|
UNIVERSITE OFFICIELLE DE BUKAVU
B.P. 570 BUKAVU
FACULTE DES SCIENCES
DEPARTEMENT DE BIOLOGIE
Année académique 2009-2010
REPUBLIQUE DEMOCRATIQUE DU CONGO
Taxonomie et écologie des Bryophytes dans les
forêts de montagne de l'Est de la R.D.Congo. Cas du Parc National de
Kahuzi - Biega
Mémoire présenté en vue de l'obtention du
titre de Licencié en Sciences
Option : Biologie
Orientation : Ecologie et gestion des ressources
végétales (Sciences Botaniques)
DIRECTEUR : Prof. Dr. NYAKABWA M. CODIRECTEUR :
Prof. Dr. E. FISCHER ENCADREUR : C.T. BALEZI
Z.
Par WABIKA DUMBO PAZO
DEDICACE
A mafille ?osalie ICAVVKA DWMOO;
A mon épouse Ange LWANGO 7vtWAZI -- 5t1WA5lIBA
;
A ma mè e ?osalie 7ABv7A WANZA;
A mon pè e !mmanuel JYTY1v1BO
?CIRON-UV ;
WABiV1 DV!M 3O "AZo
~!~!~#i!i!w$%
L'achèvement de ce présent travail est
un concours de plusieurs personnes qui, de près ou de loin, nous ont
aidé matériellement ou moralement. Ainsi, nous exprimons nos
gratitudes aux Prof. Dr. NYAKABWA M., Prof. Dr. FISCHER E., C.T. BALEZI Z. ;
respectivement directeur, codirecteur et encadreur de ce travail.
Nous remercions les autorités tant
scientifiques qu'administratives de l'Université Officielle de Bukavu
qui ont contribué à notre formation pendant ces cinq
ans.
A nos chers parents DUMBO K. et Rosalia KABUKA ainsi
que nos frères : ZABINGWA D., WEMA D., BONNY D. et son épouse
Milchelline M., BIKALA D., MISANZILA D., PAULIN I. pour l'amour fraternel
affiché à notre égard.
A mon épouse Ange LWANGO et ma fille Rosalie
KABUKA pour avoir accepté d'être sacrifiées
matériellement et abandonnées au profit de ces
études.
Nous restons également reconnaissants envers le
C.T. BALAGIZI K. et MAHANGAIKO N. pour l'appui tant financier que
moral.
A nos cousins et neveux WALEMBA F., WABULA W., ISONGA
P. et MUKUBI D. qui ont matériellement contribué.
A nos amis : AKILI DEAZ, ITHE MW. et son épouse
Claire, IMANI K. et son épouse SIFA, KOKO H. et son épouse
Francine, Romain M. et son épouse ZABIBU, SIMON K. et CIZA
N.
A nos collègues : CIRHUZA Sylvie, MUGARUKA
Patient, MILENGE Héritier, MUGHOLI Elysée, MUTAYONGWA BABUNGIRE,
MUTAYONGWA BIKALIRA et SHUKURU Alliance pour les cinq ans de peines
passées ensemble.
WJBI~~ çDt)MBO "J4ZO
RESUME
Le présent travail a porté sur
l'étude taxonomique et écologique des Bryophytes des forêts
de haute montagne de l'Est de la RDC, plus précisément dans le
Parc National de Kahuzi - Biega. Les méthodes de transects et de
relevés ont été utilisées pour réaliser un
inventaire des Bryophytes et la caractérisation de leurs habitats
respectifs. L'étude taxonomique a permis de grouper les 48
espèces identifiées en 3 embranchements, 6 classes, 7 ordres et
35 familles. La famille de Bryaceae a été la plus
dominante. Par ce fait, nous avons déterminé pour chaque
espèce le type de gamétophyte, le type de substrat, le type
d'habitat ainsi que la tranche altitudinale. Ainsi, pour le type de substrat,
on a relevé la dominance des Bryophytes saprophytes et épiphytes
avec un taux de 30,79%, elles sont suivies des Bryophytes ripicoles et
saprophytes pour lesquelles chaque groupe a 15,38%, viennent ensuite les
Bryophytes terrestres avec 7 espèces (13,46%). Le secteur Tshibati a
été le plus diversifié, avec 30 espèces.
L'interprétation de types d'habitat a montré une dominance des
Bryophytes dans la forêt secondaire. Enfin, pour ce qui est de la tranche
altitudinale, il a été trouvé que les Bryophytes sont
reparties dans diverses tranches altitudinales, en tout, 7tranches ont
été reconnues. Seule l'espèce Symphyogyna cfr
podophylla s'est retrouvée dans 6 tranches d'altitude.
ABSTRACT
The present work is about the taxonomic and ecological
study of bryophytes of high land forests of the eastern part of the D.R.Congo,
especially Kahuzi - Biega National Park. The methods of transect and
relevé has been used in order to carry out a survey of bryophytes and
the characterization of their respective habitats. The taxonomic study has
allowed to group 48 species identified in 3 embranchments, 6 classes, 7 orders
and 35 families. The family of Bryaceae has been the most dominant, by
distinguishing the kind of gametophytes, the kind of substrat, the kind of
habitat, and the altitude for each collected species. Thus, the substrat kind
has been dominated by saprophytes bryophytes and epiphytic bryophytes of 40,
79% each; before rupicolous bryophytes of 15, 38%. Terricolous bryophytes are
the last of 13, 48%. Tshibati area has been the most diversified in species of
30 species. The analysis of habitat kind has shown the dominance of secondary
forests of 59, 3%. Focusing on the altitudinal gradient, we have noticed that
bryophytes are in different slices of altitude. At all, 7 slices have been
divided. Only the species Symphyogyna cfr podophylla has been met in 6
slices of altitude.
CHAPITRE 1. INTRODUTION
1.1. PROBLEMATIQUE
La République Démocratique du Congo est
comptée parmi les sept nations du monde riches en biodiversité ;
à côté du Mexique, de la Colombie, du Brésil, du
Madagascar, de l'Indonésie et de l'Australie selon Basabose et al.
(2001)
Le Parc National de Kahuzi-Biega (PNKB) est l'un de 7
parcs de la R.D.Congo. De part ses caractères, la forêt de
montagne se définit comme un peuplement pluristrate constituée
des arbres de taille basse et une grande abondance des bryophytes
épiphytiques. Le PNKB hébergerait une diversité des
bryophytes, mais malheureusement les données biologiques sur ces
dernières dans la région du Grands lacs sont encore
fragmentaires. Cette constatation est particulièrement vraie dans les
zones montagneuses de l'Est du Congo, incluant le Parc National de Kahuzi Biega
(PNKB). Alors, que plusieurs travaux scientifiques ont été
déjà effectués dans le PNKB sur les plantes, aucune
information n'est disponible sur les Bryophytes de cet
écosystème. L'herbarium du Centre de Recherche en Sciences
Naturelles de Lwiro qui englobe la bonne partie de spécimens
végétaux de la région, ne donne aucune information sur les
bryophytes de la contrée pourtant constituant un groupe taxonomique de
grande valeur écologique. Par ailleurs, il faut dire que les Bryophytes
jouent le rôle d'indicateur de pollution. Mais aussi comme toutes les
plantes chlorophylliennes, les Bryophytes sont une source d'oxygène et
sont absorbateurs du dioxyde de carbone ; ils constituent aussi un abri pour
des petits êtres vivants, ainsi qu'un réservoir d'eau dans des
tourbes. Pour essayer de donner une information sur la diversité de
Bryophytes, nous nous sommes fixé comme objectif de faire une
étude taxonomique et écologique de ces organismes dans la partie
de haute altitude du PNKB.
Pour bien s'imprégner de la diversité
des Bryophytes au PNKB, les questions suivantes ont animé notre
curiosité:
- Quelle est la diversité taxonomique des
bryophytes au Parc National de Kahuzi-Biega?
- La distribution des Bryophytes est-elle la
même dans tous les étages de la végétation du Rift
Albertin? particulièrement ceux du PNKB ?
- Les Bryophytes montrent-ils de préférence
quant à ce qui est du substrat?
- Les lieux de fixation des Bryophytes sur les arbres
différent-ils d'un substrat à l'autre ou d'un étage de
végétation à l'autre?
- Enfin, les Bryophytes du PNKB montrent-ils une plus
forte tendance au regroupement de leurs individus?
C'est donc à ces questions fondamentales que
cette étude voudrait apporter des éléments de
réponse par une approche quantitative et descriptive. Pour aborder ces
questions, des travaux de terrain et de laboratoire ont été
réalisés. Les travaux de terrains consisteront en des
relevés systématiques le long des transects dans
différents types de végétations et à
différentes tranches d'altitude.
1.2. Objectifs du travail
En initiant ce travail, nous nous sommes fixé
comme objectifs de faire une
étude taxonomiques et écologique des
Bryophytes dans divers habitats du PNKB.
Etude taxonomique :
> réaliser un inventaire des espèces de
Bryophytes présentes dans divers
habitats ;
> contribuer à la description morphologique des
Bryophytes.
Etude écologique :
> caractériser la spécificité des
relations bryophytes-plante hôte et biotope ;
> caractériser la spécificité des
bryophytes-statuts biologiques de la plante hôte
(substrat mort ou vivant).
> estimer et comparer la biodiversité attendue
et observée dans divers habitats.
1.3. Intérêt du travail
Les résultats du présent travail vont nous
permettre de :
- contribuer à la connaissance des
espèces des Bryophytes des forêts d'altitude du PNKB.
- constituer un herbier de bryophytes que nous allons
garder à l'herbarium de l'U.O.B qui servira d'outils de
référence pour les prochaines études de ce taxon au PNKB
en particulier et dans la région du Rift Albertin en
général.
1.4. Généralités sur les
Bryophytes
Les bryophytes constituent un groupe naturel des
plantes chlorophylliennes ; elles occupent une position intermédiaire
entre les algues et les plantes vasculaires en ce qui concerne
l'évolution bien qu'aucun lien ne les unit. Ils différent des
algues par leur important rôle de sporophyte et des plantes vasculaires
par le fait que le sporophyte diploïde est la plante entière
d'après Wigginston (2004).
Selon Letouzey (1982), les mousses sont formées
des petites racines (rhizoïdes) et tiges de quelques centimètres de
hauteur au maximum, serrées les unes contre les autres et garnies de
minuscules feuilles très simples n'ayant qu'une nervure. A certaines
saisons, au dessus de la tige feuillée, naissent des minuscules organes
mâles ou femelles et de ces derniers s'élève ensuite un
filament (petit) qui fait sortir à l'extrémité un
renflement (sporogone) d'où s'échappent les spores comme du
chapeau de champignon qui, en germant donneront une nouvelle
bryophyte.
Ces feuilles ne sont pas des vraies feuilles. Il est
plus juste de parler d'appendices phyllidiens. La mise au point d'un cormus
(tige et feuilles) augmente considérablement la surface
réceptrice aux rayons nourriciers du soleil chez les Bryophytes.
Toutefois, n'ayant aucune structure rigide et pas de système
circulatoire efficace, les bryophytes n'ont pas pu croître
énormément, la surface supérieure de l'appendice
phyllidien est couverte de cuticule qui présente plusieurs
caractéristiques importantes :
- elle est plus rigide ; ce qui favorise le port
dressé des petites tiges des bryophytes,
- elle les protège contre les radiations
nocives du soleil,
- elle limite la perte d'eau par évaporation.
(bryo2hb.en ligne)
Selon Petiot (2003), le cycle de vie des bryophytes
commence par une spore se développant en une structure filamenteuse
connue comme protonéma à partir du - quel apparaît un jeune
bouton qui se développe en un gamétophyte qui peut être une
plante feuillée ou thalloïde. Quand la plante feuillée ou
thalloïde atteint la maturité, elle produit des structures
reproductrices : l'anthéridie (organe reproducteur mâle) et
l'archégone (organe reproducteur femelle). Ces deux organes
reproducteurs sont protégés par un appendice feuillé
périgonial (protégeant anthéridie) et les feuilles
périchantiales (protégeant l'archégonie). Une
anthéridie est constituée d'une tige et une structure
sphérique contenant des anthérozoïdes biflagellés.
