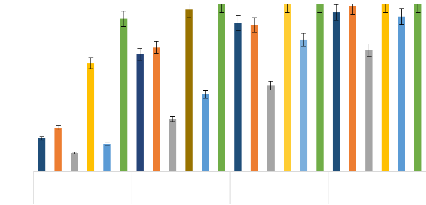Alternative à la résistance aux pyréthrinoà¯des au Burkina Faso: évaluation des activités insecticides et répulsives-irritantes d'huiles essentielles de plantes locales seules et en combinaison sur les populations de anopheles gambiae et de aedes aegypti (diptera: culicidae)par Mahamoudou BALBONE Université Joseph Ki-Zerbo - Doctorat 2022 |

PARTIE 3 : RESULTATS ET DISCUSSIONSThèse de Doctorat unique - 51 - Dans cette partie, nous présenterons les différents résultats obtenus à la suite de nos travaux de recherches puis nous les discuterons. Dans chaque chapitre, nous ferons une introduction où nous indiquerons le contexte scientifique qui a motivé chaque étape de nos recherches. Nous décrierons la méthodologie spécifique utilisée pour l'atteinte des objectifs à chaque niveau de nos travaux. Thèse de Doctorat unique - 52 - Thèse de Doctorat unique - 53 - Chapitre 1 : Activités larvicides de cinq huiles essentielles extraites de cinq plantes locales vis-à-vis des populations de Aedes aegypti et de Anopheles gambiae collectés à l'Ouest du Burkina Faso. Les résultats présentés dans ce chapitre ont fait l'objet d'un article publié dans le journal Frontiers in Tropical Diseases (article 1). 3- 1-1. Introduction Les stratégies actuelles de lutte antivectorielle sont basées principalement sur l'utilisation des insecticides chimiques. Cependant, l'émergence de la résistance aux insecticides chimiques chez les principales espèces de vecteurs de maladies, la pollution de l'environnement et les effets toxiques de ces insecticides chimiques pour l'homme et d'autres organismes non ciblés, en raison de leur large spectre d'activité, compromettent le succès de la LAV (Minjas et Sarda, 1986 ; OMS, 1992 ; Hemingway, 2004). Ce constat a suscité un intérêt accru dans la recherche de produits naturels dérivés de plantes comme alternatifs (Gnankiné et al., 2013 ; Redwane et al., 2002). Parmi les nombreux produits naturels, les huiles essentielles (HE) et leurs constituants chimiques ont fait l'objet d'une attention considérable dans la recherche de nouveaux pesticides. En effet, des études menées au Burkina Faso ont montré qu'ils possédaient un potentiel insecticide en général (Bassolé et al., 2003 ; Drabo et al., 2017 ; Wangrawa et al., 2016 ; Gnankiné et Bassolé, 2017) et des propriétés larvicides en particulier (Yaméogo et al., 2021, Wangrawa et al., 2022a). Par contre, très peu d'études ont concerné la sensibilité des HE aux larves de An. gambiae et de Ae. aegypti, respectivement vecteurs de la dengue et du paludisme, de la partie occidentale du pays. De plus, les études précédentes n'ont pas pris en compte le pyriproxyfène comme contrôle positif dans les différents tests larvaires. Notre travail avait pour objectif d'étudier les propriétés larvicides des HE de cinq (5) plantes aromatiques locales du Burkina Faso, que sont Cymbopogon citratus (D.C.) Stapf., Cymbopogon nardus (Linn.) Rendle, Eucalyptus camaldulensis Dehn., Lippia multiflora Moldenke, Ocimum americanum (Wild.) A.J. Paton, en termes de concentrations létales (CL50 et CL90) sur des populations de terrain de Aedes aegypti et de Anopheles gambiae collectées respectivement à Bobo-Dioulasso et à la Vallée du Kou (VK). 3-1-2. Méthodologie : tests larvaires Les tests ont été réalisés dans l'un des laboratoires d'entomologie médicale de l'IRSS/DRO dans les conditions décrites dans la méthodologie générale. Les tests ont consisté à évaluer la mortalité des larves de An. gambiae et de Ae. aegypti en contact avec des solutions diluées d'HE suivant une méthodologie inspirée du protocole de l'Organisation Mondiale de la Santé (OMS, 2017). Des expériences préliminaires ont permis de retenir une gamme de concentrations devant servir aux tests proprement dits. Des larves de stades 3 et 4 ont été utilisées pour ces tests. Ces larves ont été isolées (Figure 16A) grâce à des tamis et aux critères morphologiques caractérisant celles-ci, puis triées à l'aide des pipettes Pasteur et gardées en observation pendant 30 minutes. Pendant ce temps, les larves qui ont présenté des difficultés de mouvement ou un aspect anormal ont été remplacées. Les tests ont été réalisés dans des gobelets en plastique transparents (Figure 16B) d'une capacité de 200 ml. Une quantité appropriée de chaque HE a été dissoute dans de l'acétone pour préparer 1 ml de solution mère, en fonction des concentrations voulues, 50, 100, 150 et 200 ppm. Dans chaque gobelet contenant initialement un volume d'eau distillée de 149 ml avec 20 larves de moustiques (Figure 16B), 1 ml de la solution mère d'HE préparée avec l'acétone a été ajouté (Figure 16C) afin d'obtenir les concentrations finales voulues dans un volume total de 150 ml. Pour chaque concentration, quatre (4) répétitions ont été faites plus quatre (4) témoins. Dans les témoins, 20 larves ont été introduites dans 149 ml d'eau distillée et 1 ml d'acétone. Le pyriproxyfène a été utilisé comme témoin positif. Les larves mortes et les larves moribondes (celles qui bougeront lentement après agitation de l'eau) considérées comme mortes ont été dénombrées pour l'évaluation de la mortalité larvaire 24 heures après leur mise en contact avec les HE. Thèse de Doctorat unique - 54 -
A B C Figure 16: Quelques étapes du test larvaire A : Tri des larves de stades 3 et 4 ; B : Les larves au repos dans l'eau distillée ; C : Ajout du mélange d'acétone et l'huile essentielle aux concentrations 50, 100, 150 et 200 PPM (Cliché Balboné, 2020). 3- 1-3. Résultats 3- 1-3.1. Caractérisation chimique des huiles essentielles Les différents composés de chacun des 5 huiles essentielles sont résumés dans le tableau II. L'HE de C. citratus s'avère riche en géranial (48,1%), néral (35,8%) et myrcène (11%). Celle de C. nardus est principalement composée de citronellal (41,7%), de géraniol (20,8%) et de f3-élémène (11%). L'HE de E. camaldulensis était riche en 1,8-cinéole (59,5%) et en á-pinène (9,17%). L'HE de L. multiflora était caractérisée par du p-cymène (25,27%), du f3-caryophyllène (12,7%) et du thymol (11,88%). L'HE de O. americanum se caractérise par un pourcentage élevé de 1,8-cinéole (31,22%) suivi du camphre (12,73%). Thèse de Doctorat unique - 55 - Tableau II : Composition chimique des huiles essentielles des 5 plantes Cymbopogon citratus, Cymbopogon nardus, Eucalyptus camaldulensis, Lippia multiflora et Ocimum americanum
NB : Les composes en gras représentent les composes majeurs de chaque HE ; JR : Jndice de rétention Thèse de Doctorat unique - 56 - 3- 1-3.2. Activités larvicides des huiles essentielles sur Anopheles gambiae Les huiles essentielles (HE) des cinq (5) espèces végétales ont démontré une activité larvicide vis-à-vis des larves de stades 3 et 4 des deux (2) souches de An. gambiae, la souche sensible aux insecticides chimiques (Kisumu) et celle collectée à la Vallée du Kou (VK). Dans l'ensemble, toutes les cinq (5) HE ont été plus toxiques sur les larves des stades 3 et 4 de la souche sensible Kisumu avec des valeurs de CL50 et de CL90 qui ont été respectivement de 3,9 à 75,8 ppm et de 29,5 à 193,5 ppm (Tableau III). Sur les larves de An. gambiae de VK, les valeurs des CL50 et CL90 étaient comprises entre 39,5 et 138,1 ppm et 90,2 et 328,9 ppm, respectivement (Tableau IV). Tableau III : Les concentrations létales à 50 et 90 % (CL50 et CL90) et leurs intervalles de confiance des huiles essentielles de plantes contre les larves de la souche sensible de An. gambiae Kisumu
CL50 et CL90 : Concentration létal à 50 et 90 ; IC : Intervalle de confiance ; ppm : Partie par million. Les produits dont les CL50 sont suivies de lettres alphabétiques identiques ne diffèrent pas statistiquement par le test (HSD) de Tukey p<0,001. Tableau IV : Les concentrations létales à 50 et 90 % (CL50 et CL90) et leurs intervalles de confiance des huiles essentielles des plantes contre les larves de la souche résistante de An. gambiae de VK
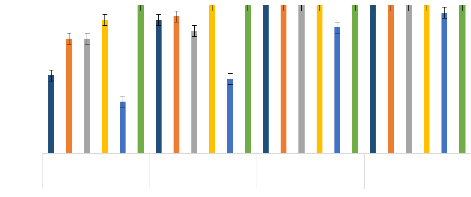
CL50 et CL90 : Concentration létal à 50 et 90 ; IC : Intervalle de confiance ; ppm : Partie par million. Les produits dont les CL50 sont suivies de lettres alphabétiques identiques ne diffèrent pas statistiquement par le test (HSD) de Tukey p<0,001. Thèse de Doctorat unique - 57 - Thèse de Doctorat unique - 58 - L'HE de L. multiflora a été plus efficace sur les 2 souches de An. gambiae avec des valeurs de CL50 et de CL90, qui ont été respectivement de 3,9 et 29,5 ppm pour les larves de Kisumu et, de 39,5 et 90,2 ppm pour les larves de VK. L'HE de O. americanum a montré la plus faible activité larvicide sur Kisumu la souche sensible aux insecticides chimiques avec des valeurs de CL50 = 75,8 ppm et de CL90 = 193,5 ppm, tandis que celle de C. citratus a montré la plus faible activité larvicide sur les larves de la souche de terrain VK avec des valeurs CL50 = 138,1 ppm et CL90 = 328,9 ppm. En outre, sur les larves de Kisumu, les HE de E. camaldulensis, C. nardus et C. citratus ont montré de fortes activités larvicides après l'HE de L. multiflora, avec des valeurs CL50 < 50 ppm et CL90 < 91 ppm. Sur les larves VK, les HE de C. nardus, de O. americanum et de E. camaldulensis ont montré une forte activité larvicide après l'HE de L. multiflora avec des valeurs CL50 <140 ppm et une valeur CL90 < 270 ppm. Seule l'HE de L. multiflora a donné des valeurs CL50 et CL90 proches de celles du témoin positif, le pyriproxyfène, sur les larves de Kisumu et de VK. En effet, au niveau des valeurs CL50 et CL90, aucune différence significative n'a été observée entre L. multiflora et le pyriproxyfène sur les deux souches de An. gambiae testées (p = 0,95) avec un chevauchement des intervalles de confiance. En ce qui concerne les mortalités obtenues sur les larves de Kisumu la souche sensible aux insecticides chimiques, l'HE de L. multiflora a montré une mortalité de 90 et 100% respectivement à 50 et 100 ppm (Figure 17). Elle a été suivie par les HE de C. citratus, E. camaldulensis et C. nardus qui ont donné des mortalités supérieures à 50% déjà à 50 ppm et supérieures à 80% à la concentration 100 ppm. L'HE de O. americanum avait donné la mortalité la plus faible (35% à 50 ppm et 50% à 100 ppm). Toutes les HE ont donné 100% de mortalités à partir de 150 ppm (Figure 17). Sur les larves de la souche VK, seule l'HE de L. multiflora a donné une mortalité supérieure à 65% à 50 ppm, pendant que les autres HE ont donné des mortalités inférieures à 30% (Figure 18). A 100 ppm, l'HE de L. multiflora a donné des mortalités de 87,1% suivie de l'HE de C. nardus qui a donné des mortalités de 64,58%. Tandis que les trois (3) autres HE ont donné des mortalités inférieures à 50%. A 150 ppm, l'HE de L. multiflora a donné des mortalités de 100% et celles de C. nardus et O. americanum ont donné des mortalités supérieures à 80% suivies de l'HE de E. camaldulensis dont les mortalités produites ont été supérieures à 50%. A 200 ppm, toutes les cinq (5) HE testées ont donné des mortalités supérieures à 85% (Figure 18). Souche sensible de An. gambiae "Kisumu"
Mortalités 100 40 90 70 20 60 50 30 80 10 0
Traitements et concentrations Figure 17: Mortalités des larves de la souche de An. gambiae « Kisumu » sensible aux insecticides chimiques en fonction des concentrations des HE. Les barres d'erreurs représentent l'intervalle de confiance à 95%. Les mortalités moyennes (#177;intervalle de confiance) ont été obtenues au seuil de probabilité de 5% d'après le test LSD de Fisher (p?0,05).
Souche de An. gambiae collectée à VK
Traitements et concentrations Figure 18: Mortalités des larves de la souche de terrain de An. gambiae « VK » en fonction des concentrations des HE. Les barres d'erreurs représentent l'intervalle de confiance à 95%. Les mortalités moyennes (#177;intervalle de confiance) ont été obtenues au seuil de probabilité de 5% d'après le test LSD de Fisher (p?0,05). Thèse de Doctorat unique - 59 - 3- 1-3.3. Activités larvicides des huiles essentielles sur Aedes aegypti Toutes les cinq (5) HE testées ont montré une toxicité contre les larves de stades 3 et 4 des deux (2) souches de Ae. aegypti, sensibles (Bora Bora) et collectée sur le terrain à Bobo-Dioulasso (Tableaux V et VI ; Figures 19 et 20) . Comme dans le cas des populations de An. gambiae, parmi les HE, l'HE de L. multiflora a été la plus toxique sur les deux (2) souches de Ae. aegypti testées. Cette toxicité est perçue à travers les valeurs des concentrations létales (CL50 et CL90) et les mortalités produites sur les différentes souches de moustiques testées. Tableau V : Les concentrations létales à 50 et 90 % (CL50 et CL90) et leurs intervalles de confiance des huiles essentielles contre les larves de la souche de Ae. aegypti sensible Bora Bora aux insecticides chimiques
Les différences significatives entre les produits testés par le test LSD de Fisher (P<0,05) ; Les produits dont les CL50 sont suivies de lettres alphabétiques identiques ne diffèrent pas statistiquement par le test (HSD) de Tukey p<0,001. CL50 et CL90 : Concentration létal à 50 et 90 ; IC : Intervalle de confiance ; ppm : Partie par million Tableau VI : Les concentrations létales à 50 et 90 % (CL50 et CL90) et leurs intervalles de confiance des huiles essentielles contre les larves de la souche de Ae. aegypti de Bobo-Dioulasso
Les différences significatives entre les produits testés par le test LSD de Fisher (P<0,05) ; Les produits dont les CL50 sont suivies de lettres alphabétiques identiques ne diffèrent pas statistiquement par le test (HSD) de Tukey p<0,001. CL50 et CL90 : Concentration létal à 50 et 90 ; IC : Intervalle de confiance ; ppm : Partie par million Thèse de Doctorat unique - 60 - Thèse de Doctorat unique - 61 - En effet, les valeurs de CL50 et CL90 ont été respectivement de 42 et 74,6 ppm pour l'HE de L. multiflora testées sur les stades larvaires 3 et 4 de la souche de terrain de Ae. aegypti collectée à Bobo-Dioulasso. Ces valeurs ont été légèrement plus faibles que celles obtenues sur les larves des mêmes stades de la souche sensible Bora Bora (Tableaux V et VI). Également, les valeurs CL50 et CL90 obtenues avec l'HE de L. multiflora sur les larves de Bobo-Dioulasso ont été significativement inférieures à celles obtenues avec les autres HE testées pour lesquelles les valeurs de CL50 et CL90 ont varié respectivement de 63,5 à 103,8 ppm et de 148,5 à 311,3 ppm sur la souche collectée à Bobo-Dioulasso (Tableau VI). Sur la souche Bora bora, les valeurs de CL50 et CL90 ont varié respectivement de 61,5 à 101,3 ppm et de 132 à 188,8 ppm pour les HE de C. citratus, C. nardus, E. camaldulensis, et O. americanum (Tableau V). L'HE de E. camaldulensis reste la moins toxique sur les deux souches de Ae. aegypti au regard des valeurs CL50 et CL90 qui ont été respectivement de 101,3 à 188 ppm pour Boro Bora et de 103,8 à 311,3 ppm pour la souche collectée à Bobo-Dioulasso. L'HE de L. multiflora et le pyriproxyfène à la CL90 sur Bora Bora ont présenté un niveau de toxicité élevé avec un chevauchement des limites de confiance à 95%, leurs taux de mortalités a été supérieur à 95% à 100 ppm. Nous constatons qu'il n'y a pas de différence significative entre l'HE de L. multiflora et le pyriproxyfène sur la souche sensible de Ae. aegypti, Bora Bora (F= 0,02; DDL=8, p= 0,97). De même, il n'y a pas eu de différences significatives entre C. citratus et C. nardus concernant les valeurs CL50 et CL90 sur les deux souches de Ae. aegypti testées. De même, il n'y a pas eu de différences significatives entre les HE de E. camaldulensis et O. americanum concernant la CL90 sur les larves des deux souches de Ae. aegypti testées et la CL50 sur les larves de Bora Bora. Aussi, sur les larves de la souche sensible Bora Bora, l'HE de L. multiflora a montré une mortalité de 67,5 et 98,75% respectivement à 50 et 100 ppm. A partir de 150 ppm, cette HE a donné 100% de mortalités sur les mêmes larves (Figure 19). Par contre, les autres HE ont donné des mortalités inférieures à 40% à 50 ppm. A 100 ppm, les HE de C. citratus et C. nardus ont donné respectivement des mortalités de 77,5 et 90% sur les larves de Bora Bora. Cependant, les mortalités des HE de E. camaldulensis et O. americanum ont été inférieures à 50% à 100 ppm. A partir de 150 ppm, toutes les HE ont donné des mortalités supérieures à 80% (Figure 19). Thèse de Doctorat unique - 62 - Sur les larves de la souche de Bobo-Dioulasso, les mortalités obtenues à 50 ppm avec l'HE de L. multiflora ont été de 64,65% tandis que celles des autres HE ont été inférieures à 30%. Les mortalités obtenues à 100 ppm avec l'HE du L. multiflora étaient supérieures à 95% et à partir de 150 ppm, elles ont été de 100%. Les mortalités obtenues avec les HE de C. citratus et C. nardus ont été respectivement de 70 et 74,13% à 100 ppm et à partir de 150 ppm, elles ont été supérieures à 85% (Figure 20). Tandis que celles obtenues avec les HE de E. camaldulensis et O. americanum ont été inférieures à 50% à 100 ppm et à 150 ppm, elles ont été respectivement de 51,25 et 78,75% (Figure 20). A 200 ppm, les mortalités de toutes les HE ont été supérieures à 90% sauf celles de l'HE de E. camaldulensis. Souche sensible de Ae. aegypti "Bora Bora" Traitements et concentrations
Taux de mortalité 100 40 20 90 70 60 30 80 50 10 0
Figure 19: Mortalités en fonction des concentrations des HE des larves de la souche de Ae. aegypti Bora Bora sensible aux insecticides chimiques. (Les barres d'erreurs représentent l'intervalle de confiance à 95%). Les mortalités moyennes (#177;intervalle de confiance) ont été obtenues au seuil de probabilité de 5% d'après le test LSD de Fisher (p?0,05). Souche de Ae. aegypti collectée à Bobo-Dioulasso
Taux de mortalité 100 40 20 90 70 60 50 30 80 10 0
