|
REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET
POPULAIRE
Ministère de l'enseignement supérieur et de la
recherche scientifique
|
Institut des Sciences Vétérinaires- Blida
|
Université Saad Dahlab-Blida 1-
|
|
Année : 2015/2016
Projet de fin d'études en vue de l'obtention
du
Diplôme de Docteur Vétérinaire
Le lapin de population locale
algérienne
(Oryctolagus cuniculus) :
Performances de
reproduction, de croissance et état
sanitaire
(Synthèse
bibliographique)
Présentés par
Mazari Fetheddine et Kaci Sidahmed
Soutenu le
|
Devant le jury :
|
|
|
|
|
Président(e) :
|
Boumahdi Merad Z
|
Maître de Conférences A
|
USDB
|
|
Examinateur :
|
Salhi O
|
Maître-assistant A
|
USDB
|
|
Promoteur :
|
Belabbas R
|
Maître-assistant A
|
USDB
|
Merci à tous. ?
Remerciements
La réalisation d'un travail de fin
d'études n'est pas seulement un travail de longue
haleine mais aussi une formidable expérience scientifique.
Bien que délicate, l'écriture des
remerciements est un élément indispensable pour
témoigner notre profonde
reconnaissance. Nous tenons tout d'abord
à exprimer nos sincères remerciements aux membres du jury :
A Dr Boumahdi Merad Z. Maître de
Conférences A. à l'Institut des Sciences
Vétérinaires, Université de Blida, qui nous a
fait l'honneur d'accepter la présidence de notre jury de mémoire
de fin d'études. Qu'elle reçoive ici nos hommages respectueux.
A Dr Salhi O. Maitre-Assistant A à l'Institut
des Sciences Vétérinaires, Université de Blida,
qui nous a fait l'honneur d'accepter de faire partie de notre jury de
thèse, pour avoir accepté de juger notre travail. Qu'il trouve
ici l'assurance de nos très sincères remerciements.
A Dr Belabbas R. Maitre-Assistant A à
l'Institut des Sciences Vétérinaires, Université de Blida,
qui nous a fait l'honneur d'accepter notre sujet et d'encadrer notre
travail. Pour son soutien sans faille mais aussi pour son authentique
humanité qui fait écho à notre sujet. Qu'il reçoive
ici l'expression de notre reconnaissance ainsi que l'assurance de notre plus
profonde gratitude.
Nous voulons aussi adresser nos remerciements
à tous les enseignants de l'Institut
des Sciences Vétérinaires de l'Université
de Blida. Qu'ils reçoivent ici l'expression de notre reconnaissance et
nos chaleureux remerciements.
Nous tenons à remercier aussi les agents
de la bibliothèque de l'Institut des
Sciences Vétérinaires de l'Université de
Blida notamment Ms Riad, pour leur aide, leur patience, leur
sourire et leur compréhension.
Enfin, nous adressons ce remerciement
à toutes les personnes qui ont contribué de loin ou de
près à la réalisation de ce travail.
Dédicace
Je dédie ce modeste travail :
A ma raison de vivre, mes chers parents Tayeb
et Djamila pour leur confiance, tout leur sacrifice,
patience et tendresse.
A
tous mes chères soeurs : Malika, Akila,
Ghania, Bouchra, Soussou, à tous mes neveux et mes
nièces : Maria, Melissa, Nazim, Ranim, Anes et
Nihal, a tous mes beaux-frères, avec mes souhaits de
bonheur, de santé et de succès, je vous aime.
A ma très chère grande mère
Tounes que dieu la garde en vie , à la
mémoire de Menad, Bouzid et Tounes :
mes grands-parents et aussi à la mémoire de ma tante
Samia et ma cousine Aicha : que dieu les
accueille dans son vaste paradis.
A mes tantes Fatima, Fatiha
et Zohra et leurs maris, à tous mes oncles :
Azzedine, Noureddine, Ahsen, Hocine, Belkacem, Nacer-eddine, Mohamed
et Ali, ainsi que leurs femmes.
A tous mes cousins : Samir, Hamid, Mustapha,
Abd-samia, Ahmed, Mourad, Mohamed et
leurs femmes, Taoues et Nacera, Khadîdja, Amel,
Hakima et leurs maris, à Brahim, Mohamed, Kheiro,
Khalil, Ghanou, Abdou, Khaled, Akram, Amina, Hassina, Zineb, Khalida, Loubna,
Dounia, Weissem, Farida,
Soumia et la petite Hadil, que dieu vous
préserve longue vie et prospérité.
Dédicace spécial pour mon cher frère,
mon alter ego : Islam
A mes très chers amis :Redha, Med Boubaker,
Saleh, koukou , Amine, Da Meziane, Sid Ahmed, Nabil, Med boukhari , Ali, Latif,
Mounir, Chérif, , Meriem, Ahlem ,
Anissa , Bouchra et spécialement
Saïd, aucun mot, aucune dédicace ne peut exprimer
mon énorme amour.
A tous les membres de l'association Jil El
ihsan, merci pour tous les bons moments qu'on a passé
ensemble.
Et enfin à tous ceux qui m'aiment et à tous ceux
que j'aime.
Feth-eddine ?
Sid-ahmed?
Dédicace
Au nom d'Allah le très miséricordieux, je tiens
à le remercier de m'avoir donné le courage et la patience
d'arriver à ce stade afin de réaliser ce modeste travaille que je
dédie à:
Mes parents, tous les mots du monde ne sauraient exprimer
l'immense amoure que je vous porte, ni la profond gratitude que je vous
témoigne pour tous les efforts et les sacrifices que vous n'avez jamais
cessé de consentir pour mon instruction et mon bien-être.
Mon frère Mohamed pour son
soutien moral , qui a été une source de joie et de bonheur
m'encourageant à fournir plus d'efforts.
Mon beau frère Abdelkader et mes
soeur Meriem et Amina et
ses filles Naziha ,Wafae ,
jomana. . Spéciale dédicace à
mes grands-parents paternelles et maternelles, puisse dieu vous protéger
de tout mal.
Mes oncles AbdElwahabe
,khaled ,Laid
et Rabeh et mes tantes
Houria Zohra ,Fatma
,Luiza , Razika
et spécialement Khadija ,mes
cousins Abd allah Sidali , Younas
et mes cousines Soumia ,
Sara , Faiza ,
Yasmin ,Souad et
Marwa veuillez accepter l'expression de ma profonde
gratitude pour votre soutien ,encouragements et affection.
Mon Ami Abd el Rahman tu occuperas
toujours une partie de ma vie et de mon coeur. A Lazhar
et Rodwan .
Mes amis d'enfance:
Hamoudat
Abdo, Moustafa , Amine , Nassim
,Chawki , Sadam
,Rafik Djamal ,Mohamed ,merci pour
votre générosité et votre soutien.
Mes amis de l'institut vétérinaire:
Nabil ,Fethi , Latif , Ali ,Belkacem ,Mounir, Mohamed
,Cherif le grand Abd el rezak merci
pour l'ambiance et les bons moments qu'on a passé tout au long des cinq
années d'étude .J'espère que nous saurons entretenir cette
belle amitié. Docteurs Sidahmed
,Brahim et ,Bouzid
pour leur précieux conseils .
Tous ceux que j'aime et qui m'aiment, je dédie ce
mémoire qui j'espère être à la hauteur de leur
espérance et la mienne aussi.
L'objectif de notre travail est de faire le point sur les
travaux réalisés sur la croissance, la reproduction ainsi que
l'état sanitaire du lapin de population locale algérienne
(Oryctolagus cuniculus).
Le lapin local se caractérise par un taux moyen de
fertilité de 87 %, une réceptivité de 89 %, une
prolificité de 7,2 et une mortinatalité de 18,9% avec cependant,
une grande variabilité. Les performances zootechniques liées
à la reproduction du lapin local, sont influencées par des
facteurs liés à l'animal notamment l'âge, état
physiologique et le type génotype de celui-ci et des facteurs
liées à l'environnement à l'exemple de l'alimentation, la
température environnementale, la lumière et la saison. Enfin, des
facteurs liés à la conduite d'élevage notamment le mode et
le rythme de reproduction peuvent influencer également ses performances.
Le lapin local se caractérise aussi par un poids moyen de 49,4g à
la naissance, de 463,7g au sevrage, 621g à l'âge de 6 semaines et
de 1433g à l'âge de la 11ème semaine. Durant
toute la période d'engraissement, l'indice de consommation moyen est de
3,00 avec un gain moyen quotidien de 22,72 g/J. A l'abattage, le rendement de
la carcasse chaude est considéré comme satisfaisant (69,23%). La
croissance du lapin local est influencée par les mêmes facteurs
qui contrôlent ses performances de reproduction.
Enfin, le taux d'infection par Escherichia coli et
par Clostridium perfringens est de 36,11% et de 25% respectivement.
L'infestation par Cryptosporidium spp est assez fréquente chez
le lapin local (83,33%). Le lapin local a fait l'objet de plusieurs
études sur la coccidiose et qui ont montré un taux
d'excrétion oocystale faible ou nulle au cours de la gestation mais
très importante au cours de la lactation avec une prédominance
d'Eimeria magna (43%).
Mots clés : Lapin local,
fertilité, prolificité, croissance, facteurs de variation,
rendement, carcasse, pathologies.
ÉíáÍãáÇ
ÉááÇÓáÇ äã
ÈäÑáá
ÉíÍÕáÇ
ÉáÇÍáÇæ
ÑËÇßÊáÇæ
æãäáÇ ìáÚ
ÒÌäãáÇ
áãÚáÇ
ãííÞÊá æå
ÇäáãÚ äã
ÏåáÇ
ÉíáÍãáÇ
ÉááÇÓáÇ äã
ÈäÑáá
ÉíÍÕáÇ
ÉáÇÍáÇæ
ÑËÇßÊáÇæ
æãäáÇ
ìáÚ
ÒÌäãáÇ
áãÚáÇ
ãííÞÊá æå
ÇäáãÚ
äã ÏåáÇ
.)
Óáíßíäíß
ÓíÞáÇæÊßíÛæÇ
(
ÉíÑÆÇÒÌáÇ
.)
Óáíßíäíß
ÓíÞáÇæÊßíÛæÇ
(
ÉíÑÆÇÒÌáÇ
|
ÏäÚ Êæã
áÏÚã æ
|
7,2 ÈÇÌäÇ
áÏÚã ,%89 áæÈÞ
áÏÚã
|
,
|
%
|
87
|
ìáÅ
|
ÉÈæÕÎáÇ
|
áÏÚã
|
ØÓæÊãÈ
|
íáÍãáÇ
|
ÈäÑáÇ
ÒíãÊí
|
ÈäÑáÇÈ
ÉÞáÚÊã
áãÇæÚÈ
ÉíáÍãáÇ
ÈäÇÑáÇ äã
ÈÇÌäáÅÇÈ
ÞáÚÊí Çãí
äÇæíÍáÇ ÁÇÏ
ÑËÊÊ .ÊÇÊæÇÊ
ÏæÌæ Úã %18,9
ÉÏáÇæáÇ
íÆÇÐÛáÇ
ãÇÙäáÇ áËã
ÉÆíÈáÇÈ
ÉÞáÚÊã
áãÇæÚæ åá
íäíÌáÇ
ØãäáÇ
ÉÞáÚÊãáÇ
áãÇæÚáÇ
ÇÑíÎæ
|
ÏÞ
ÑËÇßÊáÇ
áÏÚãæ
|
ÉÞíÑØ
|
ßáÐ
|
í
|
ÇãÈ
ÈäÇÑáÇÇ
ÉíÈÑÊ ÑíÓÈ
|
. ãÓæãáÇ
ÁæÖ æ
ÉíÆíÈáÇ
|
Úæäæ
ÉíÌæáæíÓ
|
ÉáÇÍ
äÇæíÍáÇ
|
äÓ ßáÐ
|
í
|
ÇãÈ
|
|
ÉÑÇÑÍáÇ
|
ÉÌÑÏæ
|
ìáÚ ÇÖí
ÑËÄÊ
ÚíÈÇÓ 6 äÓ
í Û 621 Û7 463 ãÇØáÇ
ÉÏáÇæáÇ ÏäÚ
Û4 49 äÒæ
ØÓæÊãÈ ÇÖí
íáÍãáÇ
ÈäÑáÇ ÒíãÊí
.Á ÇÏáÇ
ÈÓÇßã Úã 3.00
ßáÇåÊÓáÇÇ
ÑÔÄã
ÉíáÍãáÇ
ÈäÇÑáÇ æãä
ÑËÊí .)% 69.23
íáÍãáÇ
ÈäÑáÇÇ ÛáÈ
äíãÓÊáÇ
ÉÑÊ
1433æ
ØÓæÊã
áÇæØ .1 1 áÇ
ÚæÈÓáÇ äÓ í
Û
(
|
ÉíÖÑã
|
ÉäÎÇÓáÇ
ÉÍíÈÐáÇ ÁÇÏ
|
ÑÈÊÚí
|
ÍÈÐáÇ ÏäÚ
|
. ãæí
|
/ Û 22.72
ØÓæÊãáÇ
|
í Éíãæí
|
|
.
íáÓÇäÊáÇ
|
ÇåÆÇÏ
|
í ãßÍÊÊ
íÊáÇ
|
áãÇæÚáÇ
|
Óä
|
áÈÞ äã
|

ÉÈÇíäáÇ
ÛÇæÈáÇ
( íáÍãáÇ
ÈäÑáÇ
íáÍãáÇ
ÈäÑáÇ äÇß.)%83.33
Ñã æå
ÇÚæÖæã
í ÇÏÌ ÚÆÇÔ
Çåäßáæ
áãÍáÇ ÁÇäË
ÑÕáÇ
æ
ÊÑåÙæ
ÇíÏíÓßæßáÇ
ÉÖÎäã
ÉÓíßÊãáÇ
ÒÇÑÅ
ÇÏÌ ÉÚÊÑã
ÊáÇÏÚã
( ÇäÌÇã
.) %43
ÉíÑãíáÇ
ÉÈáÛ
ÏÆÇÚáÇ
,æãä ,
,
íáÍãáÇ
ÈäÑáÇ :
ÑíÛÊáÇ
áãÇæÚ
4..%L:o1I c :+Lo151I
ÈÇÌäÇ
áÏÚã
ÉÈæÕÎáÇ
|
äã
|
ÊÇÓÇÑÏáÇ
|
äã
|
ÏíÏÚáá
|
|
Úã
|
ÉÚÇÖÑáÇ
|
ÉÑÊ
|
ááÇÎ
|
|
.ÖÇÑãÇ ,
|
ÉÍíÈÐ
|
,
ÉíÎ ìæÏÚ
.íáÇæÊáÇ
ìáÚ %25 æ %36.11 æå
ÉíËØãáÇ
ãæíÏíÑÊÓæáßæ
ÉíäæáæÞáÇ
ÉÈÇÕáÅÇ
áÏÚã äÅ
ÇÑíÎæ
The objective of our work is to take stock of the work done on
growth, reproduction and the health of the local Algerian population of rabbits
(Oryctolagus cuniculus).
