Contribution à l'étude de la niche écologique de la Cigogne blanche Ciconia ciconia L. , 1758 (Aves, Ciconiidae ) et du Héron garde- boeufs Bubulcus ibis L. , 1758 (Aves, Ardeidae ) dans la région de Batna( Télécharger le fichier original )par Naoual BOUKHTACHE Université El Hadj Lakhdar Batna Algérie - Magister en agronomie 2008 |

D'après ETCHECOPAR et HÜE (1964) la Cigogne blanche est appelée encore dans les régions Nord de l'Afrique : -Arabe parlé (Algérie, Maroc, Tunisie, et régions septentrionales de la Mauritanie et du Sahara Occidental) : Bellaredj, Berraredj et Hadj-Kacem. -Berbère (Kabylie, Gourara et Aurès) : Bellaredj. -Libye, Egypte et Soudan septentrional : Laklak et Hadj laklak. 2- Systématique et sous espèces 2.1- SystématiqueGEROUDET (1978), SCHIERER (1981), DARLEY (1985), CREUTZ (1988) BOCK (1994), MAHLER et WEICK (1994) et WHITFIELD et WALKER (1999) classent la Cigogne blanche dans les taxons suivants : Règne : Animalia Sous règne : Metazoa Super embranchement : Cordata Embranchement : Vertebrata Sous embranchement : Gnatostomata Super classe : Tetrapoda Classe : Aves Sous classe : Carinates Ordre : Ciconiiformes Famille : Ciconiidae Genre : Ciconia Espèce : Ciconia ciconia L., 1758 Synonyme : Ciconia alba Bechstei 2.2- Sous espèces de C. ciconia et leur distributionIl existe actuellement dans le monde trois sous-espèces de la Cigogne blanche (CRAMP et SIMMONS, 1977 ; COULTER et al., 1991) : - Ciconia ciconia ciconia Linné, 1758 : niche dans une partie de l'Asie mineure, en Europe centrale (Autriche, Bulgarie, Portugal), en Afrique du Nord (du Maroc à la Tunisie), en Afrique du Sud (province du Cap). Rencontrée en Afrique de l'Ouest tous les mois de l'année sauf au mois de juin (DEKEYSER et DERIVOT, 1966). - Ciconia ciconia asiatica Severtzov, 1872 : son aire de reproduction se situe en Asie centrale et niche donc au Turkestan, l'ancienne URSS, Ouzbékistan, Tadjikistan et à l'extrême ouest de Sin-Kiang en Chine : 59° et 79° E, 38° et 43° N (CREUTZ, 1988). - Ciconia ciconia boyciana Swinhowe, 1873 : considérée souvent comme une espèce propre, nidifie en Asie Orientale, de l'Ussuri à la Corée et au Japon (COULTER et al., 1991). D'après LOWE (1994), la Cigogne orientale Ciconia boyciana figure sur la liste des oiseaux menacés dressée par le CIPO (Conseil International de la Protection des Oiseaux). 3- IdentificationOiseau peu farouche envers l'homme, la Cigogne blanche est l'échassier le plus facile à observer. Les adultes (Fig. 1) sont facilement reconnaissables à leurs plumages blanc et noir, ailes robustes et larges, à leur grand cou et brève queue, bec rouge vif et long, droit et très pointu et pattes hautes minces de couleur rouge vif, rémiges primaires et secondaires noires et doigts reliés par une petite membrane (BURTON et BURTON, 1973 ; PETERSON et al., 1986-2006 ; CREUTZ, 1988). Les jeunes ressemblent beaucoup aux adultes, sauf que le plumage est blanc avec du brun sur les ailes, le bec et les pattes sont de couleur brun rougeâtre (HAYMAN et BURTON, 1977 ; HANCOCK et al., 1992). Il est très difficile de distinguer le mâle de la femelle dans la nature, ils ont un plumage identique. En principe, le mâle est légèrement plus corpulent et son bec plus long et plus haut à la base avec un relèvement de l'arête inférieure avant la pointe (BOUET, 1950 ; GEROUDET, 1978 ; SILLING et SCHMIDT, 1994).
Figure 1- La Cigogne blanche (C. ciconia) dans
son nid avec sa progéniture sur une Les cigognes blanches pratiquent le vol à voile. Ce sont d'excellents planeurs. Elles utilisent les courants d'air ascendants qu'on ne rencontre qu'au dessus de la terre ferme (SILLING et SCHMIDT, 1994). La Cigogne blanche vole le cou tendu en avant un peu incliné au dessous de l'horizontale et les pattes dépassent la queue. En fait, elle profite au départ des courants ascendants pour prendre de la hauteur sans effort (ETCHECOPAR et HÜE, 1964 ; MOALI et MOALI-GRINE, 2001 ; PETERSON et al., 2006). Elles regagnent souvent la terre après une descente acrobatique (GEROUDET, 1978). Les cigognes ne sont pas des oiseaux chanteurs, mais lorsqu'elles prennent leur tour sur le nid, elles exécutent un salut rituel, avec des claquements de bec qui produisent un bruit caractéristique. Elles détournent en même temps la tête, et donc le bec, comme s'il s'agissait d'une épée. Ce geste, à l'opposé de la posture de menace, bec en avant, désamorce toute agressivité entre partenaires (ETCHECOPAR et HÜE, 1964 ; WHITFIELD et WALKER, 1999). Toutefois, les petits font entendre des sifflements et des cris aigus : ouyeh...eche...eche...urh...qui sont de curieux miaulements et grincements pour mendier leur pitance (GEROUDET, 1978). 4- Répartition géographique4.1- Dans le MondeLa Cigogne blanche est une espèce Paléarctique, sa distribution englobe une partie de l'Europe, le moyen Orient, le centre Ouest Asiatique, le Nord-ouest de l'extrême Sud Africain (DUQUET, 1990 ; HANCOCK et al., 1992). La sous espèce Ciconia ciconia ciconia se trouve dans les régions tempérées méditerranéennes d'Europe, dans le Sud et l'Est du Portugal, l'Ouest et le centre de l'Espagne, l'Est de la France, les Pays-Bas, le Danemark, la région de Saint Petersburg, la Turquie, le Nord de la Grèce, l'Est de la Yougoslavie et sporadiquement le Nord de l'Italie, elle a niché dans le Sud de la Suède, l'Ouest de la France et en Belgique (HEIM DE BALSAC et MAYAUD, 1962 ; CRAMP et SIMMONS, 1977) (Fig. 2). En Afrique du Nord, on rencontre la même sous espèce dans le Nord-est de la Tunisie en passant par l'Algérie jusqu'au Sud du Maroc (ETCHECOPAR et HÜE, 1964 ; LEDANT et al., 1981 ; DUQUET, 1990). Au moyen Orient, elle se rencontre en Turquie, l'Azerbaïdjan, l'Ouest de l'Iran, le Nord de l'Iraq et en Asie de Sud-Ouest (BURTON et BURTON, 1973 ; MAHLER et WEICK, 1994).
Aires de reproduction Aires d'hivernage Voies de migration Figure 2- Repartition geographique de la Cigogne
blanche (Ciconia ciconia ciconia) dans le monde : aires
de 4.2- En AlgérieEn Algérie, la Cigogne blanche est bien plus commune ; elle est répandue dans toute la région tellienne et descend jusqu'à l'Aurès (commune à Batna). Plus au sud encore, un nid inoccupé en 1923 à Djelfa, une colonie à El Kreider (Chott-Ech-Chergui), un nid en 1966 à Aflou et un autre en 1974 à El Idrissia, mais la nidification signalée au XIXe siècle dans le M'zab par Loche a été mise en doute (BOUET, 1956 ; HEIM DE BALSAC et MAYAUD, 1962 ; THOMAS et al., 1975 ; LEDANT et al., 1981) Au Nord, elle est présente dans les régions de Béjaia, Sétif, dans le Nord du Hodna (M'sila) et sur les plateaux de Bouira jusqu'à Sour-El Ghozlane. On la trouve également dans la dépression de Lakhdaria. Elle peuple aussi toute la vallée du Sébaou jusqu'à la lisière du massif forestier d'Akfadou, à Azazga ; ainsi que sur les plaines entre Ouadhias et Draâ El Mizan. Un nombre réduit de couples nichent près de Boufarik, de Rouiba, de Hadjout et de Mouzaia (MOALI-GRINE, 1994). D'après ce dernier auteur, la cigogne reste abondante dans la région humide d'El Kala et se trouve également dans le Constantinois. A l'Ouest, l'espèce peuple la vallée de Chlef et Miliana, sa répartition continue jusqu'à Mostaganem et plus loin qu'Oran sur la bande littorale jusqu'à Beni Saf. Ayant besoin de zones humides ou cultivées, la Cigogne blanche est plus abondante dans la partie orientale que dans la partie occidentale du pays (ISENMANN et MOALI, 2000). 5- Etat actuel des populations5.1- Dans le MondeLa situation de la population européenne et nord-africaine de la Cigogne blanche a fait objet de plusieurs congrès internationaux durant ces dernières décennies. Depuis 1934 jusqu'à nos jours 6 recensements internationaux sont organisés pendant les années : 1934, 1958, 1974, 1984, 1994-95 et 2004-2005 (THOMSEN et HÖTKER, 2006). Ces recensements internationaux organisés ont permis de constater un déclin général de l'espèce et surtout de sa population occidentale (RHEINWALD et al., 1989 ; BIBER et al., 1995). En Europe occidentale, la Cigogne blanche se porte moins bien que sa signification symbolique pourrait nous le faire espérer, car ses populations se sont dramatiquement raréfiées et elle a failli disparaître en Alsace (neuf couples en 1974). Dans d'autres régions d'Europe, elle est menacée par l'intensification des pratiques agricoles et l'assèchement des marais (WHITFIELD et WALKER, 1999 ; DUBOURG et al., 2001, MASSEMINCHALLET et al., 2006). Les résultats du symposium international pour la Cigogne blanche qui s'est tenu à Hambourg en 1996 ont montré que le recensement international des couples nicheurs comparé à celui de 1984 révèle un développement positif des populations dans la plupart de ses pays de distribution. Le nombre de couples nicheurs est passé de 140.300 en 1984 à 168.000 en (1994-1995), donc la population a augmenté de 20 % (SCHULZ, 1999). THOMSEN et HÖTKER (2006), constatent que les populations de cigognes blanches ont décliné de 20 % entre 1974 et 1984 puis elles ont augmenté de 23 % entre 1984 et 1994- 95, et que la population occidentale a augmenté de 75 % depuis 1984, alors que la population orientale a augmenté seulement de 15 %. Un ensemble de 40 états européens, nord-africains, du Proche-Orient et de l'Asie centrale, ont participé au dernier recensement de 2004-2005. Les résultats préliminaires recueillis de 13 états montrent que la population est encore en augmentation et qu'il y a des pays où cette augmentation est de l'ordre de 100 % (Tab. II) (THOMSEN et HÖTKER, 2006). Tableau II- Résultats préliminaires du sixième recensement international de la Cigogne blanche (C. ciconia) dans quelques pays participants.
Cette augmentation des populations de la Cigogne blanche est constatée aussi par HAMDI et al. (2007) en Tunisie où le nombre de nids est passé de 358 entre les années 1980- 1999 à 489 nids en 2002-2005. 5.2- En AlgérieEn Algérie, des recensements nationaux ont donné 6.400 à 6.500 couples nicheurs en 1935 (BOUET, 1936) et 8.844 en 1955 (BOUET, 1956). Mais cet effectif a beaucoup décliné de 1955 à 1993. Ainsi, pour MOALI et MOALI-GRINE (1995) et MOALI-GRINE et al. (1995), l'effectif nicheur de l'essentiel de l'aire de distribution de la Cigogne blanche en Algérie s'élève à 1.195 couples en 1992-1993, soit un déclin de l'ordre de 86,49 % par rapport au recensement de 1955. A cet effet, l'espèce reste encore relativement abondante mais le déclin est évident par rapport aux recensements de 1935 et de 1955 (ISENMANN et MOALI, 2000). Selon MULLIÉ et al. (1995), le déclin continu des couples nicheurs d'Algérie et de Tunisie se reflète par le nombre faible d'hivernant au Tchad. A cet effet, un autre recensement de Cigognes réalisé par MOALI et al. (1998) in ZENNOUCHE (2002), effectué dans le cadre du recensement international, révèle une tendance positive de la population nicheuse algérienne. D'après MOALI-GRINE (2005), les récents dénombrements (1998 et 2001) ont tous les deux confirmé la tendance à l'augmentation de la population qui est passé de 1.195 couples en 1993 à près de 6.000 couples en 2001, ce qui rejoint les synthèses du colloque organisé à Hambourg en 1996 concernant l'ensemble des populations de Cigognes blanches dans le monde. Dans la région de Batna, un recensement minutieux fait récemment par DJADDOU et BADA (2006) donne 490 nids recensés dont 7 nids seulement sont non occupés. Ce recensement assez exhaustif dénote des lacunes au niveau des recensements réalisés par les services des conservations des forêts qui rapportent généralement des chiffres en deçà de la réalité. 6- Migration et hivernage en Afrique6.1- MigrationLe phénomène grandiose et passionnant de la migration s'effectue chaque année entre la fin du mois de juillet et la deuxième décade du mois d'août, où les cigognes quittent leur lieu de reproduction et se rendent en Afrique pour y passer l'hiver (SCHIERER, 1963 ; GORIUP et SCHULZ, 1991 ; ISENMANN et MOALI, 2000 ; JONSSON et al., 2006). Cependant, METZMACHER (1979), DUQUET (1990) et SKOV (1991a) signalent que quelques individus s'attardent jusqu'à la mi-octobre. SKOV (1991a), signale encore qu'il y a des cas très rares de cigognes qui ne migrent pas et passent l'hiver sur les lieux de reproduction et supportent même des températures de -25 °C au Danemark, c'est la cas d'une cigogne mâle observée pendant les hivers 1985-86, 1986-87 et 1987-88. Les cigognes blanches d'Europe se scindent en deux parties bien distinctes pour migrer (Fig. 2), l'une suivant une voie orientale passant par le Bosphore, la Turquie et la Palestine pour rejoindre l'Est africain (les plateaux de l'Ouganda), l'autre emprunte une voie occidentale passant par la France, l'Espagne, le détroit de Gibraltar survole le Maroc, puis la Mauritanie pour qu'elle aboutit et se dissémine entre le Cameroun et le Sénégal (DORST, 1962 ; SCHÜZ, 1962 ; THOMAS et al., 1975 ; GRASSE, 1977 ; GEROUDET, 1978 ; WHITFIELD et WALKER, 1999). Une zone de démarcation qui traverse les Pays-Bas et l'Allemagne occidentale, sépare les deux courants migratoires et au milieu de laquelle passe une ligne virtuelle où le partage se fait à 50 % (DORST, 1962). Le départ des lieux de reproduction vers les aires d'hivernage a lieu au Maroc et en Algérie au début d'août, époque semblable à celle observée en Europe centrale (HEIM DE BALSAC et MAYAUD, 1962). La migration des cigognes d'Algérie semble se faire sur un large front à travers le Sahara, bien qu'il se dégage une voie privilégiée empruntant l'est de l'Algérie par El Goléa, Ain Salah, Arak et Tamanrasset pour rejoindre le Sahel (ISENMANN et MOALI, 2000). L'espèce a aussi été signalée, parfois avec des effectifs importants, dans l'ouest du Sahara algérien, notamment à Tindouf (HEIM DE BALSAC et MAYAUD, 1962). Après un séjour de quelques mois sur le continent africain, l'instinct rappelle peu à peu les cigognes vers le Nord et la migration reprend. Les voies de retour sont sensiblement les mêmes que celles de l'automne que ce soit à l'Est ou à l'Ouest (GEROUDET, 1978). Ces différentes voies de migration sont constatées dans les premières études par les méthodes basées sur le baguage. Les études récentes utilisent le suivi satellitaire et arrivent à des résultats beaucoup plus détaillés. Par exemple, BOSSCHE et al. (2002), signalent que la période de vol varie de 8 à 10 heures par jour séparées par des périodes de repos de 14 à 16 heures. La région d'Israël-Palestine est une importante voie de migration des oiseaux de l'Europe vers l'Afrique (Cigognes, Pélicans...). A cet effet, les Israéliens ont développé différents systèmes tels les radars pour la prédiction des différentes altitudes des vols des oiseaux migrateurs. Le plus récent "The convection model ALPTHERM" est développé pour éviter les éventuels accidents aériens avec les avions militaires qui sont en permanence dans cette région. "The convection model ALPTHERM", basé sur les conditions climatiques et les données de la topographie, montre que la Cigogne blanche, lors de sa migration, atteint des altitudes variant entre 488 m et 1.615 m (SHAMOUN-BARANES et al., 2003). En Espagne, GORDO et al. (2007), ont essayé de modéliser la migration de cette espèce tout en reliant ce phénomène aux conditions de l'environnement, aux conditions géographiques et notamment aux comportements sociaux de l'espèce, mais leurs résultats restent encore embryonnaires. 6.2- Hivernage en AfriqueLa Cigogne blanche n'a pas de quartiers d'hivernage bien définis. Les cigognes partent en troupes d'importance variable vers les quartiers d'hivernage qui s'étendent d'une part, dans l'Ouest entre la zone désertique et celle des forêts tropicales du Sénégal au Soudan, et d'autre part dans l'Est sur les steppes et savanes échelonnées depuis le Soudan et l'Ethiopie jusqu'au Cap (CREUTZ, 1988 ; SILLING et SCHMIDT, 1994). Les cigognes blanches algériennes, par exemple, semblent hiverner de la région du fleuve Niger à la République Centre Africaine, quoique des exemplaires bagués aient aussi été repris au Zaïre et en Ouganda (HEIM de BALSAC et MAYAUD, 1962). Pendant la période d'hivernage, les oiseaux vagabondent et suivent le plus volontiers les essaims de grandes sauterelles rouges (GEROUDET, 1978). La cigogne est attirée par les cendres qui en résultent des feux de brousse, qui sont des terrains véritablement riches en cadavres d'invertébrés et surtout d'insectes (THIOLLAY, 1971 in AMARA, 2001). 7- Habitat :Les cigognes blanches sont grégaires et familières de la présence de l'homme (ETCHECOPAR et HÜE, 1964 ; WHITFIELD et WALKER, 1999). Elles habitent avec prédilection les paysages ruraux à forte proportion de prairies, de cultures et de pâtures, des bas fonds humides, des eaux peu profondes, des paysages découverts, des mares temporaires, les territoires humides et les champs qui lui fournissent sa subsistance (ETCHECOPAR et HÜE, 1964 ; BOUKHEMZA, 2000 ; DUBOURG et al., 2001 ; MOALI et MOALI-GRINE, 2001). L'espèce fréquente les steppes et les savanes, mais ne pénètre guère dans les zones forestières. Elle ne montre aucun intérêt pour les rivages maritimes. Sa crainte de la mer l'empêche en général de parvenir sur les îles éloignées (BOUET, 1938 ; ETCHECOPAR et HÜE, 1964 ; GEROUDET, 1978). Les bonnes conditions de détection des proies ainsi que la possibilité de se déplacer sans être entravée par la végétation, sont des facteurs importants quant au choix des habitats d'alimentation (GEROUDET, 1978 ; NICOLAI et al., 1985 ; PETERSON et al., 1986 ; HANCOCK et al., 1992 ; THOMSEN, 1995). 8- Biologie de la reproduction
8.1- Maturité sexuelleA l'âge de première année la jeune cigogne blanche ne rentre jamais à son aire natale et elle est souvent observée dans ses quartiers d'hivernage en été. A l'âge de deux ans, le mécanisme de l'activité reproductive est mieux développé, mais ne se reproduit pas encore. A l'âge de trois ans la Cigogne se reproduit, mais habituellement avec un nombre moindre de petits par rapport aux Cigognes âgées. A quatre ans, la Cigogne blanche est bien mature (SCHÜZ, 1936). Selon ZINK (1960), les jeunes cigognes blanches se reproduisent à partir de la troisième année jusqu'à la sixième année. Pour DORST (1971a) et BARBRAUD et al. (1999), l'âge de première nidification est en moyenne de trois ans. 8.2- Formation des couples et parade nuptialeLe mâle arrivant généralement une semaine avant la femelle prend possession d'un nid qu'il défend contre tout autre concurrent (SCHÜZ, 1936 ; ETCHECOPAR et HÜE, 1964 ; GEROUDET, 1978 ; GORIUP et SCHULZ, 1991). La première femelle qui arrive est souvent acceptée et un couple saisonnier monogame se forme. Ceci se manifeste par un grand bruit de claquettements de bec (GORIUP et SCHULZ, 1991). Dans tous les cas observés, c'est la femelle qui prend l'initiative et va au devant de son partenaire, le mâle reste passif, très excité, claquette en effectuant de lents et amples battements d'ailes (il pompe) ; trois phases sont observées durant la formation d'un couple, une phase d'approche, une phase intermédiaire et une phase terminale (SCHMITT, 1967 in AMARA, 2001). 8.3- AccouplementL'accouplement a lieu sur le nid, et il n'existe qu'une seule nichée par an (WHITFIELD et WALKER, 1999). Les accouplements sont exécutés sur l'aire, debout le mâle sautant sur la femelle en s'accrochant les pattes sur les épaules avant de s'accroupir en battant des ailes, tandis que caresse du bec le cou de l'autre (CREUTZ 1988 ; SILLING et SCHMIDT 1994). Selon SCHMITT (1967) in AMARA (2001), un accouplement dure à peu près sept secondes, le couple peut procéder à deux accouplements successifs. 8.4- Sites de nidification et construction du nidLa Cigogne blanche niche généralement en colonies sur les constructions humaines, où elle est assez bien accueillie. Elle installe son nid sur des endroits élevés, sur les cimes d'arbres, mais souvent sur une enfourchure de branches ou de tronc (Peuplier, Eucalyptus, Platane...), sur les toits, les tours, les édifices, les poteaux électriques, les bâtiments, les minarets, les églises et les grosses fermes (HEIM DE BALSAC et MAYAUD, 1962 ; YEATMAN, 1976 ; HEINZEL et al., 1985-2005 ; DUBOURG et al., 2001 ; BROWN, 2005). Le nid (900-1.500 mm de diamètre) est une énorme construction de branchages, à base de branchettes, mottes de terre, de touffes d'herbe, réutilisé chaque année, sur lesquelles les oiseaux aménagent une coupe peu profonde garnie de foin et de plumes, parfois de papier et de chiffons (ETCHECOPAR et HÜE, 1964 ; GEROUDET, 1978 ; BOLONGNA, 1980 ; GORIUP et SCHULZ, 1991 ; WHITFIELD et WALKER, 1999). Chaque année, à son retour, la Cigogne blanche renforce son nid avec de nouvelles branches et rembourre l'intérieur avec de l'herbe fraîche, du duvet, végétaux et même de vieux chiffons (GEROUDET, 1978). D'année en année, ces édifices peuvent atteindre des dimensions et poids très importants (SIGNOLLET et MANSION, 2002). La fidélité au nid est considérée comme une stratégie adaptative pour l'augmentation du taux de succès de la reproduction. Par conséquent, un échec dans une nichée précédente a un effet sur le changement du nid dans la nichée suivante (VERGARA et al., 2006 ; VERGARA et al., 2007). Ces derniers auteurs suggèrent que l'âge des Cigognes blanches est un facteur majeur et a une relation étroite avec cette fidélité, ceci s'explique par leur expérience. ISENMANN et MOALI (2000), signalent que lors du recensement de 1995, en Algérie, 59 % des couples ont niché dans des agglomérations, 25 % sur des pylônes et des poteaux, 38 % sur des toits de maisons et 37 % sur des arbres. Selon BOUET (1936) et GEROUDET (1978), très souvent quelques couples de moineaux (Passer domesticus et P. hispaniolensis), de même que des bergeronnettes grises et des étourneaux occupent fréquemment le substratum du nid et y construisent leurs propres demeures sans être jamais molestés par leurs puissants voisins. 8.5- PonteLa taille de la ponte varie entre 2 et 6 oeufs, assez fréquemment de 4 (ETCHECOPAR et HÜE, 1964 ; GORIUP et SCHULZ, 1991 ; WHITFIELD et WALKER, 1999), rarement de 7 (BOLOGNA, 1980). SKOV (1991b), signale des cas de 8 oeufs au Danemark. Ces oeufs sont pondus à 24 ou 48 heures d'intervalle (RIGHI, 1992 ; BOUKHEMZA, 2000). En cas de la destruction de la couvée, une deuxième ponte de remplacement peut rarement avoir lieu (GEROUDET, 1978). La ponte est déposée au mois de février dans les plaines marocaines et elle se déroule entre le mois de mars et le mois d'avril en Algérie et en Tunisie (HEIM DE BALSAC et MAYAUD, 1962 ; ETCHECOPAR et HÜE, 1964). Cependant, la ponte est plus tardive en Europe centrale où elle s'étale surtout sur le mois de mai et parfois même jusqu'au mois de juin (SCHÜZ, 1936 ; GEROUDET, 1978). Le nombre des oeufs par ponte paraît varier sensiblement et sans doute est-il en rapport avec l'abondance de la nourriture, singulièrement des criquets (HEIM DE BALSAC et MAYAUD, 1962). VALVERDE et al. (1960) in AMARA (2001), ont remarqué que les années où la sécheresse et la plus accusée, le nombre des pontes diminue, alors que les années caractérisées par d'abondantes précipitations corrèlent avec l'augmentation du nombre d'oeufs par ponte. 8.6- Couvaison et éclosion des oeufsLa couvaison commence après la ponte du deuxième oeuf ou avant que le dernier oeuf soit pondu (SCHÜZ, 1936 ; DORST, 1971a ; GEROUDET, 1978 ; HAMADACHE, 1991). Elle est assurée alternativement par les deux partenaires pendant 30 à 34 jours (SCHÜZ, 1936 ; DORST, 1971a ; BOLONGNA, 1980 ; WHITFIELD et WALKER, 1999). Ils se relaient à peu près toutes les deux heures, sauf la nuit où la femelle reste d'habitude sur les oeufs (GEROUDET, 1978). D'après BOUKHEMZA (2000), 15 relais sont notés en 50 heures d'observation, soit 1 relais toutes les 3 heures et 30 minutes environ. Les éclosions s'échelonnent sur une dizaine de jours à l'abri des adultes (GEROUDET, 1978), moment à partir duquel on observe un surcroît d'activité dans le nettoyage est l'élargissement du nid et une accélération dans les allées et venues au nid pour la recherche de la nourriture qui se fait tantôt individuellement tantôt en couple, cas le plus fréquent (BOUKHEMZA, 2000). 8.7- Nourrissage et élevage des jeunesLes parents apportent la nourriture dans le jabot et la dégorgent toujours sur le nid où les petits la picorent, encore enrobée de salive (GEROUDET, 1978 ; BOUKHEMZA, 2000). Peu à peu, cependant, les jeunes se développent et passent leur temps à se quereller, assis sur leurs tarses, ils accueillent l'arrivée du nourricier avec le bec ouvert, en miaulant et en agitant leurs moignons d'ailes. Accroupis en cercle, ils se hâtent d'engloutir la provende vomie en leur milieu dont le surplus éventuel est mangé par l'adulte. Par temps chaud, celui-ci apporte aussi de l'eau et la déverse directement dans leurs becs et asperge à gros bouillons, trempés par la chaleur (GEROUDET, 1978 ; SILLING et SCHMIDT, 1994 ; BOUKHEMZA, 2000). Comme le dernier né a un retard assez important, il n'est pas rare qu'il demeure chétif et dépérisse, victime de ses frères et soeurs qui le réduisent à la famine, ou même de ses parents qui le tuent en le malmenant à coups de bec, il est alors jeté en bas de l'aire ou même dévoré par ses parents (GEROUDET, 1978). 8.8- EnvolLes jeunes commencent à battre les ailes vers l'âge de trois semaines mais ne volent qu'à deux mois. A six semaines, les plumes noires apparaissent aux ailes, à sept semaines la station debout est régulière et on voit des exercices de battements qui prépare les muscles à voler. Au bout de la neuvième semaine ou dixième semaine, les jeunes accomplissent leurs premiers vols (SCHÜZ, 1936 ; ARNHEM, 1980 ; BOLOGNA, 1980 ; WHITFIELD et WALKER, 1999 ; BOUKHEMZA, 2000). 9- Ecologie trophique9.1- Composition du régime alimentaireSelon GEROUDET (1978) et SKOV (1991b), la nourriture de la Cigogne blanche est exclusivement animale, elle se compose en somme de tout ce qui se présente et qui peut être avalé. La Cigogne blanche récolte une grande variété d'insectes, tout spécialement des coléoptères et des orthoptères qui constituent une bonne part du régime alimentaire, aussi bien sur les lieux de nidification que dans les quartiers d'hiver en Afrique centrale et méridionale. Elle consomme aussi des reptiles, des petits mammifères, des grenouilles, des poissons, des vers de terre et même des jeunes oiseaux (ETCHECOPAR et HÜE, 1964 ; DORST, 1971a ; BURTON et BURTON, 1973 ; NICOLAI et al., 1985 ; JONSSON et al., 2006). Elle récolte les mollusques, notamment les escargots dont elle casse la coquille avant de les ingurgiter, elle glane beaucoup de vers de terre, surtout en début de saison quand les autres aliments sont encore rares et prend à l'occasion des crustacés, par exemple le Crabe chinois, dans les cours d'eau qu'il a envahis (GEROUDET, 1978 ; SKOV, 1991b). En Algérie, des études concernant le régime alimentaire de la Cigogne blanche ont été menées à Tizi-ouzou dans la région du bas et du moyen Sébaou par BOUKHEMZA et al. (1995), FELLAG (1995-2006), BENTAMER (1998) et BOUKHEMZA (2000) ; dans la région de Tébessa par AMARA (2001) et SBIKI (2008) ; dans la région de Béjaia par DOUADI et CHERCHOUR (1998) et ZENNOUCHE (2002) et dans la région d'Annaba par SAKER (2006). A Batna, une seule étude a été faite par DJADDOU et BADA (2006). Toutes ces études basées sur la décortication des pelotes de réjection ont montré que la Cigogne blanche est presque exclusivement insectivore avec une grande préférence aux coléoptères et aux orthoptères. 9.2- Milieux d'alimentationVu son mode d'alimentation, la Cigogne blanche fréquente les milieux ouverts avec une végétation assez basse pour n'entraver ni sa marche ni sa vue (GEROUDET, 1978 ; PETERSON et al., 1986 ; HANCOCK et al., 1992 ; LATUS et KUJAWA, 2005). La Cigogne blanche, étant de plus en plus anthropophile, fréquente actuellement une large gamme de milieux : marais, labours, friches, prairies. BLANCO (1996), GARRIDO et FERNÁNDEZ-CRUZ (2003), PERIS (2003) et BLÁZQUEZ et al. (2006), signalent qu'en Espagne, les décharges publiques constituent une nouvelle source humaine de gagnage pour la Cigogne blanche. Ceci a été également noté en Algérie par BOUKHEMZA (2000) et SBIKI (2008). 9.3- Association avec d'autres animauxLa Cigogne blanche chasse seule ou en groupe (ETCHECOPAR et HÜE, 1964 ; THOMSEN, 1995). Mais elle ne donne pas d'importance à l'association avec les machines agricoles (RACHEL, 2006). L'espèce est souvent observée dans les aires de gagnage en compagnie de hérons garde-boeufs (Bubulcus ibis) avec qui elle partage, dans certaines localités le même support de nidification tels l'Eucalyptus, le Cyprès, le Pin, le Platane et les résineux (BOUKHEMZA, 2000 ; SI BACHIR, 2007). 9.4- Mode de chasseLa Cigogne ne chasse jamais à l'affût (GEROUDET, 1978). C'est en position courbée, le cou sinueux et le bec abaissé que la cigogne chasse. Elle avance lentement, le regard attentif, piquant de côté et d'autre et relevant la tête après chaque capture pour avaler avec secousse (GEROUDET, 1978). 9.5- Capture et digestion des proiesLes sucs gastriques des cigognes sont très actifs et peuvent dissoudre complètement les os des proies si bien que l'on n'en trouve que peu ou pas de traces dans les pelotes. Les matières non digérées, poils, os et cuticules sclérotinisées sont régurgités sous la forme de pelotes de réjection. Ces dernières sont des agglomérats de résidus indigestes, qui s'accumulent dans l'estomac où les mouvements péristaltiques les assemblent en boulette que l'oiseau crache plus au moins régulièrement (BANG et DAHLSTROM, 1987-2006). Le degré de digestion est variable : des parties osseuses peuvent être rendues intactes ou plus ou mois digérées, les élytres plus ou moins écrasés. Chaque pelote ne résulte pas d'un seul repas (SCHIERER, 1962). 9.6- Recherche de nourriture et rythme d'activités alimentairesLa distance parcourue par cet échassier pour la recherche de la nourriture semble être différente et dépendante ainsi de sa disponibilité, elle peu atteindre jusqu'à 14 km (SCHIERER, 1967 ; SKOV, 1998 ; JOHST et al., 2001). D'après PINOWSKI et al. (1986), le temps consacré à la recherche de la nourriture constitue 59 % de l'activité de la Cigogne blanche dépendant ainsi du type de l'habitat et de la saisonnalité. Pour SKOV (1991a), les cigognes adultes cherchent la nourriture 7 fois par jour. Les juvéniles (moins de 4 semaines d'âge), observés dans 7 nids par STRUWE et THOMSEN (1991), sont nourris par leurs parents 7 à 9 fois par jour, ce qui correspond à un intervalle moyen de nourrissage de 141 minutes. Le taux de nourrissage est influencé par les disponibilités de l'habitat et le besoin respectif de chaque couple reproducteur, ce dernier (besoin) dépendant de l'âge et du nombre de juvéniles à nourrir (STRUWE et THOMSEN, 1991). 10- Facteurs de menace et de mortalitéLes fluctuations des effectifs de la Cigogne blanche sont attribuées à plusieurs facteurs qui sont essentiellement : > La perte des habitats et des sites de nidification A partir des années 1960, le développement économique accompagné par le changement des pratiques culturales (utilisation d'herbicides, de pesticides et de la motorisation) ont induit l'homogénéisation et la stérilité des territoires qui ont été intensifiés. Par conséquent, la perte de la biodiversité s'est traduite par un impact négatif sur l'écologie trophique de la Cigogne blanche (JACOB, 1991 ; SENRA et ALÉS, 1992 ; CARRASCAL et al, 1993 ; MARTINEZ et FERNÁNDEZ, 1995 ; JONSSON et al., 2006). Selon RANDIK (1989), GORIUP et SCHULZ (1991) et SKOV (1998), l'urbanisation incluant l'extension de l'industrie a affecté négativement les populations de cigognes blanches par la démolition des anciennes constructions qui servaient de support de nidification et qui sont ainsi perdues. Spoliée de ses aires traditionnelles, la Cigogne blanche a dû chercher d'autres endroits pour y construire son nid volumineux ; elle les a trouvés sur les mâts des conduits électriques. Le problème ne s'arrête pas à ce niveau car même les nids construits sur les poteaux électriques sont détruits par les services de maintenance (MARTINEZ et FERNÁNDEZ, 1995). > Le changement des conditions d'hivernage Les fluctuations des effectifs de la population occidentale étaient corrélées aux conditions climatiques sur les quartiers d'hiver qui déterminent les potentialités alimentaires. Celles-ci ayant un effet direct sur le taux de survie plutôt que sur le succès de la reproduction. Ceci est la conséquence d'une longue sécheresse sudano-sahélienne qui a fait disparaître des zones humides importantes en 1960-1970, additionnée aux divers systèmes de contrôle des eaux effectués dans les rivières au Sénégal et au Niger (DALLINGA et SCHOENMAKERS, 1984-1989 ; KANYAMIBWA et LEBRETON, 1991 ; SYLLA, 1991 ; SCHULZ, 1995). > Les empoisonnements massifs par les antiacridiens dans le Sahel Les quartiers d'hivernage des deux sous populations de cigognes, orientale et occidentale, semblent se croiser avec les régions souvent affectées par des invasions de criquet migrateur (Locusta migratoria), de criquet marron (Locustana pardolina), de criquet rouge (Nomadacris septumfasciata) et le criquet du désert (Schistocerca gregaria). Les essaims de ces criquets ont été contrôlés par l'utilisation des insecticides (Dieldrin) depuis les années 50 jusqu'à son interdiction en 1980 (DALLINGA et SCHOENMAKERS, 1989 ; SCHULZ, 1988 in GORIOP et SCHULZ, 1991). Les cigognes représentent un agent efficace pour le contrôle des populations de locustes dans certaines régions devant les grandes invasions. D'autre part, l'inhibition de ces locustes prive les cigognes d'une importante source d'alimentation tout spécialement dans la partie orientale. Il semble important de savoir qu'un début de déclin régulier de la sous population occidentale nichant en France et en Allemagne débute en 1961 quant des grands essaims de criquets ont été éradiqué de l'Afrique occidentale (DALLINGA et SCHOENMAKERS, 1989). > La chasse D'après THAURONT et DUQUET (1991), SYLLA (1991) et GORIOP et SCHULZ (1991), la chasse et la capture des cigognes blanches sur le chemin de migration et aux quartiers d'hivernage viennent en tête des causes de déclin. D'après l'analyse des bagues retrouvées, il est certain que la chasse serait la cause majeure de mortalité. > Le baguage Les cigognes blanches sont connues pour leur pouvoir de réguler leur température en déféquant sur leurs pattes ; l'évaporation de l'humidité à partir des déjections aide à refroidir le corps. Cependant, l'accumulation de ces déjections entre la patte et la bague stimule la formation de l'acide urique qui provoque de sérieuses blessures pouvant même conduire jusqu'à la mort. Le taux de mortalité induit par le baguage s'avère important surtout dans quelques pays européens, environ 70 % des poussins sont bagués et 5 % de ces derniers sont perdus chaque année (SCHULZ, 1987 in GORIUP et SCHULZ, 1991). > L'électrocution L'électrocution est considérée comme l'une des causes principales de mortalité des cigognes blanches, elle s'effectue par la collision avec les câbles découverts et les poteaux métalliques qui deviennent de plus en plus dangereux lorsqu'ils sont installés dans des zones rurales. Elle est assez importante chez les jeunes cigogneaux qui quittent leur nid pour la première fois (GORIUP et SCHULZ, 1991 ; BIBER, 1995 ; SKOV, 1998 ; GARRIDO et FERNÁNDEZ-CRUZ, 2003 ; DOLATA, 2006). JACUBIEC (1991) in DOLATA (2006), en se basant sur les observations dans la nature, les données des recensements internationaux et le baguage, a conclu que 74,5 % de mortalité de cigognes blanches est causée par électrocution entre 1970 et 1987. > La contamination par les métaux lourds La Cigogne blanche est exposée aux différents polluants évacués dans ses milieux de gagnage, tels que les métaux lourds, les polluants organiques (les amines aromatiques) et les organochlorés (pesticides), par leur accumulation dans les oeufs en affectant sa productivité (HERNANDEZ et al., 1988) et ses différents organes (foie et rein) (MEHARG et al., 2002 ; SMITS et al., 2005 ; BLÁZQUEZ et al., 2006). Ces derniers auteurs ont fait des études dans ce sens et ont prouvé des taux élevés de métaux lourds (Pb, Co, Cr, Ti, Zn, Sn, V, Ba, Sr) qui ont des effets dangereux sur la santé de cette espèce tels des mal formations dans le squelette (jambe) des jeunes cigognes et leur exposition aux différentes pathologies. > L'impact de la téléphonie mobile BALMORI (2004-2005), a consacré ses recherches sur les effets des champs électromagnétiques émis par les antennes et les pylônes de la téléphonie mobile sur la Cigogne blanche en Espagne. Il a trouvé que dans une colonie distante de 200 m de ces antennes téléphoniques, 40 % de nids n'ont pas eu de poussins alors que dans une autre colonie éloignée de plus de 300 m, 3,3 % de nids seulement n'ont pas eu de poussins. Les micro-ondes des champs électromagnétiques qui sont plus intenses au voisinage des antennes ont ainsi un grand effet sur la productivité de la Cigogne blanche. Partie III :Présentation générale du Héron garde-boeufs (Bubulcus ibis)1- NomenclatureLe Héron garde-boeufs B. ibis est désigné par différents synonymes vernaculaires en plusieurs langues, nous retiendrons ceux cités par ETCHECOPAR et HÜE (1964), GEROUDET (1978) et PETERSON et al., (1986-2006) (Tab. III) : Tableau III- Noms vernaculaires donnés au Héron garde-boeufs dans plusieurs langues.
