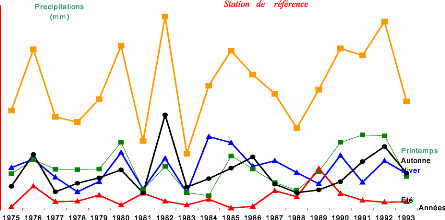
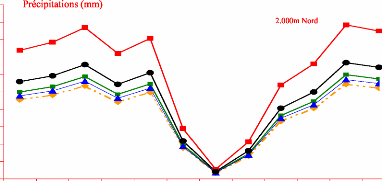
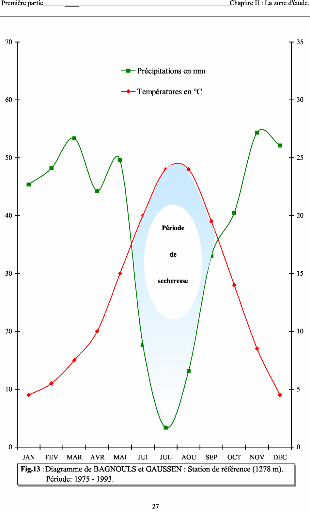
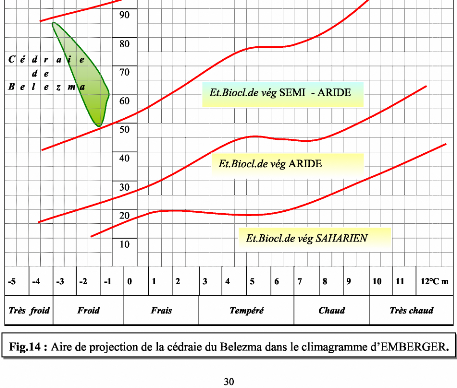
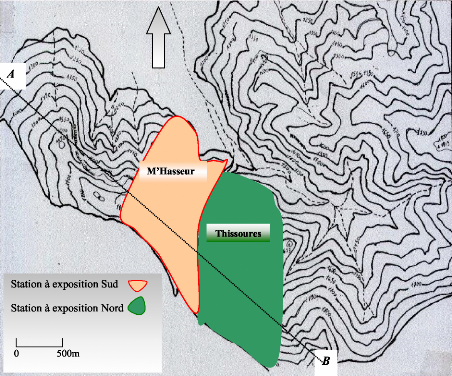
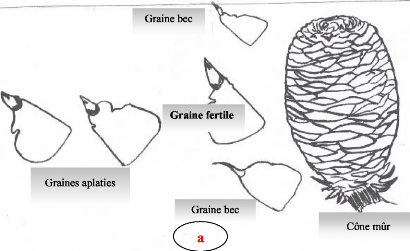
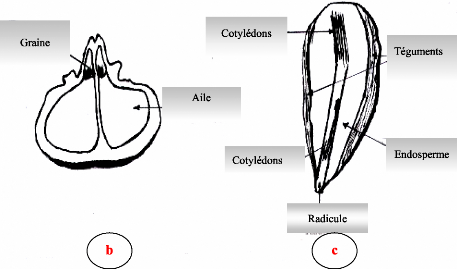
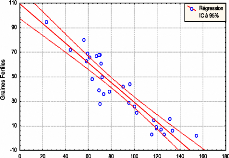
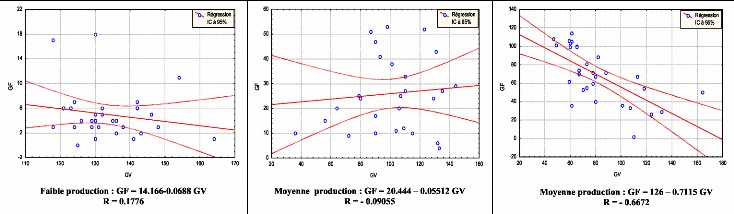
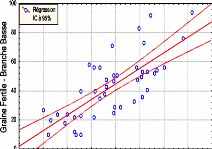
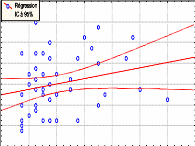
Contribution a l'étude de la fructification et de la régénaration du Cèdre de l'Atlas( Télécharger le fichier original )par KHANFOUCI Med Said Université hadj lakhdar-batna - magister en science forestière 2005 |

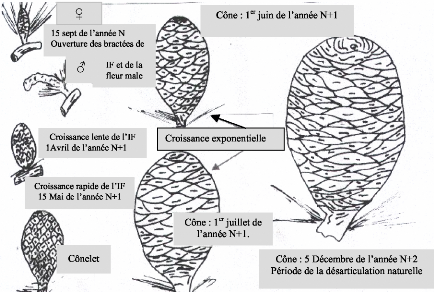
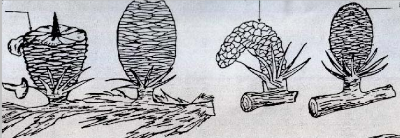
Fig. 18 : Diagramme phénologique du cèdre de l'Atlas (Cedrus atlantica M) au Belezma. 49 3.1.1 - L'année N : C'est l'année de la floraison et de la pollinisation. Elle débute avec l'apparition des fleurs mâles vers la mi - Juin. Les inflorescences femelles apparaissent vers la 2ème semaine du mois d'Août. 3.1.1.1 - Les fleurs mâles : Depuis leur apparition, elles deviennent de plus en plus nombreuses vers la fm de la 3ème semaine du mois de Juin. Au début, elles se présentent sous la forme d'un petit bourgeon arrondi de quelques mm au milieu d'une rosette d'aiguilles, insérées sur rameau court. Leur développement qui était au départ lent, s'accélère et ce n'est que vers la troisième semaine du mois de Septembre, qu'elles atteignent leur taille définitive : 2 à 5 cm de long pour 0,5 à 1 cm de diamètre. Leur couleur d'un vert tendre devient alors vert foncée avec des taches jaunâtres (grains de pollen). Le chaton mâle qui à son apparition (mi-juin) était de forme arrondie et rigide devient à sa maturité cylindro - conique et souple. Il se courbe légèrement, la libération des grains de pollen (sec et poudreux) est entamée. Après pollinisation, les chatons mâles se fanent, se dessèchent et commencent à se détacher des rameaux. Leur chute débute à partir du mois de Septembre, période qui correspond à l'ouverture des bractées de la quasitotalité des chatons mâles. Par ailleurs, quelques-uns - uns demeurent au-delà du mois d'Avril. L'apparition des chatons, leur croissance et la libération des grains de pollen diffèrent selon les expositions et également d'un arbre à autre selon les expositions et au sein d'une même exposition. Ils sont plus précoces sur l'exposition Sud et les arbres isolés que l'exposition Nord et les sujets groupés. Ce décalage entre les arbres qui ne dépasse pas une semaine, serait dû à la température et la lumière. Par ailleurs, la manifestation précoce des fleurs aboutie à la production des chatons mâles les plus dimensionnés et, à une production précoce et importante des grains de pollen et donc une pollinisation optimale. 3.1.1.2 - Les inflorescences femelles :Ce sont des fleurs composées ; chacune comprend une soixantaine de fleurs et chaque fleur (écaille) est composée de deux carpelles soudés (TOTH, 1978 a; 1984). Ces dernières sont susceptibles de donner deux graines une fois la fécondation achevée. Leur apparition est beaucoup plus tardive par rapport aux fleurs mâles ; elles ne se manifestent en quantité importante qu'à partir de la 3ème semaine du mois d'Août, chez les sujets les plus précoces. Leur nombre est extrêmement faible par rapport aux chatons mâles ; toutefois, elles sont plus nombreuses sur l'exposition Sud que Nord. Elles se présentent sous la forme d'un bourgeon vert pâle au milieu d'une rosette d'aiguille. La couleur verte devient de plus en plus foncé au fur et à mesure qu'elles s'accroissent. Leur développement est extrêmement rapide par rapport aux fleurs mâles. A partir de la deuxième semaine de Septembre, elles commencent à recevoir les grains de pollen. Elles sont alors dressées, les bractées disposées en hélice autour d'un axe (rachis) permettant aux grains de pollen de se déposer donnant un aspect jaunâtre aux inflorescences. Pendant la période de pollinisation maximale, l'ouverture des écailles forme des angles pratiquement perpendiculaires au rachis. A la fin du mois de Septembre on assiste à la fermeture des écailles ; les grains de pollen ainsi emprisonnés, seront protégés à l'intérieur des inflorescences femelles pendant la période hivernale. 50 3.1.2 - L'année N+1 : 3.1.2.1 - Les fleurs mâles : Durant cette année on assiste à la chute de la quasitotalité des fleurs mâles, particulièrement durant le mois de Janvier. Quelques-unes, les plus grosses, peuvent subsister au-delà du mois d'Avril. 3.1.2.2 - Les inflorescences femelles :C'est l'année la plus importante pour la production de graines fertiles, car les principales phénophases se passent durant cette période. En effet, si au cours des deux premiers mois (Janvier et Février) les inflorescences s'accroissent lentement, elles atteignent des dimensions appréciables par rapport à celles de l'année N. Il faut noter que juste avant la fécondation, (1' Mai - 20 Mai) au moment du débourrement, on assiste à un grossissement de l'inflorescence. Cet accroissement est le résultat de la remontée de la sève (TOTH, 1982-1984). C'est pendant cette période que se produit l'attaque par Megastigmus suspectus var pinsapini (Cf. fig. 3). La femelle insère sa tarière dans le cônelet et dépose directement les oeufs dans les graines, car les écailles sont très molles. Le cèdre de l'Atlas débourre fin Avril -- début Mai selon AUSSENAC et al (1981). NEDJAHI (1988) rapporte qu'à Chréa, le débourrement se situe vers le 13 Mai pour l'année 1984 et le 22 Avril pour 1985. Sur le même site, LAIMOUCHI (1996) note que le débourrement a débuté le 6 Mai et s'est terminé le 20 Mai pour l'année 1995. Le décalage du débourrement est en rapport direct avec la température (AUSSENAC et al, 1981 ; NEDJAHI, 1988 et LAIMOUCHI, 1996) A partir du printemps, la croissance s'accélère. Ainsi, dès le mois de Mai il y a fécondation qui se traduit par une croissance exponentielle des chatons femelles. Ces dernières deviennent cônelets vers la fin du mois d'Août. Le cônelet devient cône et atteint presque sa taille définitive dès la deuxième semaine de juillet.. Cette dernière se traduit par des changements de couleur, les cônes d'une année passeront progressivement du vert au vert violet puis au marron, couleur défmitive. Elle est due selon TOTH (1978 a, 1982-1984) à l'oxydation de la résine suintante, déposée sur la face externe des écailles La variation de la couleur commence à apparaître au niveau de la partie supérieure (sommet) du cône. Les mieux dégagés, c'est à dire ceux qui reçoivent le plus de lumière, changent de couleur les premiers. Cette modification de couleur débute à partir de la dernière semaine du mois d'Août et s'achève à la fin de Septembre. Une fois que les cônes sont totalement marron, leur maturité morphologique est achevée; les graines sont totalement formées. 51
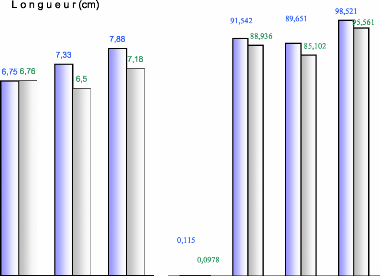
15 sept de l'année N Ouverture des bractées de Cône : 1er juin de l'année N+1 IF et de la Croissance lente de l'IF 1Avril de l'année N+1 Croissance exponentielle Croissance rapide de l'IF Cône : 1' juillet de Cône : 5 Décembre de l'année
N+2 Fig. 19 : Evolution morphologique des organes de reproduction femelles. 3.1.3 - L'année N+2 : Les cônes de l'année (N+1) continuent à se développer pendant l'année suivante (N+2) où ils seront mûrs physiologiquement durant l'Eté (Juillet - Août), soit une période de l'ordre de 10 mois. Cette maturité s'accompagne d'une déshydratation des cônes (TOTH, 1978 a et 1984).Elle se traduit par une perte de poids et rend les cônes plus secs causant souvent des dommages aux graines DOGRA (in DERRIDJ, 1990) ; les fissurations sur les écailles apparaissent à partir du sommet des cônes. Ces derniers commencent à s'ouvrir, dès que les conditions climatiques sont réunies. Elle débute lentement à partir de la mi -- Septembre puis s'accélère et se poursuivra jusqu'au mois de Novembre, on assiste alors à la désarticulation. Pour qu'il y est désarticulation, il faut donc que l'absorption de l'eau par le cône soit suivie d'un gel, puis d'une élévation de la température (dégel). En effet, durant les précipitations automnales, les cônes s'imbibent d'eau, leurs écailles deviennent souples et commencent à s'ouvrir progressivement grâce à l'amplitude thermique. Ils commencent à se désarticuler vers la mi--Novembre : les écailles se détachent du rachis qui reste solidaire à la branche pendant longtemps, en emportant les graines. Durant la dernière semaine du mois de Décembre les températures devenant très basses, l'eau qui a imbibé les cônes devient glace, les écailles s'ouvrent largement dès que les températures remontent (fonte de la neige), la désarticulation devient optimale. Elle s'achève généralement vers la fin de la première semaine de Janvier de l'année N+3. 52 TOTH (1982-1984) note que si les cônes du cèdre de l'Himalaya, s'ouvrent sous l'effet de la chaleur, ceux du cèdre de l'Atlas se désarticulent sous l'influence de l'humidité et de la chaleur. Par ailleurs, l'ouverture des cônes est précoce sur les expositions Sud. Elle est également variable selon la position des cônes dans l'arbre : cônes dégagés et ceux des branches hautes se désarticulent les premiers. Notons enfin que selon TOTH (1982-1984 et1984) le cèdre de l'Atlas possède un cycle long de 29 mois et que si les conditions ne sont pas favorables, il empiète sur la 3ème année. En outre, il faut signaler que la levée de dormance des graines est entamée avant la désarticulation et après la dissémination. (Cf. Troisième partie Chapitre I : La germination des graines) 3.2 - Evolution du poids et de la longueur :Les figures n°20 et 22 représentent l'évolution du poids et de la longueur pendant le cycle de reproduction 3.2.1 - L'année N : 3.2.1.1 -- Poids : L'évolution pondérale durant l'année N reste faible : un mois après leur apparition, le poids moyen enregistré n'est que de 0,0695g pour l'exposition Nord, et 0,0793g pour celle du Sud. Ceci ne présente que 0,070 % et 0,085 % par rapport au poids des cônes mûrs. A la fin de l'année N, le poids moyen des inflorescences femelles est de l'ordre de 0,1g, ce qui représente à peine 1% par rapport aux cônes mûrs. 3.2.1.2 -- Longueur : Un mois après leur initiation, la longueur moyenne des inflorescences femelles est de 1,08 cm pour l'exposition Nord et 1,01 cm pour l'exposition Sud.Ceci représente respectivement 13,70 % et 14,06 % par rapport aux cônes murs. L'accroissement se stabilise autour de 1,20cm à partir de la mi--Octobre ; il reprendra vers le printemps de l'année suivante 3.2.2 - L'année N+1 : Rappelons que c'est l'année des principales phénophases : formation de cônelets et de cônes. C'est également une année cruciale pour la formation des graines : durant la fécondation beaucoup de cônelets se dessèchent à ce stade, suite à des problèmes liés probablement à la station et l'état de l'arbre (absence de la pollinisation et à la formation d'ovules anormaux). Pour l'étude des phénophases durant l'année N+1, l'échelle adoptée est la suivante : - Phase Une : croissance lente. - Phase Deux : croissance rapide. - Phase Trois : croissance lente. 53 3.2.2.1 - Phase Une : Cette phase qui dure trois mois et demi, débute au mois de Janvier et se termine à la mi -- Avril.Durant cette période l'accroissement du poids et de la longueur des inflorescences femelles reste modeste, cependant il est plus important sur l'exposition Nord que l'exposition Sud. Par ailleurs, l'accroissement en longueur est plus important que celui du poids. En effet, au mi - Avril, le poids moyen enregistré est de 0,19g pour l'exposition Sud et 0,33 g pour celle du Nord ; ce qui représente 0,21 à 0,34 % par rapport aux cônes mûrs. Quant à la longueur, elle atteint 1,90 cm pour l'exposition Nord et 1,15 cm pour l'exposition Sud soit un taux supérieur à 20 %. 3.2.2.2 - Phase Deux : Elle se caractérise par la transformation des inflorescences femelles en cônelets puis en cônes. Formation du cônelet : Elle dure 45 jours environ. Elle débute vers la mi - Avril et se termine à la fin du mois de Mai. Durant les deux dernières semaines d'Avril, la croissance des inflorescences femelles s'accélère : elles se préparent à la fécondation. Elle a lieu vers la première semaine du mois de Mai. La période de la fécondation varie selon les arbres et l'exposition. Elle est plus précoce sur l'exposition Nord et les branches hautes (cônes dégagés). Après la fécondation, l'inflorescence femelle devient cônelet. Le poids et la longueur sont respectivement multipliés par 5 et par 2 pour l'exposition Nord, vers la fin du mois de Mai (15 g pour 4,30 cm). Pour l'exposition Sud, l'accroissement enregistre un certain retard, puisque le poids obtenu à la même date n'est que de 7g pour 3 cm. Ce retard qui sera rattrapé par la suite (vers le 10 Juin) s'explique par les difficultés que rencontre le cèdre à prospérer sur les expositions défavorables. Formation du cône d'une année : Du mois de Juin à la mi - Juillet la croissance du cônelet est extrêmement rapide, il atteint presque ses dimensions définitives et devient cône en 45 jours : c'est la maturité morphologique.Cette croissance exponentielle permettra aux cônes d'atteindre des poids de l'ordre 90 g et des longueurs dépassant les 6,70 cm à l'état frais. 3.2.2.3 - Phase trois : Les cônes gorgés d'eau continuent à se développer en longueur mais très lentement. Leur couleur vire au violet puis au marron -- roux ; à ce moment là, le poids commence à diminuer très lentement par déshydratation. 54 55 3.2.3 - L'année N+2 : C'est l'année de la maturité physiologique, elle se traduit par une déshydratation des cônes N+1. Cette dernière commence à partir de Septembre - Octobre de l'année N+1 jusqu'au mois d'Août de l'année N+2. Les pertes d'eau enregistrées pour l'exposition Nord sont de l'ordre de 16 % et 10 % pour celle du Sud. Dès l'Automne, les cônes (N+2) s'imbibent d'eau, leur poids augmente de 10 % environ, c'est le prélude à la désarticulation et à la dissémination des graines. 3.2.4 - l'année N+3 : Les cônes fortement imbibés se désarticulent facilement ; dès la première semaine de Janvier, la dissémination des graines est pratiquement achevée. 3.3 - Etude des variations :3.3.1-Analyse de la variance :Au cours d'une série de mesures et d'oservations sur le processus de développement des cônes dans deux stations distinctes et en vue de comparer le poids et la longueur durant leur cycle, nous avons procéder à l'analyse des données pendant l'année N, (N+1), (N+2) et [(N) + (N+1) + (N+2) + (N+3)] afin de mettre en évidence, l'existence d'éventuelles différences par exposition. Le tableau n°7 montre que l'influence de la station, sur l'évolution des inflorescences femelles n'est pas significative pour les deux caractères selon les critères adoptés. En effet, l'analyse de la variance au seuil de 5 % montre que le F test est inférieur à F théorique pour les années. Ainsi, l'exposition n'a pas d'influence sur la croissance des cônes, les dimensions augmentent en poids et s'allongent de la même manière. Tableau N°7 : Evolution du poids et de la longueur des inflorescences femelles au cours du temps. Influence de l'exposition.