Local rabbit is characterized by an average fertility rate of
87%, a receptivity of 89%, a prolificacy of 7.2 and a stillbirth with 18.9%,
however, great variability. Animal performance related to reproduction of the
local rabbit are influenced by factors including the animal's age,
physiological condition and genotype type thereof and the factors related to
the environment to the example of power, environmental temperature, light and
season. Finally, factors related to the conduct of breeding including the
method and rate of reproduction may also influence performance. Local rabbit is
also characterized by an average weight of 49,4g at birth, weaning 463,7g, 621G
at the age of 6 weeks and 1433g at the age of the 11th week. Throughout the
fattening period, the average consumption index is 3.00 with an average daily
gain of 22.72 g / J. At slaughter, the performance of the hot carcass is
considered satisfactory (69.23%). The growth of local rabbit is influenced by
the same factors that control their reproductive performance.
Finally, the rate of infection by Escherichia coli and
Clostridium perfringens is 36.11% and 25% respectively. Infection with
Cryptosporidium spp is quite common in the local rabbit (83.33%). The local
rabbit was the subject of several studies of coccidiosis and showed oocyst
excretion rates low or zero during pregnancy but very important during
lactation with a predominance of Eimeria magna (43%).
Keywords: Local Rabbit, fertility,
prolificacy, growth, change factors, performance, carcass, pathologies.
01
03
03
03
04
06
08
08
09
09
10
11 11 13 13
13
13
14
14
15
16
16
18
19 19 21 21
TABLE DE MATIERES
PARTIE BIBLIOGRAPHIQUE
INTRODUCTION GENERALE
CHAPITRE I : PERFORMANCES DE REPRODUCTION
I.1. La réceptivité
I.2. La fertilité
I.3. La prolificité
I.4. La mortinatalité
I.5. Les facteurs de variation des performances de
reproduction
I.5.1. Facteurs liés aux femelles
L'effet de l'âge
L'effet de l'état physiologique de la femelle
L'effet génétique
I.5.2. Facteurs lies aux mâles
I.5.3.Facteur lié à l'alimentation
I.5.4. Facteurs liés à l'environnement
L'effet de la température
L'effet de la lumière
L'effet de la saison
I.5.5. Autres Facteurs
Facteurs liés à la conduite d'élevage
Facteurs liés aux maladies et aux troubles de
reproduction
CHAPITRE II : LES PERFORMANCES DE CROISSANCE
II.1. La croissance de la naissance au sevrage
II.1.1. Evolution de la quantité du lait
consommé par lapereau en fonction de l'âge
II.1.2. L'influence de l'effectif de la portée sur la
quantité de lait consommée par
lapereau
II.2. La croissance entre le sevrage et l'âge adulte
II.3. La vitesse de croissance
II.3.1. Gain moyen quotidien
22 22 22 22 24 24
24
25
26
26
27
27
27
28
28
28
29
29
30
32
32
32
33
33
33
34 36 36 38 40
II.3.2. L'indice de consommation
II.3.3. La croissance relative et coefficient
d'allométrie
II.3. Facteurs de variation des performances de croissance
II 3.1. L'influence du facteur génétique
II 3.2. L'influence de de l'exercice physique
II 3.3. L'influence du numéro de portée
II.3.4.Influence de l'alimentation
L'effet de l'apport des protéines
L'effet du rapport protéines / énergie
L'effet de l'apport de lest
II 3.5. Influence de l'environnement
L'effet de la température ambiante
L'effet de la saison
L'effet de l'hygrométrie
L'effet de la densité
L'effet du mode de logement
II.4. Le rendement de la carcasse
II.4.1. Définition de la carcasse
II.4.2. Définition du rendement
CHAPITRE III : ETAT SANITAIRE
III.1. Pathologies bactériennes
III.1.1. La colibacillose
III.1.2. L'entérotoxémie
III.2. Pathologies parasitaires
III.2.1. La cryptosporidiose
III.2.2. Les coccidioses
III.3. Pathologies virales
III.3.1 VHD ou Maladie hémorragique virale
CONCULSION
REFERENCES BIBLIOGRAPHIQUES
Liste des tableaux
Titre du tableau Pages
Tableau 1 Taille de la portée en fonction
de l'origine de l'animal 06
Tableau 2 Variation de la mortinatalité
en fonction de l'origine de l'animal 08
Tableau 3 Effet de l'âge à la
première saillie sur le taux de fertilité 09
|
Tableau 4 Résultats de performances de
reproduction obtenus à la station de
Baba Ali sur 3 différents génotypes
|
11
|
Tableau 5 Effet du niveau alimentaire sur le
taux de gestation 12
Tableau 6 Le poids moyen des lapereaux à
la naissance (portée et individuel) 17
Tableau 7 Poids moyen des lapereaux au sevrage
18
20
23
26
Tableau 8 Poids moyen de lapereau pendant la
période de l'engraissement, le
gain moyen quotidien et la
consommation moyenne quotidienne
Tableau 9 Variation de poids adulte et de gain
moyen quotidien selon
différente phénotype, un essai à
Toulouse
Tableau 10 L'effet du niveau protéique et
de la concentration en énergie digestible de l'aliment sur les
performances de l'abattage du lapin âgé de 90 jours
Tableau 11 Effet des basses et hautes
températures sur la croissance 27
29
Tableau 12 Effet de saison sur les
caractères de croissance 28
Tableau 13 L'incidence du mode de logement sur
les performances zootechniques du lapin (souche Hyplus)
Tableau 14 le rendement à l'abattage du
lapin local 31
|
Tableau 15
|
Pouvoir pathogène comparé des différentes
coccidies intestinales du lapin
|
35
|
Tableau 16 Oocystes sporulés d'Eimeria
isolés 36
Liste des figures
Titre des figures Pages
Figure 1 Position de lordose chez la lapine
réceptive 03
|
Figure 2 Evolution de la consommation
alimentaire de la femelle au cours de
cycle reproductif
|
12
|
Figure 3 Variation de poids vif en fonction de
l'âge de lapereau 16
Figure 4 Paramètres de la croissance
pondérale globale 20
Figure 5 Evolution des gains moyens quotidiens
en fonction de l'âge 21
23
Figure 6 Evolution moyenne du poids vif entre 4
et 12 semaines des lapins de
population locale algérienne
élevés à Tizi Ouzou et Baba Ali
Figure 7 Rendement en viande d'un lapin de
format moyen de 2,3kg 30
Liste des abréviations
°C : Degré celsius.
CC : Carcasse chaude.
CF : Carcasse froide.
CMQ : Consommation moyenne quotidienne.
EPEC : Entéropathogenic Escherichia
coli (colibacilles entéropathogènes).
g/J : Grammes/jour.
GMQ : Gain moyen quotidien.
GPR : Gras péri rénal.
h/J : Heurs/jour.
INRA: Institut National de la Recherche
Agronomique.
Itavi : Institut Technique de l'Aviculture.
Itelv : Institut technique des
élevages.
J : Jour.
kcal : Kilocalories.
n : Nombre.
P : Poids.
P/E : Protéine/énergie.
PL : Population locale.
PM : Poids de manchons.
PP : Poids de la peau.
Pva : Poids vif à l'abattage.
REPEC : Rabbit entéropathogenic
Escherichia coli (Escherichia coli entéropathogènes
du
lapin).
RHDV : Rabbit hemorragic disesese virus (Virus
de la maladie hémorragique du lapin).
S : Semaines.
t: Temps.
TDP : Poids du tube digestif plein.
Ufc : Unité formant colonie.
Ummto : Université de Mouloud Mammeri
Tizi-ouzou.
vs : Versus.
VHD : Viral hemorraic disesese (Maladie
hémorragique virale).
Partie
bibliographique
|
|
Introduction
|
1
Introduction
|
|
|
L
|
a cuniculture peut représenter pour l'Algérie
une source de protéines non négligeable compte tenu de
l'important déficit en ce nutriment. Le recours au
|
lapin est justifié par ses nombreux atouts, entre
autres, son cycle biologique court, sa forte prolificité (ce qui
représente une importante quantité de viande), sa capacité
à valoriser plusieurs ressources végétales et
sous-produits des IAA même riche en fibres et enfin, sa viande de bonne
qualité organoleptique.
Comme le faisait remarquer JAFFE, le manque des données
fondamentales sur le lapin était très apparent dans le monde, il
y a plus d'une cinquantaine d'années, mais dès le début
des années soixante (20ème siècle), cette
branche de zootechnie a enregistré une croissance considérable
tant du point de vue qualitatif que quantitatif, et surtout par rapport
à certaines autres espèces. Les fédérations de
cuniculture, la presse, et les centres d'études ont exercé une
propagande massive en faveur de cette espèce, l'opinion publique et
privée commencent à connaître et à
réévaluer le lapin qui est de" moins en moins"
considéré comme un animal de basse-cour.
La situation n'est pas la même en Algérie
où l'élevage du lapin reste une activité millénaire
des ruraux, cette espèce est intégrée aussi bien dans le
système "autarcique" des basses-cours que les systèmes
d'élevage ouverts au marché et les pratiques
cynégétiques, elle n'apparaît pas dans les statistiques et
ne fait pas l'objet d'un enseignement similaire aux autres zootechnies
spéciales (bovine, ovine, aviaire ...).
En plus, l'examen de l'élevage du lapin en
Algérie a révélé que ce dernier ayant reposé
essentiellement sur les souches hybrides (Hyplus), importées de France,
résultat d'une politique d'élevage "productiviste" visant
à assurer un approvisionnement régulier des marchés
urbains en protéines animales de moindre coût mais qui, dans les
faits, a eu pour conséquence la marginalisation de la population locale
tant du point de vue de sa connaissance que de son intégration dans les
systèmes d'élevage. Mais cette opération a rapidement
échouée (moins de deux années) en raison d'une
alimentation de mauvaise qualité qui a provoqué une importante
mortalité (Berchiche et Lebas, 1990). Une décennie plus tard, la
cuniculture rationnelle est de nouveau relancée mais avec une
stratégie
2
Introduction
favorisant la valorisation des reproducteurs de population
locale (Berchiche et al, 2000 b). Cette opération disposera donc d'un
aliment industriel de qualité mais en s'orientant sur le marché
de matières premières importées.
Cette situation renvoie à l'absence d'un capital de
connaissance suffisant susceptible de servir de base à un
développement, ainsi qu'une ignorance massive des qualités
précieuses de cette population, du point de vue de son adaptation aux
conditions alimentaires et climatiques (résistance avérée
à la chaleur et adaptation à des conditions rigoureuses et
à une alimentation de qualité médiocre), autrement dit,
toutes les caractéristiques souhaitables pour une agriculture durable
à faibles besoins d'intrants contribuant à la
sécurité alimentaire.
Cependant, dans tous projets de développement d'une
production cunicole utilisant le lapin local, une logique d'ensemble doit
être suivie comprenant, en premier lieu, l'identification de la
population locale existante, la connaissance de ses aptitudes biologiques et
zootechniques et son adaptabilité, ainsi que ses performances de
croissance et de reproduction avant de désigner les programmes de
sélection ou les systèmes de production convenables. C'est ainsi
que depuis 1990 l'ITELV de Baba Ali et certaines universités notamment
l'université de Mouloud Mammeri de Tizi-Ouzou ont mis en place des
programmes de caractérisation de ces populations et de contrôle de
leurs performances.
Cette synthèse bibliographique fait le point sur les
travaux réalisés sur le lapin de population local
algérienne. Nous aborderons dans le premier chapitre, les performances
de reproduction et leurs facteurs de variation. Dans le 2ème
chapitre, nous décrirons la croissance et ses principaux facteurs de
variation chez le lapin local en comparaison avec d'autres races et souches
étrangères. Enfin, nous terminerons avec une description des
principales pathologies rencontrées dans les élevages cunicoles
et leurs situations en Algérie.

Chapitre I
Les performances de
reproduction
1
3
Chapitre I Partie bibliographique
Chapitre I : Les performances de reproduction.
I.1. La réceptivité :
Une lapine dite réceptive, lorsqu'en présence du
male adopte la position de lordose et accepte l'accouplement (Fortun et
al., 1995). Chez la lapine, la réceptivité est
très élevée pendant les heures qui suivent la mise bas
(environ 100%), elle décroit ensuite 4 à 6 jours après,
pour atteindre 40 à 60% puis augmente 10 à 14 jours après
la mise bas et atteint son maximum initiale après le sevrage.

Figure 1 : Position de lordose chez la
lapine réceptive (Anonyme, 2015)
Chez les lapins de population locale, les travaux
effectués par Moulla et Yakhlef (2005) montrent que les
taux moyens de réceptivité s'établissent à 89 %.
Ces valeurs sont supérieures à celles enregistrées par
Zerrouki et al. (2001) chez des lapines issues de la même
population, avec des taux de réceptivité de 80 %. La bonne
réceptivité de la lapine locale s'expliquerait par le mode de
reproduction pratiqué (saillie naturelle) et le rythme de reproduction
adopté (La remise à la reproduction s'est faite 10 à 12
jours après la mise bas) (Moulla et Yakhlef, 2005).
Les travaux effectués par Gacem et al. (2009)
montrent que les taux de réceptivité chez les lapins de
population locale, de population synthétique et chez la souche blanche
sont respectivement : 64, 64,5 et 69,2% donc les lapines de la souche
synthétique sont significativement moins réceptives que celles de
la population blanche, mais du même niveau que celles de la population
locale. Ces résultats sont inférieurs à ceux
enregistrées
4
Chapitre I Partie bibliographique
par Zerrouki et al. (2005) sur les lapines de la
population locale (77%) et par Zerrouki et al. (2009) chez les lapines
de la souche blanche (92%).
I.2. La fertilité :
La fertilité est la capacité d'un individu
à se reproduire. Elle est définie par le nombre de femelles
palpées positives rapporté au nombre de femelles saillies
(Blocher et Francher, 1990). C'est également le nombre des femelles
mettant bas rapporté au nombre de femelles mises à la
reproduction (Chmitelin et al., 1990).
Chez les lapines de population locale, les travaux
effectués par Moulla et Yakhlef (2005) montrent que les
taux moyens de fertilité s'établissent à 87 %. Et tout
comme la réceptivité, ces valeurs sont supérieures
à celles enregistrées par Zerrouki et al. (2001) chez
des lapines issues de la même population locale soit 80%. La
différence entre le taux de fertilité observée serait
liée à la perte de portées entières, aux
mortalités des lapines avant la mise bas, à l'élimination
des femelles pour des raisons de santé mais aussi aux quelques cas
d'avortements (Moulla et Yakhlef, 2007).