L'archégone est quant à lui, constitué d'un canal
cellulaire. Quand l'archégone atteint la maturité, le canal
cellulaire se désintègre et devient mucilagineux. Grâce
à l'eau de pluie, un
anthérozoïde se dépose sur
l'archégone, nage à travers la substance mucilagineuse dans le
canal et finalement atteint l'oosphère, s'unit avec cette
dernière et forment avec elle le zygote. Le zygote subit la division
mitotique pour produire un embryon multicellulaire qui éventuellement se
développe en sporophyte. Ce dernier a trois parties : le pied, le zeta
et la capsule. Le calyptra est souvent présent au sommet de la capsule.
Le sporophyte reste attaché au gamétophyte par le pied et
à partir du quel il obtient des nutriments. La capsule peut relativement
être simple ou complexe en structure. Dans la capsule, les cellules de
sporogone produisent les spores tétrades par méiose ou division
réductionnelle. Quand le sporophyte atteint la maturité, la
capsule s'ouvre, libère les spores. Les spores sont soit
apportées par le vent, dispersées et s'éloignent de la
plante - mère. Sous les conditions climatiques favorables et un substrat
souhaitable, elles germent pour produire le protonéma et le cycle se
répète.
Il y a ainsi alternance de génération,
la génération gamétophytique haploïde alterne avec la
génération sporophytique diploïde comme le montre le cycle
représenté dans la figure 1 ci - dessous.
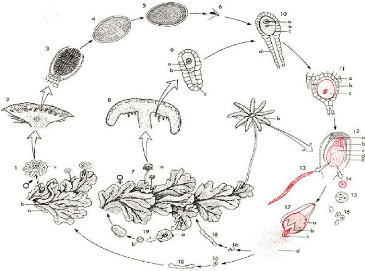
Figure 1 : Cycle de vie de l'hépatique
Marchantia polymorpha.
Légende de la Figure 1.
1 Thalle (gamétophyte)
a Rhizoïdes
b Corbeille à propagules
c Anthéridiophore
2 Anthéridiophore 3-5 Anthéridie
6 Anthérozoïde
7 Thalle (gamétophyte)
a Jeune archégoniophore
b Archégoniophore mature
8 Archégoniophore
9 Archégone
a O(v)osphère
b Cellule ventrale du canal
du col
c Cellule apicale du canal du
col
10 Archégone
a Noyau de l'o(v)osphère
b Noyau de l'anthérozoïde
c Ventre de l'archégone
d Col de l'archégone
11 Archégone
a Coiffe (involucre)
b Ventre
c Zygote
12 Archégone développé
a Pied
b Soie
c Capsule
d Ventre
e Coiffe (involucre)
13 Elatère
14 Cellule-mère de spore
15 Tétrade
16 Spores
17 Sporophyte déhiscent
a Coiffe (involucre)
b Capsule
c Elatères
d Spores
18 Spore germant
19 Cycle de reproduction
asexuée
a Corbeille à propagules
b Propagule
Notons que les bryophytes peuvent aussi se reproduire
asexuellement ou végétativement par une variété de
propagules : le sommet du bourgeon peut se détacher de la plante,
tombé sur la terre et se développer en une plante
mature.
Selon Wolfgang et al. (2009), les Bryophytes,
abusivement appelées mousses, est un groupe des plantes invasculaires
constitué de trois embranchements dont : Marchantiophyta, Bryophyta
et Anthocerophyta.
Ces trois embranchements différent l'un de
l'autre par des caractéristiques données dans le tableau
1.
Tableau 1 : Caractères distinctifs des trois
embranchements
|
Organes
|
Marchantiophyta
|
Anthocerophyta
|
Bryophyta
|
|
Protonéma
|
Peu développé
|
Peu développé
|
Bien développé
|
|
Gamétophyte
|
Thalle ou tige
feuillée
|
Thalle
|
Tige feuillée
|
|
Symétrie
|
Dorsi-ventrale
|
Dorsi-ventrale
|
Radiale
|
|
Rhizoïde
|
Unicellulaire non
ramifié
|
Unicellulaire non
ramifié
|
Multicellulaire ramifié
|
|
Anthéridie
|
Epidermique
|
Hypodermique
|
Epidermique
|
|
1ère division du
zygote
|
Transversale
|
Verticale
|
Transversale
|
|
Croissance du
sporophyte
|
Limité
|
Illimité
|
Limité
|
|
Capsule
|
pas de columelle pas de coiffe pas d'opercule pas de
péristome présence de valve présence
d'élatère
|
columelle présente pas de coiffe pas d'opercule
pas d'opercule pas de valve pseudo-élatère
|
columelle présente coiffe présente opercule
présente péristome présente pas de valve pas
d'élatère
|
Le tableau 1 présente les caractères
distincts de 3 embranchements de bryophytes : en se basant sur le
protonéma, les Marchantiophyta et les Anthocerophyta
ont un protonéma peu développé mais les Bryophyta
ont un protonéma bien développé. Le
gamétophyte des Marchantiophyta est soit une thalle ou une tige
feuille, mais les
Anthecerophyta et les Bryophyta ont
respectivement un thalle et une tige feuillée; Marchantiophyta
et Anthocerophyta ont une symétrie dorsiventrale mais
celle des Bryophyta est radiale ; les rhizoïdes sont
unicellulaires non ramifiées chez les Marchantiophyta et les
Anthocerophyta, mais multicellulaire ramifiée chez les
Bryophyta. L'anthéridie est épidermique chez les
Marchantiophyta et chez les Bryophyta mais hypodermique chez
les Anthocerophyta. La 1ère division du zygote est
transversale chez les Marchantiophyta et chez Bryophyta ;
elle est ventrale chez les Anthocerophyta. Il faut dire qu'ils
ont plusieurs autres caractères en commun, mais aussi de dissemblances
(voir tableau 1).
CHAPITRE 2. MILIEU D'ETUDE ET METHODES DE RECHERCHE
2.1. Milieu d'étude
Par ordonnance n° 81/Agriculture du 27/07/1937,
la réserve zoologique et forestière de Kahuzi-Biega fut
instituée par l'administration coloniale belge afin de
réglementer l'exploitation des ressources naturelles dans cette partie
de la forêt tropicale humide située dans le Kivu montagneux. En
1970, cette réserve située en haute altitude (1200-3308 m) prend
le statut du Parc National en vue d'une protection intégrale de la
biodiversité avec une attention particulière sur le gorille de
plaine de l'Est, Gorilla berengei graueri, une espèce
endémique dans les forêts de l'Est du Congo. Ce parc couvrait
60.000 ha.
En 1975, il est élargi vers la basse altitude
(600-1200m) et couvre 600.000 ha. C'est en 1980, par la loi Congolaise n°
80/008 du 18/07/1980, que le PNKB fut inscrit sur la liste des sites du
patrimoine mondial de l'UNESCO (Shalukoma 2008).
2.1.1. Situation géographique
Le PNKB est situé à l'Est de la RDC
(voir figure 1) à cheval sur trois provinces : le Sud-Kivu, le Nord-Kivu
et le Maniema. Il est cerné par le territoire administratif de Kabare,
Walungu, Kalehe, Shabunda, Punia et Walikale. Géographiquement, il est
situé à 1°36'-2°27' de latitude Sud et
27°33'-28°46' de Longitude Est (Fischer, 1993).

Figure 2. Parc National de Kahuzi-Biega
2.1.2. Végétation
Selon Mühlenberg et al. (1995), près
de 90% du PNKB se trouve dans la
partie basse altitude (600 - 1200m) et est
dominé par la forêt ombrophile équatoriale très
humide. Elle possède une grande densité d'espèces
végétales dominées par les grands arbres ramifiés
aux sommets ; leurs pénombres ne favorisent pas le développement
des plantes herbacées. Dans la partie de haute altitude (15001300m), on
a la région des monts Kahuzi et Biega.
On y distingue 6 formes de végétations
:
- forêt ombrophile de montagne : en se basant
sur leur composition floristique et leur physionomie, on distingue
:
- forêt de basse montagne à
Pantadesma lebrunii et Lebrunia bushae avec comme
espèces caractéristiques : Gossera multinervis
- forêt de moyenne montagne à
Drypetes divsp., cet étage est beaucoup plus
hétérogène que le précédent. Newtonia
buchananii et Lebrunia bushae se rangent parmi les espèces
dominantes ;
- forêt de haute montagne à
Podocarpus, à partir de 2000 m apparaissent Podocarpus
milanjianus et Podocarpus usambarensis qui peuvent former des
véritables peuplements. Viennent ensuite les espèces comme
Ficalhoa laurifolia, Parinari excelsa et Parinari
holstii. Le sous-bois est riche en fougères.
- Aningeria adolfi-frederici se rencontre
à tous les étages.
- formation à Sinarundinaria alpina :
au Kahuzi, jusqu'à plus de 3000 m d'altitude, la strate
supérieure atteint 20 m de hauteur. Elle est composée uniquement
de Sinarundinaria alpina. Dans le sous-bois règne une
demi-obscurité peu favorable au développement d'une strate
inférieure dont le recouvrement ne dépasse jamais 15 à 20
%. On peut y trouver Pilea, Ceratomera et diverses Polypodiaceae
et des jeunes Podocarpus.
- Forêt mésophile de montagne : cette
forêt, à base de Newtonia buchananii, a jadis recouvert
d'importantes surfaces en bordure du Lac-Kivu. Saccagée par l'homme,
elle a pratiquement disparue du versant oriental, mais subsiste encore sur le
versant occidental entre 1400 et 1600 m d'altitude.
- Recrus et forêt secondaire de montagne : Aux
altitudes moyennes, c'est un recru à base de Pennisetum purpureum
qui s'installe après l'abattage de la
forêt aux altitudes plus élevées.
Les stades initiaux sont caractérisés par Lobelia
gibberoa. On note, cependant, des dominances locales de Croton
macrostachys, Dombeya goetzenii, Neoboutonia macrocalyx et Markhamia lutea.
Hagenia abyssinica est très abondant vers 2000m. Les forêts
secondaires que l'on rencontre aux altitudes supérieures à 2000m
sont dominées par Polyscias fulva et Macaranga neomildbraediana.
Xymalos monospora est l'espèce caractéristique du
sous-bois.
- Formation sclérophile : ces formations sont
nettement d'origine secondaire.
Elles apparaissent lors de l'abattage du manteau
forestier primitif dans les endroits où une faible pluviosité, un
relief accidenté ou un sol poreux crée des conditions de
sécheresse élevée. Agauria salicifolia, Hypericum
lanceolatum et Myrica salicifolia en sont les espèces les plus
courantes.
Aujourd'hui, la partie de haute altitude du PNKB est
constituée des îlots de forêts primaires et des plages des
forêts secondaires de terre ferme. On y remarque aussi des taches
(clairières ou trouées) dues à la dominance de
Sericostachys scandens et d'autres espèces qui lui
succèdent ; il s'agit de Mimulopsis solmsii, Mimulopsis
arborescens et plusieurs autres espèces des familles Urticaceae
(du genre Pilea), Acanthaceae (Barleria et
Hypoestes), Lamiaceae (Solanestemon), Malvaceae
(Triumfetta) ainsi que des fougères (Pteridium)
et autres herbacées. A plusieurs endroits, on observe l'émergence
de Sericostachys scandens allant jusqu'à la hauteur de grands
arbres et par la fin, cette dernière étouffe la plante en lui
privant d'une bonne absorption de l'oxygène. Au fil de temps, l'arbre
sèche et cède à la fin au poids des individus et
ramifications enchevêtrées de S. scandens. On peut
observer par ci par là, des taches parfois avec corridor et matrice,
donnant ainsi une physionomie non forestière à cette
phytocénose. Il faut aussi ajouter que dans cette partie, à part
les marais, aucun endroit n'est épargné par l'action
dévastatrice de Sericostachys scandens. Elle attaque tous les
habitats dans tous les biotopes, que ce soit une végétation
climax fixée sur une surface plane ou sur une pente et/ou dans les
vallées. Même dans les îlots de forêt primaire la
présence de cette espèce est probable, mais peut être
encore en adaptation. La présence de Sericostachys scandens est
plus intense dans la forêt secondaire (jeune ou vieille) et dans la
forêt de bambous (Balezi 2008).
2.1.3. Faune
L'espèce des gorilles de plaine de l'Est, une
espèce endémique des forêts de l'Est de la R.D.Congo
constitue le principal objectif de création du PNKB. A part cette
espèce, le PNKB renferme plusieurs grands mammifères dont les
éléphants, les buffles, les hylochères, les
potamochères, plusieurs espèces d'oiseaux, de petits
mammifères et des amphibiens. (Shalukoma op. cit.).