D'après ETCHECOPAR et HÜE (1964), le Héron garde-boeufs est appelé dans les régions Nord de l'Afrique : - Arabe (Tunisie, Algérie et Maroc) : Tir-elbgar ou bien Dadjadj El B'gar. - Berbère (Kabylie, Gourara et Aurès) : Asaboua et Tir-Amellal. - Libye, Egypte et Soudan septentrional : Abou Kardan, Abou Bakar, Abou Ghanam. 2- Systématique et sous espèces 2.1- SystématiqueGRASSE (1950), GEROUDET (1978), DARLEY (1985), VOISIN (1991), BOCK (1994) et WHITFIELD et WALKER (1999) classent le Héron garde-boeufs dans les taxons suivants : Règne : Animalia Sous règne : Metazoa Super embranchement : Cordata Embranchement : Vertebrata Sous embranchement : Gnatostomata Super classe : Tetrapoda Classe : Aves Sous classe : Carinates Ordre : Ciconiiformes Famille : Ardeidae Genre : Bubulcus Espèce : Bubulcus ibis Linné, 1758 Synonymes : Ardeola ibis, Ardea veranyi Roux et Ardeola bubulcus Gray. 2.2- Sous espèces de B. ibis et leur distributionSelon VOISIN (1991), il existe actuellement dans le monde trois sous-espèces voisines du Héron garde-boeufs : - Bubulcus ibis ibis Linné, 1758 : nidifie dans le Sud de l'Europe (seulement dans la Péninsule Ibérique et la Camargue), en ancienne U.R.S.S. et en Iran, en Isräel et au Yémen. En Afrique, elle niche en Egypte, au Maroc et dans toute l'Afrique au Sud du Sahara. Elle niche aussi à Madagascar, à Sào-Thomé, aux Iles du Cap-Vert, aux Iles Comores et Mauritius (DEKEYSER et DERIVOT, 1966 ; VOISIN, 1991). Dans le Nouveau Monde, l'espèce niche au quartier Nord, où elle s'y est établie jusqu'en Colombie, Californie, la côte Est des Etats-Unis et le Sud-est du Canada. Dans le quartier Sud elle niche dans le delta Amazonien en Uruguay et en Argentine. Elle a été introduite à Hawaï où actuellement elle niche également (VOISIN, 1991). - Bubulcus ibis coromandus Boddaert, 1783 : niche en Asie, du Pakistan et l'Inde à l'Est de la Chine, à la Corée du Sud et au le Sud du Japon. Elle niche également dans les îles environnant l'Inde (Iles Maldives; Sri Lanka, Iles Nicobar ; ...). Elle habite aussi le Nord et le Nord-Est de l'Australie (VOISIN, 1991). - Bubulcus ibis seychellarum Salomonsen, 1934 : est une forme intermédiaire entre les deux premières et vit dans les Seychelles (HANCOCK et KUSHLAN, 1989). Les sous espèces de Bubulcus ibis peuvent être distinguées les unes des autres au mieux par leurs couleurs durant la phase de reproduction. Les plumes nuptiales varient de l'or sombre à la cannelle foncé. La race Bubulcus ibis seychellarum a tendance à avoir des ailes courtes et Bubulcus ibis coromandus un tarse plus long que les autres sous espèces (HANCOCK et KUSHLAN, 1989). La systématique du Héron garde-boeufs a de tout temps connu des changements. Actuellement, l'espèce est dénommée Ardea ibis et classée dans la sous famille des Ardeinae, tribu des Ardeini (KUSHLAN et HANCOCK, 2005). 3- IdentificationB. ibis est un petit héron de couleur blanche. Les sexes sont semblables. On peut le confondre avec d'autres petits hérons de tailles presque égales comme l'Aigrette garzette. Son bec court est jaune. L'iris est jaune pâle. Les jambes plutôt courtes et vert foncé peuvent de loin paraître noires. Sa longueur varie entre 50 et 56 cm et son envergure de 90 à 96 cm (ETCHECOPAR et HÜE, 1964 ; GEROUDET, 1978 ; PETERSON et al., 1986-2006 ; JONSSON, 1994 ; WHITFIELD et WALKER, 1999 ) (Fig. 3). Les adultes en été ont un plumage entièrement blanc sauf de longues plumes sur la calotte, le bas de la gorge et au centre du manteau qui sont chamois rosâtre ou chamois orangé. En hiver, ces plumes palissent beaucoup au point de donner un ensemble blanc. Les pattes sont rouges vineux sombres en été et vert plus au moins sombre en hiver (ETCHECOPAR et HÜE, 1964 ; DUBOURG et al., 2001). Chez les jeunes, le plumage est blanc sans plumes roussâtres, bec jaune, pattes brun verdâtre (PETERSON et al., 2006). Dès l'âge de deux à cinq mois, une crête érectile de plumes roux pâle se développe et la couleur du bec devient jaune (BREDIN, 1983).
Figure 3- En haut : B. ibis en dehors de la
saison de reproduction en association avec un Le dimorphisme sexuel est peu net, les deux sexes sont pratiquement semblables. En dehors de la période de reproduction, les adultes ressemblent aux jeunes, seule une variation dans les mensurations existe, les femelles sont plus petites, mais ceci n'est pas décelable sur le terrain (BLAKER, 1969 ; VOISIN, 1991). En hiver, la femelle se différencie du mâle par la longueur moindre des plumes du manteau et du bas de la gorge (ETCHECOPAR et HÜE, 1964). Comme tout Ardéidé, le garde-boeufs vole le cou replié en forme de « S », les pattes sont tenues en arrière dépassant fortement la queue, les doigts allongés et le pouce généralement dressé (BARRUEL, 1949 ; DORST, 1971a). Les garde-boeufs se manifestent vocalement par des émissions vocales et nasales rauques, émises lors des cérémonies de salutations et des cris d'alarmes parfois légers et parfois assourdissants de type «kok et kaah» (VOISIN, 1991). Par ailleurs, les jeunes héronneaux font entendre des cris de peur et d'inconfort comme «eeeh» (BLAKER, 1969). 4- Répartition géographique4.1- Dans le mondeD'après SIEGFRIED (1978) et FRANCHIMONT (1986a), le Héron garde-boeuf est d'origine faunique Indo Africaine, c'est à dire que ses lieux d'origine sont les suivants : toute l'Asie méridionale et le Japon et toute l'Afrique tropicale au sud du Sahara y compris le Madagascar (Fig. 4). A partir des lieux d'origine, la distribution du héron garde-boeufs, au cours de ces cinquante dernières années, est passée par un processus de continuel accroissement dont la conséquence a été une aire de distribution presque cosmopolite (HANCOCK et KUSHLAN, 1989 ; LOWE, 1994 ; WHITFIELD et WALKER, 1999 ; DUBOURG et al., 2001). Cette espèce habite toute l'Afrique, à l'exception du Sahara aride, ainsi que le Madagascar (GEROUDET, 1978 ; HANCOCK et KUSHLAN, 1989). D'après HEIM DE BALSAC et MAYAUD (1962), le Héron garde-boeufs se rencontre communément dans la région tellienne en Tunisie, la plaine des Trifa au Maroc orientale et les plaines du Maroc espagnol jusqu'à Marrakech. En Asie, on le trouve dans la région Iranienne qui s'étend vers l'est et le sud-est à partir du Pakistan, de l'Inde, du Népal et du Bangladesh jusqu'aux îles méridionales des Figure 4- Repartition geographique mondiale du Heron garde-bceufs, Bubulcus ibis ibis (BREDIN, 1983, modifiee).
Maldives et au Sri Lanka. Son aire continue vers l'est à travers le sud-est de la Chine, la Birmanie et les Philippines. Vers le nord, le Héron garde-boeufs se trouve en Corée et dans le sud du Japon (HANCOCK et KUSHLAN, 1989). Il est observé aux îles Maldives, à Aldabra Atoll, pour la première fois par ABBOTT (RIDGWAY, 1895 in BENSON, 1967) et indiqué comme arrivé récent dans cette région par GAYMER depuis 1964 (GAYMER, 1967). En Europe, son aire de distribution couvre l'Espagne, le Portugal et la France, et vers l'est Israël et la Turquie (GEROUDET, 1978 ; HANCOCK et KUSHLAN, 1989). En Amérique, on le trouve au nord et au sud. Il se reproduit sur tout le littoral du Mexique, en Amérique centrale et aux Antilles. Le Héron garde-boeufs a établi tout d'abord des populations reproductrices en Floride et au Texas au début des années 50. En quelques années, il a étendu son aire nord Américaine le long de la côte atlantique jusqu'au Canada ; il s'est installé en Californie et jusqu'au Chili et l'Argentine au sud (HANCOCK et KUSHLAN, 1989 ; HAFNER, 1994). HANCOCK et KUSHLAN (1989), délimitaient l'aire de distribution de l'espèce à une longitude d'environ 45° N sur les deux masses terrestres de l'Amérique du Nord et de l'Eurasie et à une latitude méridionale de 35 à 40° S environ. Cette limite sud a été dépassée, atteignant récemment le sud des îles Falkland et la Terre de Feu à 55° S (MORALES in KUSHLAN et HAFNER, 2000) et même près de l'Antarctique à 61°23' S 63°39' O à une température de l'air de 1° C (CLARK, 1985). GAUTHIER-CLERC et al. (2002), ont observé un adulte harcelé sur une colonie du Pingouin Royal Aptenodytes patagonicus à Crozets (48°35'S, 68°43'E) au sud de l'océan Indien. 4.2- En AlgérieEn Algérie, l'espèce était jadis nicheuse aux anciens lacs Halloula et Fetzara (extrême nord-est) et peut être ailleurs dans le Tell (HEIM DE BALSAC et MAYAUD, 1962). Sur les plaines littorales un peu humides (régions d'Oran et d'El Kala), c'est l'Ardéidé le plus commun à toutes les époques de l'année (LEDANT et al., 1981). Il hiverne dans les mêmes zones citées ainsi que dans la vallée de Chlef jusqu'à Khemis Melyana, dans la Mitidja, à Mascara, dans les plaines côtières de Béjaia, au cap de Fer et dans la plaine de Fetzara (LEDANT et al., 1981). METZMACHER (1979) in ISENMANN et MOALI (2000), a signalé plusieurs colonies entre El Asnam et Hassi-El-Ghella dans les années 1970, alors que CHALABI et al. (1985) in ISENMANN et MOALI (2000), ont observé plusieurs centaines d'individus en juillet 1976 et mai 1984 dans la région d'El Kala, au lac Tonga, mais sans preuve de nidification. Plus récemment, l'espèce est devenue nicheuse en grand nombre dans plusieurs régions du pays, notamment à Tizi-Ouzou, à Bouira, à Jijel, dans le Constantinois, sur les Hauts Plateaux et à M'Sila (MOALI et ISENMANN, 1993 ; MOALI, 1999 ; ISENMANN et MOALI, 2000). Selon SI BACHIR (2007), jusqu'en 1999, la colonie la plus méridionale est celle de Saïda située à environ 34°50' N. 5- Evolution des populations du Héron garde-boeufs5.1- Causes de l'expansion du Héron garde-boeufsPlusieurs auteurs (SKEAD, 1956 ; SIEGFRIED, 1965 ; HAFNER, 1977-1980 ; HAFNER et MOSER, 1980 ; BREDIN, 1984 ; HANCOCK et KUSHLAN, 1989), s'accordent à dire que l'expansion géographique mondiale du Héron garde-boeufs est favorisée non seulement par des facteurs propres à l'espèce mais aussi par des facteurs liés aux modifications des conditions du milieu. Ils citent : 5.1.1- Facteurs liés à l'espèce elle-même- Une période d'immaturité courte : le garde-boeufs est capable de se reproduire dès la première année (SIEGFRIED, 1971a et b). - La taille moyenne des pontes est importante. Dans la majorité des régions entre 3 et 4 oeufs par nid et éventuellement plus d'une nichée par an (HAFNER, 1970-1977). - La dispersion des jeunes, lorsqu'ils sont rejetés par un groupe par les adultes à la suite d'un surpeuplement (BLAKER, 1971 ; SIEGFRIED, 1971b). - Le développement d'un comportement migratoire, sur de longues distances afin d'éviter les effets néfastes d'un hiver rigoureux (HOPKINS, 1972 ; BOCK et LEPHTIEN, 1976). -La migration et l'erratisme chez le Héron garde-boeufs et d'autres membres de la famille des Ardeidae pourraient être sous un contrôle génétique et programmés pour fournir des mécanismes pour la dispersion des espèces et pour le contrôle des populations (BROWDER, 1973). 5.1.2- Facteurs du milieuD'après BLAKER (1971), SIEGFRIED (1971b), SALLEE (1982), ARENDT (1988), PAREJO et SANCHEZ-GUZMAN (1999), l'intensification de l'agriculture, y compris l'augmentation des surfaces irriguées, l'intensification de l'élevage et l'anthropisation des milieux naturels par la déforestation, seraient des causes responsables de l'extension démographique et géographique du héron garde-boeufs. Selon SIEGFRIED (1978), la diversification du régime alimentaire de B. ibis et sa plasticité alimentaire lui ont valu le nom d'espèce «généraliste opportuniste».
5.2- Etat actuel des populations
5.2.1- Dans le mondeDepuis le début du XXème siècle, le garde-boeufs est une espèce en pleine expansion tant par l'évolution de son aire de répartition que par l'augmentation locale de ses effectifs (SIEGFRIED, 1978 ; BREDIN, 1983). En Afrique, à partir de l'Afrique tropicale, l'espèce s'est répondue à travers toute l'Afrique méridionale dès la fin du siècle dernier (BREDIN, 1983). Ainsi, la colonisation a débuté à l'extrême sud entre la fin du XIXème et le début du XXème siècle avec une augmentation des effectifs et des territoires occupés depuis la fin des années 1920 (VINCENT, 1947 ; SKEAD, 1956 ; SIEGFRIED, 1965-1966a). En Afrique du Nord, le Héron garde boeufs a également colonisé de nouvelles régions de plus en plus méridionales. Au sud du Maroc, par exemple, cette espèce se reproduit jusque dans la région de Ouarzazat (THEVENOT et al., 1982) et à Tiznit depuis 1994 (KUSHLAN et HAFNER, 2000). En Europe, l'espèce n'était jadis commune qu'en Andalousie au XIXème (IRBY, 1895 in SARASA et al., (1994) ; RIDDEL, 1944 ; VOISIN, 1991). Ce n'est qu'à la fin des années 1960 que l'on observa une progression vers le nord (FERNÁNDEZ-CRUZ, 1975). Selon KUSHLAN et HAFNER (2000), depuis les années 1980, la distribution et les effectifs des couples nicheurs du Héron garde-boeufs ont connu un grand essor, estimés à la fin des années 1990, ils dépassent les 100.000 couples. En France, la première observation du garde-boeufs en Camargue remonte à 1953 et sa première reproduction à 1969 (HAFNER, 1970). Il a atteint la Brenne depuis 1992 où il devient aujourd'hui une espèce banale (TROTIGNON, 2005). En Drôme et Ardèche, FATON et coll. (2001), ont recensé deux couples nicheurs en 1995 et 30 couples en l'an 2000. Il a niché pour la première fois en Grèce en 1991 (GOUTNER et al., 1991) et en Roumanie depuis 1997 (MUNTEANU, 1998). L'espèce, nichant depuis les années 80 en Sardaigne, compte actuellement plusieurs centaines de couples (GRUSSU, 1997) et un effectif augmentant en Italie (GRUSSU et al., 2000, GUSTIN et al., 2001). En zone semi-aride, en Israël, plusieurs nouvelles colonies ont été signalées (SHY, com. pers. in KUSHLAN et HAFNER, 2000) et même dans le delta du Nil depuis 1990 (MEININGER et al., 1994). Selon KUSHLAN et HAFNER (2000), en sus de ces nouvelles colonisations, les populations déjà implantées dans le bassin circumméditerranéen ont connu une nette progression des effectifs au cours de la dernière décennie. Sur le continent américain, bien que la première capture de cette espèce remonte à 1937 (BLAKER, 1969), ce n'est qu'à partir des années 1950 que l'on commença à assister à une véritable conquête du nouveau monde (BREDIN, 1983). D'après plusieurs auteurs, le Héron garde-boeufs a colonisé le nouveau monde durant ce dernier siècle. Il est observé aux Etats-Unis aux années 1940 (SPRUNT, 1953 in FOGARTY et HETRICK, 1973 ; PALMER, 1962 in MILLER, 1979) ; déclaré aux états d'Est, en Pennsylvanie, par GRUBE, 1962 (GRUBE, 1963), et à l'Ouest, au Colorado, en 1964 (BAILEY et NIE'DRACH, 1965 in MILLER, 1979). Par ailleurs, le garde-boeufs est observé en Amérique centrale, à Costa Rica en 1954 (SLUD, 1957) et au Mexique par ZIMMERMAN en 1971 tout en citant qu'il est déclaré déjà en 1964 par ALDEN (ZIMMERMAN, 1973). En Amérique du Sud, B. ibis est signalé pour la première fois au Pérou par STOTT en 1957 (MC FARLANE, 1975). Ce dernier auteur ajoute qu'à cause de sa grande dispersion, le Héron garde-boeufs est observé dans plusieurs régions du Pérou, à l'Est et à l'Oust, même à des altitudes de 3.500 m et le long de toute la zone côtière, bien qu'il est plus abondant dans les régions du nord. En effet, sa première nidification à Cuba est notée en 1957 (ARENDT, 1988) et en Colombie en 1958 (LEHMANN, 1959). KREBS et al. (1994) et BROOKS et DA'VALOS (2001), signalent sa dispersion aux îles Caraïbes (Antilles) à partir de ces régions (Amérique du Sud) durant cette même période (aux années 1950) tout en soulignant que sa dispersion est continue jusqu'à nos jours (observé pour la première fois à l'Île-à-Vache aux années 1990-2000). Selon BREDIN (1983), en plus de l'expansion naturelle, certains individus furent introduits en Australie par des éleveurs pensant qu'ils débarrasseraient le bétail de ses parasites. Au Japon, IKEDA (1956), a noté un accroissement sensible de ses effectifs dès les années 1950. Parallèlement à cette expansion, la sous espèce B. i. coromandus à commencer à coloniser l'Australie, effectivement à partir des années 1940 (MORRIS, 1970 in MADDOCK, 1990 ; MADDOCK et BRIDGMAN, 1992) et par la suite elle a atteint la Nouvelle Zélande aux années 1960 (TURBOTT et al. 1963 ; GRANT, 1964), période depuis laquelle le nombre d'oiseaux n'a pas cessé d'augmenter (HEATHER, 1978-1980-1982-1986-1991 ; PRATT, 1979 ; CHILD, 1985 ; MORRISON, 1987 ; JACKSON et OLSEN, 1988 ; MADDOCK, 1990 ; MADDOCK et GEERING, 1993 ; DOWDING, 1991). 5.2.2- En AlgérieLe nord Algérien montre une dynamique évidente des populations du héron gardeboeufs tant en hivernation qu'en nidification ; ainsi, cette espèce qui ne peuplait que certaines zones est devenue très répandue dans plusieurs régions du pays. On la trouve actuellement hivernante et nicheuse à Tizi-Ouzou (FELLAG, 1995 ; BENTAMER, 1998 ; BOUKHEMZA, 2000), à Béjaia (BENALLAOUA et BENAIDA, 1997 ; DOUADI et CHERCHOUR, 1998 ; SI BACHIR, 2007), à Sétif, à El-Khroub et à Ain-Smara (SI BACHIR, 2007). Selon SI BACHIR (2007), en moins de 30 années, l'aire de nidification de l'espèce a connu une expansion de près de 2° de longitude vers le sud (Fig. 5). 6- Migration, erratisme et sédentarisationIl est difficile de distinguer la migration et la dispersion chez les garde-boeufs, car ces derniers ont une tendance marquée pour le vagabondage. Cette tendance est sans doute en grande partie responsable de la forte extension de l'aire de distribution qui peut être caractérisée par des incursions répétées, des colonisations temporaires, des régressions et, selon les conditions, l'établissement dans des régions de plus en plus distantes (HANCOCK et KUSHLAN, 1989). Les garde-boeufs se déplacent du sud-ouest de l'Europe vers le sud pour hiverner en Afrique du Nord (HANCOCK et KUSHLAN, 1989). Dans les principaux quartiers de reproduction, tels la Péninsule Ibérique ou le sud de la France, l'espèce est aujourd'hui largement sédentaire et ne s'éloigne qu'en automne par des distances relativement faibles, quelques centaines de kilomètres au plus (KUSHLAN et HAFNER, 2000 ; SARASA et al., 1994). Le détroit de Gibraltar constitue le seul endroit où des mouvements de migration sont observés entre l'Europe et l'Afrique du Nord mais les migrateurs espagnols ne dépassent vraisemblablement pas le Maroc (HEIM DE BALSAC et MAYAUD, 1962). Les reproducteurs nord-africains sont considérés comme résidants mais certainement pas sédentaires. Ils se dispersent en direction du sud le long de la bande côtière, suivant peut être davantage les fluctuations des ressources alimentaires qu'un logique schéma migratoire saisonnier. Dans leurs déplacements, les populations nord-africaines ainsi que celles de l'Ethiopie évitent le Sahara (HANCOCK et KUSHLAN, 1989 ; HEIM DE BALSAC et MAYAUD, 1962). Selon HEIM DE BALSAC et MAYAUD (1962), les quelques sujets observés au Sahara ou en deçà des Atlas ne représentent que des cas d'erratisme.
