INTRODUCTION.
La forte variabilité spatio-temporelle des
précipitations dans les régions sahéliennes a
été marquée par une longue période de
sécheresse depuis la fin des années 60 jusqu' au milieu des
années 90. Cette période de sécheresse a
entraîné d'importantes fluctuations dans la composition
floristique et la production végétale. Les causes de cette
variabilité sont à rechercher dans le phénomène de
la mousson ouest africaine. En effet lorsque le continent se réchauffe,
en été, il attire l'air qui s'est chargé en
humidité au dessus du Golfe de Guinée. Ce flux d'air humide
appelé mousson remonte vers le nord et, une fois au dessus du continent,
se transforme en systèmes orageux. Ces systèmes se
déplacent d'est en ouest, arrosant toute la région avant
d'arriver sur l'Atlantique, où ils se transforment parfois en cyclones.
Pour le Sahel c'est le seul période de pluies de l'année. Toutes
les ressources en eau en dépendent, ainsi que les ressources
végétales naturelles et cultivées.
L'un des objectifs du programme AMMA (Analyse
Multidisciplinaire de la Mousson Africaine) est d'améliorer la
compréhension de la mousson d'Afrique de l'Ouest et de son impact sur
l'environnement physique, chimique, et biologique à l'échelle
régionale et mondiale. Plusieurs disciplines sont donc mises à
contribution dans ce programme : hydrologie, océanographie, sciences de
l'atmosphère, bioclimatologie, écologie, et plusieurs
régions d'études ont été définies notamment
au Bénin, au Niger et au Mali.
Au Mali, la zone retenue pour les recherches est le Gourma. Le
programme dispose d'un site méso-échelle qui regroupe 27 sites de
suivi de la végétation, d'un super site instrumenté
à Hombori et d'un site de mesure intensif à Agoufou.
Ces sites font l'objet d'un suivi régulier de la
végétation depuis une vingtaine d'années et sont
maintenant instrumentés par le programme AMMA afin de
caractériser l'environnement climatique (pluviométrie,
rayonnement, températures,...) ainsi que le contenu en eau du sol.
Notre stage se situe dans le cadre du suivi de la
végétation et avait pour objectif de :
10
- suivre et caractériser l'évolution de la
dynamique saisonnière de la végétation herbacée et
ligneuse en relation avec les précipitations pendant la saison des
pluies 2006,
- comparer les données de mesure 2006 aux données
de 2005.
- comparer les données de biomasse aux mesures par
photographie hémisphérique. - valider un logiciel d'analyse de
photographies hémisphériques, le logiciel can_eye (INRA, Avignon)
et comparer les résultats estimés par can_eye et les
résultats de surface foliaire estimés par le planimètre
à partir des mêmes échantillons.
Les paramètres suivis au cours du stage étaient
:
- l'indice foliaire ou LAI (Leaf Area Index) : c'est la surface
totale d'une face des feuilles par unité de surface de sol (WATSON,
1947). Il s'exprime en m2 de feuilles par m2 de sol.
- le recouvrement végétal ou Fcover : c'est le
pourcentage de la surface au sol masquée par la végétation
en visée verticale.
- le PAI (Plant Area Index) : c'est la surface
végétale totale (tronc, branches, feuilles) par m2.
Le logiciel can_eye est un logiciel qui permet de
déterminer l'indice foliaire ou LAI à partir des photographies
hémisphériques, et le recouvrement ou Fcover à partir des
photographies verticales.
Une photographie hémisphérique est une photographie
réalisée avec un objectif « fisheye » dont le champ de
vue est de 180° en zénit et de 360° en azimut. La photographie
verticale est une photographie prise sans objectif « fish-eye ».
Parmi les variables d'étude, le LAI est une variable
clé du fonctionnement des couverts végétaux. Il intervient
dans l'interception du rayonnement, la photosynthèse et
l'évapotranspiration, il permet de caractériser l'état de
la végétation.
De nombreuses méthodes ont été
développé pour déterminer le LAI : les méthodes
directes et indirectes.
Les méthodes directes sont destructives et coûteuses
en main d'oeuvre et en temps. Néanmoins, elles demeurent indispensables
pour l'évaluation et la validation des
11
méthodes indirectes. Les méthodes indirectes par
exemple à l'aide de photographies sont non destructives et plus rapides.
Cependant ces méthodes reposent sur l'hypothèse que la
végétation est homogène avec des éléments
distribués uniformément. Dans la pratique, ceci est rarement
vérifié, particulièrement pour les couverts discontinus.
Dans ce contexte nous avons utilisé le logiciel can_eye pour traiter nos
données. L'intérêt du logiciel can_eye est qu'il estime le
LAI de deux façons différentes : soit par l'utilisation du
modèle de Poisson qui considère que les feuilles sont
distribuées uniformément ce qui permet d'avoir une estimation du
LAI effectif, soit par l'utilisation du modèle de Poisson modifié
qui permet de prendre en compte la disposition non uniforme des
éléments du couvert et d'estimer le LAI vrai.
Trois sites font l'objet de notre stage. Ce sont les sites 17
(Agoufou), 21 (Kelma), et 40 (Egérit).
La première partie de ce mémoire porte sur la
présentation de la région d'étude, la deuxième sur
la présentation de la zone d'étude la troisième sur les
sites d'étude. La quatrième sur la méthodologie
utilisée. La cinquième porte sur les résultats et
discussion. Enfin la sixième et dernière partie constitue une
conclusion
12
1- Présentation de la région
d'étude: Le Sahel
1-1- Position géographique (fig.
01)
Le sahel est une zone de transition entre le domaine saharien
au nord et le domaine des savanes humides soudaniennes au sud. Il
s'étend sur environ 6000km entre l'Océan Atlantique et la mer
Rouge (LE HOUEROU, 1989). D'ouest en est, la bande sahélienne large de
400 à 600km traverse le Cap-Vert, la Mauritanie, le
Sénégal, le Mali, le Niger, le Tchad, le Soudan, et effleure le
Nord du Burkina-faso, du Nigeria et du Cameroun, elle couvre environ 3 millions
de km2. (Fig.01).
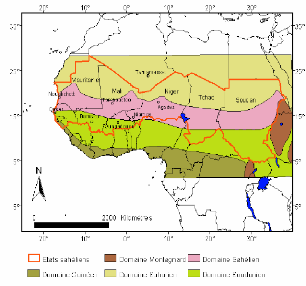
Fig.01: Les grands domaines climatiques et bioclimatiques
(d'après AUBREVILLE, 1949)
1-2 Milieu naturel
1-2-1 Le relief
Le sahel se caractérise par un relief relativement
faible (200-600m) dû à l'extension des ensembles dunaires et des
cuirasses qui occultent les diverses phases d'érosion et de
sédimentations successives depuis l'ère primaire. Cependant,
quelques exceptions sont à noter comme la frontière soudano-
tchadienne (1000m),le plateau de Bandiagara (700m) et la région des
Monts Hombori au Mali avec (1155m). Les autres reliefs
13
importants se trouvent en bordure du Sahara et du Sahel :
l'Adrar de Mauritanie (700m), l'Adrar des Ifôghas au Mali (900m),
l'Aïr au Niger (1900m), le Tibesti (3400m) et l'Enedi (1500m) au Tchad.
