bon outil de gestion des pâturages savaniens (Campbell,
1954 ; Dolidon, 2005) riches en Graminées surtout pérennes. Dans
nos milieux cependant, où les savanes sont fortement
dégradées avec un envahissement des écosystèmes
savaniens par des graminées annuelles, il apparaît de plus en plus
évident que la mise à feu des pâturages constitue une
menace quant à leur équilibre et une perte de fourrage
(même de faible valeur) pour le bétail (Kièma S., 2007).

Par ailleurs, il importe de noter que, si les feux sont
favorables au maintien de groupements savanicoles pyrophiles originaux,
lorsqu'ils sont intenses et répétés par contre, ils
homogénéisent la végétation en réduisant la
richesse spécifique de la strate ligneuse et/ou de la strate
herbacée (Gill, 1975 ; Briggs et al. 1998 ; Achard et
al. 2001 ; Cochrane, 2003 ; Western & Maitumo, 2004 ; Archibald et
al. 2005). Dans une certaine mesure, il en est de même pour les
effets de la pâture notamment intense (Yates et al. 2000 ;
Achard et al. 2001 ; Archibald et al. 2005) sur lesquels nous
revenons dans le point 2.2.2 suivant.
2.2.2. Les perturbations des milieux dues à la
pâture
La compréhension de la variation des réponses
des écosystèmes pastoraux à la pâture est un
préalable nécessaire à leur aménagement (Boudet,
1978 & 1991; César, 1994 ; Daget & Godron, 1995; Adler et
al. 2004). Les feux et les sols (Gaucherand, 2005) sont des facteurs
majeurs qui structurent la végétation (distribution spatiale,
traits de vie, composition floristique, etc.) des milieux en savane. Il est par
ailleurs possible de suivre les effets ou perturbations dus aux facteurs
secondaires d'origine exclusivement anthropiques comme la mise à la
culture et la pâture (César, 1994 ; Boutrais, 1996).
Il est unanimement reconnu que l'action du bétail
(pâture et broutage) provoque des modifications dans les milieux
fréquentés par les animaux (Boudet, 1978 ; César, 1992
& 1994 ; Fournier, 1994 & 1996; Boutrais, 1994 & 1996 ;
Carrière, 1996; Steinfeld et al. 1997 ; Devineau, 1999 ; Woldu
& Saleem, 2000; Gaucherand, 2005 ; Turner et al. 2005 ; Peco et
al. 2006) et modifie l'état initial (César, 1992 &
2005). La composition floristique, la richesse floristique et l'organisation
structurale des pâturages sont affectées : suivant le niveau de
charge animale, les espèces appréciées peuvent
disparaître au profit des espèces non consommées plus
résistantes ou plus adaptées aux nouvelles conditions (Daget
& Godron, 1995 ; Boutrais, 1996). La présence animale en milieu
ouvert (système culture-jachère) génère par exemple
des perturbations mécaniques, la dispersion des graines, principalement
d'adventices et d'espèces ligneuses du groupe des légumineuses
selon Devineau (1999), par endozoochorie ou épizoochorie et le
changement dans la fertilité des sols par l'apport de nutriments par
leurs excréments.
Les effets du bétail sur les milieux, au-delà
des changements observés sur la dynamique de la
végétation, touchent donc également le support
édaphique, notamment sa texture, sa structure et sa composition
chimique. Tous ces effets dépendent de l'intensité de la
pâture (charge animale et rythme) mais aussi de la saison de
présence animale et du type de sol.
En ce qui concerne la végétation, on
appréciera la perturbation causée par le bétail en
distinguant l'espèce animale (Boutrais, 1994 & 1996), le type de
milieu, et bien sûr, la zone agro écologique où le
phénomène est analysé (Boudet, 1978). Les travaux conduits
par Hoffmann (1985) dans le nord de la Côte d'Ivoire et par et
Guérin et al. (1989) au Sénégal, par exemple
illustrent le fait bien connu que les bovins composent surtout leurs rations
à partir des herbacées (75 % environ), alors que les ovins et
surtout les caprins ont des rations comprenant plus d'espèces ligneux.
Même au sein de chaque strate, la pression de pâturage n'est pas la
même sur toutes les espèces végétales ou tous les
groupes d'espèces. A ce propos,

31
les travaux de Akpo et al. (1995) et Kièma S.
(2007) indiquent que, dans la strate herbacée, l'effort de
prélèvement du bétail est ciblé sur les
graminées vivaces ou annuelles mais moins sur les légumineuses et
les autres familles ou phorbes, de sorte qu'il en résulte un changement
dans la composition spécifique des milieux et dans la diversité
végétale. Ce prélèvement orienté favorise
l'abondance relative des espèces non désirées ou non
attrayantes (les phorbes généralement) et il se produit en
quelque sorte une colonisation des milieux pâturés par cellesci au
détriment des graminées pérennes. De ce fait, les
proportions respectives des graminées et des phorbes peuvent renseigner
sur la qualité globale des pâturages (Hoffmann O., 1985). Une
confirmation de ce phénomène est donnée dans les cas
où, à l'inverse, les terres pâturées sont
abandonnées. Comparant des milieux soumis à la pâture
à des milieux homologues qui venaient d'en être soustraits, Peco
et al. (2006) montrent que jusqu'à 50 % d'espèces
originelles des milieux pâturés sont perdues du fait de cette
situation d'abandon. En ce qui concerne la strate ligneuse, la pâture
entraîne une homogénéisation (baisse de diversité
des espèces) et une densification (Boutrais, 1994).
Par ailleurs, Daget & Godron (1995) ainsi que Toutain et
al. (2001) montrent que l'action du bétail peut provoquer des
changements dans la structure de la végétation herbacée
suite à l'étalement des espèces qui la composent,
celles-ci réagissant ainsi au piétinement
répété.
Devineau (1999), étudiant le rôle
disséminateur du bétail dans le cycle culture-jachère en
région soudanienne du Burkina Faso, rapporte que les fortes variations
de la végétation du milieu suite au dépôt des
graines d'adventices et de plantes rudérales, n'est possible que dans
des milieux ouverts ou à forte emprise pastorale, ce qu'indiquent les
traces de piétinement et de grandes quantités de bouses. Ceci est
confirmé par Kièma S. (2007), qui montre que dans les milieux de
savane où la végétation est relativement dense et
où les graminées vivaces sont encore
prépondérantes, les espèces introduites ne peuvent
résister à la compétition. Notons que selon ces auteurs,
la nature des semences observées dans les fèces des troupeaux
(phorbes, adventices de culture, céréales et espèces
rudérales) serait due au fait qu'au sortir de la saison pluvieuse, les
animaux se détournent des graminées devenues pauvres en azote
(César, 1994) au profit de ces autres groupes d'espèces qui
offrent du fourrage plus appétible : plus frais avec de meilleures
teneurs en protéines. Les effets de la pression du bétail sur la
végétation sont parfois accentués par leur influence sur
les sols. Le bétail exerce sur le sol des actions physiques
(piétinements) et biologiques ou chimiques (apport de manière
organique par les excréments ou exportation par les
prélèvements sur la végétation) (Fournier et
al. 2001; Besse & Toutain, 2002). Carrière (1996) montre
que les effets du piétinement dépendent du type de sol, ils sont
généralement moins importants sur les sols secs de nature
sableuse alors que leur incidence est parfois spectaculaire sur les sols
humides riches en éléments fins comme les limons et argiles non
gonflantes. Sur de tels sols, il s'ensuit parfois un compactage (Toutain et
al. 1983 ; Gaston, 1981 ; Audru et al. 1987 in
César, 1994 ; Daget & Godron, 1995) qui se traduit par un
accroissement de leur densité, ce qui induit alors une baisse de
l'infiltrabilité (Boutrais, 1994). On comprend dès lors l'effet
nocif du piétinement intense sur les milieux car l'infiltrabilité
est un facteur écologique d'importance pour l'entretien des
activités biologiques du sol et pour les plantes (Devineau &
Fournier, 1998). En effet, selon Stark (1994), la variation de structure du
sol, fortement dépendante de sa

teneur en eau, influence sensiblement la diffusion des
nutriments, l'activité biologique et la disponibilité et
l'hétérogénéité desdits nutriments. Par
ailleurs, en ouvrant les formations végétales et en rendant
meuble la couche superficielle des sols, le bétail participe à la
baisse de leurs valeurs organiques par érosion soit éolienne,
soit hydrique. Selon Hiernaux et al. (1999), les sols sont de bons
indicateurs du niveau de pâture par les variations dans leurs teneurs en
nutriments (azote, phosphore et carbone). La teneur en éléments
chimiques des sols est en effet sensible à la pression animale
(Kièma S., 2007) puisque l'on observe une réduction de tous ces
nutriments et une augmentation du pH de l'horizon supérieur en cas de
forte présence animale, mais l'érosion éolienne et
hydrique augmente et avec elle le lessivage chimique (Devineau et al.
2009). Mais ces nutriments, surtout l'azote et le carbone, sont à
nouveau massivement produits suite à l'installation de phorbes
ubiquistes, notamment les légumineuses fixatrices d'azote,
adaptées à ces nouvelles conditions (César, 1991 ; Daget
& Godron, 1995). Begon et al. (1996) notent que les cas où
des espèces exotiques s'installent à la faveur des nouvelles
conditions créées par une perturbation sont fréquents.
L'effet modificateur du bétail sur les milieux, qui
peut être bénéfique (Boutrais, 1994 & 1996), est
parfois notable, mais bien souvent il n'est pas le seul responsable des
changements constatés (Bartolomé et al. 2000 ; Botoni,
2003). En général, il est associé à la variation
d'autres conditions notamment la mise en culture et le régime du feu de
brousse. Beaucoup d'études conduites dans des milieux et conditions
différents (Woldu & Saleem, 2000; Devoto et Medan, 2003; McIntyre et
al. 2003 ; Mysterud, 2006 ; Loeser et al. 2006; Andrieu et
al. 2007) indiquent qu'en situation de faible charge, la richesse
spécifique ne diminue pas alors que la composition floristique se trouve
modifiée (Steinfeld et al. (1997) ; ils en concluent que,
lorsque la pression de pâture est raisonnable et bien répartie
dans le temps, les animaux contribuent à la bonification des sols et
augmentent la biodiversité végétale et animale. Leur
constat est une confirmation que la diversité des espèces est
maximisée à des niveaux intermédiaires de perturbation
(Connell, 1978). Les résultats obtenus par Saré (2003) et
Kièma S. (2007) en zone soudanienne du Burkina Faso tendent
effectivement à montrer qu'en partant d'un niveau d'emprise animale
faible, la richesse floristique des milieux s'accroît avec la charge
animale, mais ces auteurs restent réservés et attribuent
plutôt leurs observations aux variations interannuelles du climat. Par
ailleurs, Boutrais (1994) fait observer qu'en zone de savane, une pâture
régulière et raisonnable enrichit les formations
herbacées, maintient les espèces pâturables et rend plus
abondants et plus verdoyants les feuillages des arbres fourragers. Ce que
confirment Boudet (1978 & 1991) et Hatfield & Davies (2006) dans des
études conduites respectivement en zone tropicale ou dans divers
horizons arides, le premier auteur constate d'ailleurs que «la
pâture crée le pâturage» et l'explique par le fait
que l'action animale est source d'amélioration au plan fourrager d'un
pâturage. Mais ceci reste valable seulement jusqu'à un certain
niveau de charge ou seuil de rupture à partir duquel le
pâturage se trouve au contraire engagé dans un cycle de
dégradation pastorale (Boutrais, 1994 & 1996 ; César, 1994)
qui peut être très rapide. C'est aussi le point de vue de Beani et
Dessi (1984) qui indiquent qu'à condition de ne pas être
excessive, la pâture en zone de savane stimule la productivité
primaire en éliminant, par une défoliation partielle, les tissus
les plus anciens qui freinent la photosynthèse. Du point de vue pastoral
donc, la pâture, si elle est intense et répétitive,
appauvrit les milieux. En effet,

33
dans un tel cas, les espèces pérennes
appétées sont remplacées par des espèces moins
recherchées (César, 1992 & 1994; Daget & Godron, 1995 ;
Boutrais, 1996), généralement à large distribution et qui
sont indicatrices de végétation perturbée (Devineau &
Fournier, 1998 ; Fournier & Devineau, 2009 ; Djenontin, 2010).
Si une telle notion de seuil de dégradation est
claire, déterminer en pratique les conditions précises à
partir desquelles le fonctionnement des écosystèmes commence
à être compromis, est difficile. Pour une bonne gestion de
l'exploitation des pâturages il serait très utile de
connaître ces seuils (Boudet, 1978), mais, au plan écologique, il
n'existe pas de méthode pour définir à partir de quand les
capacités de régénération des milieux sont
compromises. On sait seulement qu'en zone de savane, la pression du
bétail, en éliminant surtout les graminées pérennes
héliophiles, prive le feu du combustible nécessaire à la
régulation de la végétation. Rappelons que (voir
paragraphe 2.2.1.3), sur les bons sols, le résultat en est
l'embroussaillement (César, 1992; Boutrais, 1994 & 1996 ;
Yaméogo, 2005) de ces formations par la densification du couvert ligneux
notamment de la strate arbustive (on parle alors d'embuissonnement), produisant
ainsi une biomasse ligneuse plus importante. Sur les sols légers,
pauvres ou secs, les herbes vivaces disparaissent, ce qui s'accompagne d'un
épuisement du sol conduisant à une régression des herbes
dans leur ensemble et même des ligneux et conduit à une chute de
la biomasse globale produite. Il faut noter que, dans des conditions plus
arides comme au Sahel, cette baisse de biomasse s'accompagne de la
fragmentation du tapis herbacé (Boutrais, 1994 ; César, 1994).
Les préoccupations liées aux transformations
des milieux et surtout à leur gestion ont conduit les gestionnaires des
parcours à chercher à déterminer cette valeur seuil
à l'aide de la notion de capacité de charge. Ce concept, qui
définit une pression de pâture en équilibre avec les
capacités de régénération de la
végétation (Boutrais, 1994), ne fait pourtant pas
l'unanimité au sein de la communauté scientifique (Encadré
II-1) (Carrière, 1996 ; Meuret, 1993 ; Allen et al. 2011). Le
modèle n'intègre en effet pas certains facteurs comme le
comportement alimentaire des animaux au pâturage (Meuret, 1993 ;
Boutrais, 1992 & 2002), il n'est, en outre, pas opérant dans les
écosystèmes en déséquilibre des zones subarides
dans lesquels les mécanismes classiques de rétroaction (impact
négatif sur la végétation lorsque le cheptel devient
important) sont compromis ou alors se déroulent anormalement (Breman
& De Ridder, 1991 ; Illis, 1999 ; Scoones, 1995 & 1999).
Du point de vue de la biodiversité, les implications
liées à la pression de pâture en zone de savane sont en
réalité difficiles à saisir. En effet, sauf en cas de
surpâturage, la richesse floristique des milieux n'est pas toujours
diminuée, même si les cortèges floristiques sont
généralement profondément remaniés. Lorsque la
pression de pâture devient très importante, il se produit une
banalisation de la flore dans laquelle des espèces envahissantes,
généralement des ubiquistes remplacent une partie de celles des
communautés d'origine18. On
18 Une faible pression de pâture
accompagnée d'un régime de feu faible conduit au même
résultat (Daget & Godron, 1995 ; Boutrais, 1996) même si la
flore qui en résulte est différente. Dans ce cas en effet, les
ligneux finissent, grâce au jeu de la compétition, par s'imposer
et la strate graminéenne est alors dominée d'espèces
sciaphytes peu productives (Daget & Godron, 1995) et de moindre
qualité. La charge potentielle se trouve ainsi abaissée.

parle de dégradation19 verte des
pâturages, la perte de valeur pastorale (par perte des herbes de bonne
qualité) des pâturages s'accompagnant d'une densification de leur
couvert ligneux (Boutrais, 1994 & 1996 ; Daget & Godron, 1995) et donc
de leur embroussaillement. Or, comme le rappelle Boutrais (1996), ce qui
apparaît comme une dégradation aux yeux des éleveurs peut
s'interpréter comme la première étape d'un processus de
reforestation aux yeux des forestiers.
Encadré II-1: La capacité de charge, un indicateur
changeant, peu pertinent en milieu ouvert (adapté de
Boutrais, 1994).
La capacité de charge est le concept qui a
été le plus utilisé comme indicateur-clé par les
pastoralistes et les développeurs dans le cadre du suivi des espaces
pastoraux. « Elle équivaut à la quantité de
bétail que peut supporter le pâturage sans se
détériorer, le bétail devant rester en bon état
d'entretien, voire prendre du poids ou produire du lait pendant son
séjour sur le pâturage » (Boudet, 1978 & 1991).
Selon Breman & De Ridder (1991), elle correspond « au nombre
d'animaux qui peuvent être alimentés par unité de surface,
de telle manière que la production atteigne un niveau
déterminé tout en préservant la capacité de
production des pâturages ». De nos jours, beaucoup de griefs
sont faits à ce concept, qualifié d'ambigu ou de changeant (Allen
et al. 2011).
En effet, quel que soit l'objectif poursuivi, son estimation
comporte de telles difficultés que son utilisation comme base de
décision doit être considérée avec réserve
(Carrière & Toutain, 1995).
La capacité de charge prend en compte la surface du
pâturage (S), la production primaire totale (P), la part de
prélèvement qui préserve la capacité de
régénération de ce pâturage qui est exprimée
sous forme de coefficient (K) et les besoins d'un animal (Ba) Or, on estime que
les imprécisions sur l'estimation de S, P, K et même de Ba sont
toutes de l'ordre de 20 %. (Carrière, 1994; Godard, 1991 ; Boudet, 1984
; Grouzis, 1988 ; De Wispelaere & Peyre, 1988). Ces
imprécisions se cumulant, la charge animale d'un pâturage ne peut
être estimée qu'avec une forte incertitude, la fourchette de
valeurs variant « du simple au quintuple ». On voit bien que la
précision du diagnostic n'est pas suffisante.
Carrière & Toutain (1995) et Allen et al. (2011)
rappellent que la capacité de charge doit être
appréhendée en fonction des objectifs de production de
l'éleveur qui peuvent être la production de lait, la production de
viande de qualité, la croissance numérique du troupeau ou le
maintien de la biodiversité ; pour un même pâturage, et
selon ces objectifs, le nombre optimal d'animaux qu'il peut héberger va
différer. Même si l'on se borne à une vision «
éco-centrique » du problème, l'évaluation sera
différente selon que l'on recherchera en priorité:
~ le maintien de la couverture herbeuse du sol (lutte contre
l'érosion),
|
|
19 Les auteurs invitent à nuancer cette
dégradation qui n'en serait une que lorsqu'elle se
révélait irréversible. Ils rappellent en effet que la
végétation en zone aride et subaride manifeste des
capacités étonnantes de reconstitution. La dégradation
intervient seulement lorsque le système d'élevage ne
préserve plus les ressources nécessaires à leur
perpétuation (Boutrais, 1994). Par ailleurs, Daget & Godron (1995)
et Boutrais (1996) font observer que la notion en elle-même dépend
du point de vue adopté, c'est-à-dire des objectifs de
production.
· le maintien de la diversité floristique
(conservation du patrimoine biologique),
· le maintien de la valeur fourragère des parcours
(conservation des productions animales ponctuelles).
Les calculs de capacités de charge tiennent davantage
compte de la quantité de biomasse fourragère que de sa
qualité. De fait, si l'on considère que la qualité
fourragère diminue du nord au sud en Afrique au sud du Sahara (Breman
& De Ridder, 1991 ; Sallah, 1999), il est fort probable que les
capacités de charge définies de manière classique en zone
de savane soient surestimées.
Un autre facteur non moins important mais rarement pris en
compte par la capacité de charge est le comportement alimentaire de
l'animal. Or, selon Meuret (1993), ignorer cet aspect des choses c'est «
manger à la place de l'animal », c'est confondre « valeur
alimentaire » et « valeur nutritive ». Bien souvent en effet,
les valeurs des parcours sont estimées en attribuant des valeurs
calorifiques aux espèces fourragères présentes. Mais,
comme le montrent différents auteurs comme Dumont et al. (2001)
et Magda et al. (2001) dans leur travail sur des brebis au
pâturage, des plantes même de faible valeur nutritive sont
consommées en même temps que d'autres de bonne qualité.
Selon leurs observations « les brebis ne privilégient pas, comme on
pourrait le croire, les espèces les plus nutritives, mais cherchent
plutôt à maintenir une vitesse d'ingestion constante, en associant
au cours du repas, des plantes de nature très différente ».
Par ailleurs, les choix alimentaires des animaux ne sont pas absolus, ils sont
variables et renvoient à des échelles de temps allant d'un repas
(séquence ininterrompue de consommation sur parcours), à la
journée ou à des journées successives (Dumont et al.
2001).
Boutrais (1994 & 1996) de même que Dumont (1996)
notent que l'ampleur de l'impact de la pâture animale sur les parcours
dépend des espèces de ruminants considérés, des
races, du stade physiologique et de l'état de faim des animaux. Ainsi,
les petits ruminants, notamment les caprins, sont plus incriminés que
les bovins dans la dégradation des parcours. Ils ont, en effet, des
besoins énergétiques plus importants que les espèces de
grand format ramenés à leur volume digestif, ce qui les
emmène à rechercher les aliments de plus grande concentration
énergétique (Demment & Greenwood, 1988). Boutrais (1994 &
1996) a observé au sein de l'espèce bovine que certaines races
avaient des comportements plus dommageables que d'autres. Par exemple, les
vrais zébus (Bos indicus), dégraderaient moins les
pâturages que les pseudos zébus (Red Bororo, White
Fulani) très exigeants au plan alimentaire.
|
|

35
2.2.3. L'agriculture et les successions post-culturales
L'agriculture est reconnue comme un facteur majeur de
modification des milieux. Cette modification est d'autant plus marquée
que la mise en culture est encore récente (Kièma S., 2007).
L'exploitation des terres à des fins agricoles donne lieu, en effet,
à des défrichements qui occasionnent la destruction parfois
totale de la couverture végétale. Souvent ne sont
préservés que les arbres et herbes utiles comme Vitellaria
paradoxa Gaertn.f, Parkia biglobosa (Jacq.) Bent ou
Andropogon gayanus Kunth (Devineau & Fournier, 1998).

Dans leur stratégie de gestion de la fertilité
des terres cultivées, les communautés rurales des zones de savane
de l'Afrique soudanienne ont de tout temps intégré dans leurs
systèmes de production, la pratique de la jachère dans l'objectif
de permettre la reconstitution du potentiel perdu (éléments
minéraux du sol, végétation, etc.). Le système
courant d'utilisation des terres consiste en une phase de culture à
laquelle succède une phase d'abandon dès que le rendement du
travail devient faible (Fournier et al. 2001). La durée de la
jachère est variable et liée au niveau de la pression
foncière. En contexte de faible pression sur les terres, cette
durée peut atteindre 30 ans ou plus alors que dans les contrées
où la demande en terre se fait forte, certaines parcelles sont
exploitées de façon permanente sans phase de repos. La
jachère permet le retour à la savane originelle dans un temps
plus ou moins long (de 20 à 40 ans selon ces auteurs). Dans le texte
« Écologie d'une savane africaine » consacré à
la région de Bondukuy en zone soudanienne du Burkina Faso, Devineau
& Fournier (1998) permettent de comprendre les mécanismes de retour
vers le stade originel et notent que trois à quatre phases sont à
considérer entre l'abandon du champ et la pleine reconstitution.
- Dans les jeunes jachères de moins de six ans la
végétation uniforme est caractérisée, à un
an d'abandon, par des espèces herbacées adventices ou messicoles
comme Eragrostis tremula Hochst, Digitaria horizontalis
Willd, Pennisetum pedicellatum Trin et Dactyloctenium
aegyptium (L.) P. de B. Ensuite et surtout entre deux et cinq ans on
commence à observer l'installation des graminées vivaces de
transition comme Andropogon gayanus et Cymbopogon
schoenanthus (L.) Spreng alors que les premières espèces
messicoles régressent et sont remplacées par d'autres comme
Eragrostis turgida (Schum.) De Wild, Brachiaria ramosa (L.)
Stapf., etc.
- Dans les jachères d'âge moyen (de six à
vingt ans), les milieux sont plus diversifiés que les
précédents, avec une strate ligneuse plus importante en taille et
en recouvrement. Deux situations y sont observées : la jachère
présente soit des formations à Cymbopogon schoenanthus
aux endroits fortement pâturés sur sol pauvres soit des formations
à Andropogon gayanus ailleurs. Les jachères d'âge
moyen à Andropogon gayanus sont généralement les
plus répandues. Ces observations sont confirmées par Le Mire
Pêcheux et al. (2000) qui notent que, lorsque la durée de
jachère se situe entre dix et vingt ans, le milieu réunit alors
les conditions biologiques les plus favorables Andropogon gayanus.
- Les vieilles jachères (plus de vingt ans) sont de
plus en plus rares et représentent les savanes reconstituées ou
proches de l'être dans lesquelles les herbacées vivaces comme
Andropogon gayanus, Andropogon ascinodis C.B. Cl ou Schizachyrium
sanguineum (Retz.) Alst sont dominantes ou déjà bien
présentes. La végétation ligneuse, qui peut comprendre en
son sein l'espèce Isoberlinia doka Craib et Stapf, est
très développée et diversifiée. Le Mire
Pêcheux et al. (2000) font observer cependant qu'à ce
stade, l'espèce Andropogon gayanus est absente ou très
faiblement représentée, elle est éliminée sous
l'effet de la concurrence avec d'autres herbacées pérennes.

37
2.3. Les interactions entre le bétail et la
faune sauvage: quelle possibilité de compromis ?
Les aires protégées sont de nos jours presque
les seules zones où la biodiversité est encore importante et la
faune abondante. Elles mobilisent actuellement la communauté mondiale
surtout depuis le sommet de Rio de 1992. Les efforts de préservation de
la biodiversité qui mobilisent déjà bon nombre d'acteurs
en certains endroits du monde comme en Afrique de l'Est, vont être
démultipliés et des fonds (GEF, FFEM, etc.) et programmes mis en
place à cet effet. De nombreuses études (Benoit, 1999a &1998
; Toutain et al. 2001 ; Kièma S., 2007 ; Binot et al.
2006 ; Boutrais, 2008) montrent pourtant que ces écosystèmes sont
depuis longtemps partie prenante de la chaîne de pâturage
annuel20 des populations pastorales environnantes et mêmes
lointaines. Les tendances indiquent même que l'exploitation pastorale des
aires protégées est appelée à se renforcer car
l'espace ouvert est de plus en plus rare et de mauvaise qualité du fait
de l'accaparement agricole des pâturages et de la pression anthropique
d'ensemble (Binot et al. 2006).
A l'évidence, cette tendance à l'exploitation
pastorale des aires fauniques, qui n'est pas prête de s'estomper, n'est
pas favorable à la conservation; de nombreux spécialistes de la
conservation considèrent même le bétail comme une menace
pour la faune et son habitat. Wambwa (2004) montre par exemple, à partir
de ses observations au Kenya, qu'il existe des échanges de pathologies
diverses entre le bétail et la faune lorsque ceux-ci cohabitent.
Boutrais (2008) rapporte qu'au parc W du Niger, la population de buffles a
fortement chuté en 1984 par suite d'une épidémie de peste
bovine introduite par des zébus transhumants. De même, les
conservateurs des aires protégées indiquent que le bétail
serait en compétition alimentaire avec la faune sauvage, du moins avec
les espèces avec lesquelles il partage la même niche
écologique21. Ceci semble vérifié entre les
bovins et certains herbivores paisseurs comme les buffles ou les gnous (Fritz,
1995) qui ont un régime alimentaire très proche. L'argument qui
fait du bétail une menace pour la faune n'est donc pas, dans l'absolu,
faux, mais il vaut dans les deux sens. En effet, Binot et al. (2006)
ont observé que la faune sauvage exerce des influences négatives
directes (prédation, vecteurs ou réservoirs de maladies, etc.) ou
indirectes avec la restriction à l'accès aux ressources
naturelles (eau, fourrage, cures salées, etc.) sur le bétail
domestique. Par ailleurs, les éleveurs incriminent la faune sauvage,
notamment le buffle, dans la survenue de la fièvre aphteuse dans leurs
troupeaux.
Même si à partir de ces observations, des
arguments existent pour soutenir les politiques d'exclusion (Homewood &
Rodgers, 1984), des expériences de cohabitation plutôt
réussies sont également rapportées. Si la
compétition entre faune et bétail domestique ne fait l'objet
d'aucun doute, il est des cas où il n'en existe cependant pas : les
bovins cohabitent bien avec les koudous, des brouteurs avec lesquels ils ont un
recouvrement de niche très limité (Fritz, 1995). Par ailleurs,
les herbivores à régime intermédiaire, comme les
impalas,
20 Concept désignant l'ensemble des
pâturages qui sont successivement exploités par le bétail
domestique ou sauvage au cours de l'année.
21 Selon Grinnell (1914) "the niche is the
habitat requirements of one species". Pour Elton (1927) "the niche is
what is doing by a species within a community". La niche écologique
peut donc être comprise comme l'ensemble des conditions environnementales
telles qu'une espèce donnée peut former des populations
viables.

consomment les mêmes herbacées que les bovins
(Fritz, 1995 ; Fritz et al. 1996), mais les bovins sont de meilleurs
compétiteurs sur ce type de fourrage, et les impalas semblent modifier
leur régime alimentaire en fonction de la présence des bovins
(Fritz et al. 1996). Les auteurs ajoutent que les conclusions seraient
sensiblement différentes si l'on considérait les petits
ruminants, notamment les caprins, qui consomment une grande part de ligneux
dans leur alimentation. Bayer et al. (2008), par des exemples assez
illustratifs, mettent l'accent sur les complémentarités. Ainsi,
ils montrent que la girafe qui broute sur les étages supérieurs
(à partir de 5m), maintient les écosystèmes savaniens
ouverts, favorisant ainsi les arbustes, les petits arbres et les
herbacées utilisables par le bétail domestique. Ils ajoutent que
le bétail, en pâturant dans les milieux humides, y apporte des
nutriments consommés par les oiseaux aquatiques et le poisson. Dans le
même sens, Touré et al. (2001) ont observé une
coexistence sans gêne réciproque entre oiseaux et bétail
dans le parc national du Djoudj au Sénégal. Boutrais (2008) rend
compte des résultats d'une expérience de cohabitation entre la
faune et le bétail domestique conduite entre 1970 et 1980 dans une zone
au Kenya qui a entraîné une augmentation des effectifs
d'éléphants de cette localité alors que les zones
environnantes voyaient plutôt les leurs chuter. Par ailleurs, Bayer &
Ciofolo (2004) affirment que le rapport coûts/bénéfices de
la coexistence entre faune et animaux domestiques est favorable.
Tout ceci incite donc à rechercher un compromis entre
les actions de conservation et les activités pastorales, les enjeux
actuels étant d'assurer les besoins légitimes des populations
pastorales sans compromettre la préservation du patrimoine que
constituent les aires protégées dans leur diversité.
2.4. Le pastoralisme : un genre de vie, un rapport
particulier à l'espace
La sociologie du pastoralisme et celle de la
société peule qui lui est associée ont fait l'objet de
nombreuses recherches en Afrique soudano sahélienne. En effet, la
pratique pastorale a été pendant longtemps, dans cette
région comme un peu partout dans le monde, l'affaire de groupes
socioculturels précis (Lhoste et al. 1993) dont la vie est
entièrement organisée autour du troupeau. Dans la région
qui nous intéresse, elle est l'apanage du groupe ethnique Peul et de
groupes apparentés.
Pendant de nombreuses années, une abondante
littérature sur les peuples pasteurs tendait à démontrer
leur archaïsme et l'improductivité de leurs systèmes. Ainsi,
bien après les indépendances, le pastoralisme était encore
jugé inopérant dans la gestion des espaces des jeunes
États en Afrique sahélienne. Le point de vue dominant dans les
années 1970 considérait les pasteurs comme des individus
dénués de bon sens économique, adoptant des
systèmes de tenure des terres communales intrinsèquement
néfastes (Dahl, 1983 ; Moorehead & Lane, 1995 ; Steinfeld et
al. 1997 ; Wane, 2006 ; Nori, 2007) qui aboutissaient
inéluctablement au surpâturage et à la dégradation
de l'environnement. Cette posture, soustendue par la théorie de la
« tragédie des communs » (Hardin, 1968) ajoutée au
désir des jeunes états de fixer les pasteurs pour mieux les
contrôler (Jaubert, 1997), allait servir de base à des politiques,
aujourd'hui reconnues d'une impertinence totale (au moins dans leur
conception), de fixation des pasteurs et de leurs animaux (Moorehead &
Lane, 1995; Touré,

39
1997 ; Baroin, 2003)22. Le credo était en
effet que toute ressource à "accès libre" (ici l'espace pastoral)
était inéluctablement vouée à se dégrader
(Moorehead & Lane, 1995), l'hypothèse étant faite que les
différents utilisateurs de cette ressource (ici les pasteurs),
fonctionnaient comme des agents économiques de manière
rationnelle, et seraient tentés, chacun de son côté, de
tirer un profit maximal du caractère gratuit de la ressource. Cette
approche a fait son temps et il est maintenant reconnu que partout où
les institutions traditionnelles sont restées intactes ou peu
déstructurées, comme le Macina au 19è siècle
(Boutrais, 1994), les espaces pastoraux ont rarement été à
accès libre (Boutrais, 1994 & 2002 ; Riegel, 2002). Selon
Thébaud (1995), Touré (1997) et Bary (1998), dans un tel
contexte, si l'accès aux ressources est collectif, leur gestion n'en est
pas moins rationnelle et définie entre groupes sociaux.
On a montré depuis que les sociétés
pastorales ont de tout temps disposé dans leurs principes, d'une
certaine rationalité économique (Bonfiglioli, 1988) se traduisant
par un rapport particulier à l'espace et aux ressources naturelles.
L'inexistence de règles d'accès et de gestion en milieu pastoral
était une idée fausse. Il faut comprendre que, même si le
pastoralisme est basé sur la ponction et le libre accès aux
ressources naturelles (Benoit, 1998; Riegel, 2002), la réalité
est que sans règles d'accès et d'usage de l'espace et de ses
ressources, les sociétés pastorales auraient difficilement
survécu (Thébaud, 1995). Comme le fait remarquer Pélissier
(1995), « les pratiques foncières expriment la projection de la
société sur l'espace et sont largement le reflet de son
organisation et de son histoire ». Or, compte tenu du fait que
l'élevage pastoral marque faiblement l'espace, ces pratiques, qui sont
d'une exceptionnelle souplesse, ont mis du temps à être mises en
évidence. En fait, le pasteur, par le truchement de ses animaux,
entretient avec la nature un certain type de rapport excluant un processus
d'appropriation de l'espace qui pourrait compromettre son accessibilité
par tous (Benoit, 1979; D'Amico et al. 1995 ; Boutrais, 2002). Riegel
(2002) note qu'on relève rarement auprès des bergers peuls des
termes qui exprimeraient un sentiment d'appartenance à un lieu
géographique donné ou une appropriation d'un espace bien
défini. En outre, il n'existe pas de transformation volontaire de la
nature et d'accumulation de biens dans leurs projets. Au contraire, il y a une
relation directe et égalitaire avec la richesse naturelle et
l'épuisement du stock est perçu comme normal et assumé en
conséquence par le déplacement. C'est pourquoi le pasteur ne
comprend pas qu'on veuille lui refuser le droit d'accès aux aires
protégées qu'il considère comme des stocks fourragers et
comme des maillons de sa stratégie (Benoit, 1998).
Touré (1997) et Thébaud (1995), étudiant
des sociétés peules respectivement au Sénégal
(Ferlo) et au Niger oriental, rapportent que la mobilité des hommes et
des troupeaux repose sur un principe de réciprocité entre les
éleveurs ou agriculteurs sédentaires et les arrivants, des
alliances durables favorisant le partage des ressources entre plusieurs
utilisateurs suivant des accords. En outre, l'occupation et l'utilisation de
l'espace font l'objet d'un contrôle collectif qui engage la
responsabilité de tous les usagers dans la gestion des problèmes
fonciers. En réalité, dans les zones traditionnellement
d'élevage comme au Sahel,
22 Si dans le milieu des chercheurs, la pertinence
de l'élevage mobile en zone aride et semi aride est reconnue, cela ne
semble toujours pas être le cas au niveau des politiques et des services
techniques qui le considèrent toujours comme dégradant à
l'égard de l'environnement (Kossoumna Liba'a, 2008 ; Kossoumna Liba'a et
al. 2010) et mettent tout en oeuvre pour sa sédentarisation.

l'espace pastoral était « divisé » en
secteurs selon les potentialités pastorales et l'accès à
chacun des secteurs, dont certains sont soumis à des droits prioritaires
mais non exclusifs, était soumis à des règles
(périodes et ordre d'accès) (Touré, 1997 ; Riegel,
2002).
Selon donc ces auteurs, contrairement aux apparences, toutes
les ressources, dans les communautés pastorales, étaient sous des
systèmes à « accès surveillé »
jusqu'à ce que le droit de regard de celles-ci sur lesdites ressources
leur soit retiré par l'administration coloniale et les jeunes
États (Babin et al. 2002).
Un autre aspect à prendre en compte est que le
pastoralisme relève d'un système de vie (Bonfiglioli, 1988 ;
Lhoste et al. 1993 ; Daget & Godron, 1995 ; Bovin, 1999). En
effet, la mobilité, en tant que mouvement vers l'inconnu (Benoit, 1979)
ou dans un environnement hostile (Landais, 1990) et
célébré comme tel, est source de valorisation sociale et
vue comme un fait nécessaire indispensable à la survie du groupe.
Dans son rapport direct au milieu naturel, le pasteur se confronte à un
certain nombre de risques, dont les risques sanitaires du bétail, les
risques liés à la prédation par les fauves, les risques
liés aux contraventions résultantes des infractions et les
risques de conflits avec les agriculteurs (Toutain, 2001; Paris, 2002; Tamou,
2002 ; Kagoné, 2004). De nombreux éleveurs interrogés de
nos jours considèrent qu'au-delà de la recherche du
bien-être pour leur bétail, la transhumance dans les aires
protégées constitue un défi que seuls peuvent relever les
bergers braves et authentiques (Convers, 2002; Riegel, 2002 ; Paris, 2002).
Au plan écologique, la mobilité tant
reprochée à ces peuples est assimilée par Daget &
Godron (1995) et Faure (1997) à un phénomène historique
dans les zones arides et semiarides réputées difficiles ou
pauvres en ressources pastorales. Benoit (1976 & 1998) et Touré
(1997), précisant que ce phénomène s'opère dans des
milieux en « équilibre instable» où les ressources sont
dispersées, pensent qu'il s'agit d'une stratégie opportuniste
d'exploitation de la diversité et de la variabilité de l'offre en
ressource en relation avec la diversité écologique. Loin donc de
traduire seulement des stratégies de survivance, la mobilité
consisterait en un véritable « programme de nutrition »
centré sur des objectifs à atteindre.
Nous avons indiqué plus haut que l'un des reproches
majeurs faits aux peuples pasteurs, c'est de manquer de rationalité
économique. Or, cette rationalité existe, mais elle n'est pas
orientée vers le marché (Pratt et al. 1997), elle
combine des biens privés (bétail) avec des ressources publiques
(les pâturages) et réside dans la capacité des
systèmes pastoraux à s'appuyer sur la mobilité (Boutrais,
2002), Rester sur place reviendrait en effet à accepter la baisse des
rendements (chute des productions et des effectifs) et à empêcher
le milieu de se restaurer (Benoit, 1979). L'itinérance, selon Bernus
(1981), autorise la conservation d'un effectif animal maximal, objectif
essentiel pour les pasteurs qui adoptent ainsi une stratégie de gestion
du risque en contexte aléatoire (Boutrais, 1996). Ils s'assurent ainsi
d'une reconstitution plus rapide des troupeaux, donc du capital, après
d'éventuelles épizooties par rapport à des conditions
d'élevage sédentaire.
2.5. Les grandes étapes dans l'histoire de la
conservation
La volonté de protection de la nature, dont la
manifestation visible est la mise en place d'aires protégées, est
récente. Elle se matérialise pour la première fois par la
création du parc

41
national de Yellowstone (1872) aux États-Unis
(Barbault, 2002). Mais c'est véritablement dans la première
moitié du 20ème siècle que l'idée de la protection
de la nature par la « mise sous cloche » de certaines étendues
de forêts, s'impose. Ainsi, apparurent des parcs nationaux pour la
première fois en Suède (1909), en Suisse (1915), en Grande
Bretagne (1949) et en France (1928 mais surtout 1963).
La volonté émergente d'érection des
aires protégées va être traduite au plan international par
la mise en place en 1948 de l'Union Internationale de Protection de la Nature.
Cette union se transforme huit ans plus tard en Union Internationale pour la
Conservation de la Nature et des Ressources Naturelles (IUCN) avec une
évolution dans la pensée conservatrice. La vision
réductrice de la nature (milieux naturels équilibrés
à préserver) est de plus en plus abandonnée au profit
d'une perspective plus large d'utilisation sage de celle-ci et de ses fruits au
bénéfice des humains. Mais cette révolution dans la
philosophie de conservation se fait véritablement jour à partir
de 1982 (Stockholm) mais surtout de 1992 à Rio au sommet mondial pour
l'environnement et le développement durable (Weber, 2002). Un lien fort
est désormais fait entre conservation et développement à
travers la recherche de la satisfaction des besoins socio-économiques
des populations locales. Une stratégie mondiale de la conservation est
mise en place et il y est souligné la nécessité de
sauvegarder le fonctionnement des processus écologiques dans la marche
vers le développement. L'homme est ainsi mis au centre de la
réflexion à travers la reconnaissance de son droit d'accès
légitime aux ressources naturelles qu'il a souvent contribué
à gérer (Aubertin, 2005).
Cette approche rejoint celle de l'UNESCO qui, en
197223, à travers son programme MAB (Man And
Biosphère) met l'homme et ses activités au coeur des
écosystèmes à conserver (Benoit, 1997; Barbault, 2002;
Weber, 2002; Fournier, 2004) et les aires de conservation prennent
désormais le statut de réserves de biosphère. A travers ce
concept, cette structure des nations unies se préoccupait
déjà de la conciliation entre conservation des ressources
biologiques et leur utilisation durable. Le concept des réserves de
biosphère, répondant pleinement à l'approche
préconisée par la Convention de Rio, trouve alors un nouvel essor
et est réactualisé en 1995 lors de la conférence de
Séville. « Le lien entre la conservation de la biodiversité
et les besoins en développement des communautés locales y est
reconnu comme un facteur clé dans la plupart des parcs nationaux,
réserves naturelles et autres zones protégées »
(UNESCO, 1996).
C'est de cette vision qu'est née le concept de
développement durable («sustainable development») vu comme une
utilisation des ressources naturelles qui permette aux
générations présentes de satisfaire leurs besoins tout en
garantissant la possibilité pour les générations futures
d'assurer les leurs (Weber, 2002).
La mise en application de ces principes ne semble cependant
pas encore évidente dans de nombreux espaces protégés dans
lesquels l'exploitation des ressources par les populations est plutôt
considérée comme un obstacle à la conservation. Il y a une
contradiction entre « le pacte » préconisé entre les
gestionnaires et les communautés locales (UNESCO, 1995) et les
23 Pour des raisons liées notamment au niveau
des connaissances du moment sur les processus écologiques, cette
approche de la conservation (dite éco développement) n'a pas
été adoptée tout de suite.

interdits d'accès et d'usages qu'on leur
impose24. Beaucoup d'espaces protégés sont, par
ailleurs, classés sous divers statuts, qui semblent parfois en
contradiction : il paraît, en effet, difficile de gérer de
façon cohérente un site classé à la fois Patrimoine
Mondial25 de l'Humanité - donc soumis à des
règles et des normes traduisant une appropriation étatique ou
internationale (Luxereau, in Cormier-Salem et al. 2002) - et
également Réserve de Biosphère - donc impliquant
l'adhésion et la participation des communautés locales au
processus de conservation.
L'autre saut majeur dans l'évolution
épistémologique de la pensée conservatrice est le
changement d'échelle. Il ne s'agit plus de protéger des
espèces sauvages symboliques dans des îlots de réserves
naturelles mais plutôt de sauvegarder les grands
écosystèmes (Barbault, 2002; Génot & Barbault, 2004;
Aubertin, 2005). Un courant de pensée considère en effet que,
face aux changements climatiques, le monde de la conservation devrait
être conçu comme un système de zones complémentaires
reliées entre elles par des corridors et permettant à la faune
d'exploiter toute la diversité des milieux et donc de s'adapter
(Fournier, 2004).
24 Kaboré(2010) parle d'ailleurs, à
ce propos de résurgence à l'orée du
21ème siècle d'un courant conservationniste dont les
tenants seraient Oates (1999), Terborgh (1999) et Chapin (2004) et qui, arguant
des échecs des approches participatives de gestion, prône un
retour à la protection stricte.
25 Dans l'approche éthique de la
conservation, la biodiversité est considérée comme un
patrimoine naturel. Un patrimoine est un bien chargé d'une forte valeur
à caractère utilitaire et symbolique et qui doit être
transmis aux générations futures (Boutrais, 2002). Il a une
valeur identitaire pour le groupe en assurant le lien
intergénérationnel.
CHAPITRE III

43
CARACTERES PHYSIQUES, HUMAINS ET SOCIO -
ECONOMIQUES DE LA PROVINCE DE LA TAPOA

44
3.1. La zone d'étude et sa localisation
Le terroir de Kotchari (carte III-1), notre zone
d'étude, est situé dans la région Est du Burkina Faso,
précisément dans la partie méridionale de la province de
la Tapoa dans la commune rurale de Tansarga entre les 11,68° et
11,95° de latitude Nord et les 1,87° et 2,03° de longitude Est.
C'est un terroir qui s'étend de la chaîne du Gobnangou au nord, au
poste forestier de Kondio situé entre le parc W et la réserve
partielle de la Kourtiagou. Il est adossé sur sa partie Est à la
réserve de biosphère transfrontalière du W, la plus vaste
des aires protégées de la zone avec laquelle il partage une
trentaine de kilomètres de frontière. La superficie
estimée26 est d'environ 350 km2 et, sur la base de
la définition du terroir empruntée à Bonnet
(1990)27, on peut l'assimiler à un
Sokun28 (ensemble de villages organisés en
réseaux plus ou moins complexes) flou (Turco, 2004). On y rencontre un
village-mère qui est Kotchari-centre à partir duquel se sont
constitués les autres quartiers ou hameaux à la faveur de la
recherche des terres de cultures. Cependant, si la majeure partie des villages
du terroir est issue de Kotchari-centre dans lequel beaucoup de migrants
agricoles conservent des résidences secondaires et même des
champs, on remarque que certains hameaux surtout du sud du terroir (Gnimboama,
Mangouel, etc.) ont été créés par des migrants
venus de divers horizons, notamment des terroirs de Madaga, Tambaga et
Logobou.
Sur le plan phytogéographique, le terroir appartient
au centre régional d'endémisme soudanien (White, 1986) et est
à cheval entre le secteur Nord-soudanien où la pluviosité
annuelle varie entre 700 et 900 mm et le district de la Pendjari (zone
sud-soudanienne) où elle excède 900 mm (Fontes & Guinko,
1995) (carte III-2).
26 Les limites du terroir ont été
déterminées de manière participative en associant des
personnes ressources du village. Elles restent en conséquence
approximatives.
27 Selon cet auteur, le terroir, désigne un
ensemble spatial agronomiquement homogène caractérisé par
une même structure et une même dynamique écologique, ainsi
que par un même type d'aménagement agricole
28 L'auteur distingue trois types de Sokun
ou réseaux villageois allant du plus simple au plus complexe. Ce sont :
- le réseau villageois classique : centré sur un
village-mère et basé sur des relations de type
foncier,
politique et religieux en principe hiérarchiques et
fondées sur l'antériorité.
- le réseau éco-fonctionnel : ensemble de villages
et terroirs villageois dont les relations sont produites et
entretenues par une ressource naturelle commune exploitée
ou gérée par les différents acteurs. Ce Sokun est
le plus souvent centré sur un ou plusieurs villages- pivot.
- le réseau flou : Sokun dont les villages
constitutifs sont liés entre eux par des relations d'origine et de
stabilité extrêmement variables allant de la
dépendance hiérarchique aux relations d'homologie. Le terroir de
Kotchari est un Sokun structuré en réseau flou.
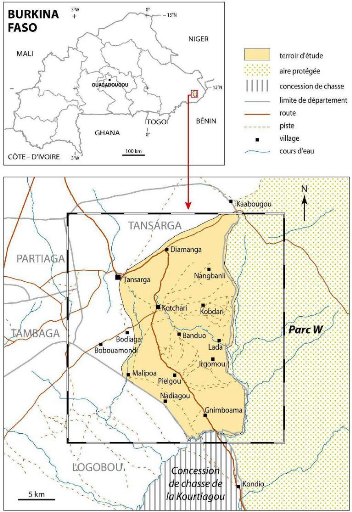

Carte III-1. Localisation du terroir d'étude

46
Carte III-2. Carte des domaines phytogéographiques du
Burkina Faso (Kièma S., 2007)
3.2. Le milieu biophysique
3.2.1. Les caractéristiques cimatiques
Le climat de la province de la Tapoa est de type
Nord-soudanien et se caractérise par l'alternance d'une saison
sèche et d'une saison pluvieuse. Ces saisons sont de durées
différentes et très variables généralement de sep
à neuf mois et de trois à cinq mois respectivement. La saison
sèche elle-même comprend une saison sèche froide et une
saison sèche chaude.
- La saison sèche froide
(novembre-mars) correspond à la période de la saison sèche
pendant laquelle le temps est frais. Les mois les plus frais sont les mois de
janvier et février et la température minimale moyenne
enregistrée est d'environ 15°C à Diapaga.
- La saison sèche chaude (avril- mai)
est la période la plus chaude de l'année avec des
températures moyennes maximales tournant autour de 35°C et des pics
dépassant parfois 40°C.
- La saison pluvieuse (ou hivernage) est la
période de survenue des averses, elle correspond à la saison
agricole. La courbe de la figure III-1 montre une répartition
inégale des précipitations dans le temps qui se concentrent
pendant et autour du mois

d'août. A côté de cette variabilité
temporelle, il est aussi observé d'un endroit à l'autre une
variabilité spatiale.
- Notons par ailleurs que la saison
sèche se caractérise par des vents venant du nord-est
chargés de poussière (harmattan) tandis que la saison pluvieuse
est surtout marquée par des vents de mousson qui repoussent le front
intertropical vers le nord et qui sont à l'origine des
précipitations.
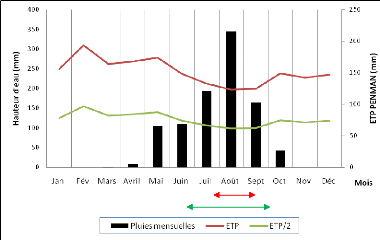
PH
PA
Figure III-1 : Bilan hydrique et période active de
végétation de l'année 2009 de la province de la Tapoa
(Sources : DPAHRH/Tapoa et DMN).
PH : période humide ;
PA : période active de végétation ; ETP :
évapotranspiration potentielle.
La période active de végétation
correspond par définition à la période pendant laquelle la
pluviosité mensuelle est supérieure à la moitié de
l'ETP (FAO, 1978). Durant l'année 1999 (figure III-1) elle a
été de trois mois et demi, soit de mi-juin à septembre.
Elle se superpose presque à la saison pluvieuse et l'on considère
qu'en cette période, les besoins des plantes sont couverts. La
période humide qui correspond à la période pendant
laquelle la pluviosité est supérieure à l'ETP a
été, quant à elle, d'environ deux mois allant de
mi-juillet à miseptembre. Rappelons que l'ETP correspond à la
quantité totale d'eau rejetée dans l'atmosphère par la
transpiration de la végétation (phénomène
physiologique) et l'évaporation du sol humide (phénomène
physique) (Breman et De Ridder, 1991).
La pluviosité est donc un facteur déterminant
pour l'activité de la végétation et pour la
productivité primaire des milieux (Schnell, 1971), d'où
l'importance de suivre son évolution pendant un temps suffisamment long
afin de comprendre les fluctuations de la disponibilité
fourragère. Dans notre zone d'étude et pendant la période
allant de 1997 à 2008 (figure III-2), les hauteurs d'eau annuelles
enregistrées ont varié entre 652,8 mm en 2001 et 1113 mm en 1999,
la moyenne de toute la période se situant à 791,3mm pour un
nombre moyen de jours de

48
pluies de 43. Le nombre d'années enregistrant des
hauteurs d'eau annuelles inférieures à la moyenne de la
période est plus élevé (sept années sur douze) que
celles pour lesquelles les écarts à la moyenne sont positifs. La
région est donc engagée dans un processus chronique de baisse de
la pluviosité qui est du reste confirmé par la courbe de tendance
mais aussi par les travaux de Santoir (1999) et Kpoda (2010) portant
respectivement sur l'expansion de l'élevage dans la zone et sur la
vulnérabilité et l'adaptation des éleveurs aux changements
climatiques. Le premier auteur montre en effet que la tendance à la
baisse est assez ancienne et qu'entre 1920 et 1960, l'isohyète 900 mm
qui matérialise la limite nord de la zone soudanienne (limite d'avec la
zone soudano-sahélienne) s'est déplacée de plus de 200 km
vers le sud (carte III-3).
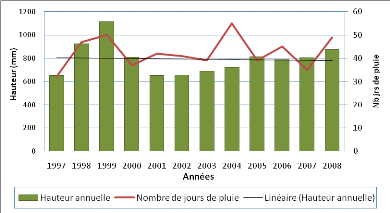
Figure III-2 : Variation interannuelle de la pluviosité
et du nombre de jours de pluies dans la commune de Tansarga entre 1997 à
2008 (sources : DPAHRH et DMN)
Kpoda (2010), s'appuyant sur les normales 1941-1970 et
1971-2000 dans les communes de Diapaga (50 km au nord de notre terroir) et de
Kantchari (100 km plus au nord) confirme le discours tenu par ailleurs par les
éleveurs eux-mêmes sur le climat qui tendrait à
s'assécher. Il a montré (figures III-3 & III-4) que de la
première à la deuxième normale, il y avait une chute de la
pluviosité moyenne (de 869 mm à 748 mm à Diapaga et 813 mm
à 674 mm à Kantchari) plus importante à Kantchari (17,1%)
qu'à Diapaga (13,7%) accompagnée d'un recul du moment de la
survenue des premières pluies et, par voie de conséquence, de la
période de début d'activité de la
végétation. L'auteur observe par ailleurs que les saisons
pluvieuses, qui s'installent progressivement dans le sens sud-nord, sont de
plus en plus capricieuses (forte variabilité) et de moins en moins
longues.
Chapitre III. Caractères physiques, humains
et socio-économiques
1200
400
200
0
3941 3942 1943 1944 1945 1946 3947 1948 3949 1950
1951 1952 1953 1954 1955 1956 3557 1958 1959 1960 3.961 3962 3963 3964 3965
3966 3967 3968 3969 1970
Années
Haters (rrrri
1000
800
600
Diapaga (mm) Moy. Diapaga (mm) Kantchari (mm) Moy. Kantchari
(mm)



Figure III-3. Évolution des hauteurs annuelles de pluies
de la normale 1941-1970 (source : Kpoda, 2010).
1971 1972 1973 1974 1975 195 1977 1978 1979 1933 1931
1932 1933 1934 1935 1595 1937 1593 1933 1930 1931 3592 1933 1934 1935 1936 1997
1933 1939 20:0
Années
Hatear(mr)
1400
1200
1000
800
400
600
200
0
Diapaga (mm) Moy. Diapaga (mm) Kantchari Moy. Kantchari (mm)



Figure III-4. Évolution des hauteurs annuelles de pluies
de la normale 1971-2000 (source : Kpoda, 2010).
Par ailleurs, les moments de sécheresse entre les deux
périodes de référence (1941- 1970 et 1971-2000) sont
devenus plus fréquents et ont tendance à être plus longs
(tableau III-1); parallèlement, les précipitations de faible
ampleur (0 - 20 mm) prennent un peu d'importance au détriment de celles
à ampleur moyenne (20 - 40 mm). Enfin, l'auteur a noté une
tendance à l'accroissement des températures moyennes mensuelles
entre les deux normales.
Tableau III-1. Caractéristiques des séquences
climatiques sèches sur les normales 1941-1970 et 1971-2000 pour une
saison pluvieuse (source : Kpoda, 2010).
Normale Stations Séquence sèche de
Total
7 jours ] 7-10 Jrs] ] 10-15 jrs] > 15 jrs
Diapaga 12 4 4 2 22
1941-1970

49
Kantchari 14 6 3 3 26

50
Diapaga 12 14 6 5 37
1971-2000
Kantchari 15 9 3 4 31
La variabilité climatique et la péjoration
climatique ainsi observées montrent que la zone est vulnérable et
menacée par un assèchement de son climat. Signalons cependant que
cette baisse n'est pas continue (Kièma S., 2007) et que par moments,
comme entre 1990- 1996, on observe une remontée des moyennes
pluviométriques annuelles (Santoir, 1999).
Notons enfin que l'insolation tourne entre 7-8 h/jour en moyenne
tandis que l'humidité de l'air est soit moyenne, soit faible (Kuela,
2000).
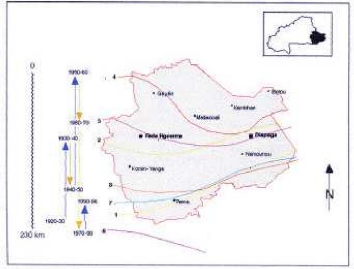
Carte III-3. Variation en latitude de l'isohyète 900 mm
(source : Santoir, 1999)
3.2.2. La géomorphologie et les sols
Le relief de la province de la Tapoa fait partie de l'immense
pénéplaine dont les roches datent du Précambrien (Hottin
& Ouédraogo, 1976). Il est dans l'ensemble faiblement ondulé
et dominé par des glacis, des buttes cuirassées et des collines
de grès. Deux sous-ensembles caractéristiques peuvent être
distingués :
- la moitié septentrionale, étendue
relativement plane dont l'altitude varie entre 220 et 250 m, - la moitié
sud, moins monotone, présentant de nombreuses buttes gréseuses et
cuirassées, les falaises de Gobnangou. Le point culminant de la province
se trouve dans cette partie et est de 351 m.

Les premières caractérisation de la
géomorphologie et des sols de la région de l'Est et
précisément de la province de la Tapoa ont été
conduites par Boulet (1968), Kaloga (1968), Boulet & Leprun (1969) à
de petites échelles (1/200 000 à 1/500 000). S'en sont suivies
celles à échelles plus détaillées menées par
le bureau national des sols (BUNASOLS) (1979, 1980, 2000 & 2007).
La nature des sols est le reflet des facteurs
géomorphologiques et de la nature du substrat d'altération. Par
ailleurs, les processus pédogénétiques dominants sont le
lessivage et les altérations géochimiques marqués par la
kaolinisation, la vertisolisation et la ferruginisation (Boulet, 1975 ). Ces
processus qui résultent des transformations verticales et
latérales conduisent dans la Tapoa à une juxtaposition de roches
sédimentaires et de roches cristallines (Kuela, 2000). à partir
de ce substrat géologique se sont développés plusieurs
types de sols. Ainsi, six unités distinctes sont rencontrées sur
le territoire provincial. Ce sont:
- les sols à minéraux bruts ;
- les sols peu évolués ;
- les vertisols ;
- les sols à sesquioxydes ;
- les sols à mull des pays tropicaux ; - les sols
hydromorphes.
Ces sols sont géographiquement inégalement
répartis en trois ensembles.
· Les sols à sesquioxydes principalement de types
ferrugineux tropicaux lessivés dominent l'est et le nord de la province.
Ils sont d'une fertilité médiocre et à vocation surtout
pastorale.
· Les sols peu évolués dominent le
Nord-ouest, le centre et l'ouest de la province.
· Au sud, on rencontre des sols à sesquioxydes, des
sols à minéraux bruts, et des sols hydromorphes de
fertilité moyenne propices aux cultures.
Les sols du terroir de Kotchari, qui est situé
justement dans cette partie méridionale de la province, sont de
plusieurs types avec des caractéristiques, des aptitudes et
potentialités agronomiques assez diverses mais
généralement faibles à moyennes (cf. tableau III-2). Ils
sont présents presque partout dans l'espace du terroir, excepté
les lithosols sur cuirasse et buttes qui se concentrent dans la partie nord et
qui correspondent à la chaine du Gobnangou (carte III-4).
Chapitre III. Caractères physiques, humains et
socio-économiques Tableau III-2. Caractéristiques
des classes de sols dans le terroir de Kotchari (Sources : BUNASOL, 2007)
Type de sols Description Type Aptitudes agronomiques
Superficies et
géomorphologique proportions
dans le terroir
BEHV (58%), BEF (30%), BEPE (7%) Haut de pente de Bruns Eutrophes
glacis
Hydromorphes à faciès BEHV (47%), BEF (31%), VV
(13%), Bas de pente de
Vertique BEH (9%) glacis
6,97 km2
(1,99%)
Moyennement apte aux cultures pluviales de
céréales, d'oloprotéagineux, de coton et de cultures
fourragères
Marginalement apte au riz pluvial et irrigué et à
la maraîcheculture
|
Ferrugineux tropicaux Lessivés indurés à
Tâches et Concrétions
|
FLTC (63%), FLM (26%), FLI (11%) Bas de pente de
glacis
|
Moyennement apte aux cultures pluviales d'oloprotéagineux
(sésame, arachide, niébé) et de cultures
fourragères Marginalement apte aux céréales et au riz
pluvial et irrigué
|
62,56 km2
(17,89%)
|
FLI (82%), Lithosols (18%) Plateau cuirassé Moyennement
à marginalement apte aux
|
Ferrugineux tropicaux Lessivés Indurés
|
FLI (78%), FLC (11%), FEER (6%), PEACM (5%)
FLI (63%), FLTC (27%), FLM (6%), FLC (4%)
|
Haut de pente de glacis
Moyenne pente de glacis
|
cultures pluviales de céréales,
d'oloprotéagineux et de cultures fourragères
Permanemment inapte aux cultures pluviales ou irriguées
de riz et à la maraîcheculture
|
197,15 km2
(56,38%)
|
HPGS (82%), BEH (13%), VV (3%), BEHV (2%)
HPGS (74%), PEAAH (21%), FLH (5%)
HPGS (52%), PEAAH (33%), FLH (12%), PEACAM (3%)
Plaine alluviale Apte à moyennement apte aux cultures
de
riz irrigué et pluvial de maïs, au sorgho, Bas-fonds
au niébé, aux cultures fourragères et à
la
maraîcheculture
Vallons colluviaux Marginalement apte au sésame et inapte
au mil, arachide et coton
Hydromorphes peu Humifères à Pseudogley de
Surface
24,11 km2
(6,89%)
Lithosols sur cuirasse ou sur Lithosols (100%) Butte
cuirassée ou Permanemment inadapté à toute 58,92
km2
rocheuse rocheuse exploitation (16,85%)
BEHV = sols bruns eutrophes hydromorphes faciès vertique;
BEF = sols bruns eutrophes ferruginisés; BEPE = sols bruns eutrophes peu
évolués; BEV = sols bruns eutrophes vertiques; FLC = sols
ferrugineux lessivés à concrétion; FLH = sols ferrugineux
lessivés hydromorphe; FLI = sols ferrugineux lessivés
indurés; FLM = sols ferrugineux lessivés modal; FLTC = sols
ferrugineux lessivés à taches et concrétions; HPGS = sols
hydromorphes peu humifère à pseudogley de surface; PEAAH = sols
peu évolués d'apport alluvial hydromorphe; PEACAM = sols peu
évolués d'apport colluvial alluvial modal; PEER = sols peu
évolués d'érosion régosolique; VV = vertisols
vertiques.

54
Chapitre III. Caractères physiques, humains et
socio-économiques Ces sols sont présentés
ici par ordre d'importance décroissante.
- Sols ferrugineux lessivés indurés.
Généralement superficiels (sur les plateaux
cuirassés et les hauts glacis) et parfois profonds (sur les moyens
glacis), ils sont les plus dominants (56,38%). Sur les moyens glacis, ils sont
très souvent associés à des taches et concrétions.
Ce type de sol est apte aux cultures pluviales de céréales, de
sésame, d'arachide et de niébé.
- Sols ferrugineux tropicaux lessivés indurés
à taches et concrétions
Couvrant 17,89% du terroir ils sont situés dans les bas
glacis avec une texture sablo limoneuse en surface et argilo sableuse à
argileuse en profondeur. L'état de surface est sableux. Ces sols
présentent une aptitude pour les cultures pluviales
d'oloprotéagineux (sésame, arachide, niébé). Ils
sont par contre peu aptes à la céréaliculture et à
la riziculture pluviale ou irriguée.
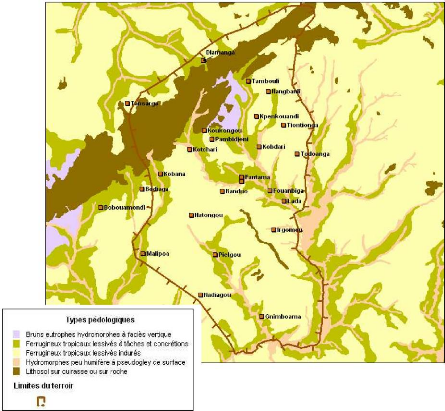
Carte III-4 : Sols de la région du terroir de Kotchari
(Données : BUNASOLS, 2007)

55
- Lithosols/r
Couvrant 16,85% de la superficie et situés sur les
buttes rocheuses de la chaine du Gobnangou et sur les cuirasses environnantes,
ils sont généralement inaptes aux activités agricoles et
servent de zone de pâture villageoise traditionnelle. Cependant, avec la
pression sur les terres en contrebas, on observe que quelques champs agricoles
sont mis en place sur les parties basses de ces sols.
- Sols hydromorphes peu humifères à pseudogley
de surface (6,89%) des bas-fonds, plaines et vallons colluviaux.
Ils sont, de par leur position topographique, les plus
profonds et les plus fertiles. Ils sont en conséquence indiqués
pour les cultures de riz pluvial et irrigué, le sorgho et le
niébé. Ils sont par ailleurs modérément aptes pour
le mil, le maïs et le sésame et inaptes pour l'arachide et le
coton.
- Sols bruns eutrophes hydromorphes à faciès
vertique
Ils sont marginalement rencontrés sur le terroir de
Kotchari (1,99%). Rencontrés surtout sur les hauts et bas glacis, ils
peuvent supporter des spéculations comme les céréales, les
oloprotéagineux et le coton et parfois le maraichage dans les parties
les plus basses.
3.2.3. Le réseau hydrographique
Deux grands bassins versants collectent les eaux de surface de
la province de la Tapoa. Ce sont le bassin versant du fleuve Niger au Nord et
celui de la Pendjari au sud. La ligne de partage se situe aux environs de la
latitude de Tansarga.
Dans le Nord, on rencontre les rivières suivantes :
- le Diamangou et ses affluents dont le plus important est la
Boudiéri dans la partie septentrionale de la province,
- la Tapoa dont la province tient son nom, orienté d'Est
en ouest avec ses deux affluents que sont : le Kpenboanga le Borofwanou. Elle
est située dans la partie centrale de la province,
- le Goulbi et son affluent le Bargou,
- le Mékrou,
- et le Tyénétiégal.
Dans le sud de la province, la Pendjari (180 km de long) qui
constitue une frontière naturelle entre le Burkina d'une part et le
Bénin et le Togo d'autre part, se compose du réseau suivant : -
le Bokouongou,
- le Doubodo qui collecte les eaux du Bopiéna et du
Moribonga,
- la Kourtiagou dont les affluents sont le Bonulogodi et le
Pendjo,
- et l'Arly.
Malheureusement, tout ce réseau hydrographique ne
présente qu'une activité saisonnière, à l'exception
de la Tapoa et du Mékrou qui ont, par endroits, des points d'eau
stagnante tout au long de la saison sèche (Paris,
2002).

Le terroir de Kotchari (carte III-5) qui relève du
bassin de la Pendjari est traversé par quelques cours d'eau saisonniers
relevant tous de la Kourtiagou situé à la limite sud du terroir.
La plupart de ces cours d'eau s'assèche au cours de la saison
sèche froide. Seuls les plus importants, c'est-à-dire le
Kotchalpoundougou (Kotchari-centre), la Kourtiagou et le
Pendjo, deux affluents du fleuve Pendjari, atteignent la saison
sèche chaude avant de s'assécher.
3.2.4. La végétation et la flore
On doit à Guinko (1984), Fontes & Guinko (1995),
Thiombiano (1996), Thiombiano (2005) et Hahn-Hadjali (1998), les principales
études de caractérisation de la végétation de la
région de l'Est du Burkina Faso. On en retient que les formations
végétales sont de type savanicole et que leur physionomie est
souvent tributaire de l'impact des activités anthropiques (Mbaygone,
2008), de la nature physicochimique des sols et de la géomorphologie.
Les savanes arborées mais surtout arbustives y sont les plus
répandues ; elles correspondent à des groupements
végétaux dominés par les espèces de la famille des
Combretaceae. Des savanes herbeuses y sont aussi observées dans les
endroits à haut degré d'anthropisation.
Par ailleurs, des reliques de forêts sèches
à Anogeissus leiocarpa, se présentant sous-forme
d'îlots de boisement dense ou de bandes étroites de forêts
claires, sont enregistrées le long des cours d'eau (Traoré, 2008
; Ouédraogo O., 2009).
La végétation de la commune rurale de Tansarga,
de laquelle relève notre terroir d'étude, est très
contrastée en liaison avec un relief également diversifié.
Les savanes arbustives y dominent, mais on rencontre aussi des savanes
herbeuses (sur les plaines et bas-fonds où la très forte pression
anthropique a fini d'éliminer ou de réduire fortement la
couverture ligneuse) et arborées (dans les bas-fonds et plaines
marécageuses à sols profonds favorables aux espèces
hydromorphes des genres Terminalia et Cyperus sp.).
Les ligneux les plus courants appartiennent à la
famille des Combretaceae (Combretum collinum, Combretum nigricans,
Combretum glutinosum, Terminalia sp.). Ils sont accompagnés
d'espèces de la famille des Mimosaceae (Acacia gourmaensis), de
celle des Rubiaceae (Feretia apodanthera, Gardenia ternifolia, Mitragyna
inermis et Nauclea latifolia), des Anacardiaceae (Sclerocarya birrea,
Lannea acida et Lannea microcarpa) et des Caesalpiniaceae (Burkea
africana, Detarium microcarpum, Piliostigma reticulatum). Les
herbacées sont fortement dominées par les Graminées
annuelles avec lesquelles cohabitent les Cyperaceae (Cyperus sp.,
Abildgaardia sp., Kyllinga
sp. et Sclerea sp.) et les
Fabaceae.
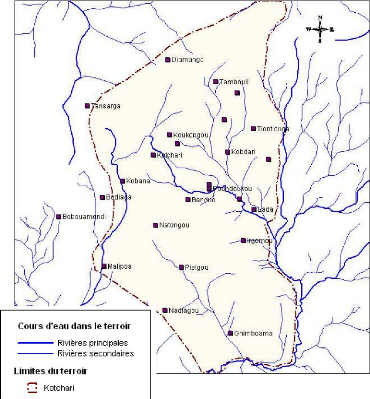

57
Carte III-5. Réseau hydrographique dans le terroir de
Kotchari
3.2.5. Les réserves fauniques : le parc national du
W et la concession de chasse de la Kourtiagou.
3.2.5.1. Le parc national du W
Le parc du W (carte III-6) est un ensemble transfrontalier
vaste de plus de 10 000 km2 que se partagent le Bénin (5
632,8 km2), le Burkina Faso (2 350 km2) et le Niger (2
260 km2). Il doit son nom à la sinuosité en "W" que
forme le fleuve Niger à la limite nord de la réserve du
coté nigérien (Convers, 2002). Comme le rappelle Paris (2002), la
partie burkinabè du parc est limitée par l'axe Tapoa-Djerma,
Kabougou, Kondio à l'Ouest, la piste allant de Tapoa-Djerma à
Tapoa (Niger) au Nord, les rivières Tapoa, Mékrou et la
chaîne de l'Atakora au Sud.
L'histoire de la partie Burkinabé du parc W est presque
similaire à celle de la plupart des aires protégées de la
sous-région ouest-africaine. Déjà en 1913, les premiers
actes juridiques du pouvoir colonial français introduisaient un
règlement provisoire pour la chasse à l'éléphant
dans la zone. Ensuite, par l'arrêté du 16 avril 1926, était
créé le parc refuge des cercles de Say

et de Fada sur une superficie de 5 430 km2 qui
allait devenir par la suite successivement Réserve Totale de Faune par
arrêté n°2606/SE/F du 14 avril 1953 puis Parc National du W
par décret du 4 août 1954 du gouvernement de l'Afrique Occidentale
Française (Benoit, 1998).
Ce parc est inscrit sur la liste du Patrimoine Mondial de
l'Humanité de l'UNESCO et classé en 2002 comme Réserve de
Biosphère de l'UNESCO. Il est, depuis le 27 octobre 1990, classé
comme site de Ramsar.
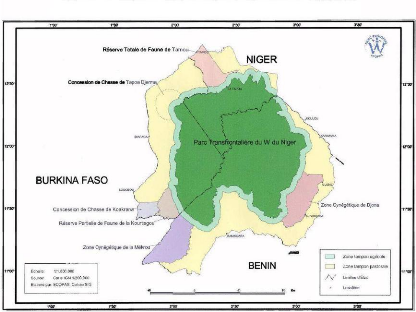
Carte III-6 : L'organisation spatiale du Parc W et de sa
périphérie Burkinabé
3.2.5.2. L a concession de chasse de la Kourtiagou
La concession de chasse de la Kourtiagou (ou de Kondio) qui
est vaste de 51.000 km2, correspond à la réserve
partielle de faune de ladite localité concédée en gestion
privée et qui a été créée en 1957 (Babin et
al. 2002) par l'administration coloniale. Elle est située
à cheval entre les parcs nationaux du W et d'Arly et est bordée
au sud par le parc national de la Pendjari du Bénin (ACRA, 2009). La
faune de grand port est peu importante à l'exception de l'hippotrague,
par contre on y rencontre en nombre important des antilopes de moyenne et de
petite taille, sauf le cobe de Buffon, le céphalophe à flancs
roux et l'ourébi. La forte prévalence du braconnage dans la zone
et la position géographique de cette aire (elle est contiguë au
couloir de transhumance officiel n°3 qui passe par Kotchari en direction
du Bénin) l'expose à une forte pression de chasse et de
pâturage.

59
3.3. Le milieu humain
La province de la Tapoa est peuplée essentiellement par
deux groupes socioculturels que sont les Gourmantchés (84,44%) et les
Peuls (10,92%) (DRED-Est, 2008). A côté de ces groupes dominants
on rencontre les Djerma, les Haoussa et les Mossi beaucoup plus
rencontrés dans la partie nord de la province dans les communes de
Botou, Kantchari et Diapaga dans une moindre mesure (Sanou, 2005). La
population, dont la croissance annuelle est supérieure à la
moyenne régionale (3,8% contre 3,65%), était estimée en
2006 à 342 305 habitants, soit 23,45 habitants/kilomètre
carré29. Elle est essentiellement rurale avec 97,55% de
ruraux et à dominante féminine (50,46% de femmes) surtout en
milieu rural.
Le tableau III-3 rend compte de la situation
démographique actuelle des différentes communes de la Province de
la Tapoa. Les données générales sur la population sont
obtenues par estimation en partant des données du recensement
général de la population et de l'habitat de 2006. Afin de mieux
rendre compte de la situation foncière réelle, la superficie des
aires protégées ont été déduites pour
effectuer les calculs.
Les données montrent un gradient Nord-sud avec une
pression démographique beaucoup plus forte au Sud de la province, zone
où la population se concentre. La densité moyenne y est en effet
assez forte et largement au-dessus de la moyenne provinciale, elle
représente plus du double de celle observée au Nord. Dans la
commune de Tansarga dont dépend le terroir de Kotchari, l'estimation de
la population est de 42 190 habitants avec une densité moyenne de 71,19
habitants/km2. La population de cette commune, la
3ème plus peuplée par unité de surface
(après les communes de Namounou et Logobou), représente, à
ce jour, un peu moins de 11 % de la population totale de la province.
Tableau III-3. Densité de population dans la province de
la Tapoa
|
Communes
|
Superficie
utile
(km2)30
|
Population en
2010
|
Densité
(hts/km2)
|
Densité
moyenne
(hts/km2)
|
|
|
Nord de la Province
|
|
|
|
Botou
|
1750,8
|
54514
|
31,14
|
|
|
Diapaga
|
1282,6
|
37978
|
29,61
|
28,78
|
|
Kantchari
|
3620
|
68451
|
18,91
|
|
|
Partiaga
|
1648
|
58467
|
35,48
|
|
|
|
Sud de la Province
|
|
|
|
Logobou
|
893
|
71299
|
79,84
|
|
|
Namounou
|
56,1
|
17262
|
307,69
|
70,34
|
|
Tambaga
|
790,5
|
47408
|
59,97
|
|
29 Cette valeur doit être
reconsidérée puisqu'elle avait pris en compte les superficies des
réserves qui ne sont pourtant pas accessibles.
30 Superficie totale moins superficie des aires
protégées

Tansarga 592,6 42190 71,19
Tapoa 10634 397570 37, 39
(Sources: D'après BNDT/IGB, 2003; DREP-Est, 2008 et
MEF, 2008)
Il faut noter, par ailleurs, que la province était
jusqu'à une période récente un lieu d'immigration agricole
surtout pour les cotonculteurs à la recherche de terres fertiles. De nos
jours, cette forme d'immigration n'existe pratiquement plus probablement
à cause de la raréfaction des terres, mais aussi à cause
de la baisse de l'engouement pour la culture du coton.
3.4. Les systèmes d'élevage
3.4.1. Importance du secteur
L'élevage est la seconde activité
socio-économique de la province de la Tapoa, il vient après
l'agriculture. C'est une importante activité économique qui
occupe presque tous les ménages ruraux de la province (Sanou, 2005)
auxquels il apporte des revenus considérables. Les espèces
animales élevées sont : les bovins (zébus et taurins), les
ovins, les caprins, les porcins, les asins et la volaille (poules, pintades, et
autres oiseaux de la basse-cour) et le cheptel y est numériquement
important et diversifié. Le tableau III-4 fait la synthèse des
effectifs de bétail pour les principales espèces
élevées dans la zone.
Tableau III-4. Effectif du cheptel et charge animale dans la
Province de la Tapoa
|
Communes
|
|
Effectif 2009
|
|
|
Total UBT31
|
Sup (km2)
|
32
Charge réelle
|
|
Bovins
|
Ovins
|
Caprins
|
Asins
|
Équins
|
(km2/UBT)
|
(UBT/ km2)
|
|
Botou
|
23799
|
43236
|
55441
|
6608
|
658
|
46835
|
1751
|
0,037
|
26,75
|
|
Diapaga
|
14726
|
38187
|
46700
|
5583
|
209
|
34145
|
1283
|
0,038
|
26,62
|
|
Kantchari
|
41753
|
38227
|
55435
|
5586
|
668
|
63388
|
3620
|
0,057
|
17,51
|
|
Tambaga
|
18909
|
21805
|
31415
|
5587
|
262
|
32050
|
791
|
0,025
|
40,54
|
|
Logobou
|
32819
|
27240
|
37415
|
5430
|
365
|
48287
|
893
|
0,018
|
54,07
|
|
Partiaga
|
43345
|
43657
|
60914
|
5731
|
564
|
67115
|
1648
|
0,025
|
40,73
|
|
Namounou
|
11509
|
36078
|
44503
|
4614
|
121
|
29592
|
56
|
0,002
|
527,48
|
|
Tansarga
|
18996
|
27267
|
39033
|
5587
|
262
|
34753
|
593
|
0,017
|
58,64
|
1 UBT = unité de bétail tropical = 1 bovin de
250 kg
En rapportant les effectifs animaux de ce tableau convertis en
UBT à la superficie de chacune des communes de la Province, on obtient
des charges variant entre un peu plus de 17 UBT/km2 (commune de
Kantchari) et 527 UBT/km2 (commune de Namounou). La valeur
exceptionnellement très élevée de la densité
animale dans la commune de Namounou est due au fait que celle-ci est une
commune « sans terre ». Dans les faits, les animaux de cette
31 Sur la base de 1 bovin = 1 équin = 1UBT ; 1
ovin adulte = 1 caprin adulte = 0,2 UBT et 1 asin = 0,4 UBT (Ministère
de la Coopération et du Développement, 1991)
32 Seules sont prises en compte les espèces de
ruminants (bovins, ovins & caprins)

61
commune exploitent les terres des communes voisines. On peut
observer que, la commune de Namounou mise à part, celle de Tansarga est
celle qui a la plus grande charge de bétail par unité de surface
avec près de 59 UBT/km2, et cela compte non tenu des animaux
transhumants et des porcins dont les statistiques n'étaient pas
disponibles. Précisons, et cela est important, que l'élevage
porcin mobilise une frange non négligeable de la population
gourmantché, surtout féminine. Remarquons par ailleurs que les
densités animales les plus fortes se rencontrent dans les communes de la
moitié sud de la province (Partiaga, Tambaga, Logobou, Tansarga et
Namounou).
Les systèmes d'élevage dominants dans la zone
sont de type traditionnel extensif. C'est un élevage essentiellement
mobile qui peut être sédentaire (élevage sur parcours
villageois) ou transhumant et qui se confronte de nos jours à de
nombreuses contraintes, dont l'insécurité foncière
pastorale et le déficit fourrager. Parallèlement à
l'élevage traditionnel sur parcours villageois (mode dominant) ou
transhumant, il émerge de nos jours, grâce aux actions des ONG de
soutien et aux projets de développement rural, un élevage de type
intensif avec stabulation. Ce type d'élevage est surtout à
objectif d'embouche qu'elle soit ovine, bovine ou porcine.
3.4.2. Les systèmes extensifs
3.4.2.1. L'élevage sédentaire
L'élevage sédentaire est pratiqué par les
groupes socioculturels autochtones (les Gourmantchés) ou anciennement
installés (les peuls sédentaires). Ce type d'élevage est
présent dans la zone depuis bien longtemps. Benoit (1998) et Santoir
(1998 & 1999) signalent sa présence dès le début du
siècle dernier. Les caractéristiques de cet élevage,
qualifié aussi d'agropastoral, sont différentes d'un groupe
socioculturel à l'autre.
Chez les autochtones gourmantchés, agriculteurs
traditionnels, les troupeaux étaient initialement de petite taille et
constitués essentiellement de petits ruminants dont la garde se faisait
surtout en saison pluvieuse et au piquet. L'intégration des bovins dans
les exploitations s'est opérée lentement avec le temps et s'est
accélérée avec les sécheresses qui ont
suscité des échanges entre éleveurs peuls et agriculteurs
gourmantchés sous-forme de troc (bétail contre
céréales) et l'adoption de la mécanisation agricole (il
fallait des boeufs de trait) plus récemment grâce à la
généralisation de la cotonculture. En 1989, 25% de bovins et 10%
de petits ruminants étaient détenus par les autochtones (Santoir,
1999). Ces taux ont dû évoluer de nos jours avec le coton. Cette
culture, en procurant des revenus importants aux agriculteurs à partir
de la fin des années 90, leur a permis d'accroitre leur cheptel
composé essentiellement à l'origine de
Berbaji33 et de Gurmaji (zébu introduit par
les peuls sédentaires) ou des produits de leur métissage
(Gurmaji x Berbaji).
Les Peuls sédentaires ou semi-sédentaires, les
Gurmaabe selon l'appellation de Santoir (1998), se sont
installés dans la zone à la suite d'un long processus migratoire
soutenu par les crises climatiques successives qu'ont connu les régions
sahéliennes. Ces éleveurs
33 La plupart des termes locaux ici utilisés
pour désigner les ethnies et les races animales sont de Santoir (1999).
Dans la suite du présent manuscrit notamment dans les chapitres V &
VI, beaucoup de termes de ce genre reviennent. Dans ce cas, leur transcription
suit les normes proposées par Olivier de Sardan (2003).

migrants se sont adonnés à l'agriculture,
certains pour la première fois (Santoir, 1998), à laquelle ils
restent de nos jours fortement attachés. On sait en effet qu'à la
suite de sécheresses et famines, les groupes pastoraux qui ont perdu une
grande partie de leur bétail se fixent ainsi à proximité
d'espaces cultivés (Bonfiglioli, 1990). Dans le terroir de Kotchari, les
Peuls sédentaires continuent d'être fortement engagés dans
l'élevage qui constitue d'ailleurs leur principale activité. La
taille des troupeaux est variable, elle est liée au rythme de
reconstitution d'un cheptel qui avait été
presqu'entièrement décimé. Ce cheptel est en
général plus nombreux que celui des troupeaux
gourmantchés, mais il est courant que, dans ces élevages,
certains animaux soient confiés en gardiennage par des agriculteurs
(constat fait également par Lhoste et al. (1993)), des
fonctionnaires ou des commerçants locaux. Les troupeaux sont à
l'origine composés majoritairement de Gurmaji mais on rencontre
aussi des Kiwali34, des Jaliji (race originaire de Torodi
et du Nigéria), des Gudali (venant de Sokoto au
Nigéria) et des Boboroji. Par ailleurs, c'est au sein de ce
groupe de Peuls que les grands éleveurs gourmantchés recrutent
leurs bergers salariés. En effet, Santoir (1999) fait observer que
« le bétail est confié par les Gurmance à leurs
Peuls et le confiage demande que Peuls et Gurmance se connaissent et aient
cohabité pendant un certain temps ».
Encadré III-1. La sédentarité des pasteurs :
entre reconstitution des forces et voyage sans retour
La question de la sédentarisation des éleveurs
pasteurs est un sujet qui fait débat de nos jours, après avoir
été expérimentée sans succès au début
des indépendances (Boutrais, 1994). En effet, les tenants de cette
solution arguent du fait qu'il y a bien des éleveurs pasteurs
aujourd'hui sédentarisés ou que de toute façon l'espace
étant en manque cette alternative s'imposerait d'elle-même. On
peut toutefois se demander si la sédentarisation évoquée
en est réellement une et, si c'est le cas, si elle est
définitive. La question sous-jacente est de savoir à partir de
quelle durée d'installation on peut commencer à parler de
sédentarité ? Plusieurs exemples montrent que la
sédentarité des pasteurs est vécue comme un mal être
par les principaux concernés. Bonfiglioli (1990) et Santoir (1999)
abordent la question en mettant l'accent sur la complexité du sujet pour
lequel toute réponse doit être nuancée. Le premier auteur,
s'intéressant aux mouvements des communautés pastorales de
l'Afrique sahélienne après les crises alimentaires de 1973 et de
1984 (Woodabe du Niger, Touareg du Mali et du Niger, arabes
Missiriye du Tchad, Maures de Mauritanie) ou à une
crise de peste bovine survenue dans un lointain passé (Woodabe
du Nigéria), qualifie les phases de sédentarisation
observées au sein de ces peuples « d'agropastoralisme d'attente
». Pour lui en effet, cette sédentarité provisoire ou de
transition des pasteurs survient après des pertes importantes du
cheptel, et elle vise juste à permettre, par la fixation des habitats,
la réduction des mouvements et la pratique de l'agriculture, d'assurer
la survie du groupe et de reconstituer le cheptel ainsi perdu avant de
reprendre, en temps opportun, la liberté. La sédentarité
apparait alors comme une stratégie de survie. A ce propos, Baroin (2003)
et Dongmo (2009) évoquent
34 Cette race bovine a été introduite
dans la zone par les peul Siwalbe (ou Kiwabe) (un lignage
relevant du grand groupe d'éleveurs Woodabe aussi
appelés Mbororo originaires du Nigeria qui se sont
installés après les indépendances dans les terroirs de
Kotchari et de Mardaga.
des stratégies d'adaptation au contexte
écolimatique et socioéconomique. Ces observations sont
corroborées par les constats faits par Santoir (1999) qui s'est
intéressé au processus d'occupation de l'espace Gurmance
par les Peuls Gurmaabe dans l'Est du Burkina. L'auteur note un recul
de la sédentarité peule dans la Tapoa dès après
1985, puisque beaucoup de Peuls qui s'y étaient installés
à la suite des différentes sécheresses sont repartis vers
leurs régions d'origine mais surtout vers l'étranger. Il avait,
en effet, observé que, parmi les familles recensées dans la zone
entre 1983 et 1986, seulement 39 % d'entre elles étaient restées
sur place ou à proximité. Finalement cette sédentarisation
reste relative et entrecoupée de déplacements plus ou moins
longs. Ceci amène Boutrais (1984) à considérer que chez
lez peuls, on rencontre rarement une immobilisation complète, ce que
confirment Lhoste et al. (1993) qui constatent que « les
pasteurs forcés de changer de mode de vie après la disparition de
leur troupeau tentent de reprendre leur vie nomade, après avoir
reconstitué leur cheptel ».
Il existe cependant un noyau plus ou moins important de Peuls
initialement pasteurs qui ont « réussi » leur
sédentarisation en devenant agropasteurs notamment dans le sud-ouest du
Niger (Bolé et Gosso par exemples ; Riegel, 2002) et dans la partie sud
de la Tapoa (communes de Tansarga et Logobou), en particulier à
Kotchari. Raimond (1999) a noté une tendance à la
sédentarisation d'éleveurs dans certains terroirs du bassin du
lac Tchad dans un contexte de surcharge animale des parcours. Ces
éleveurs qui s'adonnent à l'agriculture, intensifient leurs
élevages grâce à l'embouche bovine et ovine. Ce type de
sédentarisation a été qualifié «
d'agropastoralisme de sécurité » ou de « voyage sans
retour » par Bonfiglioli (1990), il est l'aboutissement d'une crise
chronique et s'opère généralement par un changement
radical (changement dans les races comme l'a observé Boutrais (1984) au
Nord Cameroun et au Nord Côte d'Ivoire), qui n'est pas toujours
positivement assumé puisqu'il ne résulte pas toujours d'un projet
réfléchi mais relève plutôt d'une logique de
survie.

63
3.4.2.2. L'élevage transhumant
L'élevage transhumant dans la province de la Tapoa a
été étudié par plusieurs auteurs dont Benoit (1998
& 1999a), Santoir (1998 & 1999), Toutain & al. (2001),
Paris (2002), Kagoné (2004), Sawadogo (2004), Ouédraogo (2008),
Kpoda (2010) et Bambara (2010). Toutes ces études montrent que
l'activité y est importante et que le phénomène trouve ses
sources dans un lointain passé. On peut situer ses débuts dans la
province au milieu du 20ème siècle quand les Peuls et
leurs troupeaux franchissent pour la première fois le fleuve Tapoa.
Depuis, grâce à l'amélioration des conditions
environnementales et sanitaires locales et au potentiel en ressources
fourragères de la zone, de nombreux éleveurs en ont fait une zone
de séjour pendant la saison sèche marquée
généralement par des crises hydraulique et fourragère. La
campagne de transhumance, commandée par l'état des ressources
(Kagoné, 2000 & 2004), est variable. Paris (2002) montre que les
périodes d'arrivée ou de départ des transhumants sont
étalées, les arrivées se situant de décembre
à mai alors que les départs commencent avec les premières
pluies (juin-juillet).
Les zones d'origine des transhumants qui arrivent dans la
partie sud de la province sont diverses : région sahélienne du
Burkina Faso (Yagha, Séno, Komondjari), nord de la province (Botou,
Kantchari, Matiacoali, etc.) et sud-ouest du Niger (Torodi, Gueladio, Say,

Tamou, Téra, Tillabéry, etc.). Les transhumants
empruntent en général des itinéraires choisis sur la base
des expériences des campagnes précédentes,
itinéraires devant être indemnes de maladies et permettre de
nourrir et d'abreuver les animaux en cours de chemin (Riegel, 2002 ; Paris,
2002).
Les terroirs de Kotchari et de Logobou sont les derniers
endroits de repos avant la traversée des frontières nationales en
direction du Bénin et du Togo. Mais depuis quelques années,
beaucoup de ces transhumants y passent entièrement leur campagne (Paris,
2002).
Ces transhumants, qui sont essentiellement du groupe ethnique
peul35, sont, dans le terroir de Kotchari, majoritairement (84%)
d'origine burkinabé. Les troupeaux sont de composition spécifique
ou raciale assez peu diversifiée, (généralement mono
spécifiques à 70% - 75%. Ils se composent essentiellement de
zébus peuls à grande bosse (les Puli ou Puli
Puli selon Santoir, 1999) ou de Gurmaji pour ceux venant de la
partie nord de la province, et leurs effectifs sont élevés
(rarement moins de 100 têtes). L'importance des effectifs d'animaux
transhumants dans la province n'est pas connue avec précision, mais
certains terroirs comme celui de Kotchari voient leurs effectifs en bovins
passer du simple au double (Paris, 2002) pendant la période de pointe de
la transhumance située entre mai et juin.
3.4.3. L'élevage intensif et semi-intensif
L'élevage pratiqué en mode intensif ou
semi-intensif existe dans la province de la Tapoa, mais il reste marginal. Il
concerne essentiellement l'embouche traditionnelle ovine, caprine ou porcine
des femmes, à laquelle il faut ajouter l'embouche conduite grâce
aux actions des ONG et projets de développement et qui concerne les
bovins en plus des ovins et porcins. Un certain nombre de structures de
soutien, comme l'ONG RECOPA et les projets d'envergure comme le PADAB II, le
PICOFA, sont les plus actifs dans le domaine. Si l'engouement semble visible,
il est en revanche difficile de connaître le poids économique
réel de ce système par rapport au système pastoral ou
agropastoral. Une étude plus spécifique serait nécessaire
pour le déterminer.
3.5. Les autres activités socio-
économiques
3.5.1. Les activités agricoles
L'agriculture est la première occupation en milieu
rural dans la Tapoa (DREP-Est, 2010) où elle occuperait environ 93,7% de
la population rurale (Sanou, 2005). A l'instar de l'élevage, c'est une
activité conduite de manière itinérante et sur
brûlis. Jusqu'en 1997, période de retour du coton dans la zone,
cette agriculture était essentiellement orientée vers la
subsistance. Les principales spéculations agricoles sont le sorgho, le
mil, le maïs (qui a pris un grand essor avec la
généralisation de la cotonculture). Le riz est en essor avec
l'aménagement des bas-fonds entrepris par les projets (PADL/T, PICOFA).
Les spéculations de rente sont
35 Pendant tout le temps de nos enquêtes nous
n'avons rencontré que deux troupeaux d'éleveurs
gourmantché venant de la commune de Botou.

65
l'arachide, le coton, le niébé, la patate, le
soja, le sésame et le manioc. D'autres spéculations plutôt
marginales et maraîchères (pastèque, choux, oignon, pomme
de terre, tomate, aubergine) sont conduites par endroits (ACRA, 2009).
Le retour du coton dans la région pendant la campagne
1996/97 a occasionné de grands bouleversements dans le paysage agricole
et dans l'économie rurale. La pression sur l'espace a été
accrue avec l'arrivée de migrants agricoles et l'installation de grandes
exploitations qui ont accéléré le taux d'occupation des
sols dans toutes les communes cotonnières comme celle de Tansarga. La
corrélation entre la cotonculture et l'accélération de
l'occupation de l'espace est si forte que lorsqu'en 2007 l'engouement pour
cette culture a baissé, faute de prix rémunérateur
(Renaudin, 2007)36, on a observé une déprise agricole
après une longue période de forte progression du front agricole ;
cette déprise semble se poursuivre actuellement (figure
III-5)37. Il faut signaler que, dans cette partie du Burkina Faso,
seule la commune de Botou, où l'élevage représente la
première activité économique et qui présente des
traits physiques soudano-sahéliens, ne connaît pas la culture du
coton.
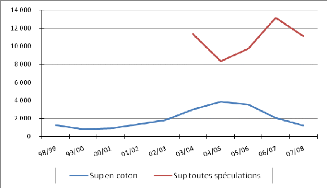
Figure III-5 : Évolution des superficies totales
emblavées (1999 à 2008) et des superficies en coton (2004
à 2008) dans la commune de Tansarga (Données:
DPAHRH/Tapoa38, 2008 ; SOCOMA39, 2010).
3.5.2. Les activités extractives : la chasse et la
pêche.
La province de la Tapoa dispose d'un réseau
hydrographique assez important. Trois plans d'eau d'importance, la Tapoa, la
Boudieri et l'Arly, peuvent être signalés. Ils offrent un
potentiel halieutique assez important, dont l'exploitation reste cependant au
stade rudimentaire (Sanou, 2005). Les acteurs du secteur sont en effet
très peu organisés et les
36 A l'échelle du pays, selon l'auteure, ce
recul a occasionné une baisse de production du coton graine de l'ordre
de 10% (de 715000 à 660000 tonnes).
37 Les données sur les superficies totales
emblavées avant 2003/2004 n'étaient pas exploitables.
38 Direction provinciale de l'agriculture, de
l'hydraulique et des ressources halieutiques de la Tapoa
39 Société cotonnière du
Gourma.

produits issus de cette activité sont destinés
essentiellement à l'autoconsommation et au marché local (Zouri,
2003).
La province dispose, par ailleurs, d'un potentiel faunique
très important dû à la concentration en aires
protégées et surtout de leur structuration en réseau. Un
recensement aérien conduit par Bouché et al. (2003) en
2002 avait donné les statistiques suivantes pour l'ensemble du complexe
WAPOK (ensemble d'aires protégées W, Arly, Pendjari, et
OtiKéran): 442 éléphants, 399 buffles et 421 hippotragues.
La présence du buffle (Syncerus caffer), du bubale
(Alcelaphus buselaphus), du cobe defassa (Kobus defassa), du
phacochère (Phacochoerus africanus), etc., témoigne
d'une faune sauvage assez riche qui attire chaque année beaucoup de
touristes en période de chasse. Par ailleurs, quatre espèces de
primates s'y rencontrent: le cynocéphale ou babouin (Papio
hamadryas), le singe rouge (Erythrocebus patas), le singe vert ou
vervet (Chlorocebus aethiops) et le galago du Sénégal
(Galago senegalensis). Notons en outre que le parc W est classé
zone d'importance pour les oiseaux et qu'on y a dénombré en 1979,
278 espèces d'oiseaux. Quelques rapaces comme l'oricou, le gyps
africain, le gyps de Rüppel, le percnoptère brun, le vautour
huppé, l'aigle martial, le messager serpentaire, le bateleur, ainsi que
de nombreux échassiers et passereaux ont été
enregistrés. Certains estiment même que plus de 70% des ressources
fauniques du Burkina Faso se trouvent dans le complexe d'aires
protégées de la région (Tankoano et al. 2010).
Ce potentiel faunique permet l'émergence d'un secteur
touristique qui commence à être dynamique. De nombreuses
réserves partielles de faune ont, en effet, été
concédées à des particuliers (exemple de la réserve
partielle de faune de la Kourtiagou)40 et chaque année la
saison de chasse draine de nombreux touristes pour la chasse sportive et la
vision. Par ailleurs, pour aller dans le sens des recommandations de la
Convention sur la biodiversité (CBD) et des différentes
conventions internationales et programmes (notamment le programme MAB (Man And
Biosphere) de l'UNESCO de 1971) qui suggèrent que, dans un souci
d'équité et de développement durable, les
communautés riveraines soient associées dans la gestion des
ressources naturelles (concept de gestion participative ou inclusive),
l'État et ses partenaires promeuvent localement la mise en place de
zones villageoises d'intérêt cynégétique (ZOVIC).
Une ZOVIC est définie comme "une partie du terroir d'une
communauté de base, affectée par elle à l'exploitation des
ressources cynégétiques" (article 99 ; code forestier) ou comme
"une aire de protection faunique créée sur le terroir d'une
communauté de base" (article 4 ; décret 2008-312). Selon
Kaboré (2010) « l'idée de ZOVIC a été
introduite dans la législation forestière nationale comme mesure
d'accompagnement des nouvelles décisions de concéder les
réserves de chasse aux opérateurs privés ». Les
ZOVIC sont régies à la fois par le code forestier (loi 006/97/ADP
du 31 janvier 1997), le décret 2008- 312/PRES/PM/MECV/MATD/MEF portant
conditions de création et de gestion et par la loi 055-2004/AN portant
code général des collectivités territoriales (CGCT).
Ces zones de chasse, lorsqu'elles sont adossées aux aires
fauniques, peuvent jouer le rôle de zones tampons
40 Cela à la faveur des nouvelles
orientations (à partir de 1987 puis de 1996) en matière de
politique de gestion des aires fauniques (Baillon & Sournia, 1987 ;
Kaboré, 2010) qui veut que, pour mieux assurer leur surveillance,
celles-ci soient confiées à des opérateurs privés
(guides de chasse puis concessionnaires).