(A)
(B) Figure 5- Evolution de l'aire de nidification avec
importance des colonies (A) et 7- Biologie de la reproduction
7.1- Maturité sexuelleSelon BREDIN (1983), les garde-boeufs sont généralement monogames et sont capables de se reproduire dès la première année d'âge. Pour SIEGFRIED (1971a et b), la maturité sexuelle de B. ibis est atteinte à l'âge d'un an alors que le plumage adulte n'est pas encore complètement acquis. Toutefois, la proportion d'oiseaux se reproduisant dès cet âge n'est pas connue. 7.2- Choix des colonies de nidification et du site du nidComme les dortoirs, les colonies de nidification sont situées dans des arbres et une colonie peut accueillir plusieurs milliers de couples de B. ibis uniquement ou de plusieurs espèces de Ciconiiformes (HAFNER, 1977 ; BLAKER, 1969). En Camargue, d'après TOURENQ et al. (2004), le Héron garde-boeufs choisi les sites de reproduction et préfère ceux entourés de rizières, milieux utilisés en alimentation. En dehors de la période de reproduction, la colonie est soit désertée soit utilisée comme dortoir (BLAKER, 1969 ; GEROUDET, 1978). Toutefois, les colonies de nidification ne sont construits que dans des sites sécurisés contre les aléas climatiques et les prédateurs, généralement à proximité de milieux de gagnages et où des branches sèches procurent des matériaux de construction (FRANCHIMONT, 1985 ; SIEGFREID, 1972 ; SI BACHIR, 2007). Les essences d'arbres utilisées diffèrent d'une région à une autre. La héronnière peut s'établir sur des jujubiers épineux (GEROUDET, 1978), parfois dans des bosquets de Pins, d'Eucalyptus et même de Chêne liège (ETCHECOPAR et HÜE, 1964). Parmi les arbres porteurs de nids on trouve aussi le Peuplier blanc, l'Ormeau, le Frêne ainsi que le Figuier, en Camargue (HAFNER, 1980) ; des Caroubiers, des Pistachiers et des Oliviers à Asjène, au Maroc (FRANCHIMONT, 1985). Ils s'installent sur l'Acacia (Acacia confusa) et l'Eucalyptus, (Eucalyptus sp.) en Chine (LIANG et al., 2006) ; l'Eucalyptus, le Platane, le Frêne, le Cyprès et l'Araucaria en Algérie (BOUKHEMZA et al. 2006). Selon HAFNER (1980), SAMRAOUI et al. (2007) et SI BACHIR et al. (2008) l'installation des nids dans les arbres de reproduction a lieu d'abord dans les zones centrales ensuite elle s'étale vers la périphérie. Ces derniers auteurs rajoutent que le centre offre de meilleures conditions aux nicheurs ainsi qu'une meilleurs protection pour leurs nouveaux nés. En Inde, HILALUDDIN et al. (2003), et en Camargue, DAMI et al. (2006), dans une colonie mixte du héron garde-boeufs et de l'Aigrette garzette, le garde-boeufs occupe les meilleurs et les plus hauts emplacements et déplace l'aigrette aux zones périphériques. SI BACHIR (2007), démontre que lors de leur nidification, les garde-boeufs s'installent de préférence sur les arbres les plus hauts dans la position la plus haute et la plus proche du tronc et réussissent, de ce fait, à avoir un meilleur succès de reproduction. 7.3- Formation des couples et parades nuptialesAu stade de l'association nuptiale des deux sexes, le mâle délimite un territoire. Il choisit une place qu'il défend et à ce moment il devient agressif (GEROUDET, 1978 ; HANCOCK et KUSHLAN, 1989). La femelle initie la formation du couple en battant ses ailes sur le dos du mâle (VOISIN, 1991). Dans les salutations d'accueil, le mâle parcours lentement ses perchoirs ou se balance d'un pied sur l'autre en émettant des cris. Attirées par ce comportement, les femelles viennent roder autour du séducteur, se perchant tout près et l'observent en silence, le plumage collé au corps (BLAKER, 1969 ; GEROUDET, 1978 ; VOISIN, 1991). Il n'est pas rare qu'un mâle s'accouple avec plusieurs femelles (BLAKER, 1969 ; GEROUDET, 1978 ; FUJIOKA et YAMAGISHI, 1981 ; MC KILLIGAN, 1990). 7.4- AccouplementLa copulation a lieu au nid ou à moins de 50 cm de ce dernier. La femelle s'accroupit respire à petits coups. Le mâle saute sur cette dernière en s'accrochant des pieds et saisissant les plumes de la calotte ou du cou et en exhibant des mouvements rythmiques avant de baisser sa queue en vue d'établir le contact cloacal en battant des ailes pour maintenir son équilibre. Durant cet acte, le mâle caresse du bec les plumes du cou et de la tête de la femelle (BLAKER, 1969 ; VOISIN, 1991). 7.5- Construction du nidLa construction du nid est assurée principalement par la femelle, tandis que la collecte des matériaux est surtout le travail du mâle qui collecte des branchettes et des petits bâtons morts et séchés de 1 à 30 cm de longueur généralement trouvés par terre, ramassés avec le bec dans les endroits les plus proches de la héronnière, mais peut aussi arracher des brindilles d'arbres environnant la héronnière (JENNI, 1969 ; BLAKER, 1969 ; HAFNER, 1977 ; BREDIN, 1983 ; WHITFIELD et WALKER, 1999 ; DUBOURG et al., 2001). 7.6- PonteLa ponte a généralement lieu environ 7 jours après la formation des couples (BLAKER, 1969 ; HAFNER, 1980). Le nombre d'oeufs par ponte varie entre 2 et 7 avec une moyenne de 3 à 4 oeufs par nid. Les oeufs blancs avec une nuance vert pâle ou bleu, sont de forme ovale large et un peu pointue (HEIM DE BALSAC et MAYAUD, 1962 ; ETCHECOPAR et HÜE, 1964 ; GEROUDET, 1978). DARMALLAH (1989), estime la taille moyenne de la ponte à 3,3 oeufs dans la région d'El Kala. En Albufera, en Espagne, PROSPER et HAFNER (1996), signalent des moyennes de 3,4 à 3,9 oeufs et FRANCHIMONT (1985), à Asjène au Maroc, l'estime à 3,3 oeufs par nid. En Inde, HILALUDDIN et al. (2003), notent que la taille moyenne de la ponte et de 3,03 oeufs, contre 2,92 oeufs par nid dans la région de Béjaia (SI BACHIR et al., 2000). Selon SAMRAOUI et al. (2007) la taille moyenne de la ponte dans la colonie de Sidi Achour, à Annaba, est de 3,1 oeufs par nid. La ponte est déposée parfois dès janvier, comme au Maroc (FRANCHIMONT, 1985). En Camargue, elle est réalisée en avril mai (HAFNER, 1977), alors qu'à El Kala, DARMALLAH (1989), note que la ponte est déposée dès le début du mois de mars. Dans la région de Béjaia, selon KASRI et LALOUNI (1998) et SI BACHIR (2007), le nombre d'oeufs par ponte varie de 1 à 5 avec exceptionnellement des pontes de 6 et 7 oeufs. 7.7- Couvaison et éclosion des oeufsLa couvée chez le Héron garde-boeufs commence après la ponte d'un oeuf, les deux sexes font l'incubation et les oeufs ne sont jamais laissés sans surveillance (BLAKER, 1969 ; BREDIN, 1983 ; VOISIN ,1991). Chez un couple observé par BLAKER (1969), l'un couvait de 9h 00' à 16h 00' et l'autre de 16h 00' à 9h 00' et les oeufs n'étaient jamais laissés non gardés. Le temps de couvaison peut varier légèrement, il dure de 22 à 24 jours (JENNI, 1969 ; HAFNER, 1977 ; RUIZ et JOVER, 1981 ; WHITFIELD et WALKER, 1999). A El Kala, DARMALLAH (1989), a noté une durée de couvaison moyenne de 26 jours. Les oeufs de B. ibis sont éclos, généralement, à des intervalles d'un à deux jours, ce qui signifie, pour une ponte de 5 oeufs, que les éclosions s'échelonnent sur presque une dizaine de jours (BLAKER, 1969). A El Kala, la période d'éclosion se situe entre la fin mai et le début juin (DARMALLAH, 1989) et entre la fin avril et le début mai en Camargue (HAFNER, 1977). En Espagne, PROSPER et HAFNER (1996), situent la période d'éclosion à la fin du mois d'avril. 7.8- Nourrissage et élevage des jeunesAprès la naissance des poussins, on note deux phases principales d'activité. Elles correspondent à l'époque de gardiennage qui dure jusqu'à l'âge de 15 jours et à la phase où les poussins restent seuls dans les nids après avoir dépassé 15 jours d'âge (HAFNER, 1980). Ce sont les deux parents qui élèvent leurs poussins durant leur première semaine d'âge. Les adultes, debout, bec pointé en bas, laissent tomber les proies entre les jeunes poussins dont les becs sont ouverts en quête de nourriture. Le premier né de ces poussins reçoit la plus grande partie de nourriture, car il est le plus fort et le plus volumineux et arrive le premier à la rencontre des parents qui nourrissent indifféremment leur progéniture (VOISIN, 1991). 7.9- EnvolLes jeunes commencent leurs excursions hors du nid dès le 9ème jour. A 15 jours d'âge, les poussins peuvent quitter le nid mais pas la héronnière, dans le but d'attendre le retour des adultes vers les nids (BLAKER, 1969 ; GEROUDET, 1978 ; FRANCHIMONT, 1986a ; VOISIN, 1991). Ils commencent alors à grimper les branches avoisinantes du nid et à battre des ailes à l'âge de 25-30 jours. Les premiers vols sont observés à 25 jusqu'à 30 jours d'âge. Les poussins volent jusqu'à une distance de 4 m et volent d'avantage et commencent à visiter les environs, jusqu'à 600 mètres du nid, à partir de 40 à 45 jours d'âge (BLAKER, 1969). 8- Ecologie trophique8.1- Composition du régime alimentaireDe nombreux auteurs dans le monde ont montré que le Héron garde-boeufs se nourrit essentiellement d'insectes : KADRY-BEY (1942) en Egypte, IKEDA (1956) au Japon, BURNS et CHAPIN (1969) en Louisiane (U.S.A.), SIEGFRIED (1966b, 1971c, 1978) en Afrique du sud, FOGARTY et HETRICK (1973) en Floride (U.S.A.), BREDIN (1983, 1984) en Camargue (France) et RUIZ et JOVER (1981) en Espagne, RICHARDSON et TAYLOR (2003) en Australie. En Algérie, les résultats les plus notables, obtenus suite à l'analyse des pelotes de réjection des adultes ou des régurgitats de poussins montrent que l'espèce a principalement un régime alimentaire insectivore. Selon la région d'étude et la période de l'année, l'espèce se nourrit essentiellement d'orthoptères et de coléoptères (DOUMANDJI et al., 1992-1993 ; BENTAMER, 1998 ; HARIZIA, 1998 ; BOUKHEMZA et al., 2000-2004 ; SI BACHIR et al., 2001). Le garde-boeufs est un oiseau insectivore par excellence. Toutefois, au sein d'une même région, sa nourriture subit des variations au cours des différentes saisons et des variations au cours des années. Parmi les invertébrés non aquatiques, SIEGFRIED (1966b-1971c), note l'importance qu'ont les vers de terre dans l'alimentation du garde-boeufs pendant la saison des pluies en Afrique du sud. En période de gel, les vers de terre, indisponibles, sont remplacés par des petits mammifères en Camargue (BREDIN, 1983-1984) et dans le delta Del Ebro en Espagne (RUIZ, 1985). Des données assez ponctuelles, signalent également la prédation sur des poissons, des reptiles, des oiseaux et des mammifères (DUXBURY, 1963 ; CUNNINGHAM, 1965 ; HERRERA, 1974 ; TAYLOR, 1979). La consommation d'ectoparasites telles les tiques paraît également très occasionnelle voire même exceptionnelle (BATES, 1937 ; BEVEN, 1946 ; HOLMAN, 1946 ; SKEAD, 1963). 8.2- Milieux d'alimentationContrairement aux autres Ardéidés, le Héron garde-boeufs est un oiseau semiaquatique et dans certaines régions il est essentiellement «terrestre». Parmi les milieux aquatiques fréquentés, seuls les zones d'eau douce ou légèrement saumâtres sont exploitées. Selon VOISIN (1978), aucun garde-boeufs n'a jamais été vu dans les salins (exploitations de sel) de Camargue ; l'espèce niche pourtant non loin de là. Le garde-boeufs chasse et pêche, le plus souvent, dans des milieux pourvus d'une végétation assez abondante. Le Héron garde-boeufs fréquente principalement les marais, les garrigues dégradées, les dépôts d'ordures, les champs labourés, les cultures basses, les mares temporaires, les plaines basses, les deltas ou les larges vallées, où cet échassier jouit des ressources abondantes pendant toute l'année, comme il fréquente les prairies, les zones boisées et les marécages (ETCHECOPAR et HÜE, 1964 ; DORST, 1971a ; VOISIN, 1979-1991 ; BREDIN, 1983 ; FRANCHIMONT, 1986b ; DOUMANDJI et al., 1988). Cela n'exclut nullement l'exploration des collines et des zones arides quand elles sont parcourues par le bétail (GEROUDET, 1978). L'espèce fréquente également des lieux d'importance mineure, comme les jardins cultivés près des agglomérations (CRAUFURD, 1965) ainsi que les bords des ruisseaux. Ces derniers ne sont fréquentés que pour s'alimenter en eau (FRANCHIMONT, 1986b). Dans plusieurs régions du monde, les dépôts d'ordures sont de plus en plus fréquentés par le garde-boeufs (HAFNER, 1977 ; RENCUREL, 1972 ; DEAN, 1978 ; FRANCHIMONT, 1986b ; DOUMANDJI et al., 1988 ; BOUKHEMZA, 2000 ; YORIO et GIACCARDI, 2002 ; SI BACHIR, 2007). 8.3- Association avec d'autres animauxLes garde-boeufs peuvent se nourrir isolément, mais l'association est la règle générale qui peut être intra ou interspécifique. Dans le cas d'association intraspécifique les groupes sont alors composés d'individus de différents âges, des deux sexes et avec d'autres échassiers (SKEAD, 1956). Par ailleurs, l'association avec d'autres animaux domine très largement l'écologie et le comportement alimentaire du Garde-boeufs. Ce dernier est le plus souvent associé à des mammifères à l'origine sauvages et maintenant à du bétail domestique (BLAKER, 1969). Grâce à cette technique, ils absorbent 50 % de plus de nourriture que les autres oiseaux, pour un effort moindre (HEATWOLE, 1964 in BLAKER, 1971 ; WHITFIELD et WALKER, 1999). Plus récemment, les garde-boeufs ont pris l'habitude de suivre même les machines agricoles dans toutes les régions couvertes par son aire de répartition (BREDIN, 1983 ; WHITFIELD et WALKER, 1999 ; DUBOURG et al., 2001). 8.4- Mode de chasseChez le garde-boeufs, avant «le coup de bec», le cou est tendu puis le bec est propulsé vers l'avant légèrement ouvert et se ferme sur la proie. Les petites proies sont avalées d'un seul coup avec un rapide hochement de la tête (BREDIN, 1983). Les grosses proies sont tuées par une compression au niveau du cou, le bec agissant comme une puissante paire de pinces avec une agitation latérale violente (BLAKER, 1969). Le Héron garde-boeufs est spécialiste dans la marche sur la terre. En suivant le bétail et les machines agricoles, il accroît l'efficacité de la recherche de nourriture et se procure d'avantage d'aliments que s'il se nourrissait seul. En suivant les commensaux, les gardeboeufs utilisent des comportements variés, incluant la marche rapide, la course, la capture à l'affût des insectes, le glanage, le balancement latéral de la tête et du cou, la marche lente, les sauts successifs et les captures aériennes des insectes (BREDIN, 1983 ; HANCOCK et KUSHLAN, 1989). 8.5- Digestion des proiesLes hérons sont connus pour avoir des sucs digestifs très acides et très puissants, certains peuvent même digérer les os de poissons (GEROUDET, 1978). BREDIN (1983), a montré que lors du passage dans le tractus digestif du garde-boeufs, des dents de moutons ont été totalement digérées ne laissant que peu de traces et uniquement dans les pelotes. La vitesse de digestion est élevée et varie suivant les proies : les poissons et les grenouilles, par exemple, sont digérés dans les six heures (VINOKUROV, 1960 in BREDIN, 1983). Les hérons, régurgitent des pelotes qui contiennent des parties non digestibles des proies consommées : poils, os, plumes, chitine (HIBBERT-WARE, 1940 ; KUSHLAN, 1978). Généralement, une seule pelote est rejetée par jour avant le lever du soleil (BREDIN, 1993). 8.6- Accessibilité des proies et rythmes d'activités alimentairesPour les proies, le terme «d'accessibilité» est préféré à celui de « disponibilité » qui est généralement réservé pour la nourriture. Il tient compte de la présence et de l'absence des «espèces proies» donc de leur cycle biologique, mais aussi de certains de leurs comportements qui les rendent accessibles aux garde-boeufs (BREDIN, 1983). Aussi, les garde-boeufs adoptent leur mode de chasse en fonction du type de proies accessibles : à la chasse à l'affût par exemple correspond la capture de mammifères. Le phénomène d'association est également en relation étroite avec l'accessibilité des proies que délogent le bétail et les machines agricoles (BREDIN, 1983). En saison défavorable (saison sèche et hiver), les gardes-boeufs cherchent à passer le plus de temps sur les gagnages, ils quittent alors les dortoirs plus tôt. Une fois sur les gagnages, ils consacrent la plus grande partie de la journée à s'alimenter que ce soit derrière les tracteurs ou non. Juste après la période d'élevage des jeunes, le temps consacré à l'alimentation est très important (BREDIN, 1983). 9- Facteurs de menace et de mortalitéLes populations de B. ibis sont contrôlées par plusieurs facteurs naturels et bien d'autres reviennent à l'effet de l'homme. Ces facteurs interviennent essentiellement aux stades poussins, par contre, les garde-boeufs adultes sont moins confrontés aux menaces de mortalité. Parmi les facteurs naturels nous citons : > L'asynchronisation des éclosions Selon LACK (1968), la différence d'âge des poussins permettrait d'ajuster au mieux le taux de mortalité aux disponibilités alimentaires. Par manque de proies, les parents ne peuvent subvenir aux besoins alimentaires de leur progéniture ce qui accentue l'agressivité entre les poussins. Parmi ces derniers, les plus âgés sont bien avantagés lors du nourrissage et les plus jeunes, se trouvant quasi privés de nourriture et meurent par inanition (FUJIOKA, 1985 ; INOUE, 1985 ; FRANCHIMONT, 1986c ; MOCK et PARKER, 1986). > La prédation Dans la littérature, les données relatives à la prédation par les oiseaux dans différentes régions du monde concernent généralement aussi bien les oeufs que les jeunes poussins. Ce sont, essentiellement, des prédateurs aériens tels des corvidés (FRANCHIMONT, 1986c), des rapaces diurnes et nocturnes (BURGER, 1981 ; HAFNER, 1977), des Goélands (SIEGFRIED, 1972) et d'autres espèces d'Ardéidés (BURGER, 1981 ; HANNANE, 1981 ; SIEGFRIED, 1972). SIEGFRIED (1972), signale également la prédation par la Poule d'eau, Gallinula chloropus, la Poule sultane, Parphyrio parphyrio et par l'Ibis sacré, Threskiornis aethiopicus. Les prédateurs terrestres, peu fréquents, s'attaquent surtout aux nids près du sol. Ils sont représentés essentiellement par des mammifères et des reptiles (RIDDEL, 1944 ; TAYLOR et MICHAEL, 1971 ; HANNANE, 1981 ; BURGER, 1981 ; VOISIN, 1991 ; HAFNER, 1977). > Les aléas climatiques Beaucoup de nids ainsi que leurs contenus sont détruits par les vents violents, les orages, les ouragans et les fortes pluies (MEANLEY, 1955 ; JENNI, 1969 ; HAFNER, 1977- 1978, HANNANE, 1981). HAFNER (1994), a montré que l'effectif de la population nicheuse est affecté par l'action des facteurs climatiques sévissant en hiver sur et à proximité des sites de reproduction en Camargue et par la hauteur des pluies reçues en automne et en hiver au sud de l'Espagne. > Le cannibalisme En période de famine, les hérons garde-boeufs adultes peuvent manger leurs propres poussins, ce phénomène est également appelé Kleptonisme (SIEGFRIED, 1972 ; FRANCHIMONT, 1986c). > Le parasitisme Plusieurs types de parasites se développent sur les hérons garde-boeufs. Certaines espèces peuvent entraîner la mort de ces derniers à tout âge. RENCUREL (1972), décrit des vers du genre Heterakis qui parasitent les garde-boeufs au Maroc ; la tique Argas persicus est notée par SKEAD (1956) en Afrique du sud. MURATA et al. (1998), ont dénombré un total de 22 trématodes du genre Pegosomum dans le canal biliaire de B. ibis. En outre, l'homme est également à l'origine de perturbations pouvant constituer des facteurs de menace : > Les dérangements humains En cas d'intrusion humaine dans la héronnière, les adultes quittent leurs nids et n'y reviennent qu'une fois le danger passé. Les oeufs et les poussins se trouvent alors exposés aux prédateurs, à la pluie, au vent et au soleil, ce qui accroît leurs taux de mortalité (FRANCHIMONT, 1986c). Ce dernier auteur rajoute, suite à l'intrusion humaine, des vols de brindilles par les voisins causent également la destruction de certains nids abandonnés et dont les contenus tombent sur le sol. Aussi, le dérangement humain pousse les poussins à pénétrer dans les territoires d'adultes qui les reçoivent avec de violents coups de bec parfois mortels. L'intrusion de l'homme dans une colonie provoque également des régurgitations trop fréquentes des poussins, cela pourrait accélérer la mort des plus faibles sujets par inanition (FRANCHIMONT, 1985). > La dégradation du milieu naturel Les garde-boeufs, autant que le reste des Ardéidés et autres oiseaux, sont susceptibles d'intoxications dues à l'absorption de produits chimiques et ils peuvent être, à cet effet, utilisés comme bio-indicateurs de l'état de contamination de l'environnement par l'homme (BURGER et GOCHFELD, 1993-1997). Au Pakistan, BOSTAN et al. (2007), ont constaté des concentrations importantes de métaux lourds, Plomb (Pb), cadmium (Cd), Chrome (Cr), Cobalt (Co), Argent (Ag), Nickel (Ni), dans les oeufs et les régurgitas de deux colonies de hérons garde-boeufs, dépassant la limite maximale de résidus standardisée par l'Agence de la Protection de l'Environnement (A.P.E.). Ainsi, l'intoxication des adultes par le D.D.E. est à l'origine de la fragilité des oeufs qui se brisent facilement au nid. Ces derniers présentaient une mince coquille (OHLENDORF et al., 1979). La diminution des effectifs du garde-boeufs dans le Delta du Nil et de la région du canal de Suez est un résultat direct de l'épandage du D.D.T sur les cultures du coton (MULLIÉ et al., 1992). Dans ce chapitre nous présenterons le cadre d'étude qui est la région de Batna et les deux zones échantillons, El Madher et Merouana, dans lesquelles s'est déroulée la présente étude. Partie I : Présentation générale de la région de Batna1- Situation géographiqueLa wilaya de Batna est localisée dans la partie orientale de l'Algérie entre les 4° et 7° de longitude Est et 35° et 36° de latitude Nord (ANONYME, 2005) (Fig. 6). D'une Superficie de 12.038,76 km2, le territoire de la wilaya de Batna s'inscrit presque entièrement dans l'ensemble physique constitué par la jonction de deux Atlas Tellien et Saharien ce qui représente la particularité physique principale de la région et détermine, de ce fait, les caractères du climat et les conditions de vie humaine (ANONYME, 2005). Administrativement, la wilaya de Batna est composée de 21 daïras et 61 communes, elle est limitée au nord par les wilayas d'Oum El Bouaghi, Mila et Sétif, à l'est par la wilaya de Khenchela, au sud par la wilaya de Biskra et à l'ouest par la wilaya de M'sila (ANONYME, 2005) (Fig. 6). 2- OrographieLa topographie de la région de Batna est subdivisée en trois grands ensembles : les Hautes plaines Telliennes, les reliefs Montagneux et les Hautes plaines Steppiques. > Hautes plaines Telliennes Au nord de la wilaya de Batna se distingue le domaine des hautes plaines telliennes qui matérialisent la limite nord de la wilaya, particulièrement avec une série de petits chotts. D'une manière générale, ces hautes plaines sont composées de vastes glacis plus ou moins encroûtés. Les altitudes dans cette zone varient entre 800 et 950 m en moyenne et les pentes excèdent rarement 3 %, c'est la raison pour laquelle elle prend le nom des hautes plaines (ANONYME, 2005). > Reliefs Montagneux (la jonction des deux Atlas) L'Atlas Tellien par l'ensemble des monts du Hodna, du Bou Taleb et les monts de Bellezma, et l'Atlas Saharien par les Aurès, le Dj. Metlili et les monts du Zab, forment « l'ossature physique » principale de la région. Les altitudes varient de 700 m (la plus basse altitude localisée dans cet ensemble est située au Dj. Rebaa sur le versant nord des monts du
Figure 6- Situation, limites et subdivisions
administratives de la wilaya de Batna et situation des zones d'etude Zab) à 2.328 m au Dj. Chélia considéré comme le plus haut point de la wilaya et troisième plus haut point de l'Algérie (ANONYME, 2005). > Hautes plaines Steppiques Situées dans la partie ouest de la wilaya de Batna, elles se trouvent encadrées au nord par les monts du Hodna, à l'est par les monts de Bellezma et le Dj. Metlili et au sud par le versant nord des monts du Zab constitué par les Djebels Ammar, Mekmizane et Bouzokma (ANONYME, 2005). 3- HydrographieL'ensemble de la wilaya de Batna est réparti sur 3 grands bassins versants selon la répartition faite par l'A.N.R.H. (Agence Nationale des Ressources Hydriques) (ANONYME, 2005) : - Le Bassin du Constantinois : Situé au nord et nord-est, caractérisé par la présence d'une série de Sebkhas, Garâats et Chotts qui marquent toute une série de cuvettes au passage vers les hauts plateaux Constantinois. - Le Bassin versant du Hodna : Situé à l'ouest, caractérisé par la présence du Chott El Hodna, point d'arrivée de certains Oueds qui se dirigent vers le sud-ouest, en particulier l'Oued de Barika et celui de Bitam. - Le Bassin des Aurès-Nememcha : Occupe la partie sud et sud-est de la wilaya et dont les cours d'eau ont leur exutoire dans le Chott Melrhir en dehors des limites de la wilaya. Tout le réseau hydrographique est étroitement lié aux précipitations ainsi qu'il est surmonté par une atmosphère à très fort pouvoir évaporant (ANONYME, 2005). En effet, il est à signaler la vulnérabilité des ressources en eau dans les trois bassins versants, que ce soit les eaux superficielles ou les eaux renouvelables des nappes souterraines (ANONYME, 2005). 4- Flore et formations végétales caractéristiquesA l'image de la diversité de ses écosystèmes et sous climats, la région de Batna abrite une flore riche est diversifiée. Le parc national de Belezma, aire protégée représentative de la région et qui couvre une superficie de 26.250 ha, renferme un total de 510 espèces végétales, ce qui représente 14 % du potentiel national (ANONYME, 2007a). Les formations végétales sont composées essentiellement de forêts et de maquis. Les forêts de la région de Batna couvrent une superficie d'environ 290.038 ha, soit 24 % de la superficie totale. Une partie très importante de la superficie forestière est localisée au niveau des massifs de Bellezma, Beni Fedhala, Z'gag, Bouarif, Beni Emloul et Aurès (ANONYME, 2005). Le patrimoine forestier est constitué de formations végétales issues essentiellement des peuplements naturels à base de Pin d'Alep (Pinus halepensis), Chêne vert (Quercus ilex), Genévrier oxycèdre (Juniperus oxycedrus), Genévrier de Phénicie (Juniperus phoenicea), Frêne dimorphe (Fraxinus dimorphe), Genévrier Thurifère (Juniperus thurifera) et Cèdre (Cedrus atlantica) (ANONYME, 2005). Les maquis sont d'origine naturelle et sont surtout à base de Chêne vert qui représente près de 80 %. Ils se localisent au niveau de l'ensemble des massifs forestiers de la région, à l'état pur ou mélangé avec d'autres espèces comme le Pin d'Alep et le Cèdre où ils forment un sous étage de ces essences (ANONYME, 2005). Le couvert végétal forestier de la région de Batna se trouve à différents niveaux de dégradation ce qui est le fait d'une forte pression humaine exercée durant plusieurs décennies. Actuellement, cette situation a atteint un stade avancé de dégradation et en certains terrains, l'équilibre écologique est totalement rompu et le couvert végétal forestier a cédé la place à l'installation d'espèces steppiques telles l'Armoise, Artemisia vulgaris et l'Alfa, Stipa tenacissima, particulièrement en zones semi-arides localisées au sud et sud-ouest du territoire de la wilaya qui subissent l'influence désertique (ANONYME, 2005). 5- FauneLa région de Batna possède un milieu biologique assez riche et varié en composantes faunistiques. A titre d'exemple, le parc national de Belezma abrite 387 espèces animales dont 18 espèces de mammifères, 106 espèces d'oiseaux, 19 espèces de reptiles, 3 espèces d'amphibiens et 241 espèces d'invertébrés. Parmi ces espèces, on note la présence de 62 espèces protégées en Algérie (ANONYME, 2007a). Il est à signaler que la région de Batna est limitrophe de la région Sud-Constantinoise, riche en zones humides, avec 5 sites classés Ramsar par leur importance. BECHIM et BACHA (2005), en se penchant sur l'avifaune de ces zones humides, ont recensé 63 espèces aviaires dont la Cigogne blanche et le Héron garde-boeufs. 6- Agriculture6.1- Répartition générale des terresLa superficie totale de la wilaya de Batna est estimée à 1.203.876 ha, répartie sur 744.026 ha pour la superficie agricole totale, 290.038 ha pour les forêts, 166.812 ha pour les terres improductives et 3.000 ha sont occupés par des terres alfatières (Fig. 7) (ANONYME, 2007b).
S.A.U. Bois et forêts Parcours Terres improductives Terres alfatières 0,25 % 20,83 % 19,72 % 24,09 % 35,11 % Figure 7- Répartition des terres de la wilaya de Batna. Cette répartition des terres montre l'importance qu'occupe la superficie agricole totale, qui représente 35,11 % de la superficie totale de la wilaya de Batna. La superficie agricole totale est occupée comme suit : 422.677 ha de surface agricole utile (S.A.U.), 237.426 ha de parcours et 83.923 ha de terres improductives (ANONYME, 2007b). 6.2- Répartition de la S.A.U. par type de cultureLa rigueur climatique de la région de Batna justifie la dominance des cultures extensives en association avec l'élevage et limite l'introduction de quelques types de cultures (Tab. IV). Tableau IV- Répartition de la S.A.U. par type de culture.
(ANONYME, 2007b) La céréaliculture est la principale spéculation cultivée dans la région de Batna, avec 137.162,86 ha soit 32,45 % de la S.A.U. totale. A noter que les emblavures restent étroitement liées à la pluviométrie et sont par conséquent variables d'une année à une autre. La jachère occupe une place non négligeable dans la S.A.U. représentant 58,46 % de celle-ci, soit 247.105 ha. La pratique de la jachère s'impose comme un impératif technique surtout en zones des hautes plaines et montagneuses à vocation céréalière à cause des conditions climatiques. La superficie consacrée aux cultures fourragères, cultures maraîchères et à l'arboriculture et la viticulture reste très réduite et ne représente que 8,76 % de la S.A.U. 6.3- ElevageL'effectif du cheptel dans la wilaya de Batna, jusqu'à la compagne 2006-2007, est évalué à 34.412 têtes de bovins dont 17.431 vaches ; à 398.146 têtes d'ovins dont 214.929 brebis ; et à 156.300 têtes de caprins dont 92.853 chèvres (ANONYME, 2007b). 7- ClimatSelon LACOSTE et SALANON (2001), le climat est l'une des composantes fondamentales d'un écosystème terrestre. A cet effet, il est particulièrement connu que l'influence de la nourriture, comme une ressource, et du climat, comme un agent, affectent la distribution, la migration et la reproduction des oiseaux (ELKINS, 2004 in DENAC, 2006). Beaucoup d'études ont démontré ces dernières années des tendances temporelles à long terme dans les paramètres biologiques qui peuvent être expliqués seulement par le changement du climat. La phénologie des oiseaux a reçu la grande attention, comme ils sont populaires et facilement observables dans la nature (CRICK, 2004 ; GORDO et SANZ, 2006). L'effet des changements climatiques sur les oiseaux a fait l'objet de plusieurs études récentes CARRASCAL et al. (1993), WATKINSON et al. (2004) et ROBINSON et al. (2007) en Bretagne, NILSSON et al. (2006) en Suède, JIGUET et al. (2007) en France et LEMOINE et al. (2007) en Europe centrale. SAINO et AMBROSINI (2007), dans leur étude sur la connectivité climatique entre l'Afrique et l'Europe et son effet sur la migration des oiseaux, ont constaté que, à cause du réchauffement climatique, la plupart des oiseaux migrateurs européens hivernant au Sahel (Afrique), ont anticipé leurs dates d'arrivées aux aires de reproduction pendant ces dernières décennies. Toutefois, cette étude a révélé que les oiseaux migrateurs sont capables de prédire les conditions météorologiques au début de la saison de la reproduction et ajuster, de ce fait, leurs dates de migration et optimiser les dates d'arrivées. A partir de la littérature publiée depuis 1960 sur les premières dates d'arrivées et dernières dates de départs des oiseaux migrateurs de l'Australie, BEAUMONT et al. (2006), assurent que les oiseaux de l'hémisphère sud ont répondu aux changements climatiques de la même manière qu'à l'hémisphère nord. 7.1- Caractéristiques climatiques de la région d'étudeDans la présente analyse, nous avons pris en considération les observations homogènes sur une période de 25 ans allant de 1913 à 1937 selon les données de SELTZER (1946) comparées aux données climatiques récentes recueillies au niveau de la station météorologique d'Ain Skhouna sur une période de 27 ans, allant de 1980 jusqu'à 2006. 7.1.1- TempératuresD'après BARBAULT (2000), la température a une action majeure sur le fonctionnement et la multiplication des êtres vivants et comme elle varie selon un schéma géographique net, les espèces animales et végétales se distribuent selon des aires de répartition souvent définissables à partir des isothermes. Les données thermométriques caractérisant la région de Batna durant les périodes 1913-1937 et 1980-2006 sont reportées dans le Tableau V. Tableau V- Données thermométriques moyennes en (°C) de la région de Batna calculées sur les périodes 1913-1937 et 1980-2006 (M : moyenne mensuelle des températures maximales absolues quotidiennes ; m : moyenne mensuelle des températures minimales absolues quotidiennes ; (m + M) /2 : moyenne mensuelle).
(SELTZER, 1946 ; Station météorologique d'Ain Skhouna) En analysant les données récentes, de la période 1980-2006, nous constatons que janvier est le mois le plus froid avec une température moyenne de 5,36 °C et juillet est le mois le plus chaud avec une moyenne de 25 °C. Les moyennes maximales dépassent 30 °C pour les mois de juillet et août. Les valeurs thermiques maximales comprises entre 20 °C et 30 °C sont enregistrées de mai à octobre. Les amplitudes thermiques variant entre 5 °C et 10 °C sont enregistrées entre les mois de novembre et mars, puis elles augmentent entre les mois d'avril et octobre pour atteindre 17 °C au mois de juillet. En comparant entre les deux périodes (1913-1937 et 1980-2006), il ressort immédiatement que la température a augmenté de 0,46 °C pour janvier, le mois le plus froid et de 0,3 °C pour juillet, le mois le plus chaud dans une période de près de 7 décennies. Ceci montre une véritable tendance à une augmentation des températures de la région. 7.1.2- PluviométrieLa répartition spatiale de la pluviométrie moyenne annuelle varie selon plusieurs paramètres locaux caractéristiques de chaque région dont l'altitude, l'exposition et l'orientation jouent le rôle principal. Ainsi, pour la région de Batna, la répartition des pluies sur les différents mois varie en général dans les trois grands bassins versants considérés (ANONYME, 2005). Les données pluviométriques de la région de Batna durant les périodes 1913-1938 et 1980-2006 sont mentionnées dans le Tableau VI. Pour la pluie, SELTZER (1946) a pris comme point de départ de l'année le premier septembre et non le premier janvier, et la période de 25 ans s'étend de septembre 1913 à août 1938. Tableau VI- Précipitations moyennes mensuelles de la région de Batna durant les périodes allant de 1913 à 1938 et de 1980 à 2006 (M : moyenne mensuelle des précipitations).
(SELTZER, 1946 ; Station météorologique d'Ain Skhouna). La comparaison des données des précipitations montre qu'elles ne sont pas réparties d'une manière homogène sur les différents mois et sur les deux périodes prises en considération. Pendant la période allant de 1913 à 1938, mars et janvier sont les mois les plus pluvieux, avec respectivement 43 mm et 40 mm, alors que pour la période 1980-2006 on note un maximum de pluies durant les mois de mai et septembre, avec respectivement 38,3 mm et 36,9 mm. Par ailleurs, juillet reste le mois qui reçoit le minimum de précipitations pour les deux périodes. La remarque la plus importante dans cette comparaison c'est la différence dans le total des précipitations des deux périodes. La période 1913-1938 est plus arrosée avec une différence de 15,2 mm. Ce qui dénote d'une baisse de précipitations au cours des dernières décennies. 7.1.3- Humidité relative de l'airL'humidité relative de l'air indique que l'état de l'atmosphère est plus ou mois proche de la condensation ; c'est à la valeur de l'humidité relative que correspond la sensation d'humidité ou de sécheresse de l'air (SELTZER, 1946). Les données caractérisant l'humidité relative de l'air de la région de Batna au cours de la période allant de 1995 à 2006 sont reportées sur le Tableau VII. Tableau VII- Humidité relative de la région de Batna au cours de la période 1995-2006 (HR : Humidité relative de l'air).
(Station météorologique d'Ain Skhouna) Nous signalons une humidité relative haute durant les mois de décembre et de janvier avec respectivement 74,58 % et 74,08 % et une humidité relative basse durant le mois le plus chaud, juillet, avec 34,91 %. Les températures moyennes de la région de Batna, ainsi que le gradient thermique correspondent aux climats secs notamment pour les basses altitudes. Une confirmation de cette caractéristique est fournie par les données de l'humidité relative qui dépasse 60 % pendant 4 mois seulement, alors que les températures accusent une forte amplitude thermique annuelle, saisonnière et journalière pour la période chaude, soulignant de ce fait, la continentalité de la région. 7.1.4- VentsLe vent est caractérisé par sa vitesse et sa direction. Les vents les plus dominants dans la région de Batna sont de direction ouest et sud-ouest avec des vitesses moyennes variant entre 2,9 et 3,8 m/s (Tab. VIII). Tableau VIII- Moyennes mensuelles de la vitesse du vent (m/s) avec leurs directions dominantes pour la période allant de 1974-2006 (W : ouest ; SW : sud-ouest ; S : sud ; N : nord).