Plus à l'est, au Soudan le Gourgeil et le Marra
d'origine volcanique culminent respectivement à 2300 et 3100 m.
1-2-2 Le climat
Le climat en zone Sahélienne est tropical avec une
courte saison des pluies et une longue saison sèche. La saison des
pluies débute avec l'installation de la Zone de Convergence
Intertropicale (ZCIT) et les pluies régulières vers la fin du
mois de juin, et se termine au mois de septembre. Les premières pluies
enregistrées au mois de mai en zone sahélienne ne
résultent pas de l'installation de la mousson mais plutôt de
l'instabilité atmosphérique due à l'arrivée du FIT
(Front de Convergence Intertropical) situé à environ 10° au
nord de la ZCIT. Un maximum de précipitation s'observe en Août. La
saison est dite précoce si les pluies apparaissent
régulièrement dès le mois de mai et tardive si elles
subsistent régulièrement au mois d'octobre.
La moyenne annuelle des températures de l'air est
d'environ 28°C (PENNING et DJITEYE, 1982).
Durant l'hivernage, les variations de température
dépendent fortement des précipitations mais pas seulement, le
vent influe aussi sur les changements des températures et la
saisonnalité.
Les valeurs minimales et maximales des températures
moyennes journalières sont enregistrées respectivement en
décembre- janvier (10-15°C) et en fin de saison sèche vers
les mois d'avril- mai (35-40°C). Un minimum et un maximum secondaires
peuvent aussi être observés au mois d'août en raison du
rafraîchissement de l'air entraîné par les pluies et autour
du mois d'octobre après l'hivernage.
1-2-3 Les sols
Trois ensembles pédologiques peuvent être
reconnus dans le Sahel (PENNING et DJITEYE, 1982) ce sont : Les sols sableux,
les sols détritiques sur grès ou latérites, et les sols
fluviatiles ou lacustres.
14
- Les premiers possèdent une texture grossière,
avec une capacité d'infiltration importante et un ruissellement
limité. Ces sols sont peu profonds et homogènes sur de grandes
superficies, représentant environ 50% de la surface totale du sahel. Ils
sont légèrement acides (pH=6.5) dans la moitié nord du
Sahel. Dans la partie sud du Sahel, les sols sont plus acides, avec un pH
compris entre 5 et 6. Dans l'ensemble, les sols sableux contiennent peu de
matière organique, d'azote et de phosphore.
- Les surfaces latéritiques occupent 30% du Sahel et
donnent généralement des sols limoneux.
- Enfin, les 20% restant du Sahel correspondent à
l'ensemble lacustre/fluviatile dont les sols sont d'origines
sédimentaires et plus riches en argile que les deux
précédents.
1-2-4 La végétation.
Trois composantes peuvent être identifiées dans
le Sahel (LE HOUEROU, 1989) :
- les herbacées annuelles couvrent de 20 à 80%
de la surface avec une hauteur moyenne comprise entre 40 et 80 cm et se
composent principalement de graminées comme Cenchrus biflorus,
Aristida mutabilis, Brachiaria xantholeuca, Echinochloa colona, Panicum
laetum, et de dicotylédones comme Zornia glochidiata, Tribulus
terrestris, Corchorus tridens, Alisicarpus ovalifolius, et Trianthema
pilosa .
- la strate arbustive ou buissonnante d'une hauteur de 0,5
à 3m peut occuper de 0 à 20% de la surface. Parmi les
espèces les plus caractéristiques, on trouve : Leptadania
pyrotechnica, Boscia senegalensis et Calotropis
procera.
- la strate arborée se compose de petits arbres de 3
à 6m et couvre de 1 à 5% de la surface. Ils sont répartis
de façon dispersée et possèdent des
caractéristiques leur permettant de lutter contre l'aridité,
telles que la petite taille et les feuilles réduites (microphyllie chez
les Acacia) ou recouvertes par une épaisse cuticule. Les
espèces les plus fréquentes sont des épineux de la famille
des Mimosaceae (Acacia senegal ou gommier, Acacia raddiana),
accompagnés ici et là d'espèces à feuilles plus
grandes, épaisses et cireuses comme Balanites aegyptiaca et
Combretum glutinosum.
15
2- Présentation de la zone d'étude :
Le Gourma malien
2-1- La position géographique.
Le Gourma est une région naturelle de 130.000 km2 dont
la partie malienne couvre près de 90.000km2. Situé dans la partie
Nord Est du pays entre les isohyètes 200 et 400mm et les latitudes 15 et
17°N, le Gourma malien se trouve à cheval sur trois régions
administratives du Mali, celle de Gao, de Tombouctou et de Mopti (Ag Mahmoud,
1992). Il est délimité par la boucle du Niger à l'0uest,
au Nord et à l'Est, la République du Niger et le Burkina Faso au
Sud Est, puis le plateau de Bandiagara au Sud Ouest (fig.02).
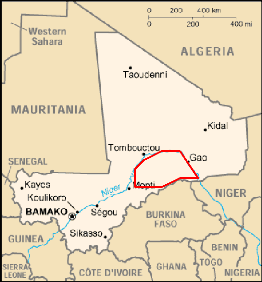
Fig. 02 : Position géographique du Gourma
malien.
2-2 Le milieu naturel
2-2-1 Le relief
Le relief du Gourma est bas. On y observe des groupes de
collines rocheuses atteignant 350m. Le point culminant est localisé dans
la zone des monts entre Douentza-Hombori c'est le mont Hombori avec une hauteur
de 1155m. Les dunes de sable ont généralement une hauteur
comprise entre 3 et 10m.
16
2-2-2 L'hydrographie:
On trouve dans le Gourma les eaux de surface et les eaux
souterraines superficielles. - Les eaux de surface :
Pendant l'hivernage l'eau s'accumule dans les
dépressions situées entre les roches et les inter - dunes sous
forme de mares permanentes comme celle de Gossi et Benzena ou temporaire comme
celles de Tessit, Ekia, Adiora, Niangaye, Fintrou, Idamane, Inadiatafane et
N'daki.
Des puisards de cinq à dix mètres sont
creusés dans le lit des mares asséchées.
- Les eaux souterraines:
Les nappes phréatiques sont profondes compte tenu de la
nature géologique du sol. On y compte peu de forages et de puits. Les
puits et forages sont creusés dans les bas fonds à une profondeur
de 15 à 25m.
2-2-3 Le climat.
Le climat est sahélien. Les précipitations
annuelles sont faibles avec une grande variabilité spatio-temporelle. La
pluie commence en juin et se termine en septembre. Un maximum est
observé en août. La saison sèche dure 7 à 8 mois et
se divise en 2 périodes. Une période sèche et
fraîche (décembre - février) et une période
sèche et chaude (mars- mai). Le mois d'octobre est humide et chaud et le
mois de Novembre humide et frais.