Traitement et concentrations Thèse de Doctorat unique - 63 - Figure 20: Mortalités en fonction des concentrations des HE des larves de la souche de Ae. aegypti de Bobo-Dioulasso. (Les barres d'erreurs représentent l'intervalle de confiance à 95%). Les mortalités moyennes (#177;intervalle de confiance) ont été obtenues au seuil de probabilité de 5% d'après le test LSD de Fisher (p?0,05). 3- 1-4. Discussion A l'heure actuelle, les moustiquaires imprégnées d'insecticide à longue durée d'action (MILD) et les pulvérisations intra domiciliaires à effet rémanent (PID) restent les deux principaux outils ciblant principalement les adultes de An. gambiae et, dans une moindre mesure, Ae. aegypti. Ainsi, il y a un besoin urgent de développer des outils et des techniques innovantes, y compris l'utilisation des HE contre les oeufs, les stades larvaires et les adultes afin d'augmenter le succès de la lutte antivectorielle (Bassolé et al., 2003 ; Yaméogo et al., 2021). Le statut de résistance des populations du complexe An. gambiae aux insecticides recommandés pour la lutte anti vectorielle a été signalé dans la partie occidentale du Burkina Faso (Dabiré et al., 2014 ; Namountougou et al., 2019). Aussi, d'autres études ont confirmé le statut de résistance de la population de Ae. aegypti de la ville de Bobo-Dioulasso (Namountougou et al., 2020). Thèse de Doctorat unique - 64 - Notre travail visait à étudier la bio-efficacité de cinq (5) HE de plantes locales sur les larves de Ae. aegypti et de An. gambiae selon les méthodes standard de l'Organisation Mondiale de la Santé (OMS). Il est connu que la plupart des études sur les tests larvicides, n'ont pas pris en compte le Pyriproxyfène comme témoin chimique, un Régulateur de Croissance des Insectes (RCI). Les RCI agissent en imitant l'une des hormones de croissance des insectes ou en perturbant directement la formation de la cuticule ou la biosynthèse des lipides. Le pyriproxyfène est recommandé par l'OMS contre les larves de moustiques à des concentrations comprises entre 0,02 et 0,05 mg/L (5 à 10 g/ha) avec un temps d'activité de 6 semaines (OMS, 2001, 2006). Dans cette étude, toutes les cinq HE testées ont montré une activité insecticide variable contre les larves de An. gambiae et de Ae. aegypti. En ce qui concerne les valeurs CL50 et CL90, les HE de C. citratus, C. nardus, E. camaldulensis, L. multiflora et O. americanum, ont présenté une activité larvicide plus élevée contre les larves de troisième et quatrième stade des souches de laboratoire et de terrain de An. gambiae et Ae. aegypti après 24 heures d'exposition. La plus efficace de toutes ces HE a été celle de L. multiflora. La CL50 de l'HE de E. camaldulensis a été la moins efficace sur Ae. aegypti, alors que l'HE de C. citratus reste la moins efficace sur An. gambiae. Ces données ne sont pas en accord avec celles trouvées par Ho Dung Manh et al. (2020) qui ont rapporté que sur les larves de Ae. aegypti, l'HE de C. citratus (CL50 = 120,6 ppm) est moins efficace que celle de E. camaldulensis (CL50 = 33,7 ppm). Nos résultats obtenus avec l'HE de C. nardus sur An. gambiae dans cette étude sont inférieurs à ceux obtenus par Ahouansou et al. (2019) qui ont obtenu une valeur de 97,3 ppm pour la CL50 sur An. gambiae. Nos résultats sont similaires à ceux rapportés par Cavalcanti et al. (2004) sur les populations de Ae. aegypti. Dans leur étude, les HE de O. americanum et C. citratus ont donné des valeurs de CL50 égales à 67 et 69 ppm respectivement. Cependant, nos résultats obtenus avec l'HE de O. americanum sont inférieurs à ceux rapportés par Wangrawa et al. (2016) sur les lerves de An. gambiae dont la CL50 était de 209,84 ppm. Les valeurs de CL50 et CL90 obtenues avec l'HE de L. multiflora dans notre étude sont plus faibles que celles rapportées par Bassolé et al. (2003). Récemment, la même tendance a été observée avec Yameogo et al. (2021) qui ont également montré l'effet toxique de l'HE de L. multiflora contre les populations de Ae. aegypti de Tabtenga à Ouagadougou. Il est intéressant de noter que l'activité élevée de l'HE du L. multiflora peut résulter de la présence de trois composants majeurs : le thymol, le p-cymène et le â-caryophyllène. Selon Thèse de Doctorat unique - 65 - Folashade et Omoregie (2012), le thymol, le germacrène D, le ñ-cymène, l'acétate de thymyle et le sabinène ont été trouvés dans l'HE de L. multiflora au Ghana et semblent être impliqués dans son activité contre les larves de moustiques. Inversement, l'activité larvicide plus faible des HE de C. citratus, C. nardus, E. camaldulensis et O. americanum peut s'expliquer par l'effet mineur de la pipéritone, du géraniol et du 1,8-cinéole sur les larves de Ae. aegypti et de An. gambiae (Bossou et al., 2013). La diversité de composition chimique des HE testées pourrait expliquer les différences observées dans leurs activités larvicides observées dans cette étude. En ce qui concerne les valeurs CL50 et CL90, la bio-efficacité de l'HE de L. multiflora sur les larves des souches de An. gambiae est plus proche de celle du pyriproxyfène, le contrôle positif utilisé dans cette étude qui est le larvicide de référence. Cela ouvrirait une nouvelle alternative pour le contrôle des moustiques. Globalement, dans les populations des deux (2) souches de An. gambiae, les larves de la souche Kisumu se sont avérées les plus sensibles aux HE que celles de la population de terrain (VK), connues pour leur résistance aux pyréthroïdes (Namountougou et al., 2019 ; Chandre, et al., 1999 ; Poda et al., 2018). Comme obtenu avec An. gambiae, les tests avec les larves de Ae. aegypti ont aussi montré une sensibilité de l'HE de L. multiflora proche de celle du pyriproxyfène. Les valeurs de CL50 trouvées dans cette étude ont été légèrement inférieures à celles observées dans les populations de Ae. aegypti de Ouagadougou (Yaméogo et al., 2021) qui pourrait s'expliquer par le développement très prononcée de la résistance phénotypique chez les populations de Ae. aegypti de Ouagadougou aux principaux produits chimiques. Cette résistance phénotypique aux insecticides chimiques chez les populations de Ae. aegypti de Ouagadougou ont été montré par Namountougou et al. (2020) à travers les tests réalisés avec la deltaméthrine sur des populations de Ae. aegypti collectées à Bobo-Dioulasso et à Ouagadougou. En effet, ces études ont montré des taux de mortalité atteignant 89,62% et 82,72% respectivement pour 2013 et 2014, avec les populations de Ae. aegypti collectées à Bobo-Dioulasso et 50,7% et 20,7% respectivement pour 2013 et 2014, avec Ae. aegypti collectées à Ouagadougou. Toutes ces valeurs étaient inférieures à 90%, suggérant une résistance. Il est très urgent de rechercher des alternatives aux produits chimiques. Nos données permettront de développer des stratégies innovantes à base d'HE. 3- 1-5. Conclusion partielle Nos résultats ont montré que l'utilisation des HE comme larvicides est une stratégie prometteuse. Par conséquent, les HE pourraient faire l'objet d'une attention particulière dans la recherche de nouveaux produits larvicides naturels, sélectifs et biodégradables qui peuvent être utilisés dans les programmes de santé publique de lutte antivectorielle contre An. gambiae et Ae. aegypti en particulier. Dans la perspective de valorisation de ces HE, il serait intéressant d'évaluer l'efficacité de ces HE sur les adultes de ces deux populations de moustiques. Thèse de Doctorat unique - 66 - Thèse de Doctorat unique - 67 - Chapitre 2 : Évaluation des propriétés adulticides des huiles essentielles de cinq plantes locales et des combinaisons de deux d'entre elles sur des populations de Aedes aegypti et de Anopheles gambiae (Diptera : culicidae) de l'Ouest du Burkina Faso. Les résultats présentés dans ce chapitre ont fait l'objet de deux articles publiés dans Journal of Medical Entomology (Article 2) et Scientific report (Article 3). 3- 2-1. Introduction Les moustiquaires et la PID à effet rémanent sont les méthodes sur lesquelles se basent la LAV actuelle (OMS 2017). Les insecticides chimiques de la famille des pyréthrinoïdes sont ceux utilisés en santé publique pour l'imprégnation des moustiquaires et la PID (OMS, 2019). Cependant, la résistance aux pyréthrinoïdes constitue une menace pour l'efficacité de ces outils de protection, surtout lorsque la résistance est présente à des niveaux élevés (Churcher et al. 2016 ; Hemingway et al. 2016). Et au Burkina Faso, nous crayons que l'objectif de l'utilisation des MILDA à base de pyréthrinoïdes ne soit pas atteint (Dabire et al. 2012b). Les extraits de plantes tels que les HE peuvent être utilisés comme alternative aux insecticides chimiques auparavant utilisés dans la LAV (Zoubiri et Baaliouamer 2014. Tripathi et al., 2009 ; Balbone et al., 2022a). Des études antérieures avaient montré les propriétés adulticides des HE sur des moustiques (Chansang et al., 2018; Wangrawa et al., 2018; Yaméogo et al., 2021). Tout récemment, les études de Balboné et al. (2022a) ont montré des activités larvicides intéressantes sur les populations de Anopheles et Aedes au Burkina Faso. D'autres études avaient montré que la toxicité des HE est liée à certains de leurs composés bioactifs (Sarma et al., 2019 ; Isman, 2000). Certaines études avaient également tenté de formuler des insecticides efficaces en combinant différents extraits de plantes avec des insecticides chimiques pour augmenter la toxicité globale et minimiser les effets secondaires (Mansour, Foda et Aly, 2012). Bien que des études récentes aient montré que des combinaisons des HE de L. multilora et de Cymbopogon schoenanthus ont donné des effets synergiques contre les moustiques des populations de An. gambiae de la souche sensible Kisumu (Wangrawa et al., 2022b), peu de connaissances sont disponibles sur l'effet du mélange d'HE sur les populations de An. gambiae résistantes et les populations sensibles et résistantes de Ae. aegypti. Thèse de Doctorat unique - 68 - Dans cette partie nous présenterons les principaux résultats des tests adulticides des HE de Cymbopogon citratus, Cymbopogon nardus, Eucalyptus camaldulensis, Lippia multiflora, et Ocimum americanum et des combinaisons des HE de Cymbopogon nardus et Ocimum americanum sur des populations sensibles et résistantes de Ae. aegypti et de An. gambiae, respectivement de la ville de Bobo-Dioulasso et de la Vallée du Kou, à l'Ouest du Burkina Faso. 3- 2-2. Méthodologie : test de sensibilité en tubes OMS Les tests de sensibilité des HE sur les populations de An. gambiae et de Ae. aegypti ont été réalisés à l'aide des kits de tests de sensibilité aux insecticides de l'OMS et selon les procédures standard (OMS, 2017) (Figure 21). Pour chaque concentration d'HE, quatre (4) lots de 20 à 25 femelles de moustiques à jeun âgés de 2 à 3 jours ont été introduits dans des tubes OMS contenant les papiers imprégnés où ils ont été mis en contact forcé avec le produit pendant une (1) heure, temps pendant lequel le nombre de moustiques femelles ayant subi un effet Knock Down (KD), c'est-à-dire les moustiques qui sont tombés sur le dos ou sur le côté, a été évalué à des intervalles de 5 minutes. Pour chaque concentration, un lot de 25 femelles de moustiques à jeun âgés de 2 à 3 jours, servant de témoin, a toujours été placé dans un tube contenant du papier neutre. Après le temps d'exposition aux HE ou aux combinaisons d'HE, les moustiques ont été transférés dans les tubes d'observation, contenant des papiers neutres, puis nourris avec du jus sucré (glucose 5%) et conservés pendant 24h. L'effet létal des HE ou des combinaisons d'HE testé a été observé à partir des taux de mortalité déterminés. Les concentrations appliquées sur les adultes lors de ces tests en tubes OMS sont 0,1%, 0,5% et 1%. La perméthrine a été utilisée comme témoin positif. Pour chaque HE ou combinaison d'HE testée, les souches de An. gambiae « Kisumu » et de Ae. aegypti « Bora bora » ont servi de souches de référence sensible. Lorsque la mortalité des moustiques témoins était inférieure à 5%, le test était validé. Par contre, si elle était comprise entre 5 et 20%, la mortalité des moustiques exposées aux insecticides était corrigée par la formule d'Abbott (Abbott, 1925). Si la mortalité des témoins était supérieure à 20%, le test était repris. Pour la combinaison des deux (2) HE, neuf (9) combinaisons ont été obtenues selon les proportions de chaque HE dans la combinaisons : i) C.n 10 % : O.a 90 % ; ii) C.n 20 % : O.a 80 % ; iii) C.n 30 % : O.a 70 % ; iv) C.n 40 % : O.a 60 % ; v) C.n 50 % : O.a 50 % ; vi) C.n 60 % : O.a 40 % ; vii) C.n 70 % : O.a 30 % ; viii) C.n 80 % : O.a 20 % et ix) C.n 90 % : O.a 10 % correspondant respectivement à C1, , C3, C4, C5, C6, C7, C8 et C9.
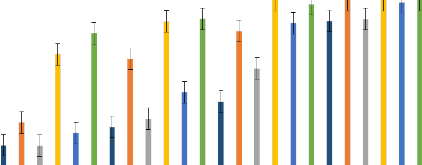
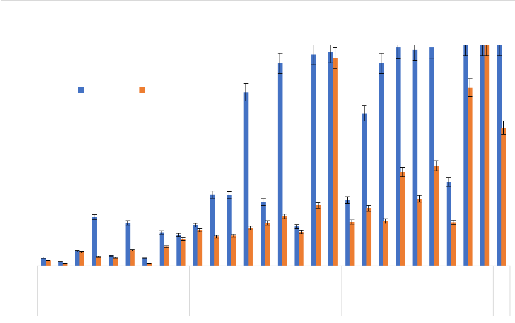
A B C Figure 21: Quelques étapes du déroulement des tests bioessais A : Aspiration des moustiques femelles de 3 à 5 jours non gorgés ; B : Mise en contact des moustiques avec les papiers imprégnés ; C : Mise en observation des moustiques pour 24 heures après le contact avec les papiers imprégnés (Cliché Balboné, 2020) 3- 2-3. Résultats 3- 2-3.1. Caractérisation chimique des huiles essentielles Les différents composés chimiques de chacune des cinq huiles essentielles sont résumés dans le tableau II. La description de leurs composés majeurs a été faite dans le chapitre 1 partie 31.3.1. Thèse de Doctorat unique - 69 - Thèse de Doctorat unique - 70 - 3- 2-3.2. Tests adulticides sur les populations de Anopheles gambiae Les HE et les combinaisons d'HE testées ont présenté une activité adulticide plus ou moins efficaces sur les deux souches de An. gambiae. Cette activité a été perçue à travers les mortalités des HE/combinaisons et leurs concentrations létales. 3- 2-3.2.1. L'activité des huiles essentielles Avec les HE seules, les mortalités obtenues sur la souche de An. gambiae de terrain VK ont varié de 0,0 à 13,16 % à la concentration 0,1 % ; de 1,12 à 23,56 % à la concentration 0,5% et de 3,16 à 96,88 % à la concentration 1 % (Figure 22). L'HE de L. multiflora seule a montré une forte activité avec une mortalité de 100 et 96,88 % à la concentration de 1 %, respectivement sur la souche sensible Kisumu et la souche de terrain VK. De même, l'activité élevée de l'HE de L. multiflora a été observée avec des valeurs CL50 de 0,21 et 0,67 %, de CL99 de 0,74 et 1,17 %, respectivement sur les souches Kisumu et VK, (Tableau VII). Également, cette HE a donné les plus faibles valeurs de KDT50 et KDT95 sur les deux souches de An. gambiae testées (KDT50 = 19,8 et 58,8; KDT95 = 87,2 et 142,7 min, respectivement pour Kisumu et VK à la concentration plus faible, 0,1 %) (Tableau VIII). L'HE de E. camaldulensis a été la moins toxique avec une mortalité inférieure à 10 % à la concentration de 1 % sur la souche sensible Kisumu (Figure 22), des valeurs de KDT50 et KDT95 élevée (KTD50 = 144,8 et 166,1 min, respectivement pour les souches Kisumu et VK à la concentration la plus élevée, 1 %) et des valeurs CL50 de 2,86 et 2,94 %, CL99 de 5,05 et 5,26 %, respectivement sur les souches Kisumu et VK. De plus, les HE de C. nardus (mortalités de 86,36 et 40,63 %, CL50 de 0,63 et 1,12 %, respectivement sur Kisumu et VK) et de O. americanum (mortalités de 41,49 et 6,38 %, des CL50 de 2,18 et 2,21 %, respectivement sur Kisumu et VK) ont des activités adulticides plus élevées que celle de l'HE de C. citratus (mortalités de 35,29 et 4,76 %, CL50 de 2,3 et 2,7% sur Kisumu et VK, respectivement) (Figure 22 et Tableau VII). Pour la souche sensible Kisumu, aucune différence significative n'a été observée avec la CL50 et la CL99 pour les HE de C. citratus, E. camaldulensis et O. americanum. Cependant, sur la souche de terrain VK, il y avait une différence significative avec toutes les HE (Tableau VII). La concentration diagnostique a été obtenue à partir de deux fois la CL99 sur la souche sensible (OMS, 2017). La concentration diagnostique la plus faible (1,48%) a été obtenue avec l'HE de L. multiflora. Cette concentration diagnostique n'a pas été significativement différente de la concentration diagnostique de la perméthrine 0,75%, le contrôle positif. Les concentrations diagnostiques les plus élevées ont été obtenues avec les HE de O. americanum (9,18%), C. citratus (9,4%) et E. camaldulensis (10,1%). Mortalité des adultes de Anopheles gambiae aux huiles essentielles 100 Kisumu VK 90 80 70 60 50 40 30 Taux de mortalité
10
Les traitements et les concentrations (%)
0
Figure 22: Taux de Mortalité des adultes de An. gambiae Kisumu et VK en fonction des concentration des HE. (Les barres d'erreurs représentent l'intervalle de confiance à 95%). Les mortalités moyennes (#177;intervalle de confiance) ont été obtenues au seuil de probabilité de 5% d'après le test LSD de Fisher (p?0,05). Thèse de Doctorat unique - 71 - Tableau VII : Concentrations létales 50 et 99 (CL50, CL99) et concentrations diagnostiques pour les huiles essentielles et les combinaisons de C. nardus et O. americanum testées sur Anopheles gambiae
CL = Concentration létale ; IC = Intervalle de confiance ; Cn = Cymbopogon nardus; Oa = Ocimum americanum Thèse de Doctorat unique - 72 - Tableau VIII: KDT50, KDT95 obtenus avec les huiles essentielles des 5 plantes sur les deux souches de Anopheles gambiae (Kisumu et de VK) Traitements Anopheles gambiae « Kisumu » Anopheles gambiae « VK » Doses KDT50 KDT95 KDT50 KDT95 (%) (min) (min) (min) (min) C. citratus 0,1 103,24 204,66 140,2 318,7 0,5 77,10 155,86 124,9 275,8 1 60,19 132,39 116,5 233,6 C. nardus 0,1 88,37 196,99 148,6 241,8 0,5 33,3 108,9 90,2 149,5 1 5,8 11,5 12,4 70,7 E. camaldulensis 0,1 192,2 289,7 227,3 309,7 0,5 157,1 275,1 189,9 286,7 1 144,8 266,5 166,1 280 L. multiflora 0,1 19,8 58,8 87,2 142,7 0,5 10,5 19 32 57,7 1 -28,3 -8,2 -5,9 1,5 O. americanum 0,1 137,5 225,6 156,4 267,5 0,5 87,06 146,64 144,7 248,9 1 30,04 60,76 116,3 204,6 Permethrine 0,75 16,3 30 41,2 100 KDT50 : temps nécessaire pour assommer 50% des moustiques et KDT95 : temps nécessaire pour assommer 95% des moustiques 3.2.3.2.2. L'activité des combinaisons des huiles essentielles En utilisant les combinaisons sur les deux souches de An. gambiae (Kisumu et VK), les mortalités sur la souche de terrain VK ont varié de 1,15 à 12,15 %; 5,21 à 94 % et 9,62 à 100 %, respectivement aux concentrations de 0,1 ; 0,5 et 1 % (Figure 23). Les mortalités sur la souche Kisumu ont été plus remarquables aux mêmes concentrations (Figure 23). Thèse de Doctorat unique - 73 -
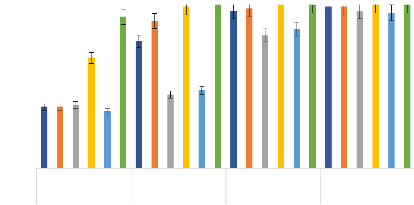
100 90 Mortalité des adultes de Anopheles gambiae aux combinaisons d'HE de C. nardus et de O. americanum Kisumu VK 80 70 Taux de mortalité 60 50 40 30 20 10 0 C1 C3 C4 C5 C6 C7 C8 C9 C1 C3 C4 C5 C6 C7 C8 C9 C1 C3 C4 C5 C6 C7 C8 C9 0,5 Traitements et concentrations (%) 1 0,75 0,1 Perm Figure 23: Taux de Mortalité des adultes de An. gambiae de Kisumu et VK en fonction des concentration des combinaisons des HE de C. nardus et O. americanum. (Les barres d'erreurs représentent l'intervalle de confiance à 95%). Les mortalités moyennes (#177;intervalle de confiance) ont été obtenues au seuil de probabilité de 5% d'après le test LSD de Fisher (p?0,05). La combinaison C9 présentait une toxicité élevée sur toutes les souches. En effet, cette combinaison a donné une mortalité de 100% à la concentration de 1% sur les deux souches de An. gambiae testées, 96,7 et 94 % à la concentration de 0,5 %, respectivement sur les souches Kisumu et VK. De plus, C9 a présenté les plus faibles valeurs de KDT (Tableau IX) et les plus faibles valeurs de CL50 et CL99 sur les deux souches (CL50 de 0,22 et 0,32 %, CL99 de 0,52 et 0,59 %, respectivement sur Kisumu et VK) (Tableau VIII). Également, C9 a été la seule combinaison qui a produit un effet synergique sur les deux souches (Tableaux X et XI). En outre, les combinaisons C6 et C8 ont présenté une toxicité élevée avec des taux de mortalité supérieurs à 98% à la concentration de 1 % sur la souche Kisumu. Sur la souche VK, la toxicité de ces deux combinaisons était relativement élevée avec des mortalités de 45,21 et 80,7 Thèse de Doctorat unique - 74 - Thèse de Doctorat unique - 75 - % à la concentration de 1%, respectivement pour C6 et C8. Avec ces deux combinaisons, les valeurs de KDT étaient légèrement faibles sur les deux souches (KDT95 = 20.7 min 12.2 min à la concentration de 1% sur la souche VK, respectivement pour C6 et C8) aussi, les CL50 et CL99 étaient légèrement élevées par rapport à C9 sur les deux souches (CL50 = 1.08 et 0.89%, CL99 =1.3 et 2.15%, respectivement). Sur la souche Kisumu, C8 a produit un effet synergique et C6 un effet additif (Tableau X). Les combinaisons , C4 et C6 ont donné des valeurs CL50 et CL99 respectivement inférieures à 1,5 et 2,5% avec la souche VK. Toutes les autres combinaisons ont donné des CL50 supérieures à 2 et des CL99 supérieures à 5%, sauf C5 qui a donné 1,36% comme CL50 et 3,26% pour CL99 sur la souche VK (Tableau VIII). Les plus faibles concentrations diagnostiques de 1,04%, 1,16% et 1,48% ont été obtenues respectivement avec les C9 et C8, et l'HE de L. multiflora (Tableau VIII). Les CL50 et CL99 obtenues sur les deux souches de An. gambiae ont été significativement plus faibles pour les combinaisons C9, C8 et C6 que celles obtenues avec chacune des 2 HE qui ont servi à la combinaison (Tableau VIII). Les valeurs de CL50 obtenues avec les combinaisons C8 sur la souche VK n'ont montré aucune différence avec celles obtenues avec l'HE de C. nardus avec des intervalles de confiance qui se chevauchent, de même les CL50 obtenues avec C3, C4, C5 et C7 n'ont montré aucune différence avec celles obtenues avec l'HE de O. americanum sur la souche VK (p < 0,0001) (Tableau VIII). Les ratios de résistance obtenus avec toutes les HE ainsi qu'avec toutes les combinaisons étaient inférieurs à 5 (Tableau VIII). Cela indique que même s'il y a une résistance avec certaines HE et les combinaisons, elle serait très faible. L'HE L. multiflora et la combinaison C9 ont produit des mortalités plus élevées et des KDT plus faibles à la concentration de 0,5% que la perméthrine 0,75%. Les concentrations diagnostiques les plus faibles ont été obtenues respectivement avec C9 et C8 avec des valeurs de 1,04 et 1,16 % respectivement. Ces deux concentrations diagnostiques n'ont pas été significativement différentes de la concentration diagnostique de perméthrine 0,75%, le contrôle positif (p = 0,91). Les concentrations diagnostiques les plus élevées ont été obtenues avec C1 (5,9%) et C7 (8,6%). Thèse de Doctorat unique - 76 - Tableau IX: KDT50, KDT95 avec les combinaisons d'huiles essentielles de C. nardus et de O. americanum sur les souches de Anopheles gambiae (Kisumu et VK)
KDT50 : temps nécessaire pour assommer 50% des moustiques et KDT95 : temps nécessaire pour assommer 95% des moustiques Thèse de Doctorat unique - 77 - Tableau X: Effets des interactions des HE de Cymbopogon nardus (C.n) et Ocimum americanum (O.a) dans les combinations sur les adultes de la souche sensible de An. gambiae Kisumu (n = 150).
FIC = FICCn + FICOa; Synergiste: FIC < 0.5; Additif; 05 = FIC = 1; Sans effet: 1 = FIC = 4; Antagoniste: FIC > 4. Tableau XI: Effets des interactions des HE de Cymbopogon nardus (C.n) et Ocimum americanum (O.a) dans les combinations sur les adultes de la souche de An. gambiae de VK (n = 150).