Les vents du sud-ouest (Sirocco) sont secs et chauds. Ils provoquent une chute brutale de l'humidité et une augmentation notable de la température, généralement supérieur à 30 °C (SELTZER, 1946). Le maximum de sa fréquence a lieu généralement entre juin et juillet. Les vents du nord-ouest soufflent surtout en hiver apportant des pluies à la région (ANONYME, 2005). 7.2- Synthèse climatique7.2.1- Diagramme ombrothermique de GAUSSENLe diagramme ombrothermique de GAUSSEN permet de déterminer les périodes sèches et humides de n'importe quelle région à partir de l'exploitation des données des précipitations mensuelles et des températures moyennes mensuelles (DAJOZ, 2003). D'après FRONTIER et al. (2004), les diagrammes ombrothermiques de GAUSSEN sont constitués en portant en abscisses les mois et en ordonnées, à la fois, les températures moyennes mensuelles en (°C) et les précipitations mensuelles en (mm). L'échelle adoptée pour les pluies est double de celle adoptée pour les températures dans les unités choisies. Un mois est réputé «sec» si les précipitations sont inférieures à 2 fois la température moyenne, et réputé «humide» dans le cas contraire (FRONTIER et al., 2004). Pour localiser les périodes humides et sèches de la région de Batna, nous avons tracé deux diagrammes ombrothermiques pour les périodes allant de 1913-1938 et 1980-2006 (Fig. 8). - A partir de ces diagrammes nous pouvons faire les observations suivantes : Chapitre II- Cadre d'étude (A)
Température 30 Pluviométrie 60 (mm) 50 40 30 20 10 0 25 20 15 10 5 0 I II III IV V VI VII VIII IX X XI XII Mois Pluviométrie I II III IV V VI VII VIII IX X XI XII (B) Température (°C) 30 25 20 15 10 5 0
Mois Température (°C) Pluviométrie (mm)
Figure 8- Diagrammes ombrothermiques de GAUSSEN de la
région de Batna pendant Le diagramme ombrothermique de la région de Batna pour la période allant de 1913 à 1938 fait apparaître deux périodes bien distinctes, l'une sèche et chaude s'étalant sur 5 mois, depuis juin jusqu'à octobre, et l'autre humide et froide allant de novembre jusqu'à mai. Concernant la période récente, allant de 1980 à 2006, nous observons également les mêmes périodes, l'une sèche et chaude et l'autre humide et froide. La période sèche et chaude s'étale sur 5 mois, allant de juin jusqu'à octobre, alors que la période humide et froide s'étale sur les mois allant de novembre jusqu'à mai. 7.2.2- Climagramme d'EMBERGERLe système d'EMBERGER permet la classification des différents climats méditerranéens (DAJOZ, 1985-2003). Cette classification fait intervenir deux facteurs essentiels, d'une part la sécheresse représentée par le quotient pluviothermique Q en ordonnées et d'autre part la moyenne des températures minimales du mois le plus froid en abscisses. Il est défini par la formule simplifiée suivante (STEWARTE, 1969) :
P : pluviométrie annuelle en mm. M : température moyenne maximale du mois le plus chaud en °C. m : température moyenne minimale du mois le plus froid en °C. Le quotient pluviothermique est d'autant plus élevé que le climat est plus humide (DAJOZ, 1985). FAURIE et al. (1998-2003) avancent que cet indice n'est vraiment établi que pour la région méditerranéenne et qu'en fonction de la valeur de ce coefficient on distingue les zones suivantes : - humides pour Q > 100 ; - tempérées pour 100 > Q > 50 ; - semi-arides pour 50 > Q > 25 ; - arides pour 25 > Q > 10 ; - désertiques pour Q < 10. Afin de déterminer l'étage bioclimatique de la région de Batna et la situer dans le climagramme d'EMBERGER, nous avons calculé le quotient pluviothermique Q avec des données climatiques calculées sur deux périodes, de 25 ans Q1 (1913-1938) et de 27 ans Q2 (1980-2006). 346 Q1 = 3,43 = 35,62 3 3,44 0,1 3 - 3 3 0,8 Q2 = 3,43 = 34,38 3 3,3 0,3 - Malgré la différence de 1,24 entre Q1 et Q2, la région de Batna est toujours classée dans l'étage bioclimatique semi-aride à hiver frais (moyenne des températures minimales du mois le plus froid janvier, est de 0,3 °C pour la période 1913-1937 et 0,13 °C pour la période 1980- 2006). En conclusion, nous pouvons dire que, le climat de la région de Batna est de type semi-aride à hiver frais pour toutes les cuvettes et reliefs, à l'exception de la zone des hautes plaines de l'Est qui sont plus ou moins exposées au nord, alors qu'il est sub-humide à hiver frais, grâce à l'avancée de l'imposant massif des Aurès (ANONYME, 2005). Selon HUNTLEY et al. (2006), les changements climatiques potentiels sont donc une cause pour une inquiétude considérable quant à leur impact sur la biodiversité. Les oiseaux, avec les autres organismes, sont supposés répondre aux changements climatiques de l'une des deux manières : ils peuvent faire des adaptations aux nouvelles conditions de vie, ou ils ont une réponse spatiale en ajustant leur distribution géographique. L'espèce incapable d'accomplir une réponse suffisante par l'un ou l'autre mécanisme ou par les deux sera à risque d'extinction (HUNTLEY et al., 2006). Ces auteurs rajoutent que sans mise en oeuvre de nouvelles mesures de conservation, ces impacts seront sévères et vraisemblablement aggravés par les changements de la répartition des terres associée à la fragmentation de l'habitat. Partie II : Présentation des zones échantillons1- Choix des zones échantillonsAvant de commencer nos sorties de terrain, nous avons réalisé des sorties de prospection afin de choisir les zones échantillons sur lesquelles nous travaillerons. Nous nous sommes également basé sur une étude réalisée sur la bioécologie de la Cigogne blanche dans la région de Batna par DJADDOU et BADA (2006). Ces enquêtes nous ont relevé principalement trois zones dans lesquelles ces derniers auteurs ont dénombré les plus grands effectifs de cigognes blanches et de hérons garde-boeufs. Ces zones sont AinTouta, El Madher et Merouana, qui sont particulièrement caractérisées par des plaines et des vallées très peu accidentées. Ain-Touta, abrite une grande colonie mixte de cigognes blanches et de hérons gardeboeufs construite sur de hauts arbres de Pin d'Alep, située au centre ville. La hauteur des pins qui atteignent jusqu'à 25 m et leur situation au centre ville (dérangements et piétinement des pelotes de réjection des oiseaux), nous ont dissuadé à travailler dans cette zone. Nous avons alors opté pour les zones d'El Madher et de Merouana, qui sont peu éloignée de l'Université. Nous avons pris en considération que les aires d'étude sont beaucoup plus grandes que les données qui concernent seulement les communes d'El Madher et de Merouana. Dans la zone échantillon d'El Madher l'aire d'étude peut s'élargir sur les communes d'El Madher, Boumia, Djarma et Fesdis et dans la zone échantillon de Merouana les communes de Merouana, Oued El Ma, Ksar Bellezma et El Hassi (Fig. 6). 2- Description générale de la zone échantillon d'El MadherLa daïra d'El Madher est située dans la willaya de Batna à une distance de 22 km à l'est du chef lieu de la willaya. La zone échantillon d'El Madher couvre la majorité du territoire de la daïra d'El Madher, notamment les communes d'El Madher, de Boumia et de Djerma et la commune de Fesdis (daïra de Batna). L'ensemble de ces communes couvre une superficie de 438, 69 km2 dont 19.980 ha de S.A.U. (Fig. 6). Se situant à 900 m d'altitude, la zone d'El Madher est caractérisée par des terres fertiles situées au pied du Djebel Bouarif point culmunant Ras fourrare 1.741 m. Cette zone est dominée par un relief simple et relativement plat. La pente représente environ 8 %, c'est pourquoi elle est souvent dite « la plaine d'El Madher » (ANONYME, 2005). C'est au sein de cette plaine que se sont déroulées nos études sur les disponibilités alimentaires, le recensement des populations et la fréquentation des milieux de gagnage. Le suivi de la biologie de la reproduction et la collecte des pelotes de réjection destinées à l'étude du régime alimentaire sont effectués comme suit : > Cas du héron garde-boeufs En dehors de la période de reproduction du héron garde-boeufs, nous avons travaillé sur deux dortoirs. Le « dortoir d'El Madher », bâti sur 4 arbres de cyprès (Cupressus sempervirens), est situé à la limite nord du village d'El Madher. Le « dortoir de Djerma », bâti sur 2 arbres de cyprès et un arbre de Pin d'Alep, est situé sur la route nationale n° 3, à 7 km du chef lieu de la commune de Djerma en allant vers Constantine. Pendant la période de reproduction du garde-boeufs, nous avons travaillé dans une colonie de nidification située au niveau de la psychiatrie d'El Madher, qui est construite sur 3 arbres de pin d'Alep. > Cas de la Cigogne blanche Concernant la Cigogne blanche, nous avons travaillé dans une colonie construite dans une ferme distante de 7 km à l'ouest du village d'El Madher. Dans la « clonie d'El Madher », la Cigogne blanche a construit 12 nids sur une maison de premier étage à toiture en tuiles, 2 nids sur deux cyprès et 5 nids sur des poteaux électriques en béton de 8 m de hauteur. 3- Description générale de la zone échantillon de MerouanaLa daïra de Merouana est située à 48 km au nord-ouest de la ville de Batna et à 60 km en passant à l'est par la commune de Seriana. La zone échantillon de Merouana est représentée par l'ensemble des communes de Merouana, d'Oued El Ma et de Ksar Belezma appartenant à la daïra de Merouana et la commune d'El Hassi (daïra d'Ain Djasser). Cette zone échantillon couvre une superficie de 418,15 km2 dont 23.603 ha de S.A.U. (Fig. 6). La zone de Merouana est entourée de montagnes, les plus imposantes sont le Dj. Tikelt au sud, le Dj. Refâa à l'ouest et le Dj. Fakhra à l'est. L'altitude moyenne en plaine est de 850 m, elle peut atteindre 1.400 m en zone montagneuse, alors que la ville de Merouana est située à une altitude moyenne de 990 m (ANONYME, 2005). Dans cette zone, nous avons suivi et dénombré les effectifs des modèles biologiques étudiés dans leurs milieux de gagnage dans la plaine de Belezma, dans une aire passant par les communes de Merouana, d'Oued El Ma, d'El Hassi et de Ksar Belezma. Pour le suivi des cycles biologiques des deux échassiers étudiés et la collecte des pelotes de rejection, nous avons travaillé au centre ville sur une colonie mixte de cigognes blanches et de hérons garde-boeufs, représentant en même temps un dortoir et une colonie de nidification pour le Héron garde-boeufs. Les oiseaux étudiés ont construit leurs nids, sur 3 arbres de Pin d'Alep plantés dans la cours de la mairie de Merouana et dans la cours d'une entreprise privée (SARL Réalisation Services Merouana "RSM") qui sont limitrophes. Chapitre III : Matériel et méthodes d'étude1- Méthode d'étude de la phénologie des deux échassiers et du statut actuel du héron garde-boeufs1.1- Méthode d'étude du cycle biologique des deux espècesDurant nos sorties de terrain, allant de novembre 2006 jusqu' à novembre 2007, nous avons opté à enregistrer les différentes dates concernant les stades phénologiques des deux échassiers dans les deux zones d'étude. Ainsi, nous avons noté pour la Cigogne blanche les dates d'arrivées et de départs, la date de ponte, la période de couvaison et d'élevage des jeunes et la date d'envol des cigogneaux. De même pour le Héron garde-boeufs, nous avons noté les dates de première observation du plumage nuptial, de première observation d'accouplements et de pontes, les périodes de couvaison et d'élevage des jeunes et la date de première observation d'envols de héronneaux. 1.2- Méthode d'étude du statut actuel des populations du Héron garde-boeufs1.2.1- Objectif de cette étudeL'objectif principal de cette démarche est de mettre la lumière sur la nouvelle aire de répartition du héron garde-boeufs en période d'hivernage et de reproduction jusqu'en 2007 sur tout le territoire de la wilaya de Batna avec la mise en évidence des principaux éléments caractérisants la biologie et l'écologie de l'espèce. Cette étude, contribuerait à la connaissance de la place écologique qu'occupe cette espèce et le rôle qu'elle peut jouer dans le fonctionnement des écosystèmes qu'elle a nouvellement colonisés. 1.2.2- Enquête localePendant toute la période d'étude et particulièrement durant une dizaine de jours au mois de juillet 2007, nous avons interrogé une cinquantaine de personnes de différents âges et niveaux intellectuels (paysans, adolescents, universitaires), dans les centres villes et environs (Douars) des communes suivantes : Tazoult, Oyoun El Assafir, El Madher, Djerma, Seriana, Zana El Beida, Ain Djasser, Merouana, Ksar Belezma, Ain Touta, Tilatou, Sefian, Boumagueur, N'Gaous, Barika, Bitam et Seggana. La démarche suivie est celle des entretiens selon la méthode semi-directive, en se basant sur un questionnaire typique de 15 questions (Fig. 9). Chapitre III- Matériel et méthodes Enquête locale sur le Héron garde-boeufs Date :
Figure 9- Prototype du questionnaire utilisé dans l'enquête locale. Selon TREPPOZ et VAIS (1988) in OUELMOUHOUB (2005), ce type de méthode permet à la personne interrogée d'aborder des thèmes et sous-thèmes que l'enquêteur lui propose ouvertement ou bien qu'il aborde lui-même. Dans ce cas, les questions posées s'enchaînent en fonction des informations apportées par l'interviewé. Cette enquête est réalisée auprès des habitants, en particulier, des agriculteurs et des paysans vivants en contact direct avec l'espèce. Elle a porté sur le recueil d'informations concernant d'une part, le statut phénologique de l'espèce (historique, périodes d'arrivée et de départ, année de première nidification), la localisation exacte des colonies et des dortoirs, les types des milieux d'alimentation, le type des proies habituellement consommées et d'autre part, sur le type de relation existant entre l'espèce et l'homme (Fig. 9). 2- Méthode d'étude des disponibilités alimentairesLa disponibilité des proies est définie par l'abondance des types de proies potentielles vivants dans les micro-habitats fréquentés par l'oiseau prédateur lors de la recherche de nourriture (WOLDA, 1990). Afin de donner une image fidèle de la qualité systématique des proies potentielles que renferment les différents milieux trophiques fréquentés par les modèles biologiques étudiés et dans le but de donner un aperçu quantitatif de l'évolution temporelle des peuplements de proies disponibles, nous avons mis en place un dispositif de piégeage et réalisé des captures à l'aide du filet fauchoir. L'échantillonnage des disponibilités alimentaires offertes aux cigognes et aux gardeboeufs de la région de Batna est réalisé dans 3 types de milieux localisés dans la plaine d'El Madher, choisis à cause de leur facilité d'accès et de leur sécurité. Ces stations échantillons sont représentées par un milieu cultivé (cultures maraîchères, grandes cultures au stade levé, les chaumes de céréales et même parfois dans un labour), un milieu humide (marais temporaire d'eau douce) et une pelouse naturelle. Ces milieux représentent généralement les principaux milieux de gagnages exploités par les deux échassiers étudiés (BREDI N, 1983- 1984 ; BOUKHEMZA, 2000 ; SI BACHIR, 2007). Chaque mois, durant la période allant de décembre 2006 à novembre 2007, nous avons appliqué un dispositif de collecte constitué d'une parcelle carrée de 400 m2 (20 m x 20 m de côté) dans laquelle sont implantés 9 pots Barber au ras du sol distants de 5 m sur trois rangées, et 4 assiettes de couleur jaune au centre de chaque carré formé par les pots Barber (Fig. 10). Les pièges trappes sont en fait des boîtes de conserve, cylindriques, de 1 décimètre cube de contenance, alors que les assiettes colorées utilisées sont des récipients peu profonds, en matière plastique de 15 cm de diamètre. Tous les pièges utilisés sont remplis à 2/3 d'eau additionnée d'un mouillant et sont retirés du sol avec leur contenu au bout de 3 jours à une
20 m 5 m 5 m Pot Barber 20 m Piège coloré Figure 10- Dispositif appliqué dans l'échantillonnage des disponibilités alimentaires. semaine après leur installation (BENKHELIL, 1991). Les pièges trappes ont permis d'échantillonner principalement des invertébrés se déplaçant sur le sol. Nous citons en particulier parmi les innombrables groupes recensés, des coléoptères, des arachnides, des crustacés et des mollusques. Un grand nombre d'insectes volants qui viennent se poser à la surface ou y tombent sont également colletés. Les pièges colorés ont servi à la capture d'une grande variété d'insectes ailés, notamment des hyménoptères, des diptères et des lépidoptères. Parallèlement à ce dispositif, nous avons réalisé un fauchage à l'aide d'un filet fauchoir de 30 cm de diamètre le long d'un transect de vingt mètres, totalisant en général 15 à 20 coups dans les mêmes carrés réservés à l'échantillonnage par les pièges trappe. Cette méthode est appliquée sur une végétation basse et non humide. Les principaux groupes récoltés sont les orthoptères, les lépidoptères, les diptères et les hyménoptères. 3- Méthode d'étude de l'exploitation des milieux de gagnageDans le but de mettre en évidence les principaux milieux d'alimentation fréquentés par la Cigogne blanche et le Héron garde-boeufs et les modalités d'exploitation de ces gagnages au cours des diverses saisons phénologiques que connaissent les deux espèces, nous avons réalisé des recensements réguliers sur deux itinéraires échantillons. 3.1- Description et choix des itinéraires Les itinéraires choisis sont :> Itinéraire de la zone d'El Madher : regroupe les tronçons de la route nationale 3 depuis la ville de Batna jusqu'à Djerma, puis les tronçons de la route départementale 26 de Djerma à El Madher en revenant sur la ville de Batna tout en passant par Fesdis (Fig. 11). > Itinéraire de la zone de Merouana : parcourt les tronçons de la route nationale 86 allant de la ville de Merouana à Oued El Ma et les tronçons de la route départementale 153 passant par El Hassi et Ksar Bellazma, tout en revenant à la ville de Merouana par la cité de Chiddi, puis le tronçon de la route nationale 77 allant de Merouana à Ksar Belezma (Fig. 11). La longueur totale de ces deux itinéraires est respectivement de 35 km et 50 km, parcourus à l'aide d'un véhicule roulant à vitesse modérée, en moyenne en 2 heures de temps chacun. Ces transects, comprennent une partie de la plaine d'El Madher et celle de Belezma qui représentent les principales régions où la cigogne et le garde-boeufs sont le plus fréquemment rencontrés dans la région de Batna.
Ksar Belezma N77 N86 Oued El Ma W153 El Hassi N86 N3 N77 N3 Fesdis N31 N86 N3 W26 Djerma N88 Figure 11- Cheminement des
itinéraires-échantillons parcourus lors des recensements (En jaune : réseau routier ; en rouge : Itinéraires-échantillons ; route nationale ; N3 W26 route départementale, sens du déplacement). Les tronçons prospectés sont représentés par des zones à activités agricoles très diversifiées mettant à profit les eaux des oueds et les eaux sous terraines. Ces régions sont caractérisées par de vastes prairies, des cultures basses et par l'élevage de bovins, d'ovins et d'aviculture. Nous avons parcouru ces itinéraires une fois par mois au cours de la période allant de décembre 2006 jusqu'à fin septembre 2007 totalisant 10 recensements par chacun des deux itinéraires. 3.2- Méthode de recensement des groupesLes dénombrements sont réalisés le matin, vers 10 heure de matin, au cours de journées sans pluie battante ou interrompue ni vents violents (BREDIN, 1983). Lors de nos sorties nous avons évité les matinées où les hérons se rassemblent après l'envol des dortoirs et le soir où ils se réunissent dans les dortoirs. A chaque contact avec un groupe de cigognes ou de garde-boeufs, nous avons noté, systématiquement :
> Prairies : ce sont des terrains caractérisés par une couverture permanente de végétation herbacée, généralement pâturés. Deux types de prairies sont observés, naturelles ou cultivées. > Friches et parcours : ce sont des milieux relativement secs, les friches sont caractérisées par une végétation arbustive clairsemée, alors que les parcours résultent des activités agricoles notamment après les moissons. > Bordures de ruisseaux : les ruisseaux naturels de la région d'étude sont à écoulement momentané pendant la saison des pluies et sont généralement soumises aux déversements des eaux usées. > Marais temporaires (pelouses inondées) : comprennent de petites étendues d'eau peu profondes envahies par la végétation. > Labours > Cultures basses : elles sont représentées essentiellement par des cultures céréalières non irriguées, fréquentées seulement au cours de leurs premiers stades phénologiques, ainsi que par diverses cultures maraîchères exploitées en irrigué. > Immondices : représentées particulièrement par les dépôts d'ordures ménagères. > Dépôts d'animaux domestiques : résultant de l'aviculture ou de l'élevage de bovins et d'ovins. 3- Les types d'associations observés, regroupés en 4 types :
Ces trois dernières associations correspondent principalement au comportement d'association décrit chez le Héron garde-boeufs (BREDIN, 1984 ; SI BACHIR, 2007). 4- La présence ou l'absence d'eau. 4- Méthode d'étude du régime alimentaire4.1- Choix de la méthodeLes études réalisées sur le régime alimentaire des oiseaux sont basées sur différentes méthodes : l'analyse des contenus stomacaux, l'analyse des pelotes de rejections chez les adultes ou des régurgitations chez les poussins et les méthodes d'observation directe (BREDIN, 1983). Dans notre étude, nous avons choisi la méthode de l'analyse des pelotes de rejection des adultes, pour la plus ou moins simplicité de son emploi et pour le fait que les pelotes régurgitées par la Cigogne blanche et le Héron garde-boeufs contiennent les parties non digestibles des principales proies consommées. Ce sont en particulier, les proies arthropodologiques à partir des fragments chitineux, et mammalogiques grâce aux restes de leurs poils. Néanmoins, cette méthode reste incomplète du fait qu'une certaine proportion des proies échappe totalement à la détection suite à leur digestion complète, comme c'est le cas des vers de terre. 4.2- Collecte et conservation des pelotesPendant la période de séjour de la Cigogne blanche, nous avons collecté un total de 136 pelotes de cigognes, dont 92 pelotes de la colonie d'El Madher et 44 pelotes de la colonie de Merouana. La collecte des pelotes est faite avec une fréquence régulière d'une à trois fois par mois, et des lots variant entre 2 et 18 pelotes. Le nombre important des pelotes de la cigogne collectées dans la zone d'El Madher revient au fait que l'accès aux nids s'est fait grâce à un camion-échelle. Pour le Héron garde-boeufs, durant une période étalée sur 11 mois, allant de novembre 2006 à septembre 2007, nous avons collecté mensuellement et avec des lots variant entre 25 et 60 pelotes, un total de 957 pelotes, 481 pelotes de la zone d'El Madher et 476 pelotes de la zone de Merouana. En fait, la collecte des pelotes du garde-boeufs dans la zone d'El Madher a lieu au niveau des deux dortoirs « dortoir d'El Madher » et « dortoir de Djerma » en dehors de la période de reproduction et à « la colonie de la psychiatrie » d'El Madher en période de reproduction. Les pelotes récoltées sont conservées dans des cornets en papier et mises dans des sachets portant une étiquette sur laquelle sont mentionnés la date, la région et le nom de l'espèce. 4.3- Mensurations et pesées des pelotesUne fois au laboratoire, la première étape du traitement des pelotes de régurgitation est la mensuration de leurs tailles. Chaque pelote collectée est soumise à des mesures de longueur et de largeur à l'aide d'un papier millimétré (Fig. 12a et b). Le poids humide est pris à l'aide d'une balance électronique de 0,0001 g de précision (Fig. 12c). Elles sont ensuite déshydratées à 120 °C pendant 24 h dans une étuve pour éviter l'effet des moisissures qui gênerait leur analyse et tout risque d'infection par des microorganismes pathogènes (DOUMANDJI et SETBEL, 2001). Les pelotes desséchées sont ensuite repesées à sec aussitôt sorties de l'étuve. Après les mensurations et les pesées, les pelotes sont numérotées, datées puis conservées dans des cornets en papier en vue de leur analyse ultérieure (Fig. 12d). 4.4- Macération des pelotes par voie humideChaque pelote est macérée séparément dans une boite de Pétri contenant de l'eau additionnée de quelques gouttes d'alcool pour aseptiser le milieu et éviter les mauvaises odeurs (Fig. 12e et f). A l'aide de pinces entomologiques et sous une loupe binoculaire, sont séparés les différents fragments que contient la pelote triturée (têtes, thorax, élytres, pattes, fémurs, mandibules, écailles, agglomérat de poils, ...) et mis dans une autre boite de Pétri sur laquelle sont mentionnées les indications de l'espèce, le numéro de la pelote, la date et la région de collecte (Fig. 13).
Figure 12- Différentes étapes suivies
dans le traitement au laboratoire des
pelotes
Figure 13- Exemple des fragments de proies
détectés dans les pelotes de rejection des Sur l'ensemble des lots récoltés (136 pour la cigogne et 957 pour le garde-boeufs), seules les pelotes fraîches et non fragmentées (96 pour la cigogne et 110 pour le garde-boeufs) ont fait l'objet de la trituration. 4.5- Détermination et comptage des itemsLe suc gastrique très fort des modèles biologiques étudiés ne laisse que peu de traces des restes des proies vertébrés tels les poissons, les reptiles et les mammifères (BREDIN, 1983 ; BANG et DAHLSTROM, 1987-2006). Aussi, les proies à téguments mous comme les annélides, les mollusques sans coquilles et certaines larves d'insectes sont peu décelables. La détermination des fragments retrouvés dans les pelotes reste très délicate du fait que les parties observées soient souvent incomplètes et/ou dénaturées. L'identification des premiers fragments a été rendu possible grâce à leur comparaison avec la collection des fragments des insectes confectionné au laboratoire d'écologie et environnement de l'université de Béjaia avec la précieuse collaboration de Mme GHARBISALMI R. et du Dr MOULAI R., et grâce à leur comparaison avec les individus entiers collectés par la méthode d'échantillonnage des disponibilités alimentaires. La détermination de la majorité des items a été faite par le Dr SI BACHIR du département de Biologie de l'université de Batna, qui a poussé la détermination systématique des items jusqu'à l'ordre, la famille et parfois même jusqu'au genre et l'espèce. L'identification des proies invertébrées, notamment celle des insectes, la classe la plus dominante tant en espèces qu'en nombre, repose sur la reconnaissance simultanée de plusieurs fragments chitineux comme les têtes, les thorax, les élytres, les cerques, les fémurs, les tibias et les mandibules. Le nombre d'individus de chaque espèce est déterminé principalement par le nombre de têtes ou de thorax. Aussi, lors du comptage des élytres, des mandibules et des cerques, le nombre d'appendices gauches et droits est pris en considération. Pour les proies non entomologiques, les coquilles restent le seul critère d'identification des mollusques, chaque fois que cette présence est révélée dans une pelote nous comptons un seul individu. La présence de plusieurs apex de coquilles permet de compter le nombre d'individus où chaque apex représente un individu (BOUKHEMZA, 2000). Les arachnides se différencient des autres classes par la présence d'un céphalothorax muni de chélicères, de pédipalpes et de pinces (VACHON, 1952). Les myriapodes sont reconnaissables grâce au nombre important des pattes portées par l'abdomen et des mandibules fines et pointues. Pour les batraciens, les reptiles et les mammifères, la présence respectivement d'un tégument, d'une écaille ou d'un agglomérat de poils correspond à un individu. L'identification des petits mammifères s'appuie sur l'analyse micrographique des poils qui se base sur la spécificité des empreintes des poils (DAY, 1966 ; KELLER, 1978-1980 ; FALLIU et al., 1979 ; DEBROT et al., 1982 ; HAMDINE, 1990). Pour identifier l'espèce proie à laquelle appartiennent les poils retrouvés dans les pelotes de régurgitation, nous avons procédé à des montages de poils. Afin d'effectuer de bonnes observations, nous avons suivi les trois étapes suivantes (ANNA et al., 1993 in ZENNOUCHE, 2002) : - Macération du poil à l'eau chaude pendant 5 à 10 mn puis séchage à l'aide de papier filtre pour éliminer les impuretés et la matière grasse et afin de séparer les poils les uns des autres. - Immersion du poil dans l'alcool à 70 % pendant 60 mn afin de dissoudre les impuretés et éliminer les corps gras résiduels. - Séchage du poil avec de l'éther afin de faire disparaître toute trace de graisse. Après le rinçage, nous déposons les poils sur du vernis à ongles transparent étendu en fine couche sur une lame de verre. Une fois le vernis est séché, on arrache les poils et on observe l'empreinte laissée sous le microscope optique au grossissement (X10 et X40). La détermination est réalisée par le Dr SI BACHIR avec consultation d'un Atlas des poils de mammifères (DEBROT et al., 1982). La consommation des poissons se traduit par la présence d'écailles dans les pelotes, alors que les plumes révèlent la consommation d'oiseaux. Un seul individu est pris en considération quand des écailles ou des plumes sont présentes dans une pelote de réjection. 5- Méthodes de traitement des donnéesIl est d'une importance capitale de préciser le sens à accorder à certains paramètres que nous prendrons en considération lors de l'exploitation de nos données. Ces paramètres généralement utilisés par d'autres auteurs, permettront de comparer nos résultats à ceux obtenus dans d'autres régions du monde et à mieux comprendre les variations éventuelles de certaines mesures dans l'espace et dans le temps.
5.1- Traitement des données par l'application
d'indices écologiques
5.1.1- Structure de la composition du régime alimentaire> Fréquence en nombre La fréquence centésimale en nombre représente l'abondance relative et correspond au pourcentage du nombre d'individus d'une espèce (ni) par rapport au total des individus recensés (N). Elle peut être calculée pour un prélèvement ou pour l'ensemble des prélèvements d'une biocénose (DAJOZ, 1985).
La fréquence d'occurrence (C), appelée aussi fréquence d'apparition ou indice de constance est le pourcentage du rapport du nombre de relevés contenant l'espèce i (ri) au total de relevés réalisés (R) (DAJOZ, 1985). La constance est calculée selon la formule suivante : C r i × 100 = i R Ce paramètre nous a servi pour calculer la fréquence d'apparition des divers types de proies disponibles dans les gagnages et dans la composition de l'alimentation des deux échassiers. Celle-ci correspond au pourcentage du nombre de pelotes ou de capture contenant le taxa i (ri) par rapport au nombre total de pelotes ou de captures (R). BIGOT et BODOT (1973), distinguent des groupes d'espèces en fonction de leur fréquence d'occurrence : - les espèces constantes sont présentes dans 50 % ou plus des relevés effectués ; - les espèces accessoires sont présentes dans 25 à 49 % des prélèvements ; - les espèces accidentelles sont celles dont la fréquence est inférieure à 25 % et supérieure ou égale 10 % ; - les espèces très accidentelles qualifiées de sporadiques ont une fréquence inférieure à 10 %. > Analyse de similitude ou indice de SORENSON Afin de pouvoir statuer sur la similitude ou la différence existante dans la composition du peuplement des invertébrés dans l'espace d'une part et dans le temps d'autre part, nous avons calculé l'indice de SORENSON ou le coefficient de similitude de SORENSON (Qs) (MAGURRAN, 1988) : a b + Qs = 2 C× 100 a : Nombre d'espèces proies mentionnées dans le premier échantillon ; b : Nombre d'espèces proies décrites dans le deuxième échantillon ; C : Nombre d'espèces proies recensées simultanément dans les deux échantillons. 5.1.2- Diversité et équirépartition des peuplements de proies consommées > Richesse spécifique totale (S) C'est le nombre total d'espèces qui composent un peuplement considéré dans un écosystème donné (RAMADE, 1984). Dans notre cas, la richesse totale représente le nombre total d'espèces recensées lors de l'analyse d'un échantillon de pelotes ou de proies capturées dans les milieux de gagnage. > Richesse moyenne (Sm) La richesse spécifique moyenne (Sm) est utile dans l'étude de la structure des peuplements. Elle est calculée par le nombre moyen d'espèces présentes dans un échantillon (RAMADE, 1984). nombre total d espèces Sm ' recensées lors de chaque relevé nombre de relevés réalisés > Indice de diversité de SHANNON L'indice de SHANNON (H') est capable de traduire la diversité des peuplements (BLONDEL, 1979 ; MAGURRAN, 1988). Il est calculé par la formule suivante : H' = -? Pi log2 Pi Où : Pi représente le nombre d'individus de l'espèce i par rapport au nombre total d'individus recensés (N) : p i
n i = N > Indice d'équirépartition ou équitabilité L'équitabilité correspond au rapport de la diversité observée (H') à la diversité maximale (H'max), (RAMADE, 1984 ; MAGURRAN, 1988).
H'max Avec H' : est l'indice de diversité de Shannon H'max = log2 S (S : la richesse totale). 5.1.3- Electivité des proies ou indice d'IVLEVPlusieurs indices ont été proposés pour mesurer le choix alimentaire des espèces animales prédatrices en faisant une comparaison, à l'aide d'indices différents, entre l'utilisation et la disponibilité des proies. Dans la présente étude, nous avons utilisé l'indice de sélection ou d'électivité d'IVLEV. Symbolisé par (E), cet indice permet de comparer l'abondance relative des proies disponibles dans le milieu et le choix des proies consommées par le prédateur. Il se calcule par la formule suivante :
Où : r représente la fréquence d'un item dans le spectre alimentaire d'une espèce et p la fréquence du même item dans le milieu environnant. (E) varie de -1 à 0 pour une sélection négative et de 0 à +1 pour une sélection positive (JACOBS, 1974). 5.2- Traitement des données par les méthodes d'analyses statistiques5.2.1- Analyses statistiques univariéesNous avons calculé pour chaque variable les paramètres de base qui sont les statistiques descriptives : la moyenne (x'), l'écart-type (s) et les valeurs maximales et minimales (DAGNELIE, 2000). La comparaison de la variation du régime alimentaire des deux espèces dans les deux zones d'étude a été réalisée à l'aide de l'analyse de la variance à un critère de classification (AV1). - Si p > 0,05 il n'existe pas de différences significatives ; - Si P = 0,05 il y a des différences significatives ; - Si P = 0,01 il y a des différences hautement significatives ; - Si P = 0,001 il y a des différences très hautement significatives. 5.2.2- Analyses statistiques bivariées (Tests de corrélation)Pour mettre en évidence la relation entre la variation de la composition du régime alimentaire des deux espèces dans les deux zones d'étude, nous avons utilisé le coefficient de corrélation linéaire de BRAVAIS-PEARSON (DAGNELIE, 2000). Ce coefficient de corrélation (r), mesure l'intensité du lien qui existe entre deux caractéristiques ou variables quantitatives quelconques, pour autant que cette liaison soit linéaire ou approximativement linéaire. Ce coefficient est compris entre -1 et +l. Il est en valeur absolue, d'autant plus proche de 1 que la liaison entre les deux séries d'observations est nette, pour autant que cette liaison soit linéaire ou approximativement linéaire. Au contraire, si le coefficient est nul ou approximativement nul c'est que les deux variables ne sont pas corrélées entre elles. D'autre part, le signe du coefficient de corrélation indique si la relation entre les deux variables (séries d'observations) est croissante ou décroissante. En effet, lorsque le coefficient de corrélation est positif, les valeurs élevées d'une variable correspondent, dans l'ensemble, aux valeurs élevées de l'autre variable, et les valeurs faibles d'une variable correspondent aux valeurs faibles de l'autre variable. Par contre, lorsque la corrélation est négative, les valeurs élevées d'une variable correspondent, dans l'ensemble, aux valeurs faibles de l'autre variable et vice-versa. - Si p > 0,05 il n'existe pas de corrélations ; - Si P = 0,05 il existe une corrélation significatives ; - Si P = 0,01 il existe une corrélation hautement significatives ; - Si P = 0,001 il existe une corrélation très hautement significatives. Tous les calculs uni et bi variés sont réalisés à l'aide du logiciel d'analyse et de traitement statistique des données, MINITAB version 13.31 pour Windows (Anonyme, 2000). Chapitre IV : Résultats
1- Cycle biologique et statut des populations des deux échassiers dans la région de BatnaNos sorties de terrain étalées de novembre 2006 à novembre 2007, nous ont permis de noter les différents stades phénologiques des deux modèles biologiques étudiés et de statuer sur la chronologie leurs cycles biologiques. Les sorties réalisées dans le but de faire une enquête locale sur le Héron garde-boeufs dans la région de Batna nous ont permis également d'avoir un aperçu sur l'historique de l'implantation des populations de cette espèce et de leurs états actuels dans la région de Batna. 1.1- Cycle biologique de la Cigogne blancheLes premières arrivées de la Cigogne blanche sur les sites de reproduction sont enregistrées à la fin du mois de janvier. La première cigogne arrivée est observée en date du 28 janvier 2007, simultanément, dans les colonies d'El Madher, de Merouana et d'Ain Touta. Cependant, Batna et Arris ont vu leurs premières cigognes arriver au début du mois de février, respectivement les 2 et 8 février 2007. A partir de la deuxième décade du mois de février, les couples nicheurs commencent à se former sur les sites de reproduction. La date de première observation d'accouplements dans la colonie d'El Madher coïncide avec le 12 février 2007. Généralement, un seul individu arrive et occupe le nid en premier, puis sera suivi par son partenaire une semaine plus tard. L'accès aux nids grâce à un camion-échelle utilisé dans la colonie d'El Madher nous a permis de décrire les échéances des différentes étapes caractérisant le cycle biologique de la Cigogne blanche (Fig. 14). Ainsi, le 24 mars 2007 correspond à la date de première observation d'oeufs et le 23 avril 2007 à la date d'observation des premières éclosions. L'envol des jeunes cigognes blanches est observé le 25 juin 2007 et leur départ vers les aires d'hivernage au 27 juillet 2007. Par ailleurs, nous signalons la présence curieuse d'une cigogne blanche observée en date de 23 août 2007 à moins de 500 m au nord de la colonie d'El Madher. 1.2- Cycle biologique du Héron garde-boeufsA Merouana, nous avons observé les premiers hérons garde-boeufs en plumage nuptial à la troisième décade du mois de janvier et la formation des couples à la troisième décade du mois de mars. La ponte commence au début du mois d'avril, les premières éclosions à la fin du même mois et les premiers envols à la mi-juin. La période de nidification de cet échassier prend fin en cette zone à la première décade d'août (Fig. 15A).
Arrivée des cigognes 28/01/2007 Première observation Première observation Première
observation Départ des cigognes 27/07/2007 Première observation d'envols Pré-reproduction Période d'accouplements Ponte et couvaison Nourrissage et élevage des jeunes Post reproduction Hivernage Figure 14- Cycle biologique de la Cigogne blanche dans
la colonie d'El Madher en l'an (A) Merouana
Apparition du Observation des premières pontes Observation des premiers envols (B) El Madher Observation des premiers couples formés Désertion totale de la colonie Observation des premières éclosions
Apparition du Observation des premières pontes Observation des premiers envols
Pré- reproduction Période d'accouplements Ponte et couvaison Nourrissage et élevage des jeunes Hivernage Figure 15- Cycle biologique du Héron
garde-boeufs dans les deux colonies d'étude : (A) Dans la zone d'El Madher, B. ibis revête son plumage nuptial à la troisième décade du mois de février et commence à s'accoupler à la troisième décade d'avril. Les premières pontes son notées au début mai et les premières éclosions à la fin de ce même mois. Les envols commencent à la deuxième décade de juillet. Le cycle de l'espèce s'achève à la fin août suite à la désertion totale de la colonie (Fig. 15B). 1.3- Historique et état actuel des populations du Héron garde-boeufs dans la région de BatnaLa synthèse des résultats de l'enquête réalisée dans plusieurs localités de la région de Batna est établie comme suit : > Année de première observation et dénomination de l'espèce 37 % de réponses obtenues signalent l'espèce depuis l'an 2005, 32,6 % depuis l'an 2004, 15,2 % depuis l'an 2002, 13 % depuis l'an 2003 et 2,2 % de réponses depuis l'an 1997. Pour son appellation, 85,7 % des réponses rapportent qu'il n'a pas d'appellation locale bien connue (en arabe ou en chouia), 6,1 % de personnes signalent l'appellation de « Djaj El Ma » ou « poule d'eau », 4,1 % de personnes donnent l'appellation de « Farkh Labhar » ou « oiseau de la mer » et 4,1 % donnent l'appellation d'« Aigrettes ». > Statut phénologique Concernant le statut phénologique de l'espèce dans la région de Batna, 64,4 % de personnes indiquent que c'est une espèce sédentaire, observée durant toutes les saisons de l'année, et 35,6 % de réponses signalent avec précision que c'est une espèce nicheuse. > Milieux d'alimentation Les principaux milieux d'alimentation cités par les différentes personnes questionnées sont : les labours, les parcours, les prairies, les milieux cultivés, les dépôts d'aviculteurs, les eaux usées et les décharges publiques comme celle d'Ain Touta. Les milieux aquatiques fréquentés sont représentés par des barrages et des retenues d'eau comme ceux : de Timgad, la retenue collinaire d'Ain Touta, le barrage d'El Ataouta à Barika et Sebkha de Medoukel, la retenue de Seggana nommée « Khalij Sakhri », et près des forages comme à Zana El Beida et à Sefian. 60 % des réponses obtenues rapportent la présence de l'eau dans les milieux d'alimentation, 26 % notent la présence du bétail, surtout les bovins et les ovins et 18 % de réponses mentionnent la présence d'engins agricoles tels les tracteurs. > Répartition des dortoirs et des colonies L'enquête menée dans la région de Batna nous a révélé l'existence de 16 dortoirs et de 10 colonies de nidification. La répartition des différents dortoirs et colonies sur le territoire de la wilaya de Batna est reportée sur le Tableau IX. Tableau IX- Localisation et année de première implantation des colonies de nidification et des dortoirs de B. ibis dans le territoire de la wilaya de Batna.
> Intérêt de l'espèce et relation Homme-oiseau Concernant, l'utilité ou la nuisance du héron garde-boeufs, 48,8 % de réponses rapportent que l`espèce n'est considérée ni utile ni nuisible ; 26,8 % de personnes interrogés pensent qu'il est plutôt utile pour l'agriculture parce qu'il consomme les vers de terre et les insectes nuisibles, et 24,4 % rapportent que l'espèce est nuisible à cause des fientes et du bruit qu'il émet au niveau des dortoirs et des colonies. Toutes les personnes interrogées (100 % de réponses) rapportent que le Héron gardeboeufs n'est pas dissuadé ni chassé ou consommé dans la région de Batna. 2- Disponibilités des ressources alimentaires2.1- Composition taxinomique du peuplement de proies potentielles pour les deux échassiersLe dispositif d'échantillonnage appliqué dans trois milieux de gagnage connus pour être fréquentés par la Cigogne blanche et le Héron garde-boeufs : un milieu humide (marais temporaire), une pelouse naturelle et un milieu aménagé (cultures basses), durant la période allant de décembre 2006 jusqu'à novembre 2007, nous a permis de dresser une liste systématique des proies potentilles pour les modèles biologiques étudiés. Cet inventaire est dressé suite à la consultation de plusieurs guides et clés : PERRIER (1961-1963-1964), LUQUET (1984), ZAHRADNIK (1988), LERAUT (1992), AUBER (1999) et BERLAND (1999a et b) (Tabl. X). La détermination des espèces réalisée par le Dr SI BACHIR A. est poussée autant que possible jusqu'à l'espèce. Tableau X- Liste systématique des invertébrés recensés dans les divers milieux de gagnage de C. ciconia et B. ibis dans la région de Batna.