2.2. Matériel et Méthodes
d'étude
2.2.1. Matériel
La réalisation de ce travail a requis
l'utilisation d'un lot de matériels variés
d'après
leurs différents usages.
- une machette : pour tracer le transect ;
- un décamètre : pour la
détermination de la distance de chaque transect ;
- un GPS GARMIN (Oregon 300) : pour la prise des
coordonnées
géographiques et l'orientation du transect suivant
le Nord magnétique ;
- une latte graduée : pour tracer le relevé
;
- un canif : pour récolter les échantillons
;
- une loupe : pour observer les échantillons sur
le terrain ;
- des enveloppes en papier : pour conserver les
échantillons ;
- un sac: pour transporter les échantillons
;
- un microscope de lame et lamelles pour observer les
échantillons.
Signalons que notre matériel biologique comporte
différents échantillons récoltés qui
sont séchés et gardés dans le
laboratoire de Biologie de l'Université Officielle de
Bukavu.
2.2.2. Méthodes de travail
2.2.2.1. Zones d'échantillonnage
Après une prospection générale du
milieu d'étude, nous avons déterminé les zones à
étudier profondément ; ainsi, nos transects ont été
tracés de manière à ce qu'ils traversent différents
habitats dans les quatre secteurs où notre étude a
été réalisée. Il s'agit des secteurs Madiriri,
Mugaba, Tshibati et Tshivanga.
Pour réaliser ce travail, nous avons
utilisé la méthode de transect afin d'analyser qualitativement la
végétation, c'est-à-dire y relever les différents
types d'habitats et la méthode de relevé pour faire une analyse
quantitative des Bryophytes.
2.2.2.2. Analyse qualitative de la
végétation
La méthode de transect a été
utilisée. Elle consiste à tracer une ligne droite de 20 m de
largeur le long de laquelle les observations, les mesures et récoltes
sont réalisées. Nos transects ont été
orientés de manière à traverser divers types d'habitats
avec une dénivellation importante. A chaque changement d'habitat et/ou
après une dénivellation de 50 m, une description qualitative de
la végétation était faite. Les transects que nous avons
tracés sont :
Madiriri
- transect 1 NW 153° de 500 m de longueur
;
- transect 2 NE 22° de 500 m de longueur
;
Mugaba
-transect 3 NE 73° de 1000 m de longueur
;
Tshivanga
- transect 4 : N 33° de 500 m longueur ;
- transect 5 : SE 300° de 500 m de longueur
;
Tshibati
-transect 6 : SW 280° de 1000 m de
longueur.
Signalons que pour chaque secteur, 2 ha ont
été échantillonnés en tenant compte des variables
tels que :
types d'habitats : forêt primaire non
perturbée : définie selon White et al. (2000) comme
étant une forêt mixte à sous - bois ouvert avec de nombreux
arbres, une canopée élevée et continue, une
végétation au sol éparse constituée principalement
de buissons. Quelques espèces d'arbres nous ont servi pour
reconnaître cet habitat : Parinari excelsa, Chrysophyllum
gorungozanum, Strombosia schefflera, Symphonia globulifera, Carapa grandiflora,
etc.
Forêt primaire perturbée : avec les
mêmes arbres, mais ici, le sous-bois est formé par des
espèces comme : Sericostachys scandens, Rubus sp.,
Clerodendron sp. Forêt secondaire : selon White et al.
(2000), c'est une forêt avec grands arbres ou jeunes arbres montrant
des signes de dégradation humaine passée. Les espèces
suivantes nous ont servi comme indicatrices : Macaranga neomildbraediana,
Neoboutonia macrocalyx, Dombeya goetzenii, Alangium chinense,
etc.
Marais : c'est une zone humide ou inondée de
façon permanente dominée par les monocotylédones. Nous
avons observé des espèces comme : Cyperus latifolius,
Eulophia holsfalii et ces arbustes comme Hypericum
revolutum.
Forêt des bambous : c'est la forêt
dominée par le Sinarundinaria alpina.
La longueur et la couverture de la canopée ont
été aussi évaluées ; ainsi, nous avons
observé : une canopée fermée, une canopée ouverte
et une canopée intermédiaire (semi-fermée).
L'inclinaison de la pente était
évaluée en : pente faible, pente moyenne, pente forte et pente
nulle.
2.2.2.3. Analyse quantitative des Bryophytes
Nous avons utilisé la méthode des
relevés qui est une zone de surface connue dans laquelle chaque
élément est compté. Elle peut être circulaire,
carrée ou rectangulaire (White et al. op. cit.).
Nos relevés avaient chaque fois une surface de
20 cm x 20 cm. En nous basant sur le type de substrat, nous avons
observé :
· les espèces épiphytes :
espèces vivant sur des arbres ;
· les espèces saprophytes : vivant sur le
bois mort ;
· les espèces épilithes : vivant sur
la surface des rochers et pierres ;
· les espèces saprophytes ou terrestres :
vivant sur la surface du sol.
2.2.2.4. Récolte et conservation des
échantillons
Nous référant à Wigginston
(2004), nous avons récolté un nombre important de Bryophytes.
Notre attention a porté plus sur toutes les parties nécessaires
pour l'identification (le gamétophyte et le sporophyte). Deux
espèces affichant une petite différence étaient
récoltées différemment. Pour des échantillons
mouillés, il était nécessaire de les compresser doucement
pour dégager l'eau et les emballer dans deux ou trois enveloppes. Des
soins spéciaux ont été entrepris pour des
échantillons d'Anthécérotes à thalles car elles
perdent des caractères si elles sont séchées. Pendant la
récolte, chaque échantillon devrait avoir sur son emballage, des
indications relatives à ses caractères, à son habitat,
à son substrat. Il devait porte aussi un numéro.
Une fois séchés, les échantillons
sont alors placés dans une nouvelle enveloppe pliée, selon le
modèle décrit par Wigginston (Op. Cit.).
2.2.2.5. Types morphologiques de Bryophytes
Le type morphologique est l'aspect extérieur
que présente un être vivant. Ainsi, nous avons observé les
types suivants :
y' les mousses acrocarpes : les bourgeons
gamétangiaux se trouvent en position terminale. Les gamétanges
sont toujours accompagnés de paraphyses ;
y' les mousses pleurocarpes : les bourgeons
gamétangiaux sont en position latérale et sont placés sur
une seule feuille ;
y' les bryophytes à thalles : le
gamétophyte se présente en thalle, il n'y a pas de tige
;
y' les bryophytes à tige - feuillée : le
gamétophyte est un cormus se différentiant en tige et
feuilles.
2.2.2.6. Identification des échantillons
L'identification s'est faite par observation au
microscope des caractères de chacun des échantillons en se
référant aux caractères décrits dans des
clés : Min S. Pétiot (2003) et Wigginton (Op. Cit.). Les
échantillons ont été aussi envoyés à
Monsieur Fischer, un spécialiste des Bryophytes de University Koblenz-
Landau en Allemagne.
2.2.2.7. Mesure de DBH
Le DBH a été mesuré seulement sur
les plantes sur lesquelles nous avons récolté les bryophytes.
Pour cela, la mesure de la circonférence à la hauteur de 130 cm
(au niveau de la poitrine) a été effectuée, ainsi que la
distance à laquelle est situé l'échantillon par rapport au
sol.
2.2.2.8. Analyse des données
Le traitement des données issues du terrain
s'est réalisé suivant différents facteurs en utilisant les
logiciels Past et MVSP.
CHAPITRE 3. RESULTATS
3.1. Etude floristique
Les espèces inventoriées dans les quatre
différents secteurs (Madiriri, Mugaba, Tshibati, et Tshivanga) de la
partie haute altitude du PNKB sont rangées dans la liste floristique
présentée par ordre alphabétique des familles et des
espèces au sein des genres. Pour chaque espèce
inventoriée, les types de gamétophyte et de substrat ont
été donnés (tableau 2).
Tableau 2 : Liste floristique des espèces
inventoriée
Tf= tige feuillée, Th= thalle, Bm= saprophyte
sur bois mort, T=terrestre, R= rupicole (sur une roche), Epi= épiphytes,
Acro=acrocarpe, P= pleurocarpe.
|
N°
|
Familles, Genres et Espèces
|
Gamétophytes
|
Substrats
|
|
1
|
Adelanthaceae
|
|
|
|
1
|
Adelanthus sp.
|
Tf, Acro
|
Bm
|
|
2
|
Aneuraceae
|
|
|
|
2
|
Riccardia fastigiata (Lindenb) Berd
|
Th
|
T, R
|
|
3
|
Riccardia sp.
|
Th
|
T, R
|
|
3
|
Brachytheciaceae
|
|
|
|
4
|
Eurhynchium sp.
|
Tf, Pleuro
|
Epi,
|
|
4
|
Bryaceae
|
|
|
|
5
|
Brachymenium acuminatum Harv.
|
Tf, Acro
|
T
|
|
6
|
Bryum argenteum Hewd. Var lanatum
|
Tf, Acro
|
Bm.T
|
|
(P.Beauv.) Hampe
|
|
|
|
7
|
Bryum sp.
|
Tf, Acro
|
Bm, T
|
|
5
8
|
Calymperraceae Syrrhopodon sp.
|
Tf, Acro
|
Bm, Epi
|
|
6
|
Cyatothophorelaceae
|
|
|
|
9h
|
Cyathophorum africanum Dix.
|
Tf, Pleuro
|
Bm, Epi
|
|
7
|
Dicranaceae
|
|
|
|
10
|
Dicranoloma billardierei (Brid.)
Par.
|
Tf, Acro
|
Bm, Epi
|
|
11
|
Dicranum johnstonii Mitt.
|
Tf, Acro
|
Bm, Epi
|
|
8
|
Dilaenaceae
|
|
|
|
12
|
Jensenia spinosa (Lindeb. &Gott.)
Grolle
|
Th
|
R
|
|
9
|
Dumortieraceae
|
|
|
|
13
|
Dumortiera hirsuta (SW) Nees
|
Th
|
T
|
|
10
|
Fabroniaceae
|
|
|
|
14
|
Fabronia sp.
|
Tf, Pleuro
|
R
|
|
11
|
Fissidentaceae
|
|
|
|
15
|
Fissidens sp.
|
Tf, Acro
|
R
|
|
12
|
Frullaniaceae
|
|
|
|
16
|
Frullania angulata Mitt.
|
Tf, Acro
|
Bm, Epi
|
|
17
|
Frullania sp.
|
Tf, Acro
|
Bm, Epi
|
|
13
|
Funariaceae
|
|
|
|
18
|
Funaria hygrometrica Hewd.
|
Tf, Acro
|
R
|
|
14
|
Hookeriaceae
|
|
|
|
19
|
Cyclodictyon sp.
|
Tf, Acro
|
Epi, R
|
|
15
20
|
Hypnaceae
Hypnum cupressiforme Hedw.
|
Tf, Acro
|
Bm
|
|
16
|
Lejeuneaceae
|
|
|
|
21
|
Lejeunea sp.
|
Tf, Acro
|
R
|
|
22
|
Lejeuneaceae sp.
|
Tf, Acro
|
R
|
|
17
|
Lepidoziaceae
|
|
|
|
23
|
Lepidozia cupressina (SW) Lindend
|
Tf, Acro
|
Bm
|
|
18
|
Leucobryaceae
|
|
|
|
24
|
Athractilocarpus sp.
|
Tf, Pleuro
|
Bm, Epi
|
|
25
|
Orchobryum sp.
|
Tf, Pleuro
|
Bm, Epi
|
|
19
|
Leucodontaceae
|
|
|
|
26
|
Pterigonium sp.
|
Tf, Acro
|
Bm
|
|
20
|
Marchantiaceae
|
|
|
|
27
|
Marchantia polymorpha L.
|
Th
|
T
|
|
28
|
Marchantia planiloba STEPHANI
|
Th
|
T
|
|
21
|
Meteoriaceae
|
|
|
|
29
|
Pilotrichella ampullacea (C Müll)
Jaeg.
|
Tf, Pleuro
|
Bm, Epi
|
|
30
|
Pilotrichella sp.
|
Tf, Pleuro
|
Bm, Epi
|
|
22
|
Metzgeriaceae
|
|
|
|
31
|
Metzgeria agnewii Kuwahara
|
Tf, Pleuro
|
Epi
|
|
23
|
Lophoziaceae
|
|
|
|
32
|
Andrewsianthus sp.
