Etudes bibliographique de la génétique de résistance à drechslera teres( Télécharger le fichier original )par Salim DIFALLAH Université Mohamed Boudiaf de M'sila Algérie - Diplome d' études supérieures 2009 |

ÉÜÜÜÜÜÜÜÜÜÜÜÜÜÜÜÜÜÜÜÜÜ?ÈÚÔáÇ
ÉÜÜ?ØÇÑÞã?ÏáÇ
ÉÜÜÜÜÜÜÜÜ?ÑÆÇÒÌáÇ
ÉÜÜÜÜÜÜÜÜÜÜÜÜÜÜÜÜÜÜÜ?ÑæãÌáÇ MEMOIRE Présenté Le diplôme des études supérieures en Biologie (DES) OPTION : MICROBIOLOGIE THEME
Encadré (e)(s) par : Remerciements Nous remercions tout d'abord dieu tout puissant de nous avoir donné la force et la connaissance pour accomplir une action qui lui plaise. Dans le cadre de ce mémoire de recherche, nous tenons à remercier, profondément, notre directrice de recherche « Mme BENSEMANE L » pour la qualité d'encadrement, la rigueur scientifique et le soutien affectif dont nous sommes bénéficiés tout au long de la période d'élaboration de ce mémoire. Notre sincères remerciements s'adressent aussi à l'ensemble des enseignants de département de biologie pour la qualité de formation et d'encadrement dont nous avons bénéficiés tout au long de notre études. A tous ceux et à toutes celles dont les acronymes n'apparaissent pas sur cette page, ils sont nombreux, qu'ils demeurent convaincus, que nous ne les avons point oublié et qu'ils soient assurés de notre profonde gratitude. Merci. Dédicaces A Allah la miséricorde divine, le tout puissant de m'avoir donné la volonté et la capacité de présenter ce travail et à son prophète Mohamed (PSL). A ma très chère mère, dont aucun mot n'est assez fort et suffisant pour exprimer l'amour que je lui voue, elle que je dois tout et qui sera toujours pour moi un exemple de réussite et de courage, je lui témoigne mon affection profonde en reconnaissance de tout ce qu'elle a fait pour moi. pour l'ame pure de mon père que dieu le blesse A tous mon cher frère Said et mes soeurs pour le soutien et l'estime qu'ils portent à ma modeste personne Pour les enfants Bochra, Insaf, Baraa, Bahaa ... A toutes les familles: DIFALLAH, SERAICHE, A mes amis : Omar, samir, Mohamed, Abdelmalek, Taib, Hicham, Abdrrahim, Abdrahman, Said, Farid, Fayçal, Hamza, Raouf, Adel, Mohcin, Smail, Saleh, fateh, Malek Elaid, Salim, djamal, Fowaz, Jalal, Abdellatif, Ali, Belkacem, Youcef, Brahim, Fares, Issam, Nabil, ... A mes camarades de promotion avec qui j'ai partagé les moments joyeux de la réussite et à qui je souhaite un avenir radieux A tous ceux que j'aime et m'aime. Pour mes collègues dans ce recherche Fatima et Assia.
Salim Liste des figures Figure no 01: description de l'orge A- schéma descriptife de l'orge 3 B- orge a 2 rangs et 6 rangs 3 Figure no 02 : les symptômes de l'helminthosporium sur des feuilles de d'orge 9 Figure no 03 : voie de transmission de maladie et l'évolution de l'épidémie 10 Figure no 04 : Conidiophore et spore de helminthosporium teres 14 Figure no 05: les conidies de helminthosporium teres 15 Figure no 06 : cycle de développement de l'agent pathogène 16 Figure no 07: les cinq approches de la protection des cultures 20 Liste des tableaux Tableau no 01: Quelques variétés d'orge cultivées en Algérie et leurs caractères 4 Tableau no 02: liste des
helminthosporium avec leur synonyme et leur forme sexué
11 de l'un de groupes du Helminthosporium Link 12 Liste des abréviations JNO : jaunisse nanaisant de l'orge. BYDV : barley yellow dwarf virus. VMJO : virus de la mosaïque jaune de l'orge. VMMO : virus de la mosaïque modérée de l'orge . CTPS : comité technique permanent de la sélèction . BYMV : bean yellow mosaic virus . BMML : Bach Musicology Mailing List . OGM :Orca Green Marine . SOMMAIRE
4.1. La structure génétique de l'orge 17 4.2. Virulence du champignon 17 4.3. La resistance chez l'orge (hordeum vulgar L ) 18 4.4. La résistance aux maladies 19 4.4.1 Gène de résistance au BYMV et BMML 19 4.4.2 Résistance aux maladies 19 5. La protection des plantes 20 5.1. Lutte biologique 20 5.2. Une autres Moyens de lutte 21 III . Conclusion 23 IV. REFERENCES BIBIOGRAPHIQUEI. Introduction En Algérie, l'orge est la 2eme céréale cultivée aprés le blé (Anonyme 2007). L'orge occupe avec le blé dur 80% de la surface ensemencée en céréales chaque année ( Bouzerzour, Benmahamed 1995). Elle reste un pays importateur de toutes les céréales malgré la place