Chapitre IV- Résultats Scarites sp.
Syrphidae sp. ind.
(*) : Espèce protégée par l'arrêté du 17 janvier 1995 paru dans le journal officiel de la république Algérienne n° 19 du 12 avril 1995 complétant la liste des espèces animales non domestiques protégées en Algérie (décret n° 83-509 du 20/08/1983 relatif aux espèces animales non domestiques protégées). Cet inventaire réalisé dans la plaine d'El Madher englobe une riche variété d'espèces invertébrées, les insectes en particulier. La liste taxinomique totalise 234 espèces, réparties en 6 classes : les annélides, les gastéropodes, les arachnides, les myriapodes, les crustacés et les insectes. Ces classes sont représentées par 17 ordres, 94 familles et 194 genres. La classe des annélides n'est représentée que par l'ordre des oligochètes avec une seule espèce (Lumbricus sp.), les gastéropodes avec 4 espèces (Albea candidissima, Albea sp., Helix sp. et Helicidae sp. ind.), les arachnides avec 2 espèces (Araneus sp. et Araneidae sp. ind.), les myriapodes avec 2 espèces et les crustacés avec une seule espèce. Les insectes dominent largement avec 223 espèces, réparties en 11 ordres et 87 familles, représentent 95,3 % de l'ensemble des invertébrés recensés. Parmi les insectes, l'ordre des coléoptères réparti sur 26 familles différentes, est le plus important (84 espèces, 35,90 %), suivi par les hyménoptères (48 espèces, 20,51 %), les diptères (31 espèces, 13,25 %), les lépidoptères (21 espèces, 8,97 %) et les orthoptères (16 espèces, 6,84 %). 2.2- Variation spatio-temporelle des peuplements de proies potentielles2.2.1- Variation de l'abondance relative totaleEn termes d'abondances des proies potentielles recensées, la classe des insectes est la plus dominante, elle englobe un total de 4.520 individus soit 83,92 % de l'ensemble des invertébrés capturés, suivie par les arachnides avec 466 individus (8,65 %), les crustacés avec 281 individus (5,22 %), les gastéropodes avec 112 individus (2,08 %), les myriapodes avec 6 individus (0,11 %) et les annélides avec 1 seul individu (0,02 %). L'abondance totale des invertébrés récoltés (5.386 individus) est relativement similaire dans les trois milieux étudiés (Fig. 16A). Elle fluctue entre 31,47 % dans la pelouse naturelle, 32,45 % dans le milieu humide et 36,08 % dans la culture basse. Par contre, la répartition temporelle montre une grande variation lors des quatre saisons climatiques. Le plus grand nombre d'invertébrés est capturé au cours des saisons printanière et estivale avec respectivement 44,54 % et 24,27 % du total des captures (Fig. 16B). Concernant les méthodes de piégeage, ce sont les pots Barber qui permettent de capturer un nombre plus important d'invertébrés, soit 79,22 % du total des captures, contrairement aux pièges colorés qui n'ont capturés que 18,47 % et le filet fauchoir qui nous a permis de capturer un nombre très réduit d'invertébrés, soit 2,3 % (Fig. 16C). 2.2.2- Variation spatiale de la fréquence d'abondance et d'occurrence des différents ordres d'invertébrés recensés dans les gagnages.L'abondance relative des différents ordres recensés ne marque pas une grande variation dans les trois milieux étudiés. Sur le Tableau XI ci-après, nous notons que les hyménoptères et les coléoptères sont les ordres les plus abondants dans les trois milieux et également les plus constants dans le temps. Les diptères et les aranéides viennent en second lieu avec des abondances dépassant rarement les 10 % et sont de constance réduite. Les dermaptères, les orthoptères et les lépidoptères dont l'échantillonnage est accidentel dans les trois milieux présentent des valeurs d'abondance moins importantes. (A)
Milieux d'étude
Pelouse naturelle Milieu humide Culture basse Saisons climatiques
Hiver Printemps Eté Automne Types de pièges
Pots Barbers Pièges colorés Filet fauchoir Figure 16- Variation de l'abondance relative totale des peuplements de proies potentielles : (A) dans les milieux étudiés, (B) suivant les saisons climatiques et (C) suivant les méthodes de piégeage. Tableau XI- Fréquences d'abondance et d'occurrence des différents ordres d'invertébrés recensés en fonction des milieux étudiés.
(-) : Absence ; Ab. : Abondance ; Occu. : Occurrence ; Ech. : Echelle de constance ; C : Constant ; Ac : Accessoire ; A : Accidentel ; Tac : Très accidentel. 2.2.3- Variation temporelle de la fréquence d'abondance et d'occurrence des différents ordres d'invertébrés recensés dans les gagnages.Il est à signaler que les hyménoptères, les coléoptères, les diptères et les aranéides sont les ordres les plus abondants au cours des différentes saisons climatiques, néanmoins leur abondance en nombre varie d'une saison à une autre (Tab. XII). En été, les hyménoptères marquent une augmentation en nombre d'individus (54,09 %) contrairement aux coléoptères qui ont régressé (11,09 %) mais restent tous deux les ordres les plus occurrents en cette saison. C'est seulement pendant la saison printanière que nous enregistrons des ordres constants tels les hyménoptères, les coléoptères, les aranéides et les diptères. Par ailleurs, nous notons l'apparition de la plupart des invertébrés recensés lors de la saison automnale, mais au cours de laquelle nous notons un faible degré d'occurrence. Tableau XII- Fréquences d'abondance et d'occurrence des différents ordres d'invertébrés recensés en fonction des saisons cimatiques.
(-) : Absence ; Ab. : Abondance ; Occu. : Occurrence ; Ech. : Echelle de constance ; C : Constant ; Ac : Accessoire ; A : Accidentel ; Tac : Très accidentel. 2.3- Variation des peuplements de proies potentielles pendant les saisons phénologiques (nidification et hors nidification).Afin de mettre en évidence la variation de l'abondance en nombre dans l'espace (milieux étudiés) et dans le temps (saisons phénologiques), nous avons représenté ces variations dans la Figure 17. Dans les trois types de milieux, les hyménoptères, les coléoptères et les aranéides marquent une présence plus forte tant en période de nidification qu'en période de hors nidification. Les autres ordres ne sont présents qu'en fréquences faibles, relativement semblables pendant les deux périodes phénologiques. 2.4- Diversité et équirépartition des peuplements de proies potentielles dans les différents gagnages2.4.1- Diversité stationnelleDans le but de caractériser la diversité spécifique du peuplement d'invertébrés recensés, nous avons calculé plusieurs paramètres écologiques (richesse totale (S), richesse Pelouse naturelle
Aranea Diptera Homoptera Orthoptera Oligocheta Diplopoda Isopoda Coleoptera Chilopoda Blattoptera Isoptera Heteroptera Lepidoptera Hymenoptera Dermaptera Odonotoptera Stylommatophora Milieu humide
Aranea Diptera Isoptera Isopoda Dermaptera Coleoptera Chilopoda Oligocheta Blattoptera Orthoptera Lepidoptera Diplopoda Homoptera Heteroptera Hymenoptera Stylommatophora Odonotoptera Culture basse
Aranea Diptera Isoptera Isopoda Heteroptera Coleoptera Oligocheta Lepidoptera Diplopoda Orthoptera Chilopoda Blattoptera Homoptera Hymenoptera Dermaptera Odonotoptera Stylommatophora Nidification Hors-nidification
Figure 17- Disponibilité des proies
invertébrées en période de nidification et
hors moyenne (Sm), indice de diversité de SHANNON (H') et équirépartition (E)) pour les trois stations d'étude (Tab. XIII). La richesse totale la plus élevée est notée au niveau de la pelouse naturelle avec 157 espèces, dans laquelle est enregistrée aussi la richesse moyenne la plus élevée avec 13,08. Les valeurs les moins importantes sont mentionnées dans la culture basse. Tableau XIII- Richesse totale (S), richesse moyenne (Sm), indice de diversité de SHANNON (H') et équirépartition (E) des peuplements d'invertébrés recensés dans les trois milieux d'étude.
Par contre, l'indice de diversité de SHANNON montre, que le milieu humide abrite les peuplements les plus diversifiés avec une valeur de 5,63 bits, suivi par la pelouse naturelle avec une valeur de 5,32 bits et la culture basse avec seulement 4,76 bits. La valeur de l'équitabilité (équirépartition) varie entre 0,67 dans la culture basse et 0,78 dans le milieu humide. Cette valeur est dans l'ensemble assez proche de l'unité ce qui reflète que les peuplements des trois milieux sont assez bien équilibrés. 2.4.2- Diversité temporelleNous avons également calculé les mêmes paramètres de diversité pour les quatre saisons climatiques dont les valeurs sont reportées dans le Tableau XIV. Tableau XIV- Richesse totale (S), richesse moyenne (Sm), indice de diversité de SHANNON (H') et équirépartition (E) des peuplements d'invertébrés recensés au cours des différentes saisons climatiques.
Les saisons du printemps et de l'automne représentent les plus grandes valeurs de richesse totale avec respectivement 142 et 121 espèces, suivies par la saison d'été avec 84 espèces. De même, les valeurs de la richesse moyenne sont plus importantes au printemps (15,78) et en automne (13,44), tandis que l'indice de SHANNON révèle une diversité plus importante en automne avec 5,5 bits bien qu'elle est proche de celle du printemps (5,14 bits). Nous notons aussi que l'équirépartion varie de 0,69 à 0,79 durant toute la période d'étude. Ceci reflète un bon équilibre de la structure des peuplements durant toute la période d'étude. 2.5- Similitude des peuplements de proies potentielles recensées dans les gagnagesEn vue de comparer la composition des peuplements d'invertébrés échantillonnés entre les trois milieux pris deux à deux (Tableau XV), et également entre les saisons climatiques (Tableau XVI), nous avons calculé l'indice de similitude de SORENSON en se basant sur la présence ou l'absence des espèces. Cet indice est obtenu à partir du nombre des espèces communes entres les relevés pris deux à deux. Tableau XV- Indice de similitude de SORENSON (% de similitude) dans les trois milieux d'échantillonnage pris deux à deux.
Les valeurs de l'indice de similitude de SORENSON entre les stations varient de 58,25 % à 65,77 %. Ceci revient à dire qu'il y a une plus ou moins grande similitude des peuplements d'invertébrés entre les différents milieux. La plus grande similitude est notée entre la culture basse et la pelouse naturelle (Tab. XV). Tableau XVI- Indice de similitude de SORENSON (% de similitude) dans les saisons climatiques prises deux à deux.
La similitude de la composition des peuplements d'invertébrés suivant les saisons climatiques est la plus importante entre l'été et le printemps avec 54,87 %. Ce qui nous permet de constater une plus faible similitude entre les saisons climatiques qu'entre les milieux étudiés où cette similitude dépasse les 50 %. 3- Evolution des effectifs et modalités de fréquentation et d'exploitation des milieux d'alimentation3.1- Evolution des effectifs3.1.1- Evolution des effectifs de la Cigogne blancheL'évolution des effectifs de C. ciconia recensés mensuellement sur les deux itinéraires échantillons (zones d'El Madher et de Merouana) suivant les stades phénologiques de l'espèce sont reportés dans la Figure 18.
El Madher Merouana Effectif 60 50 40 35 49 35 23 11 3 3 30 20 10 0 7 Pr. Rep. P. C. Elev. Pt. Rep. (Pr. Rep. = période de pré-reproduction « formation des couples » ; P. C. = période de ponte et couvaison ; Elev. = période d'élevage des poussins ; Pt. Rep. = période de post reproduction). Figure 18- Evolution des effectifs de la Cigogne blanche suivant les saisons phénologiques sur deux itinéraires échantillons dans la région de Batna. Les effectifs totaux des Cigognes comptés sont comparables dans les deux zones d'étude avec 84 individus recensés à El Madher et 82 individus à Merouana. L'évolution des effectifs suivant les saisons phénologiques de l'espèce est relativement comparable aussi entre les deux zones où nous enregistrons un pic en période de ponte et couvaison des oeufs avec 49 individus notés à Merouana et 35 à El Madher. Une diminution des effectifs est enregistrée en période d'élevage dans les deux zones. L'augmentation du nombre d'individus recensés reprend à El Madher mais reste en diminution à Merouana pendant la période post reproductrice (Fig. 18). 3.1.2- Evolution des effectifs du Héron garde-boeufsL'évolution des effectifs de B. ibis pendant la période allant de décembre 2006 à septembre 2007 suivant les saisons phénologiques de l'espèce dans chacune des zones d'étude (El Madher et Merouana) sont reportés dans la Figure 19.
240 208 66 48 El Madher Merouana Effectifs 300 265 180 150 92 109 110 94 0 250 200 100 50 Hiver. Pr. Rep. P. C. Elev. Pt. Rep. (Hiver. = Période d'hivernage ; Pr. Rep. = période de pré-reproduction « apparition de plumage nuptial » ; P. C. = période de ponte et couvaison ; Elev. = période d'élevage des poussins ; Pt. Rep. = période de post reproduction). Figure 19- Evolution des effectifs du Héron garde-boeufs suivant les saisons phénologiques sur deux itinéraires échantillons dans la région de Batna. L'évolution des effectifs en relation avec les périodes phénologiques est relativement similaire dans les deux zones d'étude. L'effectif total d'oiseaux comptés s'élève à 696 individus à El Madher et 716 à Merouana. Les effectifs les plus importants sont notés pendant la période d'élevage avec 265 individus à El Madher et 240 individus à Merouana. Les gardeboeufs comptés pendant les périodes d'hivernage et de post reproduction sont moins importants, néanmoins, les effectifs les plus faibles sont notés durant les périodes de pré- reproduction et de ponte et couvaison avec 48 à El Madher et 92 à Merouana (Fig. 19). 3.2- Fréquentation et exploitation des milieux d'alimentation3.2.1- Fréquentation des milieux d'alimentation par la Cigogne blancheDurant nos sorties de terrain nous avons relevé la fréquentation de 7 types de milieux d'alimentation différents fréquentés par la Cigogne blanche. Il s'agit de : (1) Bordures d'eaux usées, (2) Prairies, (3) Friches, (4) Labours, (5) Cultures basses, (6) Déchet d'aviculteurs et (7) Milieux fauchés. Les taux de fréquentation de chacun de ces types de gagnage calculé dans les deux zones d'étude sont illustrés dans la Figure 20. A El Madher, la Cigogne blanche est beaucoup plus rencontré dans les friches durant toute la période de son séjour, notamment en période de pré-reproduction avec un taux de 100 %. Les prairies sont plus exploitées en périodes de ponte et couvaison (51,43 %), en période d'élevage (54,55 %) et de post reproduction (2,86 %). Les cultures basses ne sont fréquentées qu'en période de ponte et couvaison (14,29 %) et en période post reproductrice (40 %). En faibles taux (de 2 à 9 %), ce sont les bordures d'eaux usées qui sont utilisés en période de reproduction (ponte, couvaison et élevage des jeunes), et les labours surtout en période de ponte et couvaison (8,57 %). Les milieux fauchés sont particulièrement exploités en période d'élevage (27,27 %) et de post reproduction (51, 43 %). C'est à cette dernière période que nous avons également observé un plus grand nombre de milieux exploités par C. ciconia. En effet, elle a fréquentée en même temps les milieux fauchés, les cultures basses, les labours, les friches et les prairies (Fig. 20). Or à Merouana, durant les périodes de pré-reproduction, de ponte et couvaison et d'élevage des poussins, la Cigogne blanche fréquente principalement les prairies et les cultures basses, avec des taux allant de 30,43 % à 66,67 %. Contrairement à la zone d'El Madher, les déchets d'aviculteurs sont plus exploités dans cette zone où on les a noté depuis la période de ponte jusqu'au départ des Cigognes. Les labours sont surtout exploités en période d'élevage (8,7 %) alors que les milieux fauchés sont largement exploités en cette période ainsi qu'en période post reproductrice (71,43 %) (Fig. 20). 3.2.2- Fréquentation des milieux d'alimentation par le Héron garde-boeufsL'observation saisonnière des Hérons garde-boeufs dans leurs milieux d'alimentation de part et d'autre des itinéraires échantillons parcourus, pendant la période allant de décembre 2006 à septembre 2007, nous a permis d'enregistrer 10 types de milieux de gagnages exploités par l'espèce. Il s'agit de : (1) Bordures d'eaux usées, (2) Prairies, (3) Friches, (4) Labours, (5) Cultures basses, (6) Immondices, (7) Prairies inondées, (8) Dépôts de fumier, (9) Déchet d'aviculteurs et (10) Milieux fauchés. Les taux de fréquentation de chacun de ces types de gagnage calculé dans les deux zones d'étude suivant les saisons phénologiques de l'espèce sont représentés dans la Figure 21.
Merouana
Pr. Rep. P. C. Elev. Pt. Rep.
Bordures d'eaux usées Prairies Friches
Labours Cultures basses Déchets d'aviculteurs Milieux fauchés (Pr. Rep. = période de pré-reproduction « formation des couples » ; P. C. = période de ponte et couvaison ; Elev. = période d'élevage des poussins ; Pt. Rep. = période de post reproduction). Figure 20- Taux de fréquentation des milieux
d'alimentation par la Cigogne blanche
Chapitre IV- Résultats El Madher
Hiver. Pr. Rep. P. C. Elev. Pt. Rep. Merouana
Hiver. Pr. Rep. P. C. Elev. Pt. Rep.
Bordures d'eaux usées Prairies Friches Labours Cultures basses Immondices Prairies inondées Dépôts de fumier Déchets d'aviculteurs Milieux fauchés (Hiver. = hivernage, Pr. Rep. = période de pré- reproduction « formation des couples » ; P. C. = ponte et couvaison ; Elev. = période d'élevage des poussins ; Pt. Rep. = période de post reproduction). Figure 21- Taux de fréquentation des milieux
d'alimentation par le Héron garde-boeufs Dans la plaine d'El Madher, nous avons observé les garde-boeufs s'alimentant essentiellement dans les prairies, durant toute la période d'étude, mais avec une plus forte proportion en dehors de la période de reproduction (87,88 % en post reproduction). Les prairies inondées sont surtout exploitées en période de reproduction (18,35 %) et post reproduction. Par contre, les cultures basses, avec des taux dépassant 28 %, et les labours, avec des taux allant de 14 % à 26 %, sont surtout fréquentés en périodes d'hivernage, de préreproduction et d'élevage des poussins. C'est en période de ponte et couvaison que B. ibis exploite le plus grand nombre de types de milieux tout en utilisant en particulier les milieux fauchés, les déchets d'aviculteurs et les friches (Fig. 21). Dans la zone de Merouana, les prairies, avec des taux allant de 5 % à 47,87 %, et les labours, avec des taux allant de 2,13 % à 43,33 %, sont les milieux les plus fréquentés par les garde-boeufs tout au long de la période d'étude. Les cultures basses sont utilisées par cette espèce en période d'hivernage (20,91 %), de pré-reproduction (52,17 %) et en périodes de ponte et couvaison (3,19 %). Par ailleurs, le garde-boeufs s'alimente dans des immondices jetées dans un dépotoir à l'entrée de la ville de Merouana durant les périodes d'hivernage, de pré-reproduction et en période de reproduction (Ponte, couvaison et élevage des jeunes). De même, nous l'avons observé utilisant les déchets d'aviculteurs depuis la période de ponte et couvaison jusqu'en post reproduction (Fig. 21). 3.3- Types d'association du Héron garde-boeufs dans les milieux d'alimentationAu niveau des gagnages, les garde-boeufs sont parfois retrouvés seuls ou en association active avec le bétail et les machines agricoles, comme ils peuvent être rencontrés en compagnie d'autres espèces aviaires utilisant les mêmes milieux d'alimentations. Les taux des différents types d'associations, calculés dans les deux zones d'étude, suivant les saisons phénologiques de l'espèce sont représentés dans la Figure 22. Pour les deux zones d'étude, la proportion de hérons garde-boeufs non associés est relativement plus forte durant toutes les périodes phénologiques de l'espèce sauf en période de pré-reproduction à El Madher (Fig. 22). La période d'hivernage est aussi comparable entre les deux zones d'étude. A part les hérons garde-boeufs non associés qui dépassent 50 %, le reste des garde-boeufs, en cette période, sont distribués en association avec les bovins (15 %), les ovins (15 %) et les machines agricoles (12 %) (Fig. 22).
Merouana
Hiver. Pr. Rep. P. C. Elev. Pt. Rep.
(Hiver. = hivernage, Pr. Rep. = période de pré-reproduction « formation des couples » ; P. C. = période de ponte et couvaison ; Elev. = période d'élevage des poussins ; Pt. Rep. = période de post reproduction). Figure 22- Taux d'association du Héron
garde-boeufs au bétail, aux machines agricoles Dans la zone d'El Madher, le Héron garde-boeufs s'associe aux ovins durant toute la période d'étude. Alors qu'à Merouana, à l'exception de la période de pré-reproduction, B. ibis s'associe tout au long de la période d'étude aux bovins et aux ovins (Fig. 22). Par ailleurs, le Héron garde-boeufs est observé avec la Cigogne blanche partageant les mêmes milieux d'alimentation en périodes de pré-reproduction et d'élevage des jeunes poussins à El Madher, avec respectivement 16,44 % et 9,36 %, et en périodes de préreproduction et de ponte et couvaison à Merouana, avec respectivement 19,57 % et 4,17 % (Fig. 22). Aussi, B. ibis manifeste une association aux machines agricoles dans les deux zones d'étude en périodes d'hivernage et de pré-reproduction, avec un taux allant de 5 % à 12 % (Fig. 22). 4- Analyse qualitative et quantitative du régime alimentaire4.1- Caractérisation des pelotes de réjection4.1.1- Caractérisation des pelotes de réjection de la Cigogne blancheLes résultats relatifs à la caractérisation physique des pelotes de réjection de la Cigogne blanche (taille, poids humide et sec et nombre de proies) dans les deux zones d'étude, sont consignés dans le Tableau XVII. Tableau XVII- Mensurations et pesées des pelotes de rejection de la Cigogne blanche (N = 136 pelotes).
En moyenne, une pelote de la Cigogne blanche mesure 5,1 cm de long et 3,8 cm pour son petit diamètre et pèse 9,7 g en poids sec et contient 165,4 proies (Tab. XVII). Le nombre minimal de proies recensées dans une pelote est de 7 alors que le nombre maximal est de 1.022 items. Par ailleurs, les pelotes contiennent, en plus des fragments d'insectes et de poils agglomérés entre eux, de petites pierres sous forme de cristaux transparents ou blancs, des graines de végétaux, des fragments en plastique et de petits morceaux de bois. Nous ne relevons pas de grandes différences entre les caractéristiques des pelotes analysées des deux zones d'étude (Tab. XVII). 4.1.2- Caractérisation des pelotes de réjection du Héron garde-boeufsLes résultats concernant la caractérisation physique des différentes pelotes du Héron garde-boeufs collectées dans les deux zones d'étude, El Madher et Merouana, sont consignés dans le Tableau XVIII. Tableau XVIII- Mensurations et pesées des pelotes de rejection du Héron garde-boeufs (N = 957 pelotes).
En moyenne, une pelote de héron garde-boeufs mesure 3,7 cm de long, 2,1 cm de diamètre et pèse 1,8 g en poids sec (Tab. XVIII). Ces pelotes sont généralement de forme cylindrique, légèrement effilées sur un coté et de taille et de couleur très variables. De même que pour les pelotes de la Cigogne blanche, celles du Garde-boeufs contiennent aussi, en plus des fragments d'insectes et de poils agglomérés entre eux, de petites pierres, des graines de végétaux, des fragments en plastique et de petits morceaux de bois. Les pelotes du garde-boeufs contiennent en moyenne 85,3 de proies. Le nombre minimal de proies recensées est de 3 et le nombre maximal est de 586 proies (Tab. XVIII). 4.2- Spectre alimentaire global des deux espècesLes deux échassiers étudiés ont un large spectre alimentaire composé de proies invertébrées et vertébrées. Ainsi, nous avons recensé au total 146 espèces proies différentes réparties en 9 classes, 19 ordres et 57 familles, dont 79 espèces proies de la Cigogne blanche et 134 espèces proies du Héron garde-boeufs (Tableau XIX). Les insectes dominent largement le régime alimentaire des deux échassiers, avec 129 espèces différentes, représentant ainsi 88,36 % de l'ensemble des espèces recensées, suivi par la classe des mammifères avec 8 espèces (5,48 %) et la classe des arachnides avec 3 espèces (2,05 %). Les classes des gastéropodes, myriapodes, poissons, amphibiens, reptiles et des oiseaux ne sont représentées que par une seule espèce chacune, soit 0,68 % du spectre alimentaire global des deux échassiers. Tableau XIX- Liste systématique des espèces proies recensées dans les pelotes de rejection de la Cigogne blanche (C. ciconia) et du Héron garde-boeufs (B. ibis) dans la région de Batna.
(+) présence, (-) absence, (*) : Espèce protégée par l'arrêté du 17 janvier 1995 paru dans le journal officiel de la république Algérienne n° 19 du 12 avril 1995 complétant la liste des espèces animales non domestiques protégées en Algérie (décret n° 83-509 du 20/08/1983 relatif aux espèces animales non domestiques protégées). Le régime alimentaire du Héron garde-boeufs est riche et varié en proies avec une richesse spécifique totale de 134 espèces différentes réparties en 9 classes, 19 ordres et 56 familles. Par contre, le régime alimentaire de la Cigogne blanche révèle une richesse spécifique moins importante, avec 79 espèces réparties en 5 classes, 11 ordres et 34 familles. Les proies invertébrées représentent la majorité des espèces consommées, avec 97,47 % pour la Cigogne blanche et 91,04 % pour le Héron garde-boeufs. Parmi les invertébrés consommés, la classe des insectes domine largement avec 74 espèces, soit 93,67 %, pour la Cigogne blanche, et 117 espèces, soit 87,31 %, pour le Héron garde-boeufs. Il est à signaler également l'importance relative des ordres des coléoptères, orthoptères, dermaptères, hyménoptères et aranéides chez B. ibis, avec respectivement 48,21 %, 10,71 %, 5,36 %, 3,57 % et 3,57 % du total des espèces proies consommées. Ainsi que pour C. ciconia qui consomme 47,06 % de coléoptères, 17,65 % d'orthoptères, 8,82 % de dermaptères, 5,88 % d'hétéroptères et 2,94 % pour le reste des ordres. 4.3- Analyse de la composition et de la structure du régime alimentaire4.3.1- Composition et structure globales du régime alimentaire des deux échassiersEn termes d'abondance totale des proies consommées, c'est la Cigogne blanche qui s'attaque à un plus grand nombre de proies avec un total global de 15.881 individus recensés dans 96 pelotes dépassant ainsi, largement le nombre de proies consommées par le Héron garde-boeufs qui est de 9.387 individus dans 110 pelotes. Pour la Cigogne blanche, la classe des insectes est la plus dominante en nombre d'individus avec 15.759 soit 99,23 %, suivie par celle des arachnides avec 79 individus (0,5 %). De même pour le Héron garde-boeufs, la classe des insectes englobe un total de 9.234 individus soit 98,37 % suivie par la classes des arachnides avec 74 individus soit 0,79 % et la classe des mammifères avec 39 individus soit 0,42 %. La composition et la structure globales du régime alimentaire des deux modèles biologiques étudiés, à travers le calcul de la fréquence d'abondance et de la fréquence d'occurrence des différentes familles de proies composant leur régime alimentaire, sont reportées dans le Tableau XX. Tableau XX- Fréquences d'abondance et d'occurrence des différentes familles de proies composant le régime alimentaire de C. ciconia et B. ibis dans la région de Batna.
(-) : Absence ; Ech. : Echelle de constance ; C : Constante ; Ac : Accessoire ; A : Accidentelle ; Tac : Très accidentelle. Le spectre alimentaire de la Cigogne blanche est composé de 34 familles différentes mais seulement quelques familles sont les plus occurrentes et apparaissent régulièrement dans les pelotes de réjection (8 familles constantes et 7 familles accessoires). Parmi les familles les plus abondantes en nombre dans le régime alimentaire de C. ciconia nous en citons les Caelifera ind. avec une abondance relative de 37,08 %, suivie par les familles des Scarabaeidae (12,1 %), Ensifera ind. (11,47 %), Labiduridae (6,86 %) et celle des Silphidae (6,65 %). Nous remarquons également l'importance numérique des familles des Tenebrionidae, Carabidae et Harpalidae (Tab. XX). Parallèlement, le spectre alimentaire du Héron garde-boeufs est plus diversifié que celui de la Cigogne blanche, par le fait qu'il est constitué de toutes les familles recensées à l'exception de celle des Pentatomidae. Cependant, la structure du régime alimentaire de B. ibis est comparable avec celle de C. ciconia, dans la mesure où seulement 8 familles sont constantes et 9 familles sont accessoires. Parmi les familles les plus abondantes en nombre, nous citons en tête de toutes les familles celle des Formicidae avec une abondance de 14,52 %, suivie par les familles des Caelifera ind. (12,79 %), Acrididae (12,65 %), Carabidae (11,87 %) et celle des Labiduridae (10,5 %). De même, il est à remarquer l'importance des familles des Harpalidae, Pterosrichidae, Scarabaeidae et les Gryllidae avec, toutefois, des fréquences moindres (Tab. XX). 4.3.2- Variation spatio-temporelle du régime alimentaire de la Cigogne blanche > Variation spatialeLes fréquences d'abondance et d'occurrence des différents ordres de proies consommées par C. ciconia dans chacune des zones d'étude et dans la région de Batna en général, sont consignées dans le Tableau XXI. Tableau XXI- Fréquences d'abondance et d'occurrence des différents ordres de proies consommées par la Cigogne blanche dans chacune des zones d'étude et dans la région de Batna.
(-) : Absence ; Ab. : Abondance ; Occu. : Occurrence ; Ech. : Echelle de constance ; C : Constant ; Ac : Accessoire ; A : Accidentel ; Tac : Très accidentel. Les coléoptères, les orthoptères et les dermaptères sont les ordres les plus abondants et les plus constants dans les deux zones d'étude (Tab. XXI). A El Madher, l'ordre des coléoptères enregistre une abondance plus élevée avec 56,21 % de l'effectif total des différents ordres, et est signalé avec une occurrence de 100 %, c'est-àdire qu'il est présent dans toutes les pelotes de rejection triturées. L'ordre des orthoptères vient en second lieu avec 30,09 % d'abondance en nombre et 79,31 % d'occurrence, suivi par l'ordre des dermaptères avec 11,67 % d'abondance et 93,1 % de fréquence d'occurrence (Tab. XXI). Par ailleurs, dans la zone de Merouana, c'est l'ordre des orthoptères qui est noté avec la plus grande abondance (75,4 %) et une occurrence de 81,58 %, suivi par l'ordre des coléoptères avec une abondance de 22,5 % et une occurrence de 100 % et l'ordre des dermaptères qui est présent dans 50 % des pelotes avec une abondance relative totale de 1,31 % (Tab. XXI). En général, dans la région de Batna les pelotes de C. ciconia contiennent en moyenne 50,75 % d'orthoptères avec une occurrence dépassant 80 %, et 40,84 % de coléoptères avec une apparition absolue, et 6,95 % de dermaptères avec une occurrence de 76,04 %. L'ordre des hyménoptères est accessoire dans les deux zones d'étude avec une abondance moyenne de 0,49 % et une occurrence moyenne de 32,29 % (Tab. XXI). > Variation temporelle L'étude de la variation spatiale des différents ordres consommés par la Cigogne blanche nous a permis de mettre en évidence cinq ordres importants dans son régime alimentaire. A cet effet, nous avons étudié la variation de ces ordres en fonction des six mois de séjour da la cigogne dans la région d'étude comme illustré dans la Figure 23. Dans la zone d'El Madher, l'ordre des coléoptères est le plus dominant lors des quatre premiers mois, notamment aux mois de mars (77,42 %), avril (75,91 %) et mai (63,48 %). Aux mois de juin et de juillet, ce sont les orthoptères qui dominent largement avec un taux dépassant 52 % (Fig. 23). Les dermaptères, présents dans les pelotes de la Cigogne durant les six mois d'observation, sont consommés avec des taux fluctuant entre 5,03 % au mois de juin et 19,64 % au mois de mars. Les hyménoptères enregistrent leurs taux les plus élevés aux mois de février (0,62 %) et de juin (0,89 %), alors que les reptiles sont surtout présents au mois de mars avec un taux de 0,44 % (Fig 23). Cependant, à Merouana, c'est l'ordre des orthoptères qui domine durant toute la période de séjour de la Cigogne avec des taux fluctuant entre 51,16 % au mois de juin et 94,93 % au mois de février. En fait, les coléoptères enregistrent des taux plus élevés aux mois d'avril (39,75 %) et de juin (40,45 %). Nous notons aussi l'importance des hyménoptères dans la zone de Merouana où ils sont surtout présents au cours des mois d'avril (0,89 %), de mai (1,91 %) et de juin (1,75 %). Les dermaptères enregistrent un taux plus élevé au mois de juin (6,37 %) alors que les reptiles sont totalement absents (Fig. 23). Globalement, dans la région de Batna les taux des coléoptères et des orthoptères consommés varient d'une façon inverse. En effet, le taux des coléoptères augmente à partir du mois de février (30,89 %) pour atteindre un maximum au mois d'avril (65,71 %) puis redescend jusqu'au mois de juillet (25,66 %). Par contre, les orthoptères marquent le taux El Madher
Merouana
Région de Batna
Février Mars Avril Mai Juin Juillet
Figure 23- Variation mensuelle de l'abondance relative
des cinq ordres de proies les plus le plus élevé aux mois de février (60,77 %) et de juillet (70,04 %) et des taux moyens aux mois de mars (39, 94 %), de mai (30,97 %) et de juin (52,42 %). Le taux le plus faible est noté au mois d'avril avec 22,32 % (Fig. 23). Les dermaptères qui sont présents dans le régime alimentaire de la Cigogne tout au long de la période d'étude, sont notés avec des taux fluctuants entre 3,84 % au mois de juillet et 12,39 % au mois de mai. Les hyménoptères et les reptiles sont surtout consommés respectivement aux mois de juin (1,14 %) et d'avril (0,30 %) (Fig. 23). > Variation en fonction des périodes de reproduction et hors reproduction Afin de mieux comprendre l'importance des principaux ordres de proies consommées par la Cigogne blanche, nous avons calculé leurs taux en fonction des périodes de reproduction et hors reproduction (Fig. 24). Ciconia ciconia
Hors reproduction Reproduction Rodentia
Figure 24- Fréquence d'abondance relative des
principaux ordres de proies composant Pendant la période de reproduction, C. ciconia consomme un taux plus élevé de coléoptères avec 51,35 % suivis par les orthoptères avec 38,02 % et les dermaptères avec 8,54 %. Les hyménoptères, reptiles et mammifères sont consommés avec de très faibles taux avec respectivement 0,64 %, 0,13 % et 0,06 % (Fig. 24). A l'opposé, en période hors reproductrice, la cigogne consomme un taux plus élevé d'orthoptères avec 67,14 % suivi par les coléoptères (27,29 %) et les dermaptères (4,89 %) (Fig. 24). 4.3.3- Variation spatio-temporelle du régime alimentaire du Héron garde-boeufs > Variation spatialeLes fréquences d'abondance et d'occurrence des différents ordres de proies entrant dans la composition du régime alimentaire de B. ibis dans chacune des zones d'étude et globalement dans la région de Batna sont reportées dans le Tableau XXII. Tableau XXII- Fréquence d'abondance et d'occurrence des différents ordres de proies consommées par le Héron garde-boeufs dans chacune des zones d'étude et dans la région de Batna.