3.3.2- Corrélation entre l'évolution le poids et la longueur. Cette matrice renferme 4 variables, poids et longueur de l'exposition Nord et de l'exposition Sud, à 16 observations chacune correspondant à 16 périodes de mesures. Le tableau n°8 montre qu'il existe des corrélations hautements significatives entre les poids et la longueur aussi bien pour l'exposition Nord que Sud (Coefficient de corrélation > 0,9). La valeur critique du coefficient de corrélation est de 0.742 à 1 °/o o Tableau N°8 : Matrice de corrélation entre les dimensions des organes de reproduction femelle.
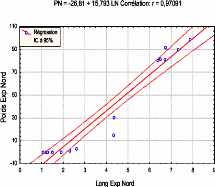
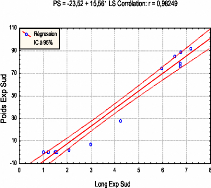
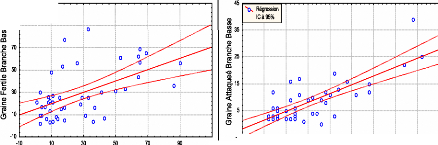
La figure ci-dessous montre l'existence de fortes relations linaires entre l'accroissement en poids et en longueur des inflorescences femelles depuis leurs apparitions jusqu'à leur désarticulation. 56 Fig.21: Corrélation entre le poids et la longueur des organes de reproduction au cours de leur évolution.
7 8 9 PN = -26,81 + 15,793 LN Corrélation: r = 0,97091 110 90 10 -10 2 3 4 5 Long Exp Nord 1
7 8 PS = -23,52 + 15,561 LS Corrélation: r = 0,98249 2 3 4 5 Long Exp Sud 10 0 1 110 90 7 -a0 rn X x 50 50 3 a_ 10 En prenant en considération l'évolution du poids et de la longueur par année nous remarquons qu'il n'y a pratiquement pas de différence de poids et de longueur entre les deux expositions (Cf. fig n° 20 et 22)
Poids(g)
Longueur(cm) 91,542 88 Q16 7,88 7,33 7,18 6,75 6,76 6,5 98,521 --95,561 89,651 85,102 0,115 0,0978 1,28 1,21 N+1 N+2 N+3 N N+1 N+2 N+3
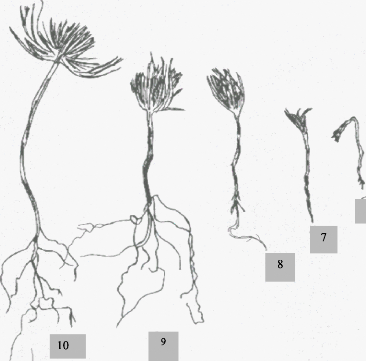
57 3.4 -- Discussion :Le cycle de reproduction représenté par la figure N° 23 montre que dure 32 mois depuis l'apparition des inflorescences femelles jusqu'à la fin de la désarticulation des cônes. La première année il y eut l'apparition des fleurs mâles puis celles des inflorescences femelles. La floraison ayant eu lieu en été, la pollinisation n'aura lieu qu'au début de l'automne de la même année. La période printanière voit l'activité végétative reprendre, on assiste alors au développement du chaton mâle et à la fécondation. La période comprise entre de la pollinisation à la fécondation est décisive quant à la production qualitative des graines. L'influence de certaines conditions climatiques particulièrement les gelés printanières peuvent causer de graves dégâts aux chatons, ainsi BONNETMASSIMBERT (1984) dans son étude sur la biologie florale et le cycle de reproduction des arbres, a remarqué l'importance des pertes causées par les températures basses durant la période printanière. TOTH (1985) indique que l'intervalle entre la pollinisation et la fécondation est la conséquence directe d'une floraison automnale qui se produit lors de l'arrêt de la végétation. Durant la seconde année, après la dormance hivernale il y eut de nouveau un changement, qui se traduisit par une augmentation assez lente du poids et des dimensions. Vers la mi - Mai la fécondation se produisit, la croissance devint très importante, l'inflorescence femelle devint cônelet. Les cônelets continuent à se développer rapidement. A partir de la fin du mois de Septembre, la couleur verte du cônelet vire progressivement est devient marron.: la maturité morphologique est atteinte c'est le cône N+1. Ce dernier continue son développement, sa maturité physiologique ne sera atteinte que pendant les deux derniers mois de l'été de l'année N+2. A partir de Décembre, la désarticulation atteint son optimum, elle dure jusqu'au début de Janvier. Durant ce cycle, nous assistons à l'augmentation du poids et de la longueur dans des proportions variables selon les phénophases, tantôt elle est lente, tantôt elle est rapide. C'est durant l'année N+1 que l'augmentation en poids et en volume est importante (Cf. Fig. n° 20). Notre étude nous a permis de déterminer le cycle de reproduction, de situer chronologiquement toutes les phénophases et de proposer une échelle de notation comportant les stades d'observation de l'appareil reproducteur. Cette échelle nécessite de fréquentes sorties. 22-Fécondation et formation de 32- Ouverture des écailles. cônelets. 1-L 'année N : 11- Bourgeons floraux 12- Gonflement des bourgeons floraux. 2-L'année N+1 : 21- Croissance lente des 3-L'année N+2 : 4-L'année N+3 41- Fin de la 31- Déshydratation des cônes. 13- Apparition des fleurs 23- Croissance rapide, formation mâles. des cônes et des graines. 33- Désarticulation des cônes.
58 Juillet Septembre Novembre
Début d'ouverture Ouverture totale
Chute totale Apparition
Année N+1
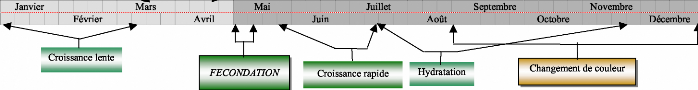
Octobre Novembre Décembre III Septembre Mars Janvier Mai Juillet Croissance rapide ti: Juin 1 I f___ t Août FÉCONDATION Croissance lente Hydratation Changement de couleur
_ f I 1 1 Février 1 I 1 1 Avril Année N+2
Décembre ........... e ........... ...... ................................ Février Avril Juin Août Octobre
Année N+3
Février 11H Avril
Inflorescence Fin de la désarticulation Apparition des semis Fig. 23 : Le cycle de reproduction du cèdre de l'Atlas (Cedrus atlantica M) au Belezma : principales phénophases. Notons également que nos observations, ne convergent toujours pas avec celles de TOTH (1978 a et 1984), ni celles de DERRIDJ (1996). Ainsi, ces deux auteurs indiquent par exemple que la désarticulation peut aller jusqu'au mois de Mars. Pour la cédraie du Belezma, la désarticulation s'est toujours achevée dès la première semaine de Janvier. Ceci est dû aux conditions écologiques (températures et précipitations) qui influencent tout le cycle de développement. 3.5 -- Conclusion :Lors de nos différentes observations, nous avons reconstitué le cycle de reproduction avec les principales phénophases où nous avons remarqué un décalage entre les arbres, même ceux d'une seule exposition. Ceci est du à la qualité du site aux peuplements (densité, âge...) au climat (LAHMADI et al, 1994). Selon nos observations, les fleurs males et les inflorescences femelles se présentent toujours en position terminale sur respectivement des rameaux courts et des rameaux longs portés par des axes rigoureux. Les rameaux courts se répartissent autour de ceux portant des inflorescences femelles. La floraison du cèdre de l'Atlas est précoce sur l'exposition Sud. C'est probablement la température qui influence cette précocité. Cependant il n'y a pas de grandes différences entre les deux expositions quant à l'évolution des cônes, quelle que soit l'année. NEDJAHI (1988) note que les bourgeons situés sur exposition Sud, présentent un débourrement plus précoce. La tardiveté ou la précocité du débourrement est généralement d'ordre génétique (OSWALD, 1984) Par ailleurs, les changements annuels de l'appareil aérien sont en étroite relation avec les conditions écologiques (températures moyennes égales à 12,40°C) et l'âge des arbres (AUSSENAC et al 1981) Durant l'année N+1, la formation des cônelets est légèrement tardive sur l'exposition Sud. Ce retard qui sera rattrapé dès la deuxième semaine du mois de Juin; ainsi on enregistre à cette date, pratiquement le même poids et la même longueur (30g pour 4.25 cm). Ces cônelets qui évoluent en cônes atteindront en Automne de l'année N+1 leur poids optimum (supérieur à 40 g). Le poids des cônes commence à baisser progressivement surtout en été de l'année N+2. Cette déshydratation se traduit par une désarticulation dès le mois de Décembre ; elle est précoce sur l'exposition Sud où elle dure un mois, sur exposition Nord, elle est plus longue et dure un mois et 20 jours. De point de vue biométrique les variations sont très importantes. Elles dépendent de la station, de l'arbre, de l'année de fructification, de l'exposition des arbres et des branches ainsi que de la position du cône dans l'arbre. Notons, qu'au sein d'une même branche, nous avons noté d'importantes variations : une initiation précoce des inflorescences femelles donne des cônes les plus dimensionnés et les plus fertiles. Les cycles de reproduction dépendent des variations de l'environnement climatique et microclimatique : la pollinisation et la fécondation peuvent jouer un rôle important dans le caractère irrégulier de la fructification. Il existe de grande variabilité dans la phénologie de la floraison à l'intérieur d'une même espèce. 60 CHAPITRE III : Estimation de la fructification et de la productiondes organes de reproduction femelles.
r 61 1-- Introduction et problématiqueLa production semencière des essences forestières varie d'une année à une autre. Pour le cèdre de l'Atlas elle se caractérise par une année de production abondante suivie généralement d'une ou plusieurs années de moyenne à faible production. Cette périodicité est un facteur important qu'il faut déterminer et le prendre en considération lors de la planification de la récolte des graines. La récolte des semences forestières est actuellement extensive, il est souhaitable sinon donc indispensable de concentrer la cueillette des graines sur les années de forte fructification afin de constituer des stocks pour les années de faible production. Ceci permettra un approvisionnement régulier, grâce au stockage, des pépinières à moindre frais tout en préservant les possibilités de régénération naturelle. En effet, une année de forte fructification, se caractérise par une récolte facile, peu coûteuse et surtout de bonne qualité. BARITEAU et al (1994) rapportent que le prix des graines de cèdre de l'Atlas est de 150 à 200 $ le kg L'estimation de la fructification est une opération indispensable pour la récolte des graines et la reconstitution des peuplements. En Suède par exemple les prévisions des cônes chez Pinus sylvestris sont établies chaque année depuis 80 ans et qu'aux USA chaque année, on estime la production moyenne de cônes par essence et par zone géographique (F.A.O, 1982). Si l'estimation de la production quantitative globale n'est pas encore mise au point à cause de l'influence de divers facteurs (climat, altitude, année de fructification...), celle permettant son importance qualitative avec une année ou même deux années par un comptage de cônes ou d'inflorescences femelles a été déterminée par TOTH (1984) ; pour notre cas, nous avons utilisé cette méthode, puis nous l'avons adapté aux cédraies sèches. 2 -- Méthodologie :La méthode d'estimation adoptée est celle qui déterminé par TOTH, après une décennie d'étude sur la biologie de la fructification. Elle est utilisée actuellement pour apprécier l'estimation de la fructification des cédraies françaises. 2.1 -- Echantillonnage : 2.1 - Les stations : Les observations se feront sur deux stations: station à exposition Nord et station à exposition Sud (Cf. tableau n°6 et figures n°16 et n°17). 2.1.2 - Les arbres : Les arbres - échantillons choisis sur lesquels se feront toutes les observations, pendant un cycle complet de fructification sont : - Permanents : tous les comptages seront réalisés uniquement sur les mêmes échantillons. 62 - Représentatifs : arbres bien venants, d'âges moyens et surtout les plus fructifères. Dans chaque station (parcelle - échantillon), nous avons choisi 15 arbres, les plus fructifères en cônes de deux années qui porteront les numéros de 7 à 21. Leur distribution au niveau de chaque station est la suivante:
Afin d'éviter l'influence du facteur altitudinal entre les deux versants, les arbres choisis sont situés dans une tranche altitudinale comprise entre 1.600 et 1.650 m. 2.1.3 - Les branches : Comme pour les arbres, les deux branches choisies sont
également les plus fructifères en cônes de deux
années. La première branche est localisée dans la
partie haute de l'arbre, généralement au 3/4 du
houppier: c'est la branche haute (BH).La seconde branche est située
à hauteur d'homme Afin d'éviter toute confusion entre les différentes branches, celles choisies sont peintes en rouge au niveau de leur base. Ce sont en réalité des sous- branches et non des branches complètes qui partent du tronc. Leur longueur est supérieure à 1,50 m. 2.2 - Les observations : Toutes les observations, sont effectuées pendant trois années sur les mêmes arbres et les mêmes branches. Il s'agit de compter les organes de reproduction: les inflorescences femelles, les cônes d'une année et les cônes de deux années. Pour le dénombrement des cônes aucune difficultés n'est à signaler (différence de couleur, consistance et taille) entre ceux d'une année (N+1) et de deux années (N+2) ; quant aux inflorescences femelles leur couleur qui épouse celles des aiguilles et leur petite taille posent des problèmes de comptage. D'où la nécessité d'effectuer au mois 3 répétitions par branches. Le mois d'Octobre est préconisé par TOTH (1984) pour le comptage de tous les organes de reproduction. Ceci permettra de faire un seul comptage pour les trois catégories d'organes d'une part, et de l'autre part les inflorescences avortées après pollinisation sont facilement reconnaissables (sèches) ne seront pas prise en compte. Signalons qu'il est possible de réaliser des comptages en Novembre, mais des problèmes d'exécution se posent en raison du froid. Cependant selon, nos différentes 63 observations, l'intervalle compris entre la mi-Septembre et le mois d'Octobre est la période la plus indiquée pour le comptage dans la cédraie du Belezma parce que: - Les inflorescences femelles sont relativement grosses donc facilement reconnaissables. Celles qui sont avortées sont de couleur brune, sèches et rigides ne seront pas comptabilisées. - Les cônes N+1, dont leur couleur a viré au violet- marron, avec des taches vertes à la base. Ils sont très consistants au toucher. - Les cônes N+2 un peu plus gros que les cônes N+1, sont de couleur marron foncé avec des écailles molles, légèrement entrouvertes au niveau de la partie haute. Notons enfin, que pour faciliter les observations il faut de préférence s'installer au- dessus des branches car les organes sont dressés. Les données brutes sont reportées en annexes V 2.3 - Estimation de la fructification : Cette étude, qui a pour objet de prévoir les possibilités de récolte par l'utilisation de la méthode de TOTH (1984).Néanmoins, nous pensons que cette méthode doit être ajuster aux cédraies algériennes, qui sont plus sèches. La fructification est en relation directe avec le climat. Elle dépend de plusieurs facteurs externes et internes à l'arbre et de leurs interactions. La prévision quantitative et qualitative se heurte à de nombreux obstacles qui en découlent en partie, du cycle de reproduction lui-même. En effet, TOTH (1984) rapporte que le cycle de reproduction n'est pas immuable et que l'abjectif « forte », moyenne ou « faible » appliquée à une année peut être affectée de variations importantes (en nombres de cônes récoltables) d'un cycle à un autre. 2.3.1 - Méthode de TOTH : La production des graines fertiles du cèdre de l'Atlas dure en général trois ans. Mais il peut se prolonger jusqu'à quatre ans dans certaines conditions. C'est cette longue période, qui a été utilisée par TOTH (1984) pour prévoir l'importance de la fructification et l'évaluation des possibilités de récolte par simple comptage des inflorescences femelles et / ou des cônes d'une année, sur 30 branches à raison de deux branches par arbre. Les cônes d'une année permettent une prévision d'une fiabilité totale de 14 mois. Celles des cônelets de 9 mois (avant fécondation), dès le mois de Mai de l'année N+1, permet une prévision d'une fiabilité excellente de 17 mois à l'avance. Pour les inflorescences femelles, après pollinisation, dès Octobre de l'année N, la prévision est d'une bonne fiabilité de 24 mois à l'année (Cf. tableau n° 9 et annexe IV). TABLEAU N°9: Prévision qualitative de la possibilité de récolte 1 ou 2 ans à l'avance, à partir du nombre d'inflorescences femelles ou des cônes de 1 an comptés sur 30 branches- échantillon dans une parcelle (2 branches par échantillon, 15arbreséchantillons dans la parcelle).