importante qu'occupe ces dernières, de fait qu'elles servent de base à l'alimentation humaine. Selon Catton 2000, ces importations sont passé de 4.9 millions de tonnes (moyenne compagnes 1978/1979/1980) à 15 millions de tonnes (moyenne compagnes 1995/1996;1996/1997;1997/1998 ) soit plus de 220% Cette insuffisance se trouve maintenant confrontée à deux problèmes essentiels : - une production céréalière insuffisante dûe particulièrement à la faiblesse des rendements. - une démographie galopante. Dans le cas particulier de l'orge, étant donné que les variétés locales (Saïda et Tichedrett ) ont un potentiel de production assez limité, ceci a conduit à l'importation de nouvelles variétés à haut rendement en vue d'améliorer la production et d'en intensifier l'espèce, (Abdelgnerfi, Rahal 2003) Parmi les facteurs qui accentuent la faiblesse et la limitation des rendements et de la production céréalière, les maladies representent un facteur important. En Algérie, malheureusement, les maladies ne sont pas considérées comme importantes du point de vue économique, donc elles sont placées au second plan. Les pertes annuelles, dans le monde, causées par les maladies sont de l'ordre de 135 millions de tonne (Benbelkacem 2004). Mais dans notre pays, elles n'ont jamais été évaluées. Parmi les maladies qui touchent l'orge, la rayure reticulée causée par Drechslera teres synonyme de Helminthosporium teres. L'agent causal, pyrenophora teres transmis par les semences et persiste dans les résidus de culture. Le champignon se conserve localement sur les pailles pendant environ 2 ans sous forme de clamidospores et de périthèces. Les conidies formées à partir des chlamydospora à l'automne sont responsables des attaques primaires sur les feuilles. Il se conserve également dans les semences sous forme de mycélium, les semences pourront donc jouer un rôle fondamental dans l'extention de la maladie dans des nouvelles zones de production. Les objectifs de ce travail est : - Description de cette maladie. - Connaître la virulence de pyrenop ho ra teres sur les orges. - Trouver des solutions autres que la lutte chimique . II. ANALYSE BIBLIOGRAPHIQUE : 1. Généralité sur l'orge : Orge est le nom commun des plantes du genre Hordeum, de la famille des GRAMINÉES. Il existe plusieurs espèces sauvages et cultivées. Étant donné que toutes les variétés cultivées et certaines des espèces sauvages sont capables de pollinisation croisée, on suppose qu'elles appartiennent à l'espèce Hordeum vulgare. L'orge, qui s'accommode q'une grande variété de sols et de climats, est cultivée dans plusieurs régions du monde. Elle est, d'ailleurs, l'une des premières céréales à l'être . Quoique l'on ne connaisse pas son lieu d'origine, l'orge est récoltée depuis des milliers d'années au Proche-Orient, en Extrême-Orient et dans le Nord et le Centre-Est de l'Afrique .
Figure no 01 : description de l'orge A- Schéma descriptif de l'orge. B- Orge à 2 rangs ,6 rangs Selon les variétés, l'orge peut être semée en hiver ou au printemps. Les orges de printemps: sont sensibles au gel et ont un cycle végétatif plus court. Elles se sèment en février-mars. La récolte s'effectue en été. Les orges d'hiver : se sèment fin septembre - début octobre. Après avoir passées l'hiver sous terre, elles sont récoltées juste avant les orges de printemps. Elles peuvent supporter des températures allant jusqu'à -15°C. Bon à savoir : Il existe deux types d'orges d'hiver : - Hordeum. Vulgare distichum celles à épis plats à 2 rangs de graines . - Hordeum. Vulgare hescastichum celles à épis cylindriques à 6 rangs de graines. Ces variétés d'hiver à 6 rangs sont communément appelées escourgeon. (Anonyme 2008). 1.1. Les principales variétés d'orge cultivées en Algerie : L'orge est cultivée en Algérie là où le blé ne peut donner de bon rendement, c'est-à-dire dans les zones semi-arides. Elle occupe les moins bonnes terres, parmi celles réservées aux blés. Comme on peut trouver dans les zones marginales à sol plus ou moins pauvres et cela grace à sa rusticité (Oufroukh et Hamadi 1988; Khaldoune 1989). Les variétés d'orge cultivées en Algérie sont indiquées dans le tableau 1. Tableau 1: Quelques variétés d'orge cultivées en Algérie et leurs caractères .
