|
|
UNIVERSITE DE MARADI
|
|
|
|
|
|
|
N0 d'ordre : 20
|
UNIVERSITE DE MARADI
FACULTE DES SCIENCES ET TECHNIQUES
M E M O I R E
Présenté pour l'obtention du
DIPLOME DE MASTER II
Biodiversité et Gestion de l'Environnement
Soudanien et S ahélo-S aharien

Sur le thème
Etude de l'effet de coupes de la partie
aérienne sur la croissance et l'utilisation d'eau chez deux
espèces fourragères (Cenchrus biflorus Roxb et
Zornia glochidiata Reichenbach ex de Candolle)
Présenté Par : Oumarou Abdoulaye Moussa,
Sous la direction de :
Dr Mainassara Zaman Allah
Soutenu publiquement le 09/02/2013 devant le jury composé
de : Président : Pr Ali Mahamane
Membres : Dr Mainassara Zaman Allah
Dr Elh Mahamadou Gounga

Dédicace
Je dédie le présent travail à mon
père Oumarou Abdoulaye et ma mère Assamaou Ibrahima qui m'ont
soutenu financièrement et moralement durant tout mon parcours
académique.

REMERCIEMENTS
A l'issus de ce stage, nous adressons nos remerciements les
plus sincères à tous ceux qui de près ou de loin n'ont
ménagé aucun effort pour sa réussite.
C'est l'occasion pour nous de leur exprimer notre profonde
gratitude et notre reconnaissance. Ainsi nos remerciements vont
particulièrement à :
· Dr Mainassara Zaman-Allah, Enseignant-chercheur
à la Faculté des Sciences pour avoir accepté d'encadrer ce
travail et pour ses conseils éclairés dans la conception du
présent document,
· Mr Karim saley, Enseignant-chercheur à la
Faculté des Sciences qui a bien voulu accepté nous aider
malgré ces préoccupations,
· Pr Saadou Mahamane, Recteur de l'Université de
Maradi qui nous a initiés à la recherche,
· Pr Ali Mamane, vice-recteur de l'Université de
Maradi et Doyen de la Faculté des Sciences qui nous a tenus pendant 4
ans au cours de notre formation,
· Pr Bakasso Yacouba, vice doyen de la Faculté
des Sciences et Techniques de l'Université Abdou Moumouni de Niamey, qui
nous a tenus tout le long de notre formation,
· Dr Amadou Oumani Abdoulaye, Enseignant-chercheur et
chef du service stage à l'Université de Maradi,
· Dr Morou Boubé, Enseignant-chercheur à
la Faculté des Sciences et chef du département de Biologie,
· Nous voudrions enfin exprimer nos sentiments de
cordialité à tous ceux qui nous ont aidés de près
ou de loin, et tous les camarades de Master 2 en BGE/SSS pour toute la
collaboration. A tous ceux que nous aurions oubliés de citer, nous
formulons nos sentiments de profonde gratitude et d'indéfectible
attachement.
iv
SOMMAIRE
Dédicace ii
REMERCIEMENTS iii
SIGLES ET ABBREVIATIONS vi
Liste du tableau vii
Liste des figures vii
RESUME ix
ABSTRACT x
INTRODUCTION 1
I . PREMIERE PARTIE: REVUE BIBLIOGRAPHIQUE
3
1. Caractéristiques bioécologiques de
Cenchrus biflorus 3
1.1 Systématique et description botanique 3
1.2 Écologie 3
1.3 Importance 4
2. Caractéristiques bioécologiques de
Zornia glochidiata 5
2.1 Systématique et description botanique 5
2.2 Écologie 6
2.3 Importance 6
3. Effet de la sécheresse et du broutage sur les
parcours 6
3.1 Effet de la sécheresse sur les parcours 6
3.2 Effet du broutage sur les parcours 7
4. Conclusion partielle 9
II. DEUXIEME PARTIE : MATERIELS ET METHODES
10
1. Etude anatomique 10
2. Matériel végétal 10
3.
V
Matériels techniques 10
4. Sites d'étude, dispositif expérimental
et conduite des cultures 11
4.1 Dispositif expérimental au niveau des placettes du
site IUT 11
4.2 Dispositif expérimental au niveau de l'essai en
pots 11
4.3 Traitements 13
4.4 Paramètres mesurés 14
4.5. Analyse des données 14
III. TROISIEME PARTIE : RESULTATS
15
3.1 Etude de l'anatomie de Cenchrus et Zornia
15
3.2. Réponse de la croissance au broutage
simulé 19
3.2.1 Effet des coupes sur la production de biomasse chez
Cenchrus biflorus cultivé sur placettes 19
3.2.2 Effet des coupes sur la production de biomasse chez
Cenchrus biflorus cultivé dans des pots 20
3.2.3 Effet des coupes sur la production de biomasse chez Zornia
cultivé sur placettes 21
3.2.4 Effet des coupes sur la production de biomasse chez Zornia
cultivé dans des pots 22
3.3. Réponse de la transpiration à la
variation journalière du déficit de pression de vapeur
chez
Cenchrus 23
IV. QUTRIEME PARTIE : DISCUSSION
26
CONCLUSION 28
REFERENCES BIBLIOGRAPHIQUES
29
Annexes a
Annexe 1 : Dispositif de culture au niveau de L'IUT
a
Annexe 2 : Données du Mesure de la transpiration
par voie gravimétrique b
Annexe 3: Données du Mesure de biomasse du
cenchrus Biflorus c
Annexe 4: Données du Mesure de biomasse du Zornia
glochidiata e

SIGLES ET ABBREVIATIONS
DVP: déficit de pression de vapeur
FAO: Organisation des Nations Unies pour l'Alimentation et
l'Agriculture
INRAN : Institut National de Recherche Agronomique du Niger
ICRISAT : International Crops Research Instituted for the
Semi-Arid Tropics IUT : Institut
Universitaire de Technologie
UM : Université de Maradi
vii
Liste du tableau
Tableau 1 : Calendrier des coupes chez les
deux espèces 11
Liste des figures
Figure 1 : Schéma d'une parcelle
élémentaire. 11
Figure 2 : Production de biomasse chez Cenchrus biflorus
pour différents niveaux de coupes
(3 cm et 5 cm) 19
Figure 3 : Effet des coupes à 3 cm
et 5 cm du sol sur la production de biomasse totale chez
Cenchrus biflorus. 20
Figure 4 : Production de
biomasse chez Cenchrus biflorus pour différents niveaux de
coupes
(3 cm et 5 cm) 20
Figure 5 : Effet des coupes à 3 cm
et 5 cm du sol sur la production de biomasse totale chez
Cenchrus biflorus. 21
Figure 6 : Effet des coupes
à 3 cm et 5 cm du sol sur la production de biomasse totale chez
Zornia cultivé sur placette 22
Figure 7 :
Effet des coupes à 3 cm et 5 cm du sol sur la production de biomasse
totale chez
Zornia cultivé en pots 22
Figure 8 :
Variation du déficit de pression de vapeur (A) au cours d'une
journée nuageuse et (B) au cours d'une journée très
ensoleillée. Les données ont été
enregistrées au niveau du site
de l'Université de Maradi. 23
Figure 9 : Variation
de la transpiration en fonction du temps au cours d'une journée
nuageuse
chez Cenchrus 24
viii
Figure 10 : Variation de la transpiration en fonction du temps au
cours d'une journée très
ensoleillée chez Cenchrus 25
Liste des photos
Photo 1 : (A) semis de Cenchrus en
ligne sur une placette et (B) Dispositif expérimental
des plants de Cenchrus dans les pots 12
Photo 2 : (A) culture de Z.glochidiata
sur les placettes et (B) dans les pots 13
Photo 3 : Mesure de la transpiration pour le
génotype de Cenchrus .13
Photo 4 :
Observation de l'épiderme de Cenchrus biflorus au microscope
photonique (a- x100
et b-x400) ..15
Photo 5 : Observation de
l'épiderme de Zornia glochidiata au microscope photonique
(a-x100 et
|
b-x400)
|
16
|
|
Photo 6 : Coupe anatomique de la tige de
Z.glochidiata (x400)
|
17
|
|
Photo 7 : Coupe anatomique de la tige de
C.biflorus (x400)
|
..18
|
|
Photo 8 : (A) portion de la structure anatomique
de Z.glochidiata et (B) de C.biflorus
(x400)
|
18
|
iX
RESUME
Afin d'étudier l'impact du broutage (simulé par
des coupes sur la partie aérienne) sur la production de biomasse, deux
espèces fourragères, Cenchrus biflorus et Zornia
glochidiata ont été cultivées sur les placettes et
dans des pots et soumises à des opérations de coupes à
différents niveaux (3cm et 5 cm du sol). D'autre part, l'effet du
broutage sur la tolérance à la sécheresse a
été évalué chez Cenchrus biflorus en
étudiant la réponse de la transpiration à la variation
journalière du déficit de pression de vapeur (DPA).