67
pour celles-ci41, et permettre ainsi de les
sécuriser en réduisant la pression anthropique sur leur noyau
central. Elles permettent la valorisation du petit gibier local qui procure des
revenus, peu importants pour l'instant42, aux populations. Dans le
terroir de Kotchari, quatre ZOVIC sont enregistrés (Gnimboama :
162 ha ; Lada : 279 ha ; Nangbanli : 221 ha et
Pielgou) mais elles sont à des stades différents de
développement et certaines comme celle de Pielgou sont d'ailleurs
contestées alors que celle de Lada attend d'être consensuellement
délimitée (Sawadogo, 2011)43. La contestation des
ZOVIC, qui s'inscrit dans la continuité de celle des aires fauniques
elles-mêmes44, est généralement du fait
d'agriculteurs faisant face à un besoin crucial en terres ou devant
déguerpir ou alors d'éleveurs qui craignent que les ZOVIC ne
constituent une astuce supplémentaire des agriculteurs pour les exclure
de certaines portions du terroir45. La majorité des
agriculteurs locaux (plus de 89%) (Zombra, 2008 ; Kaboré, 2009)
montrent, quant à eux, un grand intérêt à la mise en
place de ces entités de gestion et de valorisation de la petite faune
villageoise.
Il faut signaler par ailleurs, qu'une grande portion du
terroir (423 ha) située dans le secteur sud du côté du
village de Gnimboama (Sawadogo, 2004 ; Sawadogo, 2011) constituant initialement
la ZOVIC dudit village est passée depuis une dizaine d'années
sous la gestion de l'ONG Nature et Vie46. C'est une zone assez
particulière en ce qu'elle regorge une importante diversité
faunique, notamment aviaire et végétale.
41 L'état encourage en effet qu'autour des
aires fauniques, soient mises en place des zones tampons, définies comme
des bandes ceinturant les aires et dans lesquelles les aménagements
socioculturels et économiques doivent être compatibles avec les
objectifs de l'aire protégée (article 79, code forestier). Les
ZOVIC, en tant que zones tampons, sont un compromis efficace en ce qu'elles
permettent de répondre à ces objectifs tout en mobilisant les
communautés qui tirent profit de leur gestion (il y a alors une sorte de
compensation face aux interdictions d'accès au noyau central des
réserves); ce qui rejoint l'esprit du programme MAB (Man And Biosphere)
de l'UNESCO de 1971.
42 Kaboré (2010) note par exemple qu'au titre de la
campagne de chasse 2006-2007 seulement une somme de 50000 FCFA à 200000
FCFA ont été redistribués à chacun des villages
bénéficiaires dans la zone de la réserve de Pama Nord
(province du Gourma)
43 Selon les dispositions réglementaires en
la matière (code forestier, décret 2008-312, CGCT), la ZOVIC est
mise en place et gérée par le CGF « Commission de gestion de
la faune » (article 6, décret 2008-312) sous le contrôle du
conseil villageois de développement (CVD) et du Conseil communal
(article 11, décret 2008-312) et qui doit, avec l'appui technique des
services étatiques de gestion faunique (article 3, décret
2008-312), assurer la gestion de la zone à travers des actions
d'aménagement comme la surveillance, l'ouverture des pistes, les
reboisements.
44 Kaboré (2010) montre que le rejet des
aires fauniques n'est pas seulement mû par la seule considération
de l'accès aux ressources qu'elles regorgent. Des considérations
comme le désir d'exercer un droit de contrôle ou de la poursuite
de la pratique des rites sacrificiels sont causes aussi des revendications.
45 La loi dispose que c'est le CGF (représentant les
communautés de base) qui définit les activités
autorisées (article 101, code forestier du 31 janvier 1997) avec l'appui
des services techniques. On peut craindre alors qu'il ne naisse des
velléités tendant à soustraire ces zones à
l'exploitation animale.
46 La cession de cette zone à l'ONG ne s'est
pas faite par consentement. Les villageois, se sont vus obligés de le
faire car la promotrice est l'épouse du premier Ministre de
l'époque qui est par ailleurs, natif de Tansarga, chef-lieu de la
commune.
CHAPITRE IV

68
LES UNITES PAYSAGERES PASTORALES DANS LE
TERROIR ET DANS L'AIRE PROTEGEE VOISINE :
DEFINITION, DISTRIBUTION SPATIALE ET
CARACTERISTIQUES

69
4.1. Introduction
Comme on l'a indiqué en introduction
générale le terroir de Kotchari constitue un des nouveaux
territoires qu'investissent les pasteurs dans leur descente vers le sud
à la recherche de zones plus clémentes et mieux fournies en
ressources fourragères (Boutrais, 1996 & 2007 ; Convers, 2002).
Localement, avec les grands bouleversements socioéconomiques (notamment
individualisation des exploitations, diversification des activités,
montée de la culture du coton, etc.), il s'y déroule par ailleurs
une course à l'occupation des terres qui se manifeste par des
défrichements conservatoires47 (Sawadogo, 2004).
L'état des ressources pastorales qui attirent les éleveurs dans
la région apparaît comme une question centrale.
L'objectif du présent chapitre est de fournir une
typologie des différentes unités agrostologiques du terroir de
Kotchari et de la partie contigüe du parc W, elle aussi utilisée
par les pasteurs. Il s'agit également d'évaluer leur état
écologique et leurs potentialités pastorales en ayant recours aux
méthodes classiques de l'écologie et de l'agrostologie, certaines
faisant intervenir des savoirs locaux. Cette étude a fait l'objet d'un
article (Sawadogo et al. sous presse).
Rappel de l'hypothèse de recherche
L'afflux observé de nombreux troupeaux transhumants
chaque année sur le terroir de Kotchari et ses environs ne s'explique
pas, comme il est souvent dit, par la disponibilité en ressources
pastorales dans les espaces légalement accessibles
(périphérie du Parc). L'intérêt pastoral du terroir
de Kotchari est plutôt lié à l'opportunité qu'il
peut offrir d'accéder illégalement aux ressources du Parc du
W.
4.2. Matériel et mé thodes
Pour définir puis caractériser les unités
paysagères pastorales (UPP sensu DeWispelaere & Waksman,
1977) et produire ainsi une carte des pâturages du terroir, trois
approches complémentaires ont été mises en oeuvre :
- l'analyse d'une image satellitaire ;
- une analyse phytoécologique par relevés de
végétation au sol ;
- une évaluation de la valeur pastorale fondée sur
un indice de qualité apprécié par les éleveurs.
4.2.1. Approche des unités paysagères
pastorales par la télédétection
Un document cartographique offre l'avantage de
présenter la répartition géographique des pâturages
et leur importance relative sous une forme synthétique. Le travail de
cartographie a consisté à réaliser une carte des
ressources pastorales en se basant sur des images satellitales. Ce travail a
permis de faire une esquisse des pâturages du terroir que nous avons
ensuite validé par l'analyse phytoécologique. De manière
concrète, il s'agissait de
47 Pratique qui consiste à défricher un
terrain sans avoir l'intention d'y implanter un champ, l'objectif étant
d'empêcher son exploitation par autrui.
regrouper les pixels (la plus petite surface homogène
d'une image numérique) de toute l'image selon leurs valeurs
radiométriques (Devineau et al. 1997).
Une classification de l'espace du terroir en différents
types de pâturages a été faite grâce à une
image Aster acquise le 10 octobre 2006. Les images Aster ont une
résolution spatiale de 15 m dans le domaine du visible et proche infra
rouge, qui est bien adaptée à une étude
phytoécologique de la végétation comme à
l'étude de l'occupation des sols dans un territoire à petit
parcellaire.
Comme notre connaissance du terrain nous permettait de bien
différencier les différents thèmes d'occupation du
paysage, nous avons opté pour une interprétation visuelle d'une
composition colorée de l'image Aster. L'exploitation visuelle d'une
composition colorée permet une interprétation globale des
paysages et l'accès rapide à des informations d'ordre spectral,
structural et textural, dont l'intégration avec des méthodes
numériques serait complexe (De Wispelaere & Waksman 1977 ; Lambin,
1988 ; Lainé & Paré 1996 ; Dedieu et al. 1997). Elle
permet en particulier de repérer par leurs formes les zones de champs et
de jachères, dont les signatures spectrales se différencient
souvent mal de celles des milieux savanicoles environnants (Lainé &
Paré 1996 ; Devineau et al. 1997 ; Ba et al. 2000).
La composition colorée a été
établie en mode fausses couleurs avec les bandes spectrales du visible
et proche infrarouge (VNIR), les bandes 1 (0,52-0,60um) (coloré en
bleu), 2 (0,63-0,69 um) (coloré en vert) et 3 (0,76-0,86 um)
(coloré en rouge). L'interprétation s'est fondée sur les
couleurs rendues par chaque surface ou thème au sol (tableau IV-1). Les
formations végétales apparaissent dans divers tons de rouge ; le
rouge vif correspond à la végétation naturelle notamment
aux savanes arbustives à arborées ou les forêts claires
(activité photosynthétique élevée entraînant
une forte réflectance dans le proche infrarouge); et le rouge clair
correspond aux savanes herbeuses et aux cultures (activité
photosynthétique faible à moyenne compte tenu de la
période). Les sols nus se traduisent par des teintes blanches à
grises, les cuirasses et les buttes rocheuses par du bleu et l'eau libre
apparaît en noir.
En octobre, mois d'acquisition de l'image, les cultures sont
toutes au stade de l'épiaison avec un feuillage dont le vert commence
à diminuer d'intensité et la strate herbacée des zones non
cultivées, dominée par les annuelles, a également
déjà bien entamé son assèchement. Ainsi, les zones
de culture et de savane herbeuse dont l'activité photosynthétique
est en forte baisse, apparaissent moins rouges dans la composition
colorée que des formations plus fermées où la strate
ligneuse est encore très verte bien que la chute des feuilles soit
déjà amorcée (Mahamane et al. 2007).
Tableau IV-1. Expression colorée des différents
thèmes du paysage dans l'image ASTER en composition colorée,
infrarouge fausse couleur utilisée pour définir les unités
paysagères pastorales du terroir de Kotchari.
Bande
spectrale
(capteur)
Couleur
sur
l'écran
Longueur
d'onde (um)
Couleur rendue par thème
Sol nu Eau Cuirasse Cultures Végétation
et butte naturelle

Vert Bleu 0,520 - 0,600
Rouge Vert 0,630 -0,690 Blanc à
Proche Rouge 0,760 -0,860 gris
Infrarouge
Noir Bleu Rouge-
clair
Rouge-vif

71
Les unités paysagères pastorales qui ont
été définies par l'analyse d'image ont été
ensuite rapprochées à une base de données des sols
produite par le Bureau National des Sols (BUNASOLS, 2007), ce qui a permis leur
caractérisation pédologique.
4.2.2. Analyse phytoécologique par relevés au
sol
Le but de l'analyse phytoécologique était
d'établir une typologie des pâturages du terroir de Kotchari en
validant les unités paysagères issues de l'analyse d'image mais
aussi en caractérisant leur végétation et leur valeur
pastorale. Cette analyse s'est fondée sur des relevés
phytoécologiques, chacun étant un ensemble d'observations
phytoécologiques qui concernent un milieu déterminé
(Boudet, 1978 ; CEPE, 1983), il produit le portrait d'une portion de
végétation choisie dans ce milieu (Grouzis, 1982).
Ces relevés ont été effectués sur
le terroir de Kotchari et dans la partie adjacente du Parc du W dont l'usage
par les pasteurs est avéré. L'échantillonnage a
visé à rendre compte de la diversité
végétale présente dans chacun de ces deux espaces. Un
total de 45 relevés (21 dans l'aire protégée et 24 en
dehors) ont été faits sur des placeaux de
végétation les plus homogènes possible.
Les relevés ont consisté en un inventaire
séparé des strates végétales herbacée et
ligneuse - comme le préconisent Trochain (1940), Gounot (1969) et
Zoungrana (1991) - ainsi qu'en des observations sur la nature des sols et la
topographie du milieu. La superficie des placeaux, 2500 m2 (50 m x
50 m), était conforme aux surfaces généralement
utilisées par les auteurs qui travaillent en zone soudanienne : 1200
m2 pour Hahn-Hadjali (1998), 1600 m2 pour Devineau (2005)
et 3000 m2 pour Hiernaux (1975).
Par ailleurs, pendant l'exécution des relevés,
des herbiers, représentant les espèces végétales
dont les noms scientifiques ne sont pas connus, ont été
récoltés et ont servi à des identifications
ultérieures à l'aide de flores. La conformité,
l'orthographe ainsi que l'actualité des noms ont été
vérifiés sur le site web du Conservatoire des Jardins Botaniques
de Genève.
Toutes les activités d'inventaire ont eu lieu entre 2007
et 2009, en fin septembre.
4.2.2.1. Relevés de la strate
herbacée
La composition et la structure spécifique de la strate
herbacée ont été étudiées par analyse
linéaire selon la méthode quantitative des points quadrats (Daget
& Godron, 1982 ; Daget & Poissonet, 1971) au moment du maximum de
phytomasse, c'est-à-dire quand la végétation
herbacée est au stade fin floraison-début fructification dans
cette région.
La méthode des points quadrats (ou des points
contacts), mise au point en NouvelleZélande, développée en
Australie et en France (CEPE de Montpellier) a été
appliquée aux

pâturages africains avec succès par bon nombre
d'auteurs notamment Daget & Poissonet (1971), Poissonet & César
(1972), Daget & Godron (1982), Fournier (1982, 1983), Akpo et al.
(1995), Kagoné (2000), Botoni (2003), Kièma S. (2007).
Cette méthode est beaucoup plus longue à mettre
en oeuvre que la méthode de notation d'abondance-dominance de
Braun-Blanquet. Cependant, elle est particulièrement recommandée
lorsqu'une précision des mesures est recherchée, notamment
lorsqu'on cherche à caractériser et à quantifier, comme
c'est le cas ici, l'état des milieux soumis aux actions anthropiques
(Kièma S., 2007). En effet, (i) elle est performante dans l'inventaire
des espèces situées sur les lignes de relevés, (ii) elle
permet de déterminer les « espèces productrices »48 de
biomasse et leurs fréquences, (iii) enfin, lorsque le nombre de
relevés est important, le nombre de contacts par point donne des
biovolumes pouvant s'assimiler à la phytomasse (Poissonet &
Poissonet, 1969).
Dans chaque placeau, un ruban de 20 m de long, tenu par deux
piquets fixés au sol, est placé horizontalement à la
surface du tapis herbacé ou en son sein. Les lectures sont faites d'un
bout à l'autre du ruban tous les 20 cm et il est noté la
présence-absence des espèces interceptées par la tige
métallique, chacune étant comptée une seule fois par point
de lecture (Boudet, 1978 & 1991). L'opération a été
répétée quatre fois par placeau sur des lignes de
relevés disposées parallèlement tous les 10 m, ce qui a
donné 400 points de lecture. On considère en
général que pour avoir une bonne précision
d'échantillonnage (de 5 % environ)49, le nombre de points de
lecture nécessaires doit être d'au moins de 200 (Rippstein, 1985 ;
Grouzis, 1988 ; Zoungrana, 1991 ; Sinsin, 1993 ; Sawadogo, 1996). Plus
généralement, selon Daget & Poissonet (1971), à mesure
que le nombre d'observations s'élève, l'intervalle de confiance
de la fréquence centésimale se réduit rendant la mesure
plus sûre.
La liste floristique a ensuite été
complétée en inventoriant les espèces situées en
dehors des lignes de relevé.
4.2.2.2. Évaluation de la phytomasse
herbacée
La valeur d'un pâturage prend en compte la
quantité d'herbe offerte (production primaire) et sa qualité
comprenant sa valeur bromatologique (valeur fourragère) ou son
degré de palatabilité défini par observation du
comportement alimentaire des animaux ou par déclaration des
éleveurs (indice de qualité) et sa teneur en
macroéléments et oligoéléments (Boudet, 1978 ;
Zoungrana, 1991).
On appelle phytomasse ou biomasse végétale, le
poids par unité de surface, exprimé en matière
sèche, du total de la matière vivante et morte des
végétaux (Duvigneaud, 1974 ; Daget
48 Kièma (2007) explique clairement cette
notion déjà évoquée par Sawadogo (1986) et dont les
auteurs sont Daget & Poissonet (1971). Il simplifie l'expression en
considérant comme « espèce productrice » dans un
groupement végétal, toute espèce dont la contribution
spécifique dépasse 5%.
49 La précision d'échantillonnage est
variable d'un pâturage à un autre en fonction du degré
d'hétérogénéité de la strate
herbacée. C'est pourquoi, pour se rapprocher le plus possible du nombre
de points nécessaires sur le pâturage étudié, on
recommande de recourir à la détermination de l'intervalle de
confiance (IC) au risque d'erreur á à partir de la
fréquence de l'espèce dominante (Grouzis, 1982 ; Boudet, 1984).
IC = p #177; t v?p*q/N avec p = fréquence de l'espèce
dominante ; q = (1-p) ; N = nombre total de contacts ; t= t de Student pour le
risque á.
& Godron, 1995). Il faut y distinguer la biomasse
proprement dite équivalente aux parties vivantes, et la
nécromasse constituée des parties mortes (Duvigneaud, 1974). En
général, pour plusieurs raisons (faible participation du fourrage
ligneux à la ration des animaux pendant une bonne partie de
l'année, estimation difficile de sa production primaire, etc.), la
biomasse évaluée porte uniquement sur la strate herbacée.
La production tant quantitative que qualitative de cette biomasse
herbacée est influencée en zone soudanienne par nombre de
facteurs comme la pluviosité (Fournier, 1991 ; Kabore-Zoungrana, 1995),
le sol (Breman & De Ridder, 1991), le rayonnement solaire (Cocheme &
Franquin, 1968) et les propriétés des végétaux
eux-mêmes (Breman, 1975 ; Boudet, 1975 & 1978). La nature
hétérogène des pâturages en termes de composition
floristique et de contribution spécifique et la variabilité
climatique fait que la biomasse produite varie au cours de l'année.
Ainsi, en saison sèche, la quantité de biomasse herbacée
diminue considérablement et sa qualité se déprécie
avec la perte de certains constituants cellulaires (matière
azotée notamment) et le développement des tissus de soutien tels
que la lignine (Kabore-Zoungrana, 1995 ; Kagoné, 2000).
La biomasse végétale peut être
estimée de plusieurs manières avec des résultats forts
différents (fauchages successifs à des intervalles
réguliers durant la période active de végétation ou
fauchage en un seul temps à la fin de la période active
correspondant à la période de fructification). Toutefois, la
biomasse mesurée à la fin de la période active,
correspondant au maximum de production, est une bonne expression de la
productivité des pâturages de la savane car elle donne des
indications sur son potentiel de productivité (Boudet, 1978). Nous avons
retenu cette approche et seule la partie épigée de l'herbe a
été récoltée intégralement au stade de
maturation des semences à l'intérieur de 30 placettes
carrées de 1 m2 (1m x 1m) (Grouzis 1988 ; Boudet, 1978 &
1991 ; Fournier, 1991), ce qui permet d'atteindre un taux de précision
d'environ 20 % (Levang, 1978). Les placettes ont été
disposées tous les 5 m sur 5 lignes parallèles espacées de
10 m. Les prélèvements par fauchage à 2 cm du sol ont
été effectués début octobre, période
où la phytomasse est maximale.
La précision (P) des mesures a été
déterminée selon la formule de Daget et Poissonet (1971) :
tó
P ( % ) = ×100
xN
Avec
t, coefficient de Student (P = 0,5)
ó, écart-type
x, poids moyen par placette d'1 mètre
carré N, nombre de placettes par placeau

73
Notons que, même si elle est destructrice, longue et
souvent fastidieuse, la méthode de récolte intégrale a
l'avantage d'être simple, particulièrement fiable (Fournier, 1991
& 1994) et économique. Par ailleurs, un éventuel effet de
bordure lié à la forme des placettes peut être
minimisé en utilisant des placettes circulaires.

4.2.2.3. Relevés de la strate ligneuse
L'intérêt porté à l'analyse de la
strate ligneuse (arbres, arbustes et arbrisseaux) se justifie par plusieurs
raisons. Tout d'abord, on sait que les espèces ligneuses et
herbacées réagissent différemment, en savane soudanienne,
vis-à-vis des facteurs anthropiques (Devineau et al. 2009), les
premières étant les témoins des évolutions sur le
long terme (César, 1992). Par ailleurs, Akpo (1992) montre qu'au niveau
du système d'exploitation, l'arbre améliore les conditions
d'élevage en agissant positivement sur la strate herbacée
(production, matière azotée digestible, énergie) et en
produisant des feuilles vertes pendant au moins 9 mois de l'année.
L'apport de la strate ligneuse dans l'alimentation des animaux apparait donc
inestimable et prépondérante pendant la saison sèche
chaude au cours de laquelle le fourrage herbacé est rare et de
très mauvaise qualité (Lapeyronie & Le Floc'h, 1995). Le
Houérou (1980) estime, en effet, que la consommation de fourrage ligneux
en saison sèche est indispensable pour compenser les déficits en
phosphore et carotène de la paille. Ce que confirme Rivière
(1991) qui montre qu'un troupeau exclusivement nourri en paille pendant la
saison sèche, présentait des carences assez graves en vitamine A
pouvant aller jusqu'à la cécité. Des études ont pu
montrer que certains ligneux, disponibles en Afrique, tels que Acacia
albida, Acacia seyal, Balanites aegyptiaca, Leucaena leucocephala, Pterocarpus
erinaceus, Afzelia africana, Ficus spp., etc. présentaient des
valeurs nutritives intéressantes pour les animaux et étaient
aptes à améliorer qualitativement des rations à base de
fourrages pauvres de saison sèche (Le Houérou 1980; Koné
1987; Fall 1993, KaboréZoungrana, 1995) malgré les
problèmes d'accessibilité physique et la présence de
certains facteurs limitant de nature anti métabolique et
anti-nutritionnelle comme les tanins et les autres composés secondaires
que sont les phénols, l'anthocyane et la mimosine (Skearman, 1982; Fall,
1993 ; Baumer, 1997).
La part des ligneux dans les rations est variable en fonction
de leur accessibilité physique, de l'espèce animale, de la
période de l'année et même du contexte
écoclimatique. En effet, Breman & De Ridder (1991) notent que
seulement 25 % de la production annuelle des ligneux est disponible pour les
animaux qui peuvent avoir de 85, 50 et 25 % MS (respectivement pour les
caprins, les ovins et les bovins) de leur régime constitués de ce
type de fourrage. Les travaux de Bourbouze (1982) effectués dans les
montagnes marocaines montrent qu'en janvier, les rations des caprins
étaient presque exclusivement constituées de ligneux (91 %) alors
qu'en juillet la part de ce type de fourrage n'atteignait plus que 3%. Daget
& Godron (1995) ont par ailleurs noté que chez les caprins la ration
pouvait comprendre entre 50% et 90% de fourrage ligneux alors que chez les
ovins et bovins elle tournait respectivement entre 20 et 40% et entre 5 et 10%.
Selon Von Maydell (1983), au moins 75 % des ligneux sont broutés au
Sahel et en zone nord soudanienne de manière temporaire ou
permanente.
Par ailleurs, les ligneux ont des cycles phénologiques
assez diversifiés (MuellerDombois & Ellenberg, 1974 ; Grouzis &
Sicot, 1980 ; Guérin & al. 1991 ; César, 1992 ;
Devineau, 1997 & 1999), ce qui rend disponible le matériel
végétal tout au long de l'année pour être mis
à contribution dans la ration des animaux.

75
La structure de la strate ligneuse - arbres, arbustes et
sous-arbustes ou buissons (Descoings, 1975) - a été
analysée par recensement exhaustif sur les placeaux pour permettre un
calcul de densité (nombre de pieds par hectare). Le recouvrement ligneux
(R), qui exprime le quotient de l'aire de projection des couronnes ligneuses
sur le plan horizontal par l'aire totale du placeau (Descoings, 1975), a
été évalué par la méthode de l'interception
linéaire (Canfield, 1941; Mueller-Dombois & Ellenberg, 1974 ; Kaiser
1983) avec, par placeau, 3 lignes parallèles matérialisées
au sol, de 50 m de long et espacées de 20 m. La méthode a
consisté à projeter, à l'aide d'une perche verticale
longue de 5 m environ, la couronne des ligneux sur la ligne au sol. Dans les
cas où les ligneux formaient une touffe ou lorsque les couronnes se
chevauchaient ou se superposaient, le diamètre du houppier au sol a
été mesuré pour l'ensemble de la touffe (ou des couronnes
superposées) et non séparément, contrairement à
Kièma S. (2001). Cet auteur dans ses travaux conduits dans les
forêts classées du Maro et du Tuy, a en effet
considéré individuellement chaque ligneux formant les touffes et
a par moment obtenu des taux de recouvrement supérieurs à 100%
à cause du chevauchement ou de la superposition des couronnes entre
individus.
4.2.3. Enquêtes sur la qualité des
espèces fourragères : la valeur pastorale
La valeur pastorale constitue avec la capacité de
charge, deux indicateurs qui permettent d'appréhender globalement la
valeur des pâturages (Zoungrana, 1991). La valeur pastorale est une
méthode d'évaluation de la qualité fourragère des
parcours (Daget & Poissonet, 1971 ; Daget & Godron, 1995 ;
César, 2005), elle dépend à la fois de la contribution
spécifique des espèces (CSi) et de leur indice de qualité
spécifique (ISi). Cette dernière correspond soit aux valeurs
bromatologiques des espèces végétales des milieux
analysés ou alors aux notes attribuées par des éleveurs
(bergers, propriétaires de troupeaux, personnes ressources)
enquêtés (encadré IV1).
Encadré IV-1. L'indice de qualité
spécifique, une valeur relative
L'indice de qualité spécifique, est subjectif et
inconstant car il dépend à la fois de
l'appétibilité, de la valeur alimentaire, de
l'accessibilité de l'espèce végétale et de
l'espèce animale. Il est obtenu selon les caractéristiques
bromatologiques des espèces ou selon la classification des bergers
(Zoungrana, 1991 ; Kièma S., 2007) par détermination de classes
d'appétence. Le rapprochement qui est fait entre l'indice de
qualité et la composition chimique est cependant, souvent
inopérant. En effet, l'appétibilité bien qu'étant
un paramètre important, est une notion relative car les animaux
choisissent différemment selon le disponible fourrage et/ou les
espèces qui sont à leur disposition dans le pâturage
(Boudet, 1978 ; Zoungrana, 1991).
Par ailleurs, les animaux peuvent être amenés
à consommer des espèces de moindre appétence ou
ordinairement délaissées si le pâturage manque des
espèces les plus appréciées ou est de qualité
pauvre. Ceci est confirmé par Le Houerou (1980) à partir
d'observations faites sur les ligneux. Il note en effet, que des espèces
comme Boscia senegalensis et Calotropis procera, qui sont de
bonne valeur chimique, sont peu ou nullement consommées,
alors que des espèces comme Combretum acculeatum,
Grewia tenax, Stereospermum kunthianum ou Oxytenanthera
abyssinica sont recherchées malgré une valeur nutritive
moins bonne ou médiocre.
Nous avons en conséquence adopté la
classification des bergers qui, en plus, a été utilisée
avec succès par beaucoup d'auteurs comme César & Coulibaly
(1993), Daget & Godron (1995), Akpo & Grouzis (2000) et Kièma S.
(2007). L'indice obtenu de cette manière est fiable mais seulement dans
le cadre d'une étude visant à comparer divers pâturages
d'un même territoire comme dans notre cas.

Le nombre de classes utilisées dans le cas de la
classification des bergers dépend de la capacité des
interlocuteurs à bien classer les espèces. Ainsi, Akpo (2000),
Dahlberg (2000), Akpo et al. (2002), Kièma S. (2007),
Rakotoarimanana & Grouzis (2006), travaillant dans des contextes
différents, ont considéré quatre classes alors que Le
Houérou (1980) et César & Coulibaly (1993), dans le cadre de
leurs travaux en Côte d'Ivoire, en distinguent six (0 à 5). Pour
leur part, Grouzis (1982) et Daget & Godron (1995) définissent cinq
classes allant de 0 (non appété) à 4 (très bien
appété). C'est également ce même nombre de classes
que nous avons défini à l'issue de nos entretiens exploratoires
(tableau IV-2).
Tableau IV-2. Classes de qualité fourragère des
espèces établies avec les éleveurs de Kotchari
Classe d'indice de Appréciation de la Niveau
d'appétence par les bovins
qualité spécifique qualité
fourragère
0 Nulle Habituellement non appété
1 faible Habituellement peu appété
2 Moyenne Habituellement moyennement appété
3 Bonne Habituellement bien appété
4 Très bonne Habituellement très bien
appété
La classification des espèces fourragères des
cortèges floristiques des différentes unités
paysagères dans ces différentes classes de qualité a
permis de déterminer les valeurs pastorales desdites unités.
4.2.4. Analyse des données
4.2.4.1. Cartographie des unités
paysagères pastorales
L'analyse d'image satellitaire a été faite par le
logiciel ENVI 4.0 tandis que les sorties cartographiques ont été
effectuées dans le logiciel cartographique MapInfo version 8.1.

77
4.2.4.2. Constitution et caractérisation des
groupements agrostologiques, lien avec les unités pastorales
paysagères
L'intérêt du travail d'analyse de
végétation est d'aboutir à une caractérisation
agrostologique du terroir de Kotchari. C'est pourquoi, contrairement aux
pratiques courantes qui consistent à partir de la strate ligneuse, nous
avons concentré notre analyse sur la strate herbacée ; celle-ci
constitue, en effet, l'aliment principal du bétail et apparaît,
par ailleurs, comme le meilleur révélateur des perturbations
anthropiques qui se produisent sur le court terme (César, 1992).
La typologie des groupements agrostologiques que nous
proposons se fonde donc uniquement sur la strate herbacée. La
présence des ligneux, qui constituent une ressource
complémentaire pour le bétail (Lhoste, 2004), est prise en compte
ensuite pour expliquer certains traits de la strate herbacée dans les
unités paysagères.
Les groupements agrostologiques ont été
constitués par classification ascendante hiérarchique
agrégative des relevés floristiques à l'aide du logiciel
XlStat version 2010 4.03. C'est l'indice de similitude de Jaccard (J)
sur des données de présence-absence des espèces dans les
relevés qui a été utilisé pour construire le
dendrogramme.
Les herbacées caractéristiques des groupements
ont été recherchées par la méthode
d'évaluation de la valeur indicatrice des espèces (IndVal) de
Dufrêne & Legendre (1997). Cette méthode se fonde sur la
fidélité des espèces (présence dans un seul des
groupements agrostologiques) et leur constance (présence dans la plupart
des relevés de ce groupement). Pour chaque espèce (i) et
chaque groupement agrostologique (k) l'indice se calcule comme suit
:
|
IndVal
|
= 100 × , × ,
A k i B k i
Groupe k Espèce i
,
|
(Dufrêne & Legendre, 1997); avec
|
Aki, fidélité de l'espèce =
NEki/NEi (moyenne de la contribution de l'espèce
i dans le groupe k / somme des moyennes des contributions de
l'espèce dans tous les groupes);
Bk,i, constance de l'espèce = NRki
/NRk (nombre de sites du groupe k dans lesquels l'espèce
i est présente / total des sites du groupe k);
IndValEspèce i = max (IndValki)
Seules sont prises en compte les espèces
herbacées dont la contribution spécifique moyenne est
supérieure ou égale à 4% dans au moins l'un des
groupements agrostologiques. L'espèce végétale est
estimée caractéristique d'un groupement agrostologique quand sa
valeur indicatrice y est maximale (Indvalmax), mais ceci seulement
si son lien avec ce groupement est statistiquement significatif (p = 0,05).
À partir de là il a été possible
d'évaluer l'homogénéité floristique des
unités pastorales paysagères par l'examen des groupements
agrostologiques qu'elles contiennent.
L'évaluation a par la suite été
complétée notamment au niveau des unités paysagères
pastorales, par la recherche des espèces ligneuses dominantes sur les
principales unités paysagères pastorales. Pour ce faire seules
les quatre espèces les plus abondantes de chaque relevé ont
été retenues. Les abondances moyennes de ces espèces ont
ensuite été comparées entre unités
paysagères. Sont alors considérées comme dominantes
caractéristiques, les espèces dont les moyennes sont
significativement les plus élevées.

Chaque fois que, comme ici, les effectifs des groupes à
comparer étaient faibles, les comparaisons de moyennes ont
été effectuées grâce à une analyse de
variance (ANOVA) non paramétrique. Dans ce sens, nous avons
utilisé le test de Kruskal-Wallis (Scherrer, 2007) suivi d'un post-hoc
de comparaison multiple ou par paire de Dunn-Bonferroni dans le logiciel
XlStat.
4.2.4.3. Détermination de la diversité
spécifique des espèces herbacées
La diversité spécifique est une notion qui
permet d'avoir rapidement, en un seul chiffre, une évaluation de la
biodiversité du peuplement étudié (Nshimba, 2008). C'est
un bon indicateur de son niveau d'équilibre écologique qui
synthétise à la fois la richesse spécifique et
l'abondance. Il existe plusieurs expressions de la diversité
(Mueller-Dombois & Ellenberg, 1974 ; Magurran, 2004) : diversité
locale (richesse spécifique locale S, diversité
á), diversité régionale ou diversité gamma
(ã) et diversité inter-formations ou diversité bêta
(â).
Nous avons tenu compte de ces trois niveaux de diversité
dans l'analyse de la diversité spécifique de la
végétation herbacée des différentes unités
paysagères pastorales.
4.2.4.3.1. La diversité intra-placeau ou
diversité á.
La diversité á (ou diversité intra
placeau ou encore diversité locale) se réfère à la
diversité spécifique au niveau de l'échantillon de la
formation végétale étudiée. Cette diversité
locale peut être estimée par l'indice de diversité (H) de
Shannon. H, déterminée selon la formule ci-dessous, est fonction
de la probabilité Pi de présence de chaque espèce dans un
ensemble d'individus. Elle est indépendante de la taille de
l'échantillon et repose sur la théorie de l'information (Dajoz,
1982 ; Burel & Baudry, 2003). En effet, H suppose que la diversité
dans un écosystème peut être mesurée comme
l'information contenue dans un message (Akpo & Grouzis, 2004).
H = - ( Pi log 2 Pi)
(Mueller-Dombois & Ellenberg, 1974; Dajoz, 1982 ; Magurran, 2004), avec
Pi, Contribution spécifique de chaque
espèce au recouvrement ; log2, logarithme à base 2.
On démontre que pour un nombre constant
d'espèces, plus il y a d'espèces d'abondance voisine (groupement
peu organisé ou peu hiérarchisé), plus
élevée est la valeur de l'indice de diversité de Shannon.
H atteint son maximum (c.-à-d. Hmax = log2 N ; N = nombre total
d'individus) lorsque toutes les espèces présentes sont
équiprobables (Dajoz, 1982 ; Burel & Baudry, 2003). L'usage de
l'indice de Shannon est cependant déconseillé en particulier
lorsqu'on cherche à comparer deux formations, des valeurs identiques de
celle-ci pouvant traduire des situations bien différentes. En effet,
bien que H soit indifférent à la taille de l'échantillon,
il reste très sensible à la richesse spécifique.
En lieu et place, nous avons fait appel à l'indice
d'équitabilité ou d'équirépartition (ou encore de
régularité) de Pielou noté E (Daget, 1976 ;
Mueller-Dombois & Ellenberg,

79
Chapitre IV. Les unités agrostologiques
1974; Dajoz, 1982 ; Magurran, 2004) compris comme le rapport
entre la diversité observée et la diversité maximale. Dans
une perspective d'exploitation pastorale, l'indication sur la dominance des
espèces est une donnée importante pour évaluer l'apport de
chacune d'elles au disponible fourrager. E est maximale quand les
espèces ont des abondances identiques dans le peuplement, minimale quand
une seule espèce domine tout le peuplement (Mueller-Dombois &
Ellenberg, 1974; Dajoz, 1982 ; Magurran, 2004). Insensible à la richesse
spécifique, E est très utile pour comparer les
dominances d'espèces entre stations ou entre dates
d'échantillonnage (Devineau et al 1984 ; Akpo et al.
1999 ; Grall & Hily, 2003).
L'équitabilité (E) a été
calculée à l'aide du logiciel PAST (PAleontological
STatistics) version 2.03 (Hammer & al. 2001) selon la
formule de Pielou (1966).
H
E = (0 = E = 1), avec
H max
H, entropie ou indice de diversité de Shannon
Hmax = log2N ; N, nombre total d'individus
Pi, Contribution spécifique de chaque
espèce au recouvrement (0 = Pi = 1);
log2, logarithme à base 2.
4.2.4.3.2. La diversité inter-placeaux ou
diversité â
Pour quantifier le degré d'association ou encore le
niveau de similitude entre placeaux, nous avons fait recours au coefficient (ou
indice) de similarité ou de similitude (ou encore de communauté),
il correspond à l'inverse de la diversité â (similitude = 1
- â).
La diversité â traduit la diversité inter
formations (richesse en espèces communes entre plusieurs formations
végétales). Elle peut être par exemple le taux de
remplacement des espèces végétales dans un gradient
topographique, climatique ou d'habitat dans une région donnée
(Whittaker, 1972 ; Condit et al. 2002). Il existe plusieurs indices
dont les plus courants et les plus utilisés sont ceux de Jaccard
(J) et de Sorensen (Cs). Nous avons retenu l'indice de
Jaccard qui est la plus simple des expressions mathématiques de la
similarité des communautés végétales
(Mueller-Dombois & Ellenberg, 1974), il se base sur la
présenceabsence des espèces, indépendamment de leur
abondance, entre deux sites.
L'indice J, qui exprime le rapport entre le nombre
d'espèces communes et le nombre total des espèces pour les
placeaux pris deux à deux (Boudet, 1978), est élevé
lorsque les deux milieux ne présentent pas de grande différence
et atteint son maximum (c'est-à-dire 1), lorsque les deux milieux
comparés sont exactement identiques en terme floristique
(diversité nulle). Il équivaut à 0 lorsqu'à
l'inverse les deux n'ont aucune espèce commune (diversité
maximale).
a b c
+ +
a, nombre d'espèces propres au premier placeau
b, nombre d'espèces propres au deuxième placeau
c, nombre d'espèces communes aux deux placeaux

80
J a été calculé par le logiciel
PAST.
4.2.4.3.3. La richesse spécifique totale ou
diversité ã dans et en dehors du parc
W.
Le recensement de l'ensemble des espèces
herbacées rencontrées dans les relevés ne peut donner
qu'une idée par défaut des espèces effectivement
présentes dans le terroir de Kotchari : moins l'effort
d'échantillonnage est important, plus la liste obtenue est
incomplète. C'est pourquoi, vu le faible nombre de relevés
(trois) dans certaines unités paysagères (tableau IV6 plus loin),
la richesse réelle des unités paysagères (ou
diversité ã) n'a pas été calculée pour
chacune, mais globalement pour les deux grandes unités que constituent
l'intérieur et l'extérieur du parc W. Il a été
possible de simuler cette richesse réelle par des méthodes
d'extrapolation de la courbe aire-espèce - ou courbe d'accumulation ou
d'enrichissement des espèces et des relevés- et des coefficients
de variation. De tels estimateurs de richesse sont fondés sur
l'idée que l'information sur les espèces manquantes est
donnée par les espèces rares. Nous avons eu recours à la
méthode de Chao (1984) mise en oeuvre dans le logiciel EstimateS 7.5
(Colwell, 2004).
4.2.4.4. Le recouvrement ligneux
Le recouvrement ligneux par ligne a été
calculé selon la formule suivante : Dh
R(%) = × 100
L
Dh, Projection du houppier sur la ligne au sol
L, longueur de la ligne.
Les valeurs moyennes ont ensuite été
recherchées par placeau puis comparées entre elles par
unité paysagère pastorale.
4.2.4.5. La phytomasse herbacée et la
capacité de charge
A l'issue de la récolte, la nécromasse
(phytomasse ancienne) a été éliminée pour ne
prendre en compte que la production herbacée de la saison en cours. Les
30 échantillons collectés sur chaque placeau au moment de la
biomasse maximale ont immédiatement été pesés
(peson de sept kilogrammes de portée) pour en déterminer le poids
de matière fraîche. Le poids moyen de matière sèche
a été déterminé par calcul, après
évaluation de la teneur en eau de trois échantillons de 500 g par
placeau, séchés à l'étuve à 65 °C
pendant 48 heures (Akpo & Grouzis, 2004). Les résultats sont
exprimés en tonnes de matière sèche par hectare
(tMS.ha-1).
La production herbacée par unité
paysagère a été utilisée par la suite pour
déterminer leurs capacités de charge. En rappel, cet indicateur
dont le mode de calcul est varié (Boudet, 1978, Zoungrana, 1991 ;
Kagoné, 2000) est pris avec réserve particulièrement en
contexte de libre accès ou de conditions imprévisibles (Pratt,
1975 ; Carrière & Toutain, 1995 ; Allen et al. 2011)
(encadré II1, chapitre II). De nos jours, la capacité de charge
reste cependant une référence acceptable en particulier pour les
gestionnaires des parcours.

La capacité de charge (CC) saisonnière des
pâturages a été estimée de la manière
suivante :
BMkgMS ha CU
( / ) ×
CC UBT ha dp
( / / ) avec
= 6,25( / )
kgMS UBT dp
×
CC, capacité de charge en UBT/ha/période
(ou ha/UBT/période)50 ;
BM, biomasse maximale ;
CU, coefficient d'utilisation;
6,25, consommation journalière de l'UBT en kg de
matière sèche ;
dp, durée en jours de la période de la
saison de pâturage concernée, (soit 365 pour la capacité de
charge annuelle ; période de végétation active soit 105
jours pour la saison pluvieuse ; 260jours soit l'année moins la
période de végétation active pour la saison
sèche).
Nous avons recherché la capacité de charge
moyenne annuelle (dp = 365 j) pour toutes les unités légalement
accessibles (hors aires protégées) en toute saison et celle de
saison sèche (dp = 260 j) pour les unités qui ne sont accessibles
qu'en cette seule saison.
Le coefficient d'utilisation (CU) représente la portion
de la production potentielle qui peut être réellement
consommée en tenant compte des pertes dues à la consommation par
d'autres herbivores et différents insectes, aux moisissures, aux feux,
au piétinement (Breman & De Ridder, 1991) et à la part qui
doit être nécessairement préservée pour assurer la
couverture du sol le protégeant ainsi contre l'érosion (Boudet,
1978). En appliquant ce coefficient à la phytomasse on obtient le
disponible fourrager (DF). Il est par ailleurs fonction de la saison, du
domaine climatique, du statut hydrique du substrat (Boudet, 1991 ; Breman &
De Ridder, 1991) ou de la qualité du fourrage (Baars & Jeanes,
1997). Diverses estimations de CU sont données (Boudet, 1978 ; Breman
& De Ridder, 1991 ; Bourbouze, 1995 ; Baars & Jeanes, 1997), mais pour
le calcul de la capacité de charge annuelle et celle de saison
sèche (année moins période active de la
végétation) nous avons utilisé un CU unique de 35%), taux
le plus souvent utilisé (Boudet, 1978; Toutain et Lhoste, 1978). Il peut
cependant être moindre ou plus élevé - respectivement 20%
et 50% d'après Breman (1975), Boudet (1975) et Breman & De Ridder
(1991) ou même 75% (Compère et al. 1993) - mais notre
choix permet la comparaison avec une gamme de travaux conduits dans la
région.
La capacité de charge ainsi évaluée reste
cependant théorique, elle est plus faible si on tient compte du niveau
bas de la qualité moyenne de la matière sèche et de la
perte de production à plus ou moins longue échéance qui
survient nécessairement à la suite de l'action animale (Boudet,
1975 ; Pratt, 1975 ; Breman & De Ridder, 1991). Elle est par ailleurs
sujette à une grande variabilité suivant divers facteurs
notamment la pluviosité (Rivière, 1975 ; Pratt, 1975), ce qui
implique une réévaluation régulière. Une autre
manière, beaucoup plus pratique (procédé
expérimental) d'évaluer la capacité de charge peut
être de suivre les performances, en particulier l'évolution de
poids, du bétail exploitant directement les parcours (Boudet, 1975 &
1978).
50 La capacité de charge s'exprime
généralement en UBT/ha/an (ou en ha/UBT/an), en rapport avec un
animal théorique de référence (l'Unité
Bétail Tropical) équivalent en Afrique à un bovin de 250
kg de poids vif consommant 6,25 kg de matière sèche par jour.
Notons que, parfois, pour faire référence
à la capacité d'exploiter le potentiel fourrager existant, une
autre notion, la capacité de pâturage est utilisée (FAO,
1991). Elle prend en compte des facteurs comme la qualité du fourrage
(teneur en azote digestible) mais surtout la disponibilité d'un point
d'abreuvement du bétail. En l'absence de point d'abreuvement, cette
capacité de pâturage peut dont être déclarée
nulle (Baars & Jeanes, 1997).
4.2.4.6. La valeur pastorale des unités
paysagères pastorales
Pour déterminer la valeur pastorale des
différents pâturages, seules les espèces dites productives
(Daget & Poissonet, 1971) ou structurantes, c'est-à-dire celles dont
la contribution spécifique était supérieure ou
égale à 5%, ont été prises en compte (Sawadogo,
1996 ; Kièma S., 2007).
La valeur pastorale (VP) est une évaluation
globale de la qualité fourragère des parcours (Daget &
Poissonet, 1969 &1971 ; Grouzis, 1982 ; Daget & Godron, 1995). Elle
intègre à la fois l'abondance des espèces (contribution
spécifique CSi) et leurs qualités nutritionnelles (indice de
qualité spécifique ISi). Couramment utilisée pour comparer
les différents herbages consommés par les bovins (Daget, 2004),
elle se détermine selon la formule suivante :
VP 1
= × ( CSi ×
ISi) (Daget et Poissonet, 1969), avec
k
k, plus fort indice de l'échelle;
ISi, indice de qualité spécifique ou
coefficient de valeur de l'espèce i ; CSi, contribution
spécifique de l'espèce i
FSi
(Boudet, 1991)
CSi, contribution spécifique de l'espèce
i, égale à 100
×
n
FSi
i = 1
FSi, fréquence de l'espèce i ; n,
nombre d'espèces sur la ligne.

82
4.3. Résultats et discussion
4.3.1. Définition des unités
paysagères pastorales
La partie du terroir située en dehors de l'aire
protégée pour laquelle il n'existait pas de carte de
végétation, a été étudiée uniquement
par analyse de l'image ASTER du 10 octobre 2006. Pour la partie du terroir
située dans l'aire protégée nous avons utilisé les
résultats d'une carte de végétation de l'ensemble du parc
du W établie par le programme Régional ECOPAS
(Écosystèmes protégés en Afrique soudano
sahélienne) à partir de huit scènes Landsat ETM7 acquises
entre le 31 octobre 1999 et le 19 mai 2000 (Arbonnier et al., 2002);
cette carte comporte certaines zones externes à l'aire
protégée (zones cultivées) ou non classées (zones
brûlées).
Six unités paysagères pastorales (UPP) ont
été reconnues en dehors de l'aire protégée, (Carte
IV-1 & tableau IV-3). Les unités 4 (hauts glacis et plateaux sur
sols ferrugineux

lessivés indurés) et 1 (bas-fonds) les plus
étendues couvrent chacune environ un quart de la superficie du terroir.
Elles sont suivies de l'unité de buttes et cuirasse
représentée essentiellement par la chaîne du Gobnangou.
La classification de la partie du terroir située dans
le Parc du W a été faite en se basant sur les unités que
nous venions de définir dans la périphérie du Parc et sur
la carte ECOPAS. Parmi les huit unités originelles de cette carte, les
formations de savane arbustive claire à herbacées annuelles et de
savanes arbustives à annuelles ont tout d'abord été
regroupées, les zones cultivées (externes à l'aire
protégée) ou brûlées étant ignorées,
pour définir cinq unités paysagères pastorales dans le
parc du W (UPw) (tableau IV-4 et carte IV-1).
Unités paysagères pastorales hors aire
protégée
UPP1 (80,10 km2; 22,90%)
UPP2 (28,53 km2; 08,20%)
UPP3 (39,95 km2; 11,40%)
UPP5 (42,73 km2; 12,20%)
UPP4 (93,59 km2; 26,80%)
UPP6 (64,82 km2; 18,50%)
Unités paysagères dans l'aire
protégée
UPw1
UPw2
UPw3
UPw4
UPw5
Zones brulées


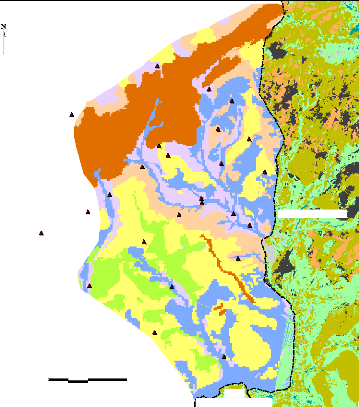
Bobouamondi
0 3 6
Kilomètres
TANSARGA
Bodiaga
Malipoa
Kobana
Kotchari-Centre
Natongou
Nadiagou
Diamanga
Koukongou
Pambidjeni
Pielgou
Banduo
de chasse Kourtiagou
Fantama
Tambouli
Kpenkouandi
Conc
Kobdañ
Gnimboama
Nangbanli
Fouanbiga
Irgomou
Tiontionga
Lada
Todoanga
Parc National du W
Carte IV-1. Les unités paysagères pastorales du
terroir de Kotchari

84
Le rapprochement des deux classifications paysagères,
étayé par les observations au sol notamment lors des
relevés a permis de mettre en évidence concordances suivantes
(tableau IV-5):
· Les unités paysagères de bas-fonds
(UPP1) et de plaine hydromorphe (UPP2) de la périphérie sont
assez proches des unités de savane arborée à
herbacée pérenne (UPw3), des cordons ripicoles (UPw2) et des
prairies hydromorphes (UPw5) de l'aire protégée. Ce sont des
unités paysagères qui, au plan topographique, se trouvent dans
les parties les plus basses du terroir (bas-fonds et plaines) et aux sols
généralement argileux à argilo-limoneux hydromorphes et
profonds.
· Les unités paysagères de bas et moyens
glacis (UPP3) ainsi que celles de haut glacis et de plateaux (UPP4) peuvent
être rapprochées dans l'aire protégée aux savanes
arbustives à herbacées annuelles (UPw4). Au plan topographique
ces entités ont en commun de se situer entre les parties les plus basses
du terroir (bas-fonds et plaines) et les parties les plus hautes (Buttes
rocheuses ou cuirassées); en plus les sols y sont peu à
moyennement profonds.
· L'unité paysagère des buttes et
cuirasses (UPP6) équivaut dans l'aire protégée aux
affleurements rocheux et cuirassés à végétation
clairsemée (UPw1). Ces unités représentent les sols les
plus superficiels trouvés dans l'aire d'étude.
Tableau IV-3. Unités paysagères reconnues dans le
terroir de Kotchari (hors Parc du W)
|
|
Superficie
|
% du
|
|
Nom
|
Topographie et sol
|
(km2)
|
terroir
|
|
UPP1
|
Bas-fonds sur sol hydromorphe à pseudogley de surface
|
80,10
|
22,90
|
|
UPP2
|
Plaine inondable sur sol hydromorphe à pseudogley de
surface
|
28,53
|
8,20
|
|
UPP3
|
Bas et moyen glacis sur sols ferrugineux lessivés
indurés à taches et concrétions
|
39,95
|
11,40
|
|
UPP4
|
Haut glacis et plateaux sur sols ferrugineux lessivés
indurés
|
93,59
|
26,80
|
|
UPP5
|
Mosaïque agroforestière sur sols ferrugineux
lessivés indurés
|
42,73
|
12,20
|
|
UPP6
|
Buttes et cuirasses
|
64,82
|
18,50
|
Tableau IV-4. Regroupement des unités paysagères de
la carte du Parc du W établie par le programme ECOPAS (Arbonnier et
al. 2002)
Nom Description
UPw1 Affleurements rocheux et cuirasses à
végétation clairsemée
UPw2 Cordon ripicole, galerie forestière et
fourré
UPw3 Savane arborée et boisée dominée par
les herbacées pérennes
UPw4 Savane arbustive dominée par les herbacées
annuelles

UPw5 Savane herbeuse, prairie hydromorphe,
végétation aquatique
Tableau IV-5. Correspondances entre unités de paysages
définies dans la périphérie et dans le Parc du W
UPP périphérie UPw d'après carte
Commentaire
du Parc (1) d'ECOPAS (2)
|
UPP1 et UPP2 UPw2, UPw3 et
UPw5
|
Parties les plus basses du terroir, sols
généralement argileux à argilo-limoneux hydromorphes et
profonds
|
UPP3 et UPP4 UPw4 Sols peu à moyennement profonds
UPP6 UPw1 Sols les plus superficiels (buttes et
cuirasses)
(1) Cette étude (voir tableau IV3)
(2) Arbonnier et al. (2002)
4.3.2. Les groupements agrostologiques et leurs
espèces caractéristiques
La classification hiérarchique a permis d'isoler six
grands ensembles à l'intérieur desquels le degré de
similitude (J) entre relevés est supérieur à 0,5.
Cependant, l'examen de ces ensembles, en particulier l'analyse de la valeur de
l'indice IndVal à chacun des niveaux du dendrogramme, a conduit à
scinder certains et à reconnaître au total dix groupements
agrostologiques (figures IV-1 & IV-2).
Les groupements floristiques du Parc du W se séparent
nettement de ceux du terroir de Kotchari. Ainsi, un premier ensemble (Ga5) qui
se dessine dès la deuxième partition n'est composé que de
relevés effectués dans le Parc. Il se caractérise par cinq
espèces dont l'indice Indval y est maximum: Diheteropogon
amplectens, Hyparrhenia smithiana, Andropogon chinensis, Hyparrhenia
subplumosa et Andropogon gayanus. À la partition suivante
Ga5 éclate en Ga5-1 caractérisé par Schizachyrium
sanguineum, Hyparrhenia subplumosa, Hyparrhenia smithiana et
Andropogon gayanus et Ga5-2 plutôt caractérisé par
Andropogon chinensis et Diheteropogon amplectens.
A ce premier ensemble s'oppose la totalité des autres
relevés, caractérisés principalement par Andropogon
pseudapricus et Schizachyrium exile, qui se séparent en
deux ensembles.
Le premier d'entre eux, caractérisé par
Aristida funiculata et Loudetia togoensis, va d'une part conduire
à Ga4 et Ga6 (partition 4). Ga4, caractérisé par
Eragrostis tremula et Eragrostis tenella, correspond à des
zones exploitées dans le terroir de Kotchari. D'autre part, les
relevés de Ga6, en revanche plutôt liés au Parc, se
caractérisent par Loudetia simplex, Loudetia togoensis, Aristida
funiculata et Andropogon tectorum. À la partition 5 ces
derniers se séparent en Ga6-1 avec L. simplex et L. togoensis
et Ga6-2 avec Aristida adscensionis, A. funiculata et Andropogon
tectorum.
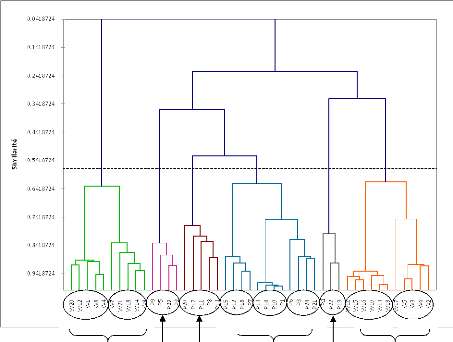
Ga5-2
Ga5-1
Ga3-1
Ga3-2
Ga3-3
Ga6-1 Ga6-2

86
Ga5 Ga2 Ga3
Figure IV-1. Dendrogramme des groupes agrostologiques des
pâturages de Kotchari
Légende
Ga1 : Groupement à Oryza longistaminata, Echinochloa
stagnina et Echinochloa colona ;
Ga2 : Groupement à Sorghastrum bipennatum,
Abildgaardia hispidula, Microchloa indica et Kyllinga pumila ;
Ga3-1 : Groupement à Brachiaria jubata, Sporobolus pyramidalis
et Melochia corchorifolia ;
Ga3-2 : Groupement à Andropogon pseudapricus,
Schizachyrium exile ;
Ga3-3 : Groupement à Digitaria debilis, Dactyloctenium
aegyptium et Setaria pallide-fusca ; Ga4 : Groupement à
Eragrostis tremula et Eragrostis tenella ;
Ga5-1 : Groupement à Schizachyrium sanguineum,
Hyparrhenia subplumosa, Hyparrhenia smithiana et Andropogon gayanus
;
Ga5-2 : Groupement à Andropogon chinensis et
Diheteropogon amplectens ; Ga6-1 : Groupement à Loudetia
simplex et Loudetia togoensis ;
Ga6-2 : Groupement à Aristida funiculata,
Aristida adscensionis et Andropogon tectorum.
Le second permet de différencier trois groupements Ga1,
Ga2, et Ga3 situés sur le terroir de Kotchari. Ga1 est nettement
caractérisé par Oryza longistaminata et Echinochloa
stagnina et Echinochloa colona. Ga2, nettement individualisé, est
caractérisé par Sorghastrum bipennatum, Abildgaardia
hispidula, Microchloa indica et Kyllinga pumila, toutes ces
espèces y présentant leur valeur maximale de l'indice Indval. Au
sein de Ga3 caractérisé principalement par la rudérale
Melochia corchorifolia, apparaissent trois sous-groupes.
Ga3-1 se caractérise nettement par Brachiaria jubata,
Sporobolus pyramidalis et Melochia corchorifolia et s'oppose
à un ensemble à Andropogon pseudapricus, Schizachyrium exile,
et

Sida acuta. Au sein de ce dernier il est possible de
différencier le groupement Ga3-2, qui reste caractérisé
par les deux dernières espèces, alors que le Ga3-3 se
différencie surtout par les adventices Digitaria debilis,
Dactyloctenium aegyptium et Setaria pallide-fusca.
4.3.3. Caractéristiques des unités
paysagères pastorales
4.3.3.1. La richesse en groupements agrostologiques des
unités paysagères
Pour évaluer leur qualité, notamment leur
composition en groupements agrostologiques, les unités paysagères
pastorales obtenues par analyse d'image ont été comparées,
à l'aide d'une matrice de confusion, aux groupements agrostologiques
obtenus par analyse floristique (tableau IV-6). Au total, la matrice de
confusion montre une bonne cohérence entre les deux typologies.
Tableau IV-6. Matrice de confusion entre unités
paysagères et groupement agrostologiques
|
Groupements
agrostologiques
|
|
|
|
Unités paysagères
|
|
|
|
|
Dans l'aire protégée
UPw 1 UPw
3 UPw 4
|
UPP1
|
En dehors de l'aire
protégée
UPP2 UPP3 UPP4 UPP5
|
UPP6
|
Total
|
|
Ga1
Ga2
|
|
|
|
|
5
|
4
|
|
|
|
|
5
4
|
|
Ga3-1
|
|
|
|
|
|
3
|
|
|
1
|
4
|
|
Ga3
|
Ga3-2
|
|
|
|
|
|
|
2
|
|
2
|
4
|
|
Ga3-3
|
|
|
|
|
|
|
4
|
|
|
4
|
|
Ga4
|
|
|
|
|
|
|
|
|
3
|
|
3
|
|
Ga5-1
|
|
4
|
1
|
|
|
|
|
|
|
5
|
|
Ga5
|
Ga5-2
|
|
2
|
3
|
|
|
|
|
|
|
5
|
|
Ga6-1
|
|
|
6
|
|
|
|
|
|
|
6
|
|
Ga6
|
Ga6-2
|
3
|
|
2
|
|
|
|
|
|
|
5
|
|
Total
|
|
3
|
6
|
12
|
5
|
4
|
3
|
6
|
3
|
3
|
45
|
Les chiffres dans les cases correspondent au nombre de
relevés dans le groupement agrostologique ou dans l'unité
paysagère pastorale correspondante.
Le résultat indique que l'unité UPw4 est la plus
hétérogène (4 groupements agrostologiques), suivent UPw3,
UPP4 et UPP6 (deux groupements agrostologiques chacune). Les autres
unités, les plus homogènes, se composent de seulement un
groupement agrostologique chacune.
|
|
Partition 2
|
|
|
|
Ga 5
S = 44
cv = 9
n = 10
|
Diam (100, 6) Hysm (100, 4)
Anch (99, 37) Hysu (90, 4)
Anga (89, 11) Scsa (48, 3)
|
Anps (92, 60) Loto (64, 42)
Scex (60, 12) Arfu (60,8)
|
S = 144 cv = 5 n = 35
|
|
|
|
|
|
|
|
|
Arfu (90, 7) Losi (81, 20)
Loto (80, 31) Ante (61, 11) Arad (37, 2)
|
|
|
|
Anps (67, 47) Ecco (43, 4) Ecst (43, 3) Miin (41, 7) Meco (33,
5) Orlo (26, 4)
|
S = 137 cv = 7 n = 21
|
|
|
|
S = 86
cv = 8
n = 14
|
|
Partition 4
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
S = 22 cv = 9 n = 5
|
Scsa (88, 3) Hysu (86, 4) Hysm (81, 4) Anga
(53, 9)
Ga 5-1
|
|
|
Anch (76, 28) Diam (67, 4)
Ga 5-2
|
|
|
|
|
|
|
|
Losi (61, 14)
Loto (48, 22)
Ga 6-1
Partition 7
Ga 3-3
Arad (70, 2) Arfu (61, 5) Ante (54,7)
Ga 6-2
S = 29 cv = 8 n = 5
S = 38 cv = 3 n = 6
S = 66 cv = 3 n = 4
S = richesse spécifique ;
cv = coefficient de variation ;
n = nombre de relevés (taille du
groupe)
Partition 3
|
S = 65 cv = 11 n = 3
|
Erte (92, 2)
Ertr (89,
4)
Ga 4
|
|
Losi (92, 20) Arfu (83, 7) Ante
(81, 11) Loto (61, 29)
Ga 6
|
S = 47 cv = 10 n = 11
|
Sobi (94, 6) Ecst (59, 3) S = 135
Abhi (71, 3) S = 46 Ecco (55, 3) cv = 8
Miin (67, 5) cv = 12 Anps (44, 42) n = 17
Kypu (61, 2) n = 4 Scex (41, 6)
Meco (41, 5)
Ga 2
Partition 5


Orlo (96, 4)
Ecst (74,
3)
Ecco (70, 3)
Ga 1
Meco (58, 5) Scex (37, 6) Anps (35, 33)
Ga 3
S = 111 cv = 7 n = 12
Partition 6
|
Brju (100, 4) Sppy (100, 2) Meco (52, 2)
Ga 3-1
|
S = 61 cv = 9 n = 4
|
Siac (97, 3) Scex (37, 6) Anps (27, 24)
|
S = 83 cv = 5 n = 8
|
S = 52 cv = 12
n = 5
S = 37 cv = 10 n = 5
S = 51 cv = 9 n = 4
Siac (90, 2) Scex (31, 4)
Ga 3-2
Dide (100, 2) Daae (90, 2) Sepa (89, 3) Anps
(25, 14)
Légende
Seules les espèces statistiquement significatives
à p = 0,05 sont retenues. Entre parenthèses, les valeurs d'IndVal
et les contributions spécifiques moyennes ; les espèces en gras
sont celles qui ont leur maximum d'indice dans la partition ; S = richesse
spécifique du groupe, cv = coefficient de variation (% de la moyenne), n
= taille du groupe (nombre de relevés dans le groupe). La signification
des abréviations des noms des espèces est donnée en annexe
3
Figure IV-2. Les espèces caractéristiques des
différents groupements agrostologiques des pâturages

89
Par ailleurs, chaque unité paysagère a
été caractérisée par le recouvrement et la
densité des espèces ligneuses les plus abondantes (tableau
IV-7).
- L'unité paysagère UPP1 des bas-fonds sur sols
hydromorphes correspond au groupement agrostologique Ga1 constitué
d'espèces herbacées caractéristiques de milieux
humides. Les espèces ligneuses principales de cette
unité sont Mitragyna inermis et Anogeissus leiocarpa.
- L'unité paysagère UPw3 de savane arborée et
boisée est dominée par les herbacées pérennes. Elle
correspond surtout au groupe agrostologique Ga5-1 caractérisé par
les graminées vivaces de sols profonds. Le groupement
agrostologique Ga5-2 bien que moins bien représenté, s'y
rencontre également. L'espèce ligneuse dominante ici est
Terminalia laxiflora.
Dans une certaine mesure, ces deux unités UPP1 et UPw3
s'apparentent aux savanes herbeuses de bas-fonds à Mitragyna
inermis de Gampela avec cependant une plus grande densité
arborée (Zoungrana, 1991) ou aux formations ripicoles de bas-fonds
à Combretum nigricans, Brachiaria lata et
Sporobolus pyramidalis de Luili-Nobéré (Kagoné,
2000).
- L'unité paysagère UPP2 des plaines inondables
sur sols hydromorphes correspond au groupement agrostologique Ga2
caractérisé également par des espèces
herbacées hydrophiles. Ici, Terminalia macroptera et
Combretum molle sont les espèces ligneuses dominantes. C'est
une unité qui présente de fortes similitudes avec les savanes
arborées de vallée hydromorphe à Vitellaria paradoxa,
Andropogon gayanus et Pennisetum pedicellatum identifiées
par Zoungrana (1991).
- L'unité paysagère UPP3 des sols ferrugineux
à taches et concrétions de bas et moyens glacis correspond au
groupement Ga3-1 qui se caractérise également par des
espèces herbacées de milieux humides ou à inondation
temporaire Les observations au sol montrent en effet que cette unité
représente des zones de transition entre milieux nettement humides ou
inondés et milieux secs. Les principales espèces ligneuses sont
Terminalia mollis et Gardenia aquala. Cette unité, par
sa position topographique surtout, s'apparente aux diverses savanes
arborées et arbustives de glacis rencontrées à Gampela et
dans la forêt du barrage de Ouagadougou (Zoungrana, 1991) ou aux
jachères d'âge variable sur bas glacis rencontrées au
Centre-sud du Burkina (Kagoné 2000).
- L'unité paysagère UPP4 de plateaux et hauts
glacis sur sols peu profonds correspond aux groupements agrostologiques Ga3-3,
caractérisé par des espèces adventices ou de jeunes
jachères et Ga3-2 caractérisé par des espèces
indicatrices de dégradation pastorale. Cette unité peut
être considérée comme très anthropisée. Dans
cette unité, Vitellaria paradoxa, Combretum glutinosum et
Khaya senegalensis sont les principales espèces ligneuses.
- L'unité paysagère UPw4 de savane arbustive de
l'aire protégée est dominée par des herbacées
annuelles. Relativement hétérogène, elle correspond
principalement au groupement agrostologique Ga6-1, mais aussi au groupement
Ga5-2. Cette unité peut être vue comme une transition entre les
sols profonds situés dans les bas de toposéquence (UPw3) et les
sols peu profonds sur élévations (UPw1). Elle est dominée
par l'espèce ligneuse Acacia gourmaensis.
Les unités UPw4 et surtout UPP4, qui présentent
une grande similitude au plan topographique et même de la composition
botanique surtout ligneuse, sont couramment rencontrées ailleurs en zone
soudanienne. Ainsi, une unité pastorale similaire à ces
unités, mais de stade évolutif différent au regard de son
cortège floristique - savane arborée à Vitellaria
paradoxa et

Andropogon gayanus - a été
identifiée au sein de la forêt du barrage de Ouagadougou
(Zoungrana, 1991). Une autre variante, les savanes arborées sur glacis
de versant cuirassé à Anogeissus leiocarpa et
Pennisetum pedicellatum, a été rencontrée
à Luili-Nobéré par Kagoné (2000).
- L'unité paysagère UPP5, consiste en des
mosaïques agroforestières dans les endroits les plus
cultivés du terroir. Elle correspond au groupement agrostologique Ga4
qui est caractérisé par des adventices annuelles messicoles et
par Diospyros mespiliformis. Les espèces ligneuses principales
de cette unité sont cependant Vitellaria paradoxa et Acacia
gourmaensis. Ces unités typiques des milieux cultivés de
zone nord-soudanienne ont été définies à Gampela
(savane arborée à Vitellaria paradoxa et
Schizachyrium exile) (Zoungrana, 1991) et à
LuiliNobéré (savane-parc anthropique sur bas-glacis à
Vitellaria paradoxa et Parkia biglobosa) (Kagoné,
2000).
- L'unité paysagère UPP6 des buttes et cuirasses
abrite le groupement agrostologique Ga3-2, déjà présent
dans l'unité paysagère UPP4. On y rencontre aussi le groupement
agrostologique Ga3-1, dans les parties les plus basses où le sol peut
être moyennement profond et humide. L'espèce ligneuse principale
de cette unité paysagère est Combretum micranthum.
- L'unité paysagère des affleurements rocheux de
l'aire protégée (UPw1) correspond au groupement agrostologique
Ga6-2 caractérisé par les graminées annuelles de sols
superficiels. Les espèces ligneuses principales de cette
unité paysagère sont Combretum nigricans et Detarium
microcarpum.
Ces deux dernières unités, typiques des buttes
et cuirasses, correspondent à peu près à la savane
arbustive sur glacis de versant rocheux à Combretum glutinosum
et Brachiaria lata de Kagoné (2000).
La répartition des différents groupements
agrostologiques dans les unités paysagères permet de
définir les types de biotope auxquels ils sont liés. Il existe,
en effet, un gradient de distribution des relevés allant de la gauche
(Ga5, Ga1 et G2 : relevés de milieux humides de bas de
toposéquence) vers la droite (Ga4, Ga6 : relevés de milieux secs
de haut de toposéquence). Les facteurs pédologiques,
topographiques et anthropiques, moteurs de cette répartition, ont
été tour à tour prépondérants, agissant
parfois à force égale, aux différentes partitions. Ainsi,
à la première partition des relevés, le groupement
agrostologique de milieux les plus humides, les plus bas et les moins
anthropisés (Ga5) s'oppose au reste des groupements. A la
deuxième partition par contre, les facteurs anthropiques semblent
prépondérants. On voit, en effet, que le groupement Ga4 de
l'unité de mosaïque agroforestière, milieu assez
fragmenté par le parcellaire agricole, est associé au groupement
Ga6 situé majoritairement sur les affleurements. Au-delà du
caractère sec de leurs substrats, leur proximité pourrait
s'expliquer par la faible emprise du feu qui autoriserait la présence au
sein de ces groupements d'espèces non pyrotolérantes comme
Pennisetum pedicellatum (César, 1991) ou Diospyros
mespiliformis. Les autres groupements présents en dehors de l'aire
protégée ont en commun d'être soumis à plusieurs
facteurs anthropiques (feu, exploitation animale, etc.), qui selon leur
ampleur, déterminent, en sus des facteurs édaphiques
sous-jacents, leur structuration spatiale (Bourlière & Hadley, 1983
; Cole, 1986 ; Scholes & Walker, 1993).

91
Certains de nos groupements surtout ceux des parties hautes du
terroir, ont été déjà décrits dans la
même zone ou ses environs. Ainsi, Hahn-Hadjali (1998) et Wittig et
al. (2000), travaillant respectivement à l'échelle de la
grande région ou simplement sur la chaine du Gobnangou, partiellement
incluse dans notre terroir, ont pu décrire le groupement à
Loudetia togoensis de sols squelettiques à peu profonds
identique au groupement Ga6-1. Ce groupement de milieux incultes (Wittig et
al. 2000) ou de vieilles jachères (Hahn-Hadjali, 1998), peut
s'étendre en contrebas de la chaîne sur les cuirasses
latéritiques ou, comme dans le cas présent, sur les plateaux
parfois cuirassés.
Wittig et al. (2000) distinguent aussi sur la
chaîne, dans les endroits à sols plus épais ou plus
humides, le groupement à Terminalia laxiflora et Combretum
glutinosum qui pourrait être vu comme une variante du
précédent dans laquelle les espèces Loudetia
annua et Andropogon pseudapricus s'associent à
Loudetia togoensis.
A bien des égards ce groupement est assimilable
à Ga3-2 qui peut, lorsqu'il se retrouve sur les plateaux aux sols
variés, comprendre à la fois le groupement à
Schizachyrium exile de très jeunes jachères et le
groupement à Andropogon pseudapricus de jachères
d'âges variés de Hahn-Hadjali (1998). Le groupement Ga3-2
présente également une certaine proximité avec un
groupement déjà identifié par Boudet (1975) ailleurs en
zone soudanienne et dans lequel l'espèce Andropogon
pseudapricus est souvent accompagnée de Diheterepogon
hagerupii et de graminées sciaphiles notamment Pennisetum
pedicellatum plutôt localisées.
Les groupements Ga3-3 de jachères d'âge moyen
(Hien, 1996 ; Fournier et al. 2001) et Ga4 de milieux cultivés
(différencié par des espèces messicoles ou de jeunes
jachères) (César, 1991; Fournier et al. 2001) sont des
groupements courants des milieux exploités en zone soudanienne. Par
rapport à la classification de Hahn-Hadjali (1998), Ga4 peut être
identifié comme la variante à Eragrostis tremula
située sur les sols profonds à texture sableuse du groupement
à Schizachyrium exile. Ga3-3 y apparait par contre comme une
variante d'un groupe plus large à Schoenefeldia gracilis sur
substrats généralement argileux (vertisols). Ce dernier
présente, par ailleurs, de bonnes similitudes avec Ga6-2, ses autres
espèces différentielles étant des espèces du genre
Aristida. Il faut noter que le groupement Ga4 est associé dans
notre étude au groupement Ga6 situé majoritairement sur les
affleurements. Audelà du caractère sec de leurs substrats, leur
proximité pourrait s'expliquer par la faible emprise du feu qui
autoriserait la présence au sein de ces groupements d'espèces non
pyrotolérantes comme Pennisetum pedicellatum ou Diospyros
mespiliformis (César, 1991).
Le groupement Ga1, de terrain limono-argileux à
inondation importante plus ou moins prolongée situé
généralement dans les marécages ou les bas-fonds, se
rencontre aussi dans les savanes du nord ivoirien (César, 1991) ou dans
divers autres pâturages soudaniens (Boudet, 1975). Aussi, sur certains
faciès à tendance monospécifique dans des
dépressions, on peut l'assimiler au groupement à Echinochloa
stagnina identifié en zones humides du nordCameroun, sous climat
relativement plus humide (1000 mm/an) (Onana et al. 2003). En outre,
le groupement Ga2 de sol hydromorphe argilo-sableux sur plaine inondable, est
proche du groupement à Heteropogon contortus de Onana et
al. (2003), au regard des espèces qui le caractérisent,
il marque la transition entre prairies semi-aquatiques et groupements
mésophiles.
Chapitre IV. Les unités agrostologiques
Tableau IV-7. Espèces ligneuses dominantes (nombre de
pieds.ha-1) des unités paysagères pastorales du
terroir de Kotchari
|
Espèces
|
UPP1
|
UPP2
|
UPP3
|
UPP4
|
UPP5
|
UPP6
|
UPw3
|
UPw4
|
UPw1
|
|
Acgo
|
132,2a
|
258,75ab
|
343,67b
|
192,83ab
|
363b
|
330,67b
|
214,17ab
|
550,08c
|
397,17b
|
|
Anle
|
1228,2c
|
569,5b
|
---
|
135a
|
249,33ab
|
---
|
300ab
|
119,75a
|
---
|
|
Cogl
|
---
|
---
|
209,67a
|
556c
|
333,33ab
|
245,33a
|
---
|
447,38b
|
294,44a
|
|
Comi
|
---
|
---
|
---
|
166,17b
|
---
|
405,33c
|
---
|
35,75a
|
292,5b
|
|
Como
|
---
|
662,25c
|
---
|
---
|
---
|
---
|
120,67b
|
41,33a
|
---
|
|
Coni
|
---
|
---
|
310a
|
311,83a
|
---
|
303,67a
|
---
|
296,36a
|
579,69b
|
|
Demi
|
---
|
---
|
---
|
295,17c
|
---
|
152,33b
|
---
|
101,01a
|
440,99d
|
|
Dime
|
16a
|
---
|
---
|
---
|
255,6c
|
---
|
32a
|
113,5ab
|
168,06ab
|
|
Gaaq
|
---
|
---
|
232,67b
|
61,33a
|
---
|
62,67a
|
---
|
---
|
---
|
|
Khse
|
8,6 a
|
---
|
30,33b
|
387,33c
|
66b
|
1,33a
|
---
|
---
|
---
|
|
Miin
|
1376,8b
|
---
|
---
|
---
|
---
|
---
|
351,67a
|
---
|
---
|
|
Tela
|
185,2a
|
706,5c
|
---
|
---
|
---
|
---
|
1003d
|
341,42b
|
---
|
|
Tema
|
149,4a
|
2477,5d
|
183ab
|
---
|
---
|
121a
|
542,17c
|
184,83ab
|
---
|
|
Temo
|
---
|
166b
|
1669,33c
|
---
|
---
|
---
|
11a
|
45,25a
|
---
|
|
Vipa
|
65,2a
|
210b
|
384,67b
|
1182d
|
632,33c
|
26,33a
|
93,67a
|
284,03b
|
67a
|
Sur la même ligne, les valeurs portant des lettres
différentes sont significativement différentes au seuil de 5%.
Les espèces dont les valeurs de densité sont en gras sont
liées aux unités paysagères correspondantes. Les cases
marquées en pointillé indiquent l'absence de l'espèce dans
l'unité paysagère correspondante.
Légende :
Acgo = Acacia gourmaensis, Anle = Anogeissus leiocarpus,
Cogl = Combretum glutinosum, Comi = Combretum micranthum, Como = Combretum
molle, Coni = Combretum nigricans, Demi = Detarium microcarpum, Dime =
Diospyros mespiliformis, Gaaq = Gardenia aquala, Khse = Khaya senegalensis,
Miin = Mitragyna inermis, Tela = Terminalia laxiflora, Tema = Terminalia
macroptera, Temo = Terminalia mollis, Vipa = Vittelaria paradoxa.

93
Le groupement Ga3-1, anthropisé comme en
témoigne la présence de Sporobolus pyramidalis, une
graminée vivace rudérale (César, 1991 ; César &
Zoumana, 1993), est proche du groupement à Aspilia
helianthoides de Hahn-Hadjali (1998). L'espèce Sporobolus
pyramidalis est en effet bien représentée dans ce
groupement.
Le groupement Ga5-1, de graminées vivaces
héliophytes à large amplitude écologique (César,
1991) sur sols profonds sableux, limoneux ou argilo-sableux, parfois
hydromorphes est un groupement de savane peu anthropisée, mais la
présence d'Andropogon gayanus y traduit sans doute un
passé cultural (César, 1991 ; Fournier et al. 2001). Il
présente une proximité avec le groupement Ga5-2 également
à héliophytes de sols sableux à caillouteux peu profonds
(César, 91) identifié par Fournier et al. (2001) sur des
substrats édaphiques plus secs et sur sol ferralitique.
4.3.3.2. La diversité floristique des
unités paysagères pastorales : les
diversités alpha (richesse S,
équitabilité E) bêta (J) et gamma (S Chao2)
La richesse moyenne locale (S) en espèces
herbacées des unités paysagères est de 19 à 52
espèces (tableau IV-8). Bien que la richesse spécifique soit
très variable entre relevés, on observe qu'elle est plus faible
dans l'aire protégée (19 à 23) qu'en dehors de l'aire
protégée (29 à 52). En dehors de l'aire
protégée, cette richesse en espèces est plus
élevée sur sols moyennement ou peu profonds (UPP3, UPP4, UPP5)
que sur sols profonds hydromorphes (UPP1 et UPP2). Toutefois, la richesse des
buttes et cuirasses (UPP6) est relativement faible et équivalente
à celle des unités sur sols profonds (UPP1 et UPP2). Dans l'aire
protégée l'unité UPw4 peut être
considérée comme unité de transition, sa richesse en
espèces herbacées est plus grande que celles des unités de
savane arborée et boisée à pérennes (UPw3) et des
affleurements rocheux et cuirassés (UPw1).
L'indice d'équitabilité E de Pielou
varie d'un minimum de 0,62 pour les savanes arborées et boisées
à pérennes de sols profonds de l'aire protégée
(UPw3) à un maximum de 0,89 pour les mosaïques
agroforestières sur sols ferrugineux lessivés indurés ou
unités cultivées (UPP5) (tableau IV-8). Les valeurs
enregistrées indiquent que les unités sont presque toutes
homogènes car elles sont soit légèrement
inférieures ou alors supérieures à 0,80, valeur
considérée comme traduisant une forte
homogénéité de la végétation d'un milieu
(Daget, 1976). De manière générale, la composition
floristique des unités paysagères est plus équitablement
répartie en périphérie (0,79 à 0,89) que dans
l'aire protégée (0,62 à 0,73). Enfin, que ce soit dans
l'aire protégée ou en dehors de celle-ci, plus les sols sont
superficiels et secs, plus la valeur de l'équitabilité
augmente.
L'indice de similitude de Jaccard (J) qui traduit la
similitude spécifique entre les relevés d'une même
unité paysagère varie entre 0,46 (forte diversité â)
pour l'unité de savane arbustive de l'aire protégée (UPw4)
qui est la plus hétérogène et 0,91 (faible
diversité â) pour l'unité d'affleurements et de cuirasse de
la même entité (UPw1) qui est la plus homogène. La
diversité entre placeaux reflète ici la diversité des
unités paysagères en termes de groupes agrostologiques, les
unités les plus hétérogènes (UPW4, UPW3, UPP4,
UPP6) étant composées de plusieurs groupements agrostologiques,
les plus homogènes (UPW1, UPP1,
Chapitre IV. Les unités agrostologiques
UPP3, UPP2, UPP5) n'en comportant qu'un seul. Ceci confirme les
observations faites précédemment (tableau IV-6).
Tableau IV-8. Valeurs de diversités intra placeau (S,
E) et inter placeaux (J) des unités paysagères
pastorales du terroir de Kotchari (moyenne #177; écart-type).
|
Unité paysagère
pastorale
|
Richesse
spécifique (S)
|
Equitabilité
de Pielou
(E)
|
Indice de similitude
de Jaccard
(J)
|
|
UPP1
|
35,20 #177; 1,48ab
|
0,79 #177; 0,01abc
|
0,88 #177; 0,06c
|
|
UPP2
|
29,25 #177; 3,30 ab
|
0,81 #177; 0,01abc
|
0,80 #177; 0,08bc
|
|
UPP3
|
45,33 #177; 1,53b
|
0,83 #177; 0,02bc
|
0,85 #177; 0,21bc
|
|
UPP4
|
45,50 #177; 12,88b
|
0,84 #177; 0,04c
|
0,66 #177; 0,11b
|
|
UPP5
|
51,67 #177; 1,53b
|
0,89 #177; 0,01c
|
0,82 #177; 0,06bc
|
|
UPP6
|
32,00 #177; 7,55ab
|
0,82 #177; 0,01abc
|
0,73 #177; 0,04b
|
|
UPw3
|
19,00 #177; 3,16a
|
0,62 #177; 0,06 a
|
0,55 #177; 0,19ab
|
|
UPw4
|
23,00 #177; 3,91a
|
0,69 #177; 0,03ab
|
0,46 #177; 0,14 a
|
|
UPw1
|
20,00 #177; 1,73a
|
0,73 #177; 0,07abc
|
0,91 #177; 0,03c
|
Sur la même colonne, les valeurs portant les mêmes
lettres ne sont pas significativement différentes au seuil de 5%.
La diversité régionale, ou diversité
ã, évaluée par le calcul de la richesse estimée
(Schao2) (tableau IV-9) est plus élevée en dehors de
l'aire protégée (143,6) qu'à l'intérieur 80,6). Il
est ainsi confirmé à cette échelle d'observation que la
diversité spécifique des herbacées est moindre dans l'aire
protégée.
Tableau IV-9. Diversité gamma (ã) dans le terroir
de Kotchari. Comparaison des secteurs dans et hors du Parc du W (valeurs
moyennes et écart-type)
|
Unités
Paysagères
pastorales
|
SChao2
|
|
SChao2 moyen SChao2 min (95%
IC) SChao2 sup (95% IC)
|
Parc W 80,56 #177; 4,62a 76,34 98
Hors Parc W 143,64 #177; 1,91b 142,27 152,1
Les valeurs portant les mêmes lettres ne sont pas
significativement différentes au seuil de 5%.
Nos résultats indiquent que la richesse
spécifique herbacée est plus élevée à
l'extérieur qu'à l'intérieur de l'aire
protégée. Des observations analogues ont été faites
par plusieurs auteurs dans l'est burkinabé et dans le terroir même
de Kotchari (Hahn-Hadjali et al. 2006; Ouédraogo, 2008 ;
Sodré, 2009), mais aussi dans l'ouest burkinabé (Kièma S.,
2007). Dans cette dernière région, Devineau et al.
(2009) donnent des résultats plus nuancés pour de petites aires
protégées très anthropisées.


95
La richesse la plus forte en espèces herbacées
est observée sur les terres situées entre les parties les plus
basses (bas-fonds et plaines) et les plus hautes (buttes rocheuses ou
cuirassées) du terroir, qui correspondent aux milieux les plus
anthropisés (UPP5, UPP4, UPP3) (milieux les plus aptes à la
culture, jachères de divers âges bien pâturées). La
richesse spécifique des milieux équivalents à
l'intérieur du Parc est moindre, ainsi que leur
équitabilité. Ce dernier indice atteint des valeurs
particulièrement élevées dans l'unité la plus
cultivée (UPP5). Ce qui indique qu'il s'y déroule une faible
compétition entre espèces se traduisant par une dominance
d'espèces peu affirmée. Par ailleurs, la grande richesse
floristique qui y est observée serait la résultante de
perturbation modérée ou intermédiaire (Connell, 1978 ;
Sousa, 1979). D'après Connell (1978), en effet, dans les formations
naturelles ou peu transformées, quelques espèces les plus
compétitives dominent sous le seul effet de l'exclusion
compétitive51, à l'inverse, une communauté
continuellement perturbée, n'atteint jamais l'équilibre et les
espèces endémiques ont une forte probabilité d'extinction.
Les deux cas de figure conduisent à une faible diversité
floristique. Entre les deux extrêmes (état de perturbation
intermédiaire), la diversité est généralement, mais
pas toujours (Lubchenco, 1978), plus élevée. Travaillant dans un
terroir voisin de Kotchari, Hahn-Hadjali et al. (2006) font des
observations comparables. Hors de l'aire protégée, la richesse en
herbacées est plutôt faible sur les sols à hydromorphie
superficielle, argileux à argilolimoneux humides (UPP1, UPP2) comme
cuirassés (UPP6). Devineau et al. (2009) parviennent à
des résultats semblables dans l'ouest burkinabé : ils mettent en
évidence l'influence des facteurs édaphiques mais aussi celle de
l'usage des terres sur la diversité des peuplements herbacés. Ils
montrent que la perte de fertilité chimique par lessivage n'affecte pas
la richesse de la strate herbacée, celle-ci reste élevée
sur les terres les plus exploitées. César (1991) explique
d'ailleurs que les phorbes ubiquistes, notamment les légumineuses
fixatrices d'azote, s'accommodent de tels milieux épuisés (baisse
de la richesse en azote) tandis que les graminées ne peuvent y
subsister. L'effet d'enrichissement de la strate herbacée qui
résulte de la pratique de la jachère a par ailleurs
été clairement démontré. En zone soudanienne en
effet, la reconstitution de la strate herbacée des jachères, sur
10 à 30 ans selon le contexte, se fait par une succession
d'espèces messicoles (les adventices) puis savanicoles ; les mêmes
observations ont été faites dans les savanes du nord de la
Côte d'Ivoire par César (1991) et dans l'ouest burkinabé
par Hien (1996), Fournier et al. (2001) et Botoni-Liehoun et al.
(2006). À chaque étape de ce processus, dont la
première phase (1 à 10 ans) est marquée par un remaniement
(Zoungrana, 1993), le milieu s'enrichit en espèces diverses, qui
cohabitent momentanément sans dominance affirmée
(phénomène de chevauchement) avant de se substituer aux
précédentes. La dissémination par le bétail y joue
un rôle important comme vectrice d'espèces colonisatrices
d'espaces ouverts (Devineau 1999). Ce qui pourrait être le cas de
certaines espèces comme Zornia glochidiata, Tephrosia
pedicellata, Crotalaria spp., Indigofera spp.
(Légumineuses), Spermacoce spp. Sida spp.,
Commelina spp. (C. diffusa, C.
forskalaei & C. benghalensis) qui sont bien
représentées avec parfois de fortes
51 Le principe d'exclusion compétitive ou
principe de Gause (ou encore principe d'exclusion réciproque) (Gause,
1935 ; Dajoz, 1982) est un concept central en écologie fondamentale
(biologie des populations) selon lequel des populations de deux espèces
partageant exactement la même niche écologique dans un milieu
stable ne sauraient cohabiter indéfiniment. A plus ou moins long terme,
l'espèce la plus adaptée (ie : la plus compétitive)
éliminera l'autre.

96
contributions spécifiques. Dans l'aire
protégée, nos observations indiquent que l'unité UPw4 est
l'unité la plus pâturée illégalement, en
témoigne la présence d'espèces herbacées de milieux
anthropisés comme Digitaria spp., Brachiaria
spp. et même Tephrosia
pedicellata. La contribution de ces espèces est cependant infime
indiquant alors que cette présence animale est nettement moindre qu'en
périphérie du parc. Ces espèces indicatrices
d'anthropisation s'observent également en des proportions encore plus
faibles sur les affleurements (UPw1), Ceci suggère que les
éleveurs fréquentent surtout les plateaux puis les affleurements
et évitent les bas-fonds. L'enrichissement en espèces
herbacées est perceptible tant à l'échelle locale
(á) lors des phases de chevauchement de la succession post culturale
qu'à l'échelle du paysage (â et ã) par
diversification des milieux qui présentent chacun un stade
différent de reconstitution.
L'ensemble de ces observations confortent l'idée
souvent exprimée que la pression anthropique, jusqu'à un certain
niveau, est un facteur d'accroissement de la richesse spécifique
(Connell, 1978 ; Sousa, 1979; Bourlière & Hadley, 1983; Cole, 1986;
César 1992; Scholes & Walker, 1993; Huston, 1994; César 1994
& 2005; Rakotoarimanana et al. 2001; Rakotoarimanana et
al. 2008). Sous d'autres climats ou avec une anthropisation -
notamment une pression de pâture - plus intense une diminution de la
richesse floristique peut être observée comme par exemple au Sahel
(Hiernaux, 1998) ou dans une savane humide de Madagascar (Rakotoarimanana et
al. 2008). Quand aux valeurs plus élevées de
l'équitabilité hors des espaces protégés, elles
s'expliquent par une perturbation relativement plus importante qui limite la
dominance des espèces.
Le rôle des aires protégées dans la
conservation de la diversité biologique en région de savane est
diversement apprécié. Si des études mettent en
évidence une dégradation floristique due à la pression
anthropique dans les terres exploitées et la préservation de la
flore dans les aires protégées (Guinko et al. 2000),
d'autres montrent que la diversité végétale, et plus
précisément le nombre d'espèces, ne diffère pas ou
est même supérieure en dehors des aires protégées
(Shackleton, 2000; Hahn-Hadjali et al. 2006; Gardner et al.
2007). Il importe cependant de ne pas oublier que l'évaluation de la
diversité végétale peut varier en fonction de
l'échelle à laquelle elle est étudiée ou encore
selon les critères pris en compte (Devineau et al. 2009).
Ainsi, dans le cas de Kotchari, malgré le nombre d'espèces
herbacées plus important hors de l'aire protégée, on y
note la régression drastique des espèces caractéristiques
de la savane soudanienne, notamment les herbes pérennes, sous l'effet de
la pression d'exploitation, en particulier la pâture. Cette pression
favorise l'installation des herbes annuelles qui, par ailleurs, s'adaptent
mieux aux conditions pédoclimatiques médiocres ou
perturbées (César, 1992 ; Fournier & Nignan, 1997).
4.3.3.3. La phytomasse herbacée et la
capacité de charge
Il faut noter d'emblée que les évaluations de
phytomasse épigée qui sont conduites en milieu réel,
c'est-à-dire en libre accès, ne représentent que des
valeurs par défaut de la production potentielle dont une partie est
consommée par le bétail. Pour avoir cette production potentielle,
il est nécessaire que le pâturage considéré soit
soustrait de la pâture animale par un dispositif de mise en défens
(Boudet, 1975). La grande majorité des données

disponibles dans la littérature ont cependant
été acquises dans les mêmes conditions que les nôtres
et la comparaison paraît donc pertinente. Par ailleurs, pour des besoins
d'analyse de la pression saisonnière du bétail nous avons
recherché la capacité de charge de saison sèche pour les
unités dont l'exploitation est impossible (UPP1 & UPP2) ou
très modérée (UPP5) en saison pluvieuse. Les
capacités de charge des unités de l'aire protégée
n'ont pas été calculées, ces unités étant
légalement interdites au bétail domestique.
Les phytomasses (6,5 tMS.ha-1) et, en
conséquence, les capacités de charge annuelle les plus fortes
(1,06 UBT/ha/an) correspondent aux savanes arborées et boisées
à pérennes sur sols profonds du Parc (UPw3) (tableau IV-10). En
dehors de l'aire protégée, les valeurs de phytomasse (de
même que celles de capacité de charge) les plus
élevées sont observées sur les sols les plus profonds et
les plus humides (UPP1 & UPP2). La phytomasse et la capacité de
charge (en UBT à l'hectare ou en nombre jours pour un UBT) apparaissent
d'autant plus faibles que le sol est plus mince et plus sec.
Tableau IV-10. Biomasse herbacée (moyenne #177;
écart-type) et capacité de charge moyenne dans les
différentes unités paysagères
|
Unités
paysagères
Pastorales
|
Biomasse
(tMS.ha-1)
|
|
Capacité de charge
|
|
|
En saison sèche
|
Toute l'année
|
|
(UBT.ha-1)
|
(nb jour /
UBT.ha-1)
|
(UBT.ha-1)
|
(nb jour /
UBT.ha-1)
|
|
UPP1*
UPP2*
UPP3
UPP4 UPP5* UPP6 UPw3** UPw4** UPw1**
|
4,78 #177; 0,43c 4,59 #177; 0,53c 3,42
#177; 0,58bc 3,27 #177; 0,50b 1,41 #177; 0,50a
1,84 #177; 0,85a 6,50 #177; 0,63d 4,32 #177;
1,27c 2,55 #177; 0,56a
|
1,03 #177; 0,09b
0,99 #177;
0,11b
----
----
0,30 #177;
0,10a
----
----
----
----
|
152, 30 #177; 13,70b
146,24 #177;
16,89b
----
----
44,92 #177; 15,93a
----
----
----
----
|
----
----
0,52 #177; 0,09 b
0,50 #177; 0,08
b
----
0,28 #177; 0,13 a
----
----
----
|
----
----
81 #177; 14 b
77 #177; 12
b
----
43 #177; 21a
----
----
----
|
Sur la même colonne, les valeurs portant les mêmes
lettres ne sont pas significativement différentes au seuil de 5%.
* Unités paysagères pastorales peu ou pas
exploitées en saison pluvieuse
** Unités paysagères de l'aire
protégée à accès illégal.
nb, nombre
Il importe de noter qu'en étant évitées
ou peu pâturées en saison de pluies les unités UPP1, UPP2
et UPP5 voient leur potentiel de phytomasse mieux préservé et
leur capacité de charge plus importante que si elles n'étaient
pas épargnées.
Les tendances observées dans nos données de
phytomasse tout comme dans celles de capacité de charge en dehors de
l'aire protégée ont déjà été
enregistrées dans le même terroir (Zouri, 2003) ou ailleurs
(Boudet, 1978 ; Zoungrana, 1991 ; Sawadogo, 1996 et Kagoné, 2000)
(tableau IV11). Boudet (1975) notamment, à partir de travaux conduits
dans la bande soudanienne d'Afrique de l'Ouest, obtient une valeur de
phytomasse allant de 0,8 tMS.ha-1sur
les sols cuirassés à 2,5 tMS.ha-1pour
les terrasses colluviales ou savanes inondables en zone nord-soudanienne. Par
ailleurs, les plus fortes valeurs de phytomasse sont enregistrées dans
les sites ombragés non embroussaillés (3 tMS.ha-1),
toutefois les valeurs de phytomasse observées à Kotchari (y
compris dans l'aire protégée) sont plus élevées que
celles enregistrées par Boudet (1975). Elles sont, en outre, assez
proches mais plus variées que celles obtenues par Kagoné (2000)
et Zoungrana (1991) en zone nord-soudanienne du Burkina Faso. Par contre nos
données sont légèrement supérieures à celles
de Tiogo (Sawadogo, 1996), bien meilleures que celles enregistrées plus
au Nord dans la Tapoa (Barpoa, Botou et Kanlayenou) mais plus faibles que
celles de sites plus méridionaux (Nikki Kalalé au
Nord-Bénin et la province du Kénédougou à l'Ouest
du Burkina Faso). Ceci est une indication que la quantité de biomasse
produite suit le gradient climatique. Plus on va vers le sud, horizon plus
humide, plus celle-ci est importante. Par ailleurs, les données de
l'aire protégée enregistrées sont légèrement
plus faibles que celles enregistrées dans des aires
protégées voisines, la réserve de faune de Pama-Nord et le
ranch de gibier du Singou (2,84 à 7,25 tMS.ha-1, Savadogo,
2004). Si on revient aux données de Zouri (2003) obtenues dans le
même terroir de Kotchari, on peut observer, relativement à nos
données, que les capacités de charge des unités
paysagères ont depuis connu une baisse, cette baisse est notamment plus
importante sur les unités les moins productives.
Les capacités de charge enregistrées par
Kagoné (2000), en saison pluvieuse sont nettement supérieures aux
nôtres, mais l'auteur a utilisé un coefficient de 75% pour cette
saison.
Tableau IV-11. Différentes valeurs de phytomasse et de
capacité de charge en zone soudanienne
|
Lieux
|
phytomasse
(tMS.ha-1)
|
Capacité de charge
annuelle
(UBT.ha-1)
|
Sources
|
|
Kotchari (hors aires protégées)
|
1,41 - 4,78
|
0,28 - 0,99
|
Présente étude
|
|
Kotchari (aires protégées)
|
2,55 - 6,50
|
----
|
Présente étude
|
|
Kotchari
|
2,92 - 4,92
|
0,43 - 0,72
|
Zouri, 2003
|
|
Botou
|
1,87 - 2,8
|
0,27 - 0,41
|
Zouri, 2003
|
|
Kanlayenou (Botou)
|
0,32 - 2,07
|
0,05 - 0,32
|
Bambara, 2010
|
|
Barpoa (Diapaga)
|
0,73 - 2,36
|
0,11 - 0,36
|
Bambara, 2010
|
|
Réserve de faune de Pama-Nord
|
2,84 - 7,25
|
0,41 - 5,4
|
Savadogo, 2004
|
|
Luili-Nobéré
|
1,94 - 3,87
|
0,6 - 1,6*
|
Kagoné, 2000
|
|
Luili-Nobéré
|
1,94 - 3,87
|
0,2 - 0,4**
|
Kagoné, 2000
|
|
Nikki-Kalalé (Nord Bénin)
|
1,98 - 12,4
|
----
|
Oloulotan, 1988
|
|
Secteur nord-soudanien
|
0,8 - 3
|
0,11 - 0,37
|
Boudet, 1975
|
|
Secteur nord-soudanien
|
2,68 - 4,58
|
0,77 - 1,11
|
Zoungrana, 1991
|
|
Secteur Sud-soudanien
|
2,4 - 8,13
|
----
|
Zoungrana, 1991
|
|
Tiogo (Centre- ouest)
|
1 - 4,30
|
----
|
Sawadogo, 1996
|
|
Sidi, Banfoulaguè, Guèna
|
2,98 - 4,78
|
0,45 - 0,73
|
Yanra, 2004
|
|
(Kénédougou, Ouest)
|
|
|
|
* Capacité de charge de saison pluvieuse avec un
coefficient d'utilisation de 75% ; ** Capacité de charge de saison
sèche.