FIC = FICCn + FICOa; Synergiste: FIC < 0.5; Additif; 05 = FIC = 1; Sans effet: 1 = FIC = 4; Antagoniste: FIC > 4. Thèse de Doctorat unique - 78 - 3- 2-3.3. Tests adulticides sur les populations de Aedes aegypti 3- 2-3.3.1. L'activité des huiles essentielles Le tableau XII résume les temps nécessaires pour assommer 50 et 95% de moustiques (KDT50 et KDT95) avec les intervalles de confiance de 95% (IC95%) obtenus sur les deux souches de Ae. Aegypti testées (Bora bora et de Bobo-Dioulasso) avec les HE. Sur la souche de Bobo-Dioulasso, les valeurs de KDT50 et KDT95 variaient respectivement de 234,1 à 113,5 min et 343,9 à 182,8 min à la concentration 0,1%, de 209,4 à 43 min et 297,2 à 75 min à 0,5% et de 190,5 à 20,5 et 287 à 32,8 à 1%. Sur cette souche, les valeurs de KDT les plus faibles ont été obtenues avec l'HE de L. multiflora, suivies par les HE de C. nardus et O. americanum, et les valeurs de KDT les plus élevées ont été obtenues avec l'HE de E. camaldulensis. À la concentration la plus élevée (1%), il n'y a pas de différences significatives entre les valeurs de KDT obtenues avec les HE testées sur les deux souches de Ae. aegypti, à l'exception de celles obtenues avec les HE de C. nardus et de L. multiflora. Sur la souche sensible Bora Bora, les valeurs du KDT50 et du KDT95 des HE de L. multiflora et de C. nardus ont été respectivement de -3,9 ; 7,8 min et 24 ; 33,7 min à une concentration de 1%. Sur la souche sensible Bora Bora, les KDT ont été les plus faibles et les mortalités les plus élevées que celles obtenues sur la souche Bobo-Dioulasso. Tableau XII : les temps nécessaires pour assommer 50 et 95% (KDT50 et KDT95) avec les intervalles de confiance à 95% (IC95%) obtenus sur les deux souches de Ae. Aegypti testées (Bora bora et de Bobo-Dioulasso) avec les HE. Aedes aegypti "Bora bora" Aedes aegypti "Bobo-Dioulasso" KDT50 KDT95 (min) (min) (min) (min) Huiles essentielles Concentrations KDT50 KDT95 C. citratus 0,1 193,3 291,1 221,2 311,2 0,5 192,6 290,2 228,1 298,5 1 140,6 230,4 142,5 244,8 C. nardus 0,1 88,6 134 179 278 0,5 35,6 53,8 88,7 143,9 1 24 33,7 35,6 57,2 E. camaldulensis 0,1 211,3 299,1 234,1 343,9 0,5 178,1 285,7 209,4 297,2 1 156,5 267,5 190,3 287,2 L. multiflora 0,1 107 178,7 113,5 182,8 0,5 40,9 72 43 75 1 -3,9 7,8 20,5 32,8 O. americanum 0,1 131,57 194,82 151,7 264,3 0,5 128,1 188,4 145,3 247,5 1 125,7 210,6 133,3 249 Permethrin 0,75 16,6 29,1 144 188,7 KDT50 et KDT95 : Temps de knock down de 50 et 95% de moustiques ; min : minute Les mortalités (Figure 24) et les valeurs de CL50 et CL99 (Tableau XIII) ont varié en fonction des HE testées, de leurs concentrations et de la souche de Ae. aegypti sur laquelle les HE ont été testées. Comme dans le cas de An. gambiae, L'HE de L. multiflora a été toujours celle qui a l'activité la plus élevée quelle que soit la souche de Ae. aegypti sur laquelle elle a été testée. En effet, elle a produit les mortalités les plus élevées et les concentrations létales les plus faibles (mortalité = 85% à la concentration de 1% ; CL50 = 0,79% et CL99 = 1,25 pour la souche de Bobo-Dioulasso et une mortalité de 100% à la concentration 1% avec pour CL50 = 0,57% et CL99 = 1,03 pour la souche Bora Bora). A la concentration 1%, l'HE de C. nardus a produit des mortalités de 20,34% et 12,75%, respectivement sur les souches Bora Bora et Bobo-Dioulasso. L'HE de E. camaldulensis a l'activité la plus faible avec les plus faibles mortalités sur les deux Thèse de Doctorat unique - 79 - souches (7,14 et 0,7% à la concentration 1%, respectivement sur les souches de Bora Bora et Bobo-Dioulasso). La concentration diagnostique la plus faible de 2,06 %, a été obtenue avec l'HE de L. multiflora. La concentration diagnostique la plus élevée a été obtenue avec l'HE de E. camaldulensis (15,62 %). Sur les deux souches de Ae. aegypti, les mortalités obtenues avec l'HE de L. multiflora ont été significativement plus élevées que celles obtenues avec la perméthrine 0,75%.
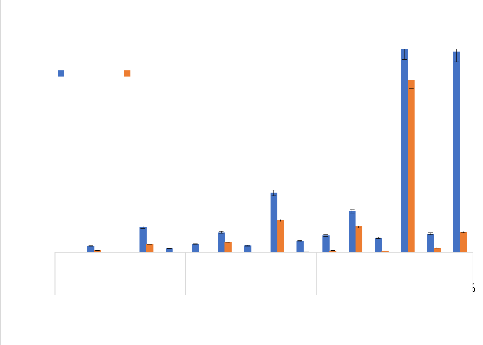
Mortalités des adultes de Aedes aegypti aux HE 100 Bora Bora Bobo-Dioulasso 90 80 70 60 50 40 30 20 Mortalités
Traitements et concentrations 10 0 Figure 24: Mortalité des adultes de Ae. aegypti en fonction des concentrations des HE. (Les barres d'erreurs représentent l'intervalle de confiance à 95%). Les mortalités moyennes (#177;intervalle de confiance) ont été obtenues au seuil de probabilité de 5% d'après le test LSD de Fisher (p?0,05). Thèse de Doctorat unique - 80 - Tableau XIII : Concentrations létales 50 et 99 (CL50, CL99) et concentrations diagnostiques pour les huiles essentielles et les combinaisons de C. nardus et O. americanum testées sur Aedes aegypti
CL = Concentration létale ; IC = Intervalle de confiance ; Cn = Cymbopogon nardus; Oa = Ocimum americanum Thèse de Doctorat unique - 81 - 3- 2-3.3.2. L'activité des combinaisons des huiles essentielles Les valeurs des KDT50 et KDT95 (Tableau XIV) et les mortalités obtenues avec les combinaisons préparées à partir des HE de C. nardus et de Ocimum americanum sur les deux souches de Ae. aegypti ont varié selon les proportions des HE combinées et les concentrations des combinaisons (Figure 25). Mortalité des adultes dAedes aegypti aux
combinaisons d'HE de C.
Taux de mortalité 70 60 50 40 30 20 10 0 100 90 Bora Bora Bobo-Dioulasso 80 C1 C3 C4 C5 C6 C7 C8 C9 C1 C3 C4 C5 C6 C7 C8 C9 C1 C3 C4 C5 C6 C7 C8 C9 1 0,7 0,1 0,5 Perm Traitements et les concentrations (%) Figure 25: Mortalité des adultes de Ae. aegypti en fonction des concentration des combinaisons des HE de C. nardus et O. americanum. (Les barres d'erreurs représentent l'intervalle de confiance à 95%). Les mortalités moyennes (#177;intervalle de confiance) ont été obtenues au seuil de probabilité de 5% d'après le test LSD de Fisher (p?0,05). Thèse de Doctorat unique - 82 - Thèse de Doctorat unique - 83 - Sur les deux souches de Ae. aegypti, les valeurs de KDT les plus faibles ont été obtenues par la combinaison C8, suivie de . Par contre, les valeurs les plus élevées ont été obtenues avec C3 et C1. En effet, les valeurs de KDT50 et KDT95 avec C8 ont été respectivement de 48,9 et 77 min à la concentration 1% sur la souche Bora Bora, 60,6 et 79,8 min sur la souche de Bobo-Dioulasso. Globalement, toutes les combinaisons ont montré de faibles valeurs de KDT. En effet, sur les deux souches de Ae. aegypti, les valeurs de KDT95 ont été inférieures à 60 min à 1%, sauf pour les combinaisons C3 et C1 qui ont montré des valeurs de KDT95 respectivement de 186 et 172.4 min à 1%. Les valeurs de KDT obtenues avec les combinaisons C8, et C9 ont été significativement plus faibles que celles des deux (2) HE testées individuellement à toutes les concentrations. En ce qui concerne les mortalités, les valeurs les plus élevées à la concentration de 1% ont été de 100 et 86,67% sur la souche Bora Bora et 95,09 et 81,99% sur la souche Bobo-Dioulasso obtenues respectivement avec les combinaisons C8 et . Quant à la combinaison C9, elle a donné une mortalité de 51,76% à la concentration de 1%. C'est seulement avec ces trois (3) combinaisons, , C8 et C9, que les mortalités obtenues ont été significativement plus élevées que celles obtenues avec les 2 HE testées séparément. Les mortalités les plus faibles ont été obtenues avec la combinaison C1 (2,66 et 1,6% à la concentration de 1%, respectivement sur les souches de Bora Bora et de Bobo-Dioulasso). Les effets interactifs entre les deux HE dans les combinaisons (Tableaux XV et XVI) ont été obtenus à partir des indices de concentration inhibitrice/létale fractionnelle (FIC). Ces effets de combinaison ont été classés comme synergistes, additifs, indifférents ou antagonistes. Les combinaisons les plus efficaces en ce qui concerne les KDT, les concentrations létales et les mortalités ont été celles qui donnaient des effets synergiques et additifs. En effet, l'effet synergique a été obtenu sur les deux souches avec la combinaison C8 et l'effet additif a été trouvé avec les combinaisons et C9. Le tableau XIII résume les CL50 et CL99 ainsi que les concentrations diagnostiques pour toutes les cinq (5) HE testées et les combinaisons préparées avec les HE de C. nardus et O. americanum. Sur les deux souches de Ae. aegypti, les CL50 obtenues avec les combinaisons C8, et C9 ont été inférieures à 1%, tandis que celles de C4 et C6 ont été légèrement inférieures à 2%. Les concentrations diagnostiques les plus faibles de 2,16 % et 2,56 % ont été obtenues respectivement avec les combinaisons C8 et . Les concentrations diagnostiques les plus élevées ont été obtenues avec C5 (19,4 %) et C1 (12,72 %). A l'exception de C5 et C1, toutes les combinaisons ont donné des concentrations diagnostiques plus faibles que celles des HE de C. nardus et O. americanum. Tableau XIV : KDT50, KDT95 produits par les combinaisons d'huiles essentielles de C. nardus et de O. americanum sur les souches de Aedes aegypti (Bora bora et de Bobo-Dioulasso)
KDT50 et KDT95: temps de knock down de 50 et 95% de moustiques ; min : minute Thèse de Doctorat unique - 84 - Tableau XV : Effets des interactions des huiles essentielles de C. nardus (C.n) et O. americanum (O.a) dans les combinations sur les adultes de la souche sensible de Aedes aegypti Bora Bora (n = 150).
FIC = FICCn + FICOa; Synergiste: FIC < 0.5; Additif; 05 = FIC = 1; Sans effet: 1 = FIC = 4; Antagoniste: FIC > 4.
FIC = FICCn + FICOa; Synergiste: FIC < 0.5; Additif; 05 = FIC = 1; Sans effet: 1 = FIC = 4; Antagoniste: FIC > 4. Thèse de Doctorat unique - 85 - Thèse de Doctorat unique - 86 - 3- 2-4. Discussion Jusqu'à présent, la gestion de la résistance aux insecticides reste un défi majeur pour parvenir à une élimination efficace du paludisme (OMS, 2012). En effet, la résistance aux pyréthrinoïdes a été signalée dans 27 pays d'Afrique sub-saharienne, soulignant l'urgence de trouver des alternatives à ces insecticides (Tchoumbougnang et al., 2009 ; Ranson et Lissenden, 2016). La recherche d'alternatives aux insecticides chimiques à base d'extraits de plantes peut représenter de nouvelles pistes pour le contrôle des populations de moustiques vecteurs du paludisme. Ici, nous avons évalué le niveau d'efficacité de cinq (5) HE, à savoir les HE de Cymbopogon citratus, Cymbopogon nardus, Eucalyptus camaldulensis, Lippia multiflora et Ocimum americanum sur des populations de An. gambiae et de Ae. aegypti. Aussi, l'efficacité des combinaisons de C. nardus (C.n) et O. americanum (O.a) et des effets de ces HE dans la combinaison ont été examinés dans cette étude. Globalement, presque toutes les HE testées ont montré un effet adulticide sur les souches de An. gambiae sensible Kisumu et résistante collectée à VK et sur les souches de Aedes aegypti sensible Bora Bora et résistante collectée à Bobo-Dioulasso. Cet effet insecticide, mis en évidence par des effets « knock-down », des concentrations létales et des mortalités des moustiques adultes testés, a varié significativement selon les concentrations et les HE utilisées. Les résultats de la présente étude confirment partiellement les travaux de Yameogo et al. (2021) qui ont montré les effets adulticides de certaines HE identiques testées sur des populations de Ae. aegypti collectées à Ouagadougou. Ils confirment également les travaux réalisés par Drabo et al. (2017) qui ont montré les propriétés adulticides des HE de C. citratus et O. americanum sur des populations de Bemisia tabaci, confirmant ainsi l'efficacité de ces HE sur une large gamme d'insectes. En effet, parmi les HE testées, celle de L. multiflora a été la plus efficace sur les adultes des deux souches de An. gambiae testées, suivie de l'HE de C. nardus. L'HE de E. camaldulensis a été la moins efficace. Il est intéressant de noter que l'HE de L. multiflora peut constituer une bonne alternative au regard de la mortalité à la concentration 1% qui atteignait 96,88% et 85%, respectivement sur les souches résistantes de An. gambiae collectée à VK et de Ae. aegypti collectées à Bobo-Dioulasso. L'efficacité de l'HE de L. multiflora dans la présente étude a été significativement supérieure à celles des HE de Lantana camara, Hyptis spicigera, Hyptis Thèse de Doctorat unique - 87 - suaveolens évaluées sur des souches de An. gambiae, Kisumu et VK dans les travaux de Wangrawa et al. (2018). La différence relative de l'efficacité entre les HE testées dans nos travaux pourrait s'expliquer par leur composition chimique. En effet, plusieurs études antérieures ont montré que les composés majeurs étaient le facteur responsable de la bio-activité d'une HE, car ils constitueraient la partie la plus importante des composés totaux de l'HE (Lucia et al., 2007 ; Gbolade et Lockwood, 2008 ; Vani, Cheng et Chuah, 2009). Ainsi, l'efficacité de l'HE de L. multiflora peut être due à ses principaux composés qui sont le ß-caryophyllène, le p-cymène, l'acétate de thymol et le 1,8 cinéol. En effet, selon des études antérieures réalisées par Bassolé et al. (2003), la toxicité de l'HE de L. multiflora sur les larves de An. gambiae et de Ae. aegypti est due à la présence de trois composants majeurs : le thymol, le p-cymène et l'acétate de thymol. L'HE de Hyptis suaveolens contenant également du â-caryophyllène et 1,8 cinéol (Peerzada, 1997) a une activité insecticide élevée (Ilboudo et al. 2010). Ainsi, l'activité insecticide élevée de l'HE de L. multiflora a été attribuée au â-caryophyllène et au 1,8 cinéol qui sont des composés majeurs dans l'étude actuelle. Cependant, la faible activité insecticide de l'HE de E. camaldulensis peut être expliquée par l'absence de â-caryophyllène, dans la mesure où cette HE était riche en 1,8 cinéol. Également, l'HE de C. nardus a montré un effet adulticide intermédiaire sur les populations de An. gambiae testées contrairement à ce qui a été trouvé par Zulfikar, Aditama et Sitepu (2019). Selon ces auteurs, l'activité la plus élevée de C. nardus est due à la présence de géraniol, le même composé trouvé dans C. nardus testé dans notre étude. D'autres études antérieures ont rapporté que les propriétés insecticides de l'HE de O. americanum est dues aux composés 1,8-cinéole, camphre et á-pinène trouvés dans cette HE (Koul, Walia et Dhaliwal, 2008). Ces composés ont été les mêmes que ceux trouvés dans l'HE de O. americanum testée dans la présente étude et pour lesquels nous avons obtenu la faible efficacité. Selon Ntonga et al. (2017), le 1,8-cinéole contribue fortement à l'efficacité de l'HE de Callistemon citrinus contre les adultes femelles de An. coluzzii. Cependant, l'HE de E. camaldulensis, riche en 1,8-cinéole, était la moins efficace sur les adultes femelles de Ae. aegypti. Cette faible efficacité peutt être due à l'absence du composé sabinène parmi les principaux composés de l'HE de E. camaldulensis. En effet, Liu et al. (2020) ont rapporté qu'il y a un effet additif entre le 1,8-cinéole et le sabinène sur Sitophilus oryzae, un ravageur du riz. Thèse de Doctorat unique - 88 - Les HE qui ont été moins efficaces comme adulticides dans la présente étude pourraient être efficaces comme répulsifs. En effet, des études précédentes ont montré l'effet répulsif de ces HE sur les moustiques (Ojewumi, Adeyemi et Ojewumi, 2018 ; Gnankiné et Bassolé, 2017 ; Bossou et al., 2013). Les valeurs de KDT de ces HE principalement à la concentration de 1% peut justifier cette propriété répulsive mise en évidence par l'activité excito-répulsive. Si la préexposition des moustiques au PBO peut restaurer totalement ou partiellement la susceptibilité aux pyréthrinoïdes (Hien et al., 2021), il est fort probable que l'efficacité d'une HE peut être améliorée par ses combinaisons avec d'autres HE. Dans la présente étude, cette amélioration a été mise en évidence par les valeurs KDT, CL50, CL99 et les mortalités obtenues avec certaines combinaisons qui ont été supérieures à celles des HE testées seules. En termes d'efficacité après l'HE de L. multiflora, les HE de C. nardus et O. americanum ont donné des taux de mortalités relativement élevées. C'est pourquoi, dans cette étude, nous avons examiné les effets de leurs combinaisons à travers des concentrations binaires. En effet, les combinaisons qui ont été plus efficaces que les HE de C. nardus et O. americanum testées individuellement étaient C6, C8 et C9 sur les populations de An. gambiae et, et C8 sur Ae. aegypti. Cela peut etre dû à l'effet toxique simultané des composés des HE combinées. Ceci est en accord avec les travaux de Bekele et Hassanali (2001) et de Pavela (2014) qui ont rapporté que l'activité biologique des HE dépend non seulement de leur composition qualitative, mais aussi du rapport quantitatif de leurs composants. On peut penser que l'HE de C. nardus a besoin d'une certaine quantité des principaux composés de l'HE de O. americanum pour être plus efficace. Inversement, l'HE de O. americanum a besoin de certains constituants de l'HE de C. nardus pour agir efficacement. En effet, des études ont montré que l'ajout d'un peu de citronellal renforce l'effet du mélange citral-myrcène contre Ae. aegypti. Cependant, le citronellal seul ne peut pas réduire de manière significative le comportement de recherche d'hôte des moustiques (Hao et al. 2008). L'amélioration de l'efficacité par les combinaisons des 2 HE peut être expliquée par l'effet toxique combiné des principaux composés. En effet, des études ont montré que la toxicité des HE est due aux effets combinés de différents composants, avec ou sans action toxique individuelle significative contre les insectes (Tanprasit, 2005 ; Park et al. 2011). Selon Burt (2004), les HE individuelles contiennent des composants complexes qui, lorsqu'ils sont combinés entre eux, peuvent entraîner des effets indifférents, additifs, synergiques ou antagonistes. Dans Thèse de Doctorat unique - 89 - une étude antérieure, Abbassy, Abdelgaleil et Rabie (2009) ont rapporté que dans certains cas, l'HE entière peut avoir une activité insecticide plus élevée que ses composants majeurs isolés. Pour ces auteurs, les composés mineurs sont essentiels pour l'activité et peuvent permettre un effet synergique ou une influence potentielle. A partir de ces informations, nous pouvons dire que les combinaisons qui ont montré un effet synergique ou additionnel auraient comme composés majeurs, d'une part certains composés majeurs de l'HE de C. nardus et d'autre part ceux de l'HE de O. americanum, mais aussi une variété de composés mineurs. Des études précédentes ont montré que la bio-activité de l'HE est une conséquence de l'interaction entre les composants majeurs, mais aussi d'autres composés, éventuellement des oligoéléments. Cela peut être dû à des effets combinés, à une action additive entre les classes chimiques et à une synergie ou à un antagonisme (Bassolé et al., 2003 ; Chiasson et al., 2001 ; Luz et al., 2020). En général, il semble que l'effet d'un composé actif puisse être renforcé par d'autres composés majeurs et/ou stimulé par des composés mineurs pour donner des effets additifs ou synergiques (Deletre et al., 2015). Par conséquent, selon Berenbaum et Neal (1985), les composants mineurs présents en faibles pourcentages peuvent agir comme des synergistes, améliorant la bio-efficacité des composants majeurs par divers mécanismes. En effet, des combinaisons de 1,8-cinéole et de citronellal ainsi que de plusieurs composés mineurs et de citronellal ont donné des effets synergistes sur les larves de Spodoptera littoralis (Pavela, 2014). Selon Hyldgaard, Mygind et Meyer (2012), chaque composé d'une HE avait un mode d'action spécifique, mais ces composés pouvaient interagir avec les composés d'une autre HE dans une combinaison pour donner d'autres modes d'action. Ainsi, la combinaison de composés avec différents modes d'action pourrait augmenter les actions létales totales dans les mélanges (Sarma et al., 2019). Il est intéressant de noter que seules les combinaisons C8 et C9 ont fourni un effet synergiste respectivement sur les populations résistantes de Ae. aegypti et de An. gambiae. L'effet synergiste obtenu par cette combinaison peut également s'expliquer par l'action simultanée des principaux composés. De plus, cet effet synergiste peut être dû à l'apparition de nouveaux composants qui n'existaient pas dans les HE individuelles (Muturi et al., 2017; Wangrawa et al., 2022b). Des études antérieures ont montré que certaines HE ou leurs composés peuvent interagir pour créer un effet synergiste à certains ratios et un effet antagoniste à d'autres ratios (Intirach et al., 2012; Pavela, 2015). 3- 2-5. Conclusion partielle Notre étude a examiné les propriétés adulticides des huiles essentielles de cinq (5) espèces de plantes traditionnellement utilisées au Burkina Faso, Cymbopogon nardus, Cymbopogon citratus, Eucalyptus camaldulensis, Lippia multiflora et Ocimum americanum, et des combinaisons de deux (2) de ces HE contre les adultes de moustiques de Anopheles gambiae, vecteur du paludisme et de Aedes aegypti, vecteur de la dengue au Burkina Faso. Les HE de Cymbopogon nardus et Ocimum americanum ont été moins toxiques, mais leurs toxicités ont été améliorées par leurs combinaisons, à certaines proportions. L'HE de L. multiflora et les combinaisons de C9 (Cn 90% : Oa 10%), C8 (Cn 80% : Oa 20%), C6 (Cn 60% : Oa 40%) et dans une moindre mesure (Cn 20% : Oa 80%) peuvent être des alternatives prometteuses dans les stratégies de contrôle contre les adultes de Ae. aegypti, le vecteur de la dengue et de An. gambiae, vecteur du paludisme. Pour une valorisation de ces HE testées, il serait bon d'examiner leur activité excito-répulsive sur les deux populations de moustiques, comme c'est le cas des pyréthrinoïdes qui malgré la présence de résistance vis-à-vis de certains insectes, peuvent procurer une activité excito-repulsive ou irritante qui peuvent être exploitée dans les stratégies de lutte antivectorielle. Thèse de Doctorat unique - 90 - Thèse de Doctorat unique - 91 - Chapitre 3 : Activité excito-répulsive de cinq huiles essentielles extraites de plantes locales contre les vecteurs de la dengue et du paludisme au Burkina Faso 3- 3-1. Introduction partielle Dans les chapitres précédents, nous avons expliqué le contexte qui a motivé l'utilisation des huiles essentielles comme insecticides alternatifs ou complémentaires. Vu que l'homme n'est pas permanemment sous les MILDA et surtout que certains moustiques ont des activités diurnes (Ae. aegypti), il est donc nécessaire de chercher des outils de protection individuelle tels que les répulsifs. Les répulsifs sont des produits qui agissent localement ou à distance afin d'empêcher un arthropode de se poser sur la peau d'un homme ou d'un animal (ou sur toute autre surface en général), de voler vers lui ou de le piquer (Boulanger et Gentile, 2012). Des études ont montré que la perméthrine et le DEET, deux produits chimiques, sont des répulsifs (Badolo et al., 2004; Deletre et al., 2019; Achee et al., 2009). D'autres études ont montré des pouvoirs répulsifs des plantes productrices d'HE sur les espèces d'arthropodes nuisibles (Deletre et al., 2013, 2016; Tawatsin et al., 2001; Gillij, Gleiser et Zygadlo, 2008; Omolo et al., 2004; Oyedele et al., 2002) et les propriétés de dissuasion de l'alimentation des moustiques (Hummelbrunner et Isman, 2001; Moretti, Zerba et Alzogaray, 2013). Cinq phénomènes répulsifs ont été identifiés par Deletre et al. (2016) : répulsion-expulsion, répulsion-masquage, répulsion-irritation, répulsion-anti-appétence et répulsion visuelle. Un répulsif-expulsion est une substance qui agit à distance, faisant fuir l'insecte du stimulus (Bernier et al., 2006; Nerio, Olivero-Verbel et Stashenko, 2009 ; Nolen et al., 2002). Un répulsif masquant est défini comme un inhibiteur volatil qui empêche la recherche et la localisation de l'hôte (OMS, 2013b). Un répulsif irritant appelé «inhibiteur d'atterrissage» selon l'OMS (Koul, Walia et Dhaliwal, 2008), provoque un mouvement de fuite du stimulus après un contact physique avec celui-ci (Deletre et al., 2016). Les répulsifs anti-appétant agissent sur le comportement alimentaire de l'insecte en l'empêchant, l'interrompant ou l'interférant (Schoonhoven et Van, 2002). Bien que de nombreuses études aient été menées sur les propriétés des HE au Burkina Faso, peu d'études ont été menées sur les propriétés répulsives des HE sur les moustiques encore moins sur les activités répulsives-irritantes au Burkina Faso. Thèse de Doctorat unique - 92 - Dans ce chapitre, nous présenterons les principaux résultats des tests répulsifs d'irritation de contact des HE de cinq plantes (Cymbopogon citratus, Cymbopogon nardus, Eucalyptus camaldulensis, Lippia multiflora et Ocimum americanum), du DEET (N, N-diethyl-3-methylbenzamide) et de la perméthrine, deux (2) témoins positifs, réalisé sur des souches de Ae. aegypti collecté à Bobo-Dioulasso et de An. gambiae de la Vallée du Kou. 3- 3-2. Méthodologie : test répulsif-irritant par contact La technique d'imprégnation des papiers Whatman avec HE a été décrite par Balboné et al. (2022b). La procédure de test répulsif irritant de Grieco et al. (2005) mise en place pour Ae. aegypti a été adaptée pour An. gambiae (Figure 26). Pour ce test, trois (3) concentrations ont été testées : 0,1%, 0,5% et 1%. Ces concentrations ont été retenues après des tests préliminaires. Pour chaque concentration et le témoin (acétone pur), 3 lots de 20 moustiques ont été testés. En effet, des lots de 20 moustiques femelles non nourris au sang et âgés de 3 à 7 jours ont été aspirés (Figure 28A) puis introduits dans trois tubes contenant du papier imprégné (Tube rouge) (Figure 27A). Ensuite, chaque tube «imprégné» contenant des moustiques a été fixé sur un tube de contrôle (Tube vert) contenant du papier neutre (Figure 27B). Après 30 secondes d'exposition (Figure 28B), le tiroir séparant le tube imprégné du tube neutre (contenant le papier neutre) a été ouvert (Figure 27C). Les moustiques ont ensuite été laissés libres pendant 10 minutes. Les moustiques passant du tube imprégné au tube neutre étaient considérés comme «échappés». A l'inverse, les moustiques qui restaient dans le tube imprégné étaient considérés comme des moustiques «restés». Après la fermeture du tiroir, on a compté le nombre de moustiques «échappés» et «restés» dans chaque tube (Figure 28C). Parmi les moustiques «restés», certains ont été assommés. Pour le témoin, les moustiques ont été mis dans un tube neutre qui a été fixé sur un autre tube neutre. Le test est validé lorsque la proportion de moustiques «échappés» dans le témoin a été inférieure à 50%. L'effet répulsif-irritant de contact d'un produit a été évalué sur la base de la proportion de moustiques qui s'échappent. Les moustiques assommés ont été enregistrés. Le DEET et la perméthrine ont été utilisés comme témoins positifs. Les proportions du nombre de moustiques «échappés» ont été comparées par paire avec le témoin par le test de Fisher et corrigées avec la méthode de Bonferroni-Holm. Un modèle linéaire généralisé (distribution binomiale) a été étudié pour connaître l'effet du produit, de la concentration et de l'interaction concentration-produit (dose dépendante) sur l'effet irritant sur le moustique. Enfin, une classification ascendante hiérarchique a été effectuée pour classer les extraits de plantes.
