|

UNIVERSITE D'ABOMEY-CALAVI
*=*=*=*=*=*=*=*
FACULTE DES SCIENCES AGRONOMIQUES
*=*=*=*=*=*=*=*=*
DEPARTEMENT DE PRODUCTION ANIMALE
Gestion et conservation de quatre
espèces
de ligneux fourragers (Khaya
senegalensis
A. Juss., Afzelia africana Smith ex
Pers.,
Pterocarpus erinaceus Poir. et
Daniellia
oliveri Hutch. et Dalz.) des terres
de
parcours en zone soudanienne au Bénin
:
cas des communes de Banikoara, Kandi,
Malanville et Karimama
THESE
Pour l'obtention du diplôme d'Ingénieur
Agronome
Option : Sciences et Techniques de Production
Animale
Présentée et soutenue par : Alfred
HOUNGNON Le 19 Décembre 2008
Superviseur : Co-Superviseur :
Prof. Dr. Ir. Brice SINSIN Dr. Ir. Marcel
HOUINATO
Professeur titulaire d'Ecologie Tropicale Maître
Assistant des universités du CAMES
Membres du Jury
Dr ADANDEDJAN Claude, Président
Dr HOUINATO Marcel, Rapporteur
Dr KINDOMIHOU Valentin, Examinateur
Dr ADJOLOHOUN Sébastien, Examinateur
|
UNIVERSITY OF ABOMEY-CALAVI
|
|
|
*=*=*=*=*=*=*=*
|
|
|
FACULTY OF AGRONOMIC SCIENCES
|
*=*=*=*=*=*=*=*=*
DEPARTMENT OF ANIMAL PRODUCTION

Management and conservation of four woody
fodders species (Khaya senegalensis A.
Juss.,
Afzelia africana Smith ex Pers.,
Pterocarpus
erinaceus Poir. et Daniellia oliveri
Hutch. et
Dalz.) in Benin sudanian rangeland:
case of Banikoara, Kandi, Malanville and
Karimama townships
THESIS
Submitted to obtain the degree of Ingénieur
Agronome
Option: Animal Production
Presented and defended by
Alfred HOUNGNON
December, the19th 2008
Supervisor: Co-Supervisor:
Prof Brice SINSIN Dr Marcel HOUINATO
Senior lecturer of Tropical Ecology Lecturer of CAMES
universities
Members of Jury
Dr ADANDEDJAN Claude, Chairman Dr HOUINATO Marcel,
Reporter Dr KINDOMIHOU Valentin, Examiner Dr ADJOLOHOUN Sébastien,
Examiner
CERTIFICATION
Je certifie que ce travail a été
entièrement conduit et dirigé par HOUNGNON Alfred
du Département des Sciences et Techniques de Production Animale
(STPA), Faculté des Sciences Agronomiques (FSA) de l'Université
d'Abomey - Calavi (UAC)
i
Le Directeur de thèse :
Dr. Ir. Marcel HOUINATO
Maître Assistant (CAMES)
Chef Département de Production Animale (FSA - UAC)
ii
DEDICACES
Aux sans soutien,
A tous ceux qui ont franchi le rubicon, sans
gîtes et sans couvert dans une course folle vers la
connaissance,
A tous ceux qui aspirent encore au savoir
malgré les douloureuses et humiliantes expériences de
l'école,
A tous ceux qui croient encore au
développement en Afrique au Sud du Sahara malgré la
rigidité des systèmes.
iii
REMERCIEMENTS
A l'issue des présentes recherches, je me dois de
témoigner ma gratitude aux :
> Dr. Ir. Marcel HOUINATO mon superviseur, chef du
Département de Production Animale à la Faculté des
Sciences Agronomiques de l'Université d'Abomey - Calavi. Qu'il veuille
bien trouver ici l'assurance de mes sentiments dévoués et de ma
haute considération.
> Professeur Brice SINSIN, responsable du Laboratoire
d'Ecologie Appliquée (LEA) pour m'avoir accueilli dans son laboratoire
et, à ses collaborateurs notamment monsieur Sylvain GBOHAYIDA du LEA
pour m'avoir accordé toutes les facilités.
J'exprime également ma reconnaissance aux:
> Ingénieur agronome Méryas KOUTON pour son
dévouement lors du traitement de mes données;
> Dr Ismaël TOKO pour sa fructueuse collaboration;
> Ingénieur agronome Thierry D. HOUEHANOU pour ses
orientations avant le démarrage de mes travaux de recherche, ses
conseils et pour la lecture attentive et éclairée du document
malgré ses multiples préoccupation;
> Ingénieur des travaux Christian ADJAHOSSOU pour son
soutien
Je remercie aussi le Dr Pierre DOSSOU-YOVO, pour m'avoir
aidé à me réinscrire à la Faculté des
Sciences Agronomiques de l'Université d'Abomey-Calavi et tous ceux qui
m'ont soutenu, aidé en prenant à coeur mes problèmes
malgré les leurs pesant.
Dans cet ordre d'idée, je dois remercier Mr Chakirou
LAWANI pour m'avoir chaque fois écouté et conseillé
pendant les dures épreuves; monsieur François LABE pour m'avoir
offert l'opportunité de passer le baccalauréat dans les
meilleures conditions; mes tutrices Hélène TOSSOU HOUNNOU et
Mariam LAWANI.
Je remercie Corinne LABE pour m'avoir toujours soutenu,
Félicité AGUEGUE pour m'avoir encore une fois donné raison
de croire à nouveau en moi au cours de ces deux dernières
années d'étude à Abomey - Calavi. Veuillez trouver ici
l'expression de ma profonde gratitude.
Toute ma gratitude va à Madame Cécile NAGNICHEDE
pour dire combien je demeure encore très sensible à sa protection
et aux sacrifices qu'elle a consentis en son temps.
Je remercie aussi Houeffa V. Ghislaine NAGNICHEDE, Laurencia,
Diane, Romaric et Pépé pour leur soutien, de même que
Lucien HOUNGNON, Daniel BOCOSSA, Jean - Claude HOUNGNON, et Valérie
KOUADIO, pour leur attention et enfin Moukaram LAWANI, Serges Kamal Deen HABIB,
Hugues LABE et Anicet AÏNA pour leur amitié.
Mes remerciements vont aussi à tous les membres du Jury
qui ont apporté des critiques à l'amélioration de la
qualité de ce travail ; je veux nommer Dr. ADANDEDJAN Claude
(Président), Dr. HOUINATO Marcel (Rapporteur), Drs ADJOLOHOUN
Sébastien et KINDOMIHOU Valentin (examinateurs).
iv
RESUME
Le plus grands défis de la recherche scientifique en
Afrique subsaharienne est la gestion durable et la conservation efficace des
ressources naturelles. Cette étude aborde la gestion et la conservation
des ligneux fourragers dans les terres de parcours communautaire entre les
parallèles 11°20' et 12°50' de latitude Nord et les
méridiens 2°17 et 3°05 de longitude Est au Bénin.
L'objectif est de déterminer les paramètres
dendrométriques et caractériser les populations de Khaya
senegalensis, Afzelia africana, Pterocarpus erinaceus et Daniellia
oliveri en raison de leur intérêt fourrager. Les
potentialités et pratiques d'exploitation pastorale des quatre
espèces ligneuses fourragères ont été
évaluées à partir de relevé de la
végétation, d'observations des indicateurs qui rendent compte des
mutilations et du suivi phénologique. Pour atteindre cet objectif, 60
relevés ont été faits sur des aires de 900 m2
chacun dans des peuplements des quatre espèces ligneuses. Une DCA sur
cette matrice a permis de constituer quatre groupements végétaux
:
- Le groupement à K. senegalensis et
Mimosa pigra;
- Le groupement à P. erinaceus et
Anogeissus leiocarpus;
- Le groupement à A. africana et Balanites
aegyptiaca;
- Le groupement à D. oliveri et Combretum
ghasalense.
Les caractéristiques structurales de ces groupements
dans les formations naturelles montrent que la richesse spécifique des
ligneux varie de 34 à 81 espèces. La densité des arbres
(pour un diamètre à hauteur de poitrine, dbh >10 cm) varie de
66,67 à 93,44 pieds/ha et la surface terrière de 10,71 à
25,87 m2/ha. La densité de la
régénération et la probabilité de reconstitution
des peuplements sont généralement faibles. L'analyse de
l'abondance et de la qualité de la régénération
pour une reconstitution des quatre espèces montre que la survie de la
dynamique forestière est hypothéquée. Les données
sur l'ébranchage, l'étêtage, l'écorçage et la
phénologie ont été collectées et les
résultats révèlent que 98% de K. senegalensis,
91% de P. erinaceus et 86% de A. africana dans les
populations étudiées, sont mutilés alors que les
éléments appétibles à savoir feuilles et fleurs
sont disponibles pendant la période d'étude. Ces trois
espèces sont menacées par une surexploitation pastorale ; les
sujets adultes sont sévèrement émondés et les
jeunes sont généralement affectés par le broutage direct.
Toutefois, la répartition phytogéographique des espèces
montre que le milieu conserve son caractère naturel. La gestion durable
et la conservation des ligneux fourragers dans cette éco région
requiert donc la conservation in situ ou la domestication par la
promotion des jardins botaniques communautaires ; l'agroforesterie par
l'enrichissement des peuplements et leur suivi ainsi que la mise en oeuvre des
mesures restrictives concernant l'exploitation de certaines espèces
telles que K. senegalensis, A. africana, P. erinaceus et D.
oliveri.
Mots clés : Gestion durable,
conservation, ligneux fourragers, zone soudanienne, Bénin.
v
ABSTRACT
The biggest challenge of scientific research in sub saharian
Africa is the sustainable management and effective conservation of natural
resources. This dissertation accesses the management and the conservation of
woody fodders in communities range land located between 11°20 - 12°50
N and 2°17 - 3°05 E. The objective is to determine dendrometrical
parameters and characterise Khaya senegalensis, Afzelia africana,
Pterocarpus erinaceus and Daniellia oliveri populations owing to
their fodder prized by the livestock. The potentialities and practical of
pastoral exploitation of the four species were evaluated starting from
statement of the vegetation, threat sign observations and of follow-up
phonologic. To achieve this objective, 60 statement of vegetation have been
settled on 900 m2 area of each one size in the four woody species
populations. A Detrented Correspondence Analysis (DCA) on this matrix permitted
to identify four plants communities:
- The group of K. senegalensis and Mimosa
pigra;
- The group of P. erinaceus and Anogeissus
leiocarpus;
- The group of A. africana and Balanites
aegyptiaca ;
- The group of D. oliveri and Combretum
ghasalense.
The structural characteristics of the plant communities show
that species richness among woody plants varies from 34 to 81 species. Tree
density (dbh > 10 cm) varies from 66.67 to 93.44 stems/ha and basal area
from 10.71 to 25.87 m2/ha. Regeneration density and probability of
stand reconstitution are generally low. An analysis of the abundance and
quality of regeneration of the four species shows that the survival of forest
dynamics is mortgaged. Data on foliage pruning, and bark high harvests have
been collected and the results show that 98% of K. senegalensis, 91%
of P. erinaceus and 86% of A. africana in populations
studied, are debarked while appetible elements as leaves and flowers are
available during field study. These three species are threatened by a pastoral
overexploitation; the adult subjects are severely pruned and the young people
generally affected by direct chattering. However, the phytogeographical types
show that the area retains its natural character. Sustainable management and
conservation of woody fodders in this ecological region thus requires the
conservation in situ or the domestication by local botanical gardens
promoting; the agroforestory by enrichment planting and their monitoring as
well as measures to restrict felling of certain species such as K.
senegalensis, A. africana, P. erinaceus and D. oliveri.
Keywords: Sustainable management, conservation,
woody forages, sudanian area, Benin.
vi
LISTE DES SIGLES ET ABREVIATIONS
ASECNA : Agence pour la Sécurité
de la Navigation Aérienne en Afrique et au Madagascar
CENAGREF : Centre National de Gestion des
Réserves de Faune
Dbh: Diameter at breast height
ECOPAS : Ecosystèmes
Protégés en Afrique Soudano-Sahélienne
ETP: Evapo-transpiration Potentielle
FAO : Organisation des Nations Unies pour
l'Agricullture et l'Alimentation
FSA : Faculté des Sciences
Agronomiques
LEA : Laboratoire d'Ecologie Appliquée
MEPN : Ministère de l'Environnement et de
la Protection de la Nature
RBP : Réserve de Biosphère de la
Pendjari
RGPH : Recensement Général de la
Population et de l'Habitat
RTB W : Réserve Transfrontalière
de Biosphère du W
TP : Types phytogéographiques
UAC : Université d'Abomey Calavi
UICN : Union Internationale pour la Conservation
de la Nature
vii
LISTE DES FIGURES
Figure 01: Localisation de la zone d'étude 4
Figure 02: Diagramme climatique de Kandi (1978-2007) .
5
Figure 03: Variations mensuelles moyennes de l'insolation et de
l'humidité relative (HR)
à Kandi (1978-2007) . 7
Figure 04: Cours d'eau dans la
Réserve de Biosphère Transfrontalière du W (Zone
soudanienne du Bénin) 9
Figure 05: Localisation des placeaux dans le milieu
d'étude 21
Figure 06: Répartition des groupements
végétaux dans le plan des axes 1 et 2 de la
DCA 28
Figure 07: Dendrogramme des groupements végétaux
29
Figure 08: Distribution par classe de diamètre des individus du
groupement à Daniellia
oliveri et Combretum ghasalense (Groupement
G1) 30
Figure 09: Distribution par classe de diamètre de
Daniellia oliveri au sein du groupement
G1 31
Figure 10: Distribution par classe de hauteur de
Daniellia oliveri au sein du groupement
G1 31
Figure 11: Spectres phytogéographiques bruts
du groupement à Daniellia oliveri et
Mimosa pigra (Groupement G1) .. 32
Figure 12:
Distribution par classes de diamètre des individus du groupement
à Khaya
senegalensis et Mimosa pigra (Groupement G2) .
33
Figure 13: Distribution par classes de diamètre de Khaya
senegalensis au sein du
groupement G2 . 33
Figure 14: Distribution par classe des
hauteurs de Khaya senegalensis au sein du
groupement G2 34
Figure 15: Spectres phytogéographiques bruts du groupement
G2 34
Figure 16: Distribution par classes de diamètres des individus
du groupement à
Pterocarpus erinaceus et Anogeissus leiocarpa
(Groupement G3) .... 35
Figure 17: Distribution par classe de
diamètre de Pterocarpus au sein du groupement
G3 . 36
Figure 18: Distribution par classe des hauteurs de
Pterocarpus au sein du groupement
G3 .. 36
Figure 19: Spectres phytogéographiques bruts du groupement
G3 37
Figure 20: Distribution par classes de diamètres des
individus du groupement Afzelia
africana et Balanites aegyptiaca (Groupement
G4) 38
Figure 21: Distribution par classe de diamètre de Afzelia
africana au sein du groupement
G4 .. 38
Figure 22: Distribution par classe des hauteurs de Afzelia
africana .. 39
Figure 23: Spectres phytogéographiques bruts du groupement
G4 ... 39
Figure 24: Pourcentage des individus de K. senegalensis ayant
subi des mutilations ... 40
Figure 25: Pourcentage des individus de D. oliveri ayant
subi des mutilations 40
Figure 26: Pourcentage des individus de P. erinaceus
ayant subi des mutilations 41
Figure 27: Pourcentage des individus de A. africana
ayant subi des mutilations . 41
viii
LISTE DES TABLEAUX
Tableau 1: Valeurs propres et pourcentage de variance
expliqué par les 4 premiers axes. 26
Tableau 2 : Résultat des observations phénologiques
43
ix
LISTE DES ANNEXES
Annexe1: Liste des photos des espèces ligneuses
fourragères étudiées 62
Annexe 2: Liste des espèces végétales des
terres de parcours communautaires de la Zone
soudanienne (Bénin) 66
Annexe 3: Liste des
espèces du groupement à K. senegalensis et M. pigra
(Forêt galerie)...... 68
Annexe 4: Liste des espèces du groupement à D.
oliveri et C. ghasalense 69
Annexe 5: Liste des espèces rencontrées dans le
groupement à P. erinaceus et A. leiocarpa... 71
Annexe 6 Liste des espèces rencontrées dans le
groupement à A. africana et B. aegyptiaca 73
Annexe 7: Quelques espèces de plantes domestiquées
dans les terres de parcours 75
Annexe 8: Tableau Phénologique des quatre ligneux
fourragers 76
TABLES DES MATIERES
CERTIFICATION
i
x
DEDICACES ii
REMERCIEMENTS iii
RESUME iv
ABSTRACT v
LISTE DES SIGLES ET ABREVIATIONS vi
LISTE DES FIGURES vii
LISTE DES TABLEAUX viii
LISTE DES ANNEXES ix
TABLES DES MATIERES x
1. INTRODUCTION GENERALE 1
1. 1 Contexte et justification 1
1.2 Objectifs de recherche 2
1.3 Hypothèses de recherche 3
2. MILIEU D'ETUDE 4
2.1 Situation géographique 4
2.2 Milieu physique 5
2.2.1 Facteurs climatiques 5
2.2.2 Précipitations, températures et
évapotranspiration potentielle 5
2.2.3 Vents 6
2.2.4 Insolation et humidité relative 6
2.3 Types de sols 7
2.4 Réseau hydrographique 8
2.5 Végétation 10
2.5.1 Les savanes soudaniennes 10
2.5.2 Les forêts galeries 11
2.5.3 Les forêts claires 11
2.5.4 Les marécages 11
2.6 Faune 12
2.7 Milieu humain 13
2. 8 Activités économiques et culturelles
13
2.8.1 L'élevage 13
2.8.2 L'agriculture, l'exploitation des produits
forestiers 14
3. CARACTERISTIQUES GENERALES DES ESPECES ETUDIEES 15
3.1 Daniellia oliveri (Rolf) Huch et Dalz. 15
3.1.1 Taxonomie 15
3.1.2 Biologie 15
3.1.3 Distribution géographique 15
3.1.4 Usages 15
3.2 Pterocarpus erinaceus Poir. 16
3.2.1 Taxonomie 16
3.2.2 Biologie 16
3.2.3 Distribution géographique et
phénologie 16
3.2.4 Usages 16
3.3 Afzelia africana Smith ex Pers. 17
3.3.1 Taxonomie 17
3.3.2 Biologie 17
3.3.3 Distribution géographique 17
xi
3.3.4 Phénologie 18
3.3.5 Usages ethno médicinaux 18
3.4 Khaya senegalensis (Desv.) A. Juss. 18
3.4.1 Taxonomie 18
3.4.2 Biologie 18
3.4.3 Distribution géographique 19
3.4.4 Usages 19
4. MATERIELS ET METHODES 20
4.1 Matériels 20
4.2 Méthodes 20
4.2.1 Collecte des données 20
4.2.2 Traitement et analyse des données 22
5. RESULTATS 26
5.1. Analyse globale de la végétation
26
5.2 Caractéristiques dendrométriques
30
5.2.1 Caractéristiques du groupement à
Daniellia oliveri et Combretum ghasalense (G1) 30
5.2.2 Caractéristiques du groupement Khaya
senegalensis et Mimosa pigra (G2) 32
5.2.3 Caractéristiques du groupement P. erinaceus
et Anogeissus leiocarpus (G3) 35
5.2.4 Caractéristiques du groupement Afzelia
africana et Balanites aegyptiaca (G4) 37
5.3 Exploitations pouvant compromettre la
viabilité des espèces 40
5.3.1 Khaya senegalensis 40
5.3.2 Daniellia oliveri 40
5.3.3 Pterocarpus erinaceus 41
5.3.4 Afzelia africana 41
5.4 Phénologie des quatre espèces
ciblées 42
5.4.1 Khaya senegalensis 42
5.4.2 Daniellia oliveri 42
5.4.3 Pterocarpus erinaceus 43
5.4.4 Afzelia africana 43
5.4.5 Comparaison des observations phénologiques
entre espèces 43
6. DISCUSSION 44
6.1 Facteurs du milieu et répartition des
groupements végétaux 44
6.2 Caractéristiques floristiques des
espèces ligneuses fourragères 44
6.2.1 Chorologie 44
6.2.2 Diversité de Shannon et
équitabilité de Piélou 45
6.3 Comparaison des paramètres
dendrométriques 46
6.3.1 Groupement à Daniellia oliveri et Combretum
ghasalense 46
6.3.2 Groupement à Afzelia africana et Balanites
aegyptiaca 47
6.3.3 Groupement à Khaya senegalensis et Mimosa
pigra 47
6.3.4 Groupement à P. erinaceus et A. leiocarpus
48
6.4 Analyse de la stratification 48
6.5 Régénération et reconstitution
des espèces cibles 48
6.6 Phénologie 49
6.7 Analyse des facteurs de Viabilité des
espèces étudiées 49
6.7.1 Prélèvement des organes 51
6.7.2 Les espèces menacées 52
6.7.3 Efficacité des méthodes de
conservation endogène des plantes dans la zone d'étude
52
6 .8 Les possibilités de gestion optimales des
ligneux 53
7. CONCLUSION GENERALE ET PERSPECTIVES 54
REFERENCES BIBLIOGRAPHIQUES 56
1
1. INTRODUCTION GENERALE
1. 1 Contexte et justification
La seconde moitié du 20e siècle est
marquée par une modification impressionnante des
écosystèmes sous une pression anthropique remarquable que dans
n'importe quelle autre période de l'histoire humaine. De ce fait, la
dégradation de l'environnement et de ses ressources naturelles si
préoccupante appelle une gestion rationnelle de la flore et de la faune
en vue de préserver la biodiversité et l'équilibre
écologique des systèmes naturels. En 1992, à l'issue de la
convention de Rio sur la biodiversité, la gestion durable des ressources
naturelles et la conservation de la diversité biologique sont devenues
une préoccupation majeure pour les Etats Parties (Kokou & Sokpon,
2006). Nombre d'espaces forestiers qui étaient auparavant
négligés intéressent désormais les scientifiques et
les organismes de conservation qui ont engagé des actions de sauvegarde
et de gestion durable, en particulier dans les pays en développement
où l'évolution et la viabilité de la diversité
biologique sont encore critiques. Dans cette nouvelle approche, les formations
végétales et ressources menacées en Afrique au Sud du
Sahara sont devenues le lieu d'intérêt particulier et concurrent.
A l'extrême Nord du Bénin, les forêts, savanes, pelouses ou
bosquets constituent pour la plus part, les formations végétales
naturelles pâturées. Souvent utilisés dans les
systèmes agraires, ces espaces naturels appartiennent à des
collectivités de droit public et sont exploités
traditionnellement et conjointement par des propriétaires de
bétail, résidents et transhumants (Agri.site, 1999).