|
Tf, Acro
|
Bm
|
|
24
33
34
|
Neckeraceae
Neckera submacrocarpa Dix. Porotrichium
sp.
|
Tf, Acro Tf, Acro
|
Bm, Epi R
|
|
25
|
Notothylaceae
|
|
|
|
35
|
Notothylas corbicularis
(Schwein.)Sull
|
Th
|
T
|
|
36
|
Phaeoceros laevis (L.) Prosk.
|
Th
|
T
|
|
26
|
Pallavîcînîaceae
|
|
|
|
37
|
Symphyogyna cfr podophylla
|
Th
|
Bm, Epi
|
|
(Thunb).Mont. &Nees.
|
|
|
|
27
|
Plagîochîllaceae
|
|
|
|
38
|
Plagiochilla bifara (SW)
Lindenb.
|
Tf, Acro
|
Bm, Epi
|
|
39
|
Plagiochilla sp.
|
Tf, Acro
|
Bm, Epi
|
|
28
|
Polytrîchîaceae
|
|
|
|
40
|
Polytrichium commune Hedw.
|
Tf, Acro
|
T
|
|
29
|
Pottîaceae
|
|
|
|
41
|
Desmatodon sp.
|
Tf, Acro
|
R
|
|
42
|
Tortula sp.
|
Tf, Acro
|
R
|
|
30
|
Pylaîsîadelphaceae
|
|
|
|
43
|
Pterogonidium sp.
|
Tf, Pleuro
|
Bm, Epi
|
|
31
|
Racopîlaceae
|
|
|
|
44
|
Racopilum africanum Mitt.
|
Tf, Acro
|
R
|
|
32
|
Radulaceae
|
|
|
|
45
|
Radula cfr evelynae Yamada
|
Tf, Acro
|
Epi
|
|
33
|
Ricciaceae
|
|
|
|
46
|
Riccia stricta (Lindenb) Berd
|
Th
|
T
|
|
34
|
Rhizogoniaceae
|
|
|
|
47
|
Pyrrhobryum spiniforme (Hedw.) Mitt.
|
Tf, Pleuro
|
Bm, Epi
|
|
35
|
Splachnaceae
|
|
|
|
48
|
Tayloria kilmandscharica
|
Tf, Acro
|
Epi
|
|
49
|
Inconnue sp.1
|
Tf, Acro
|
Bm, R
|
|
50
|
Inconnue sp.2
|
Tf, Acro
|
R, T
|
|
51
|
Inconnue sp.3
|
Tf, Acro
|
Bm
|
|
52
|
Inconnue sp.4
|
Tf, Acro
|
Bm
|
3.1. Analyse taxonomique des espèces
inventoriées
3.1.1. Classification taxonomique des espèces
inventoriées
Nous avons utilisé la systématique selon
Qiu et al. In Wolfrang (Op. Cit.). Tableau 3. Classification taxonomique des
espèces inventoriées
|
Embranchement
|
S/ Embran.
|
Classe
|
Sous-classe
|
Super-ordre
|
Ordre
|
Sous-ordre
|
Famille
|
Nbre de
genres
|
Nbre d'espèces
|
|
Marchantiophyta
|
|
Marchantiopsida
|
Marchantiidae
|
|
Marchantiales
|
Marchantiinae
|
Marchantiaceae
|
1
|
2
|
|
Dumortieraceae
|
1
|
1
|
|
Ricciales
|
|
Ricciaceae
|
1
|
1
|
|
Pallaviciniopsida
|
|
|
Pallaviciniales
|
|
Pallaviciniaceae
|
1
|
1
|
|
Jungermanniopsida
|
Jungermanniidae
|
Jungermannianæ
|
Jamessoniellales
|
|
Adelanthaceae
|
1
|
1
|
|
Lophoziales
|
Lophoziinae
|
Lophoziaceae
|
1
|
1
|
|
Lepidoziales
|
|
Lepidoziaceae
|
1
|
1
|
|
Lophoceleales
|
|
Phagiochilaceae
|
1
|
2
|
|
Radulales
|
|
Radulaceae
|
1
|
1
|
|
Jubulales
|
|
Frullaniaceae
|
1
|
2
|
|
Lejeuneaceae
|
2
|
2
|
|
Metzgeriidae
|
|
Aneurales
|
|
Aneuraceae
|
1
|
2
|
|
Metzgeriales
|
|
Metzgeriaceae
|
1
|
1
|
|
|
|
|
Bryophyta
|
Brophytina
|
Politrichiopsida
|
|
|
Polytrichiales
|
|
Politrichiaceae
|
1
|
1
|
|
Bryopsida
|
Funariidae
|
|
Funariales
|
|
Funariaceae
|
1
|
1
|
|
Dicraniidae
|
|
Dicranales
|
|
Calymperaceae
|
1
|
1
|
|
|
|
|
|
|
|
Dicranaceae
|
2
|
2
|
|
Fissidentaceae
|
1
|
1
|
|
Leucobryaceae
|
2
|
2
|
|
Pottiales
|
|
Pottiaceae
|
2
|
2
|
|
Bryiidae
|
|
Splachnales
|
|
Splachnaceae
|
1
|
1
|
|
|
|
|
|
Bryales
|
|
Bryaceae
|
2
|
3
|
|
Rhizogoniales
|
|
Rhizogoniaceae
|
1
|
1
|
|
Hypnodendrales
|
|
Racopilaceae
|
1
|
1
|
|
Hookeriales
|
|
Hukeriaceae
|
1
|
1
|
|
Hypnales
|
|
Brachytheciaceae
|
1
|
1
|
|
|
|
|
|
|
|
Fabroniaceae
|
1
|
1
|
|
Hypnaceae
|
1
|
1
|
|
Leucodontaceae
|
1
|
1
|
|
Meteoriaceae
|
1
|
2
|
|
Neckeraceae
|
2
|
2
|
|
Pylaisiaceae
|
1
|
1
|
|
Antocerotophyta
|
|
Antoceropsida
|
Notothylatidae
|
|
Notothyladales
|
|
Notothyladaceae
|
2
|
2
|
|
Total 3
|
1
|
6
|
7
|
1
|
22
|
2
|
35
|
42
|
52
|
Les tableaux 2 et 3 montrent que 48 espèces de
Bryophytes ont été inventoriées. Elles sont reparties dans
42 genres, 35 familles, 22 ordres, 7 sous classes, 6 classes et 3
embranchements (Marchantiophyta, Bryophyta et
Antocerotophyta). L'embranchement des Marchantiophyta est
celui qui a plus des classes (Marchantiopsida, Pallaviciniopsida et
Jungermanniopsida) ; il est suivi des Bryopsida qui ont 2
classes (Politrichiopsida et Bryopsida). L'embranchement des
Antocerotophyta n'a qu'une seule classe (Anteceropsida). La
classe des Bryopsida est celle qui a 3 sous-classes (Funariidae,
Dicraniidae et Bryiidae). La classe des Jungermanniopsida
compte 2 sous-classes (Jungermanniidae et Metzgeriidae)
; les autres classes n'ont qu'une sous - classe chacune. La sous - classe des
Bryiidae est celle qui a beaucoup d'ordres (7). Elle est suivie des
Jungermanniidae avec 6 ordres. L'ordre Hypnales est celle qui
a beaucoup de familles (7) ; viennent ensuite les Dicranales avec 4
familles. La famille Bryaceae est celle qui a beaucoup
d'espèces (3)
3.1.2. Brève description morphologique de certaines
espèces inventoriées
1. Dumortiera hirsuta (SW) Nees
Plante à gamétophyte thalloïde,
avec des chambres d'air (corbeille), présence des rhizoïdes sur la
surface ventrale des gamétangiophores.
2. Riccia stricta (Lindenb) Berd
Plante à gamétophyte thalloïde,
à ramification dichotomique, de couleur vert clair à verte
brillante, thalle linéaire comme un ruban de 3 à 4 cm de long et
0,5 mm de large, sur un sol mouillé ou flottant sur l'eau.
3. Symphyogyna cfr podophylla (Thunb) Mont.
&Nees
Plante à gamétophyte
érigé, provenant d'un rhizome ou stipe, à
archégones groupés sur la surface dorsale délimité
par un involucre.
4. Plagiochila bifara (SW)
Lindenb
Plante robuste formant un tapis, tige
feuillée, simple ou ramifiée, de 9 cm de long. Feuilles à
lobes de 2,75 cm de long et de 2,75 cm de large.
5. Adelanthus sp.
Plante à gamétophyte constitué
d'une tige feuillée non lobée et bidentée.
6. Andrewsianthus sp.
Plante à tige feuillée, feuille
poussant sur un axe flagiliforme, feuilles très petites.
7. Radula cfr evelynae Yamada
Petite plante verdâtre à chocolate,
à petite tige prostrée de 1,5 cm de long,
ramifiée.
8. Frullania angulata Mitt.
Plante robuste rougeâtre, jaunâtre
à noirâtre ; tige prostrée de 3 cm de long à
plusieurs branches ; feuilles petites, ovales, imbriquées,
entières avec amphigastre.
9. Frullania sp.
Plante à tige feuillée
légèrement ramifiée, feuilles petites mais subovales,
entières et avec amphigastre.
10. Polytrichium commune Hedw. Var
commune
Plante large de #177; 20 cm de long, tige
feuillée à feuilles linéaires à
lancéolées, sommet acuminé.
11. Dicranum johnstonii Mitt.
Plante large, robuste, vert - jaunâtre au
dessus. Tige érigée, simple ou ramifiée ; feuilles
lancéolées de 10 mm de long, à marge
irrégulièrement denticulée.
12. Syrrhopodon sp.
Plante robuste, érigée, tige de 4
à 5cm de long, ramifiée, à feuilles
lancéolées.
13. Bryum argenteum Hedw. Var. lanatum
(P.Beauv.) Hampe
Plante de couleur argentée, tige
feuillée de #177;1 cm de long souvent avec des branches à la
base.
14. Bryum sp.
Plante de couleur verte ; tige feuillée de
#177; 2 cm de long souvent avec des branches à la base.
15. Hypnum cupressiforme Hedw
Plante mince à robuste, verte, tige
prostrée à subérigée de 3 à 4 cm de long ;
tige feuillée à petites feuilles de 2,5 mm de long ; branches
prostrées à ascendantes.
16. Pilotrichella ampullacea (C.Mull)
Jaeg
Plante robuste, verte, la plupart de fois pendantes
sur des branches des arbres. Feuilles de 1,5 mm de long, auriculées
à la base ou fortement cordées, sommet
mucroné.
17. Pilotrichella sp.
Plante robuste, verte. Tige feuillée, feuilles
de 3mm de long fortement cordées.
18. Neckera submacrocarpa Dix
Plante brillante, vert pâle ; tige et branches
aplaties, la tige primaire rampante, la tige secondaire allongée,
aplatie souvent pendante. Feuilles fortement ondulées, transversalement
asymétrique, oblong - lancéolées.
19. Lepidozia cupressina (S.W.)
Lindenb
Petite plante verte pâle à
vert-jaunâtre, à branches rampantes. Feuilles très petites.
Plante formant une touffe sur des roches et des bois morts.
20. Riccardia fastigiata (Lehm).
Trev.
Plante thalloïde, vert sombre à branches
subopposées.
21. Riccardia sp.
Plante petite à robuste, thalloïde de
couleur vert clair à branches subopposées.
22. Metzgeria agnewii Kuwahara
Petite plante thalloïde, dichotomiquement
ramifiée.
23. Marchantia polymorpha L.
Plante thalloïde, verte, dichotomiquement
ramifiées de 5 à 6 cm de long et de 6 à 8 cm de large ;
chambres d'air (corbeilles) présentes, thalles à formes diverses
et charnus.
24. Marchantia planiloba L.
Plante thalloïde, verte, dichotomiquement
ramifiée, chambres d'air (corbeilles) présentes, thalles presque
vert pâle.
25. Notothylas corbicularis (Schwein.)
Sull.
Plante thalloïde de 5 à 7 mm, sporophytes
en paires fusiforme à ellipsoïdale.
26. Lejeunea sp.1
Plante petite, verte, tige prostrée de
quelques millimètres de long ; feuilles imbriquées, ovales
à sommet rond, marges entières.
27. Lejeunea sp.2
Plante petite à tige érigée ;
àfeuilles imbriquées et à marges
entières.