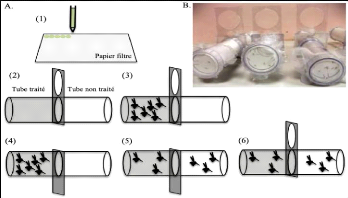
Figure 26: Dispositif de Grieco et al. (2005) adapté à Anopheles gambiae (Deletre et al. 2013) A : Séquence de test ; B : Photo du dispositif (1) : un papier filtre (12*15cm) est traité avec 2mL de solution ; (2) : le papier est placé dans le tube traité ; (3) : les moustiques sont introduits dans le tube traité ; (4) : la porte est ouverte après 30s d'acclimatation ; (5) : les moustiques sont laissés libres pendant 10 min ; (6) : après fermeture de la porte, le nombre de moustiques échappés est calculé. Thèse de Doctorat unique - 93 -
p to osy o t e Rrc pour e Déve oppement ntpe Figure 27: Présentation des tubes OMS (OMS 2022b) (A : Tube rouge, B : Tube vert, C : Tube o rouge fixé au tube vert.) St 2 Mito
Am C Bub Figure 28: Déroulement des tests répulsifs irritants de contact Transfer mosquitoes into the holdng tubes A : Aspiration des moustiques femelles de 3 à 7 jours non gorgés ; B : Les moustiques libres dans Provide them access to a water-sugar le tube contenant les papiers imprégnés après l'ouverture du tiroir ; C : Dénombrement des solution. Read mortality at the end of the 2h moustiques échappés (Cliché Balboné, 2020) Thèse de Doctorat unique - 94 - Thèse de Doctorat unique - 95 - 3.3.3. Résultats L'effet répulsif irritant de contact a été différent selon la souche de moustique et le produit testé. De plus, il a été dose-dépendant. Globalement, chez les populations de An. gambiae, les HE testées ont montré un effet irritant de contact (taux d'échappement supérieur à 30%). Ce qui n'a pas été le cas avec les populations de Ae. aegypti. Les HE des cinq extraits de plantes ainsi que les deux produits chimiques synthétiques, le DEET et la perméthrine, ont montré des effets irritants de contact significatifs à de faibles concentrations pour les HE et à des concentrations élevées pour les produits chimiques (Figures 29 et 32). Dans toutes les populations de moustiques testées, la classification ascendante hiérarchique (CAH) est résumée en 3 classes en fonction de la similarité de la réponse comportementale du moustique (Figures 29-32). 3.3.3.1. Réponses comportementales avec les populations de Anopheles gambiae Sur la souche sensible de An. gambiae «Kisumu», la classe 1 est constituée de l'HE de C. citratus et le DEET présentant des taux d'échappés légèrement supérieurs à 50 % à la concentration la plus faible (0,1 %) et des taux d'échappés, respectivement de 84,63 et 85 % à la concentration de 0,5 % (Figure 29). À la concentration la plus élevée, l'HE de C. citratus a été moins répulsif-irritante (31,16 % de taux d'échappés) par rapport au DEET (56 % de taux d'échappés), tandis que les effets knock-down (KD) ont été élevés chez les moustiques restants (Tableau XVII). La classe 2 a été constituée par les HE de C. nardus et O. americanum qui ont été significativement plus répulsif-irritantes à faible concentration avec une efficacité décroissante avec la concentration et leurs activités diminuaient avec l'augmentation de la concentration (60,05 et 51,88% à la concentration 0,1%, 32,27 et 40,53% à 0,5%, 19,76 et 36,82% à 1%, respectivement pour C. nardus et O. americanum). En même temps, ces deux HE ont produit des effets KD croissants en fonction de la concentration du produit au niveau des moustiques restés (Tableau XVII). La classe 3 a été constituée par les HE de E. camaldulensis, de L. multiflora et la perméthrine qui ont été les produits les moins irritants à la faible concentration (taux d'échappement inférieur à 45 % à la concentration de 0,1 %) mais les activités de l'HE de E. camaldulensis et de la perméthrine ont varié en fonction de la concentration du produit (taux d'échappement compris entre 52,3 et 69,72%, 58,33 et 78,84 %, respectivement aux Thèse de Doctorat unique - 96 - concentrations de 0,5 et 1%). L'HE de L. multiflora a produit des effets KD plus élevés que les 2 autres et cet effet KD augmentait avec la concentration (Tableau XVII). Sur la souche de An. gambiae de terrain VK, la classe 1 a regroupé l'HE de C. citratus et le DEET qui ont été répulsif-irritants dès la faible concentration avec une efficacité qui augmentait avec la concentration. La classe 2 a été constituée par les HE de C. nardus, L. multiflora et O. americanum qui ont présenté une activité décroissante de la faible à la forte concentration. Au même moment, l'HE de C. nardus a produit des effets KD qui augmentaient avec la concentration (taux KD = 29,46% à 0,1%, 32,33% à 0,5% et 37,62% à 1%). D'autre part, l'HE de L. multiflora a produit des effets KD élevés. La classe 3 a été constituée de l'HE de E. camaldulensis et de la perméthrine qui n'ont pas été répulsif-irritantes à faible concentration mais sont devenues plus répulsif-irritantes lorsque la concentration augmentait (Figure 30). Hormis l'HE de L. multiflora, toutes les HE testées ont été plus répulsif-irritantes sur la souche VK que sur la souche Kisumu à la concentration la plus élevée, 1% (Figures 29 et 30). Car sur la souche Kisumu, l'effet KD a été plus perçu. Tableau XVII: Taux de KD obtenus parmi les moustiques qui ne se sont pas échappés
KD: Knock down Thèse de Doctorat unique - 97 -
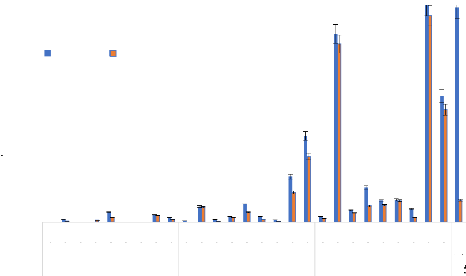
Figure 29: Taux d'échappés des femelles de la souche sensible de Anopheles gambiae «Kisumu» aux 5 HE, au DEET et à la perméthrine et, le dendrogramme représentant les réactions comportementales obtenues par le regroupement hiérarchique de ces insecticides selon leurs effets irritants de contact. (Les intervalles de confiance ont été calculés par la méthode de Wald).
10 8 6 4 2 0 VK Dissimilarity Figure 30: Taux d'échappés des femelles de la souche de terrain de Anopheles gambiae «VK» aux 5 HE, au DEET et à la perméthrine et, le dendrogramme représentant les réactions comportementales obtenues par le regroupement hiérarchique de ces insecticides selon leurs effets irritants de contact. (Les intervalles de confiance ont été calculés par la méthode de Wald). Thèse de Doctorat unique - 98 - Thèse de Doctorat unique - 99 - 3- 3-3.2. Réponses comportementales avec les populations de Aedes aegypti Pour la souche sensible de Ae. aegypti Bora Bora, le groupe de classe 1 a été constituée par les HE de C. citratus, E. camaldulensis, O. americanum et le DEET qui ont donné des taux d'échappés croissant avec la concentration, sauf l'HE de O. americanum et le DEET (Figure 31). Tous les produits de cette classe ont produit des effets KD faibles (Tableau XVII). La classe 2, constituée des HE de C. nardus et de L. multiflora, a regroupé les produits dont les effets irritants ont été décroissants à mesure que la concentration augmentait. Par contre, ils ont produit des effets KD élevés et croissants avec la concentration. Pour la classe 3, la perméthrine n'a présenté aucun effet répulsif-irritant à faible concentration (0,1 % et 0,5 %) mais elle est devenue répulsif-irritante à la concentration (1 %) (Tableau XVII). Pour la souche de Ae. aegypti de Bobo-Dioulasso, la classe 1 a été constituée par les HE C. citratus, E. camaldulensis, O. americanum, le DEET et la perméthrine qui ont présenté des effets répulsif-irritants faibles (taux d'échappé inférieur à 50 %) à la concentration de 1% même si leurs effets répulsif-irritants augmentent avec la concentration. (Figure 32). Ces substances ont produit de très faibles effets KD. La classe 2 a été constituée de l'HE de C. nardus qui a été la plus répulsif-irritante à la plus faible concentration (72,22 % de taux d'échappés à la concentration de 0,1 %), mais cette activité diminuait au fur à mesure que la concentration augmentait. Inversement, cette HE a produit des effets KD élevés lorsque la concentration augmente. La classe 3 a été constituée de l'HE de L. multiflora qui a été plus répulsif-irritante à des concentrations plus élevées (taux d'échappés de 52,79 et 58,79, respectivement aux concentrations de 0,5 et 1%) ; son activité augmentait avec la concentration (Figure 32). De plus, cette HE a produit des effets KD qui augmentaient avec la concentration (Tableau XVII). Aux faibles concentrations, l'HE de C. nardus a produit des effets répulsifs de contact les plus élevés sur toutes les souches de Ae. aegypti (Figure 32). Les effets de cette HE ont été significativement supérieurs à ceux du DEET et de la perméthrine.
Thèse de Doctorat unique - 100 -
Figure 31: Taux d'échappés des femelles de la souche sensible de Aedes aegypti «Bora Bora» aux 5 HE, au DEET et à la perméthrine et, le dendrogramme représentant les réactions comportementales obtenues par le regroupement hiérarchique de ces insecticides selon leurs effets irritants de contact. (Les intervalles de confiance ont été calculés par la méthode de Wald).
12 10 8 6 4 2 Dissimilarity Bobo-Dsso Figure 32: Taux d'échappés des femelles de la souche de terrain de Aedes aegypti «Bobo-Dioulasso» aux 5 HE, au DEET et à la perméthrine et, le dendrogramme représentant les réactions comportementales obtenues par le regroupement hiérarchique de ces insecticides selon leurs effets irritants de contact. (Les intervalles de confiance ont été calculés par la méthode de Wald). Thèse de Doctorat unique - 101 - 3- 3-4. Discussion Le contrôle des maladies à transmission vectorielle passe par une approche qui implique une protection personnelle contre les piqûres de moustiques (Debboun et Strckman, 2012). Cette approche utilise des répulsifs commerciaux qui sont principalement basés sur le DEET (N, N-diethyl-3-methylbenzamide) (Badolo et al., 2004; Calafat et al., 2012). Cependant, le DEET peut avoir des effets secondaires nocifs (Briassoulis, Narlioglou et Hatzis, 2001; Clem, Havemann et Raebel, 1993) et semble être toxique. Il est donc urgent d'évaluer l'effet d'autres répulsifs ou ingrédients irritants, en particulier les HE qui possèdent des propriétés répulsives, irritantes et toxiques sur les moustiques. Ici, les propriétés répulsif-irritantes de contact des HE de Cymbopogon citratus, Cymbopogon nardus, Eucalyptus camaldulensis, Lippia multiflora et Ocimum americanum ont été mesurées, en utilisant le procédé de Grieco et al. (2005), sur des populations de Anopheles et de Aedes, respectivement de Bobo-Dioulasso et de la «Vallée du Kou» où la résistance aux pyréthrinoïdes était répandue. Dans la présente étude, l'exposition des moustiques aux 5 HE a montré des degrés variables d'activité irritante par contact de ces HE. L'effet irritant d'un produit pourrait être dû à son action sur le système nerveux par l'intermédiaire des tarses des insectes (White, 2007). Certains composés individuels des HE sont clairement détectés et évités par les moustiques grâce à leurs antennes. Pourtant, l'influence physiologique des HE conduisant à la répulsion reste largement inconnue (Dekker et al., 2011; Ramirez et al., 2012). En effet, l'HE de L. multiflora s'est avérée moins répulsif-irritante sur les deux souches de An. gambiae et sur la souche sensible de Ae. aegypti «Bora Bora», surtout aux concentrations les plus élevées. Cependant, des études précédentes ont montré les propriétés répulsives par irritation du â-caryophyllène et du p-cymène (Nararak et al., 2019; Ndirangu, Opiyo et Ng'ang'a, 2020; Azeem et al. 2019), 2 composés majeurs de l'HE de L. multiflora qui a été testée dans la présente étude. Cette faible activité irritante de l'HE de L. multiflora est en accord avec son effet «knockdown» élevé (90, 88,24 et 87,88 % KD à une concentration de 1%, respectivement pour Kisumu, VK et Bora Bora) qui a empêché les moustiques de s'échapper. De même, dans d'autres études récentes, cette HE a donné des effets KD élevés sur les mêmes populations de Ae. aegypti et An. gambiae (Balboné et al., 2022b, 2022c). En plus de son effet toxique montré récemment par Balboné et al. (2022a, 2022b, 2022c), l'HE de L. multiflora a exercé un effet irritant sur les Thèse de Doctorat unique - 102 - populations de Ae. aegypti. Cela s'avérerait intéressant dans le cadre de son utilisation dans les stratégies de lutte antivectorielle. Dans la plupart des cas, l'HE de C. citratus dont les activités irritantes sont proches de celles du DEET, a eu un effet irritant significatif à faible concentration sur les 2 souches de An. gambiae, alors que sur la souche sensible de Ae. aegypti Bora Bora, elle a été efficace à forte concentration. Nos résultats corroborent ceux de Deletré et al. (2013) qui ont également montré des effets répulsifs irritants de l'HE de C. citratus sur An. gambiae. Selon ces auteurs, l'effet irritant était dû à la présence de géranial, néral et l'activité de cette HE augmentait avec la concentration. L'effet répulsif-irritant obtenu avec les HE de C. citratus et aussi de C. nardus serait facilité par le grand nombre de moustiques non paralysés, connus pour produire de faibles effets KD sur Ae. aegypti et An. gambiae (Balboné et al., 2022b, 2022c). Selon ces auteurs, ces deux HE testées récemment pour leur toxicité sur les mêmes souches de moustiques ont donné des taux de mortalité inférieurs à 5% et à 50% à la concentration de 1%, respectivement pour l'HE de C. citratus et l'HE de C. nardus confirmant ainsi leur faible toxicité (Balboné et al., 2022b, 2022c). Globalement, l'HE de O. americanum, une autre HE moins toxique (Balboné et al., 2022b, 2022c), présente un effet irritant de contact significatif sur les souches sensibles de Ae. aegypti Bora Bora et de An. gambiae Kisumu. La bio-activité irritante de contact relativement élevée de l'HE de O. americanum pourrait être attribuée à ses principaux composés, le 1,8 cinéole et le camphre. Des études antérieures ont signalé que le 1,8 cinéole et le camphre possédaient des effets répulsifs contre les insectes, respectivement Blattella germanica et Sitophilus zeamais (Cansian et al., 2015 ; Chooluck et al., 2019). De plus, cette HE a eu de faibles effets KD à forte concentration. En outre, certains auteurs ont montré dans leurs travaux que l'HE de O. americanum a produit des effets KD plus élevés à forte concentration sur les souches sensibles, Bora Bora et Kisumu, que sur les souches de terrain de Ae. aegypti et de An. gambiae collectées, respectivement à Bobo-Dioulasso et à VK (Balboné et al., 2022b, 2022c). D'autre part, des études menées en Guinée Bissau, Afrique de l'Ouest, avaient rapporté que l'HE de O. americanum avait également un effet répulsif sur les moustiques du genre Anopheles (Pålsson et Jaenson 1999). Les deux témoins positifs, le DEET et la perméthrine, se sont également révélés irritants. En effet, la perméthrine a montré un effet irritant de contact légèrement élevé sur toutes les souches de An. gambiae (Kisumu et VK) et un faible effet irritant de contact sur toutes les Thèse de Doctorat unique - 103 - souches de Ae. aegypti (Bora Bora et Bobo-Dioulasso) à toutes les concentrations, avec de faibles effets KD sur toutes les souches des deux espèces de moustiques testés, par rapport aux HE de C. nardus, L. multiflora et O. americanum. Nos résultats sont similaires à ceux de Deletre et al. (2013) qui ont montré que la perméthrine avait un faible effet irritant de contact sur les populations résistantes de An. gambiae à des concentrations élevées. On pourrait penser que le faible pouvoir répulsif de la perméthrine serait dû à la résistance accrue des moustiques aux insecticides chimiques. Cependant, des études antérieures ont signalé que la perméthrine peut encore être appliquée comme répulsif contre les piqûres de moustiques dans de nombreux endroits, même si la résistance aux pyréthrinoïdes était répandue (Bowman et al., 2018). Alors, selon la présente étude, il serait intéressant d'utiliser la perméthrine comme un répulsif irritant au moins à la concentration (0,5%). Quant au DEET, il a été répulsif-irritant pour toutes les souches de An. gambiae et a provoqué moins d'effet KD. Ces résultats obtenus sont en accord avec ceux obtenus par Deletre et al. (2013) qui ont montré que le DEET est irritant à des concentrations élevées sur toutes les populations de An. gambiae testées. Cependant, sur la souche de Ae. aegypti de Bobo-Dioulasso, le DEET a été moins irritant que toutes les HE testées et n'a pas provoqué de KD. Ces résultats ne sont pas en accord avec ceux trouvés par Licciardi et al. (2006) qui ont montré que le DEET donne un effet répulsif-irritant significatif et provoque des KD à des concentrations élevées. Cela pourrait s'expliquer par le fait que nos concentrations étaient plus faibles que celles utilisées par Licciardi et al. (2006). De plus, nos résultats montrent que le DEET était plus irritant sur les souches de An. gambiae que sur les souches de Ae. aegypti testées alors que des études précédentes avaient montré des données contraires (Travis 1947; Rutledge et al., 1983; Curtis et al., 1987; Badolo et al., 2004). Cette contradiction pourrait être due aux différents protocoles expérimentaux utilisés. 3- 3-5. Conclusion partielle Toutes les huiles essentielles testées dans cette étude ont montré des effets répulsifs-irritants. Sur An. gambiae, les effets répulsifs irritants de contact des HE de Cymbopogon citratus, Cymbopogon nardus et Eucalyptus camaldulensis ont été très proches de ceux du DEET, le répulsif de référence. Sur Ae. aegypti, toutes nos HE ont des effets répulsifs-irritants supérieurs pour certaines et très proches du DEET pour les autres. Les HE testées dans la présente étude pourraient être des répulsifs potentiels contre les moustiques en tant qu'alternatives aux répulsifs synthétiques, les pyréthroïdes et le DEET. Thèse de Doctorat unique - 104 - Thèse de Doctorat unique - 105 - DISCUSSION GENERALE, CONCLUSION GENERALE ET PERSPECTIVES Thèse de Doctorat unique - 106 - Cette étude s'inscrit dans le cadre général de la recherche d'alternatives de lutte contre les moustiques résistants aux pyréthrinoïdes. En effet, les pyréthrinoïdes, insecticides de synthèse, constituent jusqu'à présent les principaux insecticides dans la lutte contre les maladies qui peuvent être transmises par piqûres de moustiques. Cela, à cause de leurs propriétés qui permettent un contrôle efficace de la transmission de ces maladies lorsqu'ils sont utilisés en pulvérisations spatiales ou sur des moustiquaires (Zaim, Aitio et Nakashima, 2000). La recherche d'alternative aux pyréthrinoïdes est un axe de recherche majeur depuis quelques années car l'usage massif des pyréthrinoïdes à des fins de santé publique et surtout en agriculture a entraîné la sélection de moustiques résistants, ce qui compromet l'efficacité des campagnes de LAV (Cohuet, 2020). L'efficacité d'un insecticide repose sur sa capacité à prévenir la piqûre de l'utilisateur ou à tuer les moustiques vecteurs. Ainsi, les nouveaux insecticides à base des HE qui sont utilisées comme alternative doivent avoir des propriétés larvicides ou adulticides soit répulsives pour assurer une bonne gestion de la résistance pyréthrinoïdes. Nos objectifs opérationnels nous ont amené à conduire des études au laboratoire avec des HE sur des moustiques An. gambiae et Ae. aegypti. Dans cette partie nous rappellerons et discuterons les principaux résultats obtenus avant de développer les perspectives ouvertes par ce travail. 1. Discussion générale 1.1. Les huiles essentielles comme des larvicides A travers cette étude, nous avons mis en évidence les propriétés larvicides des huiles essentielles de cinq (5) plantes locales qui sont C. citratus, C. nardus, E. camaldulensis, L. multiflora et O. americanum. Parmi ces 5 HE testées, l'HE de L. multiflora a été la plus efficace sur les larves des souches de Aedes aegypti et de Anopheles gambiae sensibles et résistantes aux insecticides de synthèse. En effet, cette HE n'a pas présenté de différence significative avec le pyriproxyfène, un larvicide de référence (p = 0,86). Récemment, Yameogo et al. (2021) et Wangrawa et al. (2022a) ont montré l'effet toxique de l'HE de L. multiflora sur des populations de An. gambiae, Ae. aegypti, An. funestus et Culex quinquefasciatus. Aussi, excepté l'HE de E. camaldulensis, toutes les autres HE ont montré une toxicité considérable. Cette toxicité de ces HE a été rapportée par des études antérieures (Dayananda et Ranaweera, 1996 ; Yaméogo et al., 2021 ; Ntonga et al., 2014). Thèse de Doctorat unique - 107 - Les HE ont été plus toxiques sur les larves de An. gambiae que sur celles de Ae. aegypti. Cependant, il y a plus de 19 ans, Bassolé et al. (2003) ont rapporté que les larves de Ae. aegypti étaient plus sensibles que celles de An. gambiae s.l. aux HE de L. multiflora et O. americanum. Ce changement de tendance de la toxicité vis-à-vis des 2 populations de moustiques testées dans notre étude peut être due à la différence de localité de collecte des larves de moustiques et aux facteurs favorisant l'apparition de la résistance surtout au sein des populations de Ae. Aegypti, vu le nombre d'années passées entre ces deux (2) études. En effet, Namountougou et al. (2020) ont rapporté que Ae. aegypti de Bobo-Dioulasso était plus résistant que celui de Ouagadougou aux insecticides chimiques (Deltamethrine, bendiocard, chlorpyrifos-methyl). L'HE de L. multiflora et dans une moindre les quatre autres HE utilisées dans cette étude peuvent constituer en cela de bons outils de gestion de la résistance des moustiques des moustiques aux insecticides de synthèse. 