Les résultats ont une variation de la production de
biomasse en réponse aux coupes selon l'espèce, le milieu de
culture, le niveau ou le nombre de coupes. De façon globale, la
production de biomasse a augmenté significativement surtout au niveau
des placettes pour les coupes à 5 cm du sol par rapport aux coupes
à 3 cm et au traitement témoin. Par ailleurs, la transpiration
chez Cenchrus biflorus a montré une forme augmentation en
réponse à l'accroissement du DPV. Les coupes ont
significativement réduit cette transpiration, quel que soit le niveau,
du fait surtout de la réduction de la surface foliaire qu'elles
occasionnent. En définitive, un broutage modéré (coupe
à 5cm) peut améliorer la production de biomasse et permettre une
meilleure adaptation au déficit hydrique par une réduction
sensible de la perte d'eau par transpiration.
Mots-clés : Cenchrus biflorus,
Zornia glochidiata, biomasse, déficit de pression de vapeur, coupe
de la partie aérienne, broutage.
X
ABSTRACT
To investigate the impact of grazing (simulated by shoot cuts)
on biomass production, two forage species, Cenchrus biflorus and
Zornia glochidiata were grown on plots and in pots and subjected to
shoot cuts at different levels (3cm and 5 cm from soil surface). On the other
hand, the effect of grazing on drought tolerance was evaluated in Cenchrus
biflorus by assessing the response of transpiration to the diurnal
variation of vapor pressure deficit (VPD).
Results showed that the biomass production varied in response
to shoot cuts depending on the species, the culture system, the level or number
of cuts. Overall, biomass production increased significantly especially in
plots for cuts at 5 cm compared to those at 3 cm and the control treatment. In
addition, transpiration of Cenchrus biflorus was highly increased in
response to the VPD increase. Shoot cuts significantly reduced transpiration,
whatever the level, largely because they reduce leaf surface. We concluded that
moderate grazing (cuts to 5cm) can improve biomass production and allow better
adaptation to water deficit as they significantly reduced water loss through
transpiration.
Keywords: Cenchrus biflorus, Zornia
glochidiata, biomass, vapor pressure deficit, cutting the aerial parting,
grazing.
INTRODUCTION
Le Niger est un pays sahélien appartenant à une
zone d'Afrique semi-aride, avec un climat qui se caractérise par
l'alternance de deux saisons aux caractères fortement contrastés
: une saison humide courte (2 à 3 mois) et une saison sèche
longue (9 à 10 mois). Au cours des dernières décennies,
ces régions ont connu une forte variabilité annuelle et spatiale
de la pluviosité; ce qui s'est traduit par une tendance à
l'aridité progressive du climat (Banoin et al. 1996) avec de
graves conséquences notamment dans le domaine de la production animale
liée à la rareté de fourrage. La sécheresse de
1973-1974 a décimée près de la moitié du cheptel
nigérien (MAE, 1992). Ces changements climatiques ont beaucoup
modifiés les systèmes de production agropastoraux, en particulier
les règles de gestion des ressources naturelles par les populations
(Thébaub B., 1999). Ces modifications ont été
accentuées par un taux de croissance démographique de 3,5%
(Rapport Nations Unies, 1996) entraînant une extension des terres de
culture au détriment des aires de parcours. L'élevage est de type
extensif, donc fortement dépendant de la végétation
naturelle. La zone sahélienne connaît de profondes mutations et,
avec la sécheresse qui y sévit depuis plus d'une décennie,
on assiste à une dégradation graduelle des
écosystèmes et une diminution de surfaces pâturables. Dans
ce contexte, un intérêt particulier devrait être
porté sur les espèces locales ayant une bonne valeur
fourragère et adaptées aux conditions pédoclimatiques du
Sahel. Cependant, l'utilisation de telles espèces nécessite une
meilleure connaissance de leur biologie et de leurs mécanismes de
régénération et de croissance. C'est dans ce cadre que
s'inscrit la présente étude qui concerne deux espèces
fourragères Cenchrus biflorus Roxb et Zornia glochidiata
Reichb ex DC, très appréciées par le bétail.
Ces espèces ont fait l'objet de quelques études mais,
malgré une excellente valeur fourragère et une relative
adaptation à la sécheresse, la biologie de ces espèces
n'est pas encore très bien connue. A ce titre, la sélection
d'espèces fourragères est encore nécessaire pour rehausser
la valeur des pâturages naturels ou créer de nouveaux
pâturages ensemencés. La légumineuse (Zornia
glochidiata) constitue à cet égard une excellente plante
fourragère du fait de sa teneur élevée en protéines
et en vitamines et de sa capacité à fixer l'azote
atmosphérique par suite d'une association symbiotique avec les
bactéries du genre Rhizobium. Ainsi, elle est souvent
utilisée pour l'amélioration des pâturages (N. Mbaye et
al. 2002). La graminée Cenchrus biflorus constitue aussi
une excellente plante fourragère du fait de
2
sa teneur très élevée en matières
minérales (protéine, lipide, Ca, K...) et en acides aminés
qui sont des éléments très importants pour l'alimentation
du bétail (FAO, 1970). Le travail présenté dans ce
mémoire étudie quelques paramètres relatifs à la
réponse de ces deux espèces au broutage simulé et au
déficit hydrique. L'objectif est d'apporter des éléments
permettant une meilleure compréhension des stratégies de
résistance au déficit hydrique et au broutage par ces
espèces. Le mémoire est articulé en quatre grandes
parties. La première partie comporte une synthèse bibliographique
sur la taxonomie des deux espèces fourragères, leur
écologie ainsi que leur importance. La deuxième partie
décrit le matériel biologique et les méthodes
utilisés, ainsi que les protocoles d'essais, les dispositifs et les
conditions de culture appliqués. La troisième partie est
consacrée aux résultats obtenus et la dernière partie
englobe la discussion générale des résultats et les
principales conclusions.
3
I . PREMIERE PARTIE: REVUE BIBLIOGRAPHIQUE
1. Caractéristiques bioécologiques de
Cenchrus biflorus
1.1 Systématique et description botanique
Cenchrus biflorus est une plante de la classe des
Monocotylédones, ordre des Cypérales et de la famille des
Poaceae. C'est une graminée annuelle, en touffes lâches, à
tiges (chaumes) ascendantes jusqu'à 1 m de haut. Les feuilles sont
alternes, simples et entières ; la ligule est constituée d'une
ligne de poils ; le limbe est linéaire, plat, de taille 2-25(-35) cm
× 2-7(-10) mm, et l'apex filiforme. L'inflorescence est une panicule
spiciforme de 2-15 cm × 9-12 mm, avec 1-3 épillets enserrés
par un involucre de soies hérissées. Le rachis est anguleux,
sinueux ; involucre ovoïde, de 4- 11 mm de long, portant de nombreuses
épines. Les épines internes sont érigées,
soudées à la base, à poils dirigés vers le bas sur
l'apex piquant et recourbé. Les épines externes sont plus courtes
et étalées. L'épillet est lancéolé de 3,5-6
mm de long, aigu, composé de 2 glumes et généralement de 2
fleurs ; les glumes étant plus courtes que l'épillet. La fleur
inférieure mâle ou stérile, a un lemme aussi long que
l'épillet et membraneux. La fleur supérieure bisexuée est
à lemme aussi long que l'épillet, légèrement
coriace ; avec 3 étamines, un ovaire supère, glabre à 2
stigmates poilus. Le fruit est un caryopse (grain) comprimé dorsalement,
de 2-2,5 mm× 1,5-2 mm.