Malheureusement, ils ne font pas partie du réseau national d'Aires
protégées alors qu'il s'avère particulièrement
difficile et en réalité peu probable de conserver la
biodiversité en dehors des Aires protégées. A chaque
année, cette région du Nord-Bénin, représentative
des aires protégées naturelles soudaniennes (Chabi, 1994)
connaît une forte pression agropastorale, surtout en saison sèche
(Houinato et al., 2000). On estime que plus de 100 000 têtes de
gros et petits ruminants transhumants provenant du Sahel, s'ajoutent aux
1.300.000 têtes de bovins autochtones et plus de 2.000.000 de têtes
d'ovins /caprins pour les parcours de saison en zone soudanienne au
Bénin (Tchiwanou 1999). En 2008, ces effectifs sont passés
à 1.905.000 têtes de bovins avec un taux de croît de 2,60 %
et à 2.265.000 têtes d'ovins/ caprins pour un taux de croit de
2,2% (Zoungrana; Adoum et Ajavon, 2008). De Mars à Juin, les
pâturages graminéens très recherchés deviennent
rares dans la zone soudanienne. Après le passage des feux de
végétation annuels de saison sèche, les savanes sont
nettement plus fréquentées en raison des réserves en
fourrage ligneux, principale source d'alimentation des troupeaux en cette
période (Sinsin et al., 2006). L'essentiel du fourrage
consommé par le bétail domestique en saison sèche provient
de l'émondage et de l'étêtage des arbres fourragers
(Houinato, 1996). S'ils subsistent encore dans quelques
écosystèmes pâturés, ils sont transformés en
paille riche en énergie mais présentant une teneur en
protéine assez faible. Les pâturages herbacés, essentiels
pour le bétail, sont régulièrement parcourus par les feux
(Houinato et al., 2001) . Vers la fin de la saison des pluies et en
saison sèche, le fourrage herbacé des savanes perd sa valeur
azotée. Cela réduit davantage non seulement leur
disponibilité mais aussi et surtout leur qualité. A cette
même période, la végétation ligneuse composée
d'arbres et d'arbustes apporte sous forme de fourrage vert un
complément
2
alimentaire intéressant, riche en protéines,
sels minéraux et vitamines indispensable à l'équilibre
alimentaire du bétail (Fall - Touré, 1993 ;
Zoungrana-Kaboré, 1995). Le bétail compte essentiellement sur ces
ligneux fourragers qui constituent aussi un appoint alimentaire
extrêmement important pour relever les teneurs en protéines,
phosphore (P), calcium (Ca) et vitamines A de leurs rations (Le Houérou,
1974 et 1980b, Miranda, 1989). Les arbres et les arbustes des terres de
parcours soudaniens servent de pâturage aérien en saison
sèche, permettant ainsi de pallier le caractère aléatoire,
instable et saisonnier du tapis herbacé (Sinsin et al., 1989 ;
Akpo et al., 1995 ; Brisso et al., 2007). Par ailleurs, les
effets combinés du surpâturage et des feux ont
considérablement accentué l'élagage sévère
des arbres, arbustes et lianes pour l'obtention du fourrage aérien en
Zone soudanienne du Bénin. Certaines espèces comme Khaya
senegalensis, Pterocarpus erinaceus, et Afzelia africana
qui subissent régulièrement cette coupe
répétée de leur frondaison finissent par ne plus
fructifier; ce qui compromet toute possibilité de leur
régénération naturelle. Même les habitats en sont
affectés puisque les grands oiseaux notamment les rapaces et les
charognards y perdent du coup une partie des possibilités de
nidification (Houinato, 1996). Cette forme d'exploitation menace gravement la
conservation des espèces ligneuses fourragères et compromet la
régénération naturelle des écosystèmes
forestiers voire la préservation de la biodiversité.
Malgré les rôles multiples des arbres et arbustes fourragers,
leurs potentiels quantitatif et qualitatif sont encore mal ou peu connus. Il en
résulte que la majorité de nos écosystèmes et de
nos ressources biologiques se trouvent, à un degré ou à un
autre, dégradée; certaines espèces et ressources
génétiques se seraient même éteintes comme en
témoigne la liste rouge de l'IUCN. Par exemple, A. africana,
K. senegalensis et P. erinaceus sont vulnérables alors
qu'elles seraient en danger critique d'extinction au Bénin (UICN, 2008).
Leurs multiples utilisations augmentent le risque de leur disparition puisque
Daniellia oliveri et P. erinaceus dans les savanes
soudaniennes sont de plus en plus menacées de nos jours. Satisfaire nos
besoins et ceux des générations futures est un crédo de la
convention relative à la diversité biologique, qui exige la
planification et l'utilisation de ces ressources pour en assurer
l'équilibre et la pérennité. Il importe, donc d'adopter
des approches et des mécanismes de prévention, d'utilisation
rationnelle et de promouvoir la restauration et la réhabilitation des
ressources détruites, leur valorisation par la mise en oeuvre des
mesures incitatives d'une part et d'autre part, la participation et
l'engagement des communautés. Face à cette situation, il urge de
connaître l'état actuel des populations de ces espèces
ligneuses fourragères et leur diversité spécifique dans
les peuplements ligneux des terres de parcours afin de comprendre, de
prévoir et d'anticiper sur leur gestion et leur conservation. C'est ce
qui justifie l'importance de la présente étude qui s'intitule :
« Gestion et conservation de quelques ligneux fourragers (Khaya
senegalensis, Daniellia oliveri, Pterocarpus erinaceus
et Afzelia africana,) dans les terres de parcours en zone
soudanienne du Bénin.
1.2 Objectifs de recherche
L'objectif principal de cette recherche est d'évaluer
l'importance fourragère et alimentaire des ligneux fourragers dans les
terres de parcours. Les objectifs spécifiques sont :
1.
3
Faire la caractérisation floristique des populations de
chaque espèce ciblée;
2. Déterminer les paramètres
dendrométriques de Khaya senegalensis, Daniellia
oliveri, Pterocarpus erinaceus et Afzelia africana;
3. Caractériser l'exploitation qui compromet la
viabilité de ces espèces;
4. Etudier la phénologie de chaque espèce
ciblée.
1.3 Hypothèses de recherche
1. Les groupements végétaux au sein desquelles
évoluent ces espèces varient d'une formation
végétale à une autre;
2. La distribution par classe de diamètre et le potentiel
de régénération de Khaya senegalensis,
Daniellia oliveri, Pterocarpus erinaceus et Afzelia
africana permettent de prédire leurs tendances futures;
3. Les espèces ciblées sont surexploitées
et menacées de disparition,
4. La phénologie des ligneux fourragers
étudiés présente un intérêt majeur du point
de vue agropastoral.
4
2. MILIEU D'ETUDE
2.1 Situation géographique
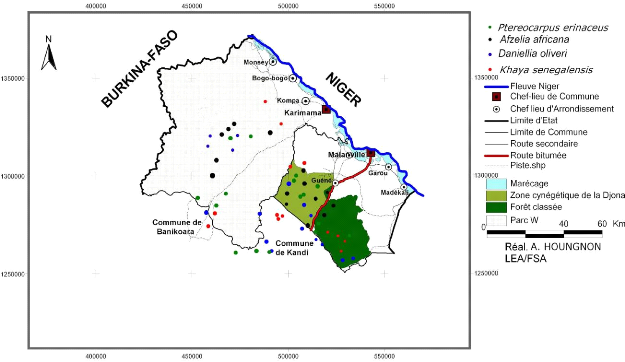
Cette étude a été conduite dans le
Département de l'Alibori au Nord du Bénin. La zone d'étude
est limitée au Nord et au Sud respectivement par les Communes de
Karimama et Banikoara, à l'Est par les Communes de Kandi et Malanville
et à l'Ouest par la rivière Mékrou faisant
frontière avec le Burkina-Faso et le Niger.
Elle renferme le Parc National du W (zone
protégée) s'étendant sur 762 438 ha déjà
classé comme Réserve de Biosphère Transfrontalière
(RBT) du W depuis 2001 et d'une vaste étendue de parcours (zone libre)
considérées comme des pâturages naturels
gérés par les communautés riveraines elles même.
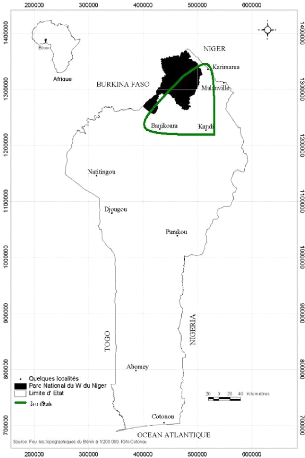
Source : CENAGREF Kandi (2008)
Figure 1 : Localisation de la zone
d'étude.
5
2.2 Milieu physique
2.2.1 Facteurs climatiques
Le climat du milieu d'étude est du type soudanien, bien
que les communes de Banikoara, Malanville et Karimama connaissent des
influences sahéliennes se traduisant par la présence de beaucoup
d'épineux tels que Balanites aegyptiaca, Acacia
gourmaensis, etc (Kpèra, 2008). Le milieu d'étude
présente une saison sèche et une saison pluvieuse qui dure
respectivement de novembre à début-mai et de mai à octobre
marqué par un gradient pluviométrique négatif sud-nord
avec environ 785 mm, 1008 mm et 1015 mm de pluie moyenne annuelle
respectivement à Malanvillle, Banikoara et Kandi (Ibouraïma,
2005).Toutes les données climatiques utilisées proviennent de la
station météorologique de Kandi couvrant la période
normale (1978 - 2007). Ces données montrent que le régime
pluviométrique connaît une grande variation d'année en
année, mais qui ne s'améliore guère puisque le milieu
devient de plus en plus sec. La moyenne annuelle des précipitations est
de 793 mm (1978-2007).
2.2.2 Précipitations, températures et
évapotranspiration potentielle
La répartition des pluies dans l'année permet de
connaître les périodes pendant lesquelles les animaux sont
confrontés aux problèmes de déficit en eau et en fourrage.
La construction sur une même figure des courbes de pluviométrie
(P) et de l'évapotranspiration potentielle (ETP) détermine la
position de deux évènements de nature purement climatique (la
saison sèche et la saison humide). La figure 2 présentant le
diagramme climatique de la région de Kandi permet de scinder ces deux
bioclimatiques successifs..
Précipitation ETP ETP/2
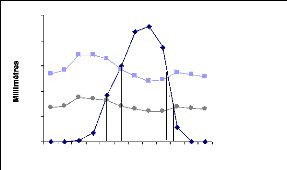
250
200
150
100
50
0
J F M A M J J A S O N D
Mois
Source : ASECNA Kandi (2007)
Figure 2 : Diagramme climatique de Kandi
(1978-2007)
Le diagramme combine la pluie et l'ETP qui exprime la
pluviométrie saisonnière. Il présente une distribution
uni-modale caractéristique de la zone soudanienne avec une saison
sèche et une saison pluvieuse. Le cycle de végétation est
compris entre 180 et 270 jours/an. La moyenne annuelle des
précipitations est de 793 mm. Cette valeur détermine une moyenne
incluse dans la fourchette 700 mm à 1200 mm avancée pour la
même zone par Ibouraïma (2005), après avoir calculé
les pluies moyennes annuelles
6
sur une période de 30 ans (1969-1998) aux stations de
Kandi, Malanvilles et Banikoara.
Ce graphe montre quatre phases importantes. La période
de végétation correspondant aux périodes humide et
subhumide pendant lesquelles la pluviométrie est au moins
supérieure à la moitié de l'évapotranspirati on,
est subdivisée en trois phases. La phase 1 correspond à
l'apparition des premières pluies (mai à juin) où les
hémicryptophytes (graminées vivaces) très
recherchées par les ruminants s'installent. Les graminées
repoussent grâce à la réserve d'eau dans le sol et par
succion racinaire. La phase 2 correspond à la période active de
végétation caractérisée par les mois où la
pluie est au dessus de l'ETP. Cette période pendant laquelle les
hémicryptophytes abondent après la reprise
végétative correspond sur la courbe aux mois de juin, juillet
aout et septembre. Le pic de biomasse s'observe en septembre et marque la fin
de la période humide. Le mois d'août est le mois le plus
arrosé. La pluviosité au cours de ce mois dépasse
généralement 227 mm et la température est de l'ordre de
26°C. A la fin de la période active de végétation, on
note l'apparition des feux de végétation favorisant la repousse
des graminées car, il ya déjà la lignification qui les
rend peu appétibles. Cette phase correspond à la troisième
phase de la période végétative. La phase 1 et la phase 3
sont les phases subhumides comprises entre l'ETP et l'ETP/2. La
quatrième phase correspond à la sécheresse et
s'étend de mi-octobre à mi-mai. Les pluies sont en dessous de la
moitié de l'évapotranspiration. Les mois de novembre,
décembre, janvier et février sont complètement secs.
Pendant cette période, les ressources alimentaires du bétail se
réduisent aux ligneux fourragers et résidus de récolte.
Quant à la température moyenne, la plus basse (24,9°C) est
observée en décembre. La température moyenne la plus
élevée est enregistrée au mois d'avril à
32,2°C.
2.2.3 Vents
Deux types de vents soufflent dans la zone d'étude :
- L'alizé maritime d'avril à novembre suivant
une direction Sud-ouest. Sa vitesse moyenne est de 2 m/s ou de 3m/s. Sa vitesse
maximale oscille entre 23 m/s et 30 m/s suivant les mois. Il porte la pluie
dans la région.
- L'harmattan de décembre à mars provient de
l'anticyclone des Açores au Sud de l'Espagne. Ce vent sec augmente le
déficit hygrométrique de l'air entre décembre et janvier
et accentue encore les conditions d'aridité de la zone en saison
sèche.
2.2.4 Insolation et humidité
relative
L'insolation représente le paramètre essentiel
du rayonnement global et joue à ce titre un rôle très
important à la fin de l'hivernage en intensifiant le pouvoir
évaporant de l'air (Sinsin, 1993).
La figure 3 présente les variations moyennes mensuelles
de l'insolation de la localité de Kandi (1978-2007).
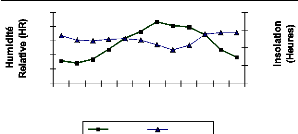
100
40
80
60
20
0
J E M A M J JASON D
HR(%) Insolation
Mois
400
300
200
0
100
7
Source : ASECNA Kandi (2007)
Figure 3 : Variations mensuelles moyennes de l'insolation
et de l'humidité relative (HR) à Kandi (1978-2007)
saton
La durée d'insolation maximale est observée en
novembre (286,13 heures) et celle minimale en août (187,55 heures). Sa
durée annuelle est en moyenne de 3000 heures. La période la plus
ensoleillée va d'octobre à juin et la moins ensoleillée de
juillet à septembre.
L'humidité relative joue le rôle
atténuateur du déficit hydrique. Elle est inversement
proportionnelle à l'insolation et est plus faible pendant la saison
sèche. Les moyennes mensuelles varient de 26% en février à
86% en juillet. De décembre à avril, sa valeur moyenne est
inférieure à 50%, le pouvoir évaporant de l'air est alors
fort.
o s
Heures)
ure
2.3 Types de sols
Le département de l'Alibori présente comme
principaux sols, les sols ferrugineux tropicaux, les sols hydromorphes, les
sols minéraux bruts et les sols peu évolués. Sur le plan
écologique, la répartition de ces sols est la suivante (Viennot,
1978) : On distingue :
? des sols minéraux bruts lithiques, d'origine non
climatique, sur cuirasses;
? des sols peu évolués d'origine non climatique
d'érosion lithique sur quartzite et micaschiste atacoriens, sols modaux
d'apport sur alluvions du fleuve Niger, sols hydromorphes sur matériau
alluvio-colluvial limono-argileux et alluvions argileuses du fleuve Niger;
? des sols à sesquioxyde de fer et de manganèse
;
? des sols peu évolués d'origine non climatique
d'apport hydromorphe
? des sols ferrugineux tropicaux peu lessivés en
argile, lessivés en sesquioxydes de fer et de manganèse sur
matériau kaolinique issu de roche basique.
8
Dans la zone d'étude, les principaux types de sols sont
les sols ferrugineux sur le socle cristallin, les sols alluviaux dans
l'extrême Nord et les sols ferrugineux tropicaux dans le Sud du
département (Ibouraïma, 2005).
2.4 Réseau hydrographique
La Région nord soudanienne est parcourue par de
nombreux cours d'eau (figure 4) dont le fleuve Niger, les confluents des
grandes rivières Alibori et Mékrou. Les rivières Alibori
et Mékrou drainent entièrement cette région et le Parc
National du W.
A ces deux importantes rivières, s'ajoutent leurs
affluents dont les principaux sont : Kpako, Kompagarou, Bédarou, Djiga
et Konékoga qui sont tous des intermittents. Ils reçoivent de
part et d'autre des cours d'eau secondaires.
Les formations rocheuses de la région, disposées
en longues bandes de direction NNE - SSO, sont traversées en deux
endroits par la Mékrou donnant naissance aux chutes de Koudou et
à une série de rapides et des passages étroits dont la
gorge de la Mékrou est la plus représentative.
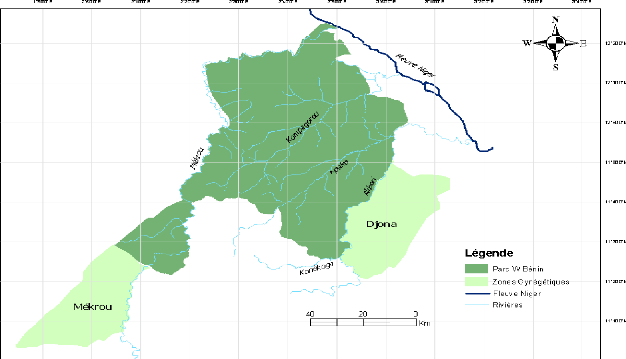
9
Source : CENAGREF Kandi (2007)
Figure 4 : Cours d'eau dans la Réserve de
Biosphère Transfrontalière du W (Zone soudanienne du
Bénin)
10
2.5 Végétation
La végétation de la zone d'étude comprend
quatre écosystèmes : les savanes, les forêts galeries, les
forêts claires et les marécages (Houngnon, 1996).
2.5.1 Les savanes soudaniennes
Les formations savanicoles couvrent la majeure partie de la
zone. Elles occupent près de 90.000 ha de la superficie totale de la
zone cynégétique de la Djona (118.000 ha). Elles sont
composées de savanes boisées, arborées, arbustives et
herbeuses.
-Savane boisée : Elle est
constituée d'arbres de recouvrement important (35 à 60%) mais
dont le couvert n'est pas continu. Leur hauteur est de 8 à 16 m. La
strate herbacée est abondante. Les espèces fréquemment
rencontrées sont : Daniellia oliveri, Anogeissus leiocarpa,
Terminalia spp, Vitellaria paradoxa, Isoberlinia tomentosa, Afzelia
africana.
-Savane arborée : Le couvert des
arbres est de 5 à 35 % ; leur hauteur est généralement de
5 à 10 m, mais peut atteindre 15 m. Les arbustes sont plus abondants
avec une couverture de 5 à 60 %. Dans la strate arborée, on y
rencontre les espèces ligneuses Vitellaria paradoxa, Adansonia
digitata, Diospyros mespiliformis, Burkea africana, Sterculia setigera, Vitex
doniana, Tamarindus indica; en peuplements clairsemés.
Acacia polyacantha et Terminalia glaucescens
occupent les sols relativement frais en formant de petits peuplements.
Acacia sieberiana s'installe tantôt sur les terres basses
tantôt sur les terres exondées, en petits peuplements très
souvent fréquentés par les éléphants.
Pseudocedrala kotschyi colonise les bas-fonds
où il se développe par drageonnage. Il est dispersé sur
les terres exondées.
Mitragyna inermis et Terminalia macroptera
préfèrent les bas-fonds tout en fuyant les forêts
galeries. Khaya senegalensis, Borassus aethiopum, Ostrioderris stuhlmannii,
Lophira lanceolata, Afzelia africana, sont disséminés mais,
Borassus aethiopum malgré sa dispersion par les
éléphants et les singes, se fait de plus en plus rare. Ce fait
serait - il dû soit à sa surexploitation dans un passé
récent ou à la modification de l'habitat ou du régime
alimentaire de ces animaux? Afzelia africana et Khaya senegalensis
sont très recherchés par les éleveurs pour leur
bétail si bien qu'elles sont toujours émondées.
-Savane arbustive : Le couvert des arbres
n'excède pas 5 % ; celui des arbustes est de 5 à 60 %. Les
arbustes sont de petites tailles (3 à 5 m). Les espèces les plus
rencontrées sont : Crossopteryx febrifuga, Combretum hypopilinum,
Combretum micranthum, Combretum glutinosum, Combretum nigricans, Combretum
ghasalense, Pteleopsis suberosa, Balanites aegyptiaca, Piliostigma reticulatum,
Piliostigma thonningii, Bridelia ferruginea, Sarcocephallus latifolius, Feretia
apodanthera, Gardenia ternifolia, Acacia seyal, Acacia macrostachya, Acacia
macrothyrsa, Acacia gourmaensis, Dichrostachys cinerea, Strychnos spinosa,
Stereospermum kunthianum, Ziziphus mauritiana, Sclerocarya birrea, Guiera
senegalensis, Ficus platyphylla,
11
Ficus gnaphalocarpa, Prosopis africana, Terminalia
avicennioides, Pericopsis laxiflora, Lannea acida, Lannea kerstingii,
etc.
Combretum glutinosum, Guiera senegalensis sont
souvent en peuplements. Les individus de Combretum micranthum et de
Feretia apodanthera se déveleppent également sur les
termitières dans cette savane. Les individus de Gardenia sont
disséminés à travers la savane.
-Savane herbeuse : Elle est essentiellement
composée de tapis graminéen. Les espèces dominantes sont :
Andropogon gayanus, Andropogon pseudapricus, Andropogon chinensis,
Hyparrhenia involucrata, etc. (graminées) et Aframomum
spp, Spermacoce scabra, Spermacoce stachydea,
Murdania simplex, Barleria bornuense. Ces herbacées atteignent
leur croissance maximale entre mai et septembre.
2.5.2 Les forêts galeries
Elles ont une composition floristique élevée et
présentant physionomiquement l'aspect de forêt ombrophile, avec
des essences atteignant 30 m de hauteur (Toko, 2008). L'Alibori et ses
affluents forment l'essentiel des cours d'eau. On n'y rencontre des arbres et
arbustes plus denses sur les rives des rivières. Les espèces les
plus rencontrées sont Khaya senegalensis, Diospyros
mespiliformis, Kigelia africana, Pterocarpus santalinoides,
Lecaniodiscus cupanioides etc. Combretum micranthum,
à l'allure lianescente forme par endroits des peuplements à
feuilles sempervirentes alors qu'en savane, même sur les
termitières, les feuilles jaunissent dès Octobre. Les arbres et
arbustes des berges des rivières tels que : Cola, Saba, Albizia
glaberrima, etc. forment les forêts ripicoles. Les lianes ligneuses
et certaines plantes sarmenteuses comme Saba senegalensis,
Lonchocarpus cyanescens, montent sur les arbres tandis que Mimosa
pigra protège les berges aux endroits dégradés.
2.5.3 Les forêts claires
C'est un peuplement ouvert avec des arbres de petites et
moyennes tailles (8 à 16 m) dont les cimes sont plus ou moins jointives
(80 % du couvert). On y rencontre des espèces comme Anogeissus
leiocarpa, Daniellia oliveri, Lannea microcarpum, Terminalia spp,
Combretum spp, Tamarindus indica, Isoberlinia doka etc.
La présence des termitières est souvent
remarquée au pied de Tamarindus indica. Les forêts
claires sont très peu représentées dans la zone la zone
d'étude. Souvent, c'est le prolongement des galeries de la berge vers la
savane. Toutefois, dans l'arboretum de la zone cynégétique de la
Djona, on retrouve une tâche de forêt claire dans les
éboulis de latérite, qui serait le vestige d'une ancienne
forêt dense sèche. On y observe : Horrhena
floribunda, Combretum nigricans, Strychnos innocua. Dans
la forêt claire issue de la galerie, on note : Celtis senegalensis,
Malacantha alnifolia, Combretum nigricans, Dombeya quinqueseta. Quelques
herbacées telles que Pennisetum unisetum, Andropogon tectorum,
Rottboellia exaltata, Indigofera dendroides, Desmodium velutinum,
etc. se contentent de la lumière
diffuse des clairières.
2.5.4 Les marécages
Ils sont disséminés dans la savane et sont
constitués par des dépressions plus ou moins profondes qui
viennent rompre la monotonie du paysage. On note certaines espèces
12
qui préfèrent ces zones humides : Mitragyna
inermis, Pseudocedrela kotschyi, Mimosa pigra, Sesbania spp, Vetiveria
nigritana etc.
2.6 Faune
La faune de la région nord soudanienne du Bénin
est essentiellement représentée par celle de la Réserve de
Biosphère Transfrontalière du W (RTB-W) Bénin où
l'on rencontre presque toutes les espèces de grands mammifères de
la savane soudanienne de l'Afrique de l'Ouest à l'exception des girafes,
des élans de Derby et des gazelles rufifrons
(Szaniawsky, 1982). Lamarque (2004),
signale que 51 espèces de grands mammifères ont été
identifiées dans le WAP. Les mammifères les plus courants sont :
Syncerus caffer (Buffle), Panthera leo (Lion),
Loxodonta africana (Eléphant), Kobus
defassa (Cob defassa), Phacochoerus aethiopicus
(Phacochère), Papio anubis (Babouin doguera),
Erythrocebus patas (Singe rouge), Tragelaphus scriptus (Guib
harnaché). Le Lycaon (Lycaon pictus) a été
signalé « en danger critique d'extinction par le CENAGREF selon une
étude réalisée par Di Silvestre et
al.(2003).Toutefois, Di Sylvestre (2004) note que toutes les
espèces de grands carnivores typiques de la savane soudanienne se
trouvent encore dans le Complexe du W. »
La faune aviaire compte 221 espèces selon une
étude dans la RBT-W (Bénin) initiée par le CENAGREF et
réalisée par Adjakpa (2003). Les oiseaux les plus courants sont :
Pintade, Francolin (Francolinus sp),
Charognards, Macrodipterys longipennis (Engoulevant
à balanciers), Crapimulgus olimacurus (Engoulevant à
longue queue) ; Buphagus africanus (Pique-boeuf à bec jaune)
Adjakpa (2003).