28. Jensenia spinosa (Lindenb. &Gott.)
Grolle
Plante thalloïde, de forme
irrégulière, jeune plante simple, dichotomiquement
ramifiée, plante adulte avec stipe à portion
palmée.
29. Phaeoceros laevis (L.)
Prosk.
Plante thalloïde de couleur verte,
dichotomiquement lobée de manière irrégulière.
Capsules longues et cylindriques.
30. Funaria hygrometrica Hedw.
Plante petite, solitaire, parfois ramifiée
à la base ; feuilles contortées, lancéolées
à ovales formant une rosette terminale.
31. Cyathophorum africanum Dix
Plante robuste, vert pétrole. Tige principale
simple, subérigée, de 3,4 cm de long ; feuilles
asymétriques.
32. Brachymenium acuminatum
Harv.
Petite plante de couleur vert-pétrole. Tige de
7 mm de long, ramifiée. Feuilles érigées, ovales à
acuminées.
33. Plagiochila sp.
Plante petite, irrégulièrement
ramifiée ; feuilles ovales de couleur verte.
34. Atractylocarpus sp.
3.1.3. Types de gamétophytes des espèces
inventoriées
Plante dicranoide, costa 1/2 - 2/3 de la base
de la feuille. Seta flexible.
35. Orchobryum sp.
Plante à gamétophyte ressemblant à
un petit Leucobryum à capsules
hémisphériques.
36. Fissidens sp.
Plante petite en touffe, verte, à seta petit,
capsule exserte, érigée et symétrique.
37. Tortula sp.
Plantes petites, feuilles vertes, ovales, marge
entière à péristome variante.
38. Desmatodon sp.
Plante petite de couleur verdâtre, à
feuille ovale, marge entière.
39. Tayloria kilmandscharica
Plante vert jaunâtre, en touffe ; feuille ovale,
à base décurrente, marge entière. Seta mince et
flexible.
40. Pyrrhobryum spiniforme (Hedw.)
Mitt.
Plante petite, en touffe ; tige feuillée,
à feuille longuement spiralée.
41. Cyclodictyon sp.
Plante petite, verte ; à feuille courtement
acuminée.
42. Fabronia sp.
Plante petite ; tige irrégulièrement
ramifiée, avec une partie de branche subascendante. Feuille
entièrement marginée.
43. Eurhychium sp.
Plante petite, à gamétophyte
érigé, régulièrement ramifiée.
44. Pterogonidium sp.
Plante verte à tige feuillée ; tige
irrégulièrement ramifiée ; feuille avec une marge
entière. Capsule érigée.
45. Porotrichium sp.
Plante large, à tige érigée,
irrégulièrement ramifiée. Feuilles superficiellement
rangées à huit.
46. Racopilum africanum Mitt
Plante robuste, ramifiée de couleur vert
foncée, tige longue. Feuilles nombreuses dimorphes.
47. Pterogonium sp.
Plante petite, à tige feuillée, de couleur
verte, feuille avec marge entière.
48. Inconnue sp.1
Plante petite ; de tige de #177;1,5cm de long, poilue ;
à feuilles lancéolées, séparées sur la
tige.
49. Inconnue sp.2
Plante à tige ramifiée ; à feuilles
lancéolées, espacées ; longuement
acuminées.
50. Inconnue sp.3
Plante robuste ; à tige provenant d'un rhizome ;
à feuilles groupées au sommet, mais légèrement
séparées à la base.
51. Inconnue sp.4
Plante verte à jaunâtre à la partie
supérieure ; feuilles groupées au dessus ; tige densément
poilue à la base.
3.1.4. Diversité des secteurs
prospectés
Tableau 4 : Spectre de types de
gamétophyte
|
Gamétophytes
|
Nbres d'espèces
|
Pourcentage (%)
|
|
Tige feuillée (Tf)
|
42
|
81
|
|
Acrocarpes (Acro)
|
32
|
62
|
|
Pleurocarpes (Pleuro)
|
10
|
19
|
|
Thalle (Th)
|
10
|
19
|
|
Total
|
52
|
100,00
|

Série1;
Pleurocarpes
(Pleuro); 10;
19%
10; 19%
Types de gamétophytes des
espèces
Série1; Thalle
Acrocarpes (Acro) Pleurocarpes (Pleuro) Thalle
(Th)
(Th);
inventoriées
Série1;
Acrocarpes
(Acro); 32; 62%
Figure 2. Types de gamétophytes des espèces
inventoriées
L'interprétation du tableau 6 et de la figure 2
montre une dominance des Bryophytes à tige feuillée avec 42
espèces (soit 81 %) dont les Bryophytes acrocarpes ont 32 espèces
(soit 62%) et les Pleurocarpes 10 espèces (soit 19 %), les Bryophytes
à thalle n'ont aussi que 10 espèces (soit 19 %).
Tableau 5. Diversité des secteurs
prospectés
|
Madiriri
|
Tshibati
|
Mugaba
|
Tshivanga
|
|
Dominance D
|
0,04545
|
0,03571
|
0,04762
|
0,03704
|
|
Shannon H
|
3,091
|
3,332
|
3,045
|
3,296
|
|
Simpson 1-D
|
0,9545
|
0,9643
|
0,9524
|
0,963
|
Dans le tableau 5, on remarque la diversité
pour chaque secteur et c'est le secteur Tshibati qui a une grande
diversité et cela par les indices de diversité calculés
à savoir Shannon et Simpson.
Tableau 6. Similarité et degré de distance
entre secteurs
|
Madiriri
|
Tshibati
|
Mugaba
|
Tshivanga
|
|
Madiriri
|
1
|
|
|
|
|
Tshibati
|
0,40909
|
1
|
|
|
|
Mugaba
|
0,38095
|
0,47619
|
1
|
|
|
Tshivanga
|
0,36364
|
0,48148
|
0,66667
|
1
|
On voit bien une bonne similarité entre les
différents secteurs, chacun se ressemble à 10 %, mais il y a une
bonne différence entre les secteurs. Par exemple, les secteurs Tshibati
et Madiriri ont un degré de distance d'à peu près 41 %
(voir tableau 6).
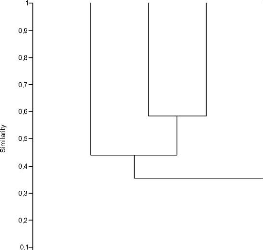
Figure 3. Similarité entre secteurs
La lecture de la figure 3 montre la similarité
entre les secteurs. Suivant la loi de Curtis, on voit que les secteurs Madiriri
et Tshivanga ont une similarité de 60 % tandis que Tshibati et les deux
précités ci-haut ont une similarité d'à peu
près 41 %. Mugaba et tous les autres secteurs ont une similarité
de 35 %.
3.2. Ecologie des espèces des Bryophytes
étudiées
Les espèces étudiées ont
été trouvées dans plusieurs habitats et sur plusieurs
types de substrats. L'analyse de l'habitat et du substrat nous a permis de bien
connaître l'écologie de chaque espèce. Ensuite, la tranche
altitudinale pour chaque espèce a été aussi
déterminée.
Tableau 7. Types d'habitats parcourus dans la partie de
haute altitude du PNKB Légende : F.S. = Forêt secondaire, F.P. =
Forêt primaire
|
N
°
|
Type d'habitats
|
Secteur
|
Transects
|
Sous- bois
|
Hauteur de la végétation
|
Couverture Canopée
|
Topographie (Pente)
|
Altitude (m)
|
|
1
|
F.S. à Markhamia lutea
|
Madiriri
|
1
|
Fermé
|
#177;30 m
|
Fermée
|
Moyenne
|
2007
|
|
2
|
F.S. à Croton et Alangium
|
Madiriri
|
1
|
Fermé
|
#177;25 m
|
Fermée
|
Forte
|
2034
|
|
3
|
Clairière à Dombeya et
Alangium
|
Madiriri
|
1
|
Ouvert
|
20 m
|
Ouvert
|
Forte
|
2060
|
|
4
|
F.S. à Dombeya et
Myrianthus
|
Madiriri
|
1
|
Fermé
|
15 m
|
Fermée
|
Forte
|
2110
|
|
5
|
F.P. à Chrysophylum,
Strombosia et Entadophragma
|
Madiriri
|
1
|
Fermé
|
40 m
|
Semi - fermée
|
Forte
|
2160
|
|
6
|
F.S. à Macaranga et
Myrianthus
|
Madiriri
|
1
|
Semi -
fermé
|
15 m
|
Semi - fermée
|
Forte
|
2189
|
|
7
|
F.S. à Macaranga et
Polycias
|
Madiriri
|
1
|
Semi -
fermé
|
20 m
|
Semi - fermée
|
Forte
|
2241
|
|
8
|
F.P. à Chrysopyllum, Strombosia
et Parinari
|
Madiriri
|
1
|
Fermé
|
30 m
|
Semi - fermée
|
Faible
|
2263
|
|
9
|
F.S. à Mytragina et
Sapium
|
Madiriri
|
2
|
Fermé
|
25 m
|
Fermée
|
nulle
|
2019
|
|
10
|
F.P. à Chrysophylum et
Strombosia
|
Madiriri
|
2
|
Fermé
|
35 m
|
Fermée
|
Moyenne
|
2030
|
|
11
|
F.P. à Chrysophylum et
|
Madiriri
|
2
|
Ouvert
|
35 m
|
Fermée
|
Forte
|
2016
|
|
Strombosia
|
|
|
|
|
|
|
|
|
12
|
F.P. à Chrysophylum
|
Madiriri
|
2
|
Fermé
|
35 m
|
Fermée
|
Forte
|
2084
|
|
13
|
F.S. à Macaranga
|
Tshibati
|
3
|
Fermé
|
20 m
|
Semi - fermée
|
Moyenne
|
2015
|
|
14
|
F.S. à Macaranga et
Sapium
|
Tshibati
|
3
|
Fermé
|
25 m
|
Semi - fermée
|
Moyenne
|
2020
|
|
15
|
Clairière à Dombeya et
Sapium
|
Tshibati
|
3
|
Fermé
|
20 m
|
Ouverte
|
Moyenne
|
2033
|
|
16
|
F.S. à Sapium
|
Tshibati
|
3
|
Fermé
|
25 m
|
Semi - fermée
|
Moyenne
|
2041
|
|
17
|
Marais à Cyperus latifolius
|
Tshibati
|
3
|
Fermé
|
5 m
|
Ouverte
|
Nulle
|
2030
|
|
18
|
F.S. à Dombeya et
Neoboutonia
|
Tshibati
|
3
|
Fermé
|
10 m
|
Ouverte
|
Moyenne
|
2016
|
|
19
|
F.S. à Macaranga
|
Tshibati
|
3
|
Fermé
|
20 m
|
Fermée
|
Forte
|
2278
|
|
20
|
F.P. à Ficalhoa
|
Mugaba
|
4
|
Semi -
fermé
|
15 m
|
Semi - fermée
|
Nulle
|
2205
|
|
21
|
F.P. à Symphonia et
Carappa
|
Mugaba
|
4
|
Semi -
fermé
|
40 m
|
Fermée
|
Moyenne
|
2215
|
|
22
|
F.S. à Macaranga
|
Mugaba
|
4
|
Fermé
|
40 m
|
Fermée
|
Forte
|
2222
|
|
23
|
F.P. à Chrysophylum et
Symphonia
|
Mugaba
|
4
|
Fermé
|
40 m
|
Fermée
|
Forte
|
2330
|
|
24
|
F.S. à Neoboutonia et
Dombeya
|
Tshivanga
|
5
|
Fermé
|
20 m
|
Semi - fermée
|
Faible
|
2175
|
|
25
|
Galerie forestière à
Cyathea
|
Tshivanga
|
5
|
Fermé
|
20 m
|
Ouverte
|
Forte
|
2194
|
|
26
|
F.S. à Macaranga et
Syzygium
|
Tshivanga
|
5
|
Fermé
|
25 m
|
Semi - fermée
|
Faible
|
2284
|
|
27
|
F.S. à Macaranga et
Harungana
|
Tshivanga
|
5
|
Fermé
|
30 m
|
Fermée
|
Forte
|
2297
|
|
28
|
F.S. à Macaranga et
Dombeya
|
Tshivanga
|
6
|
Fermé
|
20 m
|
Semi - fermée
|
Nulle
|
2146
|
|
29
|
F.P. à Prunus
|
Tshivanga
|
6
|
Fermé
|
30 m
|
Semi - fermée
|
Moyenne
|
2155
|
|
30
|
F.S. à Hagenia
|
Tshivanga
|
6
|
Ouvert
|
13 m
|
Fermée
|
Moyenne
|
2186
|
|
31
|
F.S. à Macaranga
|
Tshivanga
|
6
|
Fermé
|
20 m
|
Fermée
|
Moyenne
|
2208
|
La lecture du tableau 7 montre que 31
différents habitats ont été observés dans les 4
secteurs (Madiriri, Mugaba, Tshibati et Tshivanga). Pour chaque habitat, il a
été tenu compte de la hauteur et la couverture de la
canopée, le type de sous-bois, la topographie, ainsi que
l'altitude.