2-2-4 Les sols.
Comme pour l'ensemble du Sahel, le Gourma est dominé
par des sols sableux qui occupent 59,6 % de la surface (KAMMERUD, 1993). Les
sols limoneux (16,1%), argileux (3,4%) et les sols gravillonnaires 20,9% sont
les autres grands types de sols.
Les sols sableux s'observent au sein de plusieurs formations
comme les dunes stabilisées et les plaines dunaires.
Le pH de ces sols est généralement compris entre
4 et 6. La teneur en azote et carbone y est faible. Le phosphore se rencontre
également en très faible quantité avec cependant une
localisation préférentielle dans les 10 premiers
centimètres.
17
A l'exception du calcium, les autres éléments
comme le potassium, le magnésium ne sont pas non plus très
présents dans le sol quelle que soit la profondeur de la couche
étudiée (GJESSING et DIALLO, 1999)
2-2-5 La végétation.
La formation végétale majoritairement
rencontrée dans le Gourma est la « steppe à épineux
».
Ainsi on distingue dans le Gourma quatre types de
végétations classées en fonction de la morphologie des
sols.
1) - végétation des dunes : dominée par
des graminées annuelles telles que Cenchrus biflorus et
Aristida mutabilis et des ligneux composés essentiellement de
Balanites aegyptiaca, Acacia senegalensis et Acacia
raddiana.
2) - végétation des ensablements et des
surfaces cuirassées : c'est le domaine de la brousse tigrée avec
les espèces ligneuses telles que Commiphora africana, et
Boscia senegalensis et des graminées comme Cenchrus
biflorus, et Aristida mutabilis.
3) - La végétation des forêts claires :
c'est un peuplement dense d'Acacias dans les dépressions
argilo-limoneux. On y observe principalement Acacia seyal, quelques
Balanites aegyptiaca et Acacia nilotica en nombre plus
réduit.
4) - végétation des mares et du fleuve :
dominée par le bourgou (Echinocloa stagnina, Echinochloa
colonna). Autour des mares et dans les dépressions poussent
Bracharia lactum, Panicum subaloidium, Salvadora percica et Acacia
laeta.
2-3 La populations et les systèmes de
production.
2-3-1 La population
La population du Gourma est difficile à estimer
à cause de sa grande mobilité. Selon le recensement
général effectué au Mali en 1998, la population de Gourma
Rharous qui couvre la majeure partie du Gourma malien est de 67.717 habitants.
Les densités varient de 8,4 habitants/km2 à 0,49
habitants/km2.
Cette population présente une complexité
ethnique. Elle est composée de Kel Tamasheq (Berbères venus du
Nord), de Songoy (habitants de la vallée du fleuve Niger), de Peul
(transhumants) et de Dogons (agro- pasteurs)
18
2-3-2 Les systèmes de production
:
Selon la nature des activités des populations, il
existe quatre systèmes de production
(UNESCO / PPOOMLI 507, 2001)
- Le système pastoral pur pratiqué par les Kel
Tamacheq.
-Le système agropastoral pratiqué par les Peuls,
les Songoy, et les Dogons.
Du fait de la régression de la pluviométrie, les
populations ont abandonné les dunes et
ont installé leurs champs près des bas fonds
où convergent les eaux de ruissellement.
- Le système agricole pur pratiqué par les
Peuls, les Dogons et les Songoys.
- Le système maraîcher pratiqué par la
population sédentaire autour des mares
pérennes comme Gossi, Dimamou.
Plusieurs familles pratiquent le maraîchage autour des
mares à cause du caractère
aléatoire de l'agriculture pluviale.
Pendant l'hivernage, elles cultivent du mil, du sorgho, du
maïs et le reste de l'année des
légumes.
19
3- Les sites d'étude:
Plusieurs sites ont été mis en place dans le
Gourma malien par l'équipes du CIPEA (P. HIERNAUX et al, 1984) dans le
but d'étudier les systèmes de production pastoraux mais aussi
d'effectuer des essais de télédétection spatiale de 1984
à 1993. Le CESBIO a repris ces suivis en 1999 et depuis 2002 le projet
AMMA mène ses recherches sur ses sites pour étudier les
mécanismes de la mousson africaine et essayer de mieux comprendre les
causes de la sécheresse.
Les trois sites qui font l'objet de notre étude
regroupent les différentes sortes de substrats rencontrés dans le
Gourma : surfaces sableuses (site 17), plaines argileuses (site 21) et surfaces
gravillonnaires (site 40)
3-1 Agoufou (site 17)
Le site d'Agoufou est le site principal du programme AMMA. Il
est situé sur un cordon dunaire sableux, à environ 30km à
l'ouest d'Hombori
3-1-1 Les réseaux de mesure :
Des appareils automatiques sont installés sur ce site.
Parmi ceux-ci il y a :
1- La station météo muni de plusieurs appareils
qui fournissent chaque 15mm des informations sur les paramètres
météorologiques et l'humidité du sol. Parmi ces appareils
nous avons :
- les sondes de température et d'humidité relative
de l'air,
- le baromètre qui mesure la pression
atmosphérique,
- l'anémomètre qui mesure la vitesse du vent,
- la girouette qui mesure la direction du vent,
- le radiomètre CNR1 qui mesure le rayonnement incident et
réfléchi par la surface,
- Le Radiomètre PARGLOB et PARDIFF (Rayonnement
Photosynthétique Actif Global et Diffus).
- des sondes d'humidité de sol qui sont enfuies sous
terre jusqu'à 2,5m de profondeur et mesurent avec précision
l'infiltration de l'eau.
- le pluviomètre mesure la quantité de pluie
20
- Les thermo radiomètres IRT (Infra rouge thermique) :
ce sont des récepteurs passifs qui collectent le rayonnement émis
par la surface (dans le domaine de l'infrarouge lointain
2- les flux de sève : mesurent la consommation en eau
des arbres en fonction des conditions climatiques environnementales
(rayonnement, température, humidité relative), à
l'échelle horaire, journalière ou mensuelle.
3-1-2 La végétation
La végétation de ce site est une steppe à
épineux typique du Gourma sur sol sableux profond d'origine
éolienne (dune fixée). La strate ligneuse est composée des
espèces couramment rencontrées en zone sahélienne comme :
Acacia (Acacia raddiana, Acacia senegal), Combretum
glutinosum, Balanites aegyptiaca etc. A cela il faut noter la
présence très abondante de Leptadenia pyrotechnica
apparu après la sécheresse de 1984. La strate
herbacée (Fig. 04) est composée essentiellement d'espèces
annuelles monocotylédones (des graminées comme Cenchrus
biflorus, Tragus Berteronianus, Aristida mutabulis,
Bracharia xantholaca, Bracharia deflexa, Bracharia
latan, etc) et des dicotylédones (légumineuses comme
Zornia Glochidiata, Alysicarpus ovaifolius, Indigofera
strobilifira ou non légumineuses comme Gisekia
pharnacioïdes).