(Anonyme 2006) 1.2. Les principales maladies qui attaquent l'orge : 1.2.1. Les maladies bacteriennes : Pseudomonas : est un genre pathogène pour les céréales tel que le blé, l'orge, seigle et triticale. la maladie de porriture de la base des glumes est provoqué par pseudomonas syringea pathovare atrofaciens et l'apparaition de nécrose bactérienne causé par pseudomonas syringea pathovare syringea. xanthomonas: ce genre attaque l'orge et le blé et provoque des rayures bactériennes par l'agent causal xanthomonas campestris PV translicens (Soltner, 1995. Cavelier, 1992). Clavibacter : cause la mosaïque bacteriènne du blé et de l'orge dont l'agent causal est clavibacter michiganensis sub espéce sptesselerius (Boukhris 1992 ) 1.2.2. les maladies virales : Les maladies les plus importantes en incidence et en sévérité sur l'orge sont : La jaunisse nanisante de l'orge (JNO) est une maladie virale transmise par les pucerons bien connue des agriculteurs. En piquant la plante pour se nourrir, le puceron transmet le virus BYDV (barley yellow dwarf virus) aux plantes. L'importance de la maladie dépend de plusieurs facteurs, la douceur du climat, la précocité de l'attaque et la durée de présence des pucerons dans la parcelle, mais aussi du nombre de pucerons ailés et de leur pouvoir infectieux. La présence de repousses de graminées ou de parcelles de maïs à proximité des parcelles d'orge, aggrave également les risques d'apparition de la maladie. Lorsque l'orge est attaquée par la JNO, les dernières feuilles prennent une couleur rouge lie de vin et les plantes sont rabougries par foyer .( Amandine L, 2003) Les mosaïques sont aussi des maladies virales transmises par un vecteur, non pas par un ravageur aérien cette fois mais par le Polymixa graminis, une « plasmodiophorale », parasite des racines des céréales, très voisin des champignons de sol. Deux types de mosaïques s'attaquent à l'orge d'hiver, la mosaïque jaune, VMJO et la mosaïque modérée, VMMO. Malheureusement, il n'existe jusqu'à présent aucun moyen de lutte contre le vecteur des mosaïques, le Polymixa, ni contre les virus eux-mêmes. La seule solution efficace et disponible actuellement, pour éviter le développement de ces maladies virales, est d'avoir recours à des variétés résistantes. C'est grâce à la sélection de variétés résistantes à la fois à la mosaïque jaune et à la mosaïque modérée, que les agriculteurs peuvent continuer à cultiver aujourd'hui de l'orge d'hiver dont les parcelles touchées par ces deux maladies virales. (Amandine L. 2003) 1.2.3. Les maladies fongiques : La rhynchosporiose (Rhynchosporium secalis) La rhynchosporiose apparaît souvent en foyers. Elle se voit sur les limbes et à l'insertion des feuilles. En fin de cycle, le centre des tâches se dessèche et devient blanc. Les effets « pulvérisation » sur le feuillage sont assez nets courbures des feuilles, nombreux impacts de gouttes, etc. (Anonyme 2001) La strie foliaire ( Pyrenophra graminea). Pyrenop ho ra graminea (anamorphe Drechslera graminea) est transmis par les semences. Des lignes longitudinales jaunes, devenant brunes, apparaissent sur les feuilles des plantes infectées. L'épiaison est médiocre et les grains sont mal remplis. Il s'agit de la maladie transmise par les semences la plus importante de l'orge. Elle attaque les cultures d'hiver et de printemps. L'oïdium (Erysiphe gramini). Erysiphe graminis forme des plages de mycélium superficiel blanc puis gris sur les feuilles, les gaines et les épis d'orge. Les feuilles restent vertes et actives pendant un certain temps après l'infection, puis les zones infectées meurent progressivement. Les conidies sont formées en grand nombre et se présentent sous forme de poudre blanche à la surface du mycélium. Elles sont dispersées par le vent à des distances considérables et infectent les feuilles saines. L'oïdium est donc favorisé par l'alternance de conditions humides et sèches. (Anonyme ,1997) La ramulariose (Ramularia collo-cygni) Les tâches de la ramulariose sont visibles sur les 2 côtés de la feuille. La ramulariose a été identifiée pour la 1 ère fois en France en 2002. Les tâches de la ramulariose se présentent comme de « mini-tâches » d'helminthosporiose, mais elles sont plus claires, marrons, rectangulaires et suivent les nervures. (Anonyme 2001) Le charbon (Ustilago hordei , Ustilago nuda ). Ustilago hordei cause le charbon couvert de l'orge. Les plantules sont contaminées de manière systémique par des spores portées à l'extérieur des semences ou persistant dans le sol. Peu après la floraison, les talles contaminées produisent des épis infectés. Les plantes malades sont rabougries et les grains sont remplis d'une masse de spores noires enveloppées par une membrane transparente jusqu'à la récolte. Ustilago nuda provoque le charbon nu de l'orge. L'infection est transmise par les semences. Le champignon pénètre l'endosperme au cours de la formation du grain. Les semences infectées produisent des plantes qui sont infectées de manière systémique. Les épis malades sont visibles juste après l'épiaison. Des spores noires sont libérées entre les glumes et les grains, et produisent une poussière noire libre (charbon nu). Elles sont transportées par le vent vers des épis sains qu'elles infectent. (Anonyme ; 1997) Ergot L'ergot est une maladie des plantes provoquée par un champignon, Claviceps purpurea, qui infecté les grains en développement des céréales et des graminées. Les symptômes de la maladie se manifestent au stade de gonflement du grain, lorsque l'ergot proprement dit, aussi appellé sclérote, pousse dans l'épi à la place des grains. Les sclérotes sont formés d'un corps dur issu du mycélium du champignon. Leur capacité à survivre aux conditions hivernales