64
> 250 > 100 Forte (récolte assurée et conseillée) TOTH (1984) 2.3.2 - Adaptation de la méthode de TOTH à la cédraie du Belezma : Les travaux de TOTH (1984), sur la production grainière de cèdre de l'Atlas ont mis en évidence l'influence de l'année de fructification sur la variabilité qualitative et quantitative des graines. Toutefois, le facteur station pourrait être également aussi important sinon plus quant à la capacité des arbres à produire des semences. En effet, selon les études de DERRIDJ (1990) les facteurs écologiques jouent un rôle primordial sur la qualité des graines. Pour adapter la méthode de TOTH, à la cédraie du Belezma, nous nous sommes basés sur le principe suivant : pour avoir les mêmes possibilités de récoltes, il faut que la cédraie du Belezma, produise le même nombre de graines fertiles que celle du Mont- Ventoux. Pour cela, nous avons calculé pour deux stations du Belezma le nombre d'inflorescences femelles et de cônes, en fonction de l'importance de la fructification (Forte- Moyenne- Faible) en utilisant les données suivantes : -Nombre moyen de graines fertiles par cône (N+2) pour le Belezma et le Mont-Ventoux. -Le nombre de cônes (N+2) selon l'importance de la fructification pour le Belezma. -Le taux de pertes des inflorescences femelles par rapport aux cônes (N+1) pour le Belezma. En outre, les données utilisées sont les moyennes d'un cycle de reproduction pour le Belezma et de trois cycles le Mont -- Ventoux. 2.4 - Les productions des organes de reproduction : Apres l'estimation de la fructification les résultats obtenus, sont ensuite traités dans un premier temps par des paramètres de tendance centrale et de dispersion, afin de rechercher les relations entre les différents organes de reproduction et leurs distributions. Dans une deuxième étape, nous essayerons de mettre en évidence l'influence des critères de classification à savoir la station, la branche et l'année de fructification et leurs interactions sur la production des inflorescences femelles, des cônes d'une année et deux années par une analyse globale. A ce titre, nous avons utilisé l'analyse de la variance et le test de NEWMAN et KEULS. Par ailleurs, TOTH (1984) a noté que selon l'expérience acquise, le nombre d'arbres (15 par station) et le nombre de branches (deux par arbre) sont statistiquement convenable. 3 -- Résultats :3.1 - Prévisions et estimation de la fructification : 3.1.1 - Méthode de TOTH : Les travaux de TOTH (1978 a et 1984) ont montré que le cycle de reproduction qui dure en général 3 années, se caractérise par : - Une année de floraison abondante, conduisant à une forte possibilité de récolte deux années plus tard. - Suivie d'une année moyenne, avec une possibilité de récolte. 65 - Elle-même suivie d'une année faible à nulle. Néanmoins le même auteur signale que la fructification pourrait être affectée par de grande variation d'un cycle à un autre. En effet, aussi bien le cycle de reproduction, que l'importance de la fructification ne sont pas stables et dépendent des facteurs du milieu. Afin de prévenir la fructification du cèdre d l'Atlas nous avons utilisé le " tableau de référence " : Tableau n° 9) Les résultats obtenus sont reportés dans le tableau n°15, qui montre l'alternance de la fructification selon les années de production. Les possibilités de récolte sont :
3.1.2 - Adaptation de la méthode de TOTH : 3.1.2.1 - Evolution du nombre de cônes : 3.1.2.1.1 - La cédraie du Mont - Ventoux : Les résultats des travaux de TOTH sur la prévision des possibilités de récolte des cônes au Mont- Ventoux (plaine de l'Escalier) durant une décennie (1974-1983) sont reportés en annexe IV. Ils nous permettent de suivre le nombre des inflorescences femelles, des cônelets et des cônes (N+1) au cours de leur évolution. Le nombre total des inflorescences femelles (Octobre N) et celui des cônes (Août N+1) durant la période de 1974 à 1983 est reporté sur le tableau n° 10 qui indique que : - Le nombre d'inflorescences femelles ayant atteint le stade de cônes (N+1) est de 1041 soit une perte de 1660 individus ce qui représente 61,25 %. - Quant aux cônes (N+1), aucune perte n'a été enregistrée durant leur passage de la maturité morphologique à la maturité physiologique (N+2) Tableau N°10 : Evolution du nombre d'inflorescences femelles et des cônes au Mont- Ventoux. Période 1974- 1983.
(TOTH, 1984) 66 Ces chiffres montrent que la période comprise entre l'initiation florale et la maturité morphologique se traduit par d'énormes pertes. Celle allant de la maturité morphologique à la maturité physiologique se fait sans dommage. 3.1.2.1.2 - La cédraie du Belezma : Les données du Belezma, (Cf. tableau n° 11) indiquent que pour l'exposition Nord, les pertes subies par les inflorescences femelles depuis leur apparition jusqu'au stade cône N+1 sont de l'ordre de 52.22%.Celles de l'exposition Sud atteignent 73,44 %. Ce qui correspond à un taux équivalent à celui du Mont-Ventoux pour les deux expositions. Par ailleurs, le passage des cônes (N+1) au stade cônes (N+2) se fait en principe sans aucun problème. Autrement dit, quand le cône atteint sa maturité morphologique, il parviendra après 11 mois à sa maturité physiologique sans aucune perte. Tableau N°11 Evolution du nombre des inflorescences femelles pendant un cycle complet du Belezma. Influence de l'exposition.
Il faut noter que l'importance des pertes de l'exposition Sud en inflorescences femelles est compensée par sa grande production. Ceci peut s'expliquer par la capacité de l'arbre à moduler son investissement dans la fonction femelle, en fonction de son état et la disponibilité des ressources. 3.1.2.2-Estimation des organes de reproduction au Belezma selon de l'importance de la fructification : 3.1.2.2.1 - Principe : Il s'agit de déterminer le nombre de cônes mûrs et celui des inflorescences femelles produit par 30 branches sur 15 arbres les plus fructifères d'un peuplement pour estimer les possibilités de récoltes. Rappelons que nous utilisons le nombre moyen de graines fertiles par cône et le nombre de cônes produit selon l'importance de la fructification et les pertes des inflorescences par apport aux cônes pour le Mont-Ventoux. Ceci nous permettra de calculer le nombre de graines fertiles en fonction de l'importance de la fructification. Cette estimation sera calculée pour les deux expositions, car ces dernières ont une influence sur la production et le processus de l'évolution des organes de reproduction ainsi que la qualité des graines et donc sur les possibilités de récolte. 67 Nous pensons que l'appréciation de la production des graines fertiles est plus précise que celle basée sur la production des cônes étant donnée l'importance de la variation de ces graines par cône (CV > à 40 % quelle que soit la station. Cf. chapitre V : Etude des cônes et des graines) 3.1.2.2.2 - Les cônes : En estimant le nombre de cônes TOTH (1984) qualifia la fructification de faible si ce nombre est inférieur à 50, moyenne s'il est compris entre 50 - 100 et forte s'il est supérieur à 100(Cf. tableau n° 9). Par ailleurs, selon les données du même auteur, le nombre moyen de graines fertiles par cône est de 87,40 au Mont --Ventoux. Celui du Belezma n'est que de 33.21 pour l'exposition Sud et de 53.60 pour l'exposition Nord. Ainsi, l'estimation des cônes au Belezma est obtenue de la façon suivante: - Pour le Mont- Ventoux, nous déterminons le nombre moyen de graines fertiles en fonction des possibilités de récolte. - Puis, nous calculons le nombre de cônes que doit produire la cédraie de Belezma, selon l'exposition pour avoir le même nombre de graines que le Mont- Ventoux en prenant en considération l'importance de la fructification. Exemple: Si la fructification est considérée, comme faible au Mont- Ventoux, le nombre de graines fertiles doit être inférieur à 4.370 (50 x 87,40). Au Belezma, pour avoir le même nombre de graines fertiles : L'exposition Nord doit produire 81,52 cônes (N+2) (4.370/53,60). Quant à l'exposition Sud le nombre de cônes serait 131,58 (4.370/33,21). La même démarche est appliquée, quand la fructification est moyenne ou forte. Les résultats obtenus, sont reportés sur le tableau n°12 qui montre que pour avoir les mêmes possibilités de récolte que le Mont -Ventoux, le Belezma doit produire plus de cônes (N+2) : le nombre de graines des cédraies sèches par cône est relativement peu important. Tableau N°12: Estimation qualitative de la fructification et possibilités de récolte des cônes au Mont- Ventoux et au Belezma par le nombre de cônes et de graines.
68
3.1.2.2.3 - Les inflorescences femelles : Le nombre d'inflorescences femelles est déterminé en fonction de leurs pertes. Pour le Mont- Ventoux, elles sont estimées à 50 % (TOTH, 1984). Les données du tableau n°11 relatif à l'évolution des inflorescences femelles et celles du tableau n°12 sur l'estimation qualitative de la fructification par le nombre de cônes nous permettent de déterminer le nombre des inflorescences femelles en fonction de l'importance de la fructification. Exemple: Supposant que la fructification est faible, le nombre d'inflorescences est de 100, celui des graines fertiles correspondant est de 8.740 (100 x 87.40). Au Belezma pour avoir le même nombre de graines fertiles sur l'exposition Sud, sans perte, il faut 263.17 cônes (8.740/33.21).Les pertes estimées sont de 73.44% ce qui représente 193.27 cônes. Le nombre des inflorescences femelles est de 263.17+193.27 = 456.44 Pour déterminer le nombre d'inflorescences maximal quand la fructification est moyenne il suffit de multiplier celui de la fructification faible par le coefficient 2,5. Le nombre d'inflorescences femelles obtenu selon l'estimation de la fructification pour l'exposition Nord et Sud est porté sur le tableau n° 13. Tableau N°13 : Estimation de la floraison selon l'exposition par le nombre d'inflorescences femelles.
3.1.2.3 - Estimation de la fructification : Les données relatives à l'estimation de la fructification et aux possibilités de récolte dans la cédraie de Belezma selon les inflorescences et les cônes, sont reportées dans le tableau ci- dessus. 69
Tableau n°14 : Estimation de la fructification et possibilités de récolte à partir du nombre des inflorescences femelles, des cônes (N+1) et (N+2) sur 30 branches.
En comparant le tableau n° 9 et n° 14 nous constatons que pour avoir les mêmes que pour avoir les mêmes possibilités de récolte, le Bélezma doit d'une part produire beaucoup plus d'inflorescences femelles et de cônes que le Mont-Ventoux, d'autre part les stations de l'exposition Sud 'doivent' produire plus de d'organes de reproduction que celles Nord. Les possibilités de récoltes selon nos estimations et ceux obtenues par TOTH, sont reportées dans le tableau ci-dessus. Tableau N° 15 : Prévision qualitative de la possibilité de récolte.
3.1.3 -- Discussion - conclusion : A partir de cette étude, nous constatons que pour avoir les mêmes possibilités de récolte, les stations écologiquement défavorables doivent « produire » plus de cônes que les stations favorables. En effet, au cours de son cycle de reproduction certaines fleurs des inflorescences femelles subissent des avortements qui se traduisent soit, par une diminution qualitative et quantitative des graines soit par leur perte après dessiccation. Si au Mont - Ventoux, ces pertes sont de l'ordre de 61,46 % pour les inflorescences femelles, celles des cônes (N+1) sont nulles. Au Belezma, les pertes varient selon l'exposition : - Pour l'exposition Nord, elles atteignent une moyenne de 50,55 % pour les inflorescences femelles. - Dans la station à exposition Sud, les pertes enregistrées sont de l'ordre de 73 % La variation de ces pertes serait due à la valeur de la station. En effet les facteurs écologiques influent tout le processus du cycle de reproduction, les plus importantes pertes 70 Deuxième partie Chapitre III : Estimation de la fructification et de la production des organes de reproduction femelles touchent surtout les premières phénophases (Pollinisation -- Fécondation) : période ou il y a une concurrence entre le cycle végétatif et reproducteur. Mais une fois le cône atteint sa maturité morphologique, il n'y a plus de déperditions. Le cèdre de l'Atlas, aurait donc tendance à accommoder sa capacité dans la production des inflorescences femelles et des fleurs mâles selon la disponibilité des ressources (Eau, éléments minéraux...). Beaucoup d'inflorescences femelles tombent au cours de la pollinisation et de la fécondation ajustant apparemment le nombre de fruits aux disponibilités des rameaux en ressources (MC. CARHY in KROUCHI, 1995). Comme le montre nos résultats, l'incapacité des stations du Belezma, en particulier les expositions Sud de mener terme la totalité des inflorescences au stade de la maturité, serait due à la faiblesse de l'intensité de pollinisation, au déficit de l'humidité... Le nombre de graines fertiles est lié à plusieurs facteurs : (Cf. chapitre W) - La limitation des ressources. - La position des cônes dans l'arbre et celle des graines dans le fruit. - La source et la disponibilité du pollen. - Les paramètres écologiques exceptionnels. - Les insectes. La floraison et la fructification du cèdre de l'Atlas au Belezma varient énormément d'un endroit à un autre, les micro-climats semblent jouer un rôle substantiel. En effet nous avons noté que les arbres d'une même exposition ont des cycles différents, certains fleurissent une année et d'autres l'année suivante. La production grainière est en relation avec la production des cônes et varie en fonction des conditions écologiques qui influencent l'initiation florale, la pollinisation, la fécondation, la croissance et la maturité des cônes ; cependant selon TOTH (1978 a ; 1978 b et 1984) ces variations individuelles annuelles, ne perturbent pas l'ensemble de la récolte. Ainsi la production qualitative et quantitative du Mont- Ventoux est bien meilleure que celle du Belezma. En outre la station Nord et Nord - Ouest du Belezma sont les plus fertiles. La méthode la plus proche de la réalité pour la récolte des cônes est reportée dans le tableau n°14, elle serait la plus indiquée pour les cédraies sèches. Toutefois, il faut continuer à récolter les données pendant au moins une décennie, pour avoir les meilleurs résultats. Par conséquent, il n'est pas judicieux d'utiliser les données du tableau n° 9 pour étudier la prévention de la fructification et les possibilités de récolte de la cédraie du Belezma, étant donné les conditions écologiques qui diffèrent considérablement de celles de la cédraie du Mont-Ventoux. 3.2 - Les productions des organes de reproduction :Les résultats obtenus par branche, pour les deux branches (B.H et B.B) et la somme des deux branches, selon la station et l'année de fructification sont reportés dans le tableau n° 16. 3.2.1 - Les productions sur une année : 3.2.1.1 -- Production par branche: 71
Les principaux paramètres statistiques sur la production des inflorescences femelles, des cônes (N+1) et (N+2) montrent qu'il existe de grandes variations selon l'année, la branche et la station. Tableau N° 16 : Paramètres statistiques sur la production des inflorescences femelles des cônes (N+1) et des cônes (N+2).