|
Site 17(Agoufou), 02 /07/2006
|
Site 17, 29/08/2006
|
Fig. 03 Vue d'ensemble de Agoufou en juillet et
août 2006
21

|
Fig.04 : Végétation herbacée,
Agoufou le 12 août 2006
|
3-2- Kelma (site 21)
Le site 21 ou « acacière », situé
à 11km du site 17, correspond à une forêt claire d'acacias
située dans une dépression argilo- limoneuse. On y trouve
principalement Acacia seyal, ainsi que quelques Balanites
aegyptiaca et Acacia nilotica. Le couvert ligneux reste cependant
assez ouvert et assez bas avec 45% de recouvrement. Les individus les plus
âgés ont 7-8m de haut ; la taille moyenne est autour de 4-5m.
La feuillaison se fait en saison pluvieuse. Cette
acacière devient souvent inondée (jusqu'à 70cm d'eau par
endroit) au cours de la saison pluvieuse (fig. 05)

|
Site 21 (Kelma), 03/07 /2006
|
|
Site 21, 26/08/2006
|
22
Fig.05 : Vue d'ensemble de Kelma en juillet et août
2006
23
3-3 Egerit (site 40)
Le site 40 situé à 15km du site 17 est une vaste
surface gravillonnaire avec une végétation limitée aux
ensablements (fig. 06)
La strate ligneuse est composée à majorité
d'Acacia erhenbergiana. On y trouve également Acacia laeta,
Maerua angolensis, Acacia senegal, Balanites aegyptiaca etc. La strate
herbacée est composée de : Penisetum violaceum, Tragus
racemosus, Bracharia xantholeuca, Aristida mutabilis, Panicum laetum, Cenchrus
biflorus, Schoenfeldia gracilis etc.

Site 40(Egerit), 12/08/2006
Fig. 06 : Vue d'ensemble de Egerit en août
2006
24
4- Matériel et méthodes.
En dehors de l'utilisation de photographies
hémisphériques, la méthode classiquement utilisée
pour l'estimation des masses du couvert herbacé combine une
stratification du couvert herbacé à un échantillonnage de
12 placettes de 1x1m le long d'un transect d'1 km2.
4-1 Echantillonnage stratifié aléatoire
du couvert herbacé.
Il se fait en 2 étapes :
1) La stratification de la végétation se fait
le long d'un transect de 1000m Est-Ouest, matérialisé par un
ruban hectométrique tendu entre deux bornes. La distinction des strates
est basée sur la densité apparente de la végétation
appréciée à l'échelle du mètre
carré.
Quatre catégories de strates sont identifiées
pendant les mesures : les plages de sol nu (0), la densité basse (B),
moyenne (M), et la haute (H). Les strates se définissent de façon
relative à chaque date d'observation exceptées les plages de sol
nu.
2) Echantillonnage de 12 placettes de 1x1m le long du même
transect :
Les mesures se font sur 12 placettes de 1x1m réparties
à raison de 3 dans chacune des strates B et H et 6 dans la strate M. La
strate O, sans végétation, n'est pas
échantillonnée. Les emplacements des strates sont
préalablement tirés de façon aléatoire. Les
mesures, prélèvements et observations se font sur les
échantillons de 1x1m le long de l'axe de stratification. Les mesures
sont les suivantes :
- appréciation visuelle du taux de recouvrement
herbacé (%), de la proportion des organes végétaux verts,
soit le rapport entre biomasse et phytomasse (%), et du taux de recouvrement de
la litière herbacée (%).
- prélèvement par fauche (mesures destructives)
de la phytomasse en frais (g/m2) et (prélèvement par ratissage)
de la masse de litière en matière sèche (g/m2)
après séchage de l'échantillon à l'air.
- identification des espèces herbacées
présentes sur l'échantillon.
25
4-2 Estimation du LAI et du Fcover (recouvrement)
sur le même transect par photographie hémisphérique et
verticale.
Dans le but de suivre l'évolution saisonnière
des herbacées et de préciser la relation LAI/Fcover, des mesures
du LAI et du Fcover sont faites aux mêmes dates et sensiblement aux
mêmes endroits (fig.07)
Une seule méthode est utilisée pour ces deux
paramètres :
Un ruban gradué tiré le long du transect de
1000m (Est-Ouest), permet de se placer tous les 10m pour faire les photos. La
seule différence consiste à utiliser ou ôter le fish eye
selon que l'on veut faire des photos hémisphériques pour la
mesure du LAI, ou des photos verticales pour la mesure du Fcover.

Fig. 07 : photo hémisphérique des
herbacées, site 17, 2006
4-3 Essai de validation des mesures par photographie
hémisphérique, site 17. 4-3-1 Prélèvement
des échantillons.
L'objectif principal de ce travail est de comparer
l'estimation du LAI par photographie de la mesure par planimétrie sur
une même surface.
Le principe est le suivant :
Nous choisissons des placeaux d'1m2 de façon
à avoir les placeaux de biomasse faible, moyen et fort. Les placeaux
sont choisis visuellement et délimités à l'aide d'un
grand
26
cadre métallique de 1m2. A l'intérieur de ce
placeau, un petit placeau de 400cm2 est aussi choisi de façon
à être représentatif du grand placeau.

Fig.08 : Le grand carré et le
petit carré représentatif du grand carré
Ensuite nous faisons les photos hémisphériques
et verticales du grand et petit placeau avant de procéder à une
estimation visuelle du taux de recouvrement et de la hauteur de la
végétation herbacée en distinguant les
monocotylédones des dicotylédones. Nous établissons
également la liste de toutes les espèces présentes dans le
placeau, nous comptons toutes les individus du petit placeau et nous les
prélevons pour mettre dans les sacs plastiques. Les
monocotylédones sont séparées des dicotylédones.
Nous enregistrons sur un coupon toutes les informations sur
l'échantillon et le mettons ensemble avec l'échantillon. Pour
éviter que les feuilles ne se recroquevillent pendant que nous sommes
sur le terrain nous mettons les échantillons au froid dans une
glacière. Le reste de l'herbe dans le grand placeau est aussi
coupé et mis dans le sac en tissu coton et destiné
ultérieurement au séchage et à la pesée pour avoir
la masse de l'échantillon. Là aussi les monocotylédones
sont séparées des dicotylédones. Une fois les
opérations finies sur le terrain nous ramenons les échantillons
dans la salle de mesure pour déterminer les surfaces foliaires à
l'aide d'un planimètre.
27
4-3-2 Mesure de la surface foliaire par
Planimétrie :
Le planimètre utilisé pour nos mesures est un
planimètre Li 3100, Licor, NE.USA. C'est un appareil muni d'un tapis
roulant qui pendant son mouvement déplace les feuilles au dessus d'une
source lumineuse fluorescente et détecte automatiquement leurs
surfaces.