1.2. Les cinq (5) HE et les combinaisons de deux (2) d'entre elles comme adulticides Dans cette étude, la toxicité des 5 HE (HE de C. citratus, C. nardus, E. camaldulensis, L. multiflora et O. americanum) et les combinaisons de C. nardus (C.n) et O. americanum (O.a) ont été évaluées sur les adultes de An. gambiae et de Ae. aegypti. Dans l'ensemble, presque toutes les HE et les combinaisons testées ont montré des effets adulticides plus ou moins intéressantes sur les souches de An. gambiae, sensible (Kisumu) et de terrain (VK) résistante aux pyréthrinoïdes. Ces effets adulticides, mis en évidence par des effets knock-down (KD), des concentrations létales faibles et des taux de mortalité élevés des moustiques adultes testés, ont été par contre plus faibles sur Ae. aegypti. Cela montre que les populations de An. gambiae sont plus sensibles aux HE testées que les populations de Ae. aegypti. Des études menées par Amerasan et al. (2012) et Murugan et Kumar (2012) ont trouvé des résultats similaires avec les HE de Citrus sinensis et de Cassia tora en Taïwan et en Inde où Anopheles sp est plus sensibles que Ae. aegypti. L'HE de L. multiflora a été la plus efficace sur les adultes de An. gambiae et de Ae. aegypti testées, suivie de l'HE de C. nardus. L'HE la moins toxique était celle de E. camaldulensis. Il est intéressant de noter que L. multiflora peut constituer une bonne alternative au regard du taux de mortalité qui atteint 96,88% sur An. gambiae, largement supérieure au taux de mortalité obtenu avec la perméthrine. Thèse de Doctorat unique - 108 - L'efficacité des 5 HE testées dans cette étude serait due à leurs composés majeurs. En effet, plusieurs études antérieures ont montré que les composés majeurs sont le facteur responsable de la bio-activité d'une HE, car ils constitueraient la partie la plus importante des composés totaux de l'HE (Lucia et al., 2007; Gbolade et Lockwood, 2008; Vani, Cheng et Chuah, 2009). En effet, des études ont montré que le p-cymène et le f3-caryophyllène présentent des propriétés adulticides sur An. gambiae et Ae. aegypti (Almadiy, 2020; Bossou et al., 2013). Ainsi, l'activité adulticide élevée de l'HE de L. multiflora serait attribuée au f3-caryophyllène et au p-cymène qui font partie de ses composés majeurs dans la présente étude. Globalement, toutes les combinaisons obtenues avec les 2 HE (C. nardus et O. americanum) ont amélioré l'efficacité globale des HE par rapport aux HE individuelles. Dans la présente étude, l'amélioration du potentiel adulticide par la combinaison des deux HE peut être due par leur proportion dans la combinaison. Nos résultats corroborent ceux des travaux antérieurs qui ont rapporté que l'activité biologique des HE dépend non seulement de leur composition qualitative, mais aussi du rapport quantitatif de leurs composants (Bekele et Hassanali, 2001 ; Pavela, 2008). L'amélioration du potentiel adulticide par les combinaisons des HE a été prouvée par les effets synergiques et additifs obtenus. Cette amélioration de la toxicité par les combinaisons des 2 HE serait due à l'effet toxique combiné des composés majeurs des 2 HE prises individuellement. Des travaux antérieurs ont montré que l'action toxique des HE est due aux effets combinés de différents composants, avec ou sans action toxique individuelle significative contre les insectes (Tanprasit, 2005; Park et al., 2011). D'autres études ont montré que des combinaisons de 1,8-cinéole et de citronellal ainsi que de plusieurs composés mineurs et de citronellal ont donné des effets synergiques sur les larves de Spodoptera littoralis (Abbassy, Abdelgaleil et Rabie, 2009). Recement, Wangrawa et al. (2022b) ont montré que l'amélioration des HE par les combinaisons est due à l'apparition de nouveaux composés préalablement absents dans les HE individuelles. La mise en évidence de l'effet adulticide des HE seules et des combinaisons de certaines de ces HE est une étape importante dans la recherche de nouveaux types d'insecticides. En effet, nos études ont montré qu'il y avait un gain d'efficacité à combiner les HE, en termes de mortalité induite, d'effet Knock Down (KD) et de concentration létale induite. Il faut noter que, pour la première fois, une interaction synergique a été observée en combinant les HE de C. Thèse de Doctorat unique - 109 - nardus et de O. americanum sur des populations de Ae. aegypti et de An. gambiae au Burkina Faso. Le système nerveux est généralement le principal site cible des insecticides utilisés dans les programmes de lutte contre les vecteurs du paludisme (Rants'o et al. 2022). Ces auteurs ont montré l'existence d'un lien entre les modes d'action de certains insecticides chimiques et les HE. En effet, selon ces auteurs, les HE agissent au niveau du système nerveux central sur l'acétylcholinestérase (AChE) comme les organophosphorés et les carbamates. Kiendrebeogo et al. (2011) ont montré qu'en utilisant des conditions expérimentales et des concentrations d'enzyme AChE similaires, les HE de Ocimum canum (Sims.) du Burkina Faso ont inhibé l'AChE. De plus, Selon d'autres auteurs, les HE et leurs constituants inhibent les enzymes de détoxification P450 monooxygénase et glutathione-S-transférase impliquées dans la résistance à de multiples insecticides dont les pyréthroïdes, les organophosphorés et les carbamates (Norris et al., 2018 ; Gaire et al., 2021). 1.3. Les huiles essentielles comme répulsifs anti moustiques Toutes les 5 HE testées ont montré des effets répulsif-irritants de contact. Cependant, l'HE de L. multiflora s'est montrée moins répulsive sur les deux souches de An. gambiae et la souche sensible de Ae. aegypti « Bora Bora ». Cette faiblesse de l'effet répulsif-irritant de l'HE de L. multiflora a été plus remarquable surtout aux concentrations les plus élevées. Cela serait dû au taux élevé de KD que produit cette HE sur les différents types de moustiques testés. Nos études antérieures ont montré que l'HE de L. multiflora produit des effets KD élevés sur les populations de Ae. aegypti et An. gambiae (Balboné et al. 2022b; Balboné et al. 2022c). Il est évident que les moustiques assommés ne peuvent pas s'échapper. L'HE de L. multiflora peut être un bon répulsif à faible dose. Compte tenu du fait que les HE de C. citratus, C. nardus et de E. camaldulensis ont montré des effets répulsif-irritants de contact très proches ou dans certains cas supérieurs à ceux causés par le DEET, ces HE peuvent être de bons répulsif-irritants de contact. En effet, des auteurs ont montré que les HE de C. nardus et de Eucalyptus spp qui ont été citées dans de nombreuses inventions brevetées ont un pouvoir répulsif contre les espèces de Anopheles et de Aedes, dans certains cas comparables à celui du DEET (Tawatsin et al., 2001; Moore, Lenglet et Thèse de Doctorat unique - 110 - Hill, 2002 ; Sultana et al., 2008). En outre, d'autres auteurs ont montré que l'HE de C. citratus a une propriété répulsive assez significatives (Amer et Mehlhorn, 2006 ). L'HE de O. americanum a produit des effets répulsifs irritants de contact élevés ou modérés sur les souches de terrain de Ae. aegypti et de An. gambiae (respectivement de Bobo-Dioulasso et VK) à toutes les concentrations, mais de manière croissante. De plus, cette HE a produit de faibles effets KD même à la concentration élevée. Ces résultats corroborent ceux des études antérieures menées en Guinée Bissau, en Afrique de l'Ouest, qui ont rapporté que l'HE de O. americanum a un effet répulsif sur les moustiques Anophèles (Palsson et Jaenson, 1999). Les différences dans la spécificité d'espèce des profils de répulsion de ces HE peuvent motiver leur utilisation en combinaison afin de générer des formulations à large spectre pour la répulsion simultanée des deux espèces de moustiques. Il faut noter que, pour la première fois, l'effet répulsif irritant par contact des HE a été testé sur des populations de moustiques des espèces de Ae. aegypti et de An. gambiae au Burkina Faso. 2. Conclusion générale et perspectives L'objectif général de cette thèse était d'évaluer les propriétés insecticides de 5 huiles essentielles et des combinaisons de deux (2) d'entre elles sur des populations de Ae. aegypti et de An. gambiae reconnues résistantes aux insecticides chimiques. Dans cette thèse, nous avons évalué trois propriétés insecticides, à savoir les propriétés larvicides, adulticides et répulsives irritantes de contact. Concernant les propriétés larvicides, les travaux ont permis de montrer que l'HE de L. multiflora serait un bon larvicide dans la mesure où cette HE a donné une toxicité élevée sur les larves de Ae. aegypti et de An. gambiae, sensiblement proche de celle produite du pyriproxyfène, un larvicide de référence. Cependant, les HE de C. nardus et de O. americanum ne seraient pas négligeables comme des larvicides. En termes d'efficacité adulticide, l'HE de L. multiflora serait un candidat privilégié, puisqu'elle a donné des taux de mortalités et de knock-down très élevés. Aussi, les combinaisons aux rations 90 : 10, 80 : 20 et 20 : 80, respectivement des HE de C. nardus et de O. americanum ont produit des effets adulticides élevés. Toutes les 5 HE testées ont produit des effets répulsifs irritants de contact modérés pour certaines et élevés pour d'autres. Les HE de C. nardus, C. citratus et E. camaldulensis peuvent être de bons répulsifs. Compte tenu de ces différentes propriétés utiles de ces HE dans la lutte antivectorielles, il serait intéressant de vulgariser leur utilisation. Cette étude nous a donné un autre goût. Donc, en perspective, nous envisageons : - Entreprendre des tests larvicides, répulsifs et adulticides avec les huiles essentielles en milieu semi contrôlé pour la phase II de l'évaluation ; - Tester les combinaisons intéressantes des HE sur une large gamme de populations naturelles de moustiques ; - Tester les combinaisons d'HE avec des insecticides de la famille des pyréthrinoïdes sur une large gamme de populations naturelles de moustiques ; - Déterminer les compositions chimiques des combinaisons ayant donne des effets synergistes ; - Mettre au point des formulations insecticides à base des HE de plantes locales microencapsulés pour améliorer l'efficacité dans le temps en collaboration avec les firmes locales. Thèse de Doctorat unique - 111 - Thèse de Doctorat unique - 112 - REFERENCES BIBLIOGRAPHIQUES Abbassy, M.A., Abdelgaleil, S.A., & Rabie, R. (2009). Insecticidal and synergistic effects of Majorana hortensis essential oil and some of its major constituents. Entomologia Experimentalis et Applicata, 131 (3): 225-32. https://doi.org/10.1111/j.1570-7458.2009.00854.x. Abbott, W S. (1987). A Method of Computing the Effectiveness of an Insecticide. 1925. Journal of the American Mosquito Control Association 3 (2): 302-3. Achee, N. L., Sardelis M. R., Dusfour I., Chauhan K. R. & Grieco J. P. (2009). Characterization of spatial repellent, contact irritant, and toxicant chemical actions of standard vector control compounds. Journal of the American Mosquito Control Association, 25(2), 156-167. Adams, R. P. (2007). Identification of essential oil components by gas chromatography/mass spectrometry (Vol. 456, pp. 544-545). Carol Stream: Allured publishing corporation. Agence Nationale de Sécurité Sanitaire de l'alimentation, de l'Environnement et du travail (ANSES), 2020. Rôle des arthropodes dans la transmission de la peste porcine africaine. Avis de l'Anses, Rapport d'expertise collective. Saisine n° « 2019-SA-0028 ». France. Agence Nationale de Sécurité Sanitaire de l'alimentation, de l'Environnement et du travail (ANSES), (2013). Avis relatif à la recherche d'insecticides potentiellement utilisables en lutte antivectorielle (Classement des 32 substances actives sélectionnées par l'analyse multicritère SIRIS en trois classes selon le niveau de connaissances sur leur efficacité contre les moustiques). Saisine n° 2012-SA-0028 (Maisons-Alfort, Anses). Ahouansou, A. C., Fagla, S. R. M., Tokoudagba, J. M., Toukourou, H., Badou, Y. K., & Gbaguidi, F. A. (2019). Chemical composition and larvicidal activity of the essential oil of Cymbopogon nardus (L.) Rendle on Anopheles gambiae. International Journal of Biological and Chemical Sciences, 13(3), 1861-1869. https://doi.org/10.4314/ijbcs.v13i3.53. Akrout, A. (2004). Étude des huiles essentielles de quelques plan tes pastorales de la région de Matmata (Tunisie). In : Ferchichi A. (comp.), Ferchichi A. (collab.). Réhabilitation des pâturages et des parcours en milieux méditerranéens. Zaragoza : CIHEAM, 2004. p. 289- ; n. 62). 292 (Cahiers Options Méditerranéennes http://om.ciheam.org/om/pdf/c62/04600172.pdf Thèse de Doctorat unique - 113 - Aliota, M. T., Peinado, S. A., Velez, I. D., & Osorio, J. E. (2016). The WMel Strain of Wolbachia Reduces Transmission of Zika Virus by Aedes aegypti. Scientific Reports 6 (June): 1-7. https://doi.org/10.1038/srep28792. Aldridge, W. N. (1950) `Some properties of specific cholinesterase with particular reference to the mechanism of inhibition by Diethyl p-Nitrophenyl Thiophosphate (E 605) and Analogues', Biochemical Journal, 46(4), pp. 451-460. doi: 10.1042/bj0460451. Almadiy, A. A. (2020). Chemical Composition, Insecticidal and Biochemical Effects of Two Plant Oils and Their Major Fractions against Aedes aegypti, the Common Vector of Dengue Fever.» Heliyon 6 (9): e04915. https://doi.org/10.1016/j.heliyon.2020.e04915. Alphey, L. (2014). Genetic Control of Mosquitoes. Annual Review of Entomology 59: 205-26. https://doi.org/10.1146/annurev-ento-011613-162002. Amer, A., & Mehlhorn, H. (2006). Repellency effect of forty-one essential oils against Aedes, Anopheles, and Culex mosquitoes. Parasitol Res; 99: 478-490 Amerasan, D., Murugan, K., Kovendan, K., Mahesh Kumar, P., Panneerselvam, C., Subramaniam, J., ... & Hwang, J. S. (2012). Adulticidal and repellent properties of Cassia tora Linn. (Family: Caesalpinaceae) against Culex quinquefasciatus, Aedes aegypti, and Anopheles stephensi. Parasitology research,
111(5), 1953-1964. Azeem, M., Zaman, T., Tahir, M., Haris, A., Iqbal, Z., Binyameen, M., ... Mozûraitis, R. (2019). Chemical composition and repellent activity of native plants essential oils against dengue mosquito, Aedes aegypti. Industrial Crops and Products, 140, 111609. https://doi.org/10.1016/j.indcrop.2019.1116. https://doi.org/10.1016/j.indcrop.2019.111609. Badolo, A., Ilboudo-Sanogo, E., Ouédraogo, A. P., & Costantini, C. (2004). Evaluation of the sensitivity of Aedes aegypti and Anopheles gambiae complex mosquitoes to two insect repellents: DEET and KBR 3023. Tropical Medicine & International Health, 9(3), 330-334. https://doi.org/10.1111/J.1365-3156.2004.01206.X. Badolo, A., Sombié, A., Pignatelli, P. M., Sanon, A., Yaméogo, F., Wangrawa, D. W., Sanon, A., Kanuka, H., McCall, P. J., & Weetman, D. (2019). Insecticide resistance levels and mechanisms in Aedes aegypti populations in and around Ouagadougou, Burkina Faso. PLoS Thèse de Doctorat unique - 114 - neglected tropical diseases, 13(5), e0007439. https://doi.org/10.1371/journal.pntd.0007439. Balboné, M., Soma, D. D., Namountougou, M., Drabo, S. F., Konaté, H., Toé O., Bayili, K., Meda, G. B., Dabiré, R. K. and Gnankine, O. (2022a). Essential oils from five local plants: an alternative larvicide for Anopheles gambiae s.l. (Diptera: Culicidae) and Aedes aegypti (Diptera: Culicidae) Control in Western Burkina Faso. Front. Trop. Dis. 3:853405. doi: 10.3389/fitd.2022.853405. Balboné, M., Diloma Soma, D., Fogné Drabo, S., Namountougou, M., Konaté, H., Benson Meda, G., Sawadogo, I., Romba, R., Bilgo, E., Nebié, R. C. H., Bassolé, I. H. N., Dabire, R. K., & Gnankine, O. (2022). Alternatives to Pyrethroid Resistance : Combinations of Cymbopogon nardus and Ocimum americanum Essential Oils Improve the Bioefficiency Control Against the Adults' Populations of Aedes aegypti (Diptera: Culicidae). Journal of medical entomology, 59(6), 2102-2109. https://doi.org/ 10.1093/jme/tjac148. Balboné, M., Sawadogo, I., Soma, D. D., Drabo, S. F., Namountougou, M., Bayili, K., Romba, R., Meda, G. B., Nebié, H. C. R., Dabire, R. K., Bassolé, I. H. N., & Gnankine, O. (2022). Essential oils of plants and their combinations as an alternative adulticides against Anopheles gambiae (Diptera: Culicidae) populations. Scientific reports, 12(1), 19077. https://doi.org/ 10.1038/s41598-022-23554-6. Banks, S. D., Murray, N., Wilder-Smith, A., & Logan, J. G. (2014). Insecticide-treated clothes for the control of vector-borne diseases: a review on effectiveness and safety. Medical and veterinary entomology, 28 Suppl 1, 14-25. https://doi.org/10.1111/mve.12068 Barreaux, A. M. G., Oumbouke W. A., Tia I. Z., Brou N'G., Koffi A. A., N'Guessan R. and Thomas M. B. (2019) `Semi-field evaluation of the cumulative effects of a lethal House Lure on malaria mosquito mortality', Malaria Journal, 18(1), pp. 1-9. https://doi.org/ 10.1186/s12936-019-2936-2 . Bassolé, I.H.N., Ouattara, A.S., Nebieb, R. C. H., Ouattara, C. A. T., Kabore, Z. I., Traore, S.A. (2003). Ovicidal and Larvicidal Activity against Aedes aegypti and Anopheles gambiae complex mosquitoes of essential oils extracted from three spontaneous plants of Burkina Faso. Parasitologia 45 :23-26. Parasitologia 45 (1): 23-26. Bassole, I. H. N., Nebie, R., Savadogo, A., Ouattara, C. T., Barro, N., & Traore, S. A. (2005). Thèse de Doctorat unique - 115 - Composition and antimicrobial activities of the leaf and flower essential oils of Lippia chevalieri and Ocimum canum from Burkina Faso. African Journal of Biotechnology 4 (10): 1156-60. https://doi.org/10.5897/AJB2005.000-3230. Bassolé, I.H.N. and Juliani, H.R. (2012). Essential Oils in Combination and Their Antimicrobial Properties. Molecules, 17 :3989-4006. https://doi.org/10.3390/molecules17043989. Bayili, K., Namountougou M., Sanou R., Ndo S., Ouattara A., Dabiré R. K., Malone D. and Diabaté A. (2015). `The sterilizing effect of pyriproxyfen on the malaria vector Anopheles gambiae: physiological impact on ovaries development', Malaria Journal, 14(101), pp. 1-8. https://doi: 10.1186/s12936-015-0609-3. Bekele, J., and Hassanali, A.. (2001). Blend Effects in the toxicity of the essential oil constituents of Ocimum kilimandscharicum and Ocimum kenyense (Labiateae) on two postharvest insect pests. Phytochemistry 57 (3): 385-91. https://doi.org/10.1016/S0031-9422(01)00067-X. Bendali, F., Djebbar, F. et Soltani, N. (2001). Efficacité comparée de quelques espèces de poissons à l'égard de divers stades de Culex pipiens L. dans des conditions de laboratoire résumé. Parasitica 57: 255-65. Benelli, G., Jeffries, C. L., & Walker, T. (2016). Biological Control of Mosquito Vectors: Past, Present, and Future. Insects, 7(4), 52. https://doi.org/10.3390/insects7040052 Benz, G. W., Roncalli R. A. and Gross S. J. (1989) `Use of ivermectin in Cattle, Sheep, Goats, and Swine', in Ivermectin and Abamectin. New York, NY: Springer New York, pp. 215229. https://doi: 10.1007/978-1-4612-3626-9_15. Berenbaum, M. A. Y., and Neal J. J. (1985). Synergism between myristicin and xanthotoxin, a naturally cooccurring plant toxicant. Journal of Chemical Ecology 11 (10): 1349-58. Bernier, U. R., Kline, D., Posey, K. H., Ernier, U. B., Kline, D. L. and Posey, H. (2006). Human emanations and related natural compounds that inhibit human emanations and related natural compounds that inhibit mosquito host-finding abilities mosquito host-finding abilities human emanations and related natural compounds that inhibit mosquito ho. Insect repellent: principales, methods and uses. p 77-100.USA Thèse de Doctorat unique - 116 - daarsfacpub/948. Bharati, M., & Saha, D. (2018). Multiple insecticide resistance mechanisms in primary dengue vector, Aedes aegypti (Linn.) from dengue endemic districts of sub-Himalayan West Bengal, India. PloS one, 13(9), e0203207. https://doi.org/10.1371/journal.pone.0203207 Bhatt, S., Gething, P. W., Brady, O. J., Messina, J. P., Farlow, A. W., Moyes, C. L., Drake, J. M., Brownstein, J. S., Hoen, A. G., Sankoh, O., Myers, M. F., George, D. B., Jaenisch, T., Wint, G. R., Simmons, C. P., Scott, T. W., Farrar, J. J., & Hay, S. I. (2013). The global distribution and burden of dengue. Nature, 496(7446), 504-507. https://doi.org/10.1038/nature12060. Bilgo, E., Lovett, B., Fang, W., Bende, N., King, G. F., Diabate, A., & St Leger, R. J. (2017). Improved efficacy of an arthropod toxin expressing fungus against insecticide-resistant malaria-vector mosquitoes. Scientific reports, 7(1), 3433. https://doi.org/10.1038/s41598-017-03399-0. Bøgh, C., Pedersen, E. M., Mukoko, D. A., & Ouma, J. H. (1998). Permethrin-impregnated bednet effects on resting and