L'analyse des performances, par types
génétiques, étudiées par Abida Ouyed (2006) montre
que dans le cas des femelles de races pures, ce sont la race
Néo-Zélandaise blanche et la race Californienne qui
présentent des taux de fertilité les plus élevés
(92,1 et 93,6 % respectivement), suivis de la race de Chinchilla et de
Géant des Flandres avec une fertilité de 63% et 50%
respectivement. Concernant les femelles de lignées hybrides, ce sont les
Néo-Zélande blanc x Chinchilla et Néo-Zélande blanc
x Géant des Flandres qui arrivent au premier rang pour la
fertilité avec 88,6 et 88,9 % respectivement, suivis des femelles
Néo-Zélande blanc x Californienne avec un taux de
fertilité de 81,8 %. (Ouyed, 2006)
I.3. La prolificité :
La prolificité est le nombre de lapereaux nés
par mise bas (Armero et al., 1995). Elle résulte d'une
série d'événements, qui vont de la maturation des
gamètes jusqu'à la
5
Chapitre I Partie bibliographique
naissance : ovulation, fertilisation, développement
embryonnaire et foetal (Bidanel, 1998 ; Mattaraia et
al., 2005).
La population locale Algérienne de lapin se
caractérise par une prolificité relativement moyenne à la
naissance. D'après les travaux réalisés par Remas (2001),
Saidj (2006) et Moulla et Yakhlef (2007), le nombre total de lapereaux
nés par portée chez la population Kabyle est en moyenne 7,2. Ces
résultats sont nettement supérieurs à ceux obtenus sur des
femelles de même origine mais exploitées à un niveau
fermier avec une moyenne de 5 (Berchiche, 1998 cité par Berchiche et
Zerrouki (2000) ; Djellal et al., 2006). Ces faibles performances dans
les élevages fermiers pourraient être attribuées au rythme
de reproduction adopté.
Les lapines des populations locales Marocaine et Egyptienne se
caractérisent par une prolificité plus modeste, qui est en
moyenne de 6,4 (Bouzekraoui, 2002 ; Barkok et Jaouzi; 2002 ; Khalil, 2002a et
2002b ; Afifi, 2002).
Toutefois, la prolificité de la population locale
Algérienne est inférieure à celle des races
Européennes notamment le Fauve de Bourgogne (Bolet, 2002a), le
Géant d'Espagne (Lopez et Sierra, 2002) et le Gris de Carmagnola
(Lazzaroni, 2002), estimée en moyenne à 8,8. Par ailleurs, les
souches sélectionnées à l'exemple d'INRA 2066 (Bolet,
2002b) et Hyplus (Verdelhan et al. 2005) se caractérisent par
des prolificités encore supérieures avoisinant en moyenne 10,3
lapereaux (Tableau 1).
|
Chapitre I Partie bibliographique
|
|
Tableau 1: Taille de la
portée en fonction de l'origine de l'animal
(Synthèse des
références bibliographiques).
Auteurs Origine de l'animal Taille de la
portée
Remas (2001) Population locale 7,4
Saidj (2006) Population locale 7,2
Moulla et Yakhlef (2007) Population locale
7,1
6,2
6,7
Bouzekraoui (2002) Population locale
(Tadla, Maroc)
Barkok et Jaouzi (2002) Population locale
(Zemmouri, Maroc)
Khalil (2002b) Giza White (Égypte) 6,7
Khalil (2002a) Baladi White (Égypte)
5,3
Afifi (2002) Gabali (Égypte) 6,3
6
Bolet (2002a) Fauve de Bourgogne 9
Lopez et Sierra (2002) Géant d'Espagne
8,8
Lazzaroni (2002) Gris de Carmagnola (Italie)
8,5
Bolet (2002b) INRA2066 (France) 10
Verdelhan et al. (2005) Hyplus 10,6
I.4. La mortinatalité :
Dans les élevages, la mortinatalité des
lapereaux dépend de la qualité maternelle des lapines, de la
taille de portée et du poids des lapereaux à la naissance
(Rashwan et Maria, 2000).
Chez la population locale, les travaux effectués par
Berchiche et Zerrouki (2000) ; Berchiche et Kadi (2002) ; Remas (2001) ;
Zerrouki et al. (2005a) et Moulla et Yakhlef (2007), montrent que la
mortinatalité est en moyenne de 18,9% avec cependant une grande
variabilité (Tableau 2). Cette situation pourrait
être liée à la perte de portées entières
à la naissance chez certaines femelles avec un comportement maternel
déficient (mises bas sur
7
Chapitre I Partie bibliographique
le grillage, abandon des portées, cannibalisme) et aux
mauvaises conditions d'ambiance (température élevée durant
l'été et basse en hiver).
En Egypte, la mortinatalité chez les lapines de
population locale (Baladi White et Giza White) est plus faible, que celle
observée sur les lapines de population locale au Maroc (5,4 vs
11,8) (Khalil, 2002a ; 2002b ; Barkok et Jaouzi, 2002 ; Bouzekraoui,
2002). Cette mortinatalité est nettement supérieure à
celle des races Européennes notamment le Fauve de Bourgogne et le
Chinchilla (France) (Bolet et al., 2004) et aux souches (Caldes et
Prat) et aux lignées (lignée A et lignée V) d'origine
Espagnole (Gomez et al., 2002a ; 2002b ; Baselga, 2002a ; 2002b).

8
Chapitre I Partie bibliographique
Tableau 2 : Variation de la
mortinatalité en fonction de l'origine de l'animal
|
(Synthèse des résultats bibliographiques).
|
|
|
Origine de l'animal
|
Mortinatalité (%)
|
|
Population locale (Algérie)
|
13,6
|
|
Population locale (Algérie)
|
12,8
|
|
Population locale (Algérie)
|
16,4
|
|
Population locale (Algérie)
|
21
|
|
Baladi (Egypte)
|
5,6
|
|
Giza White (Egypte)
|
5,2
|
|
Zemmouri (Maroc)
|
14,3
|
|
Tadla (Maroc)
|
9,2
|
|
Fauve de Bourgogne
(France)
|
1,3
|
|
Chinchilla (France)
|
0,8
|
|
Souche Caldes (Espagne)
|
7,6
|
|
Souche Prat (Espagne)
|
4,3
|
|
Lignée A (Espagne)
|
6,9
|
|
Lignée V (Espagne)
|
5,8
|
I.5. Les facteurs de variation des performances de
reproduction :
I.5.1. Facteurs liés aux femelles :
Auteur
Remas (2001)
Berchiche et Zerrouki ; (2000) ; Berchiche et Kadi
(2002)
Zerrouki et al. (2005a)
Moulla et Yakhlef (2007)
Khalil (2002a)
Khalil (2002b)
Barkok et Jaouzi (2002)
Bouzekraoui (2002)
Bolet et al. (2004)
Bolet et al. (2004)
Gomez et al. (2002a)
Gomez et al. (2002b)
Baselga (2002a)
Baselga (2002b)
9
Chapitre I Partie bibliographique
? L'effet de l'âge :
Chez la lapine, la fertilité dépend de
l'âge de celle-ci. En effet, la mise à la reproduction très
tôt a une influence négative sur les performances de la
carrière de la lapine (Rommers et al, 2001). Ceci est vrai pour
les femelles présentées trop tardivement (Lebas et al.,
1996). De même, les travaux de Quastel (1984) cités par
Bousit (1989) ont révélé une baisse de fertilité
chez les lapines saillies après l'âge de 5 mois (Tableau
4).
Tableau 3 : Effet de l'âge a la
première saillie sur le taux de fertilité
(Questel, 1984)
|
Age de la lapine à la première
saillie
|
Effectif
|
Taux de fertilité (%)
|
|
-140 J
|
162
|
85
|
|
140-149 J
|
524
|
86
|
|
150-159 J
|
318
|
72
|
|
160-169 J
|
218
|
78
|
|
170-179 J
|
96
|
80
|
|
+180 J
|
195
|
79
|
Lebas (1974) montre que chez les femelles saillies pour la
1ère fois 116 J, 137 J et 158 J, le lot sailli à 137
jours présente de meilleurs performances, meilleure
longévité et une bonne productivité par portée.
Selon Lebas (1994) l'âge à la première saillie
dépend de la race et du développement corporel, soit à 80%
de leurs poids adulte.
? L'effet de l'état physiologique de la femelle
:
? L'effet de la parité :
L'ensemble des auteurs confirment que le taux de
fertilité le plus élevé est observé chez les
nullipares, les multipares sont les plus fertiles que les primipares, avec des
meilleurs
10
Chapitre I Partie bibliographique
résultats de prolificité et de viabilité
à la troisième parité Tandis que les primipares en les
moins bons résultats (Hulot et Mattheron, 1981).
Lazzaroni et al (1999) ont confirmé l'effet de
la parité sur les performances de reproduction du lapin Gris de
Carmagnola. Les primipares ont montré une faible prolificité avec
la plus basse valeur de nées vivants 6,63 et une mortalité
à la naissance très élevée 10,6%. Alors que les
femelles à la sixième parité enregistrent une meilleure
prolificité et une faible mortalité à la naissance
7,2%.
? L'effet de lactation :
Selon Fortun-Lamothe et Bolet (1995) ; Bolet (1998), la
fertilité et la prolificité des lapines allaitantes sont
globalement plus faibles que celles des lapines non allaitantes. Garcia et
Parez (1989) et Theau-Clement (2003) soulignent qu'en saille naturelle,
l'allaitement présente un effet négatif sur le taux de
l'acceptation de l'accouplement. De même, Theau-Clement et al
(1990) montrent que les lapines allaitantes en 3ème et
5ème jour de lactation sont significativement moins
réceptives que les non allaitantes (73% vs 32%).
? L'effet de la réceptivité :
Les lapines réceptives produisent 3 fois plus
d'embryons que les non-réceptives (6,2 contre 2,5 embryons) (Poujardieu
et Theau-Clement, 1994). Theau-Clement et Roustan, (1992) ont montré que
par rapport aux lapines réceptives, les conséquences de la mise
à la reproduction des lapines non réceptives sont : la diminution
de la fréquence d'ovulation, diminution de la survie embryonnaire et
donc diminution de la fertilité et la taille de la portée
à la naissance.
? L'effet génétique :
Beaucoup de chercheurs mettent en évidence l'effet de
la race de sur les performances de reproduction. Le tableau ci-dessous montre
l'effet race.
11
Chapitre I Partie bibliographique
Tableau 4 : Résultats de
performances de reproduction obtenus à la station de Baba Ali sur
3
différents génotypes (Synthèse de Lebas et Zerrouki,
2010).
|
Génotypes
|
Souche synthétique
|
Population blanche
|
Population locale
|
|
Poids des lapines, g
|
3633a
|
3434b
|
3278c
|
|
Réceptivité, %
|
64,5b
|
69,2a
|
64,0b
|
|
Fertilité, %
|
51,0
|
52,0
|
51,0
|
|
Nés totaux/Mise Bas
|
9,50a
|
7,42b
|
6,75c
|
|
Nés vivants/Mise Bas
|
8,74a
|
6,84b
|
6,23c
|
|
Sevrés / sevrage
|
7,08a
|
6,09b
|
5,45c
|
I.5.2. Facteurs lies aux mâles :
Les performances de reproduction de la femelle sont
également influencées par la qualité de la semence (Garcia
et al., 2000). La motilité massale ainsi que le nombre des
spermatozoïdes motiles par éjaculation sont les plus importants et
varient en fonction des mâles (Brun et al., 2002).
Selon une expérience de Thau-Clémet et al.
(2003), l'insémination avec seulement 6 millions de
spermatozoïdes diminue significativement la fertilité des lapines
(- 2.4%) par rapport à l'insémination avec 12 à 24
millions de spermatozoïdes totaux par dose mais sans aucune alternation de
la prolificité.
I.5.3.Facteur lié à l'alimentation
L'alimentation a un effet direct et primordial sur le niveau
de production et sur l'état de santé des animaux (mâles et
femelles) (Lebas et al., 1996). Les besoins de lapines augmentent
pendant la gestation, et la femelle simultanément allaitante et gestante
à des besoins en double que celle qui est gestante (Martinez-Gomez et
al., 2004).
12
Chapitre I Partie bibliographique
Figure 2 : Evolution de la consommation
alimentaire de la femelle au cours de cycle
reproductif (Gidenne et Lebas
2005)
L'engraissement de la future reproductrice a un effet
négatif sur les caractères de reproduction (augmentation de la
mortinatalité). Par contre une restriction énergétique
anté-partum a un effet négatif sur la réceptivité
des femelles (Quinton et Egron, 2001).
Hafez et al. (1967) (Tableau 5) ont
mis en évidence l'effet significatif du niveau alimentaire sur les
performances des femelles. En effet, en passant d'une ration alimentaire de
60g/J à 280g/J, le taux de gestation augmente de 39%. Aussi, de nombreux
auteurs (Lamming et al., 1954) ont montré l'influence
d'oligo-éléments tels que le zinc ou la vitamine A sur la
fertilité.
Tableau 5 : Effet du niveau alimentaire
sur le taux de gestation (Hafez et al., 1967)
|
Niveau alimentaire (g/J)
|
Taux de gestation (%)
|
|
280
|
74
|
|
140
|
67
|
|
60
|
45
|
13
Chapitre I Partie bibliographique
I.5.4. Facteurs liés à l'environnement :
? L'effet de la température :
Selon Arveux (1988), les fortes températures
réduisent les performances de reproduction, la réceptivité
des femelles et diminue la mortalité embryonnaire. Les
températures basses ou très élevées peuvent
occasionner des périodes de stérilité (Casting, 1979). La
chaleur affecte de façon négative la spermatogenèse,
l'hormogènes et les caractéristiques du sperme. Par ailleurs, une
dégénérescence séminale est observée chez le
lapin suite à une exposition courte à une température
élevé (Kasa et Twaites, 2001). D'après Fromont (2001), un
manque d'ardeur sexuelle des mâles lié à une chaleur
excessive provoque un refus d'accouplement.
? L'effet de la lumière :
Theau-Clement et al. (2004) montrent qu'un passage du
programme lumineux de 8 à 16 h/J n'a pas d'influence sur les
performances des reproductrices. Cependant, une durée
d'éclairement réduite par rapport à cette dernière
amène à une diminution de la réussite de la saillie.
Un programme lumineux de 16 h/J entraine une meilleure
prolificité et un poids de portée plus élevé mais
un poids moyen en sevrage plus faible par rapport à un programme
lumineux de 8h/J (Theau-Clement et Mercier, 2004).
? L'effet de la saison :
Selon Yamani et al. (1991), le taux de
réceptivité est plus élevé au printemps et en
hiver. La saison d'accouplement influence sur le nombre moyen d'ovules pondus
(1,7 corps jaunes d'écart entre l'hiver et le printemps).
14
Chapitre I Partie bibliographique
De même, la prolificité est maximale en
début d'été et son minimum est observé en
début d'automne, la différence entre le maximum et le minimum est
de 1,2 lapereaux pour les nés totaux et 0,7 lapereaux pour les
nés sevrés.