98

Les moindres phytomasses et capacités de charge
mesurées en dehors de l'aire protégée sont une
conséquence de la pâture animale et de la pauvreté en
nutriments des sols (Fournier, 1994), qui résultent toutes deux de la
pression d'exploitation (pastorale et agricole) dans ce secteur.
Cette situation d'inégale distribution de biomasse
entre la partie accessible du terroir et sa partie incluse dans l'aire
protégée, en dehors de la différence dans la
disponibilité immédiate pour le bétail, a une
conséquence sur les feux de brousse dont on connait le rôle majeur
dans l'entretien de ces milieux (César, 1991 & 1994 ; Hoffmann et
al. 2003) et dans les repousses d'arrière saison pluvieuse en
particulier pour les unités à herbacées pérennes
(Fournier, 1991 ; Kièma S., 2007). Une conséquence de cette
relative faiblesse de phytomasse herbacée hors de l'aire
protégée est que les feux de végétation y sont
moins violents que dans le Parc lui-même. Les aires
protégées de la région dans lesquelles se maintiennent les
écosystèmes de savane soudanienne sont ainsi plus
régulièrement et plus intensément parcourues par les feux
que les zones extérieures agricoles et pastorales à
végétation modifiée (Clerici 2006, Clerici et al.
2007, Devineau et al. 2010). Il faut rappeler que, du point de vue de
la gestion pastorale, le brûlage des parcours à annuelles est tout
à fait contre-productif (César, 1992 ; Kièma S., 2007)
alors que le brûlage des parcours à herbes pérennes
présente un intérêt fourrager en favorisant des repousses.
En fin de saison des pluies et début de saison sèche les pailles
sèches des pérennes sont relativement pauvres en matières
azotées et riches en silice et donc peu digestibles. En revanche leurs
repousses immédiatement après le passage du feu, s'il existe une
réserve en eau dans le sol, ou à défaut après les
premières pluies, sont très riches en nutriments et très
appétibles ; elles constituent un apport alimentaire précieux
à ces périodes difficiles « de soudure » (Fournier
1996).
4.3.3.4. Caractéristiques de la strate ligneuse
dans les unités paysagères pastorales
En dehors de l'aire protégée, la densité
la plus forte du peuplement ligneux est observée sur plateaux et hauts
glacis à sol peu ou moyennement profonds (UPP4). Le recouvrement des
ligneux (38%) apparente cependant cette unité à une formation
relativement ouverte (Emberger et al. 1983 ; Daget & Godron, 1995)
à dominance d'arbustes (tableau IV-12), autrement dit à une
savane arbustive.
Le recouvrement le plus fort (61%) est observé à
l'intérieur de l'aire protégée pour l'unité UPw3.
Les relevés montrent une strate arborée nettement dominante
(tableau IV-13) en accord avec la classification de la carte ECOPAS qui la
place parmi les savanes arborées denses.
Hors de l'aire protégée, c'est dans
l'unité UPP1 que l'on rencontre la végétation la plus
dense, une savane arborée à 53% de recouvrement. L'unité
UPP2, avec une quasi codominance des ensembles arborés et sous
arbustifs, correspond à une forme plus ouverte sensu Emberger
et al. (1983.) et peut être qualifiée de savane
boisée claire (sensu Devineau, 1997). Les autres unités
sont des savanes arbustives au recouvrement moyen (40 et 43% pour UPP3 et UPw4)
ou des savanes arbustives claires très ouvertes (12 à 24% pour
UPP5, UPP6 et UPw1). Dans les unités au recouvrement total relativement
faible (UPP6, UPP4, UPP5 et UPP3), on peut

100
observer un développement assez important de la strate
sous-arbustive qui n'est cependant jamais dominante. On peut
l'interpréter comme un début d'embuissonnement qui reste
cependant discret.
Tableau IV-12. Valeurs (moyenne #177; écart-type) de la
densité et du recouvrement ligneux dans les différentes
unités paysagères du terroir de Kotchari
|
Unités
paysagères
pastorales
|
Densité moyenne
(pieds.ha-1)
|
Recouvrement moyen (%)
|
|
UPP1
|
9917,40 #177; 524,85abc
|
52,52 #177; 3,27bc
|
|
UPP2
|
10984,75 #177; 385,18bc
|
46,55 #177; 4,12abc
|
|
UPP3
|
11920,33 #177; 125,52bc
|
39,90 #177; 2,29 abc
|
|
UPP4
|
12977,17 #177; 695,91c
|
37,73 #177; 2,47ab
|
|
UPP5
|
5441,33 #177; 172,66ab
|
14,37 #177; 1,85a
|
|
UPP6
|
4993,33 #177; 22,50a
|
12,03 #177; 2,06a
|
|
UPw3
|
6935,83 #177; 263,59ab
|
61,13 #177; 3,09 c
|
|
UPw4
|
8598,99 #177; 808,28abc
|
42,93 #177; 5,29abc
|
|
UPw1
|
7246,80 #177; 248,81ab
|
23,50 #177; 4,36ab
|
Sur la même colonne, les valeurs portant les mêmes
lettres ne sont pas significativement différentes au seuil de 5%.
Tableau IV-13. Proportions des types biologiques ligneux dans les
unités paysagères pastorales du terroir de Kotchari (spectres
pondérés des abondances)
|
Types
biologiques
|
|
|
Unités paysagères pastorales
(%)
|
|
|
|
UPP1
|
UPP2
|
UPP3
|
UPP4
|
UPP5
|
UPP6
|
UPw3
|
UPw4
|
UPw1
|
|
Arbre
Arbuste
Sous-arbuste
|
61,54c
32,36b
6,10a
|
50,60b
45,81b
3,58a
|
36,97b
44,64c
18,40a
|
24,22a
56,11b
19,65a
|
21,27a
61,22b
19,51a
|
19,13a
60,48b
20,38a
|
57,61c
37,69b
4,69a
|
36,74b
53,75c
9,51a
|
26,26b
66,60c
7,14a
|
Sur la même colonne, les valeurs portant les mêmes
lettres ne sont pas significativement différentes au seuil de 5%.
On peut expliquer cette structuration différente de la
végétation entre les deux entités du terroir
(intérieur et extérieur de l'aire protégée),
notamment sa densité, par l'action éventuelle des feux de
végétation. On sait en effet que ceux-ci se nourrissent de la
biomasse herbacée, en conséquence ils seront plus violents dans
les unités qui en produisent plus, c'està-dire les unités
du parc W. Nous avons vu par ailleurs qu'une pratique instaurée par les
forestiers durant l'exécution du programme ECOPAS (2001 - 2008),
était de mettre le feu au parc dès l'arrêt des pluies (feux
précoces). La conjugaison de ces deux facteurs ne favorise donc pas la
densification du parc étant entendu que le feu, en particulier le type
tardif, agit en

empêchant la colonisation des milieux par la
végétation ligneuse (Schnell, 1971; Monnier, 1981; Beani &
Dessi, 1984 ; Fournier, 1991 ; César, 1991 & 1994 ; Bruzon, 1995 ;
Western & Maitumo, 2004 ; Bond & Keeley ; 2005 ; Lavorel et
al. 2007). Les plus faibles densités rencontrées sur les
sites les plus anthropisés sont dues probablement à ce facteur
notamment les prélèvements humains et les défrichements
agricoles.

Cliché Sawadogo, juin 2008
Photo IV-1 . Un faciès de savane arborée de sol
inondable (UPP2): La couverture ligneuse sur ce faciès est fait
essentiellement de Terminalia macroptera.

Cliché Sawadogo, août 2009
Photo IV-2. Un faciès de savane arbustive de bas glacis
et de plaine sur sols argilo-limoneuse (UPP3). Au premier plan, des jeunes
rejets de Piliostigma reticulatum. En arrière plan, un
agroforêt fait de Vitellaria paradoxa et de Sorgum
vulgare (sorgho).

Cliché Sawadogo, août 2009

102
Photo IV-3. Un faciès de savane arbustive de hauts
glacis et plateaux sur sols moyennement profonds (UPP4) : On peut remarquer la
forte emprise animale à travers le port buissonnant des individus
ligneux et les différentes pistes à bétail
disséminées entre ceux-ci.

Cliché Sawadogo, août 2009
Photo IV-4. Un faciès de mosaïque
agroforestière (UPP5) : au premier plan, des pieds de Vitellaria
paradoxa dans un champ de Vigna inguiculata
(niébé). Au fond, une jachère d'âge
avancé.

Cliché Sawadogo, août 2009

Photo IV-5. Un faciès de savane arbustive sur buttes
rocheuses et cuirassées (UPP6). On peut observer au premier plan des
espèces comme Guiera senegalensis, Combretum nigricans et
Loudetia togoensis au premier plan ; au second plan, on note un pied de
Sclerocarya birea entouré de touffes de Combretum
micranthum.

Cliché Sawadogo, août 2009
Photo IV-6. Un faciès de savane arbustive à
herbacées annuelles à sol encroûté de l'aire
protégée (UPw4). On note une discontinuité de la strate
herbacée (en arrière plan, un fourré dans lequel on
distinge Acacia macrostachya).

Cliché Sawadogo, août 2009

104
Photo IV-7 : Un faciès de savane arbustive sur
affleurements rocheux et cuirassés de l'aire protégée
(UPw1) : ici un bosquet à Detarium microcarpum.
4.3.3.5. La valeur pastorale des unités
paysagères
Les valeurs pastorales les meilleures se rencontrent dans
l'aire protégée (63% à 80%), surtout dans les savanes
arborées à pérennes (UPw3) (tableau IV14). Les
espèces fourragères de bonne à très bonne valeur
nutritive (Is = 3 et 4) y sont en effet bien représentées (30% +
8% = 38% pour UPw1 et 28% + 15% = 43% pour UPw3).
En dehors de l'aire protégée, la proportion
d'espèces sans valeur ou à valeur faible (IS = 0 et 1) est en
revanche très forte (28% + 16% = 44% pour UPP1 et 30% + 30% = 60% pour
UPP6) ce qui entraîne de faibles valeurs pastorales. Les valeurs
pastorales les meilleures sont enregistrées sur les sols peu humides
à secs (UPP3, UPP4, UPP5) si l'on en excepte les buttes et cuirasses
(UPP6). Des constatations similaires ont été faites en Côte
d'Ivoire par César (1992) et au Burkina Faso par Kagoné (2000) au
Centre-sud et par Kièma S. (2007) dans l'Ouest. A l'opposé, les
valeurs pastorales les plus faibles s'observent sur les sols profonds, en
particulier dans les plaines hydromorphes (UPP2).
On comprend comment les activités pastorales et
agricoles en dehors de l'aire protégée induisent une baisse de la
valeur pastorale des parcours, car celle-ci dépend de la phytomasse
herbacée et de la qualité de l'herbe offerte que l'on
évalue à l'aide de l'indice de qualité spécifique
(Daget & Godron, 1995). Dans l'aire protégée où les
graminées cespiteuses bonnes fourragères dominent, la valeur
pastorale est élevée, notamment dans l'unité de savane
arborée et boisée à pérennes (UPw3), la phytomasse
y est également la plus forte. En dehors de l'aire
protégée les meilleures valeurs pastorales sont
enregistrées dans les unités les plus pâturées (UPP3
et UPP4) exception faite des buttes et cuirasses (UPP6). Ces unités,
appauvries en espèces graminéennes vivaces à très
forte qualité fourragère, se sont enrichies, à des
degrés divers, en phorbes à bonne valeur fourragère,
notamment des légumineuses (Fabaceae en particulier). La faible valeur
pastorale de l'unité de butte et cuirasse (UPP6) est à relier non
seulement à la faible productivité de ce milieu où le sol
est squelettique, mais

aussi à la forte représentation des
espèces de qualité nulle à très faible. La relative
bonne valeur pastorale de la mosaïque agroforestière (UPP5) est
tributaire d'une bonne contribution d'espèces à moyenne ou bonne
valeur (essentiellement des graminées de jeunes jachères). Par
ailleurs, dans les milieux à statut édaphique et hydrique plus
favorable de cette partie du terroir (UPP1 et UPP2) la valeur pastorale est
faible à cause de l'importance d'espèces peu
appétées.
Tableau IV-14. Qualité du fourrage dans les
unités paysagères du terroir de Kotchari : contribution des
espèces par classes d'indice spécifique et valeurs pastorales
(moyenne #177; écart-type)
|
Classes
d'indice
spécifique
(Is)
|
|
|
|
Unités paysagères pastorales
|
|
|
|
|
UPP1
|
UPP2
|
UPP3
|
UPP4
|
UPP5
|
UPP6
|
UPw3
|
UPw4
|
UPw1
|
|
0
1
2
3
4
|
0,28 ab 0,16 ab 0,31 b 0,20
ab 0,06 b
|
0,36 c 0,18 ab 0,20 a 0,22
ab 0,04 ab
|
0,31 ab 0,27 b 0,21 a 0,16
a 0,04 ab
|
0,37 c 0,17 ab 0,26 ab 0,20
a 0,01 a
|
0,31 b 0,24 ab 0,24 ab 0,19
a 0,02 a
|
0,30 b 0,30 b 0,21 a 0,18
a 0,02 a
|
0,25 ab 0,09 a 0,22 a 0,28
b 0,15 c
|
0,17 ab 0,17 ab 0,24 ab
0,32 c 0,10 b
|
0,07a 0,20 ab 0,35 b 0,30 c
0,08 b
|
|
Valeur
|
49,37
|
39,05
|
53,57
|
53,99
|
57,628
|
44,46
|
79,88
|
68,16
|
63,05
|
|
pastorale
|
#177;
|
#177;
|
#177;
|
#177;
|
#177;
|
#177;
|
#177;
|
#177;
|
#177;
|
|
brute (%)
|
4,99ab
|
5,26a
|
3,27abc
|
5,99abc
|
1,861ab
|
3,26ab
|
5,31c
|
9,67bc
|
2,29bc
|
Sur la même ligne, les valeurs portant les mêmes
lettres ne sont pas significativement différentes au seuil de 5%.
Ainsi, la valeur pastorale particulièrement faible
observée dans la plaine inondable (UPP2) est due à la faible
proportion des espèces de bonne à très bonne
qualité (Is = 3 : 20% ; Is = 4 : 6% ; total : 26%) ; les espèces
à indice spécifique nul ou faible, notamment les
Cypéracées comme Kyllinga
spp. et Abildgaardia
hispidula, sont en revanche bien représentées (Is = 0 : 36%
; Is =1 : 18% ; total 54%). La plupart des auteurs, obtiennent cependant les
meilleures valeurs pastorales dans ces types d'unités (Kagoné,
2000 ; Yanra, 2004 ; Bambara, 2010), mais ces auteurs ont noté,
contrairement à nous, une bonne représentation de
graminées pérennes bonnes fourragères dans ces sites. Dans
le terroir de Kotchari, les graminées vivaces ne sont plus bien
représentées que dans l'aire protégée notamment
dans les savanes arborées et boisées (UPw3).
Par ailleurs, pour qu'un pâturage soit
considéré de bonne qualité il faut que sa valeur pastorale
atteigne au moins 65% (Daget & Godron, 1995) ; seules les savanes
arborées et boisée à herbacées pérennes
(UPw3) et les savanes arbustives à herbacées annuelles (UPw4)
méritent ce qualificatif. Comme le rappellent cependant ces auteurs, la
valeur des pâturages évaluée par enquête est toujours
contextuelle, et, comme telle, fortement dépendante des
représentations de l'enquêté liées au contexte
environnemental local. Par exemple, Andropogon gayanus dont l'indice
de qualité spécifique a été fixé à 4,
a obtenu une meilleure appréciation, de valeur 5, dans une étude
conduite ailleurs par César (2005).

106
4.3.4. Appréciation globale des parcours de
Kotchari
Une appréciation globale des différentes
unités pastorales cartographiées peut être
faite en prenant en compte leur gestion, leur
accessibilité, leur phytomasse, la valeur pastorale de leur fourrage et,
pour les unités situées hors parc, leur étendue. Les
unités sont abordées suivant l'ordre décroissant
d'étendue.
- Les unités de savane arbustives (mosaïques
agroforestières non comprises) à l'extérieur de l'aire
protégée, localisées à mi-pente (plateaux et
glacis) (UPP3 & UPP4) et constituant ensemble une entité vaste de
133,54 km2, sont utilisables toute l'année par le
bétail. La phytomasse y est moyenne à très faible, mais de
qualité relativement intéressante. Elles sont fragmentées
dans l'espace à cause de l'exploitation agricole et connaissent un
début d'embuissonnement, ce qui rend leur utilisation pas toujours
rentable en saison sèche et risquée en saison pluvieuse à
cause des risques de dégâts champêtres et des conflits qui
en résultent. Par ailleurs, avec les buttes rocheuses et
cuirassées, ce sont les unités les plus touchées par les
feux de brousse, malheureusement leur couverture herbacée étant
composée essentiellement d'annuelles elles ne sont pas capables de
produire de repousses intéressantes;
- L'unité de savane arborée de bas-fonds en
dehors du parc W (UPP1), d'une superficie de 80,10 km2, produit une
phytomasse notable de qualité moyenne, mais elle n'est exploitable
qu'après les récoltes en début de saison sèche.
Cette unité constitue par ailleurs, un refuge pour le bétail en
période de soudure (saison sèche chaude jusqu'aux
premières pluies) à cause de sa relative fraîcheur, des
repousses et des possibilités qu'elle offre d'y creuser des puisards
;
- L'unité de savane arbustive des buttes rocheuses et
cuirassées (UPP6), assez vaste (65 km2) et surtout peu
fragmentée est intéressante en terme pastoral notamment en saison
pluvieuse bien que la phytomasse offerte y soit très faible et
nutritivement la plus médiocre du terroir. Peu convoitée par
l'agriculture, elle constitue la zone de replis de l'essentiel du bétail
villageois en saison pluvieuse, mais est malheureusement, pour le
bétail, l'une des plus sujettes au passage des feux (perte de paille non
compensée par des repousses) ;
- La mosaïque agroforestière ou savane-parc
(UPP5), vaste de 42,73 km2, est la moins intéressante
à cause de la faiblesse de la phytomasse naturelle qui y est produite et
surtout parce que les cultures la rendent inaccessible en saison des pluies.
Ces dernières deviennent cependant très utiles pour le
bétail dès la fin des récoltes grâce aux
résidus de culture exploitables en vaine pâture ;
- L'unité de savane boisée claire des plaines
inondables (UPP2) est la moins étendue du terroir (29 km2),
sa phytomasse est importante, mais de faible qualité. Il s'agit d'un
pâturage de réserve pour la saison sèche car elle est
inaccessible pendant les pluies ; certaines années, les feux accidentels
ou provoqués font cependant partir cette réserve en fumée
et les repousses y sont négligeables ;
- Les unités de savanes arborée et boisée
et celles de savane arbustive de l'aire protégée (respectivement
UPw3 et UPw4), assez vastes, offrent la plus importante phytomasse et la

meilleure valeur alimentaire grâce à la dominance
d'herbacées pérennes. Dans ces milieux, la pratique des feux
précoces à partir du mois d'octobre permet de produire
d'importantes repousses de bonne qualité qui sont susceptibles d'attirer
le bétail pendant la saison sèche froide (de novembre à
janvier) surtout dans l'unité (UPw3) ;
- L'unité d'affleurements rocheux et cuirassés
de l'aire protégée (UPw1) ne fournit qu'une faible phytomasse
herbacée de qualité moyenne, et celle-ci est vite
éliminée par une mise à feu précoce qui, vu la
pauvreté en herbes pérennes, n'est pas suivie d'une repousse
notable.
4.4. Conclusion
Dans la présente étude, les ressources
pastorales du terroir de Kotchari ont été
caractérisées. Les unités paysagères pastorales
définies à l'intérieur et à l'extérieur de
l'aire protégée sont pour l'essentiel des savanes arbustives plus
ou moins denses et des savanes arborées denses à boisées
claires. Les pâturages accessibles ne semblent pas y être trop
dégradés comme en témoignent le faible embuissonnement,
même dans les zones les plus pâturées, et la richesse
floristique plus élevée que celle de l'aire
protégée due essentiellement aux cultures (effet
jachères).
Si dans le Parc ces unités pastorales produisent une
importante phytomasse de bonne qualité, hors de cette aire
protégée la quantité fait défaut quand la
qualité est présente. De plus, les plaines inondables et, dans
une moindre mesure, les mosaïques agroforestières, largement
représentées dans le terroir sont inexploitables par le
bétail en saison des pluies. Ainsi, il faut constater que le terroir de
Kotchari n'apparait pas comme particulièrement intéressant du
point de vue pastoral si l'on s'en tient à la qualité du fourrage
disponible. Il présente pourtant une attraction particulièrement
forte de la part des transhumants. De plus c'est en avril-mai, au moment
où l'offre fourragère, alors faite de paille sèche, est la
plus médiocre en quantité comme en qualité que la campagne
de transhumance connaît son pic. Cette situation peut s'expliquer par le
fait que, relativement aux terroirs d'attache de ces pasteurs, situés
tous en zone sahélienne et subsahélienne, le terroir de Kotchari
reste attractif. Zouri (2003) évalue, en effet, la phytomasse du
département de Botou, une des zones de provenance des transhumants,
entre 1,87 et 2,8 tMS/ha. De façon générale, on estime que
la productivité des parcours sahéliens se situe dans la
fourchette de 0,5 tMS/ha et 3 tMS/ha (Boudet, 1975). Ces différentes
valeurs sont largement inférieures à celles enregistrées
sur le terroir de Kotchari (1,41 à 4,78 tMS/ha). Cette
attractivité peut aussi se comprendre par le fait d'une plus grande
disponibilité en fourrage en cette période, le terroir
étant en fin de transhumance. On peut, enfin supposer que la position de
terroir contigu au parc est un facteur supplémentaire
d'attractivité, les transhumants ayant aussi pour motivation de pouvoir
accéder aux ressources fourragères de l'aire
protégée (Kaboré, 2010) dont le statut pastoral est bien
meilleur. Toutefois, pour aller plus loin dans les investigations sur le sujet
il est intéressant de faire le bilan fourrager de la période de
l'étude, c'est-à-dire mettre en parallèle les
capacités de charge ici déterminées - desquelles il est
facile de rechercher la charge globale ou théorique - et les charges
réelles. Cette option, qui permet de mieux renseigner la question, sera
abordée dans le chapitre V suivant.
CHAPITRE V

108
LES SYSTEMES D'ELEVAGE A KOTCHARI :
PRATIQUES ET STRATEGIES D'HIER ET
D'AUJOURD'HUI

109
5 .1. Introduction
Le terroir de Kotchari est marqué par le
phénomène de la transhumance, un flux migratoire devenu habituel
et dont les origines sont variées et, pour certaines, lointaines. Toutes
les études sur le sujet (Paris, 2002 ; Kagoné, 2004 ; Bambara,
2010 et Kpoda, 2010) indiquent que la plupart des grands et petits transhumants
venant des régions plus au nord choisissent de transiter par ce terroir
lors de leur périple ou d'y passer leur saison de transhumance. Ce
processus qui touche l'ensemble de la zone si l'on en croit les travaux de
Benoit (1998 & 1999a) et Santoir (1998 & 1999), est susceptible de
bouleverser les systèmes de vie locaux. Santoir avait notamment
observé que suite à un long processus d'immigration peule
d'origine lointaine - entre le 16ème et le
19ème siècle - le Gourma (actuelle région de
l'Est du Burkina Faso) a vu accroître sa population de manière
considérable. Cet accroissement a été mal partagé
et plus sensible dans les parties sud (région de Pama) et sudest (autour
du Gobnangou dans l'actuelle province de la Tapoa). Des études plus
récentes (Doussa, 2004 & 2006 ; Guibert & Prudent, 2005) et les
données actualisées du recensement général de la
population et de l'habitat (RGPH) de 2006 (paragraphe 3.3., tableau III-2)
indiquent un accroissement général de la population de la
province de la Tapoa. Cet accroissement notamment pendant les premières
années du retour du coton dans la zone, a surtout été le
fait de migrants agricoles qui recherchaient des terres vacantes ou de grands
espaces pour la production cotonnière. À cela, d'après
Doussa (2004) il faut ajouter la baisse notable de l'émigration agricole
en direction du Bénin. Par ailleurs, avec l'attrait des agriculteurs
gourmantchés pour l'élevage (Sawadogo, 2004 ; Kaboré,
2010) et les flux importants d'éleveurs transhumants52 dont
certains se sédentarisent (Paris, 2002), les effectifs des hommes ainsi
que ceux du cheptel se sont notablement accrus. Ce phénomène
d'accroissement rapide des populations humaine et animale, observé dans
l'ensemble de la commune de Tansarga, semble accentué surtout dans les
terroirs contigus du parc W, comme Kabougou et Kotchari. Il peut engendrer de
nouvelles pratiques et provoquer des mutations ou renforcer celles
déjà en cours dans les systèmes de production locaux. Dans
une étude des pratiques et des ajustements adaptatifs des acteurs, il
est important d'identifier les contraintes notamment spatiales auxquelles ils
se confrontent.
Rappel des hypothèses de recherche
Face à la pression démographique qui semble se
faire de plus en plus forte et aux bouleversements répétés
(mutations socio-économiques en milieu rural, raréfaction et
dégradation des parcours, changements climatiques, etc.), nous avons
émis trois hypothèses rappelées dans l'encadré V-1.
Celles-ci postulent que, pour tirer le meilleur profit des ressources
pastorales et/ou pour s'adapter, les différentes catégories
d'éleveurs mettent en jeu des stratégies plus ou moins fines et
plus ou moins efficaces qui se fondent sur une perception aigüe des
caractéristiques et conditions du milieu.
52 45,7% des transhumants qui ont
fréquenté la Tapoa entre 2001 et 2002 ont séjourné
à Kotchari (Paris, 2002).

Encadré V-1. Les hypothèses de recherche
· L'afflux de nombreux troupeaux transhumants
observé chaque année sur le terroir de Kotchari et ses environs
ne s'explique pas, comme il est souvent dit, par la disponibilité en
ressources pastorales dans les espaces légalement accessibles
(périphérie du Parc). L'intérêt pastoral du terroir
de Kotchari est plutôt lié à l'opportunité qu'il
peut offrir d'accéder illégalement aux ressources du Parc du
W.
· Les systèmes d'élevage
sédentaires ou mobiles subissent des mutations perceptibles au travers
des pratiques des éleveurs qui se modifient pour
s'adapter aux nouvelles conditions.
5 .2. Matériel et méthodes
5.2.1. Méthodes d'acquisition des
données
Ce volet de la recherche a été conduit en
combinant plusieurs activités : une exploitation continue de
données bibliographiques, des recensements de cheptel et des
enquêtes prenant la forme d'entretiens directifs ou semi directifs ou de
questionnaire plus formels. Des résultats de travaux
préliminaires (Sawadogo, 2004) ont servi de base au présent
travail. Ce travail précédent portait sur les aspects
relationnels ou de conflictualité (entre agropasteurs résidents,
entre agropasteurs et transhumants, entre riverains et gestions des
réserves, entre éleveurs et administration, etc.).
5.2.1 .1. La revue des données
Pour faire le point sur la situation démographique
humaine et animale actuelle et en prendre la pleine mesure, diverses
données (recensement général de la population,
enquêtes agricoles, enquête nationale sur les effectifs du cheptel,
rapports d'étude, etc.) ont été exploitées puis
complétées par des enquêtes légères de
terrain, surtout ciblées sur le cheptel transhumant. Toutes ces
données ont permis d'avoir une idée relativement exacte de la
densité de la population du terroir, de celle des animaux
résidents et de ceux transhumants.
Pour la détermination des effectifs humains, nous nous
sommes appuyé sur les statistiques nationales (MEF, 2008 ; DREP-Est,
2008) auxquelles nous avons appliqué les taux officiels de
croît53. Par ailleurs, l'appréciation du niveau
d'occupation des sols a été faite en s'appuyant sur les
données de la base des données d'occupation des terres (BDOT) de
l'IGB, réalisée en 2002.
53 Taux de croît annuel : Botou : 3,8% ; Diapaga
: 4,1% ; Kantchari : 3,4% ; Logobou : 3,8% ; Namounou : 2,5% ; Partiaga : 4,4%
; Tambaga : 3,6% ; Tansarga : 4,6% ; Tapoa : 3,8% et Région-Est :
3,6%.

111
5 .2.2.2. Les enquêtes
Les enquêtes ont été conduites en trois
temps : une phase d'enquête exploratoire semi-directive en 2007, une
phase d'estimation des effectifs du cheptel en 2008 et 2009, et une phase
d'enquête par questionnaire en 2009.
5.2.2.2.1. Les entretiens
Ils ont été conduits de manière
semi-directive (avec guide d'entretien) et itérative tout au long de la
recherche de terrain auprès de personnes ressources d'expérience,
notamment les éleveurs possédant des troupeaux et les
Garso (Encadré V-2).
Encadré V-2. Le Garso ou guide de transhumance
La société pastorale dans la région du
parc W est organisée autour de personnages clés que sont les
Rugga et les Garso (Riegel, 2002) considérés
comme guides et porte-paroles (Benoit, 1999a)54. Le Rugga,
fait office de chef de clan. Il est chargé de la sécurité
des membres de sa communauté et de leurs animaux (Oumarou, 2004). Le
Garso est un éleveur reconnu par la communauté pour son
expérience et son savoir en matière de pâturages et de
conduite du troupeau ainsi que pour sa sociabilité et ses
qualités de médiateur (Riegel, 2002 ; Bodé, 2008). Il est
à la tête d'un groupe de transhumants (dont le nombre peut varier
de cinq à douze) qui l'élit et dont il est le porte-parole
auprès du Rugga. La réussite de la campagne de
transhumance est de sa responsabilité. C'est en général
lui qui détermine l'axe de transhumance à suivre et qui assure la
coordination et la gestion des relations avec les autres groupes
(administration et autochtones des villages de transit et d'accueil).
5.2.2.2.2. L'estimation des effectifs animaux dans le
terroir
Un inventaire du cheptel local a d'abord été
fait en 2008 par recensement exhaustif. Il a été
réactualisé en 2009 par application des taux de croît
observés en suivant cinq troupeaux de chacune des trois espèces
principales (bovins, ovins, caprins) pendant trois ans (2006- 2009). Les taux
de croît résultant de ces suivis valaient 2,2%; 2,9% et 3,3%
respectivement pour les bovins, les ovins et les caprins.
Chez les éleveurs transhumants ce type de
méthode n'était pas possible, la démarche a donc
été toute autre. Pour des troupeaux transhumants la
détermination des effectifs est toujours un exercice délicat.
Plusieurs méthodes ont été utilisées pour s'assurer
de recenser, sinon la totalité, du moins le maximum de troupeaux
transhumants. Quatre observateurs55 positionnés
quotidiennement de 6 h à 18 h aux entrées principales du terroir,
de début novembre 2007 à fin juillet 2008 puis de novembre 2008
à fin juillet 2009, se sont chargés de cette tâche. Les
données collectées ont ensuite été
agrégées pour les quatre périodes définies par
Paris (2002) par rapport au flux de transhumance à Kotchari :
début de la transhumance de décembre à mars, pleine
transhumance d'avril à mai, retour des transhumants de juin à
juillet,
54 Dans l'extrême- nord du Cameroun (sud du
bassin du lac Tchad), on rencontre plutôt le Kaydal, chef des
bergers de plusieurs villages allant en transhumance et l'Ardo, chef
de bergers au sein des campements en déplacement.
(Requier-Desjardins, 1999).
55 La surveillance étant continue, les
personnes sont choisies parmi celles qui habitent sur place.
période sans transhumants d'août à
novembre. En vue de compléter leurs informations, les mêmes
observateurs effectuaient par ailleurs des visites dans les campements de nuit
et de repos de ces éleveurs, ce qui est recommandé par Daget
& Godron (1995). Les effectifs animaux des troupeaux d'éleveurs
résidents qui partent en transhumance ont été notés
et soustraits de l'effectif total de chaque saison. Il a ainsi
été possible de se faire une idée assez précise des
effectifs animaux et des niveaux de charge ainsi que de leur évolution
au cours de l'année.
5.2.2.2.3. La typologie des systèmes
d'élevage dans le terroir
Une esquisse de typologie déjà
réalisée sur le même terrain (Sawadogo, 2004) a servi de
base pour le présent travail. Comme nous avions postulé que des
groupes différents d'éleveurs avaient des façons de faire
différentes, une typologie assez précise des éleveurs
s'imposait comme un préalable à l'étude des pratiques. Une
enquête par questionnaire a été conduite tout au long de
l'année 2009 auprès de 200 éleveurs, ce qui
représente un peu plus de 10% de la population totale estimée.
L'échantillon enquêté a été
construit de la manière suivante : dans un premier temps l'ethnie a
servi de base pour identifier deux groupes : 50 éleveurs
gourmantchés et 150 éleveurs peuls. Le nombre
d'enquêtés par quartier (ou village-satellite) a ensuite
été déterminé au prorata de leurs poids
démographiques. Enfin, le choix des chefs de ménages
enquêtés s'est fait au hasard des rencontres.
L'effort d'échantillonnage a été a priori
orienté surtout vers les Peuls, en raison de la plus grande implication
de ce groupe socioculturel dans les activités pastorales, alors qu'il
est démographiquement peu représenté. L'échantillon
ainsi constitué est un compromis entre une répartition a priori
et un tirage aléatoire, une méthode qui a montré son
caractère reproductible (Scherrer, 2007).
Pour construire le questionnaire, un certain nombre de
variables jugées pertinentes (variables d'intérêt
d'après Cibois, 2007) au vu des enquêtes exploratoires, ont
été retenues, leurs modalités ont été
définies et codées. Les thèmes abordés dans
l'enquête sur les systèmes d'élevage et des pratiques
pastorales portent sur l'ethnie et l'origine de l'éleveur, la taille la
composition et le mode de constitution du troupeau, son type de conduite, sa
mobilité, son alimentation et son agrégation (tableau V-1):
Tableau V-1. Variables et modalités constitutifs du
questionnaire
|
Variables Codes Modalités Intervalles ou contenu
des
classes correspondantes
|
Ethnie de l'éleveur Eth 1 Gourmantché
2 Peul
Lieu de résidence habituelle ou origine OrEl 1 Kotchari
de l'éleveur 2 Dans la Tapoa
3 Burkina Faso hors Tapoa
4 Non burkinabè
Effectif bovin du TaiTr 1 [0 45]
troupeau 2 [46 90]

113
|
|
Chapitre V. Les systèmes d'élevage
à Kotchari3 [91 135]
4 [136 180]
5 Plus de 180 têtes de bovins
|
|
Nombre et types d'espèces animales dans
|
NEsp
|
1
|
Bovins + Ovins + Caprins
|
|
le troupeau
|
|
2
|
Bovins + Ovins
|
|
|
3
|
Bovins + Caprins
|
|
|
4
|
Bovins
|
|
|
5
|
Pas de bovins
|
|
Nombre et types de races bovines du
|
NRaB
|
1
|
Barbadji
|
|
troupeau
|
|
2
|
Gurmaji
|
|
|
3
|
Jaliji
|
|
|
4
|
Barbadji + Gurmaji
|
|
|
5
|
Gurmaji + Jaliji
|
|
|
6
|
Autres (plus de 2 races bovines)
|
|
|
7
|
Aucun (Pas de bovins dans le troupeau)
|
|
Mode de constitution ou origine du
|
OrTr
|
1
|
Achat + emprunt
|
|
troupeau
|
|
2
|
Héritage + don
|
|
|
3
|
Achat + Héritage
|
|
|
4
|
Autres (plus de 3 formes d'acquisition)
|
|
Lieu de pâturage en saison sèche
|
LPat
|
1
|
Terroir Kotchari
|
|
|
2
|
Kotchari + terroirs voisins
|
|
|
3
|
Pays voisins: Bénin / Togo
|
|
|
4
|
Réserves voisines
|
|
Raisons de la pratique de la
|
RTra
|
1
|
Eau
|
|
transhumance
|
|
2
|
Fourrages
|
|
|
3
|
Eau+fourrage
|
|
|
4
|
Habitude
|
|
|
5
|
Ne transhume pas
|
|
Type de berger utilisé
|
NatBe
|
1
|
propriétaire ou parent
|
|
|
2
|
Salarié
|
|
|
3
|
Mixte (parent + salarié)
|
|
Nombre de bergers
|
NBe
|
1
|
Un berger
|
|
|
2
|
Deux bergers
|
|
|
3
|
Plus de deux bergers
|
|
Allotement du troupeau
|
NLot
|
1
|
Un lot
|
|
|
2
|
Deux lots
|
|
|
3
|
Plus de deux lots
|
|
Raisons dans l'allotement du troupeau
|
RLot
|
1
|
Former lots homogènes (séparer bien portants des
fatigués, malades, etc.)
|
|
|
2
|
Gérer les risques
|
|
|
3
|
Alléger les charges sur les parcours
|
|
|
4
|
Faciliter la tâche des bergers
|
|
|
5
|
Ne pratique pas
|
|
Pratiques d'alimentation du troupeau
|
PAlt
|
1
|
Pâturage naturel
|
|
|
2
|
Complément par fourrage naturel
|
|
|
3
|
Complément par fourrage cultivé
|
|
|
4
|
Complément par concentré acheté
|
|
|
5
|
Complément cultivé, collecté et
acheté
|

114
(fourrage et concentré)
Pour harmoniser et faciliter le traitement des données,
la variable «effectif de bovins dans le troupeau» a
été transformée en variable discrète. Nous avons
ainsi choisi de répartir les troupeaux dans cinq classes d'amplitude
égale (Beguin & Pumain, 1994), leur étendue (h) ayant ensuite
été définie en faisant le rapport entre l'amplitude de la
variation (écart entre l'effectif de troupeau le plus
élevé et le plus faible) et le nombre k des classes.
h max- min
=
k
L'étendue ainsi calculée était de 45
têtes. Dans notre série statistique, le plus petit effectif bovin
était de 0 alors que l'effectif le plus important était de 227
têtes.
5.2.2.2.4. Les pratiques et leur
évolution
L'étude des pratiques et de l'évolution permet
d'identifier les ajustements même minimes qu'opèrent les
éleveurs pour en déduire les stratégies qu'ils mettent en
oeuvre pour s'adapter aux changements climatiques et environnementaux. On
suivra en effet Landais & Deffontaines (1989) qui pensent que les
stratégies des éleveurs - leur nature, leurs logiques et la
manière dont elles sont mises en oeuvre peuvent être mises en
évidence au travers des pratiques que ceux-ci adoptent dans la conduite
de leurs activités pastorales.
Une période de référence (ou pas de
temps) de 20 ans a été retenue pour suivre cette évolution
dans les pratiques. Nous sommes conscient que pour juger des variations qui
s'expriment dans les pratiques, cette période est courte. Deux arguments
permettent cependant de justifier notre choix. Le premier est le souci de lier
le phénomène étudié aux grands bouleversements
fonciers qui ont accompagné la relance de la cotonculture dans la zone
d'étude. De ce point de vue, la décennie 90 (il y a vingt ans
environ) correspond à la période où la culture du coton
était à son niveau le plus bas dans le terroir, tandis que la fin
de cette décennie (il y a dix ans environ) coïncide avec le
début du boom cotonnier et du grand engouement des populations pour
cette culture qui a été suivi d'une importante occupation de
l'espace. Rappelons que cette culture très consommatrice d'espace
(Tamou, 2002 ; Doussa, 2004) a été encouragée par les
sociétés cotonnières (la SOFITEX56 puis la
SOCOMA57) qui ont facilité l'accès aux moyens de
travail et aux intrants (semences, engrais, pesticides et insecticides). Le
deuxième argument est que notre questionnaire d'enquête devait
permettre de recueillir des informations chiffrées aussi exactes que
possibles. De ce point de vue, il n'aurait pas été prudent de
chercher à remonter beaucoup plus loin dans le passé.
Dans l'étude des pratiques pastorales nous nous sommes
concentré surtout sur pratiques l'agrégation (constitution,
allotement), la conduite (gardiennage, mobilité, alimentation) et la
diversification du troupeau. Ces enquêtes structurées ont, par
ailleurs, été complétées par des entretiens
ciblés conduits auprès d'éleveurs choisis dans chacun des
groupes de notre typologie. Ces éleveurs ont été
visités régulièrement et la technique d'analyse du
discours
56 Société des Fibres Textiles
57 Société Cotonnière du
Gourma

(Mettrick, 1994) a été
privilégiée. Celle-ci, basée sur des entretiens non
directifs ou semi directifs, permet d'expliciter les ajustements que les
éleveurs opèrent dans leurs pratiques et de comprendre les
logiques qui les sous-tendent (Blanchet & Gotman, 2003). Elle permet,
in fine, à travers les questions que les acteurs
eux-mêmes se posent, de faire appel à leurs points de vue et de
donner à leurs expériences vécues, à leurs
logiques, à leur rationalité, une place de premier plan.
Dans ce chapitre, nous considérons les pratiques dans
leur globalité et leur évolution. Dans le chapitre suivant nous
reviendrons plus en détails sur les pratiques de conduite au
pâturage.
5 .2.2.3. Analyse des données : description et
tests de comparaison
Les données d'enquêtes ont été
saisies, organisées et décrites (moyenne, écart-types,
graphiques) dans Excel 2007.
La carte d'occupation des terres a été
réalisée à partir du logiciel MapInfo version 8.1
Les effectifs du cheptel ont été convertis en UBT
(Unité de Bétail Tropical, équivalent à un bovin de
référence de 250 kg de poids) sur la base de :
1 bovin = 1 équin = 1 UBT ; 1 ovin adulte = 1 caprin
adulte = 0,2 UBT ; 1 asin = 0,4 UBT (Ministère de la Coopération
et du Développement, 1991)).
Dans un premier temps, un test de corrélation entre les
variables a été effectué. Il a permis de déterminer
celles qui étaient significativement (p = 0,5) corrélées
entre elles et de choisir les plus pertinentes en vue de réaliser la
typologie des éleveurs. Une analyse factorielle de correspondances
multiples (AFCM) et une classification k-means utilisant les facteurs ou axes
à plus forte valeur d'inertie de l'AFCM ont été ensuite
réalisées. Finalement, la population d'éleveurs
enquêtés a été classée par analyse ascendante
hiérarchique (ACH) à partir des barycentres des classes issues de
l'analyse k-means. Rappelons que l'analyse factorielle de correspondance
multiple est bien adaptée au traitement d'une série statistique
multiple (Scherrer, 2007) lorsque les variables sont qualitatives comme c'est
le cas ici. C'est une méthode d'analyse multivariée qui vise
à résumer l'information d'un tableau de données en
écriture simplifiée sous forme graphique (Diatta et al.
1998). L'interprétation des résultats qui en résultent
repose sur l'examen de trois paramètres (tableau V-2).
Tableau V-2. Les paramètres significatifs dans
l'interprétation des résultats de l'AFCM
Paramètres Signification
L'inertie Quantifie l'information contenue dans
chaque axe
La contribution Représente la participation de chaque
variable à la constitution d'un axe. Permet
d'identifier les points qui interviennent significativement dans la
constitution des axes
La corrélation (cosinus carré) Permet de mesurer la
qualité de la

116
représentation d'une variable sur l'axe
Les groupes ou types d'éleveurs constitués par
analyse multivariée ont été ensuite soumis à des
tests de comparaison des moyennes. Ainsi, les moyennes des effectifs animaux
par espèce et le sex-ratio ont été comparés sous
XLSTAT version 2010 4.03 par le test non paramétrique de
Kruskal-Wallis. Dans les cas où l'hypothèse alternative (Ha) (Ha
signifie que les éleveurs sont différents pour la variable
considérée) a été retenue, les groupes ont
été ensuite comparés deux-à-deux (comparaison
multiple) par le test de Steel-Dwass-CritchlowFligner (Scherrer, 2007).
Étant donné la méthode d'échantillonnage
utilisée, ces tests étaient les plus appropriés. En effet,
notre échantillon n'était pas totalement aléatoire et le
test de normalité s'est révélé non concluant.
5 .3. Résultats et discussions
5.3.1. L'élevage face à la pression
d'occupation de l'espace du terroir
5 .3.1.1. La pression démographique
Les données générales sur la population
ont été obtenues par estimation d'après les données
du recensement général de la population et de l'habitat de 2006
et du recensement administratif de 2004. Rappelons que pour le cas particulier
de la commune de Tansarga dont dépend le terroir de Kotchari,
l'estimation de la population faite dans le chapitre III donnait 42190
habitants avec une densité moyenne de 71,19 habitants/km2.
Le recensement administratif de 2004 donnait pour le terroir
de Kotchari (le villagemère et ses hameaux et campements) une population
de 15122 personnes. Cette population peut être estimée de nos
jours à environ 19806 soit un peu moins de la moitié (46,94%) de
la population de la commune de Tansarga, ce qui correspond à une
densité de 56,45 habitants/km2. Cette densité est
inférieure à la moyenne communale mais reste supérieure
aux densités enregistrées dans la moitié nord de la
province et même à la moyenne provinciale qui est de 37,39
habitants/km2. De ce point de vue, on peut estimer avec Nebié
(1996), un auteur ayant travaillé au plateau central du Burkina Faso (de
Manga à Kaya), que la pression humaine y est forte58 en
particulier sur les terres et les ressources naturelles, notamment
végétales. Il est bien évident que cette forte pression
humaine a des répercussions sur l'occupation des terres (Boutrais,
1983). En effet, l'évaluation de l'occupation des terres que nous avons
faite à partir de la base des données d'occupation des terres
(BDOT) réalisée par l'IGB en 200259 (cf. carte V-1 et
tableau V-3), outre le fait qu'elle confirme la forte domination des formations
de savane arbustive sur les autres types de formations végétales,
montre que plus de la moitié des terres du terroir sont actuellement
cultivées ou l'ont été plus ou moins récemment
(jachères). Cette situation peut s'expliquer par le fait que dans le
terroir
58 Nebié distingue en effet, trois niveaux
de charge des milieux en fonction de la densité humaine : (i) les
milieux de peuplement moyen (20 - 40 habitants/km2), les milieux de
forte concentration humaine (41- 80 habitants/km2) et les milieux
surchargés (> 80 habitants/km2).
59 Ces données sont certainement
dépassées de nos jours, mais nous n'avons pas pu en avoir de plus
récentes.

de Kotchari où les systèmes de production sont
restés au stade extensif, la couverture des besoins alimentaires
supplémentaires qui naissent de ce surplus de population est
assurée par l'augmentation des superficies cultivées. Cela est
une constante dans les pays sousdéveloppés notamment en Afrique
sub-saharienne comme l'ont observé Dugué et al. (2004)
et comme le rapporte Kièma S. (2007).
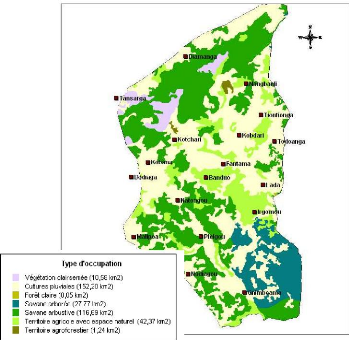
Carte V-1. Formes d'occupation des sols à Kotchari (IGB,
2002)
Tableau V-3. Types et niveau d'occupation des terres dans le
terroir de Kotchari (IGB, 2002)
|
Occupation des sols
|
Importance de la pression agricole
|
|
Type d'occupation
(nomenclature BDOT)
|
Sup (km2)
|
Terres
cultivées
|
Superficie cumulée
(km2)
|
Proportions
(%)
|
|
Cultures pluviales
|
152,20
|
|
|
|
|
Territoire agricole avec
|
|
Oui
|
195,81
|
55,81
|
|
espace naturel
|
42,37
|
|
|
|
|
Territoire agroforestier
|
1,24
|
|
|
|
|
Savane arborée
|
27,77
|
|
|
|
|
Savane arbustive
|
116,69
|
Non
|
155,04
|
44,19
|
|
Végétation clairsemée
|
10,58
|
|
|
|
|
Total
|
|
|
350,85
|
100
|

118
5.3.1.2. La pression animale : le bilan fourrager
saisonnier
En ce qui concerne le niveau de charge animale dans le terroir
et sa variation au cours de l'année, les enquêtes ont permis de
dénombrer les effectifs du cheptel (tableau V-4), toutes
catégories de cheptel prises en compte, ce qui a permis ensuite d'en
déduire le nombre d'UBT.
On observe que le cheptel local, sédentaire,
dominé par les caprins et les ovins en second lieu, est plus important
que celui qui transhume, même pendant le pic de transhumance. Si l'on
s'en tient uniquement à la race bovine, ce cheptel est constitué
de troupeaux aux tailles très variables dont les effectifs vont de
seulement quelques têtes (trois à cinq) à plus de 50
têtes dans quelques ménages considérés comme nantis,
généralement d'ethnie peule. Quant au bétail transhumant,
qui a été très important en 2008 et approchait même
par ses effectifs le cheptel local (l'effectif en bovins est même
supérieur à l'effectif local de la même espèce), il
a connu un recul assez notable en 2009.
Signalons que le nombre de troupeaux transhumants est, par
rapport à celui des troupeaux sédentaires, très faible
mais ce sont des troupeaux aux grands effectifs qui dépassent parfois
200 têtes.
Tableau V-4. Effectifs animaux en 2008 et 2009 (source :
données de recensement)
|
Types de troupeau
|
|
|
Année 2008
|
|
|
Année 2009
|
|
|
Effectifs sédentaires
|
Bovins
Ovins Caprins Asins Équins
|
|
6807
8016 9908 276 13
|
|
|
|
|
6957
8248
10235
276
13
|
|
|
|
Saisons de transhumance
|
|
S1
|
S2
|
|
S3
|
S4
|
S1
|
S2
|
S3
|
S4
|
|
Effectifs animaux transhumants entrant
|
Bovins
Ovins
Caprins
|
632
64
0
|
7004
843
22
|
|
1767
26
22
|
0
0
0
|
137
13
0
|
3954
1053
13
|
1868
136
0
|
0
0
0
|
|
Effectifs animaux transhumants sortant
|
Bovins
|
0
|
0
|
|
608
|
0
|
0
|
0
|
513
|
0
|
|
Total troupeaux locaux
|
|
|
1198 (-
|
18)
|
|
|
|
1198 (- 16)
|
|
|
|
Total troupeaux transhumants
|
|
|
95
|
|
|
|
|
52
|
|
|
Légende : S1 = [Déc. - Mars]; S2 = [Avril -
Mai]; S3 = [Juin - Juil.]; S4 = [Août - Nov.] ; les chiffres entre
parenthèse représentent les départs en transhumance
à partir du terroir.
De manière globale, le nombre d'UBT dans le terroir a
oscillé entre 10 515 et 17 692 en 2008 et entre 10 777 et 14 944 en 2009
(figure V-1). Les densités animales (ou charges instantanées),
calculées sur la base des espaces effectivement accessibles, ont
varié dans le même sens, allant de 0,32 UBT.ha-1
à 0,68 UBT.ha-1 en 2008 contre 0,31 UBT.ha-1
à 0,70 UBT.ha-1 en 2009 (tableau V-5). La période
où les effectifs animaux sont les plus élevés se situe
entre avril et mai, période de pointe de la transhumance, alors qu'ils
sont les plus bas en saison des pluies, notamment entre août et novembre.
C'est pourtant en cette période
pluvieuse où les transhumants allochtones sont tous ou
presque repartis vers leurs terroirs d'attache que les pâturages sont les
plus chargés par unité de surface. En cette saison une grande
partie du terroir est inaccessible du fait des activités agricoles
(UPP5) ou des risques d'embourbement (UPP2) ou des deux à la fois
(UPP1).
Les charges en bétail les plus faibles sont
enregistrées de décembre à mars au moment où tous
les espaces commencent à être accessibles mais où la
transhumance commence timidement.
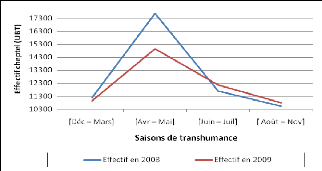
Figure V-1. Variation des effectifs animaux (en UBT) dans le
terroir en 2008 et 2009
Tableau V-5. Variation des charges de bétail (en UBT) dans
le terroir en 2008 et 2009 (Sources : Enquêtes terrain)
Périodes Densités
(UBT.ha-1)
Année 2008 Année 2009
[Déc. - Mars] 0,32 0,31
[Av - Mai] 0,50 0,43
[Juin - Juil.] 0,33 0,35
[Août - Nov.] 0,68 0,70
La recherche des capacités de charge théoriques
permet, par comparaison avec les charges réelles enregistrées, de
suivre au fil des saisons la situation réelle et de déceler les
saisons de grand risque pour les unités pastorales (surpâturage)
ou pour le cheptel (baisse des productions ou même perte de poids).
Rappelons que certaines unités, sont peu (UPP5 : zones les plus
cultivées) ou pas (UPP1 : bas-fonds et UPP2 : plaines inondables)
pâturables en saison des pluies. Nous les avons considérées
comme non exploitées en cette saison. Les autres unités (UPP3 :
bas-glacis et plaines argileuses; UPP4 : plateaux, hauts glacis et plaines
sableux à sablo-limoneux ; UPP6 : buttes rocheuses et cuirassées)
sont exploitées en toute saison. Ainsi, pour l'ensemble des
unités (tableau V-6), les capacités de charges théoriques
moyennes journalières ont été de 8 622
UBT.jour-1 (soit de 0,28 à 0,52
UBT.ha-1) et de 20 986
UBT.jour-1 (soit de 0,28 à 1,03
UBT.ha-1) respectivement en saison pluvieuse et en
saison sèche.

120
Tableau V-6. Capacité de charge théorique globale
du terroir de Kotchari
|
Unités
paysagères
pastorales
|
BM (kgMS.ha-1)
|
DF (kgMS.ha-1)
|
Sup (ha)
|
Capacité de charge
théorique (UBT.
j-1)
|
|
à l'hectare
|
sur l'unité
|
|
UPP1
|
4780
|
1673
|
8010
|
1,03
|
8246,60
|
|
UPP2
|
4590
|
1606,5
|
2853
|
0,99
|
2820,52
|
|
UPP3
|
3420
|
1197
|
3995
|
0,52
|
2096,23
|
|
UPP4
|
3270
|
1144,5
|
9359
|
0,50
|
4695,40
|
|
UPP5
|
1410
|
493,5
|
4273
|
0,30
|
1297,68
|
|
UPP6
|
1840
|
644
|
6482
|
0,28
|
1829,88
|
|
Total
|
19310
|
|
34972
|
|
20986,30
|
Pour les unités en gras, CC théorique moyenne
journalière de saison sèche; pour le reste CC théorique
moyenne journalière pendant toute l'année.
BM, biomasse ; DF, disponible fourrager ; Sup, superficie de
l'unité
DF = BM*Sup*0,35
Légende
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
Le bilan fourrager, qui équivaut à la
différence entre la capacité de charge théorique et la
charge globale réelle (figure V-2) confirme que le risque se situe bien
en saison pluvieuse (juin à novembre) pour toute la période de
référence (2008 & 2009). En cette saison en effet, le bilan
est négatif pour les pâturages accessibles qui sont alors en
surpâturage. On peut être tenté de penser que la relative
bonne santé des parcours du terroir dans sa partie accessible
légalement (seules quelques unités sont en dégradation
légère), est imputable aux aires protégées qui
résorbent cet excédent de charge. Cette hypothèse est
à première vue invalide, les deux aires protégées
voisines (parc W et réserve partielle de la Kourtiagou) étant en
effet insalubres (il y sévit la mouche tsé-tsé) et
impraticables en cette saison (leurs parties qui bordent le terroir
étant vite inondées avec les premières pluies). Mais les
observations faites ailleurs en contexte presque similaire par Kaboré
(2010) permettent de ne pas exclure totalement cette possibilité.
L'auteur a, en effet, constaté que la réserve de Pama Nord
devenait pour les éleveurs peuls, un refuge pour prévenir les
conflits à l'égard des agroéleveurs gourmantchés et
mossi ; conflits consécutifs à la trop grande concentration
d'animaux en périphérie causée par le retour des troupeaux
transhumants.
En saison sèche, au contraire, et
particulièrement en début de cette période
(décembre à mars), les charges réelles sont nettement
inférieures aux capacités théoriques et le bilan est
très satisfaisant. Il le serait davantage si, en plus de
l'évaluation des disponibilités fourragères naturelles, on
avait pu prendre en compte les résidus culturaux qui sont
exploités en vaine
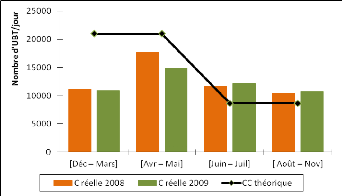
pâture en cette période de même que le
fourrage ligneux dont la contribution dans les rations en fin de saison
sèche est très importante (Boudet, 1991 ; Lhoste et al.
1993 ; Daget & Godron, 1995 ; Kaboré-Zoungrana, 1995 ; Raimond,
1999 ; Kagoné, 2000 ; Lhoste, 2004 ; Yanra, 2004 ; Kièma S., 2007
; Dongmo, 2009).
Figure V-2. Evolution du bilan fourrager au cours des
années 2008 & 2009
Ce bilan reste indicatif, il ne s'appuie en effet que sur une
évaluation ponctuelle effectuée au maximum de biomasse en
septembre. Par ailleurs, malgré le risque encouru, les interstices entre
les parcelles agricoles des mosaïques agroforestières (UPP5) sont
pâturés, en particulier par le bétail local, notamment
gourmantché. Inversement, les unités du terroir
considérées comme accessibles en saison pluvieuse y compris les
collines (UPP3, UPP4 et UPP6), hébergent par endroits des parcelles
cultivées. Ces unités ne sont donc pas, dans leur
entièreté, accessibles en cette saison. La surcharge ainsi
constatée en saison pluvieuse pourrait en conséquence avoir
été surestimée dans l'hypothèse où le
territoire exploré dans UPP5 produisait une biomasse beaucoup plus
importante que les portions non accessibles de UPP3, UPP4 et UPP6 ou
sous-estimée si, au contraire, elle en produisait moins.
La très forte variation du niveau de charge animale
dans le terroir au cours de l'année n'est pas seulement attribuable aux
activités agricoles qui occupent plus de la moitié (55,81%) de
l'espace du terroir, en effet les flux et reflux annuels de transhumants y
tiennent un rôle majeur. Ceci confirme les observations faites dans la
même zone par Paris (2002) qui rapporte qu'en avril-mai (période
de pic de transhumance), on pouvait compter autant d'animaux transhumants que
de locaux. De plus, en 2008, de décembre à mars et d'avril
à mai, l'afflux de transhumants a été plus important que
l'année suivante. La plupart de ces transhumants sont cependant plus
vite repartis vers leurs terroirs d'attache en 2008 qu'en 2009, comme l'atteste
la charge de bétail moins importante pendant la période de
juin-juillet de la première année. A cette période en
effet, l'effectif d'animaux transhumants a été inférieur
à celui enregistré l'année d'après.

122
Si de telles charges en bétail peuvent paraître
très élevées et inquiétantes, les craintes doivent
se porter surtout sur une tendance confirmée à l'accroissement
des effectifs car le terroir continue « paradoxalement » d'attirer de
nouveaux transhumants. De tels niveaux de charge peuvent en effet induire des
changements très importants dans la végétation ou
même dans les sols des parcours, sans pourtant toujours affecter
immédiatement le niveau de la production primaire, comme l'ont
observé Kamuanga et al. (2003) dans l'extrême Nord du
Cameroun. Il faut cependant rester prudent à propos de ce dernier point
car, à Kotchari, le dépassement de charge a lieu au moment
où la strate herbacée, principale composante du fourrage, est en
pleine croissance. On imagine en effet mal comment, dans ces conditions, les
capacités productives ne seraient pas affectées et comment le
niveau de production nette potentielle ne serait pas influencé
négativement.
Malgré le bilan fourrager positif en saison
sèche, on est en droit de s'interroger sur les facteurs qui ont
favorisé et continuent de favoriser la mise en place d'effectifs animaux
aussi élevés dans le terroir. En effet la densité humaine
y est notable, la pression sur les terres se fait importante et, par ailleurs,
les valeurs des pâturages ne sont pas si exceptionnelles que cela (voir
chapitre IV précédent et Sawadogo et al., sous
presse). Il est possible de trouver une explication à un tel niveau
de charge animale. Selon certains auteurs (Boutrais, 1983 & 1996 ;
Dugué, 1998 ; Santoir, 1999 ; Dugué et al. 2004), dans
une certaine fourchette de densité humaine, tant que l'espace n'est pas
encore saturé, la taille du cheptel croît presque toujours avec la
densité démographique. Cette fourchette se situerait entre 20 et
50 à 60 habitants/km2 de densité humaine ou entre 15%
à 50% voire 60% de niveau d'occupation agricole des terres (Dugué
et al. 2004). À partir de ses travaux conduits au Cameroun et
au Nigeria, Boutrais (1983) a constaté par contre que dès 30-40
habitants/km2, le nombre de têtes de bétail par
habitant ainsi que la densité de bétail au km2
amorçaient une tendance à la baisse. Les mêmes auteurs ont
observé, par ailleurs, qu'en dessous de 20 habitants/km2 le
milieu, dans ces zones, est insalubre à cause des glossines et qu'il
n'attire donc pas les éleveurs. Il faut savoir que les agriculteurs, par
leurs activités, sont reconnus comme de bons agents d'assainissement des
milieux (Bonfiglioli, 1990 ; Boutrais, 1996 ; Dongmo et al. 2007), de
sorte que les zones de concentration des éleveurs et donc du
bétail coïncident, en général, avec celles de
concentration agricole (Boutrais, 1983 & 1996 ; Dugué et
al. 2004).
Les conditions d'installation des éleveurs étant
favorables à partir de 20 habitants/km2 si les conditions
sanitaires sont favorables (notamment faible risque de trypanosomose), les
effectifs de bétail augmentent aussi bien au niveau des troupeaux des
éleveurs que chez ceux des agriculteurs (rappelons que ceux-ci
capitalisent leurs revenus agricoles dans le bétail), ceci tant que les
possibilités d'extension des parcelles agricoles restent possibles. Ce
qui ne serait plus le cas à partir de 40-60 habitants/km2
(Boutrais, 1983), densité à partir de laquelle les relations
auparavant cordiales deviennent conflictuelles et les éleveurs se
trouvent contraints d'adopter de nouvelles stratégies, soit en se
sédentarisant60 soit en allongeant leur saison de
transhumance comme l'ont observé Dugué et al. (2004) en
pays Sereer au Sénégal et dans l'extrême Nord du Cameroun.
Notons cependant avec Boutrais (1983) qu'en situation
60 Ce qui implique, dans bien de cas, le passage du
système vers le mode intensif de production animale (Raimond, 1999 ;
Requier-Desjardins, 1999).

d'intégration véritable entre l'agriculture et
l'élevage, ce qui l'est moins à Kotchari, le bétail peut
continuer à croître jusqu'à un niveau de densité
humaine atteignant 100 habitants/km2.
Si l'on s'appuie sur les observations de ces auteurs, on ne
peut raisonnablement expliquer la venue massive et cyclique des éleveurs
dans ce terroir particulier par la prétendue richesse en ressources
pastorales et par la disponibilité en espace qui sont souvent
évoquées par ces derniers. On voit bien qu'avec une
densité humaine de 56,45 ha/km2, un taux d'occupation des
sols de 55,81% et une faible intégration de l'agriculture à
l'élevage, les conditions ne sont objectivement pas réunies,
même si, par ailleurs, les charges en bétail restent raisonnables
durant la saison sèche concernée par la transhumance.
Visiblement à Kotchari, le point de rupture est atteint
ou presque et, en toute logique et suivant le schéma observé
ailleurs, on devrait s'attendre à ce que les départs en
transhumance soient de plus en plus importants ou que les effectifs de
bétail amorcent leur baisse comme ce fut le cas entre 2008 et 2009. A la
lumière de nos observations récentes cette baisse apparaît
comme un simple artefact puisqu'au contraire, on enregistre un accroissement
continu des effectifs locaux et transhumants. Aucune explication rationnelle
basée sur les potentialités du terroir ne peut justifier cette
tendance et on peut logiquement penser que les ressources pastorales des
réserves voisines sont mises à contribution. Nous reviendrons sur
la question dans la discussion générale dans le chapitre VII.
5.3.2. Les races bovines dans le terroir
Une étude sur les pratiques et stratégies des
éleveurs mobiles se doit de porter une attention particulière aux
espèces et surtout aux races animales (notamment bovines) en
présence. Les races servent en effet souvent, de marqueur identitaire au
groupe pastoral (Boutrais, 1996). On observe une diversité relativement
grande dans les races bovines au sein des troupeaux d'élevage à
Kotchari. Les races les plus courantes sont la Barbaji et la
Gurmaji, cependant avec la forte fréquentation du terroir par
les transhumants, la race Jaliji (ou Djeli) y est
également importante à certaines périodes de
l'année, notamment pendant la saison sèche chaude de mars
à mai. À côté de ces races dominantes, on rencontre
aussi des Boboroji (ou Borheji), des Gudali et des
Kiwali.
Il faut toutefois noter qu'il existe un fort taux de
métissage entre ces races et qu'en l'absence de toute étude de
caractérisation génétique, il est impossible de juger de
la pureté de
celles-ci. On sait par exemple que la race Barbaji, ou
race Borgou, rencontrée au Bénin
d'oüelle a d'ailleurs été importée, est
issue du métissage entre le zébu Fulani et la race
taurine
Somba (taurin de petite taille à courtes
cornes) (Dehoux et Hounsou, 1993). Il est possible qu'il en soit de même
pour les autres races. Certaines des races rencontrées à Kotchari
ne sont peut-être que des écotypes locaux de races existantes ou
des hybrides entre elles. Nous ne cherchons pas ici à identifier et
à caractériser précisément les races localement
rencontrées. Un tel objectif aurait nécessité un
dispositif plus lourd avec des travaux de laboratoire assez coûteux et
des compétences en génétique qui sortent largement du
cadre du présent travail. Il s'agit ici uniquement d'identifier et de
comprendre les représentations que les éleveurs font

124
des différentes "races" supposées de leur cheptel
et des stratégies qu'ils mettent en oeuvre en conséquence.
Les données présentées ici sur les
caractéristiques productives et reproductives sont issues
d'enquêtes auprès d'éleveurs expérimentés que
nous avons conduites avec D. Ouédraogo dans le cadre de son
mémoire d'ingénieur (2008) dont nous avons assuré
l'encadrement. Ces données ont été
complétées par la littérature. Ne résultant donc
pas d'études génétiques, elles doivent être
considérées avec précaution. Cependant, comme elles
résultent d'observations sur une longue période, la
hiérarchisation faite entre les races par les éleveurs pour
chaque paramètre peut être considérée comme à
peu près correcte. Malgré son caractère empirique, cette
classification suffit largement pour comprendre les choix qu'opèrent les
éleveurs dans les races bovines qui composent leurs troupeaux.
5.3.2.1. La race Barbaji
Elle est, avec la Gurmaji, la race la plus importante
du terroir. Originaire du Bénin, elle serait entrée dans le
terroir il y a plus de 60 ans. Son nom est dérivé de «
Bargoube », appellation gourmantché d'une ethnie du Nord
Bénin (les Bariba), propriétaire de cette race. Cette
race résulterait d'un métissage (Vissac, 1994) entre le
zébu White Fulani et le taurin Somba (Dehoux et
Hounsou, 1993) ou entre le zébu et le Muturu, une race taurine
rencontrée au Nigeria (Kagoné, 2000).
De petite taille, elle est caractérisée par une
quasi-absence de bosse, de petites cornes et une queue peu
développée. La couleur de la robe est variable. L'âge au
premier vêlage varierait entre 3 et 4 ans et l'intervalle de mise bas
serait d'un an dans les conditions de Kotchari.
Pour les éleveurs, l'intérêt de cette race
est lié à sa trypanotolérance, à sa
rusticité alimentaire qui lui permet de s'adapter à l'état
actuel des ressources et à sa prolificité. Elle est peu exigeante
en alimentation, et se déplace peu au pâturage. Cette
rusticité lui permet, par ailleurs, de garder sa production
laitière à un bon niveau à toute période de
l'année.

Cliché Sawadogo
Photo V-1: Race Barbaji 5.3.2.2. La race Gurmaji(ou
Guiabo)
Cette race tire son nom de « Gurmaabe » qui
signifie en langue peule, les Peuls qui vivent en territoire gourmantché
(ou Peul du pays gourmantché). Selon les éleveurs, la race

Gurmaji serait originaire du pays gourmantché
précisément de la zone de Kantchari. Mais, alors que
Kaboré (2010) la considère comme résultant du
métissage entre de multiples races fréquentant la zone
(Keteji, Pulli, Kiwali, Jaliji, Gudali), les travaux de Santoir (1999)
indiquent plutôt qu'en réalité elle a été
introduite par les premiers immigrants peuls, c'est-àdire les
Gurmaabe.
Grande de conformation, la Gurmaji a une bosse
très développée et bien dressée chez le mâle.
La queue est également bien développée et a tendance
à frôler le sol. Le cornage, lui aussi bien
développé, est fait de longues cornes larges et peu robustes. Le
pelage est variable. L'âge au premier vêlage varierait entre trois
et quatre ans et l'intervalle de mise bas serait de deux ans environ dans les
conditions actuelles. Avec la Barbaji, cette race est la plus
représentée dans les effectifs ce qui s'explique par sa
très bonne adaptation aux conditions locales. Elle combine en effet la
trypanotolérance observée chez les Barbaji avec les
performances productives (viande et lait) des zébus sahéliens,
mais les informations sur son adaptation à la marche sont
contradictoires. En effet, alors que la population la considère comme
bonne "marcheuse", (Kaboré, 2010) avance le contraire, ce qui lui fait
dire d'ailleurs que cette race est signe de
sédentarité61.

Cliché Guiré R.
Photo V-2: Race Gurmaji 5.3.2.3. La race Kiwali
Cette race est originaire du Nigeria, plus
précisément du pays haoussa, où résiderait une
communauté d'éleveurs d'ethnie « Kiwabe ».
Elle a été introduite dans le terroir il y a environs 50 ans par
des migrants peuls et ne se rencontrerait que dans quelques troupeaux des
descendants de ceux-ci.
De très grande taille, cette race doit surtout sa
particularité à son cornage fait de longues et robustes cornes
tournées vers l'arrière et à sa longue queue. La bosse
n'est pas développée. De nos jours, la race pure est pratiquement
inexistante et l'on rencontre surtout des métis présentant des
traits intermédiaires soit entre la Kiwali et la Barbaji,
soit entre la Kiwali et la Gurmaji.
Dans les conditions actuelles, l'âge au premier
vêlage serait de quatre ans et l'intervalle de
mise bas de deux ans.
Très exigeante en alimentation, la Kiwali se distingue surtout
par sa
capacité à parcourir de longues distances. Sa faible
rusticité alimentaire la rend cependant
61 Dans la suite du document nous retenons le point de
vue de la population compte tenu de la démarche adoptée.

126
vulnérable au manque de ressources et aux maladies.
Elle serait une bonne laitière selon les éleveurs et
présente une très bonne rentabilité économique au
regard de sa grande conformation.

Cliché Guiré R.
Photo V-3: Race Kiwali
5.3.2.4. La race
Boboroji(Mbororoojiou Borheji
Cette race, très esthétique, de robe
généralement rouge, est originaire du pays Woodabe (ou
Bororo), un groupe de Peuls pasteurs rencontré entre l'Est du
Niger, le Nord-est du Nigeria, le Sud-ouest du Tchad (Bovin, 1999), et du fait
d'une migration ancienne, dans le Nord Cameroun et en Centrafrique (Lhoste,
1969). Elle a été introduite dans le terroir de Kotchari via le
Niger. Elle est de grande taille et se caractérise par son pelage
généralement roux, son fourreau pendant, son cornage
développé. La bosse est assez développée et
tombante. La hauteur au garrot oscille entre 75-120 cm pour un poids variant
entre 250-300 kg (femelle) et 350-500 kg (mâle). L'âge au premier
vêlage et l'intervalle de mise bas se situeraient respectivement à
42 mois et à 11 mois alors que la production laitière est faible
: 1- 1,6 l/jour pendant 180-200 jours. Le rendement carcasse est de 40%- 50%.
Réputée pour ses capacités à parcourir de longues
distances au pâturage, elle est cependant exigeante et sélective
en alimentation (Amadou, 1999 ; Boutrais, 1996 & 2002) et très
sensible aux trypanosomoses (Boutrais, 2002). Elle résiste à la
soif, mais est très sensible à la faim selon les éleveurs.
Ceci pourrait expliquer sa faible représentation dans les troupeaux.
Boutrais (1996) note par ailleurs que le Boboroji (Red
Bororo) a un comportement sélectif au pâturage alors que la
White Fulani (Akuuji) qui lui est proche (ce sont toutes deux
des pseudo-Zébus) et qui est plus rustique a une pâture
plutôt rasante cependant dommageable pour l'équilibre des
pâturages. Pour les éleveurs de Kotchari, même si cette race
est économiquement rentable, la difficulté de son élevage
réside dans sa faible prolificité et son caractère sauvage
qui fait d'elle une race à la contention difficile. Cela n'est pourtant
pas toujours négativement perçu dans certaines communautés
pastorales car l'attitude « guerrière » (na'i
kono)62 des vaches Boboroji (Boutrais, 2002) est
propice à des activités pastorales en contexte
d'insécurité (vol, prédation, etc.).
62 Signifie vaches guerrières (Boutrais, 2002).

Cliché Convers (2002)

Photo V-4: Race Boboroji
5.3.2.5. La race Jaliji(Djeli)
Cette race de zébu, très voisine de la race
Gurmaji, est venue du Niger ; elle est la plus fréquente dans
les troupeaux transhumants nigériens. Elle serait rustique au plan
alimentaire mais supporterait peu la marche, en particulier sur les longues
distances (Amadou, 1999). D'une hauteur au garrot 115 à 130 cm, elle
pèse entre 250 et 300 kg et produit 2 à 3 litres de lait par jour
pendant environ 270 jours. Le rendement carcasse varie entre 48 et 50%)
(Geesing & Djibo, nd; MRA, 2003).

Cliché Sawadogo
Photo V-5: Race Jaliji près d'un campement de
transhumant
5.3.2.6. La race Gudali
La race Gudali est une race traditionnelle
d'éleveurs sédentaires (les Foulbés) de l'Adamaoua (nord
Cameroun) (Boutrais, 2002 ; Kossoumna Liba'a, 2008). Elle est très peu
présente à Kotchari. C'est une race trapue au cornage peu
développé avec une bosse dorsale retombante. Le lait produit est
de 4 - 5 l/j sur une durée de 246 à 300 jours et le rendement
carcasse se situe entre 59% et 63%, ce qui lui confère de bonnes
qualités bouchères lui conférant le surnom de charolais
d'Afrique (Kossoumna Liba'a, 2008). Sa prolificité est cependant l'une
des plus faibles (Boutrais, 2002). Par ailleurs, elle a un comportement
sélectif au pâturage. Compte tenu de cette qualité, on
observe que de plus en plus d'éleveurs modernes commencent à
l'importer du Nigéria aux fins d'amélioration de leurs
troupeaux.

Cliché Sawadogo

128
Photo V-6: Race Gudali
5.3.3. Les acteurs et leurs pratiques d'élevage
5.3.3.1. Résultats généraux
La figure V-3 et le tableau V-7 synthétisent les
résultats généraux issus de l'analyse des données
issues des enquêtes.
Les éleveurs enquêtés sont essentiellement
des autochtones de Kotchari (150 éleveurs, soit 75%). Seulement 18 (soit
9%) ne sont pas des nationaux, les autres viennent des différentes
communes de la province (19 éleveurs, soit 9,5%) ou des provinces de la
Komondjari ou du Yagha (13 éleveurs, soit 6,5%), situées plus au
Nord-ouest de la Tapoa.
L'effectif du cheptel sur lequel a porté
l'enquête est de 14 631 têtes ; il comprend 109 73 bovins (soit
75%), 2 634 ovins (soit 18%) et 1 097 caprins (soit 7%) (figure V-3). Par
ailleurs, parmi les éleveurs enquêtés, seulement quinze
(soit 7,5%), tous Gourmantchés, ne possèdent pas l'espèce
bovine dans leurs troupeaux. Toujours au sujet des bovins, on peut noter que
49,5% des troupeaux ont un effectif de plus de 50 têtes et que 27% ont au
moins 100 têtes. En outre, huit troupeaux de plus de 200 bovins ont
été enregistrés qui appartiennent tous à des
éleveurs transhumants d'ethnie peule. En ce qui concerne les ovins, 50
éleveurs (soit 25%) n'en disposent pas, parmi lesquels on
dénombre sept Gourmantchés (soit 3,5%) et 43 Peuls (soit 21,5%).
Les caprins sont les moins représentés dans les troupeaux pris en
compte. En effet, 110 éleveurs (soit 55%) dont 102 Peuls (51%) et
seulement trois Gourmantchés (4%) n'en disposent pas au sein de leurs
élevages.
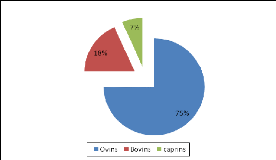
Figure V-3. Proportions des espèces dans l'effectif de
bétail enquêté (source : cette thèse, données
d'enquête).
Les troupeaux à effectifs majoritairement (48%) bovins
(TaiTr) de moins de 45 têtes sont surtout plurispécifiques (80%)
le plus souvent ils ne comportent que des bovins et ovins (35,5%) ou les trois
espèces bovines, ovines et caprines (33,5%). On note toutefois une bonne
représentation des troupeaux monospécifiques bovins (18%).
Dans les troupeaux, il est courant d'avoir plusieurs races
bovines (2 races et plus) (68%) avec notamment une combinaison de
Barbaji et de Gurmaji (33%) ou bien de Gurmaji et de
Jaliji (20%). Les troupeaux à plus de deux races (15%) ou
composés seulement de Gurmaji (11%) sont moins nombreux, mais
sont toutefois plus représentés que ceux qui n'ont que des
Barbaji (6%) ou que des Jaliji (7,5%).
Tableau V-7. Données générales sur
l'échantillon enquêté.
Variables
|
Modalités
|
Intervalles ou contenu des
classes
correspondantes
|
Effectif
d'éleveurs
|
Fréquence par
modalité (%)
|
|
Variables actives
|
|
|
|
|
|
TaiTr
Effectif bovin du troupeau
|
1
2
3
4
5
|
[0 45]
[46 90]
[91 135]
[136 180]
Plus de 180 têtes de bovins
|
97
39
37
14
13
|
48,50 19,50 18,50 7,00 6,50
|
|
NEsp
Nombre et types d'espèces animales dans le troupeau
|
1
2
3
4
5
|
Bovins + Ovins + Caprins Bovins + Ovins
Bovins + Caprins
Bovins
Pas de bovins
|
67
71
11
36
15
|
33,50 35,50 5,50 18,00 7,50
|
|
NRaB
Nombre et types de races bovines du troupeau
|
1
2
3
4
5
6
|
Barbadji
Gurmaji
Jaliji
Barbadji + Gurmaji
Gurmaji + Jaliji
Autres (plus de 2 races bovines)
|
12 22 15 66
40
30
|
6,00 11,00 7,50 33,00 20,00 15,00
|

130
|
7
|
Chapitre V. Les systèmes d'élevage
à Kotchari
Aucun (Pas de bovins dans le
troupeau) 15 7,50
|
|
OrTr
|
1
|
Achat + Emprunt
|
49
|
24,50
|
|
Mode de constitution ou origine du troupeau
|
2
3
|
Héritage + Don Achat + Héritage
|
74
69
|
37,00
34,50
|
|
4
|
Autres (plus de 3 formes d'acquisition)
|
8
|
4,00
|
|
LPat
|
1
|
Terroir Kotchari
|
75
|
37,50
|
|
Lieu de pâturage en saison sèche
|
2
3
|
Kotchari + terroirs voisins Pays voisins: Bénin / Togo
|
59
58
|
29,50
29,00
|
|
4
|
Réserves voisines
|
8
|
4,00
|
|
RTra
|
1
|
Eau
|
28
|
28,00
|
|
Raisons de la pratique de la transhumance
|
2
3
|
Fourrages Eau+fourrage
|
5
64
|
5,00
64,00
|
|
4
|
Habitude
|
3
|
3,00
|
|
5
|
Ne transhume pas
|
100
|
----
|
|
NatBe
|
1
|
propriétaire ou parent
|
170
|
85,00
|
|
Type de berger utilisé
|
2
|
Salarié
|
13
|
6,50
|
|
3
|
Mixte (parent + salarié)
|
17
|
8,50
|
|
NBe
|
1
|
Un berger
|
110
|
55,00
|
|
Nombre de bergers utilisés
|
2
|
Deux bergers
|
45
|
22,50
|
|
3
|
Plus de deux bergers
|
45
|
22,50
|
|
NLot
|
1
|
Un lot
|
48
|
24,00
|
|
Allotement du troupeau
|
2
|
Deux lots
|
112
|
56,00
|
|
3
|
Plus de deux lots
|
40
|
20,00
|
|
RLot
Raisons dans l'allotissement du troupeau
|
1
|
Former lots homogènes (séparer bien portants des
fatigués,
malades, etc.)
|
39
|
26,53
|
|
2
|
Gérer les risques
|
48
|
32,65
|
|
|
Alléger les charges sur les
|
|
|
|
3
|
parcours
|
33
|
22,45
|
|
4
|
Faciliter la tâche des bergers
|
27
|
18,37
|
|
5
|
Ne pratique pas
|
53
|
----
|
|
1
|
Pâturage naturel
|
43
|
21,50
|
|
PAlt
|
2
|
Complément par fourrage naturel
|
15
|
7,50
|
|
Pratiques d'alimentation du troupeau
|
3
|
Complément par fourrage cultivé
|
62
|
31,00
|
|
4
|
Complément par concentré acheté
|
25
|
12,50
|
|
|
Complément cultivé, collecté et
|
|
|
|
5
|
acheté (fourrage et concentré)
|
55
|
27,50
|
|
Variables supplémentaires
|
|
|
|
|
|
Eth
|
1
|
Gourmantché
|
50
|
25,00
|
|
Ethnie de l'éleveur
|
2
|
Peul
|
150
|
75,00
|
|
1
|
Kotchari
|
150
|
75,00
|
|
OrEl
|
|
|
|
|
|
2
|
Dans la Tapoa
|
19
|
9,50
|
|
Lieu de résidence habituelle
|
3
|
Burkina Faso hors Tapoa
|
13
|
6,50
|
|
ou origine de l'éleveur
|
4
|
Non burkinabè
|
18
|
9,00
|

131
Les troupeaux enquêtés ont été
constitués par plusieurs voies notamment par héritage
associé soit au don (37%) ou alors à l'achat (34,50%). L'achat
auquel s'associe l'emprunt est aussi une forme bien représentée
(24,5%). Signalons que l'héritage est la forme dominante d'acquisition
chez les Peuls, alors que l'achat est la voie dominante chez les
Gourmantchés.
En saison sèche c'est dans l'espace du terroir
villageois que pâturent les troupeaux. Au total 67% d'éleveurs
adoptent cette pratique, 37,5% exploitent exclusivement les pâturages du
terroir et 29,5% mettent aussi à contribution les pâturages des
terroirs voisins. Une proportion importante d'éleveurs (29%)
déclare aller en transhumance, notamment au Bénin, alors que
seuls huit éleveurs (soit 4%) reconnaissent exploiter le parc W en plus
des pâturages du terroir.
Les motifs généralement évoqués
par les éleveurs qui transhument pour justifier leurs
déplacements parfois lointains sont la recherche combinée de
l'eau et du fourrage (64% des éleveurs) ou seulement de l'eau (28%).
Seulement 5% des éleveurs avancent la question fourragère comme
argument exclusif de déplacement alors que 3% d'entre eux inscrivent le
fait qu'ils pratiquent la mobilité par simple héritage familiale,
bien qu'ils n'ignorent pas par ailleurs, les avantages que la pratique procure
aux animaux.
La garde des animaux est d'ordinaire assurée par 1
berger (soit 55%), parfois 2 bergers (soit 22,5%) ou davantage (22,5%). Il
existe un lien étroit entre la taille du troupeau et le nombre de
bergers. En effet, les troupeaux suivis par au moins deux bergers sont
généralement de grande taille. Les bergers sont
généralement de proches parents (85%) (l'éleveur
lui-même, ses fils ou ses neveux), ou des proches aidés de bergers
salariés (8,5%). Il est plus rare (6,5% de cas) que le troupeau soit
gardé exclusivement par des bergers salariés.
L'allotement (division du troupeau en lots plus ou moins
homogènes) est une pratique bien ancrée dans les habitudes, elle
touche 76% des éleveurs. La préférence est de former deux
lots (56%) plutôt que trois ou plus (20%). Parmi les éleveurs qui
ont adopté la pratique d'allotement, les motivations sont
généralement le souci de gérer les divers risques
(dégâts de champs ou perte d'animaux, mortalités dues aux
longues marches pour les animaux malades ou épuisés par
l'âge par exemple) (32,65%). Mais il s'agit également de
constituer des lots homogènes (malades/bien portants ; adultes/petits ;
mâles/femelles) (26,53%), d'alléger les charges sur les parcours
(22,45%) ou encore d'alléger la tâche des bergers (19,05%).
En ce qui concerne les pratiques d'alimentation, la
complémentation (apport de compléments alimentaires
achetés, cultivés ou collectés et conditionnés) est
de rigueur. Elle est mise en oeuvre par 78,50% des éleveurs
enquêtés. Ainsi, seulement 21,50% d'entre les éleveurs
disent continuer de se contenter des fourrages directement
prélevés par les animaux sur les pâturages. Dans le groupe
d'éleveurs engagés dans la pratique de complémentation, la
majorité (40%) d'entre eux soit achète seulement du
concentré (12,50%), soit utilise diverses sources de compléments
(fourrage collecté, cultivé et achat de fourrage et de
concentré) (27,50%). Un grand nombre cependant (31%) de ces
éleveurs, généralement de grands agriculteurs, utilise
uniquement le fourrage issu des parcelles cultivées tandis que seulement
7,50% complémentent leurs animaux avec du fourrage naturel
collecté et mis en stock.
5.3.3.2. Les types d'élevages dans le terroir
et leurs caractéristiques
La typologie des élevages dans le terroir a
été établie pour elle-même à titre
descriptif, mais surtout afin de fournir une base d'échantillonnage pour
le suivi ultérieur des pratiques et stratégies des
éleveurs au pâturage. Rappelons que la population du terroir de
Kotchari est constituée majoritairement de Gourmantchés à
côté desquels on note une forte présence de Peuls
sédentaires. Par ailleurs, ce terroir est une destination de Peuls
transhumants qui y constituent presque la moitié des effectifs pendant
la saison de transhumance. Tous ces groupes pratiquent l'élevage de nos
jours. Le système d'élevage pastoral, notamment sa forme
extensive sur parcours villageois, est le système le plus courant. C'est
la forme d'élevage pratiquée traditionnellement et qui reste
prédominante chez les Peuls comme chez les Gourmantchés. Comme le
terroir est, adossé au parc W et à la réserve partielle de
faune de la Kourtiagou (ou Concession de chasse de la Kourtiagou), la question
de la conservation y est très sensible ; il est donc nécessaire
de s'interroger sur la manière dont les systèmes d'élevage
s'accommodent des restrictions d'usage associées aux réserves et
comment les gestionnaires de ces dernières gèrent les
éventuelles incursions des troupeaux. De ce point de vue, nous sommes
parti de l'hypothèse que les différents groupes n'ont pas les
mêmes pratiques et notamment pas les mêmes rapports à
l'espace et à ses ressources. Il était en conséquence
nécessaire de définir ces groupes.
5.3.3.2.1. Le tri et la catégorisation des
variables en vue de la typologie
Le test de corrélation entre variables, en vue de
déceler une redondance éventuelle, a montré que la
variable "taille du troupeau bovin" (TaiTr) était associée
à la variable "nombre de bergers" (NBe) (R2 = 0,66) et
à la variable "principale motivation dans l'allotement" (RLot)
(R2 = 0,52) ; la variable "ethnie de l'éleveur" (Eth) est,
quant à elle, corrélée aux variables "mode de constitution
du troupeau" (OrTr) (R2 = 0,76) et "nombre de lots dans le troupeau"
(NLot) (R2 = 0,56) qui est, par ailleurs, associée à
RLot (R2 = 0,55). Les variables OrTr, NBe, RLot et NLot, qui sont
moins importantes que TaiTr et Eth, n'ont donc pas été prises en
compte dans la typologie. Le nombre de lots et de bergers sont en effet
fonctions de la taille du troupeau comme l'a aussi observé Kagoné
(2000). De même, l'origine du troupeau est de nature socioculturelle, il
est généralement mis en place par héritage chez les
éleveurs peuls (Bonfiglioli, 1988) ; ce qui l'est moins chez les
Gourmantchés.
Deux groupes de variables parmi celles retenues ont ensuite
été formés : des variables actives (sept) et des variables
supplémentaires (deux). Nous avons considéré comme actives
les variables qui sont plus révélatrices d'évolution ou de
changement dans les pratiques, et comme supplémentaires, celles qui
permettent de saisir la structure de la distribution en fonction des
caractéristiques qui nous intéressent (Pollak, 2009). Ces
variables, qui vont permettre d'éclairer la situation
étudiée, sont aussi dites explicatives ou illustratives (Cibois,
2007). Elles sont projetées dans le nuage de points mais ne participent
pas à sa construction.
5.3.3.2.2. Les axes factoriels, les variables et les
modalités associées
Les pourcentages d'inertie des différents facteurs issus
de l'analyse factorielle des correspondances multiples (AFCM) (figure V-4)
réalisée avec neuf variables (sept actives et


133
deux supplémentaires) (tableau V-7 ci-dessus) montrent
que l'essentiel (78,01%) de l'information est fourni par le plan F1-F2. F1
explique 63,88% de l'information et F2 en explique 14,13%. La forme
particulière, en parabole, sous laquelle se présente le nuage de
modalités est courante et désignée dans la
littérature sous le terme d'effet Guttman (Cibois, 2007). D'un point de
vue statistique, l'effet Guttman met en évidence l'existence de
corrélations entre variables. Dans le cas de notre jeu de données
il montre que les catégories d'éleveurs ne sont pas
discrètes, mais qu'il y a au contraire une continuité entre elles
avec des intermédiaires : il existe clairement des liens multiples entre
les réponses des éleveurs. On peut réduire l'effet Guttman
en jouant sur un meilleur équilibrage du nombre de modalités trop
différent entre les variables et sur un regroupement éventuel
pour que celles-ci n'aient pas un poids trop différents (Cibois, 2007).
Dans ce sens, les variables "effectif bovin du troupeau" (TaiTr) et "nombre et
types d'espèces animales dans le troupeau" (NEsp) comprenant
initialement neuf et sept modalités respectivement ont été
revues pour ne former que cinq modalités chacune. Il n'a cependant pas
été possible d'en faire autant pour la variable "nombre et types
de races bovines du troupeau (NRaB)" qui comprend sept modalités dont le
regroupement faisait perdre beaucoup d'informations. L'effet Guttman bien que
réduit à l'issue de cette opération, a néanmoins
persisté, mais le nuage tel qu'il se présente permet de bien
expliquer nos données.
La forte valeur enregistrée pour le facteur F1 dans
l'histogramme des valeurs propres (environ 0,29) (figure V-5) confirme, comme
montré plus haut, que ce facteur résume l'essentiel de
l'information ; en effet, le seuil empirique de forte liaison pour cet
indicateur est estimé à 0,1 (Cibois, 2007). Un tel cas traduit un
phénomène pratiquement unidimensionnel (Pollak, 2009),
l'interprétation du nuage peut donc reposer uniquement sur cet axe F1
comme nous l'avons fait.
Pour définir les différents pôles de l'axe
F1 par les variables qui leurs sont associées, nous avons
recherché celles qui contribuent le plus à ces axes. Ensuite,
parmi ces variables les modalités qui sont corrélées aux
axes ont été identifiées.
Ainsi, les variables les plus importantes (plus contributives)
pour cet axe sont la variable "effectif bovin du troupeau" (TaiTr) et la
variable "raisons de la pratique de la transhumance" (RTra) ; elles contribuent
toutes les deux à hauteur de19,40% dans la construction de F1.
En observant ensuite l'association ou la corrélation
entre les modalités des deux variables plus fortes contributrices et F1
(tableau V-8), on voit bien que les modalités RTra-5 (troupeau ne
transhumant pas) et TaiTr-1 (effectif bovin du troupeau inférieur
à 45 têtes de bovins) sont celles qui expliquent mieux l'axe
notamment son pôle négatif. Elles sont en effet, à la fois
fortes contributrices et bien corrélées à ce pôle.
Ces deux modalités sont par ailleurs proches sur le plan factoriel, ce
qui indique qu'elles sont associées entre elles et que des individus
exprimant l'un des deux caractères sont fortement susceptibles
d'exprimer l'autre.
134
Chapitre V. Les systèmes d'élevage
à Kotchari
Figure V-4. Graphique des variables de l'analyse factorielle des
correspondances multiples
Légende :
Les points et étiquettes en noir représentent les
variables actives ;
Les points et étiquettes en rouge représentent les
variables supplémentaires. TaiTr, effectif bovin du troupeau;
RLot, raison de l'allotement ;
RTra, raison de la transhumance ;
PAlt, pratiques alimentaires ;
NEsp, nombre et types d'espèces du troupeau ;
NRaB, nombre de races bovines du troupeau ;
LPat, Lieu de pâturage en saison sèche;
Eth, ethnie de l'éleveur ;
OrEl, origine de l'éleveur
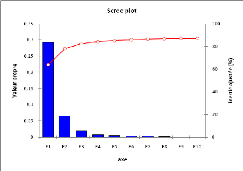
135
Figure V-5. Histogramme des valeurs propres Tableau V-8.
Modalités expliquant le plan factoriel F1-F2
|
|
|
Modalités
|
F1
|
|
F2
|
|
|
Contribution
|
Cosinus carré
|
Contribution
|
Cosinus carré
|
|
RTra-5
|
0,10
|
0,82
|
0,00
|
0,01
|
|
TaiTr-1
|
0,09
|
0,75
|
0,00
|
0,02
|
|
LPat-3
|
0,08
|
0,47
|
0,00
|
0,00
|
|
PAlt-1
|
0,08
|
0,44
|
0,05
|
0,16
|
|
NEsp-4
|
0,07
|
0,34
|
0,03
|
0,11
|
|
PAlt-3
|
0,06
|
0,38
|
0,02
|
0,08
|
|
RTra-3
|
0,06
|
0,38
|
0,00
|
0,00
|
|
LPat-1
|
0,03
|
0,23
|
0,08
|
0,34
|
Légende :
Modalités en gras, modalités à la fois
fortes contributrices et bien corrélées à F1
Pour le reste des modalités, modalités non
corrélées alors qu'elles contribuent bien à l'un ou
à l'autre des axes ou aux deux axes à la fois.
RTra, raison de la transhumance ;
TaiTr, effectif bovin du troupeau;
LPat, Lieu de pâturage en saison sèche; PAlt,
pratiques alimentaires ;
NEsp, nombre et types d'espèces du troupeau ;
Par ailleurs, si on considère ces modalités une
à une et, en s'appuyant sur le fait que deux modalités s'opposent
lorsque l'angle formé entre elles, à partir de l'origine de
l'axe, est supérieur à 90° (l'opposition parfaite est
à 180°), et s'attirent lorsque cet angle est inférieur
à 90 ° (lien parfait à 0°) (Cibois, 2007), on peut
faire plusieurs déductions.
- La modalité Taitr-1 (plus petit
effectif bovin dans le troupeau) qui est liée au pôle
négatif du facteur F1, s'oppose aux modalités TaiTr-2, TaiTr-3,
TaiTr-4 et TaiTr-5. Cette

opposition est très nette avec les plus grands effectifs
bovins dans le troupeau (TaiTr-4 à TaiTr-5).
- La modalité RTra-5 (troupeau ne
transhumant pas), elle aussi négativement corrélée
à F1, s'oppose nettement (RTra-4) à très nettement
(RTra-3, RTra-2, RTra-1) aux modalités qui décrivent des
troupeaux en transhumance qui, elles, sont associées au
côté positif de l'axe.
A la lumière de ces constats, on peut déduire
que le facteur F1 exprime la taille et le degré de mobilité des
troupeaux. L'extrémité de son pôle négatif exprime
des troupeaux peu mobiles (RTra-5; ne transhument pas) et aux petits effectifs
bovins (TaiTr-1 ; nombre de bovins inférieur à 50 têtes)
tandis que celle du pôle positif est significative de troupeaux aux
grands effectifs bovins (TaiTr-4 à TaiTr-5 ; nombre de bovins
supérieur à 135 têtes) pratiquant la transhumance. Entre
les deux pôles, la situation est assez diverse, elle exprime des cas
intermédiaires entre les deux cas extrêmes.
5.3.3.2.3. Les catégories
d'éleveurs
Le comportement des modalités, leur caractère
ordonné le long de l'axe F1, qui est nettement
prépondérant indique que ce facteur F1 semble bien structurer
l'information en opposant les groupes d'éleveurs les plus
différents. L'axe F2, comme indiqué plus haut, semble par contre
peu informatif et ne permet pas d'expliquer notablement l'organisation des
variables et, en conséquence le comportement des éleveurs.
En conclusion, l'axe F1 distingue un premier groupe
d'éleveurs (pôle négatif de l'axe) ayant les
caractéristiques suivantes:
- éleveurs qui ne transhument pas (RTra-5),
- éleveur à troupeaux à effectif bovin de
petite taille (TaiTr-1 : 0 à 45 têtes)
Le groupe le plus opposé à ce premier groupe
(extrémité positive de l'axe) regroupe les éleveurs qui
:
- ont des troupeaux à effectif bovin de grande taille
généralement supérieure à 135 têtes (TaiTr-4
et TaiTr-5),
- pratiquent la transhumance pour diverses raisons notamment la
recherche de l'eau et du fourrage à la fois (RTra-3) ou seulement de
l'eau (RTra-1).
Les autres éleveurs se retrouvent dans une situation
relativement diversifiée plutôt intermédiaire par rapport
à ces deux groupes, ils se regroupent autour de l'origine du plan
factoriel. Dans cet ensemble d'éleveurs dont la taille du troupeau va de
46 à 135 têtes (TaiTr2 à TaiTr-3), la transhumance est
pratiquée et motivée surtout par la recherche de l'eau et du
fourrage (RTra-3 ; Rtra-1) mais aussi, pour quelques éleveurs, par
héritage culturel (RTra-4).
Par ailleurs, le positionnement dans le nuage de points des
modalités des variables supplémentaires Eth et OrEl (figure V-4)
est assez significatif et permet de donner une identité à ces
groupes d'éleveurs (figure V-6). Ainsi donc, les éleveurs du
groupe lié à la partie négative extrême de l'axe F1
sont des éleveurs résidents (OrEl-1), ils peuvent être
d'ethnie gourmantché (Eth-1) ou peule (Eth-2) ; ceux du groupe
lié à l'autre extrémité de F1 (pôle positif)
sont des non résidents venant d'horizons divers (autres zones du Burkina
: OrEl-
137
2 et OrEl-3; Niger : OrEl-4) de l'ethnie peule (Eth-2). Le groupe
intermédiaire comprend des Peuls (Eth-2) résidents (OrEl-1).
L'analyse hiérarchique (figure V-7) après une
classification k-means des éleveurs à partir de leurs
coordonnées sur les facteurs F1 et F2 (résultats de l'AFCM),
confirme ces comportements et le recodage a permis d'affecter chacun des
éleveurs dans les classes ainsi constituées : la catégorie
C1 qui comprend 98 éleveurs résidents d'ethnie gourmantché
(50 éleveurs) et peule (48 éleveurs), la catégorie qui
comprend des peuls essentiellement résidents pratiquant la transhumance
(51 éleveurs) et la catégorie C3 constituée de 51
éleveurs peuls essentiellement allochtones et transhumants (48
éleveurs).
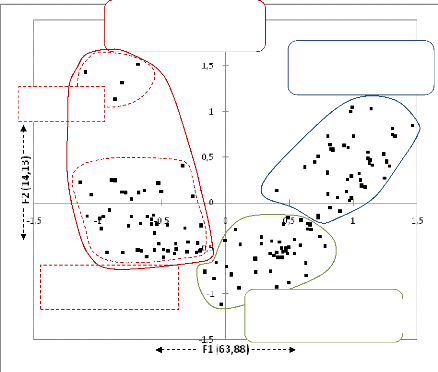
Sous-catégorie C1-1 - Gourmantché
;
- Pas de bovins
Sous-catégorie C1-2
- Peul et Gourmantché ;
- Présence de bovins aux effectifs inférieurs
à 46 têtes
Catégorie 1 (C1)
- Éleveurs résidents et sédentaires -
Effectif bovin inférieur à 46 têtes - Non transhumants
Catégorie 2 ()
- Éleveurs peul résidents
- Effectif bovin entre 46 et 135 têtes - Transhumants
Catégorie 3 (C3)
- Éleveurs peul allochtones - Effectif bovin > 135
têtes - Transhumants
Figure V-6. Graphique de projection des éleveurs (analyse
factorielle des correspondances multiples) montrant leur répartition en
classes.
Figure V-7. Dendrogramme des groupes d'éleveurs
5.3.3.2.4. Caractéristiques des catégories
d'éleveurs
Les différents groupes d'éleveurs ainsi
constitués, nous avons fait appel aux autres modalités pour mieux
les caractériser, en tenant toujours compte des corrélations qui
existent entres les nouvelles modalités et celles qui ont permis de
séparer les groupes.
Le groupe C1 : constitué de 98 éleveurs
résidents non transhumants d'ethnie gourmantché (50
éleveurs) et peule (48 éleveurs), est un groupe dans lequel la
transhumance n'est pas pratiquée (figure V-6). Les troupeaux, quand ils
comprennent des bovins, ceux-ci sont à effectif faible (au plus 45
têtes). Ils sont majoritairement plurispécifiques (48% sont
formés à la fois de bovins, ovins et caprins) et comprennent
presque autant de bovins (19 têtes en moyenne) que d'ovins (17
têtes en moyenne) avec un effectif caprin moyen (dix têtes) le plus
élevé par rapport aux deux autres catégories
d'éleveurs (tableau V-9). Le sex-ratio (0,54) dans l'effectif bovin
indique qu'il y a plus de vaches que de taureaux (un peu plus de 1 taureau pour
2 vaches). Par ailleurs, ces éleveurs exploitent en saison sèche
le terroir de Kotchari ainsi que les terroirs avoisinant et le
complément fourrager distribué aux animaux en cette saison sont
issus des résidus de culture (tiges de sorgho, diverses fanes notamment
d'arachide et de niébé). Il faut noter que ce groupe
d'éleveurs, plus que les deux autres groupes suivants, a comme
activité principale l'agriculture mais l'apport de l'élevage dans
l'économie des ménages peut être considérable
surtout dans les ménages peuls.
L'analyse du comportement de ce groupe montre qu'il est assez
diversifié, ce qui autorise à le séparer en deux
sous-groupes. Pour ce faire, nous avons repris l'ACH en fixant cette fois la
troncature (nombre attendu de classes) à quatre, ce qui a permis de
reclasser les éleveurs du groupe dans deux sous-groupes. On a ainsi, la
sous-catégorie C1-1 qui comprend seulement quinze éleveurs tous
d'ethnie gourmantché et dont le troupeau ne comprend que des ovins et
des caprins exclusivement complémentés en saison sèche par
du fourrage cultivé. Cette souscatégorie d'éleveurs dont
les troupeaux sont conduits par de jeunes bergers familiaux (un berger par
troupeau) n'exploite que les pâturages du terroir de Kotchari en saison
sèche.
La sous-catégorie C1-2, quant à elle, regroupe le
reste des éleveurs de ce groupe (83 éleveurs
dont 48 Peuls et
35 Gourmantchés) chez lesquels l'espèce bovine est
rencontrée dans les

139
troupeaux ; qui sont, par ailleurs, complémentés
à la fois en fourrage cultivé surtout mais aussi naturel
collecté et mis en stock. En saison sèche, les troupeaux de ce
groupe peuvent exploiter à la fois les pâturages du terroir de
Kotchari ainsi que ceux voisins.
Le groupe : catégorie d'éleveurs peuls
(51 éleveurs) résidents et transhumants dont l'effectif bovin du
troupeau oscille entre 45 et 135 têtes avec une moyenne de 88 têtes
(tableau V-9). Ici, l'effectif ovin (moyenne : 20 têtes) est
statistiquement le même que celui de la catégorie
d'éleveurs précédente (C1) et nettement plus
élevé que dans les troupeaux du groupe suivant (C3). En
général, les troupeaux comprennent soit l'espèce bovine
associée aux ovins (51%) ou les trois espèces (bovines, ovines et
caprines) (35%). L'espèce bovine comprend plusieurs races notamment les
associations des trois races courantes (Barbaji + Gurmaji et
Gurmaji + Jaliji) ou de ces deux races associées à
d'autres races moins importantes. Le sex-ratio (0,36), montre que ces troupeaux
se composent d'à peu près 1 mâle pour 3 femelles. C'est un
élevage à tendance « naisseur » dans lequel les vaches
semblent nettement privilégiées. Ici, l'allotement
majoritairement à deux lots (68%) se pratique avec comme motivations
premières l'allègement des charges sur les parcours (45%) et la
gestion des risques éventuels (43%). Ces éleveurs qui
transhument, sont, en saison sèche présents sur les
pâturages des pays voisins (53%) mais certains (16%) reconnaissent
fréquenter les aires protégées voisines. Enfin, pour
complémenter leurs animaux en saison sèche, ces éleveurs
font appel à diverses sources alimentaires (mise en stock de fourrage
naturel et cultivé, achat de concentré). Les bergers sont
essentiellement de la famille des propriétaires mais quelques troupeaux
(4%) associent à ces bergers des salariés. Au sein de ce groupe
les bergers peuvent être solitaires (39%), ou en binôme (33%) ou
même plus (27%) pour les plus grands effectifs.
Le groupe C3 : ensemble de 51 éleveurs peuls
transhumants allochtones (non résidents, ils viennent des terroirs et
départements voisins, des provinces plus ou moins voisines notamment le
Yagha et du Niger et seuls trois d'entre eux sont résidents de
Kotchari). Se consacrant presque exclusivement à l'élevage
(l'activité agricole est marginale), les effectifs bovins de leurs
troupeaux sont assez importants allant de 135 à plus de 200 têtes.
L'essentiel du bétail ici est donc constitué de l'espèce
bovine (effectif moyen : 135 têtes) avec une quasi absence de caprins
(effectif moyen : 0 têtes) (tableau V-9). Le sex-ratio (0,48) (moins de
un mâle pour deux femelles) statistiquement identique à celui
observé dans C1, permet de dire que les troupeaux de cette
catégorie ont également une tendance « naisseur » mais
moindre que précédemment (). Les races bovines du troupeau sont
la Gurmaji (Éleveurs du Burkina) (41%) ou la Jaliji
(éleveurs du Niger surtout) (29%) avec une forte combinaison des deux
(24%). Ces éleveurs qui sont généralement de passage pour
les pays voisins (Bénin et Togo), transhument à la rencontre de
l'eau, du fourrage ou des deux à la fois. La complémentation,
quand elle existe, est basée sur l'achat de concentré, la grande
taille des troupeaux ne permettant pas d'acheter suffisamment du fourrage
complémentaire. Notons qu'à cause justement de cette grande
taille, les troupeaux sont toujours subdivisés en deux lots (57%) ou en
trois lots (43%).