feeding behaviour of lymphatic filariasis vector mosquitoes in Kenya. Medical and veterinary entomology, 12(1), 52-59. https://doi.org/10.1046/j.1365-2915.1998.00091.x. Bossou, A. D., Mangelinckx, S., Yedomonhan, H., Boko, P. M., Akogbeto, M. C., De Kimpe, N., Avlessi, F., and Sohounhloue, D. C. K. (2013). Chemical composition and insecticidal activity of plant essential oils from benin against Anopheles gambiae ( Giles ). Boulanger, N., & de Gentile, L. (2012). Les répulsifs cutanés. In Duvallet G., de Gentile L. Ed., Protection personnelle antivectorielle, IRD Éditions, Marseille : 50-116. Boulanger, N., & Lipsker, D. (2015). Protection contre les piqûres de tiques [Protection against tick bites]. Annales de dermatologie et de venereologie, 142(4), 245-251. https://doi.org/ 10.1016/j.annder.2014.11.018. Boussès, P. and Granouillac, B. (2011). Les Anopheles de La région Afro-Tropicale, IRD éditions. ISBN 2-7099-1380-1. http://vmpinsarim-dev.mpl.ird.fr/xper2/web/index.html. Bouyer, J., Gentile L. and Chandre F. (2017) `Chapitre 5- La lutte antivectorielle', in Entomologie médicale et vétérinaire IRD éditions. Marseille, pp. 89-120. http://doi: Thèse de Doctorat unique - 117 - 10.4000/books.irdeditions.22031. Bowman, N. M., Akialis, K., Cave, G., Barrera, R., Apperson, S. and Meshnick, S. R. (2018). Pyrethroid insecticides maintain repellent effect on knock-down resistant populations of Aedes aegypti mosquitoes.» PLoS ONE 5 (13): 1-14. Boyce, R., Lenhart A., Kroeger A., Velayudhan R., Roberts B. and Horstick O. (2013). Bacillus Thuringiensis israelensis (Bti) for the control of dengue vectors: systematic literature review.» Tropical Medicine and International Health 18 (5): 564-77. https://doi.org/10.1111/tmi.12087. Boyer, S., Toty C., Jacquet M., Lempérière G. and Fontenille D. (2012). Evidence of multiple inseminations in the field in Aedes albopictus. PLoS ONE 7 (8). https://doi.org/10.1371/journal.pone.0042040. Brady, O. J., Gething, P. W., Bhatt, S., Messina, J. P., Brownstein, J. S., Hoen, A. G., Moyes, C. L., Farlow, A. W., Scott, T. W., & Hay, S. I. (2012). Refining the global spatial limits of dengue virus transmission by evidence-based consensus. PLoS neglected tropical diseases, 6(8), e1760. https://doi.org/10.1371. Briassoulis, G., Narlioglou M. and Hatzis T. (2001). Toxic encephalopathy associated with use of deet insect repellents: a case analysis of its toxicity in children. Human & Experimental Toxicology, 20 (October 2000): 8-14. Bruneton, J. (1993). Pharmacognosie: phytochimie plantes médicinales (No. 581.634 B7). Burt, S. (2004). Essential oils: their antibacterial properties and potential applications in foods - a review. International Journal of Food Microbiology 94 (3): 223-53. https://doi.org/10.1016/j.ijfoodmicro.2004.03.022. Calafat, A. M. (2012). The U.S. National Health and Nutrition Examination Survey and human exposure to environmental chemicals. International journal of hygiene and environmental health, 215(2), 99-101. https://doi.org/10.1016/j.ijheh.2011.08.014. Caly, L., Druce, J. D., Catton, M. G., Jans, D. A., & Wagstaff, K. M. (2020). The FDA- approved drug ivermectin inhibits the replication of SARS-CoV-2 in vitro. Antiviral research, 178, 104787. https://doi.org/10.1016/j.antiviral.2020.104787 Thèse de Doctorat unique - 118 - Campbell, W. C., Fisher M. H., Stapley E. O., Albers-Schönberg G. and Jacob T. A. (1983) `Ivermectin: A potent new antiparasitic agent', Science, 221(4613), pp. 823-828. doi: 10.1126/science.6308762. Cansian, R. L., Astolfi V., Cardoso R. I., Paroul N., Roman S. S., MIelniczki-Pereira, A. A., Pauletti G. F. and Mossi A. J. (2015). Insecticidal and repellent activity of the essential oil of Cinnamomum camphora Var . Linaloolifera Y . Fujita ( Ho-Sho ) and Cinnamomum camphora ( L .) J Presl . Var . Hosyo ( Hon-Sho ) on Sitophilus zeamais Mots . ( Coleoptera , Curculionedae ).» Revue Bras. Pl. Med. 17: 769-73. Carnevale, P, and Robert, V. (2009). Les Anopheles: biologie, transmission du plasmodium et lutte antivectorielle, Marseille: IRD Editions, 391 p. (Didactiques). ISBN 978-2-7099-16622. ISSN 1142-2580. Casida, J. E. and Quistad G. B. (1998) `Golden Age of Insecticide Research: Past, Present, or Future?' Annual Review of Entomology, 43(1), pp. 1-16. doi: 10.1146/annurev.ento.43.1.1. Cavalcanti, E. S. B., Morais, S. M. de, Lima, M. A. A., & Santana, E. W. P. (2004). Larvicidal Activity of essential oils from Brazilian plants against Aedes aegypti L. Memórias Do Instituto Oswaldo Cruz, 99(5), 541-544. doi:10.1590/s0074-0276200400050. Chaccour, C., Lines, J. and Whitty, C. J. M. (2010) `Effect of Ivermectin on Anopheles gambiae Mosquitoes Fed on Humans: The potential of oral insecticides in malaria control', The Journal of Infectious Diseases, 202(1), pp. 113-116. doi: 10.1086/653208. Chaccour, C. J., Kobylinski, K. C., Bassat, Q., Bousema, T., Drakeley, C., Alonso, P. and Foy, B. D. (2013) `Ivermectin to reduce malaria transmission: a research agenda for a promising new tool for elimination', Malaria Journal, 12(153): 1-8. doi: 10.1186/1475-287512-153. Chandre, F., Darrier, F., Manga, L., Akogbeto, M., Faye, O., Mouchet, J., & Guillet, P. (1999). Status of pyrethroid resistance in Anopheles gambiae sensu lato. Bulletin of the World Health Organization, 77(3): 230-234. Chansang, A., Champakaew D., Junkum A., Jitpakdi A., Amornlerdpison D., Aldred A. K., Riyong D. and Pitasawat B. (2018). Synergy in the adulticidal efficacy of essential oils for the improvement of permethrin toxicity against Aedes aegypti L. (Diptera: Culicidae).» Parasites & Vectors 11 (1): 417. https://doi.org/10.1186/s13071-018-3001-7. Thèse de Doctorat unique - 119 - Chapman, R F. (1969). The Insects, Structure and Function 1th Edition, American Elsevier Pub. Co. 819 pages. ISBN-10- 0444197583 ISBN-13- 978-0444197580. Chippaux, J-P., Boussinesq M. and Prod'hon J. (1995) `L'ivermectine dans le contrôle', Cahiers Santé, 5: 149-158. Chiasson, H., Bélanger A., Bostanian N., Vincent C., & Poliquin A. (2001). Acaricidal properties of Artemisia absinthium and Tanacetum vulgare (Asteraceae) essential oils obtained by three methods of extraction. Journal of economic entomology, 94(1), 167-171. https://doi.org/10.1603/0022-0493-94.1.167. Chooluck, K., Teeranachaideekul V., Jintapattanakit A., Lomarat P., & Phechkrajang C. (2019). Repellency effects of essential oils of Cymbopogon winterianus, Eucalyptus globulus, Citrus hystrix and their major constituents against Adult German cockroach (Blattella germanica Linnaeus (Blattaria: Blattellidae). Jordan Journal of Biological Sciences Short Communication, 12(4), 519-523. Churcher, T. S., Lissenden N., Griffin J. T., Worrall E. and Ranson H. (2016). The impact of pyrethroid resistance on the efficacy and effectiveness of bednets for malaria control in Africa. ELife 5 (AUGUST). https://doi.org/10.7554/eLife.16090. Clem, J. R, Havemann D. F, and Raebel M. A. (1993). Insect repellent (N,N-Diethyl-m-Toluamide) cardiovascular toxicity in an adult. Annals of Pharmacotherapy 27 (March): 289-93. Clevenger, J. F. (1928) Apparatus for Volatile Oil Determination, Description of New Type. The Journal of the American Pharmaceutical Association (1912), 17, 345-349. https://doi.org/10.1002/jps.3080170407. Centre National d'Expertise sur les Vecteurs (CNEV), (2014) Rapport relatif à l'utilisation des insecticides et gestion de la résistance https://www.anses.fr/fr/system/files/CNEV-Ft Fev2014-Rapport_Utilisation_insecticides_gestion_resistance.pdf. Cohuet, A. (2020). L'efficacité des principaux répulsifs contre les moustiques les plus dangereux. Les cahiers de la Recherche. Santé, Environnement, Travail, (15), 39-42. Corbel, V., Fonseca, D. M., Weetman, D., Pinto, J., Achee, N. L., Chandre, F., Coulibaly, M. B., Thèse de Doctorat unique - 120 - Dusfour, I., Grieco, J., Juntarajumnong, W., Lenhart, A., Martins, A. J., Moyes, C., Ng, L. C., Raghavendra, K., Vatandoost, H., Vontas, J., Muller, P., Kasai, S., Fouque, F., ... David, J. P. (2017). International workshop on insecticide resistance in vectors of arboviruses, December 2016, Rio de Janeiro, Brazil. Parasites & vectors, 10(1), 278. https://doi.org/10.1186/s13071-017-2224-3 . Costantini, C., Badolo, A., & Ilboudo-Sanogo, E. (2004). Field evaluation of the efficacy and persistence of insect repellents DEET, IR3535, and KBR 3023 against Anopheles gambiae complex and other Afrotropical vector mosquitoes. Transactions of the Royal Society of Tropical Medicine and Hygiene, 98(11),
644-652. Cuisance, D., Politzar H., Clair M., Sellin E., and Tazé Y. (1978). Impact des lâchers de mâles stériles sur les niveaux de deux populations sauvages de <em> Glossina palpalis Gambiensis</Em> En Haute-Volta (Sources de La Volta Noire). Revue d'élevage et de Médecine Vétérinaire Des Pays Tropicaux 31 (3): 315. https://doi.org/10.19182/remvt.8126. Curtis, C. F., Lines J. D., Ijumba J., Callaghan A., and Hill N. (1987). The relative efficacy of repellents against mosquito vectors of disease. Medical and Veterinary Entomology 1: 109- 19. Dabire, K. R., Diabaté, A., Agostinho, F., Alves, F., Manga, L., Faye, O., & Baldet, T. (2008). Distribution of the members of Anopheles gambiae and pyrethroid knock-down resistance gene (kdr) in Guinea-Bissau, West Africa. Bulletin de la Societe de pathologie exotique (1990), 101(2), 119-123. Dabiré, K. R., Diabaté, A., Namountougou, M., Toé, K. H., Ouari, A., Kengne, P., Bass, C., and Baldet, T. (2009). Distribution of pyrethroid and ddt resistance and the l1014f kdr mutation in Anopheles gambiae s.l . from Burkina Faso ( West Africa ). Transactions of the Royal Society of Tropical Medicine and Hygiene 103: 1113-20. https://doi.org/10.1016/j.trstmh.2009.01.008. Dabiré, K. R., Diabaté, A., Namountougou, M., Djogbenou, L., Wondji, C., Chandre, F., Simard, F., Ouédraogo, J., Martin, T., Weill, M., & Baldet, T. (2012a). Trends in Insecticide Resistance in Natural Populations of Malaria Vectors in Burkina Faso, West Africa: 10 Thèse de Doctorat unique - 121 - Years' Surveys. In (Ed.), Insecticides - Pest Engineering. IntechOpen. https://doi.org/10.5772/28749. Dabiré, R. K., Namountougou, M., Sawadogo, S. P., Yaro, L. B., Toe, H. K., Ouari, A., Gouagna, L-C., Simard, F., Chandre, F., Baldet, T., Bass, C. and Diabate, A. (2012b). Population Dynamics of Anopheles gambiae s.l. in Bobo-Dioulasso City: Bionomics, Infection Rate and Susceptibility to Insecticides. Parasites and Vectors 5 (1): 1-9. https://doi.org/10.1186/1756-3305-5-127. Dabire, R. K., Namountougou, M., Diabaté, A., Soma, D. D., Bado, J., Toé, H. K., Bass, C. & Combary, P. (2014). Distribution and frequency of kdr mutations within Anopheles gambiae sl populations and first report of the ace. 1 G119S mutation in Anopheles arabiensis from Burkina Faso (West Africa). PloS one, 9(7),
e101484. Darriet, F. (1998). La lutte contre les moustiques nuisants et vecteurs de maladies : l'évaluation de nouveaux insecticides utilisables contre les moustiques en Afrique tropicale. Paris (FRA) ; Paris : Karthala ; ORSTOM, 114 p. (Economie et Développement). ISBN 2-86537-864-0. Dayananda, K. and Ranaweera, S., (1996). Mosquito-larvicidal activity of Ceylon citronella {Cymbopogon Nardus (L.) Rendle} oil fractions. Journal of the National Science Foundation of Sri Lanka, 24(4), pp.247-252. http://doi.org/10.4038/jnsfsr.v24i4.5557 de Araújo, A. P., Paiva, M., Cabral, A. M., Cavalcanti, A., Pessoa, L., Diniz, D., Helvecio, E., da Silva, E., da Silva, N. M., Anastácio, D. B., Pontes, C., Nunes, V., de Souza, M., Magalhães, F., de Melo Santos, M., & Ayres, C. (2019). Screening Aedes aegypti (Diptera: Culicidae) populations from Pernambuco, Brazil for Resistance to Temephos, Diflubenzuron, and Cypermethrin and Characterization of Potential Resistance Mechanisms. Journal of insect science (Online), 19(3), 16. https://doi.org/10.1093/jisesa/iez054 Debboun, M., & Strickman, D. (2013). Insect repellents and associated personal protection for a reduction in human disease. Medical and Veterinary Entomology, 27(1), 1-9. https://doi.org/10.1111/j.1365-2915.2012.01020.x. Dekker, T., Ignell, R., Ghebru, M., Glinwood, R., & Hopkins, R. (2011). Identification of mosquito repellent odours from Ocimum forskolei. Parasites & vectors, 4, 183. Thèse de Doctorat unique - 122 - https://doi.org/10.1186/1756-3305-4-183. Deletre, E., Martin, T., Campagne, P., Bourguet, D., Cadin, A., Menut, C., Bonafos, R. and Chandre, F. (2013). Repellent, irritant and toxic effects of 20 plant extracts on adults of the malaria vector Anopheles gambiae mosquito. PloS One, 8(12) : e82103. https://doi.org/10.1371/journal.pone.0082103 Deletre, E., Chandre, F., Williams, L., Duménil, C., Menut, C., & Martin, T. (2015). Electrophysiological and behavioral characterization of bioactive compounds of the Thymus vulgaris, Cymbopogon winterianus, Cuminum cyminum and Cinnamomum zeylanicum essential oils against Anopheles gambiae and prospects for their use as bednet treatments. Parasites & vectors, 8, 316. https://doi.org/10.1186/s13071-015-0934-y Deletre, E., Schatz, B., Bourguet, D., Chandre, F., Williams, L., Ratnadass, A., & Martin, T. (2016). Prospects for repellent in pest control : current developments and future challenges. Chemoecology, 26(4), 127-142. https://doi.org/ 10.1007/s00049-016-0214-0. Deletre, E., Martin, T., Duménil, C., & Chandre, F. (2019). Insecticide resistance modifies mosquito response to DEET and natural repellents. Parasites & vectors, 12(1): 1-10. https://doi: 10.1186/s13071-019-3343-9. Dell'Agli, M., Sanna, C., Rubiolo, P., Basilico, N., Colombo, E., Scaltrito, M. M., ... & Bosisio, E. (2012). Anti-plasmodial and insecticidal activities of the essential oils of aromatic plants growing in the Mediterranean area. Malaria journal, 11(1): 219. https://doi: 10.1186/14752875-11-219. Dhadialla, T. S., Carlson, G. R. and Le, D. P. (1998) `New insecticides with ecdysteroidal and juvenile hormone activity', Annual Review of Entomology, 43(1), pp. 545-569. https://doi.org/10.1146/annurev.ento.43.1.545 . Diabate, A., Baldet, T., Chandre, F., Akoobeto, M., Guiguemde, T. R., Darriet, F., Brengues, C., Guillet, P., Hemingway, J., Small, G. J., & Hougard, J. M. (2002). The role of agricultural use of insecticides in resistance to pyrethroids in Anopheles gambiae s.l. in Burkina Faso. The American journal of tropical medicine and hygiene, 67(6): 617-622. https://doi.org/10.4269/ajtmh.2002.67.617. Drabo, S. F., Gnankiné, O., Bassolé, I. H., Nébié, R. C., & Laurence, M. (2017). Susceptibility of Thèse de Doctorat unique - 123 - MED-Q1 and MED-Q3 Biotypes of Bemisia tabaci (Hemiptera: Aleyrodidae) populations to essential and seed oils. Journal of economic entomology, 110(3), 1031-1038.: 1031-38. https://doi.org/10.1093/jee/tox100. Dua, V. K., Pandey, A. C., and Dash, A. P. (2010). Adulticidal activity of essential oil of Lantana camara leaves against mosquitoes. Indian Journal of Medical Research 131 (3): 434-39. Duvallet, G., Fontenille, D., and Robert, V. (2017) Entomologie médicale et vétérinaire. IRD éditions. Edited by Éditions Quae. https://doi: 10.4000/books.irdeditions.22031. Enayati, A. A., Ranson, H., and Hemingway, J. (2005). Insect glutathione transferases and insecticide resistance. Insect Molecular Biology 14 (1): 3-8. https://doi.org/10.1111/j.1365-2583.2004.00529.x. Fegan, G. W., Noor, A. M., Akhwale, W. S., Cousens, S., & Snow, R. W. (2007). Effect of expanded insecticide-treated bednet coverage on child survival in rural Kenya: a longitudinal study. The Lancet, 370(9592): 1035-1039. Ffrench-Constant, R. H., Daborn, P. J., & Le Goff, G. (2004). The genetics and genomics of insecticide resistance. Trends in genetics: TIG,
20(3), 163-170. Foley, D. H., Bryan, J. H. and Lawrence, G. W. (2000) `The potential of ivermectin to control the malaria vector Anopheles farauti', Transactions of the Royal Society of Tropical Medicine and Hygiene, 94(6), pp. 625-628. doi: 10.1016/S0035-9203(00)90211-6. Flannery, D. J., Todres, J., Bradshaw, C. P., Amar, A. F., Graham, S., Hatzenbuehler, M., ... & Rivara, F. (2016). Bullying prevention: A summary of the report of the National Academies of Sciences, Engineering, and Medicine. Prevention Science, 17(8), 1044-1053. 1044-53. https://doi.org/10.1007/s11121-016-0722-8. Folashade, K. O., & Omoregie, E. H. (2012). Essential oil of Lippia multiflora Moldenke: A review. Journal of Applied Pharmaceutical Science, (Issue), 15-23. Fournet, F., Kassié, D., Dabiré, R. K., & Salem, G. (2015). Analyse de la distribution socio-spatiale du paludisme dans une ville moyenne ouest africaine, Bobo-Dioulasso (Burkina Faso). Dynamiques environnementales. Journal international de géosciences et de l'environnement, (36), 146-165. https://doi.org/10.4000/dynenviron.1004. Thèse de Doctorat unique - 124 - Fradin, M. S., & Day, J. F. (2002). Comparative efficacy of insect repellents against mosquito bites. The New England journal of medicine, 347(1), 13-18. https://doi.org/10.1056/NEJMoa011699 Gaire, S., Zheng, W., Scharf, M. E., & Gondhalekar, A. D. (2021). Plant essential oil constituents enhance deltamethrin toxicity in a resistant population of bed bugs (Cimex lectularius L.) by inhibiting cytochrome P450 enzymes. Pesticide biochemistry and physiology, 175, 104829. https://doi.org/10.1016/j.pestbp.2021.104829 Gbolade, A. A., and Lockwood, G. B. (2008). Toxicity of Ocimum sanctum L. Essential oil to Aedes aegypti larvae and its chemical composition. Journal of Essential Oil-Bearing Plants 11 (2): 148-53. https://doi.org/10.1080/0972060X.2008.10643611. Georges, S. O. (1979) « Les huiles essentielles - généralités et définitions », dans Traité de phytothérapie et d'aromathérapie, éd. Maloine, p. 141-142. Gillies, M. T., & Coetzee, M. (1987). A supplement to the Anophelinae of Africa South of the Sahara. Publ S Afr Inst Med Res, 55, 1-143. Gillij, Y. G., Gleiser, R. M., and Zygadlo J. A. (2008). Mosquito repellent activity of essential oils of aromatic plants growing in Argentina.» Bioresource Technology 99 (7): 2507-15. https://doi.org/10.1016/J.BIORTECH.2007.04.066. Gnankiné, O., & Bassolé, I. H. N. (2017). Essential oils as an alternative to pyrethroids' resistance against Anopheles species complex giles (Diptera: Culicidae). Molecules, 22(10), 1321. https://doi.org/10.3390/MOLECULES22101321. Gnankiné, O., Bassolé, I. H., Chandre, F., Glitho, I., Akogbeto, M., Dabiré, R. K., & Martin, T. (2013). Insecticide resistance in Bemisia tabaci Gennadius (Homoptera: Aleyrodidae) and Anopheles gambiae Giles (Diptera: Culicidae) could compromise the sustainability of malaria vector control strategies in West Africa. Acta Tropica, 128(1), 7-17. https://doi.org/10.1016/j.actatropica.2013.06.004. Grieco, J. P., Achee, N. L., Sardelis, M. R., Chauhan, K. R., & Roberts, D. R. (2005). A Novel high-throughput screening system to evaluate the behavioral response of adult mosquitoes to chemicals1. Journal of the American Mosquito Control Association, 21(4), 404-411. https://doi.org/10.2987/8756-971X(2006)21[404:ANHSST]2.0.CO;2. Thèse de Doctorat unique - 125 - Guarrera, P. M. (1999). Traditional antihelmintic, antiparasitic and repellent uses of plants in Central Italy. Journal of ethnopharmacology, 68(1-3), 183-192. https://doi.org/10.1016/s0378-8741(99)00089-6. Hao, H., Wei, J., Dai, J., & Du, J. (2014). Host-seeking and blood-feeding behavior of Aedes albopictus (Diptera: Culicidae) exposed to vapors of geraniol, citral, citronellal, eugenol, or anisaldehyde. Journal of Medical Entomology, 45(3), 533-539. https://doi.org/10.1603/0022-2585(2008)45[533:HABBOA]2.0.CO;2. Harbach, R. E. (2007). The Culicidae (Diptera): a review of taxonomy, classification and phylogeny. Zootaxa, 1668(1), 591-638. doi.org/10.11646/zootaxa.1668.1.28 Hardstone, M. C., Leichter, C. A., and Scott, J. G. (2009). Multiplicative interaction between the two major mechanisms of permethrin resistance, kdr and cytochrome p450-monooxygenase detoxification, in mosquitoes. Journal Evol. Biol 22: 416-23. https://doi.org/10.1111/j.1420-9101.2008.01661.x. Heath, H. B. (1981). Source Book of Flavors: (AVI Sourcebook and