L'effet de la saison sur la prolificité est
significatif. Arveux (1988) rapporte que les meilleures tailles des
portées sont observées en hiver et en automne. En
été, la prolificité diminue et les faibles
températures d'automne et d'hiver traduisent un accroissement de la
prolificité.
I.5.5. Autres Facteurs :
? Facteurs liés à la conduite
d'élevage :
? Effet de mode de reproduction :
Les deux modes existants sont la saillie naturelle et
l'insémination artificielle. Theau-Clement et Poujardieu (1994) montrent
que les lapins inséminés ovulent plus que les lapins saillies.
Les lapines réceptives ovulent plus que les non réceptives. Ces
auteurs confirment qu'il existe une interaction entre le mode de reproduction
et la réceptivité. La réceptivité n'a pas
d'influence sur la fréquence d'ovulation en insémination
artificielle. Par contre, en saillie naturelle, l'ovulation semble être
conditionnée par la réceptivité sexuelle des lapines en
moment de l'accouplement.
? Effet de rythme de reproduction :
La viabilité foetale augmente lorsque les femelles sont
inséminées plus tardivement après la parturition. En
effet, chez les femelles primipares allaitantes, une saillie à 25 jours
post partum réduit la mortalité foetale de 12,5 à
8,7 % par rapport à une saillie à J11 post partum
(Feugier et al., 2005 ; 2006).
15
Chapitre I Partie bibliographique
? Facteurs liés aux maladies et aux troubles de
reproduction
Selon Fromont (2001), les reproducteurs qui présentent
des irritations de l'appareil génital, atteintes de mammites et des
maladies bactériennes (pasteurellose, staphylocoques ou Chlamydiose)
peuvent refuser l'accouplement.

Chapitre II
Les performances de
croissance
2
16
Chapitre II Partie bibliographique
Chapitre II : Les performances de croissance
La croissance est l'ensemble de modification de poids, de la
forme et de composition anatomique et biochimique depuis la conception
jusqu'à l'âge adulte (Prud'hon, 1976). Elle est
conditionnée par des phénomènes de multiplications, de
développement et de différenciation cellulaire, tissulaire et
organique (Prud'hon, 1970).
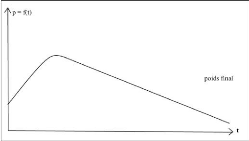
La croissance pondérale entre la naissance et
l'âge adulte correspond à l'évolution du poids de
l'organisme en fonction du temps (t) : P = f(t) (Figure 1).
Cette évolution est continue. La courbe de croissance est
généralement sigmoïde, avec un point d'inflexion, elle tend
asymptotiquement vers une valeur finale qui est assimilée au poids
adulte (Ouhayoun, 1983)
Figure 3 : Variation de poids vif en
fonction de l'âge de lapereau (Gidenne, 2015).
II.1. La croissance de la naissance au sevrage :
La croissance des lapereaux avant le sevrage est
conditionné par la production laitière de la lapine. Le lait de
lapine est très riche en éléments minéraux totaux.
Parmi ceux-ci, le calcium a la plus grande importance pondérale avec 3
à 7 mg/g de lait devant le phosphore (2 à 4 mg/g de lait) et le
potassium (2 mg/g de lait). Au cours de la lactation, le lait s'enrichit en
calcium, en phosphore et en magnésium, tandis que les teneurs en sodium
et en phosphore sont relativement stables, et que celle en zinc tend à
diminuer. Par rapport à la vache ou à la truie ; la lapine
exporte de grande quantités de minéraux qui peuvent
17
Chapitre II Partie bibliographique
atteindre la moitié des réserves totales de
l'organisme en 2 a 5 jours, tant pour le calcium que pour le phosphore
(Besançon et al., 1971).
A la naissance, avant d'ingérer le lait de sa
mère dans les minutes suivant sa « sortie » de l'utérus
maternel, un lapereau pèse de 50 à 55g environ avec de
fluctuations assez fortes en fonction de la taille de la portée. Le
poids moyen des lapereaux à la naissance varie en fonction de la race de
sa mère (Tableau 6).
Tableau 6 : Le poids moyen des
lapereaux à la naissance (portée et
individuel)
(Synthèse de Moulla, 2006).
|
Auteurs
|
Race/souche/ population
|
Poids à la naissance (g)
Portée Individuel
|
|
Gallal et al. (1994)
|
Californienne
Néo-zélandaise
|
520
490
|
-
-
|
|
Alliance et al. (2002)
|
Lapin local algérien
|
269
|
49,4
|
Sa croissance est ensuite linéaire pendant 3 semaines
(11-13 g par jour au sein d'une portée de 10), puis elle
s'accélère, pour atteindre 35-38 g par jour à partir de 25
jours quand la part de l'alimentation solide devient conséquente (Lebas,
2002).
La seconde semaine, les lapereaux ont un joli duvet, on peut
reconnaitre la couleur et le marquage de leur fourrure définitive. Ils
commencent petit à petit à ouvrir leurs yeux. Certain sont plus
précoces que d'autres mais cela se produit généralement
entre le 8ème et le 11ème jour.
A partir de la troisième semaine, les lapereaux
commencent à être assez autonomes. Leur alimentation se
diversifie. Ils consomment la nourriture de leur maman. Ils grignotent tout
d'abord du foin et des granulés mais continuent à téter
leur mère. Jusqu'à 8 semaines, ils vont continuer à
grandir et à devenir de plus en plus agiles. C'est la période
pendant
18
Chapitre II Partie bibliographique
laquelle la socialisation est importante. Si les lapereaux ne
sont pas habitués aux humains ils risquent d'être assez craintifs
plus tard. Il ne faut pas hésiter à les manipuler.
Au cours d'un allaitement de 6 semaines un lapereau multiplie
son poids de naissance par 20, le poids des lapereaux au sevrage varie en
fonction des races (Tableau 7).
Tableau 7 : Poids moyen des lapereaux
au sevrage
(Synthèse de Moulla, 2006)
|
Auteurs
|
Races/souches/ population
|
Poids au sevrage (g)
Portée Individuel
|
|
Poujardieu et al.
|
Néo-zélandaise
|
-
|
840*
|
|
(1984)
|
|
|
|
|
Gallal et al. (1994)
|
Californienne
|
3390
|
520*
|
|
Néo-zélandaise
|
2810
|
490*
|
|
Khalil (1998)
|
Baladi rouge
|
1550
|
322
|
|
Baladi blanc
|
1145
|
320
|
|
Baladi noire
|
1320
|
258
|
|
Giza blanc
|
1700
|
330
|
|
Alliance et al. (2002)
|
Lapin local algérien
|
2130
|
463,7*
|
*l'âge au sevrage : 28 jours.
II.1.1. Evolution de la quantité du lait
consommé par lapereau en fonction de l'âge :
La quantité du lait consommée par lapereau
augmente jusqu'à la 3ème semaine. Elle atteint alors
un maximum représentant 2 fois celle de la 1ère
semaine et diminue ensuite (Lebas, 1969). Elle est plus faible au sein d'une
portée nombreuse que dans une portée à effectif
réduit. Ainsi, pour l'ensemble de la période, un lapereau
appartenant à une portée de 4 reçoit théoriquement
près de 1000 grammes de lait alors qu'un jeune faisant parti d'une
portée de 12 n'en reçoit que 705 grammes de lait (Lebas,
1969).
19
Chapitre II Partie bibliographique
II.1.2. L'influence de l'effectif de la portée sur
la quantité de lait consommée par lapereau :
Les coefficients de corrélation entre l'effectif de la
portée et le lait consommé par lapereau pour les périodes
0 à 3 semaines, 4 à 6 semaines et 0 à 6 semaines sont dans
l'ordre : 0,35, 0,31 et 0,38 (Lebas, 1969).
II.2. La croissance entre le sevrage et l'âge
adulte :
Le comportement alimentaire du lapin est très
particulier comparé à d'autres mammifères, avec une
spécificité qui est la pratique de la caecotrophie,
associée à une physiologie digestive « mixte »
monogastrique et herbivore. Le lapin peut consommer une grande
variété d'aliments, et peut ainsi s'adapter à des
environnements alimentaires très divers. La bonne connaissance du
comportement d'ingestion du lapin est nécessaire pour permettre au point
des aliments équilibrés et adaptés à chaque stade
physiologique (Gidenne et Lebas, 2005)
La période de l'engraissement commence à 4
semaines d'âge et prend fin entre l'âge de 10 à 11 semaines
avec poids vif de 2,3 kg. Ce qui correspond à un taux de maturité
de 55% du produit adulte d'un lapin âgé de 2 ans (4 kg) (Blasco,
1992).
Durant la période de l'engraissement, le poids des
lapereaux s'accroît et varie, ainsi que l'indice de consommation, en
fonction de la race (Tableau 8).
20
Chapitre II Partie bibliographique
Tableau 8 : Poids moyen de lapereau
pendant la période de l'engraissement, le gain moyen
quotidien et la
consommation moyenne quotidienne (synthèse des résultats).
|
1-
|
2-
|
3-
|
4-
|
5-
|
6-
|
|
PI
|
PI
|
P blanche
|
P blanche
|
P blanche
|
Souche
|
|
Ummto
|
Itelv
|
Itmas/
|
Eleveur/
|
Djebla /
|
synthétique
|
|
|
Ummto
|
Ummto
|
Ummto
|
Itelv
|
|
Poids vif 5 s (g)
|
453
|
621
|
564
|
445
|
590,86
|
-
|
|
Poids vif 11 s (g)
|
1975,83
|
1433
|
1909
|
1997,21
|
2199,57
|
1506
|
|
Poids vif 12 s (g)
|
2157,39
|
1576
|
2011
|
-
|
2506,5
|
-
|
|
GMQ 5-11(g)
|
31,11
|
25,33
|
-
|
36,8
|
38,30
|
24
|
|
CMQ 5-11(g)
|
120,07
|
69,7
|
-
|
107,2
|
121,72
|
-
|
PL : population locale ; S : semaines, 1 -
Berchiche et al 2000 ; 2 - Chaou T., 2006 in Mefti- Kortby
2010 ; 3 - Lounaouci 2008; 4 -
Lakabi 2009 ; 5 - Kadi et al., 2011
; 6 - Gacem et al.,2008 et 2009)
Durant cette période qui va du sevrage à
l'abattage, ce sont les potentialités génétiques
transmises par les parents en interaction avec le milieu qui s'expriment. Le
lapin doit toujours avoir des aliments à sa disposition. Si
l'éleveur utilise un aliment granulé complet, la consommation
journalière moyenne est 100 à 130 g pour de animaux de format
moyen. La croissance permise dans des bonnes conditions sera alors d'environ 30
à 40 g par jour, soit une consommation de 3 à 3,5 kg d'aliment
pour un gain de poids vif de 1 kg.
La courbe de croissance pondérale du lapin est une
courbe sigmoïde avec un point d'inflexion qui est situé entre la
5ème et la 7ème semaine d'âge de la
vie post-natale (Figure 4).
Figure 4 : Paramètres de la
croissance pondérale globale (Ouhayoun, 1983).
21
Chapitre II Partie bibliographique
II.3. La vitesse de croissance :
Au point d'inflexion de la courbe de croissance (5-7 semaines)
jusqu'à l'âge de 11 semaines, la vitesse de croissance est
maximale puis ralentit progressivement, notamment après 11 semaines. La
vitesse de croissance tend vers zéro à partir de 6 mois
d'âge (Baumier et Retailleau, 1986 ; Blasco, 1992).
Il est à signaler que des infléchissements de la
vitesse de croissance instantanée dus le plus souvent aux modifications
de l'alimentation et l'environnement au sevrage, se manifeste entre la
5ème et la 6ème semaine d'âge
(Ouhayoun, 1983).
II.3.1. Gain moyen quotidien :
Chez le lapin de chair de souche améliorée,
placé dans une ambiance de 18 à 22°C, le GMQ est de 35,8 g/J
avec un maximum au cours de la 8ème semaine ; soit 45,5 g/J
(Laffolay, 1985).
La moyenne des gains moyens quotidiens chez les souches
améliorées est de 23,18 g/J. Elle est proche de celle
trouvée par Lounaouci (2001) (22,72 g/J) chez le lapin local de
population, mais qui reste inférieure à celle du lapin
amélioré rapporté par Laffolay (1985).
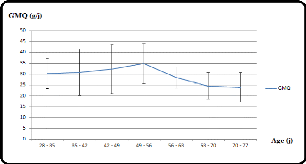
Figure 5 : Evolution des gains moyen
quotidiens en fonction de l'âge
(Lounaouci et al., 2012
modifié)
22
Chapitre II Partie bibliographique
II.3.2. L'indice de consommation :
Il correspond à la quantité totale d'aliment
dépensé dans un élevage pour obtenir un kilogramme de
lapin vivant bon à vendre (Lebas, 1991). L'indice de consommation moyen
pour toute la période d'engraissement est de 3,00 trouvé (Moulla,
2008). Ce dernier est proche de celui obtenu par Lounaouci (2001) chez le lapin
local soit, 3,12.
II.3.3. La croissance relative et coefficient
d'allométrie :
La croissance pondérale globale de l'organisme
résulte de la croissance particulière de ses différentes
composants qui ne se développe pas tous au même rythme : C'est ce
que définie l'allométrie (Ouhayoun et al., 1986).
L'existence de l'allométrie signifie que les différents organes
de l'organisme ne réalisent une même fraction de leurs poids que
successivement dans le temps, ils sont plus au moins précoces (Ouhayoun,
1983). Le classement des différents organes et tissus par ordre
croissant des coefficients de l'allométrie permit de mettre en
évidence de véritable gradient de précocité.
Chez le lapin en engraissement, la croissance est prioritaire
d'abord pour le tissu osseux, tissu musculaire et en fin le tissu adipeux
(Cantier et al., 1969). La vitesse de croissance relative du squelette
diminue quand le poids vif atteint 1 kg et celle de la musculature quand ce
poids atteint 2450 g. Entre ces deux poids, le rapport muscle/os du membre
postérieur, qui prend compte du développement relatif de la
musculature et de squelette de la carcasse dans son ensemble, croit donc
très vite (Ouhayoun, 1989).
II.3. Facteurs de variation des performances de
croissance :
II 3.1. L'influence du facteur génétique
:
La croissance du lapereau avant le sevrage dépend de
l'influence maternelle qui est la résultante du génotype de la
mère et des facteurs environnant (milieu utérin, taille de la
portée, aptitude laitière de la mère et comportement
maternel post natal de la mère). Le
|
Chapitre II Partie bibliographique
|
|
poids du lapin à 11 semaines subit encore une influence
maternelle, mais résulte de l'expression des potentialités
génétiques transmises par le mâle de divers souches ou
races (Henaff et Jouve, 1988).
La vitesse de croissance enregistrée à Baba Ali
(19 g/J) et à Tizi Ouzou (25g/J) n'est probablement que la
conséquence de l'utilisation d'un aliment mal équilibré.