3.2.2. Distribution des bryophytes selon les types
d'habitat
Tableau 8 : Liste des Bryophytes inventoriées
selon les différents habitats
|
N°
|
ESPECES
|
F.S.
|
F.P.
|
Marais
|
Clairière
|
Galerie forestière
|
|
1
|
Adelanthus sp.
|
1
|
1
|
0
|
0
|
0
|
|
2
|
Andrewsianthus kilmandscharica
|
1
|
0
|
0
|
0
|
0
|
|
3
|
Athractiloparcus sp.
|
1
|
1
|
0
|
0
|
1
|
|
4
|
Brachymenium acuminatum
|
0
|
1
|
0
|
1
|
0
|
|
5
|
Bryum argenteum
|
1
|
0
|
0
|
0
|
0
|
|
6
|
Bryum sp.
|
1
|
1
|
0
|
1
|
0
|
|
7
|
Cyathoforela africana
|
1
|
0
|
0
|
1
|
0
|
|
8
|
Cyclodictyon sp.
|
0
|
0
|
0
|
1
|
1
|
|
9
|
Desmatodon sp.
|
1
|
0
|
0
|
0
|
0
|
|
10
|
Dicranoloma billartieri
|
0
|
0
|
0
|
1
|
0
|
|
11
|
Dicranum johnstonii
|
1
|
1
|
0
|
0
|
0
|
|
12
|
Dumortiera hirsuta
|
1
|
0
|
0
|
0
|
0
|
|
13
|
Fabronia sp.
|
1
|
0
|
0
|
0
|
0
|
|
14
|
Fissidens sp.
|
1
|
0
|
0
|
0
|
0
|
|
15
|
Funaria hygrometrica
|
1
|
0
|
0
|
0
|
0
|
|
16
|
Frullania angulata
|
1
|
1
|
0
|
1
|
0
|
|
17
|
Frullania sp.
|
1
|
0
|
0
|
0
|
0
|
|
18
|
Eurhynchium africanum
|
1
|
0
|
0
|
0
|
0
|
|
19
|
Hypnum cupressiforme
|
1
|
1
|
0
|
0
|
0
|
|
20
|
Jensenia spinosa
|
1
|
0
|
0
|
0
|
0
|
|
21
|
Lejeunea sp.
|
1
|
0
|
0
|
0
|
0
|
|
22
|
Lejeuneaceae sp.
|
1
|
0
|
0
|
1
|
0
|
|
23
|
Lepidozia cupressina
|
1
|
0
|
0
|
0
|
0
|
|
24
|
Marchantia planiloba
|
1
|
0
|
0
|
0
|
0
|
|
25
|
Marchantia polymorpha
|
0
|
0
|
1
|
0
|
0
|
|
26
|
Metzgeria agnewii
|
0
|
1
|
0
|
0
|
0
|
|
27
|
Neckera submacrocarpa
|
1
|
0
|
0
|
0
|
0
|
|
28
|
Notothylas corbiculata
|
1
|
0
|
1
|
0
|
0
|
|
29
|
Pilotrichella ampulacea
|
1
|
0
|
0
|
0
|
0
|
|
30
|
Pilotrichella sp.
|
1
|
10
|
0
|
0
|
0
|
|
31
|
Plagiochila bifaria
|
1
|
0
|
0
|
0
|
0
|
|
32
|
Plagiochila sp.
|
1
|
0
|
0
|
1
|
1
|
|
33
|
Orchobryum sp.
|
1
|
0
|
0
|
0
|
0
|
|
34
|
Polytrichium commune
|
1
|
0
|
0
|
1
|
1
|
|
35
|
Pterogonidium sp.
|
1
|
1
|
0
|
1
|
0
|
|
36
|
Pterigonium sp.
|
1
|
0
|
0
|
0
|
0
|
|
37
|
Porotrichium sp.
|
1
|
1
|
0
|
0
|
0
|
|
38
|
Pyribryum spiniforme
|
1
|
1
|
0
|
0
|
1
|
|
39
|
Racopilum africanum
|
1
|
0
|
0
|
0
|
0
|
|
40
|
Riccardia fastigiata
|
1
|
1
|
0
|
0
|
0
|
|
41
|
Riccardia sp.
|
0
|
0
|
0
|
1
|
0
|
|
42
|
Riccia stricta
|
0
|
0
|
1
|
0
|
0
|
|
43
|
Radula cfr evelynae
|
1
|
1
|
0
|
1
|
0
|
|
44
|
Symphyogyna cfr podophylla
|
1
|
1
|
0
|
0
|
0
|
|
45
|
Syrrhopodon sp.
|
1
|
1
|
0
|
0
|
0
|
|
46
|
Tayloria kilmandscharica
|
|
1
|
0
|
0
|
0
|
|
47
|
Phaeoceros laevis
|
1
|
0
|
0
|
0
|
0
|
|
48
|
Tortula sp.
|
1
|
0
|
0
|
1
|
0
|
|
49
|
Inconnue sp.1
|
1
|
1
|
0
|
0
|
0
|
|
50
|
Inconnue sp.2
|
1
|
1
|
0
|
0
|
0
|
|
51
|
Inconnue sp.3
|
0
|
0
|
0
|
1
|
0
|
|
52
|
Inconnue sp.4
|
1
|
0
|
0
|
0
|
0
|
Il ressort du tableau 8 qu'aucun habitat n'a eu la
présence de toutes ces espèces. Six espèces se rencontrent
dans trois habitats à la fois (Bryum sp., Frulania
angulata, Pterogonidium
sp. et
Radula cfr evelynae) dans la forêt secondaire, forêt primaire
et clairières. L'espèce Athractiloparcus sp. est
à la fois dans la forêt secondaire, forêt primaire et
galerie forestière. Par ailleurs, l'espèce Plagiochila
sp. est dans la forêt secondaire, clairière et galerie
forestière.
Les forêts secondaires ont plus des bryophytes
spécifiques avec 18 espèces (soit 34 %). Seule l'espèce
Tayloria kilmandscharica est spécifique pour les forêts
primaires (soit 2 %). Deux espèces (soit 4 %) dont Marchantia
polymorpha et Riccia stricta n'ont été
trouvées que dans les marais. Deux autres espèces sont
rencontrées seulement dans les clairières (Dicranoloma
billartieri et Riccardia sp.).
3.2.3. Types de substrats des espèces
inventoriées
Tableau 9. Liste des plantes hôtes et leurs types
Morphologiques (TM)
|
N°
|
Plantes hôtes
|
TM
|
Famille
|
|
1
|
Chrysophylum gorungosanum
|
A
|
Sapotaceae
|
|
2
|
Dombeya goetzenii
|
A
|
Malvaceae
|
|
3
|
Ficalhoa laurifolia
|
A
|
Theaceae
|
|
4
|
Macaranga neomildbraediana
|
A
|
Euphorbiaceae
|
|
5
|
Mytragina stipulosa
|
A
|
Rubiaceae
|
|
6
|
Neoboutonia macrocalyx
|
A
|
Euphorbiaceae
|
|
7
|
Pleiocarpa pychnanta
|
Arb
|
Apocynaceae
|
|
8
|
Polyscias fulva
|
A
|
Apiaceae
|
|
9
|
Sapium ellipticum
|
A
|
Euphorbiaceae
|
|
10
|
Sericostachys scandens
|
L
|
Amaranthaceea
|
|
11
|
Strombosia schefflera
|
A
|
Olacaceae
|
|
12
|
Tabernaemontana johnstonii
|
A
|
Apocynaceae
|
|
13
|
Xymalos monospora
|
A
|
Monimiaceae
|
|
14
|
Zanthophyllum macrophylla
|
Arb
|
Rutaceae
|
Il ressort du tableau 9 que les Bryophytes
récoltées (espèces épiphytes) avaient comme
substrat 14 espèces des plantes vasculaires. Parmi ces espèces,
une est lianescente, deux sont des arbustes et les autres sont des arbres. La
famille Euphorbiaceae en a eu à elle seule 3 espèces,
les Apocynaceae sont représentées par 2 espèces.
Les autres familles sont représentées par une seule
espèce.Ici, nous devons rappeler que les autres espèces sont
terrestres (voir tableau 8).
Tableau 10 : Classe de diamètre des individus
des plantes hôtes à DBH ~10 cm par
secteur.
|
Classes de
|
Madiriri
|
Tshibati
|
Mugaba
|
Tshivanga
|
|
diamètre
|
Nbre
|
%
|
Nbre
|
%
|
Nbre
|
%
|
Nbre
|
%
|
|
10 - 20
|
2
|
15,4
|
0
|
0
|
2
|
28,6
|
0
|
0
|
|
21 - 30
|
2
|
15,4
|
2
|
28,5
|
3
|
42,8
|
2
|
40
|
|
31 - 40
|
5
|
38,4
|
3
|
42,9
|
1
|
14,3
|
3
|
60
|
|
41 - 50
|
0
|
0
|
1
|
14,3
|
0
|
0
|
0
|
0
|
|
51 - 60
|
1
|
7,7
|
1
|
14,3
|
0
|
0
|
0
|
0
|
|
61 - 70
|
2
|
15,4
|
0
|
0
|
1
|
14,3
|
0
|
0
|
|
71 - 80
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
81 - 90
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
91 - 100
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
101 - 110
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
111 - 120
|
1
|
7,7
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Total
|
13
|
100
|
7
|
100
|
7
|
100
|
5
|
100
|
Tshivanga Mugaba Tshibati Madiriri
Classes de diamètre des individus des plantes
hôtes des Bryophytes
Nombre d'individus
Figure 4. Classes de diamètre des individus des
plantes hôtes des Bryophytes Le tableau 10 et la figure 4 montrent que
les classes de diamètres des individus des plantes hôtes des
Bryophytes à Madiriri, Mugaba, Tshivanga et Tshibati sont comprises
entre 10 - 20 cm et 111 - 120 cm. Aucun individu n'a été
trouvé dans les classes de 71 - 80 cm à 101 - 110 cm. C'est la
classe 31 - 40 cm qui a beaucoup d'individus, 12 sur 32 individus (soit 37,5%).
Cette classe est représentée dans les quatre secteurs. Elle est
constituée des espèces d'arbres suivantes :
Chrysophylum
gorungosanum, Dombeya goetzenii, Macaranga
neomildbraediana, Neoboutonia macrocalyx, Polyscias fulva, Sapium ellipticum,
Strombosia schefflera et Tabernaemontana johnstonii. Elle est
suivie de la classe 21 - 30 cm avec 9 individus, (soit 28,1%) ; elle est
constituée des espèces suivantes : Dombeya goetzenii, Ficalhoa
laurifolia, Macaranga neomildbraediana, Mytragina stipulosa, Neoboutonia
macrocalyx, Zanthophyllum macrophylla et Polyscias fulva. Cette
classe est aussi représentée dans les quatre secteurs. La classe
111 - 120 cm est la moins représentée car ne se trouve
qu'à Madiriri et représentée par une seule espèce
et un seul individu (Chrysophyllum gorungosanum).
C'est le secteur Madiriri qui a beaucoup d'individus
hôtes à DBH ~10 cm avec 13 individus (soit
40,6%), ces individus sont comprises dans 5.