72
3.2.1.2-Production des deux branches La production des inflorescences femelles et des cônes (N+1) et (N+2) pour les deux branches et par année est portée sur le tableau ci-dessus. Tableau N°17:Paramètres statistiques sur la production des inflorescences femelles, des cônes (N+1) et (N+2) selon les deux branches. Effet de l'année de fructification.
-Les inflorescences femelles La production des inflorescences femelles est plus importante sur l'exposition Sud que Nord. En effet, leur nombre oscille entre un minimum de 01 et un maximum de 192 individus avec une moyenne de 54.46 pour la station à exposition Sud. Quant à celle du Nord, le nombre d'individus varie entre 00 et 152, la moyenne est de 44.06. Les valeurs des coefficients de variation, montrent l'existence d'écart dans la production des inflorescences femelles selon les arbres pour chaque année. Ainsi, quand la floraison est importante l'écart entre les arbres est relativement peu élevé, quand elle est faible l'écart devient considérable. Nos résultats montrent, que certains arbres sont incapables de produire des inflorescences femelles en quantité par rapport à d'autres. Autrement dit tous les arbres ne sont pas en état d'engendrer chaque année le même nombre d'inflorescences femelles. 73 Deuxième partie Chapitre III : Estimation de la fructification et de la production des organes de reproduction femelles OSWALD (1988) rapporte l'existence de grandes variabilités dans la phénologie de reproduction pour une même espèce. -Les cônes (N+1) La production varie pour l'Exposition Sud entre un minimum de 03 cônes et un maximum de 51 cônes avec une moyenne de 15.44 cônes. Celle du versant Nord est comprise entre 01 et 84 cônes. La moyenne obtenue sur les 03 années et de 22.48 cônes. Contrairement aux inflorescences femelles, la production des cônes N+1 est plus forte sur l'exposition Nord que sur l'exposition Sud. Par ailleurs, l'année de production moyenne à été la plus fructifère, suivi de l'année de forte production. Celle à faible production a été la plus mauvaise. La variation est fonction surtout de l'année de fructification. Les valeurs du coefficient de variation sont peu élevées par rapport à celles obtenues pour les inflorescences femelles. Pour une même année les écarts de la production des cônes d'une année entre les arbres sont plus importants sur l'exposition Nord que l'exposition Sud. Ces résultats indiquent une tendance de certains arbres à assurer une production de cônes (N+1) pour chaque année.Ces arbres qu'on peut qualifier de fertiles, sont plus nombreux sur l'exposition Nord que Sud. En effet, quant la fructification est faible, la production est uniforme pour l'ensemble des arbres ; Forte, l'ensemble des arbres contribue à la production des cônes (N+1) de la même manière. Quant elle est moyenne il y a une proportion d'arbres qui continuent à produire fortement de cônes par rapport à d'autres. Il y a une alternance de la production. -Les cônes (N+2) La moyenne obtenue est de 10.28 cônes (N+2) avec un minimum de 0 cône et un maximum de 49 cônes, pour la station à exposition Sud. Celle exposée au Nord varie entre 01 et 84 cônes (N+2) avec une moyenne de 17. Comme pour les cônes (N+1) l'exposition Nord est la plus fructifère. L'année à forte production a été de loin la plus prolifique par rapport aux années à faible et moyenne production. 3.2.2- La production sur trois années :La production des inflorescences femelles, des cônes (N+1) et des cônes (N+2) pendant un cycle de reproduction (trois années) sur les deux expositions est indiquée sur le tableau n°18. Tableau N°18: Paramètres
statistiques sur la production des inflorescences femelles, des
cônes
74
En prenant en considération la station, les résultats obtenus indiquent pour la floraison que le nombre des inflorescences est compris entre 0 et 192 individus, avec une moyenne de 49.12. Quant à la fructification, le nombre de cônes (N+1) varie entre 1 et 84 pour une moyenne de 24.86, celui des cônes (N+2) est compris entre 0 et 84 avec une moyenne de 13.64. Les valeurs élevées du coefficient indiquent l'existence de grands d'écarts de la production des inflorescences femelles, des cônes (N+1) et (N+2) selon les branches pour les trois années consécutives. Nous notons que la production cumulée pour deux branches par arbre est maximale pendant une année donnée, l'année suivante et /ou l'année d'après, la production est nulle. Il y a donc des arbres qui sont dans l'incapacité de produire des cônes chaque année. 3.2.3 -- L'alternance de la production Chez les conifères, l'abondance de la fructification annuelle n'est pas constante dans le temps. La production est alternée, cette alternance qui est due à des facteurs intrinsèques et extrinsèques a été soulignée par TOTH (1978a; 1980; 1984 ; 1985) DERRIDJ (1990), F.A.O (1992), KROUCHI (1995), BONNET-MASSIBMBERT (1984) et OSWALD (1983). Chez les arbres fruitiers, les variations peuvent avoir selon COURANDJOU (1983) des origines diverses principalement climatiques. Notre étude sur la fructification du cèdre de l'Atlas a permis de montré l'importance de l'amplitude sur la production quantitative et qualitative des graines et donc sur la l'ensemble du processus de la régénération. Pour apprécier l'alternance de la production pou les deux branches les plus fructifères et donc celle de l'arbre durant les trois années, nous avons calculé l'amplitude d'alternance (AP) en utilisant la formule de COURANDJOU (1982).
Quantité de fruits MAX Si la variation de l'alternance est égale à 100%, cela voulait dire que les branches n'ont pas assuré de production au moins une année sur trois. Une valeur faible signifie une tendance à la diminution de l'écart entre les productions annuelles des deux branches. Les résultats du tableau n°19 montrent que les branches appartenant aux arbres n°16 et 2lévoluant sur l'exposition Sud, n'ont pas assuré de production au moins une année sur trois (faible, moyenne et forte production) et une année sur deux (faible et moyenne production). Pour les années à moyenne et à forte production toutes les branches ont assuré une production au moins une année sur deux. Cependant, nous remarquons que sur l'exposition favorable, les arbres n°13 et 20 ont soutenu une production durant 2 années consécutives (faible et moyenne production). L'étude de la population des différents organes de reproduction par branche et pour les deux branches montre que les arbres de Cedrus atlantica M alternent leurs productions. Ainsi, 75
TOTH (1978) a noté que sur une période d'observation de 5 années, un nombre très faible d'arbres arrivent à produire des cônes chaque année. Tableau N° 19 : Amplitude d'alternance de la fructification selon les années. Année : Faible, moyenne et forte production.
Année : Faible et moyenne production
Année : Moyenne et forte production.
76
3.2.4- Corrélation entre les productions annuelles Cette matrice a pour objet de tester les liaisons qui peuvent exister entre les organes de reproduction pendant trois années de fructification. Elle renferme 18 variables, chacune est mesuré pendant chaque année et sur deux expositions. Elle montre que la production des différents organes de reproduction pour les trois années présente des corrélations au seuil de 5 %, 1 % 1 %o. Les résultats sont significatifs s'ils sont supérieurs ou égales à 0.482 (5%) ,0.605(1%) et 0.724 (1%o ). Tableau N°20 : Matrice de corrélation sur la production des inflorescences femelles, des cônes (N+1) et (N+2)
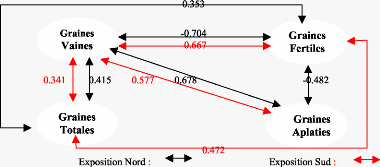
Légende: IF: inflorescences femelles - C 1 : cônes (N+1) - C2 cônes (N+2) - S : Exposition Sud - N : exposition Nord Année de - faible production (f) - moyenne production (M) - forte production (F). Sur exposition Nord: - Durant l'année à faible, les inflorescences femelles sont corrélées positivement production aux cônes (N+1) au seuil de 5% et (N+2) au seuil de 1%. - En outre, durant toujours l'année à faible production, nous enregistrons aussi des liaisons positives entre les inflorescences femelles et les cônes (N+1) et ( N+2) de moyenne production avec respectivement des valeurs de r égales à 0.549 et 0.599. Nous relevons également une liaison au seuil de 5%, entre les inflorescences femelles et les cônes (N+2) de l'année à forte production (r = 0,549 P =5%) ainsi qu'une liaison entre les cônes (N+2) de l'année à faible production et les cônes - Notons également que durant une année moyenne de production les cônes d'une année sont fortement corrélée aux inflorescences femelles (r = 0.801, P = 1%0) L'exposition Sud: 77
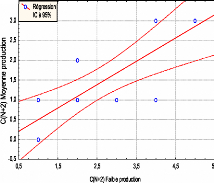
- Au seuil de 5%, la production des cônes (N+1) de l'année à forte production est positivement corrélés à ceux de l'année à faible production et aux cônes (N+2) de l'année à moyenne production (r = 0.588) O. Régression o 8,5 r 3,5 7,5
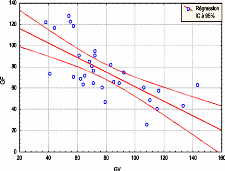
3,5 2,5 ..../0 0 0 10 20 30 40 50 60 Cone (N+1) forte production Forte production C(N+1)= 3.6454 + 0.0566 C(N+1) Faible production Moyenne production C(N+1)= 0.0920 +0.058896 C(N+1) Faible production R = 0.58807 R = 0.58807
1o, Régression o Q o
20 -2 2 20 6 10 14 18 05 1,5 25 3,5 4,5 5,5 Cone(N+1) faible production C(N+21 Faible production Faible production If = 52,589 +3.9018 C(N+1) Faible
production Faible production IF= 35.086 +16.150 C(N+2) Faible production R = 0.63669 78
140 `0, Régression IC à 95% 60 (7) C c 120 o lao 40 o 20 o 10 20 30 40 50 60 70 80 90 Deuxième partie
Fig. 24:C,orrélations mue les organes de reproduction selon leur stade.1nfluence de l'année de fructificationet de -Les cônes (N+1) de l'année à forte production de l'exposition Nord sont liés à ceux de l'année à faible production et forte production de l'exposition Sud (r = 0,624) et aux cônes (N+2) de l'année à moyenne production (r = 0,639). -Les cônes (N+2) de l'année à faible production de l'exposition Sud sont liés positivement au inflorescences femelles de ceux de l'exposition Nord produit également durant l'année à faible production ( r = 0,569 , P = 5% ). 3.2.5- Influence de la station, de l'année de fructification et des branches sur la production des inflorescences femelles et des cônes :3.2.5.1 - L'analyse de la variance : Pour étudier les variations de la production des inflorescences femelles, des cônes d'une année et de deux années selon leur position dans l'arbre : Branches hautes et branches basses et La somme des deux branches nous avons utilisé l'analyse de la variance : La première analyse, à deux critères de classification (Cf. tableau n°21 qui a but de comparer les stations et les années de fructification. Tableau N°21: Etude de la production des inflorescences femelles des cônes (N+1) et (N+2) pour les deux branches.Analyse de la variance.
79
La deuxième analyse à trois critères de classification qui étudie l'effet de la station, de l'année de fructification et des branches ainsi que leurs interactions est consignée dans le tableau n° 22. Tableau N°22: Etude de la production des inflorescences femelles et des cônes. Analyse de la variance.
Pour ces facteurs, l'analyse au seuil de 5 % fait apparaître selon les caractères étudiés les résultats suivants : Par branche Pour les deux branches Inflorescences Cônes Cônes Inflorescence
Cônes Cônes FACTEURS Facteur 1 : Année - 1-E +HF 1-E +1-E Facteur 2 : Exposition +++ +++ +++ +++ +++ +HF Facteur 3 : Branche 1-E 1-E 1-E INTERACTIONS DES FACTEURS Facteur 1 - Facteur2 + 1-E 1-E+ 1-E Facteur 1 - Facteur3 Facteur 2 - Facteur3 Facteurl-Facteur2- Facteur3 ... - non significatif + significatif -HF Très significatif -HE-Ellautement significatif 3.2.5.2 - Le test de NEWMAN et KEULS : 80 Deuxième partie Chapitre III : Estimation de la fructification et de la production des organes de reproduction femelles L'analyse de la variance, complétée par le test de NEWMAN et KEULS. 3.2.5.2.1 - Les deux branches : Pour les deux branches nous obtenons les résultas suivant. 3.2.5.2.1 -- Les facteurs étudiés: - Les stations TableauN°23:Détermination des groupes homogènes de station par les inflorescences Femelles.
Le tableau n° 23 et 24 montre que si pour le caractère inflorescence femelle les différences ne sont pas significatives, celui des cônes (N+1) et (N +2) donne deux groupes homogènes qui se différencient où l'exposition Nord donne les meilleurs résultats. Tableau N°24: Détermination des groupes homogènes de station par les inflorescences femelles et les cônes (N+1) (N+2).
- L'année de fructification: Pour ce facteur, nous obtenons trois groupes homogènes pour les inflorescences femelles et deux pour les cônes d'une année et de deux années (Cf. tableau n° 25) Tableau n° 25 Détermination des groupes homogènes en fonction de l'année de fructification d'inflorescences femelles, des cônes (N+1) et (N+2.
Pour les inflorescences femelles l'annee à faible production a été la plus productive suivi de l'année à moyenne production puis celle à forte production. Quant aux cônes (N+1) et (N+2) l'année à moyenne production a été la plus productive en formant le ler groupe celle à faible et à forte production compose le 2ème groupe. 3.2.5.2.1.2- Interactions des critères de classification: 81
Pour les différentes interactions, une seule est significative; elle est formée par la station --année de fructification. Les résultats obtenus pour les cônes d'une année et ceux de 2 années sont portés ans le tableau N°26. Tableau N°26 :
Détermination de groupes homogènes de l'interaction
station - année
Pour le facteur cône (N+1) le tableau n°26 montre l'existence de 4 groupes homogènes; quant aux cônes (N+2) nous obtenons 3 groupes. La meilleure fructification est donnée par l'exposition favorable est la bonne année semencière. Ces interactions montrent qu'une bonne année de fructification associée à l'exposition favorable, favorise le développement des inflorescences femelles jusqu'au stade cônes. 3.2.5.2.2- Par branche 3.2.5.2.2.1-Les facteurs de classification : - Les stations Pour chaque caractère étudié, deux groupes homogènes se dégagent : - Pour les cônes (N+1) et les cônes (N+2) la station à exposition Nord est plus productive que celle exposée au Sud.Quant aux inflorescences femelles, c'est la station Sud qui produit plus que la station Nord avec respectivement une moyenne de 27.23 individus pour 22.03.Cependant, il n'y a pas de différence entre les deux stations pour les inflorescences femelles (Cf. tableau n° 27) Tableau N°27 : Détermination des groupes homogènes de station par la production des inflorescences femelles , les cônes (N+1) et (N+2).