(-) : Absence ; Ab. : Abondance ; Occu. : Occurrence ; Ech. : Echelle de constance ; C : Constant ; Ac : Accessoire ; A : Accidentel ; Tac : Très accidentel. Quatre ordres principaux sont considérés abondants et apparaissent régulièrement dans le régime alimentaire du Héron garde-boeufs (Tab. XXII). Dans la zone d'El Madher, les coléoptères sont en tête du régime alimentaire du gardeboeufs avec une abondance relative de 36,97 % et une occurrence de 100 %, suivis par les orthoptères avec un taux de 27,29 % avec une apparition dans 80 % des pelotes triturées. Les hyménoptères viennent en troisième position avec une abondance de 18,69 % et une occurrence dans 60 % des pelotes triturées. Les dermaptères sont notés avec une abondance de 13,87 % et une occurrence dépassant 85 % (Tab. XXII). Bien que quatre ordres (Myriapoda ind., Diptera, Cypriniformes, Mammalia ind.) ne sont pas recensés dans les pelotes de B. ibis collectées dans la zone de Merouana, les ordres des coléoptères et des orthoptères sont enregistrés avec des proportions d'abondance relativement similaires, avec respectivement 41,83 % et 39,69 %. La fréquence d'apparition des coléoptères est très importante dépassant 98 %, alors que l'occurrence des orthoptères atteint seulement 78 % (Tab XXII). Les hyménoptères et les dermaptères sont enregistrés aussi avec des proportions proches avec respectivement 8,82 % et 7,29 % représentant ainsi une abondance presque égale à la moitié de celle notée dans la zone d'El Madher (Tab. XXII). Hormis, l'ordre des rongeurs qui est classé accessoire dans les deux zones d'étude avec une apparition moyenne de 33,64 % et une abondance moyenne de 0,4 %, nous remarquons de faibles taux d'abondance des ordres des aranéides (1,24 %) et des mantoptères (0,66 %) à El Madher et des ordres des blatoptères (0,71 %) et des mantoptères (0,38 %) à Merouana. Ces types de proies restent tous accidentelles (Tab. XXII). En général, dans la région de Batna les pelotes de B. ibis sont composées essentiellement de 39 % de coléoptères avec une occurrence très élevée atteignant 99,09 %, 32,47 % d'orthoptères avec une apparition dans 79,09 % de pelotes, 14,56 % d'hyménoptères avec une apparition dans 59,09 % de pelotes et 11,12 % de dermaptères avec une occurrence dépassant 68 % (Tab. XXII). Toutefois, il ne faut pas négliger la présence des rongeurs, des aranéides et des mantoptères dans le régime alimentaire de cet échassier. > Variation temporelle L'étude de la variation spatiale des différents ordres de proies consommées par le Héron garde-boeufs nous a permis de mettre en évidence cinq ordres importants dans son régime alimentaire. L'analyse de la variation de l'abondance relative de ces cinq ordres de proies en fonction des 11 mois d'étude est consignée dans la Figure 25. Dans la zone d'El Madher, les coléoptères sont consommés par le Héron garde-boeufs durant toute la période d'étude enregistrant un taux faible seulement aux mois d'août (7,89 %) et de septembre (10,23 %), le reste des mois leurs taux fluctuent entre 34,99 % au mois de janvier et 84,59 % au mois d'avril, alors que les pourcentages des dermaptères varient entre 2,87 % au mois de septembre et 46,54 % au mois de mars (Fig. 25). El Madher
Mars Mai Aofit Juin Janvier Juillet Avril Fevrier Novembre Septembre Decembre Merouana
Mars Mai Aofit Juin Janvier Juillet Avril Fevrier Novembre Septembre Decembre Région de Batna
Mars Mai Aofit Juin Janvier Juillet Avril Fevrier Novembre Septembre Decembre
Figure 25- Variation mensuelle de l'abondance relative
des cinq ordres de proies les plus Les orthoptères sont très fluctuants dans le régime alimentaire du garde-boeufs. Absents au mois de mars, noté avec un taux faible au mois d'avril (1,26 %), et fluctuent d'un mois à l'autre de 8 % (décembre) à 70 % (août). Les hyménoptères sont surtout consommés aux mois de septembre (69,06 %), de décembre (19,78 %) et de janvier (15,74 %) (Fig. 25). A Merouana, les coléoptères et les orthoptères dominent également le régime alimentaire de B. ibis apparaissant le long de toute la période d'étude. Les coléoptères ont un taux plus faible au mois d'août avec 5,05 %, et fluctuent au cours du reste des mois entre 24,67 % au mois de septembre et 83,13 % au mois de mars. Les orthoptères ont un taux plus élevé au mois d'août avec 93,14 % et fluctuent lors du reste des mois entre 7,25 % au mois de novembre et 46,90 % au mois de décembre (Fig. 25). Les dermaptères et les hyménoptères enregistrent également une présence notable pendant la plupart des mois d'étude, en particulier en avril, en mai et en septembre où l'on enregistre respectivement 13,48 %, 16,43 % et 19,24 % de dermaptères et les mois de novembre et de février où les hyménoptères sont notés respectivement avec 35,5 % et 13,05 % (Fig. 25). Les rongeurs sont consommés surtout au mois de décembre avec un taux de 1,1 % dans la zone d'El Madher, et en particulier durant les mois de novembre et de décembre avec respectivement 1,91 % et 1,16 % dans la zone de Merouana (Fig. 25). Globalement, dans la région de Batna, les coléoptères, les orthoptères, les dermaptères et les hyménoptères sont les principaux ordres de proies consommées par B. ibis. Leur consommation fluctue d'un mois à un autre. Le taux des coléoptères varie de 6,88 % au mois d'août à 72,65 % au mois d'avril et le taux des orthoptères varie de 2,47 % au mois de mars à 78,71 % au mois d'août. Les dermaptères enregistrent un maximum au mois de mars (32,77 %) et un minimum au mois de février (3,5 %). Les hyménoptères sont notés avec des taux faibles aux mois de juillet et d'août et des taux élevés au mois de septembre (43,22 %). Par ailleurs, l'ordre des rongeurs est consommé par le garde-boeufs dans la région de Batna, surtout aux mois de novembre (1,1 %) et décembre (1,13 %) (Fig. 25). > Variation en fonction des périodes de reproduction et hors reproduction Dans le but de mieux comprendre la variation de ces principaux ordres de proies de B. ibis, nous avons opté à calculer leurs taux en fonction des périodes de reproduction et hors reproduction (Fig. 26). Chapitre IV- Résultats Bubulcus ibis Hors reproduction Reproduction
70 60 50 40 30 20 10 0 En (%) Rodentia Figure 26- Fréquence d'abondance relative des
principaux ordres de proies composant Pendant la période de reproduction, le Héron garde-boeufs consomme un taux plus élevé d'orthoptères avec 46,12 %, suivis par les coléoptères avec 37,49 %, les dermaptères avec 11 % et les hyménoptères avec 2,13 % (Fig. 26). En période hors reproductrice, B. ibis consomme un taux plus important de coléoptères avec 40,01 %, suivis par les orthoptères et les hyménoptères avec des proportions proches, avec respectivement 23,38 % et 22,84 % et les dermaptères avec 11,20 % (Fig. 26). Il est à noter que la consommation des dermaptères entre les deux périodes phénologiques est relativement égale avec un taux de 11 %, ainsi que pour les coléoptères (37,49 % et 40,01 %) (Fig. 26). 4.4- Diversité et équirépartition des peuplements de proiesLes paramètres de diversité de la composition du régime alimentaire des deux espèces étudiées, pris d'une manière globale et en période de reproduction, sont consignés dans le Tableau XXIII. La richesse totale est plus élevée au mois de mars pour la cigogne avec 67 espèces et au mois de juin pour le garde-boeufs avec 80 espèces. Parallèlement, les taux les plus élevés de richesse moyenne sont également notés pendant ces deux mois (Tab. XXIII). Tableau XXIII- Variation mensuelle des paramètres de diversité calculés pour la composition du régime alimentaire de C. ciconia et B. ibis.
L'indice de SHANNON s'élève à 4,3 bits chez la Cigogne blanche au mois d'avril et à 5,2 bits chez le Héron garde-boeufs au mois de juin (Tab. XXIII). L'équirépartition des différentes catégories de proies consommées enregistre pendant la période de reproduction des valeurs un peu plus élevées chez le garde-boeufs avec 0,9 au mois d'avril, que chez la Cigogne avec 0,7 aux mois d'avril et de mai, et des valeurs respectivement de 0,7 et 0,6 en analyse totale reflétant ainsi une plus forte équirépartition des proies consommées par B. ibis qui sont aussi plus diversifiées que celles consommées par C. ciconia (Tab. XXIII). Dans l'ensemble, la composition du régime alimentaire est plus équilibrée chez le Héron garde-boeufs que chez la Cigogne blanche. 4.5- Similitude des proies consommées par les deux échassiers4.5.1- Similitude des proies consommées par C. ciconia entre les deux zones d'étudeLe nombre d'espèces recensées dans le régime alimentaire de la Cigogne blanche dans la zone d'El Madher est de 78 espèces, alors qu'à Merouana, il est de 49 espèces. Le nombre d'espèces recensées simultanément (dans les deux zones) est de 48 espèces. De ce fait, l'indice de SORENSON calculé entre les deux zones est de 75,59 ce qui signifie qu'il existe une grande similitude entre les deux zones d'étude. 4.5.2- Similitude des proies consommées par B. ibis entre les deux zones d'étudeLe nombre d'espèces recensées dans le régime alimentaire du Héron garde-boeufs dans la zone d'El Madher est de 117 espèces, alors qu'à Merouana, il est de 114 espèces. Le nombre d'espèces recensées simultanément (dans les deux zones) est de 97. A cet effet, l'indice de SORENSON calculé entre les deux zones est de 83,98 ce qui veut dire qu'il existe une forte similitude entre les deux zones d'étude. 4.5.3- Similitude des proies consommées par les deux échassiersLe nombre d'espèces recensées dans le régime alimentaires de la Cigogne blanche est de 79 espèces, alors que le nombre d'espèces recensées dans le régime alimentaire du Héron garde-boeufs est de 134 espèces. Le nombre d'espèces recensées simultanément dans les régimes alimentaires des deux espèces est de 67 espèces. Ainsi, l'indice de SORENSON calculé est de 62,91 ce qui signifie qu'il existe une assez grande similitude entre les régimes alimentaires des deux espèces. 4.6- Electivité des proies consommées par les deux échassiersAfin d'étudier la relation existante entre les disponibilités alimentaires évaluées dans les milieux de gagnage et la composition du régime alimentaire à partir de l'analyse des pelotes de réjection des deux espèces étudiées, nous avons calculé l'indice d'IVLEV (E), qui permet de comparer l'abondance relative des proies disponibles dans le milieu par rapport à l'abondance de proies retrouvées dans les pelotes (Fig. 27). Pour la Cigogne blanche, les valeurs de cet indice varient de + 0,12 à + 0,96 pour la sélection positive des proies et de - 0,93 à - 0,98 pour la sélection négative (Fig. 27A). Les valeurs positives sont enregistrées pour les dermaptères, orthoptères et coléoptères, alors que les valeurs négatives sont enregistrées pour les aranéides et les hyménoptères. Pour le Héron garde-boeufs, les valeurs de l'indice d'IVLEV (E) varient de + 0,1 à + 0,94 pour la sélection positive des proies et de - 0,46 à - 0,86 pour la sélection négative (Fig. 27B). De même que pour la cigogne, chez le garde-boeufs, les valeurs positives sont enregistrées pour les dermaptères, les orthoptères et les coléoptères, alors que les valeurs négatives sont notées pour les aranéides et les hyménoptères. Nous remarquons également que les ordres mentionnés avec des valeurs positives ou négatives sont relativement identiques dans les trois types de gagnages prospectés (Fig. 27). 4.7- Analyse du régime alimentaire par application d'une méthode d'analyse statistiqueNous avons réalisé une analyse de la variance bivariée (test à deux échantillons) de la variation de la composition du régime alimentaire de C. ciconia et B. ibis dans les deux zones d'étude pendant les deux périodes phénologies, en plus d'une analyse globale. Les résultats de cette analyse sont reportés dans le Tableau XXIV. (A) (B) Ciconia ciconia
Pelouse naturelle Milieu humide Culture basse Bubulcus ibis
Pelouse naturelle Milieu humide Culture basse Aranea Dermaptera Orthoptera Coleoptera Hymenoptera
Figure 27- Indice d'électivité d'IVLEV (E) des principales catégories de proies consommées par C. ciconia (A) et B. ibis (B) dans trois milieux de gagnages différents dans la région de Batna. Tableau XXIV- Résultats de l'analyse de variance bivariée (ANOVA) de la variation de la composition du régime alimentaire des deux échassiers.
n.s. : Non significative ; * : Significative. La lecture des résultats de l'analyse de la variance bivariée montre qu'il n'existe pas de variation significative entre tous les facteurs étudiés à l'exception de la variation de la composition du régime alimentaire des deux espèces pendant la période de nidification dans la zone d'El Madher qui est significative (d.l. = 296 ; P = 0,027) (Tab. XXIV). Nous avons également effectué une analyse monovariée (ANOVA à un facteur contrôlé) pour chacune des deux espèces étudiées dans les deux zones d'étude. Les résultats de cette analyse sont consignés dans le Tableau XXV. Tableau XXV- Résultats de l'analyse de variance monovariée (ANOVA) de la variation de la composition du régime alimentaire des deux échassiers.
n.s. : Non significative. La lecture des résultats de l'analyse de la variance monovariée révèle qu'il n'existe pas de variation significative entre tous les facteurs étudiés (Tab. XXV). Chapitre V : Discussions1- Etude de la phénologie des deux espèces1.1- Etude du cycle biologique de la Cigogne blancheEn l'an 2007, les dates d'arrivées de la Cigogne blanche dans la région de Batna sont étalées de la troisième décade de janvier (El Madher, Merouana et Ain Touta) à la première décade de février (Batna et Arris). Les dates de départ, sont enregistrées de la troisième décade de juillet (colonie d'El Madher) à la mi-août (Arris). Les dates d'arrivées et de départs de la Cigogne blanche fluctuent dans des intervalles allant jusqu'à plus de 20 jours selon la région et l'année. Sur le Tableau XXVI ci-après, nous reportons quelques dates d'arrivées et de départs de la Cigogne blanche dans quelques régions d'Algérie. Tableau XXVI- Dates d'arrivées et de départs de la Cigogne blanche dans quelques régions d'Algérie.
Selon JESPERSEN (1949), en Algérie, la date moyenne d'arrivée des cigognes est notée à partir de la première décade de février et la date moyenne de départ en migration postnuptiale coïncide avec la première décade d'août. Au Danemark, entre 1977 et 1991, l'arrivée des cigognes est enregistrée entre la fin mars et le début avril et leur départ entre la première décade d'août et la première décade de septembre (SKOV, 1991a). En plus des observations faites sur les cigognes dans les deux stations d'étude, nous avons marqué l'arrivé d'un essaim de cigognes dans la région d'Arris au coucher du soleil du 25 août 2007, supposées venues des régions nord du pays ; les cigognes ont passé la nuit à Arris et ont repris le chemin vers le sud à l'aube du lendemain. Par ailleurs, il est à signaler l'observation d'une cigogne blanche à moins de 500 m au nord de la colonie d'El Madher après le départ de ses congénères en 2007. L'hivernage des cigognes dans les lieux de reproduction est signalé par plusieurs auteurs en Europe. Nous citons entre autres, le cas d'une cigogne mâle hivernant au Danemark pendant les hivers 1985-86, 1986-87 et 1987-88 (SKOV, 1991a). En général, les couples se constituent dès la fin de l'hiver, au début d'avril (SCHIERER, 1967), ou dans les 8 jours qui suivent l'arrivée du premier membre du couple qui est le mâle le plus souvent (BOUET, 1950). Cette dernière observation est notée également dans la région de Batna où nous avons noté les premiers accouplements le 12 février 2007 dans la colonie d'El Madher. Dans la région d'étude, selon nos résultats et ceux de DJADDOU et BADA (2006), la période de nidification de C. ciconia s'étale sur une durée d'un peu plus de trois mois, allant de la troisième décade de mars à la troisième décade de juin, coïncidant ainsi avec la période printanière. Ces résultats sont obtenus également par BOUKHEMZA (2000), dans la vallée du Sébaou (Tizi-Ouzou). A Béjaia, en l'an 2001, C. ciconia a commencé un peu tôt la reproduction, dès la première décade de mars jusqu'à la première décade de juin (ZENNOUCHE, 2002). Au Danemark, la ponte ne commence qu'à la deuxième décade de mai, et l'envol des cigogneaux à partir de la deuxième décade de juillet (SKOV, 1991a). Ceci serait en relation avec la date d'arrivée et la localisation des régions en latitude. A l'instar de l'observation des périodes phénologiques de la cigogne, nous avons également noté des comportements étonnants des cigognes face aux conditions climatiques pendant la période de reproduction. Afin d'augmenter la température du nid, la cigogne apporterait des morceaux de fumier au nid (Fig. 28A), et utiliserait des sachets en plastique de couleur sombre afin de couvrir les oeufs (Fig. 28B et C) et d'augmenter leur température par « effet de serre » ! Mais, pourquoi une cigogne couvrirait-elle deux oeufs et laisserait les deux autres sans couverture ? De telles observations, non étudiées lors de ce travail, nous amèneraient à de nouveaux sujets de recherche sur l'éthologie de l'espèce notamment en période de nidification. 1.2- Etude du cycle biologique du Héron garde-boeufsQuant au héron garde-boeufs, espèce sédentaire à installation récente dans la région de Batna, la période de reproduction s'étale sur une période de près de quatre mois. A Merouana, la période de reproduction du garde-boeufs s'étale sur près de 4 mois, de la première décade d'avril à la première décade d'août. Cette période de reproduction est Chapitre V- Discussions
(A) (B) (C) Figure 28- La Photo (A) illustre une Cigogne blanche qui prend du fumier dans son bec pour le ramener au nid (El Madher, 03 avril 2007) ; les photos (B) et (C) illustrent la manière dont la cigogne couvre ses oeufs par des matières en plastique (El Madher, 23 avril 2007) (Photos personnelles). similaire à celles signalées par BOUKHEMZA (2000) dans la Kabylie du Sébaou (TiziOuzou) et PROSPER et HAFNER (1996) dans l'Albufera de Valencia en Espagne, qui ont noté des périodes de ponte plus précoces, à partir de la fin mars jusqu'à la fin juillet. La période de reproduction du garde-boeufs à El Madher est un peu tardive. La ponte n'a commencé qu'au début mai. Ceci est également le cas observé en Camargue, où la nidification ne débute qu'à la deuxième décade d'avril et se prolonge jusqu'à la fin août (HAFNER, 1982). Pour SBIKI (2008), à Tébessa, la période de reproduction du garde-boeufs est un peu plus longue, allant de la première décade de mars jusqu'à la troisième décade d'août. A El Kala, l'espèce niche de la mi-avril jusqu'à la mi-août (SAMRAOUI-CHENAFI, 2009). La reproduction chez B. ibis est fortement conditionnée par les conditions climatiques. Selon SI BACHIR (2007), dans la Kabylie de la Soummam (Béjaia), en 1997, année relativement peu pluvieuse, l'installation des nids s'est arrêtée à la fin juin. Par contre, en 1998, année bien arrosée, l'installation des nids a connu une interruption entre juillet et août pour reprendre une nouvelle fois (deuxième nichée) au début septembre. D'après ELKINS (2001), assurer l'élevage quand la nourriture est la plus abondante c'est, on l'admet en général, la raison fondamentale de la reproduction des oiseaux à un moment précis. Pour les deux échassiers étudiés, les périodes de reproduction se sont déroulées en période printanière et estivale. Selon ELKINS (2001), certaines études de populations suggèrent que chez les oiseaux insectivores (dont les échassiers), il y a corrélation entre la ponte et l'apparition d'insectes en quantité suffisante pour nourrir la femelle en train de former ses oeufs. C'est pendant ces deux périodes, printanière et estivale, que nous avons noté les plus grands effectifs d'invertébrés échantillonnés dans les milieux de gagnage, avec 44,54 % au printemps et 24,27 % en été. 1.3- Evolution des effectifs des deux espècesNous avons distingué principalement quatre phases d'évolution des effectifs correspondant aux phases phénologiques des modèles biologiques étudiés : période d'hivernage, période de pré-reproduction, période de ponte et couvaison et période de post reproduction. 1.3.1-Evolution des effectifs de la Cigogne blanchePendant la période de ponte et couvaison, nous avons noté un nombre important de cigognes dans les deux zones d'étude, à Merouana et à El Madher. En période d'élevage, nous avons enregistré une diminution des effectifs des cigognes dans les milieux d'alimentation, ceci revient au fait que les cigognes ne s'éloignent pas des colonies de reproduction et se concentrent sur les gagnages les plus proches des sites de nidification. Pendant la phase de post reproduction (juillet-août), les effectifs de cigognes ont augmenté à El Madher et ont diminué à Merouana. Ceci serait en relation avec la dispersion des adultes et des jeunes de l'année dans les gagnages. 1.3.2-Evolution des effectifs du Héron garde-boeufsPendant la période d'hivernage (novembre-décembre), nous avons noté un grand effectif de hérons garde-boeufs à El Madher (208 individus) et à Merouana (110 individus) ce qui serait probablement dû à l'arrivée de migrateurs hivernants qui se joignent aux populations autochtones. Ces deux régions sont situées dans la zone des hautes plaines ce qui leur procurent une nourriture niche et variée même pendant cette période à basses températures. Dans le complexe des zones humides d'El Kala, le nombre de garde-boeufs diminue en période d'hivernage sous l'effet des pluies intenses qui inondent la plupart des milieux d'alimentation (CHALABI-BELHADJ, 2008). Durant la période de pré-reproduction (janvier-février-mars), les effectifs des gardeboeufs diminuent par le fait que la population hivernante part vers d'autres sites de reproduction et il ne reste dans la région d'étude que les reproducteurs locaux. L'augmentation des effectifs des garde-boeufs pendant la période d'hivernage et leur diminution en période de pré-reproduction est constatée aussi par FRANCHIMONT (1986b) au Maroc et par SALMI (2001) à Béjaia. En période de ponte et couvaison (avril-mai-juin), les effectifs des garde-boeufs dans les gagnages diminuent par le fait que les colonies de reproduction sont investies et la majorité des individus se concentrent au voisinage des sites de reproduction. La période d'élevage est caractérisée par une activité de chasse très intense que montre les adultes nicheurs pour satisfaire leurs besoins alimentaires et ceux de leur progéniture et ne s'éloignent pas, de ce fait, des sites des colonies. Ceci expliquerait l'importance des effectifs de garde-boeufs en cette période à El Madher (265 individus) et à Merouana (240 individus), sachant notamment que les itinéraires choisis couvrent une aire importante des principaux milieux fréquentés et passent non loin des colonies de reproduction. Selon CHALABI-BELHADJ (2008), durant la saison de nidification de hérons gardeboeufs dans le complexe de zones humides d'El Kala, la dispersion des individus devient moins lâche et on assiste à un regroupement plus important et une concentration des individus sur les gagnages proches des sites de nidification situés dans les secteurs du marais de la Mekhada et du Lac Tonga. Pendant la phase de post-reproduction (août-septembre), les effectifs de B. ibis diminuent légèrement. Cette phase correspondrait à la fin de la période de nidification, donc à la dispersion des jeunes de l'année et des adultes. Ces résultats sont similaires à ceux obtenus par CHALABI-BELHADJ (2008), qui a noté des effectifs relativement importants de gardeboeufs en cette période. L'évolution des effectifs saisonniers de la Cigogne blanche et du Hérons garde-boeufs sur les itinéraires choisis, dépendrait non seulement du cycle biologique de l'espèce, mais également des conditions du climat et des activités humaines. 1.4- Statut actuel des populations des deux échassiers étudiés dans la région de Batna1.4.1- Statut actuel des populations de la Cigogne blancheNous n'avons pas fait une estimation de la taille de la population de la Cigogne blanche dans la région de Batna. Très récemment, un recensement de nids de C. ciconia a été réalisé dans la wilaya par DJADDOU et BADA (2006) qui l'ont estimé à 490 nids avec 7 nids non occupés. Ces nids sont répartis sur un ensemble de 15 daïras visités couvrant un total de 45 localités. Ainsi, la plus importante colonie rencontrée dans le territoire de la wilaya de Batna est celle d'El Madher avec 50 nids occupés et 3 nids non occupés, ce qui représente 10,8 % de la population de la wilaya. En deuxième position se classe la population de la ville de Batna avec 35 nids occupés et un nid non occupé et en troisième lieu vient la colonie d'Ain Touta avec 33 nids occupés (DJADDOU et BADA, 2006). Selon ces derniers auteurs, l'occupation spatiale des colonies montre que la Cigogne blanche colonise beaucoup plus la partie Nord-est de la wilaya (plaines d'El Madher et d'Ain Touta) et à un degré moindre la région nord (plaine de Belezma), par le fait que ces régions offriraient les meilleurs milieux d'alimentation habituellement fréquentés par l'espèce. Les recensements, aussi exhaustifs, réalisés en en collaboration entre le département de Biologie (Association scientifique Biologie, Biodiversité et Durabilité) et la conservation des forêts de la wilaya de Batna, montrent une évolution des effectifs ces dernières années, qui sont estimés à 593 en 2007 nids et 670 nids en 2008. 1.4.2- Statut actuel des populations du Héron garde-boeufsLes résultats de l'enquête locale menée dans une vingtaine de lieux dans le territoire de la wilaya de Batna nous ont révélé d'importantes informations sur la date d'installation de B. ibis, sur sa bio- écologie et sa relation avec l'homme dans cette région. En effet, cette enquête a montré que l'espèce est à installation récente dans cette région et les dates de première observation vont, selon les régions, de 1995 à 2005. Les premières villes investies par l'espèce sont Ain Touta en l'an 1995, Ras Layoun en l'an 2000, Merouana, El Madher et Tazoult en l'an 2002 et N' Gaous en l'an 2003. La récente installation du garde-boeufs expliquerait l'absence d'une dénomination locale de l'espèce, où nous avons enregistré un pourcentage important de réponses (85,7 %) qu'il n'a pas d'appellation. Certains ont donné l'appellation de « l'oiseau de la mer » « Farkh Labhar » tout en racontant qu'ils ont l'habitude de le voir sur les régions littorales, ou « la poule d'eau » « Djaj El Ma » parce qu'ils l'ont observé fréquentant des milieux humides de la région (Oueds, barrages...). Concernant son statut phénologique, les personnes interrogées habitant près de ses colonies de reproduction ont rapporté qu'il est sédentaire nicheur, alors que celles habitant loin de ses colonies ont rapporté qu'il est sédentaire sans préciser sa nidification. Plus de 54 % de gens questionnés, rapportent qu'il est observé durant toute l'année, confirmant ainsi son statut d'espèce sédentaire. La première année de nidification de B. ibis dans la région de Batna est notée en l'an 1995 à Ain Touta (RIGHI Y., cadre au Parc National de Belezma, com. pers.). A partir de cette année, l'expansion du garde-boeufs dans la région d'étude n'a pas cessé d'augmenter. Le nombre de dortoirs est actuellement estimé à 16 et le nombre de colonies à 10. Ceci démontre l'augmentation importante des effectifs nicheurs durant ces dernières années et l'implantation de l'espèce en tant que sédentaire nicheuse. Concernant la relation de l'oiseau avec l'homme, les personnes interrogées dans cette enquête semblent ne pas donner une grande importance à l'espèce, étant donné que la moitié d'entre eux (48,8 %) rapportent qu'il n'est ni utile ni nuisible ou peut être auxiliaire à l'agriculture. Il est toutefois considéré importun dans les dortoirs et colonies parce que c'est une espèce bruyante et qui a des fientes nauséabondes et acides qui dessèchent le support végétal qu'il utilise. Cette enquête a révélé aussi que le Héron garde-boeufs n'est pas dissuadé ni chassé ou consommé dans la région de Batna. Ceci est contrairement à ce qu'a révélé une enquête similaire réalisée à Béjaia par SI BACHIR (2007), qui a conclu que le garde-boeufs dans la Kabylie de la Soummam, connu depuis les années quarante, a une multitude de nominations : « Agtit El Male » « oiseau du bétail », « Tayazite El Male » « poule du bétail ».... Cette étude a montré également qu'il pourrait porter préjudice à la qualité du sol par le prélèvement excessif de vers de terre ou même causer des dégâts aux cultures de Cucurbitacées (pastèque et melon) en les picorant lors de la capture d'insectes. En effet, ce dernier auteur a confirmé l'utilisation de moyens de dissuasion (pièges divers, coups de feu, tir de pierres...) par les paysans de la région pour l'éloigner de leurs champs. A l'image de son expansion géographique mondiale (KUSHLAN et HANCOCK, 2005), en Algérie, l'expansion de l'espèce est signalée à partir des années 1992-1993 ; l'implantation de nouvelles colonies est notée en Kabylie, sur les hauts plateaux et en descendant jusqu'aux zones semis arides (Sétif, Batna) et mêmes aux régions arides (Biskra) (MOALI et ISENMANN, 1993 ; MOALI, 1999 ; ISENMANN et MOALI, 2000 ; SI BACHIR, 2007). 1.5- Comparaison de la phénologie des deux espècesLa comparaison des périodes phénologiques des deux échassiers, dans la colonie plurispécifique de Merouana, montre que les deux espèces mènent leurs activités de ponte, de couvaison et élevage des jeunes au cours de périodes parallèles. Ceci signifie qu'il pourrait y avoir un risque de compétition entre les deux espèces « compétition interspécifique » en plus de la « compétition intraspécifique » observée chez les garde-boeufs, en particulier lors de la période d'élevage des jeunes. Dans la zone d'El Madher, la comparaison des stades phénologiques des deux modèles biologiques étudiés, montre un décalage de près d'un mois entre les deux espèces, à partir des dates de pontes. La période d'élevage de C. ciconia s'est déroulée en 63 jours, du 23 avril 2007 (première observation d'éclosion) au 25 juin 2007 (première observation d'envol), c'est-à-dire pendant 9 semaines. En revanche, la période d'élevage du Héron gardeboeufs s'est étalée sur près de 45 jours, de la fin mai 2007 (première observation d'éclosions) à la mi-juillet 2007 (première observation d'envols). A cet effet, la période de chevauchement entre les périodes d'élevage des deux échassiers est de près d'un mois, de la fin mai 2007 jusqu'au 25 juin 2007, durée pendant laquelle pourrait se produire un risque de compétition interspécifique. Toutefois, dans les colonies étudiées, des manifestations évidentes de comportement de compétition entre les deux échassiers n'ont pas été observées. Contrairement à ce qu'a été noté dans la colonie plurispécifique située au centre ville d'Ain Touta, où le Héron gardeboeufs et la Cigogne blanche montrent de fréquentes scènes de querelles sur les sites communs des nids installés sur de hauts Pin d'Alep (RIGHI Y., cadre au Parc National de Belezma, com. pers.). Ceci ne serait-il pas le début d'une véritable compétition interspécifique. 2- Composition taxinomique et variation spatio-temporelle des disponibilités alimentaires2.1- Composition taxinomique du peuplement de proiesL'inventaire faunistique réalisé dans les trois stations d'échantillonnage, nous a permis de recenser une large variété de proies invertébrées (234 espèces) dont la classe des insectes est la plus dominante (223 espèces) tant en nombre d'espèces qu'en terme d'abondance en nombre. Dans l'espace et dans le temps, les insectes sont représentés essentiellement par les coléoptères, les hyménoptères et les diptères, suivis par les arachnides (les aranéides). Nos résultats sont comparables avec ceux obtenus à Tizi-Ouzou, à la basse vallée de la Soummam et dans l'Algérois. BENTAMER (1998), en étudiant les disponibilités alimentaires de la Cigogne blanche et du Héron garde-boeufs dans la région de Tizi-Ouzou, a noté la dominance des insectes avec un total de 60 espèces inventoriées, dont 29 espèces de coléoptères. De même, SALMI (2001), dans la basse vallée de la Soummam, a mentionné de forts taux d'insectes dans les disponibilités alimentaires du Héron garde-boeufs (183 espèces) avec une prédominance des coléoptères (101 espèces). Dans la Mitidja orientale, SETBEL (2003), a recensé un total de 236 espèces animales réparties en 7 classes, parmi lesquelles, les insectes prédominent avec 200 espèces et les coléoptères étant les plus fréquents avec 116 espèces. Bien que le dispositif d'échantillonnage n'a touché qu'une partie de la faune invertébrée de la région, nous pouvons considérer que cette dernière présente une assez bonne diversité tant invertébrée que vertébrée nécessaire à l'alimentation et au bon développement des deux échassiers étudiés. 2.2- Variation spatio-temporelle des peuplements de proies potentilles recensées dans les gagnages2.2.1- Variation spatiale des peuplements de proies potentillesL'étude de la répartition spatiale des invertébrés recensés, nous a permis de constater que l'abondance relative des invertébrés capturés ne montre pas une grande variation entre les trois milieux d'échantillonnage. Nous avons recensé 1.943 individus dans la culture basse, 1.748 dans le milieu humide et 1.695 dans la pelouse naturelle. La richesse totale la plus élevée est notée dans la pelouse naturelle, suivie par le milieu humide et la culture basse. Cependant, l'indice de diversité de SHANNON montre que le milieu humide abrite les peuplements de proies les plus diversifiés. La présence de l'eau ainsi que la diversité du couvert végétal de la zone humide, caractérisée par une couverture permanente de Medicago saliva, Malva sylvestris, Taraxacum sp., Plantago coranopus, Phalaris minor, Carex hirta, Athemis pedunculata, Melilotus sp., Buta sp., et Calendula arvens, et du couvert végétal de la pelouse naturelle échantillonnée, caractérisée par une végétation herbacée dominée par : Althaea sp., Paronychia argentea, Erodium guttatum, Anthemis montana, Hordeum annua, Adonis annua, Murrubium alyssum, Urtica dioica et Carlina acaulis, ainsi que la présence du bétail dans ces deux milieux pâturés, favoriseraient un bon développement des coléoptères, des hyménoptères et de plusieurs autres catégories d'invertébrés. En deuxième lieu, le type de végétation dans les milieux cultivés échantillonnés (céréales, cultures fourragères et parfois labours) et l'intensité du travail du sol et son enrichissement en fumier expliqueraient l'abondance des coléoptères, des hyménoptères et des orthoptères dans ces milieux. 2.2.2- Variation temporelle des peuplements de proies potentillesC'est pendant les saisons printanière et estivale que nous avons enregistré les plus grands effectifs d'invertébrés, notamment des coléoptères, des hyménoptères, des aranéides et des diptères. Ainsi, les effectifs les plus élevés sont enregistrés au printemps avec 2.399 individus suivi par l'été avec 1.305 individus, alors que les valeurs les plus élevées de la richesse totale sont notées au printemps avec 142 espèces et en automne avec 121 espèces. De même, l'indice de SHANNON révèle une diversité plus importante en automne avec 5,5 bits suivi par le printemps avec 5,14 bits. L'apparition d'invertébrés en une période et leur absence en une autre s'expliquerait par leur phénologie spécifique qui est liée aux conditions écologiques du milieu, notamment les conditions climatiques et le couvert végétal. Ainsi, en hiver nous notons l'absence de 8 ordres par le fait que la plupart des invertébrés entrent en diapause. Les ordres présents en cette saison ont un échantillonnage accidentel à très accidentel. Généralement, nos résultats sont comparables à ceux obtenus dans différentes régions d'Algérie. BENTAMER (1998), dans un terrain cultivé de blé (Tizi-Ouzou), a noté un taux élevé de coléoptères aux mois de janvier et d'avril, suivis par les orthoptères, et des taux assez fréquents de crustacés, arachnides et hyménoptères. Pour ce même auteur, ce sont les orthoptères qui sont notés avec des taux élevés dans la prairie échantillonnée, surtout aux mois d'octobre, de novembre et de juillet. BOUKHEMZA (2000), signale également un maximum d'abondance des orthoptères en été et au début de l'automne. SALMI (2001), dans la basse vallée de la Soummam, mentionne un taux élevé de coléoptères qui atteignent 73,1 % en avril 1999 dans une friche contre 21,3 % dans une prairie. Pour SI BACHIR (2007), dans la région de la Kabylie de la Soummam, les effectifs d'invertébrés recensés lors de la période printanière et estivale sont plus importants que ceux enregistrés en dehors de ces périodes. 3- Modalités d'exploitation des milieux de gagnage3.1- Exploitation des milieux de gagnage par la Cigogne blancheDans la région de Batna, C. ciconia s'alimente essentiellement dans des milieux ouverts à végétation basse comme les prairies naturelles ou aménagées (32,53 %) et les cultures basses (28,31 %), telles les cultures céréalières et fourragères à leurs premiers stades de développement et ainsi qu'après les moissons (Fig. 29A). Elle fréquente aussi les friches (7,83 %), qui sont des milieux naturels caractérisés par une végétation arbustive ouverte et les labours même s'ils sont signalés en moindre importance (3,61 %) (Fig. 29A). Nos résultats rejoignent ceux de BOUKHEMZA (2000) en Kabylie, de LATUS et KUJAWA (2005) en Allemagne et ceux de PINOWSKA et PINOWSKI (1989) et RACHEL (2006) en Pologne. Tous ces auteurs rapportent que la Cigogne blanche a une préférence aux prairies, aux milieux cultivés à végétation basse, les milieux pâturés, les jachères, les terrains labourés et les mares temporaires. Nos résultats sont également comparables à ceux de SBIKI (2008) dans la région de Tébessa, où la cigogne fréquente essentiellement les friches (48,89 %), les immondices (18,01 %), les cultures basses (12,86 %) et les milieux fauchés (12,01 %). En plus des milieux naturels fréquentés par C. ciconia, nous l'avons observé plusieurs fois s'alimentant dans des déchets d'aviculteurs (8,43 %) (Fig. 29A), notamment dans la zone de Merouana caractérisée par le développement de l'aviculture, et même dans des dépotoirs en dehors des itinéraires échantillonnés, dans la ville d'Arris. La fréquentation des dépotoirs et des décharges publiques est également signalée en Espagne par GARRIDO et FERNÁNDEZ-CRUZ (2003), PERIS (2003) et BLÁZQUEZ et al. (2006). (A) Ciconia ciconia