Compte tenu du poids adulte (2,9 kg), la vitesse de croissance devrait se
situer aux environs de 28 à 30 g/J (Lebas, 2007).
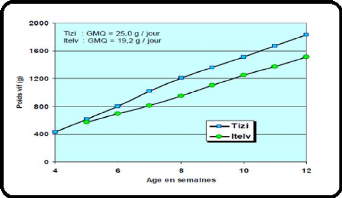
Figure 6 : Evolution moyenne du poids vif
entre 4 et 12 semaines des lapins de
population locale algérienne élevés à
Tizi Ouzou et Baba Ali (Lebas et al., 2007).
Il à signaler que la vitesse de croissance augmente
sensiblement avec le poids vif adulte (Lebas, 2007) (Tableau
9).
Tableau 9 : Variation de poids adulte
et de gain moyen quotidien selon différente
phénotype, un
essai à Toulouse (Lebas, 2007)
|
Génotype
|
A
|
B
|
C
|
|
Poids Adulte
|
3350
|
3850
|
4350
|
|
GMQ Aliment Equilibré
|
32,9
|
36,9
|
40,2 g/j
|
23
24
Chapitre II Partie bibliographique
II 3.1. L'influence de de l'exercice physique :
Combes et al. (2005), on étudie les effets de
l'exercice (sauts obligatoire entre mangeoires et abreuvoirs), effectués
durant toute la durée de l'engraissement. Au sevrage, les animaux on
était reparti en 2 lots. Les lapins du groupe exercice (EXE) on
était élevé collectivement dans des cages géantes
(1.32 m2) munies de 2 obstacles verticaux, éparant les
sources d'alimentation et l'abreuvement. Les lapins du groupe secondaire (SED)
ont était placé individuellement dans des cages. Les lapins des
lots EXE présentent au cours de la période d'étude, une
vitesse de croissance similaire à celle des lapins des lots SED mais
l'indice de consommation est amélioré. En effet la consommation
d'aliment du lapin du lot EXE tend à être inférieure
à celle des lapins du lot SED. Dans les dispositifs a plateforme, jehl
et al (2003), ont précédemment observé une croissance
similaire ou légèrement détériore par apport au
lapin élevé en cage collective classique.
II 3.2. L'influence du numéro de portée :
Selon Ouyed et al. (2007b), les lapereaux provenant
de la 4ieme et la 5ieme portée présentent les performances les
plus faible pour le GMQ (43.7 g/j Vs 45.9 g/j a la 2éme portée),
la CMQ (131.5 g/j Vs 138.7 g/j a la 1iere portée) et le poids a 63 jours
(2247 g Vs 2309 g a la 2éme portée). Ces résultats sont en
désaccord avec ceux d'Ozimba et Lukefahr (1991), que ne rapporte aucun
effet significatif du numéro de la portée sur les performances de
croissance. Orengo et al. (2004) obtient les performances les plus
faibles pour les poids en 60 jours, la vitesse de croissance et la consommation
alimentaire chez les lapins issus des 1iere portées.
II.3.3.Influence de l'alimentation :
L'effet du rationnement sur la croissance a été
rapporté plusieurs auteurs : une restriction alimentaire à
l'engraissement conduit à une réduction de la vitesse de la
croissance si la ration distribuée est inferieure a 85-90% de l'aliment
distribué à volonté (Castello et al., 1989 ;
Arveux, 1991 ; Tudela et Lebas, 2006).
25
Chapitre II Partie bibliographique
La présence ou l'absence des aliments dans la ration,
l'équilibre entre divers constituant et le niveau d'énergie et de
protéines dans la ration, sont des facteurs qui interviennent dans la
croissance des lapereaux (Ouhayoun, 1983). La vitesse de croissance est
maximisée si les équilibres recommandés sont
respectés : un aliment distribué à volonté, de 2500
kcal d'énergie digestible, 16% de protéines, 10 à 14 % de
cellulose brute et de 2 à 3% de lipides (Henaff et Jouve, 1988).
Dès qu'il y a déséquilibre, la vitesse de croissance sera
ralentie.
La distribution d'un aliment rationné de nuit
plutôt que de jour conduit à l'amélioration de l'indice de
consommation lors des 3 premières semaines d'engraissement. Ce
bénéfice se traduit sur la globalité de l'engraissement
dont la fin est menée en alimentation à volonté.
L'observation des lapins en fin d'engraissement lors du passage à
l'alimentation à volonté permettrait de savoir si le rythme
imposé aux animaux pendant les 3 semaines après sevrage est
maintenu (Weisseman et al., 2009).
? L'effet de l'apport des protéines :
Un taux élevé de protéines dans la ration
accélère la croissance (Lebas et Ouhayoun, 1987). Lorsqu'il y a
la baisse de la qualité et la quantité de ces derniers, le lapin
réduit sa consommation et donc sa croissance (Lebas et al.,
1984). L'absence d'un seul acide aminé essentiel peut être
considéré comme un manque globale de protéines (Lebas et
Colin ; 1992).
Cependant un excès de protéines peut perturber
l'équilibre dans le caecum en stimulant la flore protéolytique.
Les concentrations élevées en ammoniaque accroissent le pH
d'où le risque de troubles digestifs (Maertens et De Groote, 1987 ;
Peeters, 1988).
26
Chapitre II Partie bibliographique
? L'effet du rapport protéines / énergie
:
Après le sevrage, les équilibres alimentaires de
la ration en particulier la concentration en énergie digestible et le
taux des protéines digestibles, ont une importance
prépondérante sur la croissance des lapereaux. L'effet du niveau
protéique sur la croissance dépend de la concentration
énergétique de l'aliment. Ainsi, Martina et al. (1974)
n'observent pas la différence de croissance chez le lapin
réservant des aliments iso-énergétique de 2400 kcal / kg
et contenant 16 à 18% de protéines. Mais avec une teneur en
énergie plus élevée de 2550 kcal ED/kg, l'aliment ne
contenant que 16% de protéines diminue les performances de croissance et
d'abattage (Tableau 10).
Tableau 10 : L'effet du niveau
protéique et de la concentration en énergie digestible
de
l'aliment sur les performances de l'abattage du lapin âgé de
90 jours (Martina et al., 1974).
|
Energie (kcal/kg)
|
|
2400 2550
|
|
|
Protéines %
|
16
|
18
|
16
|
18
|
|
P/E (g/100kcal)
|
6,67
|
7,5
|
6,27
|
7,05
|
|
Poids (kg)
|
2,12
|
2,15
|
1,83
|
2,39
|
? L'effet de l'apport de lest :
Dans l'alimentation des lapines en croissance, un apport
minium de lest est considéré comme nécessaire pour assurer
un bon fonctionnement du tube digestif. La croissance est sensiblement
réduite lorsque l'apport en fibre est déficient (<16% ADF)
(Peinheiro et Gidenne, 1999). Perez et al. (1996) suggèrent
qu'un taux assez élevé en cellulose est nécessaire en
début de la croissance pour réduire la mortalité, alors
qu'un taux de 12% semble suffisant en fin d'engraissement, s'il renferme au
moins 4,5% de lignine. Cependant, l'excès de cellulose brute (>16%)
peut réduire la teneur en énergie digestible et la faire passer
en dessous du seuil de régulation des aliments (Lebas, 1984). Le lapin
sera simultanément en carence en protéines et en énergie.
Un déficit <12% entraine un ralentissement du transit digestif.
27
Chapitre II Partie bibliographique
II 3.7. Influence de l'environnement :
? L'effet de la température ambiante :
Les performances de croissance sont affectées à
partir de 25 °C (Grazzani et Dubini, 1982 ; Samoggia, 1987).
L'augmentation de la température ambiante entraine une réduction
de l'ingestion alimentaire, d'où baisse des performances car l'animal se
trouve en déficit nutritionnel et donc en brusque ralentissement de la
croissance (Colin, 1985 ; 1995).
Par contre, une baisse de la température engendre une
consommation accrue de l'aliment et donc une augmentation de la vitesse de
croissance mais un mauvais indice de consommation.
Tableau 11 : Effet des basses et hautes
températures sur la croissance
|
(Cheiriccato et al., 1992).
|
|
|
Performance / Température °C
|
11-12
|
26-28
|
|
Poids initial (g)
|
1154
|
1171
|
|
Poids final (g)
|
3227
|
2686
|
|
GMQ (g/J)
|
32,6
|
26,6
|
|
? Effet de la saison :
|
|
|
Le poids des lapins nés en saison fraiche est plus
élevé que celui des lapins nés en saison chaude (Kamal et
al, 1994). Aussi, le GMQ est plus élevé que celui de la
période chaude avec respectivement 37 et 25 g/J (Cheiriccato et al.,
1992). Ainsi les performances de croissance sont meilleures pendant le
temps d'hiver et diminue en printemps et en été.
28
Chapitre II Partie bibliographique
Tableau 12 : Effet de saison sur les
caractères de croissance
(Baselga, 1978)
|
Critères
|
Poids moyen au
sevrage
|
Poids moyen à
l'abattage
|
GMQ (g)
|
|
Saison
|
|
|
|
|
Hiver
|
547
|
2261
|
35
|
|
Printemps
|
599
|
2152
|
31,7
|
|
Eté
|
550
|
2114
|
32,2
|
|
Automne
|
549
|
2220
|
34,1
|
? Effet de l'hygrométrie :
Le lapin est un animal très sensible aux variations de
l'hygrométrie, une faible hygrométrie favorise la formation de
poussière qui dessèche les voies respiratoire entrainant ainsi
une sensibilité accrue à l'infection, il ne l'est pas lorsque
celle-ci est trop élevée (Lebas et al., 1996). Une
humidité maintenue entre 55 et 80% est de préférence, elle
serait idéal entre 60 et 70% (Lebas et al., 1991).
? Effet de la densité :
Selon Colmin et al. (1982), une densité de
15,6 lapin /m2 permet une forte vitesse de croissance et moins de
compétition entre les animaux. Par contre, une densité
supérieure à 16 lapins /m2 réduit les
performances de croissance (Martin, 1982). Lebas et al., (1991)
précisent qu'il ne faut pas placer plus de 16 à 18 lapins /
m2 ceci dit, ne pas dépasser 40 kg de poids vif /
m2.
? Effet du mode de logement :
Le mode de logement a un effet très important sur la
croissance. En effet, Jehl et al. (2003) ont constaté que les
lapins logés en parc présentent une vitesse de croissance
29
Chapitre II Partie bibliographique
inférieure à celle des lapins logés en
cages et le poids de ces derniers à l'abattage est ainsi
supérieur de 130g (Tableau 13).
Tableau 13 : L'incidence du mode de
logement sur les performances zootechniques du lapin (souche Hyplus) (Jehl et
al., 2003).
|
Cages
|
Parc
|
|
Poids à 35 J (g)
|
907
|
904
|
|
Poids à 49J (g)
|
1651
|
1549
|
|
Poids à 63 J (g)
|
2252
|
2111
|
|
Poids à 70J (g)
|
2446
|
2251
|
II4. Le rendement de la carcasse :
II.4.1. Définition de la carcasse :
Une carcasse est un produit de l'abattage après
saignée, dépouillement et sans ses viscères abdominales
(Jaim Camps, 1983). Nous distinguons :
? Une carcasse chaude :
Obtenue après la saignée et
éviscération de l'animal. Elle comporte les
extrémités des membres (manchons sur lesquels subsistent le
pelage), les reins, les viscères thoraciques ainsi que les gras
péri-rénal et inter-scapulaire. Le poids de la carcasse chaude
est pris entre 15 et 30 minutes après l'abattage (Blasco et al.,
1993). Un lapin de boucherie de 2,2 kg fournit à l'âge de 10
à 11 semaines une carcasse chaude de 1,395 kg (Ouhayoun, 1989).
30
Chapitre II Partie bibliographique
? Une carcasse froide :
Obtenue après ressuage et réfrigération dans
une chambre froide pendant 24h à 4°C. Après la suppression
des manchons, la carcasse commerciale pèse 1.285 kg soit un rendement de
57,1%.
Les critères de valeur bouchère dont les mesures
recommandées sont : le poids de la carcasse, le rendement à
l'abattage, l'adiposité, le rapport muscle/os et la découpe
(Balasco et al., 1990 ; Larzur et Gondert, 2005).
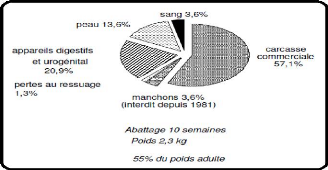
Figure 07 : Rendement en viande d'un
lapin de format moyen de 2,3kg
(Synthèse Ouhayoun, 1989)
II.4.2. Définition du rendement :
Le rendement à l'abattage est le rapport entre le poids
de la carcasse commerciale et le poids vif, il varie en fonction de plusieurs
facteurs, nous pouvons retrouver :
? Effet de la race : le rendement est plus
élevé chez les races lourdes (Fattal,
1987).
31
Chapitre II Partie bibliographique
? Effet de l'âge et du poids à l'abattage
: le rendement augmente avec l'âge. En
effet, selon Ouhayoun (1989) et Roiron (1991), le rendement
passe de 50% à 57% chez les animaux abattus à 60 jours par
rapport à ceux abattus à 70 jours.
? Effet de l'alimentation : quel que soit le
moment de son application, la réduction
quantitative ou qualitative de l'aliment réduit le
rendement de la carcasse (Ouhayoun, 1989). En effet, une réduction de la
teneur de l'aliment en certains acides aminés à l'exemple de la
méthionine réduit le rendement de la carcasse de 59% à
57,7%.
Le lapin de population locale algérienne est
caractérisé par un poids vif à l'abattage faible
comparé aux races et souches sélectionnées (Berchiche et
Lebas, 1990 ; Berchiche et al., 2000). Par contre, le rendement de la
carcasse chaude est satisfaisant (Tableau 14).
Tableau 14 : le rendement à
l'abattage du lapin local (synthèse bibliographique).
|
Berchiche et al.
(2000) (n=20)
|
Le rendement
Lounouaci (2001)
(n=16)
|
Benali (2009)
(n=17)
|
|
Pva (g)
|
1745
|
1740
|
2166
|
|
PP (g)
|
148,74
|
166,93
|
221
|
|
TDP (g)
|
27765
|
321,93
|
379
|
|
CC (g)
|
1110
|
1204,3
|
1324
|
|
CF (g)
|
/
|
1158,7
|
1295
|
|
PM (g)
|
/
|
41,73
|
/
|
|
GPR (g)
|
/
|
21,25
|
20,1
|
|
Rendement CC/Pva (%)
|
65,4
|
69,23
|
65
|
|
Rendement CF/Pva (%)
|
/
|
66,59
|
63,5
|

Chapitre III
Etat sanitaire
3
32
Chapitre III Partie bibliographique
Chapitre III : Etat sanitaire
III.1. Pathologies bactériennes :
III.1.1. Colibacillose :
Les Colibacilles (Escherichia coli) sont des
hôtes normaux de la flore intestinale de nombreuses espèces
animales. Cependant, certaines souches peuvent représenter un des agents
étiologiques les plus importants de troubles intestinaux chez les
animaux. Chez le lapin, la richesse de la flore colibacillaire est
limitée : moins de 104-105 ufc d'Escherichia coli/g de contenu
caecal ; certains lapins n'hébergeant pas d'Escherichia coli.