leur permet de compléter leur cycle de reproduction. L'enveloppe protectrice des sclérotes est dure et de couleur brun-noire violacée, et l'intérieur varie du blanc au gris. Les sclérotes ont souvent une forme allongée et dépassent des glumes dans les épis approchant le stade de maturité. Leur taille peut être jusqu'à 10 fois plus grande que celle des grains qu'ils remplacent. (Pearse, Penny 1999). Les septorioses La Septoriose provoquée par Septoria nodorum, est devenue anecdotique et bon nombre de variétés actuelles présentent un bon niveau de résistance. Par contre, depuis la fin des années 80, Septoria tritici est devenue une maladie grave et motive les sélectionneurs qui ont amorcé des programmes spécifiques.La tolérance à cette maladie n'est évaluée que depuis trois ans au CTPS et, manifestement, il existe de la variabilité génétique vis-à-vis de cette maladie. La fusariose La fusariose est l'une des maladies les plus répandues des petites céréales provoquée par fusarium graminearum où les symptômes sont décelables peu après la floraison. Des épillets atteints (glumes et fleurons) semblent avoir mûri (blanche) prématurément par comparaison aux épis sains qui sont verts. Le champignon peut s'attaquer à la totalité ou à une partie de l'épi. Le blanchiment des épis où la fusariose apparaît de 3 à 5 jours après l'infection. (Anonyme 2002). 1.2.4. Les maladies d'origine animale: Un grande nombre d'espéces animales pouvant occasionner des déprédations sur la culture d'orge, mais seulement un petit nombre d'entre elles presente une importance économique; parmi ces espèces, on peut citer : Les vers blancs, les pucerons, les mouches grises , la mineuse d'orge et les oiseaux (Oufroukh et Hamadi ,1988). Parmi toutes les maladies citées ci-dessus, notre etude s'intéresse en particulier à l'helminthosporiose teres. 1.3. Objectifs et critéres de sélèction de l'orge Quatre principaux objectifs sont recherchés dans la sélection de l'orge productivité et stabilité du rendement : L'elaboration du rendement implique l'enchaînement de multiples mécanismes liés à la croissance et au développement des peuplements végétaux en relation avec les facteurs et conditions du milieu ( Picard, 1994 ) Résistance à la verse : Selon Moule (1980), l'orge est, des trois céréales (blé, orge, avoine), la plus sensible à cet accident qui est le facteur limitant principal de la fumure. Cependant selon les régions, la résistance à divers stress doit être considérée (sécheresse, salinité du sol) (Jestin, 1992). En effet l'adaptabilité au milieu est un phénomène essentiel en raison des relations entre conditions de culture et expression des potentialités génétique (Combe et Picard, 1994 ) Resistance aux maladies : la maitrise des maladies par les traitements chimiques n'est pas sans inconvénients (Simon, 1989), l'objectif est de rechercher des génotypes qui présentent avec l'agent pathogène un rapport d'incompatibilité ( Seilleur, 1989) 2.description et symptomatologie de L'helminthosporiose de l'orge 2.1. description Tâche de Pyrenop ho ra teres. Drechs. Smedeg. est une maladie importante et économiquement de l'orge (Hordeum vulgare) dans le monde et en particulier dans les régions tempérées et humides, des zones de production dans l'Europe du Nord et les pays méditerranéens (Shipton, et al 1973) . La maladie provoque des pertes de rendement jusqu'à 50% et réduit la qualité du grain (Mahmoud M. Saker). L'helminthosporiose est la maladie la plus préjudiciable au rendement de l'orge causée par le champignon pyrenophora ( ou drechslera = helminthosporium) teres affecte les cultures d'orge en zones humides, où la culture revient souvent sur elle même. En fonction de la sensibilité des variétés, les pertes de rendement atteignent couramment 10 à 30 %. Les lésions sur les feuilles dues à pyrenophora teres se manifestent sous deux formes bien distinctes : - des symptômes en réseau brun foncé, diffus sur un fond chlorosé brun clair . - des tâches brunes ovoïdes de 3×5 mm environ, entourées d'un halo chlorosé, ou a bords paralléles entre 2 nervures avec en haut et en bas de la tâche un point de chlorose plus clair, la maladie s'etend a partir de semences infectées ou de résidus de récolte (Shipton et al 1973) Le champignon survit après la récolte sur les débris de paille mal incorporés ou les repousses d'orge. Une contamination primaire peut se produire à partir des ascospores produites sur les résidus de récolte. Les conidiophores, qui se développent ensuite peuvent être transportées par le vent dans les parcelles éloignées et provoquer des contaminations secondaires. Les semences infectées peuvent aussi être à l'origine de contaminations primaires. Le champignon gagne les différents étages foliaires en progressant du bas vers le haut.
Figure no 02 : symptôme de l'helminthosporium sur feuilles d'orge . 2.2. les sources de contamination L'inoculum de départ a plusieurs origines, qui sont les suivantes : - la semence mère qui était contaminée avant sa mise en terre (Champion, 1997), Le champignon survit sur les pailles laissées sur place (Yves Morvan, 2006 ). - Résidus de moissons infestés et insuffisamment incorporés au sol, repousses d'orge ou semences infestées dès le départ. - La succession d'orge d'hiver sur orge entraîne un danger d'infection primaire par les ascospores présentes sur les chaumes. L'infection secondaire se fait dans la parcelle ainsi que sur les parcelles éloignées par les conidiospores dispersées par le vent. Les conidiospores se forment dans des sporanges spécifiques sur les feuilles nécrosées (Yves Morvan, 2006 ). - des travaux culturaux ou de récolte qui favorisent le transport des spores et l'infection des semences (Champion, 1997).