Le genre Cenchrus comprend environ 20 espèces
dans les régions tropicales et tempérées chaudes,
essentiellement en Afrique et aux Amériques. Il est très proche
de Pennisetum, qui se distingue par ses soies involucrales internes
non épineuses et libres jusqu'à la base. Les épillets
épineux de Cenchrus biflorus s'accrochent aux poils des animaux
et aux vêtements, facilitant ainsi amplement leur dispersion.
Cenchrus biflorus a une photosynthèse en C4. (Burkill, et al.,
1994).
1.2 Écologie
On trouve habituellement Cenchrus biflorus dans les
régions semi-arides et arides où les précipitations
annuelles atteignent 250-650 mm, jusqu'à 1300 m d'altitude,
généralement sur des sols sableux secs et dans des zones
cultivées, sur
4
pâturées ou autrement perturbées. Il est
extrêmement abondant au Sahel et au sud du Sahara, où il peut
former des peuplements massifs. Une étude menée dans l'ouest du
Niger a montré qu'il était devenu beaucoup plus abondant et
dominant à la fin des années 1980 qu'il ne l'était au
début des années 1960. (Burkill, et al. 1994).
1.3 Importance
Les grains de Cenchrus biflorus sont comestibles et
très nutritifs (Burkill, et al. 1994).Dans les zones où les gens
ont à peine de quoi vivre, les graines sont régulièrement
récoltées. Ailleurs, il est considéré comme un
aliment de famine. Au Sahel, il est ramassé comme une
céréale sauvage, par exemple par les Touaregs. Les grains sont
écrasés et consommés crus, transformés en bouillie,
ou bien mélangés et cuits avec d'autres aliments. Ils servent
aussi à préparer une boisson. Au Soudan, on fabrique une galette
plate («kisra») avec les grains et en Mauritanie des gâteaux
à partir des grains moulus. Les grains de Cenchrus biflorus
servent aussi d'aliment de famine en Inde, où ils sont
consommés crus ou bien utilisés, mélangés à
du mil, pour la fabrication du pain. En temps normal, ils sont
consommés, mélangés avec du sucre et du «ghee»,
comme aliment pour les enfants
Cenchrus biflorus est considéré comme
une plante fourragère très utile au Sahel. Elle est
broutée essentiellement lorsqu'elle est jeune et une fois que les grains
sont tombés. On peut la faucher à plusieurs reprises pendant la
saison des pluies pour en faire du foin ou de l'ensilage. Les involucres
épineux sont suffisamment ramollis par l'ensilage pour rendre possible
la consommation de la plante entière. Cenchrus biflorus
persiste jusqu'à la fin de la saison sèche, ce qui contribue
à en faire une source sûre de fourrage. En Inde également,
la plante sert de fourrage et elle est semée pour lutter contre la
désertification. Dans le nord de l'Australie, on la cultive comme plante
fourragère. Ses feuilles sont consommées en période de
famine dans le désert du Thar en Inde. La racine de Cenchrus
biflorus est un ingrédient qui figure dans les ordonnances
aphrodisiaques traditionnelles. (Burkill, et al, 1994)
5
2. Caractéristiques bioécologiques de
Zornia glochidiata
2.1 Systématique et description botanique
Zornia glochidiata est une plante de la classe des
Dicotylédones, ordre des Leguminosae et famille des Fabaceae. C'est est
une espèce herbacée annuelle à tiges érigées
ou décombrâtes atteignant 45 (voire 70) cm de long. Les feuilles
sont alternes, bifoliolées à stipules lancéolées,
atteignant 1,5 cm de long, munies d'un éperon avec un pétiole de
0,5-2 cm de long. Les folioles lancéolées à
ovales-lancéolées, atteignent 4,5 cm × 1,5 cm, sont
aiguës et brièvement mucronées à l'apex. Elles sont
glabres à pubescentes au-dessous et légèrement
ponctuées-glanduleuses.
L'inflorescence est en forme d'épi atteignant 20 cm de
long avec un pédoncule de 1-5 cm de long. Les bractées sont
appariées, ovales à elliptiques, atteignant 1,5 cm de long. Les
fleurs sont bisexuées, papilionacées à calice hyalin et
tube d'environ 1,5 mm de long, 5 lobes, atteignant 1,5 mm de long. Les 2 lobes
supérieurs sont connés et les pétales sont jaunes, roses
ou pourpres, souvent à veines rouges et un étendard atteignant 6
mm de long. Les étamines au nombre de 10 sont en filets réunis
à la base en un tube fermé. L'ovaire est supère,
uniloculaire avec un style courbé. Le fruit est une gousse
articulée atteignant 2 cm de long, couverte de soies
étalées, contenant 2-5 graines. Les graines sont
comprimées réniformes, d'environ 1,5 mm de long et brunes.
Zornia est un genre pantropical comprenant environ 80
espèces, dont 13 sont indigènes en Afrique et une introduite.
Zornia glochidiata et les autres espèces de Zornia
africaines ont été longtemps confondues avec Zornia
diphylla (L.) Pers., dont la répartition est limitée au Sri
Lanka et au sud de l'Inde. Il est probable que d'autres espèces de
Zornia soient utilisées en Afrique tropicale comme
légume de la même façon que Zornia glochidiata.
Les plantes de Zornia glochidiata fanent après
la saison des pluies et se désintègrent rapidement. Les racines
sont pourvues de nodules fixateurs d'azote. (Berhaut, 1967.)
6
2.2 Écologie
Zornia glochidiata est commune dans les zones
sablonneuses où la pluviométrie annuelle atteint au moins 300 mm
pendant la saison des pluies. Il constitue un élément important
des steppes herbeuses sahéliennes et soudano-sahéliennes. Autour
des points d'eau au nord du Sénégal, il peut former un tapis de
végétation ininterrompue pendant la saison des pluies. En Afrique
orientale et australe, il se rencontre dans les savanes herbeuses et
boisées, les terrains vagues et les friches, jusqu'à 1800 m
d'altitude. (Berhaut, 1967)
2.3 Importance
Les feuilles de Zornia glochidiata sont
récoltées dans la nature et consommées cuites en sauces
avec du riz ou du couscous. Zornia glochidiata est une bonne plante
fixatrice des sols, par ex. sur les digues des rizières.
Considérée comme une bonne plante fourragère, elle est
importante surtout dans la région du Sahel, malgré les risques de
météorisation qu'elle occasionne au bétail. Ses feuilles
se prennent comme laxatif. Au Kenya, les feuilles cuites sont
administrées aux enfants qui souffrent de kwashiorkor. Au Congo, le jus
de la plante est utilisé en collyre pour combattre l'épilepsie,
et la racine est consommée comme aphrodisiaque. Au Zimbabwe, on emploie
les racines pour traiter les maladies vénériennes,
prévenir les fausses couches et faciliter les accouchements (Berhaut,
1967).
3. Effet de la sécheresse et du broutage sur les
parcours
3.1 Effet de la sécheresse sur les parcours
Une sécheresse peut être définie comme un
déficit hydrique dans le continuum sol-plante atmosphère
dommageable à la production. Ce déficit hydrique correspond
à une réduction de la teneur en eau, du potentiel hydrique ou du
potentiel de turgescence de la plante, et dont l'expression dans les tissus
végétaux détermine un certain nombre de réponses
des plantes. La notion de sécheresse recouvre souvent à la fois
des stress hydriques et thermiques, qui induisent tous deux des stress ioniques
(Monneveux, 1997). Un stress est défini comme l'action de tout facteur
du milieu potentiellement préjudiciable ou défavorable aux
organismes vivants (Levitt, 1980).