Tableau V-9. Caractéristiques des élevages dans le
terroir de Kotchari (Données d'enquête)
Effectifs moyens des espèces de
ruminants
|
Groupes d'éleveurs
dans le
terroir
|
Bovins
|
Ovins
|
Caprins
|
Sex-ratio
(mâles/femelles)
|
|
C1 (n = 98)
|
19,12
|
#177; 16,69a
|
17,39
|
#177; 10,75b
|
9,68
|
#177; 9,77c
|
0,54
|
#177; 0,21b
|
|
(n = 51)
|
87,84
|
#177; 36,77b
|
19,90
|
#177; 11,90b
|
4,20
|
#177; 5,31b
|
0,36
|
#177; 0,12a
|
|
C3 (n = 51)
|
174,71
|
#177; 46,34c
|
10,18
|
#177; 13,78a
|
0,12
|
#177; 0,84a
|
0,48
|
#177; 0,11b
|
Les valeurs situées sur la même colonne et portant
des lettres distinctes sont significativement différentes au seuil
á = 0,05 à p = 0,001 ; n, nombre d'éleveurs de la
catégorie.
5.3.4. Les pratiques pastorales : évolution depuis
20 ans
L'approche diachronique adoptée ici privilégie
les tendances d'ordre général, cependant, nous accordons de
l'importance aux variations mineures, étant entendu que tout changement
aussi minime soit-il n'est pas anodin, il peut être
révélateur d'ajustements plus ou moins importants au sein des
élevages ou du groupe d'élevages. Rappelons également que
nous ne perdons pas à l'esprit que la période de
référence choisie peut être insuffisante pour percevoir de
changements notables ou pour permettre la claire lisibilité d'une
quelconque tendance.
Pour aborder la question de l'évolution du comportement
général des éleveurs présents dans le terroir ou le
fréquentant nous nous appuierons, sans les reprendre, sur les
données générales présentées dans le tableau
V7 (paragraphe 5.3.3.1).
5 .3.4.1. Constitution et allotement des troupeaux
En rappel (voir paragraphe 5.3.3.1), les formes courantes de
mise en place de troupeau ou d'acquisition d'animaux sont l'héritage
(Sendereeji bibe et Sukkadi)63
associé ou non à l'achat. Presque tous les troupeaux
enquêtés ont, en effet, été constitués par
ces voies seulement ou associées à d'autres, plus marginales,
comme les emprunts ou les dons. L'acquisition par héritage est propre
aux éleveurs transhumants résidents () ou non (C3), tandis que
celle par achat domine chez les éleveurs résidents non
transhumants (C1). Dans cette dernière catégorie cependant, un
grand nombre d'éleveurs, ceux surtout de C1-2 dominés par des
peuls, acquièrent leurs animaux par héritage. L'acquisition par
achat dans le
63 Ces deux pratiques d'héritage uniquement
présentes chez les Peul, ont déjà été
rencontrées par Thébaud (1999) respectivement en pays Woodabe du
Niger (Diffa) et au Yagha burkinabè. Sendereeji bibe fait
référence à une forme de constitution d'un troupeau
embryonnaire dans laquelle les petits garçons de la famille
reçoivent une dotation d'animaux en pré héritage. Dans
notre échantillon la pratique est citée par les transhumants
nigériens.
Sukkadi ou Sukkaaji ou encore
halalji (Boutrais, 2002) est une pratique dans laquelle tout nouveau
né reçoit automatiquement en donation un animal en
héritage du vivant de son ascendant. C'est une pratique
rencontrée chez les éleveurs peul du terroir qu'ils soient
transhumants ou non. Les bovins Sukkadi constituent un patrimoine qui
sert de lien intergénérationnel dans les familles pastorales
peules.
groupe est surtout rencontrée chez les
Gourmantchés du groupe C1 (le sous-groupe C1-1 et une bonne partie du
sous-groupe C1-2).
La manière dont on acquiert les animaux dans le terroir
de Kotchari a été étudiée et expliquée en
détail par Ouédraogo (2008). Les faits qu'il a observés
correspondent aux normes culturelles en vigueur dans les groupes socioculturels
qui caractérisent les 3 groupes d'éleveurs que nous avons
reconnus. En effet, chez les Peuls on obtient généralement son
troupeau par héritage car « dans la tradition peule, un enfant
a droit à un animal le jour de son baptême ». Chez les
agropasteurs gourmantchés (le groupe C1-1 et une bonne partie du groupe
C1-2), les animaux sont acquis avec les ressources issues de l'agriculture.
Notons que l'emprunt d'animaux est aussi rencontré dans ces
élevages, en particulier ceux de C1-2, il se résume souvent
à quelques boeufs de trait.
De manière générale, le cheptel local
notamment bovin s'est accru depuis la décennie 1990. Chez les non
transhumants (C1) (figure V-8a), la taille des troupeaux est restée
faible après avoir connu une hausse passagère. Cette hausse
notable mais momentanée des effectifs est liée au boom cotonnier
enregistré dans la zone dans la première moitié de la
décennie 2000. Comme nous l'avons déjà signifié
plus haut, les Gourmantchés de la zone se sont beaucoup adonnés
à la culture du coton depuis la relance de cette activité en 1997
dans la zone et les ressources qui sont générées sont
capitalisées dans l'élevage qui joue ainsi le rôle de
« banque traditionnelle » confirmant ainsi les observations de Lhoste
et al. (1993). La situation, du reste, n'est pas spécifique
à Kotchari ; un peu partout en zone soudanienne, les agriculteurs
thésaurisent leurs revenus dans l'achat du bétail. Dans les
autres groupes d'éleveurs (figures V-8b et V-8c), on constate que,
durant la période de référence, l'augmentation des
effectifs du cheptel s'est effectuée par l'accroissement du nombre de
troupeaux de taille intermédiaire (TaiTr-2, TaiTr-3) ou
supérieure (TaiTr-3, TaiTr-4 et TaiTr5) respectivement pour et C3 et au
détriment des effectifs en troupeaux de taille inférieure
(TaiTr-1 pour ; TaiTr-1 et TaiTr-2 pour C3). Il faut noter que les troupeaux
de très grande taille (TaiTr-5) n'existent chez les éleveurs
résidents transhumants () que depuis quelques années (on n'en
rencontrait pas il y a seulement une dizaine d'années). Il faut
comprendre que la plupart des éleveurs peuls résidents qu'ils
soient transhumants () ou non transhumants (une partie des éleveurs de
C1-2), sont dans une phase de reconstitution de leur cheptel. La plupart
s'étaient sédentarisés après avoir perdu, suite aux
grandes sécheresses, une grande partie de leurs animaux.
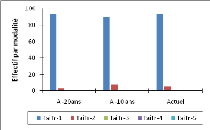
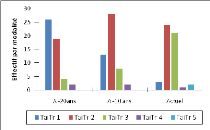
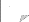
141
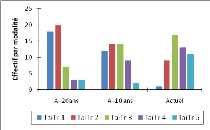
c. Groupe C3
Figure V-8. Évolution des effectifs bovins des troupeaux
enquêtés
Légende:
TaiTr-1 : 0-45 têtes ; TaiTr-2 : 45-90 têtes ;
TaiTr-3 : 90-135 têtes ; TaiTr-4 : 135-180 têtes ; TaiTr-5 : plus
de
180 têtes
Si la hausse du cheptel est certaine de manière globale
pour chacune des catégories d'éleveurs, une analyse plus fine
permet de voir cependant que certains éleveurs (31%, tous groupes
confondus), ont vu la taille de leur troupeau diminuer, parfois notablement.
Des constatations assez différentes mais nuancées sont faites par
Kièma S. (2007) dans l'Ouest burkinabè. Cet auteur indique qu'au
sein des éleveurs qu'il a enquêtés, seulement 38,5% d'entre
eux ont vu leurs troupeaux s'accroître ces dernières années
alors que, contrairement à nos observations à Kotchari, il a
noté concomitamment un recul de l'effectif du cheptel global.
L'allotement (constitution de sous-groupes dans le troupeau) est
une pratique de plus en plus courante, en particulier chez les éleveurs
transhumants ( et C3).
Chez les non transhumants (C1) (figure V-9a), l'allotement
était une pratique marginale il y a une vingtaine d'années, elle
implique cependant de plus en plus d'éleveurs de cette catégorie
depuis une dizaine d'années. Ici, la subdivision du troupeau en 3 lots
n'est cependant qu'à son début. Les motifs
généralement évoqués par ces éleveurs pour
expliquer la division de leurs troupeaux en sous-lots étaient à
l'origine de faciliter la tâche des bergers (RLot-4) ; mais depuis une
dizaine d'années d'autres arguments (avoir des lots homogènes :
RLot-1, gérer d'éventuels risques : RLot-2 et gérer les
charges sur les parcours : RLot-3) de plus en plus importants sont
avancés (figure V-9b) cela du fait surtout d'éleveurs
(généralement des éleveurs peuls dont les troupeaux sont
en reconstitution) qui se sont nouvellement mis à la pratique.
L'argument majeur demeure tout de même la constitution de lots
gérables par les bergers (RLot-4).
Chez les transhumants ( et C3), les troupeaux ont toujours
été subdivisés en plusieurs lots ; mais alors que la forme
à trois lots n'a commencé à être connue qu'il ya une
dizaine d'années chez les résidents () (figure 10a), elle est
rencontrée il ya au moins une vingtaine d'années chez les
allochtones (C3) (figure 11a). De toutes les catégories
d'éleveurs, c'est ici que les troupeaux à trois lots sont les
plus importants. L'allotement, chez les éleveurs de type se justifiait,
il y a une vingtaine d'années, surtout par le souci d'avoir des lots
homogènes (RLot-
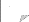
1), mais une proportion notable d'éleveurs
étaient, eux, préoccupés par la bonne gestion
d'éventuels risques (RLot-2) (figures V-10b) ou par la bonne
répartition des charges animales sur les parcours (RLot-3). Depuis, les
deux derniers arguments (RLot-2 et RLot-3) ont pris régulièrement
de l'importance au détriment du premier. Depuis une dizaine
d'années, s'y ajoute le souci d'alléger les charges des bergers
par constitution de troupeaux de taille raisonnable. Dans la catégorie
d'éleveurs C3, la constitution de lots homogènes (RLot-1), ainsi
que, dans une moindre mesure, la gestion des risques potentiels (RLot-2) sont
des motifs constants (figure V-11b). Depuis un certain temps, un nombre plus
important de ces éleveurs, qui font face de plus en plus à
l'adversité, se préoccupent prioritairement de la gestion des
risques éventuels qui sont susceptibles de se présenter à
eux lors de leur transhumance. Toutefois, avoir des lots homogènes reste
le motif premier évoqué.
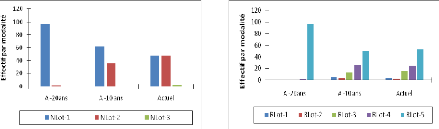
a. Allotement du troupeau b. Motivations
Figure V-9. . Évolution des pratiques d'allotement du
troupeau dans le Groupe C1
NLot-1: 1 lot Motivations
RLot-1: Séparer en lots homogènes
Légende: Allotement
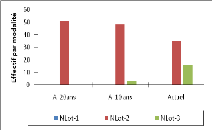
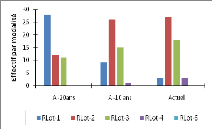
NLot-2: 2 lots
NLot-3: plus de 2 lots RLot-2: Gérer les risques
RLot-3 : Gérer charges sur parcours RLot-4 : Faciliter
tâche des bergers RLot-5 : N'est pas concerné
143
a. Allotement du troupeau b. Motivations
Figure V-10. . Évolution des pratiques d'allotement du
troupeau dans le Groupe
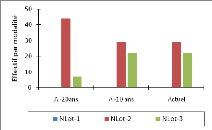
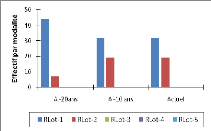
a. Allotement du troupeau b. Motivations
Figure V-11. Évolution des pratiques d'allotement du
troupeau dans le Groupe C3
L'allotement est une pratique courante au sein des
élevages peuls ou mobiles en général (Kagoné, 2000
; Riegel, 2002 ; Paris, 2002 ; Botoni, 2003 ; Kièma S., 2007;
Ouédraogo, 2008), mais il a pris de l'ampleur face aux contraintes que
rencontre de nos jours le bétail. Il est par ailleurs, fortement
dépendant de la taille du troupeau de l'éleveur (Kagoné,
2000). Chez les agroéleveurs gourmantchés (ils
représentent une bonne partie des éleveurs de C1),
l'intérêt de diviser le troupeau en lots est bien perçu
mais la pratique n'est pas systématique même lorsque le troupeau
dépasse la cinquantaine de têtes. Nos données indiquent en
effet, qu'en dessous de 49 têtes, aucun des troupeaux ne compte plus d'un
berger à la fois. Cet intérêt reconnu à la pratique
se confronte cependant à un manque de main d'oeuvre prioritairement
consacré en général aux travaux champêtres, ces
agroéleveurs étant fortement impliqués dans la culture du
coton. La configuration des lots dépend de la composition du troupeau,
mais ils sont en général hétérogènes : s'il
y a un équilibre numérique entre bovins et petits ruminants
(ovins et/ou caprins), chacun des deux lots sont ainsi constitués : lot
un : bovins ; lot deux : ovins et/ou caprins. Si par contre il ya un grand
déséquilibre, les lots constitués vont comprendre à
la fois un peu de chaque espèce, ils demeurent cependant
équilibrés en taille. Cette façon de faire est rendue
possible par le fait que bien des fois, ces troupeaux sont
hétérogènes à l'origine, chacun des membres de la
famille y apportant quelques têtes.
Il semble que pour les éleveurs qui sont
concernés par l'allotement, ce ne sont pas les mêmes contraintes
qui commandent la pratique selon que l'on appartient à un groupe
d'éleveurs ou à un autre. Chez ce type C1, les éleveurs
sont surtout soucieux d'assurer une meilleure surveillance de leurs troupeaux,
devenus de plus en plus importants, par des bergers peu spécialistes en
témoigne le propos de cet éleveur (Tankoano Nindia) du village
Malipoa « Avec le coton, nous n'avons que l'élevage pour mettre
de côté l'argent que nous gagnons, or nos enfants sont peu
habitués à suivre de gros troupeaux et les risques qu'ils
commettent des dégâts sont importants. Diviser le troupeau en lots
de petite taille apparait comme une obligation, mais elle se confronte souvent
au manque de bergers, nos enfants nous étant très utiles au champ
».
Chez les Peuls de toutes les catégories, l'allotement
est systématique (généralement deux
lots : bien
portants vs malades et allaitants) quelle que soit la taille du troupeau. Il
devient
fortement dépendant de la taille du troupeau lorsque celui-ci
tend à dépasser 150 têtes (aucun

145

troupeau de taille inférieure ne
bénéficie de plus de deux bergers). Au-delà de cet
effectif, la plupart des troupeaux sont subdivisés en trois sous-lots
(troupeau d'allaitants et/ou de malades, troupeau de veaux et troupeau de bien
portants) et ce cas de figure est plutôt courant chez les éleveurs
transhumants ( & C3). Dans ces catégories, les éleveurs sont
soucieux de faciliter leurs déplacements (constitution de lots
homogènes) ou de le sécuriser (gestion des risques), eux qui sont
très souvent partis vers l'inconnue ou l'incertain (Benoit, 1979 ;
Landais, 1990). Cette subdivision du troupeau, qui se produit
généralement en saison sèche comme l'a aussi
observé Kièma S. (2001 & 2007), notamment lors de la campagne
de transhumance (Kagoné, 2000), généralement
précédée d'une organisation minutieuse (Toutain et
al., 2001), répond, selon Riegel (2002) et Paris (2002), ainsi
que quelques transhumants interrogés, à une stratégie
préventive : épargner aux animaux fatigués, gestants,
malades ou trop jeunes de dures conditions de marche et dont la présence
retarderait, par ailleurs, l'avancée du troupeau et l'exposerait
énormément à divers risques; éviter les pertes en
chemin dues aux attaques de prédateurs. On peut penser aussi comme
Convers (2002) à une stratégie de répartition des risques
surtout qu'un des éleveurs du groupe (Diadié Diallo, transhumant
nigérien du département de Say), questionné, nous a tenu
le propos suivant « nous faisons attention à ne pas mettre tous
nos oeufs dans le même panier. Lorsque nous devons aller loin, nous
laissons une partie de nos animaux à la maison ». Certains
transhumants évoquent l'idée de permettre à une partie de
la famille restée sur place de bénéficier du lait des
vaches allaitantes. L'argument est aussi avancé par ce pasteur,
transhumant habituellement vers la Kompienga et le Togo, interrogé par
Kaboré (2010) « ... Nous pâturons au Togo en passant par
la Kompienga (...) On n'envoie jamais tout le troupeau pour qu'il y ait du lait
pour ceux qui restent à la maison... ». Bary (1998), à
la suite d'ateliers organisés avec des éleveurs de Gorom-Gorom,
Djibo, Solenzo et Fada N'Gourma, rend compte du fait que lors du départ
en transhumance, les vaches allaitantes et les animaux faibles sont
laissés sur place. Ce constat est également fait par Dongmo et
al. (2007), Harchies et al. (2007) ainsi que Kossoumna Liba'a
et al. (2010), ces auteurs rapportent que les éleveurs
transhument avec un lot principal et laissent en place un lot constitué
de vaches allaitantes. Dans l'échantillon d'éleveurs que nous
avons enquêtés, certains de ceux qui répartissent leur
troupeau en plus de deux lots (trois en général), ont
évoqué ce dernier argument comme motif principal. L'argument peut
être pertinent quand on fait un lien avec la composition en genre du
troupeau. En effet, on constate (tableau V-9) que, par rapport au groupe
constitué d'éleveurs peuls transhumants résidents, les
troupeaux des transhumants non résidents (C3), comprennent beaucoup
moins de femelles que de mâles (les sex-ratios sont de 0,36 et 0,48
respectivement). Or, il est connu que le caractère naisseur, qui est une
des marques des élevages mobiles notamment peuls (Doutressoulle, 1947 ;
Veyret, 1951 ; Sere & Steinfeld, 1996 ; Sounkéré, 2003 ;
Kièma S., 2007), se traduit dans le troupeau par un sex-ratio
très bas, c'est-à-dire un nombre très faible de taureaux
par rapport aux vaches. Sur cette base on penser que, dans les troupeaux de ce
groupe d'éleveurs, aux caractéristiques assez voisines de ,
quelques femelles sont restées en terroirs d'attache au moment du
départ des troupeaux. Nous revenons plus bas sur cette question
spécifique du sex-ratio dans les troupeaux des groupes d'éleveurs
dans le paragraphe sur les pratiques de diversification.
Si les éleveurs non transhumants (C1), dont certains
ont abandonné la transhumance ou ne pratiquent plus que la petite
transhumance64, sont préoccupés, comme nous l'avons
dit, par la capacité des bergers à bien gérer leur
troupeaux pour prévenir les éventuelles pertes et surtout les
dégâts qu'ils pourraient être amenés à
commettre, argument les emmenant à repartir leurs troupeaux en lots plus
réduits, ils sont aussi assez regardants sur la question de la
préservation des ressources locales. Ils pensent comme Lassissi
(éleveur peul sédentaire, quartier Kobana) que « de
petites têtes partout procurent plus de santé aux parcours et
préservent plus longtemps le fourrage que plusieurs têtes en un
seul lieu ». Cet argument, assez pertinent du point de vue des
spécialistes de l'écologie des parcours ou des gestionnaires des
ressources (Breman & De Ridder, 1991 ; César, 1992 ; Lhoste et
al. 1993 ; Boutrais, 1997 ; Scoones, 1999 ; Nori, 2007 ; Nori et
al. 2008), est peu ou pas évoqué par les transhumants
allochtones (éleveurs du groupe C3), alors que les transhumants
résidents, tout en en tenant compte de plus en plus, le relèguent
au second plan. L'explication à cela peut se retrouver dans ce propos de
Sadio Sondé (transhumant nigérien venant de Makalondi) «
chez nous, nous ne perdons pas de vue que lorsque les animaux sont en grand
nombre en un même lieu, ils perdent en embonpoint et ce milieu est perdu
pour longtemps. Mais lorsque nous devons aller loin, nous sommes plutôt
préoccupés à nous assurer que des conditions de voyage
moins périlleuses sont réunies ». Notons enfin que les
lots d'animaux malades (ou fatigués ou jeunes) restés en terroir
d'attache sont parfois amenés à aller loin dans les terroirs
voisins (cas de petite transhumance) suivant que la saison pluvieuse
précédente a été bonne ou mauvaise.
5.3.4.2. Garde des troupeaux
Dans des travaux antérieurs (Sawadogo, 2004), nous
avons montré que les animaux étaient conduits différemment
selon que l'on était en saison sèche où l'espace
était ouvert et de libre accès ou en saison humide, saison
agricole par excellence, où les parcours étaient
fragmentés et les risques de conflits accrus. En saison sèche, si
les bovins sont surveillés, les petits ruminants sont laissés en
libre pâture. En saison pluvieuse en revanche, tous les animaux sont sous
la surveillance de bergers. Ceci mérite cependant d'être
nuancé car Ouédraogo (2008) a observé que, de nos jours
à Kotchari, certains éleveurs faisaient surveiller leurs moutons
et leurs caprins même en saison sèche surtout lorsqu'ils
étaient proches des réserves, en effet, ils risquent à
tout moment d'y pénétrer. Des différences sensibles dans
la garde des animaux sont observées selon le groupe ethnique. Ainsi les
animaux sont abreuvés une seule fois par jour vers midi chez les
Gourmantchés, tandis que les Peuls les abreuvent dès les
premières heures de la matinée au sortir du parc ou campement de
nuit puis à nouveau vers 13 ou 14 heures. La pâture de nuit,
reconnue comme très bénéfique car moins épuisante
pour le bétail, est propre aux éleveurs peuls, ce sont en
général les bouviers les plus âgés et les plus
expérimentés qui s'en chargent.
Chez les éleveurs non transhumants (C1), lorsque le
troupeau ne comporte pas de bovins, le gardiennage est assuré par des
mineurs (garçons et filles) de huit à dix ans. Par
64 Parmi ces éleveurs, certains
délocalisent leurs troupeaux dans la partie sud plus humide du terroir.
Ceci s'apparente à de la petite transhumance (Yawtooru en
langue peule), nous avons cependant confondu ces éleveurs aux non
transhumants car cela se passe dans l'espace du terroir.
contre, dès que des bovins sont présents dans le
troupeau, sa garde relève de la responsabilité de jeunes
adolescents d'un âge dépassant généralement douze
ans (Sawadogo, 2004). Aujourd'hui, on dénombre un berger par troupeau
(NBe-1), alors qu'il y a une dizaine d'années, correspondant alors au du
boom cotonnier, ce nombre était passé à deux (NBe-2) pour
les troupeaux de quelques uns de ces éleveurs (figure V-12a). Depuis
lors, on observe une tendance au retour à 1 seul berger pour ces
troupeaux. Parallèlement, quelques élevages ont, de plus en plus,
depuis un certain temps, recours à des bergers salariés seuls
(confiage ou employé) à qui on associe désormais des
bergers internes aux exploitations (figure V-12b). On peut faire
l'hypothèse que si l'expérience qui consiste à utiliser
plusieurs bergers ou des salariés exclusivement est en train de tourner
court, c'est que quelques troupeaux locaux (troupeaux gourmantchés
notamment), qui avaient gagné en taille du fait de l'explosion de la
culture cotonnière, ont depuis diminué en effectif avec le recul
"momentané" de cette dernière. En effet, comme signalé
plus haut (voir paragraphe 5.3.3.2.1) ou comme nous le détaillons dans
les parties qui suivent, le nombre de bergers est fortement dépendant de
la taille du troupeau.
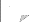
147
a. Nombre de bergers b. Types de bergers
Figure V-12. La garde des animaux et son évolution dans le
groupe C1
Légende:
NBe-1 : un seul berger NatBe-1 : propriétaire ou proche
parent (fils, neveu, etc.)
NBe-2 : deux bergers NatBe-2 : berger salarié (berger
employé ou animaux confiés)
NBe-3 : plus de deux bergers NatBe-3 : les deux à la fois
(parent et salarié)
Chez les transhumants résidents (), la garde des
animaux est réservée le plus souvent à des adolescents et
jeunes adultes (12 à 20 ans) et même à des adultes pouvant
être le propriétaire lui-même (20 à 40 ans). On voit
ici que, du fait de l'accroissement en taille du cheptel, le recours à
des bergers supplémentaires (d'abord un puis deux) est constant au cours
de la période de référence (figure V-13a) mais ces
bergers, dans les cas où ils sont des salariés de l'exploitation,
viennent toujours en appui à un membre de la famille du
propriétaire (figure V-13b).
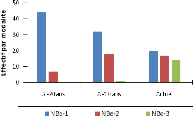
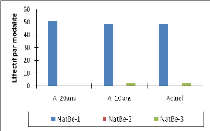
a. Nombre de bergers b. Types de bergers
Figure V-13. La garde des animaux et son évolution dans le
groupe
Chez les transhumants étrangers au terroir de Kotchari
(C3), les troupeaux sont depuis bien longtemps conduits par au moins deux
bergers avec une tendance à l'augmentation du nombre de ceux-ci (figure
V-14a). Ces bergers qui sont généralement les
propriétaires des troupeaux, se voient cependant associer des
salariés ces dernières années. Certains éleveurs du
groupe, la plupart de ceux qui utilisent la main d'oeuvre extérieure de
nos jours, ont dans un passé lointain eu recours exclusivement à
des bergers salariés mais ont dû mettre fin à cette
expérience suite, selon leur dire, à diverses insatisfactions
(vols, dépenses excessives) (figure V-14b).
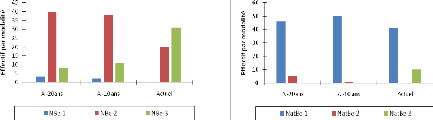
a. Nombre de bergers b. Types de bergers
Figure V-14. La garde des animaux et son évolution dans le
groupe C3
On peut remarquer le recul du confiage et du salariat dans les
élevages de ce terroir là où Botoni (2003), dans ses
travaux à l'Ouest (Ouara, Torokoro) a noté plutôt un
engouement vers ces pratiques de gardiennage, en particulier le confiage.
Diverses explications sont données par les éleveurs pour
expliquer ce recul, surtout la garde des troupeaux exclusivement par des
personnes étrangères à leurs exploitations. Deux arguments
majeurs reviennent : (i) l'insatisfaction dans la prestation du berger
salarié au regard des coûts de plus en plus insupportables que
cela nécessite ; (ii) la perte de confiance entre les parties
contractantes.

149
Les insatisfactions résultent, selon les
éleveurs, des faibles performances de gardiennage des bergers, elles
seraient liées à une négligence sélective envers
les animaux confiés. Ainsi, d'après Combary Tadjoa
(éleveur résident gourmantché du quartier Tambouli) «
depuis maintenant quelques années, les bergers que nous sollicitons
ne nous donnent plus satisfaction. Il est facile de reconnaître dans un
troupeau mixte, nos animaux des leurs ; leurs animaux sont mieux
conformés. Pourtant, nous avons recours à eux pour leurs
qualités de bons bergers mais visiblement les bonnes manières de
faire (sic) ils les réservent à leurs propres animaux. En outre,
dès qu'un berger obtient la garde de tes animaux, non seulement il te
fait dépenser régulièrement, mais en plus tu es
obligé de contribuer à l'alimentation de sa famille. Mais le plus
écoeurant dans tout ça ce sont les pertes (animaux morts et
égarés) que nous enregistrons de plus en plus ». Cet
argumentaire est surtout valable dans les cas de confiage où le troupeau
de l'éleveur se trouve fusionné avec celui du berger. Dans ce cas
de figure en effet, les compléments alimentaires fournis par le
propriétaire seraient distribués à l'ensemble du troupeau
alors que les apports du berger sont prioritairement distribués à
ses propres animaux. En ce qui concerne les bergers salariés, les
reproches se situent dans ce qui est qualifié de manque d'engagement et
de lisibilité dans la gestion des troupeaux (nombreuses pertes et morts
d'animaux) alors que le service est fortement rémunéré
parfois à la limite du supportable pour les ressources du
propriétaire. Ces récriminations, si elles étaient
avérées, justifient pourquoi certains propriétaires de
troupeaux, s'ils continuent à vouloir de ce type de bergers dont
l'expérience et le savoir-faire sont reconnus et recherchés, leur
adjoignent de proches parents (fils ou autres personnes de confiance). Par
ailleurs, au sein des élevages des transhumants ( et C3), les
conversations ont montré qu'il n'est pas courant pour un éleveur
peul de confier ses animaux à la garde d'un autre ; les rares cas de
confiage sont liées à des contraintes familiales notamment
lorsque dans la famille il manque un garçon d'un âge
adéquat et que le propriétaire lui-même est d'un âge
avancé65.
Les grands éleveurs sont ceux qui ont le plus souvent
recours à 2 ou 3 bergers. Ceci a été aussi observé
par Thébaud (2002) chez les Peuls et Riimaaybe du Yagha (Burkina Faso)
et chez les Wodaabe de Diffa (Niger). Par ailleurs, un nombre plus
élevé de bergers est observé dans les troupeaux
transhumants qui sont souvent regroupés (un troupeau est très
souvent une agrégation de plusieurs troupeaux de petite taille) (Riegel,
2002). Cette pratique qui consiste à rassembler le troupeau au
départ de la campagne de transhumance, contribue à alléger
la tâche du Garso et à renforcer la
sécurité au cours de la campagne de transhumance. Elle permet de
plus un partage de savoir-faire technique ainsi que la formation des plus
jeunes qui s'entraînent ainsi à la vie de privation et à
l'endurance. D'après Baadjo Idrissa (transhumant nigérien de
Makalondi), en effet « sur le terrain, les épreuves sont
nombreuses et en nous regroupant, nous arrivons à les juguler en nous
répartissant les tâches. Par ailleurs, beaucoup de jeunes
subissent leur examen de passage de bons bergers en participant à une
campagne de transhumance, mais il est trop risqué pour eux d'y aller
seuls. Enfin, de cette façon nous sommes moins dispersés et notre
chef (le garso) peut facilement intervenir car, il a alors moins de troupeaux
sous sa coupe». Ce récit rejoint parfaitement celui des
éleveurs de la région de Tamou (Niger) rapporté par Riegel
(2002) et confirme le fait que la transhumance
65 Boutrais (1996) évoque d'ailleurs
l'âge comme pouvant être une cause de baisse des effectifs animaux
au sein des familles pastorales.

est une occasion de rencontre, de partage et de renforcement de
liens sociaux qui permettent l'intégration des communautés
pastorales (Bary, 1998).
5.3.4.3. M obilité des éleveurs
Dans le contexte actuel de raréfaction de l'espace et
des ressources, la mobilité peut être révélatrice
des conditions et contraintes que vit l'éleveur ainsi que de ses
objectifs de production. Elle peut être considérée comme
l'expression de sa manière d'utiliser l'espace pour exploiter des sites
potentiellement différents. Il s'agit donc d'un paramètre
très important. La mobilité est décrite à travers
l'ampleur des déplacements des troupeaux, leur fréquence et leur
éventuel caractère cyclique, éléments qui
caractérisent le degré de dépendance de ses animaux
vis-à-vis des pâturages naturels. Dans la présente
étude nous aborderons ces points, mais aussi la nature des lieux
fréquentés et les motivations des choix opérés par
les éleveurs.
Pendant la période de référence, les non
transhumants (C1) ont, dans un passé récent, eu à
transhumer à la recherche surtout de fourrage et accessoirement d'eau.
Dans un passé plus lointain, cette pratique était
également connue mais elle impliquait peu d'éleveurs et elle
l'était seulement pour des besoins de recherche d'eau (figure V-15a). Il
faut dire que, d'après ces éleveurs, il ya une dizaine
d'années le terroir a connu une crise fourragère alors que la
saison pluvieuse n'a pas été particulièrement mauvaise. Il
y a qu'en cette saison la pression agricole sur les terres avait atteint son
maximum, ce qui avait entamé la production fourragère naturelle
causant un déficit en la matière. Quand on regarde les zones
pâturées, en saison sèche, par les troupeaux de ces
éleveurs (figure V-16a), on se rend compte qu'en réalité
il s'agit de petite transhumance, ces troupeaux ne pâturant pas
au-delà des terroirs voisins. A ce sujet, on notera qu'avec le temps,
ces troupeaux qui ne fréquentaient que les pâturages du terroir,
mettent de plus en plus à contribution les pâturages des terroirs
proches.
Les transhumants résidents dans le terroir de Kotchari
(), avec le temps, fréquentent des territoires de plus en plus
lointains. De nos jours ils vont majoritairement dans les pays voisins mais se
rencontrent aussi dans les terroirs voisins tout comme dans les aires
protégées voisines, en l'occurrence le parc W et la concession de
chasse de la Kourtiagou (figure V-16b). Leur départ en transhumance est
motivé surtout par la recherche combinée de l'eau et du fourrage
alors que par le passé, l'argument de l'eau prévalait (figure
V-15b).
La plupart des transhumants venant du Nord du terroir (C3),
comme les éleveurs de la catégorie précédente, se
destinent majoritairement vers les pays voisins, un grand nombre cependant
demeure sur place ou dans les terroirs riverains de Kotchari (figure V-16c). Un
fait notable à signaler est que bon nombre de ces éleveurs,
reconnaissent avoir par le passé régulièrement
fréquenté les aires protégées. Les mêmes
motivations que celles des éleveurs de type , justifient les
départ en transhumance de ces éleveurs (figure V-15c) : la
recherche à la fois de l'eau et du fourrage de nos jours ou seulement de
l'eau par le passé.
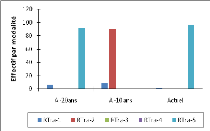
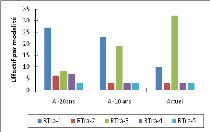
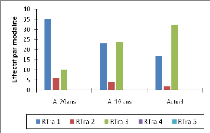
a. Groupe C1 b. Groupe
c. Groupe C3
Figure V-15. Pratique de la transhumance : raisons principales
évoquées
Légende:
RTra-1 : recherche de l'eau d'abreuvement RTra-4 : par simple
habitude
RTra-2 : recherche de fourrage RTra-5 : n'est pas concerné
par la pratique
RTra-3 : recherche d'eau et de fourrage
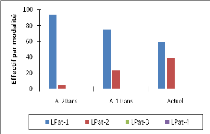
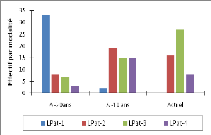
151
a. Groupe C1 b. Groupe
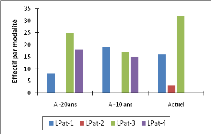
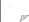
c.Groupe C3
Figure V-16. Milieux de pâture habituelle en saison
sèche
Légende:
LPat-1 : dans le terroir de Kotchari LPat-2 : Kotchari et
terroirs voisins LPat-3 : pays voisins (Bénin/Togo) LPat-4 :
réserves voisines (Parc W et CC Kourtiagou)
Dans la région où se situe notre terrain, la
question de la mobilité, en particulier de la transhumance a fait
l'objet de nombreuses études. Dans le programme régional ECOPAS
coordonné par une équipe du CIRAD tout un volet de la composante
« recherche scientifique » lui était consacré. Ce
programme qui a porté sur les 3 pays qui accueillent le parc W
(Bénin, Burkina Faso et Niger) a produit une impressionnante masse
d'informations. Ainsi, sans que cela ne soit exhaustif, nous citerons les
travaux de Convers (2002), Paris (2002), Tamou (2002), Kabirou (2003) et
Kagoné (2004) qui ont permis de spatialiser la mobilité des
éleveurs autour du Parc.
Nos résultats confirment le statut de terroir d'accueil
et de transit attribué au terroir de Kotchari (Paris, 2002 ; Kpoda,
2010). La tendance actuelle est à davantage d'accueil, les
Gourmantchés et les Peuls résidents (catégories C1 & )
partageant désormais de plus en plus l'espace de leur terroir avec des
transhumants allochtones (C3) qui, cependant, continuent d'aller plus au sud,
notamment au Bénin. Si les Gourmantchés qui relèvent du
groupe d'éleveurs C1 affirment ne jamais fréquenter les
réserves, (affirmation confirmée par notre guide Y. Diallo), les
autres groupes reconnaissent les avoir utilisées surtout dans le
passé (C3 par le passé et par le passé et de nos jours).
On peut être étonné que si quelques éleveurs
résidents reconnaissent fréquenter encore les réserves,
aucun transhumant non résident ne l'avoue alors que dans un passé
récent, la plupart d'entre eux y allaient. La réponse à
nos questions insistantes sur la cause de cette différence est que le
parc est devenu plus dangereux que par le passé. Par « dangereux
» les riverains entendent que le parc W comme la réserve de la
Kourtiagou sont, depuis 2001, soumis à une surveillance accrue. Dans la
pratique, cette surveillance se traduit par la présence dans les
terroirs riverains de pisteurs, personnes du crû qui connaissent bien les
réserves. Ils évoquent largement des campagnes rapprochées
de « ratissage » par les forestiers parfois organisées
conjointement par les 3 pays,

153
des fortes amendes et des abus divers (racket66,
abattage ou « vaccination67 » d'animaux, etc.). Si de tels
faits sont bien réels, ils ne suffisent pourtant pas à expliquer
entièrement la situation. En effet Bary (1998), Toutain et al.
(2001) Tamou (2002) et Kagoné (2000 & 2004) indiquent que ces
contraintes sont connues et acceptées par les éleveurs en
échange du bien être de leurs animaux. En vérité, la
question reste taboue et une longue préparation de mise en confiance a
été nécessaire pour qu'elle soit évoquée de
manière franche par le berger transhumant ou même résident.
Il n'est pas toujours possible de parvenir à une telle confiance,
surtout avec des éleveurs très mobiles même dans l'espace
villageois. Ils restent d'ailleurs assez méfiants à
l'égard de l'impertinent inconnu (c'est ainsi que nous étions
perçu) qui se comporte comme le "Toubaku" (terme peul
désignant une personne de race blanche) et qui, en plus, est familier
des forestiers, leurs ennemis jurés. Nous n'avons réussi à
lever cette barrière qu'avec certains éleveurs résidents
dont certains étaient nos interlocuteurs au sein de leur
communauté. Ils ont alors pu avouer. Bien que nous n'ayons jamais
été témoin de ces infractions, elles permettent
d'expliquer pourquoi, comme nous l'avions déjà dit (Sawadogo,
2004) les éleveurs peuls placent leur campements de nuit mobiles
à proximité des réserves. Par cette mobilité ils
évitent d'être repérés par les forestiers et
pisteurs ou d'éventuels indicateurs villageois. Il est fort probable que
parmi les éleveurs qui affirment continuer leur transhumance au
Bénin, certains aboutissent finalement dans l'une ou l'autre des deux
réserves (parc W et Concession de chasse de la Kourtiagou). Pour
vérifier cette hypothèse, assez logique, il aurait fallu pouvoir
vérifier les flux réels et les identités des
éleveurs réellement accueillis de l'autre côté de la
frontière.
5.3.4.4. Complémentation des anim aux
La forte pression animale sur les ressources pastorales, qui
découle de l'accroissement permanent des effectifs animaux dans le
terroir et de l'allongement de la saison sèche, a conduit à
l'émergence de nouvelles pratiques d'alimentation. Celles-ci viennent en
appoint à l'exploitation directe du fourrage naturel des parcours qui,
à elle seule, ne suffit plus à satisfaire aux besoins d'entretien
et de production des animaux en saison sèche.
De nos jours chez les éleveurs non transhumants (C1),
de grands agriculteurs aux petits effectifs, la complémentation est
courante ; les pratiques de complémentation dominantes consistent
à la valorisation des résidus de cultures (tiges de
céréales, fanes d'arachide et de niébé), cependant
un bon nombre d'entre eux (ceux qui ont de grands effectifs) collectent en plus
du fourrage naturel et même en achètent. Par le passé,
surtout il ya une vingtaine d'années, les éleveurs se
contentaient majoritairement, du fourrage directement prélevé par
les animaux au parcours, mais ils pouvaient également en faire un stock
pour une distribution ultérieure (figure V-17a). Notons que ces
agroéleveurs ont expérimenté dans un
66 Courant 2007, une vaste escroquerie a
été dénoncée à l'autorité par les
éleveurs qui, une fois n'est pas coutume, ont pris leur courage à
deux mains encouragés en cela par les Garso locaux. Il semble
que cette dénonciation suivie d'une marche de protestation dans la ville
de Diapaga, ait été pour quelque chose dans l'affectation de
nombreux agents forestiers soupçonnés d'indélicatesse.
67 Ce terme serait un qualificatif que les
forestiers béninois donnent à leur campagne de ratissage du parc
W béninois et qui conduit le plus souvent à des abattages parfois
d'effectifs importants de bétail rencontrés en son sein (Convers,
2002 ; Binot et al. 2006).
passé récent la culture de diverses
espèces de légumineuses fourragères telles que le pois
d'angole (Cajanus cajan), le mucuna (Mucuna rajada
et Mucuna deeringiana), la dolique (Dolichos lablab) et
certaines plantes à double usage (sorgho, niébé et
arachide fourragers) avec l'appui technique de la station de recherche agricole
de l'INERA de Fada N'Gourma et des projets de développement, mais
l'engouement n'a duré que le temps de vie de ces projets. Les aliments
achetés sont généralement des concentrés
alimentaires (divers sons, graines et tourteaux de coton, sel).
Les transhumants résidents () quant à eux, plus
impliqués dans la complémentation que le groupe
précédent (depuis une dizaine d'années, on ne trouve
d'ailleurs pas dans ce groupe d'éleveurs des personnes qui ne soient
concernées), l'usage seulement de concentrés achetés ou
accompagnés de fourrage divers mis en stock, sont les pratiques
dominantes de nos jours. Par le passé ces éleveurs utilisaient
surtout des compléments fourragers naturels (pratique dominante il y a
une vingtaine d'années) ou cultivés (pratique dominante il y a
une dizaine d'années). L'achat d'aliments concentrés est une
habitude qui s'installe depuis une dizaine d'années (figure V-17b).
Les transhumants allochtones (éleveurs de la
catégorie C3) se sont toujours contentés des ressources
fourragères prélevées directement sur parcours dans le
terroir de Kotchari (figure V17c). Depuis un moment cependant, ils commencent
à acheter des concentrés dans les marchés locaux ou
directement dans les concessions chez les résidents.
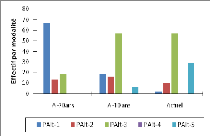
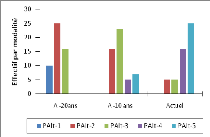
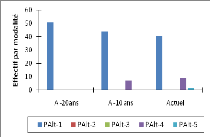
a. Groupe C1 b. Groupe
c.Groupe C3
Figure V-17. Pratiques de complémentation

155
Légende:
PAlt-1 : pâturage naturel (pas de
complémentation)
PAlt-2 : collecte fourrage naturel PAlt-3 : collecte fourrage
cultivé PAlt-4 : achat de concentré (sons et tourteaux)
PAlt-5 : achat d'aliments (fourrage et concentré) et
collecte (fourrage naturel et cultivé)
Chez les éleveurs résidents (C1 & ), il y a
un début d'intégration de l'agriculture à
l'élevage, qui est favorisée par la culture du coton (utilisation
des charrues asines et bovines, enrichissement des champs par la fumure
organique produite à partir de fèces animaux, etc.).
Révélatrice de la tendance à la diversification des
activités agricoles (Broussard, 2001) au sein des exploitations, cette
nouvelle organisation permet aux éleveurs d'accroître la
production de fourrage d'appoint au sein de leurs unités de production.
Certains Gourmantchés parviennent même à constituer des
stocks de résidus agricoles plus importants que leurs besoins dans le
but de les revendre.
Un point important est que la complémentation
bénéficie prioritairement aux animaux mal en point ou allaitant.
On comprend dès lors pourquoi les éleveurs transhumants sont
longtemps restés peu concernés par la pratique localement, ce
type d'animaux étant resté dans les terroirs d'attache. Il est
vrai que lorsque les grands transhumants arrivent dans le terroir, il est trop
tard pour collecter le fourrage naturel local tandis que le fourrage
cultivé a déjà été collecté ou est
exploité en vaine pâture par les propriétaires des champs.
En outre, il peut être financièrement difficile de
complémenter de grands effectifs avec des produits achetés. A ce
propos, Sadio raconte, avec un ton amer, les contraintes qu'il rencontre dans
ce terroir auparavant si attractif « avant, quand on venait les champs
étaient récoltés et le Gnagnical (résidus de
culture) laissé en place. Chacun avait un ou des tuteurs et pouvait
librement faire paître ses animaux sur leurs parcelles. Maintenant,
à notre arrivée, tout est déjà fini et en plus
l'accès aux parcelles vides est souvent refusé, les
propriétaires y ayant déjà leurs propres animaux. Par
moments nous sommes contraints de supplémenter nos animaux, mais on n'y
arrive pas toujours car cela nous revient cher ». Toutes ces raisons
expliquent que la complémentation, surtout par les résidus de
récolte, pratique très répandue chez les agropasteurs
locaux (Gourmantchés comme Peuls), soit restée très
marginale chez les transhumants qui se contentent jusqu'à présent
de la paille naturelle et de concentrés (son et tourteau).
Toutes ces catégories d'éleveurs, en particulier
ceux des groupes et C3, ont recours aux ligneux68 disponibles sur
les pâturages du terroir en saison sèche notamment chaude.
68 Les bergers émondent les ligneux à
la machette, mais c'est illégal sans autorisation préalable (cf.
l'article 118 alinéa 4 de la loi portant Réforme Agraire et
Foncière). Les éleveurs, s'ils ne sont pas pris sur le champ, ne
reconnaissent donc pas toujours se livrer à cette pratique. Pourtant,
beaucoup d'auteurs (Zouri, 2003 ; Ouédraogo, 2008) et nos propres
observations sur le terroir de Kotchari montrent que de nombreux arbres sont
mutilés, en tout premier lieu les espèces Ptereocarpus
erinaceus et Afzelia africana.
Notons que la distribution de ces compléments
alimentaires aux animaux se fait à l'auge et surtout dans la
deuxième moitié de la saison sèche (mars à juin)
période de pointe de la transhumance et de disette pour l'ensemble des
animaux.
Le recours de plus en plus fréquent de tous les
éleveurs à la complémentation animale avec achat de
fourrage cultivé et/ou de concentré alimentaire montre que les
pâturages naturels n'arrivent plus à satisfaire les besoins des
animaux dont les effectifs sont sans cesse croissants.
De plus on observe que le fourrage ligneux est aussi mis
à contribution toute l'année, mais surtout en saison
sèche. Ce type de fourrage, meilleur que la paille en saison
sèche (biomasse verte et plus grande teneur en azote) (Breman & De
Ridder, 1991 ; KaboréZoungrana, 1995 ; Baumer, 1997), est fourni
localement par des espèces comme Afzelia africana, Pterocarpus
erinaceus, Khaya senegalensis, Lonchocarpus laxiflorus, Acacia seyal,
Balanites aegyptiaca, Combretum aculeatum, etc. qui, du fait de la
pression d'utilisation, se raréfient en dehors des réserves. Il
est exploité directement par les animaux (arbustes de moins de 2 m de
hauteur)69, ou grâce à l'intervention des bergers qui
émondent les grands arbres pour le troupeau (photos V7).

Cliché Sawadogo Cliché Sawadogo

a. Ptereocarpus erinaceus b. Afzelia africana
Photos V-7. Deux espèces de ligneux fortement
émondés par les bergers
5.3.4.5. Diversification des troupeaux
La composition des troupeaux villageois est assez
variée, mais le plus souvent ils regroupent bovins, ovins et caprins.
Cependant les troupeaux composés uniquement de bovins ou à la
fois de bovins et d'ovins sont de plus en plus nombreux. Quelle que soit la
69 Les travaux de Ouédraogo (2008) montrent
qu'il n'existe pas à Kotchari de difficulté d'exploitation du
fourrage ligneux, l'essentiel (63 à 94%) des types ligneux ayant une
taille inférieure à 2 m, hauteur limite à partir de
laquelle l'intervention du berger est nécessaire pour l'accès au
matériel végétal par les animaux (Hiernaux, 1980 ;
Kièma, 2007).
composition d'ensemble du troupeau, les mélanges de races
bovines sont de plus en plus fréquents, notamment les associations
Barbaji + Gurmaji et Gurmaji + Jaliji.
Les troupeaux des éleveurs non transhumants (C1)
étaient, il ya une vingtaine d'années, pour une grande part
d'entre eux, dépourvus de l'espèce bovine. Les troupeaux qui en
disposaient, comprenaient aussi des ovins et des caprins. Il ya une dizaine
d'années, la part de troupeaux sans bovins ainsi que celle des troupeaux
plurispécifiques (bovins, ovins, caprins) ont diminué, à
leur place se forment des troupeaux à deux espèces (bovins et
ovins) ou monospécifiques à bovins. Ces derniers temps cependant,
la tendance s'inverse presque : les troupeaux sans bovins ou à trois
espèces reprennent du poids dans ces élevages alors qu'il
apparaît quelques troupeaux comprenant à la fois des bovins et des
caprins (figure 18a). Au niveau de la structuration raciale bovine, ces
troupeaux se caractérisent par un poids de plus en plus important des
troupeaux composés à la fois de la Barbaji et de la
Gurmaji (NRaB-4) surtout mais aussi de ceux qui sont à trois
races au moins (NRab-6). L'émergence de ces deux profils de troupeaux se
fait aux dépens de ceux constitués de la seule race
Barbaji (NRab-1) Gurmaji (NRab-2) ou même
Jaliji (NRab-3) (figure V-18b).
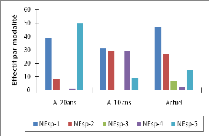
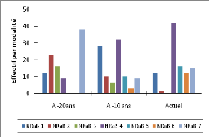
157
a. Nombre et types d'espèces du troupeau b. Nombre et
types de races bovines
Figure V-18. Composition spécifique et raciale des
troupeaux du groupe C1 Légende:
NEsp-1 : bovins + ovins + caprins NRaB-1 : Barbaji
NEsp-2 : bovins + ovins NRaB-2 : Gurmaji
NEsp-3 : bovins + caprins NRaB-3 : Jaliji
NEsp-4 : bovins NRaB-4 : Barbaji + Gurmaji
NEsp-5 : Pas de bovins NRaB-5 : Gurmaji + Jaliji
NRaB-6 : Autres cas de figures (plus de deux races)
NRaB-7 : pas de bovins dans le troupeau
Chez les transhumants résidents (), l'évolution
de la composition spécifique au cours des 20 dernières
années montre un déclin des troupeaux comprenant à la fois
bovins, ovins et caprins alors que sont en train d'émerger des troupeaux
à bovins et ovins (figure V-19a). Alors que quelques troupeaux
monospécifiques à bovins se rencontrent actuellement dans ces
élevages, il n'en existe plus qui ne contiennent pas de bovins. Les
troupeaux comprenant seulement la race bovine Gurmaji, fortement
représentés par le passé, n'existent plus, alors qu'il
devient de plus en plus possible de rencontrer des troupeaux ayant deux
à plus
de deux races bovines. De nos jours, les configurations
raciales des troupeaux qui sont les plus représentées sont les
associations Barbaji + Gurmaji (NRaB-4) surtout et Gurmaji +
Jaliji (NRaB-5) ou alors les associations de plus de deux races bovines
(NRaB-6) (figure V-19b).
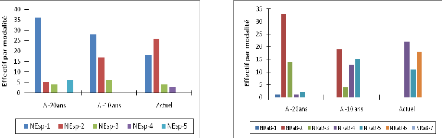
a. Nombre et types d'espèces du troupeau b. Nombre et
types de races bovines
Figure V-19. Composition spécifique et raciale des
troupeaux du groupe
Les troupeaux du groupe des éleveurs transhumants non
résidents (C3), montrent une évolution (figure V-20a) qui se fait
en défaveur de ceux composés à la fois des espèces
de bovins et d'ovins (NEsp-2), beaucoup plus présents par le
passé, et de ceux à trois espèces (bovins, ovins et
caprins) (NEsp-1) et en faveur de ceux ne comprenant que la seule espèce
bovine (NEsp-4). L'espèce caprine est peu présente dans ces
élevages. Au sein de ces troupeaux, caractérisés par la
présence d'une seule race bovine en général, au fil du
temps, l'ordre d'importance s'est inversé en faveur de la race
Gurmaji et au détriment de la race Jaliji (figure
V-20b). Par ailleurs, une part de plus en plus importante de troupeaux
comprenant à la fois les races Gurmaji et Jaliji
(NRab-5) est enregistrée alors qu'au contraire on ne
rencontre plus de troupeaux à plus de deux races bovines (NRaB-6).
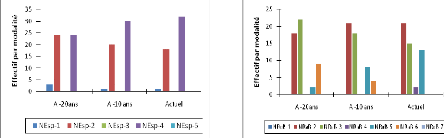
a. Nombre et types d'espèces du troupeau b. Nombre et
types de races bovines
Figure V-20. Composition spécifique et raciale des
troupeaux du groupe C3
On le voit, dans les différents troupeaux, quel que
soit le type d'élevage, l'évolution se fait vers la
réduction du nombre d'espèces animales. Cette tendance est nette
chez les transhumants résidents ou non ( et C3) et moins nette et
même en inversion chez les agroéleveurs résidents (tendance
en légère inversion il ya une dizaine d'années). Ce

159

comportement particulier est à relier au
caractère assez diversifié de ce groupe d'éleveurs qui
comprend des Gourmantchés dépourvus ou non de bovins (C1-1 et une
partie de C1-2) et des peuls sédentaires. Seuls les premiers s'adonnent
à la culture du coton, activité déterminante dans la
taille de leur cheptel, la capitalisation des revenus agricoles se faisant par
l'achat de bétail. De manière globale, le cheptel des
Gourmantchés est dans une phase de baisse des effectifs et de
déstockage des espèces bovines qui avaient pu être mis en
place à la faveur de l'envol de l'activité cotonnière. La
tendance constatée vers le retour à la
plurispécificité chez C1 pourrait signifier que plus le troupeau
perd en taille, moins l'éleveur est enclin à spécialiser
son troupeau vers une ou quelques espèces et inversement. Cette
même raison expliquerait donc les constats faits dans les troupeaux des
Peuls résidents ou non chez lesquels le cheptel est en constante
augmentation et où le nombre d'espèces animales par troupeau tend
à baisser.
On note par ailleurs, que dans les élevages de type C1
et (éleveurs résidents), il y a une tendance à la
diversification, notamment au doublement, des races bovines dans les troupeaux
(Barbaji + Gurmaji au lieu de Barbaji ou
Gurmaji à l'origine pour C1 et respectivement). Boutrais
(2002), étudiant le rapport des populations pastorales à leurs
races bovines, avait fait les mêmes observations qui semblaient
répondre à une stratégie de reconstruction des troupeaux
pour les éleveurs ayant connu une crise pastorale. Il a cependant aussi
observé une tendance, dans certains groupes pastoraux, à
l'uniformisation de la race bovine vers les types les plus adaptés
à l'écologie des aires d'élevage.
De manière générale, il y a un recul de
la race Jaliji en faveur des races Gurmaji et Barbaji,
ce qui traduit un choix stratégique. En effet, alors que la
Jaliji, bien que rustique, est peu trypanotolérante, la
Gurmaji qui semble être son adaptation locale de très
longue date (Santoir, 1999), tolère le climat local (elle est
trypanotolérante) tout en exprimant des propriétés
zootechniques (âge au premier vêlage, intervalle de mise-bas,
lactation, etc.) peu différentes de celle-ci. Elle est, en outre,
adaptée à la longue marche et serait disciplinée rendant
son gardiennage aisé. Les mêmes raisons d'adaptation aux
conditions locales (trypanotolérance et rusticité alimentaire)
expliqueraient le maintien et le renforcement de la présence de la race
Barbaji. Cette dernière, mauvaise marcheuse cependant, est
indisciplinée (troupeau généralement dispersé sur
parcours) et sa garde commande beaucoup plus d'attention de la part du
berger.
Chez les non résidents (C3) la tendance à la
diversification des races, quoique moins rapide, est également
observée. Même si les troupeaux à Gurmaji + Jaliji
gagnent en importance, il subsiste majoritairement des troupeaux à une
seule race bovine dans leur cheptel ; cependant, la race Gurmaji y
supplante la race Jaliji originelle. Ces éleveurs, sans doute
plus exposés aux contraintes du milieu, du fait des distances
parcourues, sont en fait en train de délaisser, comme c'est le cas chez
les Peuls résidents transhumants, la race Jaliji au profit de
la race Gurmaji. Si les motifs ci-dessus évoqués restent
valables ici, il faut ajouter en plus l'incapacité de la Jaliji
à supporter les longs déplacements (Amadou, 1999) alors que,
selon les éleveurs, la Gurmaji s'en adapte mieux.
De manière générale, on peut relever avec
grand intérêt le fait que les configurations raciales dans les
espèces bovines des troupeaux s'opèrent avec pour
préoccupation sous-
jacente de les adapter aux exigences du moment faites de longs
et pénibles déplacements sur des parcours
généralement très appauvris. Ceci exige des
éleveurs de choisir non pas toujours les races les plus productives
et/ou les plus esthétiques (Boboroji par exemple) mais surtout
celles qui s'adaptent le mieux notamment au climat ou qui sont peu exigeantes
au plan alimentaire (rusticité) face à la rareté de la
ressource. Les races Barbaji et Gurmaji semblent
réunir toutes ces exigences. Cette tendance est confirmée par
Amadou (1999) à partir d'observations enregistrées dans le Boboye
au Niger. L'auteur rend compte du fait que « le choix des
espèces chez les peuls du Boboye n'est plus uniquement guidé par
les impératifs de production (...) ou d'affection (...) mais aussi par
leur capacité d'adaptation. En effet, l'aptitude à supporter
certaines situations difficiles (pâturages médiocres,
sécheresse, maladies) compte de plus en plus dans la sélection du
troupeau ». Pour les mêmes raisons, les éleveurs
expliquent l'abandon ou la faible présence de certaines races par leur
grande conformation (Boboroji et Kiwali notamment) qui
implique de grands besoins alimentaires (Boutrais, 1994)70 pas
toujours aisés à satisfaire. Notons cependant que, comme les
éleveurs transhumants qui continuent d'entretenir des troupeaux bovins
à Jaliji, il n'est pas toujours aisé pour
l'éleveur de se départir de sa race initiale à laquelle il
reste lié et qui constitue bien souvent, comme l'a signalé
Boutrais (2002), un patrimoine intergénérationnel.
Le rapport du nombre mâles/femelles (sex-ratio) dans la
composante bovine des troupeaux est relativement (C3) ou statistiquement ()
plus faible dans les élevages purement peuls (tableau V-9, paragraphe
5.3.3.2.4 plus haut). Des différences plus nettes que celles que nous
observons ont été notées par Tamou (2002) et
Sounkéré (2003) au nord-Bénin et Kièma S. (2007)
dans l'ouest burkinabè. Le premier auteur a observé que, alors
que le sex-ratio des groupes résidents (Gourmantché, Dendi,
Haoussa) oscillait entre 0,55 et 0,84, celui du groupe Peul était
seulement de 0,27. Kièma S., quant à lui, a constaté que
les troupeaux d'éleveurs résidents qu'il a enquêtés
se composaient d'environ 1 mâle pour 10 femelles (soit un sex-ratio de
0,1). Ceci est révélateur des objectifs des élevages peuls
(ou mobiles en général) qui sont tournés beaucoup plus
vers le type « naisseur ». Rappelons que les éleveurs de type
« naisseur » garantissent des grands effectifs capables de subsister
aux catastrophes naturelles comme les sécheresses et les
épizooties (Bernus, 1981 ; Boutrais, 1996). Une autre explication est
que les peuples pasteurs, qui dépendent beaucoup du lait pour leur
alimentation ou comme produit d'échanges, sont amenés à
favoriser les femelles, en particulier les vaches reproductrices, dans leurs
troupeaux (Boutrais, 1996).
Le groupe d'éleveurs peuls transhumants ou non mettrait
donc l'accent sur l'exploitation des mâles de leurs troupeaux, ce qui
d'après Tamou (2002), permettrait en même temps de gérer
les risques de combats mortels en leur sein. Au contraire, dans bien des
élevages résidents surtout chez les Gourmantchés, le
réélevage71 est développé et explique la
plus forte proportion de taureaux en leur sein. Si nos observations corroborent
bien ce qui se constate en zone sahélienne, elles contredisent la
tendance notée par Boutrais (1996) en contexte savanien
70 L'auteur distingue les pseudo-zébus
(Mbororroji et Akuuji ou White Fulani) à
longues cornes et silhouette élancée qui ont des grandes
exigences fourragères des vrais zébus (Bos indicus :
Gudali, Azawak, etc.) à courtes cornes, stature ramassée
et grosse bosse. Ces derniers et les taurins, qui ont moins d'exigences
fourragères et qui ont de fortes capacités d'adaptation, sont
moins redoutables pour les parcours par rapport aux premiers.
71 Le réélevage est une pratique qui
consiste à importer dans le troupeau des jeunes veaux achetés
pour leur finition (alimentation généralement plus intensive)
(Touré, 2010)

161


163


165


167


169
d'Afrique subsaharienne. Cet auteur rapporte qu'en zone de
savane, l'économie des pasteurs ne reposerait plus essentiellement sur
le lait, ce qui ne les oblige pas à entretenir un grand effectif de
femelles au sein de leurs troupeaux. Par ailleurs, les troupeaux des
agriculteurs résidents y seraient généralement en phase
d'accumulation, avec des effectifs importants de femelles.
5.3.5. Les rapports sociaux entre les acteurs et leurs
conséquences sur les pratiques pastorales
Les acteurs de l'élevage (transhumants et
sédentaires) et de la conservation (agents forestiers) entretiennent
entre eux un certain nombre de rapports de collaboration ou d'échanges,
tantôt de méfiance et surtout de plus en plus hostiles et
même conflictuels. Aborder la question des rapports conflictuels dans un
milieu donné revient à déterminer les parties en conflits
(les catégories de protagonistes qui se disputent une ressource
spécifique), les enjeux et les objets (Barrière, 1996).
Ce chapitre des rapports entre les acteurs dans le terroir a
été largement abordé par le passé (Sawadogo ;
2004). La question a été approfondie par des entretiens
ciblés avec quelques personnes ressources avec une faveur
accordée à la question de la conflictualité, vue comme un
révélateur de contraintes rencontrées par les populations
dans leurs activités agropastorales (Boutrais, 1983 ; Dugué et
al. 2004). Notons que la question a été également
largement abordée par le programme ECOPAS (tableau V-10) mais nous avons
orienté notre investigation sur quatre niveaux de rapports : les
relations (i) transhumants - populations sédentaires, (ii)
éleveurs résidents gourmantchés - éleveurs
résidents peuls, (iii) agroéleveurs - administration
forestière et (iv) entre éleveurs transhumants.
Tableau V-10: Typologie des conflits dans la
périphérie du WAP
|
Type de conflit
|
|
Principales causes
|
|
Éleveurs transhumants vs
|
·
|
Occupation des espaces pastoraux (pistes à bétail,
aires
|
|
Agriculteurs
|
|
de pâturage, voies d'accès aux points d'eau)
|
|
·
|
Dégâts de cultures et/ou de récoltes dans les
champs
|
|
Éleveurs transhumants vs
|
·
|
Inexistence ou non aménagement de couloirs de
|
|
Services forestiers
|
|
transhumance obligeant les animaux très affaiblis à
de grands détours
|
|
·
|
Exploitation pastorale des aires protégées,
dégradation de la faune et de son habitat
|
|
·
|
Mauvais traitement infligé aux animaux saisis, abattages
systématique d'animaux (surtout du côté Bénin)
|
|
Éleveurs transhumants vs
|
·
|
Dégradation de la faune et de son habitat
|
|
Concessionnaires de zone
|
·
|
Dépréciation de la valeur touristique de la
concession du
|
|
de chasse
|
|
fait de la présence de bétail domestique dans les
concessions
|
|
Éleveurs transhumants vs
|
·
|
Pâturage nocturne, dégâts de champs et
exacerbation des
|
|
Éleveurs sédentaires
|
|
conflits avec les agriculteurs
|
|
·
|
Concurrence sur l'exploitation des rares ressources
|
|
|
pastorales conduisant à des déplacements
obligatoires des éleveurs résidents
|
|
Éleveurs transhumants vs
|
·
|
Tracasseries administratives, y compris raquettes et taxes
|
|
corps habillés (Police,
|
|
sauvages
|
|
Douane, Gendarmerie,
|
·
|
Non respect de la réglementation
|
|
Forestiers)
|
·
|
Dégâts humains (viols de femmes, mort d'hommes,
etc.)
|
|
·
|
Tentatives de contournement des mesures de suspension de la
transhumance transfrontalière au Bénin et au Togo
|
(Source :
http://www.cirad.bf/fr/anx/parc-w-transhumance.php
consulté le 15 mai 2010)
5.3.5.1. Les relations entre les transhumants et les
acteurs locaux
5.3.5.1.1. Des liens de réciprocité parfois
anciens, mais de plus en plus fragiles
Avec les populations du terroir de Kotchari comme avec celles
des terroirs traversés, les transhumants entretiennent des relations de
nature diverse, parfois multiséculaires. Ces rapports qui prennent
parfois l'allure d'alliance, peuvent être durables, ils reposent sur le
principe de réciprocité qui se caractérise par des
échanges divers (échanges d'informations, de biens et de
services) entre partenaires. Ce type de relations entre sociétés
peules et sociétés autochtones généralement
agricultrices a été déjà décrit par bien
d'auteurs notamment Thébaud (1995 & 2002) qui a étudié
les pasteurs au Yagha burkinabé et au Niger oriental et par Touré
(1997) qui s'est intéressé aux pasteurs du Ferlo
sénégalais. De nos jours ces relations connaissent de profondes
mutations et c'est avec un brin de nostalgie teintée d'amertume que
Sondé Sadjo, un transhumant de Makalondi (Sud-ouest du Niger), nous
dépeint la situation telle qu'il la vit actuellement : «j'ai
commencé à fréquenter ce terroir il y a plus de 20 ans.
Les premières années je venais avec mon grand frère qui
est maintenant fatigué. Grâce à lui j'ai connu la famille
de Mr Couldiaty Tadjoa mon tuteur actuel. Les premiers liens entre nos familles
ont été établis par nos pères respectifs. Lors de
mes premiers séjours, je passais la nuit tout près de la
concession des Couldiaty dans un campement de fortune établi sur leurs
champs de case. Le soir j'avais droit à de l'eau de toilette servie par
les femmes et ils me faisaient partager leurs repas. Et après on passait
une bonne partie de la soirée à échanger. C'est vrai
qu'à cette époque, il m'arrivait parfois, avant mon départ
pour le Niger, de leur laisser un taurillon sans compter les rations
journalières en lait72 qui leur étaient
distribuées ainsi que le fumier que nos animaux apportaient
gracieusement à leurs champs. Aujourd'hui, quand nous arrivons à
Fantama ici (c'est le quartier de la famille Couldiaty), l'accueil manque de
chaleur et selon l'année on peut même être amené
à établir notre campement loin de leurs concessions».
Si la situation de Sadjo peut être déplorée, il faut
signaler que ce cas de figure, où des liens aux origines lointaines sont
encore entretenus, est devenu rare à Kotchari. La plupart des
transhumants n'ont pas ou plus d'attache sur place et établissent leurs
campements, devenus plus mobiles, loin des concessions des résidents.
L'une des explications serait liée à l'accroissement des
effectifs locaux qui épuisent rapidement les fourrages proches des
concessions et qui « repoussent » le
72 De nos jours, les troupeaux produisent moins de
lait du fait de dégradation des conditions fourragères pendant la
campagne de transhumance. Les possibilités de donner une partie de ce
lait sont amoindries en conséquence surtout les bergers qui accompagnent
les troupeaux sont de plus en plus nombreux.
bétail transhumant, devenu par ailleurs plus important.
Par le passé, en effet, les animaux transhumants étaient
relativement très peu nombreux (rarement plus d'une dizaine par an) et
les bergers transhumants pouvaient avoir chacun un tuteur local avec qui ils
partageaient services et difficultés. Les tuteurs gourmantchés,
dont le bétail était auparavant très embryonnaire et
dominé par de petits ruminants, installaient « leurs
étrangers » à proximité de leurs concessions et, en
plus de leur apporter nourriture et eau de boisson, leur servaient
d'intercesseurs en cas de problèmes particuliers avec les autres
villageois.
De nos jours donc les rapports entre partenaires d'autrefois
sont de moins en moins cordiaux, voire hostiles. Fuyant cette hostilité
ambiante, générée selon eux par la concurrence pour
l'accès aux ressources (les tuteurs sont devenus, en effet, de grands
éleveurs), les transhumants ont de moins en moins des relations avec les
locaux et les échanges tendent vers un mode purement commercial
(échanges rémunérés de biens sur le marché
local). Les liens de réciprocité sont devenus superficiels et
ceux qui subsistent pourraient être qualifiés d'opportunistes ; il
n'existe, en effet, plus de réseaux constitués dans le temps et
dans l'espace. Par exemple, de plus en plus, le transhumant change de tuteur
d'une année sur l'autre ou le temps d'un aller-retour au cours de la
même saison de transhumance. De manière générale on
ne rencontre plus que quelques familles résidentes, notamment
gourmantchés, qui acceptent d'accueillir les transhumants sur leurs
parcelles agricoles surtout en début de saison sèche au moment de
la vaine pâture. Ce qui surprend dans tout cela c'est que les
transhumants n'établissent jamais leurs campements auprès des
concessions peules sédentaires avec qui, pourtant, ils interagissent.
Une explication subtile est donnée par l'un d'entre eux, en l'occurrence
Baadio Idrissa. Selon ce dernier, il n y a pas intérêt pour un
transhumant à se faire héberger chez un peul sédentaire ou
autochtone puisque ce dernier a aussi besoin de faire pâturer son champ
(généralement de petite taille relativement à celui du
gourmantché) par son troupeau qui au contraire est de plus grande
taille. La concurrence y serait donc plus importante. A cette raison
liée à l'accès aux ressources se greffe une autre raison
de nature socioculturelle. Ordinairement, il n y a pas d'échanges de
femmes en mariage entre Peuls et Gourmantchés dans cette région,
au contraire cela est possible entre Peuls transhumants et
résidents73. Ainsi, le transhumant, en restant loin des
autres Peuls, accroît ses chances en gardant la liberté de pouvoir
courtiser n'importe quelle fille peule du village. Un autre argument non moins
important est que les Peuls résidents seraient gênés
d'héberger un étranger qui pourrait alors avoir une idée
de la taille de leur cheptel : « un bon Peul est toujours
gêné qu'un autre sache qu'il a moins de têtes de bovins dans
son troupeau », nous a confié Idrissa.
Les relations avec les résidents peuls restent
cependant importantes. Si, les transhumants sont hébergés par les
Gourmantchés, ils passent le clair de leur temps, surtout la
journée lorsqu'ils ne sont pas au pâturage, chez les Peuls
résidents de qui ils se sentent plus proches. Par ailleurs, ces derniers
leur servent d'intermédiaires sur diverses questions. En effet, pour
mener une transaction commerciale ou pour régler un conflit avec
l'administration (forestière ou générale) le transhumant a
recours aux éleveurs sédentaires de son groupe social notamment
les Garso locaux.
73 Kaboré (2010) a noté dans la zone de
la réserve de Pama Nord que des échanges de femmes entre Peul et
autochtones existaient par le passé, ils sont cependant en net recul de
nos jours.
5.3.5.1.2. Des relations de plus en plus
conflictuelles
Pour la grande majorité des autochtones, une
véritable hostilité est entretenue à l'égard des
transhumants et des pasteurs en général. Cette hostilité
est plus manifeste chez les rares non-éleveurs et elle épouse
plusieurs formes allant des feux volontairement mis aux restes des
résidus de culture et à la brousse et parfois à des
menaces voilées proférées à l'endroit des tuteurs
de ces éleveurs allochtones. Cette hostilité peut même
parfois déboucher sur des conflits ouverts que nos entretiens n'ont
malheureusement pas pu révéler74. Le type de conflits
que nous avons pu répertorier a déjà été
rapporté par Barrière et Barrière (1997) à la suite
d'études qu'ils ont conduites dans le delta du fleuve Niger au Mali. Ils
tournent tous autour du foncier et de l'exploitation de l'espace-ressource.
- Les conflits pour l'accès aux points d'eau. La
période de pointe de la transhumance correspond à la
période où l'eau de boisson et d'abreuvement provient
essentiellement des puits et des forages. En conséquence, les pasteurs
surtout transhumants mettent beaucoup de temps aux points d'eau puisqu'ils sont
généralement les derniers à passer leur tour. Pour
remédier à cela, ils creusent des puisards dans les
différents lits de rivières rencontrés sur le terroir. Ces
puisards sont également pris d'assaut par les autochtones qui
considèrent de toute façon qu'ils sont les propriétaires
des lieux, ce qui occasionne des bagarres, cependant vite circonscrites, dont
nous avons souvent été témoin.
- Les conflits liés aux dégâts
provoqués aux champs. Même si lors des entretiens, les
pasteurs peuls affirment prendre leurs dispositions pour éviter ce genre
de situation, la réalité est tout autre. Les transhumants pour
des raisons évidentes (retard des pluies dans leurs terroirs d'attache)
s'attardent de plus en plus dans les terroirs d'accueil comme cela a
été le cas en 2008. Or les terres agricoles du terroir de
Kotchari sont, du fait de la culture du coton, ensemencées très
tôt75. Par ailleurs, du fait de la pression agricole sur les
rares terres arables, l'activité agricole se déporte sur les
terres jadis délaissées pour leur faible valeur agronomique et
utilisées comme zones de pâture traditionnelles (photo V-8). Pour
toutes ces raisons, les risques de conflits sont accrus, tout
spécialement pour ceux des transhumants qui sont allés plus bas
notamment au Bénin. En effet, leur repli vers les terroirs d'attache
devient problématique surtout que même les pistes à
bétail sont ignorées par les agriculteurs qui y placent leurs
champs. Les conflits qui résultent de l'obstruction des voies
d'accès du bétail ou de l'occupation des parcours sont,
d'après la plupart des personnes ressources interrogées, les plus
fréquents dans le terroir.
74 La méfiance manifestée à
notre égard, malgré nos efforts de mise en confiance, fait que
nos interlocuteurs disent toujours ne pas connaître de problèmes
entre eux et avec les villageois malgré souvent des faits qui trahissent
ce discours.
75 En 2008 par exemple, certains agriculteurs ont
ensemencé les parcelles de coton dès mi-mai. Cette culture exige
d'être semée très tôt dès les premières
pluies au moment même où les sites de repli pour les troupeaux
même locaux ne connaissent pas encore de repousses attrayantes.

Cliché Sawadogo, août 2009
Photo V-8. Zone de pâture traditionnelle prise d'assaut
par les agriculteurs : en arrière plan, en haut à gauche, un
champ de maïs incrusté dans une zone de pâture traditionnelle
(plateau gravillonnaire) couverte de Loudetia togoensis.
5.3.5.1.3. Une présence des transhumants qui reste
bénéfique à l'économie locale
Sur le plan économique, pendant la période de
présence des transhumants dans le village, le commerce local, surtout
alimentaire, connaît un véritable dynamisme. Certaines
commerçantes de denrées alimentaires comme le couscous à
base de millet, en profitent d'ailleurs, de manière abusive, pour
maximiser leurs gains en majorant parfois les prix. Dans le même sens, le
passage des transhumants est l'occasion pour les éleveurs et
commerçants locaux de faire de bonnes affaires en s'achetant, parfois
à vil prix76, des animaux saisis et vendus aux
enchères ou fatigués et ne pouvant poursuivre le chemin.
5.3.5.2. Relations entre les populations
résidentes
Autrefois, dans le terroir de Kotchari la cohabitation entre
les groupes socioculturels peuls et gourmantchés était
marquée par un niveau relativement faible d'échanges et
frôlait l'indifférence: pas d'échange de femmes en mariage
(ceci reste valable de nos jours), ni de biens ou de services à
l'exception de la vente de lait de bovins par les femmes peules sur la place du
village. Ces relations ont connu une évolution particulière qui
semble sortir quelque peu du schéma observé ailleurs par Guillaud
(1994), Thébaud (1995), Boutrais (1999), Requier-Desjardins (1999) et
Kaboré (2010) dans les régions d'implantation peule plus ancienne
comme le Gourma (Santoir, 1998 ; Benoit, 1998). Kaboré (2010) montre, en
effet, qu'autour de la réserve de Pama Nord, la construction des
maîtrises territoriales avait été marquée par une
inclusion sociale exhaustive qui prenait en compte tous les étrangers,
y
76 On est parfois choqué par l'attitude et
le manque de compassion que les commerçants affichent en de telles
occasions. Accourus d'abord pour jouer aux « conciliateurs », ils
n'hésitent pas à miser des sommes insignifiantes dès que
la vente est ouverte sachant que de toute façon le berger n'a pas le
choix que de vendre quelques têtes pour s'acquitter des amendes qui
dépassent le plus souvent l'entendement.
compris les Peuls. Ces derniers assuraient des fonctions
précises en rapport avec leur statut de pasteurs, ils fournissaient le
lait et le bétail pour les rites, tissaient les toits dans les cours
royales et gardaient les troupeaux des tuteurs. Par ailleurs, ils assuraient
les fonctions de marabouts, de guérisseurs ou de circonciseurs. Il y
avait donc une complémentarité entre systèmes de
production différents (Nori et al. 2008 ; Kaboré, 2010),
ce qui participait à assurer une bonne cohabitation entre la
communauté peule et celle des autochtones notamment les
Gourmantchés.
La situation assez singulière77
d'indifférence, à l'époque, dans notre terroir pouvait
s'expliquer par (i) une certaine méfiance qui était
observée entre ces groupes aux coutumes et rites totalement
différents et qui, en plus, se connaissaient alors à peine, (ii)
le positionnement des campements peuls, d'installation relativement
récente, en des endroits assez éloignés des villages, et
(iii) une relative autosuffisance des autochtones gourmantchés, ceux-ci
ne pratiquant que le petit élevage et le fumier produit suffisait bien
souvent à enfumer les champs de case (les champs de village et de
brousse étaient auparavant laissés tels quels). Par ailleurs, les
Peuls semblaient se suffire au niveau des besoins céréaliers
à partir de leurs propres champs. Avec le temps, la situation a connu
des changements notables. La diversification locale des activités de
production qui émergeait déjà s'est exacerbée (les
Peuls, pour reconstituer un cheptel perdu et aussi pour s'adapter, sont devenus
de grands producteurs agricoles, alors que les Gourmantchés, grâce
à la culture du coton, entretiennent désormais parfois de gros
effectifs de bovins) dans un contexte de forte demande en terre
consécutive à la pression démographique. Cette situation
nouvelle a engendré des besoins d'échange de service (garde des
animaux par exemple) mais aussi et surtout généré de
fortes concurrences pour l'accès aux ressources naturelles du terroir.
Tout ceci évolue dans un contexte de dégradation (baisse de la
quantité et de la qualité du fourrage produit, tarissement plus
rapide des points d'eau, etc.) et de la raréfaction (pression agricole
amenuisant les espaces pâturables) desdites ressources. Cette situation
de rivalité entre Peuls et Gourmantchés a très souvent
été abusement jugée comme résultant d'une
compétition entre pasteurs et agriculteurs pour l'accès aux
ressources naturelles. Pourtant, comme le font observer divers auteurs
notamment Thébaud (2002) et Turner (2004), l'identification de ces
groupes par leur profession n'est plus de nos jours opérante, ceux-ci
s'étant "déspécialisés" (Boutrais, 1999 ;
Kaboré, 2010) avec le développement de l'élevage bovin
chez les agriculteurs gourmantchés et celui de l'agriculture chez les
pasteurs peuls. Pour ces deux auteurs il faut rechercher les raisons à
ces rivalités, dans la baisse des complémentarités
(confiage, contrat de fumure, etc.) et dans les préjugés
qu'entretiennent les autochtones gourmantchés à l'égard
des peuls "étrangers".
Ces dernières années, en effet, les
Gourmantchés à Kotchari, sont de moins en moins enclins à
donner en gardiennage leurs animaux aux bergers peuls dont pourtant ils louent
les qualités. Ils trouvent, comme nous l'avons signalé plus haut,
que ces derniers font montre d'un relâchement coupable lorsque le
bétail ne leur appartient pas et qu'en outre des pertes parfois
77 Cette singularité est cependant à
relativiser car chronologiquement la situation décrite par Kaboré
(2010) est plus ancienne que la notre. L'implantation des premiers Peul
à Kotchari est en effet plus récente et rien ne dit qu'autour de
Pama Nord les mêmes constats dans les relations entre communautés
n'ont pas été observés à un moment donné de
l'évolution.
énormes et aux causes douteuses (attaque de
prédateurs, saisies, vols) sont enregistrées. Toutes ces raisons
entretiennent un climat désormais tendu entre ces groupes, situation
accentuée par les fréquents dégâts causés aux
champs et les empiètements des agriculteurs sur les pâturages et
les pistes à bétail.
5.3.5.3. Relations agroéleveurs -
administration forestière : la question de la fréquentation des
aires protégées.
Kaboré (2010), dans ses recherches autour de la
réserve de Pama Nord, a montré que les populations riveraines de
cette réserve nourrissent un certain nombre de revendications qui ne
sont pas toutes de nature économique et qui traduisent parfois un refus
de l'existence de celle-ci. D'après l'auteur, bien souvent, les
maîtres territoriaux (aînés des lignages) vivent mal la
négation de leur autorité de contrôle sur les terres de ces
aires et l'impossibilité d'y effectuer les rites sacrificiels
ancestraux. Si ces raisons ne sont pas à exclure dans les contestations
observées dans le terroir de Kotchari, celles qui sont affirmées
sont de nature foncière (Zombra, 2008 ; Kièma & Fournier,
2007 ; Ouédraogo K., 2009), c'est-à-dire l'accès aux
terres et aux riches ressources du parc W et de la réserve de la
Kourtiagou. Ces observations sont également rapportées par
Fournier et Toutain (2007) et Kièma S. (2007). Les populations
résidentes qui voient ces aires protégées environnantes
comme une contrainte78 (Zombra, 2008), sont en effet très
dépendantes de celles-ci pour leurs ressources (terres, paille,
pâturages, bois de construction et divers produits forestiers non
ligneux, etc.) qui se font rares en dehors. L'existence de ces entités
nourrit d'ailleurs, à leurs yeux, la compétition foncière
entre agriculture et élevage et est source d'amenuisement des ressources
à l'extérieur par suite de fortes et diverses pressions
d'exploitation79. Cependant, du fait des dispositions
spéciales prévues par la loi forestière (autorisations
saisons saisonnières et collectives d'accès à ces
ressources : produits forestiers non ligneux, paille, etc.) (Kaboré,
2010) qui ne manque cependant pas de limites évidentes (les
périodes d'autorisation ne coïncident pas forcément avec les
périodes de besoin ou de disponibilité de main d'oeuvre compte
tenu des tâches à accomplir au sein des exploitations), les
conflits sur l'exploitation humaine desdites ressources sont limitées
relativement à celles qui ont cours en ce qui concerne l'exploitation
directe (pâturage) par les animaux.
S'il y a un facteur déterminant dans le
déroulement des activités pastorales et dans ses manifestations
actuelles, c'est bien les relations qu'entretiennent les éleveurs
transhumants ou non avec l'administration forestière et qui sont
généralement de nature conflictuelle. Les éleveurs
à Kotchari, surtout les transhumants, reconnaissent rarement
fréquenter les réserves environnantes bien que des indices
semblent indiquer du contraire. En effet, bien souvent les campements sont
érigés à une distance raisonnable (pas assez proche pour
aiguiser les soupçons, pas assez lointaine pour occasionner de grandes
dépenses d'énergie en
78 Ouédraogo K. (2009) invite toutefois
à nuancer cette affirmation. L'auteur en mettant en balance les
bénéfices que procurent les aires protégées aux
populations riveraines (accès aux ressources, revenus
redistribués, etc.) et les coûts additionnels qu'elles
génèrent (saisies et amendes diverses) arrive à la
conclusion qu'en définitive les bénéfices tirés
sont supérieurs aux coûts.
79 Le Mire Pecheux et al. (2000) repris par
Kaboré (2010) montrent, en effet, que Andropogon gayanus, une
espèce utilitaire dans l'artisanat local et comme fourrage ne peut
subsister à une forte pression anthropique.
déplacement) et les animaux, en saison sèche
sont gardés au ras des réserves. Au-delà de leurs
attitudes assez révélatrices, c'est surtout leurs
représentations des aires protégées, aux antipodes de
celles de l'administration forestière, qui rendent leurs rapports
malsains. Voyant dans ces réserves, d'énormes ressources
fourragères « inutilement » préservées et dont
ils contestent la pertinence de la mise sous cloche, ceux-ci imaginent des
scénarios devant leur permettre de les valoriser. Cette posture, aussi
rencontrée autour de la réserve de Pama Nord par Kaboré
(2010), a été d'autant plus renforcée que l'accroissement
du cheptel et l'avancée du front agricole ont été
importants ces dernières années. Dans la pratique, la question de
la fréquentation des réserves reste un mystère difficile
à percer. Cependant, la plupart des Peuls avec qui nos fréquentes
interactions ont fini par instaurer un rapport de confiance, reconnaissent
fréquenter les réserves voisines80. Ils expliquent
même (voir aussi Kpoda, 2010) qu'il existe dans le parc W, des sites
refuges difficilement accessibles par les forestiers et vers lesquels ils se
dirigent automatiquement une fois qu'ils y ont pénétré. Il
existe ainsi un jeu de cache-cache entre les agents veillant à
l'atteinte des objectifs de conservation et les éleveurs pour qui le
salut du bétail en saison sèche réside dans l'accès
à ces aires (Parc W, Réserves et Concessions de chasse de la
zone) où les pâturages sont de meilleure qualité et la
biomasse nettement plus intéressante. Cette situation qui oppose des
parties aux objectifs divergents a rendu délétères les
rapports entre elles, ils se manifestent le plus souvent par des abus divers
notamment le racket81. Les éleveurs, pour atteindre leurs
objectifs utilisent diverses armes comme la corruption82 ou des
schémas de contournement de la vigilance des agents comme les fausses
déclarations. En effet, bon nombre d'entre eux, après
s'être acquittés des frais de CIT (certificat international de
transhumance), simulant ainsi leur désir d'aller en transhumance
transfrontalière au Bénin, se retrouvent ensuite dans les aires
protégées une fois qu'ils se sont éloignés du poste
forestier de Kondio ou lorsqu'ils ont franchi la frontière
béninoise.
5.3.5.4. Relations entre transhumants
Elles sont presque inexistantes. Cependant, avant d'entamer la
campagne de transhumance, les troupeaux sont parfois regroupés en un
seul. Dans ce cas, l'ensemble des bergers avec à leur tête le
Garso et les Mawdo (le berger le plus âgé)
constituent une famille tout au long du parcours et la prise de
décisions tout comme la gestion des difficultés sont collectives.
Il arrive cependant, comme ce fut le cas en 2007 (Fournier et Toutain, 2007)
que,
80 Quand ils le reconnaissent, ils s'empressent
d'ailleurs de préciser qu'ils ont cessé la pratique.
81 Des animaux sont souvent pris hors des
réserves comme ayant pénétré illégalement
dans celles-ci. Cela peut être vrai mais bien souvent il s'agit de
troupeaux pâturant à proximité. Bien souvent la manoeuvre
n'est pas entourée du minimum de précaution nécessaire, le
forestier considérant que de toute façon c'est sa parole contre
celle du berger. Nous avons été personnellement témoin
d'une situation de ce genre et avons tenté, sans succès, de
plaider la cause des bergers qui étaient tous en pleurs. Le dernier
incident en date, relaté par le journal quotidien "Le Pays" du 13 juin
2011, est une parfaite illustration du déficit de confiance qui s'est
installé entre populations et agents forestiers, les derniers ayant
été interdits de séjour dans le terroir par les premiers.
D'après le compte rendu qui en est fait dans le journal, les forestiers,
sachant que la coupe de certains arbres est interdite par la législation
forestière (article 46 du code forestier), s'adonnent eux-mêmes
à la pratique qu'ils mettent ensuite sur le dos des bergers qui
viendraient à s'y aventurer avec leurs animaux.
82 D'après Paris (2002) certains agents
forestiers, contre forte rémunération, acceptent de laisser le
bétail pénétrer dans le parc.
face aux exactions des agents forestiers notamment, des
transhumants isolés fassent front commun. En cette saison en effet, les
éleveurs sédentaires et transhumants, lassés par les
excès des forestiers se sont organisés pour dénoncer la
situation aux autorités administratives de la province.
5 .4. Conclusion
Le terroir de Kotchari dispose de ressources pastorales qui,
sans être exceptionnelles notamment en dehors des réserves, sont
relativement meilleures que dans bien des contrées de la région
notamment les parties plus au nord de la province de la Tapoa. Pour cette
raison et aussi pour sa position stratégique pour les transhumants qui
vont plus bas, il attire de grands troupeaux en saison sèche qui, avec
ceux des troupeaux locaux, valorisent ces ressources en usant de
manières de faire assez différentes.
L'étude des systèmes d'élevage à
Kotchari montre que 4 catégories et sous-catégories
d'éleveurs cohabitent dans un climat relationnel de moins en moins
cordial et de plus en plus suspicieux ou même conflictuel. Ces groupes
d'éleveurs qui vivent des réalités propres, mettent en jeu
des stratégies ayant pour finalité de leur permettre de mieux
tirer profit des potentialités du terroir et au-delà (terroirs
voisins, réserves et Nord-Bénin) en s'adaptant à des
conditions climatiques, écologiques et socioéconomiques
très changeantes
Les éleveurs non transhumants, des agropasteurs et
agroéleveurs résidents, aux effectifs de troupeaux bovins
réduits mais globalement en croissance et ayant pour certains
l'agriculture comme activité principale, arrivent, en combinant
pâturage naturel local et résidus de culture, produits
essentiellement dans leurs parcelles, à assurer les besoins essentiels
de leurs animaux dans le seul espace du terroir de Kotchari ou aux alentours.
Les éleveurs de cette catégorie, la plus importante des trois,
qui ressentent de plus en plus de contraintes foncières et
d'alimentation du bétail, commencent à aller au-delà du
territoire villageois mais s'abstiennent de fréquenter les
réserves voisines. De plus en plus les troupeaux sont divisés en
deux lots pour diverses raisons, la principale étant d'avoir des tailles
raisonnables facilement gérables par les bergers.
Les transhumants résidents ont des troupeaux de taille
importante pouvant dépasser la centaine, ils sont amenés à
transhumer (y compris à fréquenter les réserves) à
cause de l'insuffisance des fourrages naturels, cultivés ou
achetés pour complémentation. La division du troupeau en deux ou
trois lots (bien portants, moins bien portants, petits) est ici motivée
par le besoin de gérer les risques potentiels (particulièrement
les dégâts de champs) et aussi d'alléger ou de mieux
repartir les charges animales sur les parcours. C'est une pratique pertinente
en contexte de morcellement des surfaces pâturables que de grands
troupeaux ne pourraient exploiter sans difficulté.
Les transhumants non résidents, aux troupeaux à
grands effectifs, voient leurs contraintes augmenter d'année en
année avec l'affaiblissement des relations de réciprocité
naguère fortes et les tracasseries croissantes de l'administration
forestière. Les ressources locales naturelles ou cultivées,
soumises à forte concurrence, ne sont plus suffisantes, mais les
éleveurs, qui complémentent peu leurs animaux, restent là,
bien qu'un grand nombre continue

d'aller plus loin, au Bénin notamment. Exposés
à divers risques au cours de leur déplacement et désireux
d'optimiser le rythme de marche, ces éleveurs sont amenés
à scinder leurs troupeaux à deux ou parfois trois lots comme le
groupe précédent.
Tous ces groupes éleveurs voient leurs troupeaux
évoluer vers l'uniformisation de leurs espèces (exception des
Gourmantchés dont les troupeaux sont en recul de taille) et
l'homogénéisation des races de l'espèce bovine vers les
types le plus adaptés localement, notamment ceux qui sont rustiques au
plan alimentaire et qui sont trypanotolérants. Le stock fourrager des
réserves voisines a été et demeure un recours important en
saison sèche notamment pour les transhumants résidents et aussi,
comme on peut le soupçonner, pour les transhumants non résidents.
Par ailleurs, pour diverses raisons liées aux expériences
passées et à leurs traditions ou encore pour obtenir de
meilleures performances, ils font de plus en plus garder leurs troupeaux par de
proches parents qui peuvent cependant être aidés par des bergers
salariés.