Les reptiles sont assez représentés.
Différentes espèces de serpents, de tortues aquatiques, de
sauriens telles que : Crocodylus niloticus (Crocodile), Varanus
niloticus (Varan) sont rencontrées. Selon les travaux de
Kpéra (2008), deux espèces de crocrodiles (Crocodylus
niloticus et Osteolaemus tetraspis) ont été
observées dans 88% des points d'eau du grand complexe de Réserve
de Biosphère Transfrontalière du W (RBTW). Les plus petits
lézards des familles de Gekkonidae, Agamidae, Scincidae et
Chamaeleunidae sont rencontrés partout dans le Parc.
Les principaux cours d'eau de la zone d'étude sont
poissonneux. Les poissons les plus courants sont : Lates niloticus
(Capitaines), Alestes baremoze, Distichodus rostratus,
Labeo senegalensis, Citharinus citharinus et Hydrocynus
sp (Poissons chats) (Lalèyè et al., 2001)
Une étude récente (Projet 7 ACP/NIR/082
financé par l'UE en 1995 - 96) montre que le lamantin semble être
présent dans tout le fleuve Niger et particulièrement au niveau
des zones riches en bourgou (Echinochloa stagnina), très
appréciée pour ses qualités fourragères. Le
Lamantin d'Afrique est un animal peu connu car extrêmement difficile
à observer, fort peu étudié sur le continent, et, en voie
de disparition. Au Bénin comme au Niger, le lamantin semble être
confronté à deux grandes menaces : le braconnage et la perte de
son habitat, due à l'ensablement et à l'occupation des zones
d'épandage par les aménagements hydro-agricoles. L'état
des connaissances actuelles est trop fragmentaire pour pouvoir proposer des
actions précises en vue de sauvegarder l'espèce. La composante
Niger du Programme comprendra donc une sous-composante chargée des axes
de recherche - action sur le lamantin non seulement au Niger mais aussi au
Bénin où l'espèce est également présente.
13
2.7 Milieu humain
La population des quatre communes concernées par notre
étude est de 452 459 habitants selon le recensement de 2002 (RGPH2).
Celle des communautés riveraines du Parc National du W et de la zone
cynégétique de la Djona est estimée à 254 286
habitants au recensement de 2002 (RGPH3). Ces communautés sont
essentiellement rurales à 91 %. Les religions islamiques surtout,
chrétiennes et animistes, etc. sont celles rencontrées. Les
différentes religions collaborent de façon pacifique dans la
zone. Plusieurs groupes socio-culturels peuplent les villages des terres de
parcours communautaire (zone libre) et les villages riverains du Parc W (zone
protégée). Selon Szaniawsky (1982), il s'agit des Bariba qui
dominent à Banikoara (52 %) et à Kandi (32 %), des Dendi et
apparentés qui dominent à Karimama (64 %) et Malanville (62 %),
des Peuhl qui représentent 18 % des habitants de Karimama, 14 % à
Malanville, 35 % à Banikoara et 30 % à Kandi. On note aussi dans
le milieu la présence des Haoussa, Mokolé, Djerma,
Gourmantché, Tchinga, Fon, Yoruba et Nago.
2. 8 Activités économiques et
culturelles
Les principales activités économiques
pratiquées par les populations des terres de parcours de la zone
soudanienne sont l'agriculture, l'élevage ; toute deux demandeurs
d'espace. Le commerce est peu développé et concerne notamment la
commercialisation des produits agricoles et de quelques articles
importés des pays voisins. A ces activités s'ajoutent l'artisanat
et la chasse. En général, les activités culturales
pratiquées sont de type traditionnel (Adjahossou, 2008).
2.8.1 L'élevage
L'élevage constitue la seconde spéculation
économique dont la pratique demeure aussi traditionnelle. Il est peu
intégré à l'agriculture dans la zone d'étude mais
occupe une place de choix tant au plan social qu'économique dans la vie
des populations riveraines. On rencontre dans toutes les exploitations des
animaux domestiques tels que les bovins, ovins, caprins et volailles. Du point
de vue écologique, c'est l'élevage bovin qui a un impacte
facilement appréciable sur la zone d'étude. Le système
d'élevage de ruminant et plus particulièrement de bovins le plus
largement répandu est un élevage extensif basé sur
l'exploitation directe du pâturage naturel. On note le déplacement
saisonnier des éleveurs et de leurs troupeaux de ruminant (transhumance
nationale et transhumance transfrontalière) à la recherche de
pâturage et d'eau pendant les périodes de pénurie en
ressources naturelles. La transhumance nationale concerne les troupeaux
basés au Bénin dont les propriétaires en saison
sèche les déplacent vers les points d'eau. Quant à la
transhumance transfrontalière, elle est pratiquée par les en
provenance du Sahel qui séjournent durant la saison sèche sur le
territoire béninois. Seuls les Peuhls et les Gando pratiquent
l'agro-élevage, ils obtiennent les meilleurs rendements agricoles. La
typologie des éleveurs de la zone révèle trois grands
groupes à savoir : les sédentaires, les anciens transhumants et
les transhumants de saison sèche. Les races dominantes sont les
zébu sahéliens adaptés à
la marche, les métis stabilisés borgou
issus du croisement zébu et
white Foulani qui ont conservé le
caractère
14
dominant zébu. La zone
d'étude est convoitée surtout pour sa disponibilité
relative en ligneux fourragers. Toutefois, certains cultivateurs sont devenus
également éleveurs de bovins pour des besoins de traction.
2.8.2 L'agriculture, l'exploitation des produits
forestiers
C'est la culture itinérante sur brûlis
basée sur des moyens de production rudimentaire qui est en vogue. Les
principales cultures vivrières sont : le sorgho, le mil, le maïs,
le manioc et l'igname. La principale culture de rente est le coton.
Aujourd'hui, l'évolution de la biodiversité pose de
sérieux problèmes face à la persistance dans l'utilisation
abusive des pesticides. La concentration des résidus de pesticides dans
les eaux et l'occupation anarchique des terres à des fins agricoles
constitue les types de menace les plus destructeurs pour l'homme, les habitats
de la faune et les animaux domestiques.
Divers produits forestiers sont exploités dans les
localités de collecte de données. L'exploitation des produits
forestiers se limite à l'extraction de la lignine sous forme de madrier
et du bois - énergie dans une moindre mesure. Quand aux produits
forestiers non ligneux leurs exploitations se limitent essentiellement au
prélèvement du fourrage, des plantes médicinales et de la
récolte des fruits tropicaux (Karité, Néré, Baobab
etc.)
15
3. CARACTERISTIQUES GENERALES DES ESPECES ETUDIEES
3.1 Daniellia oliveri (Rolf) Huch et Dalz.
3.1.1 Taxonomie
Daniellia oliveri appartient à la famille des
Leguminosae et à la sous famille des Caesalpinoideae. Le genre Daniellia
comporte plusieurs espèces dont Daniellia oliveri (Rolf) Huch
et Dalz., Daniellia thurifera Benn., D. thurifera var.
chevalieri Léonard. Dans la zone d'étude, Daniellia
oliveri (Rolf) Huch et Dalz est connue sous le nom de Falmey en Dendi,
Gnankonrongou en Bariba et Gounmé en Peulh.
3.1.2 Biologie
- Morphologie
Daniellia oliveri est un grand arbre à cime
étalée (cône renversé) et assez dense, de 15
à 20 voire 25 m de haut, souvent bas branchu, à fût droit
blanc gris. Le feuillage vire du rose au rouge au moment de la feuillaison. Son
écorce écailleuse se desquame par plaques plus ou moins
circulaires, grise à tranche rouge foncée striée de blanc
(Arbonnier, 2002).
-Caractéristiques botaniques
Les feuilles : le feuillage est rose ; les
feuilles alternes, paripennées atteignant 45 voire 50 cm de long,
portent, 4 à 11 paires de folioles opposées ou
subopposées.
L'inflorescence : c'est une panicule
axillaire courte d'environ 25 cm ; les fleurs sont blanches ou
blanc-verdâtres devenant crèmes, comprenant 4 sépales et 1
à 2 grands pétales et 10 étamines dépassant les
pétales.
Les fruits : ce sont des gousses plates
ovales à 2 valves papyracées rigides de 5-10 x 2,5-5 cm, beiges.
La graine est brune, ovale et plus ou moins plate, pendante et retenue à
une des valves par un funicule de 12 à 15 mm.
3.1.3 Distribution
géographique
L'espèce Daniellia oliveri a une aire de
répartition qui s'étend du Sénégal au Cameroun,
jusqu'au Soudan, au Congo et en Angola. Souvent grégaire sur tout type
de sol, l'espèce est localement très commune et ne pousse
qu'à découvert avec une distribution
irrégulière.
3.1.4 Usages
L'espèce Daniellia oliveri présente un
vaste domaine d'usages notamment en pharmacopée, dans la fabrication des
objets d'arts, dans la lutte contre l'érosion mécanique et dans
l'agriculture. En ce qui concerne la pharmacopée, l'espèce est
utilisée seule ou en combinaison avec d'autres plantes pour
guérir diverses maladies (Babatoundé, 2003). Ces feuilles sont
entre autres utilisées contre la fièvre, l'ictère et la
toux. Quant à l'écorce, elle est surtout utilisée contre
les maux d'yeux chez les ruminants (Fagnissè, 2006) et les racines sont
utilisées contre la stérilité féminine et la folie
(Arbonnier, 2002).
16
3.2 Pterocarpus erinaceus
Poir.
3.2.1 Taxonomie
De son nom français, Palissandre du
Sénégal, l'espèce appartient à la famille des
Leguminosea et à la sous famille des Papilionioideae. Dans le milieu
d'étude, différents noms sont utilisés pour
désigner l'espèce. Il s'agit de Tonan en Bariba et Banoui en
Peulh.
En fon il est connu sous le nom de Kosso. 3.2.2
Biologie
- Morphologie
C'est un arbre haut de 8 à 12 (- 15) m, de haut
à fût droit et cylindrique atteignant 1 m de diamètre,
à cime arrondie et ouverte. L'écorce est noirâtre,
crevassée et très lamelleuse, à tranche brune
striée de rouge exsudant une résine translucide rougeâtre
qui durcit rapidement avec un rameau densément pubescent devenant
glabre, gris.
- Caractéristiques botaniques
- Les feuilles : sont alternes,
imparipennées, à 3 - 5 (-7) paires de folioles alternes. Folioles
finement pubescentes dessous devenant glabres de forme variable, ovales
à elliptiques, de 6 - 11 * 3,6 cm, à sommet arrondi ou obtus
généralement échancré à base arrondie. Les
nervures sont pennées, peu saillante sur les deux faces.
- L'inflorescence : est une grappe
lâche disposée en bout de branche de 10 - 20 cm de long. Les
fleurs jaune d'or devenant jaune pâle, asymétrique, odorante, sont
de 10 - 12 mm de long à calice pubescent à 5 dents courtes.
- Le fruit est une samare plate
entourée d'une aile circulaire membraneuse et plus ou moins
plissée portant sur les deux faces de la graine, de nombreux poils
épineux, rigides et enchevêtrés, de 4 - 7 cm de
diamètre, jaune paille à maturité, persistant longtemps
sur l'arbre. La floraison et la fructification ont lieu en saison sèche
toujours avant la feuillaison. (Arbonnier, 2002).
3.2.3 Distribution géographique et
phénologie
Pterocarpus erinaceus est présente dans les
formations ouvertes et fermées. En régions
soudano-sahélienne et soudano-guinéenne, on la rencontre en
savanes et dans les forêts sèches, sur tous types de sols y
compris latéritiques. En Afrique, elle s'étend du
Sénégal au Cameroun et à la république
centrafricaine. Elle est assez commune, localement grégaire et
disséminée.
3.2.4 Usages
Les racines de l'espèce Pterocarpus erinaceus
sont surtout utilisées pour le traitement des plaies et de la toux.
La cendre des racines et des feuilles traite les maladies mentales. Aussi
l'écorce est-elle utilisée dans le traitement des plaies, carie
dentaire, toux, ulcère chronique, anémie hématurie,
dysménorrhée et la diarrhée chez les volailles. Les
feuilles et l'écorce sont fébrifuges dans certaines
régions, sous forme de lotions chaudes et d'infusions. La sève
est un vermifuge. Elle traite également la taie, les affections
oculaires, l'impuissance sexuelle et la blennorragie. Les feuilles sont
très
17
appréciées par le bétail. C'est un bois
de coeur brun foncé veiné lourd, durable très dur et bien
utilisé pour le charbon.
3.3 Afzelia africana Smith ex Pers. 3.3.1
Taxonomie
Le genre Afzelia Sm. appartient
à la famille des leguminoseae, sous famille des Ceasalpinioideae. Ce
genre comporte plusieurs espèces dont les plus connues sont Afzelia
africana Smith., Afzelia africana bracteata T. Vogel.,
Afzelia africana bella Harms (Aubreville, 1950). Afzelia africana
est la seule signalée au Bénin. Elle est connue sous le nom
de Lingué (français), African oak, African mahogany (anglais),
Kpakpagidé (fon). Dans le milieu d'étude il est appelé
Hitchinémoïgo en Dendi, Gbéou ou Gbébou en Bariba et
en Peulh.
3.3.2 Biologie - Morphologie
Afzelia africana est un grand arbre de 25 à 30
voire 35 m de haut, à cime dense, large et étalée ; le
fût est droit et le diamètre pouvant franchir les 2 m. C'est un
arbre à branches massives et à écorce grise plus ou moins
foncée, relativement fissurée et lamelleuse, s'écaillant
en plaques, laissant apparaître des zones gris claires, à tranche
rose à brun-claire granuleuse (Ahouangonou & Bris, 1997).
- Caractéristiques botaniques
Les feuilles : le feuillage est vert brillant
; les feuilles alternes, paripennées mesurant jusqu'à 30 cm de
long, portent 3 à 8 paires de folioles opposées ou
subopposées, et luisantes, vert-foncées au dessus, largement
espacées sur les rachis.
L'inflorescence : c'est une panicule
terminale d'environ 20 cm de long ; les fleurs sont blanches, striées et
pourpre, à trois pétales supérieurs elliptiques de 10
à 12 mm de long et un pétale inférieur à deux lobes
arrondis divergents.
- Les fruits : ce sont des gousses aplaties
ligneuses, glabres, de 10 à 18 cm x 6 à 8 cm, épaisses de
2 à 5 cm, à deux valves noires, contenant 7 à 10 graines
noires, lisses plus ou moins brillantes.
3.3.3 Distribution
géographique
Afzelia africana est une espèce africaine
disséminée dans toute l'Afrique tropicale. On la trouve en
savane, dans les forêts humides et forêts sèches et sur les
collines (Akoègninou et al., 2006). L'espèce est
présente en Afrique avec une aire de répartition
s'étendant du Sénégal au Cameroun jusqu'en Ouganda et
Tanzanie. Au Bénin Afzelia africana se rencontre aussi dans
certains boqueteaux de type xérophile ou tropicale sec que dans les
savanes boisées de type guinéen (Aubreville, 1937).
L'espèce est également signalée dans la zone de la Lama,
dans la région de Pobè, dans la région des Monts
Kouffé, dans la zone soudanienne (Adomou, 2005) et dans le bassin du
fleuve Niger. Elle est commune, localement abondante et grégaire
(Arbonnier, 2002).
18
3.3.4 Phénologie
Afzelia africana ne montre pas une feuillaison
permanente. En zone sahélienne, les feuilles connaissent une chute
partielle de novembre à février. En zone nord-soudanienne,
l'intensité de la défeuillaison est plus forte (Cuny &
Sanogo, 1998). La floraison a lieu généralement entre mars et
avril en Afrique occidentale. Les fruits arrivent à maturité vers
décembre ou janvier au Bénin (Ahouangonou & Bris, 1997). La
durée de dissémination des graines est courte et a lieu
généralement en décembre et janvier.
3.3.5 Usages ethno médicinaux
Le décocté de l'écorce de tronc seul ou
associé avec d'autres plantes telles que Berlinia grandiflora,
Bombax costatum et Burkea africana, est administré per os
dans le traitement des oedèmes, épilepsie, névralgies
intercostales, dysménorrhées, convulsions hyperthermiques et du
retard staturo-pondéral (Adjanohoun et al., 1989). Les racines
sont utilisées pour soigner la blennorragie et la trypanosomiase
(Arbonnier, 2002) ; elles sont employées contre l'hypertension, en
mixture avec celle de Annona senegalensis, et pour le traitement des
plaies extérieures, en mixture avec celle du Morinda lucida
(Ahouangonou & Bris, 1997).
3.4 Khaya senegalensis (Desv.) A.
Juss.
3.4.1 Taxonomie
Vulgairement appelé Caïlcédrat ou Acajou du
Sénégal, Khaya senegalensis (Desr.) A. Juss. est l'une
des plus importante espèces de la famille des Meliaceae en Afrique. Dans
la zone d'étude, Khaya senegalensis (Desv.) A. Juss. est connue
sous le nom de Gbira, Gbirou, ou Gbiribou en Bariba, et Kahi en Peulh.
3.4.2 Biologie
- Morphologie
Khaya senegalensis (Desv.) A. Juss est un grand arbre
de 25 à 35 m de haut présentant un fût
généralement court et trapu mais pouvant atteindre 10 m de haut
et 2 m de diamètre, parfois avec un faible empattement à la base.
Il présente une cime arrondie avec les feuilles disposées au bout
des rameaux. Son écorce, grise et lisse devient plus ou moins
ferrugineuse et écailleuse, exsudant un peu de liquide rougeâtre
(Arbonnier, 2002).
-Caractéristiques botaniques
Les feuilles : Le Caïlcédrat
possède des feuilles alternes, paripennées ou parfois
imparipennées, glables avec 3-4-6 paires de folioles opposées ou
subopposées à sommet arrondi. Ses pétioles sont longs de
12 à 25 cm.
L'inflorescence : C'est un fascicule de
panicules à l'aisselle des jeunes feuilles. Les fleurs quant à
elles sont blanches, pédicellées, à pétales
étalés portant au centre les étamines rouges
réunies en tube.
Les fruits : Ils sont dressés vers le
haut, et sont globuleux et ligneux. A maturité, les fruits de couleur
gris clair s'ouvrent en 4 valves en commençant par le sommet. Ils
19
contiennent des graines plates, rectangulaires, ailées
et plus ou moins boursouflées. Sa floraison commence plutôt en
première partie de saison sèche.
3.4.3 Distribution
géographique
Le Caïlcédrat est une espèce des savanes
soudaniennes à guinéennes qui préfère les sols bien
profonds et bien drainés mais qui s'adapte aussi aux sols superficiels
et latéritiques. Il est réparti du Sénégal au
Cameroun jusqu'au Soudan.
3.4.4 Usages
C'est une plante à usage multiple. Ainsi, outre l'objet
de cette étude, le Khaya est sollicité pour le traitement du
paludisme, de l'ictère, de la fièvre, des coliques, de
l'ulcère, de la blennorragie, des conjonctivites, de la gale, la
contusion, la lèpre, l'anémie etc. à travers ses feuilles,
ses racines, son écorce. (Arbonnier, 2002).
20
4. MATERIELS ET METHODES
4.1 Matériels
Le matériel utilisé pour la réalisation de
nos travaux de recherche est constitué:
- d'un GPS (Global Positionning System) pour la prise des
coordonnées géographiques des différents placeaux ;
- d'un coupe-coupe pour la confection de piquets, et
l'ouverture des layons de passage ;
- d'un ruban de 30 mètres pour dimensionner les placeaux
;
- d'un ruban de 1,50m pour les mesures de
circonférence;
- des banderoles pour une bonne délimitation des placeaux
;
-des sécateurs pour le prélèvement des
échantillons à herboriser ;
- des sachets pour garder temporairement les spécimens
;
- d'une presse et des papiers journaux pour la confection des
herbiers ;
- d'un bloc notes pour l'enregistrement des différentes
données de terrain ;
- d'une jumelle pour les observations phénologiques et les
mutilations ;
- des guides d'entretien. 4.2
Méthodes
Elle concerne la recherche documentaire qui nous a surtout
permis d'obtenir des éléments de comparaisons et de mieux
interpréter les résultats obtenus, la collecte et le traitement
des données relatives à chaque objectif spécifique. A cet
effet quelques centres de documentation ont été visités
notamment : les bibliothèques du CENAGREF (Kandi et Banikoara), de la
FSA (BIDOC-FSA/UAC) et de l'Agence pour la
Sécurité de Navigation Aérienne en Afrique et à
Madagascar (ASECNA. Les informations recueillies sont complétées
par celles relevées sur le terrain et sur divers sites Internet (
www.ixquick.com,
http://www.redlist.org).
4.2.1 Collecte des données a) Données
floristiques
Ces données concernent l'inventaire de tous les ligneux
dans des placeaux des terres de parcours communautaires de la zone
d'étude. Les relevés ont été effectués
à partir d'un échantillonnage orienté sur des placeaux de
30 m x 30 m installés dans différents types de formations (savane
saxicole, savane arborée, savane arbustive et savane herbeuse sur
bowé). Ainsi, avec le concours des pisteurs et éco-gardes et sur
le principe de présence-abondance des espèces cibles (D.
oliveri, A. africana, P. erinaceus et K.
senegalensis), 60 placeaux dont 16 placeaux pour
l'espèce D. oliveri, 16 pour P. erinaceus, 15 pour
A. africana et 13 pour K. senegalensis ont été
installés au sein des différents types de formations
végétales rencontrées. La figure 5 montre la localisation
des placeaux
21
Figure 5 : Localisation des placeaux dans le milieu
d'étude (2008)
22
b) Données
dendrométriques
Les données dendrométriques collectées
pour chaque relevé étaient essentiellement : - Le diamètre
de tous les individus à 1,30 m du sol;
- La hauteur de tous les individus ayant au moins 1,30 m de
hauteur de poitrine (dbh); - Le nombre d'individus de la
régénération (hauteur inférieur à 1,30 m) de
tous les brins du sous bois dans des quadrats de 5 m x 5 m. A cet effet, le
placeau de 30 m x 30 m a été quadrillé en des quadrats de
5 m x 5 m dont les quatre des coins et celui du centre ont été
considérés.
c) Données pouvant compromettre la
viabilité des espèces
Ces données concernent la présence ou non de
mutilation sur les individus (écorçage, émondage,
étêtage). La méthode adoptée lors de la collecte des
données est l'appréciation directe des impacts observés
sur toutes les espèces ciblées dans les peuplements de la zone
d'étude. Des codes ont été attribués à
chaque type de mutilation rencontré. Selon le type de mutilation :
- 0 correspond aux individus n'ayant subi
aucun impact
- 1 correspond aux individus
émondés
- 2 correspond aux individus
écorcés
- 3 correspond aux individus
étêtés
- 4 correspond aux individus
étêtés + écorcés
- 5 correspond aux individus
émondés + étêtés
- 6 correspond aux individus
émondés + écorcés + étêtés
d) Données
phénologiques
La méthode adoptée au cours du suivi
phénologique a permis de suivre l'évolution dans le temps de la
disponibilité en élément appétibles (Feuilles,
Fleurs, Gousses). Pour ce faire, dix (10) pieds de chacune des quatre
espèces (A. africana Smith ex Pers., K. senegalensis P. erinaceus,
D. oliveri) numérotées de 1 à 10 ont
été régulièrement suivis tous les 15 jours pendant
deux mois (du 15 septembre au 15 novembre 2008). Les observations ont
porté sur les phases de feuillaison (Fe), Floraison (Fl) et
Fructification (Fr). Les critères d'observation ont été
inspirés de ceux décrits par Grouzis et Sicot (1980).
4.2.2 Traitement et analyse des données a)
Données floristiques
Les différents relevés sont encodés
à l'aide du logiciel Excel. Les relevés phytoécologiques
ont été traités en fonction de la présence-absence
des espèces. L'ensemble de ces relevés a été soumis
à une « Detrented Correspondence Analysis » (DCA) qui est une
forme améliorée de l'Analyse Factorielle des Correspondances
(AFC), méthode qui permet une discrimination dans un espace
réduit de nuage constitué par r relevés (objets) et de
celui de n espèces. Elle autorise ainsi une compréhension plus
facile des différentes structures (groupes de relevés, groupes
d'espèces...), grâce à l'examen des projections des nuages
relevés et espèces dans le
23
plan factoriel. Cette méthode a l'avantage de corriger
la configuration arquée (effet Gutman) qui traduit un gradient
élevé dans les données soumises à une AFC.