7
La sécheresse entraîne un assèchement des
cours d'eau et de la végétation des parcours, ce qui conduit
à des effets néfastes dont entre autre une réduction des
ressources végétales et animales et une réduction des
parcours et des cultures. Les sols deviennent souvent décapés et
érodés.
Après une série d'années sèches,
les espèces végétales des parcours les plus
pâturées cèdent la place à des espèces de
mauvaises herbes moins profondément enracinées et moins
productives. La croissance limitée des racines réduit la
capacité des plantes fourragères d'atteindre le peu
d'humidité qui subsiste dans le sol. Le faible taux d'humidité du
sol limite la croissance des plantes et réduit le rendement fourrager.
Les effets de la sécheresse peuvent être plus rapides sur les
pâturages dont le sol est d'une texture grossière ; ce qui a pour
conséquence une réduction de la capacité de charge de ces
sols en période de sécheresse, une réduction de la
production primaire, une modification de la structure du couvert
végétal et une réduction massive de la faune sauvage et du
cheptel.
3.2 Effet du broutage sur les parcours
La sensibilité des écosystèmes pastoraux
à la pression de pâture est conditionnée par des
particularités écologiques de l'écosystème
sahélien. Cette pression peut avoir des effets positifs ou
négatifs sur les parcours.
- Effets positifs du broutage
Le broutage dans les zones des parcours qui ont
été protégées sur une longue période peut
stimuler la couverture foliaire (Popolizio et coll., 1994). Habituellement, en
l'absence de broutage, les feuilles des végétaux s'allongent,
s'amincissent et se dressent et les grandes talles sont moins nombreuses, ce
qui réduit leur capacité de photosynthèse (Briske et
Richards, 1995). Si une partie des herbages au-dessus du sol est
éliminée, les autres feuilles sont exposées à des
intensités lumineuses plus fortes, ce qui augmente leur capacité
de photosynthèse et stimule leur croissance. La croissance des racines
et les processus racinaires bénéficient de la même
façon de la disparition d'un excès de fourrage. L'augmentation de
l'intensité lumineuse favorise la respiration des racines et
l'assimilation des éléments nutritifs (Manske, 1998; Briske et
Richards, 1995). Le broutage peut également stimuler la croissance
végétale par l'élimination de la dominance apicale, ce qui
force les végétaux à se ramifier et à
8
développer des talles secondaires, augmentant ainsi le
rendement et la couverture des herbages. Après chaque
défoliation, des périodes de repos suffisantes sont
nécessaires pour permettre la reconstitution des éléments
nutritifs qui alimenteront le nouveau feuillage.
Le broutage intensif dans des conditions humides
tempérées ou tropicales peut abaisser et propager la couverture
herbacée, constituant un tapis ou une couche protectrice ininterrompue.
Le pâturage détruit une partie de la biomasse et empêche
l'accumulation de litière, laquelle peut gêner la repousse et les
futures germinations des graminées. En coupant les jeunes chaumes avant
la floraison, le pâturage retarde le cycle de reproduction de certaines
herbes et maintient la qualité des plantes végétatives de
la pâture, qui sont beaucoup plus nutritives que les plantes adultes.
- Effets négatifs du broutage
Le broutage des plantes annuelles pendant la saison de
croissance limite leur capacité à produire des graines. Le
pâturage sélectif intensif amoindrit la
compétitivité des espèces les plus appétées
et favorise les espèces les moins utiles. Le pâturage et le
broutage excessifs peuvent finir par causer l'affaiblissement puis la mort des
plantes appétées. La détérioration du recouvrement
de la végétation est souvent liée au piétinement du
bétail et au tassement du sol. Si le recouvrement de la
végétation est insuffisant, le sol est exposé à
l'érosion. Des vents forts sur des sols très secs emportent de
fines particules et de la poussière, qui se retrouvent dans
l'atmosphère. Ces particules affectent la pureté de l'air et la
transmission de la lumière dans l'atmosphère.
Le climat, tropical sec, est marqué par la
régularité de la saison des pluies et
l'irrégularité de la distribution des pluies au cours de la
saison, ainsi que dans l'espace. L'extrême sécheresse de l'air
pendant plusieurs mois de la saison sèche limite la place des
herbacées pérennes et des plantes succulentes et explique que les
parcours sahéliens soient dominés par des herbacées
annuelles avec des plantes ligneuses éparses. La dépendance des
herbacées annuelles de la production de semence, renforcée par le
caractère transitoire du stock de semences dans les sols, est une des
causes majeures de la sensibilité des parcours à la gestion
pastorale. Par contre, l'équilibre entre plantes herbacées et
ligneuses, arbitré par leur compétition
9
pour l'eau et les minéraux, apparaît robuste et
les risques d'embroussaillement sont minimes. L'effet de la pâture est
décomposé en processus élémentaires que sont la
défoliation, le piétinement et le dépôt des
excrétions. L'impact de la pâture sur la végétation
est confronté aux modèles de la dynamique de la
végétation. Ni le modèle "successionel", ni celui du
"non-équilibre" ne satisfont toutes les observations. Il est
proposé de les considérer comme des modèles de
comportement alternatifs à replacer dans un modèle d'ensemble,
plus empirique, de type "états et flux". Ces connaissances imparfaites
sur la dynamique de la végétation et la biologie des
espèces sont ensuite mises à l'épreuve pour
détailler les effets attendus de telle ou telle pratique de gestion :
pâture à forte charge continue ou discontinue, mises en
défens, incendies et émondage des ligneux fourragers.
4. Conclusion partielle
Cette revue bibliographique a mis en évidence
l'importance des deux espèces fourragères utilisées dans
ce travail ainsi que les effets de la sécheresse et du broutage sur les
parcours. Une meilleure connaissance de leur réponse à ces deux
contraintes pourrait être d'une contribution majeure à la gestion
des parcours qui connaissent au Niger une situation de dégradation
alarmante. Notre travail vise à étudier l'adaptation de ces
espèces fourragères au déficit hydrique ;et au broutage
simulé par des coupes à différents niveaux de la partie
aérienne de la plante. Il se base sur deux hypothèses : (i) les
coupes ont un effet négatif sur la croissance mais affectent
différemment ces 2 espèces, (ii) l'espèce Cenchrus
s'adapte au stress hydrique en ayant une stratégie conservative
d'utilisation d'eau.
10
II. DEUXIEME PARTIE : MATERIELS ET METHODES
1. Etude anatomique
Une portion de l'épiderme de la face inferieure d'une
feuille de Cenchrus a été prélevée,
placée entre lame et lamelle dans de l'eau distillée et
observée au microscope photonique. En outre, des sections à main
levée d'environ 50 um d'épaisseur ont été
réalisées sur des portions de racine et tige et montées
sans coloration entre lame et lamelle dans de l'eau distillée. Pour
chaque organe dix sections uniformes ont été choisies au hasard
pour l'observation au microscope photonique.
1. Matériel végétal
Le matériel végétal utilisé dans
cette étude est constitué de deux espèces
fourragères (Cenchrus biflorus et Zornia glochidiata). Ces
espèces d'origines différentes ont été
collectées respectivement au niveau de L'INRAN/Maradi (Cenchrus
biflorus) et de la forêt de Dan Kada dans le département
d'Aguié (Zornia glochidiata).