171
CHAPITRE VI
LE TROUPEAU AU PATURAGE : LOGIQUES,
REPRESENTATIONS ET REALITES DU TERRAIN

172
6 .1. Introduction
La province de la Tapoa, en particulier la commune de
Tansarga, depuis quelques années, est confrontée à un
accroissement constant des effectifs de bétail, une raréfaction
et une dégradation des espaces pâturables suite à la
pression anthropique et aux mises en place d'aires protégées.
Face à ce que l'on peut qualifier de péril certain pour
l'élevage, il apparaît urgent d'entreprendre des actions visant
à enrayer voire inverser la tendance ainsi observée. Comme nous
l'avons développé plus haut (voir chapitre II) pendant longtemps
le système pastoral a été l'objet de visions
stéréotypées (Moorehead & Lane, 1995 ; Steinfeld et
al. 1997 ; Bary, 1998 ; Nori 2007 ; Nori et al. 2008)
focalisées sur un prétendu manque de rationalité. Les
efforts des politiques et des développeurs ont donc pendant longtemps
été portés principalement vers l'amélioration des
ressources (exemple : ouverture de points d'eau permanents) (Baroin, 2003) et
le contrôle des grandes épizooties (Boutrais, 1999b ;
Touré, 1994 ; Thébaud, 2002), et ont fait l'impasse sur les
éleveurs, leurs modes d'occupation de l'espace, leurs pratiques et les
impacts de celles-ci sur lesdites ressources. Or aucun système
d'exploitation ne peut être bien appréhendé si on dissocie
les potentialités ou contraintes du milieu des techniques mises en
oeuvre par les acteurs (Hoffmann, 1985). Bon nombre de spécialistes de
la question pastorale (Lhoste & Milleville, 1986 ; Sinsin, 1991 ;
Kièma S., 1992 ; Daget & Godron, 1995 ; Kagoné, 2000 ;
Baroin, 2003 ; Diallo, 2006 ; Dongmo, 2009 ; Vall & Diallo, 2009 ; Vall et
al. 2009 ; Dongmo et al. 2010) sont unanimes de nos jours sur
l'intérêt d'appréhender les connaissances ou savoirs
techniques locaux ainsi que les stratégies et les motivations qui les
sous-tendent pour une intervention plus efficiente sur le système
global. Il s'agit de prendre en compte les représentations que les
éleveurs ont des ressources naturelles et d'examiner comment ils
mobilisent ce corpus de savoirs pour les valoriser par leur bétail. Par
ailleurs, les informations recueillies par enquêtes sont parfois
insuffisantes, il existe bien souvent un certain décalage entre ce que
les acteurs, en l'occurrence les éleveurs, disent faire et ce qu'ils
font réellement. A ce propos, Boutrais (1999a) fait observer qu' "un
savoir pastoral est rarement identifiable par lui-même mais à
travers des pratiques qui, souvent, ne sont pas exprimées par les
informateurs". Identifier donc l'écart qui pourrait exister entre
le discours et la réalité par l'observation des faits et les
motivations liées en interrogeant les acteurs peut être un gage de
meilleure appréhension des pratiques et donner des indications sur les
contraintes que ceux-ci rencontrent dans la mise en oeuvre de leurs
connaissances.
Rappel des hypothèses de recherche
Dans le précédent chapitre consacré aux
pratiques pastorales en général, nous avons abordé la
question de l'exploitation des parcours, notamment du choix des lieux
fréquentés par les troupeaux en saison sèche, en
interrogeant les éleveurs. Dans le présent chapitre nous
approfondissons cette question par le suivi des animaux au pâturage. Ces
observations sont faites dans l'objectif d'identifier les principaux facteurs
qui les déterminent et de mettre à l'épreuve les
hypothèses que nous avons faites (encadré VI-1). De
manière concrète il s'agit de comprendre comment les
éleveurs appréhendent et apprécient les milieux qu'ils
exploitent et comment, par leurs troupeaux, ils valorisent leurs connaissances
desdits milieux. Par

ailleurs, la nature de la biomasse végétale des
parcours arides et subarides (flore complexe, diversifiée et
hétérogène) ne renseigne pas bien sur la composition de la
ration ingérée, ce qui donne toute son importance à
l'étude du comportement alimentaire des animaux (Daget et Godron,
1995).
Encadré VI-1. Les hypothèses de recherche
· Dans une localité donnée, les
éleveurs évaluent et classent les pâturages sur des
critères écologiques, mais aussi de risques de conflits, de
risques sanitaires, etc. Cette évaluation/classification change en
fonction des périodes de l'année.
· Dans une localité donnée, le choix
des itinéraires par les animaux et/ou leurs bergers repose sur cette
évaluation/classification locale qui croise une classification des
milieux végétaux et une échelle de risque. Il se fait en
fonction de la distribution spatiotemporelle et de la valeur pastorale des
ressources végétales ainsi que du niveau d'exposition aux
différents risques évoqués ci-dessus.
|
|
6 .2. Matériel et m éthodes
6.2.1. Approche de la connaissance locale des
pâturages
Les entretiens ont été conduits en deux temps
auprès de représentants d'éleveurs, des informateurs
essentiels selon Jackson et al. (1994), choisis dans chacun des
groupes socioculturels peuls et gourmantchés. Organisées dans le
cadre d'un focus group utilisant comme supports divers outils et techniques de
travail participatif (guide d'entretien, brainstorming, matrice de classement,
carte des ressources, etc.), ces discussions de groupe ont permis d'explorer
divers thèmes. Les principaux d'entre eux ont été : (i)
les saisons des éleveurs, (ii) le vocabulaire utilisé par les
éleveurs pour désigner les différentes unités de
pâturages, (iii) les normes et paramètres qu'ils utilisent pour
les qualifier, (iv) la hiérarchisation qu'ils en font suivant la saison
et (v) les périodes optimales à leurs yeux de
fréquentation de ces unités pastorales.
6.2.1 .1. Définition, description et
classement des unités paysagères participatives
La première activité a consisté à
définir consensuellement les unités de paysage présentes
dans le terroir et de recueillir les termes qui les désignent dans la
langue des éleveurs. Après quoi, les participants au focus group
ont procédé à leur description physique (topographie et
géomorphologie) et fonctionnelle (aptitudes culturales et pastorales).
L'étape suivante a consisté à faire une liste des
paramètres utilisés pour les discriminer, à trier et
hiérarchiser ceux-ci et enfin à classer les unités
reconnues sur la base de la notation matricielle pondérée
proposée par Mettrick (1994). Dans cette méthode, à chaque
paramètre d'appréciation (ou de préférence) des
unités paysagères est associée une échelle de
notation

174
(ici de un à vingt) et un coefficient de
pondération qui indique le poids ou l'importance que les éleveurs
attribuent à ce paramètre à la saison correspondante.
Le choix de cette méthode de classement des
unités est motivé par le fait qu'à l'expérience,
nous nous sommes rendu compte que les éleveurs n'accordaient pas la
même considération pour les paramètres qu'ils utilisent.
Pour chaque paramètre à chaque saison, nous
avons recueilli un coefficient d'importance et une note puis avons
procédé au calcul de pondération. Le résultat a
été soumis à l'appréciation des membres du groupe
de discussion et validé à une rencontre ultérieure.
6.2.1 .2. Le repérage et la
matérialisation des unités identifiées
Accompagné d'un ou deux éleveurs suivant les
groupes, nous nous sommes rendu sur le terrain à la suite de chaque
entretien pour procéder au repérage et à la
caractérisation complémentaire des unités définies
en séance de discussion. Cette étape était indispensable
pour permettre de comparer les unités paysagères locales et
celles que nous avions définies par une approche scientifique.
6.2.2. Suivi de troupeaux au pâturage
6.2.2 .1. Approche de suivi de troupeau
Le suivi de troupeau, tel que nous l'avons envisagé,
se veut une étude de cas et constitue une opportunité pour
comprendre le comportement territorial des animaux (Boutrais, 1999 ;
Kagoné, 2000 ; Petit, 2000a ; Ickowicz & Mbaye, 2001 ; Botoni, 2003
; Dongmo, 2009) et de confronter la réalité au discours des
éleveurs. Il s'agit donc d'un moment clé dans la
compréhension des stratégies des éleveurs. Cette
démarche nous a permis de confronter le discours (reflet des
représentations) et la pratique des éleveurs de Kotchari, ce que
Diallo (2006) qualifie d'aller-retour entre pratiques et savoirs techniques
locaux. Le suivi a de plus permis de mieux faire apparaître le calendrier
journalier de l'éleveur et, sur la durée, de reconstituer la
chaîne de pâturage annuelle (Guillaud, 1994 ; Daget & Godron,
1995 ; Kagoné, 2000).
Un suivi diurne83 de troupeau a été
réalisé au cours de 3 jours successifs pour chacun des troupeaux
à chacune des saisons définies par les éleveurs. Le
territoire d'observation prenait en compte tout le territoire pastoral au sens
de César (1994), D'Amico et al. (1995) et Botoni (2003),
c'est-à-dire toute l'aire prospectée par les troupeaux par jour
et au cours de l'année en vue de prélever leur nourriture. Les
observations ont été néanmoins centrées sur le
terroir de Kotchari et le choix des troupeaux à suivre s'est
appuyé sur la typologie des éleveurs définie plus haut
(voir chapitre V). Ainsi, 4 troupeaux (2 éleveurs du groupe C1 et 1
éleveur pour chacun des groupes et C3) ont été
sélectionnés parmi ceux des éleveurs
83 Nous n'ignorons pas le rôle important du
pâturage nocturne et matinal (respectivement Soggunde et
Maanye d'après Diallo (2006)) qui est pratiqué par les
éleveurs peul entre minuit et 7h et qui ont des incidences importantes
sur le pâturage diurne et sur l'état du troupeau.

auxquels le questionnaire avait été
administré et ont été suivis. Initialement, nous voulions
suivre un éleveur pour chacun des quatre groupes et sous-groupes
d'éleveurs identifiés dans le chapitre précédent.
Étant donné que notre travail est beaucoup plus porté sur
les troupeaux bovins, nous n'avons pas suivi le sous-groupe C1-1. En effet,
cette sous-catégorie d'éleveurs, en plus d'être peu
représentée dans le terroir (seulement 15 éleveurs dans
notre échantillon), n'a pas de bovins dans ses troupeaux. Finalement,
tenant compte de la grande taille de la sous-catégorie C1-2 (83
éleveurs), les deux troupeaux (un troupeau gourmantché et un
troupeau peul) à suivre dans C1 ont été choisis en son
sein.
Au total, 16 suivis ont eu lieu en 48 jours (1 suivi
équivaut à 3 jours successifs de sortie) entre novembre 2008 et
octobre 2009 (tableau VI-1). Il faut noter que le troupeau des éleveurs
grands transhumants autochtones au terroir (Trp) n'a pas pu être
retrouvé pendant la saison du Kotoga (saison de transition vers
la saison des pluies) car il était parti en transhumance84.
De même, le troupeau des transhumants allochtones (type TrpC3), n'a pu
être retrouvé après le deuxième suivi qui a eu lieu
au Ceedu (saison sèche chaude)85.
Tableau VI-1. Calendrier de suivi des troupeaux au
pâturage.
Saisons en langues locales
(Peul /
Gourmantché)
|
Types de troupeaux
|
|
Année 2008
|
|
Dabunde / plein Ku fowagu 17, 18, 19 nov. 22, 23, 24
nov. 26, 27, 28 nov. 4, 5, 6 déc.
Ceedu / Ku tontogu 5, 6, 7 avril 11, 12, 13 avril 15,
16, 17 avril 19, 20, 21 avril
Kotoga / A sakoana 26, 27, 28 mai 29, 30, 31 mai
Ndungu / Ku siagu 3, 4, 5 août 6, 7, 8 août
9, 10, 11 août
Yaamde / début Ku fowagu 15, 16, 17 oct. 19,
20, 21 oct. 22, 23, 24 oct.
Les tirets signifient que le suivi n'a pas été
réalisé à la saison donnée et ce à cause de
l'absence du troupeau
TrpC1-2G, troupeau d'éleveurs gourmantchés
sédentaires non transhumants; G = Gourmantché TrpC1-2P, troupeau
d'éleveurs peuls sédentaires non transhumants) ; P = Peul
Trp, troupeau d'éleveurs peuls résidents grands
transhumants) ;
TrpC3, troupeau d'éleveurs peuls allochtones grands
transhumants)
Le détail sur les saisons est donné plus bas
(rubrique résultats).
6.2.2.2. Les activités de l'observateur lors
du suivi
Durant le temps qu'il passe au pâturage, le troupeau
effectue un certain nombre d'activités qui sont: (i) le broutage ou
broutement qui représente la phase pendant laquelle le troupeau est en
train de prélever la nourriture en étant immobile ou mobile ;
(ii) le
84 Selon le propriétaire du troupeau, il a
donné l'instruction aux bergers de l'emmener en transhumance au
Bénin pour le sécuriser pendant un temps en attendant que les
conditions alimentaires (fourrage et eau) dans le terroir
s'améliorent.
85 Nous ne pouvons dire exactement où ce
troupeau est allé à partir du mois d'avril. La première
information qui nous avait été fournie disait qu'il était
retourné vers son terroir d'attache, mais quelques jours après,
alors que nous ne l'attendions plus, il est réapparu avant de rentrer
définitivement en fin juin.

176
déplacement qui représente le moment où
l'ensemble du troupeau se déporte d'un secteur à un autre sans
brouter ; (iii) l'abreuvement et (iv) le repos-rumination, période
où le troupeau est en repos et rumine les aliments
prélevés les moments avant. Ces activités se font dans
divers lieux au cours de la journée.
Au cours du suivi qui se veut actif (méthode de
l'observation participante), l'observateur (nous-même aidé d'un
jeune qui a été recruté et formé à cela)
note divers éléments relatifs aux lieux fréquentés
et aux activités de l'éleveur et de son troupeau. Notons que
l'observation participante, ou méthode de l'observateur participant
(participant-observer en anglais), est une méthode d'étude
ethnologique et sociologique introduite par Bronislaw Malinowski au
début du XXe siècle (Arborio et Fournier, 1999). Très
utilisée par les anthropologues, cette méthode consiste à
étudier une population en réalisant un travail de terrain au
contact direct des individus et en partageant leur mode de vie. Il s'agit
d'étudier une société au plus proche d'elle-même en
se faisant accepter par ses membres et en participant aux activités de
groupe. Pour Bogdan & Taylor (1975), l'observation participante est
« une recherche caractérisée par une période
d'interactions sociales intenses entre le chercheur et les sujets, dans le
milieu de ces derniers. Au cours de cette période des données
sont systématiquement collectées (...) ». Selon Olivier
De Sardan (2003), l'observateur se doit de séjourner de façon
prolongée auprès de son sujet d'étude pour être
à la fois témoin et coacteur (imprégnation) des
phénomènes observés.
Les activités réalisées sont :
· La caractérisation visuelle de l'état
des différentes unités de pâturages traversées :
type, étendue, configuration spatiale, accessibilité, nature et
état des ressources (fourrage, points d'eau, cures salées, etc.),
emprise du bétail (régularité de la fréquentation
animale et degré d'exploitation) et état des brûlis;
· Le repérage au GPS des itinéraires
empruntés par les troupeaux;
· Le chronométrage des principales activités
: déplacement, repos-rumination, broutage, abreuvement.
· L'identification des principales espèces
végétales prélevées par les animaux ou des
espèces dominantes des sites visités.
· Le dialogue informel avec les bergers, axé
essentiellement sur leurs interventions sur les troupeaux, notamment
lorsqu'elles semblent contraires aux déclarations antérieures.
6.2.3. Identification des autres ressources et des
contraintes non fourragères
Une de nos hypothèses fortes était que les
itinéraires choisis par les éleveurs et le comportement
spatio-temporel des animaux sont déterminés par la qualité
des parcours et leur distribution dans l'espace. La qualité
dépend de la valeur (qualité intrinsèque et
quantité) du fourrage offert, mais aussi de l'accessibilité de ce
fourrage, de la présence de points d'eau, de l'absence de risques
sanitaires (épidémies, lieux maudits, etc.) (Lhoste &
Milleville, 1986 ; Guillaud, 1994 ; Lericollais & Faye, 1994 ; Bary, 1998 ;
Kagoné, 2000 ; Riegel, 2002 ; Vall et

al. 2009) et de l'absence d'interdits particuliers de
nature coutumière (bois sacré, etc.) ou légaux (aires
protégées, zones sous gestion particulière).
En conséquence nous avons procédé
à l'inventaire exhaustif et au géoréférencement
(GPS, bibliographie) des éléments suivants dans le terroir : les
aires protégées voisines, les zones villageoises de chasse, les
points d'eau, les pistes à bétail officielles, etc.
Pour les points d'eau, on a précisé leur nature
(puisard, puits traditionnel, puits moderne, forage, mare, rivière,
etc.), les types d'usage dont ils font l'objet (abreuvement des troupeaux et/ou
alimentation humaine) la durée de disponibilité de l'eau dans
l'année.
Enfin, dans une perspective de SIG, une base de données
de départs des feux dans le terroir durant la période de suivi
(2008 et 2009) a été acquise grâce à un partenariat
établi dans le cadre du projet CORUS
607586.
6.2.4. Analyse des données : approche de
système d'information géographique (SIG)
Des cartes des circuits pastoraux quotidiens ont
été réalisées par saison et par troupeau suivant
l'approche des systèmes d'information géographiques (Brunet et
al. 1998) et en utilisant le logiciel MapInfo 7.5. Chaque thème
étudié, considéré comme facteur déterminant
dans le comportement des troupeaux sur les différents secteurs du
parcours, a constitué une « couche d'information ». Au fond
cartographique, représenté par la carte des unités
paysagères pastorales écologiques87, ont ainsi
été superposées diverses « couches »
représentant les itinéraires quotidiens des troupeaux et par
saison, la distribution des points d'eau et leur état (type d'usage,
présence ou absence d'eau), les dates de départ des feux, les
pistes à bétail formelles, etc.
Pour estimer l'importance de chaque unité pastorale
pour les animaux du troupeau, nous avons comptabilisé
séparément le temps de déplacement sans broutage d'une
part et celui consacré au broutage, à l'abreuvement, au
repos-rumination d'autre part. Ceci a permis d'estimer la vitesse du troupeau
sur chacune de ces unités. En effet, un troupeau peut simplement
traverser une unité parce qu'elle se trouve sur le plus court chemin
pour rallier un autre site où se trouve la ressource (pâturage,
point d'eau) qui est convoitée, ce qui n'est pas forcément une
indication de l'intérêt particulier de celle-ci.
Par ailleurs, une analyse comparative (test statistique de
Kruskal-Wallis) a été réalisée entre les temps mis
par types d'activité du troupeau et par éleveur pour toutes les
saisons.
L'analyse du comportement des troupeaux qui s'appuie finalement
sur leurs itinéraires et les résultats des analyses statistiques
s'intéresse à la concordance ou non88 entre ceux-ci
avec
86 Le projet CORUS 6075 a été
conçu autour de notre sujet de thèse dont il a assuré, par
ailleurs, le financement du volet collecte de données. D'autres
thèses notamment celle de Caillault Sébastien de
l'Université de Caen sont conduites dans le même projet. Ce
dernier s'est intéressé à la dynamique des feux à
l'ouest et à l'est du Burkina Faso et une base de données mise en
place à cet effet nous a été bénéfique.
87 Nous aurions pu aussi partir des unités
paysagères localement définies (carte pastorale participative)
étant entendu que les équivalences entre celles-ci et les
unités écologiques ont pu être établies.
88 Nous ne perdons cependant pas de vue sur le fait
qu'un suivi de 3 jours est insuffisant pour tirer des conclusions
générales surtout pour des études de cas-types comme c'est
le cas ici.

178
les déclarations faites par les éleveurs pendant
les différents entretiens et cherche à expliquer les
écarts éventuels observés.
6 .3. Résultats et discussions
6.3.1. Les saisons de l'éleveur
Les enquêtes faites dans un travail
précédent (Sawadogo, 2004) et au cours du présent travail
ont permis de définir le calendrier annuel des éleveurs de
Kotchari. Il comprend quatre saisons principales chez les Peuls tout comme chez
les Gourmantchés (tableau VI-2).
Chez les Peuls, l'année est subdivisée en
Ndungu (juin-début octobre), Yaamde
(octobredébut novembre, saison de transition), Dabunde
(novembre-février, saison sèche froide) et Ceedu
(mars-mai, saison sèche chaude). En plus de ces quatre saisons qui
sont systématiquement citées, certains éleveurs
enquêtés individuellement évoquent deux autres saisons se
situant dans la dernière partie du Ceedu,
précisément dans la période de transition entre celle-ci
et le Ndungu ; elles marquent la rupture avec la saison sèche
et annoncent le début de l'hivernage. Il s'agit de Kotoga (fin
mai) période de survenue des premières pluies mais qui ne sont
pas encore suffisantes pour provoquer des repousses d'herbes précoces et
de Korse (début juin). Pendant le Korse, la
fréquence des pluies commence à être importante et
rapprochée et il devient possible de rencontrer sur les parcours des
repousses (ou Woulande en langue peule) intéressantes pour le
bétail.
Chez les Gourmantchés, le nombre de saisons ainsi que
le découpage de l'année sont presque identiques à ce qui
vient d'être décrit, on note même une correspondance entre
certaines saisons dans les deux groupes socioculturels. On rencontre ainsi,
Ku siagu ou saison de pluie (juin à début octobre) qui
équivaut au Ndungu chez les Peuls, Ku fowagu ou saison
sèche froide (novembre à février) qui va de
Yaamde (début Ku fowagu) à Dabunde
(Fin Ku fowagu), Ku tontogu (mars à début
mai) qui correspond au Ceedu et A sakoana (fin
mai-début juin) qui comprend à la fois Kotoga et
Korse. Précisons que le terme Li fali désigne
l'ensemble de la saison sèche (Ku fowagu + Ku tontogu + A
sakoana).
Le recueil des saisons des éleveurs a très
souvent été effectué lors d'études sur les
pratiques des éleveurs, le plus souvent peuls, par des chercheurs qui
s'intéressaient au calendrier fourrager quotidien ou à la
chaîne annuelle de pâturages. Le nombre de saisons
distinguées est en général de 5 ou 6, toujours les
mêmes, bien que quelques nuances liées au contexte local, soient
parfois observées. Certaines différences de terminologie peuvent
être notées mais on retrouve les saisons de notre calendrier avec
parfois un léger décalage dans le temps lié au contexte
climatique de la zone d'étude, mais aussi aux imprécisions dans
les limites et l'étendue de chacune de ces saisons. Les trois saisons
principales du groupe peul que sont le Ndungu, le Dabunde et
le Ceedu se retrouvent dans la plupart des calendriers
proposés.
Tableau VI-2. Calendrier pastoral comparé de
communautés d'éleveurs dans divers terroirs en Afrique de
l'Ouest.
|
Localités et sources
|
Saisons
|
|
Juin
|
Juillet
|
Août
|
Septembre
|
Octobre
|
Novembre
|
Décembre
|
Janvier
|
Février
|
Mars
|
Avril
|
Mai
|
|
Saison des pluies Saison sèche froide
|
Saison sèche chaude
|
|
Kotchari (Peul) Présente étude
|
Kor
*
|
Ndungu
|
Yaamde
|
Dabunde
|
Ceedu
|
Ko
t*
|
|
Kotchari
(Gourmantché) Présente
étude
|
Ku siagu
|
Ku fowagu
|
Ku tontogu
|
A
Sak*
|
|
Centre-sud, Burkina (Kagoné, 2000)
|
Gataaje
|
Nduungu
|
Yaamde
|
Dabbunde
|
Ceedu
|
|
Nigeria (Ferguson et al. 1979)
|
Seeto
|
Dungu
|
Yawal
|
Dabunde
|
Ceedu
|
Seeto
|
|
Koumbia (Vall & Diallo, 2009 ; Vall et al.
2009)
|
Gatadje
|
Nduungu
|
Djaamde
|
Dabuunde
|
Ceedu
|
Gatadje
|
|
Yagha/Burkina (Thébaud, 1999)
|
Korsol
|
Ndunngu
|
Yammde
|
Dabbunde
|
Ceedu
|
|
Diffa / Niger (Thébaud, 1999)
|
Kokoobe
|
Korsol
|
Ndunngu
|
Yaawol
|
Corrol
|
Ceedu
|
|
Cameroun (Dongmo et al. 2006)
|
Seeto
|
Ndungu
|
Yamde
|
Dabunde
|
Cheedu
|
Seeto
|
|
Périphérie du W, Niger (Riegel, 2002)
|
|
Loddo
|
Yamnde
|
Dabbunde
|
Ceedu
|
|
|
Périphérie du W, Bénin (Karimama) (Tamou,
2002)
|
Seeto
|
Dungu
|
Yaawol
|
Dabuney
|
Tchendwe
|
Seeto
|
|
Djenontin, 2010 (NordBénin
|
Seeto
|
Nduungu
|
Djaamdè
|
Dabuundè
|
Ceedu
|
Seeto
|
En gras, les résultats de la présente
étude; * A sak = A sakoana; Kot = Kotoga; Kor = Korse.

180
Les quelques différences notables se situent surtout au
niveau des saisons de transition vers la saison humide, Kotoga-Korse
ou vers la saison sèche, Yaamde.
6.3.2. Une valeur pastorale qui change en fonction de
plusieurs paramètres
6.3.2. 1. Les unités paysagères
pastorales participatives : types et caractéristiques
Les tableaux VI-3 & VI-4 présentent les
unités définies par les éleveurs de Kotchari, assez
similaire chez les deux principaux groupes socioculturels du terroir, les Peuls
et les Gourmantchés. La classification se fonde à la fois sur la
topographie (prépondérante chez les Peuls) et le type de sol
(prépondérante chez les Gourmantchés). Par ailleurs, le
niveau de subdivision, les toponymes utilisés et les sens qui leur sont
donnés peuvent varier selon les interlocuteurs, mais il y a en
général une plus grande diversité de termes chez les
Gourmantchés pour désigner une même unité.
Chez les Peuls, il existe six unités. Si la
majorité confond Loubare et Loubal, deux unités
de plaines hydromorphes, certains les distinguent bien. La première,
très fertile et apte à la culture du sorgho, connaît une
inondation temporaire, la deuxième est engorgée d'eau en saison
pluvieuse et donc impropre à la fois aux cultures et à
l'exploitation pastorale. Par ailleurs, entre Tilde et
Djolde, unité de haut-glacis et plaines, la différence
tient seulement à l'étendue de l'unité, la première
étant plus vaste. Dans les unités de plateaux (Banouol),
les secteurs à sols sableux sont parfois appelés Senore
(littéralement, endroit de sable), il s'agit de lieux propices
à la culture du petit mil. L'ensemble des éleveurs peuls
désignent les bas-fonds par le terme Celol, mais certains
d'entre eux y distinguent des bas-fonds particuliers ou Lougere qui
sont, en fait, de grandes dépressions caractérisées par la
présence de l'espèce Acacia seyal. Chez les Peuls,
toutes les unités cultivées forment les Gese qui
deviennent de bons pâturages à l'arrière saison,
c'est-à-dire pendant le Dabunde dès la fin des
récoltes.
Tableau VI-3. Les unités pastorales paysagères
distinguées les Peuls et leurs caractéristiques
Nom en Caractéristiques observées sur le
Principales espèces végétales citées et
Fulfuldé89 terrain retrouvées
sur le terrain
Waamde Collines (blocs rocheux) Detarium
microcarpum, Combretum
nigricans, Combretum micranthum, Schizachyrium exile,
Loudetia
togoensis, Andropogon pseudapricus
|
Banouol Plateaux et hauts glacis à sol
gravillonnaire squelettique à peu profond
|
Combretum glutinosum, Combretum molle, Melochia
corchorifolia, Loudetia togoensis, Sida sp.
|
Djolde / Hauts glacis et plaines sableuses à
Vitellaria paradoxa, Combretum
Tilde sablo limoneuses collinum, Parkia biglobosa,
Diospyros
mespiliformis, Andropogon
89 Langue des Peul

pseudapricus, Brachiaria lata, Zornia glochidiata,
Alysicarpus ovalifolius
Loubare Plaines marécageuses (inondation
Acacia seyal, Anogeissus leiocarpa,
temporaire, sols profonds argileux, Eragrostis sp., Setaria
pallide-fusca
s'asséchant rapidement et craquelant
si sec)
Loubal Plaines hydromorphes à inondation
Combretum molle, Terminalia sp.,
permanente en saison des pluies ; sols Cyperus sp.,
Sorgastrum bipennatum, variés : argileux, limono-argileux,
Schizachyrium brevifolium
limono-sableux
Celol / Bas-fonds à sols profonds Anogeissus
leiocarpa, Mitragyna
Lougere hydromorphes argilo-limoneux inermis, Oryza
longistaminata, Ipomoea
sp., Cassia mimosoides
Les Gourmantchés, comme on l'a dit plus haut,
classifient les milieux en mettant en avant les potentialités culturales
de leurs sols. Les mêmes types d'unités que chez les Peuls sont
distingués, mais huit catégories sont reconnues. Une même
unité peut être désignée par deux à trois
termes comme c'est le cas pour les glacis (Li wangeli ou Li
bangbanli) et les plaines à inondation temporaire (Li tinbuali
ou Li loubri ou encore Bualibuanli). Ici aussi, tous les
champs, généralement rencontrés sur les plaines non
inondables (Li tinmuali ou Ku tutanbigu) ou temporairement
inondables (Li tinbuali ou Li loubri), certains plateaux
(U gbanu et Ku tankiangu) et certains secteurs des bas-fonds (Ku
bagu) deviennent des pâturages (I kuanu) après les
récoltes.
Tableau VI-4. Les unités pastorales paysagères
distinguées par les Gourmantchés et leurs
caractéristiques
Nom en Caractéristiques observées sur
Principales espèces végétales citées
Goulmancema90 le terrain et retrouvées
sur le terrain
Li guali Collines (blocs rocheux) Combretum
micranthum, Detarium
microcarpum, Andropogon
pseudapricus, Schizachyrium exile
|
Ku tankiangu Plateaux cuirassés avec
affleurements rocheux à sols squelettiques
|
Combretum glutinosum,Acacia gourmaensis, Andropogon
pseudapricus, Loudetia togoensis
|
U gbanu Plateaux et glacis à sol peu
Combretum glutinosum, Andropogon
profond gravillonnaire pseudapricus, Loudetia
togoensis,
Schoenefeldia gracilis
Li wangeli /Li Glacis dégradés
(dénudés) sans Acacia seyal, Balanites aegyptiaca,
bangbanli valeur agronomique ou pastorale Loudetia
togoensis, Schoenefeldia
gracilis
90 Langue des Gourmantchés

182
|
Li tinmuali /Ku tutanbigu
|
Plaines non inondables et moyens glacis à sols
rougeâtre moyennement profonds, sableux à sablo-limoneux
|
Vitellaria paradoxa, Combretum collinum, Andropogon
pseudapricus, Schizachyrium exile, Zornia
glochidiata
|
|
Li tinbuali /Li loubri / Bualibuanli
|
Plaines hydromorphes à inondation temporaire, sol
noirâtre profond argileux à argilolimoneux
|
Acacia seyal, Lannea acida, Barchiaria jubata, Spermacoce
sp.
|
Ku pugu Plaines hydromorphes (inondation Terminalia
macroptera, Vitellaria
permanente en saison de pluie ; paradoxa, Sorgastrum
bipennatum,
sols variés : argileux, limono Pennisetum
pedicellatum, Kyllinga sp.
argileux, limono-sableux)
Ku bagu Bas-fonds à sols profonds Mitragyna
inermis, Terminalia
hydromorphes argilo-limoneux laxiflora, Echinochloa sp.,
Oryza sp.,
Scleria sp. Sporobolus pyrimidalis
6.3.2.2. Unités paysagères pastorales
participatives versus unités paysagères pastorales
écologiques
Le croisement des unités reconnues par les Peuls et
Gourmantchés entre elles, puis avec celles établies à
partir des analyses écologiques (chapitre IV), montre leur concordance
quasi-parfaite (tableau VI-5). L'unité de savane arbustive claire de
plateaux et hauts glacis (UPP4), la plus vaste du terroir est la plus
diversifiée en particulier chez les Gourmantchés. On peut
remarquer par ailleurs que l'unité Li wangeli chez les
Gourmantchés est sans valeur aucune en termes agronomiques et pastoraux
car elle est totalement dénudée.
Tableau VI-5. Correspondance entre unités
paysagères écologiques et locales
|
Unités paysagères
pastorales
écologiques
|
Unités paysagères pastorales «
participatives »
|
|
Chez les Peuls Chez les Gourmantchés
|
UPP1 Celol / Lougere Ku bagu
UPP2 Loubal Ku pugu
UPP3 Loubare Li tinbuali
UPP4 Banouol / Djolde U gbanu / Li wangeli / Li
tinmuali
UPP5 Djolde / Banouol (Gese) U gbanu / Li tinmuali (I
kuanu)
UPP6 Waamde Li guali /Ku tankiangu
Légende
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;

UPP6, unité de savane arbustive claire de buttes rocheuses
et cuirassées.
Cette division des parcours en unités pastorales par
les Peuls est largement évoquée dans de nombreux travaux conduits
en zone sahélienne (Yagha au Burkina et Diffa au Niger Oriental:
Thébaud, 1999), en zone soudanienne du Burkina Faso (Kagoné, 2000
; Diallo, 2006 ; Vall & Diallo, 2009) et du Nord Cameroun (Dongmo, 2009 ;
Dongmo et al. 2009; Dongmo et al. 2010) et en milieu lobi en
zone soudano guinéenne du Nord-est ivoirien (Hoffmann, 1985). Comme pour
les saisons des éleveurs, il existe une similitude assez forte entre nos
unités paysagères participatives et ces classifications qui
s'appuient généralement sur les mêmes critères de
géomorphologie, de toposéquence et de leur utilisation pour les
activités agropastorales. Des nuances existent cependant. Par exemple,
les pâturages de collines (ici Waamde et Li guali) sont
désignés par Calle au Centre-sud du Burkina
(Kagoné, 2000) ou par Fukkaawo / Foukah
(respectivement dans l'ouest burkinabè ou au Cameroun (Vall &
Diallo, 2009 ; Dongmo, 2009) alors que les plateaux, représentant
généralement les brousses des terroirs que ces auteurs ont
étudié, sont appelés Ferlo. Les mêmes
auteurs désignent les plaines à inondation temporaire ou non (ici
Loubal et Loubare chez les Peuls et Ku bagu et
Li tinbuali chez les Gourmantchés) par Bolaaje
(Kagoné, 2000), Bolaawo (Diallo, 2006 ; Vall & Diallo,
2009) ou encore Bolaare (Dongmo, 2009 ; Dongmo et al. 2009 ;
Dongmo et al. 2010). Dans leurs travaux, les plaines sont, comme dans
notre cas, désignées autrement selon qu'elles sont
occupées par les cultures, couvertes de résidus de
récoltes (Gese) ou mises en jachères (Jobuje
pour jeunes jachères et Soynere pour les jachères plus
âgées).
6.3.2.3. Connaissance, représentations et
usages : les catégories pastorales et les logiques de leur
fréquentation par les éleveurs
6.3.2.3.1. Les paramètres considérés
et leur importance au fil des saisons
Les éleveurs, qu'ils soient Gourmantchés ou
Peuls, classifient les pâturages selon un certain nombre de
paramètres dont l'importance varie d'une saison à l'autre. Il a
été possible, par la matrice de classement pondéré,
de mettre en évidence cette hiérarchie. Les paramètres
pris en compte par les éleveurs sont : la disponibilité en eau et
la possibilité d'y accéder, la quantité d'herbe offerte,
la qualité de cette herbe, la praticabilité (en lien avec le
risque d'embourbement) et la probabilité d'être exposé
à un risque. Les causes de ce risque peuvent être la
proximité des champs (risque de dégât et donc de conflit),
ou des aires protégées (empiètement illégal), la
présence de "lieux maudits"91 ou de tout autre risque
sanitaire. Les deux derniers paramètres de risque (lieux maudits et
risque sanitaire) semblent incertains en périphérie dans notre
terroir et ont donc été retirés de la liste après
avoir été retenus dans la liste. Tout comme la recherche de cures
salées qui a été citée par les Peuls mais
très vite retirée alors que leur existence dans le parc W n'est
pas un secret (Kpoda, 2010). Cela semble s'apparenter à une manoeuvre
maladroite des éleveurs pour cacher leur fréquentation des
91 "Lieux maudits" (ou "champs maudits") est un
terme technique en zootechnie qui qualifie les lieux infestés par
l'agent pathogène du charbon bactéridien ; l'herbe qui y pousse
est également infestée et source de contamination pour le
bétail.

184
réserves voisines. Dans des travaux similaires dans
l'ouest burkinabè (Vall & Diallo 2009) et au nord Cameroun (Vall et
al. 2009), ces paramètres en été groupés
en paramètres fonctionnels (disponibilité et accessibilité
à l'eau ; disponibilité et qualité du fourrage) et en
paramètres de risques (conflits potentiels).
Des coefficients ont été ensuite attribués
à chacun des paramètres par saison selon l'importance que les
éleveurs leur accordent.
Chez les Peuls (tableau VI-6) tout comme chez les
Gourmantchés (tableau VI-7), la praticabilité, l'absence de
risques divers notamment les possibilités de dégâts
champêtres ainsi que, dans une moindre mesure, la qualité des
fourrages sont les paramètres les plus importants en saison des pluies
(Ndungu / Ku siagu) et en fin de campagne agricole au moment
où les récoltes sont attendues ou ont commencé mais ne
sont pas encore achevées (Yaamde / Ku fowagu).
Tableau VI-6. Paramètres de classification des
unités paysagères pastorales : coefficients de pondération
chez les Peuls
|
Paramètres
|
Ndungu
(juin - début
oct.)
|
Yaamde (oct.
- début
nov.)
|
Dabunde (mi -
nov. -
févr.)
|
Ceedu (mars
- mai)
|
Kotoga/Korse
(fin mai-
début
juin)
|
|
Eau
|
1
|
1
|
3
|
5
|
5
|
|
Fourrage de bonne qualité
|
3
|
5
|
4
|
1
|
2
|
|
Fourrage en grande quantité
|
1
|
2
|
4
|
5
|
4
|
|
Lieu praticable
|
5
|
3
|
1
|
---
|
---
|
|
Lieu sans risques
|
5
|
4
|
2
|
1
|
1
|
Les tirés signifient que le paramètre n'est pas
pris en compte à la saison correspondante
Au sortir de la campagne agricole, la praticabilité
(pour cause de dessèchement progressif des parcours), le facteur risque
(pour cause de récolte de champs) sont peu ou pas
considérés alors que la quantité du fourrage et la
présence de l'eau prennent de plus en plus de l'importance aux yeux des
éleveurs, la qualité du fourrage devenant secondaire. En saison
sèche chaude (Ceedu et Kotoga ou Ku tontogu
et A sakoana), c'est surtout la présence de l'eau qui
confère à l'entité pâturée son
intérêt pastoral.
Il est important de noter que le facteur de risque prend plus
d'importance pour les Peuls que pour les Gourmantchés; il n'est
d'ailleurs pas du tout pris en compte chez ces derniers dès le
début de la saison sèche. Aux yeux des Gourmantchés, il
n'existe pas de risque dès que les récoltes sont achevées,
alors que chez les Peuls dont les troupeaux pâturent parfois au ras des
réserves, le risque demeure même s'il n'est plus aussi important
qu'auparavant. De même, en saison sèche la praticabilité
des pâturages reste un paramètre crucial pendant encore un moment
plus long pour les Peuls que pour les Gourmantchés, ceci probablement
parce que les premiers vont exploiter l'herbe verte des zones inondées
dès que c'est possible. Ils exercent sur ces unités très
humides une surveillance particulièrement serrée en vue d'y
accéder au plus vite.

Tableau VI-7. Paramètres de classification des
unités paysagères pastorales : coefficients de pondération
chez les Gourmantchés
|
Paramètres
|
Ku siagu
(juin -
début oct.)
|
Ku fowagu
(oct. -
février)
|
Ku tontogu
(mars -
début mai)
|
A sakoana
(fin mai -
début juin)
|
|
Eau
|
1
|
4
|
5
|
5
|
|
Fourrage de bonne qualité
|
4
|
5
|
1
|
1
|
|
Fourrage en grande quantité
|
2
|
3
|
4
|
4
|
|
Lieu praticable
|
5
|
---
|
---
|
---
|
|
Lieu sans risques
|
4
|
2
|
---
|
---
|
Les tirés signifient que le paramètre n'est pas
pris en compte à la saison correspondante
6.3.2.3.2. La chaîne de pâturage
saisonnière à dire d'acteurs: les représentations que les
éleveurs ont de leurs pâturages
Le résultat validé par les éleveurs de la
notation de chaque unité suivant tous les paramètres
(échelle de un à vingt), puis de la pondération est
donnée dans les figures VI-1 et VI-2.
Les collines (Waamde chez les Peuls ou Li
guali chez les Gourmantchés) et les plateaux plus ou moins
cuirassés ou gravillonnaires (Banouol chez les Peuls, Ku
tankiangu et U gbanu chez les Gourmantchés), sont les
lieux préférés pour la pâture de saison humide
(Ndungu ou Ku siagu). Les hauts glacis ainsi que les plaines
sèches aux sols sableux à sablolimoneux (respectivement
Djolde et Li tinmuali) sont également
fréquentés, mais à un degré moindre, surtout chez
les Peuls. Trois paramètres, l'impraticabilité due à
l'humidité, les risques divers (notamment conflits consécutifs
aux empiètements de champs et maladies consécutives à
l'humidité) et la qualité du fourrage, dans une moindre mesure,
sont considérés comme déterminants dans ces choix de
pâturage de saison humide. En effet, la quantité du fourrage et la
disponibilité en eau d'abreuvement sont des préoccupations
secondaires à cette période. Partout dans le terroir, le troupeau
dispose alors de suffisamment de fourrage et d'eau pour subvenir à ses
besoins. Une particularité des Gourmantchés est de
fréquenter même les abords des plaines argileuses en cette saison,
bien qu'elles fassent partie des zones les plus cultivées. Ceci semble
possible grâce à la taille modérée de leurs
troupeaux (en moyenne : cinq bovins ; treize ovins et treize caprins voir
tableau V-8, chapitre V), qui leur permet de rester dans les broussailles des
interstices inter-champs.
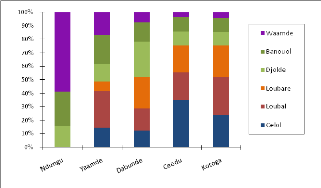

186
Figure VI-1. Chaîne de pâturage annuelle dans la
représentation des éleveurs peuls.
Ndungu : juin à début octobre
Yaamde : octobre à début novembre
Dabunde : mi-novembre à février Ceedu : mars
à mai
Kotoga : fin mai à début juin
Celol = UPP1 (unité de savane arborée sur
sol profond hydromorphe à pseudogley de surface)
Loubal = UPP2 (unité de savane boisée
claire sur plaine inondable et sol hydromorphe à pseudogley de
surface) Loubare = UPP3 (unité de savane arbustive de moyen et
bas glacis sur sols ferrugineux tropicaux à tâches et
concrétions)
Djolde / Banouol = UPP4 (unité de savane
arbustive claire de plateaux et hauts glacis sur sols ferrugineux tropicaux
lessivés indurés)
Djolde / Banouol (champs) = UPP5 (UPP 4 cultivé ;
unité de mosaïque agroforestière sur sols ferrugineux
tropicaux lessivés indurés)
Waamde = UPP6 (unité de savane arbustive claire
de buttes rocheuses et cuirassées).
Pendant le Yaamde (début Ko fowagu
dans le calendrier gourmantché), saison de transition qui correspond
à la période d'entame des récoltes, les conditions
d'ensemble ont peu évolué mais l'herbe, arrivée à
maturité depuis fin septembre, commence à perdre de sa
qualité par endroits et ce paramètre commence à
prévaloir dans la recherche des pâturages. Le début
d'assèchement du Loubal (plaine inondable non cultivée),
devenu praticable et où le retrait de l'eau libère de l'herbe
fraîche et jeune par endroits, y attire les troupeaux. Les abords du
Loubare et du Celol se sont également
asséchés et peuvent être exploités par ces
éleveurs en particulier les Peuls. Les troupeaux gourmantchés,
exploitent alors pratiquement les mêmes milieux que pendant la
période précédente mais sollicitent aussi fortement la
partie asséchée des plaines inondables (Ku pugu). Ils
sont par ailleurs moins fréquents sur les collines (Li guali).
Plus généralement, les animaux continuent d'aller sur les
hauteurs car les risques de dégâts champêtres sont encore
importants en contrebas, mais ils commencent à en descendre car la
végétation y devient répulsive : la paille y a
déjà, beaucoup plus qu'ailleurs, entamé son
jaunissement.
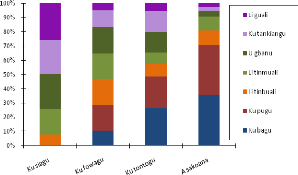

Figure VI-2. Chaîne de pâturage annuelle dans la
représentation des agroéleveurs gourmantchés.
D_Ku fowagu (début Ku fowagu):
mi-octobre à début-novembre P_Ku fowagu (plein Ku
fowagu): novembre à février
Ku tontogu : mars à début mai
A sakoana : fin mai à début juin
Ku siagu : juin à début octobre
Ku bagu= UPP1 (unité de savane arborée sur
sol profond hydromorphe à pseudogley de surface)
Ku pugu= UPP2 (unité de savane boisée
claire sur plaine inondable et sol hydromorphe à pseudogley de
surface) Li tinbuali = UPP3 (unité de savane arbustive de moyen
et bas glacis sur sols ferrugineux tropicaux à tâches et
concrétions)
U gbanu / Li tinmuali = UPP4 (unité de savane
arbustive claire de plateaux et hauts glacis sur sols ferrugineux tropicaux
lessivés indurés)
U gbanu / Li tinmuali (champs) = UPP5 (UPP 4
cultivé ; unité de mosaïque agroforestière sur sols
ferrugineux tropicaux lessivés indurés)
Li guali / Ku tankiangu = UPP6 (unité de savane
arbustive claire de buttes rocheuses et cuirassées).
Le Dabunde (plein Ku fowagu), saison
sèche froide, est la période où les récoltes sont
achevées et la baisse de la qualité ainsi que de la
quantité du fourrage naturel est alors partiellement compensée
par les résidus de culture (Gnagnical ou Nyale
d'après Kagoné, 2000) dans les unités
cultivées (Djolde, Banouol et Loubare chez les Peuls
; U gbanu, Li tinmuali et Li tinbuali chez les
Gourmantchés) alors appelées Gese (respectivement I
kuanu). À cette période, les troupeaux sont très
présents sur ces unités où la vaine
pâture92 est admise. C'est aussi vers la fin de cette
période que les feux de brousse atteignent leur pic et les
éleveurs, surtout les Peuls, retournent vers les plaines inondables
(Loubal ou Ku pugu) qui ont été
brûlées pour bénéficier des jeunes repousses
appelées Woulande.
La saison de soudure qui correspond à la saison
sèche chaude (Ceedu ou Ku tontogu) va être
marquée par l'occupation tout azimut des bas-fonds (Celol ou
Ku bagu) et de leurs environs. Ces unités, dont certaines ont
encore des filets d'eau, offrent de l'herbe fraîche mais en très
faible quantité ; elles permettent surtout d'abreuver les animaux
grâce à des puisards
92 La vaine pâture se définit comme
« un droit d'usage qui permet aux éleveurs de faire paître
gratuitement leur bétail dans des champs ne leur appartenant pas,
après les récoltes ou lors d'une jachère »
(Inter-réseaux Développement rural, 2009)

188
creusés manuellement. L'herbe offerte ne suffit plus
à assurer les besoins d'entretien des animaux et, tout autre lieu,
à distance raisonnable des points d'eau et où il est possible
d'avoir de l'herbe - de la paille sans valeur appelée Gena ou,
selon Thébaud (1999), Geene - est fréquenté.
C'est la période des transhumances et de la complémentation des
animaux. Certains éleveurs gardent leurs animaux le matin à
proximité des habitations, généralement sur les
unités de plateaux où ils bénéficient des
résidus de culture en stock.
Les premières pluies annoncent l'hivernage (saison :
Kotoga ou A sakoana) et font venir des repousses sur les
plaines hydromorphes (Loubal ou Ku pugu) et les autres
unités de bas de toposéquence, ce qui y attire les animaux. Quand
les pluies deviennent importantes (période du Korse), les
repousses le deviennent également sur tout le terroir notamment sur les
plaines plus hautes et les plateaux et elles sont facilement exploitées
avant les semis. À cette période, le terroir accueille ses
derniers transhumants, les Korseje (Sawadogo, 2004), venus à la
rencontre de cette herbe fraîche.
Il est manifeste que les éleveurs peuls et
gourmantchés ont une bonne connaissance des pâturages, de leur
dynamique et des périodes optimales de leur exploitation. Parmi les
paramètres auxquels ils se réfèrent pour qualifier et
choisir leurs pâturages, certains comme la praticabilité,
l'exposition aux risques de conflits et, dans une moindre mesure, la
qualité de la ressource pastorale (le fourrage en particulier) sont
déterminants en saison pluvieuse et vont structurer l'occupation de
l'espace. Les meilleurs pâturages sont alors ceux situés à
distance des champs et ne présentant pas de risque d'embourbement. En
saison sèche en revanche, la quantité de la ressource
fourragère dans un premier temps, puis la disponibilité en eau
vont être mises en avant dans le choix des pâturages à
fréquenter, un pâturage sera d'autant plus fréquenté
qu'il remplira les deux conditions. Les éleveurs qui font preuve d'un
bon choix d'itinéraires permettant un bon rapport coût
(énergie dépensée dans les déplacements) / avantage
(aliments et eau de boisson rencontrés sur ces parcours), ont ainsi des
animaux qui traversent cette saison sans grand dommage.
6.3.3. Les animaux au pâturage : variations
saisonnières de l'utiisation des pâturages
6.3.3 .1. Les caractéristiques des troupeaux
suivis
Les caractéristiques des quatre troupeaux qui ont fait
l'objet de suivi au pâturage sont dans le tableau VI-8.
- Le troupeau gourmantché résident (TrpC1-2G)
est constitué de bovins et ovins. Appartenant à Combary Soali, il
est gardé alternativement par deux jeunes bergers qui sont ses propres
fils (Combary Ombua, aîné et berger principal et Combary Palo,
berger suppléant). Basé dans le quartier Koukongou, le troupeau
est constitué de Barbaji et de Gurmaji ; il est
gardé nuitamment dans un campement sommaire qui se situe à
proximité de la concession familiale en saison sèche (plein
Ku fowagu, Ku tontogu et A sakoana) ou au flanc de la partie
voisine de la chaîne du Gobnangou, tout près de la carrière
de phosphate en saison de pluie (Ku siagu et début Ku
fowagu).

- Le troupeau peul sédentaire (TrpC1-2P) appartient
à un éleveur très anciennement installé dans le
terroir (quartier Kotchari-Peul), Diallo Oumarou dit Yobi. Un jeune neveu du
nom de Sondé Harouna assure la garde de ce troupeau pendant le jour.
Mais il est fréquent qu'il se fasse remplacer par le propriétaire
lui-même qui assure aussi la garde de nuit. Pendant la saison de soudure
(Ceedu) et les saisons de transition (Kotoga,
Korse), où le troupeau s'éloigne parfois de la
concession familiale (petite transhumance vers Gnimboama), ils sont deux
à en assurer la garde. Ce troupeau a la même composition que le
précédent avec cependant plus de bovins et moins d'ovins. Ici
aussi, en saison pluvieuse, le campement du troupeau est placé sur le
flanc de la chaîne du Gobnangou à proximité du couloir de
transhumance officiel. En saison sèche, par contre, il est
délocalisé plus bas auprès de la concession, endroit
devenu sec et où les animaux sont plus en sécurité.
- Le troupeau résident transhumant (Trp) est celui d'un
éleveur résident (Diawara Soumaïla) mais qui est
amené à transhumer en fin de saison sèche (Ceedu
ou Kotoga/Korse). Ce troupeau est de taille plus importante que les
précédents, quelques têtes de Kiwali (6 dont 2
mâles et 4 femelles) s'ajoutent aux races principales (Barbaji
& Gurmaji). Il est gardé par trois bergers dont le jeune
frère du propriétaire (Diawara Adou) et deux salariés
(Diallo Manga et Diallo Soumana) en saison de soudure au moment de la
transhumance, mais seulement par le jeune frère en saison pluvieuse.
Alors que la concession familiale se trouve dans le quartier Banduo, ce
troupeau se trouve dans un campement mobile situé plus bas, entre Banduo
et Pielgou, dans un endroit où la pression animale est relativement plus
faible.
- Le troupeau des transhumants non résidents (TrpC3)
est le plus important en effectif bovin, il comprend essentiellement la race
Gurmaji, mais on rencontre aussi quelques têtes de
Jaliji en son sein. C'est un troupeau de transhumants comprenant peu
de vaches (au nombre de 56 têtes) venant d'un village de la commune de
Botou au Nord de la province. Propriété de deux personnes, il est
conduit par l'un des deux propriétaires (Baadio Idrissa) et par deux
neveux (Sondé Hama, Lido Issoufi) du second propriétaire (Baadio
Issouf). Ce troupeau, présent à Kotchari seulement pendant la
saison sèche froide (Dabunde) et la saison sèche chaude
(Ceedu), a son campement érigé sur les buttes
cuirassées (UPP6) situées entre le quartier Banduo et le parc du
W.
Tableau VI-8. Caractéristiques des troupeaux suivis
|
Types de Troupeau
|
Propriétaire du troupeau
|
Composition spécifique et effectif
|
Races bovines présentes
|
Bergers (nom et âge)
|
|
TrpC1-2G
|
Combary Soali
|
Mixte : 24 bovins, 53 ovins
|
Barbadji, Gurmaji
|
Combary Ombua (20 ans) ou
|
|
|
|
|
Combary Palou (18 ans)
|
|
TrpC1-2P
|
Diallo Oumarou (Yobi)
|
Mixte : 43 bovins, 32 ovins
|
Barbadji, Gurmaji
|
Diallo Harouna (18 ans) ou Diallo Yobi (40 ans)
|
|
Trp
|
Diawara Soumaila
|
Monospécifique : 58 bovins
|
Barbadji,
Gurmaji (6 têtes de Kiwali)
|
Diawara Adou (21 ans), Diallo Manga (22 ans), Diallo Soumana (19
ans)
|
|
TrpC3
|
Baadio Idrissa, Baadio Issouf
|
Mixte : 144 bovins, 26 ovins
|
Gurmaji (35 têtes de Jaliji)
|
Baadio Idrissa (42 ans), Sondé Hama (28 ans), Lido Issoufi
(24 ans)
|

190
TrpC1-2G, troupeau d'éleveurs gourmantchés
sédentaires non transhumants; TrpC1-2P, troupeau d'éleveurs peuls
sédentaires non transhumants) ;
Trp, troupeau d'éleveurs peuls résidents grands
transhumants) ; TrpC3, troupeau d'éleveurs peuls allochtones grands
transhumants).
6.3.3 .2. Comportement des éleveurs et de leurs
troupeaux au pâturage
La gestion des ressources pastorales par les éleveurs
et leurs troupeaux se fait à deux échelles : l'échelle
régionale qui fait appel à diverses stratégies de
mobilité dont la transhumance et l'échelle locale dans les
limites, le plus souvent, des terroirs (Dongmo, 2009). Dans le chapitre
précédent, nous avons rappelé que la conduite quotidienne
différait entre les éleveurs. Les Peuls pratiquent à la
fois la pâture nocturne et diurne et, en saison sèche,
l'abreuvement se fait en deux temps, comme l'ont aussi observé D'Amico
et al. (1995) chez les éleveurs Mbororo de Centrafrique. Chez
les Gourmantchés, par contre, il n'existe pas de pâture nocturne
et l'abreuvement est unique en toute période pour le troupeau principal.
Par ailleurs, les pratiques des éleveurs évoluent au cours de
l'année, cette dynamique étant mue par les conditions
liées aux ressources (disponibilité, qualité,
accessibilité) et par les choix qui peuvent être
opérés par les troupeaux et/ou par les bergers qui les
accompagnent.
Dans les parties qui suivent nous rendons compte du
comportement des quatre troupeaux suivis au cours de l'année selon les
saisons des éleveurs (voir exemple pour tous les troupeaux pour la
saison sèche froide : carte VI-1). Notons que nous nous
référons aux unités paysagères écologiques
pour une plus grande lisibilité, leurs limites étant plus
nettement perceptibles.
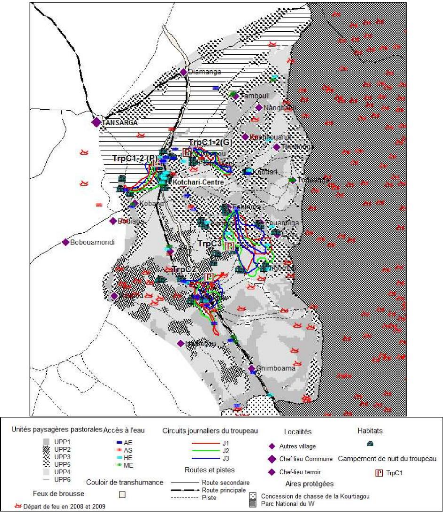

Carte VI-1. Itinéraires des quatre troupeaux en saison
sèche froide (pendant Ku fowagu ou Dabunde)
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
AE : point d'eau à usage exclusivement animal et contenant
de l'eau;

192
AS : point d'eau à usage exclusivement animal et à
sec ;
HE : point d'eau à usage exclusivement humain et contenant
de l'eau ; ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau.
6.3.3.2.1. Variations saisonnières de
l'utilisation des pâturages chez le troupeau TrpC1-2G (troupeau
gourmantché de la catégorie C1-2)
6.3.3.2.1.1. Distances et durées moyennes de
déplacement du troupeau
Le troupeau de l'éleveur Soali Combary commence sa
journée de pâture entre 8 h 13 mn et 9 h 03 mn et la termine entre
17 h 06 mn et 18 h 11 mn. Durant la période de suivi, ce troupeau a eu
un temps de présence au parcours qui a varié entre 8 h 07 mn
(D_Kufowagu : fin de saison humide) et 9 h 52 mn (Ku tontogu
: saison sèche chaude) (tableau VI-9). Par ailleurs, les distances
totales parcourues ont été également plus importantes en
saison sèche notamment pendant A sakoana (12,19 km) et
très peu importantes en saison des pluies ou Ku siagu (6,49
km). L'essentiel des trajets est consacré au déplacement sans
alimentation, surtout en saison sèche chaude (plus de 3 km au Ku
tontogu et A sakoana) alors qu'en saison humide stricte (Ku
siagu), seulement 380 m en moyenne sont parcourus sans broutage.
Notons que les animaux sortent plus tardivement du campement
de nuit en saison humide et ceci à cause de la traite matinale du lait
alors relativement plus abondant que pendant les autres saisons, mais aussi
pour éviter l'humidité matinale due à la rosée.
Tableau VI-9. Temps et distance au pâturage pour le
troupeau TrpC1-2G
|
Saisons
|
Heure départ
|
Heure retour
|
Durée pâturage
|
Distance parcourue (km)
|
|
Totale
|
Marche
|
Autres activités
|
|
P_Ku fowagu
|
8 h 35 #177; 21 mn
|
17 h 06 #177; 09 mn
|
8 h 31 #177; 16 mnb
|
7,63 #177; 0,41b
|
1,66 #177; 0,14c
|
5,97 #177; 0,29a
|
|
Ku tontogu
|
8 h 13 #177; 11 mn
|
18 h 05 #177; 16 mn
|
9 h 52 #177; 13 mnc
|
9,94 #177; 1,03c
|
3,51 #177; 0,59d
|
6,43 #177; 1,35b
|
|
A sakoana
|
8 h 39 #177; 17 mn
|
18 h 11 #177; 15 mn
|
9 h 30 #177; 31 mnc
|
12,19 #177; 1,02d
|
3,23 #177; 0,83d
|
8,96 #177; 0,19c
|
|
Ku siagu
|
9 h 01 #177; 04 mn
|
17 h 32 #177; 23 mn
|
8 h 30 #177; 27 mnab
|
6,49 #177; 0,40a
|
0,38 #177; 0,10a
|
6,11 #177; 0,31a
|
|
D_Ku fowagu
|
9 h 03 #177; 12 mn
|
17 h 08 #177; 04 mn
|
8 h 07 #177; 15 mna
|
7,13 #177; 0,20a
|
1,00 #177; 0,12b
|
6,13 #177; 0,08a
|
Les valeurs situées sur la même colonne et
portant des lettres distinctes sont significativement différentes au
seuil á = 0,05 à p = 0,001.
D_Ku fowagu (début Ku fowagu):
mi-octobre à début-novembre P_Ku fowagu (plein Ku
fowagu): novembre à février
Ku tontogu : mars à début mai
A sakoana : fin mai à début juin
Ku siagu : juin à début octobre
6.3.3.2.1.2. Importance relative des activités au
pâturage
Ce troupeau passe au moins 60% de son temps à brouter,
le reste du temps étant consacré surtout à se
déplacer, mais aussi au repos-rumination et à l'abreuvement ;
cette dernière activité apparaît cependant comme marginale
surtout en saison pluvieuse (Ku siagu) ou post-pluvieuse (début
Ku fowagu) (figure VI-3). Dans ce troupeau, le broutage occupe
davantage de temps en saison pluvieuse ou en saison sèche froide (Ku
siagu et Ku fowagu : 80%) qu'en saison sèche chaude
(Ku tontogu et A sakoana : 65%). Le repos-rumination, le

déplacement et l'abreuvement sont par contre trois
activités auxquelles plus de temps est consacré en saison
sèche chaude que pendant les autres saisons du calendrier pastoral.
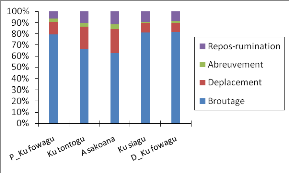
Figure VI-3 Proportions en temps consacré aux
activités au cours de l'année par le troupeau TrpC1-2G
D_Ku fowagu (début Ku fowagu):
mi-octobre à début-novembre P_Ku fowagu (plein Ku
fowagu): novembre à février
Ku tontogu : mars à début mai
A sakoana : fin mai à début juin
Ku siagu : juin à début octobre
6.3.3.2.1.3. Le circuit pastoral quotidien au fil des
saisons : unités pâturées et durée de
fréquentation
· Pendant le plein Ku fowagu (novembre
à février)
Pendant le premier suivi (17 - 19 novembre 2008) correspondant
à la saison sèche froide (plein Ku fowagu), les
récoltes sont très avancées et il ne subsiste que quelques
champs de sorgho à cycle long dans les bas-fonds (UPP1) et les
bas-glacis et plaines argileuses (UPP3). La vaine pâture, encore timide a
commencé dans certains secteurs du terroir. Les trois itinéraires
adoptés par le jeune berger Ombua et son troupeau, pendant les trois
jours d'observation, sont quasi-identiques (carte VI-2). Ainsi, en partant
tôt du parc de nuit, le troupeau entame sa journée en parcourant
brièvement les buttes rocheuses basses de la chaîne (Li
guali ou UPP6) et se retrouve rapidement en bas. Une fois à ce
niveau, il fréquente successivement les parties récoltées
et en jachère des plaines et bas-glacis. Les prises alimentaires se
composent essentiellement de résidus de mil et de quelques restes de
fanes d'arachide et de nombreux adventices comme Eragrostis
sp. et des herbes de jeunes
jachères : Digitaria sp., Pennisetum Pedicellatum,
toutes déjà à l'état de paille. Les animaux y
restent pendant une bonne partie de la matinée tout en progressant en
direction des bas-fonds (UPP1) et surtout des points d'eau situés dans
le village de Kobdari où l'abreuvement a lieu à 13 h 01mn. Par
endroits, le passage est délicat à cause des champs non encore
récoltés et le troupeau se faufile dans les interstices entre
champs ou par les sentiers empruntés par les agriculteurs. Dans ce
village, il existe encore de l'eau de surface dans certaines dépressions
au sein des bas-fonds et l'abreuvement s'y passe pendant 15 mn. Après
l'abreuvement, survient le repos de l'ensemble du troupeau. Cette phase de
repos-rumination dure 32,5 mn avant que

194
le troupeau n'entame son chemin de retour en parcourant
à peu près les mêmes unités en particulier
l'unité de type UPP3. Le soir, avant d'entrer dans le parc de nuit, le
troupeau passe un bref moment sur les plateaux environnants (UPP4) sur lesquels
la couverture herbacée est dominée par l'espèce
Schoenefeldia gracilis à l'état de paille.
L'unité UPP3 apparait donc comme la plus
intéressante dans ce secteur en cette période. En effet, le
troupeau y passe le plus clair de son temps (figure VI-4), et y montre la plus
faible vitesse de déplacement (6,13 m/mn) (tableau VI-10). Sur cette
unité, les résidus de culture sont encore abondants et surtout
relativement frais et donc de bonne qualité alimentaire. Il est à
noter que les bas-fonds abritent encore beaucoup de champs tardifs et les
animaux ne s'y attardent donc pas lorsqu'ils sont amenés à y
pâturer.
On voit par ailleurs que, le feu de brousse qui a eu lieu une
semaine avant nos observations, n'a pas occasionné de repousses notables
susceptibles d'attirer le bétail. Ce secteur est en effet couvert
essentiellement d'herbacées annuelles de jeunes jachères
(Schoenefeldia gracilis, Brachiaria sp., Digitaria sp., Pennisetum
pedicellatum, etc.) et, au plan pastoral, l'action du feu est plutôt
dommageable, il ne provoque pas de repousses fraiches comme on pourrait
s'attendre avec des graminées vivaces.
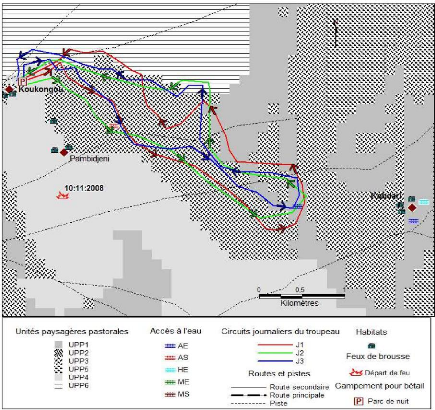
Carte VI-2. Itinéraires du troupeau TrpC1-2G en plein
Ku fowagu
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
10 :11 :2008 : date de départ du feu
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; AS : point d'eau à usage exclusivement animal et à sec
;
HE : point d'eau à usage exclusivement humain et contenant
de l'eau ; ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau. MS : point d'eau à usage mixte (humain et animal) et
à sec.
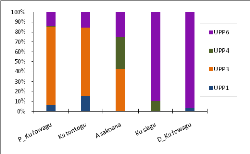
Figure VI-4. Proportions en temps de séjour dans les
unités pastorales au cours de l'année pour le troupeau
TrpC1-2G.
D_Ku fowagu (début Ku fowagu):
mi-octobre à début-novembre P_Ku fowagu (plein Ku
fowagu): novembre à février
Ku tontogu : mars à début mai
A sakoana : fin mai à début juin
Ku siagu : juin à début octobre
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes rocheuses
et cuirassées.
Tableau VI-10.Vitesse moyenne de déplacement du troupeau
TrpC1-2G suivant les unités et les saisons

Saisons
|
Vitesse (m/mn) par unité
paysagère
|
|
UPP1
|
UPP3
|
UPP4
|
UPP6
|
|
P_Ku fowagu (novembre à février)
|
7,01
|
6,13
|
9
|
11,18
|
|
Ku tontogu (mars à début mai)
|
9,85
|
13,02
|
----
|
13,81
|
|
A sakoana (fin mai à début juin)
|
----
|
16,02
|
18,75
|
18,29
|
|
Ku siagu (juin à début octobre)
|
----
|
6,44
|
7,92
|
9,81
|

196
D_Ku fowagu (mi-octobre à début-novembre)
6,19 ---- ---- 10,61
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes rocheuses
et cuirassées.
· Pendant le Ku tontogu (mars à
début mai)
En pleine saison sèche chaude (Ku tontogu), le
troupeau, partant du parc de nuit, se dirige dans la direction opposée
à celle qu'il fréquentait lors du précédent suivi.
Les circuits (carte VI-3) sont là encore quasiment identiques sur les
trois journées: ils se concentrent dans les plaines, bas-fonds et
bas-glacis désormais totalement débarrassés de toutes les
cultures. Dans cette partie du terroir, les troupeaux que nous avons
rencontrés au cours de ces journées étaient
essentiellement ceux de Gourmantchés habitant à proximité.
Chaque matin, durant les trois jours, le troupeau de Combary Soali s'est rendu
sur les bas-glacis et plaines argileuses (UPP3) à l'ouest du quartier
Koukongou où il était encore possible de trouver des
résidus de culture. Il y est resté toute la journée en les
parcourant de manière rapide (13,02 m/mn) dans tous les sens surtout le
long des bas-fonds (UPP1) qui ont été aussi, par moments, mis
à contribution. Les bas-fonds étaient les milieux les plus
intéressants comme en témoigne le temps qu'y a passé le
troupeau (figure VI-4) et le rythme adopté (9,85 m/mn) (tableau VI-10).
Cependant, la concurrence y était rude car tous les animaux basés
dans les quartiers environnants venaient s'y abreuver à cause de
nombreux filets d'eau intarissables alimentés par diverses sources qui y
sont rencontrées. Après l'abreuvement (durée moyenne: 36
mn) qui se produit un peu plus tôt que précédemment
(à 12h 48mn) et un repos de 26 mn, le troupeau est repassé
pratiquement sur ses pas, tout en restant à proximité des
habitations. Deux fois sur les trois jours de suivi, quelques instants
après le repos, les animaux ont été conduits vers le
campement de nuit où ils ont été
complémentés avec du fourrage gardé en stock
accompagné d'une boisson faite d'eau mélangée au son de
cuisine. Le retour vers le campement a eu lieu à 18h 05mn et le troupeau
y est parvenu en passant rapidement (vitesse : 13,81 m/mn) par le flanc de la
chaîne du Gobnangou (UPP6). Le premier jour de suivi, cependant, le
troupeau est rentré directement au campement en parcourant les plaines
et les plateaux (UPP4). La traversée des plateaux s'est faite à
une allure rapide sans broutage, ce qui indique son peu d'intérêt.
On peut remarquer que tout en restant non loin du parc de nuit, la distance
journalière parcourue est nettement plus importante que la saison
précédente (9,94 km contre 7,63 km).
Les feux de brousse qui étaient passés en
novembre et décembre précédents avaient notablement
réduit le disponible fourrager et l'espace exploitable par le troupeau.
En effet, le feu, parti du quartier Pambidjeni (Sud-est du secteur : carte
VI-3), a consumé une grande partie de l'importante biomasse
herbacée que produit habituellement l'unité sur laquelle il s'est
déclaré (UPP4). Il a ainsi privé le troupeau d'un fourrage
abondant et situé à proximité, qui aurait pu être
exploité en cette saison où l'économie d'énergie
est un défi important (Kagoné, 2000 ; Dumont et al.
2001).
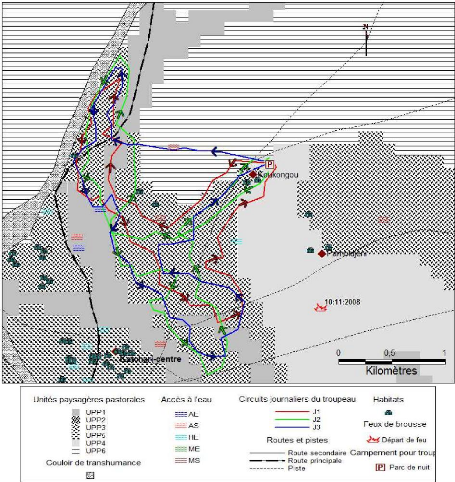

Carte VI-3. Itinéraires du troupeau TrpC1-2G pendant
Ku tontogu
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
10 :11 :2008 : date de départ du feu
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; AS : point d'eau à usage exclusivement animal et à sec
;
HE : point d'eau à usage exclusivement humain et contenant
de l'eau ; ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau. MS : point d'eau à usage mixte (humain et animal) et
à sec.

198
· Pendant A sakoana (fin mai à
début juin)
Le comportement territorial du troupeau en cette saison de
transition est assez proche de celui de la période
précédente, mais le suivi des 26, 27 et 28 mai 2009 (carte VI-4)
a montré que l'espace exploré était plus vaste et que les
animaux se déplaçaient dans tous les sens en empruntant des
itinéraires sinueux, la distance journalière moyenne parcourue
(12,19 km) étant plus longue. Les unités paysagères
parcourues restent pratiquement les mêmes que pendant Ku tontogu
avec une préférence plus affirmée pour les plateaux (UPP4)
situés à proximité du campement de nuit, maintenant
délocalisé à côté de la concession familiale.
Les animaux ont été abreuvés à 12h 44 mn pendant 26
mn en un point d'eau situé dans le même secteur et le repos qui a
suivi a été de même durée (24 mn). Sur les trois
jours qu'ont duré les observations, le troupeau est resté sur les
plaines et bas glacis pour l'essentiel de son temps de broutage, marqué
par de courts et incessants déplacements ; les prises se sont
composées de restes de résidus culturaux et de pailles
résiduelles. De manière globale, le rythme de déplacement
a été rapide (de 16,02 m/mn sur UPP3 à 18,75 m/mn sur
UPP4) (tableau VI-10), ce qui est une indication que ces parcours sont de
très faible qualité. Ici encore, l'après midi les animaux
ont reçu en complément des résidus de culture
collectés et stockés au sein de la concession de Mr Soali. A leur
retour au campement, ils ont trouvé une boisson à base de son de
cuisine et de son industriel dans de grandes bassines. Cette
complémentation et la proximité de sources d'eau permanentes dans
le bas-fond situé à l'ouest du quartier Koukongou expliquent que
le troupeau, en cette saison, ne s'éloigne pas des habitations. Notons
par ailleurs que quelques pluies ont été signalées dans la
partie sud du terroir (villages de Pielgou et de Gnimboama), mais elles
n'avaient pas eu le temps de permettre des repousses notables qui auraient pu
influencer le comportement des troupeaux.
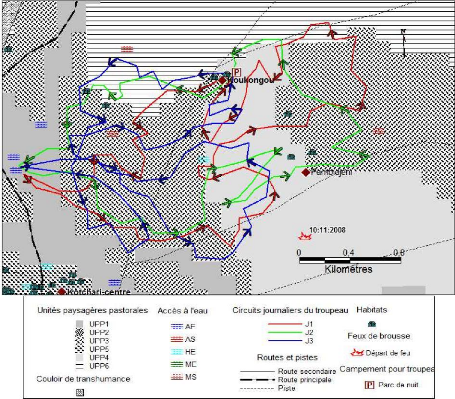

Carte VI-4. Itinéraires du troupeau TrpC1-2G pendant A
sakoana
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
10 :11 :2008 : date de départ du feu
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; AS : point d'eau à usage exclusivement animal et à sec
;
HE : point d'eau à usage exclusivement humain et contenant
de l'eau ; ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau ; MS : point d'eau à usage mixte (humain et animal)
et à sec.

200
· Pendant Ku siagu (juin à
début octobre)
La troisième journée de ce suivi fait du 3 au 5
août 2009 a été marquée par une pluie matinale. Les
circuits (carte VI-5), plus linéaires et moins longs (distance moyenne :
6,49 km) se sont concentrés sur les buttes rocheuses de la chaîne
du Gobnangou (UPP6) où les prises alimentaires se sont essentiellement
composées de Andropogon pseudapricus, Loudetia togoensis, et
parfois Schizachyrium exile en pleine croissance. Chaque matin, le
troupeau a été ramené au flanc de la chaîne et trait
par les femmes. Puis, vers 9 h, il a pris la direction du sommet de la
chaîne et y a passé pratiquement toute la journée en
progressant lentement (9,81 m/mn). L'abreuvement à 13h 12 mn dans
diverses petites retenues disséminées dans une petite
vallée située sur la chaîne a été bref (4
mn). Ensuite a suivi un long repos-rumination qui a duré 47mn, puis le
troupeau est resté encore un long moment sur les buttes avant d'amorcer
son retour vers le campement autour de 16 h. La dernière étape du
parcours, une descente vers les bas-glacis et plaines argileuses (UPP3) et
plateaux (UPP4) situés en contrebas, a permis au troupeau d'exploiter
les interstices non cultivés entre les champs, espaces riches en
certaines herbes de qualité (Eragrostis sp., Brachiaria sp.,
Pennisetum pedicellatum, Setaria pallide-fusca, Digitaria sp.) et
très appréciées par le troupeau comme en témoigne
le rythme particulièrement lent de progression (respectivement 6,44 m/mn
et 7,92 m/mn) (tableau VI-10). Cette partie du terroir étant une zone de
concentration de champs, le berger a évité d'aller plus loin avec
son troupeau.
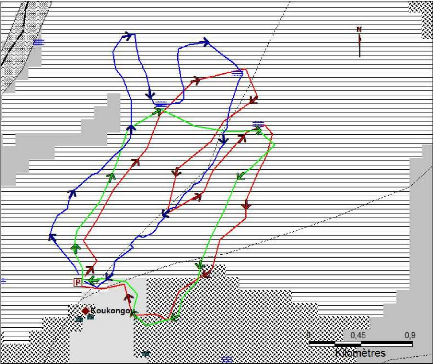

Carte VI-5. Itinéraires du troupeau TrpC1-2G pendant
Ku siagu
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; HE : point d'eau à usage exclusivement humain et contenant de
l'eau ;
ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau.
· Pendant début Ku fowagu
(octobre à début novembre)
Lors du dernier suivi (15 au 17 octobre 2009) au début
de la période Ku fowagu, seules quelques parcelles de
légumineuses (niébé, arachide) avaient déjà
été récoltées, l'essentiel des champs de
céréales attendaient d'être récoltés. Le
troupeau, après la traite matinale, a été conduit par le
bouvier sur la chaîne du Gobnangou (UPP6) en bordure de laquelle est
situé le parc de nuit (carte VI-6 & figure VI-4). L'herbe y
était encore abondante mais commençait à perdre
sensiblement en qualité car la plupart des espèces
(Andropogon pseudapricus, Schizachyrium exile, Loudetia togoensis)
avait atteint leur stade de floraison et même de fructification. Sur la
chaîne, les animaux recherchaient les dépressions où il
était encore possible de trouver des herbes tendres ou non encore
matures. L'abreuvement qui a eu lieu vers 13h pendant les 3 jours de suivi,
dans une mare de bas-fond (Ku bagu ou UPP1), a duré
environ 7 mn. Immédiatement après, il y a eu un temps de
repos-rumination pendant 41,5 mn. Vers 14 h, la recherche alimentaire a repris,
toujours sur la chaîne, mais en direction du campement que le troupeau
atteint vers 17 h (tableau VI-9). Il est à noter que juste avant et
après l'abreuvement le troupeau s'est retrouvé aux abords des
bas-fonds (UPP1), milieux appréciés (vitesse : 6,19 m/mn) mais
difficilement praticables. Les animaux se sont ensuite satisfaits dans les
parties sèches de la chaîne où ils ont cependant
progressé à un rythme plus rapide (10,61 m/mn) (tableau
VI-10).
Notons que pendant cette saison et la précédente,
le temps consacré par le troupeau au déplacement sans broutage
est plus faible que pendant les autres (figure VI-3)
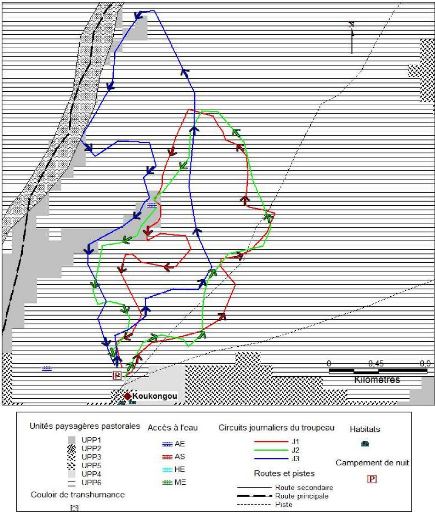

202
Carte VI-6. Itinéraires du troupeau TrpC1-2G pendant
début Ku fowagu
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; AS : point d'eau à usage exclusivement animal et à sec
;
HE : point d'eau à usage exclusivement humain et contenant
de l'eau ;

ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau.
6.3.3.2.2. Variations saisonnières de
l'utilisation des pâturages chez le troupeau de type TrpC1-2P (troupeau
peul de la catégorie C1-2)
6.3.3.2.2.1. Distances et durées moyennes du
déplacement du troupeau
Le troupeau commence sa journée de pâturage entre
8h 45mn (au Ceedu) et 9h 16 mn (au Yaamde) et la termine au
plus tôt à 18h 05mn ou plus tardivement à18h 16mn en pleine
saison humide (Ndungu). Au total, le troupeau a séjourné
au pâturage environ 9 heures à chaque saison. Les distances
moyennes parcourues ont été de 8,20 km (Yaamde) à
10,77 km (Ceedu), (tableau VI-11) et une bonne part de ces distances
reviennent aux déplacements entre plusieurs secteurs de la zone
explorée. Ces trajets ont été particulièrement
longs pendant la saison sèche (3,32 km au Ceedu et 2,66 au
Kotoga).
Tableau VI-11.Temps et distance au pâturage pour le
troupeau TrpC1-2PTrpC1-2P.
|
Saisons
|
Heure départ
|
Heure retour
|
Durée pâturage
|
Distance parcourue (km)
|
|
Totale
|
Marche
|
Autres activités
|
|
Dabunde
|
8 h 52 #177; 22 mn
|
18 h 08 #177; 09 mn
|
9 h 17 #177; 25 mn
|
8,84 #177; 0,37a
|
2,05 #177; 0,15a
|
6,79 #177; 0,25 b
|
|
Ceedu
|
8 h 45 #177; 13 mn
|
18 h 05 #177; 09 mn
|
9 h 19 #177; 19 mn
|
10,77 #177; 0,76b
|
3,32 #177; 0,45c
|
7,46 #177; 0,34c
|
|
Kotoga
|
8 h 58 #177; 16 mn
|
18 h 07 #177; 05 mn
|
9 h 10 #177; 18 mn
|
10,36 #177; 0,39b
|
2,66 #177; 0,10b
|
7,70 #177; 0,31c
|
|
Ndungu
|
9 h 12 #177; 13 mn
|
18 h 16 #177; 14 mn
|
9 h 04 #177; 23 mn
|
8,35 #177; 1,02a
|
1,80 #177; 0,38 a
|
6,54 #177; 1,33 b
|
|
Yaamde
|
9 h 16 #177; 17 mn
|
18 h 13 #177; 08 mn
|
8 h 57 #177; 21 mn
|
8,20 #177; 0,71a
|
2,28 #177; 0,25 ab
|
5,92 #177; 074 a
|
Les valeurs situées sur la même colonne et
portant des lettres distinctes sont significativement différentes au
seuil á = 0,05 à p = 0,001.
Dabunde : mi-novembre à février Ceedu
: mars à mai
Kotoga : fin mai à début juin
Ndungu : juin à début octobre
Yaamde : octobre à début novembre
6.3.3.2.2.2. Importance relative des activités au
pâturage
Le partage du temps du troupeau au pâturage (figure
VI-5) est très proche de celui du troupeau que nous venons de
présenter. Ainsi, le prélèvement alimentaire (broutage)
est l'activité la plus importante qui occupe en moyenne autour de 70% du
temps du bétail toutes saisons confondues. C'est pendant la saison
pluvieuse que le plus de temps (73,5%) lui est consacré avec le
repos-rumination tandis que le temps mis pour se déplacer ou pour
s'abreuver est le plus faible. Le repos-rumination est une activité
particulièrement peu importante en saison sèche chaude (Ceedu
+ Kotoga) : seulement 7% du temps lui est consacré contre 11%
à 16% pour les autres saisons ; en revanche, l'abreuvement y occupe la
place prépondérante (5 à 7%). C'est pendant la saison
sèche, et tout particulièrement de mars à mai (Ceedu
: 20%), que le déplacement pèse le plus dans le budget-temps
du troupeau.
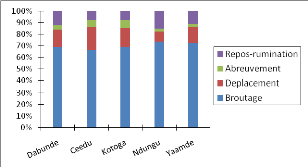

204
Figure VI-5. Proportions en temps consacrés aux
activités au cours de l'année par le troupeau TrpC1-2P.
Dabunde : mi-novembre à février Ceedu
: mars à mai
Kotoga : fin mai à début juin
Ndungu : juin à début octobre
Yaamde : octobre à début novembre
6.3.3.2.2.3. Le circuit pastoral quotidien selon les
saisons : unités pâturées et durée de
fréquentation
· Pendant la saison du Dabunde
(mi-novembre à février)
Lors du suivi du 22 au 24 novembre 2008, une grande partie des
parcelles cultivées de cette partie du terroir (flanc de la chaîne
du Gobnangou) étaient déjà récoltées mais la
vaine pâture n'était encore admise que dans certains de ces
secteurs car les résidus de culture n'avaient pas encore
été totalement collectés. Le troupeau, après la
traite matinale, s'est d'abord brièvement abreuvé dans les mares
voisines, puis s'est dirigé vers la chaîne (UPP6) au pied de
laquelle se trouve son parc de nuit (carte VI-7). Évitant
l'humidité de la rosée matinale, il a passé un long moment
à parcourir à allure rapide (13 m/mn ; tableau VI-12) les buttes
rocheuses en se dirigeant vers le bas-fond (UPP1) de la partie sud du secteur
(vers le quartier Kobana) où a eu lieu l'abreuvement de l'après
midi. Avant cet abreuvement (début à 13h 52mn; durée :
20,5 mn) et le repos-rumination de 69 mn qui a suivi, le troupeau a
exploité divers fourrages. Il s'agissait de l'herbe (Pennisetum
pedicellatum, Schoenefeldia gracilis, Oryza longistaminata) pourtant
déjà sèche des bas-fonds, bas-glacis et plaines argileuses
(UPP3), mais surtout de divers résidus de culture (tiges de sorgho,
fanes de niébé) et de jachères entre les cultures,
situées surtout sur les plaines sablo-limoneuses à sols
moyennement profonds (UPP5). Vers 16 heures, le troupeau a exploré
rapidement (12,02 m/mn) les plateaux gravillonnaires situés dans l'ouest
du secteur avant d'amorcer son retour vers le parc de nuit. De toutes les
unités pâturées, les unités de bas-fonds et de
bas-glacis sont apparues les plus attractives si l'on se fie au rythme de
progression qui y a été observé (respectivement 7,93 m/mn
et 9,44 m/mn). Pendant l'observation sur place c'était les zones
cultivées (UPP5) qui semblaient les plus recherchées, mais le
cloisonnement de l'espace par les champs et le risque de dégâts
associé obligeait le troupeau à y progresser rapidement (10
m/mn). C'est effectivement là que le plus de temps a été
passé (figure VI-6).
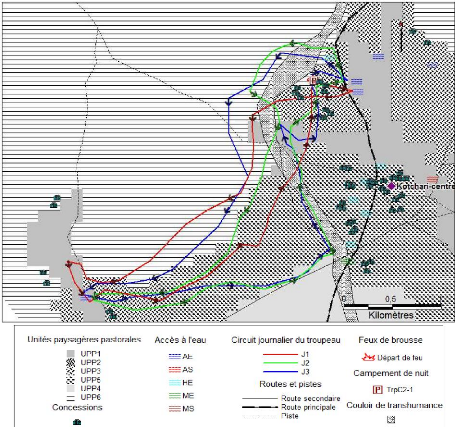

Carte VI-7. Itinéraires du troupeau TrpC1-2P pendant
Dabunde
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; AS : point d'eau à usage exclusivement animal et à sec
;
HE : point d'eau à usage exclusivement humain et contenant
de l'eau ; ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau ; MS : point d'eau à usage mixte (humain et animal)
et à sec.
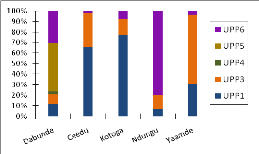

206
Figure VI-6. Proportions en temps de séjour dans les
unités pastorales au cours de l'année pour le troupeau
TrpC1-2P.
Dabunde : mi-novembre à février Ceedu
: mars à mai
Kotoga : fin mai à début juin
Ndungu : juin à début octobre
Yaamde : octobre à début novembre
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
Tableau VI-12. Vitesse moyenne de déplacement du troupeau
TrpC1-2P suivant les unités et les saisons
|
Saisons
|
|
Vitesse (m/mn) par unité
paysagère
|
|
UPP1
|
UPP3
|
UPP4
|
UPP5
|
UPP6
|
|
Dabunde (mi-novembre à
février)
|
7,93
|
9,44
|
12,02
|
10
|
13
|
|
Ceedu (mars à mai)
|
10,54
|
13
|
----
|
----
|
15,77
|
|
Kotoga (fin mai à début juin)
|
9,67
|
11,15
|
----
|
----
|
16,06
|
|
Ndungu (juin à début octobre)
|
6,02
|
7,55
|
----
|
----
|
6,89
|
|
Yaamde (octobre à début
novembre)
|
6,78
|
8
|
----
|
----
|
13,73
|
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
· Pendant la saison du Ceedu (mars
à mai)
Quelques jours après le suivi de la saison
précédente (Dabunde), un feu de brousse daté du
05 décembre 2008 (carte VI-8) s'est produit dans le secteur de la
chaîne qui fait face au parc de nuit du troupeau. Ceci n'a cependant pas
semblé influer notablement sur les itinéraires du troupeau,
l'espace villageois en bas de la chaîne étant totalement
libéré après les

récoltes et le ramassage des résidus de culture.
Au moment de notre suivi (11-13 avril 2009), la vaine pâture avait
déjà eu lieu et les zones cultivées (essentiellement UPP5
mais aussi UPP3) ont été fortement pâturées par le
cheptel local et transhumant. Durant les trois jours de suivi, le berger a
conduit son troupeau, dans une portion très restreinte du terroir
principalement constituée de plaines et bas-fonds (UPP3 et UPP1) (figure
VI-6 & carte VI-8) le long du couloir de transhumance et de la route
principale qui traverse le terroir dans le sens Nord-Sud. Chaque matin, le
troupeau passait d'abord s'abreuver dans les filets d'eau du bas-fond
situé à proximité. De là il se dirigeait vers le
nord et n'en revenait que pour le second abreuvement vers 14h (33 mn).
Après un repos de 43 mn en moyenne succédant à
l'abreuvement, le troupeau est resté sur les mêmes unités,
assez proches du parc de nuit pour attendre le moment du retour. Sur ces
unités, la progression était rapide (10,54 m/mn à 15,77
m/mn : tableau VI-12), ce qui montre qu'elles sont pauvres et la forme
très sinueuse des circuits montre des changements incessants de
direction dans la progression du troupeau et reflète la distribution
très disparate de l'offre fourragère (Dumont et al.
2001). La distance parcourue en cette saison est en moyenne
inférieure à celle de la saison précédente
(Dabunde) (10,36 km contre 10,77 km). Notons qu'en cette saison le
parc de nuit a été délocalisé juste à
côté de la concession du propriétaire.
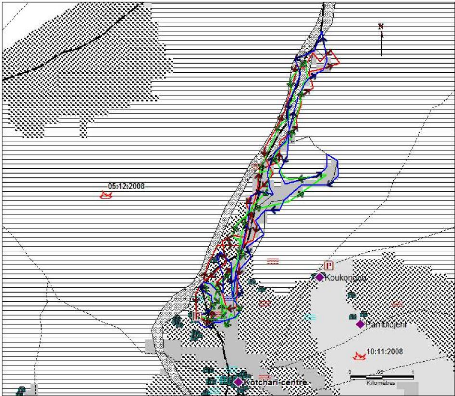


208
Carte VI-8. Itinéraires du troupeau TrpC1-2P pendant
Ceedu
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
10 :11 :2008 : date de départ du feu
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; AS : point d'eau à usage exclusivement animal et à sec
;
HE : point d'eau à usage exclusivement humain et contenant
de l'eau ; ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau ; MS : point d'eau à usage mixte (humain et animal)
et à sec.
· Pendant la saison du Kotoga (fin mai
à début juin)
Ce suivi a eu lieu un mois et demi plus tard que le
précédent, précisément les 29, 30 et 31 mai 2009,
mais le comportement du troupeau a peu changé entre les deux
périodes. Le même secteur et les mêmes unités
pastorales ont été pâturées (carte VI-9). On peut
noter cependant que la fréquentation des bas-fonds (UPP1) s'est accrue
aux dépens de celle des basglacis et plaines argileuses (UPP3). En cette
période pré humide, les quelques pluies qui ont été
enregistrées n'ont pas atteint un niveau à même de
provoquer la pousse des herbes précoces. Le fourrage herbacé des
plaines est alors totalement épuisé ou sans intérêt
alors que la concentration des points d'eau dans les bas-fonds justifie la plus
grande présence du troupeau sur ces unités qui avaient, par
ailleurs, une plus grande couverture en paille. Par ailleurs, le troupeau a
été, cette fois, plus présent sur les buttes rocheuses
(UPP6), mais cette unité a été traversée rapidement
(16,06 m/mn ; tableau VI-12) avec peu ou pas de broutage, car elle manque
d'intérêt fourrager à cette période. L'abreuvement
du troupeau s'est déroulé en deux temps (1er
abreuvement à 9h 12mn ; 2ème abreuvement à 13h
42 mn pendant 40 mn) et le repos a duré 43,5 mn.
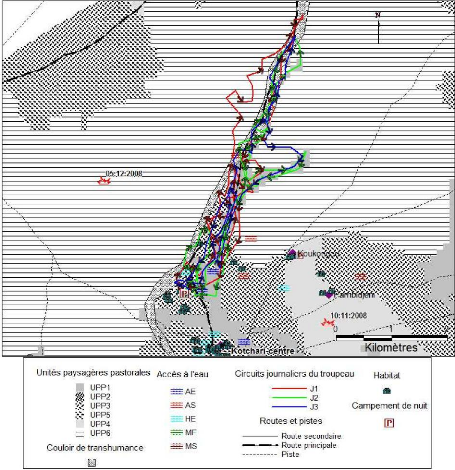

Carte VI-9. Itinéraires du troupeau TrpC1-2P pendant
Kotoga
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
10 :11 :2008 : date de départ du feu
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; AS : point d'eau à usage exclusivement animal et à sec
;
HE : point d'eau à usage exclusivement humain et contenant
de l'eau ; ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau ; MS : point d'eau à usage mixte (humain et animal)
et à sec.
·

210
Pendant la saison du Ndungu (juin à
début octobre)
Le suivi a lieu les 6, 7 et 8 août au coeur de la
saison des pluies, à cette période, le parc de nuit du troupeau
avait été ramené à son emplacement de début
Dabunde, c'est-à-dire au flanc des buttes rocheuses de la
chaîne du Gobnangou. Il s'agissait de fuir l'humidité et de
libérer l'espace pour les cultures de case (carte VI-10). Il n'y a pas
eu d'abreuvement matinal et dès la fin de la traite laitière
matinale, le troupeau a parcouru les pâturages de la chaîne (UPP6),
assez fournis et composé essentiellement de Andropogon
pseudapricus Schizachyrium exile et, par endroits, de
Loudetia togoensis. Il y est resté pendant une grande partie de
la journée avant de descendre pour s'abreuver à 12h 46 mn pendant
11,5 mn dans le lit des bas-fonds qui parsèment le couloir de
transhumance, puis profiter d'un long reposrumination (1h 24 mn). Le reste de
la journée, le troupeau a été conduit par le berger entre
les champs des plaines argileuses (UPP3) et dans les parties praticables des
bas-fonds (UPP1) du secteur. Depuis là le retour a été
amorcé vers 16 h pour arriver au parc de nuit à 18h 16mn (tableau
VI-11). L'essentiel du temps de pâturage a donc été
consacré à exploiter les pâturages des hauteurs (UPP6)
(figure VI-6) et le rythme lent du troupeau (6,89 m/mn) (tableau VI-12) montre
qu'il trouvait à s'y satisfaire. Cependant, pour diminuer les
dépenses en énergie de leur troupeau, le berger a pris le risque
de les maintenir, le soir, dans les secteurs cultivés et dans les
bas-fonds, sauf le premier jour où une pluie s'est abattue dans le
village entre 10 et 11 heures. En cette saison, la distance moyenne parcourue a
été la plus faible de l'année (8,35 km ; tableau
VI-11).
· Pendant la saison du Yaamde (octobre
à début novembre)
Pendant la saison de transition vers la saison sèche
froide (Yaamde) (carte VI-11), le secteur fréquenté a
été le même que pendant Ndungu. Cependant, le
troupeau a été plus présent dans les terres basses :
abords asséchés de bas-fonds (UPP1) et espaces entre champs
où les récoltes ont timidement commencé. Le pâturage
sur la chaîne devient exceptionnel, il ne se fait plus qu'aux
premières heures de la matinée ou pour faire passer le troupeau
d'un secteur à un autre. Les pâturages les plus recherchés
sont alors ceux des bas glacis (près de 70% de temps de présence
; figure VI-6) notamment dans leurs portions cultivées où les
résidus de culture commencent à être disponibles. Les
risques liés aux dégâts champêtres y diminuent en
effet au rythme de l'avancée des récoltes et le troupeau les
parcourt très lentement (8 m/mn) (tableau VI-12) preuve de leur
intérêt. En cette saison, tout le lit du bas fond regorge encore
d'eau de surface à plusieurs endroits et le troupeau s'y abreuve
directement vers de 13h après un premier abreuvement matinal à
proximité du parc de nuit.
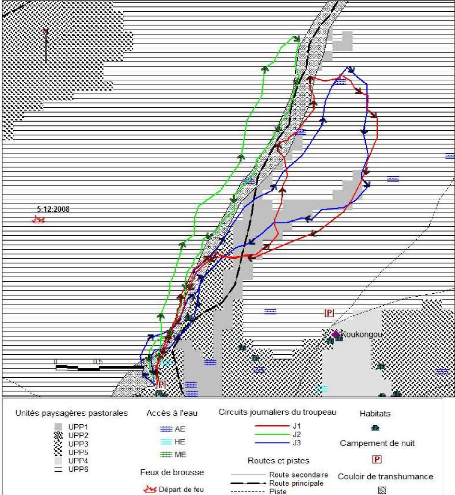

Carte VI-10. Itinéraires du troupeau TrpC1-2P pendant
Ndungu
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
05 :12 :2008 : date de départ du feu
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; HE : point d'eau à usage exclusivement humain et contenant de
l'eau ;
ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau.
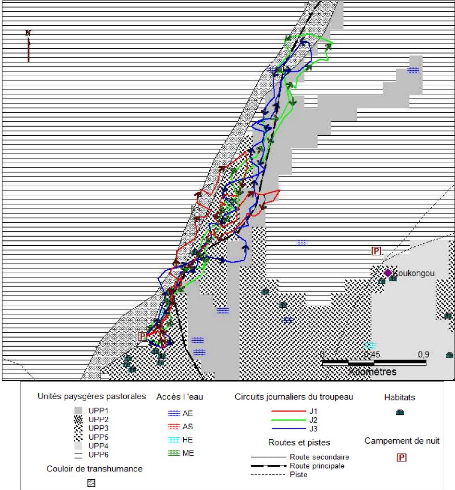

212
Carte VI-11. Itinéraires du troupeau TrpC1-2P pendant
Yaamde
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; AS : point d'eau à usage exclusivement animal et à sec
;
HE : point d'eau à usage exclusivement humain et contenant
de l'eau ; ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau.