82
Exposition Sud 7.83 5.14 B - Les branches : Pour ce critère nous obtenons également une séparation entre les branches hautes et les branches basses, pour les trois caractères étudiés. Les branches hautes sont les plus productives (Cf. Tableau n°28) Tableau N° 28 :
Détermination des groupes homogènes de branches par
la production des
- L'année de fructification : Les résultats sont portés dans le tableau n°29. Pour les inflorescences femelles, l'année à faible production a été la plus productive, suivi de l'année à moyenne production puis, celle à faible à forte production. Tableau N°29 : Détermination des groupes homogènes de station par les inflorescences femelles les cônes (N+1) (N+2).
Quant aux cônes (N+1) l'année à moyenne production a été la plus fructifère, l'année à forte production a été une année moyenne. L'année à faible production a été la plus productive pour les inflorescences femelles a donné la plus faible moyenne. Pour les cônes (N+2) l'année à forte production a été la plus productive suivi de l'année à moyenne production puis celle de l'année à faible production. 3.2.5.2.2.2 - Interactions des critères de classification. Pour les interactions entre les différents facteurs étudiés (stations- branches et années de fructification) pour les trois caractères étudiés (inflorescences femelles, cônes (N+1) et cônes (N+2), nous avons obtenu une seule interaction significative. Elle est formée entre 83 Deuxième partie Chapitre III : Estimation de la fructification et de la production des organes de reproduction femelles l'année de fructification et l'exposition pour le caractère cônes (N+1) et cônes (N+2).Les trois groupes homogènes obtenus se séparent nettement. Cônes (N+1) Le tableau n° 30 montre que pour ce caractère 3 groupes homogènes se dégagent. - Le premier groupe formé par l'année à moyenne production et l'exposition Nord est le plus productif avec 21 cônes (N+1). Il est composé par l'année la plus productive en cônes et l'exposition favorable. - Le second groupe, se compose de trois interactions : la première est formée par l'année à moyenne production et l'exposition Sud, avec 12,43 cônes, c'est à dire l'année la plus fructifère et l'exposition la plus défavorable. La deuxième interaction est formée par l'année à forte production et l'exposition Nord avec 9,50 cônes. Cette interaction se compose d'une année de fructification moyenne et d'une exposition favorable. La troisième interaction est formée par l'année à forte production (année de fructification moyenne) et l'exposition défavorable (versant Sud) avec 88.77. - Le troisième et dernier groupe se compose de deux interactions: l'année à faible fructification - exposition Nord et l'année à faible production - exposition Sud avec des moyennes respectives de 3,27 et 2,30 cônes. Tableau n°30 :
Détermination de groupes homogènes de l'interaction année
de fructification-
Cônes (N+2) Pour ce caractère le tableau ci-dessus indique l'existence de trois groupes homogènes formés par les interactions suivants : année de fructification-station Le premier groupe : année à forte production -- exposition Nord avec 20.97 cônes constitue l'interaction la plus fructifère. Le premier groupe: année à forte production--exposition Sud forme la seconde meilleure interaction avec 12.43 cônes. Le troisième groupe est composé de 4 interactions : année à moyenne production --exposition Nord (3.27 cônes), année à moyenne production -- exposition Sud (2.30 cônes), année à faible production- exposition Nord (1.20 cônes) et année à faible production - exposition Sud (0.70 cônes) Tableau n°31:Détennination de groupes homogènes de l'interaction année de fructification- station des cônes (N+2) par le test de NEWMAN et KEULS.
84
Cette interaction montre que pour les cônes (N+1) et (N+2) une forte année de fructification associée à une station favorable donnent les meilleurs résultats : c'est la somme de deux facteurs positif. Par contre l'association de deux facteurs défavorables a permis d'avoir les plus mauvais résultats. 3.2. - Discussion : Les paramètres pris en compte, en l'occurrence la station, l'année de fructification et la branche jouent un rôle très important sur l'ampleur de la floraison et de la fructification. Les peuplements de faible densité (exposition Sud) portent des arbres généralement avec des houppiers développés, d'où une surface réceptive à la lumière importante. Ce qui se traduit par une floraison plus importante sur l'exposition Sud par rapport à celle du Nord. BOUDY (1950), PUJOS (1964), TOTH (1978 a) signalent l'importance de la lumière sur la floraison du cèdre de l'Atlas. LAÏMOUCHI (1996) note que le débourrement chez le cèdre de l'Atlas semble être affecté par les températures minimales que par les températures maximales. 85 Deuxième partie 22,03 11,26 Chapitre III : Estimation de la fructification et de la D Expositon Nord 27,23 production des organes de reproduction femelles
44,77
26,4 16,72 9,13 2,73 2,78 16,7 0,98 2,78
9,5 8,77 12,43 12,43 Fructification- Exposition 3,27 2,3 3,27 2,3 1,27 0,7 Cônes( N+1) Cônes (N+2) Fig.25: Nombre moyen des inflorescences, des cônes (N +1) et (N+2) en relation avec l'exposition l'année de fructification, la position des fruits dans l'arbre et l'interaction fructification-exposition sur une période de trois mois L'influence des facteurs macro et microclimatiques et l'importance de la nutrition minérale sur la fructification est prépondérante (MATHEWS in OSWALD, 1984).Ainsi dans un essai de fertilisation LE TACON et al (1977) ont montré que l'apport de 200 kg d'azote et 150kg de phosphore par hectare à un peuplement de hêtre âgé de 120à 140 ans a un effet considérable sur la fructification. La fertilisation permet d'augmenter le nombre de fleurs males de 70% et surtout de multiplier le nombre d'inflorescences femelles par 2,7 et de multiplier le nombre de graines pleines par 3,4.Ce gain semble surtout être du à l'azote. Les années de fructification abondante sont souvent communes à tous les arbres forestiers (SEEGER in OSWALD, 1984).Tous les auteurs s'accordent sur l'effet néfaste des gelées tardives au moment de la floraison ; les fleurs et les d'inflorescences femelles sont détruites par les gelées de --1° à --3° C Une bonne année de fructification se produit une même année généralement dans une très vaste zone. Les différentes observations font apparaître que les conditions climatiques de l'été ont une grande importance sur l'abondance de la fructification. Ainsi HOLMS-GAAR et al (in OSWALD, 1984) ont démontré par expérimentation qu'un stress hydrique, mois de juin et juillet assez chauds et ensoleillés, favorisent la formation des bourgeons florifères. Par ailleurs, il apparaît d'après la valeur du coefficient de variation que l'année de forte ou moyenne floraison, se traduit par une initiation florale de l'ensemble des arbres. Mais pour une année de faible production, c'est uniquement certains arbres qui ont la capacité de produire un nombre assez appréciable des inflorescences femelles. La production des cônes, quelque soit leurs stades, est meilleure sur l'exposition Nord que Sud. La variation de cette production en fonction de l'année de production montre que les arbres du cèdre de l'Atlas alternent leur production. L'interaction entre l'année de fructification et l'exposition pour les cônes (N+1) et (N+2), nous permet de suggérer que le facteur station et significatif quand l'année de fructification est moyenne à forte, c'est à dire quand l'année de fructification est faible pour une exposition, elle l'est pour toutes les autres. 86 A la lumière de ces résultats, on peut dire que si certains arbres produisent chaque année des inflorescences femelles sur au moins 3 années, la plupart alternent leur floraison une année sur deux ou une année sur trois. L'importance de cette variation pour une même année est due probablement à la qualité de la station. En effet, les arbres de la station Nord sont les mieux pourvues en ressources et donc susceptibles des plus fortes productions.Ainsi, SABATIER et al (1993) soulignent que dans un environnement peu propice, la taille des individus et leur degré de ramification tendent à diminuer, ceci s'accompagne d'un appauvrissement de l'architecture du cèdre et donc des rameaux porteurs de cônes. La reprise végétative du cèdre de l'Atlas, coïncide avec la prise du poids du cônelet. Il peut exister une compétition entre les deux processus pour l'eau et les éléments nutritifs. Ainsi chez le cèdre de l'Atlas, on notera que les teneurs en azote et en phosphore des aiguilles, subissent un transfert vers les organes néoformés et les organes de reproduction. LAIMOUCHI (1996) rapporta que ces deux éléments ont tendance à diminuer au moment de la floraison et de la croissance.Ce transfert peut créer une compétition entre le cycle de reproduction et le cycle végétatif. Chez Pinus abies, une bonne année de fructification réduira le volume des cernes de 12 à 25 % (CHALOPKA et al in KROUCHI, 1995). Le phénomène de l'alternance de la fructification, chez le cèdre de l'Atlas a été signalée surtout par TOTH (1978 a, 1978 b et 1984). Cette irrégularité dans la production des cônes qui est favorable que tout les trois ans constitue un handicap pour la régénération. Les causes de l'alternance de la floraison et de la fructification seraient dues à l'arbre lui-même et/ou conditions écologiques. Néanmoins, il est difficile de cerner l'importance de chaque paramètre. On peut dire que les facteurs climatiques semblent jouer le rôle principal. Ils permettent à l'arbre de moduler sa fructification. La sécheresse joue un rôle extrêmement défavorable sur les possibilités de la fructification. La croissance du cèdre en hauteur et en diamètre est liée positivement aux précipitations de l'automne précédant et aux précipitations du début et de la fin d'été, ainsi qu'aux températures printanières (DUCREY, 1993) .Les paramètres dendrologiques (cimes, diamètre et hauteur) sont liés à la qualité de la station, elles agissent directement sur la fructification. Ainsi, chez le cèdre de l'Atlas, KROUCHI (1995) rapporte que le diamètre est bien corrélé à la fructification que la hauteur. La suprématie de la floraison et de la fructification des branches hautes sur les branches basses est à signaler quel que soit le stade atteint par l'organe de reproduction. Le paramètre branche est bien corrélé aux différentes fonctions femelles : floraison, pollinisation, développement et maturation. Ceci est dû à la lumière, en effet l'arbre adulte présente une cime constituée d'étages de branches de plus en plus ramifiées. A l'échelle d'un arbre ou d'un peuplement, les parties hautes de l'arbre sont les plus productives. On pourrait voir dans la séparation des branches, une certaine répartition de l'effort reproductif. Les branches sont indépendantes de point reproduction, chaque branche fournit l'énergie nécessaire au développement de ses fruits. Et, les branches les plus hautes sont les mieux pourvues en énergie lumineuse. 3.3-Conclusion: 87 Dans ce chapitre, nous avons étudié la floraison et la fructification du cèdre de l'Atlas par un comptage des inflorescences femelles, des cônes (N+1) et (N+2) en relation avec trois paramètres : exposition - branche - année de fructification pendant un cycle complet. Pour l'estimation de la fructification nous avons dans un premier temps utilisé la méthode de TOTH. Ceci nous a conduit à constater que les valeurs proposées par cet auteur pour la récolte des cônes dans les cédraies humides ne pourraient pas être appliquées à une cédraie sèche comme celle du Belezma. En effet, les conditions écologiques particulièrement l'humidité et les températures ainsi que l'état du peuplement ont une grande influence sur le nombre de cônes et la qualité des graines. Nous avons également proposé une méthode nous permettant d'estimer les possibilités de récoltes une année ou même deux années à l'avance pour l'exposition nord et pour l'exposition sud. La période comprise entre septembre et octobre constitue la meilleure phase pour le comptage. Les variations des différents organes en fonction des années, des expositions et des branches sont plus importantes. Ces variations qui ne perturbent pas l'ensemble des récoltes, peuvent avoir diverses origines principalement climatiques qui agissent aux divers stades du cycle. 88 CHAPITRE IV : Etude des cônes et des graines.