2. Matériels techniques
Les matériels techniques utilisés dans cette
étude sont constitués de :
> Microscope photonique
> Lame et lamelle
> Eau du javel
> Balance à précision pour mesurer la
transpiration
> Peseur pour mesurer la biomasse
> Piquets pour délimiter la parcelle
> Appareil numérique
> sécateur pour la coupe
> Pots de 5 litre troués pour laisser égoutter
l'eau après arrosage
> Sol sablonneux pour remplir les pots
> Files blanc e noir pour la distinction de niveau de coupe
11
4. Sites d'étude, dispositif expérimental
et conduite des cultures
4.1 Dispositif expérimental au niveau des placettes
du site IUT
L'expérimentation a été conduite suivant
un dispositif randomisé sur une parcelle de 112 m2 (8 m x14),
au sein de laquelle trois blocs de 6 placettes de 2 m2 (1 m x2 m)
ont été réalisées avec 9 répétitions
pour chaque génotype. Au niveau de chaque placette (ou parcelle
élémentaire, Figure 1) quatre lignes distantes de 0 ,20 m
creusées superficiellement ont été faites dans lesquelles
ont été semées les graines.

Figure 1 : Schéma d'une parcelle
élémentaire. 4.2 Dispositif expérimental au niveau
de l'essai en pots
Pour ce dispositif, 35 pots ont été
utilisés pour les 2 génotypes de C.biflorus et
Z.glochidiata, dans lesquels les semis ont été
effectués. Ces pots en plastics de capacité de 5L sont
troués en bas pour laisser égoutter l'eau après arrosage
et remplis avec du sol sablonneux collecté au niveau du site de l'IUT.
Les graines des deux espèces ont été semées
à la même date en poquet (2 ou 3 par espèces) à
raison de 4 à 5 graines par poquet.
Pour la simulation du broutage 6 placettes ont
été utilisées avec chacune deux lignes coupées
(dont 3 placettes pour chaque niveau de coupe) ; les deux autres lignes de
chaque placettes ont été considérées comme
étant des témoins pour les pieds coupés.
Au niveau des pots, 20 ont été utilisés
dont 10 par génotypes dans lesquels 3 pieds ont été
coupés et laissant 7 comme témoins. Des fils blanc et
noir ont été
12
utilisés, pour attacher à la base au moins 5 pieds
coupés pour chaque traitement en vue de faciliter leur distinction.
Les coupes ont été effectuées suivant le
calendrier présenté dans le tableau 1. Tableau 1 :
Calendrier des coupes chez les deux espèces
|
Espèce Date Date
levée 1èrecoupe
|
2ème
coupe
|
Date de floraison Date de
récolte
|
C.biflorus
Placettes 12-juil 11 Aout 31 Aout 18 Aout 25-sept
Pots 15-juil 11 Aout 25 Aout 07-sept
Zornia glochidiata
Placettes 16-juil 24 Aout 07-sept 25-sept
Pots 20-juil 24 Aout 02-sept 06-sept

Photo 1 : (A) Placette de
cenchrus en lignes et (B) Dispositif expérimental des plants de
cenchrus dans les pots

13
A
Photo 2 : (A) Culture de Z. glochidiata
sur les placettes et (B) dans les pots

Photo 3 : Mesure de la transpiration pour le
génotype de Cenchrus
4.3 Traitements
Les traitements ont été effectués pour
chaque génotype au niveau des placettes et dans les pots. Sur les
placettes et dans les pots toutes les deux espèces ont suivies les
mêmes traitements suivants :
(1) Lot témoin, (2) Coupe à 3 cm du sol, (3) Coupe
à 5 cm du sol
14
Pour les conditions de stress hydrique seul le génotype
de C.biflorus a subit les traitements suivants dans les pots :
(1) Stress hydrique, (2) Stress hydrique + Coupe à 3
cm, (3) Stress hydrique+ Coupe à 5 cm.
4.4 Paramètres mesurés
Au cours de cette étude les paramètres suivants ont
été mesurés et analysés:
- Biomasse aérienne et racinaire déterminée
par pesée,
- Surface foliaire estimée en utilisant le logiciel
d'analyse d'images (WinRhizo)
- Transpiration (mesurée par voie
gravimétrique): la transpiration a été
évaluée par la mesure de la perte de poids d'une plante en pot
non arrosée, en prenant la précaution d'éviter que l'eau
ne s'évapore du pot en couvrant la surface du sol avec feuille de
plastique qui ne laisse traverser que la tige.
- Déficit de pression de vapeur : il se calcule sur la
base de la température et de l'humidité relative.
4.5. Analyse des données
Les données recueillies ont été soumises
à une analyse non-paramétrique et les différences entre
les moyennes ont été appréciées sur la base de
l'erreur standard.
15
III. TROISIEME PARTIE : RESULTATS
3.1 Etude de l'anatomie de Cenchrus et Zornia
- Stomates
L'observation de l'épiderme de Cenchrus biflorus
montre des cellules stomatiques de forme allongée, plus ou moins
régulière, délimitées par les nervures. Ces
cellules entourent des stomates de très petite taille constitues deux
cellules de garde allongées entourant l'ostiole (Photo 4). Par contre
l'épiderme de Zornia glochidiata possède des cellules
stomatiques anisocytique au niveau desquelles sont logés les stomates.
Ces derniers sont de forme ovoïde et constitués de deux cellules de
garde réniformes délimitant l'ostiole (Photo 5). La
densité moyenne de stomates est de 20 stomates.mm-2 chez
C.biflorus et 12 stomates.mm-2 chez
Z.glochidiata.
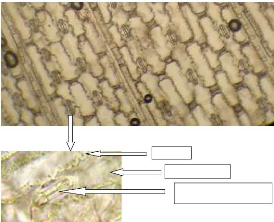
Nervure
subérifiées
Cellule Epidermique
Stomate avec lèvres de l'ostiole
Photo 4 : Observation de l'épiderme de
Cenchrus biflorus au microscope photonique (a- x100 et b-x400)
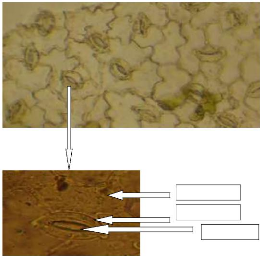
16
Photo 5 : Observation de l'épiderme de
Zornia glochidiata au microscope photonique (a-x100 et b-x400)
Cellule Epidermique
Cellule de garde
Ostiole
- Tige
Chez Z.glochidiata, la structure anatomique de la tige
montre la présence de:
- l'épiderme délimitant le parenchyme cortical vers
l'extérieur.
- le parenchyme cortical, situé sous l'épiderme. Il
est composé de cellules de
forme arrondie et moins bien organisées. Elles sont
compactées les unes contre les
autres renforçant ainsi le rôle de barrière
à l'épiderme.
- les cellules du phloème, dispersées par endroit,
se localisant au niveau de la
partie la plus interne du parenchyme corticale.
- les cellules du xylème se disposant
généralement en groupe de 3 au niveau du
parenchyme lignifié (sclérenchyme) situé en
dessous du parenchyme cortical.
- le parenchyme médullaire constituant la partie centrale
avec des cellules de
grande taille qui plus tard sont remplacées par une lacune
centrale (Photo 6).
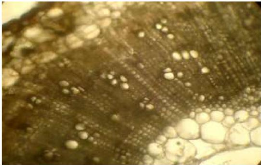
17
Photo 6 : Coupe transversale de la tige de
Z.glochidiata (x400) observée au microscope photonique.
Chez C. biflorus, la structure anatomique de la tige
montre la succession de:
- l'épiderme délimitant le parenchyme cortical vers
l'extérieur. - le parenchyme cortical, situé sous
l'épiderme. Il est très réduit.
- le parenchyme médullaire très
développé qui occupe une position centrale avec des cellules de
grande taille.
- les faisceaux libero-vasculaires en nombre
élevé et distribués dans la zone
périphérique du parenchyme médullaire (Photo 7).
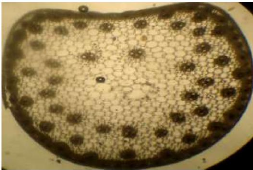
18
Photo 7 : Coupe transversale de la tige de
C.biflorus observée au microscope photonique (x400).