6.3.3.2.3. Variations saisonnières de
l'utilisation des pâturages chez le troupeau de type Trp
6.3.3.2.3.1. Distances et durées moyennes du
déplacement du troupeau
Le troupeau principal conduit par le jeune frère du
propriétaire seul pendant le Dabunde, le Ndungu et le
Yaamde et aidé par deux autres bergers pendant le
Ceedu, a été présent au pâturage pendant en
moyenne plus de 9 h par jour. Sa journée commence au plus tôt
à 8h 53mn au Ceedu (saison sèche chaude) et au plus tard
à 9h 18 mn au Dabunde. Le soir, le troupeau est rentré
au parc de nuit toujours au même moment, c'est-à-dire à 18h
20mn (tableau VI-13).
La distance moyenne parcourue a été plus
importante au Ceedu (11,16 km) mais surtout pour de trajets sans
prélèvement alimentaire (3,44 km). Par ailleurs, c'est au
Yaamde que la distance parcourue quotidiennement a été
la plus faible (8,46 km) avec peu de marche entre les milieux
fréquentés.
Tableau VI-13 Temps et distance au pâturage pour le
troupeau Trp
|
Saisons
|
Heure départ
|
Heure retour
|
Durée pâturage
|
Distance parcourue (km)
|
|
Totale
|
Marche
|
Autres activités
|
|
Dabunde
|
9 h 11 #177; 06 mn
|
18 h 22 #177; 10 mn
|
9 h 10 #177; 16 mn
|
10,24 #177; 0,27c
|
2,12 #177; 0,12b
|
8,12 #177; 0,71bc
|
|
Ceedu
|
8 h 53 #177; 09 mn
|
18 h 23 #177; 05 mn
|
9 h 29 #177; 13 mn
|
11,16 #177; 0,72d
|
3,44 #177; 0,31c
|
7,72 #177; 1,02b
|
|
Ndungu
|
8 h 59 #177; 12 mn
|
18 h 20 #177; 11 mn
|
9 h 21 #177; 04 mn
|
8,97 #177; 0,16b
|
1,66 #177; 0,08a
|
7,31 #177; 0,56b
|
|
Yaamde
|
9 h 08 #177; 07 mn
|
18 h 20 #177; 05 mn
|
9 h 11 #177; 09 mn
|
8,46 #177; 0,21a
|
1,54 #177; 0,24a
|
6,92 #177; 0,23a
|
Les valeurs situées sur la même colonne et
portant des lettres distinctes sont significativement différentes au
seuil á = 0,05 à p = 0,001.
Dabunde : mi-novembre à février Ceedu
: mars à mai
Ndungu : juin à début octobre
Yaamde : octobre à début novembre
6.3.3.2.3.2. Importance relative des activités au
pâturage
Le temps consacré au broutage a été de
plus de 65% sur toute la période de suivi. Par ailleurs, alors que la
part du temps consacré au broutage et au repos-rumination a
été plus importante au Yaamde et au Ndungu,
c'est plutôt au Dabunde et au Ceedu que l'abreuvement
et le déplacement ont bénéficié de l'allocation en
temps la plus grande. On peut aussi observer que les différentes
activités ont presque le même poids dans le temps qui leur est
consacré pendant la campagne agricole (saisons Yaamde et
Ndungu).
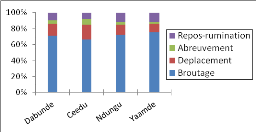

214
Figure VI-7. Proportion en temps consacré aux
activités au cours de l'année par le troupeau Trp
Dabunde : mi-novembre à février Ceedu
: mars à mai
Ndungu : juin à début octobre
Yaamde : octobre à début novembre
6.3.3.2.3.3. Le circuit pastoral quotidien selon les
saisons : unités pâturées et durée de
fréquentation
· Pendant la saison du Dabunde
(mi-novembre à février)
Le suivi de ce troupeau en cette saison s'est
déroulé du 26 au 28 novembre 2009, rappelons-le. La plupart des
champs cultivés en sorgho dans le secteur fréquenté
attendaient encore d'être récoltés. Après la traite
matinale qui se faisait très tôt, le troupeau quittait, sous la
surveillance du jeune Diawara, son parc de nuit à 9h 11mn. Il a d'abord
pris la direction des parties hautes accessibles du secteur (UPP4) en attendant
que l'humidité matinale s'assèche (carte VI-12). De là, il
a été conduit dans la vaste plaine hydromorphe (UPP2) attenante
riche en Sorgastrum bipennatum, Andropogon pseudapricus, Schizachyrium
brevifolium et surtout en de nombreuses herbes plus fraîches
désormais accessibles du fait de l'assèchement de l'unité.
Il y a passé la plus grande partie de la matinée en progressant
en direction des glacis et plaines argileuses (UPP3) puis des bas-fonds (UPP1)
sur lesquels il a pâturé les résidus culturaux disponibles
(tiges de mil ou de sorgho, fanes de niébé). Sur les portions non
cultivées de ces unités pastorales, le troupeau a exploité
une herbe aussi variée que riche comprenant Sporobus pyramidalis,
Paspalum orbiculare, Pennisetum pedicellatum, Rottboellia exaltata,
Schizachyrium brevifolium, Schizachyrium sanguineum Setaria pallidefusca,
Sclerea sp. Oryza longistaminata, Digitaria sp., Echinchloa
sp. et quelques pieds de
Vetiveria nigritana. À la mi-journée à 13h 33 mn,
le troupeau se trouvait dans la partie la plus méridionale de son
parcours, il s'est abreuvé pendant 24 mn dans l'un des nombreux points
d'eau artificiels avant un repos sous des arbres proches. Ensuite le troupeau a
repris son chemin en parcourant pratiquement les mêmes unités en
direction du campement qu'il a atteint à 18h 22 mn.
Malgré l'interêt fourrager manifeste des
bas-fonds (rythme de parcours le plus lent 9,44 m/mn ; tableau VI-14), le
troupeau a passé surtout son temps sur les plaines inondables (figure
VI-8), seul espace ouvert et librement accessible à cette période
dans cette partie du terroir. En l'espace de quelques semaines, le secteur a
connu le départ de plusieurs feux de brousse dont l'un s'est produit le
premier jour du suivi (26 novembre 2008) dans les

broussailles des plateaux (UPP4) à proximité du
parc de nuit du troupeau. Ce feu, qui a été
maîtrisé, a cependant influé sur le circuit du troupeau les
deux jours suivants. En effet, cette portion du secteur n'a plus
été que légèrement pâturée les matins
du 27 et du 28 novembre. Au total, le troupeau a parcouru 10,24 km en 9h
10mn.
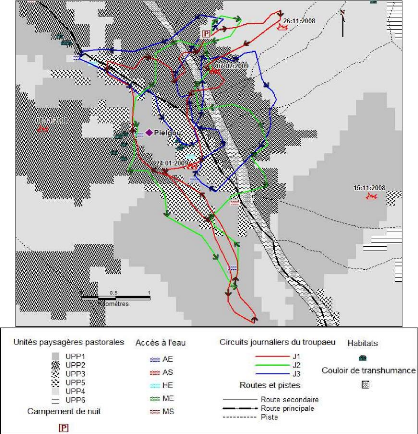
Carte VI-12. Itinéraires du troupeau Trp pendant
Dabunde
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive sur buttes rocheuses et
cuirassées ;
10 :11 :2008 : date de départ du feu
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; AS : point d'eau à usage exclusivement animal et à sec
;
HE : point d'eau à usage exclusivement humain et contenant
de l'eau ; ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau ; MS : point d'eau à usage mixte (humain et animal)
et à sec.
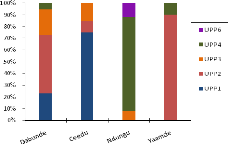
Figure VI-8 Proportions en temps de séjour dans les
unités pastorales au cours de l'année pour le troupeau Trp
Dabunde : mi-novembre à février Ceedu
: mars à mai
Ndungu : juin à début octobre
Yaamde : octobre à début novembre
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire sur buttes
rocheuses et cuirassées ;
Tableau VI-14. Vitesse moyenne de déplacement du troupeau
Trp suivant les unités et les saisons
|
Saisons
|
|
Vitesse (m/mn) par unité
paysagère
|
|
UPP1
|
UPP2
|
UPP3
|
UPP4
|
UPP6
|
|
Dabunde (mi-novembre à
février)
|
9,44
|
12,53
|
12,82
|
14,09
|
----
|
|
Ceedu (mars à mai)
|
10,78
|
13,08
|
14,35
|
----
|
----
|
|
Ndungu (juin à début octobre)
|
----
|
----
|
8,54
|
9,21
|
10,99
|
|
Yaamde (octobre à début
novembre)
|
----
|
10,77
|
----
|
9,46
|
----
|
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire sur buttes
rocheuses et cuirassées
· Pendant la saison du Ceedu (mars
à mai)
Le suivi a été effectué du 15 au 17
avril. Tout l'espace villageois était alors libre d'accès
après les récoltes et la collecte et le stockage des
résidus de cultures. Cette fois-ci, le troupeau était
encadré par 3 bergers dont deux salariés ; le parc de nuit avait
été placé plus bas dans la plaine inondable devenue
sèche et pour une bonne part consumée par un feu du 07

216

février 2009 (carte VI-13). La délocalisation du
parc de nuit en cet endroit était due au microclimat qui y était
devenu relativement plus favorable. La pâture s'est faite, comme pendant
la saison précédente dans les parties les plus basses du secteur
(UPP3 et UPP1) après un passage vers les points d'eau à usage
mixte (puits modernes et bouli) ou réservés aux animaux
(puisards) pour l'abreuvement matinal (durée moyenne: 12 mn).
Après ce premier abreuvement, le troupeau est resté pratiquement
toute la journée dans les bas-fonds (UPP1 : figure VI-8) dont le stock
fourrager était cependant fortement amoindri par de nombreux
départs de feux de brousse (15 novembre 2008, 24 janvier 2009 et 03
février 2009). Le troupeau y a exploité la paille
résiduelle épargnée par le feu. Le feu de janvier avait
été suivi de repousses fraiches de quelques pieds de vivaces,
mais celles-ci étaient en quantité insignifiante. Le
deuxième abreuvement, le plus important, s'est fait vers 14h pendant 30
mn avant un repos-rumination de 44,5 mn. Ensuite le troupeau a commencé
son retour vers le campement en passant par les mêmes unités
pastorales que celles du matin. Une grande partie des pâturages des
alentours (UPP2, UPP3 et UPP4) a été sérieusement
touchée par divers départs de feux intervenus depuis fin 2008
sans grande possibilité de repousses contrairement aux bas-fonds.
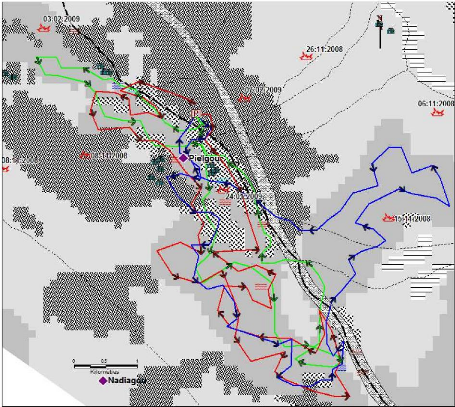


218
Carte VI-13. Itinéraires du troupeau Trp pendant
Ceedu
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire sur buttes
rocheuses et cuirassées.
10 :11 :2008 : date de départ du feu
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; AS : point d'eau à usage exclusivement animal et à sec
;
HE : point d'eau à usage exclusivement humain et contenant
de l'eau ; ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau ; MS : point d'eau à usage mixte (humain et animal)
et à sec.
Pendant son séjour dans les bas-fonds le troupeau a
progressé relativement moins rapidement que dans les autres
unités pastorales (10,78 m/mn ; tableau VI-14) ; la forme sinueuse de
son itinéraire témoigne de changements fréquents de
direction et de la recherche avide de fourrage devenu rare et surtout à
distribution très disparate. Une sorte de compensation est
réalisée au campement par un apport de résidus culturaux
collectés et de son industriel acheté. Mais ces apports restent
insuffisants au regard de la taille du troupeau. Cette pénurie va
conduire ce troupeau à la transhumance juste après notre suivi.
La distance totale moyenne parcourue en cette saison a été de
11,16 km.
· Pendant la saison du Ndungu (juin
à début octobre)
Ce troupeau, qui n'a pu faire l'objet de suivi pendant
Kotoga du fait de son absence dans le terroir, a été
retrouvé au Ndungu pour un suivi les 9, 10 et 11 août.
Son parc de nuit a de nouveau été placé sur les plateaux
(UPP4) pour fuir l'humidité et l'impraticabilité de l'ancien site
situé dans la plaine inondable voisine (UPP2) (carte VI-14). La
chaîne de pâturage est constituée principalement des
jachères et broussailles des interstices entre les nombreux champs des
plateaux. Le troupeau y a séjourné pendant les premiers moments
de la journée puis le soir au moment du retour. Le reste de la
journée, pendant les 2ème et 3ème
jours de suivi, le troupeau a progressé vers des espaces plus ouverts,
notamment les plateaux cuirassés près du quartier Banduo, en
parcourant les glacis et plaines argileuses (UPP3), unités pastorales
relativement intéressantes comme en témoigne le rythme lent de
leur traversée (8,54 m/mn ; tableau VI-14) malgré la forte
présence de champs. Les espèces qui
ont participé notablement aux prises alimentaires sont
Andropogon pseudapricus, Panicum repens, Paspalum orbiculare, Zornia
glochidiata et Eragrostis sp. A la mi-journée, le troupeau
a été positionné sur les plateaux cuirassés
où les risques étaient les plus faibles dans ce secteur, il n'en
est revenu que pour s'abreuver. Sur ces plateaux, les genres Aristida
et Alysicarpus de même que l'espèce Loudetia
togoensis fourrages assez abondants ont été les plus
pâturés. Depuis là le troupeau a amorcé son retour
vers le campement en passant par les points d'abreuvement, assez nombreux sur
UPP4. Notons qu'au premier jour de suivi, le troupeau a passé toute la
journée dans les espaces non cultivés de l'unité pastorale
UPP4 après un bref séjour dans une unité de plaine
argileuse (UPP3) située du côté nord du campement. Pour
rejoindre cette unité, il a dû emprunter à un rythme
soutenu, un sentier traversant les parties moins humides de la plaine
inondable. Selon le berger, le risque en valait la peine car l'unité
convoitée, qui apparaissait presque comme un îlot au milieu de
zones inondées était, peu pâturée par les troupeaux
des éleveurs situés dans ce secteur du terroir.
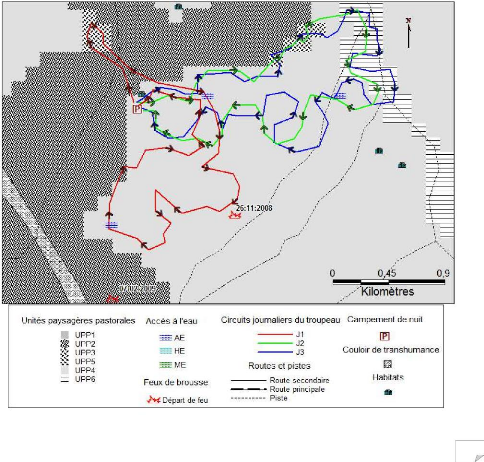
Carte VI-14. Itinéraires du troupeau Trp pendant
Ndungu
Pendant la période de suivi l'abreuvement a eu lieu
à 12 h 53 mn pendant 19 mn avant un repos de 66 mn. Le troupeau a
rejoint le campement à 18h 20 mn et la distance moyenne parcourue a
été de 8,97 km.
UPP1,

220
unité de savane arborée sur sol profond hydromorphe
à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire sur buttes
rocheuses et cuirassées.
07 :02 :2009 : date de départ du feu
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; HE : point d'eau à usage exclusivement humain et contenant de
l'eau ;
ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau.
· Pendant la saison du Yaamde (octobre
à début novembre)
Au Yaamde, le suivi du troupeau de l'éleveur
Diawara a été réalisé les 22, 23 et 24 octobre
2009. Le parc de nuit était toujours à son emplacement du
Ndungu et pendant presque toute la journée de pâturage,
le troupeau a été présent sur les plaines inondables
(figure VI-8 & carte VI-15) dont une bonne partie est devenue praticable
avec le retrait de l'eau. Celles-ci libèrent les fourrages auparavant
inaccessibles et provoquent des jeunes pousses relativement plus tendres
qu'ailleurs. Juste après la traite laitière matinale, le troupeau
a séjourné brièvement sur les plateaux en attendant la
baisse de l'humidité due à la rosée. Les plateaux
demeuraient cependant les milieux les plus intéressants (rythme de
progression moins rapide que dans les plaines ; tableau VI-14), mais les
risques y demeuraient plus élevés car de nombreux champs
étaient en phase d'épiaison-fructification et ils étaient
l'objet d'une surveillance renforcée de leurs propriétaires. Ceci
n'encourageait pas le berger à rester à proximité. Le
deuxième abreuvement, à 13h 01 mn pendant 14 mn, s'est fait dans
une mare naturelle située dans UPP2 qui contenait encore de l'eau.
L'abreuvement a duré relativement moins qu'au Ndungu parce que
cette fois il y avait eu un abreuvement matinal de 4 mn environ quelques
instants après le départ du campement. Par ailleurs, le point
d'eau semblait encore peu fréquenté, ce qui permettait au
troupeau de s'abreuver sans gêne aucune et donc de gagner du temps.
Le troupeau s'est ensuite reposé pendant 63 mn. A
partir de 15 heures le jeune berger et son troupeau ont adopté une
direction qui les a conduits progressivement vers le campement de nuit autour
de 18h 20mn après avoir parcouru une distance moyenne de 8,46km.
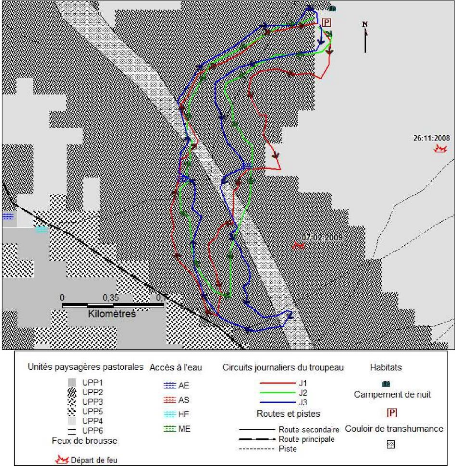

Carte VI-15. Itinéraires du troupeau Trp pendant
Yaamde
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire sur buttes
rocheuses et cuirassées.
07 :02 :2009 : date de départ du feu
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; AS : point d'eau à usage exclusivement animal et à sec
;
HE : point d'eau à usage exclusivement humain et contenant
de l'eau ; ME : point d'eau à usage mixte (humain et animal) et
contenant de l'eau.

222
6.3.3.2.4. Variations saisonnières de
l'utilisation des pâturages chez le troupeau de type TrpC3
6.3.3.2.4.1. Distances et durées moyennes du
déplacement du troupeau
Pendant les deux saisons de suivi (4, 5 & 6
décembre pour Dabunde ; 19, 20 & 21 avril pour
Ceedu), ce troupeau est resté au pâturage pendant environ
10 h de temps (9h 52 mn et 10h 19mn respectivement) et a parcouru 12,02 km et
13,03 km (tableau VI-15) dont une bonne partie a été
consacrée à la marche (3,51 à 4,02 km). Ce troupeau
transhumant est pratiquement tout le temps au pâturage (départ un
peu après 8h et retour vers 18h 30mn), ce qu'on peut expliquer par le
fait que les bergers sont entièrement concentrés sur leur
bétail, leurs familles étant au loin, en terroir d'attache.
Tableau VI-15. Temps et distance au pâturage pour le
troupeau TrpC3
Saisons Heure départ Heure retour Durée
pâturage Distance parcourue (km)
Totale Marche Autres activités
Dabunde 8 h 31 #177; 14 mn 18 h 23 #177; 11 mn 9 h 52
#177; 12 mn 12,02 #177; 0,48a 3,51 #177; 0,69a 8,51 #177;
0,92a
Ceedu 8 h 16 #177; 09 mn 18 h 34 #177; 05 mn 10 h 19
#177; 06 mn 13,03 #177; 0,65a 4,02 #177; 0,86a 9,01 #177;
1,08a
Les valeurs situées sur la même colonne et
portant des lettres identiques sont statistiquement égales au seuil
á = 0,05 à p = 0,001.
Dabunde : mi-novembre à février Ceedu
: mars à mai
6.3.3.2.4.2. Importance relative des activités au
pâturage
Au pâturage le troupeau consacre presque 70% de son
temps à brouter quelle que soit la saison (figure VI-9). Le temps
consacré au repos-rumination et, dans une moindre mesure, au
déplacement est plus long en début de saison sèche
(Dabunde) qu'en saison sèche chaude. A l'inverse, le temps
consacré à l'abreuvement est bien plus faible au Dabunde
(environ 2%) qu'au Ceedu (environ 6%).
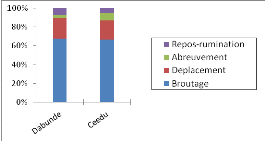
Figure VI-9. Proportions en temps consacrés aux
activités au cours de l'année par le troupeau TrpC3.
Dabunde : mi-novembre à février Ceedu
: mars à mai

6.3.3.2.4.3. Le circuit pastoral quotidien selon les
saisons : unités pâturées et durée de
fréquentation
L'éleveur considéré est un transhumant
venant du nord du pays, précisément de la commune de Botou. Assez
connu dans cette partie du terroir grâce à ses fréquents
séjours, il bénéficie d'un traitement relativement
exceptionnel qui lui permet d'exploiter les parcelles récoltées
et nettoyées (collecte de résidus culturaux) de ses tuteurs
à proximité desquels il a établi son campement de
fortune.
· Pendant la saison du Dabunde
(mi-novembre à février)
Quand nous l'avons rencontré pour demander
l'autorisation de suivre son troupeau, l'éleveur Baadio était
arrivé dans le terroir l'avant-veille. Après avoir beaucoup
hésité il nous a permis de suivre le troupeau du 4 au 6
décembre. A cette période, beaucoup de champs environnant,
notamment ceux des plateaux à sols plus ou moins profonds (UPP5) et ceux
des bas glacis et plaines argileuses (UPP3), étaient déjà
récoltés, mais le ramassage des résidus culturaux
n'était pas achevé ce qui n'autorisait encore qu'une vaine
pâture limitée. L'espace le plus ouvert et à accès
le moins risqué était donc surtout l'environnement
immédiat du campement et les jachères de divers âges des
plateaux proches (UPP4). Malheureusement, plusieurs départs de feux
avaient consumé une grande partie de sa biomasse d'herbes annuelles
produite pendant le Ndungu.
Après une traite laitière rapide, au
départ du campement le matin, le troupeau a parcouru pendant quelques
minutes les jachères puis s'est dirigé vers le bas-fond
situé plus bas, en suivant des sentiers, où il s'est rapidement
abreuvé (environ 7 mn) le long du cours d'eau (carte VI-16). De
là il a progressé, en parcourant quelques parcelles
libérées des cultures, vers les parties hautes et non
brûlées du secteur (UPP6 et UPP4) où il a passé la
première moitié de la journée en consommant un fourrage
plutôt médiocre dominé par Aristida funiculata,
Andropogon pseudapricus, Schizachyrium exile, Loudetia togoensis ainsi que
quelques espèces de jachères comme Setaria pallide-fusca
et Pennisetum pedicellatum, espèces déjà toutes
parvenues à maturité. Aux alentours de 12 h, il est redescendu
dans les zones cultivées (UPP5 beaucoup plus qu'UPP3) pour exploiter
surtout les résidus de culture et les broussailles situées entre
les champs. Le troupeau est resté à cet endroit avant et
après l'abreuvement suivi du repos puis, le soir, a regagné le
campement. Le second abreuvement a eu lieu à 13h 52mn pendant 12 mn
avant un repos-rumination de 44 mn à quelques pas du point d'eau.
Au total, le troupeau a fréquenté surtout les
plateaux cultivés (figure VI-10) mais les unités qui apparaissent
les plus attrayantes ont été les plaines cultivées (UPP3)
que le troupeau a parcouru relativement lentement (tableau VI-16). Mais il ne
pouvait y demeurer longtemps la récolte n'était pas
achevée dans ces zones de sorgho à cycle long et les
résidus de culture, assez recherchés, n'avaient pas encore
été collectés.
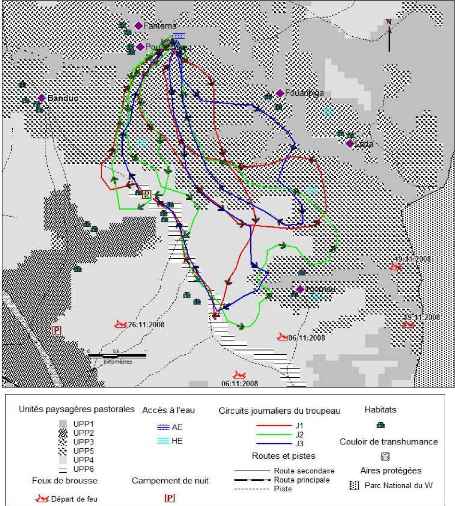

224
Carte VI-16. Itinéraires du troupeau TrpC3 pendant
Dabunde
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire sur buttes
rocheuses et cuirassées.
06 :11 :2008 : date de départ du feu
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; HE : point d'eau à usage exclusivement humain et contenant de
l'eau.

Figure VI-10. Proportions en temps de séjour dans les
unités pastorales au cours de l'année pour le troupeau TrpC3.
Dabunde : mi-novembre à février Ceedu
: mars à mai
Tableau VI-16. Vitesse moyenne de déplacement du troupeau
TrpC3 suivant les unités et les saisons
Vitesse (m/mn) par unité
paysagère
Saisons UPP1 UPP3 UPP4 UPP5 UPP6
Dabunde ---- 10,68 14,55 12,02 15,27
Ceedu 12,70 14,76 ---- 14,96 ----
Dabunde : mi-novembre à février
Ceedu : mars à mai UPP1, unité de savane
arborée sur sol profond hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire sur buttes
rocheuses et cuirassées.
· Pendant la saison du Ceedu (mars
à mai)
Au Ceedu, tout l'espace était ouvert et les
bas-fonds étaient asséchés et très appauvris par
une longue période de pâture et une forte pression d'exploitation.
De nouveaux horizons (dans l'unité UPP1 notamment) étaient alors
explorés par les bergers et leur troupeau tandis que les unités
pâturées les mois précédents étaient
désormais délaissées ou peu visitées (UPP6 &
UPP4) (carte VI-17). Après un détour matinal pour s'abreuver
(durée : 18 mn) au point d'eau (un puisard érigé par les
soins des bergers), le troupeau a donc passé toute sa journée
dans les bas-fonds où les ressources étaient relativement plus
intéressantes comme l'atteste le rythme lent de progression (tableau
VI-16). Il a brièvement visité les unités cultivées
(UPP5 & UPP3). Le second abreuvement de 31 mn, vers 13h 26mn a eu lieu dans
un secteur éloigné, au-delà des villages de Fouanbiga et
Lada. Le repos-rumination qui a suivi n'a duré que 33 mn.
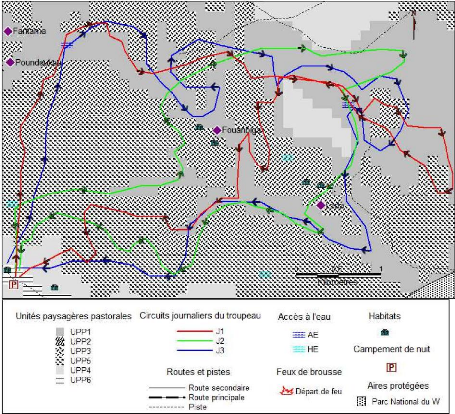

226
Carte VI-17. Itinéraires du troupeau TrpC3 pendant
Ceedu
UPP1, unité de savane arborée sur sol profond
hydromorphe à pseudogley de surface ;
UPP2, unité de savane boisée claire sur plaine
inondable et sol hydromorphe à pseudogley de surface ;
UPP3, unité de savane arbustive de moyen et bas glacis
sur sols ferrugineux tropicaux à tâches et concrétions ;
UPP4, unité de savane arbustive claire de plateaux et
hauts glacis sur sols ferrugineux tropicaux lessivés indurés ;
UPP5, unité de mosaïque agroforestière sur
sols ferrugineux tropicaux lessivés indurés ;
UPP6, unité de savane arbustive claire de buttes
rocheuses et cuirassées.
AE : point d'eau à usage exclusivement animal et contenant
de l'eau; HE : point d'eau à usage exclusivement humain et contenant de
l'eau.
Le territoire exploré aura été plus vaste
qu'au Dabunde et les itinéraires auront été moins
linéaires, ce qui montre que la qualité du parcours s'est
dépréciée avec le temps.

6.3.3.3. Synthèse sur le comportement du
troupeau au pâturage
6.3.3.3.1. Un comportement territorial,
révélateur de l'état (disponibilité et distribution
des ressources) des parcours.
Les circuits de troupeaux suivis pendant les cinq saisons du
calendrier pastoral ont montré que chacun des éleveurs exploite
un secteur particulier du terroir avec parfois une mobilité des parcs de
nuit pour prévenir les risques de vol (en saison sèche le parc
est ramené près des concessions) et d'humidité (en saison
pluvieuse le campement est délocalisé sur les hauteurs : plateaux
et flancs de la chaîne). Le troupeau transhumant résident (Trp),
qui a le plus changé l'emplacement du parc de nuit de son troupeau,
exploite le secteur le plus humide du terroir.
Le tableau VI-17 reprend de manière synthétique
les temps de présence au pâturage et les distances parcourues par
l'ensemble des troupeaux au cours de la période de suivi (novembre 2008
à octobre 2009). Les statistiques issues de notre étude de quatre
troupeaux montrent que les plus grandes distances sont couvertes en saison
sèche particulièrement en fin de saison de soudure (Ceedu /
Ku tontogu) et que plus le troupeau est de grande taille, plus grande est
cette distance exception faite au Kotoga /A sakoana où la plus
grande distance est réalisée par le plus petit (TrpC1-2G) des
deux troupeaux qui ont été suivis. Petit (2000a &
2000b), dans ses travaux à Kourouma, fait les mêmes observations
chez les agroéleveurs résidents de cette localité; il en
est de même des observations faites par Diallo (2006) à
Koumbia-Waly. Nos données s'opposent assez sensiblement à celles
obtenues dans le centresud du Burkina (Kagoné, 2000), ou auprès
des éleveurs peuls de Kourouma (Petit, 2000a) ou encore chez une
communauté Mbororo de Centrafrique (D'Amico et al. 1995) ou du
nordCameroun (Dongmo, 2009). Par ailleurs, Petit (2000a) et Dongmo (2009) ont
observé comme nous que les troupeaux des agroéleveurs
sédentaires (équivalents des troupeaux gourmantchés :
TrpC1-2G) couvraient moins de distance que ceux des éleveurs
professionnels (ici Trp à TrpC3) quelle que soit la saison. La
première explication qui peut être avancée est que les
troupeaux des agro éleveurs sont généralement de plus
petite taille que ceux des Peuls, ce qui leur permet un déplacement
moins soutenu sur parcours. Cette taille relativement petite est aussi un atout
lorsqu'il s'agit d'exploiter les interstices des unités fortement
cultivées comme UPP5, UPP3 et UPP1.
Tableau VI-17. Temps de présence au pâturage et
distances parcourues par les troupeaux au cours de la période de
suivi
|
Paramètres
|
Saisons
|
TrpC1-2G
|
TrpC1-2P
|
Trp
|
TrpC3
|
|
Dabunde
|
7,63
|
8,84
|
10,24
|
12,02
|
|
Ceedu
|
9,94
|
10,77
|
11,16
|
13,03
|
|
Distance moyenne
|
Kotoga
|
12,19
|
10,36
|
|
----
|
|
Parcourue (km)
|
Ndungu
|
6,49
|
8,35
|
8,97
|
----
|
|
Yaamde
|
7,13
|
8,2
|
8,46
|
----
|
|
Moyenne
|
8,68
|
9,3
|
9,71
|
12,53
|
|
Dabunde
|
8h 31mn
|
9h 17mn
|
9h 10mn
|
9h 52mn
|
|
Temps moyen
|
Ceedu
|
9h 52mn
|
9h 19mn
|
9h 29mn
|
10h 19mn
|

228
au pâturage Kotoga 9h 30mn 9h
10mn ----
Ndungu 8h 30mn 9h 04mn 9h 21mn ----
Yaamde 8h 07mn 8h 57 mn 9h 11mn ----
Moyenne 8h 54mn 9h 09mn 9h 18mn 10h 05mn
Dabunde = fin Ku fowagu (mi-novembre à
février) Ceedu = Ku tontogu (mars à mai)
Kotoga = A sakoana (fin mai à début
juin)
Ndungu = Ku siagu (juin à début
octobre)
Yaamde = début Ku fowagu (octobre à
début novembre)
Dans nos observations les troupeaux ont passé plus de
temps au pâturage en saison sèche qu'en saison pluvieuse, ce qui
est conforme aux observations de Dongmo (2009) mais contraire à celles
de Daget & Godron (1995) et Petit (2000a) ; ces deux derniers auteurs font
remarquer qu'en saison sèche, les animaux diminuent leurs
activités de pâturage (temps de présence au pâturage)
tout en augmentant leur durée de prise alimentaire. Il nous semble
qu'une telle affirmation doit cependant être nuancée. Nous avons,
en effet, constaté que le troupeau passait beaucoup de temps à se
déplacer d'un secteur à un autre en saison sèche et, en
conséquence, consacrait moins de temps au broutage en cette saison
comparativement à la saison pluvieuse.
Tableau VI-18. Diverses données de distance parcourue par
le troupeau au pâturage.
|
|
|
Saisons
|
|
Sources
|
|
Saison sèche
|
|
Saison humide
|
|
Dabunde
|
Ceedu
|
Kotoga
|
Ndungu
|
Yaamde
|
|
10
|
10,9
|
8
|
8
|
8,3
|
Diallo, 2006
|
|
5,22
|
4,82
|
4,38 - 9,60
|
7,99
|
5,94
|
Dongmo, 2009
|
|
|
|
|
|
|
(agroéleveurs)
|
|
Distance au
pâturage
|
9,84
|
8,30
|
----
|
9,46
|
8,60
|
Dongmo, 2009
(éleveurs)
|
|
(km)
|
----
|
12
|
----
|
9
|
----
|
Petit, 2000a
|
|
|
|
|
|
|
(agroéleveurs)
|
|
----
|
7
|
----
|
9
|
----
|
Petit, 2000a (éleveurs)
|
|
----
|
----
|
----
|
7,5 - 13,7
|
----
|
Kagoné, 2000
|
|
|
5-8
|
|
> 8
|
|
D'Amico et al. 1995
|
6.3.3.3.2. Un objectif sous-jacent : maximiser le bilan
énergétique
Nos données montrent qu'en général, la
saison sèche (Ceedu / Ku tontogu et Kotoga /A sakoana)
est la période où le troupeau, quel qu'il soit, parcourt le plus
de distance. Il y a, par ailleurs, plus de temps mis au pâturage, une
grande part de ce temps étant allouée aux activités de
déplacement et d'abreuvement également beaucoup plus importantes
en cette saison (figure VI-11). La figure montre, par ailleurs, qu'au
pâturage le troupeau consacre au moins 60% de son temps au
prélèvement alimentaire (broutage), le reste du temps
étant consacré par ordre d'importance au déplacement, au
repos-rumination et, enfin, à

l'abreuvement. Cette allocation de temps aux activités
est assez fluctuante en fonction des saisons. Le broutage et le
repos-rumination atteignent leur maximum en saison pluvieuse, période
où le déplacement et l'abreuvement, au contraire, sont à
leur minimum et inversement en saison sèche. Ainsi, selon les
périodes, le comportement des troupeaux est tributaire d'une
préoccupation essentielle relative à deux paramètres
(l'accès à l'eau et au fourrage) : subvenir à leurs
besoins au moindre coût. Pike et al. (1977) ainsi que Dumont et
al. (2001) le confirment lorsqu'ils rappellent que le choix des sites
par les éleveurs et leurs troupeaux est déterminé par la
théorie de l'alimentation optimale. D'après cette théorie
(Kagoné, 2000 ; Dumont et al. 2001), le troupeau sur parcours
cherche constamment à maximiser son bilan énergétique.
Cette préoccupation sous-jacente qui guide le troupeau, à travers
les animaux leaders, va pousser celui-ci à " être assez regardant
" sur le coût énergétique que vont exiger les
déplacements entre les différents secteurs du territoire pastoral
(Wallis de Vries, 1996). Ce qui va déterminer au final le choix de
fréquenter ou de délaisser certains secteurs du terroir, le
troupeau étant capable selon Dumont et al. (2001) de moduler
son déplacement en fonction du rapport entre la valeur du site (gain
espéré en énergie) et la distance à parcourir pour
y accéder (dépense en énergie).
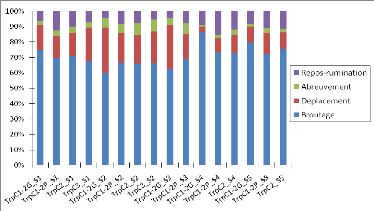

230
6.3.3.3.3. Des itinéraires dont la forme est
imprimée par la conjonction de plusieurs paramètres.
Plus les pâturages sont riches (cas de la plupart des
unités paysagères pastorales en saison pluvieuse, des
unités cultivées pendant la vaine pâture et des
unités de bas de pente en certains moments de la saison sèche),
plus le rythme de paissance (prise alimentaire) est lent. A contrario,
plus les pâturages sont pauvres (saison sèche chaude), plus ce
rythme est rapide (Bailey et al. 1996 in Kagoné, 2000 ; Diallo,
2006). Selon Petit (2000a), cette variation de rythme peut faire baisser
sensiblement l'efficacité de la pâture par suite de grandes
dépenses énergétiques en période de pénurie.
Par ailleurs, ces déplacements sont caractérisés, en cette
saison de disette (saison de rareté et d'extrême
discontinuité des ressources), par des circuits relativement plus amples
et plus sinueux témoins d'un espace plus ouvert et de fréquentes
marches des troupeaux d'un secteur à un autre du territoire afin
d'exploiter la grande diversité des milieux. Le temps plus long
consacré à l'abreuvement en saison sèche s'explique par la
pression qui s'exerce sur les points d'eau devenus moins nombreux,
localisés en des points particuliers du paysage, souvent
regroupés et le plus souvent d'usage mixte. Ceux des éleveurs,
les plus nombreux d'ailleurs, qui ne peuvent avoir accès à ces
points d'eau (c'est le cas des transhumants) ou qui fuient la concurrence
consécutive à cette pression, vont creuser des puisards dans les
lits des cours d'eau. L'abreuvement en ces points d'eau de fortune prend
énormément du temps puisque le berger doit faire remonter l'eau
avec des puisettes.
En saison pluvieuse (Ndungu / Ku siagu
surtout) quand l'herbe et l'eau (flaques et mares disséminées
dans les pâturages) sont abondantes dans l'ensemble du territoire, les
déplacements des troupeaux sont plus linéaires, moins amples et
s'effectuent à un rythme lent, preuve d'une relative abondance
(Kagoné, 2000 ; Diallo, 2006). Parallèlement, un temps de repos
plus long est nécessaire pour permettre de digérer les grandes
quantités de nourriture ingérées à la suite d'une
grande activité de broutage (figure VI-11).
La taille du troupeau semble aussi influencer son comportement
au pâturage. Nos données indiquent en effet que, plus le troupeau
est grand, plus la distance qu'il parcourt chaque jour est grande (exemple
TrpC3). Cette constatation a été aussi faite par Petit (2000a)
à Kourouma dans l'ouest burkinabè et par Dongmo (2009) au
Nord-Cameroun. En effet, les grands troupeaux épuisent plus vite le
fourrage d'un secteur donné du pâturage et passent plus rapidement
à un autre. Une autre explication logique réside dans le fait
que, plus le troupeau est grand, plus ses besoins sont importants et il lui
faut davantage de fourrage. Il se déplacera alors à un rythme
plus élevé et couvrirait en conséquence une plus grande
distance.
6.3.4. Représentations versus pratiques : les
éleveurs et leurs troupeaux à l'épreuve du terrain
Divers travaux décrivent la chaîne de
pâturage annuelle et permettent de suivre les différents milieux
que les troupeaux occupent au fil de l'année. Les spécialistes
qui se sont intéressés à la question (Guillaud, 1994 ;
Daget & Godron, 1995 ; Kagoné, 2000 ; Petit, 2000a ; Diallo, 2006 ;
Kièma S., 2007 ; Dongmo, 2009 ; Vall & Diallo, 2009) montrent qu'en
saison pluvieuse (Ndungu / Ku siagu) les éleveurs se replient
sur les hauteurs, en particulier

collines mais aussi plateaux et hauts glacis aux sols peu
profonds, endroits généralement bien secs aux sols fermes et
incultes. Ils évitent ainsi des risques notamment sanitaires
(pathologies et accidents mécaniques) liés à
l'humidité et des risques de conflits liés aux
dégâts champêtres éventuels. Dès que ces
risques commencent à diminuer du fait de la cessation des pluies et / ou
de la récolte des cultures, les éleveurs commencent à
descendre avec leurs troupeaux en direction des parties des terroirs les plus
basses (plateaux et plaines susceptibles de regorger de l'eau ou de permettre
d'en rechercher (puisards) en saison sèche chaude ou saison de soudure
(Ceedu & Kotoga / Ku tontogu & A sakoana).
En début de saison sèche (Dabunde), la vaine
pâture va maintenir pendant un bon moment les troupeaux sur les zones
cultivées (UPP5 et aussi UPP3 et UPP1). Lorsque la saison « de
soudure » arrive, les unités pastorales sont toutes appauvries en
fourrage et la plupart en eau. Les milieux pouvant proposer de l'eau ou
permettant d'en rechercher par le creusage de puisards (Mbundu en
langue peule) deviennent alors importants. Par ailleurs, les premières
pluies (pendant Kotoga-Korse / A sakoana), vont occasionner
l'émergence de jeunes pousses d'adventices très
recherchées sur ces milieux cultivés et qui vont y attirer les
troupeaux (Petit, 2000a).
Ce schéma général, qui est pratiquement
celui que nous avons pu reconstituer théoriquement en focus group avec
nos éleveurs (voir représentations ; figures VI-12a &
VI-13a), est respecté dans la plupart des circuits que nous avons
enregistrés. Quelques ajustements révèlent des situations
particulières locales relatives à la conformation du terrain, aux
pratiques et à la personnalité du bouvier.
Le troupeau gourmantché du groupe d'éleveurs
sédentaires (TrpC1-2G) se trouve dans un secteur où en saison des
pluies, la concentration des champs est importante surtout sur les bas-fonds
(UPP1 : Ku bagu), les plaines et bas-glacis (UPP3 : Li
tinbuali) et une bonne partie des plateaux et hauts glacis (UPP4 : Li
tinmuali et U gbanu). Le bouvier ne dispose alors en
définitive que de la chaîne du Gobnangou sur laquelle il passe le
maximum de son temps. La petite taille du troupeau, du coup plus facile
à contrôler, permet au bouvier de prendre le risque de
pâturer dans les espaces interstitiels surtout sur les plateaux nettement
plus intéressants (plus forte biomasse de meilleure qualité). La
prise de risque est claire puisqu'avant même les récoltes, les
parties sèches des bas-fonds, encore occupés en grande partie de
sorgho, sont visitées. De plus, ce troupeau, contrairement à ce
qu'on aurait pu attendre, n'est pas présent dans les bas-fonds en fin de
saison sèche (A sakoana). C'est parce qu'il a commencé
à être complémenté (fourrage en stock ; boisson
à base de son) dès le début de la saison sèche
chaude (Ceedu), ce qui évite alors de s'éloigner de son
campement. Le seul bas-fond situé à proximité est
très dégradé parce que l'eau d'abreuvement encore
présente y concentre un nombre élevé de troupeaux. Le
troupeau étudié l'évite donc. Globalement, les
unités les plus exploitées sont celles des buttes rocheuses
(UPP6) (saison pluvieuse) et celle des bas-glacis et plaines non inondables
(UPP3) (saison sèche).
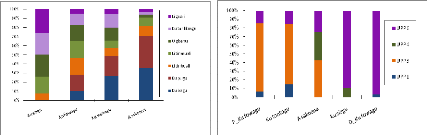
a. Représentations chez les Gourmantché b.
Pratiques chez le troupeau TrpC1-2G

232
Figure VI-12. Comparaison entre représentations et
pratiques saisonnières de conduite des troupeaux au pâturage chez
les éleveurs gourmantchés
D_Ku fowagu (début Ku fowagu):
mi-octobre à début-novembre P_Ku fowagu (plein Ku
fowagu): novembre à février
Ku tontogu : mars à début mai
A sakoana : fin mai à début juin
Ku siagu : juin à début octobre
Ku bagu= UPP1 (unité de savane arborée sur
sol profond hydromorphe à pseudogley de surface)
Ku pugu= UPP2 (unité de savane boisée
claire sur plaine inondable et sol hydromorphe à pseudogley de
surface) Li tinbuali = UPP3 (unité de savane arbustive de moyen
et bas glacis sur sols ferrugineux tropicaux à tâches et
concrétions)
U gbanu / Li tinmuali = UPP4 (unité de savane
arbustive claire de plateaux et hauts glacis sur sols ferrugineux tropicaux
lessivés indurés)
U gbanu / Li tinmuali (champs) = UPP5 (UPP 4
cultivé ; unité de mosaïque agroforestière sur sols
ferrugineux tropicaux lessivés indurés)
Li guali / Ku tankiangu = UPP6 (unité de savane
arbustive claire de buttes rocheuses et cuirassées).
Le troupeau peul du groupe d'éleveurs résidents
sédentaires (TrpC1-2P), pendant la saison pluvieuse (Ndungu), a
fréquenté les bas-fonds et plaines voisines dont l'abord est en
principe délicat à cause de nombreux champs. Ceci montre que les
éleveurs exercent une surveillance continue des ressources du terroir
qu'ils exploitent dès qu'une possibilité leur est offerte. Dans
le cas considéré, une poche de sécheresse a rendu cette
solution possible, certains secteurs de ces unités étant devenus
momentanément praticables. Ceci est une preuve que les buttes rocheuses
de la chaîne sont plutôt perçues comme une zone refuge (ou
zone de repli) que comme un pâturage à part entière,
à même de subvenir convenablement aux besoins des animaux. Par
ailleurs, la très grande distance qui sépare les plaines
inondables de ce secteur, n'encourage pas les bouviers à y conduire le
troupeau en début de saison sèche (Dabunde) alors
qu'elles sont susceptibles de regorger d'énormes ressources.
Le troupeau peul résident transhumant (Trp) se trouve
dans un secteur assez hydromorphe et cloisonné en saison pluvieuse par
la forte concentration de champs (sauf sur UPP2) et sans possibilité de
repli. L'unité UPP2 prend du coup un rôle capital dans
l'alimentation du troupeau à laquelle elle contribue
énormément notamment en fin de saison des pluies
(Yaamde) et pendant la vaine pâture (Dabunde) où
elle vient en complément des parcelles cultivées. Par ailleurs,
malgré le risque que représente la fréquentation des
unités des plateaux

(UPP4) qui sont, dans ce secteur, en grande partie
cultivées, le troupeau s'y rend pendant le Ndungu. C'est parce
que les espèces fourragères y sont assez diversifiées et
productives (forte biomasse) pour que le risque vaille la peine d'être
pris.
Le troupeau peul transhumant non résident (TrpC3) se
contente pendant le Dabunde de l'espace que veulent bien lui laisser
les agroéleveurs gourmantchés riverains dans les espaces
cultivés. Par ailleurs, avec les feux de brousse, assez présents
dans le secteur, le troupeau se trouve confiné et il est contraint
à de longs déplacements sur les espaces épargnés
par le feu ou sur des filets de broussaille entre les champs et surtout sur les
parties cultivées et ouvertes à la vaine pâture. En saison
sèche chaude, la pression sur les points d'eau du secteur est
très élevée et les puisards creusés sont
éphémères (nappe phréatique assez basse), ce qui
contribue à allonger davantage les distances journalières.
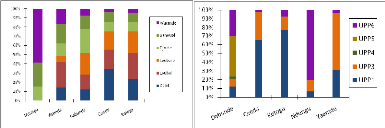
a. Représentations chez les Peul b. Pratiques chez le
troupeau TrpC1-2P
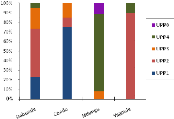
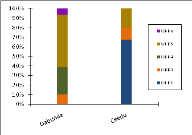
c. Pratiques chez le troupeau Trp d. Pratiques chez le troupeau
TrpC3
Figure VI-13. Comparaison entre représentations et
pratiques saisonnières de conduite des troupeaux au pâturage chez
les éleveurs Peuls
Yaamde: mi-octobre à début-novembre
Dabunde: novembre à février
Ceedu : mars à début mai
Kotoga / Korse : fin mai à début juin
Ndungu : juin à début octobre
Celol= UPP1 (unité de savane arborée sur
sol profond hydromorphe à pseudogley de surface)
Loubal= UPP2 (unité de savane boisée
claire sur plaine inondable et sol hydromorphe à pseudogley de
surface) Loubare = UPP3 (unité de savane arbustive de moyen et
bas glacis sur sols ferrugineux tropicaux à tâches et
concrétions)

234
Banouol / Djolde = UPP4 (unité de savane
arbustive claire de plateaux et hauts glacis sur sols ferrugineux
tropicaux lessivés indurés)
Banouol / Djolde (champs) = UPP5 (UPP 4 cultivé ;
unité de mosaïque agroforestière sur sols ferrugineux
tropicaux lessivés indurés)
Waamde = UPP6 (unité de savane arbustive claire
de buttes rocheuses et cuirassées).
6.4. Conclusion
L'étude des savoirs techniques et des pratiques des
éleveurs a montré que ceux-ci ont une bonne connaissance de leurs
milieux qu'ils catégorisent sur des critères topographiques,
géomorphologiques et fonctionnels (types d'usage agropastoral). Par
ailleurs, les éleveurs, qu'ils soient Peuls ou Gourmantchés,
classifient leurs catégories pastorales selon plusieurs
paramètres dont l'importance varie selon les saisons. En saison
pluvieuse, ce sont la praticabilité des sites liée à leur
degré d'humidité et à l'absence de risque de conflits
pouvant résulter de dégâts champêtres, qui sont les
paramètres mis en avant pour juger de l'intérêt d'un
pâturage. La qualité des fourrages est aussi prise en compte
notamment pendant la phase de fin de développement de la strate
herbacée (Yaamde : octobre-novembre). En saison sèche,
notamment chaude, l'accès à l'eau d'abreuvement parait être
le critère primordial avec la disponibilité en fourrage.
La mise en parallèle entre chaîne de
pâturage dans les représentations des éleveurs et
chaîne de pâturage effectivement réalisée montre que
les pratiques des éleveurs s'opèrent selon un schéma qui
prend en compte l'état de la ressource (nature, quantité et
qualité du fourrage) et les contraintes (accessibilité, pression
d'utilisation, distance au campement, feux de brousse, etc.) qui se posent
à eux. Par exemple on s'attendait à ce qu'en fin de saison
sèche chaude (Kotoga ou A sakoana : voir TrpC1-2G et
TrpC1-2P), les troupeaux soient bien présents sur les unités
cultivées (plaines sablo-limoneuses et plateaux) où des jeunes
pousses surviennent suite aux premières pluies, mais il n'en est rien.
En réalité, les pluies étaient peu tombées et n'ont
pas pu provoquer la germination notable d'adventices, bien recherchées
en cette période.
L'autre enseignement notable de cette étude, c'est que,
à cause des nombreuses contraintes (champs et sites impraticables
notamment en saison de pluies ; rareté des points d'eau, grande
pauvreté des parcours en fin de saison sèche), les
éleveurs semblent, pour ce qui est des quatre cas suivis, se cantonner
chacun dans le secteur du territoire pastoral situé à
proximité de son campement. Cette attitude semble s'opérer selon
la théorie de l'optimisation des gains ou encore de l'alimentation
optimale qui est de maximiser le bilan énergétique comme l'a
rappelé Kagoné (2000). En clair, l'éleveur évite de
faire dépenser de l'énergie à son troupeau, le
déplacement ayant un coût comme le rappelle Kièma S.
(2007), sans être sûr de la compenser. Il faut noter avec Daget
& Godron (1995) que l'éleveur sur parcours se confronte à une
double exigence : (i) produire de la viande, du lait, de la laine, etc. de la
manière la plus rationnelle et la moins coûteuse, (ii) veiller en
même temps à maintenir ou à restaurer l'équilibre du
milieu qu'il exploite. Si la première exigence est en permanence
à l'esprit des éleveurs de notre terroir, la deuxième
semble peu prise en compte (éleveurs de

type C1 : sédentaires aux petits effectifs) ou non
affirmée (éleveurs transhumants non résidents de type C3)
sauf chez les éleveurs résidents transhumants.
Tout indique finalement qu'en situation d'absence de
contraintes il est possible de traduire le comportement des troupeaux au
pâturage qui ne serait que la traduction pratique des savoirs techniques
que mettent en oeuvre les éleveurs pour faire exploiter les milieux par
leurs troupeaux.
CHAPITRE VII

236
DISCUSSION GENERALE : SYNTHESE,
CONCLUSION ET PERSPECTIVES

237
Tout le long de la présente recherche, nous avons
traité de l'état des ressources pastorales, des systèmes
d'élevage et de leur évolution récente, puis avons
insisté sur le comportement des éleveurs et de leurs troupeaux
sur parcours en fonction d'un certain nombre de déterminants à
l'échelle du terroir de Kotchari. Nous reprenons ici, de manière
synthétique, les principaux résultats, montrons
l'intérêt de l'approche utilisée, faisons un aperçu
des apports en termes de connaissances et des applications éventuelles
et terminons en proposant des scénarios de
coévolution93 durable des systèmes d'élevage et
des systèmes naturels dans le terroir.
7.1. Synthèse
Les conclusions majeures auxquelles la recherche a abouti
indiquent que :
Les ressources pastorales du terroir sont de
qualité acceptable mais elles sont en voie de
dégradation.
Le terroir de Kotchari regorge de pâturages dont les
valeurs et les qualités sont acceptables quoique pas
particulièrement attrayantes. L'ensemble des ressources du terroir qui
sont accessibles légalement (unités hors de l'aire
protégée) ou illégalement (unités dans l'aire
protégée) est assez hétérogène. Les
données qui les concernent entrent dans la fourchette de celles
observées ailleurs en zone écoclimatique similaire (transition
entre la zones sud et nordsoudanienne). Toutefois, même si cela n'est pas
encore alarmant, les signes observés sur certaines unités, en
particulier les plus cultivées et les plus pâturées (fort
remaniement de la strate herbacée avec une forte présence de
phorbes, plus grande richesse spécifique herbacée, plus grande
équitabilité, début d'embuissonnement, etc.) laissent
supposer que celles-ci sont à des stades plus ou moins avancés de
dégradation. Les six unités paysagères pastorales qui ont
été définies hors de l'aire protégée
présentent des caractères un peu différents des trois qui
se situent à l'intérieur. En dehors de l'aire
protégée, les unités ont une richesse locale et une
équitabilité plus grandes et donc une plus grande
homogénéité de poids des diverses espèces ; ceci
est surtout vrai dans les unités subissant la plus forte pression
anthropique, qui constituent généralement les zones les plus
cultivées actuellement (savane parc : UPP5) ou dans un passé
relativement proche (jachères de différents âges des
glacis, plaines et plateaux aux sols plus ou moins profonds : UPP3 et UPP4).
Ces unités, qui hébergent aussi les fourrages de meilleure
qualité et qui, par ailleurs, sont avec les buttes rocheuses et
cuirassées (UPP6) les plus pâturées, sont en revanche parmi
celles qui produisent le moins de phytomasse herbacée. L'ensemble de ces
unités exploitables en toute légalité montrent par
ailleurs une richesse spécifique et une équitabilité plus
grandes que celles de l'aire protégée voisine qui produit en
revanche plus de phytomasse de bien meilleure qualité fourragère.
Les diverses pressions liées à l'usage de ces espaces sont de
nature, d'intensité et de périodicité très
variables, elles vont croissant ce qui est source d'inquiétude.
93 Ne pas prendre ce concept dans son acception
écologique première qui signifie que lorsque deux espèces
interagissent de manière étroite celles-ci peuvent évoluer
conjointement par transformation de leur patrimoine génétique
sous l'effet de pressions de sélection réciproques (Betsch et
al. 2003). Par coévolution, nous voyons plutôt deux
systèmes qui s'adaptent les uns des autres par suite des interactions
qui se produisent entre eux.

Les ressources pastorales sont soumises à des
pressions diverses et variables, parfois fortes localement.
Les unités pastorales du terroir sont soumises à
une pression anthropique forte qui trouve son origine dans la forte
densité démographique (supérieure à la moyenne de
la province) et dans la forte densité animale. Cela a été
rendue possible par des mutations locales des systèmes de production
(immigration agricole, diversification des systèmes de production avec
des Peuls devenus agriculteurs et des Gourmantchés devenus
éleveurs) et par la transhumance. Cette pression anthropique, qui a
conduit à un taux d'occupation agricole des terres, supérieur
à 50%94, s'exprime différemment en saison sèche
et en saison pluvieuse.
En saison sèche, tout l'espace du terroir et donc
toutes les unités agrostologiques sont accessibles, le terroir attire
alors des troupeaux transhumants importants dont les effectifs bovins, au
moment de la pointe de la transhumance située en saison sèche
chaude (avril à mai notamment), peuvent être aussi
élevés ou même dépasser les effectifs locaux. Le
bilan fourrager théorique, réalisé en mettant en balance
les capacités de charge théoriques globales et les charges
observées, ne montre actuellement pas de risque pour les
différentes unités pastorales. D'ailleurs, les résidus de
culture (exploités en vaine pâture pendant la saison sèche
froide) et la biomasse ligneuse (sollicitée surtout en fin de saison
sèche chaude) (Le Houerou, 1980 ; Rivière, 1991 ; Lhoste et
al. 1993; Daget & Godron, 1995 ; Raimond, 1999 ; Kagoné,
2000 ; Petit, 2000a & 2000b, Yanra, 2004, etc.) qui sont reconnus comme
constituant des grands apports, n'ont pas été pris en compte dans
nos évaluations. Les charges de travail n'ont pas permis de faire
à temps ces évaluations qui sont en cours en ce moment. Ceci
appelle cependant deux remarques relatives à la couverture des besoins
des animaux à cette période. Premièrement, il se pose un
problème crucial de disponibilité et de qualité; en effet,
la biomasse estimée (en rappel, l'estimation se passe en septembre)
n'est plus forcément disponible pour le bétail en cette saison
sèche où certaines unités (UPP4 & UPP6 par exemples)
apparaissent totalement dénudées et le peu de paille qui y est
encore rencontrée est sans valeur et incapable d'assurer les besoins,
parfois de maintien, des animaux. Par ailleurs, comme on l'a vu en suivant les
troupeaux, certaines portions du territoire pastoral sont pratiquement
soustraites de l'exploitation animale en cette saison95 du fait de
l'absence ou de l'éloignement des points d'eau sans lesquels leur
valorisation reste impossible ou peu rentable (Lhoste & Milleville, 1986 ;
Guillaud, 1994 ; Lericollais & Faye, 1994 ; Bary, 1998). Pour toutes ces
raisons, on peut supposer que, malgré un bilan fourrager positif, les
animaux rencontrent des difficultés pour satisfaire leurs besoins
alimentaires en cette saison. Le bilan fourrager prend en effet peu compte de
la valeur fourragère et de la possibilité réelle de
valorisation du fourrage disponible.
En saison des pluies par contre, la plupart des animaux
transhumants venant du nord de la province et du Niger voisin repartent vers
leurs terroirs d'attache situés plus au nord, ce qui réduit
notablement les charges animales globales dans le terroir. Malgré cette
baisse spectaculaire des charges, c'est en cette saison où de nombreuses
unités paysagères sont peu
94 Ce taux est probablement dépassé de
nos jours, la base de donnés BDOT de l'IGB que nous avons
utilisée datant de 2002.
95 Nous sommes cependant réservé dans
cette affirmation car il ne s'agit que d'étude de cas dont les
résultats, on le sait, sont difficilement extrapolables car
l'inférence statistique n'est pas applicable (Mettrick, 1994).

239
ou pas accessibles au bétail (UPP5, UPP1 & UPP2)
à cause de risques de dégâts champêtres et/ou de
l'inondation permanente ou non en saison pluvieuse, que le bilan fourrager
révèle un déficit. Il convient cependant de
considérer avec un certain recul l'idée d'un déficit vu
que pour produire des données assez précises cette partie de
l'étude est prévue pour être conduite en deux temps. Dans
ce premier temps, parmi les unités considérées comme
totalement inaccessibles, certaines d'entre elles (UPP5 surtout ; unités
de concentration des cultures), disposent de portions (les interstices entre
champs) qui sont en réalité accessibles. A l'inverse, des
unités considérées comme totalement accessibles (UPP3,
UPP4 & UPP6) disposent des parcelles cultivées isolées en
leur sein. Notre évaluation, malgré son caractère
général à cette étape, alerte sur le risque encouru
par les pâturages du terroir sous des charges aussi
élevées. Une telle pression pendant la période de
développement de la strate herbacée peut être à
terme compromettante pour la production fourragère locale globale
(César, 1994 ; Daget & Godron, 1995 ; Kagoné, 2000). Ce
pessimisme peut cependant être tempéré parce que, en cette
période, la répartition de la pression animale sur les
pâturages est relativement homogène, la biomasse herbacée
est en effet de quantité satisfaisante presque partout et son
exploitation est rendue possible par une bonne distribution spatiale des points
d'eau comme l'a aussi relevé Kièma A. (2008) au Sahel
burkinabé.
Les ressources pastorales sont de statuts variés
aux yeux des éleveurs: notion de ressources clés.
Les catégories pastorales définies sur des
critères écologiques d'une part, d'après les
représentations des populations d'autre part diffèrent non
seulement par leurs caractéristiques fourragères mais aussi par
les contraintes auxquelles les troupeaux s'y trouvent confrontés. Nos
travaux indiquent que, pendant la saison pluvieuse, certaines unités
pastorales du terroir (les bas-fonds, les plaines inondables et les
mosaïques agroforestières) voient leurs ressources
fourragères peu ou pas exploitables à cause de l'humidité
ou des nombreux champs qui y sont disséminés et qui jouent le
rôle d'obstacles. Or, en contexte de paysage fragmenté,
l'existence des ressources fourragères n'est pas suffisante, leur
accès peut se révéler déterminant (Nori et
al. 2008) pour la couverture des besoins d'entretien et de production
des animaux. L'importance que va prendre une unité donnée de
l'espace-ressource (Daget & Godron, 1995 ; Barrière, 1996 ;
Barrière & Barrière, 1997) du terroir pour l'élevage
va donc dépendre de plusieurs facteurs notamment de leur qualité
intrinsèque (disponible fourrager) - qui combine biomasse et
qualité de celle-ci - de leur accessibilité et de la
période considérée. Toutes ces questions sont
intégrées par les éleveurs dans leur programme
d'affouragement des animaux et les catégories qu'ils distinguent en
tiennent compte. Ils considèrent certaines de ces catégories
comme capitales, ce qui correspond finalement à la notion de ressources
clés ou poches de ressources ou encore ressources stratégiques
développée par Hatfield & Davies (2006) ainsi que Nori et
al. (2008). Ces spécialistes les définissent comme
représentant toute portion de l'espace et des ressources pastorales
(fourrage et eau essentiellement) qu'elle héberge et dont la
soustraction à l'exploitation animale est susceptible de compromettre
l'existence du système dans son ensemble, en ce qu'il en constitue un
maillon essentiel à une période précise de l'année
(Hatfield & Davies, 2006). Par ailleurs, d'après ces auteurs,
l'importance de la ressource clé va au-delà de ses
capacités intrinsèques, son insertion dans le

paysage et la période de disponibilité de ses
ressources permettent de valoriser des étendues plus importantes de
pâturages, parfois de moindre qualité, qui, autrement, seraient
perdues pour le bétail.
Les éleveurs dont l'objectif principal est d'assurer la
sécurité alimentaire de leur bétail (Niamir-Fuller, 1999 ;
Kagoné, 2000) et qui sont, en conséquence, permanemment
préoccupés par la survie de leurs animaux au moment le plus
crucial de l'année (saison sèche chaude : Ku tontogu ou
Ceedu), n'ont pas tous la même définition de la ressource
clé (ou unité pastorale clé). Ainsi, du point de vue des
éleveurs peuls, la ressource clé est celle qui constitue un lieu
de séjour obligé des animaux en saison sèche chaude alors
que ce qualificatif est réservé par les agro éleveurs
gourmantchés à toute unité pastorale sur laquelle on
trouve l'eau en cette saison. Dans le fond, les deux représentations
expriment la même réalité car, comme nous l'avons
montré dans le paragraphe précédent, seuls les
pâturages offrant la possibilité d'abreuvement, soit sur place ou
à proximité, peuvent être réellement
valorisés en cette saison et ainsi aider à la survie du
bétail. Tous les groupes d'éleveurs enquêtés
considèrent qu'à Kotchari la ressource incontournable est
représentée par les bas-fonds (UPP1 : Ku bagu ou
Celol) où il est possible d'abreuver les troupeaux en cette
saison. Ceci rejoint les idées de Pratt & Gwynne (1997) et Hatfield
et Davies (2006) qui classent les bas-fonds parmi les ressources
stratégiques ou clés96, grâce auxquelles les
systèmes pastoraux arrivent à subsister dans les milieux arides
et subarides.
Nous avons vu que les bas-fonds sont l'une des meilleures
unités pastorales du terroir : ils produisent la plus forte biomasse
(4,78 tMS.ha-1) et offrent la possibilité en cette saison de
creuser des puisards pour l'abreuvement des animaux. Leur qualité
fourragère relativement faible n'est pas un handicap puisqu'à
cette période, le problème des éleveurs est surtout de
pouvoir donner suffisamment de fourrage à leurs animaux, la
qualité de ce fourrage devient alors pour eux secondaire. Ces
précieux bas-fonds sont cependant sujets à l'exploitation
agricole pluviale et de contresaison (culture maraîchère surtout)
et à l'arboriculture fruitière de mangue notamment. On n'observe
cependant pas de tendance à l'augmentation de ces usages à cause
de deux limitations imposées par le milieu: les eaux de surface sont
vite perdues après la saison pluvieuse et il n'existe pas d'ouvrages
hydrauliques adaptés (puits maraîchers par exemple).
Le niveau de concurrence entre les animaux et la culture reste
donc encore bas dans ces unités. Cependant, un certain nombre de projets
et d'ONG (ACRA97, PADAB II98, ADELE99,
PICOFA100, etc.) encouragent les activités
génératrices de revenus en finançant des microprojets
individuels ou collectifs. Il n'est donc pas exclu que sur le long terme,
l'utilisation des bas-fonds comme zone de repli en saison sèche chaude
par les animaux puisse susciter des conflits.
96 Une autre définition de la ressource
clé la désigne plutôt comme une partie de l'espace
pâturable produisant les meilleures et plus grandes quantités
d'aliments pour le bétail (Scholte & Brouwer, 2008). Cette acception
résiste cependant peu à l'analyse puisque, des unités
regorgeant de ressources fourragères de qualité peuvent se
révéler inaccessibles à un moment donné et
être alors inutiles pour le bétail.
97 Association de Cooperation Rurale en Afrique et
Amerique latine (ONG italienne)
98 Programme danois d'appui à l'agriculture au
Burkina, phase II.
99 Programme d'appui au développement local
à l'Est.
100 Programme d'investissement communautaire en fertilité
agricole.

241
Les pratiques des éleveurs reflètent des
ajustements opérés dans les exploitations et expriment des
stratégies adaptatives mises en oeuvre par eux.
Dans le terroir de Kotchari, nous avons reconnu 4
catégories et sous-catégories d'éleveurs qui cohabitent,
bien que globalement leurs troupeaux soient tous de taille importante et
croissante, ils adoptent des stratégies différentes dans leur
valorisation des ressources pastorales. Les tendances dégagées
montrent que lorsque la taille de son troupeau augmente, l'éleveur
résident (transhumant ou non) spécialise son troupeau en le
limitant à une espèce (le plus souvent l'espèce bovine) ou
à deux espèces (le plus souvent bovins et ovins) et qu'il le
remanie en favorisant les races bovines les plus adaptées localement
(Barbaji, Gurmaji : races rustiques peu exigeantes au plan alimentaire
et trypanotolérantes). On constate toutefois aussi un attachement
culturel à certaines races, pas nécessairement les mieux
adaptées comme la Jaliji et la Boboroji. En revanche,
on peut se demander pourquoi la race Gudali, pourtant bien
adaptée au contexte local, est faiblement représentée dans
les troupeaux, mais c'est probablement à cause de sa faible
prolificité (Boutrais, 2002). Les races bovines des troupeaux
transhumants non résidents sont également en voie de
diversification (Gurmaji + Jaliji) mais ceux à une seule race
bovine (Gurmaji) dominent. Il s'y déroule une sorte de
glissement du troupeau vers la race Gurmaji plus adaptée au
détriment de la race Jaliji d'origine.
Ces remaniements dans la structuration des troupeaux ne sont
pas anodins. Quand il constitue un troupeau, l'éleveur commence avec des
petits ruminants et des bovins (Barbaji) généralement
achetés (Gourmantchés) ou alors avec des bovins
hérités (Peuls résidents ou non : Gurmaji et
Jaliji respectivement). Lorsque le troupeau atteint une certaine
taille qui justifie qu'une plus grande attention lui soit portée,
l'éleveur dont le troupeau comprenait des caprins va s'orienter vers la
production d'espèces animales les plus prestigieuses et
présentant la valeur économique la meilleure: d'abord bovins et
ovins, puis seulement bovins. Parallèlement, à mesure qu'il se
professionnalise dans l'élevage, il acquiert une bonne connaissance des
races et opte autant que possible pour celles qui s'accommodent le plus des
contraintes du milieu, de jour en jour plus cruciales malgré la relative
déprise agricole actuellement observée à la faveur du
léger recul de la culture du coton (figure III-5, page 61). Ce recul
observé n'est sans doute d'ailleurs que passager, avec le prix incitatif
proposé en cette campagne agricole 2011 (245 FCFA le kg de coton graine,
prix jamais atteint auparavant), il est à craindre que, nonobstant le
renchérissement du coût des engrais, l'engouement pour cette
spéculation ne reparte de plus belle et, avec elle, l'extension de
l'occupation de l'espace.
Toutefois, dans leurs stratégies d'optimisation de
l'exploitation des ressources pastorales, les éleveurs n'agissent pas
seulement en remaniant la composition du troupeau en espèces et en
races. Ainsi, alors que le nombre de bergers s'accroît avec la taille du
troupeau, et malgré le grand besoin en main d'oeuvre de la partie
agricole de leurs exploitations, les éleveurs résidents,
notamment gourmantchés101, prennent de moins en moins le
risque de
101 L'utilisation de bergers extérieurs à la
famille était du reste peu commune chez les Peul notamment ceux qui ont
de grands effectifs.

confier leurs animaux à d'autres éleveurs ou
à des bouviers salariés. Tout au plus, lorsqu'ils tiennent
absolument à bénéficier du savoir faire de ces derniers ou
lorsque la main d'oeuvre familiale fait défaut102, ils ne
font que les associer à un membre de la famille. Les risques auxquels
ils disent s'exposer avec le confiage ou en employant uniquement des bergers
salariés sont beaucoup de dépenses et des pertes diverses aux
causes rarement élucidées. Les mêmes attitudes nourries par
les mêmes craintes sont rapportées par Raimond (1999) et
Kaboré (2010) qui se sont intéressés à la question
des rapports entre agriculteurs et pasteurs respectivement dans le bassin du
lac Tchad et dans la région de la réserve de faune de Pama Nord.
Kossoumna Liba'a (2009) rapporte par contre, à partir d'études
conduites dans deux villages peuls du groupe des Mbororo au nord-Cameroun, que
le rôle de berger pour le bétail des agriculteurs et autres
citadins (commerçants, fonctionnaires,...) joué par ceux-ci est
encore important, la moitié du bétail qui est sous leur gestion
est de ce type.
Par ailleurs, si l'allotement des troupeaux est une pratique
connue dans les élevages pastoraux, notamment peuls (Kagoné, 2000
; Riegel, 2002 ; Botoni, 2003 ; Kièma S., 2007), la division en trois
lots et certaines motivations avancées pour la justifier étaient
jusqu'alors inédites ou peu courantes. En effet, les agro
éleveurs gourmantchés qui ne dépassaient pas deux lots,
généralement hétérogènes, avaient pour seul
souci de ménager des effectifs gérables par les jeunes bergers.
Mais il faut s'interroger sur l'efficacité de ces nouvelles
façons de faire et sur leur incidence sur l'état des ressources.
Pour la plupart d'entre ces éleveurs, l'espèce bovine est
nouvelle dans leurs troupeaux et les techniques de sa conduite ne sont pas bien
maîtrisées, ils craignent donc de laisser les bergers avec de trop
nombreuses têtes. Ce groupe d'éleveurs sont les moins
exposés aux contraintes pour la gestion de leur cheptel car leurs
troupeaux sont de petite taille, ils disposent de résidus de culture
produite en grande quantité par de vastes champs et ils ont la
possibilité d'exploiter, à moindre risque, les interstices
inter-champs en saison de pluies. Ce groupe semble relativement peu
exposé aux contraintes d'accès aux ressources et ne semble pas
ressentir la nécessité de se préoccuper de leur
état, bien qu'il soit conscient des menaces qui pèsent sur
elles.
Les éleveurs peuls, résidents ou non, pratiquent
l'allotement depuis longtemps ; ils répartissent le plus souvent le
troupeau en deux lots : les, animaux bien portants d'une part, les animaux
allaitants et/ou mal en point d'autre part. Ils avancent diverses raisons pour
expliquer cette pratique. Ceux qui ne transhument pas et dépendent donc
davantage des ressources naturelles locales que les autres catégories,
cherchent à gérer au mieux leurs pâturages et ils pensent,
à juste titre (Breman & De Ridder, 1991 ; César, 1992 ;
Lhoste et al. 1993 ; Boutrais, 1997 ; Scoones, 1999 ; Nori, 2007 ;
Nori et al. 2008), qu'une répartition des charges animales sur
divers endroits du terroir est moins dommageable pour l'état des
ressources qu'une plus grande concentration en un lieu donné. Beaucoup
d'éleveurs peuls transhumants résidents ou non, qui ont les plus
grands effectifs et qui sont parfois amenés à former
jusqu'à trois lots (lot 1 : troupeau d'allaitants ou de malades ; lot 2
: troupeau de veaux et lot 3 : troupeau de bien portants destinés
à transhumer) le font pour faciliter (gain de temps)
102 La pratique du confiage chez les Gourmantché et le
fait de confier les animaux à des Peul et non à d'autres
Gourmantché n'ont pas toujours été motivés par le
seul souci de bénéficier du savoir-faire du berger peul.
Kaboré (2010) montre que la pratique a, par le passé,
relevé d'une stratégie de "camouflage" du bétail par les
Gourmantché, ceux-ci cherchant à entourer d'une certaine
discrétion l'existence de leur cheptel.

243
et sécuriser (minimisation des pertes par vol,
prédation, saisies, etc.) leur déplacement qui se
fait
très souvent vers un environnement incertain, voire hostile (Benoit,
1979 ; Landais, 1990).
La complémentation, réservée en
priorité aux animaux mal en point ou allaitant, est surtout
pratiquée dans les élevages les moins mobiles, plus
intégrés à l'agriculture et ayant par ailleurs les
effectifs les moins importants. Les éleveurs transhumants, dont les
effectifs animaux sont parfois impressionnants, n'ont pas (les non
résidents)103 ou ont insuffisamment (les locaux) de
résidus culturaux en réserve ; ils sont donc contraints d'acheter
les aliments de complémentation. Mais la plupart d'entre eux semblent ne
pas complémenter leurs animaux ou, en tout les cas, ne peuvent en
distribuer suffisamment, ce qui d'ailleurs les oblige à émigrer.
On a pu voir, en effet, par nos enquêtes (voir figure V-15b, page 145)
que, pour ce qui concerne les éleveurs résidents, il existe une
tendance à l'accroissement du phénomène migratoire. On
observe, en effet, que les éleveurs locaux élargissent leur
territoire pastoral en visitant plus que par le passé les terroirs
voisins (les résidents gourmantchés) ou lointains, y compris les
réserves (résidents transhumants). Cela est totalement en accord
avec le schéma théorique de Boutrais (1983), Santoir (1999),
Dugué et al. (2004) qui entrevoient que dans les cas qui
correspondent à la situation de Kotchari (cheptel croissant,
densité humaine de 56 habitants/km2, taux d'occupation des
sols de 56%, état moyen des ressources soumises à de fortes
pressions d'exploitation et localement en dégradation
intermédiaire), les éleveurs du terroir sont amenés
à fréquenter, avec leurs troupeaux, des pâturages plus
lointains (transhumance) ou à déstocker leurs effectifs.
Toutefois, les départs ne sont pas massifs, ceci pouvant s'expliquer par
les contraintes que les éleveurs rencontrent dans ces lieux de
délocalisation temporaire comme l'a aussi observé Raimond (1999)
dans le bassin du lac Tchad. L'auteur a noté que
l'insécurité et les tracasseries administratives que vivent les
éleveurs au cours de leur déplacement à la
frontière tchado-camerounaise, de même que les pertes et ventes de
bétail par les bouviers, n'encouragent plus les propriétaires
à laisser migrer leurs animaux. Ce sont à peu près les
mêmes arguments qui sont avancés par les éleveurs de
Kotchari pour expliquer que malgré la forte pression localement
vécue, ils soient hésitants à partir massivement.
Les éleveurs ont une bonne connaissance de leurs
milieux et disposent de savoirs techniques.
Les éleveurs définissent et appréhendent
leurs milieux selon les usages qu'ils peuvent en faire au cours de
l'année suivant un calendrier pastoral assez illustratif des contraintes
et de l'abondance du moment. Les critères topographiques et
pédologiques en lien avec le type d'usage sont utilisés pour
définir des milieux qu'ils peuvent par ailleurs caractériser par
les espèces herbacées et surtout ligneuses qui y sont
présentes. Ces milieux ou unités paysagères «
participatives » définies selon leurs critères ont à
leurs yeux une valeur variable suivant les saisons. Cinq paramètres
(disponibilité en eau, disponibilité de fourrage en
quantité, disponibilité de fourrage de qualité, milieu
praticable et milieu n'exposant pas le bétail à un
103 Ce sont pour la plupart des personnes qui s'adonnent aussi
à divers degrés à l'agriculture. Ils ont donc parfois des
stocks en résidus culturaux dans leur terroir d'attache mais qui ne peut
leur servir ici. Ces stocks sont d'ailleurs distribués aux animaux
restés sur place.

quelconque risque) sont utilisés par les
éleveurs pour les évaluer. Ainsi, alors qu'en saison pluvieuse
l'éleveur craint pour la sécurité de son troupeau
(recherche de site praticable et/ou éloigné de champs de culture)
et recherche la bonne herbe, en saison sèche, face à la
pénurie en eau et en fourrage, c'est plutôt la
disponibilité de ces deux ressources, en particulier l'eau, qui est mise
en avant. Par ailleurs, en cette saison, les éleveurs peuls aux grands
troupeaux qui pâturent au ras des aires fauniques et qui sont plus
enclins à aller vers les sites nouvellement accessibles (par exemple les
plaines inondables après assèchement de l'eau), restent attentifs
aux risques (saisie des animaux en cas d'empiètement sur l'aire
protégée, embourbement dans les milieux encore humides).
Forts de ces critères, les éleveurs ont une
vision claire des différentes unités qu'il est intéressant
de fréquenter au fil des saisons. Classiquement leur
représentation de la chaîne de pâturage saisonnière
part des milieux secs en hauteur en saison pluvieuse, pour aboutir aux sites de
bas de toposéquence en saison sèche, au moment où ceux-ci
se sont asséchés et offrent des conditions relativement
meilleures.
Ce modèle d'action, à peu près identique
chez les différents groupes d'éleveurs et mis en évidence
ailleurs par différents auteurs (Kagoné, 2000 ; Diallo, 2006 ;
Vall & Diallo, 2009 ; Dongmo, 2009), se trouve modifié en fonction
des réalités propres aux 4 types d'éleveurs que nous avons
suivi. Aussi, pour une même saison, en tenant compte de contraintes
particulières (campement entouré de champs et plus grande
difficulté d'accès aux sites de hauteur par exemple pour Trp ;
unités brûlées sans possibilité de repousses
attrayantes, unités proches des champs pour l'ensemble des quatre
troupeaux), les animaux sont conduits sur des itinéraires qui
concrétisent l'enchaînement des choix les plus judicieux
possibles. En pratique, les éleveurs opèrent des choix qui leur
permettent de minimiser les dépenses énergétiques qu'ils
engagent à la recherche de la ressource (théorie de
l'optimisation des gains). Pendant notre période d'étude les
éleveurs que nous avons suivis, ont tous exploité un seul secteur
du terroir, généralement proche de leurs campements, ce qui
indique qu'en réalité peu d'alternatives s'offrent à eux.
En saison pluvieuse les champs et le risque d'embourbement constituent des
obstacles presque infranchissables qui limitent les déplacements d'un
secteur vers un autre secteur du terroir. En saison sèche, la
rentabilité (différence entre gain et dépense en
énergie) de longs déplacements apparait incertaine, surtout que
pour les deux troupeaux sédentaires que nous avons suivi (TrpC1-2G et
TrpC1- 2P), il existe une source d'eau intarissable à proximité
de leurs campements. Par ailleurs, l'éleveur gourmantché, qui
dispose d'un stock conséquent en résidus de culture et en son de
céréales, n'hésite pas à en distribuer à son
troupeau (TrpC1-2G) qu'il maintient à proximité de sa concession
au Ku tontogu (saison sèche chaude).
Il y a une pâture inavouée dans les aires
protégées voisines mais son ampleur reste mal connue.
Alors que les troupeaux résidents élargissent
leur aire de pâturage faute de pouvoir se satisfaire localement, on
observe paradoxalement un accroissement soutenu du nombre de transhumants qui
arrivent dans le terroir. En effet, bien peu de transhumants qui y sont
rencontrés de nos jours y venaient déjà il ya dix ou vingt
ans (figure V-15c, page 145). Par

245
ailleurs, si l'on peut reconnaître que les ressources
fourragères locales peuvent être meilleures que celles de bien des
contrées d'origine de ces transhumants, on peut s'interroger sur la
disponibilité réelle de cette biomasse pendant la période
de transhumance. Indiscutablement, les explications à ces flux continus
de troupeaux se trouvent en grande partie ailleurs : le terroir, par sa
position géographique se trouve en fin de transhumance. On peut, par
ailleurs, affirmer que sa proximité avec les réserves totale
(parc W) et partielle (concession de chasse Kourtiagou) de faune aux
énormes ressources fourragères encore meilleures que celles des
parties accessibles du terroir constitue l'un des facteurs de l'attraction
observée. Beaucoup d'indices indiquent en effet que les éleveurs
locaux et transhumants exploitent les aires protégées de
manière plus ou moins régulière, certains semblent
d'ailleurs ne pas pouvoir s'en passer. En effet, des enquêtes conduites
dans des terroirs lointains104 de nos réserves (Kpoda, 2010)
ou dans d'autres régions (Kièma S. 2007 ; Kaboré, 2010)
ont montré que les éleveurs n'étaient pas
indifférents aux ressources qui y sont présentes, bon nombre
d'entre eux transhument en fait pour pouvoir les exploiter. Le travail de Kpoda
(2010) en particulier montre que la grande majorité (plus de 80%) des
transhumants nationaux ou nigériens rencontrés à Botou
(commune la plus au nord de la province) disent transhumer vers le terroir de
Kotchari. Un autre indice est que les campements des transhumants qui viennent
à Kotchari se positionnent généralement au ras des
réserves et qu'ils sont généralement mobiles, ce qui
permet de brouiller les pistes. Kaboré (2010) a décelé
autour de la réserve de Pama Nord une toute autre stratégie que
mettent en oeuvre, pour la même finalité, les éleveurs
résidents (les Gurmaabe) qui vivent une situation d'exclusion
de la part des Gourmantchés dans les terroirs villageois. D'après
cet auteur, cette stratégie consiste à camoufler leur usage des
réserves, en positionnant les campements à une distance
suffisamment éloignée pour ne pas éveiller les
soupçons mais suffisamment près pour en permettre l'"usage
à distance". Par ailleurs, lors des discussions de groupe, certains des
indicateurs de fréquentation des milieux (cure salée, champs
maudits) qui sont cités font référence à des
milieux absents de l'espace du terroir, mais bien présents dans le parc
W (Kpoda, 2010). Il est symptomatique qu'ils aient été
cités dans un premier temps avant d'être retirés de la
liste sans explication. Enfin, il peut être surprenant de constater,
à la lumière de nos données, que seuls les transhumants
locaux disent fréquenter actuellement les aires protégées
(figure V-16b, page 146) alors que par le passé, ceux qui viennent des
contrées plus au nord fréquentaient ces entités beaucoup
plus que tous les autres (figure V-16c, page 146). Cela semble contredire
d'ailleurs les constats faits par Kaboré (2010) qui indique que les
éleveurs peuls, du fait de la crise d'intégration dont ils sont
les premières victimes, sont plus fortement dépendants des aires
protégées. On peut logiquement imaginer que les transhumants
allochtones, qui sont plus victimes que leurs homologues résidents, sont
plus exposés à l'envie de pénétrer dans les aires
protégées.
Il est donc évident que les réserves qui
environnent le terroir de Kotchari sont exploitées par les
éleveurs présents à leur périphérie.
L'intensité de cette pression d'exploitation est cependant difficile
à établir à cause de la loi de silence qui est de rigueur
sur le sujet. Les éleveurs semblent s'être passé le message
et évoquer le sujet avec l'un d'entre
104 Quand les éleveurs sont loin des aires
protégées ils se montrent plus ouverts aux discussions à
ce sujet. A ces distances, les craintes se dissipent car le risque d'être
étiqueté paraît improbable.

eux peut suffire à mettre fin à une causerie bien
engagée. De toute évidence, la question risque de rester encore
non élucidée pendant longtemps.
7.2. Conclusion générale
7.2.1. Les dynamiques biologiques et
socio-économiques mises en évidence
La recherche a mis en évidence deux dynamiques qui se
déroulent de manière concomitante et qui sont fortement
dépendantes : des dynamiques biologiques au niveau de la ressource et
des dynamiques socio-économiques au niveau des éleveurs et de
leurs activités productives.
Les dynamiques biologiques
La caractérisation des écosystèmes
pastoraux du terroir de Kotchari a montré une forte diversification de
leur flore herbacée (richesse spécifique et
équitabilité élevées par rapport aux unités
de l'aire protégée voisine) en particulier dans les milieux les
plus anthropisés (plateaux plus ou moins cuirassés aux sols
superficiels ou peu profonds, plateaux et plaines cultivées à
sols plus ou moins profonds). Celle-ci s'accompagne d'une extinction locale
d'espèces, de la banalisation de la flore en particulier sur les
plateaux dans lesquels des espèces exotiques à large distribution
(phorbes : légumineuses et herbes diverses) prennent de l'importance
(voir aussi Sawadogo et al. sous presse). L'ensemble des
unités connaissent un envahissement arbustif ce qui préfigure un
début d'embuissonnement (fortes proportions d'arbustes et sous-arbustes
dans la strate ligneuse). Les causes à ces tendances n'ont pu être
clairement établies, on sait seulement que divers facteurs directs ou
indirects de fonctionnement et/ou de perturbation (le feu, les activités
agricoles, l'exploitation animale et les prélèvements
domestiques) interviennent parfois concomitamment, ils voient leur
intensité se renforcer avec la pression démographique et la
transhumance. Les travaux de Caillault (2009) montrent que, dans notre zone,
les feux de brousse sont généralement irréguliers et
peuvent être précoces (en particulier sur les milieux plus secs)
ou tardifs (dans les sites humides).
Les savanes parcs ou mosaïque agroforestière
(UPP5) sont les zones de forte concentration des parcelles cultivées,
les paysages y sont donc fragmentés et les pâturages sont peu
accessibles en saison des pluies alors que les feux de brousse qui y
surviennent ont des effets limités à cause de cette
fragmentation. Sur ces unités, il y a donc une prééminence
des activités agricoles dans l'évolution des milieux, en
témoigne leur colonisation par des espèces messicoles (adventices
annuelles) qui ont pris place au détriment des graminées
pérennes endémiques des savanes et caractéristiques de
milieux stables (Schnell, 1971 ; Daget & Godron, 1995 ; César, 2005)
et qui sont par ailleurs les plus recherchées par le bétail
(Boudet, 1978 ; César, 1994). A côté de ce
phénomène d'extinction locale d'espèces (Kièma S.,
2007),
les défrichements agricoles ont fini par éliminer
pratiquement la strate ligneuse arborée, d'oüles
très faibles densités et recouvrements observés alors que
le feu y favorise des formations buissonnantes dans lesquelles dominent des
arbustes et sous-arbustes.

247
Les glacis (UPP3), les divers plateaux et plaines (UPP4), de
même que les buttes et cuirasses (UPP6)105 dans une moindre
mesure, sont des jachères de divers âges
généralement jeunes à moyens. Ce sont les sites les plus
pâturés en saison humide et ils y sévissent aussi les feux
de brousse dont l'importance est fonction de la biomasse herbacée
(faible dans UPP6) et de son organisation spatiale, c'est-à-dire son
degré de fragmentation (la couverture herbacée est interrompue
par les champs plus nombreux sur UPP3 et UPP4 ou par les ruptures naturelles
que représentent les plages de cuirasse ou d'affleurement
rencontrées sur UPP6). Sur ces différentes unités donc,
l'évolution progressive de la végétation qui
succède à la mise en culture (succession post culturale) (Boudet,
1978 ; César, 1991 ; Zoungrana, 1993 ; Daget & Godron, 1995 ; Hien,
1996 ; Fournier et al. 2001 ; Botoni-Liehoun et al. 2006) est
influencée par l'action du feu et de la pâture qui sont parfois
complémentaires sans que l'on sache laquelle est
prépondérante (Botoni, 2003 ; Kièma S. 2007). Cette action
sur le jeu normal de la succession végétale qui, dans les
jachères d'âge avancé, permet d'ordinaire le retour des
graminées pérennes initiales, par suite de remontée
biologique (Daget & Godron, 1995) seulement possible sur les sols profonds
et humides, aboutit à des écosystèmes particuliers. En
effet, la pâture est un facteur de déséquilibre entre les
strates ligneuse et herbacée (Boudet, 1978), elle réduit la
couverture herbacée amoindrissant ainsi la force des feux de brousse, ce
qui favorise l'envahissement en ligneux de parcours (d'où la plus forte
densité ligneuse de ces unités) qui vivent moins la concurrence
des herbacées et qui, par ailleurs prennent un port arbustif, ce qui
explique le faible recouvrement ligneux; il semble donc s'y dérouler un
phénomène d'embuissonnement. Par ailleurs, les modifications
induites sur le substrat par le piétinement et par l'apport de
nutriments par les fèces, l'importation des graines principalement
d'adventices et d'espèces ligneuses du groupe des légumineuses
par endozoochorie, épizoochorie (Boudet, 1978 ; Daget & Godron, 1995
; Devineau, 1999 ; Kièma S., 2007) ou par les fèces et le
prélèvement orienté sur les graminées vivaces (Akpo
et al. 1995) vont entraîner une modification de la flore
notamment herbacée et une augmentation de la diversité
végétale (richesse, équitabilité, diversité
béta et même diversité gamma). L'importance accrue des
espèces annuelles et des espèces exotiques,
généralement des phorbes (légumineuses et herbes diverses)
indicatrices de dégradation pastorale et la forte
équitabilité (espèces présentes en des proportions
voisines) témoignent de l'instabilité de ces milieux. Sur UPP6,
où le sol est squelettique, l'action du bétail est sans effet
majeur sur le sol, elle se réduit essentiellement à l'apport
d'espèces exotiques (Kièma S., 2007).
Dans les écosystèmes sur sols profonds humides
moins anthropisés (UPP1 et UPP2) le milieu est relativement moins
perturbé, ces unités sont en effet moins diversifiées
(richesses spécifiques moindres, léger déséquilibre
dans la proportion des espèces herbacées) et les espèces
savanicoles endémiques (les graminées vivaces) y sont plus
fortement représentées.
La dégradation des écosystèmes du terroir
ne fait l'objet d'aucun doute, mais elle reste limitée et l'on peut
espérer que la capacité de résilience106 de
ceux-ci (Daget & Godron, 1995 ; Boutrais, 1996) reste
préservée. En d'autres termes on peut supposer que leur
105 Nous sommes réservé lorsque nous
considérons les buttes et cuirasses comme ayant été
cultivées par le passé comme c'est le cas maintenant à
cause de la pression foncière sur les terres arables et mêmes les
terres marginales incultes du terroir.
106 La résilience est la capacité d'un
système à se relever après une phase de
déstructuration due à une pression d'exploitation forte ou
à une variation importante des facteurs climatiques, notamment la
pluviométrie.

reconstitution serait possible si les pressions anthropiques
actuelles venaient à être allégées et la
possibilité d'accès à toutes les unités accrue par
la levée des obstacles qui entravent cela (par exemples : une
réorganisation de l'occupation de l'espace, abandon des pratiques
agricoles sur les pistes d'accès et dans les zones de pâture
traditionnelles). Ce, d'autant plus que dans ce contexte de fragmentation
importante du paysage, la fréquentation des milieux reste difficile
à prévoir (Nori et al. 2008) et des risques de surcharge
(et donc de dégradation) localisée, notamment des portions les
plus accessibles, sont à craindre.
Dans les aires protégées voisines, où
seul le feu, qui y est généralement précoce,
régulier et intense (Caillault, 2009), est important (perturbation
moindre qu'en périphérie), la végétation
herbacée est relativement stable (faibles diversité,
équitabilité et hétérogénéité)
avec une dominance marquée de quelques herbacées
hémicryptophytes reconnues comme productrices d'une biomasse importante
et de qualité. Dans ces unités, en particulier les savanes de
bas-fonds à pérennes (UPw3) où l'herbe est haute et dense,
les feux, malgré leur précocité (la mise à feu a
lieu en octobre), sont violents et limitent ainsi la densification en ligneux
du milieu (Monnier, 1981 César, 1991 & 1994 ; Bruzon, 1995 ; Western
& Maitumo, 2004 ; Bond & Keeley ; 2005 ; Lavorel et al. 2007).
Dans ces écosystèmes dont la composante ligneuse est
dominée par la strate arborée, le recouvrement est meilleur qu'en
périphérie malgré une densité plus faible.
L'absence notable de pâture maintient une importante biomasse
herbacée qui, par le jeu de la concurrence, arrive à limiter
l'envahissement du milieu par les ligneux.
Les dynamiques socio-économiques
Les éleveurs sont soucieux d'assurer l'adaptation de
leurs systèmes d'élevage dans un environnement aussi changeant.
Pour assurer la survie du bétail et donc la leur, ils mettent en place
des stratégies et adoptent des pratiques visant à mieux tirer
profit des opportunités offertes par leur environnement
socio-économique et écologique. Face donc aux mutations
économiques (pluriactivité des ménages, essor de la
culture du coton, etc.), sociales (pression démographique, recul de la
réciprocité et des complémentarités,
rivalités pour l'accès aux ressources pastorales, etc.) et
environnementales (détérioration qualitative, réduction et
obstruction des pâturages) les différents groupes
d'éleveurs émettent des réponses
révélatrices des représentations qu'ils ont de leur milieu
et témoins de la manière dont chacun vit ces changements. Ainsi,
en réponse à ces dynamiques biologiques qu'ils contribuent
à imprimer107, les 4 groupes et sous-groupes
d'éleveurs dont les élevages gardent le caractère de
banque traditionnelle (grande taille, éleveur naisseur, faible
utilisation d'intrants, etc.) (Daget & Godron, 1995 ; Boutrais, 1994 &
1996), changent, suivant l'accroissement de leurs effectifs, la configuration
des troupeaux. Ils sélectionnent les espèces les plus
économiques (ovins et surtout bovins) et diversifient les races bovines
(cette diversification est plus lente chez les transhumants non
résidents) mais seulement avec les races les plus adaptées au
contexte local. Les éleveurs restent toutefois attachés à
leurs races traditionnelles (Barbaji pour les Gourmantchés,
Gurmaji pour les éleveurs Peuls résidents ou
transhumants nationaux
107 Nous n'oublions cependant pas le rôle majeur que
joue la péjoration (chute de la pluviométrie, fortes
variabilités de la pluviosité) en contexte aride ou semi-aride
comme le nôtre (Breman et De Ridder, 19991 ; Scoones, 1995 &
1999),

249
et Jaliji pour les transhumants nigériens)
même lorsqu'elles ne sont plus adaptées (la Jaliji ne
supporte pas la longue marche, elle est par ailleurs peu
trypanotolérante, mais elle n'est présente dans le terroir qu'en
saison sèche pendant laquelle le risque est moindre). Aussi, les races
Boboroji et Kiwali restent présentes dans les
élevages peuls bien qu'étant très peu adaptées au
contexte local caractérisé par la basse des productions
fourragères. Par ailleurs, l'insécurité physique (vol,
prédation, abattage, embourbement, etc.), sociale (risque de conflits
liés aux dégâts champêtres ; exclusion par les
agroéleveurs Gourmantchés) et alimentaire (parcours pauvres,
rares et à exploitation très concurrentielle) du bétail
s'agrandissant, les éleveurs, en particulier les peuls, renforcent la
surveillance par subdivision, plus que par le passé, de leurs troupeaux
en deux voire trois lots et en augmentant le nombre de bergers issus, par
ailleurs, de leurs ménages ou comprennent un membre du ménage. La
satisfaction des besoins alimentaires est assurée par la
complémentation avec la paille naturelle et/ou cultivée interne
(résidus produits dans l'exploitation) ou externe (achat). L'achat est
caractéristique des élevages peuls résidents, mais aussi
allochtones dans une certaine mesure, il peut concerner les concentrés
(sons, graines, tourteaux divers). Malgré tout, les troupeaux n'arrivent
plus à se satisfaire localement et l'espace pâturé est de
plus en plus ample : les terroirs voisins sont de plus en plus visités
mais en deçà de ce à quoi on devait s'attendre, en
réalité les éleveurs surtout peuls exploitent les
réserves qui semblent participer à réguler localement les
charges à la fois en saison sèche qu'en saison pluvieuse. La
fréquentation plus que probable des réserves en saison des
pluies, bien que non révélée est plausible malgré
les risques (humidité et glossines) que cela comporte. Elle peut jouer
un rôle décisif dans l'évolution des pâturages
périphériques qui, en cette saison, présentent un bilan
fourrager "négatif", ce qui est porteur de menaces sur leurs
capacités productives.
En définitive, la taille du troupeau apparait comme le
facteur qui détermine toute les stratégies de
l'éleveur.
Le modèle d'action développé par les
éleveurs pour valoriser les pâturages et qui traduit leur bonne
connaissance des milieux (potentiel et dynamiques qui y ont cours) et la
rationalité dans les prises de décision (en réponse
à la triple question : quel pâturage ? à quel moment ? pour
quel gain ?), se trouve, à l'épreuve de la réalité,
confronté à des obstacles divers (obstruction de couloirs,
inaccès de pâturage, assèchement des points d'eau, etc.)
dont l'acuité est fonction de la taille du troupeau. Les aspects
fonciers sont aussi non négligeables, les transhumants non
résidents par exemple ont moins accès aux unités
cultivées pour y effectuer la vaine pâture et, si en
général les distances parcourues sont plus grandes en saison
sèche, elles le sont davantage pour ce type d'éleveurs à
l'encontre desquels une grande hostilité est
développée.
7.2.2. Des pistes d'actions pour des systèmes
d'élevage durables
Au terme de cette recherche sur les ressources pastorales, les
pratiques pastorales et les stratégies qui les sous-tendent et en
considérant le contexte tel qu'il apparaît que peut-on dire des
conditions qui pourraient permettre un élevage durable ?
L'élevage extensif (sédentaire ou mobile) est-il viable et
durable dans le terroir de Kotchari ? Si non quels sont les ajustements
nécessaires qui doivent être opérées en son sein et
dans le système global étudié ? Quelle

pourraient être les conséquences de ces
ajustements sur le devenir des écosystèmes du terroir et des
réserves voisines ? En d'autres termes, il s'agit de répondre
à la question : « comment garantir la mobilité
d'élevage tout en l'adaptant au contexte local ? »
Une gestion adaptative qui prend appui sur la
maîtrise des effectifs
En se basant sur l'état actuel des pâturages du
terroir, on peut dire que certaines des pratiques mises en oeuvre par les
éleveurs et les autres acteurs sont durables. On s'attendait, en effet,
face aux effectifs de bétail observés sur le terroir, à
des niveaux de dégradation des unités pastorales, notamment
celles les plus anthropisées (UPP3, UPP4, UPP5 et UPP6), plus
élevés. Toutefois, la situation d'ensemble ne pourrait être
durable (c'est-à-dire efficace sur le long terme) que si les effectifs
de bétail présent sur le terroir étaient
maîtrisés, comme l'explique bien César (1994). En effet, le
problème central de la gestion des écosystèmes
pâturés tient à l'équilibre entre les pressions
d'exploitation et l'état des ressources. Ce problème a longtemps
été réduit à celui d'une gestion des troupeaux et
des espaces qui se fondait sur la notion de capacité de charge.
Cependant la pertinence d'une telle approche est maintenant fortement
contestée, surtout en contexte de non-équilibre (Ellis &
Swift, 1988 ; Grouzis, 1988 ; Behnke & Scoones 1992 ; Magda et al.
2001 ; Hatfield & Davies, 2006 ; Wane, 2006 ; Allen et al. 2011)
comme c'est le cas ici108. Sans revenir sur la controverse qui
entoure une telle notion (voir l'encadré II-1, page 34 pour plus de
précision), rappelons simplement que dans les environnements instables
(César, 1992 & 1994 ; Hatfield & Davies, 2006) la
quantité de matière sèche produite varie
considérablement dans l'espace et le temps avec le régime des
pluies ; il devient alors risqué et trompeur (Daget & Godron, 1995)
de baser toute approche de gestion sur celle-ci. Nos résultats montrent,
en effet, que les unités pastorales dans le terroir n'ont pas les
mêmes potentialités et ne sont pas au même niveau de
dégradation et on ne voit pas comment en système ouvert on peut
proposer des mesures ciblées de gestion de charge.
Dans le contexte qui est le notre, marqué par une forte
variation dans la distribution spatio-temporelle des ressources et une non
appropriation de celles-ci, la gestion des écosystèmes
pâturés peut se faire selon l'approche de gestion adaptative
(CEMAGREF, 2008) ; celle-ci, mise en oeuvre dans le cadre de la gestion
intégrée de populations d'ongulés sauvages et de leur
habitat, s'appuie sur le suivi d'indicateurs biométriques (qui
renseignent sur l'état corporel des animaux composant les troupeaux) et
biologiques (qui renseignent sur l'évolution des
écosystèmes). En effet, avec le risque de dégradation des
ressources fourragères, la gestion d'ongulés sauvages circulant
librement dans des espaces limités présente certains points
communs avec celle des troupeaux dans les systèmes d'élevage
pastoral. Pour cette faune, la solution aujourd'hui adoptée en Europe
est le déstockage du surplus d'animaux dès que des signaux
négatifs viennent à être observés quant à
l'état d'équilibre des milieux. C'est pourquoi, un
préalable serait de veiller à la maîtrise des effectifs,
à partir de là les indicateurs deviennent de bons outils de
gestion. Parallèlement à
108 Selon la carte phytogéographique de Fontes &
Guinko (1995), notre terroir se trouve dans le district de la Pendjari (zone
sud soudanienne). Cependant, avec la descente des isohyètes et le
régime climatique en cours (voir chapitre 3 ; paragraphe 3.2.1 ; page
45), nous sommes plutôt en contexte de semi-aridité et les
écosystèmes évoluent selon un processus de
non-équilibre.