88 1- Introduction : Après avoir étudié :- Le cycle de reproduction du cèdre de l'Atlas ainsi que l'évolution des principaux caractères biométriques des organes de reproduction femelles, nous ont permis d'une part de déterminer l'évolution et la production des inflorescences femelles, des cônes d'une année et de deux années et de l'autre part de distinguer les principales phénophases : FloraisonPollinisation-Fécondation - Phases de croissance et de maturation -Dissémination des graines. - La production des organes de reproduction femelles ou nous avons déterminé l'estimation de la fructification et les possibilités de récolte ; les fréquences de la floraison, la production ainsi que la floraison individuelle et les prévisions de récolte. Le présent chapitre qui traite les cônes et les graines en fonction de la station, des années de fructification, des cônes et des branches. Il a pour objet l'étude de la répartition des différents types et catégories de graines dans le cône, la production des différents types de graines, la biométrie des cônes mûrs et des graines fertiles et attaquées ainsi que les relations entre les caractères biométriques des cônes (N+2) et leur production grainière. Les variables morphologiques des fruits, qui traduisent les caractères physiologiques (croissance -- capacité photosynthétique - résistance à la sécheresse...), sont très utilisées pour étudier les populations des essences forestières d'une même espèce (Cedrus atlantica M) ou d'un même genre (Cedrus) au sein d'une même station ou dans des stations différentes en relation avec certains paramètres de l'arbre et du milieu. Certaines de ces variables, comme le poids des cônes et des graines ainsi que la production qualitative des graines ont été utilisées par TOTH (1973a, 1975,1978, 1978a, 1978b) pour analyser la fructification en relation avec la régénération naturelle du cèdre de l'Atlas. TROSSET (1988) note que la régénération naturelle des forets est soumise à de nombreux aléas comme l'irrégularité de la fructification et de la production des semences, graines avortées, destructions par les insectes...CANTEGREL (1988) rapporte que la pérennité du pin à crochets est en relation avec la production grainière : destruction des graines dans l'arbre et au sol DERRIDJ (1990) en étudiant la biométrie des cônes et des graines a différencié les populations de Cedrus atlantica M. sur des territoires géographiques discontinus (Aures - Djurdjura - Ouarasenis) et au sein d'une même station (Tala - Guilef). La variable poids des graines fertiles, a été traitée par HARROUCHE (1991) pour différencier les populations de Cedrus en relation avec les paramètres du milieu. GAUSSEN (1964) a différencié le genre Cedrus en utilisant le caractère poids des cônes murs. D'autres auteurs, comme ARBEZ et al (1978), PRADAL (1979), TOTH (1982 - 1984), AUSSENAC (1984) et BARITEAU et al (1990) ont utilisé les caractères biométriques des différents organes du cèdre pour étudier sa variabilité géographique, bioclimatique et écophysiologique. KROUCHI (1995) a étudié le succès de reproduction du cèdre de l'Atlas, en utilisant comme variable la biomasse des chatons mâles, des cônelets, des cônes et la qualité grainière. 89 2- Matériel et méthode :2.1. Echantillonnage : L'étude des cônes et des graines a été réalisée sur l'exposition Nord et l'exposition Sud Elle a été subdivisée en 4 points : - La répartition des différents types et catégories de graines dans le cône. - La production des graines fertiles et vaines ainsi que les trois catégories de graines vaines (attaquées, aplaties et en forme de bec) selon l'importance de l'année de fructification et du cône lui -même. - L'influence de la position des cônes dans l'arbre sur la production des graines fertiles et attaquées. - La biométrie des cônes mûrs et celle des graines fertiles et attaquées. Pour l'étude de la production des graines fertiles et vaines ainsi que les trois catégories de graines, l'échantillonnage a porté sur deux expositions : l'exposition Nord et l'exposition Sud. Pour chacune d'elle, des récoltes ont été réalisées au hasard sur 30 arbres à raison de 4 cônes par arbre soit 120 cônes ; de cet échantillon nous tirons au hasard 30 cônes par exposition qui seront désarticulés, les graines triés et dénombrés par cône selon et l'année de fructification. La production grainière nous a permis de constater que le nombre de graines par cône varie en fonction de sa position dans l'arbre. Afin d'estimer l'importance de cette variation l'échantillonnage a été réalisé de la manière suivante : sur les mêmes arbres, n°7 à n°21 et les mêmes branches (branches hautes : BH1, BH2, BH3, ...et branches basses : BB1, BB2, BB3... ) que ceux utilisées pour les possibilités de récolte nous avons récolté trois cônes par branches soit 6 par arbre, ceci représente 90 individus par exposition. Ces cônes seront désarticulés manuellement et les graines choisies selon leurs catégories : graines fertiles et parasitées. L'étude de la production qualitative et quantitative des graines a montré que les cônes les plus dimensionnés sont les plus productifs et sont situés sur les branches les plus hautes. Ceci nous amener à étudier l'influence de la position des cônes dans les arbres sur ses caractères biométriques et la production des graines fertiles et parasitées. Après les mesures du poids et des dimensions pour chaque cône nous avons procédé à un comptage des graines fertiles et attaquées. De ces dernières, nous tirons par exposition 60 graines de chaque type pour mesurer leurs poids et leurs longueurs. Notons que les dimensions des organes de reproduction et la fructification (qualitative et quantitative) sont des variables qui reflètent les différentes stations écologiques. La notion de variables ou de caractères n'a selon BIDAUT (1971) de sens que si elle est associée à des comparaisons. Pour cela, nous avons employé principalement l'analyse de la variance. 90 2.2- Récolte des cônes :La date de récolte est importante pour l'étude des graines (poids, dimensions, germination). Elle est liée au climat (humidité et températures) et aux conditions ayant existées avant la récolte. Elle diffère donc, d'une station à une autre et même au sein d'une même station. Les récoltes se font généralement sur une surface de 5 à 10 ha lors d'une bonne année de fructification (afin d'avoir un bon brasage génétique maximum) et sur une trentaine d'arbres afin de mieux représenter la moyenne et la variabilité (PRADAL, 1979). Le même auteur note que la moyenne est bien représentée à partir d'une récolte effectuée sur au moins dix à vingt arbres. Compte tenu de la phénologie de reproduction du cèdre de l'Atlas du Belezma, la récolte des cônes murs a été réalisée durant le mois d'Octobre. Les cônes récoltés durant ce mois ont leurs écailles légèrement ouvertes, suite à l'influence répétitive de l'imbibition, du gel et du dégel. 2.3- Extraction des graines : 2.3.1- La désarticulation des cônes : C'est une opération manuelle qui consiste à tremper les cônes dans de l'eau pendant 36 h au moins. Puis, on les sèche à l'air libre durant 6 h. On les retrempe dans de l'eau une seconde fois pendant 2 h : après séchage d'une heure à l'air libre, on les désarticule ; pour cela, il faut maintenir le cône, entre les mains avec fermeté, on imprime à celui-ci un mouvement de rotation en sens inverse, afm de séparer les écailles et les graines du rachis. Cette manière de procéder, permettra au manipulateur de récupérer facilement la quasitotalité des graines. Notons que les parties hautes et basses du cône du cèdre de l'Atlas sont plus difficiles à désarticuler que la partie médiane (la plus fertile). 2.3.2- Tri de graines : Le premier tri est visuel ; il permet de séparer les graines aplaties et en forme de bec qui sont facilement reconnaissables des grosses graines. Ces dernières sont trempées par la suite dans l'éther de pétrole (1 1= 0,65 kg) afin de séparer par tri densimétrique les graines fertiles (lourdes) des graines parasitées (légères) qui surnagent. Les différentes graines ainsi récupérées sont mises dans des sachets en polyéthylène selon leurs catégories et en fonction des critères de classification pour être analysé. 2.4. Méthode : 2.4.1- Production des graines : Pour chaque cône désarticulé, nous avons procédé à un comptage après triage des différents types de graines : graines fertiles et graines vaines, puis celui des différentes catégories de graines vaines (attaquées, aplaties et en forme de bec), afin d'aborder : - L'amplitude des variations des graines selon les années de productions - Le pourcentage moyen des graines non viables (attaquées, avortées...)- L'attaque observée durant l'année de forte fructification selon la position des cones dans l'arbre. 91 2.4.2. Biométrie des cônes et des graines : Les mesures biométriques des cônes ont été faites après un séchage à l'air libre pendant 48 h. Ceci nous permettra de minimiser les risques d'erreur liés à l'humidité. En effet cette dernière diffère d'un cône à un autre selon les branches, les arbres et les stations. Quant à la biométrie des graines fertiles et attaquées après séchage à l'air pendant 2 h, nous avons déterminé la longueur et le poids, une à une de 4 lots de graines issues de l'année forte production (deux lots par exposition) : deux lots composés de graines fertiles et deux autres de graines attaquées. Le choix des caractères biométriques à déterminer est motivé par la bibliographie consultée. Ainsi pour chaque cône nous avons calculé le poids, la longueur et le diamètre dans sa partie la plus forte. Ces mesures ont été prises à l'aide d'un pied a coulisse au 1/20 mm pour les dimensions et d'une balance de précision au 1/10 du mg pour le poids. 3 - Présentation des résultats et discussion :3.1-Repartition des différents types de graines dans les cônes : La désarticulation manuelle des cônes de deux années nous a permis de relever deux types de graines : les graines fertiles et les graines vaines. Parmi ces dernières, nous avons constaté trois catégories : des graines en forme de bec, des graines aplaties et des graines parasitées ou attaquées ; elles sont d'apparence normale mais dont l'embryon est partiellement ou totalement dévoré et la présence de larve de couleur blanche. La répartition de ces graines dans le cône est la suivante : - Dans sa partie supérieure et inférieure on a trouvé surtout des graines en forme de bec. Selon OWENS et al (in DERRIDJ, 1990), les cônes des conifères sont pourvus d'écailles ovulifères à la base et dans une moindre mesure au sommet mais qui ne portent aucun ovule ou bien des ovules rudimentaires. Ces derniers ont un développement lent ou ne se développent pas et ne sont pas pollinisés. - Dans sa partie centrale, on a observé : 1-Des graines généralement de taille réduite, aplaties et sans embryon, avec des poches résinifères. Selon OWENS et al (in DERRIDJ, 1990), leur apparition serait liée à l'avortement ces ovules ou bien à des ovules anormaux. Elles pourraient être également le résultat de la non- pollinisation selon KOZLOWSKI (in ANONYME 1992). 2. Des graines de taille normale, parmi lesquelles on a : * Des graines légères qui surnagent dans l'éther de pétrole. Ce sont des graines dont l'embryon est détérioré par un Chalcidien phytophage nommé. Megastigmus suspectus var. Pinsapini HOFM, signalé notamment par TOTH (1978 a - 1980 b, 1984), ROQUES (1983) et BARITEAU (1993) ou bien probablement des graines dont l'embryon s'est desséché durant le passage du cône de sa maturité morphologique à sa maturité physiologique * Des graines lourdes et saines capables de germer si les conditions sont favorables. Chaque graine est formée par un embryon avec une tigelle, une radicule, des cotylédons et un 92
1 - Graine bec Graines aplaties Cône mûr tissu de stockage des éléments nutritifs appelé l'endosperme (tissu nourricier) et un tégument résinifère. (Cf. fig. 26)
Ces différents types et catégories de graines, produits par les cônes du cèdre de l'Atlas ont été signalés par TOTH (1978 a, 1978 b, 1980 b, 1984); (DERRIDJ 1990 et KROUCHI, 93 (1995). Nos observations nous en permis de constater que parmi les différentes graines, celles qui sont aplaties présentent le plus fort pourcentage. 3.2- Production des différents types et catégories de graines : Les données brutes sont reportées dans le tableau en annexe VI 3.2.1-Influence de l'année de fructification : Les résultats et leurs interprétations seront présentés pour les graines tout venant, fertiles, vaines et les différentes catégories des graines vaines (attaquées, aplaties et en forme de bec). Les facteurs pris en considération sont : L'exposition, L'année de fructification Le cône. 3.2.1.1- Nombre de graines par cône: 3.2.1.1.1- Graines tout venant: L'année de forte fructification a enregistre le plus grand nombre de graines par cône aussi bien sur l'exposition Nord que Sud. L'examen du tableau ci-dessus montre également que les moyennes obtenues sont variables selon l'année de fructification et l'exposition. Les faibles valeurs du coefficient de variation indiquent l'existence de faibles écarts pour une même exposition et année de production. Tableau N° 32 : Etude des principaux paramètres statistiques sur le nombre de graines total par cône.
La moyenne obtenue pour les trois années est de 136.40 graines par cône pour l'exposition Nord et 129,93 pour celle du Sud ; ce qui correspond à 133.16 pour l'ensemble de la zone. TOTH (1980 b) a obtenu pour la cédraie du Mont -Ventoux une moyenne de140 graines par cône. DERRIDJ (1990) a trouvé une moyenne de 138 graines pour l'ensemble des cédraies algériennes, celle de Tala- Guilef est de 136,48 graines tout venant par cône. Le 94 nombre de graines tout venant par cône varie d'une station à une autre et même au sein d'une station, selon DERRIDJ (1990) le nombre total de graines est en général plus important en altitude pour une même station (Babors : 110 graines par cône à 1.250 m et 172 à 1.900m). 3.2.1.1.2- Graines fertiles: Ce sont des graines saines, capables de germer et de se développer dès que les conditions sont favorables. Tableau N°33 : Paramètres statistiques sur le nombre de graines fertiles par cône.
Les résultats, mentionnés dans le tableau n°33, indiquent le nombre de graines fertiles par cône est également variable selon les années, les stations et les cônes. Il fluctue entre un minimum de 0 graine (année de faible fructification sur exposition Sud) et un maximum de 128 graines (année de forte fructification sur exposition Nord).La moyenne pour l'ensemble de la zone est de 43 graines fertiles par cône pour les 3 années. Concernant les cédraies algériennes, les valeurs données par DERRIDJ (1990) varient entre 41,25 et 138,68 graines avec une moyenne de l'ordre de 83,78 graines par cône; les cônes des hautes altitudes sont mieux pourvus en graines fertiles que les basses altitudes (Babors 59 graines par cône à 1.250 m et 107graines à 1.900 m) .Pour TOTH (1980 b) la moyenne obtenue au Mont-Ventoux est de 88 graines fertiles par cône sur une période de 7 années d'études. Le coefficient de variation montre que les écarts entre les graines fertiles par cône sont importants pour le Belezma particulièrement si l'année de production est faible et que l'exposition est défavorable. Ceci, montre que quand la fructification est faible, quelques individus seulement investissent dans les fonctions femelles. Par contre quand elle est forte, le nombre d'arbres fertiles augmente. En outre, les cônes qui ont produit peu ou pas de graines fertiles, sont maintenus sur les arbres (année de faible production - exposition Sud). TOTH (1980b) a mis en évidence, l'importance de l'année de fructification sur la production qualitative des graines : les graines sont de bonne qualité quand la fructification est bonne. Les graines fertiles constituent un paramètre intéressant pour analyser la qualité de la station et l'alternance de la fructification. Ainsi, sur les trois années d'étude l'exposition Sud (défavorable) n'a produit que 33,21 graines par cône, alors que l'exposition Nord a produit 95 53,60 graines. En outre, pour une même année de production, nous enregistrons également d'importants écarts entre les expositions. Ces graines nous permettent d'estimer la fructification et donc les possibilités de récoltes. 3.2.1.1.3- Graines vaines : Le tableau n° 34 montre que le nombre de graines vaines (graines attaquées, aplaties et en forme de bec) est plus important sur l'exposition Sud que Nord. En outre, quand la fructification est faible la production augmente : l'année de la fructification et l'exposition ont une influence certaine sur la qualité des graines. POURTET (in TOTH, 1978a) note que dans le cas des espèces résineuses, les années de mauvaise fructification correspondent à une production de graines médiocres. Tableau N°34 : Paramètres statistiques sur le nombre moyen de graines vaines par cône.
La faible valeur du coefficient de variation (CV =7.72) montre qu'une faible année de production associée à l'exposition défavorable a donné les cônes les moins fertiles pour l'ensemble des arbres. Cela montre que dans cette station, l'ensemble des arbres est dans l'incapacité de produire des graines fertiles. Alors que pour l'exposition Nord, la variabilité de la fertilité des arbres est importante (CV = 34,81 pour une faible année de production). Cela veut dire que seulement « certains arbres » sont dans l'incapacité de produire chaque année des graines fertiles ; la fertilité des arbres de la station Nord présente une variabilité : certains arbres arrivent à produire des graines fertiles par rapport à d'autres, même si l'année de fructification est faible. En outre, quand la fructification est moyenne à forte, le nombre de graines vaines diminue et la variabilité des graines entre les cônes est grande. Les résultats obtenus par TOTH (1980 b) et DERRIDJ (1990) montrent que la production des graines vaines est extrêmement variable selon les stations. TOTH (1978 a) rapporte que les années climatiques agissent sur la qualité des graines : sur une période de 5 années, le nombre moyen de graines vaines par cône est de 91 pour une mauvaise année de fructification et, il est seulement de 27 quand l'année de production est bonne. 96 Quant aux cédraies algériennes, elles sont comprises selon DERRIDJ (1990) entre 46,67 graines (Tala Guilef : 1.750m) et 131,36 (Theniet El Had). La moyenne pour les 19 stations étudiées est de 51,60 graines, ce qui représente 38,11 % par cône par rapport aux graines totales. 3.2.1.1.4 - Catégories de graines vaines: 3.2.1.1.4.1 - Graines attaquées: Les données obtenues sont indiquées dans le tableau n° 35, qui montre que globalement il n'y a pas de différence entre les deux expositions pour la même année de fructification. La moyenne obtenue pendant 3 années est de 8.37 pour la station Nord et 8.85 pour celle du Sud. Tableau N°35 : Paramètres statistiques sur le nombre moyen de graines attaquées par cône.
En outre, nous remarquons que plus la fructification est bonne, plus le nombre de graines attaquées par cône est important. Ceci serait probablement dû à une synchronisation entre le cycle des insectes et celui de la fructification. Les pullulations des insectes ravageurs de graines sont probablement en relation avec l'alternance de la fructification Notons également que l'importance des graines attaquées durant l'année de forte fructification est compensée par le nombre élevé de graines fertiles. ROQUES (1983) signale que la phénologie de reproduction des Cedrus nécessite des adaptations spécifiques au niveau du cycle des ravageurs. TOTH (1978 a et 1980 b) rapporte qu'il existe de grandes variations dans la production des graines attaquées. Ceci est confirmé par nos resultats ; ainsi les valeurs de coefficient de variation montrent que l'attaque des insectes est très variable ; elle l'est surtout sur l'exposition Sud quand la fructification est très faible (CV=130.70) : certains cônes sont plus attaqués que d'autres. Quand elle est moyenne à forte, la variation est pratiquement la même pour les deux expositions, on peut dire que les dégâts sont sensiblement les mêmes. TOTH (1978 a) signale qu'en 1973 (année de forte fructification) les dégâts causés peuvent atteindre 30 % au niveau du cône; le nombre de graines par cône varie entre 1 graine (année de forte production) et 22 graines (année de mauvaise fructification).La moyenne obtenue sur 5 années est de 48 graines. Pour le Belezma le nombre de graines attaquées est compris entre 0 (année de faible production) et 74 (année de forte production) avec une moyenne de 8,61 graines par cône. 3.2.1.1.4.2- Graines aplaties:Elles nous renseignent particulièrement sur la qualité de la fructification, c'est à dire sur l'aptitude de l'arbre à donner des fruits murs. 97 Les données portées sur le tableau n°36 montrent d'une part que les arbres ne mènent pas à terme la totalité des ovules qu'ils produisent dans la partie fertile du cône et de l'autre, que leur nombre varie en fonction de l'importance de la fructification et de l'exposition. Pour l'exposition Nord il fluctue entre un minimum de 9 et un maximum de 135 avec une moyenne de 58,26 graines par cône. Quant à l'exposition Sud, la variation est comprise entre 5 et 137 avec une moyenne de 64.06 graines. Tableau N°36 : Paramètres statistiques sur le nombre de graines aplaties.
Les valeurs du coefficient montrent l'inexistence d'écarts dans le nombre de graines aplaties quand la fructification est faible sur l'exposition défavorable ; ce qui signifie que pour l'ensemble des cônes il y a un avortement important qui serait lié à l'absence de pollinisation, à des ovules anormaux, des grains de pollen de mauvaise qualité ; c'est la résultante de deux facteurs négatifs. Quand la fructification est forte, l'investissement mâle et femelle est relativement variable selon les arbres. La moyenne nous indique par ailleurs, que plus la fructification est importante et plus le nombre de cônes fertiles augmente. On peut dire, à travers les résultats obtenus que plus l'année de production est faible, plus la qualité des graines est mauvaise; la station par son influence intervient sur la qualité. En effet TOTH (1980 b) rapporte que la qualité grainière du cèdre de l'Atlas est tributaire du milieu qui les voit s'élaborer. Pour une année de faible production ces graines représentent 48 % par cône, alors que pour une année de forte production, elles n'atteignent que 18 %. 3.2.1.1.4.3- Graines en forme de bec: Le tableau n°37 montre que leur nombre est peu important par rapport aux graines aplaties. Pour l'exposition Nord, les valeurs obtenues sur une période de trois années varient entre 02 et 59 graines par cône avec une moyenne de 15,70 graines. Quant à l'exposition Sud, le nombre moyen de graines par cône fluctue entre un minimum de 02 et un maximum de 61 avec une moyenne de 19,08. La variation de leur nombre par cône est importante. Selon l'amplitude de la fructification, la moyenne est comprise entre 11,66 et 21,60 pour l'exposition Nord.Pour l'exposition Sud, elle varie entre 14,90 et 24,60. La moyenne sur les trois années est de 17,40 graines. Ces graines sont selon OWENS et al (in DERRIDJ, 1990) une propriété inhérente à l'espèce ; leur nombre, pour le cèdre du Liban est de l'ordre de 10 par cône. Tableau N°37 : Paramètres statistiques sur le nombre de graines bec par cône.