Tableau 11. Spectre de types de substrat
|
Substrat
|
Nbres d'espèces
|
Pourcentage (%)
|
|
Saprophytes (Bm)
|
8
|
15,38
|
|
Epiphytes (Epi)
|
4
|
7,69
|
|
Ripicoles (R)
|
8
|
15,38
|
|
Terrestres (T)
|
7
|
13,46
|
|
Saprophytes et épiphytes
|
16
|
30,79
|
|
Saprophytes et terrestres
|
2
|
3,85
|
|
Saprophytes et ripicoles
|
1
|
1,92
|
|
Epiphytes et ripicoles
|
3
|
5,77
|
|
Ripicoles et terrestres
|
3
|
5,77
|
|
Total
|
52
|
100,00
|
Il ressort du tableau 11 que les Bryophytes qui sont
à la fois Saprophytes et épiphytes sont celles qui dominent avec
16 espèces (soit 30,79 %). Elles sont suivies des espèces
ripicoles et saprophytes qui ont chacune 8 espèces, soit 15,33 %.
Viennent ensuite les espèces terrestres qui sont à 7, soit 13,46
%, les autres groupes sont moins représentés.
3.2.4. Distribution des Bryophytes selon les secteurs
Tableau 12 : Liste des Bryophytes inventoriées par secteur
|
Noms Scientifiques
|
Madiriri
|
Tshibati
|
Mugaba
|
Tshivanga
|
|
1
|
Adelanthus sp.
|
1
|
0
|
0
|
0
|
|
2
|
Andrewsianthus
|
0
|
1
|
0
|
1
|
|
3
|
Athractiloparcus sp.
|
1
|
0
|
1
|
1
|
|
4
|
Brachymenium acuminatum
|
0
|
1
|
1
|
1
|
|
5
|
Bryum argenteum
|
0
|
1
|
1
|
1
|
|
6
|
Bryum sp.
|
1
|
0
|
0
|
1
|
|
7
|
Cyathoforum sp.
|
0
|
1
|
0
|
1
|
|
8
|
Cyclodictyon sp.
|
0
|
1
|
0
|
1
|
|
9
|
Desmatodon sp.
|
1
|
0
|
1
|
0
|
|
10
|
Dicranoloma billartieri
|
1
|
1
|
0
|
0
|
|
11
|
Dicranum johnstonii
|
1
|
1
|
1
|
1
|
|
12
|
Dumortiera hirsuta
|
0
|
0
|
1
|
0
|
|
13
|
Fabronia sp.
|
0
|
0
|
0
|
1
|
|
14
|
Fissidens sp.
|
0
|
0
|
0
|
1
|
|
15
|
Funaria hygrometrica
|
0
|
1
|
0
|
0
|
|
16
|
Frullania angulata
|
0
|
1
|
1
|
1
|
|
17
|
Frullania sp.
|
1
|
0
|
0
|
0
|
|
18
|
Eurhynchium africanum
|
1
|
0
|
0
|
0
|
|
19
|
Hypnum cupressiforme
|
1
|
0
|
0
|
1
|
|
20
|
Jensenia spinosa
|
0
|
1
|
0
|
0
|
|
21
|
Lejeunea sp.
|
1
|
0
|
1
|
0
|
|
22
|
Lejeuneaceae sp.
|
0
|
1
|
0
|
1
|
|
23
|
Lepidozia cupressina
|
0
|
1
|
0
|
0
|
|
24
|
Marchantia planiloba
|
0
|
0
|
0
|
1
|
|
25
|
Marchantia polymorpha
|
0
|
1
|
0
|
0
|
|
26
|
Metzgeria agnewii
|
0
|
0
|
1
|
1
|
|
27
|
Neckera submacrocarpa
|
0
|
1
|
0
|
1
|
|
28
|
Notothylas corbiculata
|
1
|
1
|
1
|
1
|
|
29
|
Pilotrichella ampulacea
|
0
|
0
|
1
|
0
|
|
30
|
Pilotrichella sp.
|
0
|
0
|
1
|
1
|
|
31
|
Plagiochila bifaria
|
0
|
0
|
1
|
1
|
|
32
|
Plagiochila sp.
|
0
|
1
|
1
|
1
|
|
33
|
Orchobryum sp.
|
0
|
1
|
0
|
0
|
|
34
|
Polytrichium commune
|
0
|
1
|
0
|
1
|
|
35
|
Pterogonidium sp.
|
1
|
1
|
0
|
0
|
|
36
|
Pterigonium sp.
|
0
|
0
|
0
|
1
|
|
37
|
Porotrichium sp.
|
1
|
1
|
1
|
1
|
|
38
|
Pyribryum spiniforme
|
0
|
0
|
1
|
1
|
|
39
|
Racopilum africanum
|
0
|
1
|
0
|
0
|
|
40
|
Riccardia fastigiata
|
1
|
1
|
0
|
0
|
|
41
|
Riccardia sp.
|
1
|
0
|
0
|
0
|
|
42
|
Riccia stricta
|
0
|
1
|
1
|
0
|
|
43
|
Radula cfr evelynae
|
1
|
1
|
0
|
0
|
|
44
|
Symphyogyna cfr podophylla
|
1
|
0
|
0
|
1
|
|
45
|
Syrrhopodon sp.
|
1
|
0
|
1
|
1
|
|
46
|
Tayloria kilmandscharica
|
1
|
0
|
0
|
0
|
|
47
|
Phaeoceros laevis
|
1
|
0
|
0
|
0
|
|
48
|
Tortula sp.
|
1
|
1
|
1
|
0
|
|
49
|
Inconnue 1
|
1
|
1
|
0
|
0
|
|
50
|
Inconnue 2
|
0
|
1
|
1
|
0
|
|
51
|
Inconnue 3
|
0
|
1
|
0
|
0
|
|
52
|
Inconnue 4
|
0
|
1
|
0
|
0
|
La lecture du tableau 12 montre que 3 espèces
(Dicranum johnstonii, Notothylas corbiculata et Porotrium sp.)
sont présentes dans les 4 secteurs ; 4
Dans le tableau 12, 1 : Présence et 0 :
Absence
espèces (Brachymenium acuminatum, Bryum
argenteum, Frullania angulata et Plagiochila sp.) sont
présentes Mugaba, Tshibati et Tshivanga, mais les espèces :
Athractiloparcus
sp. et
Syrrhopodon
sp. ne sont
pas présentes à Tshibati. L'espèce Tortula
sp. quant à elle n'est absente qu'à
Tshivanga.
D'autres espèces, cependant, sont
présentes dans 2 secteurs, Par exemple : à Madiriri et Tshibati
(Dicranoloma billardierei, Pterigonidium sp., Riccardia fastigiata
et Inconnue sp.1), à Madiriri et Mugaba (Desmatodon
sp.et Lejeunea
sp.), à Tshivanga et Tshibati (Andrewsianthus
sp., Lejeuneaceae sp., Neckera submacrocarpa et
Polytrichium commune), à Madiriri et Tshivanga (Hypnum
cupressiforme et Symphyogyna cfr podophylla).
Certaines espèces ne se retrouvent que dans un
seul secteur, le secteur Tshibati a beaucoup d'espèces plus que les
autres (8) : Funaria hygrometrica, Jensenia spinosa, Lepidozia cupressina,
Marchantia polymorpha, Orchobryum sp., Racopilum africanum,
Inconnue sp.3 et Inconnue sp.4), il est suivi du secteur Madiriri avec 5
espèces (Adelanthus sp., Frullania sp.,
Eurhynchium africanum, Phaeoceros laevis et Riccardia sp.). Le
secteur Tshivanga a, quant à lui, 4 espèces (Fabronia
sp., Fissidens sp., Marchantia planiloba et
Pterigonium sp. ) ; le secteur Mugaba n'a que 2
espèces (Dumortiera hirsuta et Pilotrichella
ampulacea).
3.2.5. Distribution des Bryophytes suivant le gradient
altitudinal
Tableau 13 : Nombre de Bryophytes par tranche
d'altitude
|
Tranche d'altitude
|
Nbres d'espèce
|
Pourcentages (%)
|
|
De 2000 à 2050
|
18
|
34,62
|
|
De 2050 à 2100
|
3
|
5,77
|
|
De 2100 à 2150
|
6
|
11,54
|
|
De 2150 à 2200
|
6
|
11,54
|
|
De 2200 à2250
|
7
|
13,46
|
|
De 2250 à 2300
|
10
|
19,23
|
|
De 2300 à 2350
|
1
|
1,92
|
|
De 2350 à 2400
|
1
|
1,92
|
|
Total
|
52
|
100
|
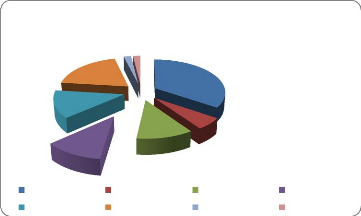
De 2000 à 2050 De 2050 à 2100 De 2100 à 2150
De 2150 à 2200
De 2200 à2250 De 2250 à 2300 De 2300 à 2350
De 2350 à 2400
Distribution des espèces suivant
l'alltitude
Figure 5 . Distribution des espèces suivant
l'altitude
Il ressort du tableau 13 et de la figure 5 que c'est
la tranche altitudinale 200-2050 qui a plus d'espèces ; elle est suivie
de celle de 2250- 2300. Les autres tranches ont eu un nombre moins
élevé d'espèces.
CHAPITRE 4. DISCUSSION
4.1. Comparaison des données sur l'étude
taxonomique des espèces étudiées
Notre étude a relevé la dominance de
l'embranchement des Bryophyta avec 27 espèces (soit 57,5%),
suivi des Marchantiophyta avec 18 espèces (soit 38,3 %).
L'embranchement des Anthocerotophyta est le moins
représenté avec deux espèces (soit 4,2%). Ces
résultats différents de ceux de Wolfrang et al. (1995)
qui ont trouvé la dominance de Marchantiophyta sur le
Bryophyta dans le Parc National de Kahuzi-Biega et à Nyungwe au
Rwanda et ceux de Tixier (1995) qui a aussi trouvé la dominance de
Marchantiophyta sur les Bryophyta dans le même site du
PNKB. Quant aux familles, nous avons trouvé 35 différentes
familles, la famille Bryaceae est la plus représentée
avec trois espèces, elle est suivie de treize familles qui ont chacune
deux espèces. A ce point, nos résultats restent toujours
différents de ceux Pitier (op. cit.) qui lui a trouvé une
dominance de Lejeuneaceae avec 7 espèces ; elle est suivie de
Frullaniaceae avec 5 espèces.
4.2. Comparaison des données sur les types de
substrat
L'analyse des types de substrat a relevé une
dominance des bryophytes qui poussent sur plusieurs substrats ; elles sont le
plus saprophytes et épiphytes avec 30,79 %, suivies des bryophytes
ripicoles et saprophytes pour lesquelles chaque groupe a 15,38 % ;elles sont
suivies des bryophytes terrestres avec 7 espèces, soit (13,46 %). Nos
résultats sont différents de ceux de Wolfrang et al.
(op.cit.) surtout du fait qu'ils se sont intéressés
particulièrement aux bryophytes épiphytes sur les feuilles
vivantes. Mais, Petiot (1996) dans son travail effectué au PNKB a
trouvé une dominance de bryophytes terrestres avec 43,7 %, avec des
bryophytes ripicoles avec 31,2 % qui arruvent en deuxième position ;
viennent ensuite les épiphytes avec 12,5 %, les bryophytes saprophytes
et épiphytes n'étaient représentées que par une
espèce (soit 6,2 %) chacune. Par ailleurs, Petiot (1996) au Kenya dans
le Saiwa Swaps National Park avait trouvé une dominance des bryophytes
épiphytes avec 64,9 %, suivies en importance des bryophytes facultatives
avec 62,8 % ; ce qui est différent de nos résultats. Cette
différence peut être dû au fait que les deux sites sont
situés dans des zones phytogéographiques
différentes.
4.3 Comparaison des données sur les types de
gamétophytes
L'interprétation de types de gamétophyte
a relevé une dominance des bryophytes à tige feuillée avec
81 %, les bryophytes thalloïdes sont représentées par 19 %.
Ces résultats ne sont pas du tout différents de ceux de Tixier
(op.cit.) qui a trouvé seulement les bryophytes épiphytes sur les
feuilles dans le Parc National de Kahuzi-Biega et à Nyungwe.
Résultats toujours semblables à ceux de Wolfrang et al.
(op.cit.) qui ont aussi trouvé une dominance des bryophytes à
tige feuillée dans le PNKB et Nyungwe. Et aussi Petiot (op.cit) qui a
trouvé seulement les ces mêmes bryophytes à tige
feuillée car il avait travaillé sur une seule famille
(Bryaceae) dans le même site du PNKB. Par contre au Kenya, dans
le Saiwa Swamps National Park, Petiot a aussi trouvé une dominance des
bryophytes à tige feuillée avec 96,5 %, suivies des Bryophytes
à thalle avec 3,5 %.