Photo 8 : (A) portion de la structure anatomique
de Z.glochidiata et (B) de C.biflorus (x400)
(A) : (1) parenchyme lacuneux, (2) bois primaire, (3) bois
secondaire, (4) cambium, (5) phloème, (6) épiderme
(B) : (1) parenchyme, (2) faisceau libero-vasculaire, (3)
sclérenchyme, (4) épiderme
Ces photos montrent une différence au niveau de
l'organisation des tissus selon que l'espèce est dicotylédone (A)
ou monocotylédone (B).
18 16 14 12
10 8 6 4 2
0
3.2. Réponse de la croissance au broutage
simulé
3.2.1 Effet des coupes sur la production de biomasse
chez Cenchrus biflorus
cultivé sur placettes
La réponse de la croissance (biomasse) au broutage
simulé a été suivie en réalisant deux coupes
successives sur les plantes à deux niveaux (3cm et 5cm du sol). Les
résultats ont montré une réponse différentielle en
fonction du niveau et du nombre de coupes (Figure 2). Au niveau de la
première coupe, la biomasse produite ne montre pas une différence
significative entre les deux niveaux de coupe. Par contre au niveau de la
deuxième coupe, la biomasse produite chez les plantes coupées
à 5 cm du sol ont produit plus du double de la biomasse produite chez
celles coupées à 3 cm du sol.
A la récolte, les données montrent que la
production de biomasse n'a pas significativement varié entre les plantes
témoins et celles coupées deux fois à 3 cm du sol (Figure
3). Par contre, chez les plantes coupées à 5 cm du sol, la
biomasse a augmenté de plus du 1/3 par rapport aux plantes
témoins.
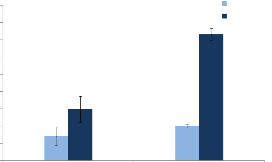
1ère Coupe
2ème Coupe
19
Coupe 3cm Coupe 5cm
Figure 2 : Production de biomasse chez
Cenchrus biflorus pour différents niveaux de coupes (3 cm et 5
cm)
2,5
2
1,5
1
45 40 35 30
25 20 15 10 5
0
Témoin Coupe à 3 cm Coupe à 5
cm
Figure 3 : Effet des coupes à 3 cm et 5
cm du sol sur la production de biomasse totale chez Cenchrus
biflorus.
3.2.2 Effet des coupes sur la production de biomasse chez
Cenchrus biflorus cultivé dans des pots
Le même type de traitement a été
effectué au niveau des plants de C. biflorus. Les
données ont montré que selon le niveau de coupe, la
réponse des plantes est variable (Figure 4). Ainsi, la production de
biomasse a été significativement plus élevée avec
les coupes à 5 cm par rapport à celles effectuées à
3 cm du sol.
0,5
0
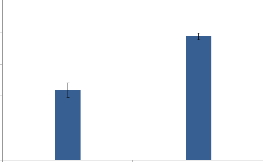
20
Coupe 3cm Coupe 5cm
Figure 4 : Production de biomasse chez
Cenchrus biflorus pour différents niveaux de coupes (3 cm et 5
cm)
A la récolte, les résultats obtenus montrent que
les plantes coupées à 3 cm ont produit moins de biomasse que les
plantes témoins. Dans le cas des plantes coupées à 5 cm du
sol, la production de biomasse tend à être supérieure
à celle des plantes témoins (Figure 5). Ces résultats
contrastent beaucoup avec ceux enregistrés chez Z.
glochidiata.
21
Témoin Coupe à 3 cm Coupe à 5
cm
Figure 5 : Effet des coupes à 3 cm et
5 cm du sol sur la production de biomasse totale chez Cenchrus
biflorus.
3.2.3 Effet des coupes sur la production de biomasse
chez Zornia cultivé sur placettes
A la récolte, les résultats montrent un effet
négatif des coupes à 3 cm qui réduisent du 1/3 la biomasse
comparée à celle des plantes témoins. Les coupes à
5cm n'ont pas montré d'effet significatif, la biomasse produite
n'étant pas différente de celle des plantes témoins
(Figure 6).
22
8 7 6 5 4
3 2 1 0
Témoin Coupe à 3 cm Coupe à 5
cm
Figure 6 : Effet des coupes à 3 cm et
5 cm du sol sur la production de biomasse totale chez Zornia
cultivé sur placette
3.2.4 Effet des coupes sur la production de biomasse
chez Zornia cultivé dans des pots
Les résultats obtenus concernant les plantes
cultivées dans les pots montrent une augmentation significative de
biomasse (presque le double) chez les plantes
coupées à 3cm par rapport aux plantes
témoins (Figure 7). Celles coupées à 5cm montrent un effet
similaire avec une plus grande biomasse que celle obtenue chez les plantes
coupées à 3cm.
3,5 3 2,5 2
1,5 1 0,5 0
Témoin Coupe à 3 cm Coupe à 5
cm
Figure 7 : Effet des coupes à 3 cm et
5 cm du sol sur la production de biomasse totale chez Zornia
cultivé en pots
2
1

3.3. Réponse de la transpiration à la
variation journalière du déficit de pression de vapeur chez
Cenchrus
La figure 8A indique qu'au cours de la journée 1, le
deficit de pression de vapeur (DPA) montre une faible augmentation entre 07h et
13h, la valeur maximale étant 2,25 kPa. Au dela de 13h, le DPV diminue
de facon graduelle jusqu'à une valeur de 1,4 kPa vers 19h. Contrairement
à la journée 1, le DPV montre une forte variation au cours de la
journée 2 (Figure 8B). De 7h à 13H, il augmente rapidement pour
atteindre un pic à une valeur de 4 kPa autour de 13-14H. Au-delà,
la valeur de ce paramètre diminue progressivement pour tomber à 2
kPa en fin de journée.

0
7-10H 10-13H 13-16H 16-19H
23
0
7-10H 10-13H 13-16H 16-19H
Figure 8 : Variation du déficit de
pression de vapeur (A) au cours d'une journée nuageuse et (B) au cours
d'une journée très ensoleillée. Les données ont
été enregistrées au niveau du site de l'Université
de Maradi.

100
90
80
70
60
50
40
30
20
10
0
La figure 9 montre l'évolution de la transpiration au
cours de la journée 1. Elle est faible tout au long de la journée
quel que soit le traitement. En début de matinée, le niveau de
coupe ne montre aucun effet significatif sur la transpiration. Par contre en
fin de matinée, les plantes coupées montrent une tendance
à une plus faible transpiration par rapport aux plantes témoins.
Le reste de la journée montre des transpirations très proches
pour tous les traitements (Figure 9).
Témoin
coupe à 5cm
coupe à 3cm

24
10h 13h 16h 19h
Figure 9 : Variation de la transpiration en
fonction du temps au cours d'une journée nuageuse chez
Cenchrus
Dans le cas de la journée 2, les résultats
contrastent beaucoup avec ceux de la journée 1 (Figure 10). En
début de journée, aucune différence n'est
enregistrée entre les différents traitements. Par contre, en fin
de matinée, les plantes témoins montrent une forte transpiration
qui atteint 90 mg plante-1. Les coupes ont eu un effet
négatif très prononcé sur la transpiration. En effet, la
transpiration des plantes coupées ne représente que moins du
tiers de celle des plantes témoins. Chez les plantes coupées
à 5 cm, la transpiration montre une augmentation significative par
rapport à celle enregistrée en début de matinée et
elle est significativement supérieure à celle des plantes
coupées à 3 cm. En début d'après-midi, les
différences de transpiration entre les plantes témoins et celles
coupées restent encore très importantes car la transpiration des
témoins est double de celle des plantes coupées. En fin de
journée, la transpiration est faible quel que soit le traitement, mais
légèrement supérieure chez les plantes coupées
(Figure 10).