Les plans factoriels sont obtenus avec le logiciel CANOCO for
Windows sur la base d'une DCA.
Le dendrogramme ou la classification ascendante
hiérarchique (CAH) est obtenu à l'aide du logiciel STATISTICA par
la méthode de Ward sur la base des distances euclidiennes.
Les types phytogéographiques retenus sont
inspirés des subdivisions chorologiques de White (1986). Ainsi on
distingue :
*Les espèces à large distribution
:
o Cosmopolites (Cos) : espèces distribuées dans
les régions tropicales et tempérées du monde ;
o Pantropicales (Pan) : espèces répandues en
Afrique, en Amérique et en Asie tropicales ;
o Paléotropicales (Pal) : espèces répandues
en Afrique tropicale, en Asie tropicale, à Madagascar et en Australie
;
o Afro-américaines (AA) : espèces répandues
en Afrique et en Amérique tropicale.
*Les espèces pluri-régionales africaines
:
o Pluri-régionales africaines (PRA) : espèces
dont l'aire de distribution s'étend à plusieurs centres
régionaux d'endémisme ;
o Afro-tropicales (AT) : espèces répandues dans
toute l'Afrique tropicale ;
o Afro malgaches (AM) : espèces distribuées en
Afrique et à Madagascar ;
o Soudano zambéziennes (SZ) : espèces
distribuées à la fois dans les centres régionaux
d'endémisme soudanien et zambézien ;
o Soudano-guinéen (SG) : espèces
distribuées à la fois dans les centres régionaux
d'endémisme soudanien et guinéen ;
o Guinéo-congolaise (GC) : espèces
distribuées dans la région guinéenne. *Les
espèces de l'élément base :
o Soudanienne (S) : espèces largement
distribuées dans le centre régional d'endémisme
soudanien.
Les données floristiques ont été
utilisées pour le calcul des paramètres de diversité
(richesse spécifique, indice de diversité de Shannon et
équitabilité de Piélou) de chaque groupement et sont
également utilisées pour l'étude de la répartition
phytogéographique des espèces ligneuses.
? La richesse spécifique (S) a été
déterminée pour les peuplements ligneux. Elle exprime le nombre
total d'espèces ligneuses de dbh = 10 cm dénombré à
chaque relevé. Cette mesure est insuffisante puisqu'elle ne permet pas
de différencier les
communautés qui comporteraient un même nombre
d'espèces mais avec des effectifs différents.
? L'indice de diversité de Shannon-Weiner (H = -? Pi
log2 Pi) où Pi désigne la proportion des individus dans
l'échantillon total qui appartiennent à l'espèce i (H
varie de 0 à 5). Un indice de diversité de Shannon
élevé correspond à des conditions du milieu favorables
à l'installation de nombreuses espèces. (Dajoz, 1985)
? L'équitabilité de Pielou élevée
correspond à une répartition régulière des
espèces avec peu ou pas de dominance marquée. C'est le signe
d'une longue évolution d'une communauté dans le milieu stable.
Les valeurs faibles correspondent à des milieux très
sélectifs comportant quelques espèces dominantes. Cet indice
varie entre 0 et 1.
i ? 1
d i
où H désigne l'indice de diversité de
Shannon et S la richesse
spécifique
1 0.000?
i ? 1
b) Données
dendrométriques
Les mesures dendrométriques ont servi au calcul des
paramètres dendrométriques et aux distributions par classe de
diamètre. Les paramètres dendrométriques calculés
sont :
- le diamètre moyen (cm) des ligneux supérieurs
(dbh = 10) pour chacune des espèces
?
Dm
n
?
cibles du placeau ;
où di désigne le diamètre de chaque sujet et
N le nombre de sujets
-la surface terrière (m2 / ha) des
groupements ainsi que celle des espèces cibles (Daniellia oliveri,
Pterocarpus erinaceus, Afzelia africana et Khaya senegalensis).
Elle est calculée pour les individus de dbh = 10 cm et a pour formule
:
Où di = diamètre à hauteur de
poitrine ( diameter at breast height = dbh) de l'individu

n
i.
2
?
G
?
4s
di
24
où S désigne l'aire du placeau, di le
diamètre de l'individu et n le nombre d'individus. La surface
terrière moyenne est : Gm = G/N,
Gm = surface terrière moyenne
(m2/ha), G = surface terrière du peuplement
(m2/ha) et
N = densité du peuplement
- la densité D (tiges / ha) des arbres. Elle
représente le nombre de pieds par hectare des individus à dbh =
10cm et effectivement présents dans les placeaux inventoriés pour
chaque espèce cible de chaque groupement.
A ce niveau, ont été calculées :
La densité des individus de chaque espèce
ciblée de chaque groupement (dbh = 10 cm) ensuite la densité de
la régénération de chaque espèce cible et de chaque
groupement (dbh < 10 cm) et enfin la densité de la
régénération de chaque espèce cible
25
et de chaque groupement pour une hauteur inférieure
à 1,30 m. Elle est calculée par la formule :
D = (N / S) x 10.000
où D désigne le nombre de plantes ligneuses et S la
superficie exprimée en hectare.
Le pourcentage de régénération des quatre
ligneux a été calculé au sein des populations et les
résultats sont présentés dans un graphe.
-Quant à la distribution par classes de
diamètre, elle a été faite pour les espèces cibles
(Khaya senegalensis, Daniellia oliveri, Pterocarpus erinaceus
et Afzelia africana) ainsi que pour leurs
groupements.
- Les hauteurs ont permis de faire la distribution et la
stratification par classe de hauteur des espèces ciblées au sein
des groupements. Ensuite, pour les quatre ligneux fourragers réunis dans
un graphe, la représentation strate par strate a été
faite.
c) Données pouvant compromettre la
viabilité des espèces
Les données physiques liées à la
présence ou non de mutilation sur les individus ciblés dans les
formations végétales ont été encodées. Les
pourcentages des individus ayant subi ou non des mutilations ont
été calculés et les diagrammes ont été
réalisés pour chaque espèce ciblée.
d) Données
phénologiques
Les résultats des observations phénologiques
faites sur les dix (10) pieds de chaque espèce ciblée ont
été analysés dans un tableau.
26
5. RESULTATS
5.1. Analyse globale de la
végétation
Globalement, 60 relevés ont été
réalisés et 123 espèces inventoriées. La «
Detrented Correspondence Analysis » (DCA) donne une inertie totale de 7,2.
Les valeurs propres et les pourcentages de variance correspondants sont
résumés dans le tableau 1.
Tableau 1: Valeurs propres et pourcentage de variance
expliqué par les 4 premiers axes.
|
Axes
|
1
|
2
|
3
|
4
|
Inertie totale
|
|
Valeurs propres
|
0,603
|
0,307
|
0,279
|
0,209
|
7,237
|
|
Longueur des gradients
|
4,416
|
3,580
|
2,724
|
2,731
|
|
|
Pourcentage cumulatif de variance expliquée
(%)
|
8,3
|
12,6
|
16,4
|
19,3
|
|
Il ressort de la DCA que les 4 premiers axes expliquent
à 19,3 % la variance totale. Cette variance est très faible et
s'explique d'une part par la durée relativement courte de l'étude
et d'autre part par la petite taille des échantillons (60
relevés) pour la zone d'étude. C'est ce qui justifie ce biais
observé. Cependant, cette valeur met en évidence, la dispersion
des informations sur les axes factoriels.
La figure 6 donne la représentation des
différents groupes de relevés dans le plan factoriel. Sur l'axe 1
se suivent de l'origine vers la partie positive les relevés des
formations végétales fermées et plus humides (forêt
galerie et forêts claires); puis ceux des formations
végétales moins ouvertes et plus sèches regroupant les
savanes boisées
27
et arbustives. Cette dynamique évolutive du couvert
végétal peut s'expliquer par l'existence d'un gradient
d'hygrométrie qui affecte la disposition spatiale des 60 relevés.
Sur l'axe 2, se suivent de l'origine vers la partie positive, les formations
fermées (forêts et savanes boisées) et les formations
ouvertes (savanes arborées et arbustives). Cette succession de
formations révèle l'existence d'un gradient de maturation de la
végétation.
La figure 7 montre le dendrogramme de la classification
hiérarchique des groupements végétaux.
A 75 % de dissimilitude, le dendrogramme révèle
quatre groupements végétaux G1, G2, G3 et G4.
- Le groupement G1 est un regroupement de 22 relevés
assez diffus constitué de 11 placeaux de D. oliveri (soit 50%
des relevés), 5 placeaux de A. africana, 4 placeaux de K.
senegalensis et de 2 placeaux de P. erinaceus dans 13 savanes
arbustives, 4 forêts claire, 3 savane boisées et 1 savane
arborés. Ce groupement est caractérisé par D. oliveri
et Combretum ghasalense dans des savanes arborées/boisées au
niveau des dépressions et des savanes arbustives sur des sols sableux et
des sols limoneux ;
- Le groupement G2 constitué de 4 relevés est
composé des relevés de K. senegalensis dans des
forêts galeries. Ce groupement est caractérisé par K.
senegalensis et Mimosa pigra.
- Le groupement G3 est composé de 18 relevés
dont 13 placeaux de P. erinaceus soit 72 % des relevés, 3
placeaux de K. senegalensis et 2 placeaux de D. oliveri dans
9 savanes arbustives, 6 savanes boisées, 2 forêts claires et 01
forêt galerie. Ce groupement est caractérisé par P.
erinaceus et Anogeissus leiocarpa, dans des savanes
boisées au niveau des savanes arbustives.
- Le groupement G4 formé de 16 relevés est le
regroupement de 10 placeaux de A. africana soit 63% des
relevés, 3 placeaux de D. oliveri, 2 K. senegalensis
et 1 P. erinaceus dans 9 savanes boisées, 4 savanes
arbustives et 3 forêts claires et Balanites aegyptiaca, Detarium
microcarpum, Burkea africana des savanes arbustives et des savanes
arborées sur des sols sableux et des sols argileux. Ce groupement est
caractérisé par Afzelia africana et Balanites
aegyptiaca.
+ Humide (Gradient) + Sec
R20
R30
G2
(Forêts Galeries)
R19
G4
(Forêts claires et Savanes
boisées)
R18
R29
R17
R44
R32
R24
R52
R27
R23
R43 R22
R R54
R16R9R5
5
R57R35
R31
R4R41R5R
R40
58
R33
R21 R8R4
R51
R46
R14
R28
R36
R6
R47
R25
R59
R50
8R42
R1
R34R7
0 R13
R15
R45
R37
11
R49R26
R39
R38
R60
G3
(Savanes arborées et
arbustives)
R3R12
G1 (Savanes arborées et
arbustives)
R2
28
R53
-1.0 5.0
Figure 6: Répartition des groupements
végétaux dans le plan des axes 1 et 2 de la
Légendes : G1 = groupement à Daniellia oliveri
et Combretum ghasalense
G2 = groupement à Khaya senegalensis et
Mimosa pigra
G3 = groupement à Pterocarpus erinaceus et
Anogeissus leiocarpa
G4 = groupement à Afzelia africana et
Balanites aegyptiaca
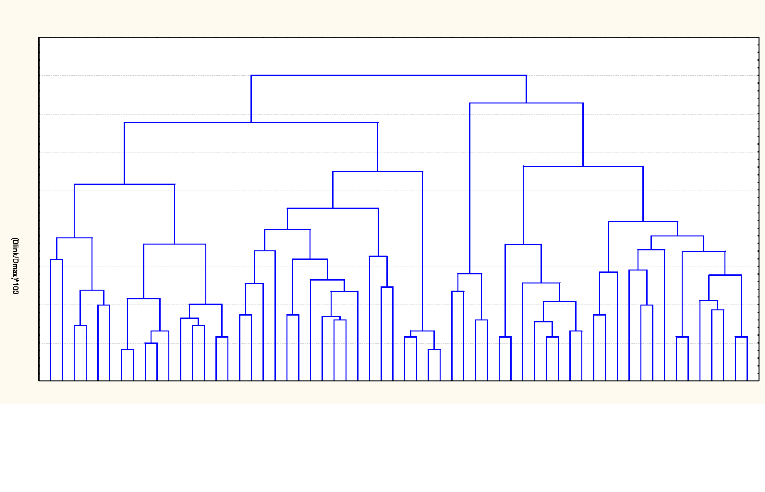
110
100
40
20
90
80
70
60
50
30
R53
R28
R43
R40
R24
R23
G4
R51
R35
R16
R14
R9
R52
R55
R42
R54
R8
R58
R44
R41
R29
R57
R47
R37
R48
R36
Tree Diagram for 60 Variables
Ward`s method
Euclidean distances
R27
R25
R60
G3
R56
R13
R11 R7
R6 R5
R30
R20
G2
R19
R18
R49
R46
R45
R59
R33
R31
R32
G1
R17
R50
R34
R21
R39
R38
R22
R12 R3
R4 R26
R15 R10
R2 R1
Figure 7 : Dendrogramme des groupements
végétaux
Légendes : G1 = groupement à Daniellia
oliveri et Combretum ghasalense
G2 = groupement à Khaya senegalensis et
Mimosa pigra
G3 = groupement à Pterocarpus erinaceus et
Anogeissus leiocarpa
G4 = groupement à Afzelia africana et
Balanites aegyptiaca
29
30
5.2 Caractéristiques
dendrométriques
5.2.1 Caractéristiques du groupement à
Daniellia oliveri et Combretum ghasalense (G1)
La composition floristique (annexe 4) est établie
à partir de 22 relevés avec 69 espèces ligneuses. Le
groupement est constitué surtout de 13 relevés de savanes
arbustives, de 03 savanes boisées, d'une savane arborée et de 04
forêts claires souvent inondées. Les espèces les plus
rencontrées sont : D. oliveri, Mimosa pigra et C.
ghasalense.
Les valeurs de la richesse spécifique et de l'indice de
diversité de Shannon sont : respectivement de 69 et de 2,51 bits.
L'équitabilité de Piélou est de 0,67. Ces
paramètres de diversité spécifique montrent une
diversité moyenne des espèces ligneuses avec une
répartition équilibrée. C'est le groupement à
D. oliveri et Combretum ghasalense
a) Structure diamétrique, distribution en
hauteur et types phytogéographiques
La répartition par classe de diamètre des
ligneux de ce groupement (dbh > 10 cm) (figure 8) présente une allure
en forme de "J" renversé caractéristique des formations
naturelles très jeunes. La distribution s'ajuste mieux à une
fonction polynôme de degré 3 avec un coefficient de
détermination élevée (R2= 0,99).
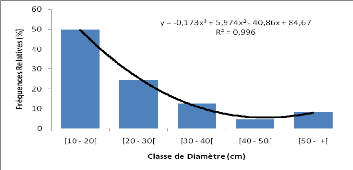
Figure 8 : Distribution par classes de diamètre
des individus du groupement à D. oliveri et C. ghasalense
(G1)
Quand à la répartition par classe de
diamètre de D. oliveri au sein de cette population (dbh >10
cm) (figure 9), l'allure de la courbe polynômiale se rapproche plus de
celle de Gauss. On remarque aussi qu'il y assez de jeunes sujets de D.
oliveri avec près de 40% de des individus ayant un diamètre
compris entre 20 et 30 cm. Le diamètre et les fréquences
relatives obtenues sont fortement corrélés (R2 =
0,94). Plus les individus sont jeunes, plus la fréquence est
élevée. Le diamètre moyen est de 30 cm pour les pieds de
D. oliveri (dbh > 10 cm).
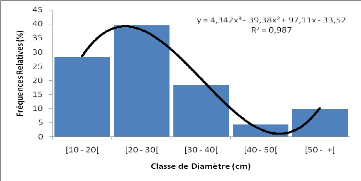
31
Figure 9: Distribution par classes de diamètre de
D. oliveri au sein du groupement G1
La figure 10 montre la distribution par classe de hauteur des
espèces de Daniellia oliveri au sein du groupement. L'allure de
la courbe montre une distribution normale avec plus de 60% des individus de
D. oliveri ayant une hauteur comprise entre 4 et 10
m.
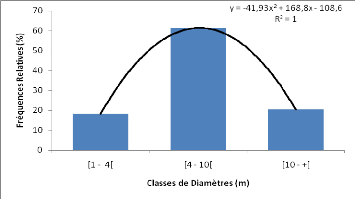
Figure 10 : Distribution par classe de hauteur de D.
oliveri au sein du groupement G1
La figure 11 donne l'importance numérique des divers
types phytogéographiques présents dans ce groupement.
Les espèces de l'élément base soudanien
(S) 40 % et les espèces Soudano-zambésiennes
(SZ) 37% sont les plus abondantes. Le milieu est soumis
à de faibles actions anthropiques et conserve un relatif état
d'équilibre. Par contre les espèces à large distribution
Pantropicales (Pan) et Paléotropicales (Pal)
et les espèces Afro - Malgache (AM) sont les
moins représentées avec un spectre total de moins de 07 %.
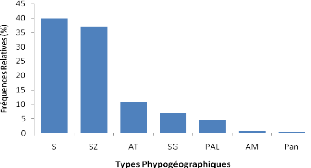
32
Figure 11 : Spectres phytogéographiques bruts du
groupement à D. oliveri et C. ghasalense
(G1)
b) Densités, potentiel de
régénération et surface terrière
La densité des ligneux dans le groupement est de 466,67
tiges/ha pour une surface terrière de 10,71 m2/ha. La
densité des pieds de D. oliveri au sein des peuplements est de
93,94 tiges/ha pour une surface terrière de 7,51 m2/ha. La
régénération naturelle (semis et repousses confondus) des
brins et perchis est de 22,63 brins/ha.
5.2.2 Caractéristiques du groupement Khaya
senegalensis et Mimosa pigra (G2)
La composition floristique (annexe 3) est établie
à partir de 04 relevés de 34 espèces ligneuses. Le
groupement est constitué uniquement des relevés de forêt
galerie. Les espèces les plus rencontrées sont : K.
senegalensis, Mimosa pigra et Kigelia africana. Les valeurs de la richesse
spécifique et de l'indice de diversité de Shannon sont :
respectivement de 34 espèces et 2,79 bits. L'équitabilité
de Piélou est de 0,73. Ces paramètres de diversité
spécifique montrent une diversité moyenne des espèces
ligneuses avec une abondance de Mimosa pigra sur l'ensemble des autres
espèces du groupement. C'est le groupement à K. senegalensis
et à M. pigra
a) Structure diamétrique, distribution en
hauteur et types phytogéographiques
La répartition par classe de diamètre des
ligneux de ce groupement (dbh > 10 cm) (figure 12) présente une
allure en forme de "J" renversé caractéristique des formations
naturelles jeunes polyspécifiques et inéquiennes. La distribution
s'ajuste mieux à une fonction polynôme de degré 3 avec un
coefficient de détermination élevée (R2 = 0,99). La valeur
élevée du coefficient de détermination (R2 =
0,99). Les individus de diamètre 40 à 50 cm sont en très
faible proportion dans ces formations. Le diamètre moyen est de 41,95 cm
pours les pieds de K. senegalensis (dbh > 10 cm).
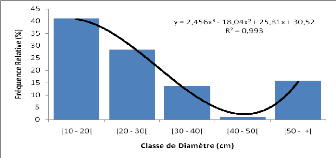
33
Figure 12 : Distribution par classe de diamètre
des individus du groupement à K. senegalensis et M. pigra
(G2)
La répartition en classe de diamètres des K.
senegalensis (figure 13) présente une allure plus ou moins
sinusoïdale au sein des formations. Cette distribution ne répond
à aucune loi normale On retrouve une proportion élevée de
gros individus et moins de jeunes arbres. La valeur élevée du
coefficient de détermination (R2 = 0, 94) explique à
94% les variations observées. La proportion des sujets âgés
est élevée dans les populations de K. senegalensis. Le
renouvellement des populations de cette espèce se trouve ainsi
hypothéqué.
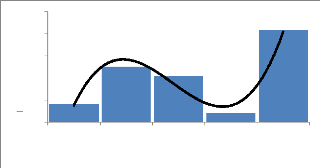
[10 - 20[ [20 - 30[ [30 - 40[ [40 - 50[ [50 - +[
Classe de Diamètre (cm)
y = 6.25x3 - 54.167x2 + 139.58x -
84.167 R2 = 0.9449
50
Fréquence Relative %
40
30
20
10
0
Figure 13 : Distribution par classe de diamètre
de K. senegalensis au
sein du groupement G2
La figure 14 montre une stratification des populations de
K. senegalensis dans le groupement. La courbe polynômiale
à l'allure de Gauss s'apparente intimement à une distribution
normale mais, on remarque que la proportion des sous arbustes dans le
peuplement est très faible. La classe modale est [4 - 10 m [
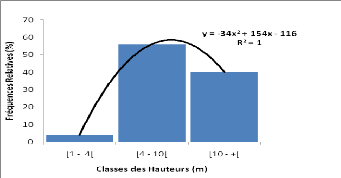
34
Figure 14 : Distribution par classe de hauteur de
K. senegalensis au sein du
groupement G2
La figure 15 montre l'importance numérique des divers
types phytogéographiques présents au niveau de la strate ligneuse
du groupement à K. senegalensis et Mimosa pigra. Les
espèces de l'élément base soudanien (S) sont les plus
abondantes avec un pourcentage de 36 % de l'effectif. Cette abondance de
l'élément base soudanien justifie un état
d'équilibre du milieu. Par contre, les espèces à large
distribution Afrique Tropicale (AT), Afro - Américaines (AA) et les
espèces Guinéo - Congolaises (GC) sont les moins
représentées avec un spectre total de moins de 6 %.
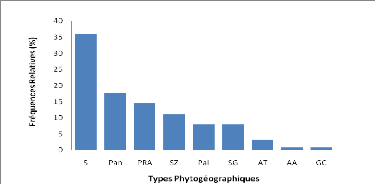
Figure 15: Spectres phytogéographiques bruts du
groupement G2
b) Densités, potentiel de
régénération et surface terrière
Au sein de ce groupement, la densité des ligneux est de
347,22 tiges/ha pour une surface terrière de 21,08 m2/ha. La
densité de K. senegalensis est de 69,44 tiges/ha pour une
surface terrière de 10,66 m2/ha. La
régénération naturelle (semis et repousses confondus) de
K. senegalensis dans ce groupement est de 3,04 brins/ha
35
5.2.3 Caractéristiques du groupement P.
erinaceus et Anogeissus leiocarpus (G3)
La composition floristique (annexe 5) est établie
à partir de 18 relevés avec 81 espèces ligneuses. Le
groupement est constitué surtout de 09 relevés de savanes
arbustives, de 06 savanes boisées, d'une forêt galerie et de 02
forêts. Les espèces les plus rencontrées sont : P.
erinaceus, Vitellaria paradoxa, Anogeissus leiocarpus dans des savanes
boisées et Terminalia avicennioides, Crossopteryx febrifuga et
Annona senegalensis, au niveau des savanes arbustives.
Les valeurs de la richesse spécifique et de l'indice de
diversité de Shannon sont : respectivement de 81 et de 3,58 bits
(moyen). L'équitabilité de Piélou (E) est de 0,90
(élevé). En chaque point, il y a une niche favorable à
plusieurs espèces. La valeur élevée de E est le
résultat d'une équitabilité parfaite. Les espèces
sont bien réparties. Elles arrivent avec des effectifs égaux
ayant tendance à une codominance relative. Cependant, Pterocarpus
erinaceus (espèce cible) vient avec des effectifs élevés
suivi de Vitellaria paradoxa, Anogeissus leiocarpa et Annona
senegalensis. C'est le groupement à Pterocarpus erinaceus
et à Anogeissus leiocarpus.
a) Structure diamétrique, distribution en
hauteur et types phytogéographiques
La répartition par classe de diamètre des
ligneux de ce groupement (dbh > 10 cm) (figure 16) présente une
allure plus ou moins oblique. La courbe décroit des classes de faibles
diamètres (10 - 20 cm) vers les classes de diamètres les plus
élevées (plus de 50 cm). La distribution s'ajuste mieux à
une fonction polynôme de degré 3 (R2 = 0,98). Plus les
individus sont jeunes, plus la fréquence est élevée. Il
s'agit d'un peuplement jeune composé d'individus d'âges
multiples.
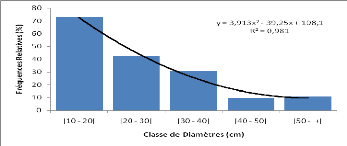
Figure 16: Distribution par classes de diamètres
des individus du groupement
P. erinaceus A.et leiocarpus
(G3)
La répartition par classe de diamètre de P.
erinaceus au sein de ces formations (dbh >10 cm) (figure 17) montre une
distribution plus ou moins normale (individus d'âges multiples)
comportant de grandes proportions de petits individus et moins de gros arbres.