La méthode de mesure par planimétrie est la
suivante :
Nous posons le planimètre sur une table,
vérifions qu'il est horizontal sur tous les côtés à
l'aide d'un niveau à bulle.
- Après avoir bien aplatie les feuilles des herbes,
nous les faisons passer une à une dans le planimètre. A la fin de
chaque manipulation s'affiche dans le cadran de comptage un nombre qui
désigne la surface totale de toutes les feuilles
planimétrées. Nous relevons ce nombre sur une feuille et
remettons le compteur sur zéro. L'échantillon est remis dans une
enveloppe sur laquelle on recopie toutes les informations notées sur le
coupon et est destiné au séchage et à la pesée.
Avant de passer à une autre manipulation on vérifie si le
compteur est sur zéro. La même méthode est utilisée
pour tous les échantillons.
4-4- Dynamique saisonnière des herbacées
du site 21.
La même méthode utilisée pour le suivi du
LAI et le Fcover au site 17 est également utilisée au site 21
sauf que la ligne est de 500m.
4-5- Suivi d'évolution du PAI (Plant Area
Index) au site 21 (Kelma)
Dans le cas de ce peuplement ligneux relativement dense et
régulier, nous faisons des séries de photographies verticales
vers le haut, prises systématiquement tous les 10 mètres sur les
500 premiers mètres de l'axe principal. L'appareil photo est
orienté perpendiculairement à l'axe principal à environ
50cm du sol. (Le photographe veille à se baisser suffisamment pour
occuper une place minimale sur le cliché (fig.09).

28
Fig. 09 : Photographie en dessous du peuplement ligneux
site 21, 2006
4-6- Dynamique saisonnière des arbres
isolés des sites 17, 21, et 40.
4-6-1 Estimation du PAI des arbres isolés par
photographie hémisphérique
Dans le but de suivre la dynamique saisonnière des
arbres isolés sur les sites précités, une même
méthode a été utilisée durant toute la
période de mesure.
La méthode consiste à prendre une série
de huit photos hémisphériques vers le haut suivant les huit
directions cardinales radiantes à partir du tronc (Nord, Nord Est, Est,
Sud Est, Sud, Sud Ouest, Ouest, Nord Ouest). L'appareil photo est placé
à une distance égale à la moitié du rayon moyen de
la couronne (mesuré dans les huit directions cardinales), et à
une hauteur qui ménage au moins 50cm entre l'objectif et la base du
houppier.
29
4-7- Logiciel et traitement des données.
Le logiciel CAN_EYE développé à l'INRA
Avignon par Frederic Baret et Marie Weiss, version 4.1 est installé sur
1 micro-ordinateur portable de marque DELL. Il est commenté en
anglais.
Le logiciel CAN_EYE permet d'analyser un lot de N
photographies à la fois (N = 20). N varie en fonction de la
résolution de l'image, de la puissance de l'ordinateur et de
l'échantillonnage angulaire choisi pour le traitement. Dans notre
étude, le nombre maximal de photographies qu'il est possible de traiter
en même temps est de 20. Les différentes phases de traitement de
photographies sont :
4-7-1- La formation des lots de
photographies pouvant être traitée en même temps dans un
même répertoire.
Les photographies des herbacées du site 17 sont
regroupées par lot de 20 parce que nous avons au total 100 photographies
par date de mesure. Ceux des ligneux et herbacées du site 21 par lot de
17, nous avons 51 photographies par date de mesure. 4-7-2-
utilisation de CAN EYE
Nous cliquons sur CAN_EYE à partir du répertoire
d'installation, cette page s'ouvre.

Dans notre cas le bouton CAN_EYE Hem permet de lancer le
traitement des photographies, nous cliquons dessus, la page Rechercher le
dossier s'affiche
30
4-7-3 - choix de l'annuaire où les photos sons
stockées.
Nous sélectionnons le lot à traiter puis ok.
La fenêtre CAN_EYE parameterization s'affiche.
4-7-4- Choix de paramètres pour le traitement.
Ici il y a deux paramètres : Sub1 pour le traitement
des lots dont le nombre de photographies est inférieur ou égal
à 8, tous les pixels (ensemble des secteurs angulaires des feuilles)
sont alors traités et Sub2 quand le nombre est supérieur ou
égal à 20. Dans ce cas le logiciel considère 1 pixels sur
2 (sous échantillonnage)
Nous cliquons sur Sub2 et 0K. Toutes les photographies contenues
dans le lot s'affichent.
31
4-7-5- Choix des images à
traiter.
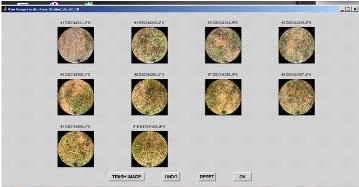
- Le bouton < TRASH IMAGE > permet de supprimer les
images jugées inexploitables,
en les sélectionnant grâce au bouton gauche de la
souris. Le bouton droit sert à sortir du
mode
<TRASH IMAGE>.
- Le bouton <UNDO> annule la dernière action.
- Le bouton < RESET> récupère toutes les
images.
- Le bouton < OK> valide pour passer à
l'étape suivante.
Nous cliquons sur OK et cette fenêtre s'ouvre.
4-7-6- Suppression des endroits jugés
inexploitables.
- le bouton <MASK> permet de réaliser un masque.
L'interface repose sur la souris : le
bouton gauche permet de délimiter la zone à
exclure et le bouton droit de valider.
- < APPLY ALL> : applique le masque présent sur
une image à toutes les images.
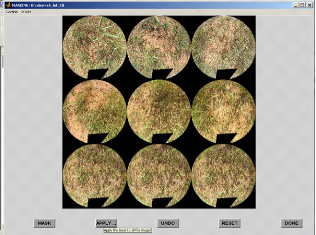
32
Le bouton < DONE> valide le masquage et permet de passer
à l'étape suivante.
La fenêtre «classification Method» s'affiche.
Nous cliquons sur «select pixels corresponding to vegetation » et OK
pour passer à l'étape suivante.
4-7-7- Classification.
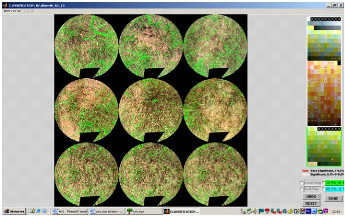
C'est la phase la plus complexe parce que les couleurs
associées à chaque classe dépendent des conditions
d'illumination et de la nature de la végétation.
- Le bouton < TRUE COLOR> permet de visualiser la
photographie en couleur. - Le bouton < CLASSIF> sert à visualiser
l'état actuel de la classification.
33
Il est possible de lancer une classification automatique avec
deux classes : feuille et ciel/sol. Il faut au préalable les avoir
sélectionnées en cliquant sur O. Une fois la classe
sélectionnée nous cliquons avec le bouton gauche sur tous les
pixels de l'image que nous voulons affecter à cette classe puis on
valide avec le bouton droit. La possibilité d'alterner l'affichage de la
classification et de l'image en couleur permet de vérifier visuellement
la pertinence de la classification.