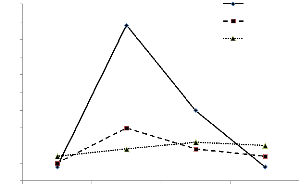
Témoin
coupe à 5cm
coupe à 3cm
70
60
50
40
30
20
10
0
10h 13h 16h 19h
100
90
80
25
Figure 10 : Variation de la transpiration en
fonction du temps au cours d'une journée très ensoleillée
chez Cenchrus
26
IV. QUTRIEME PARTIE : DISCUSSION
Les résultats de l'étude anatomique de
l'épiderme foliaire, de la tige et de la racine chez les deux
espèces étudiées montrent des différences
structurelles liées à leur appartenance phylogénique. Chez
C.biflorus les stomates sont très petits et disposés de
manière alignée par rapport à ceux de Z. glochidiata
qui sont de plus grande taille et de forme anisocytique.
La distinction entre ces deux groupes s'effectue selon
plusieurs types de critères que l'on peut hiérarchiser de la
manière suivante : l'organisation des cellules épidermiques, les
types de stomates. Les cellules épidermiques des graminées sont
généralement allongées et disposées en files
parallèles aux nervures (parallèles entre elles). En ce qui
concerne l'orientation des nervures des monocotylédones, on constate
que, contrairement aux dicotylédones, elles sont
généralement parallèles. Les stomates de
dicotylédones sont de 4 types principaux alors que chez les
monocotylédones, on distingue un type unique aux lèvres de
l'ostiole subérifiées (G. Mandret,
1989).
Au niveau de la tige, l'étude anatomique montrent une
différence nette entre la tige de graminée (C. biflorus)
et celle de légumineuse (Z. glochidiata). Cette
différence s'expliquerait par le fait que les monocotylédones ne
présentent pas des tissus secondaires, il existe uniquement des tissus
primaires tandis que chez les dicotylédones on rencontre des tissus
secondaires.
En général, chez les espèces
fourragères, la biomasse est le paramètre le plus souvent
utilisé pour caractériser la production. L'opération de
coupe menée sur les génotypes de C.biflorus et de
Z.glochidiata sur les placettes et dans les pots a donné des
résultats spécifiquement différents selon le niveau de
coupe (3 cm et 5 cm) et le nombre de coupes. Dans la plupart des cas, les
coupes ont favorisé la production de biomasse. Ces résultats sont
similaires à ceux publiés par R. TellaP, M. Oarro, M. Barbero
(1999) qui ont montré que la méthode des coupes
répétées au même endroit constitue un stimulus qui
accroît la production végétale. La production de biomasse
en réponse aux coupes à 5 cm du sol est plus importante pour les
deux espèces mais plus
27
surtout sur les placettes chez Cenchrus et dans les
pots chez Zornia. Cette différence s'expliquerait par le fait
que l'espèce Cenchrus préfère un milieu ouvert
à cause de ses racines très longues et ramifiés par
rapport au pot qui fournit des conditions défavorables à
l'expression du potentiel des plantes du fait du confinement du système
racinaire.
La faible production de ces espèces à 3cm du sol
sur les deux milieux de culture indique que ce niveau de coupe simule un
broutage relativement sévère qui ne permet pas une
régénération rapide de la biomasse. Cependant, la
limitation de la régénération de la biomasse avec les
coupes à 3 cm n'affecte pas de la même façon les deux
espèces. Chez Cenchrus, ce niveau de coupe a même eu un
effet négatif sur la biomasse comparée à celle des plantes
témoins. Ce qui traduit une plus grande sensibilité au broutage
sévère chez Cenchrus par rapport à
Zornia.
L'effet du DPV sur la transpiration des plantes témoins
montre une légère augmentation au cours de la journée
nuageuse quel que soit le traitement. Par contre au cours de la journée
ensoleillée, la transpiration a montré une importante
augmentation qui résulte de l'augmentation du DPV au cours de cette
journée. En effet, une augmentation du DPV, entraine l'ouverture des
stomates (Serge Hamon, 2007), avec une élévation
subséquente de la transpiration. Les plantes coupées montrent une
augmentation de la transpiration largement plus faible du fait d'une surface
foliaire moins importante. Ce qui peut constituer un atout en conditions de
sécheresse car la plante doit gérer le peu d'eau qui lui est
disponible.
28
CONCLUSION
Au regard de tous ces résultats, il ressort clairement
que le broutage (simulé par les coupes) peut avoir des effets positifs
comme négatifs sur la production de biomasse selon son intensité
(niveau de coupe). Un broutage sévère réduit tellement
l'appareil photosynthétique au point que la plante n'est plus en mesure
d'assurer une bonne régénération de sa biomasse. Par
contre un broutage modéré, stimule la production de biomasse qui
dans la plupart des cas dépasse celle des plantes non-broutées.
En outre, en réduisant la perte d'eau par transpiration chez les
plantes, le broutage peut leur permettre de mieux tolérer le
déficit hydrique. Ce qui implique qu'une bonne gestion des parcours peut
favoriser une bonne régénération de ceux-ci avec en plus
une réduction sensible de l'effet de la sécheresse.
Ce travail qui constitue une étude préliminaire,
mérite d'être poursuivi par l'analyse d'un large nombre
d'espèces fourragères afin de sélectionner celles qui
présentent les meilleures aptitudes à supporter les effets
conjugués de la pression du broutage et de la sécheresse au
niveau des parcours.
29
REFERENCES BIBLIOGRAPHIQUES
Berhaut, J., 1967. Flore du
Sénégal. 2nd Edition. Editions Clairafrique, Dakar,
Sénégal. 485 pp.
Banoin M. Gueye, C. Soumana, I. Ali M. Jouve P.,
1996. Péjorations climatiques et évolution des pratiques de
transhumance en zone agropastorale sahélienne cas de l'arrondissement de
Mayahi, au Niger. pp. 43-52.
Burkill, H.M., 1994. The useful plants of
West Tropical Africa. 2nd Edition. Volume 2, Families E-I. Royal Botanic
Gardens, Kew, Richmond, United Kingdom. 636 pp.
Bode S., 2004. Pratiques pastorales et
biodiversité des parcours de Dantiandou. Mémoire de fin
d'étude D.E.S.S (CRESA), Faculté d'agronomie de
L'université Abdou Moumouni de Niamey.
Briske, D.D., et J.H. Richards.1995 «
Plant Responses to Defoliation: a Physiological, Morphological and Demographic
Evaluation ». In D.J. Bedunah et R.E. Sosebee (éd.), Wildland
Plants: Physiological Ecology and Developmental Morphology. Soc. For Range
Management. Denver, Co., p. 635-710.
FAO, 1970. Amino-acid content of foods and
biological data on proteins. FAO Nutrition Studies No 24, Rome, Italy. 285
pp.
G. Mandret, 1989. Le régime
alimentaire des ruminants domestiques (bovins-ovins-caprins) sur les
pâturages naturels sahéliens et soudano-sahéliens. Revue
Sénégalaise des Recherches Agricoles et Halieutiques - Vol. 2 -
n°2 - 1989
Levitt J., 1980. Responses of plants to
environmental stresses, Water, radiation, salt and other stresses. Acad. Press
Inc., 2, 606 p.
Monneveux P., 1997. La génétique
face au problème de la tolérance des plantes cultivées
à la sécheresse : espoirs et difficultés.
Sécheresse, 2, 29-37.
30
N. Mbaye, A.T. Dioup, M. Gueye, A.T. Diallo, C.E.
Sall, P.I. Samb, 2002. Etude du comportement germinatif et essais de
levée d'inhibition tégumentaire des graines de Zornia
glochidiata Reich.ex DC., Légumineuse fourragère. Revue
Elev. Medi. vet. Pays trop. , 55(1):47-52.
Pierre Hiernaux, Henry Noël le
Houérou.2006. Les parcours du Sahel. Article scientifique.
Science et changement planétaires/Sècheresse, Volume 17,
Numéro 1, 51-71.
Popolizio, C.A., H. Goetz, et P.L. Chapman.1994
« Short-term Response of Riparian Vegetation to 4 Grazing
Treatments ». J. Range Management, vol. 47, p. 4853.
R. TellaP, M. Oarro, M. Barbero. Mai 1999.
Productivité herbacée des périmètres
améliorés du Sahel marocain : Effet des conditions stationnelles,
climatiques et du rythme de coupe. Fourrage, 158,149-156.