On note également une forte proportion (plus de 40%) d'individus ayant
un diamètre compris entre 10 - 20 cm et 30 - 40 cm. La valeur du
coefficient de détermination (R2 = 0,6) montre que seulement
60% des variations de classes de
36
diamètre sont expliquées par les variations des
fréquences relatives. Le diamètre moyen des sujets de
Pterocarpus erinaceus dans ces formations est de 26,30 cm.
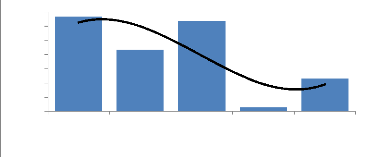
[10 - 20[ [20 - 30[ [30 - 40[ [40 - 50[ [50 -
+[
Classes de Diamètres (cm)
Fréquences Relatives (%)
35
30
25
20
15
10
0
5
y = 1.57x3 - 13.923x2 + 29.434x +
14.203 R2 = 0.6009
Figure 17: Distribution par classe de diamètre
de P. erinaceus au sein du
groupement G3
La figure 18 présente la répartition en hauteur
des individus de P. erinaceus. La distribution est normale avec des
proportions faibles d'individus dans la strate sous arbustive et très
élevées d'individus dans la strate arbustive (4 - 10 m).
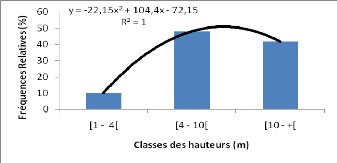
Figure 18: Distribution par classe de hauteur de P.
erinaceus au sein du
groupement G3
La figure 19 donne l'importance numérique des divers
types phytogéographiques présents au niveau de ces formations.
Les espèces de l'élément base soudanien (S) (35 %) et
soudano-guinéenne (23%) sont les plus abondantes. Par contre les
espèces pluri-régionales africaines (PRA), Afro-américaine
(AA), guinéo-congolaises (GC) et les espèces Afro - Malgache (AM)
sont les moins représentées avec un spectre total de moins de 04
%. Ceci se justifie par le fait que les actions anthropiques n'ont pas
perturbé l'état d'équilibre de ce milieu.
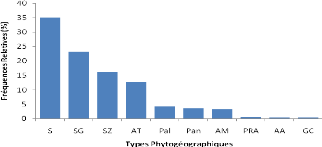
37
Figure 19: Spectres phytogéographiques bruts du
groupement G3
b) Densités, potentiel de
régénération et surface terrière
La densité des ligneux dans le
groupement est de 480,86 tiges/ha pour une surface terrière de 25,87
m2/ha. La densité des pieds de Pterocarpus erinaceus
au sein des peuplements est de 66,67 tiges/ha pour une surface
terrière de 4,99 m2/ha. La régénération
naturelle (semis et repousses confondus) des brins et perchis est de 7,90
brins/ha.
5.2.4 Caractéristiques du groupement Afzelia
africana et Balanites aegyptiaca (G4)
La composition floristique (annexe 6) est établie
à partir de 16 relevés avec 56 espèces ligneuses. Le
groupement est constitué surtout des relevés de savanes
boisées et arbustives. Les espèces les plus rencontrées
sont : Afzelia africana, Isoberlinia doka et Daniellia oliveri
dans des forêts claires et Balanites aegyptiaca, Detarium
microcarpum, Burkea africana dans des savanes arbustives et des savanes
arborées sur des sols sableux et des sols argileux.
Les valeurs de la richesse spécifique et de l'indice de
diversité de Shannon sont : respectivement de 56 espèces et 2,72
bits. L'équitabilité de Piélou est de 0,69. Ces
paramètres de diversité spécifique montrent une
diversité moyenne des espèces ligneuses avec la dominance d'un
effectif réduit d'espèces telles que Afzelia africana,
Balanites aegyptiaca, Vitellaria paradoxa et Burkea africana
sur l'ensemble des espèces du groupement. C'est le groupement
à Afzelia africana et à Balanites
aegyptiaca.
a) Structure diamétrique, distribution en
hauteur et types phytogéographiques
La répartition par classe de diamètre des
ligneux de ce groupement (dbh > 10 cm) (figure 20) présente une
allure en forme de "J" renversé caractéristique des formations
naturelles très jeunes. La distribution s'ajuste mieux à une
fonction polynôme de degré 3 avec un coefficient de
détermination élevée (R2 = 0,99) expliquant
à 99% les variations des fréquences relatives par rapport aux
classes de diamètre. La forte corrélation entre les
diamètres et les fréquences relatives montre que plus jeunes sont
les individus, plus élevée est leur fréquence relative.
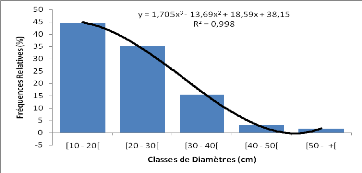
38
Figure 20: Distribution par classes de diamètres
des individus du groupement A.
africana et B. aegyptiaca
(G4)
La répartition par classe de diamètre de A.
africana au sein de ces formations (dbh>10 cm) (figure 21)
répond à une loi de distribution normale (courbe de Gauss)
d'individus d'âges multiples dans un peuplement équienne
dominé par des individus de classes de diamètres allant de 30
à 40 cm. La valeur du coefficient de détermination (R2
= 0,94) montre que 94% des variations des fréquences relatives sont
expliquées par celles des classes de diamètre. Le diamètre
moyen des sujets de A. africana dans ces formations est de 30,08
cm.
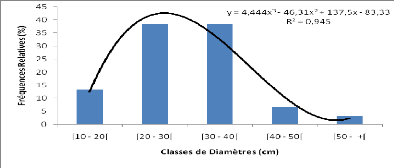
Figure 21: Distribution par classe de diamètre
de A. africana au sein du groupement G4
La figure 22 présente la répartition en hauteur
des individus de Afzelia africana. La distribution est normale avec
des proportions faibles d'individus dans la strate sous arbustive (1 - 10 m),
moyennes dans la strate arborée (+ de 10 m) et des proportions
très élevées d'individus dans la strate arbustive (4 - 10
m).
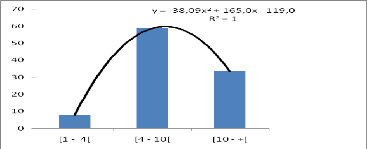
39
Figure 22: Distribution par classe de hauteur de A.
africana
La figure 23 donne l'importance numérique des divers
types phytogéographiques présents au niveau de ces formations.
Les espèces de l'élément base soudanien (S) (38 %) et
soudano - zambézienne (28%) sont les plus abondantes. Par contre les
espèces pluri - régionales africaines (PRA), pantropicales (Pan)
et les espèces Afro - Malgache (AM) sont les moins
représentées avec un spectre total de moins de 04 %. Le milieu
est faiblement assujetti aux effets des perturbations.
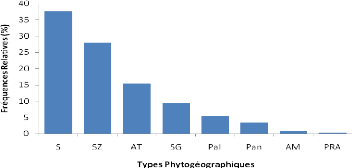
Figure 23: Spectres phytogéographiques bruts du
groupement A. africana et B. aegyptiaca (G4)
b) Densités, potentiel de
régénération et surface terrière
La densité des ligneux dans le
groupement est de 526,67 tiges/ha pour une surface terrière de 16,99
m2/ha. La densité des pieds de A. africana au sein
des peuplements est de 70 pieds/ha pour une surface terrière de 3,33
m2/ha. La régénération naturelle (semis et
repousses confondus) des brins et perchis est de 3,51 brins/ha.
40
5.3 Exploitations pouvant compromettre la
viabilité des espèces
5.3.1 Khaya senegalensis
La figure 24 illustre les différentes formes
d'exploitation et de prélèvement observées sur K.
senegalensis. Seulement 4% des sujets sont épargnés des
mutilations. L'espèce est très recherchée par les
éleveurs compte tenue des pourcentages d'émondage.
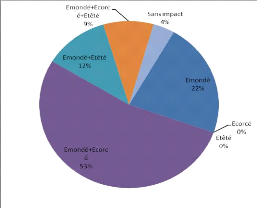
Figure 24 : Pourcentage des individus de K.
senegalensis ayant subi des mutilations
5.3.2 Daniellia oliveri
Les pourcentages des prélèvements des
différents organes sont illustrés par la figure 25. Environ 79%
des individus sont épargnés. L'espèce est peu
recherchée par les éleveurs.
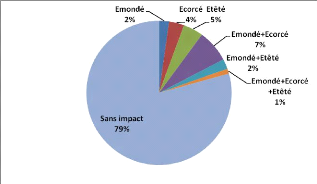
Figure 25 : Pourcentage des individus de D. oliveri
ayant subi des
mutilations
41
5.3.3 Pterocarpus erinaceus
Les pourcentages des prélèvements des
différents organes sont illustrés par la figure
26. Seulement 15% des individus sont épargnés de
toutes actions anthropiques. L'espèce est très
émondée.
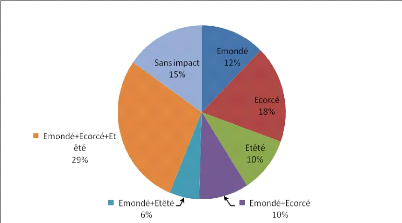
Figure 26 : Pourcentage des individus de P.
erinaceus ayant subi des mutilations
5.3.4 Afzelia africana
Les pourcentages des prélèvements des
différents organes sont illustrés par la figure
27. 28% des individus sont épargnés de toutes
actions anthropiques. L'espèce est très émondée.
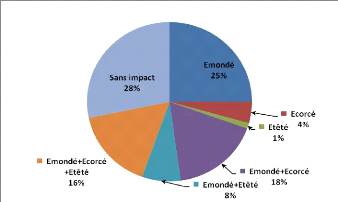
Figure 27 : Pourcentage des individus de A.
africana ayant subi des
mutilations
42
5.4 Phénologie des quatre espèces
ciblées
La phénologie est l'étude de l'apparition des
organes d'une plante au cours de l'année : bourgeons foliaires,
feuilles, floraux, fleurs, fruits, jaunissement des feuilles, chute des
feuilles, des fleurs et des fruits.
La phénologie d'un peuplement doit être
étudiée pendant 5 ans avec deux sortes environs par mois pour
établir des données fiables sur les espèces
végétales de ce peuplement (Houngnon, 1996). Dans le cadre de
notre étude, des observations ont été faites à
chaque quinzaine pendant les mois de septembre et d'octobre sur 10 pieds de
chacune des espèces ciblées dans les peuplements naturels
où elles évoluent. Deux groupes de plantes ligneuses sont
observés : les plantes de savanes (D. oliveri, P. erinaceus et
A. africana) qui perdent leurs feuilles en saison sèche et les
plantes de galerie (K. senegalensis) qui gardent pour la plupart leur
feuillage vert. Compte tenue des conditions édaphiques favorables de
certaines zones (rivière, cours d'eau...), les quatre espèces
conservent longtemps leur feuillage vert même en saison sèche. Des
observations faites, il résulte qu'A. africana et D.
oliveri porte de jeunes feuilles jusqu'au mois de novembre, incluant la
période de crise fourragère aiguë. Ces deux espèces
présentent un grand intérêt fourrager. Mais,
l'intérêt fourrager que présente A. africana de
plus que D. oliveri réside dans ses feuilles qui sont
très recherchées par le bétail. P. erinaceus perd
progressivement les feuilles vers la fin du mois de septembre tandis que la
plaine floraison a été observée depuis la première
quinzaine d'observation en septembre jusqu'à la quatrième
quinzaine au début du mois de novembre. Les résultats des
observations sont consignés dans le Tableau 2.
A défaut de cette étude, un aperçu de la
phénologie des principales espèces rencontrées est
résumé dans un tableau général (Annexe 8).
5.4.1 Khaya senegalensis
La période d'étude correspond à la phase
de floraison de K. senegalensis. Pendant les mois de septembre et
d'octobre, on a observé au moins 5 fleurs sur chaque pied. Mais,
très peu de jeunes individus issus de la germination des semences ont
été observés aux pieds des arbres (semencier). Ceci serait
dû à un faible pouvoir germinatif des conditions de germination
des semences ou à un défaut de maturation des semences.
5.4.2 Daniellia oliveri
Au cours de nos travaux, seuls les fruits verts ont
été observés sur les 10 pieds de Daniellia
oliveri. Les mois de septembre et d'octobre correspondent à la
phase de fructification. On remarque assez de jeunes petites pousses mais la
plupart, provient des repousses et non des semis.
43
5.4.3 Pterocarpus erinaceus
Les observations faites dans le mois de septembre ont
révélé que les dix (10 pieds de P. erinaceus
avaient pour la plus part du temps des feuilles vertes et par endroit des
feuilles jaunies. Une chute de feuilles de plus en plus importante
précédée de jaunissement a marqué le mois
d'octobre.
5.4.4 Afzelia africana
Les observations faites dans le mois de septembre ont
révélé que les dix (10) pieds de A. africana
sélectionnés avaient sur toute la période
d'observation, gardé leurs feuilles vertes. Aucune jeune repousse n'a
été signalée sous les grands pieds malgré la
présence des gousses éclatées issues de la
dissémination lors de la saison passée.
5.4.5 Comparaison des observations
phénologiques entre espèces Tableau 2 : Résultat des
observations phénologiques
|
Famille/Sous Famille
|
Espèce
|
SEPTEMBRE
|
OCTOBRE
|
|
Leguminosae/Caesalpinioideae
|
Daniellia oliveri
|
Fv
|
Fv
|
|
Leguminosae/Papilionioideae
|
Pterocarpus erinaceus
|
Fv
|
Ch
|
|
Leguminosae/Caesalpinioideae
|
Afzelia africana
|
Fv
|
Fv
|
|
Meliaceae
|
Khaya senegalensis
|
Fl
|
Fl
|
|
Fv = Feuilles verte Ch = Chute de feuilles Fl = Fleur
|
44
6. DISCUSSION
6.1 Facteurs du milieu et répartition des
groupements végétaux
La pluviométrie, l'hydromorphie et les types de sols
sont déterminants pour expliquer les regroupements, les variations de la
composition floristique, et de la surface terrière observés dans
les terres de parcours en zone soudanienne au Bénin. Le milieu
écologique favorable à A. africana se situe à
l'extrémité nord de la Zone d'étude, plus sec, tandis que
K. senegalensis est plus abondante dans les forêts galeries
(Alibori) plus humide (Sinsin et al., 1989). Selon Gaoué
(2008), les populations de K. senegalensis sont retrouvées dans
les galeries forestières de la zone soudanienne. D. oliveri et
P. erinaceus sont présentes dans l'ensemble de la zone
étudiée mais D. oliveri est plus abondante et surtout
dans les zones de dépression Adomou (2005), Adjahossou (2008). Les
résultats d'observation montrent que la phytocénose du milieu
d'étude subit en général plus les effets de la longue
sécheresse, des actions de la faune et également des actions
anthropiques au cours des transhumances, des patrouilles, des embuscades et de
divers travaux de pistes...etc. Les relevés phytoécologiques
révèlent en outre une grande quantité de sous bois qui est
en dessèchement et qui indique une grande vulnérabilité de
ces peuplements au dégât des feux et de la sècheresse.
Toute fois, les données quantitatives sont nécessaires pour
apprécier le niveau de menace lié à la litière du
sous bois et penser les stratégies d'aménagement et de
gestion.
6.2 Caractéristiques floristiques des
espèces ligneuses fourragères 6.2.1 Chorologie
La figure 28 montre l'importance numérique des types
phytogéographiques de la strate ligneuse des quatre groupements
étudiés. Au total, les espèces de l'élément
base soudanien (S) sont les plus dominantes. Ce résultat est en accord
avec la définition proposée par White (1986) selon laquelle
l'élément base soudanien est dominant dans cette zone.
En effet, la chorologie des espèces est un indicateur des
conditions du milieu dans la mesure où chaque espèce
réalise son abondance maximale dans sa station favorable. Ces
espèces ont une aire de répartition dominante qui s'amenuise avec
les perturbations anthropiques. Par ailleurs, les espèces à large
distribution sont en très faibles proportions ce qui montre que les
actions anthropiques ont eu de faibles impacts sur le milieu. S'il est vrai que
les espèces de la zone soudanienne dominent, il n'est pas moins vrai que
les formes d'exploitation traditionnelles rencontrées réduisent
de plus en plus leur abondance voir leur disponibilité. Les travaux
menés par Fagnissè (2006) et Adjahossou (2008) dans la même
zone d'étude montrent une forte présence des espèces
soudaniennes et soudano-zambéziennes, ce qui confirme les
résultats que nous avons obtenus. Par ailleurs, dans les biotopes
perturbés, les plantes à large distribution prédominent
(Sinsin et Oumorou, 2000 ; Houinato et Sinsin, 2001).
45
6.2.2 Diversité de Shannon et
équitabilité de Piélou
La composition floristique du groupement à K.
senegalensis et Mimosa pigra (annexe 2) uniquement
constitué des relevés de galerie forestière montre K.
senegalensis et Mimosa pigra comme espèces les plus
rencontrées.
L'analyse des valeurs de la richesse spécifique et de
l'indice de diversité de Shannon, montre une diversité moyenne
des espèces ligneuses avec une abondance de Mimosa pigra sur
l'ensemble du groupement. On note au niveau de la strate arborescente une
tendance de dominance de K. senegalensis indiquant un renouvellement
presque nulle de l'espèce dans le peuplement. Ces résultats sont
conformes à ceux de Gaoué (2008) obtenus sur six populations de
K. senegalensis dans les forêts galerie de la zone soudanienne.
C'est seulement au niveau de ces populations que l'on enregistre les forts taux
de prélèvement
(émondage+écorçage+étêtage) à plus de
50%. La régénération potentielle est presque nulle et ceci
explique aisément le faible recrutement de l'espèce et sa
présence sur la liste rouge de l'IUCN.
La composition floristique du groupement à D.
oliveri et Combretum ghasalense (annexe 3) constitué de 13
relevés de savanes arbustives, 3 relevés de savanes
boisées, d'un relevé de savane arborée et de 4
relevés de forêts claires souvent inondées ; montre que les
espèces les plus rencontrées sont D. oliveri et
Combretum ghasalense. L'analyse des valeurs de la richesse
spécifique et de l'indice de diversité de Shannon sont les
résultats d'une diversité moyenne et d'une répartition
équilibrée des espèces ligneuses du groupement. La
dispersion de Daniellia oliveri est cependant très
marquée au sein du groupement. La faible valeur de l'indice de
diversité Shannon obtenue pour le groupement à D. oliveri
et Combretum ghasalense (G1) montre une spécialisation des
espèces qui s'y trouvent avec un phénomène de dominance
d'un petit nombre d'espèces sur l'ensemble des espèces de la
communauté (Devineau, 1984). Ce résultat reste en harmonie avec
les récents travaux conduits par Adjahossou (2008) dans les aires
protégées de la même région.
Pour le groupement à P. erinaceus et A.
leiocarpus (Annexe 4), l'analyse des valeurs de la richesse
spécifique et de l'indice de diversité de Shannon concluent qu'en
chaque point, il y a moyennement une niche favorable à plusieurs
espèces. La valeur élevée de E (E > 0,8) indique des
espèces bien réparties. Elles arrivent avec des effectifs
égaux ayant tendance à une codominance relative. Cependant,
P. erinaceus vient avec des effectifs élevés (comme pour
la plupart des quatre espèces ciblées) suivi de Vitellaria
paradoxa, Anogeissus leiocarpa et Annona senegalensis. Ceci est dû
au fait que les relevés sont effectués dans les zones de
représentativité des espèces recherchées. Les
paramètres de diversité élevés du groupement
à P. erinaceus et Anogeissus leiocarpa traduisent une
grande diversité.
Pour l'ensemble des relevés constituant le groupement
à A. africana et Balanites aegyptiaca, l'analyse des valeurs de
la richesse spécifique, de l'indice de diversité de Shannon et de
l'équitabilité de Piélou révèle une
diversité moyenne des espèces ligneuses avec l'agrégation
par endroit de Balanites aegyptiaca et d'autres espèces telles
que Vitellaria paradoxa et Burkea africana sur l'ensemble du
groupement. Dans l'ensemble, les résultats d'observation, montrent que
A. africana présente une faible abondance dans les types de
formations parcourues. Ces résultats corroborent ceux de Adjahossou
(2008) dans la RBT du W. Par ailleurs, on retrouve Afzelia
africana,
46
Isoberlinia doka et D. oliveri dans des
forêts claires et Balanites aegyptiaca, Detarium microcarpum, Burkea
africana dans les savanes arbustives et arborées sur des sols
sableux et argileux.
6.3 Comparaison des paramètres
dendrométriques
Dans l'ensemble, les densités des espèces cibles
dans les groupements (dbh = 10 cm) sont faibles. Les diamètres moyens
(cm) des individus de dbh = 10 cm sont de 41,95 ; 30,08 ; 30 et 26,3 cm
respectivement pour K. senegalensis, A. africana, D. oliveri et P.
erinaceus. Comparativement aux trois autres espèces, K.
senegalensis présente la valeur la plus élevée
à cause des conditions favorables liées à son habitat
(forêt galerie). Dans la zone soudanienne du Bénin, les travaux
effectués par Sinsin et al. (2003) et Houéhanou (2008),
ont révélés respectivement 31,69 cm et 42 cm pour les
diamètres de A. africana, lesquelles valeurs sont sensiblement
supérieures à la nôtre. Or selon Sinsin et al.
(2003), le diamètre moyen de A. africana varie
significativement d'une zone climatique à une autre. Ces
différences pourraient ainsi se justifier si la zone d'étude de
ces derniers était plus large ou incluait des zones de
pluviométrie plus élevées. Dans les forêts claires
moins sèches de la zone soudano - guinéenne, plus
précisément dans le département du Borgou au Nord -
Bénin ; Sokpon et al., (2006) ont trouvé 45 cm et 35 cm
respectivement pour D. oliveri et P. erinaceus. Ces valeurs
nettement supérieures à celles trouvées dans la zone
soudanienne plus sèche confirment les travaux de Sinsin et al.,
(2003). L'analyse des mesures des surfaces terrières indicatrices
de l'importance de la biomasse végétale présente, montre
que ces quatre espèces constituent des réserves de moins en moins
importantes en tant qu'habitat (pour les arboricoles surtout), refuges, abris
et / ou sources d'aliments pour les espèces animales en saison
sèche.
6.3.1 Groupement à Daniellia oliveri et
Combretum ghasalense
La distribution par classe de diamètre du groupement
présente une allure en forme de "J" renversé
caractéristique des peuplements jeunes. Cette allure est
différente de celle des populations de D. oliveri qui
évoluent dans le peuplement suivant une loi normale et qui se rapproche
tout de même de la structure diamétrique de l'ensemble du
peuplement compte tenue de l'effectif des jeunes individus. Ce résultat
se démarque quelque peu de celui de Adjahossou (2008) qui conclut que
D. oliveri présente la même caractéristique que le
peuplement dans lequel il évolue. Ce qui du moins se justifie par le
fait que ces résultats sont ceux obtenus dans la zone
protégée (Parc national du W). La mise en défend
réduit les perturbations de l'écosystème. Plus est, cette
espèce subit beaucoup moins de pression anthropique que les trois
autres. Elle est assujettie aux perturbations liées aux feux de brousse
et au surpâturage. 79% des pieds de D. oliveri sont retrouvés
intacts. Selon Sokpon et al., (2001) dans le Complexe Parc National de
la Pendjari, zones cynégétiques de la Pendjari et de l'Atacora
(Région de Konkombri), l'examen de la distribution par classe de
diamètre des forêts claires à D.
47
oliveri révèle une forte proportion de
gros arbres. Ces résultats s'éloignent des nôtres du fait
que tous leurs relevés ont été effectués en
forêts claires.