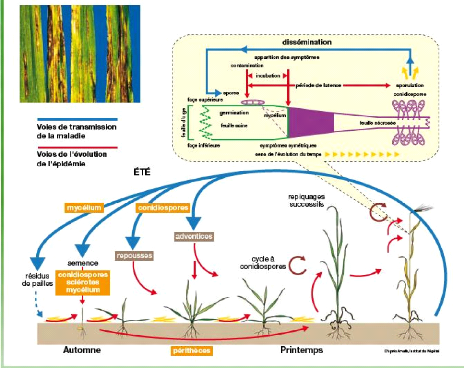
Figure no 03 : voie de transmission de la maladie et l'évolution de l'épidémie. 3. l'agent pathogène :Drechslera teres L'helminthosporiose de l'orge est une maladie fongique causée par Drechslera teres (Sacc.) amorphe de Pyrenop ho ra teres Drechs (Bendahmane et Albertini 1992). Pyrenophora teres (anamorphe Drechslera teres) est transmis par les semences et persiste dans les résidus de culture (à partir desquels les ascospores sont libérées). Les conidies formées sur les lésions foliaires sont portées par le vent et sont à l'origine des infections secondaires (Anonyme ,1997). 3.1.Critères de classification des différents genres de l'helminthosporium : Le genre helminthosporium Link, appartenant à la famille de dématiaceae (Ellis, 1976) est un champignon filamenteux (Doctorfungs), cosmopolit, isolé de sol et de plante (Indoorpol) .Ce genre érigé en 1809, comprend plus de 500 espèces, ces dernières sont surtout des pathogènes de céréales (Rapilly,1971). D'aprés Roger en 1953, il existe trois espèces d'Helminthosporium attaquant l'orge: - Helminthosporium gramineum Rabh.: cause la strie foliaire de l'orge. - Helminthosporium teres Sac.: entraine la rayure réticulée. - Helminthosporium sativum (R.K.B): responsable de la pourriture sèche de la racine . D'autres espèces d'helminthosporium sont représentées dans le tableau ci-dessous (Tableau2). Tableau 02 : liste des helminthosporium avec leur synonyme et leur forme sexué selon Rémi Champion 1997
Nisikado en 1928 a différencié, les espèces de helminthosporium en deux groupes : - Eu-helminthosporium : ayant des conidies courbées, fusiformes et germent uniquement à partir des cellules terminales. - Cylindro-Helminthosporium: à conidies cylindriques, souvent droites, qui germent par un ou plusieurs tubes à partir de n'importe quelle cellule. Tableau 03: Critères de classification des deux groupes dont deux sous genres de l'un de groupes du Helminthosporium Link.
Berrah 2004 3.2. Position systématique La forme imparfaite ( anamorph) drechslera teres Règne : champignon Embranchement : Amastigomycota. Sous/Embranchement : deuteromycota . Classe : deuteromycètes. Ordre : hyphales Famille : dématiaceae. Genre : Drechlera. Espèce : drechslera teres . La forme parfaite pyrenophora Domain : Eukaryota Kingdom : Fungi Subkingdom : Dikarya Subphylum : Pezizomycotina Classe : Ascomycètes Subclasse : Pleosporomycetidae- Order : Pleosporales Famille : Pleosporaceae Genre : Pyrenop ho ra description Specifique : teres Nom Scientifique: Pyrenop ho ra 3.3. Description de Drechslera teres L'helminthosporium est un champignon filamenteux. Ce genre contient principalement des moules de graminées (Doctorfungs). Pyrenophora teres existe sous deux formes, Pyrenophora teres f. teres causé la forme réticulée et Pyrenophora teres f. maculata causé la forme tâchetée, présentent des symptôme différents sur les feuilles. (O.M. Manninen, M. Jalli 2006 ). 3.3.1. la forme parfaite ou stade télémorphe: Le stade parfaite du parasite est connu sous le nom: pyrenophora teres drech, ( D-E. Alihaimoud et al.,1993) A ce stade, se forment les périthèces qui englobent des asques contenant des ascospores. Ces périthèces sont rarement observés dans la nature (Zyllinsky, 1983). Les ascospores (18-28 x 43- 61. ìm) sont brun clair et ont souvent 3-4 septa transversal et un septa longitudinal dans la médiane des cellules. (Afanasenko, 2006) 3.3.2. la forme imparfaite ou stade anamorphe : au stade asexué, le parasite est sous le nom helminthosporium teres où les pycnides sont globuleux à la forme de poire, 64-172. ìM de diamètre, la poire est mince, fragile et de couleur jaune à brun. Ces pycnides sont pourvues de petites spores unicellulaires , apparaissent parfois directement sur les téguments des graines (Champion, 1996), les pycnidiospores (1.0-1.9 x 1.4-3.2. ìM) sont hyalines, non septés et sphériques. Les mycélium à ce stade peuvent aussi former des conidies produites sur la surface des lésions primaires servent d'inoculum secondaire. La période d'incubation dépend des conditions météorologiques et prend à partir de 20 heures à 20 jours. Infection sévère tue prématurément les feuilles et réduit le poids des semences. Il réduit également le nombre d'oreilles et le nombre de grains par épi. Les populations d'agents pathogènes sont très hétérogènes sur la virulence.
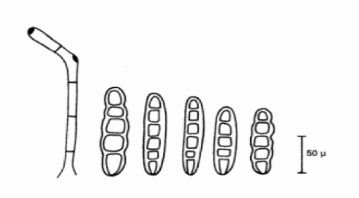
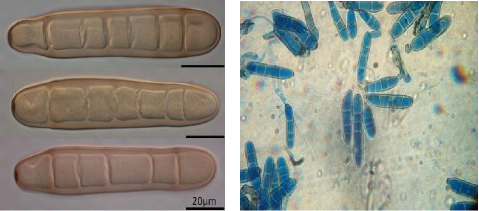
Figure no 04 :Conidiophore et spore de helminthosporium teres ( Champion 1997). Les spores sont bruns Claires et ne sont pas doré foncé comme celles d'Helminthosporium gramineum . Elles sont droites, cylindriques arrondies aux deux extrémités et présentent une légère constriction au niveau des cloisons, elles peuvent avoir jusqu'a 7 ou 8 cloisons mais en ont souvent 4 ou 5 et mesurent 60-125 × 11-20 ìM (Champion 1996).