4.4. Comparaison des données sur les
secteurs
Le secteur Tshibati est le plus diversifié en
espèces de Bryophytes, 30 espèces y sont
représentées, parmi lesquelles 7 espèces ne sont que dans
ce secteur ; il est suivi du secteur Tshivanga où 25 espèces sont
représentées, mais 3 espèces sont exclusivement dans ce
secteur. Le secteur Madiriri a 21 espèces parmi lesquelles 4 ont
été seulement présentes dans ce secteur. Par ailleurs, le
secteur Mugaba est le dernier en diversité, il a 20 espèces ;
toutes étant présentes dans les autres secteurs sauf 2
espèces qui lui sont spécifiques.
Deux espèces (Notothylas corbiculata
et Porotrichium sp.) sont présentes dans tous les 4
secteurs. Il y a de ces espèces qui ne sont que dans un seul secteur. A
ce point, le secteur Tshibati reste toujours le premier avec 7 espèces
suivi du secteur Madiriri avec 5 espèces ; vient ensuite le secteur
Tshivanga avec 3 espèces et enfin, Mugaba qui n'a que 2
espèces.
Par contre, Tixier (op.cit.) qui a travaillé au
Kivu et au Rwanda a trouvé une grande diversité au Rwanda ; ainsi
la localité des forêts marécageuses de Kamiranzovu a
été reconnue comme ayant le plus grand nombre d'espèces.
Il a partagé le PNKB en 17 localités. La localité des
forêts ombrophiles collinéennes de NNE de la vallée de
Kahuzi (secteur Mugaba) avait un grand nombre d'espèces (40
espèces), la localité de la forêt
collinéenne de pente SW du Mont Elimo à Irangi était la
moins diversifiée en espèces de bryophytes (4
espèces).
Signalons que cette différence s'expliquerait
par le fait qu'au PNKB, il n'a pas visité les secteurs Tshibati,
Tshivanga et Madiriri où nous avions trouvé une diversité
importante.
4.5. Comparaison des données sur les tranches
d'altitudes
Nous avions parcouru différents habitats avec
différentes altitudes. Le point le moins élevé que nous
avions connu était de 2007 m d'altitude et le plus élevé,
de 2330 m d'altitude (soit une dénivellation de 333 m) que nous avions
partagé en 7 tranches d'altitude. Les tranches 2000 - 2050 m et 2250 -
2300 m sont les plus diversifiées en espèces de bryophytes avec
18 et 10 espèces. Seule l'espèce Symphyogyna cfr podophylla
s'est retrouvée dans 6 tranches d'altitude, elle a
été absente seulement dans une seule tranche (2100 - 2150). A
part les espèces retrouvées dans 3 ou 2 tranches, il y a quelques
espèces qui ne sont que dans une seule tranche d'altitude.
Nos résultats ne sont pas très
différents de ceux de Tixier (op.cit.) qui a travaillé dans une
dénivellation large (de 850 - 3100 m) de 1150 m. Il a trouvé une
plus grande diversité dans la tranche 2000 - 2100m avec 62
espèces, suivie de la tranche de 2400 - 2500 m avec 27 espèces.
La tranche 3000 - 3100 m était la moins diversifiée.
4.6. Comparaison des données selon les types
d'habitats
Notre étude a montré qu'aucune bryophyte
n'est retrouvée dans les 5 types de forêts que nous avions
rencontrées au PNKB. Les forêts secondaires ont été
les plus diversifiées avec 42 espèces y
représentées dont 18 y sont spécifiques ; elles sont
respectivement suivies des forêts primaires avec 18 espèces, des
clairières avec 14 espèces, des galeries forestières avec
4 espèces et enfin, les marais avec 3 espèces.
Par contre, Tixier (op.cit.) a trouvé une
grande diversité des bryophytes dans la forêt marécageuse
de Kamiranzovu au Rwanda et dans la forêt ombrophile de montagne à
balles de mousses dans les cimes des arbres avec 35 espèces ; elle
est
suivie de la forêt ombrophile collinéenne
près de Kahuzi au PNKB avec 22 espèces, puis de la forêt
ombrophile à fougères arborescentes avec 20 espèces et
enfin, la forêt de Podocarpus-Syzygium au Mont Biega avec 14
espèces.
Signalons que les différences observées
entre notre travail et celui de Tixier s'expliquent par le fait que nous
n'avions pas les mêmes types de substrats. Nous avions travaillé
sur les bryophytes terrestres, ripicoles, saprophytes et épiphytes sur
les troncs des arbres pendant que lui a travaillé sur les bryophytes
follicoles.
CHAPITRE 5. CONCLUSION ET RECOMMANDATIONS
Le présent travail porte sur la taxonomie et
l'écologie des Bryophytes de haute montagne de l'Est de la
République Démocratique du Congo, plus précisément
dans le Parc National de Kahuzi - Biega. Durant l'inventaire
systématique dans 4 secteurs de la partie de haute altitude de cette
phytocénose, nous avons recensé 52 espèces de Bryophytes
dont 48 ont été identifiées et 4
idéterminées. Ces 48 identifiées sont reparties dans 3
embranchements, 6 classes, 7 sous-classes, 22 ordres, 35 familles et 42 genres.
L'embranchement des Bryophyta est le plus dominant avec 27
espèces (soit 57,5%).
L'analyse des types de substrat a relevé une
dominance des bryophytes rupicoles (qui vivent sur les roches), suivis des
bryophytes terrestres et enfin, les épiphytes qui sont les moins
représentées. Ces résultats nous prouvent qu'il y a une
certaine spécificité en ce qui concerne les relations entre
bryophytes et types de substrats. Quant à ce qui concerne les types de
gamétophytes, le PNKB est dominé par les bryophytes à
tiges feuillées avec 81% dont les Acrocarpes ont 62% et les Pleurocarpes
19 %. Les bryophytes thalloïdes sont les moins représentés
avec 18 %.
En se basant sur la diversité des bryophytes
selon les secteurs, le secteur Tshibati est le plus diversifié avec 30
espèces, il est suivi du secteur Tshivanga avec 25 espèces. Mais
en se basant sur la spécificité en espèces, Tshibati est
suivi de Madiriri.
L'interprétation des types d'habitats a
relevé une dominance des bryophytes des forêts secondaires au
PNKB. L'habitat qui vient en second lieu est la forêt primaire. Notre
étude a relevé que la diversité des bryophytes varie en
fonction du gradient altitudinal car aucune bryophyte n'a été
retrouvée dans toutes les tranches d'altitude malgré que la
dénivellation n'a pas été si importante.
Nous espérons, tant soit peu, avoir
contribué à la connaissance des bryophytes du Parc National de
Kahuzi-Biega surtout par la mise en évidence d' espèces n'avaient
pas encore été signalées au PNKB. Il s'agit de :
Andrewsianthus kilimanscharica, Marchantia polymorpha, Marchantia
planiloba, Notothylas corbiculata et Phaeoceros
laevis.
Ainsi, nous recommandons :
- aux chercheurs et étudiants congolais et
étrangers de bien vouloir promouvoir des recherches sur les bryophytes
du PNKB car malgré les efforts déjà consentis, beaucoup de
choses demeurent inconnues ;
- a l'Etat congolais de bien vouloir amplifier des
relations afin de disponibiliser à Bukavu un laboratoire permettant de
réaliser des études biologiques approfondies ;
- aux autorités de la Faculté des
Sciences de l'Université Officielle
de Bukavu, à orienter les
étudiants à faire de nombreux travaux sur les bryophytes du PNKB
et des ses environs afin de bien connaitre ces organismes.
REFERENCES BIBLIOGRAPHIQUES
Balezi, Z., Nyakabwa, M., Lejoly, J., 2008. Etudes
écologiques de la liane envahisante Sericostachys scandens dans la
partie de haute altitude du Parc National de Kahuzi-Biega. Annales des
Sciences de l'Université Officielle de Bukavu Vol.1 (1),
28-36p.
Basabose, A., Balagizi, I., Muhigwa, B., 2001.
Formation en parataxonomie pour les gardes du Parc National de Kahuzi -
Biega. Volet I. Plantes et Mammifères, Inédit.
Fischer, E., 1993. La végétation du
Parc National de Kahuzi - Biega (Sud - Kivu, Zaire). Botanisches Institut
Und Botanischer Garten der Rheinischen Frienrich - Wilhelms - Universitat Bonn,
Bonn, 93p.
Gabriella, K., 1996. Andean Daltonia latolimbata
Booth. In Herzog in Africa, Tropical 12: p35 - p40.
Kurshner, H., Hohenliderung,. 1995. Epiphytischer
Moose im ostlichen Kongo - becken und den angrenzenden Gebirgsstcken (Parc
National de Kahuzi - Biega / Zaire, Forêt de Nyungwe / Rwanda).
Institut fur Systematische Botanik und Pflanzengeograhie der Freien universitat
Berlin, Altensteinstr.6, D - 14195 Berlin, Deutschland.
Letouzey, R., 1982. Manuel de Botanique
forestière : Afrique tropicale. Tom 1, Paris, 193p.
Muhlenberg, M., 1995. Parc National de Kahuzi -
Biega. Sl. 52p.
Nyakabwa, M., 2005. Etude succincte de la
Biodiversité végétale : quelques notions de base. P -
BEATRA, 65p.
Petiot, C., 1996. Bryophytes from Saiwa Swamp
National Park, Kenya. Tropical. Bryology 12.
Petiot, C., 2003. Mosses, Liver worts and Horn
worts of Kenya. 273p. Tropical Bryology 12.
Petiot, C., 1996. Taxonomic Results of the
Bryotrop Expedition to Zaire and Rwanda. 32 Bryaceae, Tropical Bryology
12. p59 - p62.
Shalukoma, C., 2008. Usage des plantes
médicinales par les populations riveraines de la partie de haute
altitude du Parc National de Kahuzi - Biega, Sud - Kivu, R.D.Congo.
Mémoire du D.E.S., U.L.B., Fac. Sc., 67p.
Tixier, P., 1995. Résultats taxonomiques de
l'expédition Bryotrop au Zaire et Rwanda, 30 Bryophytes
épiphylles (récoltes de FISCHER, E.). Tropical Bryology 11.
p11 - p76.
Wolfgang, F., Michael, S., Fischer, E., 2009.
Syllabus of Plant Families 13. Borntraeger, 400p.
TABLE DES MATIERES
CHAPITRE 1. INTRODUTION 5
1.1. PROBLEMATIQUE 5
1.2. Objectifs du travail 6
1.3. Intérêt du travail 6
1.4. Généralités sur les
Bryophytes 7
CHAPITRE 2. MILIEU D'ETUDE ET METHODES DE RECHERCHE
12
2.1. Milieu d'étude 12
2.1.1. Situation géographique 12
2.1.2. Végétation 13
2.1.3. Faune 15
2.2. Matériel et Méthodes d'étude
15
2.2.1. Matériel 15
2.2.2. Méthodes de travail 15
CHAPITRE 3. RESULTATS 19
3.1. Etude floristique 19
3.1. Analyse taxonomique des espèces
inventoriées 24
3.1.1. Classification taxonomique des espèces
inventoriées 24
3.1.2. Brève description morphologique de
certaines espèces inventoriées 28
3.1.3. Types de gamétophytes des espèces
inventoriées 30
3.1.4. Diversité des secteurs prospectés
31
3.2. Ecologie des espèces des Bryophytes
étudiées 34
3.2.2. Distribution des bryophytes selon les types
d'habitat 38
3.2.3. Types de substrats des espèces
inventoriées 40
3.2.4. Distribution des Bryophytes selon les secteurs
43
3.2.5. Distribution des Bryophytes suivant le gradient
altitudinal 46
CHAPITRE 4. DISCUSSION 47
4.1. Comparaison des données sur l'étude
taxonomique des espèces étudiées 47
4.2. Comparaison des données sur les types de
substrat 47
4.3 Comparaison des données sur les types de
gamétophytes 48
4.4. Comparaison des données sur les secteurs
48
4.5. Comparaison des données sur les tranches
d'altitudes 49
4.6. Comparaison des données selon les types
d'habitats 49
CHAPITRE 5. CONCLUSION ET RECOMMANDATIONS
51
REFERENCES BIBLIOGRAPHIQUES 53
TABLE DES MATIERES 55
| 