Dans certains cas, nous forçons la classification de
certaines zones de la photo. Par exemple, sur des photos dont le sol est
classé en végétation. Il est donc possible, en cliquant
sur le bouton placé devant sol de délimiter la zone mal
classée dont tous les pixels seront affectés à la classe
sol. Une fois toutes les opérations finies, nous cliquons sur <
DONE> et la fenêtre des résultats s'affiche.
4-7-8 résultats du
traitement.
Le logiciel estime automatiquement de deux façons
différentes le LAI. Soit par l'utilisation du modèle de Poisson
qui considère que les feuilles sont distribuées
uniformément, ce qui permet d'avoir une estimation du LAI effectif, soit
par l'utilisation du modèle de Poisson modifié qui permet de
prendre en compte la disposition non uniforme des éléments du
couvert et d'estimer le LAI vrai (Weiss et al, 2004).
Nous relevons le LAI vrai et le LAI effectif et cliquons sur
YES pour passer au lot suivant.
Les résultats sont stockés dans un sous
répertoire nommé « ce_nom du répertoire de travail
». Il est créé dans le répertoire où se
situent les photographies.
La même méthode est utilisée pour
déterminer le Fcover cependant pour le Fcover nous nous arrêtons
à l'étape de la classification et relevons le pourcentage de
pixels classés en végétation. Ainsi pour le PAI la
différence réside également au niveau de la
classification. Les deux classes choisies sont la classe correspondant à
la végétation et au ciel et non à la
végétation et au sol comme les cas précédents.
34
5- Résultats et discussion.
5-1- Dynamique saisonnière des
herbacés en 2005 et 2006, site 17.
5-1-1 Estimation du LAI et du Fcover par
photographie.
Le LAI est la surface totale d'une face des feuilles par
unité de surface de sol (WASTON, 1947).
Le LAI est exprimé en m2 de feuilles par
m2 de surface et ses valeurs sont très variables d'un couvert
à l'autre, de 0 pour un sol nu, jusqu'à plus de 15 pour des
forêts tropicales très fermées ; jusqu'à 8 pour les
forêts tempérées et de 2 à 4 pour des cultures
annuelles et herbacées annuelles (BEADLE, 1993).
La pluie est un facteur primordial pour la germination et la
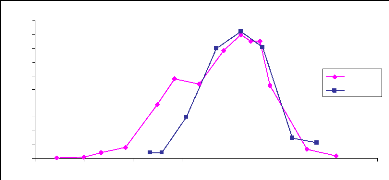
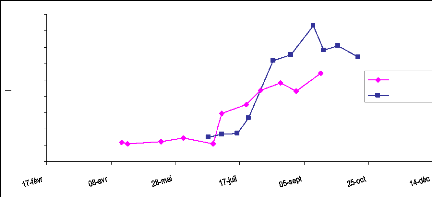
croissance des herbacées dans le Sahel. La courbe de la figure 10 montre
une évolution du LAI sur le site 17 pendant la saison des pluies 2006
(juillet- septembre). Les premières pluies sont tombées sur ce
site le 10 juillet (fig. 11). 4 jours après les premières
germinations sont mises en place et nous avons fait les mesures à cette
date. Une croissance rapide du LAI a été observée de fin
juillet à mi-août conformément à la grande
quantité des pluies enregistrées dans cette période. Le
pic de LAI est atteint le 20 août. Après cette date nous observons
une phase de décroissance du LAI. Cette décroissance marque la
sénescence des herbacées due à la rareté des pluies
et aussi à leur sensibilité au photopériodisme. A la fin
de la saison de pluies la nuit devient longue que le jour, la durée
d'exposition des végétaux à la lumière du jour
diminue, ce phénomène influence sur les végétaux et
les pousse à entrer rapidement en maturité
(photopériodisme). Le sol étant aussi sableux sa capacité
de rétention d'eau est faible et l'évaporation est rapide.
35
Fig.10 : Evolution du LAI en fonction de la
pluviométrie, site 17, 2006.
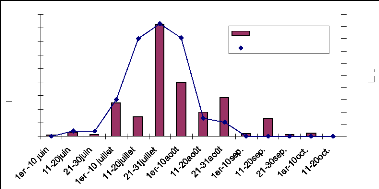
pluviométrie
180
160
140
120
100
40
80
60
20
0
pluviométrie 2006 LAI 2006
2
0,8
0,6
0,4
0,2
0
1,8
1,6
1,4
1,2
1
LAI
Fig.12 : Comparaison du LAI 2005 et 2006, site
17.
LAI
0,8
0,6
0,4
0,2
1,8
1,6
1,4
1,2
2
0
1
28-mai 17-juin 7-juil. 27-juil. 16-août 5-sept. 25-sept.
15-oct.
LAI 2005
LAI 2006
Fig.11 : Pluviométrie 2005 et 2006, site
17.
pluviométrie
180
160
140
120
100
40
20
80
60
0
pluviométrie 2005 pluviométrie
2006
Source : François LAVENU et Marc ARJOUNIN,
2006
36
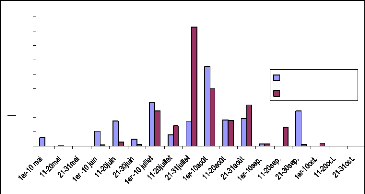
La figure 12 est une comparaison du LAI des deux années
successives 2005 et 2006. En 2005, les pluies sont tombées sur le site
17 dès début juin une phase d'installation a été
observée de mi-juin à fin juin. Une croissance normale est
observée de fin juin à mi-juillet due aux pluies
régulières, suivie d'une légère décroissance
fin juillet à cause de l'absence de pluie. En août une bonne
quantité de pluie a été enregistrée (fig.11), la
croissance a été rapide et le pic est atteint le 20. Pour
l'année 2006, les pluies ont été tardives (mi-juillet)
mais régulières. L`évolution du LAI a été
rapide et le pic est atteint à la même date que l'année
2005. La décroissance du LAI des deux années, présente la
même allure sur les courbes, ceci est dû au phénomène
de photopériodisme et à la rareté des pluies.
Fig.13 : Comparaison du Fcover, 2005 et 2006 site
17.
0,6
Fcover 2005
Fcover 2006
0,5
0,4
0,3
0,2
0,1
0
8-mai 28-mai 17-juin 7-juil. 27-juil. 16-août 5-sept.
25-sept. 15-oct.
Fcover
Le Fcover, étant le pourcentage de la surface au sol
masquée par la végétation, croît en fonction de la
dynamique de la végétation. Au fur et à mesure que les
herbacées croissent, leurs feuilles se multiplient, s'élargissent
et masquent de plus en plus le sol. Malgré les différences
observées sur la dynamique de croissance des végétaux
pendant les deux années (fig.12 et fig. 13) sous l'influence de la
précipitation, les productions végétales obtenues sont
pratiquement identiques.
37
Fig.14 : Relation LAI/Fcover, 2006, site
17.