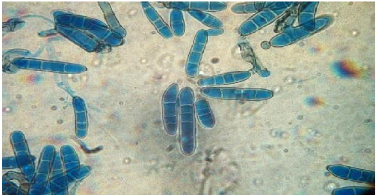
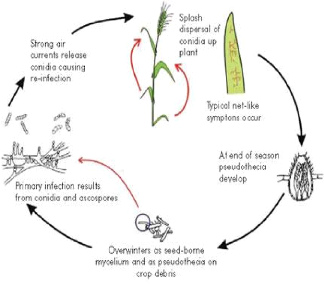
Figure no 05 : les conidies de helminthosporium teres. 3.4.Cycle de développement de l'agent pathogène Les contaminations primaires ont 3 origines : - les semences . - les repousses d'orge et certaines graminées adventices. - les débris de paille. Les deux dernières constituent la principale source d'inoculum. Le champignon se conserve localement sur les pailles pendant environ deux ans sous forme de chlamydo spores et de périthèces. Les conidies formées à partir des chlamydospores à l'automne sont responsables des attaques primaires sur feuilles. Il se conserve également dans les semences, sous forme de mycélium. Les semences pourront donc jouer un rôle fondamental dans l'extension de la maladie à de nouvelles zones. Après avoir provoqué la nécrose du tissu contaminé, le champignon produit des spores qui vont être disséminées par le vent, provoquant ainsi de nouvelles contaminations. L'enchaînement des cycles dépend fortement des conditions climatiques. Des températures se situant entre 5° et 35° C (optimum à 20° C) et une hygrométrie d'au moins 95 % constituent des conditions favorables. En conditions favorables (20° C et 100 % d'hygrométrie de l'air), le temps de latence peut se réduire à cinq jours.
. Figure no 6: cycle de developpement de l'agent pathogène. 3.5. Les facteurs contribuant au developpement de l'agent causal : Les principaux paramètres qui influencent l'évolution de la maladie sont: la température, l'humidité, la lumière et la concentration en azote dans le sol 3.5.1 Température La température et l'humidité, qui sont étroitement liée, font ensemble les principaux facteurs faisant la maladie tourner en épidimie. La plupart des pathogènes fongiques se développent bien entre 15 et 250C, si l'humidité est élevée ces températures sont energistrées pendant relativement longtemps durant l'automne et le printemps et la majorité des maladies fongiques a lieu ainsi pendant ces saisons et peut se transformer en épidimie. Durant l'hiver et l'été, la tempétrature est d'habitude trop basse ou trop élevée, pour permettre un déveleppement important des maladies. 3.5.2.L'humidité Comme la température est étroitement liée à l'humidité, celle-ci est aussi déterminante dans l'instalation du pathogéne et le développement des maladies en épidimies. L'humidité peut exister sous forme d'humidité relative dans l'air, de rosée et d'eau de pluie ou d'irrigation. ainssi leur sensibilités aux maladies, mais aussi elle favorise et accroît les activités biologiques des pathogénes comme la germination des spores, la pénétration dans la plante hôte, la sporulation, la libération des spores, etc... durant le temps ou l'humidité et de niveax élevés, ces évènements prennent place constamment, répétitivement et conduisent au développement des épidimies. Quand l'humidité exite sous forme d'éclaboussures de pluie ou d'eau courante, elle joue aussi un important rôle dans la propagule du pathogéne sur la même plante hôte et d'une plante infectée à une autre saine (Nasraoui, 2006). 3.5.3. La luminosité La reproduction est affectée par la lumière qui stimule la formation des structures reproductives et des spores. L'intensité, la durée et la qualité de la lumière sont les éléments qui déterminent l'effet général de la luminosité sur la reproduction (Nasraoui, 2006). Le nombre de conidies produites, selon Beth et al en 1975, augment avec le temps d'irradiation (lumière fluorescente aux intensité de 0.0045, 0.0075, 0.1338 cal. Cm-1S-2) et le temps d'incubation à base température. 3.5.4. La teneur en azote Toute augmentation de la concentration en azote dans le milieu de culture d'orge (sol) va favoriser l'évolution du drechslera (Raynal et al., 1989). 4. la résistance 4.1. La structure génétique de l'orge : L'orge cultivée Hordeum vulgare, est généralement une plante diploide ( 2n= 2x =14) et nettement autogame (Ramage 1960). Des formes tétraploides ( 2n =4x =28) peuvent apparaitre spontanément, ou par traitement au laboratoire, mais elle ne présente guère d'intérêt agronomique . 4.2. Virulence du Champignon Un isolat est considéré comme virulent lorsque le nombre de plants infectés par pot est superieur à 15%, mais si le nombre de ceux-ci est inférieur à 15%, le parasite est dit non virulent ou avirulent (Benbelkacem et al. 2000 ). Lors de la sélection pour la résistance à la maladie, il est important d'avoir une connaissance approfondie du niveau de variation génétique de l'agent pathogène (Zhong et al.,2003 ) 4.3. La resistance chez l'orge (hordeum vulgar L ) La résistance chez les plantes est la résistance contre les pathogènes qui est sous le contrôle d'un, ou de plusieurs gènes des plantes. (Nasraoui, 2006) La résistance aux maladies dépend de deux facteurs, l'environement, qui s'assure une résistance temporaire et le génotype de la plante, qui est une source d'élément durable à la résistance (Zahour, 1992). La génétique de la résistance varié selon les espèces végétales et le type de maladie . Dans certain cas la résistance est dominante, dans d'autre cas, elle est récessive (Anonyme, 1995) L'intéraction de la résistance des plantes et la virulence des pathogènes aboutit à une plus ou moins compatibilité entre l'un et l'autre. Deux types de résistance sont connus : résistance horizontale et résistance verticale. Les deux résistances sont généralement sous le controle de gènes logés dans les chromosomes des plantes dans le noyau cellulaire. Mais pour plusieurs plantes, cependant, la résistance est contrôlée par un matériel génétique contenu dans le cytoplasme de la cellule. Telle résistance est parfois appelée résistance cytoplasmique. Les pathogènes forment de nombreuses races, chacune d'elles différentes des autres par sa capacité d'infecter certaines variétés d'une espèce de plante mais pas d'autre espèces . La coévolution de résistance/sensibilité chez les plantes hôtes et avirulence/virulence chez les pathogènes peut être expliquée par la théorie ou le concept de gène-pour-gène suggérant qu'à chaque gène qui confère l'avirulence au pathogène correspond un gène qui confère la résistance à la plante et vice versa . Pour avoir une résistance contre toutes les races d'un pathogène, il est recommandé de combiner différents gène de résistance dans sa plante hôte. Chaque gène de résistance confère à la plante une résistance à toutes les races du pathogène qui contiennent le gène correspondant à l'avirulence. De cette façon, une espèce peut contenir plusieurs gènes contre un pathogène particulier, tel que le blé et l'orge qui ont 20 à 40 gènes de résistance contre leurs pathogènes fongiques (. Nasraoui, 2006) Les travaux de H.