Handbook Series) 1981st Edition. Hemingway, J., Hawkes, N. J., McCarroll, L., & Ranson, H. (2004). The molecular basis of insecticide resistance in mosquitoes. Insect biochemistry and molecular biology, 34(7), 653665. https://doi.org/10.1016/j.ibmb.2004.03.018. Hemingway, J., Ranson, H., Magill, A., Kolaczinski, J., Fornadel, C., Gimnig, J., Coetzee, M., Simard, F., Roch, D. K., Hinzoumbe, C. K., Pickett, J., Schellenberg, D., Gething, P., Hoppé, M., & Hamon, N. (2016). Averting a malaria disaster: will insecticide resistance derail malaria control?. Lancet (London, England), 387(10029), 1785-1788. https://doi.org/ 10.1016/S0140-6736(15)00417-1. Henry, M. C., Assi, S. B., Rogier, C., Dossou-Yovo, J., Chandre, F., Guillet, P., & Carnevale, P. (2005). Protective efficacy of lambda-cyhalothrin treated nets in Anopheles gambiae pyrethroid resistance areas of Côte d'Ivoire. The American journal of tropical medicine and hygiene, 73(5), 859-864. Hervy, J-P. and Sales, S. (1980) Évaluation de la rémanence de deux pyréthrinoïdes de synthèse Thèse de Doctorat unique - 126 - -OMS-1821 et OMS-1998 - après imprégnation de différents tissus entrant dans la confection de moustiquaires. Bobo-Dioulasso : OCCGE, pp. 1- 10. https://doi: fdi:10000. Hien, A. S., Soma, D. D., Maiga, S., Coulibaly, D., Diabaté, A., Belemvire, A., Diouf, M. B., Jacob, D., Koné, A., Dotson, E., Awolola, T. S., Oxborough, R. M., & Dabiré, R. K. (2021). Evidence supporting deployment of next generation insecticide treated nets in Burkina Faso : bioassays with either chlorfenapyr or piperonyl butoxide increase mortality of pyrethroid-resistant Anopheles gambiae. Malaria journal, 20(1), 406. https://doi.org/10.1186/s12936-021-03936-3 . Hoffmann, A. A., Montgomery, B. L., Popovici, J., Iturbe-Ormaetxe, I., Johnson, P. H., Muzzi,
suppress dengue transmission. Nature,
476(7361), 454-457. Hummelbrunner, L. A., and Isman, M. B. (2001). Acute, sublethal, antifeedant, and synergistic effects of monoterpenoid essential oil compounds on the Tobacco cutworm, Spodoptera Litura (Lep., Noctuidae). Journal of Agricultural and Food Chemistry 49 (2): 715-20. https://doi.org/10.1021/JF000749T. Hyldgaard, M., Mygind, T., & Meyer, R. L. (2012). Essential oils in food preservation: mode of action, synergies, and interactions with food matrix components. Frontiers in microbiology, 3, 12. https://doi.org/10.3389/fmicb.2012.00012. Ilboudo, Z., L. Dabiré C.B., Nébié R. C.H., Dicko I. O., Dugravot S., Cortesero A. M., and Sanon A. (2010). Biological activity and persistence of four essential oils towards the main pest of Stored Cowpeas, Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). Journal of Stored Products Research 46 (2): 124-28. https://doi.org/10.1016/j.jspr.2009.12.002. Ileke, K. D., Oyeniyi, E. A., Ogungbite, O. C., & Adesina, J. M. (2015). Nicotiana tabacum a prospective mosquitocide in the management of Anopheles gambiae (Giles). International Journal of Mosquito Research, 2(4): 19-23. Institut National de la Statistique et de la Démographie (INSD), (2020). Résultats préliminaires Thèse de Doctorat unique - 127 - du cinquième recensement général de la population et de l'habitation du Burkina Faso. Intirach, J., Junkum, A., Tuetun, B., Choochote, W., Chaithong, U., Jitpakdi, A., ... & Pitasawat, B. (2012). Chemical constituents and combined larvicidal effects of selected essential oils against Anopheles cracens (Diptera: Culicidae). Psyche, 2012. . https://doi.org/10.1155/2012/591616. Ishak, I. H., Kamgang, B., Ibrahim, S. S., Riveron, J. M., Irving, H., & Wondji, C. S. (2017). Pyrethroid resistance in Malaysian populations of dengue vector Aedes aegypti is mediated by CYP9 family of cytochrome P450 genes. PLoS neglected tropical diseases, 11(1): e0005302. https://doi.org/10.1371/journal.pntd.0005302. Isman, M. B. (2000). Plant essential oils for pest and disease management. Crop Protection 19 (8-10): 603-8. https://doi.org/10.1016/S0261-2194(00)00079-X. Iwu, M. W., Duncan, A. R., & Okunji, C. O. (1999). New antimicrobials of plant origin. Perspectives on new crops and new uses. ASHS Press, Alexandria, VA, 457, 462. Jones, J. W., Meisch, M. V., Meek, C. L. and Bivin, W. S. (1992) `Lethal effects of ivermectin on Anopheles quadrimaculatus.', Journal of the American Mosquito Control Association, 8(3), pp. 278-280. https://pubmed.ncbi.nlm.nih.gov/1402865/. Kalemba, D, and Kunicka A. (2003). Antibacterial and antifungal properties of essential oils. Current Medicinal Chemistry 10 (10): 813-29. Kamgang, B., Marcombe, S., Chandre, F., Nchoutpouen, E., Nwane, P., Etang, J., Corbel, V., & Paupy, C. (2011). Insecticide susceptibility of Aedes aegypti and Aedes albopictus in Central Africa. Parasites & vectors, 4: 79. https://doi.org/ 10.1186/1756-3305-4-79 . Katz, T. M., Miller, J. H., & Hebert, A. A. (2008). Insect repellents: historical perspectives and new developments. Journal of the American Academy of Dermatology, 58(5), 865-871. https://doi.org/10.1016/j.jaad.2007.10.005 Kawada, H., Oo, S. Z. M., Thaung, S., Kawashima, E., Maung, Y. N. M., Thu, H. M., ... & Minakawa, N. (2014). Co-occurrence of point mutations in the voltage-gated sodium channel of pyrethroid-resistant Aedes aegypti populations in Myanmar. PLoS Neglected Tropical Diseases, 8(7): e3032. https://doi.org/10.1371/journal.pntd.0003032. Kawada, H., Higa, Y., Futami, K., Muranami, Y., Kawashima, E., Osei, J. H., ... & Minakawa, Thèse de Doctorat unique - 128 - N. (2016). Discovery of point mutations in the voltage-gated sodium channel from African Aedes aegypti populations: potential phylogenetic reasons for gene introgression. PLOS Neglected Tropical Diseases, 10(6): e0004780. doi.org/10.1371/journal.pntd.0004780. Kiendrebeogo, M., Coulibaly, A. Y., Nebie, R. C., Zeba, B., Lamien, C. E., Lamien-Meda, A., & Nacoulma, O. G. (2011). Antiacetylcholinesterase and antioxidant activity of essential oils from six medicinal plants from Burkina Faso. Revista Brasileira de Farmacognosia, 21, 63-69. Knols, B. G. J., Farenhorst, M., Andriessen, R., Snetselaar, J., Suer, R. A., Osinga, A. J., Knols, J. M. H., Deschietere, J., Ng 'habi, K. R., Lyimo, I. N., Kessy, S. T., Mayagaya, V. S., Sperling, S., Cordel, M., Sternberg, E. D., Hartmann, P., Mnyone, L. L., Rose, A. and Thomas, M. B. (2016) `Eave tubes for malaria control in Africa: an introduction', Malaria Journal, 15(404): 1-7. doi: 10.1186/s12936-016-1452-x. Kobylinski, K. C., Sylla M., Chapman P. L., Sarr M. D. and Foy B. D. (2011) `Ivermectin mass drug administration to humans disrupts malaria parasite transmission in Senegalese villages', American Journal of Tropical Medicine and Hygiene, 85(1), pp. 3-5. doi: 10.4269/ajtmh.2011.11-0160. Kobylinski, K. C., Alout H., Foy B. D., Clements, A., Adisakwattana, P., Swierczewski, B. E. and Richardson, J. H. (2014) `Rationale for the coadministration of albendazole and ivermectin to humans for malaria parasite transmission control', American Journal of Tropical Medicine and Hygiene, 91(4), pp. 655-662. doi: 10.4269/ajtmh.14-0187. Kory, P., Meduri, G. U., Varon, J., Iglesias, J., & Marik, P. E. (2021). Review of the emerging evidence demonstrating the efficacy of ivermectin in the prophylaxis and treatment of COVID-19. American journal of therapeutics, 28(3), e299. doi: 10.1097/MJT.0000000000001377 Koul, O., Walia, S., & Dhaliwal, G. S. (2008). Essential oils as green pesticides: potential and constraints. Biopesticides international, 4(1), 63-84. Kpatinvoh, B., Adjou E. S., Dahouenon-Ahoussi E., Konfo C. T. R., Atrevi B., Soumanou M. M., Sohounhloue D. C. K. (2017). Efficacité des huiles essentielles de trois plantes aromatiques contre la mycoflore d'altération du niébé collecté dans les magasins de vente du Thèse de Doctorat unique - 129 - Sud-Bénin. Journal of Applied Biosciences 109 : 10680-10687. https://doi.org/10.4314/jab.v109i1.12. Kraemer, M. U., Sinka, M. E., Duda, K. A., Mylne, A. Q., Shearer, F. M., Barker, C. M., Moore, C. G., Carvalho, R. G., Coelho, G. E., Van Bortel, W., Hendrickx, G., Schaffner, F., Elyazar, I. R., Teng, H. J., Brady, O. J., Messina, J. P., Pigott, D. M., Scott, T. W., Smith, D. L., Wint, G. R., ... Hay, S. I. (2015). The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife, 4, e08347. https://doi.org/10.7554/eLife.08347 Kweka, E. J., Nyindo, M., Mosha, F., & Silva, A. G. (2011). Insecticidal activity of the essential oil from fruits and seeds of Schinus terebinthifolia Raddi against African malaria vectors. Parasites & Vectors, 4(1), 1-10. https://doi.org/10.1186/1756-3305-4-129. Lacey, L. A. (1990). The medical importance of riceland mosquito and their control using alternatives to chemical insecticides. Journal of the American Mosquito Control Association, no. 7. Lee, H. S. (2006). Mosquito larvicidal activity of aromatic medicinal plant oils against aedes aegypti and culex Pipiens pallens. Journal of the American Mosquito Control Association 22 (2): 292-95. https://doi.org/10.2987/8756-971X(2006)22[292:MLAOAM]2.0.CO;2. Lehmann, T., Dao, A., Yaro, A. S., Diallo, M., Timbiné, S., Huestis, D. L., ... & Traoré, A. I. (2014). Seasonal variation in spatial distributions of Anopheles gambiae in a Sahelian village: evidence for aestivation. Journal of medical entomology, 51(1), 27-38. Li, C., Gao, Y., Zhao, Z., Ma, D., Zhou, R., Wang, J., ... & Liu, Q. (2021). Potential geographical distribution of Anopheles gambiae worldwide under climate change. Journal of Biosafety and Biosecurity, 3(2), 125-130. doi.org/10.1016/j.jobb.2021.08.004 Licciardi, S., Hervé, J. P., Darriet, F., Hougard, J. M., & Corbel, V. (2006). Lethal and behavioural effects of three synthetic repellents (DEET, IR3535 and KBR 3023) on Aedes aegypti mosquitoes in laboratory assays. Medical and veterinary entomology, 20(3), 288293. https://doi.org/10.1111/j.1365-2915.2006.00630.x. Liu, T. T., Chao, L. K. P., Hong, K. S., Huang, Y. J., & Yang, T. S. (2019). Composition and Thèse de Doctorat unique - 130 - insecticidal activity of essential oil of Bacopa caroliniana and interactive effects of individual compounds on the activity. Insects, 11(1), 23. https://doi.org/10.3390/insects11010023. Lovett, B., Bilgo, E., Millogo, S. A., Ouattarra, A. K., Sare, I., Gnambani, E. J., Dabire, R. K., Diabate, A., & St Leger, R. J. (2019). Transgenic Metarhizium rapidly kills mosquitoes in a malaria-endemic region of Burkina Faso. Science (New York, N.Y.), 364(6443), 894- 897. https://doi.org/10.1126/science.aaw8737 Lucia, A., Audino, P. G., Seccacini, E., Licastro, S., Zerba, E., & Masuh, H. (2007). Larvicidal effect of Eucalyptus grandis essential oil and turpentine and their major components on Aedes aegypti larvae. Journal of the American Mosquito Control Association, 23(3), 299303. https://doi.org/10.2987/8756-971X(2007)23[299:LEOEGE]2.0.CO;2. Lundwall, E., Pennetier, C., Corbel, V., de Gentile, L., Legros, F. (2005) Paludisme: se protéger des piqûres d'anophèles. La Revue du Praticien. 55 Luz, T. R. S. A., de Mesquita, L. S. S., do Amaral, F. M. M., & Coutinho, D. F. (2020). Essential oils and their chemical constituents against Aedes aegypti L.(Diptera: Culicidae) larvae. Acta Tropica, 212, 105705. https://doi.org/10.1016/j.actatropica.2020.105705. Maïga, H., Damiens, D., Niang, A., Sawadogo, S. P., Fatherhaman, O., Lees, R. S., Roux, O., Dabiré, R. K., Ouedraogo, G. A., Tripet, F., Diabaté A. & Gilles, J. R. (2014). Mating competitiveness of sterile male Anopheles coluzzii in large cages. Malaria Journal, 13(1), 16. https://doi.org/ 10.1186/1475-2875-13-460. Manh, H. D., Hue, D. T., Hieu, N. T. T., Tuyen, D. T. T., & Tuyet, O. T. (2020). The Mosquito larvicidal activity of essential oils from Cymbopogon and Eucalyptus Species in Vietnam. Insects, 11(2), 128. https://doi.org/10.3390/insects11020128. Mansour, S. A., Foda, M. S., & Aly, A. R. (2012). Mosquitocidal activity of two Bacillus bacterial endotoxins combined with plant oils and conventional insecticides. Industrial crops and products, 35(1), 44-52. https://doi.org/ 10.1016/j.indcrop.2011.06.001. Marcombe, S., Fustec, B., Cattel, J., Chonephetsarath, S., Thammavong, P., Phommavanh, N., David, J. P., Corbel, V., Sutherland, I. W., Hertz, J. C., & Brey, P. T. (2019). Thèse de Doctorat unique - 131 - Distribution of insecticide resistance and mechanisms involved in the arbovirus vector Aedes aegypti in Laos and implication for vector control. PLoS neglected tropical diseases, 13(12), e0007852. https://doi.org/10.1371/journal.pntd.0007852. Marshall, J. M., White, M. T., Ghani, A. C., Schlein, Y., Muller, G. C. and Beier ,J. C. (2013) `Quantifying the mosquito's sweet tooth: Modelling the effectiveness of attractive toxic sugar baits (ATSB) for malaria vector control', Malaria Journal, 12(291), pp. 1-13. doi: 10.1186/1475-2875-12-291. Martinez-Torres, D., Chandre, F., Williamson, M. S., Darriet, F., Berge, J. B., Devonshire, A. L., ... & Pauron, D. (1998). Molecular characterization of pyrethroid knockdown resistance (kdr) in the major malaria vector Anopheles gambiae ss. Insect molecular biology, 7(2): 179-184. https://doi.org/10.1046/j.1365-2583.1998.72062.x. Mbare, O., Lindsay, S. W., & Fillinger, U. (2014). Pyriproxyfen for mosquito control: female sterilization or horizontal transfer to oviposition substrates by Anopheles gambiae sensu stricto and Culex quinquefasciatus. Parasites & vectors, 7(1), 1-12. https://doi.org/10.1186/1756-3305-7-280 Ministère de la Sante du Burkina Faso, Direction Des Statistiques Sectorielles, (2022). Paludisme Au Burkina: Bulletin Epidemiologique, 4-7. Ministère de la Santé du Burkina Faso, Direction Des Statistiques Sectorielles, (2017). Flambée de Cas de Dengue Au Burkina Faso. Sitrep Dengue. no. 26: 11 Minjas, J. N., & Sarda, R. K. (1986). Laboratory observations on the toxicity of Swartzia madagascariensis (Leguminosae) extract to mosquito larvae. Transactions of the Royal Society of Tropical Medicine and Hygiene, 80(3), 460-461. https://doi.org/10.1007/s11101-010-9170-4. Moore, S. J., Lenglet, A., & Hill, N. (2002). Field evaluation of three plant-based insect repellents against malaria vectors in Vaca Diez Province, the Bolivian Amazon. Journal of the American Mosquito Control Association, 18(2), 107-110. Moretti, A. N., Zerba E. N., and Alzogaray, R. A. (2013). Behavioral and toxicological responses of rhodnius prolixus and triatoma infestans (Hemiptera: Reduviidae) to 10 monoterpene alcohols. Journal of Medical Entomology 50 (5): 1046-54. Thèse de Doctorat unique - 132 - https://doi.org/10.1603/ME12248. Mosqueira, B., Duchon S., Chandre F., Hougard J. M., Carnevale P. and Mas-Coma S. (2010) `Efficacy of an insecticide paint against insecticide-susceptible and resistant mosquitoes - Part 1: Laboratory evaluation', Malaria Journal, 9(340): 1-6. doi: 10.1186/14752875-9-340. Mouchès, C., Magnin, M., Bergé, J. B., De Silvestri, M., Beyssat, V., Pasteur, N., & Georghiou, G. P. (1987). Overproduction of detoxifying esterases in organophosphate-resistant Culex mosquitoes and their presence in other insects. Proceedings of the National Academy of Sciences, 84(8): 2113-2116. Müller, G. C., Beier, J. C., Traore, S. F., Toure, M. B., Traore, M. M., Bah, S., Doumbia, S. and Schlein, Y. (2010) `Field experiments of Anopheles gambiae attraction to local fruits/seedpods and flowering plants in Mali to optimize strategies for malaria vector control in Africa using attractive toxic sugar bait methods', Malaria Journal, 9(1): 111. https://doi.org/10.1186/1475-2875-9-262. Murugan, K., and Kumar P. M. (2012). Larvicidal, pupicidal , repellent and adulticidal activity of citrus sinensis orange peel extract against Anopheles stephensi, Aedes Aegypti and Culex quinquefasciatus (Diptera : Culicidae). Parasitology Research 111: 1757-69. https://doi.org/10.1007/s00436-012-3021-8. Muthu, C., Ayyanar M., Raja N., and Ignacimuthu S. (2006). Medicinal plants used by traditional healers in kancheepuram district of Tamil Nadu, India. Journal of Ethnobiology and Ethnomedicine 2: 1-10. https://doi.org/10.1186/1746-4269-2-43. Muturi, E. J., Ramirez, J. L., Doll, K. M., & Bowman, M. J. (2017). Combined toxicity of three essential oils against Aedes aegypti (Diptera: Culicidae) larvae. Journal of Medical Entomology, 54(6), 1684-1691. https://doi.org/10.1093/jme/tjx168. Namountougou, M., Diabaté, A., Etang, J., Bass, C., Sawadogo, S. P., Gnankiné, O., Baldet T., Martin T., Chandre F., Simard F. & Dabiré, R. K. (2013). First report of the L1014S kdr mutation in wild populations of Anopheles gambiae M and S molecular forms in Burkina Faso (West Africa). Acta tropica, 125(2): 123-127. https://doi.org/10.1016/j.actatropica.2012.10.012. Namountougou, M., Soma, D. D., Kientega, M., Balboné, M., Kaboré, D. P. A., Drabo, S. F., Thèse de Doctorat unique - 133 - Coulibaly A. Y., Fournet F., Baldet T., Diabaté A., Dabiré R. K. & Gnankine, O. (2019). Insecticide resistance mechanisms in Anopheles gambiae complex populations from Burkina Faso, West Africa. Acta tropica, 197:
105054. https://doi.org/ Namountougou, M., Soma, D. D., Balboné, M., Kaboré, D. A., Kientega, M., Hien, A., Coulibaly A., Ouattara P. E., Meda B. G., Drabo S., Koala L., Nignan C., Kagone T., Diabate A., Fournet F., Gnankine O. & Dabiré, R. K. (2020). Monitoring insecticide susceptibility in Aedes Aegypti populations from the two biggest cities, Ouagadougou and Bobo-Dioulasso, in Burkina Faso: implication of metabolic resistance. Tropical Medicine and Infectious Disease, 5(2), 84. https://doi.org/ 10.3390/tropicalmed5020084. Nararak, J., Sathantriphop, S., Kongmee, M., Mahiou-Leddet, V., Ollivier, E., Manguin, S., & Chareonviriyaphap, T. (2019). Excito-repellent activity of â-caryophyllene oxide against Aedes aegypti and Anopheles minimus. Acta tropica, 197, 105030. Ndirangu, E. G., Opiyo, S., & Ng'ang'a, M. W. (2020). Chemical composition and repellency of Nigella sativa L. seed essential oil against Anopheles gambiae sensu stricto. Trends in Phytochemical Research, 4(2), 77-84. Nerio, L. S., Olivero-Verbel, J., & Stashenko, E. E. (2009). Repellent activity of essential oils from seven aromatic plants grown in Colombia against Sitophilus zeamais Motschulsky (Coleoptera). Journal of Stored Products Research, 45(3): 212-214. https://doi.org/10.1016/j.jspr.2009.01.002. N'Guessan, R., Corbel, V., Akogbéto, M., & Rowland, M. (2007). Reduced efficacy of insecticide-treated nets and indoor residual spraying for malaria control in pyrethroid resistance area, Benin. Emerging infectious diseases, 13(2): 199. https://doi.org/10.3201/eid1302.060631. Nikou, D., Ranson H., and Hemingway J. (2003). An adult-specific CYP6 P450 gene is overexpressed in a pyrethroid-resistant strain of the malaria vector, Anopheles gambiae.» gene 318: 91-102. https://doi.org/10.1016/S0378-1119(03)00763-7. Nolen, J. A., Bedoukian R. H., Maloney R. E., and Kline D. L. (2002). Method, apparatus and compositions for inhibiting the human scent tracking ability of mosquitoes in Thèse de Doctorat unique - 134 - environmentally defined three dimensional spaces. U.S. Patent No 6,362,235, 26 Mars 2002. Nolan, T., Papathanos P., Windbichler N., Magnusson K., Benton J., Catteruccia F., Crisanti A. (2011) Developing transgenic Anopheles mosquitoes for the sterile insect technique. Genetica. 