18,07 % 1,20 % 8,43 % 28,31 % 3,61 % 32,53 % 7,83 % (B)
Bubulcus ibis
3,90 % 0,50 % 4,25 % 12,11 % 5,59 % 2,62 % 15,79 % 5,67 % 19,90 % 29,67 % Bordures d'eaux usées Prairies Friches Labours Cultures basses Immondices Prairies inondées Dépôts de fumier Déchets d'aviculteurs Milieux fauchés Figure 29- Taux globaux de la fréquentation des
milieux de gagnage par la Cigogne La Cigogne blanche fréquente les cultures basses tout au long de ses périodes phénologiques (Fig. 30A). Elle s'alimente dans les friches surtout pendant la période de préreproduction et dans les déchets d'aviculteurs surtout en périodes de ponte et couvaison et d'élevage des jeunes. Les milieux fauchés sont particulièrement fréquentés pendant la période d'élevage et la période de post-reproduction. C'est pendant la période d'élevage des jeunes cigogneaux, que la Cigogne blanche fréquente une plus grande variété de milieux (les bordures d'eaux usées et les labours en plus des milieux cités plus hauts) (Fig. 30A). 3.2- Exploitation des milieux de gagnage par le Héron garde-boeufsQuant à B. ibis, cet échassier montre une utilisation plus diversifiés des milieux de gagnage. Nous en avons compté 10 milieux différents (Fig. 29B). Cependant, tout comme la cigogne, il a une grande préférence aux prairies (29,67 %), aux cultures basses (15,79 %), aux milieux fauchés (12,11 %) mais aussi aux labours (19,9 %). Il utilise ainsi en moindre importance les bordures d'eaux usées (5,59 %), les prairies inondées (4,25 %) et les friches (2,62 %) (Fig. 29B). Ceci dénote du fait que l'espèce utilise de plus en plus des milieux terrestres dans son alimentation. Selon SBIKI (2008), dans la région de Tébessa, le Héron garde-boeufs fréquente principalement les friches (27,38 %), les immondices (22,64 %), les cultures basses (17,41 %) et les labours (16,54 %). Par contre, les prairies, les milieux fauchés et les mares temporaires sont les moins utilisés. Les résultats de SBIKI (2008) rejoignent ceux de FRANCHIMONT (1986b), qui indique que les labours et les immondices fournissent la plus grande partie de nourriture destinée aux garde-boeufs dans la région d'Ouezzane au Maroc. Par ailleurs, en plus de ces milieux, le Héron garde-boeufs s'alimente dans différents types de déchets, tels les déchets des fermiers (fumier et déchets d'aviculteurs) et les immondices (dépotoirs de rejets ménagers). Cette observation est notée surtout dans la zone de Merouana. Nos résultats sont comparables avec ceux de FRANCHIMONT (1986b) au Maroc, BOUKHEMZA (2000) en Kabylie et SI BACHIR (2007) à Béjaia. Néanmoins, BREDIN (1883-1984) et LOMBARDINI et al. (2001) signalent qu'en Camargue le Héron garde-boeufs utilise en plus de ces milieux, les milieux halomorphes, les ronciers, les rizières et les marais d'eau douce ou salée. La fréquentation des dépôts d'ordures par le garde-boeufs n'a pas été observée en Camargue (BREDIN, 1984), contrairement à nos résultats, et à ceux obtenus par RENCUREL (1972), DEAN (1978) et FRANCHIMONT (1986b) au Maroc, SI BACHIR (2007) à Béjaia et (A) Ciconia ciconia
(B)
Pr. Rep. P. C. Elev. Pt. Rep. Bubulcus ibis
Hiver. Pr. Rep. P. C. Elev. Pt. Rep. Bordures d'eaux usées Prairies Friches Labours Cultures basses Immondices Prairies inondées Dépôts de fumier Déchets d'aviculteurs Milieux fauchés Figure 30- Variation des taux globaux de la
fréquentation des milieux de gagnage selon SBIKI (2008) à Tébessa. Selon SALMI (2001), le fumier et les rejets des poulaillers constituent une ressource renouvelable en larves d'insectes notamment de Scarabaeidae. Ce dernier auteur a déterminé aussi un fort pourcentage de larves de Scarabaeidae et de Rhizotrogus sp. dans les pelotes de rejection du Héron garde-boeufs, tout en mentionnant qu'ils ne sont jamais échantillonnées dans les milieux de gagnages. Selon ELKINS (2001), dans les habitats les plus exposés (milieux ouverts), les oiseaux bénéficient considérablement, pendant l'hiver, des ressources alimentaires crées par l'homme : cultures d'hiver, chaumes, champs d'épandage, déchets de nourriture de toutes formes. Le Héron garde-boeufs est observé dans les prairies pendant toutes les périodes phénologiques (Fig. 30B). Il fréquente les cultures basses pendant les périodes d'hivernage et de pré-reproduction et en période d'élevage des jeunes. Il s'alimente également dans les labours presque pendant toutes les périodes phénologiques. Mais, c'est pendant la période de ponte et couvaison et la période d'élevage des jeunes héronneaux, que l'espèce utilise des milieux plus diversifiés. Pendant ces deux périodes, le Héron garde-boeufs fréquente, en plus des prairies, des cultures basses, des labours et des milieux fauchés, les déchets d'aviculteurs, les prairies inondées, les immondices, les friches et les bordures d'eaux usées (Fig. 30B). La diversité des milieux exploités par le Héron garde-boeufs est en relation avec l'opportunisme de cette espèce qui fait varier son alimentation selon les disponibilités des proies dans les milieux de gagnage d'une région donnée et suivant les besoins alimentaires liés à la phénologie de l'espèce (DOUMANDJI et al., 1992-1993 ; BOUKHEMZA , 2000 ; SI BACHIR et al., 2001). Ceci serait considéré parmi les facteurs de l'expansion de cet échassier. 3.3- Association manifestée par les deux échassiers3.3.1- Association passiveCertains individus des deux échassiers étudiés s'associent d'une façon passive entre eux mêmes (Cigognes blanches et Hérons garde-boeufs), et/ou à d'autres espèces aviaires. Nous citons en particulier le grand Corbeau (Corvus corax), les Moineaux (Parus sp.) et les Pigeons (Columba sp.). Cette observation est notée presque pendant toute la période d'étude notamment en hiver et dans les déchets d'aviculteurs et dans les labours. 3.3.2- Association activeA l'opposé de l'association passive, seuls les garde-boeufs s'associent d'une façon active à différents types de bétail tels les bovins et les ovins, ainsi qu'aux engins agricoles (tracteurs en particulier) (Fig. 31). Ces types d'association sont particulièrement notées dans les prairies, les friches, les labours, les milieux fauchés et dans les cultures basses. Ce phénomène est également signalé dans la totalité de l'aire de répartition du Héron garde-boeufs par plusieurs auteurs (SKEAD, 1956 ; BLAKER, 1969 ; BREDIN, 1983 ; FRANCHIMONT, 1986b ; BOUKHEMZA, 2000 ; SI BACHIR, 2007). D'après HEATWOLE (1964) in BLAKER (1971) et WHITFIELD et WALKER (1999), cette technique permet au garde-boeufs de s'alimenter de 50 % de plus de nourriture que les autres oiseaux, pour un effort moindre. 3.4- Comparaison des modalités d'exploitation des milieux de gagnage par les deux espècesLes résultats que nous avons obtenus montrent que C. ciconia et B. ibis utilisent principalement les mêmes milieux d'alimentation : les prairies, les cultures basses, les friches et les labours. Cependant, la variation des taux de fréquentation des gagnages selon les périodes phénologiques montre une nette différence entre les deux espèces. Le garde-boeufs est une espèce à installation récente dans la région de Batna, à partir de l'an 1995. Avant son arrivée, la Cigogne blanche jouissait seule de ces milieux d'alimentation. Ceci nous amène à suggérer un risque de pression entre les deux échassiers lors de l'exploitation de ces milieux d'alimentation. Toutefois, nos résultats indiquent également que B. ibis s'alimente d'une manière énergique par la diversification des milieux d'alimentation qu'il exploite et par son association au bétail et aux engins agricoles qui mettent à jours plusieurs types de proies. Cette stratégie peut lui procurer des ressources alimentaires supplémentaires ce qui minimiserait sensiblement le risque de compétition entre les deux espèces dans les milieux d'alimentation. Chapitre V- Discussions
(A) (B) (C) Figure 31- Photos illustrant une association active du Héron garde-boeufs à un bovin (A) (Merouana, 23 janvier 2007), à des ovins (B) (El Madher, 26 septembre 2007) et à une machine agricole (C) (Merouana, 28 novembre 2006) dans la région de Batna (Photos personnelles). 4- Caractérisation qualitative et quantitative du régime alimentaire des deux espèces4.1- Caractéristiques des pelotes de rejection des deux échassiersLes pelotes de la Cigogne blanche ont une taille de 50,91 mm × 37,63 mm et pèsent en moyenne 9,7 #177; 4,6 g en poids sec. Alors que celles du Héron garde-boeufs ont une taille de 36,67 mm × 21,27 mm et un poids sec de 1,8 #177; 1 g. La taille des pelotes des deux échassiers varie d'une façon très importante d'une année à une autre et d'un mois à un autre dans la même région, et d'une région à une autre (Tab. XXVII). D'une manière générale, nos résultats sont comparables avec ceux obtenus dans différentes régions du pays. Tableau XXVII- Données comparatives de la taille et du poids des pelotes de rejection de la Cigogne blanche et du Héron garde-boeufs dans quelques régions d'Algérie.
Les résultats obtenus par SCHIERER (1967) en France (60 mm × 39mm), montrent que les pelotes de la cigogne sont un peu plus volumineuses, alors que ceux de MUZINIC et RASAJSKI (1992) aux Balkans sont proches des nôtres (50 mm × 40 mm). Quant au garde-boeufs, HAMADACHE (1991), à Draâ-El-Mizan (Kabylie) rapporte des mensurations de 45 mm x 15 mm pour la plus grosse pelote et 25 mm x 10 mm pour la plus petite. Par ailleurs, en comparant nos résultats à ceux de BREDIN (1983) en Camargue (23,8 mm x 13,8 mm), les pelotes de B. ibis dans la région de Batna paraissent plus grosses. Cette variation des dimensions des pelotes serait en relation avec le nombre de proies, leurs natures ainsi que leurs biomasses. 4.2- Analyse globale de la composition du régime alimentaire des deux échassiers4.2.1- Analyse de la composition en proies des pelotes de la Cigogne blancheLa décortication de 96 pelotes de la Cigogne blanche nous a permis d'enregistrer un minimum de 7 proies dans une pelote contenant 2 Pimelia sp., 5 individus de Curculionidae sp. ind. avec la présence de plumes de poulet domestique (Galus domesticus). Un maximum de 1.022 proies est noté dans une pelote contenant 786 Caelifera sp. ind., 197 Ensifera sp. ind., 15 Geotrupes sp., 10 Gryllus bimaculatus, 6 Coleoptera sp. ind., 3 Acinopus sp., 3 Tenebroinidae sp. ind., 1 Scarabaeus sacer et 1 Helicidae sp. ind.. Le nombre moyen de proies par pelote est de 165,4 #177; 139 proies. DAJOZ (2002), signale dans le contenu de 3 pelotes de la cigogne collectées durant la migration de printemps dans la région de Sede Boker en Israël, la composition suivante, en nombre d'individus : Adesmia dilatata : 356 ; Adesmia abbreviata : 242 ; Pimelia boehmi : 190 ; Pimelia sp. : 150 ; Pimelia grandis : 35 ; Sepidium tricuspidatum : 170 ; Erodius edomitus : 86 ; autres Tenebrionidae : 51 ; autres Arthropodes : 182 (Total 1.462). Toutefois, nos résultats dépassent largement ceux obtenus dans différentes régions du pays. Sur un lot de 350 pelotes de la cigogne BOUKHEMZA (2000), a enregistré un minimum de 4 proies, un maximum de 219 proies et une moyenne de 46,2 proies par pelote. Pour ZENNOUCHE (2002), 69 pelotes de la cigogne contiennent d'une seule proie à 399 proies et en moyenne 79,7 proies par pelote. En outre, une étude pareille à la nôtre, réalisée au cours de la même année à Tébessa, par SBIKI (2008), révèle que les pelotes de la Cigogne blanche de cette région contiennent presque 6 fois moins de proies que celles des cigognes de la région de Batna. En fait, SBIKI (2008), a mentionné 2.718 proies dans 127 pelotes de C. ciconia (21,4 proies par pelote). Cette variation importante du nombre de proies dans les pelotes, s'expliquerait non seulement par le besoin alimentaire de l'espèce, qui lui-même dépend de ses stades phénologiques, mais aussi par la richesse des milieux de gagnage et la phénologie des proies. Cependant, toutes les études faites en Algérie et dans d'autre régions du monde s'accordent à dire que C. ciconia est un échassier prédateur entomophage (SCHIERER, 1962- 1967 ; LAZARO, 1986 ; LAZARO et FERNÁNDEZ, 1991 ; PINOWSKA et PINOWSKI, 1989 ; PINOWSKI et al., 1991). La part des insectes dans son régime alimentaire dans la région de Batna est de 99,23 % de l'effectif total des proies consommées. A titre indicatif, ANTCZAK et al. (2002), mentionnent 83 % d'insectes, en Pologne. En Algérie, nos résultats sont conformes à ceux obtenus par BOUKHEMZA (2000), ZENNOUCHE (2002), DJADDOU et BADA (2006) et SBIKI (2008), qui mentionnent respectivement des taux de 94 % ; 94,3 % ; 97,3 % et 98,9 % d'insectes. Dans cette étude nous n'avons pas fait l'évaluation en biomasse des proies consommées. Néanmoins, nous jugeons utile de citer des résultats de quelques auteurs qui montrent une différence dans l'abondance en biomasse par rapport à l'abondance en nombre. BOUKHEMZA (2000), indique que la cigogne consomme, en termes de biomasse, 74,2 % d'insectes, 7,3 % d'arachnides et 17,9 % de vertébrés. ANTCZAK et al. (2002), notent 58 % de petits mammifères, 22 % d'insectes et 11,5 % de vers de terre. Dans la région de Batna, la part des vertébrés entrant dans la composition du régime alimentaire de la Cigogne blanche est de 0,13 % en termes d'abondance en nombre. Ces vertébrés, représentés principalement par des reptiles (Lacertidae) et des micromammifères (Rodentia), ne sont retrouvés que dans les pelotes collectées de la zone d'El Madher (15,63 % de fréquence d'occurrence pour les reptiles et 6,25 % pour les mammifères). Ce qui dénote d'une consommation sporadique des vertébrés. 4.2.2- Analyse de la composition en proies des pelotes du Héron garde-boeufsPour le Héron garde-boeufs, la décortication de 110 pelotes nous a permis d'enregistrer un minimum de 3 proies dans une pelote contenant 1 Forficula auricularia et 2 individus de Buprestidae sp. ind. en plus de plumes, du plastique et des pierres. Un maximum de 586 proies dans une pelote contenant 454 Formicidae sp. ind., 53 Messor barbara, 26 Anisolabis mauretanicus, 16 Caelifera sp. ind., 13 Coleoptera sp. ind., 6 Gryllus bimaculatus, 4 Carabidae sp. ind., 3 Ensifera sp. ind., 3 Chlaenius sp., 2 Ophonus sp., 2 Araneidae sp. ind. et un seul individu pour Labidura riparia, Mantidae sp. ind., Acinopus picipes, Chrysomelidae sp. ind.. Globalement, nous avons noté une moyenne de 85,3 #177; 74,5 proies par pelote. Le nombre de proies contenu dans une pelote de garde-boeufs de la région de Batna est de loin très important par rapport à celui noté dans plusieurs régions d'Algérie. En Kabylie, BOUKHEMZA (2000), dans un lot de 150 pelotes de garde-boeufs, mentionne un minimum de 4 proies, un maximum de 178 proies et une moyenne de 40 proies par pelote. DOUMANDJI et al. (1993), dans la région de Chlef, BOUKHEMZA et al. (1995), BOUKHEMZA et al. (1997) et BENTAMER (1998) dans la région de Tizi-Ouzou, notent respectivement des moyennes de 23,4 proies, 46,2 proies, 44,5 proies et 39,7 proies par pelote. En comparant nos résultats avec ceux de SBIKI (2008), nous constatons également que les pelotes du Héron garde-boeufs de la région de Batna contiennent près de 12 fois plus de proies que celles étudiées à Tébessa. SBIKI (2008), mentionne dans 124 pelotes, une moyenne de 9,5 proies par pelote. Parmi les proies consommées dans la région de Batna, nous avons noté un taux allant jusqu'à 98,37 % d'insectes. Les vertébrés, quant à eux, composent seulement 0,74 % en abondance en nombre du régime alimentaire du héron garde-boeufs, dont les mammifères (petits rongeurs) constituent 0,42 % d'abondance et 33,64 % de fréquence d'occurrence. Toutes les études faites en Algérie et dans le monde s'accordent à dire que B. ibis est un échassier prédateur entomophage. En Algérie, DOUMANDJI et al. (1992-1993), mentionnent des taux de 99,8 % et de 96,8 %, respectivement, à Draa El Mizan et à Chlef. FELLAG (1995), BENTAMER (1998), BOUKHEMZA et al. (2000) et SETBEL et al. (2004), notent plus de 95 % dans les régions de la Kabylie du Sébaou, à Boudouaou et dans la Kabylie de la Soummam. De même, SI BACHIR (2007) et SBIKI (2008), notent également que les insectes représentent les proies les plus abondantes dans la composition du régime alimentaire de l'espèce avec des taux respectifs de 92,9 % et 97,96 % dans les régions de Béjaia et Tébessa. SIEGFRIED (1971c), en Afrique du Sud, signale qu'environ 80 % du régime alimentaire du garde-boeufs est composé d'insectes. FOGARTY et HETRICK (1973), notent 90 % en Floride (Etats-Unis) ; RUIZ et JOVER (1981), rapportent un taux de 68,4 % dans le delta d'El Ebro (Espagne). Il est utile de signaler que la fréquence d'abondance en biomasse des proies ne donne pas des valeurs aussi importantes pour les insectes. BOUKHEMZA (2000), signale que 57,2 % seulement de la biomasse ingérée par le garde-boeufs est composée d'insectes et 42,5 % de vertébrés. 4.3- Analyse de la structure du régime alimentaire des deux échassiers4.3.1- Analyse de la structure globale du régime alimentaire de la Cigogne blancheCe sont les orthoptères avec 50,75 % suivis par les coléoptères avec 40,84 % et les dermaptères avec 6,95 % qui dominent le régime alimentaire de la Cigogne blanche dans la région de Batna. L'ordre des orthoptères est représenté par deux groupes non identifiés, les Caelifera ind. (37,08 %) et les Ensifera ind. (11,47 %). Les coléoptères sont représentés par les familles des Scarabaeidae (12,10 %), des Silphidae (6,65 %), des Tenebrionidae (5,43 %), des Carabidae (4,38 %) et des Harpalidae (4,46 %). Alors que les dermaptères sont représentés essentiellement par la famille des Labiduridae (6,86 %). Ces résultats sont relativement similaires à ceux soulignés par différents auteurs, notamment en Algérie (BOUKHEMZA, 2000 ; ZENNOUCHE, 2002 ; DJADDOU et BADA, 2006 ; SBIKI, 2008). A titre d'exemple, ZENNOUCHE (2002), dans la Kabylie de la Soummam, signale l'importance des Carabidae, des Silphidae, des Scarabaeidae et des Tenebrionidae parmi les coléoptères ; des Gryllidae, des Acrididae et des Tettigonidae parmi les orthoptères et les familles des Labiduridae et des Formicidae des ordres des dermaptères et hyménoptères dans le régime alimentaire de C. ciconia. L'importance des coléoptères et des orthoptères dans le régime alimentaire de la cigogne, est signalée par plusieurs auteurs en Europe, nous citons SCHIERER (1962-1967) en France, SKOV (1991a) au Danemark, MUZINIC et RASAJSKI (1992) aux Balkans. 4.3.2- Analyse de la structure globale du régime alimentaire du Héron gardeboeufsLe bol alimentaire du Héron garde-boeufs dans la région de Batna est composé principalement de 39 % de coléoptères, 32,47 % d'orthoptères, 14,56 % d'hyménoptères et 11,12 % de dermaptères. L'ordre des coléoptères est représenté par les familles des Carabidae, des Harpalidae, des Pterostrichidae et des Scarabaeidae. Les orthoptères sont représentés principalement par les Caelifera ind., les Acrididae et en moindre importance par les Gryllidae. Les hyménoptères sont essentiellement représentés par la famille des Formicidae (14,52 %) et les dermaptères par la famille des Labiduridae (10,5 %). Nos résultats sont sensiblement proches de ceux de BOUKHEMZA (2000), de SALMI (2001), de SETBEL et al. (2004), de SI BACHIR (2007) et de SBIKI (2008). SETBEL et al. (2004) mentionnent la dominance des coléoptères représentés par les Scarabaeidae, les Carabidae et les Dermestidae, des hyménoptères représentés par les Formicidae et des orthoptères représentés par les Acrididae et les Gryllidae à Boudouaou. Les coléoptères et les orthoptères constituent une part importante du régime alimentaire du garde-boeufs même dans d'autres régions du monde (HAFNER, 1977 ; VOISIN, 1991). 4.4- Variation spatio-temporelle du régime alimentaire des deux échassiers4.4.1- Variation spatio-temporelle du régime alimentaire de la Cigogne blanche> Variation du régime alimentaire de la Cigogne blanche entre les deux zones d'étudeLe régime alimentaire de la Cigogne blanche varie sensiblement entre les deux zones d'étude. A d'El Madher, ce sont les coléoptères qui dominent le régime alimentaire de l'espèce avec une abondance de 56,21 % suivis par les orthoptères (30 %) et les dermaptères (11,67 %). A Merouana, les orthoptères viennent en premier lieu avec 75,4 %, suivis par les coléoptères (22,5 %) et les dermaptères (1,31 %). L'indice de similitude de SORENSON calculé entre les deux zones d'étude est de 75,59. Ceci signifie qu'il existe une grande similitude entre les deux stations. Il est à signaler que la plupart des auteurs que nous avons consulté ont fait leurs études dans un seul lieu d'échantillonnage. Néanmoins, nos résultats concordent avec ceux de BOUKHEMZA (2000), dans la région de la Kabylie du Sébaou (Tizi-Ouzou), qui a mis en évidence la dominance des coléoptères, des orthoptères et des dermaptères tout en signalant des différences dans leur consommation entre trois localités différentes. A titre d'exemple, en 1992 les cigognes de Boukhalfa et de Tdmaït ont consommé d'importantes quantités de coléoptères tandis que celles de Drâa Ben Khedda ont préféré les orthoptères. D'après ce dernier auteur, ces variations reflètent les différences de l'entomofaune locale. > Variation temporelle En plus de la variation constatée entre localités, les principaux ordres composant le régime alimentaire de la cigogne fluctuent d'un mois à un autre dans la même localité. A El Madher, les coléoptères enregistrant le taux le plus élevé, sont surtout plus dominants aux mois de mars, avril et mai. Ils sont remplacés par les orthoptères durant les mois de juin et de juillet. Alors qu'à Merouana, les orthoptères, dont les deux sous ordres Caelifera et Ensifera, sont présents avec des fréquences presque semblables, dominent le régime alimentaire de C. ciconia durant tous les mois d'étude dépassant 51 % de l'effectif total avec des taux élevés aux mois de février et de juillet. Les dermaptères, représentés essentiellement par l'espèce Anisolabis mauritanicus noté avec une occurrence très élevée (76 %), atteignent leur maximum aux mois de mars et de mai à El Madher et au mois de juin à Merouana. Ces résultats sont en relation avec les résultats des disponibilités alimentaires, dans lesquels nous avons noté un taux élevé de coléoptères, plus de 30 % des invertébrés échantillonnés, durant les saisons hivernale, estivale et automnale, et un taux faible en été avec 11 % seulement. Contrairement aux orthoptères, qui notent un taux plus élevé en été (3,21 %) et en automne (1,99 %), un taux faible au printemps et une absence en hiver. BOUKHEMZA (2000) a noté, sur deux années successives 1992 et 1993, une augmentation considérable dans la consommation des coléoptères par la cigogne depuis son arrivée, pour atteindre un maximum au mois de mai. Les coléoptères sont relayés, par la suite, par les orthoptères, notamment au mois de juillet. Concernant les dermaptères, ce dernier auteur a mentionné encore des résultats similaires aux nôtres. Cet ordre est noté avec un taux élevé au mois de mars, en 1992, et aux mois d'avril et de juin en 1995. 4.4.2- Variation spatio-temporelle du régime alimentaire du Héron garde-boeufs> Variation du régime alimentaire du Héron garde-boeufs entre les deux zones d'étudeDe même que pour la Cigogne blanche, les coléoptères viennent en tête du régime alimentaire du Héron garde-boeufs dans la zone d'El Madher avec 36,97 %, mais leur taux n'est pas trop loin de celui des orthoptères (27,29 %), suivis par les hyménoptères, représentés essentiellement par la famille des Formicidae, avec une abondance de 18,69 %, et les dermaptères avec 13,87 %. A Merouana, les coléoptères et les orthoptères enregistrent des valeurs relativement proches avec respectivement 41,83 % et 39,69 %. De même pour les hyménoptères et les dermaptères qui ont aussi des proportions proches (8,82 % et 7,29 %). Nous avons noté également, en plus des aranéides (1,24 %) et des mantoptères (0,66 %), l'importance de l'ordre des rongeurs consommé avec 0,42 % à El Madher, et l'importance des blattoptères (0,71 %) et des mantoptères (0,38 %) à Merouana, soulignant, de ce fait, la diversification du régime alimentaire de B. ibis. Nos résultats sont comparables à ceux de BOUKHEMZA (2000) qui a signalé l'importance des coléoptères, des orthoptères, des hyménoptères et des dermaptères. Les différences dans les taux de consommation de chacun de ces ordres dans différentes localités sont également notées par cet auteur tout en les associant à l'entomofaune locale, reliée ellemême, étroitement aux types de milieux existants dans chaque localité. Ainsi, à titre d'exemple, à Drâa Ben Khedda, où les oiseaux ont consommé plus de coléoptères, la dominance des prairies dans cette région a favorisé leur abondance. Alors qu'à Baghlia, au contraire, les cultures basses et les friches sont des milieux très riches en orthoptères, ce qui a favorisé leur forte consommation. > Variation temporelle Concernant la variation mensuelle, nous avons noté un maximum de consommation de coléoptères au mois d'avril à El Madher, et aux mois de février et de mars à Merouana. Le minimum de consommation des coléoptères est enregistré au mois d'août dans les deux zones d'étude, période durant laquelle les orthoptères enregistrent leur maximum de consommation. Les dermaptères sont présents durant toute la période d'étude, alors que les hyménoptères sont surtout consommés aux mois de septembre, de décembre et de janvier à El Madher et aux mois de novembre et de février à Merouana. Généralement, la consommation de ces différents ordres est très fluctuante d'un mois à un autre ; si la consommation d'un ordre diminue elle est compensée par un autre ordre et ceci conformément à leur phénologie. Ces observations concordent avec nos résultats concernant les disponibilités alimentaires et leur variation, et sont mentionnées également par BOUKHEMZA (2000), dans la vallée de Sébaou (Tizi-Ouzou). Il est à signaler l'importante consommation des rongeurs par B. ibis, notamment aux mois de novembre et de décembre. Selon SI BACHIR (2007), pendant cette période rude (baisse de températures et rareté des insectes qui entrent en diapause), l'espèce s'alimente des proies les plus grosses et vraisemblablement plus énergiques pour compenser les efforts déployés lors des longs déplacements. Ce dernier auteur rajoute que le régime alimentaire du garde-boeufs est sujet de variations considérables, non seulement d'une région à une autre mais également d'une manière saisonnière dans la même région. 4.5- Comparaison des régimes alimentaires des deux échassiersLa comparaison de l'abondance en nombre des principaux ordres de proies consommés par les deux échassiers est faite en fonction de leurs périodes phénologiques (reproduction et hors reproduction). Pendant la période de reproduction, les coléoptères viennent en tête du régime alimentaire de la Cigogne blanche avec 51,35 %, suivis par les orthoptères (38,02 %) et les dermaptères (8,54 %). Quant au garde-boeufs, ce sont les orthoptères qui dominent son régime alimentaire en période de reproduction avec un taux de 46,1 %, mais leur taux n'est pas trop loin de celui des coléoptères qui sont noté avec 37,5 %, suivis par les dermaptères (11,0 %) et les hyménoptères (2,1 %) (Fig. 32). Durant la période hors reproductrice, le régime alimentaire de la Cigogne blanche est basé principalement sur les orthoptères (67,14 %), suivis par les coléoptères (27,29 %) et les dermaptères (4,89 %). Lors de cette même période, le Héron garde-boeufs consomme principalement des coléoptères (40,01 %), des orthoptères (23,38 %), des hyménoptères (22,84 %) et des dermaptères (11,2 %) (Fig. 32). Il est à signaler que pendant la période de reproduction, les taux de consommation des coléoptères, des orthoptères et des dermaptères sont assez proches pour les deux échassiers étudiés, ce qui dénote d'un risque de compétition vis-à-vis de ce type de proies en cette période. Par ailleurs, le bol alimentaire de B. ibis parait plus diversifié avec 134 espèces que celui de C. ciconia composé seulement de 79 espèces. Aussi, l'indice de SHANNON, révèle que le régime alimentaire du Héron garde-boeufs est plus diversifié avec 5,2 bits que celui de la Cigogne blanche avec 3,7 bits. De même, l'indice d'équitabilité indique que le régime alimentaire du garde-boeufs est relativement plus équilibré (E = 0,7) que celui de la cigogne (E = 0,6). En deuxième lieu, l'indice de similitude de SORENSON qui s'élève à 62,2 % montre qu'il existe une assez grande similitude entre les régimes alimentaires des deux échassiers. L'application de l'indice d'électivité d'IVLEV, indique une sélection positive pour les deux modèles biologiques étudiés aux orthoptères, aux coléoptères et aux dermaptères dans les trois types de milieux échantillonnés (le milieu humide, la culture basse et la pelouse naturelle). Les orthoptères notés avec une valeur de sélection positive dépassant 0,8 sembleraient être les proies les plus recherchées par les deux échassiers. Les dermaptères qui ont une valeur de sélection positive allant de 0,5 à 0,7 pour la Cigogne blanche, et de 0,7 à 0,8 pour le Héron garde-boeufs, sont également des proies nécessitant plus d'effort de capture par leur manque dans les gagnages. Reproduction
Rodentia Squamata Dermaptera Orthoptera Coleoptera Hymenoptera Hors reproduction
Ciconia ciconia Bubulcus ibis