6.3.2 Groupement à Afzelia africana et
Balanites aegyptiaca
L'analyse de la végétation ligneuse a
montré que Afzelia africana est une espèce peu abondante
dans la zone soudanienne du Bénin. La distribution par classe de
diamètre du groupement présente une même allure en forme de
"J" renversé caractéristique des populations jeunes. Par contre,
A. africana présente une allure de cloche. Ces résultats
s'apparentent intimement aux résultats de Sinsin et al. (2003),
Sokpon & Biaou (2002), Paradis & Houngnon (1997) qui ont trouvé
pour A. africana une allure en « cloche » respectivement
dans les trois zones climatiques (Guinéo-congolaise,
Soudano-guinéen et Soudanienne), dans la forêt classée de
la Lama et dans la forêt classée de Bassila au Bénin. Selon
Ouédraogo et al. (2006), la distribution par classe de
diamètre de A. africana présente une allure en "J" dans
la zone soudanienne du Burkina Faso. Or les allures en "J" et en « cloche
» des distributions en classe de diamètre traduisent des
populations vieillissantes et/ou perturbées. Selon Adjahossou (2008), la
distribution de A. africana au sein des peuplements présente
une allure de "J" renversé caractéristique des populations
jeunes. Les résultats de ces derniers auteurs s'éloignent des
nôtres car la densité des juvéniles n'est pas du tout
élevée dans notre milieu d'étude. La densité de
l'espèce dans les différentes formations est faible. Ces
résultats corroborent celles de Kagoné et al., (2006). Les
populations de A. africana sont dominées à près
de 40% des individus de 20 à 40 cm de diamètre, de moins de 15%
des individus de 10 à 20 cm de diamètre et une très faible
proportion des individus de plus de 40 cm de diamètre. Selon Sokpon
& Biaou (2002), la distribution par classe de diamètre de A.
africana montre une absence des individus de moins de 20 cm de
diamètre. Le milieu d'étude de ces différents auteurs
inclus des zones de pluviométrie plus élevée. La
distribution par classe de diamètre a été
déjà utilisée pour comprendre la dynamique des populations
d'arbres (Cunningham, 2001) et est considérée comme un outil de
prédiction très utilisé (Geldenhuys, 1992). Mais selon
Condit et al. (1998), la distribution par classe de diamètre
n'est pas un bon outil pour prédire la tendance future d'une population
d'arbres. Il renchérit en disant qu'il faut une combinaison de la
distribution par classe de diamètre avec certains paramètres
biologiques tels que le taux de production des juvéniles et le taux de
mortalité annuelle de l'espèce, le tout s'intégrant dans
un modèle. Ainsi un modèle établi à partir de cet
ensemble permettrait de mieux prédire la tendance future des quatre
espèces qui ont fait l'objet de notre étude dans les terres de
parcours en zone soudanienne au Bénin.
6.3.3 Groupement à Khaya senegalensis et
Mimosa pigra
La répartition par classe de diamètre des
ligneux de ce groupement (dbh > 10 cm) (figure 8) présente une allure
en forme de "J" renversé caractéristique des formations
naturelles jeunes polyspécifiques et inéquiennes. Par contre, la
répartition en classe de diamètres des K. senegalensis
(figure 9) au sein de ce peuplement ne répond à aucune loi
normale. On retrouve une proportion élevée de gros individus et
moins de jeunes
48
arbres. Il s'agit d'une population vieillissante,
perturbée et composée de sujets âgés dont le
renouvellement se trouve ainsi hypothéqué.
6.3.4 Groupement à P. erinaceus et A.
leiocarpus
La distribution par classes de diamètre des ligneux de
ce groupement (dbh > 10 cm) (figure 16) présente une allure plus ou
moins oblique. La courbe décroit des classes de faibles diamètres
(10 - 20 cm) vers les classes de diamètres les plus
élevées (plus de 50 cm). Il s'agit d'un peuplement jeune
composé d'individus d'âges multiples. La répartition par
classe de diamètre de P. erinaceus au sein de ces formations
(dbh >10 cm) (figure 17) ne répond à aucune loi. Pterocarpus
erinaceus se trouve ainsi perturbé dans ce peuplement (individus
d'âges multiples) comportant de grandes proportions de petits individus
et moins de gros arbres.
6.4 Analyse de la stratification
La figure 29 (Annexe7) illustre la stratification en hauteur
des quatre ligneux au sein des peuplements. Dans la strate sous arbustive (1 -
4 m) de hauteur, les quatre ligneux sont faiblement représentés
en général mais moyennement représentés dans la
strate arborée (+ de 10m) de hauteur et fortement
représentés dans la strate arbustive (4 - 10 m) de hauteur. Ceci
se justifie par le fait que le nombre de relevés effectués dans
les savanes arbustives est élevé (26 relevés). Au fait, il
est plus facile aux animaux d'exploiter les espèces basiphiles car les
cauliphiles entraînent des leviers de coup nécessitant une
dépense supplémentaire d'énergie. Les formations
végétales des terres de parcours étant très
fréquentées, plus de la moitié de la biomasse se trouvent
nettement plus exploitées par les animaux eux même dans la strate
sous arbustive que dans les strates supérieures. On constate que les
populations de D. oliveri sont beaucoup plus abondantes au niveau de
cette strate. Ceci serait dû au fait que cette espèce soit moins
appétée que les trois autres. De plus, les effets combinés
des feux et du piétinement réduisent la densité et la
croissance de ces espèces à ce stade juvénile. Les 9
relevés des forêts claires, les 6 relevés des forêts
galeries et les 17 relevés effectués dans les savanes
boisées se retrouvent au niveau des strates arbustives et
arborées. A ce stade, les ligneux sont moins exploités par les
animaux et c'est ce qui explique les variations entre les distributions en
hauteur observées au niveau des strates arbustives et arborées.
Cependant, les espèces comme K. senegalensis, P. erinaceus et
A. africana subissent régulièrement des coupes
répétées de leur frondaison. Ceci affecte sensiblement
leur croissance en hauteur.
6.5 Régénération et
reconstitution des espèces cibles
Pour beaucoup d'espèces, la
régénération naturelle, comme c'est le cas de la plus part
des formations végétales tropicales, est faible et la proportion
des essences de valeur dans la régénération
également (Sokpon et al., 2006). La
régénération naturelle (semis et repousses confondus) des
brins et perchis est de 3,04 brins/ha pour K. senegalensis; 3,51
brins/ha pour A. africana; 7,90 brins/ha pour P. erinaceus et
22,63 brins/ha pour D. oliveri.
49
Du point de vue densité, la
régénération est moins importante pour tous les
groupements. Quant aux espèces cibles, elle est très faible et
presque voisine pour les groupements de K. senegalensis (8%) et de
A. africana (9%). Le pourcentage de régénérations
naturelles de P. erinaceus est de 21%. Le pourcentage de
régénération de D. oliveri est nettement plus
élevé (61 %) que l'ensemble des quatre espèces.
L'état hypothéqué de la régénération
naturelle de P. erinaceus, A. africana et de K. senegalensis
peut se justifier par le fait que ces trois essences de valeur subissent
pendant la saison sèche une forte pression de la part des
éleveurs, qui coupent les branches, ce qui compromet dangereusement leur
fructification et leur régénération. Ces résultats
corroborent avec ceux de Houinato (1996) et de Sokpon et al., (2006). Les
formations végétales pâturées de la zone soudanienne
du Bénin sont également la proie des feux de brousse en saison
sèche (Figure 30). C'est surtout la strate sous-arbustive (1 - 4 m) qui
est souvent décimée par les feux. A l'arrivée des
premières pluies, on assiste à des rejets de souches sous forme
de touffes, très appétissantes pour le bétail et la faune
sauvage. Les effets combinés des feux et du surpâturage induisent
une mortalité très élevée lors de la
régénération, compromettant sérieusement le
recrutement des espèces dans le peuplement. La gestion et la
conservation des parcours communautaires en zone soudanienne nécessite
une bonne gestion des feux et l'enrichissement des pâturages par les
ligneux fourragers de choix. En ce qui concerne la densité et la
qualité de la régénération naturelle ; Biaou
(1999), stipule que la reconnaissance de la densité et la qualité
de la régénération naturelle permet de déterminer
les possibilités de reconstitution du peuplement naturel
étudié. Dans la présente étude, la densité
de la régénération naturelle des essences nobles telles
que P. erinaceus, A. africana et de K. senegalensis ne
représente que 13,6% de la régénération naturelle
totale, résultats conformes à ceux de Biaou (1999) et de Koty
(2002).
6.6 Phénologie
De l'analyse de la végétation ligneuse, il en
résulte que A. africana, K. senegalensis et P.
erinaceus sont des espèces peu abondantes dans les terres de
parcours communautaires de la zone soudanienne au Bénin. Cependant,
elles représentent les ligneux fourragers les plus appétés
pendant la période de soudure fourragère. L'exploitation
pastorale de l'espèce se fait par glanage des feuilles et fleurs
tombées à terre au cours de la saison sèche. La succession
dans le temps des phases végétatives et de reproduction des
quatre espèces permet d'affirmer à la suite de Onana (1995)
qu'elles appartiennent au groupe des espèces à feuillaison et
floraison décalées dans le temps. Les feuilles d'A. africana
sont très bien appétées par le bétail et elles
persistent pendant la période de soudure. Une telle phénologie
des quatre espèces présente un intérêt pastoral du
point de vue de l'exploitation optimale des feuilles pendant les
périodes de soudures.
6.7 Analyse des facteurs de Viabilité des
espèces étudiées
L'abondance et la qualité de la
régénération sont analysées en vue de
déterminer les possibilités de reconstitution de K.
senegalensis, de A. africana, de D. oliveri et P.
50
erinaceus. La régénération
naturelle totale de D. oliveri dans le milieu d'étude est
très faible mais reste nettement plus élevée que celle des
trois autres espèces. Au Bénin, selon Babatoundé (2003) la
pression anthropique est à l'origine de la régression de
l'espèce et la principale cause de la faible potentialité de
régénération de D. oliveri serait essentiellement
liée aux conditions défavorables du milieu (facteurs
édaphiques surtout). En effet, les communautés
végétales de l'espèce D. oliveri du milieu
d'étude sont le plus souvent sur des sols à affleurement rocheux.
On remarque sa présence dans les dépressions et quelques plaines
inondables. Au Tchad, selon Darnace (1998) l'utilisation des ressources
floristiques telles que D. oliveri sans connaissance approfondie des
potentialités et des capacités de
régénération, sans projection sur l'avenir est un danger
et les causes potentielles de la disparition de ces ressources sont d'ordre
anthropique.
A. africana et K. senegalensis sont des
espèces menacées de disparition. Selon l'UICN (2000a), ces
espèces sont vulnérables alors qu'elles seraient classées
au Bénin dans la catégorie des espèces en danger critique
de disparition (Adomou, 2005). En effet, les semenciers de K. senegalensis
fortement soumises aux mutilations produisent des fruits de plus petites
que ceux des populations épargnées. Ces résultats
illustrés dans la zone soudanienne sèche par les travaux de Gaoue
et Ticktin (2008) concluent que l'émonde de Khaya senegalensis
a un impact significatif sur la réduction des performances de
reproduction de l'espèce. Ces résultats selon les mêmes
auteurs s'apparentent intimement avec les travaux d'autres travaux qui ont
montré que la défoliation des ligneux peut causer la
réduction du nombre de fruits produit par inflorescence et du nombre de
graines produits par fruits (Obeso, 1993; Ratsirason, Slander & Richard
1996; Flores & Ashton 2000; Endress, Gorchov & Noble 2004 ;
cités par Gaoué, 2008). Ceci est d'autant plus vrai que ces
effets peuvent être causés par la baisse de l'activité
photosynthétique, ou à faible niveau d'utilisation des
réserves nutritives (sève élaborée)
destinées à la croissance et développement.
Plusieurs auteurs tentent de rechercher les causes d'une telle
menace en vue de prendre des dispositions adéquates pour leur
restauration. Selon Ouédraogo et al. (2006), l'aridité
climatique et la pression anthropique sont à l'origine de la
régression de nombreux ligneux en zone soudanienne.
En effet, l'exploitation du fourrage aérien suit le
calendrier phénologique des différentes espèces
fourragères. La principale espèce émondée en milieu
de saison sèche est K. senegalensis. En fin de saison
sèche, les espèces P. erinaceus et A. africana
sont utilisées presque simultanément, mais avec une
légère avance de P. erinaceus. Wala (2004) a
trouvé des résultats similaires dans la zone Tanguiéta -
Batia sur la chaîne de l'Atakora à savoir que K. senegalensis
est disponible et exploitée toute l'année, P. erinaceus
de Décembre à Mai et A. africana de Février
à Avril. De même Petit et al (2001) encore plus
précis montre qu'en zone soudanienne du Burkina -Faso, K.
senegalensis est émondée dès Janvier et jusqu'en Mai
; P. erinaceus de Mars à Mai et A. africana de
Février à Juillet Brisso et al (2007) dans leur
étude dans la forêt de Wari-Maro font également ressortir
que les espèces les plus exploitée sont K. senegalensis,
A. africana et P. erinaceus. La première est
utilisée de Janvier à Février, la seconde de
Février à Mai et la dernière de Février à
Avril.
51
6.7.1 Prélèvement des
organes
Les modes de récolte des ligneux fourragers
pratiqués par les éleveurs ne suivent aucune norme sylvicole et
par conséquent n'assure pas la pérennité des ressources.
Le prélèvement des feuilles par coupes
répétées des branches et le prélèvement des
écorces et des racines couramment utilisées dans la
pharmacopée traditionnelle semblent avoir plus d'incidences
écologiques néfastes que les effets combinés des feux et
du pâturage. Ces biomasses proviennent uniquement des formations
naturelles telles que les forêts, les savanes et les jachères en
reconstitution. Or, dans la plupart des forêts tropicales et pour
beaucoup d'espèces, la densité de la
régénération naturelle et la proportion des essences de
valeur dans la régénération sont souvent faibles (Sokpon
et Biaou 2002, Sokpon et al. 2006). Pour beaucoup d'essences, Sokpon
& al. (2006) ont signalé qu'il existe de faibles
probabilités de reconstitution des peuplements naturels. Face à
ces contraintes, le maintien des principes de la
régénération tels que la pollinisation, le
développement et la dispersion des semences, la germination et la
croissance des plantes semble être compromis.
L'exploitation pastorale de K. senegalensis, P.
erinaceus et A. africana est concurrencée par d'autres
usages socio-économiques telles que consommation humaine, bois de
chauffe, bois d'oeuvre etc. qui les exposent à de fortes pressions
accentuées par l'exploitation pastorale saisonnière se traduisant
par des émondages sévères et répétés
des mêmes sujets jeunes comme adultes.
Il ne fait aucun doute que sans une gestion raisonnable, les
ligneux fourragers soumis à cette exploitation poussée
disparaissent de leur habitat naturel. Ainsi, Cunningham (1991)
considérait que la cueuillette de plantes a des effets perceptibles sur
l'épuisement des ressources forestières. Il serait d'un important
impératif de perenniser la disponibilité de ces ressources
fourragères surtout en faveur des grand ruminants qui doivent assurer
à la fois une production mixte (viande et lait).
Le prélèvement de ces organes se fait selon deux
manières : une forme directe où l'animal broute directement sur
les arbustes et arbrisseaux qui sont à sa portée et une forme
indirecte où les bergers interviennent par émondage ou gaulage
pour mettre les fractions inaccessibles à disposition des ruminants. Ce
constat a été aussi fait par Maïga (2002).
L'émondage se fait selon 03 modes : l'émondage
du houppier à moitié (60%), l'émondage de tout le houppier
avec « sopodo » (10%), l'émondage total du houppier (30%).
L'émondage et le rythme d'ébranchage trop fréquent sont
néfastes pour la plante et réduisent fortement la fonction
chlorophyllienne ; ce qui pourraient être très
préjudiciable pour la survie de cette plante. Cette remarque a
été faite par Cissé (1985) et Bertoli (1987) cités
par Gaoué & Ticktin (2008). L'émondage réduit et/ou
supprime la floraison et ainsi la fructification ce qui compromet toute
possibilité de régénération naturelle des
espèces ainsi exploitée (Wala, 2004). Par contre, Miranda (1989)
pense que la défoliation peut avoir un effet stimulant sur les plantes
comme par exemple Acacia albida.
52
6.7.2 Les espèces
menacées
De nos travaux, il ressort deux catégories
d'espèces menacées à savoir les espèces moyennement
menacées et les espèces fortement menacées. Cette notion
tient du fait que certaines espèces, pour diverses raisons sont devenues
rares dans la région et on ne trouve que quelques pieds dans les
terroirs villageois ; d'autres espèces ont quasiment disparu des
terroirs villageois et ne se retrouvent exclusivement que dans les formations
naturelles où ils sont soumis à une forte pression. Une
espèce à savoir P. erinaceus est moyennement
menacée et deux espèces à savoir K. senegalensis
et A. africana sont fortement menacées. Ces travaux sont
confirmés par ceux de Sinsin et al (1999) au Bénin
où il est retenu que dans la région soudanienne les
espèces menacées sont K. senegalensis, A.
africana, P. erinaceus et cité dans l'ordre
décroissant de menace.
Les causes de la raréfaction de ces espèces sont
multiples. Le développement de la culture du coton a
entraîné une forte croissance des surfaces cultivées. Cette
extension engendra la mise en cause des terres de parcours qui avaient
été prêtées ou données aux éleveurs
Peulh par les premières ethnies à s'être installées
qui sont souvent les Bariba et les Dendi. Les terres ainsi saisies sont
défrichées et cultivées. Les périodes de
jachère se sont fortement raccourcies pour ne durer aujourd'hui que
trois ou quatre années ; ce qui empêche la reconstitution de la
strate ligneuse. Les agriculteurs abattaient systématiquement les
ligneux même fourragers dans leurs champs lors du défrichement. De
plus en plus cette attitude est entrain de changer et on assiste aujourd'hui
à la conservation de quelques pieds de ligneux fourragers, qui constitue
une source importante de fourrage pour leurs bêtes. L'accroissement des
troupeaux de ruminants grâce au suivi vétérinaire et
surtout la forte transhumance des troupeaux étrangers dans la zone
d'étude accentue le déficit de fourrage déjà
très important. De plus, les éleveurs étrangers ont plus
tendance à totalement émonder ces ligneux sans se soucier de leur
conservation. La forte demande en bois d'oeuvre entraîne l'abattage
abusif des ligneux, mais surtout de certains ligneux fourragers tels que K.
senegalensis, P. erinaceus qui sont très prisés.
Aussi la demande en bois de feu et en charbon de bois sans cesse croissante
occasionne l'utilisation des ligneux appétés tels que
Balanites aegyptiaca, Burkea africana, Prosopis
africana. Enfin, la médecine traditionnelle
vétérinaire et humaine engendre l'utilisation d'organes (racine,
écorce, feuille) dont le prélèvement est parfois
très préjudiciable pour la plante.
6.7.3 Efficacité des méthodes de
conservation endogène des plantes dans la zone
d'étude
Nos investigations au cours de nos travaux ont
révélé que sur les quatre espèces ligneuses
fourragères étudiées, comme pour la plupart des autres
espèces fourragères rencontrées, aucune d'entre elles n'a
fait l'objet de plantation. Celles qui sont domestiquées sont en fait
des essences colonisatrices (herbacées) cultivées ou (ligneuses)
laissées dans les champs, concessions pour les multiples usages qu'elles
offrent notamment dans le traitement des maladies. A l'issue des entretiens
directs que nous avions eus avec les différents groupes socioculturels,
le paludisme a été souvent cité comme la maladie la plus
courante traitée par les espèces de plantes conservées
(67% des recettes) suivi des maladies gynécologiques (23% des recettes).
On peut citer
53
entre autres Cymbopogon citratus, Argemone mexicana,
Ocimum sp, Cassia occidentalis, Momordica charantia, Mangifera indica,
Citrus sp, Azadirachta indica, Carica papaya, Parkia biglobosa, Vitellaria
paradoxa, Adansonia digitata, Uvaria chamae, Jatropha curcas, Commiphora
africana, Crateva religiosa, Phyllantus amarus, Hibiscus asper, Albizia
ferruginea, Albizia zygia, Cassia rotundifolia, Argemone mexicana, Tamarindus
indica Pavetta crassipes, Sarcocephallus latifolius, Cienfuegosia leptoclada
(Annexe 7). La contribution aux études ethnobotaniques et
floristiques menée au Bénin par Adjanohoun et al.,
(1989) présente 29 recettes sur le paludisme, obtenues à
partir de 37 espèces de plante.
Outre ces formes de conservation précaires, aucune
stratégie rationnelle de domestication et d'entretien de la flore n'est
mise au point. Ce qui pourrait avoir des conséquences
irréversibles sur la survie des plantes médicinales même si
une étude réalisée par le WWF et al. (1993)
montre que dans la plupart des pays africains, la cueillette pour les besoins
locaux n'est pas dommageable pour la survie des plantes en estimant que la
quantité récoltée est faible.
Cependant, il convient de souligner l'effort de conservation
des populations locales qui dors et déjà prennent eux même
l'initiative de conserver in situ, les arbres devenus rares dans la nature.
Leurs choix sont généralement portés sur les
espèces qui présentent un intérêt commercial,
médicinal, agronomique ou, écologique. C'est ce qui justifie la
densité élevée de Parkia biglobosa, de
Vitellaria paradoxa, de Tamarindus indica dans les champs et
jachères. Cette pratique également observée au
Sud-Bénin (Sokpon,1994), constitue une première démarche
à saisir pour la mise en oeuvre d'une stratégie de conservation
et de domestication des nombreuses essences de valeur.
6 .8 Les possibilités de gestion optimales des
ligneux
La gestion durable des ligneux fourragers en zone soudanienne
du Bénin passe inconditionnellement par une augmentation de la
production primaire. Mais, cette production primaire se heurte aux
difficultés liées aux types de parcours en zone soudanienne du
Bénin. Les systèmes communautaires des parcours soudaniens
béninois sont publics. Ce qui constitue invariablement un facteur
défavorable à une gestion rationnelle puisque l'accès des
animaux aux ressources communes ne peut en pratique être
contrôlé, donc géré. Par ailleurs, la gestion des
parcours contribue à modifier l'équilibre et la
compétition entre les espèces végétales (Le
Houérou, 2005). Ceci se justifie par le fait que des défoliations
fréquentes et répétées par les herbivores
réduisent ou empêchent l'accumulation de réserves, la
densité et l'élongation racinaire chez les espèces
recherchées par les animaux. Ces espèces sont donc soumises
à un désavantage sélectif vis-à-vis de leurs
concurrentes moins recherchées ou négligées des
herbivores. Il s'ensuit que les végétations soumises à une
exploitation continue pendant de longues durées tendent soit à
être dominées par des espèces délaissées des
herbivores soit à la disparition de toutes les espèces
pérennes. Pour palier à l'épuisement des ressources en
ligneux fourragers, il faudra donc envisager la plantation de réserves
d'arbres et d'arbustes fourragers palatables (Khaya senegalensis, Afzelia
africana, Pterocarpus erinaceus etc.) à des points
stratégiques comme les points d'eau dans les terres de parcours. Cette
gestion visant à favoriser le développement des arbres et
arbustes fourragers spontanés riches en protéine doit
être
54
soutenue par des interventions dans la fréquence et
l'intensité des défoliations des espèces
fourragères de manière à réduire le
désavantage et les traumatismes qu'engendrent le broutage et les coupes
anarchiques. Cette mesure permettra de respecter la physiologie du
développement et de la reproduction des espèces
désirables. Mais, cet objectif ne peut être atteint que par la
mise en oeuvre des mesures restrictives concernant le taux de charge
(Intégration de la charge par unité de surface en un temps T, sur
période donnée) et la capacité de charge (quantité
de bétail que peut supporter le pâturage sans se
détériorer). La gestion durables des ligneux fourragers fait donc
appel au rôle central de l'arbre dans les systèmes agropastoraux
ainsi qu'au suivi permanent des pâturages et à la conservation de
toutes leurs ressources naturelles.
7. CONCLUSION GENERALE ET PERSPECTIVES
La zone soudanienne du Bénin demeure encore très
viable, mais fragile compte tenue de la vitesse de dégradation du milieu
naturel sous l'effet de la pression anthropique. Elle est appelée
à jouer un rôle important dans l'intensification agropastorale.
L'étude de la gestion et de la conservation des ligneux fourragers dans
les terres de parcours en zone soudanienne au Bénin ont permis
d'étudier les caractéristiques dendrométriques, la
distribution et l'état actuel des populations de K.
senegalensis, D. oliveri, P. erinaceus et A.
africana choisis en raison de leur intérêt fourrager. Cette
étude conclut que les terres de parcours communautaires parcourues dans
la zone soudanienne du Bénin renferment environ 123 espèces.