60,00%
50,00%
40,00%
LAI/Fcover
Logarithmique (LAI/Fcover)
y = 0,1581Ln(x) + 0,3873 R2 = 0,9083
0,00%
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 2
Fcover
30,00%
20,00%
10,00%
LAI vrai
La figure 14 est une droite de régression du LAI et du
Fcover. En effet plus la valeur du coefficient de détermination est
proche de 1, plus les points sont en principe proches de la courbe de
régression et le coefficient de détermination R2
étant égal à 0.90 on déduit qu'il y a une forte
corrélation entre le LAI et le Fcover. A partir de l'équation de
droite y = 1581 Ln(x) + 0,3873 d'où y = Fcover et x= LAI on peut
déterminer un des paramètres en fonction de l'autre.
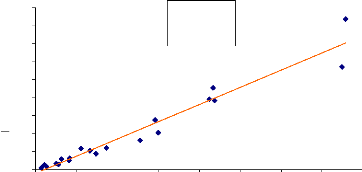
5-2- Validation de CAN EYE.
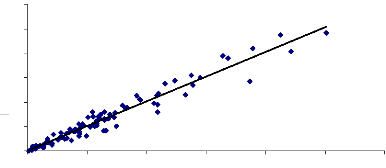
La fig. 15 et fig. 16 sont des droites de régression de
la comparaison du LAI mesuré avec le logiciel can_eye et le LAI
mesuré avec le planimètre des deux années successives 2005
et 2006. Ces droites montrent que les deux méthodes sont
équivalentes.
Y = 1.0136X + 0.0056. Y = LAI planimètre et X = LAI
can_eye.
A partir des données obtenues avec une des
méthodes on peut déduire les données de l'autre.
Et le coefficient de détermination R2= 0.95
(fig.08) montre qu'il y a une corrélation entre les LAI obtenus avec les
deux méthodes.
38
Fig. 15 : comparaison LAI can eye, LAI
planimètre site 17, 2006.
y = 0,9571x - 0,1071
R2 = 0,9479
n=20
3
2,5
2
1,5
1
0,5
0
LAI planimètre
4,5
4
3,5
0 0,5 1 1,5 2 2,5 3 3,5 4
LAI can_eye
-0,5
Fig.16 Comparaison du LAI CAN EYE et LAI
planimètre 2005 et 2006, site 17.
6
0 1 2 3 4 5
6
5
y = 1,0136x + 0,0056
R2 =
0,9536
n=106
4
3
2
LAI planimètre
1
0
LAI can-eye
39
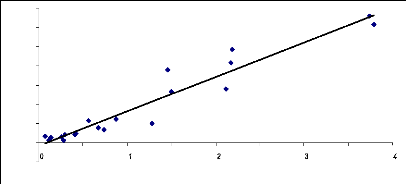
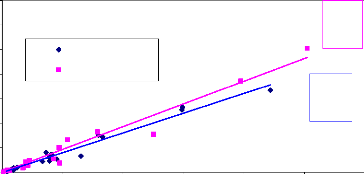
5-3- Comparaison entre le LAI et la biomasse des
grands et petits carrés Fig.17 : Relation entre biomasse et
surface foliaire des grands carrés.
350
300
250
Biomasse (g/m2)
200
150
100
50
0
Relation Biomasse / LAI can-eye
-50
LAI can-eye (m2/m2)
y = 89,584x - 7,9743 R2 = 0,9266
Fig.18 : Relation entre la biomasse et surface
foliaire des petits carrés.
Biomasse (g/m2)
14
12
10
4
2
8
6
0
Dicotylédones Monocotylédones
y = 0,008x R2 = 0,9735
y = 0,0093x R2 = 0,96
0 200 400 600 800 1000 1200
Surface planimétrée
(cm2)
Les droites de régression du LAI can_eye et de la
biomasse foliaire (fig. 17 et fig.18) montrent que les deux méthodes
sont aussi complémentaires. Connaissant le LAI des couverts
herbacés on peut déterminer la biomasse foliaire. Dans le premier
cas (fig. 20) : Y=85.31X, le deuxième (fig.21) : Y = 0.009X et Y =
0.008X
Y= biomasse foliaire et X= LAI can_eye. Et le coefficient de
détermination R2 = 0.87 montre qu'il y a une forte
corrélation entre ces deux paramètres.
40
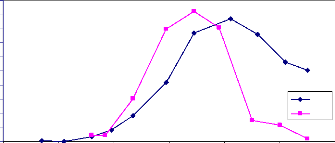
Fig.19 Evolution de la biomasse et du LAI en 2006,
site 17.
biomasse (kg/ha)_LAI
2000
1800
1600
1400
1200
1000
800
600
400
200
0
mbios LAI
2
0,8
0,6
0,4
0,2
0
1,8
1,6
1,4
1,2
1
12/06/2006 02/07/2006 22/07/2006 11/08/2006 31/08/2006 20/09/2006
10/10/2006
La figure 19 montre l'évolution de l'indice foliaire
déterminé par photographie hémisphérique et de la
biomasse par stratification. Les deux courbes présentent la même
allure pendant la phase de croissance quand les feuilles étaient vertes.
A la décroissance le LAI a chuté immédiatement parce que
le logiciel can_eye ne considère que les feuilles vertes. Tandis que la
le poids de la biomasse a décru lentement, les feuilles vertes et jaune
étant fauchées ensemble.
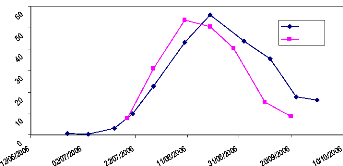
Fig. 20 : Evolution du Fcover site 17,
2006.
%
mrecs Fcover
La figure 20 montre l'évolution du Fcover estimé
à l'oeil nu et du Fcover estimé avec le logiciel can_eye.
L'allure des deux courbes montre que la méthode de détermination
du Fcover avec le logiciel can_eye est coplémentaire à la
méthode classique basée sur l'estimation visuelle.
41
5-4 Dynamique saisonnière des herbacés
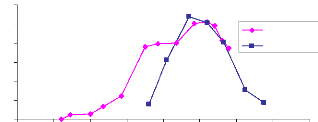
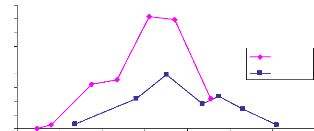
en 2005 et 2006, site 21. Fig. 21 : Comparaison du LAI 2005 et 2006,
site 21.