Par contre, tout dérèglement digestif peut se traduire par
une sévère colidysbactériose, à savoir une forte
élévation de la flore colibacillaire saprophyte jusqu'à
108-109 ufc d'Escherichia coli/g de contenu caecal. Les
Escherichia coli responsables de diarrhées primaires ont
été classés dans au moins cinq catégories. Les
Escherichia coli pathogènes du lapin appartiennent au pathovar
EPEC (entéropathogenic Escherichia coli). Ces colibacilles
entéropathogènes du lapin (REPEC) sont comparables aux EPEC
humains. Ils s'attachent à la muqueuse intestinale et provoquent des
lésions spécifiques d'attachement/effacement au niveau de la
bordure en brosse des entérocytes (effacement des
microvillosités). Ils ne produisent pas d'entérotoxines
thermostables ou thermolabiles connues et ne sont pas entéro-invasives
(Licolis, 2010).
Les travaux effectués par Bouguerroumi A et Boumahdi H
(2006), à Ain Taya (Alger) sur des lapins de race locale
algérienne, montrent que le taux de mortalité liée la
colibacillose est approximativement 36,11 %. Ce pourcentage
s'élève en particulier chez les lapereaux d'engraissement.
Alors que les travaux effectués par Belgacem (2012) au
Laboratoire de microbiologie de l'Ecole Nationale Supérieure
Vétérinaire d'Alger sur 12 lapins suspects, concernant les 8
prélèvements étudiés pour une suspicion de
colibacillose sur l'ensemble des échantillons
33
Chapitre III Partie bibliographique
analysés, 4 ont montré la présence de
colibacillose par la présence de germe sur le foie. Ce nombre laisse
apparaitre un taux de 22,22% de colibacillose parmi les 12 cas
testés.
III.1.2. Entérotoxémie :
L'entérotoxérnie est plus fréquente
surtout dans les élevages fermiers. Avec ou sans entérite
mucoïde, son évolution peut être très rapide (de un
à sept jours). Elle survient le plus souvent en fin de gestation ou au
milieu de la lactation, où elle se superpose quelquefois avec des signes
de pneumonie aigue. En élevage traditionnel, des complications de
parésie ou de paraplégie sont fréquentes surtout chez les
femelles trop grasses, car trop nourries et soumises à un rythme de
reproduction trop faible. Dans ce cas, la prophylaxie consistera à
adapter le rythme de reproduction aux capacités alimentaires de
l'élevage. Il n'existe aucun traitement (Lebas et al.,
1996).
Les travaux effectués par Bouguerroumi A et Boumahdi H
(2006), à Ain Taya sur des lapins de race locale algérienne,
montrent que les entérotoxémie sont présentées dans
9 cas soit approximativement 25 % de cas trouvés (cas de
mortalités), elles peuvent être, après l'analyse de
cellulose brute, dues au manque de cellulose dans l'aliment, ce qui a
provoqué le développement des clostridies pathogènes,
c'est le rôle de lest (Bouguerroumi et Boumahdi, 2006).
III.2. Pathologies parasitaires :
III.2.1. Cryptosporidioses :
La cryptosporidiose est une parasitose émergente
opportuniste. Le plus souvent asymptomatique, elle peut parfois se manifester
cliniquement par des troubles digestifs chez les sujets
immunodéprimés en présence d'affections intercurrentes.
L'agent causal est un protozoaire intracellulaire de la famille des coccidies
appartenant au genre Cryptosporidium (Chermette et Boufassa-Ouzrout,
1990). Plusieurs espèces ont pu être isolées d'un grand
nombre de vertébrés, y compris l'homme chez lequel
l'intérêt porté à ce parasite est
34
Chapitre III Partie bibliographique
directement lié à la survenue
d'épidémies dès les années 80. Plus de 90% des 165
épidémies liées à l'eau contaminée sont
survenues aux USA et en Europe parmi lesquelles celles de Milwaukee (USA,
1993), de Sète (France, 1998), de Dracy-le-Fort (France, 2001) et de
Divonne-les-Bains (France, 2003) (Derouin, 2010). L'épidémie la
plus récente ayant incriminé le génotype du lapin a eu
lieu au Royaume Uni en 2008. Cet épisode souligne que le risque
zoonotique dû à la contamination du réseau d'eau de boisson
ne doit pas être négligé (Robinson et Chalmers, 2009).
Plusieurs espèces animales ont fait l'objet d'investigations
limitées à l'Europe, à l'Amérique du nord et
à l'Australie (Nolan et al., 2010).
Chez le lapin de population locale, les études
effectuées par Mezali et al. (2015) montrent que sur les 102
prélèvements examinés, l'ensemble des échantillons
de matières fécales de groupe sont positifs aux cryptosporidies,
représentant ainsi une prévalence de 100%. Sur les 42 frottis de
côlon, 35 sont positifs, soit une prévalence de 83,33%. Le taux
d'infestation est de 87,5% chez les mâles contre 77,78% chez les femelles
et n'est pas significativement différent entre les deux sexes (Mezali
et al., 2015).
III.2.3. Coccidioses :
Les coccidies, ce sont des protozoaires, phylum le plus
primitif du règne animal, et des sporozoaires, presque toutes les
coccidies du lapin font partie du genre Eimeria. Ces Eimeria
se développent dans les cellules des épithéliums de
l'appareil digestif (intestin, foie), au moins 11 espèces parasitent le
lapin, une seule parasite le foie, les 10 autres sont à localisation
intestinale. Une revue complète a été faite par Coudert
(1989). Les coccidioses, comme toute maladie, peuvent laisser un certain nombre
de séquelles, rénales ou hépatiques en particulier, qui ne
sont pas sans conséquences soit sur l'état d'engraissement
l'abattage, soit sur l'avenir de l'animal si celui-ci est conserve comme
reproducteur. En outre, une maladie se complique souvent d'autres affections.
En effet, ces observations ont été obtenues sur des lapins
élevés dans des conditions particulièrement favorables,
c'est à dire qu'il n'y avait pratiquement pas de surinfections
bactériennes (Tableau 15) (Lebas et al.,
1996).
|
Chapitre III Partie bibliographique
|
|
Tableau 15 : Pouvoir pathogène
comparé des différentes coccidies intestinales du
lapin (Lebas et al., 1996).
Pathogénicité Espèce d'Eimeria
Symptômes
|
Non pathogène ou peu pathogène
|
E. coecicoia
F. oxlgua
E. perforans E. vejdovskyi
|
Aucun signe de maladie ou
légère chute du GMQ
Pas de diarrhée
|
Pathogène E. media
E. mogno E. irresidua E. piroformis
Chute du GMQ
Diarrhée inconstante Peu ou pas de mortalité
35
|
Très pathogène E. intestinalis
E. flavescens
|
Sévère chute GMQ Diarrhée importante
Mortalité élevée
|
Les travaux effectués par Hennab et Aissi (2013) dans
quatre élevages (Bordj Ménaiel (N°1), Khemis El Khechna
(N°2), Draa Ben Khedda (N°3) et Ouaguenoune (N°4)) sur 19
lapines de population locale algérienne montrent que l'excrétion
oocystale moyenne n'a pas varié significativement entre les
élevages étudiés. Cependant, elle diffère
significativement entre les deux périodes étudiées
(gestation et lactation). En effet, Chez les lapines gestantes,
l'excrétion oocystale moyenne était nulle dans l'élevage
(2) et faible dans les élevages (1), (3) et (4). Ces résultats
sont comparables à ceux obtenus par Gallazzi (1977) et Papeschi et
al. (2013) qui indiquent que les femelles gestantes, dans les conditions
naturelles, sont excrétrices d'oocystes. Une excrétion d'oocystes
durant le dernier tiers de gestation, peut être liée au bilan
énergétique négatif durant cette période, ce qui
favorise la baisse de l'immunité. Dès la mise-bas,
l'excrétion oocystale moyenne était importante dans les
élevages (1) et (3) ; cette excrétion diminuait progressivement
en allant vers la période de tarissement seulement pour l'élevage
(1). Dans les élevages (2) et (4), l'excrétion était
faible. Les mêmes résultats ont été obtenus par
Gallazzi (1977) indiquant, que pendant le début de lactation, une
augmentation de l'excrétion oocystale est observée avec un pic
vers la 3ème
36
Chapitre III Partie bibliographique
semaine de lactation. Un pic d'excrétion oocystale a
été constaté vers la dernière semaine de lactation
chez les lapines de l'élevage (3). Ceci pourrait être lié
à une baisse de l'immunité liée elle-même à
une dégradation de l'état général, engendrée
par l'existence de plusieurs infections intercurrentes, d'une part, et le
mauvais entretien de l'élevage d'autre part. Une diminution de
l'excrétion oocystale entre les périodes de gestation et
lactation a été constaté au niveau de l'élevage (4)
ce qui pourrait être lié à l'application des bonnes
conditions d'élevages. Chez les lapereaux, dès le
19ème et le 25ème jour d'âge, une
excrétion oocystale a été constatée dans les
élevages (1) et (3), avec un taux inférieur à 50 OPG. La
présence prématurée des oocystes dans les crottes des
lapereaux âgés de 19 jours, pourrait être probablement le
résultat d'une forte contamination précoce des lapereaux et/ou au
statut immunitaire déficient (Drouet-Viard et al., 1997).
Aussi, Sur les 160 oocystes sporulés et
identifiés, une prédominance des espèces intestinales et
une association de 2 à 4 espèces d'Eimeria a
été notée dans les 4 élevages (Tableau
16). L'espèce dominante est Eimeria magna (43%)
(Hennab et Aissi, 2013).
Tableau 16 : Oocystes sporulés
d'Eimeria isolés. (Hennab et Aissi, 2013)
Espèces de coccidies Pourcentage (%)
E. stiedai 23%
E. magna 43%
E. coecicola 3%
E. perforans 9%
E. media 19%
E. exigua 3%
III.3. Pathologies virales :
La maladie hémorragique virale du lapin est la maladie
virale la plus fréquente chez le lapin. Elle est hautement infectieuse
et souvent fatale pour le lapin domestique ou sauvage de l'espèce
Oryctolagus cuniculus. L'agent étiologique de la maladie, le
RHDV, est un
37
Chapitre III Partie bibliographique
Calicivirus appartenant au genre Lagovirus.
Elle est endémique dans les populations de lapins sauvages d'Europe,
d'Australie et de Nouvelle Zélande. L'apparition de la VHD a
été responsable d'importantes pertes économiques dans les
élevages industriels de lapins mais le développement de vaccins a
permis de contrôler la maladie. Cependant, celle-ci reste une menace pour
les populations de lapins de garenne (Le gal et al., 2015). Plusieurs
élevages du lapin ont été touchés en Algérie
durant l'année 2014-2015 par cette pathologie. Cette dernière
à cause la perte des élevages en entier notamment dans la wilaya
de Blida et d'Alger. Le diagnostic de cette pathologie a été
confirmé par le laboratoire central de médecine
vétérinaire cependant, aucune étude n'a été
entreprise.
|
Conclusion
&
Recommandations
|
38
Conclusion
La cuniculture à un niveau rationnel doit compter
davantage sur l'appui scientifique des programmes de recherche universitaire et
sur les programmes de développement : acquisition des connaissances
scientifiques et vulgarisation des techniques de la cuniculture rationnelle.
A la lumière des résultats bibliographiques,
nous pouvons conclure que chez la population locale algérienne :
Le taux de réceptivité
était de 89% ce qui est plus élevé par rapport
à celui de la population blanche mais du même niveau que la souche
synthétique. La bonne réceptivité de la lapine locale
s'expliquerait par le mode de reproduction pratiqué (saillie naturelle)
et le rythme de reproduction adopté (La remise à la reproduction
s'est faite 10 à 12 jours après la mise bas).
Le taux de fertilité était de
87% ce qui est plus bas que celui de la race Néo-Zélandaise
blanche et la race Californienne, mais plus élevé que la race de
Chinchilla et de Géant des Flandres.
La prolificité de la population locale
Algérienne est inférieure à celle des races
européennes notamment le Fauve de Bourgogne, le Géant d'Espagne
et le Gris de Carmagnola, estimée en moyenne à 8,8. Ces faibles
performances dans les élevages fermiers pourraient être
attribuées au rythme de reproduction adopté.
la mortinatalité était en
moyenne de 18,9% ce qui est plus élevée que celui des populations
locales égyptienne et marocaine et même plus élevée
que celui des races Européennes notamment le Fauve de Bourgogne et le
Chinchilla. Cette situation pourrait être liée à la perte
de portées entières à la naissance chez certaines femelles
avec un comportement maternel déficient (mises bas sur le grillage,
abandon des portées, cannibalisme) et aux mauvaises conditions
d'ambiance (température élevée durant l'été
et basse en hiver).
39
Conclusion
Le poids à la naissance et au sevrage
étaient de 49,4g et 463,7g respectivement, ce qui est plus bas
que celui des races Californienne et Néo-zélandaise.
Le gain moyen quotidien était de 22,
72g/J ce qui est proche à celui des souches améliorées.
Le poids vif à l'abattage était
faible comparé aux races et souches sélectionnées. Par
contre, le rendement de la carcasse chaude est
satisfaisant.
Le taux de mortalité liée à la
colibacillose était approximativement 36,11 %. Ce pourcentage
s'élève en particulier chez les lapereaux d'engraissement.
La mortalité liée à
l'entérotoxémie s'est présentées dans 9
cas soit approximativement 25 % de cas trouvés. Elle peut être,
après l'analyse de cellulose brute, dues au manque de cellulose dans
l'aliment, ce qui a provoqué le développement des clostridies
pathogènes, c'est le rôle de lest.
Le taux de morbidité liés à la
cryptosporidiose était 100% : l'ensemble des
échantillons de matières fécales de groupe sont positifs
aux cryptosporidies, représentant ainsi une prévalence de 100%.
Le taux d'infestation est de 87,5% chez les mâles contre 77,78% chez les
femelles et n'est pas significativement différent entre les deux
sexes.
L'excrétion oocystale moyenne lors de
coccidiose diffère significativement entre la période de
gestation et la période de la lactation. En effet, Chez les lapines
gestantes, l'excrétion oocystale moyenne était nulle à
faible dans les élevages mais une excrétion d'oocystes durant le
dernier tiers de gestation, peut être liée au bilan
énergétique négatif durant cette période, ce qui
favorise la baisse de l'immunité. Dès la mise-bas,
l'excrétion oocystale moyenne était importante. Et
l'espèce dominante était Eimeria magna (43%).