-L. Wu, B.J. Steffenson, et autres mettent en evidence la variation génétique de virulence de pyrenophora teres. Cette étude a été entreprise afin d'évaluer la variation génétique dans une petite collection d'isolats de Pyrenophora teres. Les phénotypes de virulence ont été déterminés pour des isolats issus de conidies uniques à l'aide de 25 génotypes différentiels d'orge. 15 pathotypes ont été identifiés dans une collection de 23 isolats de Pyrenophora teres. f. teres, et 4 pathotypes, dans une collection de 8 isolats de Pyrenophora teres. f. maculata. En général, les isolats de Pyrenophora. teres. f. teres avaient une gamme plus large et un niveau de virulence plus élevé sur les hôtes différentiels que les isolats de Pyrenophora teres. t. f. maculata. Huit génotypes d'orge étaient résistants aux 19 pathotypes identifiés et devraient être utiles en amélioration génétique de l'orge pour la résistance aux deux formes de Pyrenop ho ra teres. Selon O.M. Manninen et al, en 2006 , Plusieurs lignées d'orge affichant une résistance à la rayure réticulée conférée par des gènes majeurs ont été identifiées . les auteurs ont employé la même population pour cartographier la résistance à quatre isolats du type tâcheté et à quatre isolats du type réticulé du Pyrenop ho ra teres. 4.4. La résistance aux maladies: 4.4.1 Gène de résistance au BYMV et BMML Ces virus sont transmis par un champignon du sol polymysca gaminis, huit gènes de résistance au VMJO localisé sur les chromosomes 1 à4 ont été décrits. Ces gènes dont deux d'entre eux (ym1, ym2) sont dominant, confèrent une immunité à ce virus. Cinq de ces gènes protègent aussi les plantes viv à vis du VMJO2. La résistance au VMMO est assuré par des meme gène exeptés ym3 et ym10.(Djabbar Hariri ,1998) 4.4.2. la résistance aux maladies La résistance aux maladies est travaillée par les sélectionneurs depuis très longtemps. C'est en effet un axe de sélection majeur car une trop grande sensibilité à une maladie donnée peut avoir des conséquences catastrophiques l'année où la maladie est présente. La lutte génétique contre les maladies est souvent un travail de longue haleine et parfois un éternel recommencement ... Les maladies peuvent en effet, dans certains cas, contourner les résistances et des variétés connues pour être tolérantes à une maladie donnée deviennent sensibles. Le sélectionneur doit alors entreprendre un nouveau travail de sélection pour obtenir des résistances à la nouvelle souche de maladie.
cible 5. La protection des plantes : La protection des plantes peut être schématisée comme suit :
Figure no 07 : les cinq approches de la protection des cultures . (charles V, et al ,2000). 5.1. Lutte biologique Le terme "lutte biologique" recouvre différents concepts selon les disciplines impliquées dans la protection des cultures (Nordlund, 1996). Afin de préciser notre point de vue, nous suivrons Ferron (2000) et retiendrons la définition officielle donnée par l'Organisation Internationale de Lutte Biologique et Intégrée contre les animaux et les plantes nuisibles. La lutte biologique consiste à utiliser des organismes vivants pour prévenir ou réduire les dégâts causés par des ravageurs (Oilb-Srop, 1973). L'agent de lutte peut être un parasitoïde, un prédateur, un agent pathogène (champignon, bactérie, virus, ou protozoaire), ou un concurrent du bio-agresseur visé. On distingue trois types de lutte biologique :
Le développement d'une méthode de lutte biologique contre un ravageur nécessite quatre étapes :
5.2 Un autre Moyen de lutte qui n'existe pas sur notre schéma qui est les suivant Mesures agricoles préventives : Bien incorporer les résidus de récolte au sol. On réduira également les risques d'infection en favorisant le pourrissement de la paille, par l'élimination immédiate des repousses d'orge dans les friches et l'arrachage des repousses dans les cultures de colza. Ne pas semer orge sur orge. Ne pas semer trop tôt l'orge d'hiver. Ne pas cultiver l'orge de printemps trop près d'une culture d'orge d'hiver. Dans les principales régions à risques, employer des semences saines et désinfectées. Mesures de lutte antifongique : Pour combattre la maladie des taches brunes, on utilisera un traitement spécifique dès le début de la sporulation, à partir du stade 3-2. Le délai d'intervention avec un fongicide curatif est relativement court - un jour à un jour et demi après l'infection. Le début de la sporulation sur les étages foliaires inférieurs constitue donc le meilleur moment pour traiter. L'apparition de la sporulation permet en même temps de déterminer avec certitude l'agent pathogène en cause. 23 REFERENCES BIBIOGRAPHIQUES
net and spot blotch of barley, caused by Drechslera teres (Sacc.) Shoem , Thèse de doctorat : Sciences agronomiques , Institut national polytechnique .INPT Toulouse, FRANCE. 14. Boivin G. (2001). Parasitoïdes et lutte biologique : paradigme ou panacée ? Vertigo, 2(2) 15.Boukhris L. 1992: Identification des bactéries phytopathogènes transmissibles par la semence du blé en algérie. Mémoire Ing. Agro. Département de botanique, INA El harrach, 73 p.