Cette zone est assez riche en espèces ligneuses et leur utilisation a
été très diversifiée. La transhumance reste un
problème épineux à résoudre. Les éleveurs et
leurs animaux passent partout et séjournent dans la zone et même
dans les aires protégées. Leur présence anarchique est
surtout nuisible à la survie de nombres d'espèces. P.
erinaceus, A. africana et K. senegalensis sont les trois
principales essences recherchées par les éleveurs pour leurs
animaux. L'analyse des 60 relevés phytoécologiques ont permis de
constituer quatre grands groupements végétaux dominés par
chacune des quatre espèces ciblées à savoir :
- Le groupement à Khaya senegalensis lannea acida
et Mimosa pigra,
- Le groupement à Pterocarpus erinaceus, Anogeissus
leiocarpa et Vitellaria paradoxa,
- Le groupement à A. africana et Balanites
aegyptiaca ;
- Le groupement à D. oliveri, Mimosa pigra
et Combretum ghasalense
L'analyse des paramètres de diversité montre que
l'espèce D. oliveri, présente une large distribution
avec spécialisation dans les formations temporairement inondées
et les zones de dépression. L'espèce P. erinaceus
présente une large distribution sur tout types de sol.
L'espèce A. africana, végète dans des stations
plus ou moins stables avec une répartition irrégulière.
Quant à l'espèce K. senegalensis, elle est
retrouvée dans les galeries et préfère souvent les sols
profond et bien drainés.
L'analyse de la distribution par classe de diamètre de
D. oliveri, A. africana et de P. erinaceus et la comparaison
des diamètres moyens dans une même zone climatique montrent que
les populations des individus de dbh = 10 cm sont encore très jeunes
dans les terres de parcours communautaires de la zone d'étude avec un
faible taux de
55
sujets de dbh = 50 cm contrairement à l'espèce
K. senegalensis composée essentiellement de vieux sujets. Par
ailleurs, la régénération naturelle des espèces est
beaucoup plus faible pour l'espèce Khaya senegalensis pour une
hauteur inférieure à 1,30 m et presque nulle pour une hauteur
comprise entre [30 cm ; 1,30 m [ . Pour les trois autres espèces, la
régénération est faible mais D. oliveri conserve
un taux de régénération nettement plus
élevé. Les causes probables de cette situation sont notamment le
feu d'aménagement et les facteurs zoogènes à
l'échelle locale. A l'échelle spatiale plus grande, elles peuvent
être assimilées aux changements climatiques. Cette diagnose impose
donc la mise en oeuvre urgente pour gérer et conserver ces parcours
communautaires.
Du point de vue phytogéographique,
l'élément base soudanien (S) et les
espèces Soudano-zambéziennes sont les plus abondantes. Par
contre, les espèces à large distribution sont faiblement
représentées justifiant un faible impact des actions
anthropiques. Toutefois en dépit des efforts considérables
déployés pour le maintien des écosystèmes en
exploitation raisonnée, des parcours naturels de la zone soudanienne
malgré leur originalité demeurent encore perturbés par la
transhumance pastorale au cours des périodes critiques de
l'année. Spécifiquement, l'efficacité de la gestion et de
la conservation des ligneux fourragers des terres de parcours en zone
soudanienne indique que cinq actions majeures doivent être
engagées : elles consistent en:
- une intégration de l'arbre dans les systèmes
agropastoraux (agroforesterie) par des essais d'enrichissement des peuplements
naturels des terres de parcours de la zone d'étude. Ceci se fera
grâce au test d'aptitude des quatre ligneux étudiés
à la technique des macro boutures et des micro boutures pour la
domestication et la régénération naturelle
assistée,
- une étude permanente des bases techniques pour une
gestion durable de K. senegalensis, A. africana, P. erinaceus, D.
oliveri, et de la domestication de tous les ligneux fourragers dans les
terres de parcours de la zone d'étude,
- un contrôle de l'impact des feux d'aménagement
(feu précoce, feu tardif, feu de contre saison, protection
intégrale) sur la survie des différentes classes de
régénération,
- une intégration dans les programmes de gestion, des
programmes de conservation in situ soit par réhabilitation ou
restauration du ou des arboreta, soit par création des jardins
botaniques ou arboretum communautaire et,
- un contrôle continu et à la poursuite des
recherches écologiques à long terme pour appréhender les
phénomènes globaux (évolution climatique,
désertification et dégradation du milieu) et le rôle de ces
ligneux fourragers dans la séquestration du carbone pour une prise de
décision dans la gestion durable des ressources renouvelables.
- un contrôle sévère de la transhumance
transfrontalière et de la transhumance nationale dans une
démarche partenariale avec les communautés par la viabilisation
des couloirs de passage d'animaux d'une part et d'autre part, la stabilisation
et l'intensification de l'élevage local à travers la construction
des parcs d'élevage autour des points d'eau.
56
REFERENCES BIBLIOGRAPHIQUES
1. Adjahossou S. G. C.,
(2008). Etude phytoécologique des savanes
soudano - sahéliennes et caractéristiques dendrométriques
de Afzelia africana, Daniellia oliveri et Anogeissus
leiocarpus : Cas de la RTB -W commune de Karimama. Mémoire
d'ingénieur des travaux. Université d'Abomey - calavi,
Bénin. 65p.
2. Adjanohoun E., Adjakidjè V., Ahyi M.R.A.,
Aké Assi L., Akoègninou A., Dalmeda J., Akpovo F., Boukef K.,
Chadaré F., Cusset G., Dramane K., Eyme J., Gassita J-N, Gbaguidi N.,
Goudoté E., Guinko S., Houngnon P., Issa L., Keita A., Kiniffo H.V.,
Koné Bamba D., Musampa Nseyya A., Saadou N., Sodogandji TH., De Souza
S., Tchabi A., Zinsou Dossa C., Zohoun Th., (1989). Contribution
aux études ethnobotaniques et floristiques en République
Populaire du Bénin. ACCT. Paris. 895p.
3. Adomou C.A., (2005). Vegetation Patterns
and Environmental gradients in Benin. Implications for biogeography and
conservation. PhD Thesis Wageningen University, Wageningen. 219p + annexes.
4. Agri : site lagr., (1999). Ordonnance sur
la terminologie : Estivage. Journal Agr. http:/
www.hebdo.ch.
5. Ahouangonou S., Bris B., (1997).
Afzelia africana. Le Flamboyant n° 42: 7-10.
6. Akoègninou A., Van der Burg W. J., Van der
Maesen L. J. G., Adjakidjè V., Essou J. P., Sinsin B., Yèdomonhan
H., (2006). Flore Analytique du Bénin. 1034p.
7. Arbonnier M., (2002). Arbres arbustes et
lianes des zones sèches d'Afrique de l'Ouest. CIRAD-MNHN. 573p.
8. Aubreville A., (1937). Les forêts
du Dahomey et du
Togo. Bull. com. Etu.
His. Et Scientif. 20: 1-112pp.
9. Aubreville A., (1950). Flore
forestière soudano-guinéenne. Orstom, Paris., 533p.
10. Babatoundé L. L., (2003).
Potentialités et usages d'une plante des savanes
guinéennes: Daniellia oliveri (Rolf) Huch et Dalz. Dans la
Sous-Préfecture de Zogbodomè. Mémoire de maîtrise de
Géographie. FLASH/UAC. Bénin. 69p.
11. Brisso N., Houinato M., Adandédjan C., and
Sinsin B., (2007). Dry season woody fodder productivity in savannas.
Short communication. Ghanaian Journal of Animal Science, Vol.2
& 3 N°1: 181-185.
12. Condit R., Sukumar S. R., Hubbell R., Foster R.
B., (1998). Predicting population trends from size Distributions: A
direct test in tropical tree community, 4 (152) The
American Naturalist. 495-509pp.
13. Cunningham A. B., (1991). Development of
the conservation policy and commercially exploited medicinal plant. In
Akerele O., Heywood V. and H. Syge, 337- 354.
57
14. Cunningham A. B., (2001). Applied
ethnobotany. People wild plant use and conservation. People and plants
conservation. Earthscan Publications Ltd London. 300p.
15. Cuny P., Sanoga S., (1998).
Espèces forestières locales: Phénologies et semis
direct en zone soudanienne (Sud Mali). Le Flamboyant N° 47:
26-30pp.
16. Dajoz R., (1985). Précis
d'écologie. Troisième édition Bordas, Paris. 549p.
17. Darnace A. B., (1998). Durabilité
des usages actuels des resssources biologiques. Tchad. 49p.
http//bch-cbd..natural
science.be./Tchad.
18. Devineau J. L., Lecardier C., Vuattoux R.,
(1984). Evolution de la diversité spécifique du
peuplement ligneux dans une succession pré forestière de
colonisation d'une savane protégée des feux (Lamto, Côte
d'Ivoire). Candollea 39 ; 103-134pp.
19. Devineau J.L., (2001). Les
espèces ligneuses indicatrices des sols dans les savanes et
jachères de l'Ouest du Burkina Faso. Phytocoenologia. 31 :
325-351.
20. Di Silvestre I., Sinsin B., Daouda I., (2003).
Mission sur les grands et moyens mammifères menacés dans
les Parcs Nationaux et Zones Cynégétiques du Bénin
CENAGREF, Cotonou, Bénin. 74p + annexes.
21. Diop M., Kaya B., Niang A., & Olivier A.,
(2005). Les espèces ligneuses et leurs usages : les
préférences des paysans dans le Cercle de Ségou, au Mali.
ICRAF Working Paper no. 9. Nairobi : World Agroforestry Centre. 29p.
22. Eyog Matig O., Gaoué O.G., & Dossou
B., (2002). Réseaux "Espèces Ligneuses Alimentaire".
Compte rendu de la première réunion du Réseau tenue du
11-13 décembre 2000 au CNSF Ouagadougou, Burkina Faso 241: Institut
International des Ressources Phytogénétiques. 241p.
23. Fagnissè F. B.,
(2006).Valorisation des plantes médicinales dans le traitement
des ruminants (cas des bovins) aux alentours du Parc W: Inventaire
Ethnobotanique et Perspectives. Mémoire d'Iingénieur des Travaux
(DIT). EPAC/UAC. Bénin. 114p + annexes.
24. Fall - Touré S., (1993). Valeur
nutritive des ligneux, leur rôle dans la compréhension des
fourrages pauvres des milieux tropicaux. Doct. ESA, Montpellier, 139p.
25. FAO., (2004). Forest Genetic Resources.
Food and Agriculture Organisation of the United Nations, Rome.
26. Gaoue, O.G., & Ticktin,T.,
(2008). Impacts of bark and foliage harvest on
Khaya senegalensis (Meliaceae) reproductive performance in Benin.
Journal of Applied Ecology, 45, 000 - 000
27. Geldenhuys C. J., (1992). The use of
diameter distributions in sustained use management of forest: example from
Southern Africa. Paper presented at the SAREC Zimbabwe Forestry Commission.
Symposium: Ecology and Management of Indigenous Forest in Southern Africa,
Victoria Falls.
28.
58
Gounot M., (1969). Techniques quantitatives
d'étude de la végétation. Masson
29. Grouzis M., Sicot M.,(1980). Une
méthode d'étude phénologique des populations
d'espèces ligneuses sahéliennes. Influences de quelques facteurs
écologiques. In : Le Houérou (H.N.) éd, les fourrages
ligneux en Afrique : état actuel des connaissances.
Addis-Abéba, Ethiopie, 8-12 avril 1980, CIPEA, p.231 - 237.
30. Houéhanou T. D., (2008).
Caractéristiques dendrométriques de Afzelia africana
Smith ex Pers. et Anogeissus leiocarpa (DC) Guill. & Perr.
Comme indicateur d'appréciation de leur conservation dans la
réserve de biosphère de la Pendjari et les utilisations
ethno-médecinales. Mémoire de Diplôme d'études
Approfondies (DEA). Option : Aménagement et Gestion des Ressources
Naturelles (AGRN). FSA/UAC. Bénin. 51pages+ annexes.
31. Houinato M. R. B., (2001).
Phytosociologie, écologie, production et capacité de
charge des formations végétales pâturées dans la
région des Monts Kouffè Bénin. Thèse de doctorat.
Université Libre de Bruxelles, Belgique. 225p.
32. Houinato M., & Sinsin B., (2000). La
pression agropastorale sur la zone riveraine de la Réserve de
Biosphère de la Pendjari. Tropicultura, 18 (3) :
112-117
33. Houinato M., & Sinsin B.,
(2001a) Analyse phytogéographique de la
région des Monts Kouffé au Bénin. Syst. Georgr. Pl. 71 :
889-910.
34. Houinato M., & Sinsin B., (2001b)
Impact des feux de brousse sur la dynamique des communautés
végétales dans la forêt de Bassila (Bénin). Acta
Bot. Gallica 148 (3) : 237-251.
35. Houinato M., (1996).
Aménagement Agro-pastoral de la zone riveraine de la Réserve de
la Biosphère de la Pendjari. Rapport principal, Laboratoire d' Ecologie
Appliquée FSA/UNB, 54p.
36. Houinato M., (1999)- Contribution
à l'étude de la pharmacopée populaire dans la
région des Monts Kouffé, Bénin. Rapport de mission
d'étude. FSA / UNB. Cotonou, Bénin. 35p.
37. Houinato M., (2002). Rapport de Mission
d'étude de la végétation (aspects ethnobotaniques) du
complexe écologique du parc W, LEA/FSA, RB, UE, ECOPAS, 34p.
38. Houinato M., Kagone H., & Sinsin B., (2003).
Seasonal cycle of herbaceous plant phytomass of Monts Kouffe region in
Benin. African journal of Range & Forage Science, 2003, 20 (2): 175-193
39. Houngnon P., (1996). Etude des
espèces végétales de la zone cynégétique de
la Djona. Rapport PGRN 52p.
40. htt://
www.worldagroforestry.org/eca/downloads/ECAstrategicplan.doc3/8/2005
41. Ibouraïma S.,
(2005)- Dynamique, bilan et impact de la sédimentation
intra-cuvette : cas du département de l'Alibori.Thèse de Doctorat
(unique) en Gestion de l'Environnement,Université d'Abomey-calavi,
199p.
42.
59
ICRAF - ECA (2005). Strategic Plan for ICRAF
- ECA Programme 2000-2010.
43. ICRAF., (2005). Utilisation and
Commercialisation of dryland indigenous fruit tree species to improve
livelihoods in East and Central Africa. Proceedings of a Regional workshop,
KEFRI Kitui, Kenya 20-24 June 2005. ICRAF - ECA, ECA Working Paper No. 7, 2005.
World Agroforestry.
44. Kagone H., Obulbiga M. F.,
Zoungrana-Kaboré C. Y., Idogo S., (2006) Etude d'un
système d'exploitation adapté aux pâturages naturels en
zone Nord - Soudanienne du Burkina Faso. Bull Anim. Hlth. Prod. Afr. 54:
35 - 42
45. Kagone H., Toutain B., Dulieu D., Houinato M.,
Boureima A., & Nocker U., (2006). Pastoralisme et aires
protégées en Afrique de l'Ouest : du conflit à la gestion
concertée de la transhumance transfrontalière dans la
région du parc régional W (Bénin, Burkina Fasso, Niger).
Bulletin of Animal Health and Production in Africa Vol. 54 (1) 2006 : 43-52
46. Kent M., Coker P., (2000). Vegetation
Description and Analysis. A Practical Approach. Belhaven Press, London.
47. Kokou K., Sokpon N., (2006). Les
forêts sacrées du couloir du Dahomey. Bois et forêts des
tropiques, 288 (2) : 15 - 23
48. Koty H. I., (2002)- Contribution
à l'aménagement participatif de la forêt classées
des Monts Kouffé sur base des Produits Forestiers Non Ligneux (PFNL) :
inventaire, importance socio-économique, structure et dynamique et
stratégie de conservation durable de ces produits. Mémoire de fin
d'étude, DAGE, FSA, UAC, Cotonou, 256p.
49. Kpéra G. N.
(2008). Statut des crocodiles et Influence des facteurs
physico-chimiques de l'eau sur la répartition des crocodiles dans la
Réserve de Biosphère Transfrontalière du W du
Bénin. DEA, FSA/UNB. 68p.
50. Lalèyè P. Chikou A. et
Vandewalle P. (2001). Faune itchyologique de la rivière
Pendjari au Bénin. Rapport de mission. LHA/FSA/UAC, Bénin.
15p.
51. Le Houerou H.N., (1980). Browse in Africa,
the current state of knowledge. ILCA, région sèche.
Sécheresse 16 (2) : 89-96
52. Le Houerou H.N., (2005).
Problèmes écologiques du développement de
l'élevage en Addis Abbaba, 491p.
53. Maïga A., (2002). Sylvopastoralisme
dans le sahel occidental du Mali : Modes d'alimentation des petits ruminants
par les espèces ligneuses. Projet Dév. Elev. Sahel. Occid. (PRO.
DE. SO.) p 131-134
54. Miranda R., (1989). Rôle des
ligneux fourragers dans la nutrition des ruminants en Afrique
subsaharienne. Etude bibliogarphique Monographie n°7. CIPEA, Addis -
Abeba (Ethiopie).
55. Onana J., (1995). Les
ligneux fourragers du Nord - Cameroun. I. Inventaire et phénologie.
Revue Elev. Méd. Vét. Pays trop., 48
(2) : p213 - 219.
56.
60
Ouédraogo A., Thiombiano A., Hahn-Hadjali K.,
& Guinko S., (2006). Diagnostic de l'état de
l'état de dégradation des peuplements de quatre espèces
ligneuses en zone soudanienne du Burkina Faso. Secheresss 17
(4) : 485-491pp.
57. Paradis G., et Houngnon P., (1997). La
végétation de l'aire classée de la Lama dans la
mosaïque forêt-savane du Sud-Bénin (ex Sud-Dahomey). Bulletin
du Museum Nationale d'Histoire Naturelle, 3e serie N°503.
nov-déc. 1997. Botanique 34p.
58. Petit S., (2000). Fourrage ligneux et
parcours des troupeaux des éleveurs Peulh de l'Ouest Burkinabé.
Notes de recherche. Bois et forêts des tropiques. N°253 (3)
59. Sidi L., (1996) : Etude des facteurs
pastoraux dans les zones d'actions prioritaires, Projet de Restauration
des Ressources Forestières dans la région de Bassila,
Bénin.
60. Sinsin B., (1993). Phytosociologie,
écologie, valeur pastorale, production et capacité de charge des
pâturages naturels du périmètre Nikki-Kalalé au
Nord-Bénin. PhD. Thesis Université Libre de Bruxelles, Belguim,
396p.
61. Sinsin B., (1999). Description
floristique, structure et écologique des associations
végétales du domaine soudanien du Nord-Bénin. Document
phytosociologiques. Vol. XIX : 311-335.
62. Sinsin B., Eyog Matig O., Assogbodjo A.E.,
Gaoué O.G., & Sinandouwirou T., (2004). Dendrometric
characteristics as indicators of pressure of Afzelia africana Sm. Dynamic
changes in trees found in different climatic zones of Benin.
Biodiversity and conservation, 13: 1555-1570.
63. Sinsin B., Oloulotan S., Oumorou M.,
(2006). Les pâturages de saison sèche de la zone
soudanienne du Nord-Est du Bénin. Rev. Elev.
Méd. Vét. Pays trop., 42 (2) : 283 - 285.
64. Sinsin B., Sinandouwirou Th., & Assogbadjo
A., (2000). Caractérisation écologique de deux
essences fourragères du Bénin: Khaya senegalensis (desv) et
Afzelia africana sm. Rapport d'étude IPGRI/SAFORGEN, Cotonou,
Bénin.
65. Sokpon N., (1994)- Tenure
foncière et propriété des ligneux dans les systèmes
agroforestiers traditionnels au Bénin. Ann. Fac. Sc., Kisangani, n°
spécial pp.115-122.
66. Sokpon N., (2001). Inventaire et
caractérisation des formations végétales du Complexe Parc
National de la Pendjari, zones cynégétiques de la Pendjari et de
l'Atacora (Région de Konkombri), 48p + annexes.
67. Sokpon N., Biaou H. S., (2002). The use
of diameter distribution in sustained-use management of remnant forests in
Benin: case of Bassila forest reserve in North Benin. Forest Ecology and
Management, 161: 13-25.
68. Szaniawsky H., (1982).
Développement des Parcs Nationaux : Plan directeur du Parcs
National W du fleuve Niger Bénin. Rapport technique 3. PNUD/FAO, Rome,
98p.
69. Tchabi V., (1994) - Evaluation et
utilisation des ressources pastorales dans la réserve de faune
soudanienne de la Pendjari (Bénin). Thèse Univ. Montpellier
III, 208 p.
70.
61
Tchiwanou M., (1999) Plan forestier national
au Bénin. Rapport PGRN 127 p.
71. Toko I., (2008) - Etude de la
variabilité spatiale de la biomasse herbacée, de la
phénologie et de la structure de la végétation le long des
toposéquences du bassin supérieur du fleuve Ouémé
au Bénin. Thèse de Doctorat unique, Université
d'Abomey-calavi. 241 p + annexe.
72. UICN., (2000a). Red list of threatened
spicies. Website:
http://www.redlist.org.
73. UICN., (2000b). Red list of threatened
spicies. Compiled by craig Hilton-Taylor. The World Conservation Union. 38p.
74. Viennot M., (1978). Notice explicative
de la carte pédagogique de reconnaissance de la R.P. Bénin.
Feuille de Kandi-Karimama. ORSTOM, Paris. 47p.
75. Wala K., (2004). La
végétation de la chaîne de l'Atakora au Bénin :
Diversité floristique, phytosociologie et impact humain. Thèse de
l'Université de Lomé. FDS/UL. p 105-115
76. White F., (1986). La
végétation d'Afrique. Mémoire accompagnant la carte de la
végétation d'Afrique. UNESCO/AETFAT/UNSO, ORSTOM-UNESCO, 384p.
77. WWF., IUCN., et OM., (1993) Principes
directeurs pour la conservation des plantes médicinales. Switzerland.
35p.
78. Zoungrana - Kaboré C.Y., (1995).
Composition chimique et valeur nutritive des herbacées et
ligneux des pâturages naturels soudaniens et des sous produits existants
au Burkina Faso. Doct. Es Sc. Nat., Univ. Ouagadougou,224p.
79. Zoungrana B., Adoum A., Ajavon Y., (2008).
Mission conjointe d'évaluation préliminaire des
récoltes 2008 - 2009. MAEP - Bénin, CILSS, FEW NET, 18p.
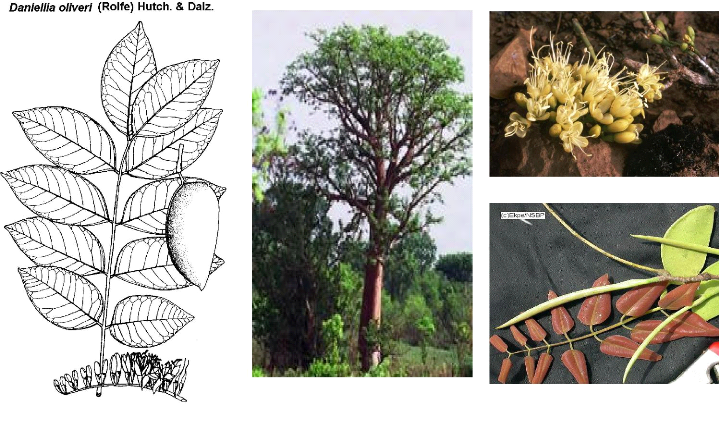
Photo 2 : Inflorescences de Daniellia
oliveri
Photo 1 : Sujet adulte de Daniellia
oliveri
Source : Adjanohoun et al. 1989 Source
:
www.pendjari.net
Photo 3 : Jeunes Feuilles de Daniellia
oliveri
62
ANNEXES 1 : Liste des photos des espèces
ligneuses fourragères étudiées
Khaya senegalensis (Desv.) A. Juss.

Photo 5 : Fruits de K.
senegalensis(Gaoué)
Photo 6 : Jeune sujet de K.
senegalensis
|
Photo 4 : Sujet adulte de Khaya
senegalensis
|
Photo 7 : Population de K. senegalensis (O.