Serge HAMON.2007. L'amélioration de la
résistance à la sécheresse peut-elle être base sur
les méthodes de sélection traditionnelle et/ ou sur les
méthodes biotechnologiques modernes Possibilités et limites
respectives. Actes des JSIRAUF, 6-9 novembre 6 p.
Thébaub B., 1999. Gestion de l'espace et
crise pastoral au Sahel. Etude comparative du Niger et du Yatanga
burkinabé. Thèse de doctorat, Ecole des hautes Etudes en sciences
sociales, Paris/France, 476p.
United Nations, 1996. World population
protected. The 1996 revision annexes I and II. Demographic indicators by major
eaea, region and country. New york. U.N. 441p.
a
Annexes
Annexe 1 : Dispositif de culture au niveau de L'JUT
|
Cenchrus
|
Kz NPK
|
K+ NPK
|
Cenchrus
|
K+ Témoin
|
Kz Urée
|
K+ Urée
|
Cenchrus
|
Kz Témoin
|
|
Kz Témoin
|
K+ Témoin
|
Zornia
|
Kz NPK
|
Zornia
|
K+ NPK
|
cenchrus
|
Kz Urée
|
K+ Urée
|
|
K+ Témoin
|
Cenchrus
|
Kz Témoin
|
K+ urée
|
Kz Urée
|
Zornia
|
Kz
NPK+ Urée
|
K+
NPK+ Urée
|
Cenchrus
|
b
Annexe 2 : Données du Mesure de la transpiration
par voie gravimétrique
|
jeudi 7h
|
Témoin Coupe à 5cm
|
|
Coupe à 3cm
|
Vendredi 7h
|
Témoin
|
coupe à 5cm
|
coupe à 3cm
|
|
P1
|
4,28
|
4,36
|
4,6
|
P1
|
4,2
|
4,34
|
4,57
|
|
P2
|
4,77
|
4,69
|
4,69
|
P2
|
4,72
|
4,66
|
4,5
|
|
P3
|
4,54
|
4,26
|
4,41
|
P3
|
4,49
|
4,23
|
4,38
|
|
P4
|
4,74
|
4,09
|
4,24
|
P4
|
4,69
|
4,06
|
4,21
|
|
P5
|
4,4
|
4,73
|
4,46
|
P5
|
4,36
|
4,69
|
4,43
|
|
10h
|
|
|
|
10h
|
|
|
|
|
P1
|
4,27
|
4,36
|
4,6
|
P1
|
4,4
|
4,34
|
4,54
|
|
P2
|
4,74
|
4,67
|
4,52
|
P2
|
4,71
|
4,64
|
4,49
|
|
P3
|
4,54
|
4,25
|
4,41
|
P3
|
4,49
|
4,22
|
4,36
|
|
P4
|
4,69
|
4,09
|
4,24
|
P4
|
4,68
|
4,05
|
4,2
|
|
P5
|
4,4
|
4,72
|
4,47
|
P5
|
4,36
|
4,68
|
4,43
|
|
13h
|
|
|
|
13h
|
|
|
|
|
P1
|
4,24
|
4,36
|
4,59
|
P1
|
4,15
|
4,3
|
4,53
|
|
P2
|
4,72
|
4,68
|
4,53
|
P2
|
4,68
|
4,61
|
4,48
|
|
P3
|
4,52
|
4,26
|
4,42
|
P3
|
4,43
|
4,2
|
4,34
|
|
P4
|
4,72
|
4,08
|
4,25
|
P4
|
4,63
|
4,03
|
4,18
|
|
P5
|
4,39
|
4,71
|
4,47
|
P5
|
4,31
|
4,64
|
4,4
|
|
16h
|
|
|
|
16h
|
|
|
|
|
P1
|
4,22
|
4,34
|
4,58
|
P1
|
4,11
|
4,28
|
4,52
|
|
P2
|
4,72
|
4,66
|
4,5
|
P2
|
4,64
|
4,6
|
4,45
|
|
P3
|
4,5
|
4,24
|
4,38
|
P3
|
4,38
|
4,18
|
4,32
|
|
P4
|
4,71
|
4,07
|
4,22
|
P4
|
4,58
|
4
|
4,16
|
|
P5
|
4,38
|
4,69
|
4,45
|
P5
|
4,29
|
4,63
|
4,37
|
|
19h
|
|
|
|
19h
|
|
|
|
|
P1
|
4,22
|
4,33
|
4,58
|
P1
|
4,1
|
4,28
|
4,51
|
|
P2
|
4,7
|
4,65
|
4,5
|
P2
|
4,63
|
4,58
|
4,44
|
|
P3
|
4,48
|
4,23
|
4,37
|
P3
|
4,37
|
4,17
|
4,29
|
|
P4
|
4,7
|
4,05
|
4,21
|
P4
|
4,58
|
3,99
|
4,14
|
|
P5
|
4,36
|
4,68
|
4,44
|
P5
|
4,28
|
4,6
|
4,34
|
C
Annexe 3: Données du Mesure de biomasse du
cenchrus Biflorus
d
|
p1
|
P2
|
P3
|
|
P4
|
|
P5
|
P6
|
|
Témoin
|
25,09
|
20,09
|
|
21,85
|
|
29,11
|
|
|
|
Témoin
|
25,66
|
23
|
|
35,32
|
|
31,3
|
|
|
|
Témoin
|
19,44
|
27
|
|
26,57
|
|
19,57
|
|
|
|
Témoin
|
34,87
|
24,79
|
|
29,33
|
|
23,33
|
|
|
|
Témoin
|
10,34
|
18,98
|
|
23,2
|
|
13,22
|
|
|
|
5 Cm
|
24,49
|
|
|
20,43
|
|
|
|
|
|
5 Cm
|
18,32
|
|
|
19,89
|
|
|
|
|
|
5 Cm
|
10,89
|
|
|
34,51
|
|
|
|
|
|
5 Cm
|
11,93
|
|
|
32,87
|
|
|
|
|
|
5 Cm
|
9,83
|
|
|
28,92
|
|
|
|
|
|
3 Cm
|
|
9,98
|
|
|
|
19,23
|
|
|
|
3 Cm
|
|
7,31
|
|
|
|
27,91
|
|
|
|
3 Cm
|
|
10,02
|
|
|
|
17,35
|
|
|
|
3 Cm
|
|
12,84
|
|
|
|
27,99
|
|
|
|
3 Cm
|
|
10,13
|
|
|
|
20,9
|
|
|
|
Nbre Pieds
|
P1:72
|
P2:88
|
P3:67
|
|
P4:34
|
|
P5:22
|
P6:13
|
e
Annexe 4: Données du Mesure de biomasse du
Zornia glochidiata
9
|
P1 P2 P3 P4
|
|
P5 P6
|
|
Témoin
|
6,09
|
7,67
|
5,96
|
6,43
|
|
|
Témoin
|
6,97
|
12,05
|
5,58
|
8,8
|
|
|
Témoin
|
5,98
|
6,45
|
4,03
|
7,63
|
|
|
Témoin
|
5,1
|
7,72
|
8,73
|
7,99
|
|
|
Témoin
|
5,31
|
5,83
|
6,26
|
6,11
|
|
|
5 Cm
|
|
|
5,48
|
4,19
|
|
|
5 Cm
|
|
|
7,63
|
5,54
|
|
|
5 Cm
|
|
|
5,71
|
3,57
|
|
|
5 Cm
|
|
|
5,27
|
3,93
|
|
|
5 Cm
|
|
|
5,02
|
2,52
|
|
|
3 Cm
|
2,57
|
4,35
|
|
|
|
|
3 Cm
|
2,72
|
2,56
|
|
|
|
|
3 Cm
|
4,49
|
3,15
|
|
|
|
|
3 Cm
|
2,36
|
2,99
|
|
|
|
|
3 Cm
|
2,84
|
2,92
|
|
|
|
|
Nbre Pieds
|
18
|
23
|
13
|
15
|
6
|
|
|