251
cette gestion adaptative, des ajustements à divers
niveaux des systèmes d'élevage sont nécessaires, ils
pourraient s'inspirer des pratiques locales dont certaines font
déjà la preuve de leur durabilité. Il convient donc d'en
faire l'inventaire pour déterminer celles qui sont à encourager
parce que compatibles avec les dynamiques biologiques en cours.
Les pratiques locales comme porte d'entrée pour
toute intervention
Les troupeaux du terroir sont de grande taille et cette
tendance se renforce. Or, on sait que les grands effectifs sont dommageables
pour les animaux composant les troupeaux en même temps que pour les
écosystèmes pâturés (Daget & Godron, 1995 ;
Boutrais, 1996). En effet, quand la concurrence est ainsi accrue, les animaux
modifient leur comportement alimentaire, ils ne sélectionnent plus les
herbes, ce qui se ressent dans la qualité des rations
prélevées et donc des performances surtout qu'en plus, en de
pareilles circonstances, les distances parcourues sont plus importantes. Par
ailleurs, aucune herbe n'est préservée et les équilibres
sont plus vite rompus.
Les différents groupes d'éleveurs
étudiés ont montré une tendance à l'allotement de
leurs troupeaux, à l'homogénéisation des espèces
(sauf chez les Gourmantchés) vers celles les plus économiques
(bovins et ovins ou bovins seuls) et à la sélection de races
rustiques comme la Barbaji et la Gurmaji qui tendent à
dominer dans les troupeaux locaux. Si cela est à encourager du point de
vue du bien être de l'animal qui arrive à se satisfaire sur de
courtes distances (gains optimisés), il n'en va pas forcément de
même pour l'état biologique de la ressource. En effet, les animaux
peu exigeants ont un comportement alimentaire peu sélectif et
prélèvent presqu'entièrement la ressource
fourragère des milieux pâturés (la pâture est alors
dite rasante ou intégrale) ; l'ampleur des dommages sera, en
conséquence, fonction du niveau de la pression (intensité et
fréquence de fréquentation) de pâture.
La question de l'impact de la composition du troupeau sur les
écosystèmes pâturés devient plus complexe lorsque
celui-ci comprend plusieurs espèces et/ou plusieurs races d'une
même espèce. En effet, alors que, d'après Boutrais (1996),
les troupeaux hétérogènes sont les plus dangereux pour
l'état des ressources en ce qu'ils exploitent une large gamme de
fourrages situés à différentes hauteurs (herbes, feuilles
et fruits d'arbres), César (1992), Lhoste (2004) et Louppe et
al. (2000) repris par Kièma S. (2007) avancent le contraire.
D'après Lhoste (2004), en situation de charge raisonnable, la
diversité animale permet de mieux valoriser les
écosystèmes pastoraux qui, eux-mêmes, restent assez divers
et moins déstabilisés. Si donc, les effectifs sont
maitrisés, il faut accompagner les éleveurs dans la
diversification spécifique de leurs troupeaux et dans l'allotement qui
permet de repartir les charges sur divers milieux et d'éviter les
surcharges localisées. Ce qui permet de prévenir ou de retarder
la dégradation des ressources et l'érosion de la
biodiversité. Les races bovines exigeantes (grandes consommatrices ou
sélectives) (Boboroji et Kiwali)
accélèrent en effet ces phénomènes, mais leur
disparition progressive dans les troupeaux présents dans notre terroir
est un bon signe109 qu'il faut encourager. Pour permettre, par
ailleurs, une diversité dans les choix des races, il convient d'aider
à la promotion de la race Gudali qui, bien que moins
prolifique, est aussi
109 Le bon signe est à voir du côté strict
de la ressource fourragère qui verrait ainsi sa dégradation
amoindrie. Mais si on se place du côté de la biodiversité
animale et de l'attachement que les éleveurs ont avec ces races, il est
clair leur disparition refléterait plutôt une perte.

adaptée. En outre, dans un contexte de cloisonnement et
de réduction des pâturages, l'allotement est une pratique efficace
(Djenontin et al. 2009), surtout lorsque que l'éleveur est mu
par le souci de pouvoir gérer les risques de dégâts
champêtres (meilleure maîtrise de son troupeau) et/ou pouvoir
exploiter tous les recoins du terroir ; ce que ne permet pas un troupeau
à grand effectif.
Le calendrier pastoral des éleveurs de même que
la chaîne de pâturage sont la preuve que ceux-ci tiennent compte
des autres acteurs (éloignement des champs et des
réserves110) dans leurs pratiques d'affouragement des
troupeaux. Toute intervention devra s'en inspirer tout en aidant à lever
les contraintes qui s'opposent à leur mise en oeuvre. Comme on l'a vu,
d'assez vastes portions du terroir ne pouvaient être exploitées en
saison sèche faute de points d'eau fonctionnels pour l'usage pastoral ou
mixte. On est alors tenté comme Binot et al. (2006) de proposer
de réaliser des points d'eau qui seraient ainsi des pôles
d'attraction (Dumont et al. 2001), mais des risques existent ; cela
peut, en effet, drainer les troupeaux des terroirs voisins et provoquer des
dégradations localisées (Touré, 1997 ; Baroin, 2003)
autour de ces points d'eau entrainant ainsi des effets contraires à ceux
attendus.
Les éleveurs résidents explorent un espace de
plus en plus vaste (transhumance, délocalisation temporaire sur les
terroirs voisins) mais le phénomène est d'ampleur moindre eut
égard à la saturation foncière locale. Ils semblent en
fait partagés entre deux options : transhumer ou se sédentariser.
En réalité ils transhument plus qu'ils ne restent sur place et un
grand nombre d'entre eux, qui se font compter parmi les "non transhumants"
fréquentent les aires protégées. Du reste, si la
présence de la race Gurmaji peut laisser penser à un
début de sédentarisation, d'après Kaboré (2010),
ceci est vite contredit par l'accroissement de la taille des troupeaux ;
d'après Boutrais (1996), en effet, la sédentarisation implique
à la fois le changement dans les races (ou leur croisement avec des
races adaptées) et l'abaissement des effectifs. L'accès
camouflé aux réserves repose la question de la valorisation de
leurs ressources fourragères déjà évoquée
par Kièma S. (2007), l'auteur suggère d'ailleurs un accès
bien encadré auxdites ressources.
Limiter les effectifs mais ne pas forcément
sédentariser
L'option de sédentarisation des élevages telle
que le recommandent bien souvent les services techniques étatiques
(Lhoste, 2004; Kossoumna Liba'a, 2009 ; Kossoumna Liba'a et al. 2010)
a déjà montré ses limites par le passé. Kossoumna
Liba'a (2009) montre d'ailleurs que même lorsque les éleveurs en
sont contraints, ils ne peuvent abandonner totalement la transhumance
saisonnière. De même, Dongmo et al. (2007) ont
observé que les tendances à la "sédentarisation" (aussi
constaté par D'Amico et al. 1995 en milieu Mbororo de
Centrafrique) qui naissent dans divers terroirs en Afrique soudano
sahélienne ne s'accompagnaient pas de décapitalisation des
troupeaux. L'option doit donc être écartée et nous
chercherons plutôt à garantir que l'élevage extensif se
maintient dans le terroir tout en s'adaptant. La limitation des effectifs dans
les troupeaux n'est donc pas synonyme pour nous de la fixation des
éleveurs. Une telle option dans les conditions actuelles à
Kotchari, où l'espace tend à manquer, serait risquée pour
l'état des ressources naturelles. Il faut en effet rappeler que des
politiques de
110 Nous avons cependant montré que cela relevait de
stratégie de camouflage

253
sédentarisation totale menées par le
passé (Touré, 1997 ; Scoones, 1999 ; Baroin, 2003) ont connu plus
d'échecs que de résultats probants, elles ont conduit à
des dégradations irréversibles localisées par suite
d'excès de charge. Il est constant en effet qu'une pâture
fréquente et prolongée sur un même secteur du territoire
(par exemple autour des points d'eau) par un nombre important d'animaux est
source d'instabilité.
Nous voyons comme durables des systèmes
d'élevage semi-sédentaires, c'est-à-dire à michemin
entre la sédentarisation et la mobilité telle qu'elle se pratique
actuellement. Nous n'ignorons cependant pas, comme Kossoumna Liba'a et
al. (2010), la difficulté d'une telle "demande" d'abaissement
des effectifs, cela appelle à un changement de paradigme qui conduirait
les éleveurs (les plus nombreux) d'un élevage extensif de
capitalisation, fondement de l'économie pastorale (Daget et Godron, 1995
; Boutrais, 1996), à un système plus marchand qui les ouvrirait
au marché (déstockage des mâles adultes). Cela suppose
cependant l'intensification du système (investissement en aliments
concentrés et en soins divers). Cette intensification suppose par
ailleurs une plus grande intégration à l'agriculture (Raimond,
1999 ; Requiers-Desjardins, 1999) : pratique de cultures à double
usage111 ; ce qui accroît la capacité des
écosystèmes pastoraux à supporter plus de pression de
charge (Harchies et al. 2007). Il est connu que le caractère
naisseur des troupeaux des éleveurs spécialisés (les Peuls
pour ce qui nous concerne) répond à deux stratégies
concourant toutes deux à l'accroissement des effectifs (Lhoste et
al. 1993; Boutrais, 1996): gérer les risques et avoir du lait
pour l'alimentation et surtout les échanges. En s'adonnant à des
activités agricoles à côté de celles pastorales, les
éleveurs seront moins dépendants de leurs troupeaux pour leur
alimentation et peuvent alors entretenir des troupeaux moins prolifiques tout
en valorisant mieux le lait qu'ils produisent comme le suggère Boutrais
(1996). Mais ce qui apparait comme un compromis nécessaire à nos
yeux entre ce type d'élevage et l'état des ressources n'est pas
évident à réaliser, le pastoralisme est en effet un
système de vie et comme tel ses transformations doivent s'inscrire sur
le long terme.
En définitive, l'argument de la maîtrise des
effectifs, même s'il ne permet pas de résorber totalement les
menaces qui pèsent sur les ressources étant donné le grand
dynamisme agricole localement observé, permet tout de même
d'abaisser notamment la pression de pâturage de saison pluvieuse et,
ainsi, de ne pas compromettre la production fourragère. Il s'agit de
maintenir le système dans une situation où la production
fourragère est telle qu'elle suffit à nourrir le cheptel sans se
détériorer.
Intervenir à d'autres niveaux
Nous sommes conscient du fait que toutes ces propositions ne
sont pas suffisantes, nous sommes en effet dans un système complexe qui
prend en compte d'autres acteurs (acteurs du mode agricole, de la conservation,
politiques, etc.) dont les activités interagissent avec les
activités pastorales. Comment alors, atteindre l'équité et
la durabilité lorsque les enjeux défendus par les uns et les
autres s'avèrent antagonistes ? (Harchies et al. 2007 ;
Djenontin, 2010).
111 Les cultures fourragères pures n'ont jamais pu
être adoptées en milieu paysan.

Il est un fait que les politiques de développement
rural, notamment les politiques foncières rurales (Kièma S., 2007
; Kaboré, 2010) privilégient les activités agricoles au
détriment des activités pastorales. Par ailleurs, selon
Kaboré (2010), la politique de gestion participative des aires
protégées et les retombées qui en résultent ainsi
que celle de gestion décentralisée des ressources naturelles dans
les terroirs, sont favorables aux velléités dominatrices des
communautés autochtones (généralement les agriculteurs),
celles-ci trouvent là, en effet, une sorte de seconde chance pour
reconquérir un pouvoir de maîtres territoriaux auparavant remis en
cause par les mêmes politiques. Les éleveurs traditionnels (les
Peuls) et leurs troupeaux, considérés comme des étrangers,
vivent ainsi une situation de précarité foncière du fait
de leur exclusion contrairement à l'esprit inclusif et d'accès
équitable aux ressources véhiculé par ces politiques ; ils
constituent alors une menace pour les objectifs de conservation et ce
malgré le renforcement en cours du dispositif de protection des
réserves (mise en place des ZOVIC comme zones tampon, plus grande
surveillance, etc.).
Cette question d'accès aux ressources des aires
protégées mérite d'être définitivement
posée. Nos résultats montrent en effet que, localement, le
système arrive à se réguler parce qu'une grande part du
troupeau accède au parc W et à la réserve partielle de la
Kourtiagou. Il est difficile d'imaginer les conséquences sur les
écosystèmes en périphérie, et donc sur les
populations et leurs troupeaux, si le dispositif de surveillance et de gestion
participative qui l'accompagne arrivait à bout de la pâture
illégale dans les réserves fauniques voisines. Sans
accéder au désir de la majorité des populations riveraines
de voir leur bétail accéder librement aux réserves (ce
qui, du reste, reviendrait à reporter les difficultés de
maintenant dans le futur), nous convenons avec Kièma S. (2007) qu'il y
va de l'intérêt même des gestionnaires des aires
protégées, d'entrevoir un accès contrôlé
à leurs ressources. Le sujet reste délicat et la forme
(accès par parcage direct ? accès indirect par fauchage de la
paille ?) reste à définir mais, de notre point de vue, il faut
anticiper sur ce qui adviendra forcément. A ce sujet, il faut signaler
qu'une expérience originale se mène dans le parc de la Pendjari
au nord-Bénin, contigu au parc W (extrait d'entretien avec Tiemoko
Djafarou, directeur national du Parc national de la Pendjari ; avril 2010). La
direction de cette réserve de biosphère, grâce à des
contrats signés avec les éleveurs résidents, permet
l'exploitation indirecte (fauchage de la paille) dans la zone tampon.
L'expérience s'est avérée concluante, elle a permis en
effet de mobiliser ces communautés, qui perçoivent
désormais l'utilité de cette entité, dans la surveillance
des ressources contre la pâture illégale. Des réflexions
sont d'ailleurs en cours pour envisager l'expérimentation du parcage
direct toujours sous-forme contractuelle.
En ce qui concerne les activités agricoles, le
défi réside dans la forte progression du front agricole imputable
à deux phénomènes : la demande naturelle en terre
résultant de la croissance démographique et la culture du coton
qui est très demandeuse d'espace. La durabilité du système
commande que des actions soient prises également à ce niveau,
l'objectif devant être de freiner à défaut de stopper
l'occupation et l'obstruction des terres de parcours par les champs. Mais cela
ne relève plus du domaine strict de la recherche.

255
7.2.3. Retour sur l'approche de recherche
utilisée
La recherche a été conduite suivant l'approche
systémique en partant du fait que le sujet étudié est
d'une certaine complexité (Lhoste, 1984; Landais 1992 &
1994 ; Daget & Godron, 1995). Cherchant à voir comment les hommes et
leurs animaux (système social) influaient sur l'état des
ressources naturelles (système naturel ou fourrager) et comment ils
s'adaptaient aux nouvelles conditions qui en résultaient, l'option
choisie nous paraissait judicieuse. Il est constant, en effet, que la
végétation et les ressources naturelles en général
réagissent à l'exploitation animale et que le bétail
s'adapte, à son tour, au nouvel état ainsi créé
(Grouzis, 1982 ; Daget & Godron, 1995). Il faut préciser que le
fonctionnement et la dynamique de l'élevage ne peuvent être saisis
que si ceux-ci sont replacés dans le contexte - entendu comme l'ensemble
formé par les ressources naturelles ainsi que les pratiques et
représentations des hommes - où ils s'opèrent. Pour
aborder cette réalité que représente ce socio -
écosystème ou système
éleveur-troupeau-environnement (Djenontin, 2010), l'approche
pluridisciplinaire s'imposait et les compétences que nous avions au
début de cette thèse se sont avérées insuffisantes.
Il faut rappeler que nous sommes zootechnicien à la base et que, pour
"passer la frontière" (Jollivet, 1992 in Djamen, 2008) de cette
discipline, il nous a fallu nous former en télédétection
et cartographie/SIG et parfaire nos connaissances en écologie
appliquée.
Ainsi donc, nous avons utilisé une batterie de
techniques et d'outils complémentaires qui s'appuient sur des
méthodes de diagnostic participatif et d'évaluation des
ressources pastorales.
L'intérêt de l'approche participative de
diagnostic utilisée c'est qu'elle mobilise les acteurs, permet de mettre
à jour des réalités ou connaissances qui, autrement,
seraient passées sous silence, elle jette, en outre, les bases d'une
relation de confiance entre les acteurs du processus de recherche. Sans compter
que lorsque le processus participatif est bien conduit, il permet
d'économiser en temps et en ressources pour des résultats presque
aussi intéressants que ceux qu'auraient générés des
dispositifs expérimentaux plus lourds. Ainsi, par exemple, nous n'avons
pas décelé de grands écarts entre les
représentations des éleveurs sur leurs milieux et leur calendrier
pastoral saisonnier par rapport à la réalité pratique
révélée par les analyses écologiques de terrain
ainsi que le suivi des troupeaux au pâturage; les éventuels
écarts étaient d'ailleurs révélateurs des
contraintes qui se posent aux éleveurs, ce qui permet d'identifier les
obstacles au fonctionnement "normal" du système.
L'approche telle que nous l'avons mise en oeuvre, demande
à être complétée pour permettre d'aller jusqu'au
bout de la logique qui la sous-tend. Le suivi de troupeau, s'il a permis de
comprendre que les choix des sites par les éleveurs et leurs troupeaux
se faisaient de manière raisonnée et bien souvent rationnelle, il
ne permet cependant pas de rendre compte de l'efficacité desdits choix.
En effet, des mesures de performances zootechniques (état corporel, gain
ou perte de poids ou d'autres productions) permettent d'établir
l'état nutritionnel (Dumont et al. 2001) des troupeaux et de
répondre à cette préoccupation qui constitue finalement le
but ultime des éleveurs. Une analyse comparée des performances
des animaux suivant les options opérées par chaque éleveur
ou groupe d'éleveurs, pouvait renseigner sur l'état des
différentes unités pastorales fréquentées par les
troupeaux (Lhoste et

al. 1993 ; Daget & Godron, 1995 ; Kagoné,
2000 ; Djenontin, 2010) et permettre de contourner la difficulté
posée par les méthodes directes d'estimation des charges.
D'après Boudet (1978), Lecrivain et al. (1993), Daget &
Godron (1995) et Kagoné (2000), les essais de charge avec bétail
permettent d'améliorer les estimations directes (mesure de biomasse) des
capacités des pâturages. Les auteurs font observer que quand le
gain de poids des animaux compense la perte de poids, les parcours sont en
situation stable et les animaux couvrent juste leurs besoins d'entretien. Si
par contre, le rapport entre la perte et le gain de poids est supérieur
à un (perte nette), cela traduit une situation de surcharge desdits
parcours. Inversement, s'il est inférieur à un (gain net), cela
indique que les charges sont inférieures aux capacités des
pâturages. Par ailleurs, le comportement alimentaire des animaux pouvait
être affiné en s'intéressant à la composition
botanique des prises alimentaires, au rythme de prise, aux quantités
ingérées, etc. (Kagoné, 2000). En outre, il apparait
nécessaire de procéder à l'évaluation du fourrage
aérien et des résidus culturaux fortement exploités en
vaine pâture où mis en stock et redistribués en
période de soudure ; ceci dans le but de mieux préciser la
situation fourragère locale en période sèche. Cependant,
le fait de ne l'avoir pas fait à ce stade signifie que les
capacités de charge ont été sous-estimées, ce qui
dans l'absolu n'est pas mal dans une approche de gestion durable qui commande
la prudence.
Ces aspects sont pris en compte dans nos projets de recherche
futures.
7.3. Perspectives de recherche futures
Notre recherche jette les bases pour un modèle pastoral
durable dans le terroir de Kotchari. Les recherches à poursuivre vont
donc participer à affiner ce modèle sur les aspects qui ont
été ignorés ou peu pris en compte.
Ainsi, on pourrait affiner la caractérisation des races
animales notamment bovines dans le terroir et aussi suivre leurs performances
sur des unités précises du terroir. Dans une perspectives de
gestion durable des terroirs, nous avons montré comment les
espèces et les races, par leur comportement, pouvaient être
favorables ou au contraire défavorables aux initiatives qui sont
développées. Cette caractérisation va surtout porter sur
leur comportement alimentaire au pâturage (composition de la ration, mode
de prélèvement alimentaire, etc.), aspect que le présent
travail n'a pas pu aborder en profondeur. Le suivi des performances
zootechniques va permettre de manière détournée de situer
l'état biologique actuel des parcours et ainsi d'en établir une
situation de référence pour ensuite pouvoir suivre leur
évolution.
Par ailleurs, comme nous l'avons indiqué plus haut,
l'établissement du bilan fourrager dans le terroir n'a pas pris en
compte la composante issue des parcelles cultivées dont on sait qu'elle
occupe une part importante dans les rations de début de saison
sèche au moment de la vaine pâture ou même dans la pratique
de complémentation chez les éleveurs surtout résidents.
Dans le même sens, on pourrait mener une étude de
caractérisation des ligneux fourragers (peuplement, importance relative,
régénération, etc.) et les possibilités
d'accès réels pour le bétail. Les travaux de
Ouédraogo (2008) et Sodré (2009) se sont intéressés
à la question mais de manière partielle, ils se sont
focalisés sur les contraintes liées à l'accès
physique au fourrage aérien;

257
Enfin, il est connu que l'efficacité des pratiques
pastorales peut se mesurer à travers les performances que celles-ci
induisent sur le bétail (Boudet, 1991 ; Lhoste et al. 1993 ;
Daget et Godron, 1995 ; Kagoné, 2000 ; Vall & Diallo, 2009 ;
Djenontin, 2010). Pour permettre d'apprécier l'efficacité des
paquets de pratiques que mettent en oeuvre les différents groupes
d'éleveurs, des activités participatives de suivi de performances
(NEC112, production en lait, impact sur les parcours, etc.) sont
à envisager, ils permettront de mieux juger de l'efficience et de la
pertinence des options techniques (races élevées, lieux
pâturés, nombre de lots, etc.) opérées par nos
éleveurs.
112 Note d'état corporel. D'après Djenontin
(2010), c'est un outil essentiel de gestion des animaux sur parcours dont
l'appropriation par les techniciens pourrait contribuer à
améliorer la communication avec les éleveurs et à
comprendre leurs objectifs, leurs stratégies de gestion du troupeau et
des pâturages.

258
REFERENCES BIBLIOGRAPHIQUES
1.

259
Achard, F., Hiernaux, P., Banoin, M., 2001. Les
jachères fourragères naturelles et améliorées en
Afrique de l'Ouest. In Floret Ch., & Pontanier, R. (eds), La
jachère en Afrique tropicale, vol 2, John Libbey Eurotext, Paris
2000, pp. 201-239.
2. ACRA, 2009. Plan communal d'action environnementale 2009-2014
de la commune de Tansarga. Analyse, 74p.
3. Adler P. B. , Milchunas D. G., Lauenroth W. K., Sala O. E.,
Burke I. C., 2004. Functional traits of graminoids in semi-arid steppes: a test
of grazing histories
4. Akpo L. E., Grouzis M., 2004. Interactions arbre/herbe en
bioclimat semi-aride: influence de la pâture. Note de recherche,
Sécheresse, 15 (3): 253-261.
5. Akpo L. E., Grouzis M., 2000. Valeur pastorale des herbages
en région soudanienne: le cas des parcours sahéliens du
Nord-Sénégal. Tropicultura, 18: 1-8.
6. Akpo L. E., 1992. Influence du couvert ligneux sur la
structure et le fonctionnement de la strate herbacée en milieu
sahélien. Les déterminants écologiques. Thèse de
doctorat de 3e cycle de Biologie Végétale, Université
Cheikh Anta Diop, Dakar, Sénégal, 175p.
7. Akpo L. E., Gaston A., Grouzis M., 1995. Structure
spécifique d'une végétation sahélienne. Cas de
Wiidu Thiengoli (Ferlo, Sénégal). Bulletin du Muséum
national d'Histoire naturelle, Paris, 4e sér., 17,
1995 Section B, Adansonia, n° 1-2 : 39-52
8. Akpo L. E., Grouzis M., Bada F., Pontanier R., Floret Ch.,
1999. Effet du couvert ligneux sur la structure de la végétation
herbacée des jachères soudaniennes. Note originale,
Sécheresse, 4 (10): 253-261.
9. Akpo L. E., Masse D., Grouzis M., 2002. Durée de
jachère et valeur pastorale de la végétation
herbacée en zone soudanienne au Sénégal. Ressources
alimentaires, Revue Élev. Méd. vét. Pays trop.,
55 (4): 275-283.
10. Alexandre, 1989. Dynamique de la
régénération naturelle en forêt dense de Côte
d'Ivoire. ORSTOM, Coll. "Etudes et Thèses", 102p.
11. Allen T. F. H., Starr, TH. B., 1982. Hierarchy perspectives
for ecological complexity. The University of Chicago Press, Chicago,
312p.
12. Allen V. G., Batello C., Berretta E. J, Hodgson J.,
Kothmann M., Li X., Mclvor J., Milne J., Morris C., Peeters A., Sanderson M.,
2011. An international terminology for grazing lands and grazing animals.
Grass and forage science, 66: 2-28.
13. Amadou B., 1999. Le pastoralisme dans le Boboye et le
Zarmaganda, continuité et rupture. In Bourgeot A. (ed.)
Horizons Nomades en Afrique Sahélienne. Karthala,
pp:327-338.
14. Andrieu N., Josien E., Duru M., 2007. Relationships
between diversity of grassland vegetation, field characteristics and land use
management practices assessed at the farm level. Agriculture Ecosystems
& Environment, 120: 359-369.
15.

Arbonnier M., Barbier N., De Wispelaere G., Dulieu D., Lejoly
J., Mahamane A., Ouadba J.M., Saadou M., 2002. Cartographie du couvert
végétal du complexe du W. ECOPAS report.
16. Arborio A-M., Fournier P., 1999. L'enquête et ses
méthodes: l'observation directe. Nathan, Sociologie, 128p.
17. Archibald S., Bond W. J., Stock W. D., Fairbanks D. H.
K., 2005. Shaping the landscape: fire-grazer interactions in an african
savanna. Ecological applications, 15 (1): 96- 109.
18. Asiedu F., Gouro A. S., Ndlovu L., Nuru H., Lametar K.,
2009. Multiplier les efforts pour soutenir le développement de
l'élevage dans les pays ACP: La science, la technologie et l'innovation
face aux défis de la sécurité alimentaire et de
l'émancipation économique. Note d'orientation ACP, CTA, 17p.
19. Aubertin C., 2005. La biodiversité: une notion en
quête de stabilité. In Aubertin C. (Ed)
Représenter la nature: ONG et Biodiversité. IRD
Editions, pp: 99-122.
20. Aubertin C., Pinton F., Rodary E., 2008. Le developpement
durable, nouvel âge de la conservation. In Aubertin C., Rodary E. (eds.)
Aires protégées, espaces durables?, IRD editions,
Objectifs Suds, pp: 17-27.
21. Aubreville A., 1957. Accord à Yangambi sur la
nomenclature des types africains de végétation, pp. 23-27.
22. Ba M., Mbaye M., Ndao S., Wade A., Ndiaye L., 2000.
Région de Diourbel: Cartographie des changements
d'occupation-utilisation du sol dans la zone agricole du Sénégal
occidental. Drylands Research, Working Paper 21, 44p.
23. Baars R.M.T. et Jeanes K.W., 1997. The grazing capacity of
natural grasslands in the Western Province of Zambia. Tropical
grasslands, 31: 561-568.
24. Babin D., Antona M., Bertrand A., Weber J. 2002.
Gérer à plusieurs des ressources renouvelables:
subsidiarité et médiation patrimoniale par récurrence.
In Cormier Salem Marie-Christine (ed.), Juhé Beaulaton
Dominique (ed.), Boutrais Jean (ed.), Roussel Bernard (ed.).
Patrimonialiser la nature tropicale: dynamiques locales, enjeux
internationaux. Paris : IRD, pp: 79-99.
25. Baillon F., Sournia G., 1987. Intégration
économique et aménagement des zones périphériques
des parcs nationaux et autres aires protégées: étude de
cas. Ministère de l'Environnement et du Tourisme, Ouagadougou, 94 p.
26. Balent G., Gibon, A, 1999. Organisation collective et
individuelle dans la gestion des ressources pastorales : conséquences
sur la durabilité agro-écologique des ressources, CIHEAM -
Options Méditerranéennes, 267-277.
27. Bambara T. G., 2010. Effet du changement climatique sur la
disponibilité des ressources fourragères. Mémoire
d'ingénieur, option élevage, IDR/UPB, 71p.
28.

261
Barbault R., 2008. Construire des visions partagées:
Pourquoi deS biodiversiteS ? communication aux Rencontres de valdeblore
"Pastoralismes, biodiversités, paysages dans les espaces
montagnards", 28-30 octobre, 6p.
29. Barbault R., 2006. Développement régional
et diversité écologique : liens et connexions ? Le point de vue
d'un écologue. In Mollard A., Sauboua E., Hirczak M.
(Eds.) Territoires et enjeux du développement
régional
30. Barbault R., 2002. La biodiversité: un patrimoine
menacé, des ressources convoitées et l'essence même de la
vie. In Barbault R. et al. (éds.). Johannesburg 2002:
Sommet mondial du développement durable, Quels enjeux? Quelle
contribution des scientifiques?, Ministère des Affaires
Etrangères, ADPF, pp. 53-87.
31. Barbault R., 1995. Le concept d'espèce-clé
de voûte en écologie de la restauration : clé ou impasse?
In : Lecomte, J. et al. (Eds.) : Recréer la
nature. Édition Nature sciences
sociétés, Hors série, pp 26-28.
32. Barbault R., 1993. Une approche écologique de la
biodiversité. Natures-SciencesSociétés, I : 322-329.
33. Baroin C., 2003. L'hydraulique pastorale, un bienfait pour
les éleveurs du Sahel? Afrique contemporaine, 1 (205): 205-224.
http://www.cairn.info/revue-afrique-
contemporaine-2003-1-page-205.htm
(consulté le 22 novembre 2010).
34. Barrière O., 1996. Gestion des ressources
naturelles renouvelables et conservation des écosystemes au Sahel: Le
foncier-environnement. Thèse de Doctorat en droit, Université de
Paris I Pantheon-Sorbonne, 99p.
35. Barrière O., Barrière C., 1997. Le
foncier-environnement: fondements juridicoinstitutionnels pour une gestion
viable des ressources naturelles renouvelables au Sahel. Editions FAO, col. "
Etudes législatives " n°60, Rome, 1997, 116p. http:/
www.c3ed.uvsq.fr/eger/publications/pdf/bibliographie.PDF
(consulté le 24 Juin 2004)
36. Bartolome J., Franch J., Plaixats J., Seligman N. G.,
2000. Grazing alone is not enough to maintain landscape diversity in the
Montseny Biosphere Reserve. Agriculture Ecosystems & Environment,
2000, 77: 267-273.
37. Bary H., 1998. Le savoir faire traditionnel des pasteurs:
exemple du Burkina Faso. PRASET, VSF, Ouagadougou, 60p.
38. Baumer M., 1997. L'agroforesterie pour les productions
animales. ICRAF, CTA, 340p.
39. Bayer W., Ciofolo I., 2004. Gestion complémentaire de
la faune sauvage et du bétail en Afrique de l'Ouest: utopie ou
perspective de développement? 64p.
40. Bayer W., Ciofolo I., von Lossau A., 2008. Complementary
management of wildlife and livestock in west africa-utopia or development
perspective?, 6p.
41. Beani, L. & Dessi, F., 1984. Les savanes africaines.
La nature et la vie animale. Editions Vinicio de Lorentiis/Debate-Itaca.
Traduction de Rémi Simon Collection « faune et flore du monde
», Larousse. Editions DURSUS, 126 p.
42.

Beerling D. J., Osborne C. P., 2006. Opinion: The origin of the
savanna biome. Global Change Biology, 12: 2023-2031.
43. Begon M., Townsend C.R., Harper J.L. 1996. Ecology,
individuals, populations and communities. Third Edition. Blackwell Science,
1068 p.
44. Beguin M., Pumain D., 1994. La représentation des
données géographiques: statistique et cartographie. Cursus,
Armand Collin, 192p.
45. Behnke R. H., Scoones I., 1992. Repenser l'écologie
des parcours: implications pour la gestion des terres de parcours en Afrique,
IED, Londres, 46p.
46. Benoit M., 1999a. Peuplement, violence endémique
et rémanence de l'espace sauvage en Afrique de l'Ouest: le no man's land
du « W » du Niger. In Espace, populations,
Sociétés, ORSTOM, pp. 29-52.
47. Benoit, M., 1999b. Opportunisme pastoral et conservation
de la savane en Afrique de l'Ouest. In Bourgeot, A. (éd.).
Horizons nomades en Afrique sahélienne. Sociétés,
développement et démocratie, éditions Karthala, pp
447-468
48. Benoit M., 1998. Dynamique des parcours pastoraux dans la
région du Parc National du W du Niger. Séminaire-atélier
sur « Utilisation durable de l'eau, des zones humides et de la
diversité biologique dans les écosystèmes partagés,
Bénin, BF, Niger, Togo. Tenu du 16 au 20 novembre 1998 à la
Tapoa, 7 p.
49. Benoit M., 1997. Conservation active et chasse au Niger.
Nature et Faune, vol 13, no 2: 21-29.
50. Benoit M., 1979. Le chemin des Peuls du Boobola:
contribution à l'écologie du pastoralisme en Afrique des savanes.
Travaux et documents de l'ORSTOM. 208 p.
51. Benoit M., 1976. Ecologie du pastoralisme et
aménagement (Haute Volta).Séminaire sur les méthodes de
planification du développement rural du 2 au 5 mars 1976,
Ministère du Plan, République de Haute-Volta, 8p.
52. Berlin S., 2002. L'écotourisme: protéger
l'éléphant et promouvoir les cultures locales? La région
de Boromo et l'aire protégée des deux Balé (au Burkina
Faso). Option Politiques du développement et de l'environnement,
Mémoire de DEA ADEn, Université d'Orléans, 128p.
53. Bernus E., 1981. Touaregs Nigériens: Unité
culturelle et diversité régionale d'un peuple pasteur. Orstom :
Paris.
54. Besse F., Toutain B., 2002. L'élevage et
l'environnement. In Mémento de l'agronome, CIRAD-GRET, pp:
1313- 1324.
55. Betsch J-M., Servan J., Drouin J-M., 2003. Introduction
à l'écologie et à la biodiversité. Cours de
DEA-EMTS 2003-2004. 140p.
56. Bierschenk T., Le Meur P.Y., 1997. Trajectoires peules au
Bénin. Six études anthropologiques. Paris : Karthala, 185 p.
57.

263
Binot A., Castel V., Caron A., 2006. L'interface
faune-bétail an Afrique subsaharienne. Sécheresse, 17
(1-2): 349-361.
58. Bird M. I., Veenendaal E. M., Moyo C., Lloyd J., Frost
P., 2000. Effect of fire and soil texture on soil carbon in a sub-humid savanna
(Matopos, Zimbabwe). Geoderma, 94:71-90.
59. Birot P., 1965. Géographie physique
générale de la zone mosaïque forêt-savane). C.D.U.,
Paris, pp. 94-106.
60. Blanchet A., Gotman A., 2003. L'enquête et ses
méthodes : l'entretien. Coll. Sociologie, Nathan Université,
Tours, France, 127 p.
61. Blanc-Pamard C., Milleville P., 1985. Pratiques
paysannes, perception du milieu et systèmes agraire." In A
travers champs, Agronomes et Géographes, Collection Colloques et
Séminaires. ORSTOM, Paris, pp: 101-138.
62. Blandin P., 1986. Bio-indicateurs et diagnostics des
systèmes écologiques. Bulletin d'Ecologie 17 (4) :
215-306.Blondel, J., 2003. Biodiversité, quels enjeux pour les
sociétés? Communication aux Journées de l'Institut
français de la biodiversité, Tours, 18-20 décembre 2003,
pp 17-19.
63. Blondel J., 1995. Biogéographie. Approche
écologique et évolutive. Masson, Paris. 297p.
64. Bodé S., 2008. Historique de l'itinéraire
et de la mobilité du groupe d'éleveurs transhumants WodaaBe Suudu
Suka'el de la commune de Tanout (Damergou), région de Zinder.
Mémoire de DEA Aménagement des espaces ruraux, UAMNiamey, LASDEL,
85p.
65. Bogdan R., Taylor S. J., 1975. Introduction to
Qualitative Research Methods. A Phenomenological Approach to the Social
Sciences. New York, John Wiley and Sons.
66. Bond, W.J., Keeley, J.E., 2005. Fire as a global
«herbivore»: the ecology and evolution of flammable ecosystems.
Trend in ecology and evolution, Vol. 20 n°7: 387-394.
67. Bonfiglioli, A. M., 1990. Pastoralisme, agro-pastoralisme et
retour : itinéraires sahéliens. Cah. Sci. Hum., 26
(1-2), pp : 255-266.
68. Bonfiglioli, A. M., 1988. Dudal. Histoire de famille
et histoire de troupeau chez un groupe de Wodaabe du Niger. Cambridge:
Cambridge University Press et Paris: Editions de la Maison des sciences de
l'homme.
69. Bonnet, 1990. Elevage et gestion des terroirs en zone
soudanienne. Cahiers Recherches Développement, no 25,
pp.43-67.
70. Botoni H. E., 2003. Interactions Elevage-Environnement.
Dynamique des paysages et évolution des pratiques pastorales dans les
fronts pionniers du Sud-Ouest du Burkina Faso. Thèse de doctorat de
l'université Paul Valery Montpellier III, Spéccialité :
Biologie des Populations et Ecologie, 293 p
71.

Botoni/Liehoun E., Daget Ph., César J., 2006.
Activités de pâturage, biodiversité et
végétation pastorale dans la zone Ouest du Burkina Faso.
Ressources alimentaires. Revue Elev. Méd. vét. Pays
trop. 59 (1-4) : 31-38.
72. Bouché, Ph., Lungren, G.C., Hein, B. et Omondi,
P., 2003. Recensement total aérien de l'écosystème W
(Arly-Pendjari-Oti-Mandouri-Kéran) (WAPOK). Rapport provisoire, 119p.
73. Boudet G., 1991. L'exploitation des parcours et la
conduite des troupeaux dans les systèmes d'élevage. In
Blanc-Pamard C., Léricollais A. (eds) A travers champs,
agronomes et géographes. Dynamique des systèmes agraires,
161-173.
74. Boudet G., 1984. Manuel sur les pâturages tropicaux et
les cultures fourragères. IEMVT, Ministère de la
Coopération, République Française, 266p.
75. Boudet, G., 1978, Manuel sur les pâturages tropicaux
et les cultures fourragères, série Manuels et précis
d'élevage, no 4, IEMVT, Ministère de la
Coopération, Paris, 258p.
76. Boudet G., 1975. Inventaire et cartographie des
pâturages de l'Afrique de l'Ouest. CIPEA, ILCA, Addis Abeba, Actes du
colloque de Bamako, Mali, 3-8 mars: 57-77.
77. Boulet R., 1975. Toposéquence des sols tropicaux en
Haute-Volta: équilibres dynamiques et bioclimat. Cahiers de l'ORSTOM,
série pédologie, 13: 3-6.
78. Boulet R., 1968. Carte pédologique de reconnaissance
de la Haute-Volta, Centre Nord, ORSTOM, Dakar, 351p.
79. Boulet R., Leprun J. C., 1969. Carte pédologique de
la Haute-Volta, Région Est, ORSTOM, Dakar, 319p + Cartes.
80. Bourbouze A.,1995. Gestion de l'animal sur le parcours.
In : Daget P., Godron M.(eds) Pastoralisme: Troupeaux, espaces et
sociétés. Hatier-AUPELF-UREF, 324-359.
81. Bourbouze A., 1982. L'élevage dans la montagne
marocaine. Organisation de l'espace et utilisation des parcours par
les éleveurs du Haut Atlas. Thèse de Doctorat Ingénieur de
l'INA Paris-Grignon. 345p. + ann.
82. Bourlière F., Hadley M., 1983. Present-day savannas:
an overview. In F. Bourlière (eds) Tropical Savannas,
Ecosystems of the World, Amsterdam, Elsevier, 13: 1-17.
83. Boutrais J., 2008. Pastoralisme et aires
protégées d'Afrique de l'Ouest en regard de l'Afrique de l'Est.
In Aubertin C., Rodary E. (eds) Aires protégées,
espaces durables ? Editions IRD, pp : 215-246.
84. Boutrais J., 2007. Crises écologiques et
mobilités pastorales: les Peuls du Dallol Bosso (Niger).
Sécheresse, 18 (1) : 5-12.
85. Boutrais J., 2002. Patrimoine animal et territoire chez
des sociétés peules. In Cormier Salem M-C.,
Juhé-Beaulaton D., Boutrais J. et Russel B. (eds.): Patrimonialiser
la nature tropicale: dynamiques locales, enjeux internationaux. Colloques et
Séminaires, IRD Editions, pp: 167-188.
86.

265
Boutrais J., 1999a. Journées de bergers au Nord-Cameroun.
In Poncet Y. (ed.) Les temps du Sahel: Hommage à Edmond
Bernus. IRD, pp. 55-80.
87. Boutrais., 1999b. Zébus et mouches
tsé-tsé: chronique de l'élevage en Adamaoua
(Caméroun). In Baroin C., Boutrais J. (eds.) L'homme et
l'animal dans le bassin du lac Tchad. IRD éditions, Colloques et
séminaires, pp:599-626.
88. Boutrais, J. 1997. Gens du pouvoir, gens du bétail.
In Boutrais J., Botte R., Schmitz J. (eds.) Figures peules.
Paris, Karthala, pp : 43-64.
89. Boutrais J., 1996. Populations et environnement pastoral
en Afrique tropicale. In Gendreau F., Gubry P. et Véron J.
(Eds): Populations et environnement dans les pays du Sud.
Karthala-Ceped, pp: 177-198.
90. Boutrais J., 1994. Les FoulBe de l'Amadoua et
l'élevage: de l'idéologie pastorale à la
pluriactivité. Cahiers d'Etudes africaines XXXIV (1-3), pp: 175-196.
91. Boutrais J., 1992. L'élevage en Afrique tropicale:
une activité dégradante? In L'environnement en Afrique
». Afrique contemporain, n°161, pp:109-125.
92. Boutrais J., 1990. Les savanes humides, dernier refuge
pastoral: l'exemple des Wodaabe, Mbororo de Centrafrique. Genève-Afrique
vet, XXVIII, 1: 66-90.
93. Boutrais J., 1984. Entre nomadisme et
sédentarité: les Mbororo à l'Ouest du Cameroun.
Systèmes agraires en mouvement, ORSTOM, 11, 32p.
94. Boutrais J., 1983. L'élevage soudanien. Des parcours
de savanes aux ranchs (Cameroun - Nigeria). Trav. et Doc. ORSTOM, n°160,
148 p. + cartes
95. Bovin M., 1999. La belle vache: chants et louange aux
animaux et aux êtres humains chez les Wodaabe du Niger.
In Baroin C. & Boutrais J. (eds.) L'homme et l'animal dans le
bassin du lac Tchad. Colloques et Séminaires, IRD Editions, pp :
203-219.
96. Breman H., 1975. La capacité de charge maximale
des pâturages maliens. In Inventaire et cartographie des pâturages
tropicaux africains. Actes Colloque Bamako, 3 - 8 mars: 249-256.
97. Breman, H. Kessler J.J., 1995. Woody plants in
agro-ecosystems of semi-arid regions, with an emphasis on the Sahelian
countries. Berlin-Heidelberg, Springer-Verlag, 340p.
98. Breman H., De Ridder N., 1991. Manuel sur les
pâturages des pays sahéliens, ACCTCTA-KARTHALA, 485 p.
99. Briggs, J.M., Nellis, M.D., Turner, C.L., Henebry &
Su, H., 1998. A landscape perspective of patterns and processes in tallgrass
prairie. In Knapp, A.K., Briggs, J.M., Hartnett, D.C. & Collins,
S.L. (eds). Grassland dynamics: long-term ecological research in tallgrass
prairie. LTER, New York, pp: 265-279.
100. Broussard F., 2001. L'intégration des
éleveurs dans la gestion d'un terroir villageois sahélien: le cas
de Mâl (Brakna-Mauritanie). Mémoire de maîtrise de
géographie, Laboratoire des Sciences de l'Environnement,
Université d'Angers, 104 p.
101.

Brunet R., Feras R., Théry H., 1998. Les mots de la
Géographie. Dictionnaire critique. Troisième édition,
Paris, RECLUS/La Documentation Française, 518 p.
102. Brunschwig Ph., Moulin C., Broqua Cl., Caramelle-Holtz
E., Lefrileux Y., 2001. Définition des principaux termes utilisés
pour caractériser l'alimentation des chèvres en production de
fromage AOC. Institut de l'élevage, UP AOC, Document de travail, 6p.
103. Bruzon V., 1995. Les feux de brousse dans les savanes
africaines. In Daget P. et Godron M (eds.). Pastoralisme :
troupeaux, espaces et sociétés, HATIER-AUPELF, UREF, pp:
269-282.
104. Bruzon V., 1994. Les pratiques du feu en Afrique
subhumide: exemples des milieux savanicoles de la Centrafrique et de la
Côte d'Ivoire. In Blanc-Pamard C. et Boutrais J.(eds.) A la
croisée des parcours: Pasteurs, éleveurs, cultivateurs.
Dynamique des systèmes agraires. ORSTOM, Colloques et
séminaires, pp : 147-162.
105. Bruzon, V., 1990. Les savanes du nord de la
Côte-d'Ivoire. Mésologie et dynamique: l'herbe, le feu et le
pâturage. Thèse de Doctorat de l'Université Paris VII,
Spécialité géomorphologie et dynamique des milieux
naturels, 301 p.
106. BUNASOLS, 2007 : Caractérisation des sols de la
province de la Tapoa. Rapport provisoire, PICOFA, 21 p.
107. BUNASOLS, 2000. Etude morphopédologique de la
province de la Gnagna, échelle 1/100000. Rapport technique 119, 110p.
108. BUNASOLS, 1980. Etude pédologique du
périmètre de Logobou - ORD de l'Est Fada NGourma, échelle
1/5000. Rapport technique 21, 40p.
109. BUNASOLS, 1979. Etude pédologique du
périmètre de Partiaga - ORD de l'Est Fada NGourma, échelle
1/5000. Rapport technique 17, 40p.
110. Burel, F. ; Baudry, J., 2003. Ecologie du paysage.
Concepts, méthodes et applications. Editions Tec & Doc,
Paris, New York, 359 p.
111. Burtt-Davy J., 1938. The classification of tropical woody
végétation types. Imp. For. Inst. Paper no. 13, 85 p.
112. Caillault S., Ballouche A., Delahaye D., 2009.
Organisation spatio-temporelle des feux de brousse. Approche comparative au
Burkina Faso. Neuvièmes rencontres de Théo Quant,
Besançon, 4 - 6 mars 2009, 10p.
113. Campbell R. S., 1954. Rapports entre le feu et le pacage en
forêt. Unasylva, vol. VIII, 2, 82 p.
114. Canfield, R. (1941). Application of the line interception
method in sampling range vegetation. J. Forestry. 39 : 338-394.
115. Carrière M., 1996. Impacts des systèmes
d'élevage pastoraux sur l'environnement en zones tropicales et
sub-tropicales arides et sub-arides d'Afrique et d'Asie, CIRADEMVT, FAO,
80p.
116.

267
Carrière M., 1994. Plantes de Guinée à
l'usage des éleveurs et des vétérinaires. Minist.
Coop. Fr., CIRAD-EMVT (eds.), 235 p. + 130 fig.
117. Carrière M., Toutain B., 1995. Utilisation des
terres de parcours par l'élevage et interactions avec l'environnement :
Outils d'évaluation et indicateurs Institut für
Biogéographie, Division Environnement, Saarbrucken, CIRAD EMVT,
MaisonsAlfort, France, 98 p.
118. CEMAGREF, 2008. Pour un meilleur équilibre
sylvo-cynégétique : des pratiques favorables aux cervidés.
Office national de la chasse et de la faune sauvage, Technique et faune
sauvage, 56p.
119. CEPE, 1983. Code pour le relevé méthodique
de la végétation et du milieu : principes et transcription sur
cartes perforées. Sous la direction de Emberger Louis, CNRS, 292 p.
120. César J., 2005. L'évaluation des ressources
fourragères naturelles. Production fourragère tropicale, Document
de synthèse, Fiche n° 17, CIRDES, CIRAD, 12p.
121. César J., 1994. Gestion et aménagement de
l'espace pastoral. In Blanc-Pamard C. et Boutrais J.(eds.) A la
croisée des parcours: Pasteurs, éleveurs, cultivateurs.
Dynamique des systèmes agraires. ORSTOM, Colloques et
séminaires, pp : 111-145.
122. César J., 1992. Etude de la production biologique
des savanes de Côte d'Ivoire et son utilisation par l'homme. Biomasse,
valeur pastorale et production fourragère. Thèse de doctorat de
l'Université de Paris VI, Sciences Naturelles, 642p.
123. César J., 1991. Typologie, diagnostic et
évaluation de la production fourragère des formations pastorales
en Afrique tropicale. Fourrages, 128, pp : 423-442.
124. César J., Coulibaly Z., 1993. Conséquence
de l'accroissement démographique sur la qualité de la
jachère dans le Nord de la Côte d'Ivoire. In Floret C.,
Serpantié G. (eds). La jachère en Afrique de l'Ouest.
Paris, ORSTOM, Atelier international 1991-12- 02/1991-12-05, Montpellier,
France, pp : 415-434.
125. Chao, A. (1984). Nonparametric estimation of the number of
classes in population. Scand J statist, 11: 265-270.
126. Cibois Ph., 2007. Les méthodes d'analyse
d'enquêtes. Que sais-je ?, puf, Paris, 127p.
127. Clerici N., 2006. Monitoring and assessing fire impacts
and land-cover change in tropical and subtropical ecosystems using satellite
remote sensing and GIS techniques. Phd thesis, 86 p.
128. Clerici N., Bodini A., Eva H., Grégoire J. M.,
Dulieu D., Paolini C., 2007. Increased isolation of two Biosphere Reserves and
surrounding protected areas (WAP ecological complex, West Africa), Journal
for Nature Conservation, 15: 26-40.
129. Cocheme J., Franquin P., 1968. Etude agroclimatologique
dans une zone semi - aride en Afrique au Sud du Sahara. Geneve : O.M.M.,
140p.
130. Cochrane, M.A., 2003. Fire science for rainforests.
Nature, 421: 913-919.
131. Cole, M.M., 1986. The savannas: biogeography and geobotany.
London, Academic Press, 438 p.
132.

Colwell, R. K., 2004. EstimateS: Statistical estimation of
species richness and shared species from samples. Version 7. (User's Guide and
application published at:
http://purl.oclc.org/estimates.).
133. Compère R., Hellemans P., Lecomte P., Lachapelle
A. et Messan F. (1993b). Les savanes guinéennes du ranch de
l'Adélé (Togo). 2. La production et la valeur alimentaire des
pâturages naturels. Bull. Rech. Agron. Gembloux, 28 (1):
5-28.
134. Condit R., Pitman N., Leigh E. G. Jr., Chave J. et al.
2002. Beta-Diversity in Tropical Forest Trees. Science 25, vol. 295
(5555): 666-669.
135. Connell, J.H., 1978. Diversity in Tropical Rain Forests and
Coral Reefs. Science, vol. 199:1302-1310.
136. Convers A., 2002. Etat des lieux spatialisé et
quantitatif de la transhumance dans la zone périphérique
d'influence du Parc National du W. Rapport de stage de DESS, UFR Sciences/Univ.
Montpellier, 41p.
137. Cormier Salem M-C., Juhe-Beaulaton D., Boutrais J.,
Russel B., 2002. Patrimonialiser la nature tropicale: dynamiques locales,
enjeux internationaux. Colloques et séminaires, IRD
éditions, 467p.
138. Da Lage A., Metailié G., 2000. Dictionnaire de
Biogéographie végétale. CNRS Editions, Paris, 579 p.
139. D'Amico F., Poussinga J.M., Le Masson C., Le Masson A.,
Cuisance D. (1995). Pratiques pastorales des Mbororo et trypanosomoses bovines
dans une zone de savanes humides de Centrafrique. Revue Méd. vét.
Pays trop., 48 (2), 203-212.
140. Daget Ph., 2004. Retour sur la valeur pastorale.
In Nouvelle PAC, quelles perspectives ?, Les journées de l'AFP,
22, 23, 24 septembre. Pastum, Bulletin de l'association Française du
Pastoralisme, 72, 2ème trimestre, pp: 21-23.
141. Daget Ph, 1976. Les modèles mathématiques en
écologie. Collection d'Ecologie 8, éditions Masson, Paris,
New-York, Barcelone, Milan, 172 p.
142. Daget Ph., Faugere E., 2003. Enquête pastorale et
enquête ethnographique: une question de symétrie, Nature
Sciences Sociétés, vol 11 no1-2003, pp: 46-50.
143. Daget Ph., Godron M., 1995. Pastoralisme : Troupeaux,
espaces et sociétés. HATIER, AUPELF, UREF, Universités
francophones, 510p.
144. Daget Ph., Godron M., 1982. Analyse de l'écologie
des espèces dans les communautés. Biogéocénoses
locales, Masson, Collection d'Ecologie, vol 18, 163p.
145. Daget Ph., Poissonet J., 1971. Méthode d'analyse de
la végétation des pâturages. Critères d'application.
Ann. Agron., 22, pp: 4-41.
146. Daget Ph., Poissonet J., 1969. Analyse phytologique des
prairies. Applications agronomiques, Ed. CNRS-CEPE Montpellier, doc 48, 67
p.
147. Dahl G., 1983. La production dans les
sociétés pastorales. In Galaty J.G., Aronson D., Salzman
P.C., Chouinard A. (eds.). L'avenir des peuples pasteurs. Compte rendu
de la conférence tenue à Nairobi, Kenya, 4-8 août 1980, pp:
220-231.
148.

269
Dahlberg C.A., 2000. Vegetation diversity and change in
relation to land-use, soil and rainfall. A case study from North-East District,
Botswana. Journal of Arid Environments, 44:19-40.
149. Dajoz R., 1982. Précis d'écologie.
4ème édition, Coll. Ecologie fondamentale et appliquée,
Gauthier-villars, Bordas, 503p.
150. Dedieu J.-P., Bernier M., Fortin J.-P., Pons I., Camps
B., 1997. Apport des données SIR-C pour évaluer les
propriétés du couvert neigeux dans les Alpes françaises :
résultats préliminaires. Proceedings of the 7th International
Symposium on Physical Measurements and Signatures in Remote Sensing, ISPRS,
A.A. Balkema, Rotterdam, pp: 703-709.
151. Dehoux J. P., Hounsou V. G., 1993. Productivité
de la race bovine Borgou selon les systèmes d`élevage
traditionnels au Nord-Est du Bénin. Revue mond. Zootech., 74/75
: 36-48.
152. Demment M. W., Greenwood G. B., 1988. Forage ingestion:
effects of sward characteristics and body size. J. of An. Sci., 66:
2380-2392.
153. Descoings, 1975. Méthode d'étude de la
structure des formations herbeuses tropicales. Colloque Bamako, 3-8 mars 1957,
ORSTOM, pp: 353-357.
154. Devineau J.-L., 2005. Generalist versus specialist : a
contrasted sociology of woody and herbaceous species in a fallow-land rotation
system in the West African savanna (Bondoukuy, Western Burkina Faso).
Phytocoenologia, 35 (1) : 53-77.
155. Devineau, J-L., 1999. Rôle du bétail dans
le cycle culture-jachère en région soudanienne: la
dissémination d'espèces végétales colonisatrices
d'espaces ouverts (Bondoukuy, sud-ouest du Burkina Faso). Revue
d'écologie (Terre et Vie), 54 : 97- 121.
156. Devineau J-L., 1997. Note sur l'analyse de la structure de
la végétation et sur la terminologie utilisée. In
Devineau J-L., Fournier A., Kaloga B. (1997), pp : 93- 96
157. Devineau J.-L., Fournier A., 1998. Ecologie d'une savane
africaine: synthèse provisoire de résultats acquis (avril, 1998).
ORSTOM/ERMES, Rapport de recherche, 77p.
158. Devineau J.-L., Fournier A., Kaloga, B., 1997. Les sols
et la végétation de la région de Bondoukuy, Sud-ouest
burkinabé: présentation générale et cartographie
préliminaire par télédétection satellitaire (SPOT).
Editions de l'ORSTOM, Paris, 117 p. + planches hors texte.
159. Devineau J- L., Lecordier C., Vuattoux R., 1984.
Evolution de la diversité spécifique du peuplement ligneux dans
une succession préforestière de colonisation d'une savane
protégée des feux (Lamto, Côte-d'Ivoire).
Candollea, 39: 103-134.
160. Devineau J-L., Fournier A., Nignan S., 2009.
«Ordinary biodiversity» in western Burkina Faso (West Africa): what
vegetation do the state forests conserve? Biodivers Conserv. 18: 2075
- 2099.
161. Devineau J.-L., Fournier A., Nignan S., 2010 . Savanna
fire regimes assessment with MODIS fire data: their relationship to land cover
and plant species distribution in

western Burkina Faso (West Africa). Journal of Arid
Environments, 74 (9):1092- 1101.
162. Devoto M., Medan D., 2003. Effects of grazing
disturbance on the reproduction of a perennial herb, Cypella herbertii (Lindl.)
Herb. (Iridaceae). Plant Systematics and Evolution, Austria, 9p.
163. De Wispelaere G., Peyre De Fabregues P, 1988. Evaluation
des ressoureges fourragères par télédétection SPOT
dans le sud Tamesna (Niger). CIRAD EMVT, Maisons Alfort, 74p + annexes + carte
1:250000.
164. De Wispelaere G., Waksman G., 1977. Contribution du
traitement des images-satellite à la cartographie des pâturages
sahéliens, Revue Elev. Méd. vét. Pays trop., 30
(4): 407- 424.
165. Diallo M.A., 2006. Savoirs locaux et pratiques de
conduite des troupeaux au pâturage: Elaboration d'une Méthode
d'Etude. DEA en gestion intégrée des ressources naturelles,
option production animale, CIRDES, IDR, UPB, 75p.
166. Diatta M., Grouzis M., Faye E., 1998. Typologie de la
végétation ligneuse en zone soudanienne. Focus, Bois et
Forêts des Tropiques, 257 (3): 23-36.
167. Djenontin J. A., 2010. Dynamique des stratégies
et des pratiques d'utilisation des parcours naturels pour l'alimentation des
troupeaux bovins au Nord-Est du Bénin. Thèse de doctorat en
aménagement et gestion des ressources naturelles, LEA/FSA,
Université d'Abomey-Calavi, Bénin, 214p.
168. Djenontin J. A., Houinato M., Toutain B., Sinsin B.,
2009. Pratiques et stratégies des éleveurs face à la
réduction de l'offre fourragère au Nord-Est du Bénin.
Sécheresse, 20 (4) : 346-53.
169. Dolidon H., 2005. L'espace des feux en Afrique de
l'Ouest: l'analyse spatio-temporelle d'un phénomène d'interface
nature-société. Thèse de doctorat de l'Université
d'Orléans, Spécialité Géographie physique, humaine,
économique et régionale, 414p.
170. Dongmo A. L., 2009. Territoires, troupeaux et biomasses:
enjeux de gestion pour un usage durable des ressources au Nord-Caméroun.
Thèse doctorat de l'Institut des Sciences et Industries du Vivant et de
l'Environnement (Agro Paris Tech), Spécialité Agronomie et
Zootechnie, 273p.
171. Dongmo A. L., Djamen P., Vall E., Koussou M. O.,
Coulibaly D., Lossouarn J., 2007. L'espace est fini! Vive la
sédentarisation? Innovations et développement durable en question
chez les pasteurs des zones cotonnières d'Afrique de l'ouest et du
centre. Renc. Rech. Ruminants, 14: 153-160.
172. Dongmo A. L., Dugué P., Vall E., Lossouarn J.,
2009. Optimiser l'usage des biomasses végétales cultivées:
une nécessité pour l'agriculture et l'élevage au
Nord-Cameroun. Accepté pour le colloque PRASAC, Avril 2009.
173. Dongmo A. L., Vall E., Dugué P., Kossoumna Liba'a
N., Bechir A. B., Lossouarn J. 2010. Le territoire d'élevage:
diversité, complexité et gestion durable. In SeinyBoukar
L., Boumard P. (eds.), « Savanes africaines en développement:
innover pour

271
durer », Actes du colloque, 20-23 avril 2009,
Garoua, Cameroun. Prasac, N'Djaména, Tchad ; Cirad, Montpellier, France,
9p.
174. Doussa S., 2006. La problématique de la culture
cotonniere dans les communes riveraines des aires classees a l'Est du Burkina
Faso: Etude de cas dans la périphérie burkinabè du Parc W.
Mémoire de DESS en Développement local, UFR/SEG, UO, 62p.
175. Doussa S., 2004. Les impacts de la culture
cotonnière sur la gestion des ressources naturelles du parc W: cas de
l'enclave de Kondio. Mémoire de Maîtrise de Géographie,
UFR/SH, ECOPAS, 123p.
176. Doutressoulle G., 1947. L'élevage en Afrique
Occidentale Française. Paris (V), éditions Larose, 298 p.
177. DRED-Est (Direction Régionale de l'Economie et du
Développement de l'Est), 2005. Tableau de bord socio-économique
de la région de l'Est, 6p.
178. DREP-Est (Direction Régionale de l'Economie et de la
Planification de l'Est), 2010. Tableau de bord socio-économique de la
région de l'Est, 10p.
179. DREP-Est (Direction Régionale de l'Economie et de
la Planification de l'Est), 2008. Atelier bilan des actions en faveur des
femmes, rapport introductif, Direction Régionale de la Promotion de la
Femme, 56p.
180. Dubar C. L., 2002. La socialisation: construction des
identités sociales et professionnelles Paris: Armand Colin, 255 p.
181. Dufrêne M., Legendre P., 1997. Species assemblages
and indicator species: the need for a flexible asymetrical approach.
Ecological Monographs, 67: 345-366.
182. Dugué P., 1998. Gestion de la fertilité et
stratégies paysannes. Le cas des zones de savanes d'Afrique de l'Ouest
et du Centre. Agriculture et développement, 18, pp : 13- 20.
183. Dugué P., Vall E., Lecomte Ph., Klein H-D.,
Rollin D., 2004. Evolution des relations entre l'agriculture et
l'élevage dans les savanes d'Afrique de l'Ouest et du Centre. OCL, Vol.
11 n°4/5, juillet-octobre, pp: 268-276.
184. Dumont B., 1996. Préférences et
sélection alimentaire au pâturage. INRA Productions An.,
9: 359-366.
185. Dumont B., Meuret M., Boissy A., Petit M., 2001. Le
pâturage vu par l'animal: mécanismes comportementaux et
applications en élevage. Fourrages, 166: 213-238.
186. Duvigneaud P., 1974. La synthèse écologique:
populations, communautés, écosystèmes, Biosphère,
Noosphère. Doin, Paris.
187. Ellis J, Swift DM. 1988. Stability of African pastoral
ecosystems. Journal of Range Management, pp: 41-450.
188. Elton C. S., 1924. Periodic fluctuations in the number of
animals: their causes and effects. Br. J. Exp. Boil. (1923-1929), 2:
119-163.
189.

Emberger L., Godron M., Daget Ph., Long G., Sauvage C., Le
Floc'H E., Wacquant JP., Poissonet J., 1983. Code pour le relevé
méthodique de la végétation et du milieu : principes et
transcription sur cartes perforées. CEPE, Montpellier, Editions du CNRS,
292p.
190. Fall T. S., 1993. Valeur nutritive des fourrages
ligneux. Leur rôle dans la complémentation des fourrages pauvres
des milieux tropicaux. Thèse Doct., ESA de Montpellier, 139 p.
191. FAO, 1991. Guidelines: land evaluation for extensive
grazing. FAO soils bulletin, 50, 82-138.
192. FAO, 1978. World soil resources report 48. Report on the
agro-ecological zones project. Methodology and results for Africa. FAO, Rome,
Italy, 158 p.
193. Faure J-F., 1997. Logique de l'utilisation de l'espace
pastoral dans le Sahel Nigérien: Modélisation et spatialisation
des interactions ressources-usages dans la zone de Banizoumbou (Sud-Ouest du
Niger). Mémoire de stage de DEA Environnement (ETES), Université
Paris VII, Université d'Orléans, Paris I, Paris IV, 127p.
194. Fischesser B., Dupuis-Tate M.F., 1996. Le guide
illustré de l'écologie. Cemagref Editions, Antony, 319 p.
195. Finegan, B. 1984. Forest succession. Nature, 312:
109-114.
196. Fontes J., Guinko S., 1995. Carte de la
végétation et de l'occupation du sol du Burkina Faso. Notice
explicative. Toulouse, Ministère de la Coopération
Française (Projet Campus 88 313 101), 65 p.
197. Fournier A., 2004. Compte rendu du Vème
Congrès mondial sur les parcs de l'Union mondiale sur la nature (IUCN).
Vie scientifique, Durban, Afrique du Sud, 8-18 septembre 2003, NSS,
12: 93-96.
198. Fournier A., 1996. Dans quelle mesure la production
nette de matière végétale herbacée dans les
jachères en savane soudanienne est-elle utilisable pour le
pâturage? In Floret Christian « Jachère, lieu de production
», Actes de l'Atelier de Bobo Dioulasso, 2-4 octobre 1996, CNRST, ORSTOM,
pp. 101-111.
199. Fournier, A., 1994. Cycle saisonnier et production nette
de la matière végétale herbacée en savanes
soudaniennes pâturées. Les jachères de la région de
Bondoukuy (Burkina Faso). Écologie, 1994, 25 (3) : 173-188.
200. Fournier, A., 1991. Phénologie, croissance et
production végétales dans quelques savanes d'Afrique de l'Ouest:
variation selon un gradient climatique. Editions de l'ORSTOM, Coll. Etudes et
Thèses, Paris, 312p.
201. Fournier, A., 1990. Variation de la dynamique foliaire
chez les Graminées pérennes le long d'un gradient climatique en
Afrique de l'Ouest. Comptes rendus de la XIIe réunion
plénière de l'aetfat, Symposium VII, Mitt. Inst. Allg. Bot.
Hamburg, Band 23 (b) : 823-839.
202.

273
Fournier, A., 1983. Contribution à l'étude de la
végétation herbacée des savanes de Ouango-Fitini
(Côte-d'Ivoire. Les grands traits de la phénologie et de la
structure. Conservatoire et jardin botaniques de Genève, 1983, 38 (1),
pp: 237-265.
203. Fournier, A., 1982. Cycle saisonnier de la biomasse
herbacée dans les savanes de Ouango-Fitini. Ann. Univ. Abidjan,
Série E (Ecologie), Tome XV, pp: 63-94.
204. Fournier A., Devineau J.-L., 2009. La plante dans
l'écosystème: une autre écologie végétale.
Bull. Soc. zool. Fr., 134(1-2) : 43-51.
205. Fournier A., Floret Ch., Gnahoua G-M., 2001.
Végétation des jachères et succession post-culturale. In
Floret Ch., Pontanier R. (eds) La jachère en Afrique tropicale. John
Libbey Eurotext, Paris, pp : 123-168.
206. Fournier A., Nignan S., 1997. Quand les annuelles
bloquent la succession postculturale... Expérimentation sur Andropogon
gayanus en savane soudanienne (Bondoukuy, Burkina Faso). Ecologie, t.
28 (1) 1997 : 13-21.
207. Fournier A., Toutain B., 2007. Suivi des
activités en matière de pastoralisme et de transhumance dans la
zone d'influence du parc régional du W du Niger. ECOPAS, CIRAD-ES, IRD,
73p.
208. Fritz H., 1995. Etude des systèmes mixtes
d'herbivores sauvages et domestiques en savane africaine. Structure des
peuplements et partage de la ressource. Thèse de doctorat,
Université Paris VI, France, 431p.
209. Fritz H., De Garine-Wichatitsky M., Lettesier
G., 1996. Habitat use by sympatric wild and domestic herbivores in an African
savanna woodland: the influence of cattle spatial behaviour. Journal of
Applied Ecology, 33: 589-598.
210. Frost P. G., Medina E., Menaut J. C., Solbrig O., Swift
M., Walker B. H., 1986. Response of savannas to stress and disturbance.
Biology International, Special Issue 10, IUBS, Paris.
211. Gardner T. A., Caro T., Fitzherbert E. B., Banda T.,
Lalbhai P., 2007. Conservation value of multiple-use areas in East Africa.
Conserv. Biol., 21: 775-787.
212. Gaston A., 1981. La végétation du Tchad
(Nord-Est et Sud-Est du Lac Tchad). Evolution récente sous des
influences climatiques et humaines. Thèse Doctorat d'Etat Sci. Nat.
Paris XII, mai, 333 p. 1 carte couleurs à 1/1 000 000 -160 000 km2. Mfe
IEMVT N°50 886.
213. Gaucherand S., 2005. Effets des pratiques pastorales sur
la biodiversité et sa dynamique dans les pelouses des alpages des alpes
du nord: apport des traits fonctionnels des plantes. Thèse de doctorat
de l'Ecole Nationale Supérieure Agronomique de Montpellier, Ecole
doctorale: Biologie Intégrative, Discipline: Biologie, 270p.
214. Gause G. F., 1935. Vérifications
expérimentales de la théorie mathématique de la lutte pour
la vie. Hermann et Cie, éditeurs, Paris, France.
215. Génot J. C., 2006. Vers un changement «
climacique »? Courrier de l'environnement de l'INRA, 53:
129-132.
216.

Génot J. C., Barbault R., 2004. Chapitre VII: Quelle
politique de conservation? In Barbault R. & Chevassus-au-Louis B. (eds)
Biodiversité et changements globaux, pp: 162-191. http//
www.adfp.asso.fr (16 septembre
2010).
217. Gill, A.M., 1975. Fire and the Australian flora: a
review. Aust. For., 38 : 4-25
218. Girard N., 1995. Modéliser une
représentation d'experts dans le champ de la gestion de l'exploitation
agricole, Thèse de Doctorat, Université Claude Bernard, INRA
SAD/IE, Lyon I, 234 pages + annexes
219. Goungounga J., 2003. Aspects techniques et humains de la
réhabilitation par semis direct de la forêt classée de Maro
dans l'Ouest du Burkina Faso. Mémoire de DEA ADEn, Université
d'Orléans, 91p.
220. Gounot M., 1969. Méthodes d'étude
quantitative de la végétation. Masson, Paris, 1-314.
221. Grall J., HilY. Ch., 2003. Traitement des données
stationnelles (faune). Fiche technique n° 10.
http://www.rebent.org//medias/documents/www/contenu/pdf/document/Fiches_techn
iques/FT10-2003-01.pdf (Consulté le 28 août 2010).
222. Grime, J. P. 1979. Plant strategies and vegetation
processes. New York: John Wiley and Sons, 222 p.
223. Grinnell, J. 1914. Barriers to distribution as regards
birds and mammals. American Naturalist, 48: 248-254.
224. Grouzis M., 1988. Structure, productivité et
dynamique des systèmes écologiques sahéliens (mare
d'Oursi, Burkina Faso). Editions de l'ORSTOM, Collection Etudes et
Thèses, 336 pages.
225. Grouzis M., 1982. Méthodes d'étude des
pâturages. ORSTOM, Ouagadougou, 43p.
226. Grouzis M., Sicot M., 1980. Une méthode
d'étude phénologique de populations d'espèces ligneuses
sahéliennes: influence de quelques facteurs écologiques.
In Le Houerou H N. (éds.) Les ligneux fourragers en
Afrique, état actuel des connaissances. Addis Abeba, Ethiopie, 8 -
12 Avril, CIPEA, pp:231 - 237.
227. Guérin G., Hubert B., 1995. Le troupeau au
pâturage. In Daget P., Godron M. (eds.) : Pastoralisme.
Troupeaux, espaces et sociétés. Universités
Francophones, pp : 361- 384.
228. Guérin H., Friot D., Mbaye N. D., Richard D.,
1991. Alimentation des ruminants domestiques sur pâturages naturels
sahéliens et sahélo-soudaniens. Etude méthodologique dans
la région du Ferlo au Sénégal. IEMVT, Maisons-Alfort, 115
p.
229. Guérin H., Friot D., Mbaye N. D., Richard D.,
1989. Le regime alimentaire des ruminants domestiques sur les pâturages
naturels sahéliens et soudano-sahéliens: essai de
détermination par l'étude du comportement alimentaire facteurs de
variations de la composition. du regime et conséquences nutritionnelles.
Revue Sénégalaise des Recherches Agricoles et
Halieutiques - Vol. 2 - n°1 - 1989
230.

275
Guibert H., Prudent P., 2005. Mise en place d'un programme
d'évaluation des impacts environnementaux exercés par le front
agricole pionnier dans l'aire protégée et la
périphérie du parc W. Mission de recherche Res/2004/050, Parc
régional W/ECOPAS, Bénin, Burkina, Niger, FED, Rapport
provisoire, 139p.
231. Guillaud D., 1994. Des pasteurs chez les villageois :
l'élevage dans l'Aribinda (Burkina Faso). In Blanc-Pamard C. et
Boutrais J.(eds.) A la croisée des parcours: Pasteurs,
éleveurs, cultivateurs. Dynamique des systèmes agraires.
Colloques et Séminaires, ORSTOM, CNRS-EHESS, Paris, pp: 215-236.
232. Guinko S., 1984. Végétation de la
Haute-Volta. Thèse de doctorat d'Etat, Université de Bordeaux
III, 394 p.+ annexes.
233. Guinko S., Ouoba P., Millogo-Rasolodimby J., 2000.
L'apport de l'inventaire des aires classées et protégées
dans la connaissance de la diversité végétale au Burkina
Faso. Berichte des Sonderforschungsbereichs 268, Band 14, Frankfurt
a.M, pp: 257-271.
234. Hahn-Hadjali K., 1998. Les groupements
végétaux des savanes du sud-est du Burkina Faso (Afrique de
l'Ouest). Études sur la flore et la végétation du
Burkina Faso et des pays avoisinants, Frankfurt a. Main/Ouagadougou, 3 :
3-79.
235. Hahn-Hadjali K., Schmidt M., Thiombiano A., 2006.
Phytodiversity dynamics in pastured and protected West African savannas. Pp
351- 359 in: S.A. Ghazanfar & H.J. Beentje (eds). Taxonomy and
ecology of African plants, their conservation and sustainable use, Royal
Botanic Gardens, Kew.
236. Hammer O., Harper D.A.T., Ryan P. D., 2001. PAST:
Paleontological Statistics Software Package for Education and Data Analysis.
Palaeontologia Electronica 4 (1).
237. Harchies M., Binot A., Wolff E., 2007. Impacts mutuels
de la conservation et de l'élevage transhumant sur l'occupation des sols
et les ressources environnementales : une étude de cas camerounaise.
VertigO, Hors Série 4, chapitre 10, 10p.
238. Hardin G. J., 1968. The tragedy of the commons: the
population problem has no technical solution; it requires a fundamental
extension in morality », Science, 162:1243-1248.
239. Hatfield R., Davies J., 2006. Revue mondiale de
l'économie du pastoralisme. L'Initiative Mondiale pour un Pastoralisme
Durable, Nairobi, 51p.
240. Heywood V. H., 1997. Information needs in biodiversity
assessments - from Genes to ecosystems. In Hawksworth D. L., Kirk P.
M., Clark S. D. (eds.) Biodiversity information: Needs and Options.
Proceedings of the 1996 international on biodiversity informations, Oxon
(UK), New-York (USA), CAB international, pp: 5- 20.
241. Hien, M., 1996. La reconstitution postculturale de la
végétation en savane soudanienne dans la région de
Bondoukuy (Burkina Faso). Les jachères de moins de six ans: flore,
persistance des adventices, lien avec le milieu et son utilisation. DEA
Sciences biologiques appliquées, option biologie et écologie
végétales, faculté des sciences et techniques,
Université de Ouagadougou, Laboratoire de botanique et biologie
végétale/ ORSTOM, 95 p.
242.

Hiernaux P., 1998. Effects of grazing on plant species
composition and spatial distribution in rangelands of the Sahel. Plant
Ecology, 138: 191-202.
243. Hiernaux P., 1980. L'inventaire du potentiel fourrager
des arbres et arbustes d'une région du Sahel Malien. Méthodes et
premiers résultats. In Le Houerou H N. (éd.) Les
ligneux fourragers en Afrique, état actuel des connaissances. Addis
Abeba, Ethiopie, 8 - 12 Avril, CIPEA, pp: 195 - 201.
244. Hiernaux P., 1975. Étude phytoécologique
du pays baoulé méridional (Côte-d'Ivoire centrale),
Th. ingén. C.N.R.S., Montpellier, 276 p.
245. Hiernaux P., Bielders C. L., Valentin Ch., Bationo A.,
Fernandez-Rivera S., 1999. Effects of livestock grazing on physical and
chemical properties of sandy soils in Sahelian rangelands. Journal of Arid
Environments, 41: 231-245.
246. Hoffmann O., 1985. Pratiques pastorales et dynamique du
couvert végétal en pays lobi (Nord-Est de la Cote d'Ivoire),
ORSTOM, Travaux et Documents n°189, Paris, 355p.
247. Hoffmann W. A., Orthen B., Vargas Do Nascimento P. K.,
2003. Comparative fire ecology of tropical savanna and forest trees. Functional
Ecology,17: 720-726.
248. Homewood K. M., Rodgers W. A., 1984. Pastoralism and
conservation. Human Ecology, 12 (4) : 431-441.
249. Hottin G., Ouédraogo O. F., 1976. Carte
géologique de la République de Haute Volta, 1/1000000. Direction
de la Géologie et des Mines, Republique de Haute Volta.
250. Hubert B., 1994. Pastoralisme et territoire.
Modélisation des pratiques d'utilisation. Cahiers Agriculture,
(3), 9-22.
251. Hubert B., Rigolot E., Turlant T., Couix N., 1993.
Forest fire prevention in the Mediterranean Region; News approachs to
agriculture-environment relations. In Brossier et al. (Eds.)
Systems Studies in Agriculture and rural development. INRA, France,
pp:63-86.
252. Huston, M. A. 1994. Biological diversity. Pages 681 in.
Cambridge University Press.
253. Ickowicz A., Mbaye M. 2001. Forêts soudaniennes et
alimentations des bovins au Sénégal : potentiel et limites.
Bois et Forêts des Tropiques, 270, pp:47-61
254. Illis J., 1999. La variabilité climatique et la
dynamique des écosystèmes complexes: implications pour le
développement pastoral. In Scoones I. (ed) Nouvelles
orientations du développement pastoral en Afrique. Vivre dans un
environnement incertain. Paris-Wageningen, CTA-Karthala, pp: 74-89.
255. IUCN, 1994. Liste des Nations Unies des parcs nationaux et
des Aires protégées 1993, Gland-Cambridge, IUCN.
256. Jackson B., Nurse M., Singh H.B., 1994. Temoignages: la
cartographie participative au service de la foresterie villageoise.
Réseau Foresterie pour le Développement Rural, Document n°17
du réseau, 7p.
257. Jacquin A., 2010. Dynamique de la
végétation des savanes en lien avec l'usage des feux à
Madagascar. Analyse par série temporelle d'images de
télédétection. Thèse de

277
doctorat de l'Institut National Polytechnique de Toulouse,
Université de Toulouse, spécialité: Fonctionnement des
écosystèmes et agrosystèmes, 144p.
258. Jaubert R., 1997. La relance du plan d'action de lutte
contre la désertification: les populations au centre des
négociations. In Tersiguel P. et Becker C. (eds.)
Développement durable au Sahel, Sociétés, Espaces,
Temps. Karthala, ClamecyFrance, pp. 31-42.
259. Jeensen, M., Michelsen, A., Gashaw, M., 2001. Responses
in plant, soil inorganic and microbial nutrient pools to experimental fire, ash
and biomass addition in a woodland savanna. Oecologia, 128: 85-93.
260. Jeltsch F., Weber G.E., Grimm V., 2000. Ecological
buffering mechanisms in savannas: a unifying theory of long-term tree-grass
coexistence. Plant Ecology, 161: 161-171.
261. Jentsch A., 2001. The Significance of Disturbance for
Vegetation Dynamics. A Case Study in Dry Acidic Grasslands. Dissertation,
Universität Bielefeld, 199 p.
262. Kabirou S., 2003. Etat des lieux spatialisé et
quantitatif de la transhumance dans la zone périphérique
d'influence du Parc National du W (Say, Kollo et Boboye). ECOPAS,
Université Abdou Moumouni (Niger), 114p.
263. Kaboré A., 2010. Brousse des uns, aire
protégée des autres, histoire du peuplement, perceptions de la
nature et politique des aires protégées dans le Gourma
burkinabè: l'exemple de la Réserve partielle de faune de Pama.
Thèse de doctorat présentée à l'Institut de Hautes
Etudes Internationales et du Développement, Genève, Suisse, 383p
+ annexes.
264. Kaboré B., 2009. Les zones villageoises
d'intérêt cynégétique de Gnimboama, Kabougou et
Saborga-Kori: perception des acteurs, enjeux écologiques et
socioéconomiques pour la conservation. Mémoire
d'ingénieur, IDR/UPB, Burkina Faso, 76p + annexes.
265. Kaboré-Zoungrana, C.Y., 1995. Composition
chimique et valeur nutritive des herbacées et des ligneux des
pâturages naturels soudaniens et des sous-produits agro-industriels du
Burkina Faso. Thèse d'Etat, Université de Ouagadougou, FAST, 201
p.
266. Kagoné H., 2004. Etat des lieux de la transhumance
dans la zone d'influence du Parc W du fleuve Niger, Rapport de synthèse,
Programme PARC-W / ECOPAS, 24p.
267. Kagoné H., 2000. Gestion durable des
écosystèmes pâturés en zone nord-soudanienne du
Burkina Faso. Thèse de doctorat. Gembloux, Faculté universitaire
des Sciences Agronomiques, 237p. + 44 tabl. 36 fig.
268. Kaiser L., 1983. Unbiased estimation in line-intercept
sampling. Biometrics, 39: 965- 976.
269. Kaloga B., 1968. Carte pédologique de reconnaissance
de la Haute-Volta, Centre Sud, 1/500000, ORSTOM, Dakar, 251p.
270. Kamuanga M., 2003. Rôle de l'animal et de
l'élevage dans les espaces et les systèmes agraires des savanes
soudano sahéliennes. Note introductive au thème 3. In
Jamin

J.Y., Seiny Boukar L., Floret C. (éds.) Savanes
africaines : des espaces en mutation, des acteurs face à de nouveaux
défis. Actes du colloque, mai 2002, Garoua, Cameroun. Prasac,
N'Djamena, Tchad - Cirad, Montpellier, France.
271. Kièma A., 2008. Effets des techniques de
restauration et d'exploitation des pâturages naturels sahéliens
sur la dynamique de la production fourragère. Thèse de doctorat
unique en développement rural, spécialité système
de productions animales, IDR/UPB, 175p.
272. Kièma S., 2007. Elevage extensif et conservation
de la diversité biologique dans les aires protégées de
l'Ouest burkinabé. Arrêt sur leur histoire, épreuves de la
gestion actuelle, état et dynamique de la végétation.
Thèse de doctorat, Université d'Orléans, 657p.
273. Kièma S., 2001. Conservation de la
diversité biologique et utilisation pastorale: la réserve de
biosphère de la mare aux hippopotames et les forêts
classées de Maro et du Tuy (Ouest Burkinabé), Mémoire de
Recherche, DEA ADEn, Université d'Orléans, 112p.
274. Kièma S., 1992. Utilisation pastorale des
jachères dans la région de Bondoukuy (Zone soudanienne, Burkina
Faso. Mémoire de D.E.S.S. « Gestion des Systèmes
AgroSylvo-Pastoraux en zone tropicale ». Université Paris XII, Val
de Marne, UFR des sciences, 89 p.
275. Kièma S., Fournier A., 2007. Utilisation de trois
aires protégées par l'élevage extensif dans l'ouest du
Burkina Faso. In Fournier A., Sinsin B. & Mensah G. A. (eds.)
Quelles aires protégées pour l'Afrique de l'Ouest?
Conservation de la biodiversité et développement. IRD
Éditions, Colloques et séminaires, pp : 498-506.
276. Koné A. R., 1987. Valeur nutritive des ligneux
fourragers des régions sahéliennes et soudaniennes d'Afrique
Occidentale: Recherche d'une méthode simple d'estimation de la
digestibilité et de la valeur azotée. Thèse 3ème
cycle, Univ. Paris VI - IEMVT, 205 p.
277. Kossoumna Liba'a N., 2008. De la mobilité
à la sédentarisation: gestion des ressources naturelles et des
territoires par les éleveurs Mbororo au Nord du Cameroun. Thèse
de doctorat en géographie, Université Paul-Valéry
Montpellier III, 260p.
278. Kossoumna Liba'a N., Dugué P., Torquebiau E.,
2010. Sédentarisation des éleveurs Mbororo et évolution de
leurs pratiques au Nord Cameroun. Etude originale, Cah. Agric., 19
(1): 60-67.
279. Kpoda C., 2010. Effet du changement climatique sur les
modes de conduite du bétail. Vulnérabilité et adaptation
des éleveurs du Burkina Faso. Mémoire d'ingérieur, option
élevage, IDR/UPB, 93p.
280. Kuela D. T., 2000. Monographie de la province de la Tapoa :
population et développement. CONAPO, PPLS, DRED-EST, CPAT/Tapoa, 97p.
281.

279
Laclan, J.P., W.S. Poumba, J.D. Nzlia, J.P. Bouillet and J.
Ranger. 2002. Biomass nutrient dynamics in a littoral savanne subjected to
annual fires in Congo. Acta Oecologica, 23: 41-50.
282. Laine, G. Pare, S., 1996. Apport de l'imagerie SPOT pour
l'évaluation statistique et cartographique des surfaces cultivées
en coton et en vivrier dans la zone cotonnière du Burkina Faso. Actes du
symposium international AISS (groupes de travail RS et DM), Ouagadougou
(Burkina Faso), du 6 au 10 février 1995, ORSTOM Éditions, p.
407-416.
283. Lambin, E., 1988. Apport de la
télédétection pour l'étude des systèmes
agraires et la gestion des terroirs en Afrique occidentale. Exemples au Burkina
Faso. Thèse de doctorat, 200p.
284. Lamotte M., 1979. Africa. Structure and functioning of the
savanna ecosystems of Lamto (Ivory Coast) in Unesco, Tropical grazing land
ecosystems, pp: 511-561.
285. Lamotte M., 1970. La participation au P.B.I. de la Station
d'Écologie tropicale de Lamto (Côte d'Ivoire). Bull. Soc.
Ecol., 1(2): 58-65.
286. Landais E., 1994. Systèmes d'élevage.
D'une intuition hooliste à une méthode de recherche, le
cheminement d'un concept. In Pamard C.B, Boutrais J. (eds.) A la
croisée des parcours : pasteurs, éleveurs, cultivateur
». Dynamique des systèmes agraires. ORSTOM, pp : 15-49.
287. Landais E., 1992. Les trois pôles des systèmes
d'élevage. Cahiers Recherches Développement, no 32-2,
pp: 3-5.
288. Landais E., 1990. Sur les doctrines des vétirinaires
coloniaux français en Afrique noire. Cahiers ORSTOM, Sér. Sc.
Hum, 26 (1-2), pp : 33-71.
289. Landais E, 1987. Recherches sur les systèmes
d'élevage. Questions et perspectives. Document de travail unité
INRA-SAD, INRA publications, 75p.
290. Landais E., Deffontaines J-P., 1989. Les pratiques des
agriculteurs: point de vue sur un nouveau contrat de recherche agronomique.
Etudes Rurales, n°109, pp: 125-158.
291. Lapeyronie A., Le Floc'H E., 1995. Pourquoi certaines
plantes sont-elles fourragères ? In Daget P. et Godron M. (eds)
Pastoralisme : troupeaux, espaces et sociétés. Paris,
Hatier-AUPELF.UREF, pp:100-110.
292. Lavorel S., Flannigan D., Lambin E. F., Scholes M.,
2007. Vulnerability of land systems to fire: Interactions among humans,
climate, the atmosphere, and ecosystems. Mitig Adapt Strat Glob
Change, Springer Science, 12: 33-53.
293. Lecomte Ph., 1995. Gestion intégrée des
ressources fourragères et de l'élevage bovin dans la savane de
basse altitude. Thèse de doctorat, Faculté Universitaire des
Sciences Agronomiques de Gembloux, 317 p. + tableaux, figures, annexes.
294. Lecrivain E., Leroy A., Savini I., Deffontaines J-P.,
1993. Les formes de troupeau au pâturage - génèse et
diversité.In Landais E. (ed.) Pratiques d'élevage
extensif, pp: 237-263.
295.

Le Guyadier H., 2006. La biodiversité: débat.
Focus, CNRS, pp: 6-11.
296. Le Houerou H N., 1980. Le rôle des ligneux
fourragers dans les zones sahélienne et soudanienne. In Le
Houerou H N. (éd.), Les fourrages ligneux en Afrique, état
actuel des connaissances. Addis Abeba, Ethiopie, 8 - 12 Avril, CIPEA,
pp:85 - 101.
297. Le Mire-Pêcheux L., Fournier A., Dugast, S., 2000,
« Andropogon gayanus et artificialisation (savane soudanienne) ».
In Gillon Y., Chaboud C., Boutrais J., Mullon C. (eds.) : Du bon
usage des ressources renouvelables, Paris, Editions de l'IRD, pp:
89-107.
298. "Le Pays" (quotidien d'informations
générales) N° 4884 du 13 juin 2011. Confidences du week-end:
Kotchari, la population en colère contre les agents des eaux et
forêts.
http://www.lefaso.net/spip.php?article42466
(consulté le lundi 13 juin 2011).
299. Lericollais et Faye 1994. Des troupeaux sans
pâturage en pays Serer Sénégal. In Pamard C.B,
Boutrais J. (eds.) A la croisée des parcours : pasteurs,
éleveurs, cultivateur ». Dynamique des systèmes
agraires. ORSTOM, pp: 165-196.
300. Levang P., 1978. Phytomasse herbacée de
formations sahéliennes. Etude méthodologique et application du
bassin versant de la mare d'Oursi. DGRST/ORSTOM, ACC. Lutte contre
l'aridité dans l'Oudalan, 34 p. +annexes.
301. Lhoste Ph., 2004. Pastoralisme et désertification:
quel avenir pour les sociétés pastorales sahéliennes?
Conférence donnée à Agropolis Museum, 15p.
302. Lhoste Ph., 1984. Le diagnostic des systèmes
d'élevage. Cahiers Recherche Développement, no 3-4, pp:
84-88.
303. Lhoste Ph., Dolle V., Rousseau J., Soltner D., 1993.
Manuel de zootechnie des Régions chaudes: les systèmes
d'élevage. Ministère de la Coopération. Collection
Précis d'élevage, 288p.
304. Lhoste Ph., Milleville P., 1986. La conduite des animaux
techniques et pratiques d'éleveurs. In Méthodes pour la
recherche sur les systèmes d'élevage en Afrique intertropicale.
Actes de l'atelier de MBour (Sénégal). Maisons-Alfort.
IEMVTCIRAD/ISRA. Coll. Etudes et synthèses de l'IEMVT.
n°20, pp: 247-268.
305. Loeser M. R. R., Sisk T. D., Crews T. E., 2006. Impact of
grazing intensity during drought in an Arizona grassland. Conservation
Biology, 21 (1): 87-97.
306. Lubchenco, J. 1978. Plant species diversity in a marine
intertidal community: importance of herbivore food preference and algal
competitive ability. American Naturalist, 112: 23-39.
307. Magda D., Meuret M., Hazard L., Agreil C., 2001.
Répondre à une politique de conservation de la
biodiversité Le pâturage des brebis pour la maîtrise des
landes à genêts. INRA, SAD, FaçSADe, 12,
septembre-décembre 2001, 4p.
308. Magurran A. E., 2004. Measuring ecological diversity.
Blackwell publishing, Malden, Oxford, Carlton, USA, UK, Australia, 256p.
309.

281
Mahamane A., Mahamane S., Lejoly J., 2007. Phénologie de
quelques espèces ligneuses du parc national du « W » du
Niger, Sécheresse, 1E (4) : 1-13.
310. Mazoyer M., Roudart L., 1998. Histoire des agricultures du
monde. Du néolithique à la crise contemporaine.
Tiers-Monde, 39 (153): 211-212.
311. Mbaygone E., 2008. Flore et végétation de la
réserve partielle de faune de Pama, sud-est du Burkina Faso.
Thèse de doctorat, Université de Ouagadougou, 137P.
312. McIntyre S., Heard K. M., Martin T. G., 2003. The
relative importance of cattle grazing in subtropical grasslands: does it reduce
or enhance biodiversity? Journal of Applied Ecology, 40: 445-457.
313. Ministère de la Coopération et du
Développement, 1991. Mémento de l'Agronome, 4è
édition. CIRAD-GRET, 1635 p.
314. MECV (Ministère de l'Environnement et du Cadre de
Vie), 2004. Rapport de suivi de la gestion des concessions. Direction des parcs
nationaux des reserves de faune et des chasses, 22 p + Annexes.
315. MEE (Ministère de l'Environnement et de l'Eau),
1997. Loi N°006/97/ADP portant Code forestier au Burkina Faso. 55p.
316. MEF (Ministère de l'Economie et des Fiances), 2008.
Recensement général de la population et de l'habitation de 2006,
résultats définitifs, 52p.
317. MRA (Ministère des Ressources animales), 2006.
Diagnostic des filières bétail-viande et petits ruminants au
Burkina Faso: rapport provisoire, Ouagadougou, Burkina Faso, Ministère
des Ressources animales, 147p.
318. MRA (Ministère des Ressources Animales), 2005.
Plan d'actions et programme d'investissements du secteur de l'élevage.
Version révisée à l'horizon 2015, Ouagadougou, Burkina
Faso, 93p.
319. MRA (Ministère des Ressources Animales), 2004. Les
statistiques du secteur de l'élevage au Burkina Faso, 54p.
320. MRA (Ministère des Ressources Animales), 2003.
Loi n°034-2002/AN portant loi d'orientation relative au pastoralisme au
Burkina Faso. Ouagadougou, Burkina Faso, Avril 2003, 27 p.
321. Menaut J.C., Lepage M., Abbadie L., 1995. Savannas,
woodlands and dry forests in Africa. In Seasonally dry tropical forests. Edited
by S.H. Bullock, H.A. Mooney & E.E. Medina. Cambridge University
Press. Cambridge, England, pp: 64-92.
322. Mettrick H., 1994. Recherche Agricole Orientée vers
le Développement: le cours ICRA, ICRA, CTA, AJ Wageningen, Pays-Bas, 288
p.
323. Meuret M., 1993. Piloter l'ingestion au pâturage.
In Landais E. (éd.): Pratiques d'élevage extensif. Identifier,
modéliser, évaluer. Etudes et Recherches sur les
Systèmes Agraires et le Développement (INRA), 27:161-198.
324.

Mills A.J., Fey M.V., 2004. Frequent fires intensify soil
crusting: physicochemical feedback in the pedoderm of long-term burn
experiments in South Africa. Geoderma, 121: 45-64.
325. Monnier Y., 1981. La poussière et la cendre.
Paysages, dynamique des formations végétales et stratégies
des sociétés en Afrique de l'Ouest. ACCT, Paris, 248 p.
326. Moorehead R., Lane C., 1995. Nouvelles orientations en
matière de politique et de tenure foncières des ressources
pastorales. In Chantal Blanc-Pamard et Luc Cambrezy (eds.): «
Dynamique des systèmes agraires: Terre, Terroir, Territoire: les
tensions foncières » éd. ORSTOM, Colloques et
séminaires, Paris, 472p.
327. Mueller-Dombois, D. & Ellenberg, H., 1974. Aims and
methods of vegetation ecology. John Wiley & sons, New York, London, Sydney,
Totonto, 547p.
328. Mysterud A., 2006. The concept of overgrazing and its role
in management of large herbivores. Wildlife Ecology, 2006, 12 (2):
129-141.
329. Naveh Z., 1975. The evolutionary significance of fire in
the Mediterranean region. Vegetatio, 9: 199-206.
330. Nebie O., 1996. Dégradation du milieu et
aménagement dans le plateau central, Burkina Faso. Berichte des
Sonderforschungsbereichs 268, Band 7, Frankfurt a.M. pp: 149- 176.
331. Niamir-Fuller M., 1999. Managing mobility in African
rangelands: the legitimization of transhumance. London, UK, Intermediate
Technology Publications Ltd, 240 p.
332. Nori M., 2007. La mobilité pastorale: une
histoire à réécrire. Cours modulaire sur le pastoralisme
au Master PARC (productions animales en régions chaudes), CIRAD,
Montpellier, 5p.
333. Nori M., 2006. Moyens mobiles d'existence, ressources
fragmentaires, droits variables: appréhender les territoires pastoraux.
Document de travail, International Land Coalition, 26p.
334. Nori M., Taylor M., Sensi A., 2008. Droits pastoraux,
modes de vie et adaptation au changement climatique. Coalition internationale
pour l'accès à la terre, IMPD, Irish Aid, iied, dossier n°
148, 28p. Source:
http://pubs.iied.org/pdfs/12543FIIED.pdf
(consulté le 22 septembre 2010).
335. Norse E.A., Rosenbaum K.L., Wilcove D.S., Wilcox B.A.,
Romme W.H., Johnston D.W., Stout M.L., 1986. Conserving biological diversity in
our national forests. Washingtin, DC: The Wilderness Society.
336. Nshimba H., 2008. Etude foristique, écologique et
phytosociologique des forêts de l'île Mbiye a Kisangani, RDCongo.
Thèse de doctorat de sciences, Faculte des sciences - Sciences
biologiques, Université Libre de Bruxelles, 272p.
337. OCDE, 1993. Corps central d'indicateurs de l'OCDE pour
les examens des performances environnementales. Rapport de synthèse du
Groupe sur l'État de

283
l'Environnement, Monographie de l'environnement, n°83,
Organisation de Coopération et de Développement
Économiques, Paris, 41p.
338. Olivier De Sardan, 2003. L'enquête
socio-anthropologique de terrain: synthèse méthodologique et
recommandations à usage des étudiants. Laboratoire
d'études et recherches sur les dynamiques sociales et le
développement local, Etudes et Travaux, n° 13, 58p.
339. Oloulotan, S. A., 1988. Productivité des
pâturages naturels du périmètre de NikkiKalalé
(Nord-Est Bénin). Thèse d'ingénieur agronome, FSA
/Université d'Abomey Calavi, Bénin, 196p.
340. Onana J., Touffet J., Alexandre D-Y., Yonkeu S.,
Asongwed-Awa A., Mvondo Awono J-P., Tarla F. N., 2003. La
végétation herbacée des zones humides du Nord-Cameroun.
In Jamin J. Y., Seiny Boukar L & Floret C. (eds.) Savanes
africaines: des espaces en mutation, des acteurs face à de nouveaux
défis. Actes du colloque, mai 2002, Garoua, Cameroun, 8p.
341. Ouédraogo D., 2008. Caractérisation des
ressources fourragères et des pratiques pastorales du terroir de
Kotchari à la périphérie du Parc W. Mémoire
d'ingénieur, IDR/UPB, Burkina Faso, 86p + annexes.
342. Ouédraogo H., 2000. Gestion des ressources
naturelles et décentralisation : dispositifs juridiques et cadres
politiques actuels. In Rapport général de l'atelier de
Cotonou sur le développement local et gestion
décentralisée des ressources naturelles, Appendice 2 du 10-16
décembre 2000, pp: 33-42.
343. Ouédraogo K., 2009. Evaluation des coûts
additionnels et des bénéfices du Parc W/Burkina Faso pour les
communautés agropastorales locales: cas des agropasteurs et des
transhumants du terroir riverain de Kotchari. Diplôme d'ingénieur
du développement rural, IDR/UPB, 52p.
344. Ouédraogo O., 2009. Phytosociologie, dynamique et
productivité de la végétation du Parc National d'Arly
(sud-est du Burkina Faso). Thèse de doctorat, Université de
Ouagadougou, 140p.
345. Oumarou S., 1994. Quelle stratégie pour une
meilleure intégration de la population locale dans la gestion des aires
protégées: cas du Parc National du W du Niger. Mémoire de
fin d'études, Faculté d'Agronomie, Université Abdou
Moumouni, Niamey.
346. Ozenda P., 1982. Les végétaux dans la
biosphère. Doin Editeurs, 431p.
347. Parguel P., Brunschwig P., Moulin C., Broqua C.,
Caramelle-Holtz E., Lefrileux Y., 2001. Définition des principaux termes
utilisés pour caractériser l'alimentation des chèvres en
production de fromage AOC. Document de travail, 6p.
348. Paris A., 2002. Etat des lieux quantitatif et
spatialisé de la transhumance dans la zone périphérique
d'influence du Parc National du W. Rapport de stage de DESS, Projet
ECOPAS/Ouaga, UFR Sciences/ Univ. Montpellier II/France, CIRAD-EMVT, 42p.
349.