98
3.2.1.2- Distribution des graines par cône: La distribution des graines tout venant (G.T.V), fertiles (G.F) et vaines (G.V) d'une part et les différentes catégories de graines vaines [graines attaquées (At), aplaties (Ap) et en forme de bec (B)] en fonction de l'année de production et de l'exposition est portée dans les tableaux n '38, n°39 et n°40. Dans ce paragraphe, nous allons étudier uniquement la distribution des graines fertiles. 3.2.1.2.1- Année de faible production: La distribution des graines fertiles durant l'année de faible production, est représentée uniquement par la classe (00-20) sur l'exposition Sud ; celle du Nord est mieux répartie, elle est partagée en 5 classes [(00-20) (20-40) (40-60 ) (60-80) et (80-100)] avec respectivement 30, 23.23 ,13.33 , 30 et 6.66 %. Tableau N°38 : Distribution des graines par exposition selon l'année à fructification faible.
3.2.1.2.2- Année de moyenne production: Pour l'exposition Sud les classes (00-20) et (20-40) sont les mieux représentées avec respectivement 43.33 et 40% soit 83.33 % ; quant à l'exposition Nord les classes les mieux représentées, sont comprises entre [(20-40), (40-60) et (60-80)] avec respectivement 30, 33.33 et 30 % ce qui représentent 93.33% Tableau N°39: Influence de l'année de fructification sur la distribution des graines par exposition.
3.2.1.2.3- Année de forte production: 99 La distribution des classes est consignée dans le tableau n°40. Les graines fertiles sont réparties en plusieurs classes. Pour l'exposition Sud, on a observe 6 classes à savoir [ (00-20), (20-40),(40-60), (80100),(100-120) (120-140)] ; quant à l'exposition Nord, le nombre de classes est 7 [(2040),(40-60),(80-100),(100-120)(120-140)(140-160)] ; nous notons l'absence de la plus faible classe et la présence d'une nouvelle grande classe .La classe la plus représentative est la (6080) avec 30% pour l'exposition Sud et 40% pour l'exposition Nord. Tableau N°40: Influence de l'exposition et de l'année à forte fructification sur la distribution des graines.
3.2.1.2.4- Conclusion : A la lumière des résultats obtenus, on peut dire que plus la fructification est forte et plus la distribution des graines fertiles est répartie en plusieurs classes d'une part et d'autre part, les classes les plus importantes sont les mieux représentées. En outre pour une même année de fructification, l'exposition Sud est indiquée par les classes les plus faibles. 3.2.1.3- Corrélation entre les différents types de graines: Cette étude a pour objet la détermination d'éventuelles relations linéaires entre le nombre de graines tout venant (NC), fertiles (NF), vaines (GV) attaquées (GA) aplaties (GP) et en forme de becs (GB) sur deux expositions pendant une durée de 3 années. La matrice de corrélation portée en annexe XXII, renferme 6 variables à 30 observations chacune correspondant à 30 cônes. Les corrélations retenues comme significatives sont égales ou supérieures à 0,3610 pour 5% ; 0,4629 pour 1% et 0,5703 pour 1 Les figures 27, 28 et 29 illustrent les différentes corrélations. La figure 30 représente graphiquement les corrélations entre les graines fertiles et les graines vaines 3.2.1.3.1- Année de faible production: Pour la station à exposition Nord, la production des graines fertiles est négativement corrélée à celle des graines vaines (r = -0,922; P = 0,001; N = 30) et à celle des graines aplaties (r = -0,935; P = 0,001; N = 30) d'une part et de l'autre celles des graines vaines est positivement corrélée aux graines aplaties au seuil de 1 %o (r =0,976; N=30). Quant à l'exposition Sud, nous avons obtenu également une liaison linéaire positive entre les graines vaines et aplaties (r = 0,870; P = 0,001; N = 30). 100 En outre, au seuil de 1 % le coefficient obtenu entre les graines vaines et les graines tout venant est de 0,505 pour l'exposition Nord. Quant à l'exposition Sud, elle est hautement significative (r = 0,925). La corrélation entre les graines aplaties et tout venant est significative au seuil pour l'exposition Nord (r = 0,423) et hautement significative pour l'exposition Sud (r = 0,781). Les graines becs (16.60 graines par cône) et attaquées (6.71 graines par cône) sont peu importantes ; la majeure partie des graines appartient à la catégorie des aplaties : 92.53 graines dont 114.56% pour l'exposition Sud et 82.56 % pour l'exposition Nord.
0.505 .925 0.976 :935
Exposition Sud : H Exposition Nord : H Fig. 27 : Corrélations entre les graines durant l'année de faible production. 3.2.1.3.2- Année de moyenne production: La corrélation entre les graines aplaties et vaines est hautement significative pour l'exposition Nord au seuil de 1 %o (r = 0,839, N = 30) et l'exposition Sud (r =0,822, N = 30) Par ailleurs, la relation entre les graines aplaties et le nombre de graines totales est positivement significative pour l'exposition Sud au seuil de 1% (r = 0,568) et significative au seuil de 5% pour l'exposition Nord (r = 0,450). En outre, sur l'exposition Sud, nous avons obtenu une corrélation négative entre les graines fertiles et les graines vaines (r = - 0,366; P = 0,05; N = 30), l'exposition nord est de (r = - 0,466; P = 0,05; N = 30). Notons l'existence de relation significative entre les graines vaines et les graines tout venant pour les deux expositions. (Exposition Nord 0.510 et Exposition Sud : 0.839) Nous constatons que les graines aplaties représentent 50% du nombre total des graines.
0.450
H H Fig. 28 : Corrélations entre les graines durant l'année de production moyenne. J.L.1.3.3- Annee ue wrte irucuncauon: Au seuil de 1%0, une liaison négative est obtenue entre les graines fertiles et les graines vaines pour l'exposition Nord (r = -704; N = 30) et celle du Sud (r = -0,677; N = 30). Aussi, les graines totales et fertiles sont fortement corrélées (r = 0,472; P = 0,01; N=30) pour l'exposition Sud. On peut aussi observer que les graines aplaties sont corrélées positivement aux graines vaines pour les deux stations (station Nord : r = 0,678 ; station Sud : r = 0,577) au seuil de 1 %o). Les graines aplaties par rapport aux graines totales ne représentent que 26.36 % pour l'exposition Nord, alors que celle du Sud indiquent un fort pourcentage avec 53.76.
Exposition Sud : H H 4-- Fig. 29 : Corrélations entre les graines durant l'année de forte production. 3.2.1.3.4 - Conclusion Les différentes relations ont mis en évidence l'importance de l'année de fructification et de la station sur l'aptitude de l'arbre à produire des graines. Cette production, qui présente une alternance annuelle, résulte de l'impossibilité pour l'arbre de mener à terme les bourgeons floraux jusqu'au stade fruit. Les arbres répartissent probablement leur énergie entre les deux cycles. Ainsi, durant l'année de faible production, l'ensemble des arbres des deux expositions sont incapables de mener à terme l'essentiel des ovules initiés : les graines vaines sont principalement constituées des graines aplaties (ovules avortés ou anormaux).Quand elle est moyenne, les graines vaines sont également des graines aplaties ; c'est surtout sur l'exposition Sud que les arbres ont été incapables de donner des graines fertiles. Durant l'année de forte production les cônes ayant initié le plus grand nombre de graines au départ, auront le taux final de graines fertiles le plus élevé ; autrement dit, la majorité des arbres des deux stations 102 ont assuré le développement optimum des ovules jusqu'au stade graine. En prenant l'année de faible production comme référence égale à 100%, nous observons que l'alternance s'applique aux cônes et aux fertiles graines. (Cf. tableau ci-dessous) Tableau n ° 41 : Alternance de la production selon l'exposition.
Les mécanismes (compétition ou inhibition ?) restent mal connus, mais l'alternance affecte tous les processus physiologiques de l'arbre (COURANJOU, 1982). 103 Exposition Nord
40 60 80 100 120 140 GV
40 60 80 GV 100 120 140 160 140 120 100 80 u_
140 160 180 110 90 70 if 50 30 10 10 120 Graines Vaines Faible production : GF = 109.42 --0.8020 GV Moyenne production : GF = 78.195 -- 0.3978 GV Forte production : GF = 129.84 --0.6842 GV
o 0 GV Faible production : GF = 14.166-0.0688
GV R = 0.1776 20 40 60 80 100 120 140 160 GV Moyenne production : GF = 20.444 -- 0.05512 GV 110 120 130 140 150 160 170 22 u_ 10 2 o 0- o o 00 0 o '0, Régression IC à 95% 40 60 80 120 140 160 180 100 GV Moyenne production : GF = 126 -- 0.7115
GV R = - 0.6672 140 100 80 LL 60 60 u_ 30 20 10 -Q o o o o o o o. Régression IC à 95% o o 0 0 Exposition Sud
3.2.1.4 - Méthodes statistiques: 3.2.1.4.1- Analyse de la variance:Cette analyse, a pour but l'étude de l'influence de trois facteurs de classification, la station, l'année de fructification et le cône (N+2) ainsi que leurs interactions sur la production grainière. Pour faciliter, l'interprétation des résultats, nous avons étudié séparément la production des : - Graines tout venant - Fertiles et vaines - Trois catégories de graines vaines : attaquées - aplaties- en forme de bec. Les résultats de l'analyse de la variance sont reportés dans le tableau ci-dessous. Tableau N°42 : La production des graines tout venant, des graines fertiles et des graines vaines. Influence de la station de l'année de production et du cône et leurs interactions.
3.2.1.4.1.1- Graines tout venant: 1- Pour les critères de classification. - Station (F 1): La production totale des graines par cône n'est pas significative. - L'année de fructification (F2) : La différence est hautement significative. - Le cône (F3) : Le troisième critère nous permet de constater qu'il n'existe pas de différence significative entre les cônes. 104 2- Pour les interactions des critères de classification. Elles nous renseignent sur l'influence simultanée de deux facteurs sur le nombre de graines tout venant. Les résultats obtenus sont : - Station - année de fructification (F1 - F2). Le F test est supérieur à F théorique. La production des graines tout venant est fortement influencée par cette interaction. - Station - cône (F1 - F3) et cône - année de fructification (F2 - F3), la différence n'est pas significative. 3.2.1.4.1.2- Graines fertiles et graines vaines: Cette analyse au seuil de 5 % fait apparaître pour les deux critères de classification a savoir la station, l'année de fructification et le cône d'une part et leurs interactions de l'autre part des différences significatives pour les deux caractères : graines fertiles et des graines vaines.
3.2.1.4.1.3- Catégories de graines vaines : attaquées - aplaties - becs. Les résultats obtenus pour les trois critères de classification et leurs interactions par l'analyse de la variance au seuil de 5 %, sont reportés dans le tableau ci-dessus. Tableau N°43: Influence de la
station, de l'année de fructification et du cône sur la
production
Les trois facteurs étudiés, la station et l'année de fructification et le cône font ressortir des différences significatives pour les trois caractères : graines attaquées, aplaties et becs
105
3.2.1.4.2- Le test de NEWMAN et KEULS : Afin de dégager des groupes homogènes les résultats obtenus par l'analyse de la variance sont soumis au test de NEWMAN et KEULS seuil de 5 %. 3.2.1.4.2.1- Les graines tout venant: Ce test a permis de déterminer des groupes homogènes uniquement pour le critère année de fructification et pour l'interaction station - année de fructification. 1-Critères de classification : La production des graines tout venant se compose pour le facteur année de fructification, de trois groupes homogènes qui se séparent nettement (Cf. Tableau n°44). - Le premier groupe, qui représente l'année de forte production, donne les meilleurs résultats avec 152,57 graines. -Le second groupe, formé par l'année de faible production, cumule une moyenne de 132,67. - Le troisième et dernier groupe, représenté par l'année de production moyenne, donne la plus faible production avec seulement 114,27 graines. Tableau N°44:Détermination des groupes homogènes de l'année de fructification par la production des graines tout venant. Méthode de NEWMAN et KEULS.
2- Interaction des critères de classification : Cette interaction a permis de déterminer trois groupes homogènes qui se discriminent L'année de fructication semble être le facteur le plus important par rapport à l'exposition. (Cf. Tableau n°45) Tableau N°45 : Détermination des groupes homogènes de l'interaction exposition - année de fructification par la production des graines tout venant.
- Le premier groupe est représenté par deux interactions: station à exposition Nord - année de forte production et station à exposition Sud - année de forte fructification avec respectivement des moyennes de 154,73 et 150,40 graines tout venant par cône. 106 - Le second groupe est formé par trois interactions : exposition Sud -- année de faible fructification, exposition Nord - année de faible fructification et exposition Nord - année de moyenne production. Les moyennes de graines tout venant par cône sont respectivement de 138,60, 127,73 et 126,73. - Le troisième et dernier groupe est composé d'une seule interaction, l'exposition Sud - année de moyenne production avec seulement 100,80 graines tout venant. 3.2.1.4.2.2- Graines fertiles et vaines: Les groupes déterminés au seuil de 5 % pour les graines fertiles et vaines en fonction de la station, de l'année de fructification et de leurs interactions sont : 1- les critères de classification: Station : Ce critère fait ressortir deux groupes homogènes (Cf. Tableau n°46). - Pour les graines fertiles, le premier est formé par la station Nord avec 53,63 graines par cône et le second par la station à exposition Sud avec 33,21 graines par cône. Tableau N°46: Détermination des groupes homogènes de station par la production des graines fertiles. Méthode de NEWMAN et KEULS.
- Quant aux graines vaines, la production est plus importante sur l'exposition Sud, qui forme le premier groupe avec 96,76 graines. Le second groupe avec 82,77 graines est constitué par la station à exposition Nord. Tableau N° 47:Utilisation de la méthode de NEWMAN et KEULS pour la détermination des groupes homogènes de station par la production des graines vaines.
- Année de fructification : Les tableaux ci-dessous, indiquent que le test de NEWMAN et KEULS a donné trois groupes homogènes pour le caractère graines fertiles et deux pour celui des graines vaines. Graines fertiles : Les trois groupes obtenus sont formés par l'année de forte production, l'année de moyenne production et l'année de faible production. Les valeurs moyennes de graines fertiles par cône correspondant aux années de production sont respectivement 72,37 , 35,73 et 22,13. Tableau N°48:Détermination des groupes homogènes de l'année de fructification par la production des graines fertiles. Méthode de NEWMAN et KEULS.
Graines vaines: 107 Nous obtenons deux groupes homogènes pour ce caractère : le premier est composé par l'année à faible et a moyenne production avec respectivement 110,53 graines par cône. Le second groupe est formé par l'année à moyenne et à forte production avec 80,17 et 78,58 graines vaines par cône. Tableau N°49: Caractérisation des groupes homogènes de l'année de fructification par la production des graines vaines. Méthode de NEWMAN et KEULS.
2- Interaction des critères de classification: Pour ces interactions, seules celles formées par la station et l'année de fructification ont permis de dégager des groupes homogènes. Les graines fertiles : Les groupes homogènes dégagés au nombre de 4 sont formés par les interactions suivantes (C.f Tableau ci- dessous): Tableau N° 50 : Détermination des groupes constituer par l'interaction station - année de fructification pour la production des graines fertiles.