Plusieurs élevages du lapin ont été
touchés en Algérie durant l'année 2014-2015 par la maladie
hémorragique virale du lapin, cependant, aucune étude n'a
été entreprise
40
Conclusion
Ainsi, les conclusions auxquelles nous avons abouti, nous ont
permis de dégager plusieurs axes de recherche intéressant
à explorer à l'avenir. A ce propos, plusieurs travaux de
recherche seraient nécessaires à réaliser à savoir
:
Étendre l'expérience à d'autres
régions du pays à titre comparatif, pour pouvoir disposer
d'autres éléments d'information, plus précis et permettant
une meilleure connaissance de performances de croissance et de reproduction du
lapin locale ainsi que les facteurs influençant sur ces derniers.
Toutefois, il faut développer la collaboration entre
équipes de recherches pour éviter la dispersion dans le choix des
thèmes. Il faut aussi activer dans le domaine de la gestion technique
d'élevage pour déterminer la rentabilité de ce type
d'élevage en Algérie et faire connaître davantage cette
production animale.
Références
bibliographiques
|
40
Références
bibliographiques
A
Afifi E.A., 2002 : The Gabali rabbits (Egypt). In rabbit
genetic resources in Mediterranean
countries. Options Méditerranéennes,
série B, CIHEAM, Zaragoza, N° 38, 51-64. Anonyme, 2015 : Position
de lordose
http://www.cuniculture.info/Docs/Biologie/biologie-
07-3.htm
Arveux P, 1988 : Production cunicole en période
estivale. Cuniculture, 82, 197-199.
Arveux P, 1991 : Le rationnement alimentaire quantitatif en
élevage cunicol. Cuniculture, N°98, 97-98
B
Barkok A, Jaouzi T, 2002 : The Zemmouri rabbits (Morocco). In
rabbit genetic resources in Mediterranean countries. Options
Méditerranéennes, série B, CIHEAM, Zaragoza, N° 38,
175-185.
Baselga M, 1978 : Analisis gentico de diversa caracetristica
de crecimiento en el conjeo de production de carne. 3éme symposium de
cunicultura. Valencia, 1-10 Nov.
Baselga M, 2002a : Line A (Spain), In rabbit genetic resources
in Mediterranean countries. Options Méditerranéennes,
série B, CIHEAM, Zaragoza, N° 38, 221-230.
Baselga M., 2002b : Line V (Spain). In rabbit genetic
resources in Mediterranean countries. Options Méditerranéennes,
série B, CIHEAM, Zaragoza, N° 38, 231-241.
Belgacem ryma, 2012 : Etude de la colibacillose chez le lapin
de population locale. Mémoire de fin d'etude en sciences
vétérinaire, ENV d'Elharache.
Berchiche M et Kadi S.A 2002 The Kabyle rabbits (Algeria).
Options Méditerranéennes,13-20.
http://ressources.ciheam.org/om/pdf/b38/02600006.pdf
Berchiche M et Lebas F, 1990 : Essai chez le lapin de
complémentation d'un aliment pauvre en cellulose par un fourrage
distribué en quantité limité : digestibilité et
croissance. 5ieme journée de la recherche cunicol. Paris 12-13
Déc
Berchiche M, Kadi S.A, Lounaouci G, 2000. Elevage rationnel de
lapin de population locale : alimentation, croissance et rendement à
l'abattage. 3ieme journée sur les productions animale « Conduit de
performance d'élevage », 13, 14, 15, Nov, P293-298
Berchiche M., Zerrouki N., 2000. Reproduction de femelles de
population locale: Essai d'évaluation de quelques paramètres en
élevage rationnel. 3émes Journées de
41
Références
bibliographiques
Recherche sur les Productions Animales : « Conduite et
performance de l'élevage » Tizi- Ouzou.13, 14, 15 Novembre,
285-291.
Besançon B, Lebas F, Abouyoub A, 1971 : composition
minérale du lait de lapine. Variation en fonction de stade de lactation,
Ann zootech 20(4), 287-495.
Bidanel P.J, 1998 : Nouvelles perspectives
d'amélioration génétique de la prolificité des
truies. INRA. Prod. Anim., 11, 219-221.
Blasco A Ouhayoun j, Maseoro G, 1993 : Harmonisation of
criteria and terminology in rabbit meat researc. World rabbit Sci, 1,3-10
Blasco A, 1992 : Croissance, carcasse et viande du lapin.
Séminaire sur les systèmes de production de viande de lapin.
Valencia, 14-25 septembre 1992.
Blasco A, Gou P, Santacreu M.A, 1990 : Effect of selection on
change in body coposition of rabbit. 4th world congress of genet. Appl. Livest.
Prod., Edenbergh, vol 16 362-365.
Blocher F et Franchet A, 1990 : Fertilité,
prolificité et productivité au sevrage en insémination
artificielle et en saillie naturelle, influence de l'intervalle mise bas
saillie sur le taux de fertilité. 5èmes Journées de la
Recherche Cunicole. Paris, France. 12-13 Décembre. Tome 1, communication
2, 1-14.
Bolet G, 2002a : Fauve de Bourgogne (France). In rabbit
genetic resources in Mediterranean countries . Options
Méditerranéennes, série B, CIHEAM, Zaragoza, N° 38,
85-92.
Bolet G, 2002b : Strain INRA 2066 (France). In rabbit genetic
resources in Mediterranean countries. Options Méditerranéennes,
série B, CIHEAM, Zaragoza, N° 38, 109-116
Bolet G, Brun J.M, Lechevestrier S, Lopez M, Boucher S, 2004 :
Evaluation of the reproductive performance of eight rabbits breeds on
experimental farms, Anim. Res. 53 (2004) 5965.
Bolet G, Brun JM, Theau-Clément M, Esparbie J and
Falieres J, 1999 : Constitution d'une souche synthétique de lapins
à l'INRA : 3. Aptitude à la combinaison avec la souche 1077 pour
produire une femelle parentale. Résultats préliminaires.
8èmes Journées de la Recherche Cunicole, France, Paris,
131-134
Bolet, G. 1998. Problèmes liés à
l'accroissement de la productivité chez la lapine. INRA Prod Anim, 11,
235-238. Juin 1998.
Bouguerroumi A et Boumahdi H, 2006 : Pathologie intestinales
du lapin locale, Mémoire de fin d'etude en sciences
vétérinaire, ISV de Blida, p78.
42
Références
bibliographiques
Bouzekraoui A, 2002 : The Tadla rabbits (Morocco). In rabbit
genetic resources in Mediterranean countries. Options
Méditerranéennes, série B, CIHEAM, Zaragoza, N° 38,
165-174.
Brun J.M ,Theau-Clement M, Bolet G, 2002 : Evidence for
heterrosis and maternal effects on rabbit semen characteristics. Ani.Res ; 51
(433-442)
C
Cantier J, Vezinhet A, Rouvier R, Dauzier L, 1969 :
Allométrie de croissance chez le lapin (Oryctolagus Cuniculus).
Principaux organes et tissus, Ann. Biol. Anim. Bioch. Biophys. 9(1) : 5-39.
Castaing J, 1979 : Aviculture et petits élevages. Ed,
J-B. Bailliére. 304p
Castello J.A, Leonart F, Luzi F, 1989 : Cité par
Tudella F et Lebas F, 2006. Experiencias de diverso tipos de restriccion en el
coneje. XIV Symposium de cuniculture, 12-14 junio, Manresa, 91-104.
Chaou T, 2006 : etude des paramètres zootechniques et
génétiques d'une ligné paternelle
sélectionnée mise en place en G0 et sa descendance, du lapin
local « Oryctolagus Cuniculus » Mémoire de magistère,
Ecole nationale supérieure veterinaire, 102p
Cheiriccato M, Bailonil L, Rizzi C, 1992 : The effect of
environmental température en the performance of growing rabbi, 5th world
rabit congress, corvalis (USA), July (1992), 2,723-731.
Chermette R et Boufassa-Ouzrout S, 1990 : Cryptosporidiosis -A
Cosmopolitan Disease in Animals and Man. Vet. Parasitology, 35 : 179-182
Chmitelin F, Rouillere R, Bureau J : 1990 Performances de
reproduction des femelles en insémination artificielle en
post-partum. 5èmes Journées de la Recherche Cunicole,
1213 Décembre, 1990, Paris, France. Tome I: Comm. 4
Colin M, 1985 : les problèmes liés à
l'été dans l'élevage du lapin. Cuniculture, N°63,
12(3), 177-180
Colin M, 1995 : comment maitriser les effets de chaleur.
L'éleveur du lapin, Juin/Juillet, 2327
43
Références
bibliographiques
Colmin G.P, Franck Y, Le loup P, Martin S, 1982 : Incidence du
nombre de lapin par gage d'engraissement sur les performances zootechniques.
3éme journée de la recherche cunicol, 9-8 Déc, Paris,
Communication N°24.
Combes S, Moussa M, Gondret F, Doutriloux J.P, Remignon H,
2005 : Influence de l'exercice physique sur les performances de croissance, la
qualité des carcasses et les caractéristiques mécaniques
de l'attachement de la viande à l'os après cuisson chez le lapin.
11éme journée de le recherche cunicole, 29-30 Novembre 2005,
Paris.
D
De Rochambeau H, 2000 : Amélioration
génétique du lapin pour la production de la viande en France.
Situation actuelle et perspectives. Jour. Cuni. 24-45 Nov , 147-159.
Djellal F, Mouhous A, Kadi S.A, 2006 : Performances de
l'élevage fermier du lapin dans la région de Tizi-Ouzou,
Algérie. Livestock Reseach for Rural Developpment, 18 (7) 2006.
Drouet-Viard F., Coudert P., Licois D., et Boivin M.,
1997.Vaccination against Eimeria magna coccidiosis using spray dispersion of
precocious line oocysts in the nest box.Vet Parasitol, (70),61-66.
F
Fattal M, 1987 : comparaison de deux types de concentré
en production cunicol. Mémoire de licence en science naturelles
appliquées, Belgique
Feugier A, Fortun-Lamothe L, 2006. Extensive reproductive
rhythm and early weaning improve body condition and fertility of rabbit does.
Anim. Res. 55 (2006) 459-470.
Feugier A, Fortun-Lamothe L, Fuin H, 2005. Réduction du
rythme de reproduction et la durée de la lactation améliore
l'état corporel et la fertilité des lapines. 11émes
Journées de la Recherche Cunicole, 29-30 Novembre, 2005, Paris,
107-110.
Fortun- Lamothe, Bolet, 1995 : Les effets de la lactation sur
les performances de reproduction chez la lapine. INRA. Prod. Anim. 1995
Fromont A 2001 : Elevage de lapins. Ed, Educargi, 2001,
123p.
44
Références
bibliographiques
G
Gacem M, Zerrouki N, Lebas F, Bolet G, 2008. Strategy for
develeloping rabbit meat production in Algeria and selection of a synthetic
strain. 9th World Rabbit Congress, June 10-13, 2008, Verona, Italy.
Gacem M, Zerrouki N, Lebas F, Bollet G, 2009 : Comparaison des
performances de production d'une souche synthétique de lapins avec deux
populations locales disponibles en Algérie. 13èmes
Journées de la Recherche Cunicole, 17-18 novembre 2009, Le Mans,
France
Gallazzi D, 1977 : Cyclical variations in the excretion of
intestinal coccidialoocysts in the rabbit.Folia Vet Latina, 7(4), 371-380
Garcia M L , Baselga M, Piero R, 2000 : Correlated response on
growth traits in the line selected for litter size at weaning. 7th W.RC,
Valencia, Spain. July 2002.P 389-395
Garcia M.L et Perez A, 1989 : Effet de lactation y
numéro de lact tante sobre la mota, evaluatos per laparoscopia, en
conejas multiparas informacion tecnica economica agararien, 20 (80), 3-10
Gidenne T et Lebas F, 2005 : le comportement alimentaire du
lapin, 11éme journée de la recherche cunicole, 9-10 Juin,
109-113
Gomez E.A, Rafel O, Ramon J, 2002a : The Caldes Strain
(Spain). In rabbit genetic resources in Mediterranean countries. Options
Méditerranéennes, série B, CIHEAM, Zaragoza, N° 38,
193-198.
Gomez E.A, Rafel O, Ramon J, 2002b : The Prat Strain (Spain).
In rabbit genetic resources in Mediterranean countries. Options
Méditerranéennes, série B, CIHEAM, Zaragoza, N° 38,
199-208.
Grazzani R et Dubini F, 1982 : Cité par Cheiriccato G.M
, Rizzi C, Rosellato V, 1996. Growth and slautering performances of three
rabbit genotyp under environemental conditions. Ann. Zootech N°43,
311-318
H
Hafez E.S.E et al, 1967 cités par Questel, 1984 :
Questel G : 1984. Contribution à l'étude de la fertilité
chez le lapin domestique. Mémoire de fin de formation, INRA
Paris-Grignon, France, 65 p
45
Références
bibliographiques
Henaff R et Jouve D, 1988 : Mémonto de l'éleveur
du lapin. 7éme édition réalisée par l'AFC et ITAVI.
P448.
Henneb M et Aissi M, 2013 : Etude cinétique de
l'excrétion oocystale chez la lapine et sa descendance et identification
des différentes espèces de coccidies, 15èmes
Journées de la Recherche Cunicole, 19-20 novembre 2013, Le Mans, France.
P222-223.
Hulot F et Matheron G, 1981 : Effet du génotype, de
l'âge et de la saison sur les composantes de la reproduction chez la
lapine. Ann. Génét. Sél. Anim. 13, 131-150.
J
Jaim Camps, 1983 : In reniff et Douala T, 2002 : Contribution
à l'étude de la croissance du lapin de population locale
algérienne. Mémoire Ingénieur, Université de
Mouloud Mammeri de Tizi-ouzou, 63p
Jehl N, Meplain E, Merabito L, Combes S, 2003 : Incidence de
trois modes de logements sur les performances zootechniques et la
qualité de la viande de lapin. 10éme journée de la
recherche cunicole, 19-20 Nov, 2003, Paris.
K
Kadi S.A, Guermah H, Bannaelier C, Berchiche M, Gidenne T,
2011 : Nutritive value of sun-dried Sulla (Hedysarum flexuosum), and its effect
on performance and carcass characteristics of the growing rabbit. World Rabbit
Sci., 19:151-159.
Kamal A, Yamani K.O, Fraghaly H.M, 1994 : Adaptability of
rabbits to the hot climate. Option mediterraniennes, serie séminaire,
n°8, 97-101.
Kasa I.W et Twaites C.J, 2001 : Relation entre la
température scrotale et la mortalité des
spermatozoïdes chez le lapin Néozélandais
blanc. World rabbit science, 9(2), 87-89. Khalil M.H, 2002a : The Baladi
rabbits (Egypt). In rabbit genetic resources in Mediterranean
countries. Options Méditerranéennes,
série B, CIHEAM, Zaragoza, N° 38, 37-50.