(hordeum vulgare L.) et leur sensibilité vis-à-vis de drechslera graminea Rab.Mém. Ing Agro.INA.El-harrach.p59.
Contribution à l'étude de l'activité toxique de l'Helminthosporium teres, parasite de l'orge (Hordeum vulgare). Net blotch in semi arid regions of Morocco. I Epidemiology . 27.H.-L. Wu, B.J. Steffenson1,2, and S. Zhong. Genetic variation for virulence and RFLP markers in Pyrenophora teres. Department of Plant Pathology, North Dakota State University, Fargo, ND 58105, U.S.A. 28.Jestin L 1992: L'orge, amélioration des espèces végétales cultivées : objectifs et critères de selection. Ed. INRA, Paris pp 5-70.
34.Pearse, Penny. 1999 : L'ergot des céréales et des graminées Agriculture et Alimentation Saskatchewan , Commission canadienne des grains.
41.Y. Li and A.E. Oleson. Department of Biochemistry, North Dakota State University, Fargo, ND 58 105, U.S.A. Department of Plant Pathology, University of Minnesota, St. Paul, MN 55108, U.S.A. Accepted 10 October 2002. 42.Yves Morvan 2006, Les maladies fongiques de l'orge - Helminthosporiose de l'orge (Drechslera teres = Helminthosporium teres = Pyrenophora teres) Responsable technique et affaires réglementaires
A B
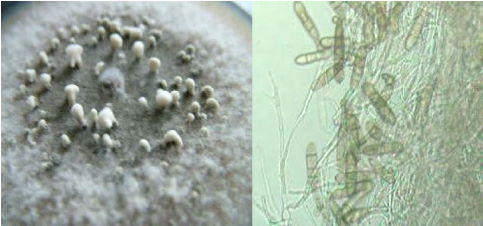
C D FIGURE 01: A : Les symptômes causés par Pyrenophora teres sur l'orge , B : sur les feuilles de blé . C et D : symptômes de l'helminthosporiose sur l'orge
A B
C Figure 2: A: Colonies de Pyrenophora teres . B: des conidies sur la plaque de gélose PDA C-D : les conidies de pyrenophora teres. Tableau :Les toxine produites par le genre drechslera
(benslimane,2003) Lexique Anamorphe : d'une forme différente de l'organisme qui lui a donné naissance. Anecdotique : recit rapide d'un evènement curieux ou méconnu (sous estimé) synonyme : secondaire, insignifiant. Asque : cellule en forme de sac, contenant gnééralement 8 ascospores, caractérisant les Ascomycètes et issue de la reproduction sexuée. Autogame : est une autofécondation de la plante : les deux gamètes sont issus du même individu. Conidiophore : filament différencié d'un champignon, spécialisé dans la production de conidies Diploïde : qui possède normalement un double assortiment de chromosomes semblables (nu). Génotype :Ensemble des constituants génétiques d'un organisme, qu'ils soient exprimés ou non. Nécrose : mortification tissulaire ou cellulaire se produisant au niveau d'un tissu, d'un organe, d'une région anatomique alors que le reste de l'organisme continue à vivre. Pathotype : fraction de la population pathogène qui préente le méme profil de virulences sur une gamme différentielle d'hotes donnée. Périthèce : le périthèce est la fructification, en forme de bouteille, renfermant les asques. C'est un organe formé après la rencontre de deux filaments mycéliens et dans lequel se développent les asques Pycnide : Les pycnides sont des structures reproductrices présentes chez certains champignons, Elles sont des structures haploïdes issues d'une mitose à partir d'hyphes végétatifs haploïdes. Résistance : capacité d'un organisme à surmonter, totalement ou suivant différents degrés, les effets d'un pathogène ou d'autres facteurs préjudiciables. Sclérote : organe asexué de conservation d'un champignon, formé de l'agrégation de filaments mycéliens riches en réserves, le plus souvent entouré d'une paroi épaisse et dure, fortement mélanisée. Téléomorphe : forme parfaite de la reproduction sexuée des Eumycètes (voir "parfait"). Tolérance : aptitude à supporter un produit, une maladie...
. ????Íæ ÉãæÇÞã áÇ?Ì ?????? ÉÓÇÓÍáÇæ ÉãæÇÞãáÇ ÇäÕ?Ç ??? ?????? Ç ä . ãæÇÞã ÓÇÓÍ Ñ?ÚÔáÇ ÇäÕ ÉÏÍ pyrenop ho ra teres íßÈÔáÇ ÚÞÈÊáÇ : ÍÇÊãáÇ ÊÇãáßáÇ |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||