Gaoué 2006)
|
63
|
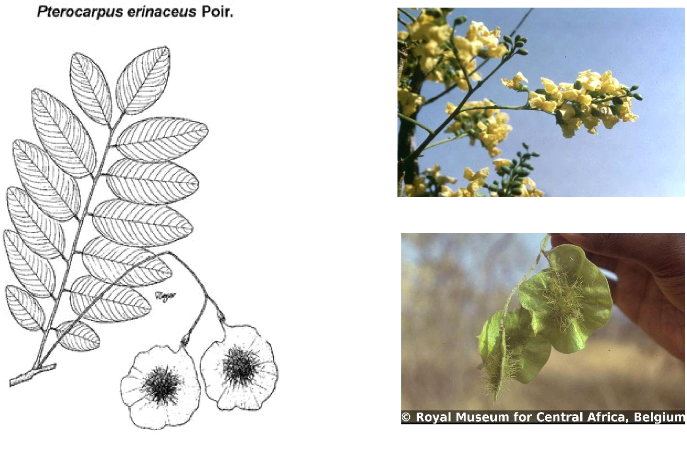
64
Source : Adjanohoun et al. 1989
Photo 9 : Fruits de Pterocarpus
erinaceus
Photo 8 : Fleur de Pterocarpus
erinaceus

Photo 10 : Inflorescences de Afzelia africana
Source :
http://www.agora.gouv.fr
65
Source : Adjanohoun et al.
|
Photo 11 : Fruits de Afzelia africana A.
HOUNGNON (2008)
|
Photo 12 : Un individu de Afzelia
africana
|
66
Annexe2: Liste des espèces végétales
des terres de parcours communautaires soudaniens(Bénin)
Familles Espèces
Mimosaceae Acacia ataxacantha
Mimosaceae Acacia dudgeoni
Mimosaceae Acacia gourmaensis
Mimosaceae Acacia hockii
Mimosaceae Acacia macrostachya
Mimosaceae Acacia macrothyrsa
Mimosaceae Acacia pennata
Mimosaceae Acacia polyacantha
Mimosaceae Acacia seyal
Mimosaceae Acacia sieberiana
Mimosaceae Acacia tortilis
Bombacaceae Adansonia digitata
Caesalpiniaceae Afzelia africana
Mimosaceae Albizia glaberrima
Mimosaceae Albizia zygia
Vitaceae Ampelocissus grantii
Annonaceae Annona senegalensis
Combretaceae Anogeissus leiocarpus
Liliaceae Asparagus africanus
Meliaceae Azadirachta indica
Balanitaceae Balanites aegyptiaca
Bombacaceae Bombax costatum
Arecaceae Borassus aethiopum
Euphorbiaceae Bridelia ferruginea
Caesalpiniaceae Burkea africana
Bombacaceae Ceiba pentadra
Vitaceae Cissus populnea
Sterculiaceae Cola laurifolia
Combretaceae Combretum collinum
Combretaceae Combretum ghasalense
Combretaceae Combretum glutinosum
Combretaceae Combretum hypopilinum
Combretaceae Combretum lecardii
Combretaceae Combretum micranthum
Combretaceae Combretum nigricans
Combretaceae Combretum paniculatum
Combretaceae Combretum sp
Burseraceae Commiphora africana
Capparidaceae Crateva adansonii
Rubiaceae Crossopteryx febrifuga
Araliaceae Cussonia arborea
Caesalpiniaceae Daniellia oliveri
Caesalpiniaceae Detarium microcarpum
67
Mimosaceae Dichrostachys cinerea
Ebenaceae Diospyros mespiliformis
Arecaceae Elaeis guineensis
Mimosaceae Entada africana
Fabaceae/Papilionaceae Erythrina senegalensis
Rubiaceae Feretia apodanthera
Moraceae Ficus capensis
Moraceae Ficus gnaphalocarpa
Moraceae Ficus ingens
Moraceae Ficus platyphylla
Moraceae Ficus sp
Flacourtiaceae Flacourtia indica
Rubiaceae Gardenia aqualla
Rubiaceae Gardenia ternifolia
Tiliaceae Grewia bicolor
Tiliaceae Grewia flavescence
Tiliaceae Grewia lasiodicus
Tiliaceae Grewia mollis
Combretaceae Guiera senegalensis
Anacardiaceae Haematostaphis barteri
Simaroubaceae Hannoa undulata
Annonaceae Hexalobus monopetalus
Hymenocardiaceae Hymenocardia acida
Caesalpiniaceae Isoberlinia doka
Caesalpiniaceae Isoberlinia tomentosa
Meliaceae Khaya senegalensis
Bignoniaceae Kigelia africana
Anacardiaceae Lannea acida
Anacardiaceae Lannea barteri
Anacardiaceae Lannea microcarpa
Anacardiaceae Lannea velutina
Sapindaceae Lecaniodiscus cupanioides
Fabaceae/Papilionaceae Lonchocarpus cyanescens
Fabaceae/Papilionaceae Lonchocarpus sericeus
Ochnaceae Lophira lanceolata
Rubiaceae Macrosphyra longistyla
Anacardiaceae Mangifera indica
Chrysobalanaceae Maranthes polyandra
Celastraceae Maytenus senegalensis
Fabaceae/Papilionaceae Millettia thonningii
Mimosaceae Mimosa pigra
Rubiaceae Mitragyna inermis
Dipterocarpaceae Monotes kerstingii
Opiliaceae Opilia celtidifolia
Fabaceae/Papilionaceae Ostryoderris stuhlmannii
Chrysobalanaceae Parinari curatellifolia
68
Mimosaceae Parkia biglobosa
Rubiaceae Pavetta crassipes
Fabaceae/Papilionaceae Pericopsis laxiflora
Caesalpiniaceae Piliostigma reticulatum
Caesalpiniaceae Piliostigma thonningii
Mimosaceae Prosopis africana
Meliaceae Pseudocedrela kotschyi
Hypericaceae Psorospermum febrifugum
Combretaceae Pteleopsis suberosa
Fabaceae/Papilionaceae Pterocarpus erinaceus
Fabaceae/Papilionaceae Pterocarpus santalinoides
Anacardiaceae Rhus natalensis
Apocynaceae Saba senegalensis
Rubiaceae Sarcocephalus latifolius
Polygalaceae Securidaca longepedunculata
Euphorbiaceae Securinega virosa
Apiaceae Steganotaenia araliaceae
Sterculiaceae Sterculia setigera
Bignoniaceae Stereospermum kunthianum
Apocynaceae Strophantus sarmentosus
Loganiaceae Strychnos innocua
Loganiaceae Strychnos spinosa
Caesalpiniaceae Tamarindus indica
Combretaceae Terminalia avicennioides
Combretaceae Terminalia glaucescens
Combretaceae Terminalia macroptera
Meliaceae Trichilia emetica
Asteraceae Vernonia colorata
Sapotaceae Vitellaria paradoxa
Verbenaceae Vitex chrysocarpa
Verbenaceae Vitex doniana
Olacaceae Ximenia americana
Rhamnaceae Ziziphus amphibia
Rhamnaceae Ziziphus mauritiana
Annexe 3: Liste des espèces du Groupement
à
Khaya senegalensis et Mimosa
pigra
(Forêt galerie)
Familles Espèces
Anacardiaceae Lannea acida
Apocynaceae Saba senegalensis
Apocynaceae Strophantus sarmentosus
Arecaceae Borassus aethiopum
Arecaceae Elaeis guineensis
Asteraceae Vernonia colorata
Bignoniaceae Kigelia africana
69
Bombacaceae Bombax costatum
Caesalpiniaceae Isoberlinia doka
Caesalpiniaceae Piliostigma reticulatum
Combretaceae Combretum lecardii
Combretaceae Guiera senegalensis
Ebenaceae Diospyros mespiliformis
Fabaceae/Papilionaceae Lonchocarpus cyanescens
Fabaceae/Papilionaceae Millettia thonningii
Fabaceae/Papilionaceae Pterocarpus erinaceus
Fabaceae/Papilionaceae Pterocarpus santalinoides
Meliaceae Khaya senegalensis
Mimosaceae Albizia glaberrima
Mimosaceae Albizia zygia
Mimosaceae Mimosa pigra
Mimosaceae Prosopis africana
Rhamnaceae Ziziphus amphibia
Rhamnaceae Ziziphus mauritiana
Rubiaceae Feretia apodanthera
Rubiaceae Gardenia ternifolia
Sapindaceae Lecaniodiscus cupanioides
Sterculiaceae Cola laurifolia
Tiliaceae Grewia flavescence
Tiliaceae Grewia lasiodicus
Tiliaceae Grewia mollis
Verbenaceae Vitex chrysocarpa
Verbenaceae Vitex doniana
Vitaceae Cissus populnea
Annexe 4: Liste des espèces des groupements
à
Daniellia oliveri et Combretum
ghasalense
Famille Espèces
Anacardiaceae Lannea acida
Anacardiaceae Lannea velutina
Annonaceae Annona senegalensis
Annonaceae Hexalobus monopetalus
Araliaceae Cussonia arborea
Arecaceae Borassus aethiopum
Balanitaceae Balanites aegyptiaca
Bombacaceae Adansonia digitata
Bombacaceae Bombax costatum
Caesalpiniaceae Burkea africana
Caesalpiniaceae Daniellia oliveri
Caesalpiniaceae Detarium microcarpum
Caesalpiniaceae Isoberlinia doka
Caesalpiniaceae Isoberlinia tomentosa
70
Caesalpiniaceae Piliostigma thonningii
Caesalpiniaceae Tamarindus indica
Celastraceae Maytenus senegalensis
Chrysobalanaceae Maranthes polyandra
Chrysobalanaceae Parinari curatelifolia
Chrysobalanaceae Parinari curatellifolia
Combretaceae Anogeissus leiocarpus
Combretaceae Combretum ghasalense
Combretaceae Combretum glutinosum
Combretaceae Combretum hypopilinum
Combretaceae Combretum micranthum
Combretaceae Combretum nigricans
Combretaceae Pteleopsis suberosa
Combretaceae Terminalia avicennioides
Combretaceae Terminalia glaucescens
Ebenaceae Diospyros mespiliformis
Fabaceae/Papilionaceae Ostryoderris stuhlmannii
Fabaceae/Papilionaceae Pericopsis laxiflora
Fabaceae/Papilionaceae Pterocarpus erinaceus
Flacourtiaceae Flacourtia indica
Hymenocardiaceae Hymenocardia acida
Hymenocardiceae Hymenocardia acida
Hypericaceae Psorospermum febrifugum
Loganiaceae Strychnos spinosa
Meliaceae Azadirachta indica
Meliaceae Trichilia emetica
Mimosaceae Acacia ataxacantha
Mimosaceae Acacia dudgeoni
Mimosaceae Acacia gourmaensis
Mimosaceae Acacia hockii
Mimosaceae Acacia macrostachya
Mimosaceae Acacia macrothyrsa
Mimosaceae Acacia polyacantha
Mimosaceae Acacia sieberiana
Mimosaceae Acacia tortilis
Mimosaceae Entada africana
Mimosaceae Mimosa pigra
Mimosaceae Parkia biglobosa
Moraceae Ficus gnaphalocarpa
Moraceae Ficus ingens
Moraceae Ficus sp
Ochnaceae Lophira lanceolata
Opiliaceae Opilia celtidifolia
Polygalaceae Securidaca longepedunculata
Rhamnaceae Ziziphus mauritiana
Rubiaceae Crossopteryx febrifuga
71
Rubiaceae Gardenia ternifolia
Rubiaceae Mitragyna inermis
Rubiaceae Sarcocephalus latifolius
Sapotaceae Vitellaria paradoxa
Simaroubaceae Hannoa undulata
Sterculiaceae Sterculia setigera
Tiliaceae Grewia mollis
Verbenaceae Vitex doniana
Vitaceae Cissus populnea
Annexe 5: Liste des espèces rencontrées
dans les
peuplements à Pterocarpus erinaceus et
Anogeissus leiocarpus
Famille Espèces
Anacardiaceae Haematostaphis barteri
Anacardiaceae Lannea acida
Anacardiaceae Lannea barteri
Anacardiaceae Lannea microcarpa
Anacardiaceae Mangifera indica
Anacardiaceae Rhus natalensis
Annonaceae Annona senegalensis
Annonaceae Hexalobus monopetalus
Apiaceae Steganotaenia araliaceae
Araliaceae Cussonia arborea
Arecaceae Borassus aethiopum
Balanitaceae Balanites aegyptiaca
Bignoniaceae Kigelia africana
Bignoniaceae Stereospermum kunthianum
Bombacaceae Adansonia digitata
Bombacaceae Bombax costatum
Bombacaceae Ceiba pentadra
Caesalpiniaceae Afzelia africana
Caesalpiniaceae Burkea africana
Caesalpiniaceae Daniellia oliveri
Caesalpiniaceae Detarium microcarpum
Caesalpiniaceae Isoberlinia doka
Caesalpiniaceae Isoberlinia tomentosa
Caesalpiniaceae Isoberlinia tomentosa
Caesalpiniaceae Piliostigma reticulatum
Caesalpiniaceae Piliostigma thonningii
Caesalpiniaceae Tamarindus indica
Capparidaceae Crateva adansonii
Celastraceae Maytenus senegalensis
Chrysobalanaceae Maranthes polyandra
Chrysobalanaceae Parinari curatelifolia
Chrysobalanaceae Parinari curatellifolia
Combretaceae Anogeissus leiocarpus
72
Combretaceae Combretum ghasalense
Combretaceae Combretum glutinosum
Combretaceae Combretum hypopilinum
Combretaceae Combretum nigricans
Combretaceae Combretum paniculatum
Combretaceae Combretum sp
Combretaceae Pteleopsis suberosa
Combretaceae Terminalia avicennioides
Ebenaceae Diospyros mespiliformis
Euphorbiaceae Bridelia ferruginea
Euphorbiaceae Securinega virosa
Fabaceae/Papilionaceae Erythrina senegalensis
Fabaceae/Papilionaceae Lonchocarpus sericeus
Fabaceae/Papilionaceae Millettia thonningii
Fabaceae/Papilionaceae Ostryoderris stuhlmannii
Fabaceae/Papilionaceae Pericopsis laxiflora
Fabaceae/Papilionaceae Pterocarpus erinaceus
Flacourtiaceae Flacourtia indica
Hymenocardiaceae Hymenocardia acida
Hypericaceae Psorospermum febrifugum
Loganiaceae Strychnos spinosa
Meliaceae Khaya senegalensis
Meliaceae Pseudocedrela kotschyi
Meliaceae Trichilia emetica
Mimosaceae Acacia polyacantha
Mimosaceae Acacia seyal
Mimosaceae Albizia zygia
Mimosaceae Entada africana
Mimosaceae Mimosa pigra
Mimosaceae Parkia biglobosa
Mimosaceae Prosopis africana
Moraceae Ficus capensis
Moraceae Ficus gnaphalocarpa
Ochnaceae Lophira lanceolata
Olacaceae Ximenia americana
Opiliaceae Opilia celtidifolia
Polygalaceae Securidaca longepedunculata
Rubiaceae Crossopteryx febrifuga
Rubiaceae Feretia apodanthera
Rubiaceae Gardenia ternifolia
Rubiaceae Macrosphyra longistyla
Rubiaceae Sarcocephalus latifolius
Sapotaceae Vitellaria paradoxa
Simaroubaceae Hannoa undulata
Sterculiaceae Cola laurifolia
Sterculiaceae Sterculia setigera
73
Tiliaceae Grewia bicolor
Verbenaceae Vitex doniana
Annexe 6: Liste complète des espèces
rencontrées
dans les groupements à Afzelia africana
et Balanites aegyptiaca
Famille Espèces
Anacardiaceae Lannea acida
Annonaceae Annona senegalensis
Annonaceae Hexalobus monopetalus
Balanitaceae Balanites aegyptiaca
Bignoniaceae Stereospermum kunthianum
Bombacaceae Adansonia digitata
Bombacaceae Bombax costatum
Caesalpiniaceae Afzelia africana
Caesalpiniaceae Burkea africana
Caesalpiniaceae Daniellia oliveri
Caesalpiniaceae Detarium microcarpum
Caesalpiniaceae Isoberlinia doka
Caesalpiniaceae Isoberlinia tomentosa
Caesalpiniaceae Piliostigma reticulatum
Caesalpiniaceae Piliostigma thonningii
Caesalpiniaceae Tamarindus indica
Celastraceae Maytenus senegalensis
Chrysobalanaceae Maranthes polyandra
Combretaceae Anogeissus leiocarpus
Combretaceae Combretum collinum
Combretaceae Combretum ghasalense
Combretaceae Combretum glutinosum
Combretaceae Combretum hypopilinum
Combretaceae Combretum nigricans
Combretaceae Combretum sp
Combretaceae Guiera senegalensis
Combretaceae Terminalia avicennioides
Combretaceae Terminalia macroptera
Dipterocarpaceae Monotes kerstingii
Ebenaceae Diospyros mespiliformis
Euphorbiaceae Bridelia ferruginea
Fabaceae/Papilionaceae Pericopsis laxiflora
Fabaceae/Papilionaceae Pterocarpus erinaceus
Fabaceae/Papilionaceae Pterocarpus santalinoides
Loganiaceae Strychnos innocua
Loganiaceae Strychnos spinosa
Meliaceae Khaya senegalensis
Mimosaceae Acacia ataxacantha
74
Mimosaceae Acacia pennata
Mimosaceae Acacia sieberiana
Mimosaceae Entada africana
Mimosaceae Mimosa pigra
Mimosaceae Parkia biglobosa
Mimosaceae Prosopis africana
Moraceae Ficus capensis
Moraceae Ficus ingens
Ochnaceae Lophira lanceolata
Polygalaceae Securidaca longepedunculata
Rubiaceae Crossopteryx febrifuga
Rubiaceae Feretia apodanthera
Rubiaceae Gardenia ternifolia
Rubiaceae Mitragyna inermis
Sapotaceae Vitellaria paradoxa
Sterculiaceae Sterculia setigera
Verbenaceae Vitex chrysocarpa
Verbenaceae Vitex doniana
75
Annexe 7: Quelques espèces de plantes
domestiquées dans les terres de parcours
|
Nom scientifique
|
Partie
|
Usages
|
Maladies traitées
|
|
Adansonia digitata,
|
Ecorce de tronc, feuilles et fruits
|
Arbre fétiche, aliments et médicaments
|
Paludisme, gynécologie, anémie et autres
|
|
Albizia ferruginea,
|
Jeunes feuilles
|
Médicaments
|
Paludisme
|
|
Albizia zygia,
|
Tige feuillée
|
Médicaments
|
Paludisme
|
|
Argemone mexicana,
|
Plante entière
|
Médicaments
|
Paludisme
|
|
Azadirachta indica,
|
Rameaux feuillées
|
Médicaments
|
Paludisme
|
|
Carica papaya,
|
Feuilles et fruits
|
Aliments et médicaments
|
Paludisme et hypertension
|
|
Cassia occidentalis
|
Feuilles, racines
|
Médicaments
|
Paludisme
|
|
Cassia rotundifolia,
|
Tige feuillée
|
Médicaments
|
Paludisme
|
|
Cienfuegosia leptoclada
|
Feuilles
|
Médicaments
|
Paludisme
|
|
Citrus sp,
|
Feuilles, Fruits
|
Aliments et médicaments
|
|
|
Commiphora africana,
|
Tige feuillée
|
Arbre protecteur contre les mauvais sorts Médicaments
|
Paludisme et aphasie
|
|
Crateva religiosa,
|
Jeunes feuilles
|
Médicaments
|
Antiparasite
|
|
Cymbopogon citratus
|
Feuilles
|
Médicaments et excipients
|
|
|
Dracaena fragrans
|
Feuilles
|
Arbre fétiche
|
Rhumatisme et anémie
|
|
Hibiscus asper,
|
Feuilles
|
Aliments et médicaments
|
Faiblesse sexuelle
|
|
Jatropha curcas,
|
Feuilles
|
Médicaments et palissade
|
Antimicrobien
|
|
Khaya senegalensis
|
Feuilles, tiges, racines
|
Médicaments et fourrage
|
Aménorrhée, diarrhée
|
|
Mangifera indica,
|
Feuilles, fruits, écorces, racines
|
Aliments et médicaments
|
Hypertension, paludisme
|
|
Momordica charantia
|
Feuilles fruits
|
Médicaments
|
Maladies éruptives et céphalées
|
|
Moringa oleifera
|
Feuilles
|
Aliment, médicament
|
Paludisme, céphalées
|
|
Newbouldia laevis
|
Feuilles
|
Protection, médicament et palissade
|
Dysménorrhée, anti-biotique
|
|
Ocimum sp,
|
Feuilles
|
Aliments et médicaments
|
Antibiotique
|
|
Parkia biglobosa,
|
Ecorces, feuilles, graines des
fruits
|
Aliments et médicaments
|
Hypertension artérielle, hémorroïdes,
dermatose et autres
|
|
Pavetta crassipes,
|
Feuilles
|
Médicaments
|
Paludisme
|
|
Phyllantus amarus,
|
Plante entière
|
Médicaments
|
Paludisme
|
|
Capsicum sp
|
Fruits
|
Epices
|
|
|
Sarcocephallus latifolius
|
Racines
|
Médicaments
|
Paludisme
|
|
Tamarindus indica
|
Fruits
|
Aliments et médicaments
|
Paludisme, gonococcie, céphalées
|
|
Uvaria chamae
|
Feuilles
|
Aliments et médicaments
|
Paludisme
|
|
Vitellaria paradoxa,
|
Ecorces et noix
|
Aliments et médicaments
|
Gastralgie, convulsion, antivenimeux, stérilité
féminine, excipient (beurre)
|
76
Annexe 8: Phénologie des quatre ligneux
fourragers
|
|
DANIELLIA
|
PTEROCARPUS
|
AFZELIA
|
KHAYA
|
|
1 ière quinzaine
|
1
|
Fv
|
Ch
|
Fj
|
Fr
|
|
2
|
Fv
|
Fv
|
Fv
|
Fl
|
|
3
|
Fv
|
Fv
|
Fv
|
Fl
|
|
4
|
Ch
|
Fv
|
Fv
|
Fl
|
|
5
|
Fv
|
Fv
|
Fj
|
Fl
|
|
6
|
Fv
|
Fv
|
Fv
|
Fl
|
|
7
|
Fv
|
Fv
|
Fv
|
Fr
|
|
8
|
Ch
|
Fv
|
Fv
|
Fl
|
|
9
|
Fv
|
Fv
|
Fv
|
Fl
|
|
10
|
Fv
|
Fv
|
Fv
|
Fl
|
|
2 ième quinzaine
|
1
|
Fv
|
Fv
|
Fv
|
Fl
|
|
2
|
Fv
|
Fv
|
Fj
|
Fl
|
|
3
|
Ch
|
Fv
|
Fv
|
Fl
|
|
4
|
Fv
|
Fv
|
Fv
|
Fj
|
|
5
|
Fv
|
Fv
|
Fv
|
Fl
|
|
6
|
Fv
|
Ch
|
Fv
|
Fl
|
|
7
|
Ch
|
Fv
|
Fv
|
Fl
|
|
8
|
Fv
|
Fv
|
Fv
|
Fj
|
|
9
|
Fv
|
Fv
|
Fv
|
Fl
|
|
10
|
Fv
|
Fv
|
Fv
|
Fr
|
|
3 ième quinzaine
|
1
|
Ch
|
Ch
|
Fv
|
Fl
|
|
2
|
Fv
|
Ch
|
Fv
|
Fl
|
|
3
|
Fv
|
Ch
|
Fv
|
Fl
|
|
4
|
Fv
|
Ch
|
Fv
|
Fl
|
|
5
|
Fv
|
Ch
|
Fv
|
Fl
|
|
6
|
Fv
|
Ch
|
Fv
|
Fl
|
|
7
|
Fv
|
Fv
|
Fv
|
Fl
|
|
8
|
Fv
|
Ch
|
Fv
|
Fl
|
|
9
|
Fv
|
Ch
|
Fv
|
Fl
|
|
10
|
Fv
|
Ch
|
Fv
|
Fl
|
|
4 ième quinzaine
|
1
|
Fv
|
Ch
|
Fv
|
Fl
|
|
2
|
Fv
|
Ch
|
Fv
|
Fl
|
|
3
|
Fv
|
Ch
|
Fv
|
Fl
|
|
4
|
Fv
|
Ch
|
Fv
|
Fl
|
|
5
|
Fv
|
Fv
|
Fv
|
Fl
|
|
6
|
Fv
|
Ch
|
Fv
|
Fl
|
|
7
|
Fv
|
Ch
|
Fv
|
Fl
|
|
8
|
Fv
|
Ch
|
Fv
|
Fl
|
|
9
|
Fv
|
Ch
|
Fj
|
Fr
|
|
10
|
Ch
|
Fv
|
Fv
|
Fl
|
|
Fl = fleurs
|
Fr = fruits
|
Ch = chute feuille
|
|
|
Fb = feu de brousse
|
Fv = feuilles vertes
|
Fj = feuilles jaunes
|
|
| 