139(1):33-9. Norris, E. J., Johnson, J. B., Gross, A. D., Bartholomay, L. C., & Coats, J. R. (2018). Plant essential oils enhance diverse pyrethroids against multiple strains of mosquitoes and inhibit detoxification enzyme processes. Insects, 9(4), 132. https://doi.org/10.3390/insects9040132 Ntonga, P. A., Baldovini, N., Mouray, E., Mambu, L., Belong, P., & Grellier, P. (2014). Activity of Ocimum basilicum, Ocimum canum, and Cymbopogon citratus essential oils against Plasmodium falciparum and mature-stage larvae of Anopheles funestus ss. Parasite, 21 (6): 33. https://doi.org/10.1051/parasite/2014033. Ntonga, A. P., Jazet, D. P. M., Tonga, C., Mache, N. P., & Kekeunou, S. (2017). Evaluation de la toxicité des huiles essentielles des feuilles de Callistemon citrinus, Cinnamomum zeylanicum et Psidium guajava sur les adultes de Anopheles coluzzii Coetzee et Wilkerson 2013, vecteur du paludisme urbain au Cameroun. Cameroon Journal of Biological and Biochemical Sciences, 25, 12-21. Ojewumi, M. E., Adeyemi, A. O., & Ojewumi, E. O. (2018). Oil extract from local leaves-an alternative to synthetic mosquito repellants. Pharmacophore, 9(2), 1-6. http://www.pharmacophorejournal.com. Omolo, M. O., Okinyo D., Ndiege I. O., and Hassanali A. (2004). Repellency of essential oils of some Kenyan plants against Anopheles gambiae 65: 2797-2802. https://doi.org/10.1016/j.phytochem.2004.08.035. Oparaocha, E. T., Iwu, I., & Ahanaku, J. E. (2010). Preliminary study on mosquito repellent and mosquitocidal activities of Ocimum gratissimum (L.) grown in eastern Nigeria. Journal of Vector Borne Diseases, 47(1), 45. Organisation Mondiale de la Santé (OMS). (1992). Vector resistance for pesticides. World Health Organization technical report serial 818, p. 62. OMS, Geneve, Suisse. Organisation Mondiale de la Santé (OMS). (2001). Report of the fourth WHOPES working Thèse de Doctorat unique - 135 - group meeting. Review of: IR3535; KBR3023; (RS)-methoprene 20% EC; pyriproxyfen 0.5% GR and lambda-cyhalothrin 2.5% CS. Document WHO/CDS/WHOPES/2001.2. OMS, Geneve, Suisse. Organisation Mondiale de la Santé (OMS). (2002). Stratégie de l ' OMS pour la médecine traditionnelle pour 2002 - 2005.» OMS, Geneve, Suisse. Organisation Mondiale de la Santé (OMS). (2006). Pesticides and their application for the control of vectors and pests of public health importance. Document WHO/CDS/WHOPES/GCDPP/2006.1. OMS, Geneve, Suisse. Organisation Mondiale de la Santé (OMS). (2012). Global plan for insecticide resistance management in malaria vectors. Organisation Mondiale de la Santé (OMS). (2013). Guidelines for efficacy testing of spatial repellents. OMS, Geneve, Suisse. Organisation Mondiale de la Santé (OMS). (2014). Entomologie du paludisme et lutte antivectorielle, 1-200, OMS, Geneve, Suisse. Organisation Mondiale de la Santé (OMS). (2017). Procédures pour tester la résistance aux insecticides chez les moustiques vecteurs du paludisme Seconde Édition. OMS, Geneve, Suisse Organisation Mondiale de la Santé (OMS). (2019). Guidelines for malaria vector control. OMS, Geneve, Suisse. Organisation Mondiale de la Santé (OMS). (2020). World Malaria Report. World Health. Vol. WHO/HTM/GM. OMS, Geneve, Suisse. https://doi.org/ISBN 978 92 4 1564403. Organisation Mondiale de la Santé (OMS). (2021). Éthique et maladies à transmission vectorielle : orientations de l'OMS. OMS, Geneve, Suisse Organisation Mondiale de la Santé (OMS). (2022a). Dengue et Dengue Sévère. OMS, Geneve, Suisse. Organisation Mondiale de la Santé (OMS). (2022b). Standard Operating Procedure for Testing Thèse de Doctorat unique - 136 - Insecticide Susceptibility of Adult Mosquitoes in WHO Tube Tests. OMS, Geneve, Suisse. Osborn, F. R., Herrera, M. J., Gómez, C. J., & Salazar, A. (2007). Comparison of two commercial formulations of Bacillus thuringiensis var. israelensis for the control of Anopheles aquasalis (Diptera: Culicidae) at three salt concentrations. Memórias do Instituto Oswaldo Cruz, 102: 69-72. Ouattara, L. P. E., Sangaré, I., Namountougou, M., Hien, A., Ouari, A., Soma, D. D., ... & Dabiré, K. R. (2019). Surveys of arboviruses vectors in four cities stretching along a railway transect of Burkina Faso: risk transmission and insecticide susceptibility status of potential vectors. Frontiers in Veterinary Science, 6, 140. https://doi.org/10.3389/fvets.2019.00140. Ouédraogo, S., Benmarhnia, T., Bonnet, E., Somé, P. A., Barro, A. S., Kafando, Y., ... & Ridde, V. (2018). Evaluation of effectiveness of a community-based intervention for control of dengue virus vector, Ouagadougou, Burkina Faso. Emerging infectious diseases, 24(10), 1859. Oyedele, A. O., Gbolade A. A., Sosan M. B., Adewoyin F. B., Soyelu O. L., and Orafidiya O. O. (2002). Formulation of an effective mosquito-repellent topical product from lemongrass oil. Phytomedicine 9 (3): 259-62. https://doi.org/10.1078/0944-7113-00120. Pagès, F., Orlandi-pradines, E., Corbel, V. (2007). Vecteurs du paludisme: biologie, diversité, contrôle et protection individuelle. Médecine Malafies Infect. 37, 153-161. https://doi.org/10.1016/j.medmal.2006.10.009. Pålsson, K., and Jaenson T. G.T. (1999). Plant products used as mosquito repellents in Guinea Bissau, West Africa. Acta Tropica 72 (1): 39-52. https://doi.org/10.1016/S0001-706X(98)00083-7. Paluch, G., Bartholomay, L., & Coats, J. (2010). Mosquito repellents: a review of chemical structure diversity and olfaction. Pest management science, 66(9), 925-935. https://doi.org/10.1002/ps.1974. Park, H. M., Kim, J., Chang, K. S., Kim, B. S., Yang, Y. J., Kim, G. H., ... & Park, I. K. (2011). Larvicidal activity of Myrtaceae essential oils and their components against Aedes aegypti, acute toxicity on Daphnia magna, and aqueous residue. Journal of medical entomology, 48(2), 405-410. https://doi.org/10.1603/ME10108. Thèse de Doctorat unique - 137 - Pavela, R. (2008). Acute and synergistic effects of some monoterpenoid essential oil compounds on the house fly (Musca Domestica L.). Journal of Essential Oil-Bearing Plants 11 (5): 451-59. https://doi.org/10.1080/0972060X.2008.10643653. Pavela, R. (2014). Acute, synergistic and antagonistic effects of some aromatic compounds on the Spodoptera littoralis Boisd.(Lep., Noctuidae) larvae. Industrial crops and products, 60, 247258. https://doi.org/10.1016/j.indcrop.2014.06.030. Pavela, R. (2015). Acute toxicity and synergistic and antagonistic effects of the aromatic compounds of some essential oils against Culex quinquefasciatus Say larvae. Parasitology Research, 114(10), 3835-3853. https://doi.org/10.1007/s00436-015-4614-9. Peerzada, M., N. (1997). Molecules chemical composition of the essential oil of Hyptis Suaveolens. Molecules 2 (11): 165-68. Plapp, F. W. (1984). The genetic basis of insecticide resistance in the house fly: evidence that a single locus plays a major role in metabolic resistance to insecticides. Pesticide Biochemistry and Physiology 201: 194-201. Poda, S. B., Soma, D. D., Hien, A., Namountougou, M., Gnankiné, O., Diabaté, A., ... & Dabiré, R. K. (2018). Targeted application of an organophosphate-based paint applied on windows and doors against Anopheles coluzzii resistant to pyrethroids under real life conditions in Vallée du Kou, Burkina Faso (West Africa). Malaria Journal, 17(1): 1-9. https://doi.org/10.1186/s12936-018-2273-x. Pohlit, A. M., Lopes, N. P., Gama, R. A., Tadei, W. P., & Neto, V. F. (2011). Patent literature on mosquito repellent inventions which contain plant essential oils--a review. Planta medica, 77(6) : 598-617. https://doi.org/10.1055/s-0030-1270723 . Pooda H. S., Rayaisse J. B., Hien D. F. D. S., Lefèvre T., Yerbanga S. R., Bengaly Z., Dabiré R. K., Belem A. M. G., Sidibé I., Solano P. and Mouline K. (2015) `Administration of ivermectin to peridomestic cattle: A promising approach to target the residual transmission of human malaria', Malaria Journal, 14(486): 1-12. doi: 10.1186/s12936-015-1001-z. Ramirez, G. I. J., Logan J. G., Loza-Reyes E., Elena Stashenko E., and Moores G. D. (2012) Repellents inhibit P450 enzymes in Stegomyia (Aedes) aegypti. PLoS One 7: e48678. https://doi.org/ 10.1371/journal.pone.0048698 . Thèse de Doctorat unique - 138 - Ranson, H., Jensen, B., Vulule, J. M., Wang, X., Hemingway, J., & Collins, F. H. (2000). Identification of a point mutation in the voltage-gated sodium channel gene of Kenyan Anopheles gambiae associated with resistance to DDT and pyrethroids. Insect molecular biology, 9(5), 491-497. Ranson, H., N'guessan, R., Lines, J., Moiroux, N., Nkuni, Z., & Corbel, V. (2011). Pyrethroid resistance in African anopheline mosquitoes: what are the implications for malaria control?. Trends in parasitology, 27(2): 91-98. https://doi.org/10.1016/j.pt.2010.08.004. Ranson, H., & Lissenden, N. (2016). Insecticide Resistance in African Anopheles Mosquitoes: A Worsening Situation that Needs Urgent Action to Maintain Malaria Control. Trends in parasitology, 32(3) : 187-196. https://doi.org/ 10.1016/j.pt.2015.11.010. Rants'o, T. A., Koekemoer, L. L., Panayides, J. L., & van Zyl, R. L. (2022). Potential of Essential Oil-Based Anticholinesterase Insecticides against Anopheles Vectors: A Review. Molecules (Basel, Switzerland), 27(20), 7026. https://doi.org/10.3390/molecules27207026. Redwane, A., Lazrek H. B., Bouallam S., Markouk M., Amarouch H., and Jana M. (2002). «Larvicidal activity of extracts from Quercus lusitania Var. Infectoria galls (Oliv.). Journal of Ethnopharmacology 79 (2): 261-263. https://doi.org/10.1016/S0378-8741(01)00390-7. Rehman, J. U., Ali, A., & Khan, I. A. (2014). Plant based products: use and development as repellents against mosquitoes: A review. Fitoterapia, 95, 65-74. https://doi.org/10.1016/j.fitote.2014.03.002. Robert, V., Ayala D. and Simard F. (2017) `Chapitre 10. Les anophèles (Diptera : Culicidae : Anophelinae)', in Entomologie médicale et vétérinaire. IRD éditions. Marseille, pp. 181241. doi: 10.4000/books.irdeditions.22031. Rodhain, F., & Perez, C. (1985). Precis d'entomologie medicale et veterinaire; notions d'epidemiologie des maladies a vecteurs. Rohrer, S. P., Birzin, E. T., Eary, C. H., Schaeffer, J. M. and Shoop, W. L. (1994) `Ivermectin binding sites in sensitive and resistant Haemonchus contortus', The Journal of Parasitology, 80(3): 493-493. doi: 10.2307/3283426. Rübsamen, H., Montgomery, M., Hess, G. P., Eldefrawi, A. T., & Eldefrawi, M. E. (1976). Thèse de Doctorat unique - 139 - Identification of a calcium-binding subunit of the acetylcholine receptor. Biochemical and Biophysical Research Communications, 70(3), 1020-1027. Rutledge, L. C., Collister, D. M., Meixsell, V. E., and Eisenberg, G. H. G. (1983). Comparative sensitivity of representative mosquitoes (DIPTERA: CULICIDAE) to repellents. Journal of Medical Entomology 20 (5): 506-10. Rutledge, L. C., Hooper, R. L., Wirtz, R. A., & Gupta, R. K. (1989). Efficacy of diethyl methylbenzamide (DEET) against Aedes dorsalis and a comparison of two end points for protection time. Journal of the American Mosquito Control Association, 5(3): 363-368. Samba, N., Aitfella-Lahlou, R., Nelo, M., Silva, L., Coca, R., Rocha, P., & López Rodilla, J. M. (2020). Chemical Composition and Antibacterial Activity of Lippia multiflora Moldenke Essential Oil from Different Regions of Angola. Molecules (Basel, Switzerland), 26(1), 155. https://doi.org/10.3390/molecules26010155. Sampaio, V. S., Beltrán, T. P., Kobylinski, K. C., Melo, G. C., Lima, J. B. P., Silva, S. G. M., Rodriguez, Í. C., Silveira H., Guerra M. G. V. B., Bassat Q., Pimenta P. F. P., Lacerda M. V. G. and Monteiro W. M. (2016) `Filling gaps on ivermectin knowledge: e ects on the survival and reproduction of Anopheles aquasalis, a Latin American malaria vector', Malaria Journal, 15(491): 1-9. doi: 10.1186/s12936-016-1540-y. Sarma, R., Adhikari, K., Mahanta, S., & Khanikor, B. (2019). Combinations of plant essential oil based terpene compounds as larvicidal and adulticidal agent against Aedes aegypti (Diptera: Culicidae). Scientific reports, 9(1): 1-12. https://doi.org/10.1038/s41598-019-45908-3. Sawadogo, I., Paré, A., Kaboré, D., Montet, D., Durand, N., Bouajila, J., ... & Bassolé, I. H. N. (2022). Antifungal and antiaflatoxinogenic effects of Cymbopogon citratus, Cymbopogon nardus, and Cymbopogon schoenanthus essential oils alone and in combination. Journal of Fungi, 8(2): 117. https://doi.org/10.3390/jof8020117. Sawadogo, S. P., Diabaté, A., Toé, H. K., Sanon, A., Lefevre, T., Baldet, T., ... & Dabiré, R. K. (2013). Effects of age and size on Anopheles gambiae ss male mosquito mating success. Journal of medical entomology, 50(2): 285-293. https://doi.org/10.1603/ME12041. Schreck, C. E., & McGovern, T. P. (1989). Repellents and other personal protection strategies against Aedes albopictus. Journal of the American Mosquito Control Association, 5(2): 247- Thèse de Doctorat unique - 140 - 250. Semmler, M., Abdel-Ghaffar, F., Schmidt, J., & Mehlhorn, H. (2014). Evaluation of biological and chemical insect repellents and their potential adverse effects. Parasitology research, 113(1), 185-188. https://doi.org/10.1007/s00436-013-3641-7. Sengottayan, S. N. (2007). The use of Eucalyptus tereticornis sm. (Myrtaceae) oil (Leaf Extract) as a natural larvicidal agent against the malaria vector Anopheles stephensi Liston (Diptera : Culicidae). Bioresource Technology 98: 1856-60. Sinka, M. E., Bangs, M. J., Manguin, S., Coetzee, M., Mbogo, C. M., Hemingway, J., ... & Hay, S. I. (2010). The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: occurrence data, distribution maps and bionomic précis. Parasites & vectors, 3(1): 1-34. https://doi.org/10.1186/1756-3305-3-117. Sinka, M. E., Bangs, M. J., Manguin, S., Rubio-Palis, Y., Chareonviriyaphap, T., Coetzee, M., ... & Hay, S. I. (2012). A global map of dominant malaria vectors. Parasites & vectors, 5(1): 1-11. https://doi.org/10.1186/1756-3305-5-69. Sinkins, S. P., and Gould, F. (2006). Gene drive systems for insect disease vectors.» Nature Reviews Genetics 7 (6): 427-35. https://doi.org/10.1038/nrg1870. Slater, H. C., Walker, P. G., Bousema, T., Okell, L. C., & Ghani, A. C. (2014). The potential impact of adding ivermectin to a mass treatment intervention to reduce malaria transmission: a modelling study. The Journal of infectious diseases, 210(12), 1972-1980. https://doi.org/10.1093/infdis/jiu351. Slater, H. C., Foy, B. D., Kobylinski, K., Chaccour, C., Watson, O. J., Hellewell, J., Aljayyoussi, G., Bousema, T., Burrows, J., D'Alessandro, U., Alout, H., Ter Kuile, F. O., Walker, P., Ghani, A. C., & Smit, M. R. (2020). Ivermectin as a novel complementary malaria control tool to reduce incidence and prevalence: a modelling study. The Lancet. Infectious diseases, 20(4), 498-508. https://doi.org/10.1016/S1473-3099(19)30633-4. Smith, David L., Perkins T. A., Tusting L. S., Scott T. W., and Lindsay S. W. (2013). Mosquito population regulation and larval source management in heterogeneous environments.» PLoS ONE 8 (8): e71247. https://doi.org/10.1371/journal.pone.0071247. Thèse de Doctorat unique - 141 - Snetselaar, J., Njiru, B. N., Gachie, B., Owigo, P., Andriessen, R., Glunt, K., Osinga, A. J., Mutunga, J., Farenhorst, M. and Knols, B. G. J. (2017) `Eave tubes for malaria control in Africa: Prototyping and evaluation against Anopheles gambiae s.s. and Anopheles arabiensis under semi-field conditions in western Kenya Lucy Tusting, Jo Lines', Malaria Journal, 16(1), pp. 1-11. doi: 10.1186/s12936-017-1926-5. Soma, D. D., Zogo, B. M., Somé, A., Tchiekoi, B. N., Hien, D. F. de S., Pooda, H. S., ... Dabiré, R. K. (2020). Anopheles bionomics, insecticide resistance and malaria transmission in southwest Burkina Faso: A pre-intervention study. PLOS ONE, 15(8), e0236920. https://doi.org/10.1371/journal.pone.0236920. Sombié, A., Saiki, E., Yaméogo, F., Sakurai, T., Shirozu, T., Fukumoto, S., Sanon, A., Weetman, D., McCall, P. J., Kanuka, H., & Badolo, A. (2019). High frequencies of F1534C and V1016I kdr mutations and association with pyrethroid resistance in Aedes aegypti from Somgandé (Ouagadougou), Burkina Faso. Tropical medicine and health, 47, 2. https://doi.org/10.1186/s41182-018-0134-5. Sternberg, E. D., Ng'habi, K. R., Lyimo, I. N., Kessy, S. T., Farenhorst, M., Thomas, M. B., Knols, B. G. J. and Mnyone, L. L. (2016) `Eave tubes for malaria control in Africa: initial development and semi-field evaluations in Tanzania', Malaria Journal, 15(447): 111. https://doi.org/10.1186/s41182-018-0134-5. Sultana, S, Ali M, Ansari, SH, Bagri, P. (2008). The effect of physical factors on chemical composition of the essential oil of Eucalyptus citriodora Hook. leaves. J Essent Oil bearing Plants; 11: 69-74. https://doi.org/10.1080/0972060X.2008.10643600. Suputtamongkol, Y., Premasathian, N., Bhumimuang, K., Waywa, D., Nilganuwong, S., Karuphong, E., Anekthananon, T., Wanachiwanawin, D., & Silpasakorn, S. (2011). Efficacy and safety of single and double doses of ivermectin versus 7-day high dose albendazole for chronic strongyloidiasis. PLoS neglected tropical diseases, 5(5), e1044. https://doi.org/10.1371/journal.pntd.0001044. Sylla, M., Gray, M., Chapman, P. L., Sarr, M. D. and Rasgon, J. L. (2010) `Mass drug Thèse de Doctorat unique - 142 - administration of ivermectin in south-eastern Senegal reduces the survivorship of wild-caught, blood fed malaria vectors', Malaria Journal, 9(365): 1-10. https://doi.org/10.1186/14752875-9-365. Tanprasit, P. (2005). Biological control of dengue fever mosquitoes (Aedes aegypti Linn.) using leaf extracts of chan (Hyptis suaveolens (l.) Poit.) and hedge flower (Lantana camara Linn.). Tawatsin, A., Wratten, S. D., Scott, R. R., Thavara, U., & Techadamrongsin, Y. (2001). Repellency of volatile oils from plants against three mosquito vectors. Journal of vector ecology: journal of the Society for Vector Ecology, 26(1), 76-82. Tchoumbougnang, F., Dongmo P. M. J., Sameza M. L., Mbanjo E. G. N., Fotso G.B. T., Zollo P. H. A., and Menut C. (2009). Activité larvicide sur Anopheles gambiae Giles et composition chimique des huiles essentielles extraites de quatre plantes cultivées au Cameroun.» Biotechnol. Agron. Soc. Environ. Vol. 13 numéro 1. Toé, H. K., Zongo, S., Guelbeogo, M. W., Kamgang, B., Viana, M., Tapsoba, M., Sanon A., Traoré A., McCall P. J., & Sagnon, N. F. (2022). Multiple insecticide resistance and first evidence of V410L kdr mutation in Aedes (Stegomyia) aegypti (Linnaeus) from Burkina Faso. Medical and Veterinary Entomology, 36(3),
309-319. Tomé, H. V., Pascini, T. V., Dângelo, R. A., Guedes, R. N., & Martins, G. F. (2014). Survival and swimming behavior of insecticide-exposed larvae and pupae of the yellow fever mosquito Aedes aegypti. Parasites & Vectors, 7(1): 1-9. https://doi.org/10.1186/1756-3305-7-195. Traore, M. M., Junnila, A., Traore, S. F., Doumbia, S., Revay, E. E., Kravchenko, V. D., Schlein, Y., Arheart, K. L., Gergely, P., Xue R. D., Hausmann A., Beck R., Prozorov A., Diarra R. A., Kone A. S., Majambere S., Bradley, J., Vontas, J., Beier, J. C. and Müller, G. C. (2020) `Large-scale field trial of attractive toxic sugar baits (ATSB) for the control of malaria vector mosquitoes in Mali, West Africa', Malaria Journal, 19(72) : 1-16. https://doi.org/ 10.1186/s12936-020-3132-0. Thèse de Doctorat unique - 143 - Travis, B. V. (1947). Relative Effectiveness of various Repellents against Anopheles farauti Laveran. Journal of the National Malaria Society, 6(3): 180-83. Tripathi, A. K., Upadhyay, S., Bhuiyan, M., & Bhattacharya, P. R. (2009). A review on prospects of essential oils as biopesticide in insect-pest management. J. Pharmacognosy Phytotherapy, 1, 052-063. https://doi.org/10.5897/JPP.9000003. Trpis, M., McClelland, G. A. H., Gillett, J. D., Teesdale, C., & Rao, T. R. (1973). Diel periodicity in the landing of Aedes aegypti on man. Bulletin of the World Health Organization, 48(5): 623. Vani, R. S., Cheng S. F., and Chuah C. H. (2009). Comparative study of volatile compounds from genus Ocimum. American Journal of Applied Sciences 6 (3): 523-28. https://doi.org/10.3844/ajas.2009.523.528. Wangrawa, D. W., Kosgei, J., Machani, M., Opala, J., Agumba, S., Yaméogo, F., ... & Ochomo, E. (2022a). Larvicidal Activities and Synergistic Effects of Essential Oils against Anopheles funestus and Culex quinquefasciatus (Diptera: Culicidae) from Kisumu, Kenya. Psyche: A Journal of Entomology, 2022. https://doi.org/10.1155/2022/8302696. Wangrawa, D. W., Ochomo, E., Upshur, F., Zanré, N., Borovsky, D., Lahondere, C., ... & Sanon, A. (2022b). Essential oils and their binary combinations have synergistic and antagonistic insecticidal properties against Anopheles gambiae sl (Diptera: Culicidae). Biocatalysis and Agricultural Biotechnology, 42, 102347. https://doi.org/10.1016/j.bcab.2022.102347. Wangrawa, D. W., Badolo, A., Ilboudo, Z., Guelbéogo, W. M., Kiendrébeogo, M., Nébié, R., Sagnon, N., & Sanon, A. (2018). Insecticidal Activity of Local Plants Essential Oils Against Laboratory and Field Strains of Anopheles gambiae s. l. (Diptera: Culicidae) From Burkina Faso. Journal of economic entomology, 111(6):
2844-2853. Wangrawa, D. W., Badolo, A., Guenne, S., & Sanon, A. (2016). Larvicidal and oviposition-deterrence activities of four local plant extracts from Burkina Faso against Anopheles gambiae S. l. (Diptera: Culicidae). Int J Mosq Res, 3(6): 11-19. Thèse de Doctorat unique - 144 - Wangrawa, D. W., Badolo, A., Guelbéogo, W. M., Kiendrébeogo, M., Nébié, R. C. H., Sagnon, N., and Sanon, A. (2015). Biological activities of four essential oils against Anopheles gambiae in Burkina Faso and their in vitro inhibition of acetylcholinesterase. International Journal of Biological and Chemical Sciences 9 (2): 793. https://doi.org/10.4314/ijbcs.v9i2.19. Weill, W., Lutfalla, G., Mogensen, K., Chandre, F., Berthomieu, A., Berticat, C., ... & Raymond, M. (2003). Corrigendum: Insecticide resistance in mosquito vectors. Nature, 425, 366. Weill, M., Malcolm, C., Chandre, F., Mogensen, K., Berthomieu, A., Marquine, M., & Raymond, M. (2004). The unique mutation in ace-1 giving high insecticide resistance is easily detectable in mosquito vectors. Insect molecular biology, 13(1): 1-7. White, G. (2007) Terminology of Insect Repellents. In: Debboun M, Frances S, Strickman D, Editors. Insect Repellents: Principles, Methods, and Uses. CRC Press. 31-46. WHOPES (2016) `WHOPES-recommended compounds and formulations for control of mosquito larvae'. Geneva: World Health Organization, Updated 18 March 2016, p. 1. Yaméogo, F., Wangrawa, D. W., Sombié, A., Sanon, A., and Badolo, A. (2021). Insecticidal activity of essential oils from six aromatic plants against Aedes Aegypti , dengue vector from two localities of Ouagadougou , Burkina Faso.» Arthropod-Plant Interactions 15 (4): 627- 34. https://doi.org/10.1007/s11829-021-09842-4. Zaim, M., Aitio, A., & Nakashima, N. (2000). Safety of pyrethroid-treated mosquito nets. Medical and veterinary entomology, 14(1): 1-5. https://doi.org/10.1046/j.1365-2915.2000.00211.x . Zoubiri, S., and Baaliouamer A. (2014). Potentiality of plants as source of insecticide principles. Journal of Saudi Chemical Society 18 (6): 925-38. Zulfikar, Aditama, W., and Sitepu F. Y. (2019). The Effect of Lemongrass (Cymbopogon Nardus) Extract as Insecticide against Aedes aegypti. International Journal of Mosquito Research 6 (1): 101-3. Thèse de Doctorat unique - 145 - |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||