Khalil M.H, 2002b : The Giza White rabbits (Egypt). In rabbit
genetic resources in Mediterranean countries. Options
Méditerranéennes, série B, CIHEAM, Zaragoza, N° 38,
23-36.
46
Références
bibliographiques
L
Laffolay B, 1985 : Croissance journalière du lapin.
Cuniculture, 12(6), 212-331.
Lakabi D, 2009 : Production de lapin en conditions de
production Algérienne, thèse de doctorat, Université de
Tizi-Ouzou, juin 2009
Lamming G.E et al, 1954 : The effect of the incipion vitamin A
deficiency on reproduction in the rabbit. I.Decidna, ova and fertilization.
Larzur C et Gondert F, 2005 : Aspect génétique
de la croissance et de la qualité de viande chez le lapin. INRA. Prod.
Anim, 18(2), 119-129.
Lazzaroni C, 2002 : The Carmagnola Grey rabbit (Italy). In
rabbit genetic resources in Mediterranean countries. Options
Méditerranéennes, série B, CIHEAM, Zaragoza, N° 38,
141-150.
Lazzaroni C, Andrione A, Luzi F, Zecchini M, 1999 :
Performances de reproduction du lapin Gris de Carmagnola : influence de la
saison et de l'âge des lapereaux au sevrage. 8émesJournées
de la Recherche Cunicole, Paris, 1999, 151-154.
Lebas et Zerrouki, 2010 : Comparaison des performances de
reproduction et de croissance d'une souche synthétique de lapins, avec
celles de lapins de 2 populations locales algériennes, dans 2 sites
expérimentaux. Atelier de travail sur la création d'une souche
synthétique, Baba Ali (Algérie) 14-15 juin 2010
Lebas F et Colin M, 1992 : World rabbit production and
research situation in 1992. 5the world rabbit congress (Orignon), July 25-30,
1-6.
Lebas F et Ouhayoun j, 1987 : Incidence du niveau
protéique de l'aliment du milieu d'élevage et de la saison sur la
croissance et les qualités bouchères du lapin. Ann. zootech, 36,
421-432
Lebas F, 1969 : Alimentation lactée et croissance
pondérale du lapin avant sevrage, Ann zootech, 18(2), 197-208.
Lebas F, 1984 : Le lapin elevage et pathologies, collection
F.O.A production et santé animal N°19.
Lebas F, 1991 : La production du lapin. A.F.C, 3éme
édition 2006p
Lebas F, 2002 : Biologie du lapin
http://www.cunicuture.info/docs/indexbiol.htm.
47
Références
bibliographiques
Lebas F, Caudert P, Rouvier R, De Réchembeau H, 1984 :
Le lapin : élevage et pathologie. Collection FAO Production et
santé animale 289p
Lebas F, Coudert P, De Rochambeau H, Thébault R.G, 1996
: Le lapin : Elevage et pathologie, Collection FAO : production et santé
animale p, 21-40-113-112-134.
Lebas F, Marionnete D, Haewaff R, 1991 : AFC (Association
française de la cuniculture). 3éme édition, p,21-40.
Lebas F, Marionnete D, Hennaf P, 1991 : La production de
lapin, technologie et documentation, LAVOISIER (3éme édition),
260p
Lebas F, Zerrouki N, Gacem M, Meftah I, Bolet G, 2007 :
Comparaison des performances de reproduction et de croissance d'une souche
synthétique de lapins, avec celles de lapins de 2 populations locales
algériennes, dans 2 sites expérimentaux. Atelier de travail sur
la création d'une souche synthétique, Baba Ali (Algérie)
14-15 juin 2010
Lebas F., 1974 : La mortalité des lapereaux sous la
mère, étude monographique. Cuniculture, 1, 8-11 et 40-45.
Licolis Dominique , 2010 : Pathologie d'origine
bactérienne et parasitaire chez le lapin :Apports de la dernière
décennie
Lopez M, Sierra I, 2002 : The Gigante de Espana Breed (Spain).
In rabbit genetic resources in Mediterranean countries. Options
Méditerranéennes, série B, CIHEAM, Zaragoza,
N°38,209-220.
Lounaouci G., Hannachi R., Berchiche M, 2012 : Elevage de
lapins descendants d'un hybride commercial en Algérie :
évaluation des performances de croissance et d'abattage .3éme
Congrès Franco-Maghrébin de Zoologie et d'Ichtyologie 6 -10
novembre 2012 Marrakech, Maroc
Lounouaci G, 2001. Alimentation du lapin de chair dans les
conditions de production algérienne. Mémoire de magistère
en sciences agronomiques, Université de Blida, 129p
M
Maertens L et De Groote G, 1987. Elevage. Revue de l'Agriculture,
N°5, V(40), 1185-1203. Martin S, 1982. En maternité, en
engraissement : les moyens d'améliorer la productivité.
Aviculture (hors-série) 19,21-24.
48
Références
bibliographiques
Martina C, Damian C, Palamaru E,1974. Retete de nutrituri
cobinate-gronulate cu diferite nivelle energo proteice pentru cresteria si
ingrasaria tineretului cunicul. Lucraril stiintiglice ale institului de
cercetari pentru nutritia animalia, 2,313-322.
Martinez-Gomez M, Distel H, Hudson R, 2004 : Overlaping
litters and reproductive performance in the domestic rabbit. Physiologie and
behavior. 82 (629-636)
Mattaraia V.G.M., Bianospino E., Fernandes S., Vasconcellos
J.L.M., Moura A.S.A., 2005. Reproductive responses of rabbit do to a
supplemental lighting program. Livest. Prod. Sci, 94 (2005), 179-187.
Mezali1 L, Mebkhout F, Saidj D, Merah S, Razali H, Larbi B,
Abdessalem L, 2015 : Premières données sur la cryptosporidiose
chez l'espèce Oryctolagus cuniculus domesticus en Algérie
16èmes Journées de la Recherche Cunicole, 24 et 25 novembre 2015,
Le Mans, France p48.
Moulla F et Yakhlef , 2007, Evaluation des performances de
reproduction d'une population locale de lapins en Algérie. 12èmes
Journées de la Recherche Cunicole, 27-28 novembre 2007, Le Mans, France
p45-48.
Moulla F, 2006. Evaluation des performances zootechniques de
l'élevage cunicol de la ferme
expérimentale de l'institut technique des
élevage de Baba Ali. Mémoire de magistère. Moulla F, 2008.
Evaluation de productivité de la lapine locale algérienne.
Institut national de
la recherche agronomique Algérie. Recherche agronomique
N°21-2008. P 72-77.
N
Nolan M.J., Jex A.R., Haydon S.R., Stevens M.A., Gasser R.B.,
2010. Molecular detection of Cryptosporidium cuniculus in rabbits in Australia.
Infect. Genet. Evol., 10 : 1179-1187.
O
Orengo J, Gomez E.A, Piles M, Rafel O, Ramon J, 2004. Growth
traits in simple crossbreeding among dam and sire lines. 8th world rabbit
congress. Puebla, Mexico 7-10
Ouhayoun J, 1983. La croissance et le développement du
lapin de chair. Cuni. Scien, V(1), 1,1-15.
Ouhayoun J, 1989. La composition corporelle du lapin, facteurs
de variation. INRA, Prod. Anim, 2(3), 215-226.
49
Références
bibliographiques
Ouhayoun J, Poujardieu B, Delmas D, 1986. Etude de la croissance
et de composition corporelle des lapins au-delà de l'âge de 11
semaines. 2/composition corporelle. J.Rech. Cunic, Paris, comm 24.
Ouyed A, 2006 Performances de reproduction et de croissance des
lapins de différents types génétiques
Ouyed A, Lebas F, Lefrançois M, Rives T.J, 2007b.
Performances de croissance de lapins de races pures et de lapins croisés
en elevage assaini au Qubec. In Proc, 12éme journ. Rech. Cunicole,
INRA-ITAVI 2007 Novembre. Le Mans, France, 149-152.
Ozimba C.E et Lukefahr S.D,1991. Compraison of rabbit breed types
for posweaning litter growth, feed efficiency, and survival performance traits.
J.Anim.Sci. 1991. 69 : 34943500.
P
Papeschi C., Fichi G., et Perrucci S. 2013. Oocyst excretion
pattern of three intestinal Eimeria species in female rabbits World Rabbit Sci.
2013, 21: 77-83
Peeters J.E, 1988. Recenent advice in intestinal pathology of
rabbit and further perspectives. 4th congress of rabbit science . Budapest
(Hangray), Oct 10-14, V(03), 293-315.
Peinheiro V et gidenne T, 1999. Conséquence d'une
déficience en fibre sur les performances zootechniques du lapin en
croissance, le développement caecal et le contenu iléal en
amondon. 8éme journée de la recherche cunicole, Paris, 1999 ,
105-109.
Prud'hon M, 1970. La reproduction des lapins, la revue
d'élevage n° spécial, la production, moderne des viandes de
poulet et de lapin 47m. 103-111.
Q
Quinton et Egron, 2001. Maîtrise de la reproduction chez
la lapine. Le point vétérinaire N° 218,
août-septembre, 28-33.
Questel G ,1984, cités par Bousit D,1989 : Boussit D.,
1989. Reproduction et insémination artificielle en cuniculture. Edition
Association Française de cuniculture. 233p.
Questel G., 1984. Contribution à l'étude de la
fertilité chez le lapin domestique. Mémoire de fin de formation,
INRA Paris-Grignon, France, 65 p
50
Références
bibliographiques
R
Rastogi P.K, 2001. Performances des lapins et leurs laperaux au
sein d une population cunicole reduite elevée en milieu sub-tropical a
trinad ( entilles) rev world rabbit science, 9(2), 65-68
Rashwan, A.A., Marai, I.F.M. 2000. Mortality in young rabbits.
World Rabbit Science, 8(3), 111-124.
Remas K., 2001. Caractéristiques zootechniques et hormones
sexuelles chez les populations locales du lapin domestique Oryctolagus
Cuniculus. Thèse de Magister, Ecole Nationale Supérieure
Vétérinaire, 89p.
Robinson G et Chalmers R.M., 2009. The European Rabbit
(Oryctolagus cuniculus), a source of zoonotic cryptosporidiosis. Zoonoses
Public Health, 57 : 7-8.
Rommers J.M., Meijerhof R., Noordhuizen J.P.T.M., Kemp B. 2001.
Effect of different feeding levels during rearing and age at first insemination
on body development, body composition, and puberty characteristics of rabbit
does. World Rabbit Sci., 9, 101108.
Roiron A, 1991. Vers une meilleure connaissance de la
composition anatomique des lapins. Cuniculture, 18(3), 147-149.
Rinaldo D., 1986. Composantes et facteurs de variation de la
carrière des femelles
reproductrices : Application au lapin. DEA de physiologie
animale. Université de Rennes I. 90p.
S
Samoggia R , 1987. Cité Benrais et Chibani, 2004. Esigen
ji sioclimatiche dei conigli nellallevamento in tensivo. Coniglicoltura, 24,
(5), 20-24.
Saidj D., 2006. Performances de reproduction et
paramètres génétiques d'une lignée maternelle d'une
population de lapin local sélectionné en G0. Mémoire de
Magister en médecine vétérinaire, Option : Zootechnie,
Ecole Nationale Supérieure Vétérinaire, 106p.
51
Références
bibliographiques
T
Theau-Clément M., Poujardieu B., Bellereaud J., 1990a.
Influence des traitements lumineux, modes de reproduction et état
physiologiques sur la productivité des lapines multipares. 5émes
Journées de la Recherche Cunicole, 12-13 Décembre, Paris
(France)
Theau-Clément M., Bolet G., Roustan A., Mercier P., 1990b.
Comparaison de différents modes d'induction de l'ovulation chez les
lapines multipares en relation avec leur stade physiologique et la
réceptivité au moment à la mise à la reproduction.
5émes Journées de la Recherche Cunicole. Paris, comm N°6.
Theau-Clément M., Roustan A., 1992. A study on
relationships between receptivity and
lactation in the doe, and their influence on reproductive
performance, 5th World Rabbit Congress, Corvallis, USA, 1992, pp. 55-62.
Theau-Clément M., Poujardieu B., 1994. Influence du mode
de reproduction, de la réceptivité et du stade physiologique sur
les composantes de la taille de portée des lapines. 6émes
Journées de la Recherche Cunicole, 6-7 Décembre, La Rochelle,
France, 1,187-194.
Theau-Clement M et Mercier P, 2004 : Influence of lighting
programs on the productivity or rabbit does of two genetic types. 8th world
rabbit congress . Puebla, Mexico, 358-365.
Theau-Clement M,2003. Advence in bio stimulation methods applieds
to rabbit reproduction. Cours approfondi. Système de reproduction de
viande de lapin. Saragosse 2003. Ciheam. P11
Tudela F et Lebas F, 2006. Modalité du rationnement des
lapins en engraissement : effet du mode de distribution de la ration
quotidienne sur la vitesse de croissance. Cuniculture, magasine, V(33), p,
21-27
Thierry Gidenne, 2015 : Quelques bases sur la croissance et la
qualité de la viande de lapin INRA Toulouse, UMR Tandem,
INRA-ENSAT-ENVT
V
Verdelhan S., Bourdillon A., David J.J., Hurtaud J., Ledan L.,
Renouf B., Roulleau X. Salaun J.M., 2005. Comparaison de deux programmes
alimentaires pour la préparation des futures reproductrices.
11émes Journées de la Recherche Cunicole, 29-30 novembre 2005,
Paris, 119-122.
52
Références
bibliographiques
W
Weisseman D, Troislouches G, Picard E, Davoust C, Leroux C,
Launay C, 2009. Amélioration de l'indice de consomation de lapin en
engraissement par une distribution nocturne de l'aliment. 13éme
journée de la recherche cunicole, 17-18 Novembre 2009, Le Mans,
France
Y
Yamani K.A.O, Dader A.H, Asckar A.A,1991. Non-genetic factor
affecting rabbit production in egypte. Option mediterrannéennes
N°17, 173-178.
Z
Zerrouki N Berchiche M, Bolet G Lebas F. 2001
Caractérisation d'une population locale de
lapins en Algérie : Performances de reproduction des
femelles. 9èmes Journ. Rech. Cunicole Paris, 2001
Zerrouki N., Kadi S.A., Berchiche M., Bolet G., 2005b. Evaluation
de la productivité des
lapines d'une population locale Algérienne, en station
expérimentale et dans des élevages. 11émes Journées
de la Recherche Cunicole. 29-30 Novembre, Paris, 11-14.
Zerrouki N., Bolet G., Berchiche M., Lebas F., 2005a. Evaluation
of breeding performance of local Algerian rabbit population raised in the
Tizi-Ouzou area (Kabylia). World Rabbit Science, 2005, 13: 29-37.
| 