4,5
4
3,5
LAI 2005
LAI 2006
3
2,5
2
1,5
1
0,5
0
17-juin 07-juil 27-juil 16-août 05-sept 25-sept 15-oct
04-nov
LAI
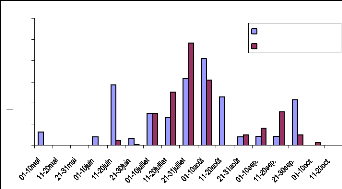
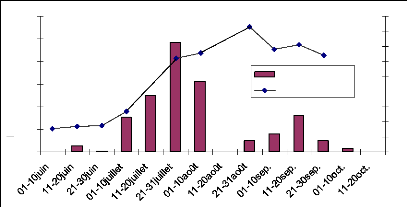
Fig. 22 : pluviométrie 2005 et 2006, site
21.
pluviométrie
120
100
40
80
60
20
0
pluviométrie 2005 pluviométrie
2006
Source : François LAVENU et Marc ARJOURNIN 2005
et 2006
Le site 21 étant situé non loin du site 17, les
précipitations sont assez semblables pour les deux sites. La croissance
du LAI en 2006 est faible et le pic de croissance est atteint le 26 août
avec une valeur de 1.96 (Annexe : tableau V). Cette phase est suivie d'une
décroissance jusqu'à 0.91 de fin août à début
septembre. En mi-septembre le LAI est encore monté à 1,17. Ce
phénomène est dû à la diminution de la
quantité d'eau. En fait le site étant inondé
jusqu'à 50cm (20 août), les herbes sont complètement
immergés dans l'eau , et l'image photographique ne considère que
les feuilles au dessus de l'eau alors le LAI est sous estimé. Fin
août quant les pluies deviennent rare l'eau disparaît et c'est en
ce moment que toutes les feuilles sont photographier permettant une bonne
42
estimation de LAI. La décroissance du LAI a
été lente, elle s'étend de fin août à
mi-octobre (fig.21). Ceci est dû à la structure du sol. Le sol
étant argileux une grande quantité d'eau à
été retenue dans le sous sol et permet aux végétaux
de subsister longtemps sans pluie. Cependant en 2005 le LAI est bien
développé par rapport à 2006. Le pic de croissance atteint
le 18 août et est de 4.1 (tableau V). Du 18 août au 30 août
le LAI a suivi une faible décroissance de 4.1 à 3.97 suivie d'une
forte décroissance du 30 au 16 septembre où le LAI est de 1,1.
Fig. 23 : Evolution du Fcover, site 21,
2006.
70,00%
60,00%
50,00%
Fcover 2006
Fcover
40,00%
30,00%
20,00%
10,00%
0,00%
7-juil. 27-juil. 16-août 5-sept. 25-sept. 15-oct. 4-nov.
Les mesures du Fcover étant faites le même jour,
l'allure de la courbe du LAI est la même que le Fcover à
l'exception d'une date manquante pour le Fcover le 01/10/2006. Aussi les
mesures du Fcover ne sont pas faites sur ce site en 2005 et donc nous ne
pouvons pas faire la comparaison.
y = 0,3159x + 0,0325
R2 = 0,9336
LAI/Fcover
Linéaire (LAI/Fcover)
0 0,5 1 LAI 1,5 2 2,5
|
Fcover
|
70,00% 60,00% 50,00% 40,00% 30,00% 20,00% 10,00% 0,00%
|
Fig. 24 : Relation LAI / Fcover, site 21,
2006.
PAI
0,8
0,6
0,4
0,2
1,8
1,6
1,4
1,2
0
1
PAI 2005 PAI 2006
43
La courbe de régression de LAI et du Fcover (fig.24)
donne un coefficient de
détermination R2 = 0.9336. On peut
déduire qu'il y a une forte corrélation entre ces deux
paramètres. A partir de l'équation de droite
y = 0.3159x + 0.0325 on peut déterminer un
paramètre par rapport à l'autre
y = Fcover et x = LAI.
5-5 Dynamique saisonnière des peuplements
ligneux en 2005 et 2006 site 21.
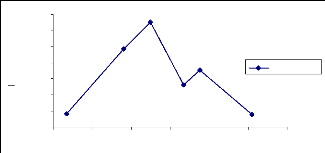
Fig. 25 : Evolution du PAI en 2006, site 21
:
pluviométrie_PAI 2006
120
100
40
80
60
20
0
pluviométrie 2006 PAI 2006
0,8
0,6
0,4
0,2
0
1,8
1,6
1,4
1,2
1
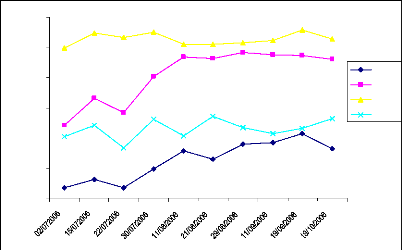
Fig.26 : Comparaison du PAI 2005 et 2006, site
21.
44
Le PAI étant la masse totale de la
végétation ligneuse sur pied, sa masse augmente pendant la saison
de pluies au moment de la feuillaison.
En 2005 les mesures du PAI ont commencé en avril. De
avril à début juin les valeurs du PAI sont restées presque
les mêmes (fig. 26). Vers fin juin après quelques pluies, la
feuillaison a commencé à se mettre en place. On observe donc une
évolution du PAI de fin juin à septembre. Un pic de croissance de
1,08 est enregistré le 29 septembre (annexe : tableau VIII) et les
mesures se sont arrêtées à cette date. En 2006 les mesures
ont commencé le 06 juin, après trois mesures successives la masse
du PAI est restée constante (fig.25). C'est en juillet qu'on observe la
croissance du PAI, due à la mise en place des feuilles sous l'effet des
pluies. Cette phase de croissance s'étale du 07 juillet au 12 septembre
et le PAI est parti de 0,54 au pic de 1,66 (annexe 01, tableau VII). Un pic de
croissance supérieur de 20% par rapport à l'année 2005.
Ensuite on observe une décroissance au bout de 10 jours de 1,66 à
1,36 suivie d'une petite croissance à 1,42 début octobre. Les
mesures du 17 octobre ont donné un PAI de 1,28 ce qui semble indiquer
que nous sommes dans la phase de décroissance du PAI.
45
5-6 Dynamique saisonnière des arbres
isolés, site 17, 2006 Fig. 27 Evolution du PAI des arbres
isolés, site 17, 2006.
PAI
40,00%
20,00%
80,00%
70,00%
60,00%
50,00%
30,00%
AR_1 AS_199 BA_192 CG_302
Les photos des arbres isolés doivent être
traitées avec un logiciel spécifique qui malheureusement
n'était pas encore prêt pour ce stage. En attendant nous avons
fait un premier traitement en utilisant la classification proposée par
can_eye. 4 arbres ont été traités : 2 caducifoliés
(Acacia raddiana N°1 et Acacia senegal N° 199) et 2
sempervirents (Balanites aegyptiaca N° 192 et Combretum
glutinosum N° 302).
Les courbes de la figure 27 montre la dynamique du PAI de ces
4 arbres pendant la saison de pluies 2006. On constate sur cette figure que la
dynamique des sempervirents est faible. Elle est de 8,75 % chez B.
aegyptiaca N° 192 et de 16,83 % chez chez C.Glutinosum
N° 302. Une bonne croissance a été observé avec
les caducifoliés et est de 55,1 % chez A.senegal N° 199 et
75,74 % chez A. raddiana N°1.
46
| 