Parker J.L., Fernandez I.J., Rustad L.E., Norton S.A., 2001.
Effects of nitrogen enrichment, wildfire and harvesting on forest-soil carbon
and nitrogen. Soil Science Society of America Journal, 65:
1248-1255.
350. Peco B., Sanchez A., Azcarate F., 2006. Abandonment in
grazing systems: Consequences for vegetation and soil. Agriculture
Ecosystems & Environment, 113: 284-294.
351. Pelissier P., 1995. Transition foncière en
Afrique Noire: du temps des terroirs au temps des finages. In Blanc-Pamard C.
et Cambrezy L. (eds.) Dynamique des systèmes agraires: Terre, Terroir,
Territoire: les tensions foncières. ORSTOM, Colloques et
séminaires, Paris, pp: 19-34.
352. Petit M., 1990. Géographie physique tropicale.
Approches aux études du milieu : morphogenèse, paysage. Paris,
Karthala-ACCT, 351 p.
353. Petit S., 2000a. Environnement des troupeaux et usage de
l'arbre chez les agropasteurs peuls de l'Ouest burkinabé. Approche
comparative et systématique de trois situations: Barani, Kourouma,
Ouangolodougou. Thèse de l'Université
d'Orléans/Laboratoire ERMES-IRD/CIRDES/CIRAD-Forêts. Tomes 1 &
2, 528 p. + 34 fiches.
354. Petit S., 2000b. Fourrage ligneux et parcours des troupeaux
des pasteurs peuls de l'ouest burkinabé. Note de recherche. Bois et
forêts des tropiques, 265 (3):77-80.
355. Pielou E.C., 1966. Shannon's formula as a measure of
species diversity: its use and misuse. Am. Nat. 100, pp: 463-465.
356. Pike G.H., Pulliam H.R., Charnov E.L., 1977. Optimal
foraging: a selective review of theory and tests. Quat. Rev. Biol.,
52: 137-153.
357. Poissonet J., César J., 1972. Structure
spécifique de la strate herbacée dans la savane à palmier
rônier de Lamto (Côte-d'Ivoire). Annales de
l'Université d'Abidjan, série E, Ecologie, tome
V, 1 : 15-18
358. Poissonet P., Poissonet J., 1969. Etude comparée
de diverses méthodes d'analyse de la végétation des
formations herbacées denses permanentes. Documents CNRS-CEPE n°50,
120 p.
359. Pollack C., 2009. Santé et pénibilité
en fin de vie active: une comparaison européenne. Document de travail,
Centre d'études pour l'emploi, 57p.
360. Pratt D. J., 1975. Problèmes qu'implique la
détermination de la capacité de charge des pâturages. In
Inventaire et cartographie des pâturages tropicaux africains.
Actes Colloque Bamako, 3 - 8 mars, pp: 259-260.
361. Pratt D. J., Le Gall F., De Haan C., 1997. Investing in
Pastoralism: Sustainable natural resource use in arid Africa and the Middle
East. World Bank Technical Paper 365, Washington DC.
362. Pratt, R.M., Gwynne, M.D., 1977. Rangeland Management and
Ecology in East Africa. Krieger Pub. Co.
363.

285
PROCORDEL, 2005. Rapport final d'exécution janvier
2000-mars 2005, Ouagadougou, Burkina Faso, CIRDES-ITC-CIRAD-ILRI.
364. Pyne, S.J., P.J. Andrews, R.D. Laven. 1996. Introduction to
wildland fire. Second edition, John Wiley & Sons, New York-Chichester, 769
p.
365. Raimond C., 1999. De la complémentarité
à la concurrence: agriculture et élevage dans les terres
d'inondation du bassin tchadien. In Baroin C., Boutrais J. (eds.)
L'homme et l'animal dans le bassin du lac Tchad. IRD éditions,
Colloques et séminaires, pp: 477- 498.
366. Rakotoarimanana V., Grouzis M., 2006. M. Influence du
feu et du pâturage sur la richesse et la diversité floristique
d'une savane à Heteropogon contortus du sud-ouest de Madagascar
(Région de Sakaraha), Candollea.
367. Rakotoarimanana V., Grouzis M., Le Floch E., 2008.
Influence du feu et du pâturage sur l'évolution de la phytomasse
d'une savane à Heterepogon contortus (Region de Sakaraha).
Tropicultura, 26 (1), pp : 56-60.
368. Rakotoarimanana V., Le Floch E., Grouzis M., 2001.
Influence du feu et du pâturage sur la diversité floristique et de
la production de la végétation herbacée d'une savane
à Heterepogon contortus (Region de Sakaraha). In Razanaka S.,
Grouzis M., Milleville P., Moizo B., Aubry C. (eds) Sociétés
paysannes, transitions agraires et dynamiques écologiques dans le
sud-ouest de Madagascar. CNRE/IRD, Antananarivo, Madagascar, pp: 339-353.
369. Ramade F., 1994. Eléments d'écologie,
écologie fondamentale. Deuxième édition. Ediscience
international, 579p.
370. Renaudin C., 2007. Etude socio-géographique sur
la zone cotonnière de l'Est de Burkina, Note intermédiaire, FARM
(Fondation pour l'agriculture et la ruralité dans le monde), 9 p.
371. Resquier-Desjardins M., 1999. L'accès aux
pâturages: une approche économique de la mobilité. In
Baroin C., Boutrais J. (eds.) L'homme et l'animal dans le bassin du lac
Tchad. Colloques et Séminaires, IRD éditions, pp:
525-523.
372. Riegel J., 2002. Utilisations pastorales et aires
protégées. Le cas des Peuls dans le Parc National du W du Niger,
Afrique de l'Ouest. Mémoire de DEA : Environnement, Milieux, Techniques
et Sociétés. Paris : MNHN, INA P-G, Université Paris VII,
67 p.
373. Rippstein G., 1985. Étude sur la
végétation de l'Adamaoua. Évolution, conservation,
régénération et amélioration d'un
écosystème pâturé au Cameroun. Études et
synthèses de l'IEMVT, n° 14, 370 p.
374. Rivière R., 1991. Alimentation des ruminants
domestiques en milieu tropical. Paris, La documentation Française.
375. Rivière R., 1975. Poblèmes de
l'évaluation de la valeur alimentaire des pâturages naturels
tropicaux. In Inventaire et cartographie des pâturages tropicaux
africains. Actes Colloque Bamako, 3 - 8 mars, pp: 269-276.
376.

Roscoe R., Buurman P., Velthorst E.J., Pereira J.A.A., 2000.
Effects of fire on soil organic matter in a «cerrado sensu-stricto»
from Southeast Brazil as revealed by changes in ä13C.
Geoderma, 95:141-160.
377. Roxburgh S. H., Shea K., Wilson J. B., 2004. The
intermediate disturbance hypothesis: patch dynamics and mechanisms of species
coexistence. Ecology, 85(2): 359-371.
378. Saidou A., 1986. Contribution a l'étude d'un
système pastoral sahélien la transhumance au Niger ses aspects,
son incidence et les perspectives d'avenir, Thèse de doctorat
vétérinaire, EISMV, Université de Dakar, 123p.
379. Sallah A., 1999. L'élevage extensif dans le
Damergou (Niger). In : Bourgeot, A. (ed.) Horizons nomades en Afrique
sahélienne. Sociétés, développement et
démocratie, édition Karthala, pp: 262-272.
380. Sankaran M., Hanan N.P., Scholes R.J., Ratnam J.,
Augustine D.J., Cade B.S., Gignoux J., Higgins S.I., Le Roux X., Ludwig F.,
Ardo J., Banyikwa F., Bronn A., Bucini G., Caylor K.K., Coughenour M.B., Diouf
A., Ekaya W., Feral C.J., February E.C., Frost P.G.H., Hiernaux P., Hrabar H.,
Metzger K.L., Prins H.H.T., Ringrose S., Sea W., Tews J., J., W., &
Zambatis N., 2005. Determinants of woody cover in African savannas.
Nature, 438: 846-849.
381. Sanou B. W., 2005. Monographie de la commune rurale de
Tansarga, province de la Tapoa. Région de l'Est, FICOD, 52 p.
382. Santoir C., 1999. Du Sahel à la savane. L'expansion
peule au sud de la Sirba [Gurma burkinabé]. IRD, 64p.
383. Santoir C., 1998. Le long voyage des Gurmaabe: la
dérive migratoire des Peuls du Gurma burkinabé. Avec la
collaboration de Guiré G. et Sy S., IRD, 36 + Cartes.
384. Saré S., 2003. Potentialités
fourragères et effets de l'élevage extensif sur la
diversité végétale dans la réserve de
biosphère de la mare aux hippopotames (Ouest burkinabé),
Mémoire d'ingérieur, IDR, UPB, 92p + annexes.
385. Savadogo S., 2004. Contribution au suivi
écologique des ressources fourragères dans la zone de chasse de
Pama Nord et le Ranch de gibier de Singou. Mémoire d'Ingénieur,
IDRlUPB, Burkina Faso, 89p + annexes
386. Sawadogo H., 2011. Le Parc National W du Burkina Faso:
Analyse des possibilités de création d'une zone tampon dans le
terroir de Kotchari. Mémoire de diplôme d'ingenieur de conception
en vulgarisation agricole, IDR, UPB, 82p + annexes.
387. Sawadogo I., 2004. Transhumance et pratiques pastorales
dans le terroir de Kotchari en périphérie du Parc W du Burkina
Faso. Mémoire de DEA EMTS Environnement : Milieux, Techniques,
Sociétés), INA P-G, ECOPAS, 63p + Annexes.
388. Sawadogo I., Devineau J-L., Fournier A. État des
ressources pastorales dans une terre d'accueil et de transit des pasteurs
transhumants : le terroir de Kotchari (sud-est du Burkina Faso)
Accepté dans Revue d'écologie (La Terre & La
Vie).
389.

287
Sawadogo, L. 1996. Évaluation des potentialités
pastorales d'une forêt classée soudanienne du Burkina Faso (cas de
la forêt classée de Tiogo). Thèse de doct. de 3è
Cycle de l'Université de Ouagadougou. Spécialité :
sciences biologiques appliquées, 147p.
390. Scholes, R.J. Walker, B.H., 1993. An African Savanna -
Synthesis of the Nylsvely Study. Cambridge University Press, Cambridge, UK.
studies, IBIS, 139 : 189-190.
391. Scholte P., Brouwer J., 2008. Relevance of key resource
areas for large-scale movements of livestock. Resource ecology, Chapiter 10A,
Wageningen UR Frontis Series, vol. 23: 211-232.
392. Scherrer, B., 2007. Biostatistique, volume 1, 2ème
édition, les éditions de la Chenelière inc., Edition
gaétan morin, Montréal, Paris, Casablanca, Canada, 816 p.
393. Schimper A.F.W., 1935. Pflanzengeographie auf
physiologischer Grundlage. 3 Aufl., hrsg von F.C.v on FABER. Fischer,
jena.,1613 p.
394. Schmitz A., Fall A. O., Rouchiche S., 1996. Contrôle
et utilisation du feu en zones arides et subhumides africaines. Ed. FAO.
147p.
395. Schnell, R., 1971. Introduction à la
phytogéographie des pays tropicaux. Vol 2: les milieux, les groupements
végétaux, Gauthiers Villars, Paris, 452 p. + fig. + photog. +
index + bibliogr.
396. Scoones I., 1999. Nouvelles orientations du
développement pastoral en Afrique: vivre dans un environnement
incertain, Khartala - CTA, Clamecy France, 362p.
397. Scoones I., 1995. Why are there so many animals? Cattle
population dynamics in the communal areas of Zimbabwe. In Behnke R.,
Scoones I., Kerven C. (eds) Range ecology at disequilibrium. News models of
natural variability and pastoral adaptation in African savanas. Overseas
Development Institute, Londres.
398. Senthilkumar K., Manian S., Udaiyan K., 1997. The effect of
burning on soil enzyme activities in natural grasslands in southern India,
Ecological Research, 12:. 21-25.
399. Seré, C., 1994. Classification and
characterization of world livestock production systems. Study document for
Interactions between livestock production systems and the environment, Rome,
FAO.
400. Seré C., Steinfeld H., 1996. World livestock
production systems. Current status, issues and trends. FAO Animal
Production and Health Paper, n°127, Rome, Italy
http://www.fao.org/WAIRDOCS/LEAD/X6101E/X6101E00.HTM
401. Shackleton, C. 2000. Comparison of plant diversity in
protected and communal lands in the Bushbuckridge lowveld savanna, South
Africa. Biological Conservation, 94: 273- 285
402. Shantz H. L., 1947. The use of fire as a tool in the
management of the brush ranges of California. California State Board of
Forestry, 156p.
403. Sinclair A. R. E., Byrom A. E., 2006. Understanding
ecosystem dynamics for conservation of biota. Journal of Animal
Ecology, 75: 64-79.
404.

Sinsin, B., 1993. Phytosociologie, écologie, valeur
pastorale et capacité de charge des pâturages naturels du
périmètre de Nikki-Kalalé au Nord-Bénin.
Thèse de doctorat en sciences agronomiques de l'Université Libre
de Bruxelles, section interfacultaire d'agronomie, 329 p.
405. Sinsin B., 1991. Exploitation des pâturages dans
un système traditionnel d'élevage bovin dans le
périmètre de Nikki-Kalalé au Bénin. Actes du
quatrième congrès international des terres de parcours,
Montpellier, France, volume 2, pp: 660-662.
406. Skearman P. J., 1982. Les légumineuses
fourragères tropicales. Collection FAO, Production
végétale et protection des plantes, 665 p.
407. Smith, J.W., Agyemang, K., Hailu, Z., Jabbar, M., Larbi,
A. et Osuji, P., 1997. Le développement du secteur laitier en Afrique
occidentale: situation actuelle et questions liées au
développement ultérieur. Acte du séminaire "Politique
pour le développement de l'élevage dans les zones humide et
subhumide de l'Afrique subsaharienne", Abidjan, Côte d'Ivoire, 5 - 9
février 1996, Wageningen, CTA.
408. Sodré, E. 2009. Caractérisation des
pâturages naturels du terroir de Kotchari et de la partie voisine du Parc
W (côté du Burkina Faso), Mémoire
d'Ingénieur, IDR/UPB, Burkina Faso, 63p + annexes.
409. Sounkéré K., 2003. Analyse des formes
d'utilisation de l'espace dans les terroirs agropastoraux de la zone
périphérique du parc National du W Bénin: cas de la
commune de Karimama, DSTPA, FSA, Université Abomey-Calavi, Cotonou,
76p.
410. Sournia G., 1987. Intégration économique et
aménagement des zones périphériques des parcs nationaux et
autres aires protégées: étude de cas. CDC, 89 p +
Annexes.
411. Sousa W., 1979. Experimental investigations of disturbance
and ecological succession in a rocky intertidal algal community. Ecological
Monographs, 49: 227-254.
412. Spinage C. A., Traoré S., 1984. Résumé
des aires de faune protégées et propositions. Document de travail
n°3, MET, FAO, 81 p.
413. Stark J.M., 1994. Causes of soil nutrient heterogeneity
at different scales. In Exploitation of environmental heterogeneity by
plants. Ecophysiological processes above- and belowground. Academic Press,
Inc., U.S.A, pp: 255-284.
414. Staver A. C, Bond J.W., STock D. W., Van Rensburg J. S.,
Waldram S.M., 2009. Browsing and fire interact to suppress tree density in an
African savanna, Ecological Applications, 19(7):1909-1919.
415. Steinfeld H., De Haan C. et Blackburn H., 1997.
Interactions entre l'élevage et l'environnement: problèmes et
propositions. Coopération française, Secrétariat d'Etat
à la Coopération, Direction du Développement, Bureau
« Gestion des ressources naturelles et environnement ».
http://www.virtualcentre.org/fr/dec/toolbox/index.htm
416. Stenning D,. 1959. Savannah Nomads. Londres, Oxford
University Press, 359 p.
417. Swift J., 1988. Dynamic ecological systems and the
administration of pastoral development
418.

289
Tamou C., 2002. Etat des lieux quantitatif et
spatialisé de la transhumance dans la zone périphérique
d'influence du Parc National du W (Bénin). Mémoire
d'Ingénieur, Sciences et Techniques de Production Animale, FSA,
Université d'Abomey-Calavi, ECOPAS, 102p.
419. Tankoano S. C. S., Ouédraogo M., Guiré T.,
Sanou F. S., 2010. Rapport d'achèvement de la troisième phase du
programme ADELE, Programme d'Appui au Développement local a l'Est,
Consortium ARCHEA (ARC Burkina Faso-ACADE Suisse), 93p.
420. Thébaud B., 2002. Foncier pastoral et gestion de
l'espace au sahel. Peuls du Niger oriental et du Yagha burkinabè, Paris,
Karthala, 319 p.
421. Thébaud B., 1999. Gestion de l'espace et crise
pastorale au Sahel: étude comparative du Niger oriental et du Yagha
burkinabé. Thèse de l'Ecole des Hautes Etudes en Sciences
Sociales (EHESS), 479 p.
422. Thébaud B., 1995. Le foncier dans le Sahel
pastoral: situation et perspectives. In Terroir, Chantal Blanc-Pamard
et Luc Cambrezy (eds.):« Dynamique des systèmes agraires: Terre,
Territoire: les tensions foncières » éd. ORSTOM,
Colloques et séminaires, Paris, pp: 37-56.
423. Thiombiano A., 2005. Les combrétacées du
Burkina Faso: taxonomie, écologie, dynamique et
régénération des espèces. Thèse d'Etat,
Université de Ouagadougou, 290p.
424. Thiombiano A., 1996. Contribution à
l'étude des Combrétacées dans les formations
végétales de la région Est du Burkina Faso. Thèse
de doctorat de 3ème cycle, Université de Ouagadougou, 220p.
425. Touré O., 2010. Pastoralisme et developpement
durable en Afrique de l'ouest: deux vieux compagnons de route.
http//
www.agriculturesnetwork.org/...durable/pastoralisme-et-developpementdurable-en-afrique/.../article
pdf (consulté le 18 juin 2011).
426. Touré O., 1997. Espace pastoral et dynamiques
foncières au Sénégal, Programme zones arides, 33p.
427. Touré O., 1994. La gestion des ressources naturelles
en afrique sahélienne: logiques étatiques et stratégies
des communautés pastorales. CRDI, BRACO, 26p.
428. Toutain B., 2001. Le risque en pastoralisme: quelques
considérations pour orienter les actions de développement. In
Tielkes E., Schlecht E., Hiernaux P. (eds.) Elevage et gestion des
parcours au Sahel, implications pour le développement. Beuren,
Stuttgart, Allemagne.
429. Touré I., Ickowicz A., Sagna C., Usengumuremyi
J., 2001. Étude de l'impact du bétail sur les ressources du Parc
National de Djoudj (PNOD-Senegal). Contribution to workshop Faune sauvage
et bétail: complémentarité, coexistence ou
compétition?, Niamey, 16-19 January.
430.

Toutain B., Bortolli L., Dulieu D.,Forgiarini G., Menaut J.C.,
Piot J., 1983. Espèces ligneuses et herbacées dans les
écosystèmes sahéliens pâturés de Haute-Volta.
ACC GRIZA (LAT), GERDAT, 124p.
431. Toutain B., Compaoré A., Ouadba J.M.,
Kagoné H., Diallo S., 2001. Mission d'appui scientifique «
transhumance ». Rapport CIRAD-EMVT n°01-43. CIRAD-EMVT, Montpellier,
France, 75p + annexes.
432. Toutain B., Lhoste Ph., 1978. Essai d'estimation du
coefficient d'estimation de la biomasse herbacée par le bétail
dans un périmiètre sahélien. Rév. Elev.
Méd. Vét. pays trop., 31 (1): 95-101.
433. Trabaud L., 1995. Le feu et ses incidences pastorales.
In Daget P. et Godron M (eds.). Pastoralisme : troupeaux, espaces et
sociétés, HATIER-AUPELF, UREF, pp. 263-269.
434. Traoré S., 2008. Les formations à Acacia
de l'Est du Burkina Faso : typologie en relation avec les facteurs
pédoclimatiques, prédictions spatiales et fonctions sur la
dynamique du carbone et de l'azote. Thèse de doctorat en Biologie et
Ecologie végétales, Université de Ouagadougou, 144p.
435. Trochain J.L., 1940. Contribution à l'étude
de la végétation du Sénégal. Mém. Inst.
fran. Afr. Noire. Vol. 2, 433 p. + 30 pl. phot.
436. Turco A., 2004. Astres et désastres: voyage dans la
conflictualité pastorale autour du parc transfrontalier de la W (Burkina
Faso-Bénin-Niger), Ponts, vol. 4, pp: 15-35.
437. Turner M. D., Hiernaux P., Schlecht E., 2005. The
distribution of grassing pressure in relation to vegetation resources in
Semi-arid West Africa: The role of herding. Ecosystems, 2005, 8:
668-681.
438. Turner M. D., 2004, «Political ecology and the
moral dimensions of «resource conflicts»: the case of farmer-herder
conflicts in the Sahel », Polical Geography, 23, pp: 863-889.
439. Turner M.G., Collins S., Lugo A. et al. 2003.
Long-term ecological research on disturbance and ecological response.
BioScience, 53: 46-56.
440. UNESCO, 1995. La stratégie de Séville pour
les réserves de biosphère. Nature et Ressources,
31(2).
441. UNESCO, 1996. Réserves de biosphère : la
stratégie de Séville et le cadre statutaire de réseau
mondial. Unesco, Paris.
442. Vall E., Diallo M. A., 2009. Savoirs techniques locaux et
pratiques: la conduite des troupeaux aux pâturages (Ouest du Burkina
Faso). NSS, 17: 122-135
443. Vall E., Blanchard M., Diallo M.A., Dongmo A.L., Bayala
I., 2009. Savoirs techniques locaux sources d'innovations? Production de
savoirs actionnables dans une démarche de recherche action en
partenariat. In Savanes africaines en développement: innover
pour durer. Actes du colloque du 21 au 24 avril 2009, Garoua, Cameroun
444.

291
Veuille M., 2006 La biodiversité: débat.
Focus, CNRS, PP: 6-11.
445. Veyret, P., 1951. Géographie de l'élevage.
Gallimard, Imprimerie Floch Mayenne, France, 254 p.
446. Vissac B., 1994. Mouvements et mémoire de
l'élevage. In Blanc-Pamard C. et Boutrais J.(eds.) A la
croisée des parcours: Pasteurs, éleveurs, cultivateurs.
Dynamique des systèmes agraires. Colloques et Séminaires, ORSTOM,
CNRS-EHESS, Paris, pp :79- 81-108.
447. Von Maydell H J., 1983. Arbres et arbustes du Sahel, leurs
caractéristiques et leurs utilisations. 531 p.
448. Wallis de Vries M. F., 1996: "Effects of resource
distribution patterns on ungulate foraging behaviour: a modelling approach".
Forest Ecology and Management, 88: 167-177.
449. Wambwa E., 2004. Diseases of Importance at the
Wildlife/Livestock Interface in Kenya. Kenya Wildlife Service, chapter 3,
pp:21-25.
450. Wan S., Hoi D., Loo Y., 2001. Fire effects on nitrogen
pools and dynamics in terrestrial ecosystems: a meta-analysis. Ecological
Applications, 11(5): 1349-1365.
451. Wane A., 2006. Economie du pastoralisme: une analyse
bibliographique (Afrique de l'Ouest). Initiative Mondiale pour un Pastoralisme
Durable (IMPD), IUCN EARO, PPZS, 23p.
452. Weber J., 2002. Enjeux économiques et sociaux du
développement durable. In Barbault R. et al. (Eds.). Johannesburg 2002:
Sommet mondial du développement durable, Quels enjeux? Quelle
contribution des scientifiques? Ministère des Affaires
Etrangères, ADPF, pp. 13-44.
453. Western D, Maitumo D. 2004. Woodland loss and restoration
in a savanna park: a 20- year experiment. African Journal of Ecology,
vol 42, Issue 2: 111-121.
454. White F., 1986. La végétation de l'Afrique:
mémoire accompagnant la carte de végétation de l'Afrique.
Paris, ORSTOM, 384p.
455. White P.S., Jentsch A., 2001. The search for generality in
studies of disturbance and ecosytems dynamics. Progress in Botany, 62:
399-449
456. White P.S., Pickett S.T.A., 1985. Natural disturbance and
patch dynamics: an introduction. In « The ecology of natural
disturbance and patch dynamics », pp: 5-13.
457. Whittaker R.H., 1972. Evolution and measurement of species
diversity. Taxon, 21, pp: 213-251.
458. Winter E., 2004. Max Weber et les relations ethniques. Du
refus du biologisme racial à l'État multinational (foreword by
Philippe Fritsch), Quebec, Presses de
l'UniversitéLaval, 214 p.
459.

Wittig R., Hahn-Hadjali K., Thiombiano A., 2000. Les
particularités de la vegetation et de la flore de la chaîne du
Gobnangou dans le sud-est du Burkina Faso. Etudes flor. Vég. Burkina
Faso, 5: 49-64.
460. Wittig R., König K., Schmidt M., Szarzynski J., 2007:
A Study of Climate Change and Anthropogenic Impacts in West Africa. Env Sci
Pollut Res 14 (3) : 182-189.
461. Woldu Z., Saleem M.A.M., 2000. Grazing induced biodiversity
in the highland ecozone of east Africa. Agriculture Ecosystems &
Environment, 79: 43-52.
462. Yaméogo U. G., 2005. Le feu, un outil
d'ingénierie écologique au Ranch de Gibier de Nazinga au Burkina
Faso. Thèse de doctorat de l'Université d'Orléans, Option
Physiologie et Biologie des organismes, populations et interactions, 268p.
463. Yanra J.D.D., 2004. Caracterisation des paturages
natcrels en zone sud soudanienne du Burkina Faso: cas des terroirs de Sidi,
Guena et Banfoulague dans la province du Kénédougou.
Mémoire d'ingénieur du développement rural,
Université Polytechnique de Bobo-Dioulasso, Burkina Faso, 77p.
464. Yates, D., T. Kittel, and R. Cannon2000. Comparing the
correlative Holdridge model to mechanistic biogeographical models for assessing
vegetation distribution response to climatic change. Clim. Change, 44:
59- 87.
465. Zombra A. W., 2008. Contribution à la
réalisation de l'intégration conservation de la
biodiversité et lutte contre la pauvreté: Cas du parc W et du
terroir riverain de Kotchari. Mémoire d'ingénieur du
développement rural, Université Polytechnique de Bobo-Dioulasso,
Burkina Faso, 49p.
466. Zoundi J. S., Hitimana L., 2008. Élevage et
marché régional au Sahel et en Afrique de l'Ouest:
Potentialités et défis. CSAO-OCDE / CEDEAO, 163p.
467. Zoungrana I., 1993. Diversité, stabilité
et évolution des communautés végétales. Les
jachères nord-soudaniennes du Burkina Faso. Ann de l'univ de
Ouagadougou, série B, 11p.
468. Zoungrana I., 1991. Recherches sur les aires
pâturées du Burkina Faso. Thèse Doc. es Sciences Naturelles
de l'Université Bordeaux III, UFR Aménagement et Ressources
Naturelles, 277p.
469. Zouri I., 2003. Impacts des pratiques pastorales sur la
végétation de la périphérie du Parc W
(côté Burkina Faso). Mémoire d'Ingénieur, IDR /UPB,
Bobo-Dioulasso, Burkina Faso, 63p + Annexes.

293
TABLE DES MATIERES
DEDICACES I
REMERCIEMENTS II
ABRÉVIATIONS, ACRONYMES ET SIGLES V
SOMMAIRE VIII
INTRODUCTION GENERALE 1
CHAPITRE I 6
CONTEXTE GENERAL ET PROBLEMES DE RECHERCHE 6
1.1. LE CONTEXTE DE LA RECHERCHE: PASTORALISME, AIRES
PROTÉGÉES ET ESSOR DU COTON 7
1.1.1. La région de l'Est et l'élevage
pastoral : entre perspectives et incertitudes 7
1.1.2. Les aires protégées de l'Est :
un potentiel en sursis ? 8
1.1.3. Des mutations agricoles porteuses de risques
pour les équilibres socio-économique et environnemental.
11
1.2. PROBLÉMATIQUE ET JUSTIFICATION DE LA RECHERCHE 11
1.3. QUESTIONNEMENT SCIENTIFIQUE ET HYPOTHÈSES DE
RECHERCHE 12
1.4. OBJECTIFS 14
1.4.1. Objectif général et
finalité 14
1.4.2. Objectifs opérationnels
14
1.5. DÉMARCHE CONCEPTUELLE 15
1.5.1. Objet de l'étude et cadre conceptuel :
le système pastoral, un système écologique complexe
et
piloté 15
1.5.2. Définition des termes et concepts
16
1.5.2.1. Les aires de conservation 16
1.5.2.2. Notions de biodiversité, de conservation
et d'écologie des parcours 17
1.5.2.3. Concepts de pastoralisme et de pratiques
pastorales 19
CHAPITRE II. ETAT DES CONNAISSANCES : FONCTIONNEMENT
DES
ECOSYSTEMES SAVANIENS, SOCIOLOGIE DU PASTORALISME ET
EVOLUTION DE LA POLITIQUE DE CONSERVATION 22
2.1. NATURE ET TYPES DE SAVANES 23
2.2. LES ÉCOSYSTÈMES SAVANIENS : FONCTIONNEMENT ET
PERTURBATION 24
2.2.1. Les feux et leurs rôles dans le
fonctionnement des écosystèmes de savane 25
2.2.1.1. Importance socio-économique et types de
feux 25
2.2.1.2. Les feux, un facteur de régulation des
savanes 25
2.2.1.3. Impacts des feux sur les
écosystèmes 26
2.2.1.3.1. Les feux et leurs impacts sur la
végétation 26
2.2.1.3.2. Les feux et leurs impacts sur les sols 28
2.2.1.3.3. La végétation de savane, une
végétation adaptée au feu. 29
2.2.2. Les perturbations des milieux dues à la
pâture 30
2.2.3. L'agriculture et les successions
post-culturales 35
2.3. LES INTERACTIONS ENTRE LE BÉTAIL ET LA FAUNE SAUVAGE:
QUELLE POSSIBILITÉ DE COMPROMIS ? 37
2.4. LE PASTORALISME : UN GENRE DE VIE, UN RAPPORT PARTICULIER
À L'ESPACE 38
2.5. LES GRANDES ÉTAPES DANS L'HISTOIRE DE LA CONSERVATION
40
CHAPITRE III. CARACTERES PHYSIQUES, HUMAINS ET SOCIO
-
ECONOMIQUES DE LA PROVINCE DE LA TAPOA 43
3.1. LA ZONE D'ÉTUDE ET SA LOCALISATION 44

3.2. LE MILIEU BIOPHYSIQUE 46
3.2.1. Les caractéristiques climatiques
46
3.2.2. La géomorphologie et les sols
50
3.2.3. Le réseau hydrographique
55
3.2.4. La végétation et la flore
56
3.2.5. Les réserves fauniques : le parc
national du W et la concession de chasse de la Kourtiagou. 57
3.2.5.1. Le parc national du W 57
3.2.5.2. La concession de chasse de la Kourtiagou
58
3.3. LE MILIEU HUMAIN 59
3.4. LES SYSTÈMES D'ÉLEVAGE 60
3.4.1. Importance du secteur 60
3.4.2. Les systèmes extensifs
61
3.4.2.1. L'élevage sédentaire
61
3.4.2.2. L'élevage transhumant 63
3.4.3. L'élevage intensif et semi-intensif
64
3.5. LES AUTRES ACTIVITÉS SOCIO-ÉCONOMIQUES 64
3.5.1. Les activités agricoles
64
3.5.2. Les activités extractives : la chasse
et la pêche. 65
CHAPITRE IV. LES UNITES PAYSAGERES PASTORALES DANS LE
TERROIR ET
DANS L'AIRE PROTEGEE VOISINE : DEFINITION,
DISTRIBUTION
SPATIALE ET CARACTERISTIQUES 68
4.1. INTRODUCTION 69
4.2. MATÉRIEL ET MÉTHODES 69
4.2.1. Approche des unités paysagères
pastorales par la télédétection 69
4.2.2. Analyse phytoécologique par
relevés au sol 71
4.2.2.1. Relevés de la strate herbacée
71
4.2.2.2. Évaluation de la phytomasse
herbacée 72
4.2.2.3. Relevés de la strate ligneuse
74
4.2.3. Enquêtes sur la qualité des
espèces fourragères : la valeur pastorale 75
4.2.4. Analyse des données 76
4.2.4.1. Cartographie des unités paysagères
pastorales 76
4.2.4.2. Constitution et caractérisation des
groupements agrostologiques, lien avec les unités
pastorales paysagères 77
4.2.4.3. Détermination de la diversité
spécifique des espèces herbacées 78
4.2.4.3.1. La diversité intra-placeau ou diversité
á. 78
4.2.4.3.2. La diversité inter-placeaux ou diversité
â 79
4.2.4.3.3. La richesse spécifique totale ou
diversité ã dans et en dehors du parc W. 80
4.2.4.4. Le recouvrement ligneux 80
4.2.4.5. La phytomasse herbacée et la
capacité de charge 80
4.2.4.6. La valeur pastorale des unités
paysagères pastorales 82
4.3. RÉSULTATS ET DISCUSSION 82
4.3.1. Définition des unités
paysagères pastorales 82
4.3.2. Les groupements agrostologiques et leurs
espèces caractéristiques 85
4.3.3. Caractéristiques des unités
paysagères pastorales 87
4.3.3.1. La richesse en groupements agrostologiques des
unités paysagères 87
4.3.3.2. La diversité floristique des
unités paysagères pastorales : les diversités alpha
(richesse S,
équitabilité E) bêta
(J) et gamma (SChao2) 93
4.3.3.3. La phytomasse herbacée et la
capacité de charge 96
4.3.3.4. Caractéristiques de la strate ligneuse
dans les unités paysagères pastorales 99
4.3.3.5. La valeur pastorale des unités
paysagères 104
4.3.4. Appréciation globale des parcours de
Kotchari 106
4.4. CONCLUSION 107
CHAPITRE V. LES SYSTEMES D'ELEVAGE A KOTCHARI : PRATIQUES
ET
STRATEGIES D'HIER ET D'AUJOURD'HUI 108
5.1. INTRODUCTION 109

295
5.2. MATÉRIEL ET MÉTHODES 110
5.2.1. Méthodes d'acquisition des
données 110
5.2.1.1. La revue des données 110
5.2.2.2. Les enquêtes 111
5.2.2.2.1. Les entretiens 111
5.2.2.2.2. L'estimation des effectifs animaux dans le terroir
111
5.2.2.2.3. La typologie des systèmes d'élevage dans
le terroir 112
5.2.2.2.4. Les pratiques et leur évolution 114
5.2.2.3. Analyse des données : description et
tests de comparaison 115
5.3. RÉSULTATS ET DISCUSSIONS 116
5.3.1. L'élevage face à la pression
d'occupation de l'espace du terroir 116
5.3.1.1. La pression démographique 116
5.3.1.2. La pression animale : le bilan fourrager
saisonnier 118
5.3.2. Les races bovines dans le terroir
123
5.3.2.1. La race Barbaji 124
5.3.2.2. La race Gurmaji (ou Guiabo)
124
5.3.2.3. La race Kiwali 125
5.3.2.4. La race Boboroji (Mbororooji
ou Borheji) 126
5.3.2.5. La race Jaliji (Djeli)
127
5.3.2.6. La race Gudali 127
5.3.3. Les acteurs et leurs pratiques
d'élevage 128
5.3.3.1. Résultats généraux
128
5.3.3.2. Les types d'élevages dans le terroir et
leurs caractéristiques 132
5.3.3.2.1. Le tri et la catégorisation des variables en
vue de la typologie 132
5.3.3.2.2. Les axes factoriels, les variables et les
modalités associées 132
5.3.3.2.3. Les catégories d'éleveurs 136
5.3.3.2.4. Caractéristiques des catégories
d'éleveurs 138
5.3.4. Les pratiques pastorales : évolution
depuis 20 ans 140
5.3.4.1. Constitution et allotement des troupeaux
140
5.3.4.2. Garde des troupeaux 146
5.3.4.3. Mobilité des éleveurs
150
5.3.4.4. Complémentation des animaux
153
5.3.4.5. Diversification des troupeaux 156
5.3.5. Les rapports sociaux entre les acteurs et
leurs conséquences sur les pratiques pastorales 161
5.3.5.1. Les relations entre les transhumants et les
acteurs locaux 162
5.3.5.1.1. Des liens de réciprocité parfois
anciens, mais de plus en plus fragiles 162
5.3.5.1.2. Des relations de plus en plus conflictuelles 164
5.3.5.1.3. Une présence des transhumants qui reste
bénéfique à l'économie locale 165
5.3.5.2. Relations entre les populations
résidentes 165
5.3.5.3. Relations agroéleveurs - administration
forestière : la question de la fréquentation des
aires
protégées. 167
5.3.5.4. Relations entre transhumants 168
5.4. CONCLUSION 169
CHAPITRE VI. LE TROUPEAU AU PATURAGE :
LOGIQUES,
REPRESENTATIONS ET REALITES DU TERRAIN 171
6.1. INTRODUCTION 172
6.2. MATÉRIEL ET MÉTHODES 173
6.2.1. Approche de la connaissance locale des
pâturages 173
6.2.1.1. Définition, description et classement des
unités paysagères participatives 173
6.2.1.2. Le repérage et la matérialisation
des unités identifiées 174
6.2.2. Suivi de troupeaux au pâturage
174
6.2.2.1. Approche de suivi de troupeau 174
6.2.2.2. Les activités de l'observateur lors du
suivi 175
6.2.3. Identification des autres ressources et des
contraintes non fourragères 176
6.2.4. Analyse des données : approche de
système d'information géographique (SIG) 177
6.3. RÉSULTATS ET DISCUSSIONS 178
6.3.1. Les saisons de l'éleveur
178
6.3.2. Une valeur pastorale qui change en fonction de
plusieurs paramètres 180
6.3.2.1. Les unités paysagères pastorales
participatives : types et caractéristiques 180

6.3.2.2. Unités paysagères pastorales
participatives versus unités paysagères pastorales
écologiques 182 6.3.2.3. Connaissance,
représentations et usages : les catégories pastorales et les
logiques de leur
fréquentation par les éleveurs
183
6.3.2.3.1. Les paramètres considérés et leur
importance au fil des saisons 183
6.3.2.3.2. La chaîne de pâturage saisonnière
à dire d'acteurs: les représentations que les éleveurs ont
de leurs pâturages
185
6.3.3. Les animaux au pâturage : variations
saisonnières de l'utilisation des pâturages 188
6.3.3.1. Les caractéristiques des troupeaux suivis
188
6.3.3.2. Comportement des éleveurs et de leurs
troupeaux au pâturage 190
6.3.3.2.1. Variations saisonnières de l'utilisation des
pâturages chez le troupeau TrpC1-2G (troupeau gourmantché de la
catégorie C1-2) 192
6.3.3.2.1.1. Distances et durées moyennes de
déplacement du troupeau 192
6.3.3.2.1.2. Importance relative des activités au
pâturage 192
6.3.3.2.1.3. Le circuit pastoral quotidien au fil des saisons :
unités pâturées et durée de fréquentation
193
6.3.3.2.2. Variations saisonnières de l'utilisation des
pâturages chez le troupeau de type TrpC1-2P (troupeau peul de la
catégorie C1-2) 203
6.3.3.2.2.1. Distances et durées moyennes du
déplacement du troupeau 203
6.3.3.2.2.2. Importance relative des activités au
pâturage 203
6.3.3.2.2.3. Le circuit pastoral quotidien selon les saisons :
unités pâturées et durée de fréquentation
204
6.3.3.2.3. Variations saisonnières de l'utilisation des
pâturages chez le troupeau de type Trp 213
6.3.3.2.3.1. Distances et durées moyennes du
déplacement du troupeau 213
6.3.3.2.3.2. Importance relative des activités au
pâturage 213
6.3.3.2.3.3. Le circuit pastoral quotidien selon les saisons :
unités pâturées et durée de fréquentation
214
6.3.3.2.4. Variations saisonnières de l'utilisation des
pâturages chez le troupeau de type TrpC3 222
6.3.3.2.4.1. Distances et durées moyennes du
déplacement du troupeau 222
6.3.3.2.4.2. Importance relative des activités au
pâturage 222
6.3.3.2.4.3. Le circuit pastoral quotidien selon les saisons :
unités pâturées et durée de fréquentation
223
6.3.3.3. Synthèse sur le comportement du troupeau
au pâturage 227
6.3.3.3.1. Un comportement territorial, révélateur
de l'état (disponibilité et distribution des ressources) des
parcours. 227
6.3.3.3.2. Un objectif sous-jacent : maximiser le bilan
énergétique 228
6.3.3.3.3. Des itinéraires dont la forme est
imprimée par la conjonction de plusieurs paramètres. 230
6.3.4. Représentations versus pratiques : les
éleveurs et leurs troupeaux à l'épreuve du terrain
230
6.4. CONCLUSION 234
CHAPITRE VII. DISCUSSION GENERALE : SYNTHESE, CONCLUSION
ET
PERSPECTIVES 236
7.1. SYNTHÈSE 237
7.2. CONCLUSION GÉNÉRALE 246
7.2.1. Les dynamiques biologiques et
socio-économiques mises en évidence 246
7.2.2. Des pistes d'actions pour des systèmes
d'élevage durables 249
7.2.3. Retour sur l'approche de recherche
utilisée 255
7.3. PERSPECTIVES DE RECHERCHE FUTURES 256
REFERENCES BIBLIOGRAPHIQUES 258
TABLE DES MATIERES 293
LISTE DES CARTES 308
LISTE DES ENCADRÉS 309
LISTE DES FIGURES 310
LISTE DES PHOTOGRAPHIES 313
LISTE DES TABLEAUX 314
ABSTRACT 316
RESUMÉ 317

297
ANNEXES 318
ANNEXE 1. ENQUÊTE SUR LES PRATIQUES ET LA
CATÉGORISATION DES PÂTURAGES 319
ANNEXE 2. GUIDE D'ENTRETIEN PERSONNES RESSOURCES 326
ANNEXE 3. ESPÈCES HERBACÉES CARACTÉRISTIQUES
DES GROUPEMENTS AGROSTOLOGIQUES 330
ANNEXE 4. LISTE FLORISTIQUE HERBACÉE 331
ANNEXE 5. LISTE FLORISTIQUE LIGNEUSE 335
LISTE DES CARTES

308
Carte III-1. Localisation du terroir d'étude .....45
Carte III-2. Carte des domaines phytogéographiques du
Burkina Faso 46
Carte III-3. Variation en latitude de l'isohyète 900 mm
50
Carte III-4 : Sols de la région du terroir de Kotchari
54
Carte III-5. Réseau hydrographique dans le terroir de
Kotchari ..57
Carte III-6 : L'organisation spatiale du Parc W et de sa
périphérie Burkinabé 58
Carte IV-1. Les unités paysagères pastorales du
terroir de Kotchari 83
Carte V-1. Formes d'occupation des sols à Kotchari 117
Carte VI-1. Comportement spatial des quatre troupeaux en saison
sèche froide (pendant Ku
|
fowagu ou Dabunde)
|
..191
|
|
Carte VI-2. Itinéraires du troupeau TrpC1-2G en plein
Ku fowagu .
|
194
|
|
Carte VI-3. Itinéraires du troupeau TrpC1-2G pendant
Ku tontogu
|
..197
|
|
Carte VI-4. Itinéraires du troupeau TrpC1-2G pendant A
sakoana
|
199
|
|
Carte VI-5. Itinéraires du troupeau TrpC1-2G pendant
Ku siagu
|
..200
|
|
Carte VI-6. Itinéraires du troupeau TrpC1-2G pendant
début Ku fowagu
|
.202
|
|
Carte VI-7. Itinéraires du troupeau TrpC1-2P pendant
Dabunde
|
..205
|
|
Carte VI-8. Itinéraires du troupeau TrpC1-2P pendant
Ceedu
|
. 207
|
|
Carte VI-9. Itinéraires du troupeau TrpC1-2P pendant
Kotoga
|
.209
|
|
Carte VI-10. Itinéraires du troupeau TrpC1-2P pendant
Ndungu
|
..211
|
|
Carte VI-11. Itinéraires du troupeau TrpC1-2P pendant
Yaamde
|
..212
|
|
Carte VI-12. Itinéraires du troupeau Trp pendant
Dabunde
|
..215
|
|
Carte VI-13. Itinéraires du troupeau Trp pendant Ceedu
|
..217
|
|
Carte VI-14. Itinéraires du troupeau Trp pendant
Ndungu
|
219
|
|
Carte VI-15. Itinéraires du troupeau Trp pendant
Yaamde
|
221
|
|
Carte VI-16. Itinéraires du troupeau TrpC3 pendant
Dabunde
|
.....224
|
|
Carte VI-17. Itinéraires du troupeau TrpC3 pendant
Ceedu
|
.....226
|

LISTE DES ENCADRÉS
Encadré II-1: La capacité de charge, un indicateur
changeant, peu pertinent en milieu ouvert 34
Encadré III-1. La sédentarité des pasteurs :
entre reconstitution des forces et voyage sans retour 62
Encadré IV-1. L'indice de qualité
spécifique, une valeur relative 75
Encadré V-1. Les hypothèses de recherche 110
Encadré V-2. Le Garso ou guide de transhumance
111
Encadré VI-1. Les hypothèses de recherche ..173

310
LISTE DES FIGURES
Figure I-1 : Schéma du modèle conceptuel
étudié 16
Figure II-1. Rôle du feu dans l'évolution des
savanes .28
Figure III-1 : Bilan hydrique et période active de
végétation de l'année 2009 de la province de
la Tapoa
46
Figure III-2 : Variation interannuelle de la pluviosité et
du nombre de jours de pluies dans la
commune de Tansarga entre 1997 à
2008 47
Figure III-3. Evolution des hauteurs annuelles de pluies de la
normale 1941-1970 48
Figure III-4. Evolution des hauteurs annuelles de pluies de la
normale 1971-2000 48
Figure III-5 : Evolution des superficies totales emblavées
(1999 à 2008) et des superficies en coton (2004 à 2008) dans la
commune de Tansarga 65
Figure IV-1. Dendrogramme des groupes agrostologiques des
pâturages de Kotchari 86
Figure IV-2. Les espèces caractéristiques des
différents groupements agrostologiques des pâturages .88
Figure V-1. Variation des effectifs animaux (en UBT) dans le
terroir en 2008 et 2009 119
Figure V-2. Evolution du bilan fourrager au cours des
années 2008 & 2009 121
Figure V-3. Proportions des espèces dans l'effectif de
bétail enquêté .129
Figure V-4. Graphique des variables de l'analyse factorielle des
correspondances
|
multiples .134
Figure V-5. Histogramme des valeurs propres ..135
Figure V-6. Graphique de projection des éleveurs (analyse
factorielle des correspondances
|
|
multiples) montrant leur répartition en classes
|
.137
|
|
Figure V-7. Dendrogramme des groupes d'éleveurs
|
.138
|
|
Figure V-8a. Evolution des effectifs bovins des troupeaux
enquêtés : Groupe C1
|
141
|
|
Figure V-8b. Evolution des effectifs bovins des troupeaux
enquêtés : Groupe
|
142
|
|
Figure V-8c. Evolution des effectifs bovins des troupeaux
enquêtés : Groupe C3
|
142
|
Figure V-9a. Evolution des pratiques d'allotement du troupeau
dans le Groupe C1 :
allotement du troupeau 143
Figure V-9b. Evolution des pratiques d'allotement du troupeau
dans le Groupe C1 :
motivations .143
Figure V-10a. Evolution des pratiques d'allotement du troupeau
dans le Groupe :
allotement du troupeau 143
Figure V-10b. Evolution des pratiques d'allotement du troupeau
dans le Groupe :
motivations 143
Figure V-11a. Evolution des pratiques d'allotement du troupeau
dans le Groupe C3 :
allotement du troupeau 144

Figure V-11b. Evolution des pratiques d'allotement du troupeau
dans le Groupe C3 :
motivations .144
Figure V-12a. La garde des animaux et son évolution dans
le groupe C1 : nombre de bergers 147
Figure V-12b. La garde des animaux et son évolution dans
le groupe C1 : types de bergers 147
Figure V-13a. La garde des animaux et son évolution dans
le groupe : nombre de bergers 148
Figure V-13b. La garde des animaux et son évolution dans
le groupe : types de bergers 148
Figure V-14a. La garde des animaux et son évolution dans
le groupe C3 : nombre de bergers 148
Figure V-14b. La garde des animaux et son évolution dans
le groupe C3 : types de bergers 148
Figure V-15a. Pratique de la transhumance : raisons principales
évoquées : Groupe C1 151
Figure V-15b. Pratique de la transhumance : raisons principales
évoquées : Groupe 151
Figure V-15c. Pratique de la transhumance : raisons principales
évoquées : Groupe C3 151
Figure V-16a. Milieux de pâture habituelle en saison
sèche: Groupe C1 151
Figure V-16b. Milieux de pâture habituelle en saison
sèche: Groupe .151
Figure V-16c. Milieux de pâture habituelle en saison
sèche: Groupe C3 .152
Figure V-17a. Pratiques de complémentation: Groupe C1
154
Figure V-17b. Pratiques de complémentation: Groupe 154
Figure V-17c. Pratiques de complémentation: Groupe C3
154
Figure V-18a. Composition spécifique et raciale des
troupeaux du groupe C1 : Nombre et
types d'espèces du troupeau 157
Figure V-18b. Composition spécifique et raciale des
troupeaux du groupe C1 : Nombre et types de races bovines 157
Figure V-19a. Composition spécifique et raciale des
troupeaux du groupe : Nombre et
types d'espèces du troupeau 158
Figure V-19b. Composition spécifique et raciale des
troupeaux du groupe : Nombre et types de races bovines 158
Figure V-20a. Composition spécifique et raciale des
troupeaux du groupe C3: Nombre et
types d'espèces du troupeau 158
Figure V-20b. Composition spécifique et raciale des
troupeaux du groupe C3: Nombre et types de races bovines 158
Figure VI-1. Chaîne de pâturage annuelle dans la
représentation des éleveurs peuls ..186
Figure VI-2. Chaîne de pâturage annuelle dans la
représentation des agroéleveurs gourmantchés ..187
Figure VI-3 Proportions en temps consacré aux
activités au cours de l'année par le troupeau TrpC1-2G 193

312
Figure VI-4. Proportions en temps de séjour dans les
unités pastorales au cours de l'année pour le troupeau TrpC1-2G
195
Figure VI-5. Proportions en temps consacré aux
activités au cours de l'année par le troupeau
TrpC1-2P 204
Figure VI-6. Proportions en temps de séjour dans les
unités pastorales au cours de l'année
pour le troupeau
TrpC1-2P .206
Figure VI-7. Proportions en temps consacré aux
activités au cours de l'année par le troupeau Trp .214
Figure VI-8 Proportions en temps de séjour dans les
unités pastorales au cours de l'année pour le troupeau Trp
..216
Figure VI-9. Proportions en temps consacré aux
activités au cours de l'année par le troupeau TrpC3 222
Figure VI-10. Proportions en temps de séjour dans les
unités pastorales au cours de l'année pour le troupeau TrpC3
..225
Figure VI-11. Proportions en temps consacré aux
différentes activités par les troupeaux au pâturage durant
la période de suivi 229
Figure VI-12. Comparaison entre représentations et
pratiques saisonnières de conduite des troupeaux au pâturage chez
les éleveurs gourmantchés ..232
Figure VI-13. Comparaison entre représentations et
pratiques saisonnières de conduite des troupeaux au pâturage chez
les éleveurs Peuls ..233

LISTE DES PHOTOGRAPHIES
Photo IV-1. Un faciès de savane arborée de sol
inondable (UPP2) 101
Photo IV-2. Un faciès de savane arbustive de bas glacis et
de plaine sur sols argilolimoneuse 101
Photo IV-3. Un faciès de savane arbustive de hauts glacis
et plateaux sur sols moyennement profonds (UPP4) 102
Photo IV-4. Un faciès de mosaïque
agroforestière (UPP5) ..102
Photo IV-5. Un faciès de savane arbustive sur buttes
rocheuses et cuirassées (UPP6) 103
Photo IV-6. Un faciès de savane arbustive à
herbacées annuelles à sol encroûté de l'aire
protégée (UPw4) 103
Photo IV-7 : Un faciès de savane arbustive sur
affleurements rocheux et cuirassés de l'aire
|
protégée (UPw1)
|
104
|
|
Photo V-1: Race Barbaji
|
124
|
|
Photo V-2: Race Gurmaji
|
.125
|
|
Photo V-3: Race Kiwali
|
.126
|
|
Photo V-4: Race Boboroji
|
.127
|
|
Photo V-5 : Race Jaliji près d'un campement de
transhumant
|
.127
|
|
Photo V-6: Race Gudali
|
.128
|
Photos V-7a. Deux espèces de ligneux fortement
émondés par les bergers : Ptereocarpus erinaceus 156
Photos V-7b. Deux espèces de ligneux fortement
émondés par les bergers : Afzelia africana ..156
Photo V-8. Zone de pâture traditionnelle prise d'assaut
par les agriculteurs (en arrière plan : en haut à gauche, un
champ de maïs incrustré dans un plateau gravillonaire couvert de
Loudetia togoensis) 165

314
LISTE DES TABLEAUX
Tableau II-1. Classification des formations de savane africaine
lors du colloque de Yangambi en 1956 .24
Tableau III-1. Caractéristques des séquences
climatiques sèches sur les normales 1941-1970 et 1971-2000 pour une
saison pluvieuse .49
Tableau III-2. Caractéristiques des classes de sols dans
le terroir de Kotchari 52
Tableau III-3. Densité de population dans la province de
la Tapoa .59
Tableau III-4. Effectif du cheptel et charge animale dans la
Province de la Tapoa 60
Tableau IV-1. Expression colorée des différents
thèmes du paysage dans l'image ASTER en composition colorée,
infrarouge fausse couleur utilisée pour définir les unités
paysagères pastorales du terroir de Kotchari .70
Tableau IV-2. Classes de qualité fourragère des
espèces établies avec les éleveurs de
Kotchari 76
Tableau IV-3. Unités paysagères reconnues dans le
terroir de Kotchari (hors Parc du W) 84
Tableau IV-4. Regroupement des unités paysagères de
la carte du Parc du W établie par le programme ECOPAS 84
Tableau IV-5. Correspondances entre unités de paysages
définies dans la périphérie et dans le
Parc du W .85
Tableau IV-6. Matrice de confusion entre unités
paysagères et groupement agrostologiques.87
Tableau IV-7. Espèces ligneuses dominantes des
unités paysagères pastorales du terroir de
Kotchari 92
Tableau IV-8. Valeurs de diversités intra placeau et inter
placeaux des unités paysagères pastorales du terroir de Kotchari
..94
Tableau IV-9. Diversité gamma dans le terroir de Kotchari.
Comparaison des secteurs dans et hors du Parc du W 94
Tableau IV-10. Biomasse herbacée et capacité de
charge moyenne dans les différentes unités paysagères
97
Tableau IV-11. Différentes valeurs de phytomasse et de
capacité de charge en zone soudanienne .98
Tableau IV-12. Valeurs de la densité et du recouvrement
ligneux dans les différentes unités paysagères du terroir
de Kotchari .100
Tableau IV-13. Proportions des types biologiques ligneux dans les
unités paysagères pastorales du terroir de Kotchari 100
Tableau IV-14. Qualité du fourrage dans les unités
paysagères du terroir de Kotchari : contribution des espèces par
classes d'indice spécifique et valeurs 105
Tableau V-1. Variables et modalités constitutifs du
questionnaire 112
Tableau V-2. Les paramètres significatifs dans
l'interprétation des résultats de l'AFCM..115 Tableau V-3. Types
et niveau d'occupation des terres dans le terroir de Kotchari .117

Tableau V-4. Effectifs animaux en 2008 et 2009
|
.118
|
|
Tableau V-5. Variation des charges de bétail (en UBT)
dans le terroir en 2008 et 2009......
|
119
|
|
Tableau V-6. Capacité de charge théorique globale
du terroir de Kotchari
|
..120
|
|
Tableau V-7. Données générales sur
l'échantillon enquêté
|
129
|
|
Tableau V-8. Modalités expliquant le plan factoriel
F1-F2
|
135
|
|
Tableau V-9. Caractéristiques des élevages dans
le terroir de Kotchari
|
140
|
|
Tableau V-10: Typologie des conflits dans la
périphérie du WAP
|
161
|
|
Tableau VI-1. Calendrier de suivi des troupeaux au
pâturage
|
..175
|
Tableau VI-2. Calendrier pastoral comparé de
communautés d'éleveurs dans divers terroirs en
Afrique de
l'Ouest ..179
Tableau VI-3. Les unités pastorales paysagères
distinguées les Peuls et leurs caractéristiques 180
Tableau VI-4. Les unités pastorales paysagères
distinguées par les Gourmantchés et leurs caractéristiques
181
Tableau VI-5. Correspondance entre unités
paysagères écologiques et locales 182
Tableau VI-6. Paramètres de classification des
unités paysagères pastorales : coefficients de pondération
chez les Peuls 184
Tableau VI-7. Paramètres de classification des
unités paysagères pastorales : coefficients de pondération
chez les Gourmantchés ..185
Tableau VI-8. Caractéristiques des troupeaux suivis
.189
Tableau VI-9. Temps et distance au pâturage pour le
troupeau TrpC1-2G 192
Tableau VI-10.Vitesse moyenne de déplacement du troupeau
TrpC1-2G suivant les unités et les saisons 196
Tableau VI-11.Temps et distance au pâturage pour le
troupeau TrpC1-2P 203
Tableau VI-12. Vitesse moyenne de déplacement du troupeau
TrpC1-2P suivant les unités et les saisons 206
Tableau VI-13 Temps et distance au pâturage pour le
troupeau Trp 213
Tableau VI-14. Vitesse moyenne de déplacement du troupeau
Trp suivant les unités et les saisons 216
Tableau VI-15. Temps et distance au pâturage pour le
troupeau TrpC3 ..222
Tableau VI-16. Vitesse moyenne de déplacement du troupeau
TrpC3 suivant les unités et les saisons 225
Tableau VI-17. Temps de présence au pâturage et
distances parcourues par les troupeaux au cours de la période de suivi
227
Tableau VI-18. Diverses données de distance parcourue par
le troupeau au pâturage .228
Abstract

316
ABSTRACT
Title: Forage resources, Breeders
representations and pastoral practices changing in the context of protected
area: the case of Kotchari territory on the periphery of the W biosphere
reserve in Burkina Faso
In a context of global changes which disrupt the living
conditions of the breeders of the sudanese and sahelo sudanese zone, what
decisions do they take, what are their seasonal strategies of adaptation on a
local and regional scale, how they implement them? It is this triple question
that this thesis tries to answer in the context of Kotchari area situated at
the border of the vast reserve of W transboundary biosphere and subjected to a
strong anthropological pressure.
Every year, from December, numerous transhumants breeders from
the North come together in Kotchari territory at the extreme southeastern part
of Burkina Faso, in the province of Tapoa. Locally, the pressure on lands and
resources is becoming more and more high and the sustainability of the system
seem to be compromised.
The research is done using methods of mapping / GIS, floral
analysis, quantitative and qualitative survey mostly participative.
The research showed that, despite the important animal influx
in particular during the dry and warm season, the forage resources of the
territory remain interesting with however a local extinction of species, an
important species disappearance and the beginning of shrub vegetation
development in particular on the tray pastoral landscaped units situated
outside the protected area. Four (4) breeders groups and subgroups were
recognized according to their strategies and practices implemented. It comes
out that the herd size is a determinant factor in the strategy of the breeders.
The research showed that the local categorization of environments is based on
topographic, geo morphological and functional criteria, and that the
pasturelands quality appreciation depends on factors of variable importance
according to the seasons. Finally, it is possible to understand the behavior of
the herds in the pasturelands that is the practical translation of the
technical knowledge implemented by the breeders to make exploit the
pasturelands by their herds.
Keywords: Farming system,
transhumance, pastoral landscaped units, plant diversity, pastoral practices,
representations, grazing chain, protected area, Kotchari, Burkina Faso.

Resumé
RESUMÉ
Face aux changements globaux qui perturbent les conditions de
vie des éleveurs de la zone soudanienne et sahélo-soudanienne,
quelles décisions prennent-ils, quelles sont leurs stratégies
saisonnières d'adaptation à l'échelle locale et
régionale, comment les mettent-ils en oeuvre ? C'est à cette
triple question que cette thèse tente de répondre dans le
contexte du terroir de Kotchari situé en bordure de la vaste
réserve de biosphère transfrontalière du W et soumis
à une forte pression anthropique.
Chaque année à partir de décembre de
nombreux éleveurs transhumants venant du nord affluent dans le terroir
de Kotchari à l'extrême sud-est du Burkina Faso, dans la province
de la Tapoa. Localement, la pression sur les terres et les ressources se fait
de plus en plus forte et la durabilité du système parait
compromise.
La recherche a été conduite en faisant appel
à des méthodes de cartographie/SIG, d'analyse floristique,
d'enquêtes quantitatives et qualitatives le plus souvent
participatives.
La recherche a montré que, malgré l'afflux
important d'animaux en saison sèche notamment chaude, les ressources
fourragères du terroir restent intéressantes avec toutefois une
extinction locale d'espèces, une banalisation de la flore et un
début d'embuissonnement en particulier sur les unités
paysagères pastorales de plateaux en dehors de l'aire
protégée. On a pu reconnaître quatre (4) groupes et
sous-groupes d'éleveurs en fonction des stratégies
développées et des pratiques mises en oeuvre. Il ressort que la
taille du troupeau est un facteur déterminant dans leurs
stratégies. La recherche a montré, par ailleurs, que la
catégorisation locale des milieux s'appuie sur des critères
topographiques, géomorphologiques et fonctionnels et que
l'appréciation en termes de qualité de pâturage
dépend de facteurs d'importance variable selon les saisons. Enfin, il
est possible de comprendre le comportement des troupeaux au pâturage qui
est la traduction pratique des savoirs techniques que mettent en oeuvre les
éleveurs pour faire exploiter les milieux par leurs troupeaux.
Mots clés : Système
d'élevage, transhumance, unités paysagères pastorales,
diversité végétale, pratiques pastorales,
représentations, chaîne de pâturage, aire
protégée, Kotchari, Burkina Faso.

318
ANNEXES

319
Annexe 1. Enquête sur les pratiques et la
catégorisation des pâturages
I. Généralités et
Caractéristiques socio-économiques
Village (quartier, hameau) Date
Nom de l'enquêté Ethnie Age
Situation matrimoniale : Marié_____ ; Célibataire
Veuf
Si marié, nombre de femmes : 1 ; 2 ; Plus de 2
Effectif du ménage ; Nombre de jeunes (8-18 ans) ; dont
Filles et
.Garçons
Nombre d'hommes adultes (18-40 ans)
Relation au troupeau: Propriétaire ____ ; Parent du
propriétaire
Principale activité : Agriculture_____ ; Elevage ;
Commerce ;
Autres (préciser)
Activités secondaires : Agriculture_____ ; Elevage ;
Commerce ;
Autres (préciser)
Rôle de l'élevage dans la subsistance du
ménage : Très important____ ; Important_____
Moyen____ ; Faible_____
Part du revenu monétaire tiré de l'élevage
(en %) : 0-25___ ; 25-50____ ; 50-75____ ; > 75
____
Autres sources importantes de revenus
II. Pratiques de configuration du troupeau
1. Constitution / Acquisition
1.1. Si propriétaire, depuis quand son troupeau a
été constitué ?
1.2. Comment le troupeau s'est mis en place : a. Héritage
; b. Achat; c. Don;
d. Emprunt
1.3. Par quelle espèce ? a. Bovin; b. Ovin; c. Caprins; d.
porcins
1.4. Pourquoi, par cette / ces espèce (s) ?
1.5. Depuis quand rencontre t-on l'espèce bovine dans le
troupeau ?
1.6. Quelles étaient les premières races de cette
espèce?
1.7. Quelles races sont rencontrées de nos jours dans le
troupeau ?
1.8. Composition spécifique du troupeau et nombre de
têtes
|
1.8.1. Maintenant
|
1.8.2. Il y a dix ans
|
1.8.3. Il y a 20 ans et plus
|

|
Bovins
|
Ovins
|
Caprins
|
Porcins
|
Bovins
|
Ovins
|
Caprins
|
Porcins
|
Bovins
|
Ovins
|
Caprins
|
Porcins
|
|
Eff.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Ra
|
|
|
|
|
|
|
|
|
|
|
|
|
Eff. = effectif ; Ra = races
Races : a = Barbaji ; b = Gurmadji ; c =
Kiwali ; d = Jaliji; e = Boboroji ; f = Gudali
Expliquez ces changements dans les races bovines du troupeau
2. Agrégation
2.1. Garde d'animaux d'autrui
2.1.1. Gardez-vous souvent des animaux qui ne sont pas de vous ?
a. Oui; b. Non;
2.1.2. Si oui, sont-ils de qui ? a. Fils ; b. Frère ; c.
Epouse; d. Autres parents; e. Autres
2.1.3. Si autres, préciser si : a. éleveur; b.
Autre secteur d'activité; c. commerçants;
d. Fonctionnaires
2.1.4. Quelles espèces vous sont données en garde ?
a. Bovins; b. Ovins; c. Caprins
2.1.5. Pendant combien de temps? a. Un bout de temps; b. Une
année; c. Plusieurs années
2.1.6. Si un bout de temps, préciser la saison :
a. Ndungu (Ku siagu); b. Guiande (D. Ku
fowagu);
c. Dabunde (F. Ku fowagu); d. Ceedu (Ku
tontogu); e. Kotoga/Korse (A sakoana)
2.1.7. Quels sont le plus souvent les termes du contrat ?
|
Questions
|
Maintenant
|
Il y a dix ans
|
Il y a 20 ans et plus
|
|
2.1.1
|
|
|
|
|
2.1.2
|
|
|
|
|
2.1.3
|
|
|
|
|
2.1.4
|
|
|
|
|
2.1.5
|
|
|
|
|
2.1.6
|
|
|
|
|
2.1.7
|
|
|
|
2.2. Confiage d'animaux à autrui
2.2.1. Avez-vous déjà confié des animaux en
garde ? a. Oui; b. Non
2.2.2. Si oui, quelle espèce ? a. Bovins; b. Ovins ; c.
Caprins
2.2.3. Quand ? a. Un bout de temps; b. Une année; c.
Plusieurs années
2.2.4. Si un bout de temps, préciser la saison :
a. Ndungu (Ku siagu); b. Guiande (D. Ku
fowagu);
c. Dabunde (F. Ku fowagu); d. Ceedu (Ku
tontogu); e. Kotoga/Korse (A sakoana)

321
2.2.5. A qui ? a. Fils; b. Frère; c. Autres parents; d.
Autre éleveur
2.2.6. Pourquoi ?
2.2.7. Quels étaient les termes du contrat?
|
Questions
|
Maintenant
|
Il y a dix ans
|
Il y a 20 ans et plus
|
|
2.2.1
|
|
|
|
|
2.2.2
|
|
|
|
|
2.2.3
|
|
|
|
|
2.2.4
|
|
|
|
|
2.2.5
|
|
|
|
|
2.2.6
|
|
|
|
|
2.2.7
|
|
|
|
3. Allottement
3.1. Nombre de lots dans le troupeau: ;
3.2. Si plus d'un lot, sur quelle (s) base (s) (ou
critères) les lots sont constitués ? a. Age;
b. Sexe; c. Etat physiologique (malade, pregnante); d. selon la
propriété
3.3. A quoi répond cette pratique ?
3.4. Durée de la séparation des lots ; a.
temporaire ; b. définitive
3.5. Si temporaire, donner la période et la durée
|
Questions
|
Maintenant
|
Il y a dix ans
|
Il y a 20 ans et plus
|
|
3.1
|
|
|
|
|
3.2
|
|
|
|
|
3.3.
|
|
|
|
|
3.4.
|
|
|
|
|
3.5.
|
|
|
|
Expliquer ces changements ?
III. Pratiques de conduite du troupeau
1. Les animaux sont-ils gardés, en saison sèche ?
a. Oui; b. Non
2. Les animaux sont-ils gardés en saison humide ? a. Oui;
b. Non
3. Si oui, sont-ils conduits : a. séparément; b.
en groupe?
4. Quand les animaux sont-ils conduits au pâturage ? a. La
nuit; b. Le jour
5. Où sont-ils conduits en saison sèche ?
a. Pâturages communs b. Parcelles du propriétaire;
c. Toutes parcelles du village

6. Qui garde le troupeau le jour? a. Fils; b. Fille; c. Autre
parent
7. Si garde de nuit, est-ce le même berger que celui du
jour ? a. Oui; b. Non
8. Si non pourquoi?
|
Questions
|
Maintenant
|
Il y a dix ans
|
Il y a 20 ans et plus
|
|
1
|
|
|
|
|
2
|
|
|
|
|
3.
|
|
|
|
|
4
|
|
|
|
|
5
|
|
|
|
|
6
|
|
|
|
|
7
|
|
|
|
|
8
|
|
|
|
9. Pourquoi ces changements
IV Pratiques d'alimentation
/Complémentation
1. Les animaux sont-ils alimentés : a. au pâturage
naturel exclusif; b. Complémentés?
2. Si complémentation, comment ? a. Aliments produits ou
conditionnés dans l'exploitation b. Aliments achetés ; c. Les
deux
3. Nature des compléments : a. Résidus de culture
(fanes, paille de riz, tige de sorgho et mil) b. Fourrage naturel (paille)
collecté et stocké c. Cultures fourragères
4. Si collecte et culture fourragère, quelles
espèces sont concernées ?
5. Quels animaux sont complémentés ? a. malades ;
b. fatigués/malnutris ;
c. allaitantes ; d. prégnantes ;
6. Quand (période de l'année) la
complémentation a-t-elle lieu ?
a. Ndungu (Ku siagu); b. Guiande (D. Ku
fowagu);
c. Dabunde (F. Ku fowagu); d. Ceedu (Ku
tontogu); e. Kotoga/Korse (A sakoana)
|
Questions
|
Maintenant
|
Il y a dix ans
|
Il y a 20 ans et plus
|
|
1
|
|
|
|
|
2
|
|
|
|
|
3.
|
|
|
|
|
4
|
|
|
|
|
5
|
|
|
|
|
6
|
|
|
|

323
V. Pratiques de déplacements /
Mobilité1. Vos animaux sont-ils toujours dans le terroir
villageois ? a. Oui ; b. Non
2. Pourquoi ?
3. Si non, quand ne sont-ils pas dans le terroir villageois ?
a. Ndungu (Ku siagu); b. Guiande (D. Ku
fowagu);
c. Dabunde (F. Ku fowagu); d. Ceedu (Ku
tontogu); e. Kotoga/Korse (A sakoana)
4. Où les trouve-t-on à cette période ? a.
Dans les terroirs environnants (Petite transhumance) b. Dans les terroirs
éloignés (Grande transhumance)
5. Fréquence des déplacements: a. chaque
année ; b. occasionnel
6. Si occasionnel, préciser les circonstances
7. Animaux et lots concernés: Si plus d'un lot, quel lot
est concerné ? a. Tous les lots; b. un lot
8. Dans les lots, a. tous les animaux; b. certains animaux
9. Si certains animaux, quels animaux : a. espèces : a1.
bovins ; a2. ovins ; a3. caprins
b. âge : b1 : jeunes ; b2. moins
jeunes ; b3. vieux
c. état physiologique : c1. bien portants ; c2. non
allaitant ; c3. Autres (préciser)
10. Qui décide des déplacements : a.
Propriétaire; b. Garso; c. Rugga; d. Bergers
|
Questions
|
Maintenant
|
Il y a dix ans
|
Il y a 20 ans et plus
|
|
1
|
|
|
|
|
2
|
|
|
|
|
3.
|
|
|
|
|
4
|
|
|
|
|
5
|
|
|
|
|
6
|
|
|
|
|
7
|
|
|
|
|
8
|
|
|
|
|
9
|
|
|
|
|
10
|
|
|
|
Quelles sont les motivations de ces changements?
Quels impacts et conséquences de ces changements dans
l'évolution du troupeau ?

Quelle est la relation de ces changements avec les aires
protégées ?
VII. Connaissance / catégorisation et
fréquentation des pâturages
7.1. Quels sont les critères utilisés pour
évaluer un pâturage et pourquoi ? C1 Explication
Explication
C3 Explication
7.2. Hiérarchie entre les critères
7.3. Sur la base de ces critères, qu'est-ce qu'un :
Excellent pâturage
Très bon pâturage
Bon pâturage
Pâturage moyen
Pâturage médiocre
7.4. Pâturages rencontrés à Kotchari, leurs
caractéristiques et leur qualité pastorale selon la saison
|
Pâturages (noms vernaculaires)
|
Caractéristiques (sols, végétation,
géomorphologie, etc.)
|
Qualité pastorale
|
7.5. Calendrier de fréquentation de ces pâturages et
raisons
|
Saisons
|
Pâturages fréquentés
|
Justification
|
1. Ndungu (Ku siagu)
|
|
|
|
|
2. Guiande (D. Ku fowagu)
|
|
|
|
3. Dabunde (F. Ku fowagu)
4. Ceedu (Ku tutongu)
5. Kotoga / Korsê (A sakoana)

325
7.6. Existe-t-il de raisons particulières pouvant
empêcher la fréquentation d'un pâturage jugé de
qualité ? a. Oui ; b. Non .
7.7. Si oui, lesquelles ?
7.8. S'il s'agit de menaces, précisez leur nature et la
distance de sécurité par rapport à chacune d'elles .
VIII. Poches de ressources
8. 1. Quels sont parmi les pâturages
énumérés, ceux qui sont déterminants à la
survie du bétail et à quelles périodes
8.2. Pourquoi ?

Annexe 2. Guide d'entretien perso nnes
ressources Terroir Enquêteur Date
Nom Personne ressource Qualité
1. Histoire personnelle dans l'élevage
2. Evolution des pratiques et de son troupeau :
insister sur les changements dans les pratiques pastorales
(distances parcourues, milieux fréquentés, changements
d'espèces ou de races animales, préparatifs si transhumance,
etc.)
3. Connaissances des pâturages et des
espèces indicatrices de richesse, de pauvreté ou de
dégradation pastorale (critères de choix des milieux
pâturés ou évités, noms vernaculaires donnés
à ces milieux, espèces végétales ligneuses ou
herbacées indicatrices de bons, mauvais pâturages ou de
pâturages en dégradation, noms donnés à ces
espèces et connaissance de leur biologie/écologie).
4. Risques encourus par les éleveurs et leurs
animaux
4.1. Risques sanitaires Existence de risques
sanitaires ?
Comment sont-ils appréhendés ?
Pathologies courantes
Dans quels types de milieux (appellation vernaculaire) sont-ils
rencontrés ?

327
Ces risques sont-ils encore importants de nos jours ?
_____
Quelle importance dans le choix des parcours ou des
itinéraires de transhumance ?
4.2. Conflits avec agriculteurs
Quels types, quelle fréquence et quelle ampleur ?
Avec quel profil d'agriculteur ? (Agriculteurs purs ?,
agropasteurs autochtones, allochtones ?, etc.)
Quelle tendance et quels impacts sur l'élevage et les
stratégies d'élevage ?
4.3. Conflits avec conservateurs
Quelle cause, quelle fréquence et quelle ampleur ?
Endroits où ces conflits sont rencontrés ou
fréquents, quelle explication ?
__
Quelle tendance et quels impacts sur l'élevage et les
stratégies d'élevage ?
4.4. Faune sauvage
Constitue t-elle une menace de nos jours ?
Comment : risque sanitaire ? Prédation ?
A quels endroits précis du terroir ou de la région
?

Quelle tendance et quels impacts sur l'élevage et les
stratégies d'élevage ?
4.5. Autres risques
Nature et importance du risque sur l'élevage et les
stratégies
5. Importance de la pâture dans la « grande
et lointaine brousse » (comme rite initiatique, dans la vie de la
communauté). Quelle importance dans la formation du jeune adolescent ?
__
6. Perception des changements globaux Quelle
lecture sur :
6.1. L'état des parcours dans la région ?
_
6.2. La dynamique agricole (défrichements, et occupation
de l'espace, coton culture, etc.) ?
6.3. Les mesures de conservation (aires
protégées)
6.4. Les attitudes des agriculteurs ou des populations des
terroirs d'accueil
7. Impacts de ces changements sur les pratiques
pastorales (au quotidien, sur la durée).
Dates/périodes importantes dans le
changement : - du calendrier pastoral
- de la composition (espèces/races) du troupeau
- de la taille du troupeau

329
- de l'organisation et le calendrier de la transhumance
(changements dans les dates de départ, dans la durée de la
campagne, direction et itinéraires, distances parcourues, etc.)
Quelles perspectives si maintient de cette tendance de
changement ?
Quel devenir pour l'élevage pastoral si cette
tendance de changement se maintient ? (Faire ressortir les craintes et
les mesures préventives prises ou envisagées)
8. Place des aires protégées dans les
stratégies pastorales Fréquentation des aires
protégées et rythme
Motivations
Période (s)
Conscience des risques encourus et dispositions prises
Quelle évolution de l'importance des aires
protégées dans les stratégies de l'éleveur
(repérer les dates ou périodes clés)
Les aires protégées, un lieu de passage
obligé ? (Pourquoi ?)
9. Le feu pastoral Quelle importance pour le
pasteur ?
Quelle période le pratique t-il et pourquoi ?
Quelle période est indiquée et pourquoi ?
Quels effets (positifs et négatifs) sur le pâturage
?

Annexe 3. Espèces herbacées
caractéristiques des groupements
agrostologiques
|
Acronyme
|
Espèces
|
Acronyme
|
Espèces
|
|
Abhi
|
Abildgaardia hispidula
|
Hysm
|
Hyparrhenia smithiana
|
|
Anch
|
Andropogon chinensis
|
Hysu
|
Hyparrhenia subplumosa
|
|
Anga
|
Andropogon gayanus
|
Kypu
|
Kyllinga pumila
|
|
Anps
|
Andropogon pseudapricus
|
Losi
|
Loudetia simplex
|
|
Ante
|
Andropogon tectorum
|
Loto
|
Loudtia togoensis
|
|
Arad
|
Aristida adscensionis
|
Meco
|
Melochia corchorifolia
|
|
Arfu
|
Aristida funiculata
|
Miin
|
Microchloa indica
|
|
Brju
|
Brachiaria jubata
|
Orlo
|
Oryza longistaminata
|
|
Daae
|
Dactyloctenium aegyptium
|
Scex
|
Schizachyrium exile
|
|
Diam
|
Diheteropogon amplectens
|
Scsa
|
Schizachyrium sanguineum
|
|
Dide
|
Digitaria debilis
|
Sepa
|
Setaria pumila
|
|
Ecco
|
Echinochloa colona
|
Siac
|
Sida Acuta
|
|
Ecst
|
Echinochloa stagnina
|
Sobi
|
Sorghastrum bipennatum
|
|
Erte
|
Eragrostis tenella
|
Sppy
|
Sporobolus pyramidalis
|
|
Ertr
|
Eragrostis tremula
|
|
|

331
Annexe 4. Liste floristique herbacée
|
Espèces
|
Familles
|
Type fourrager
|
Indice de
qualité
|
|
Abildgaardia hispidula (Vahl) Kunth
|
Cyperaceae
|
Au
|
0
|
|
Acanthospermum hispidum DC.
|
Asteraceae
|
Au
|
1
|
|
Acroceras zizanioides Kunth.
|
Poaceae
|
Ga
|
2
|
|
Alternanthera nodiflora R. Br.
|
Amaranthaceae
|
Au
|
0
|
|
Althenanthera sessilis (Linn) R. Br
|
Amaranthaceae
|
Au
|
0
|
|
Alysicarpus glumaceus (Vahl.) DC.
|
Fabaceae
|
Le
|
2
|
|
Alysicarpus ovalifolius (Schum. et Thonn.) J. Leonard
|
Fabaceae
|
Le
|
3
|
|
Alysicarpus rugosus (Willd.) DC.
|
Fabaceae
|
Le
|
1
|
|
Ampelocissus grantii (Ba.) Planch.
|
Vitaceae
|
Au
|
0
|
|
Andropogon ascinodis C.B. Cl.
|
Poaceae
|
Gv
|
4
|
|
Andropogon fastigiatus Sw.
|
Poaceae
|
Ga
|
3
|
|
Andropogon gayanus Kunth
|
Poaceae
|
Gv
|
4
|
|
Andropogon pseudapricus Stapf
|
Poaceae
|
Ga
|
3
|
|
Andropogon tectorum Schumach. & Thonn.
|
Poaceae
|
Gv
|
4
|
|
Aristida adscensionis L.
|
Poaceae
|
Ga
|
1
|
|
Aristida funiculata Trin. & Rupr.
|
Poaceae
|
Ga
|
1
|
|
Aspilia bussei O.Hoffm. & Muschl.
|
Asteraceae
|
Au
|
0
|
|
Aspilia kotschyi (Sch. Bip. Ex Hoxhst.) Oliv.
|
Asteraceae
|
Au
|
1
|
|
Boerhavia diffusa L.
|
Nyctagynaceae
|
Au
|
0
|
|
Brachiaria deflexa (Schum.) Hubb
|
Poaceae
|
Ga
|
2
|
|
Brachiaria distichophylla (Trin.) Stapf
|
Poaceae
|
Ga
|
2
|
|
Brachiaria jubata (Fig. & DeNot.) Stapf
|
Poaceae
|
Ga
|
1
|
|
Brachiaria lata (Schumach.) C. E. Hubbard
|
Poaceae
|
Ga
|
3
|
|
Cassia mimosoides L.
|
Caesalpiniaceae
|
Le
|
1
|
|
Cassia nigricans Vahl.
|
Caesalpiniaceae
|
Le
|
2
|
|
Cassia obtusifolia L.
|
Caesalpiniaceae
|
Le
|
2
|
|
Cassia occidentalis Linn.
|
Caesalpiniaceae
|
Le
|
1
|
|
Chasmopodium codatum Staff.
|
Poaceae
|
Ga
|
2
|
|
Chloris pilosa Schum. & Thonn.
|
Poaceae
|
Ga
|
1
|
|
Chrysanthellum americanum (Lin.) Vatke
|
Asteraceae
|
Au
|
0
|
|
Cissus quadrangularis Linn.
|
Vitaceae
|
Au
|
0
|
|
Cissus waterlotii A. Chev.
|
Vitaceae
|
Au
|
0
|
|
Cleome viscosa L.
|
Capparidaceae
|
Au
|
0
|
|
Cochlospermum planchoni Hook. f.
|
Cochlospermaceae
|
Au
|
1
|
|
Cochlospermum tinctorium A. Rich.
|
Cochlospermaceae
|
Au
|
0
|
|
Commelina benghalensis Linn.
|
Commelinaceae
|
Au
|
1
|
|
Commelina capitata Benth
|
Commelinaceae
|
Au
|
1
|
|
Commelina diffusa Burm. f.
|
Commelinaceae
|
Au
|
1
|

Commelina forskalaei Vahl
|
Commelinaceae
|
Au
|
2
|
|
Corchorus olitorius L.
|
Tiliaceae
|
Au
|
2
|
|
Corchorus tridens Linn.
|
Tiliaceae
|
Au
|
2
|
|
Crinum ornatum (Ait.) Bury
|
Amaryllidaceae
|
Au
|
0
|
|
Crotalaria goreensis Guill. & Perr
|
Fabaceae
|
Le
|
0
|
|
Crotalaria mucronata Desv.
|
Fabaceae
|
Le
|
0
|
|
Crotalaria ononoides Benth.
|
Fabaceae
|
Le
|
0
|
|
Crotalaria palida Ait.
|
Fabaceae
|
Le
|
0
|
|
Crotalaria senegalensis (Pers.) Bacle ex DC.
|
Fabaceae
|
Le
|
0
|
|
Ctenium elegans Kunth
|
Poaceae
|
Ga
|
2
|
|
Ctenium newtonii Hack.
|
Poaceae
|
Ga
|
2
|
|
Cucumis melo Linn.
|
Cucurbitaceae
|
Au
|
2
|
|
Cyanotis lanata Benth.
|
Commelinaceae
|
Au
|
3
|
|
Cymbopogon giganteus Chiov. var giganteus
|
Poaceae
|
Gv
|
2
|
|
Cyperus amabilis Vahl.
|
Cyperaceae
|
Au
|
0
|
|
Cyperus difformis Linn.
|
Cyperaceae
|
Au
|
0
|
|
Cyperus exaltatus Retz.
|
Cyperaceae
|
Au
|
0
|
|
Cyperus haspan Linn.
|
Cyperaceae
|
Au
|
0
|
|
Cyperus iria L.
|
Cyperaceae
|
Au
|
0
|
|
Dactyloctenium aegyptium (L.) P. de B.
|
Poaceae
|
Ga
|
2
|
|
Digitaria debilis Willd.
|
Poaceae
|
Ga
|
3
|
|
Digitaria horizontalis Willd.
|
Poaceae
|
Ga
|
3
|
|
Diheteropogon amplectens (Nees) WD Clayton
|
Poaceae
|
Gv
|
3
|
|
Diheteropogon hagerupii Hitchc.
|
Poaceae
|
Ga
|
3
|
|
Echinochloa colona (L.) Link
|
Poaceae
|
Ga
|
3
|
|
Echinochloa stagnina (Retz.) P. Beauv.
|
Poaceae
|
Gv
|
2
|
|
Eleusine indica (L.) Gaernt.
|
Poaceae
|
Ga
|
2
|
|
Eragrostis aspera (Jacq.) Nees
|
Poaceae
|
Ga
|
1
|
|
Eragrostis cilianensis (All.) Vign. ex Janchen
|
Poaceae
|
Ga
|
2
|
|
Eragrostis ciliaris (L.) R. Br.
|
Poaceae
|
Ga
|
2
|
|
Eragrostis tenella (Linn.) P. Beauv. Ex Roem
|
Poaceae
|
Ga
|
2
|
|
Eragrostis tremula Hochst. ex Steud.
|
Poaceae
|
Ga
|
2
|
|
Eragrostis turgida (Schum.) De Wild.
|
Poaceae
|
Ga
|
1
|
|
Euphorbia hirta Linn.
|
Euphorbiaceae
|
Au
|
0
|
|
Euphorbia polychmoides
|
Euphorbiaceae
|
Au
|
0
|
|
Fimbristylis ferrugineaVahl
|
Cyperaceae
|
Au
|
0
|
|
Hibiscus asper Hook. f.
|
Malvaceae
|
Au
|
1
|
|
Hygrophila auriculata (Schumach.) Heine
|
Acanthaceae
|
Au
|
0
|
|
Hygrophila barbata (Nees) T. Anders.
|
Acanthaceae
|
Au
|
0
|
|
Hyparrhenia rufa (Nees) Stapf
|
Poaceae
|
Gv
|
3
|
|
Hyparrhenia smithiana (Hook. F.) Stapf.
|
Poaceae
|
Gv
|
3
|
|
Hyparrhenia subplumosa (Hook. f.) Stapf
|
Poaceae
|
Gv
|
3
|

333
Hypparhenia involucrata Stapf
|
Poaceae
|
Ga
|
2
|
|
Hyptis spicigera Lam
|
Lamiaceae
|
Au
|
1
|
|
Indigofera bracteolata DC.
|
Fabaceae
|
Le
|
0
|
|
Indigofera macrocalyx Guill. & Perr.
|
Fabaceae
|
Le
|
0
|
|
Indigofera trichopoda Lepr. ex Guill. & Perr.
|
Fabaceae
|
Le
|
0
|
|
Ipomoea eriocarpa R. Br.
|
Convolvulaceae
|
Au
|
2
|
|
Ipomoea heterotricha F. Didr.
|
Convolvulaceae
|
Au
|
1
|
|
Ipomoea repens Lam.
|
Convolvulaceae
|
Au
|
1
|
|
Ipomoea vagans Bak.
|
Convolvulaceae
|
Au
|
1
|
|
Kaempferia aethiopica (Scheinf.) Solms-Laub.
|
Zingiberaceae
|
Au
|
1
|
|
Kyllinga erecta Schum.
|
Cyperaceae
|
Au
|
0
|
|
Kyllinga pumila Michx
|
Cyperaceae
|
Au
|
0
|
|
Kyllinga squamulata Thonn. Ex Vahl.
|
Cyperaceae
|
Au
|
0
|
|
Lepidagathis anobrya Nees
|
Acanthaceae
|
Au
|
1
|
|
Leucas martinicensis (Jacq.) Ait. F.
|
Lamiaceae
|
Au
|
1
|
|
Loudetia simplex (Nees) C. E. Hubbard
|
Poaceae
|
Gv
|
2
|
|
Loudetia togoensis (Pilger) C.E. Hubbard
|
Poaceae
|
Ga
|
2
|
|
Loudetiopsis kerstingii (Pelger) Conert
|
Poaceae
|
Ga
|
2
|
|
Ludwigia hyssopifolia (G. Don.) Exell
|
Oenotheraceae
|
Au
|
0
|
|
Melochia corchorifolia Linn.
|
Sterculiaceae
|
Au
|
1
|
|
Microchloa indica (L.) Beauv.
|
Poaceae
|
Ga
|
1
|
|
Mitracarpus scaber Zucc.
|
Rubiaceae
|
Au
|
0
|
|
Mollugo nudicaulis Lam.
|
Molluginaceae
|
Au
|
0
|
|
Monechma ciliatum (Jacq) Milne Redhead
|
Acanthaceae
|
Au
|
0
|
|
Oryza barthi A.Chev.
|
Poaceae
|
Ga
|
2
|
|
Oryza longistaminata Chev. & Roehr.
|
Poaceae
|
Ga
|
2
|
|
Pandiaka involucrata Hook.f.
|
Amaranthaceae
|
Au
|
0
|
|
Panicum anabaptistum Steud.
|
Poaceae
|
Ga
|
1
|
|
Panicum kerstingii Mez.
|
Poaceae
|
Ga
|
3
|
|
Panicum laetum Kunth.
|
Poaceae
|
Ga
|
2
|
|
Panicum repens L.
|
Poaceae
|
Ga
|
2
|
|
Paspalum orbiculare Forst. f.
|
Poaceae
|
Ga
|
3
|
|
Pennisetum pedicellatum Trin.
|
Poaceae
|
Ga
|
3
|
|
Pennisetum subangustum (Schumach.) Stapf et Hubb.
|
Poaceae
|
Ga
|
4
|
|
Pennisetum violaceum (Lam.) Rich.
|
Poaceae
|
Ga
|
3
|
|
Rottboellia exaltata Linn. f.
|
Poaceae
|
Ga
|
2
|
|
Sacciolepis africana Hubb. & Snowden
|
Poaceae
|
Ga
|
1
|
|
Schizachyrium brevifolium (Sw.) Nees ex Büse
|
Poaceae
|
Ga
|
2
|
|
Schizachyrium exile (Hochst) Pilger
|
Poaceae
|
Ga
|
2
|
|
Schizachyrium sanguineum
|
Poaceae
|
Gv
|
3
|
|
Schoenefeldia gracilis Kunth
|
Poaceae
|
Ga
|
3
|

Sclerea sphaerocarpa (E. Rob.) Napper
|
Cyperaceae
|
Au
|
1
|
|
Scleria pergracilis (Nees) Kunth
|
Cyperaceae
|
Au
|
1
|
|
Scleria verrucosa Willd.
|
Cyperaceae
|
Au
|
1
|
|
Scoparia dulcis L.
|
Scrophulariaceae
|
Au
|
0
|
|
Setaria pallide-fusca (Schumach.) Stapf et C.E. Hubb.
|
Poaceae
|
Ga
|
3
|
|
Sida acuta Burm.f.
|
Malvaceae
|
Au
|
1
|
|
Sida alba L.
|
Malvaceae
|
Au
|
0
|
|
Sida cordifolia Linn
|
Malvaceae
|
Au
|
0
|
|
Sida linifolia Juss. ex Cav.
|
Malvaceae
|
Au
|
0
|
|
Sida rhombifolia L.
|
Malvaceae
|
Au
|
0
|
|
Sorgastrum bipennatum (Hack.) Pilger
|
Poaceae
|
Ga
|
3
|
|
Spermacoce filifolia (Schum. et Thonn.) Lebrun et
Stork.
|
Rubiaceae
|
Au
|
0
|
|
Spermacoce radiata (D.C.) Sieber ex Hiern
|
Rubiaceae
|
Au
|
0
|
|
Spermacoce ruelliae DC
|
Rubiaceae
|
Au
|
0
|
|
Spermacoce stachydea DC.
|
Rubiaceae
|
Au
|
0
|
|
Sporobolus festivus Hochst. ex A.Rich.
|
Poaceae
|
Gv
|
2
|
|
Sporobolus pyrimidalis P. Beauv.
|
Poaceae
|
Gv
|
2
|
|
Striga hermontheca (Del.) Benth.
|
Scrophulariaceae
|
Au
|
0
|
|
Stylochiton hypogaeus Lepr.
|
Araceae
|
Au
|
0
|
|
Tephrosia linearis (Willd.) Pers.
|
Fabaceae
|
Le
|
2
|
|
Tephrosia pedicellata Bak.
|
Fabaceae
|
Le
|
3
|
|
Tribulus terrestris Linn.
|
Zygophyllaceae
|
Au
|
1
|
|
Tripogon minimus (A. Rich.) Hochst. ex Steud.
|
Poaceae
|
Ga
|
1
|
|
Triumfetta rhomboidea Jack.
|
Tiliaceae
|
Au
|
0
|
|
Vernonia nigritiana Oliv. et Hiern
|
Asteraceae
|
Au
|
0
|
|
Vetiveria nigritana
|
Poaceae
|
Gv
|
2
|
|
Waltheria indica Linn.
|
Sterculiaceae
|
Au
|
0
|
|
Wissadula amplissima (Linn.) R. E. Fries
|
Malvaceae
|
Au
|
0
|
|
Zornia glochidiata Reichb. ex DC.
|
Fabaceae
|
Le
|
3
|
Ga, graminées annuelles ; Gv, graminées vivaces ;
Le, légumineuses ; Au, diverses autres espèces
Annexe 5. Liste floristique ligneuse
|
Espèces
|
Familles
|
Types biologiques
|
|
Acacia ataxacantha DC.
|
Mimosaceae
|
arbuste
|
|
Acacia gourmaensis A.Chev.
|
Mimosaceae
|
arbuste
|
|
Acacia hockii De Wild.
|
Mimosaceae
|
arbuste
|
|
Acacia macrostachya Reichenb. ex Benth.
|
Mimosaceae
|
arbuste
|
|
Acacia polyacantha Willd.
|
Mimosaceae
|
arbre
|
|
Acacia sieberiana DC
|
Mimosaceae
|
arbre
|
|
Adansonia digitata Linn.
|
Bombacaceae
|
grand arbre
|
|
Albizzia chevalieri Harms
|
Mimosaceae
|
arbre
|
|
Annona senegalensis Pers.
|
Annonaceae
|
sous-arbuste
|
|
Anogeissus leiocarpus (DC.) Wall.
|
Combretaceae
|
grand arbre
|
|
Balanites aegyptiaca (Linn.) Del.
|
Balanitaceae
|
arbuste
|
|
Bombax costatum Peller. ex Vuillet
|
Bombacaceae
|
grand arbre
|
|
Bridelia ferruginea Benth.
|
Euphorbiaceae
|
arbuste
|
|
Burkea africana Hook.f.
|
Caesalpiniaceae
|
grand arbre
|
|
Calotropis procera (R. Br.)
|
Asclepiadaceae
|
sous-arbuste
|
|
Capparis corymbosa Lam.
|
Capparidaceae
|
sous-arbuste
|
|
Cassia sieberiana DC.
|
Caesalpiniaceae
|
arbuste
|
|
Cissus populnea Guill. ex Perr.
|
Vitaceae
|
sous-arbuste
|
|
Combretum aculeatum Vent.
|
Combretaceae
|
sous-arbuste
|
|
Combretum collinum Fresen.
|
Combretaceae
|
arbuste
|
|
Combretum fragans F.Hoffm
|
Combretaceae
|
arbuste
|
|
Combretum glutinosum Perr. ex DC.
|
Combretaceae
|
arbuste
|
|
Combretum micranthum G. Don.
|
Combretaceae
|
arbuste
|
|
Combretum molle R. Br. ex G. Don
|
Combretaceae
|
arbuste
|
|
Combretum nigricans Lepr. Ex Guill. et Perr.
|
Combretaceae
|
arbuste
|
|
Crossopteryx febrifuga (Afzel. ex G. Don) Benth.
|
Rubiaceae
|
arbuste
|
|
Daniellia oliveri (R.) Hutch. et dalz.
|
Caesalpiniaceae
|
arbre
|
|
Detarium microcarpum Guill. ex Perr.
|
Caesalpiniaceae
|
arbuste
|
|
Dichrostachys cinerea (L.) Wight et Arn.
|
Mimosaceae
|
sous-arbuste
|
|
Diospyros mespiliformis Hochst. ex A. Rich.
|
Ebenaceae
|
arbre
|
|
Entada africana Guill. ex Perr.
|
Mimosaceae
|
arbre
|
|
Erythrina senegalensis DC.
|
Fabaceae
|
arbre
|
|
Feretia apodanthera Del.
|
Rubiaceae
|
sous-arbuste
|
|
Ficus capensis Thumb.
|
Moraceae
|
arbre
|
|
Ficus gnaphallocarpa (Miq.) Steud. Ex A. Rich.
|
Moraceae
|
arbre
|
|
Ficus platyphylla Del
|
Moraceae
|
arbre
|
|
Fucus ingens (Miq.) Miq.
|
Moraceae
|
arbre
|
|
Gardenia aqualla Stapf et Hutch.
|
Rubiaceae
|
sous-arbuste
|
|
Gardenia erubescens Stapf et Hutch.
|
Rubiaceae
|
sous-arbuste
|
|
Gardenia ternifolia Schum. ex Thonn.
|
Rubiaceae
|
arbuste
|
|
Grewia bicolor Juss.
|
Tiliaceae
|
sous-arbuste
|
|
Grewia lasiodiscus K. Schum.
|
Tiliaceae
|
arbuste
|


336
Grewia mollis Juss.
|
Tiliaceae
|
arbuste
|
|
Guiera senegalensis J. F. Gmel.
|
Combretaceae
|
sous-arbuste
|
|
Isoberlinea doka Craib et Stapf
|
Caesalpiniaceae
|
grand arbre
|
|
Khaya senegalensis (Desr.) A. Juss
|
Meliaceae
|
arbre
|
|
Lannea acida A. Rich
|
Anacardiaceae
|
arbre
|
|
Lannea barteri (Oliv.) Engl.
|
Anacardiaceae
|
arbre
|
|
Lannea microcarpa Engl. ex K. Krause
|
Anacardiaceae
|
arbre
|
|
Lonchocarpus laxiflora Guill. & Perrott.
|
Fabaceae
|
arbuste
|
|
Maytenus senegalensis (Lam.) Exell.
|
Celastraceae
|
arbuste
|
|
Mitragyna inermis (Willd.) O. Ktze.
|
Rubiaceae
|
arbre
|
|
Nauclea latifolia Smith
|
Rubiaceae
|
arbuste
|
|
Parkia biglobosa (Jacq.) Benth.
|
Mimosaceae
|
grand arbre
|
|
Piliostigma reticulatum (DC.) Hochst.
|
Caesalpiniaceae
|
arbuste
|
|
Piliostigma thonningii (Schum.) Milne-Redhead
|
Caesalpiniaceae
|
arbuste
|
|
Pteleopsis suberosa Engl. ex Diels
|
Combretaceae
|
arbuste
|
|
Pterocarpus erinaceus Poir.
|
Fabaceae
|
arbre
|
|
Sclerocarya birea (A. Rich.) Hochst.
|
Anacardiaceae
|
arbre
|
|
Securinega virosa (Roxb. ex Willd.) Baill.
|
Euphorbiaceae
|
sous-arbuste
|
|
Sterculia setigera Del.
|
Sterculiaceae
|
arbre
|
|
Stereospermum kunthianum Cham
|
Bignoniaceae
|
arbuste
|
|
Strychnos innocua Del.
|
Loganiaceae
|
arbuste
|
|
Strychnos spinosa Lam.
|
Loganiaceae
|
arbuste
|
|
Tamarindus indica Linn.
|
Caesalpiniaceae
|
arbre
|
|
Terminalia avicennioides Guil. et Perr.
|
Combretaceae
|
arbuste
|
|
Terminalia laxiflora Engl.
|
Combretaceae
|
arbre
|
|
Terminalia macroptera Guil. ex Perr.
|
Combretaceae
|
arbre
|
|
Terminalia mollis Laws.
|
Combretaceae
|
arbre
|
|
Vitellaria paradoxa Gaertn.f.
|
Sapotaceae
|
arbre
|
|
Vitex doniana Sweet
|
Verbenaceae
|
arbre
|
|
Ximenia americana Linn.
|
Olacaceae
|
arbuste
|
|
Ziziphus abyssinica Hochst.ex A. Rich.
|
Rhamnaceae
|
arbuste
|
|
|