Figure 32- Fréquences d'abondance relative des principaux ordres de proies consommées par C. ciconia et B. ibis pendant les périodes de reproduction et hors reproduction dans la région de Batna. Les coléoptères montrent une valeur de sélection positive moins importante, variant entre 0,1 et 0,3 chez les deux échassiers, ce qui signifie qu'ils seraient plus abondants dans les gagnages par rapport aux orthoptères et aux dermaptères, mais moins prisés par les deux oiseaux étudiés. La sélection négative des aranéides et des hyménoptères s'explique par leur abondance dans les gagnages et leur moindre consommation par les deux échassiers. C'est le cas pour l'indice de sélection des hyménoptères consommés par le Héron garde-boeufs, qui est de -0,5 dans les trois milieux de gagnage. Toutefois, l'analyse statistique n'est significative que pour la similitude de la composition des régimes alimentaires des deux espèces que lors de période de reproduction et seulement dans zone d'El Madher. A cet effet, les indices de similitude et d'électivité calculés, ainsi que l'analyse statistique dénotent d'un risque d'une compétition interspécifique entre les deux modèles biologiques étudiés qui ont tendance à élire en priorité les mêmes groupes de proies (coléoptères, orthoptères et dermaptères) en particulier lors de la période de reproduction. Selon BENTAMER (1998), malgré la ressemblance apparente dans les régimes alimentaires de la Cigogne blanche et du Héron garde-boeufs, il existe certaines différences entre les proies entomologiques consommées. La Cigogne blanche consomme des proies entomologiques de tailles plus grandes que celles consommées par le Héron garde-boeufs. Bien que les mensurations des proies n'ont pas été réalisées, nous avons également remarqué ces mêmes observations lors de notre travail. BOUKHEMZA (2000), en comparant les régimes alimentaires de ces deux échassiers dans la vallée du Sébaou, a confirmé l'existence d'un certain isolement trophique entre ces deux espèces grâce à une analyse de leur partage suivant un gradient de taille des proies. Selon ce dernier auteur, la taille des proies est une dimension de la niche par laquelle ces deux espèces sont susceptibles de réaliser un isolement trophique. 5- Statut bioécologique des deux échassiers étudiés dans la région de Batna et recommandations de conservation5.1- Statut bioécologique de la Cigogne blancheDans une large partie de son aire de répartition, la Cigogne blanche a vu ses effectifs diminuer depuis les années 1930. Ce déclin s'est accentué après les années 1950 notamment parmi la population européenne occidentale (SCHULZ, 1999). Les résultats internationaux des deux derniers recensements de 1996 et 2004-2005 ont révélé un développement positif des populations de cigognes dans la majorité des sites de sa reproduction. Selon THOMSEN et HÖTKER (2006), les populations de cigognes blanches ont décliné de 20 % entre 1974 et 1984 puis elles ont augmenté de 23 % entre 1984 et 1994- 95 ; la population occidentale a augmenté de 75 % depuis 1984, alors que la population orientale a augmenté de 15 %. Aussi, les résultats préliminaires du dernier recensement de 2004-2005, recueillis de 13 états, montrent que la population est encore en augmentation et qu'il y a des pays où cette augmentation est de l'ordre de 100 % (THOMSEN et HÖTKER, 2006). Cette évolution des effectifs est constatée aussi dans le bassin méditerranéen. En Algérie, d'après MOALI-GRINE (2005), les récents dénombrements de 1998 et 2001, ont tous les deux confirmé la tendance à l'augmentation de la population qui est passée de 1.195 couples en 1993 à près de 6.000 couples en 2001, ce qui rejoint les synthèses du colloque organisé à Hambourg en 1996 concernant l'ensemble des populations de Cigognes blanches. Selon HAMDI et al. (2007), en Tunisie, le nombre de nids est passé de 358 entre les années 1980-1999 à 489 nids en 2002-2005. Dans la région de Batna, un recensement minutieux réalisé récemment en 2006 par DJADDOU et BADA (2006), donne 490 nids dont 7 nids non occupés. Deux autres recensements, aussi exhaustifs, réalisés en collaboration entre le département de Biologie (Association scientifique Biologie, Biodiversité et Durabilité) et la conservation des forêts de la wilaya de Batna, signalent un total de 593 nids occupés en 2007 et 670 nids occupés en 2008 ; ce qui donne une densité de la population variant de 11,5 couples/100 km2 en 2006 et 15, 9 couples/ 100 km2 en 2008. Les résultats de ces recensements assez exhaustifs, dénotent des lacunes observées au niveau des recensements habituellement réalisés chaque année par les services des conservations des forêts tant au niveau local que national ; qui rapportent généralement des effectifs très en deçà de la réalité. Ceci revient à dire que l'Algérie abrite des effectifs très importants par rapport à ce qui est signalé dans la littérature. Aussi, nous signalons l'observation personnelle de l'installation de nouveaux nids à Merouana, à Arris et à El Madher (Annexe, photos 10 et 13). A M'Chounèche (W. de Biskra), nous avons noté une première nidification de l'espèce en l'an 2009. Ceci montre que la région des Aurès connaît la même tendance ascendante des populations de cigognes suite à l'implantation de plus en plus importante de nouveaux couples dans la région. Le cycle biologique de la Cigogne blanche dans la région de Batna débute par les premières arrivées enregistrées à la fin janvier et au début février, et se termine par les départs en migration post nuptiale enregistrés de la fin juillet jusqu' à la mi-août. Pendant la période de son séjour, nous avons relevé 7 milieux d'alimentation différents fréquentés par l'espèce : les bordures d'eaux usées, les prairies, les friches, les labours, les cultures basses, les déchets d'aviculteurs et les milieux fauchés. Nous l'avons également observé fréquentant les décharges publiques en dehors des itinéraires suivis, notamment à Ain Touta et à Arris (Annexe, photo 5). L'étude du régime alimentaire basée sur la décortication des pelotes de réjection de la Cigogne montre que l'espèce est un prédateur entomophage. Elle s'alimente essentiellement d'insectes avec un taux de 99,23 % contre 0,13 % seulement de vertébrés. Une sélection positive est apparue pour les ordres des orthoptères, des coléoptères et des dermaptères dans les trois principaux milieux fréquentés : prairie, milieu humide et culture basse. Parmi les familles les plus abondantes et les plus constantes dans la composition de l'alimentation de l'espèce, nous citons les Caelifera ind., les Scarabaeidae, les Ensifera ind., les Labiduridae, les Silphidae, les Tenebrionidae, les Carabidae et les Harpalidae. 5.2- Statut bioécologique du Héron garde-boeufsDepuis le début du XXème siècle, le Héron garde-boeufs est une espèce en pleine expansion tant par l'évolution de son aire de répartition ou par l'augmentation locale de ses effectifs (SIEGFRIED, 1978 ; BREDIN, 1983 ; KUSHLAN et HANCOCK, 2005). Selon SI BACHIR (2007), le Héron garde-boeufs est une espèce en pleine expansion en Algérie, tant par son aire de répartition que par la dynamique de ses populations. Ce dernier auteur rajoute qu'en moins de 30 années, l'aire de nidification de l'espèce a connu une expansion de près de 2° de longitude vers le sud. Le Héron garde-boeufs, est à installation récente dans la région de Batna. L'enquête que nous avons mené sur l'historique d'installation du garde-boeufs dans cette région, révèle que l'espèce est sédentaire et se reproduit dans cette région depuis l'an 1995. Les premières régions colonisées sont celles d'Ain Touta en 1995, de Ras Layoun en 2000, de Merouana, d'El Madher et de Tazoult en 2002 et de N' Gaous en 2003. Le Héron garde-boeufs se manifeste dans la région par deux populations, l'une migratrice hivernante qui séjourne d'octobre jusqu'en mars et l'autre sédentaire nicheuse. L'importance des effectifs tant hivernants que nicheurs de B. ibis durant ces dernières années est révélée par le nombre important de dortoirs et de colonies recensés, qui est actuellement estimé à 16 dortoirs et à 10 colonies. La période de reproduction de B. ibis dans la région de Batna s'étale sur un peu plus de 4 mois. Sa reproduction est fortement conditionnée par les conditions climatiques et les différentes phases de nidification connaissent des variations locales. Une multitude de milieux d'alimentation est fréquentée par l'espèce : les bordures d'eaux usées, les prairies, les friches, les labours, les cultures basses, les immondices, les prairies inondées, les dépôts de fumier, les déchets d'aviculteurs et les milieux fauchés. Le bol alimentaire du héron garde-boeufs est très diversifié, composé par une grande variété de proies tant invertébrées que vertébrées. Son alimentation est composée essentiellement de coléoptères, d'orthoptères, d'hyménoptères et de dermaptères. Les familles de proies les plus prisées par l'espèce sont les Formicidae, les Caelifera ind., les Acrididae, les Carabidae, Labiduridae, les Harpalidae, les Pterostichidae, les Scarabaeidae et les Gryllidae. 5.3- Facteurs de menace et de mortalité des deux modèles biologiques étudiésLa Cigogne blanche et le Héron garde-boeufs sont exposés à plusieurs facteurs de menace qui aboutissent parfois même à la mortalité des deux espèces pendant les différentes étapes de leur développement (oeufs, poussins, adultes). Ces facteurs sont principalement de deux types : naturels et anthropiques. Parmi les facteurs dus à l'homme, nous citons : - Risques de brûlures dans les décharges publiques (Annexe, photo 5) ; - Risques d'intoxications lors des prises d'eau ou de l'alimentation dans les cours d'eaux polluées par les déversements des eaux usées industrielles et urbaines ; - Collision et électrocution après contact avec des câbles électriques de haute et moyenne tension (Annexe, photos 11 et 12) ; - Risques d'intoxications par les produits phytosanitaires, notamment que les deux espèces étudiées fréquentent les milieux agricoles par prédilection. - Perte des habitats favorables à cause de l'aménagement des zones humides par drainage, ce qui a pour conséquence une réduction des ressources en alimentation dans les milieux naturels pour les deux échassiers étudiés (exemple chott Djendli). - Remplacement des toitures traditionnelles en tuiles par des dalles et abattage des grands et vieux arbres servant de supports aux nids de la Cigogne blanche. Parmi les facteurs naturels, nous citons : - L'effet des conditions climatiques notamment les périodes de sécheresse prolongées et qui coïncident majoritairement avec la période de reproduction des deux échassiers étudiés. - Les maladies liées à des agents pathogènes différents (Annexe, photos 14 et 15). Des ectoparasites hématophages sont signalés sur des poussins de cigognes et de hérons (FERRAH, 2007 ; BEGHORA et DJEBIR, 2008). - Prédateurs des oeufs et des poussins aux nids (chats, reptiles). 5.4- Intérêt environnemental de C. ciconia et B. ibis Les oiseaux ont un intérêt particulier par leur situation dans le sommet des pyramides trophiques. Connaître l'écologie trophique de deux échassiers de la région de Batna contribuera à une mise en évidence de biocénoses notamment celles des insectes apportant un intérêt agro-écologique pour la région. De point de vue agronomique, nous distinguons trois catégories de familles d'insectes consommés par la Cigogne blanche et le Héron garde-boeufs. Il s'agit des insectes phytophages nuisibles, des insectes prédateurs utiles et des insectes qui paraissent indifférents, mais qui ont un intérêt écologique dans les agrosystèmes. Parmi les familles d'insectes nuisibles, nous citons des hétéroptères suceurs de sève telles les Pentatomidae et les Geocoridae. Nous avons identifié le genre Aelia dans le régime alimentaire de la Cigogne blanche, à qui appartiennent certaines punaises ravageuses de céréales qui piquent les feuilles et les grains laiteux notamment ceux du blé. Il y a lieu également de noter la prédation des deux échassiers sur des proies insectes ravageurs des céréales et des cultures maraîchères tels les Tettigonidae, les Gryllotalpidae et les Gryllidae. Mais ces familles sont d'une importance économique secondaire comparées aux familles appartenant au sous ordre des Caelifera, telle les Acrididae qui causent des dégâts considérables aux diverses cultures. Parmi les proies appartenant à la famille des Formicidae, nous notons l'espèce Cataglyphis bicolor qui est une espèce protégée. Nous signalons aussi Messor barbara, fourmi des moissons et Tapinoma simroti. Cette dernière espèce cause des dégâts aux cultures indirectement en défendant les colonies des pucerons dont elle utilise le miellat et en dissuadant, à cet effet, les coccinelles, meilleures prédateurs des pucerons. Les familles des Cicindellidae, des Carabidae, des Nebriidae, des Scaritidae, des Licinidae, des Harpalidae et des Pterostichidae appartiennent toutes à la super famille des Caraboidea et entre en partie dans la composition du régime alimentaire des deux oiseaux étudiés. En sus des Mantidae, et des Coccinellidae, toutes les espèces appartenant aux familles citées plus haut sont carnassières et sont d'une grande utilité agricole. Les adultes et les larves des ces groupes se nourrissent de larves, de chenilles et même des adultes d'insectes nuisibles. Parmi les familles considérées indifférentes à l'agriculture, nous citons les Labiduridae et les Forficulidae de l'ordre des dermaptères, les Dytiscidae, les Hydrophilodae et les Histeridae de l'ordre des coléoptères. Par exemple, de la famille des Scarabaeidae, les insectes appartenant aux genres : Copris, Gymnopleurus, Scarabaeus, Onitis, Bubas, Onthophagus, Geotrupes et Aphodius sont des coprophages et ont, de ce fait, un rôle important dans le recyclage des bouses et des crottes, ce qui enrichit le sol agricole en matières organique. Cependant, au stade larvaire, certains Scarabaeidae ont des larves phytophages qui vivent aux dépens des racines de divers végétaux et sont, a cet effet, parfois très nuisibles aux cultures maraîchères et aux pépinières. Il est utile de signaler aussi que l'inventaire réaliser pour avoir une idée sur les proies potentielles dans trois types de gagnages, comporte des espèces qui ne sont pas détectées dans le régime alimentaire des deux échassiers, exemple des annélides et de larves de lépidoptères et de diptères. Beaucoup d'espèces échappant aux pièges installés, n'échappent pas aux becs de C. ciconia et de B. ibis comme les Dytiscidae, les Hydrophilodae et les Staphilinidae. Ceci met l'accent sur l'importance des études concernant les régimes alimentaires des oiseaux insectivores dans l'approfondissement des connaissances sur la biodiversité. La Cigogne blanche et le Héron garde-boeufs sont, de ce fait, d'un intérêt agroécologique certain. Chacune occupe une place importante dans les maillons trophiques et a des fonctions dans les cycles biogéochimiques. Elles permettent à la fois de maintenir l'équilibre de la dynamique de certaines espèces et de limiter la pullulation de certains groupes ravageurs de cultures. Outre cet intérêt agronomique et écologique que présentent les deux espèces étudiées, il est à signaler l'importance des deux échassiers en tant qu'indicateurs biologiques des écosystèmes fréquentés : - Les variations apparaissant dans le cycle biologique et la dynamique des populations de ces deux échassiers, pourraient se révéler comme des témoins des changements globaux de l'environnement, les changements climatiques en particulier. - La fréquentation de plus en plus importante des milieux anthropiques créés par l'homme révèle de nouvelles adaptations des deux espèces qui exploitent ces milieux mais avec des risques de véhiculer des maladies et leurs agents pathogènes, risques pour l'Homme et pour d'autres espèces. 5.5- Recommandations de gestion et de conservationEn vu des constatations concernant la bio-écologie des deux espèces et de leurs relations avec le milieu naturel et humain et des impacts probables sur l'environnement nous avons jugé utile de dégager les recommandations et suggestions d'aménagement et de gestion des deux échassiers dans notre région. Il s'agit de recommandations d'ordre scientifique : - Suivi de la biologie de reproduction et de la dynamique des populations des deux modèles biologiques étudiés ; - Le baguage des oiseaux est également un autre moyen scientifique permettant d'avoir des données originales, tels la connaissance avec précision des voies de déplacements des oiseaux et l'approfondissement des informations sur l'état des populations comme le taux de survie et le taux de mortalité. Et d'autres suggestions d'ordre de conservation : - Mise en place de plates formes artificielles sur les poteaux électriques afin de favoriser et de sécuriser l'installation des nids de la Cigogne blanche. Cette procédure pourrait également éviter quelques risques de compétition sur les sites actuels partagés entre les deux espèces ; - Conservation des zones humides et des milieux naturels généralement utilisés par les deux espèces ; - Correction des pratiques culturales basées sur l'utilisation abusive des pesticides qui accélèrent la stérilisation des biotopes. La Cigogne blanche et le Héron garde-boeufs ont un régime alimentaire basé sur les insectes et contrôlent, de ce fait, les populations de plusieurs déprédateurs. Ce rôle ne serait aucunement remplacé par l'usage de pesticides qui, non seulement sont susceptibles d'éradiquer toutes les populations d'animaux nuisibles, mais aussi d'engendrer des conséquences écologiques dangereuses ; - Prendre en considération et remédier aux problèmes de fréquentation des décharges (dépotoirs, rejets d'aviculteurs et de fermiers...) ; - Mettre fin à la destruction des nids de cigognes qui est une espèce protégée en Algérie. Enfin, il serait utile de mettre en place un plan d'action pour ces deux espèces avec la collaboration de plusieurs intervenants, tels les scientifiques, les amateurs et les riverains des sites fréquentés, des associations et des autorités locales. Ceci aidera à mieux comprendre, conserver et gérer les deux espèces étudiées. Au terme de cette étude menée dans la région de Batna sur la niche écologique de la Cigogne blanche et du Héron garde-boeufs nous avons arrivé aux conclusions suivantes : Le cycle biologique de la Cigogne blanche dans la région de Batna débute par les premières arrivées enregistrées à la fin janvier et au début février, et se termine par les départs en migration post nuptiale enregistrés de la fin juillet jusqu'à la mi-août. La ponte débute à la troisième décade de mars, les éclosions à la troisième décade d'avril et les envols des jeunes cigogneaux à la troisième décade de juin. Le Héron garde-boeufs, est une espèce à installation récente dans la région de Batna. L'enquête menée sur la question révèle que l'espèce est sédentaire et se reproduit dans la région depuis 1995. Les premières régions colonisées sont Ain Touta et Ras Layoun. L'importance des effectifs dénombrés tant en hiver qu'en période de nidification de B. ibis révélée par le nombre important de dortoirs et de colonies recensés (actuellement 16 dortoirs et 10 colonies), dénote de l'implantation de l'espèce en tant que sédentaire nicheuse. L'espèce passe une période de reproduction de plus de 4 mois. Nous avons noté un décalage de près d'un mois de cette période entre les deux zones d'étude. Ceci est-il tributaire seulement des conditions de l'environnement de chacune des deux régions ou serait-il également en relation avec l'aspect pionnier de l'espèce. Pendant la période de nidification, les deux oiseaux étudiés investissent les milieux les plus proches des colonies de reproduction notamment en période d'élevage des jeunes. La diminution du nombre de garde-boeufs en période de reproduction par rapport à la période d'hivernage laisse à penser qu'il existerait deus types de populations, une migratrice hivernante et l'autre sédentaire nicheuse. Dans trois types de gagnages explorés dans la région d'El Madher : une prairie, un milieu humide et une culture basse, nous avons recensé 234 espèces d'invertébrés réparties en 6 classes, 17 ordres et 94 familles. Les insectes dominent largement avec 223 espèces. L'abondance totale des invertébrés récoltés est relativement similaire dans les trois milieux étudiés. La richesse totale la plus élevée est notée dans la pelouse naturelle. Cependant, l'indice de diversité de SHANNON montre que le milieu humide abrite les peuplements de proies les plus diversifiés avec une valeur de 5,63 bits. L'étude de la répartition temporelle des proies potentielles des deux échassiers, montre une grande variation au cours des quatre saisons climatiques. C'est pendant les saisons printanière et estivale, correspondant aux périodes de reproduction des deux échassiers, que nous avons enregistré les plus grands effectifs d'invertébrés. La valeur la plus élevée de la richesse totale est notée au printemps alors que l'indice de SHANNON révèle une diversité plus importante en automne. Les principaux milieux de gagnage connus et fréquentés par les deux espèces sont les prairies, les cultures basses, les friches, les milieux fauchés et les labours. Par ailleurs, nous avons noté la fréquentation de divers dépotoirs tels les déchets de fermiers et les dépôts d'ordures. Ces milieux procureraient aux modèles biologiques étudiés une ressource renouvelable de larves et de vers non décelables dans leurs pelotes, mais constitueraient également un danger pour leur santé par les risques de brûlures et d'intoxications. Dans les milieux de gagnage, nous avons noté l'association active du garde-boeufs au bétail et aux engins agricoles. Ces types d'association sont particulièrement enregistrés dans les prairies, les friches, les labours, les milieux fauchés et dans les cultures basses. En moyenne, une pelote de la Cigogne blanche mesure 5,1 cm de long et 3,8 cm pour son petit diamètre et pèse 9,7 g en poids sec. Une pelote du garde-boeufs mesure 3,7 cm de long, 2,1 cm de diamètre et pèse 1,8 g en poids sec. L'étude du régime alimentaire basée sur la décortication des pelotes de réjection des deux échassiers, montre que ce sont des prédateurs entomophages, s'alimentant essentiellement d'insectes avec des taux respectifs de 99,2 % et 98,4 % du nombre total des proies ingérées. Les proies vertébrées ne constituent qu'un faible pourcentage (0,13 % pour la Cigogne blanche et 0,74 % pour le Héron garde-boeufs). Le spectre alimentaire de la Cigogne blanche est composé de 79 espèces appartenant à 5 classes, 11 ordres et 34 familles. Parmi les familles les plus abondantes et les plus constantes dans le régime alimentaire de C. ciconia nous avons noté les Caelifera ind. suivis par les Scarabaeidae, les Ensifera ind., les Labiduridae et les Silphidae. Nous avons remarqué également l'importance des familles des Tenebrionidae, Carabidae et Harpalidae. L'étude de la variation temporelle du régime alimentaire de la Cigogne blanche montre que les taux des coléoptères et des orthoptères consommés varient d'une façon inverse. Le taux des coléoptères augmente à partir du mois de février pour atteindre un maximum au mois d'avril puis commence à décroître jusqu'au mois de juillet. Par contre, les orthoptères marquent les taux les plus élevés aux mois de février et de juillet. Le bol alimentaire du Héron garde-boeufs est plus diversifié que celui de la Cigogne blanche, constitué de 134 espèces réparties en 9 classes, 19 ordres et 56 familles. Cependant, la structure du régime alimentaire de B. ibis est sensiblement similaire à celle de C. ciconia, avec seulement 8 familles constantes et 9 familles accessoires. Parmi les familles les pus abondantes et les plus constantes, nous signalons principalement les Formicidae suivis par les Caelifera ind., les Acrididae, les Carabidae et les Labiduridae. En général, l'alimentation de B. ibis est composée essentiellement de coléoptères, d'orthoptères, d'hyménoptères et de dermaptères. Toutefois, ces ordres de proies enregistrent une consommation fluctuante d'un mois à un autre. Les coléoptères atteignent leur apogée au mois d'avril, alors que les orthoptères enregistrent leur taux le plus élevé au mois d'août. Par ailleurs, les proies vertébrées sont représentées essentiellement par les rongeurs consommés surtout aux mois de novembre et de décembre. La variation des ordres de proies consommées par les deux échassiers, entre les deux zones d'étude, relèverait de la variation de la disponibilité et de l'accessibilité de l'entomofaune locale. Le bol alimentaire de B. ibis est plus diversifié que celui de C. ciconia. Par contre, l'abondance en nombre de proies contenues dans une pelote est plus forte chez la cigogne que chez le garde-boeufs. L'indice de SHANNON, révèle également que le régime alimentaire du Héron gardeboeufs est plus diversifié que celui de la Cigogne blanche. L'indice d'équitabilité calculé indique que le régime alimentaire du garde-boeufs est relativement plus équilibré que celui de la cigogne. L'indice d'électivité d'IVLEV, indique une sélection positive par les deux modèles biologiques étudiés aux orthoptères, aux coléoptères et aux dermaptères dans les trois milieux d'échantillonnage. La comparaison des différents paramètres étudiés de la niche écologique des deux modèles biologiques (cycle biologique, exploitation des gagnages, structure du régime alimentaire) permet de suggérer un risque de « compétition interspécifique ». En effet, C. ciconia et B. ibis mènent leurs activités d'élevage des jeunes poussins au cours de périodes parallèles et utilisent principalement les mêmes sites de nidification et les mêmes milieux d'alimentation. Leurs régimes alimentaires sont basés principalement sur la consommation des coléoptères, des orthoptères et des dermaptères. C'est en particulier, pendant la période de reproduction que le taux de consommation de chacun de ces ordres par les deux échassiers est assez proche. Ceci dénoterait d'un risque de compétition vis à vis de ces types de proies en cette période. Nos résultats indiquent également que B. ibis exploite d'une manière énergique une plus grande diversité de milieux d'alimentation et recourt à l'association au bétail et aux engins agricoles qui mettent à jours plusieurs types de proies. Cette stratégie peut lui procurer des ressources alimentaires supplémentaires ce qui minimise sensiblement le risque de compétition entre les deux espèces dans les milieux d'alimentation. Encore le menu alimentaire du Héron garde-boeufs est plus diversifié, ce qui nous permettrait, pour le moment présent, d'éloigner le risque de compétition. Connaître l'écologie trophique de deux échassiers de la région de Batna, avec plus grande acuité, contribuerait non seulement à mettre en évidence les véritables risques de chevauchement des niches des deux espèces, mais également contribuerait aux études de biodiversité de mise en évidence des biocénoses notamment celles des insectes ayant un intérêt agro-écologique pour la région. En perspective et au terme de cette étude, il serait utile de compléter ce travail en mettant en oeuvre d'autres méthodes avec un protocole à long terme. Il s'agit entre autres de : - Etudier et faire le suivi régulier de la biologie de reproduction de ces deux échassiers, notamment en se penchant sur les paramètres liés au succès de reproduction ; - Associer plusieurs méthodes dans l'étude du régime alimentaire, telles la trituration des pelotes de rejection, l'observation directe de l'activité de chasse, l'évaluation des proies consommées en terme de biomasse et l'évaluation des valeurs énergétiques des proies consommée ; - Faire un suivi régulier des effectifs, des milieux d'alimentation, des dortoirs et des colonies du Héron garde-boeufs, nouvelle espèce invasive de la région en vue d'évaluer et de contrôler l'évolution de ses populations ; - Elaborer une stratégie nationale de gestion et de conservation des deux espèces étudiées. Enfin, il est souhaitable que ce travail soit suivi par d'autres études pour affiner les connaissances sur la biologie et l'écologie de ces deux espèces, pour mieux comprendre les relations qui existent entre la dynamique des populations et la qualité de l'environnement (gagnages et stocks alimentaires disponibles, sites de reproduction ...).
Annexe photographique (Photos personelles)
Photo 1- Colonie plurispécifique de cigognes blanches et hérons garde-boeufs (Merouana, 22 juillet 2007). Photo 2- Colonie de cigognes blanches (El Madher, 20 février 2007). Photo 3- Manière dont la Cigogne blanche façonne l'intérieur de son nid (El Madher, 02 avril 2007) 192
Photo 4- Cigognes blanches et hérons garde-boeufs s'alimentant dans la retenue collinaire de Bousbarka (Oued Taga, 22 juillet 2007). Photo 5- Cigogne blanche s'alimente dans une décharge publique en feu (Arris, 11 juillet 2007). Photo 6- Hérons garde-boeufs s'alimentant dans des déchets de fermiers (Merouana, 24 avril 2007).
Photos 7 et 8- Dépérissement des branches de Pins d'Alep utilisés en guise de dortoir et de colonie par B. ibis (Merouana, 22 juillet 2007). Photo 9- Vieux arbres supports de nids de cigognes (Oued Taga, 17 juillet 2007)
Photo 10- Supports artificiels utilisés par la Cigogne blanche lors de la construction de nouveaux nids (Arris, 22 juin 2009). Photo 11- Enfants capturant une cigogne blanche ayant subit une décharge électrique (Psychiatrie d'El Madher, 21 juillet 2007). Photo 12- Cigogne blanche morte électrocutée à 50 m d'un transformateur d'électricité de haute tension (Arris, 13 août 2007).
Photo 13- Nid de cigogne blanche construit sur une antenne de la téléphonie mobile (Arris, 23 avril 2007). Photos 14 et 15- Cigogneau atteint d'une pathologie au niveau des pattes (El Madher, 25 juin 2007). RésuméLa présente étude porte sur la niche écologique de la Cigogne blanche Ciconia ciconia et du Héron garde-boeufs Bubulcus ibis dans la région de Batna. Le suivi de ces deux échassiers est mené dans deux zones différentes, El Madher et Merouana, durant une période de 13 mois, étalée de novembre 2006 jusqu'à novembre 2007. Pendant cette période, nous avons élucidé la chronologie des cycles biologiques des deux espèces. En fonction des stades phénologiques des deux espèces, nous avons étudié plusieurs paramètres de leurs niches trophiques. L'étude des disponibilités alimentaires est effectuée dans la zone d'El Madher, dans trois types de gagnages : une prairie, un milieu humide et une culture basse. Nous y avons recensé 234 espèces d'invertébrés. Les insectes dominent largement avec un taux de 83,9 %. L'abondance des invertébrés récoltés est relativement similaire dans les trois milieux étudiés. Le plus grand nombre d'invertébrés récoltés est enregistré durant les saisons printanière et estivale. Le suivi des modèles biologiques étudiés dans leurs milieux d'alimentation, nous a permis de relever 7 milieux de gagnage différents fréquentés par la Cigogne et 10 milieux fréquentés par le Garde-boeufs. Les principaux milieux fréquentés par les deux espèces sont les prairies, les cultures basses, les milieux fauchés et les labours. Par ailleurs, nous avons noté la fréquentation de dépotoirs tels les déchets de fermiers et les dépôts d'ordures. Dans les milieux de gagnage, nous avons noté également l'association active du garde-boeufs au bétail et aux engins agricoles. L'analyse de 96 pelotes de C. ciconia et de 110 pelotes de B. ibis montre que les deux échassiers sont des prédateurs entomophages, s'alimentant essentiellement d'insectes avec des taux respectifs de 99,2 % et 98,4 %. Le bol alimentaire du Héron garde-boeufs est plus diversifié (134 espèces) que celui de la Cigogne blanche (79 espèces). Néanmoins, nous avons enregistré la dominance des orthoptères, des coléoptères et des dermaptères dans le régime alimentaire de la Cigogne, et la dominance des orthoptères, des coléoptères, des hyménoptères et des dermaptères dans celui du garde-boeufs. Le Héron garde-boeufs, espèce en expansion mondiale, est à installation récente dans la région de Batna, à partir de l'an 1995. La comparaison des niches écologiques des ces deux échassiers révèle des risques de compétition interspécifique, notamment en période d'élevage des poussins. Cependant, B. ibis montre des aptitudes et des stratégies tels la diversification de son menu alimentaire et des milieux d'alimentation ainsi que l'association active au bétail qui sont considérés des facteurs minimisant pour le moment actuel le risque de compétition des deux espèces. Des recommandations de conservation et de gestion sont émises à la lumière des résultats obtenues. Mots cléfs : Batna, Bubulcus
ibis, Chevauchement de niches, Ciconia ciconia,
Cigogne SummaryThis study focuses on the ecological niche of the White stork Ciconia ciconia and Cattle egret Bubulcus ibis in the region of Batna. The monitoring is conducted in two different zones, El Madher and Merouana, during a period of 13 months, from November 2006 to November 2007. During this period, we have clarified the biological cycles of both species. In the light of these stages phenological, we have studied the settings of their trophic niches. The study of food availability is carried out in the zone of El Madher, in three types of foraging sites: a grassland, a wetland and a low culture. We have identified 234 species of invertebrates. Insects dominate with 83.9%. The abundance of invertebrates was relatively similar in the three areas studied. The greatest number of invertebrates was recorded during the spring and summer seasons. The monitoring of biological models studied in their backgrounds food, has allowed us to raise 7 different foraging sites frequented by the White stork and 10 foraging sites frequented by the Cattle egret. The main food areas used by the two species are grassland, low culture, harvested areas and plowing. In addition, we have noted the attendance of dumps such waste of farmers and the deposits of garbage. In foraging sites, we have noted the association active of the Cattle egret to livestock and agricultural machinery. The analysis of 96 pellets of C. ciconia and 110 pellets of B. ibis, shows that both waders are entomophagous predators, feeding mainly on insects with rates of 99.2% and 98.4%. Feeding of Cattle egret is more diversified (134 species) than that of the White stork (79 species). Nevertheless, we have recorded the dominance of Orthoptera, Coleoptera, and Dermaptera in the diet of the White stork, and the dominance of Orthoptera, Coleoptera, Hymenoptera and Dermaptera in that of Cattle egret. The Cattle egret, species in global expansion, has recently installed in the Batna region, from the year 1995. Comparison of ecological niches of these two waders reveals interspecific competition risks, including during rearing of chicks. However, B. ibis shows skills and strategies such as diversifying its preys and feeding areas as well as active association to the cattle which are considered factors for the current time minimizing the risk of competition of the two species. Recommendations for conservation and management are made in the light of results obtained. Keywords: Batna, Biological cycle, Bubulcus ibis, Cattle egret, Ciconia ciconia, Niche overlap, Trophic niche, White stork. ~!"#~~Ç ä.2141 æ Ciconia ciconia ~~ ~~ .÷51.547Ç tijAlÇ L.ÇÑÏ ~~Å 1_41Ç Ç~å NH
. 2007J443: 41Å 2006 3,2 c54 ÉS.144 Ç_~: 13 É~+ 114i (DÇ).1Ç Baå Á~F G æ au:1-1Ç /~~ 5!6!~~~~!~~Ç 5!(Ñæ~~Ç (.Ç)4 ~66) #'~~ É.41Ç Baå á/& . 5!G.11Ç 5 ~9Ç~H~Ç ~~~~~Ç 3I#'G L-ÇÑ.. `°Ñ 1"æ ÌJA :
~åæ "*1 É}æ .% 83,9 V')
ÊÇJ.z.Ç Ïac 1441c. y1;9 Ê1,Ñ#
iN~Ç 5+ #G2 234 #'!SD ~ ~
ÑÒæ. . `!S~Çæ ~!)3~Ç S, æ 61 2Ç 1!1c ÏÏ36. ccÇ3~~Ç 5+ ÚÇ~2 7 aka~6) 1a1 :~~~ ~G3~Ç ÊÇ36, ~, al._91.1Ç a1.1å 4...41:14 äÅ : ~åæ au:1*1Ç C 141.11.
46~Ç -..16NÇ c Ç,.1Ç
CJA Ï~G ß#'å ä cÅ
-.,141Ç äj.14 4.1c ÏÏji~
ÚÇ~2 10 ä 414æ ~~
~~Ç ÏÏ51 LIÅ ÉÑ14Ç
1:14 Ña75 .
~Jæjz . ~~~46V~~Ç crÇjAlÇ cci k cÇÑ31Ç ÊcfÇæ #~#~ ä#,1k61 !D ÊÇJ.z.Ç 4.+6+ c. 1483 -.2141Ç äj.141 110 æ ~~ ~~ ÍÇ3NÅ Z. 96 c15 C144 5+ #G2 1 3 4) tzgu j -.2141Ç ä~%~) H( . % 98,4 æ % 99,2 :411-LAlÇ MV'#) ÊÇj.z1Ç 4i1c. Ê#~~~~ æ ~~'~_Ç Ê#~~~\ ~~'~_Ç Ê#~! 6V+ ä JA. .( ÊÇJ.z.Ç c. 1cj: 79) 61 ~~Ç .1c 141. (ÊÇ..z.Ç ä9.14 'G ÇÏÇ~~ jÂ_Ç 4å ~~'~_Ç Ê#!9#%\ 1~!~Å #,#Q+ Ê1r~~Z~~Ç B~å j72 æ ~~ ~~Ç .11c. H*( ~~'~_Ç . '!11.1Ç á~D #~) #'~~ 451Ç 4kÑ# 41Ç .1995 ~'+ Z1.114 ' 114 ~, #]D LD ..11c Ñ=Ç ~, 1-11-1=. ~!"#~~Ç ä~%~) Éji9 á/0 :"æ 1-~4 c+.1.-~iÇ LAU:~Ç 3*0 Ï.~æ ;4:1-AÅ ÊJil LM :1-.Ç C1)44-1 41:0~~Ç 19 41Ç
5!) j,#'6~Ç É~D 5+
411z.Ç 491Ç cci o '( C+Ç~G 364(
4åæ 44141Ç t.4
Ø14 42.0,N5 Ç.Âæ
ZA7IWA1Ç
. :41Ç1.~Ç .44Ç ~!~~~~!~~Ç ÉÑæ~~Ç ~!~~~~~~cÇ
| 
Changeons ce systeme injuste, Soyez votre propre syndic 
"Aux âmes bien nées, la valeur n'attend point le nombre des années" | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


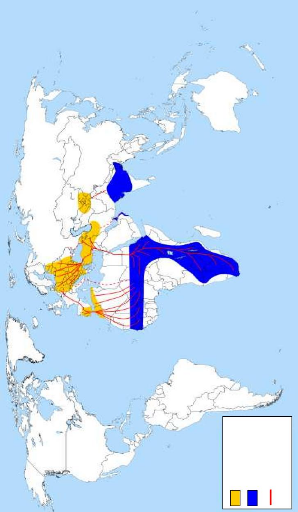


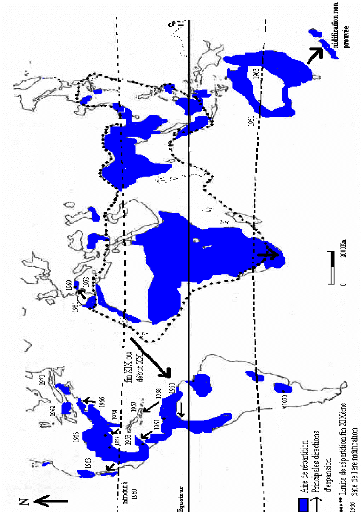
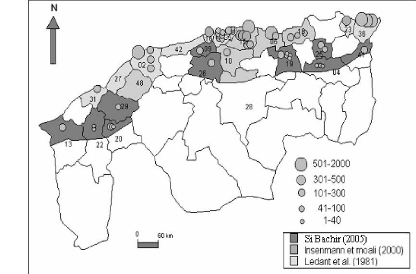
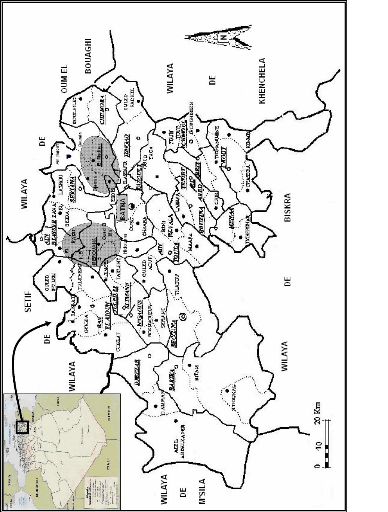
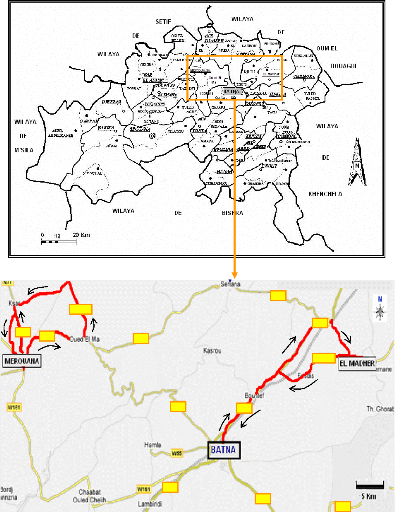















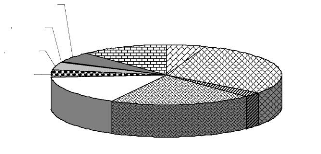









.){kind=link}