- Exposition Nord - année de forte production avec 76,03 graines fertiles par cône et exposition Sud - année de forte production avec 68,67. - L'exposition Nord - année de moyenne production et exposition Nord - année de faible production forme le second groupe avec respectivement une production de 45,47 et 39.30. - Le troisième groupe formé par l'exposition Sud et l'année de moyenne production avec 26 graines par cône. - Le dernier groupe est composé par l'exposition défavorable et la faible année de production avec seulement 04,97 graines fertiles. Les graines vaines : Les deux groupes obtenus au seuil de 5 % se discriminent nettement (Cf. Tableau n°51). - Le premier groupe est composé par une seule interaction : exposition défavorable (Sud) - année de faible fructification avec 133,63 graines. - La seconde est formée par 5 interactions : Exposition Nord - Année de faible fructification. Exposition Sud - Année de forte fructification.Exposition Nord - Année de moyenne fructification.Exposition Nord - Année de forte fructification.Exposition Sud - Année de moyenne fructification. 108 Tableau N°51 : Détermination des groupes formés par l'interaction station - année de fructification par la production des graines vaines. Méthode de NEWMAN et KEULS au seuil de 5 %.
3.2.1.4.2.3- Catégories de graines vaines: Le test de NEWMAN et KEULS au seuil de 5 % a permis de dégager des groupes homogènes pour les différentes catégories de graines vaines en fonction des critères étudies: 1- Critères de classification: - Les graines aplaties Elles se discriminent en deux groupes, pour le critère station : Le premier groupe, est formé par la station à exposition Sud (défavorable), le second par la station à exposition Nord (favorable) avec respectivement 69,87 et 58,27 graines aplaties par cône (Cf. Tableau n° 52 ). Tableau N°52 : Etude des groupes homogènes de station par la production des graines aplaties.
Le critère année de fructification. -Les graines aplaties : Elles se séparent en trois groupes homogènes : année de faible fructification, année de moyenne fructification et de forte fructification (Cf. Tableau n° 53). Tableau N°53: Détermination des groupes homogènes en fonction de l'année de l'année de fructification des graines aplaties.
- Graines attaquées : Le nombre moyen de graines attaquées par cône se compose de trois groupes homogènes : le premier groupe est représenté par l'année de forte fructification (16,33 graines) le second par l'année de moyenne production (7,75 graines), le troisième par l'année de faible production avec une moyenne de 1,77 graines (Cf. Tableau n° 54). 109 Tableau N°54 : Détermination des groupes homogènes en fonction de l'année de fructification des graines attaquées.
53,6 58,26 69,86 33,21 Graines tout venant Graines fertiles Graines aplaties 8,37 8,85 I 1 i Graines attaquées 15,71 19,08 I Graines bec
I I I i I I I I I I I I I I I 1 Poids(g) Longueur(cm) Diamètre(cm) Nbre graines Nbre graines fertiles attaquées mination des groupes homogènes en fonction de l'année de fructification des graines formes de bec. Tableau N°55: Détermination des groupes homogènes en fonction de l'année de fructification des graines formes de bec.
Le premier est formée par l'année de forte fructification avec 22,85 graines par cône, l'année de faible et de moyenne production compose le second groupe avec respectivement 16,07 et 13,28 graines par cône. 2- Interaction des critères de classification: L'interaction du critère station (F1) et celui de l'année de fructification (F2), a donné pour les graines aplaties trois groupes homogènes qui se discriminent de la manière suivante : - Le premier groupe est représenté par une seule interaction, qui se compose de l'exposition Sud - année de faible production avec une moyenne de 114,57 graines aplaties par cône. - Le second groupe représenté par deux interactions : exposition Nord - année de faible de production et exposition Nord - année de moyenne production; le nombre de graines aplaties est de 70,50 pour la première interaction et 63,50 pour la seconde. -Le troisième groupe est formé par trois interactions : Exposition Sud - année de moyenne production avec 50,43 graines aplaties par cône, Exposition Sud - année de forte production: 44,60 graines et Exposition Nord - année de forte production: 40,80 graines. Tableau N° 56 : Détermination des groupes homogènes par l'interaction station - année de fructification par la production des graines aplaties.
111 Cette interaction montre qu'un facteur de station (extrinsèque) défavorable à la production grainière comme l'exposition Sud, associé à une mauvaise année de fructification (intrinsèque) compromet la régénération. 3.2.1.4.3- Conclusion : La production des différents types de graines (graines fertiles et vaines) et catégories de graines vaines (aplaties, attaquées et bec) dépend de l'année de fructification facteur lié à l'arbre et l'exposition, variable de station ; elles constituent le meilleur critère pour l'étude de la variabilité quantitative et qualitative des graines ; le facteur cône n'est pas significatif. Ainsi, quelle que soit l'année de fructification, la qualité grainière est meilleure sur l'exposition Nord et, chaque année les arbres ajustent leurs productions selon les conditions écologiques. Ainsi, durant l'année de forte fructification, moyenne le nombre de graines fertiles par cône est respectivement de 76.03, 45.47 et 39.30 pour l'exposition Nord ; quant à l'exposition Sud , il est de 68.67, 26 et 4.97. 3.2.2 - Influence de la position des cônes dans l'arbre sur la production des graines fertiles et attaquées :Cette étude qui traite la production des graines fertiles et attaquées par cône selon leur position dans l'arbre a porté uniquement sur l'année à forte production. Rappelons que le nombre de cônes échantillonnés est de 90 par exposition. Les résultats sont portés en annexe VI. 3.2.2.1 - Production des graines par cône : 3.2.2.1.1 - Graines fertiles : L'examen du tableau n°57 indique que pour l'ensemble de la station le nombre de graines fertiles par cône selon les branches présente de grands écarts : les valeurs oscillent entre 1 et 123 graines, avec une moyenne de 38,74 graines. Pour l'exposition Nord, on enregistre une moyenne de 56,40 graines fertiles pour les cônes des branches hautes et 42,20 pour ceux des branches basses. Pour la station à exposition Sud, la moyenne par cône est pratiquement le même aussi bien pour les branches hautes que les branches basses.L'exposition Nord est 1,75 plus productive que l'exposition Sud. Les moyennes obtenues sont de 49,30 graines pour 28,19. Le facteur branche joue un rôle peu important sur la qualité des graines particulièrement pour l'exposition Sud Tableau N°57: Etude la production des graines fertiles par cône en fonction des branches.
Les valeurs du coefficient de variation sont élevées et sensiblement les mêmes pour chaque exposition. Néanmoins l'exposition Sud présente les plus importantes valeurs.Elles indiquent, que les écarts de la production des graines fertiles par cône sont importants aussi bien pour les branches hautes que basses, surtout pour la station Sud. Ceci montre que le nombre de graines fertiles par cône est très variable pour le même type de branche quelle que soient l'exposition, et les branches. Ainsi donc, on peut dire que la fertilité des arbres est variable même au sein d'une station. 112 3.2.2.1.2 - Les graines attaquées : Le tableau n°58 montre que pour l'ensemble de la station, le nombre de graines attaquées varie entre 0 et 42 individus, avec une moyenne de 9,24 graines. Tableau N°58 : Paramètres statistiques sur la production des graines attaquées par cône
Les cônes des branches hautes sont plus attaqués que ceux des branches basses, quelle que soit la station. En outre les cônes de l'exposition Sud, sont légèrement plus attaqués que ceux de l'exposition Nord. Les moyennes obtenues pour les deux branches sont respectivement de 10,35 graines par cône, pour 8,14 graines par cône.Les valeurs des coefficients de variation, montrent que certains cônes sont plus attaqués que d'autres, particulièrement ceux des branches basses de l'exposition Sud (CV = 94,71 %). ROQUES (1983) signale que le taux d'attaque peut atteindre 95 % des cônes. Les variations sont liées à la situation du cône dans l'arbre (10 fois plus d'attaques au sommet du houppier) et aux années de faible fructification. Dans le cas d'une forte production, ce taux oscille entre 1 et 9 % seulement. 3.2.2.2 - Corrélations entre les graines fertiles et attaquées : La matrice de corrélation (Cf. Tableau n°80 page 135) qui détermine la relation linéaire qui peut exister entre deux variables, est composée de 10 variables (le poids, la longueur et le diamètre des cônes; le nombre moyen des graines fertiles et attaquées pour les branches hautes et les branches basses) à 45 observations chacune, correspondant à 3 cônes par branche pour 15 arbres par exposition. Pour connaître, la valeur de r, à partir de laquelle, cette relation linéaire est significative on applique le test de STUDENT au seuil de 5% : r = 0,2875 ; 1% r = 0,3721 et r = 0,4647pour 1°/00. Il existe une liaison positive et significative entre les graines fertiles des branches hautes et celles des branches basses au seuil de 1 %o. Les valeurs de r obtenues sont égales à 0,766 pour l'exposition Nord et 0,528 pour celle Sud. Quant aux graines attaquées, la liaison entre les branches est significative à 1 %o que pour l'exposition Sud (r = 0,556). Notons enfin, que les graines fertiles des branches hautes sont corrélées à celles des branches basses (r = 0,558), pour l'exposition Sud.(Cf tableau n ° 80 ) 113 26 co) 22 ti) 18 _c 214 ..a) 10 cr < ar É 2 2 Exp Nord : GFBB = 12,529 + 0,53665 GFBH
Régression ICà95% E E 4° ' a) u_ ..... ..... a),.... .....0 c i....- "Fa 20 C9 100 ar m ar co) _c c m 0 20 40 60 80 100 120 140 Graine Fertile - Branche Haute Exp Nord : GABH = 8,4557 + 0,25957 GABB
0 0 0 `0, Régression ICà 95% r -, 0 0 0 0 o 0 o: 0 0 0 0 0 0 0 O 0 -2 2 6 10 14 18 22 26 Graine Attaquée - Branche Basse
Exp Sud : GFBB = 14,654 + 0,50890 X GFBH Exp Sud : GABB = 1,4542 + 0,65879 X GABH 110 'p., Régression ru 90 5 -5 0 5 10 15 20 25 30 110 40 Graine Fertile Branche Haute Graine Attaquée Branche Haute Fig. 32 : Corrélation entre les graines fertiles et attaquées selon les branches. 3.2.2.3.1 - L'analyse de la variance : L'étude des graines fertiles et attaquées en fonction de trois critères de classification (exposition - arbre - branche) et leurs interactions est reportée dans le tableau ci-dessous. Tableau N°59: Etude des caractéristiques des fertiles et attaquées de cèdre de l'Atlas en relation avec l'exposition les arbres, les branches et leurs interactions.
114
Les trois facteurs de classification, la station (F1), l'arbre (F2), la branche (F3) et les interactions font ressortir des différences significatives pour les deux caractères étudiés. Graines fertiles FACTEURS Graines vaines Facteur 1: Expostion Hautement significatif Très significatif Facteur 2: Arbre Facteur 3: Branche Très significatif Très significatif Significatif Significatif INTERACTIONS DES FACTEURS Facteurl - Facteur 2 Significatif Significatif Significatif 3.2.2.3.2 - Le test de NEWMAN et KEULS : Cette méthode, a permis au seuil de 5 % de dégager des groupes homogènes en fonction des critères de classification et de leurs interactions. 1 - Les facteurs de classification : - Station : Le tableau n°60 montre que pour les graines fertiles et les graines attaquées, les groupes homogènes se discriminent nettement. L'exposition Nord a une production moyenne de 49,30 graines fertiles par cône, alors que celle du Sud n'est que de 28,20. Quant aux graines attaquées, le premier groupe avec une moyenne de 9.56 graines par cône constitue l'exposition Nord. Le second groupe est formé par l'exposition Sud avec 8.14graines. Tableau N°60:Etude des groupes homogènes de stations par le nombre moyen de graines fertiles et attaquées. Test de NEWMAN et KEULS.
Les résultats obtenus montrent, que les graines attaquées représentent par rapport aux graines fertiles 19,39 % pour l'exposition Nord et 28,86 % pour celle du Sud. - Les branches : Comme pour le critère station, les deux groupes obtenus se discriminent également (Cf. Tableau n°61). 115 Tableau N°61:Etude des groupes homogènes et de branches par le nombre moyen de graines fertiles et attaquées. Test de NEWMAN et KEULS.
Le premier groupe est formé par les branches hautes avec une production moyenne de 42,03 graines fertiles par cône, le second par les branches basses avec 35,47 graines. Quant aux graines attaquées, nous obtenons les mêmes groupes homogènes : les branches hautes avec une production moyenne de 10,38 graines par cône et les branches basses avec 7,32 graines. Cette analyse indique que les graines attaquées représentent 24,69 % par rapport aux graines fertiles pour les branches hautes et 20,63 % pour les branches basses. - Arbre : Le critère arbre est constitué par la moyenne des graines fertiles et des graines attaquées des deux branches. Il s'agit donc de la moyenne de 6 cônes. Ce test donne pour ce critère, 12 groupes homogènes pour le caractère « graine fertile » et 11 pour celui des « graines attaquées » (Cf.tableau n°62). Remarque : Les groupes homogènes obtenus pour le critère arbre et l'interaction station - arbre présentent des chevauchements difficiles à interpréter, c'est l'inconvénient de la méthode de NEWMAN et KEULS Tableau N°62: Détermination des groupes homogènes des arbres et/ou de l'interaction arbre- Exposition par le nombre des graines fertiles et attaquées par cône. Méthode de NEWMAN et KEULS. 116
117 Compte tenu de l'importance des critères exposition et branche sur la production des graines fertiles, il est évident d'avoir des groupes homogènes qui se discriminent. En effet, au seuil de 5 %, le test utilisé a permis de dégager trois groupes homogènes (Cf. Tableau n°63). Le premier groupe est formé par l'interaction exposition Nord - branche haute avec une production moyenne de 56,40 graines par cône.Le second groupe se compose de l'exposition Nord- branche basse avec 42,20 graines.Le troisième groupe est représenté par deux interactions : l'exposition Sud - branche haute et l'exposition Sud - branche basse avec respectivement une production moyenne de 28,73 graines et 27,67 graines par cône. Tableau N°63: Détermination des groupes de l'interaction station - branche par le nombre moyen de graines fertiles par cône. Méthode de NEWMAN et KEULS.
- Station -- Arbre : Pour ce type d'interaction, les groupes homogènes pour les graines fertiles et pour les graines attaquées sont reportés dans tableau n°62 3.3 - La biométrie des cônes et des graines :Les travaux effectués par de nombreux chercheurs GAUSSEN (1964 et 1967), ARBEZ et al (1978),TOTH (1978 a ;1980b),PRADAL(1979) FERRANDES,(1986), DERRIDJ( 1990) et KROUCHI( 1995) ont montré l'importance des caractères biométriques pour différencier le genre Cedrus , des races ou provenance de Cedrus atlantica M et pour étudier la fructification, la production grainière , la germination des graines, en relation avec la station, la position des cônes dans l'arbre, la hauteur et le diamètre de l'arbre ... 3.3.1 - Influence de la position des cônes sur leurs caractères biométriques: Les données brutes sont reportées en annexe VII 3.3.1.1 - Le poids et les dimensions des cônes : 3.3.1.1.1 - Le poids : Les données obtenues (Cf. Tableau n°64) montrent que le poids des cônes varie selon sa position dans l'arbre. Ainsi, pour une même exposition le poids moyen des cônes des branches hautes est supérieur à celui des branches basses particulièrement pour l'exposition Nord. 118 Le poids moyen des cônes de l'exposition Nord est de 48,56 g pour les branches hautes et 43,04 g pour les branches basses. Quant à l'exposition Sud il est respectivement égal à 42,71 g et 41,93g.La moyenne générale est de 44,06 g. Le poids minimal est toujours supérieur à17g Tableau N°64 : Paramètres statistiques sur le poids des cônes par branche selon l'exposition.
Les valeurs du coefficient de variation pour ce paramètre indiquent que les écarts de la biomasse sont relativement peu importants. DERRIDJ (1990) note que le poids moyen oscille entre 41,49 g (Telmet - Aurès) et 56,91 g (Babors 1900m), avec une moyenne générale pour les cédraies algériennes de 55,83 g. A Tala- Guilef, le poids moyen obtenu par KROUCHI (1995) est de 60 g. Celui des cédraies de l'Atlas Tellien est supérieur à celui de l'Atlas Saharien (GAUSSEN, 1964; DERRIDJ, 1990). 3.3.1.1.2 - La longueur et le diamètre : Ces deux caractères sont généralement peu étudiés par rapport au poids, car ils ne sont pas très significatifs. 3.3.1.1.2.1 - La longueur : Le tableau n°65 des paramètres statistiques sur la longueur des cônes mûrs indique que les variations se comportent à peu près de la même manière que le poids : pour une même station la longueur moyenne des cônes issus des branches hautes est plus importante que celles des branches basses d'une part, d'autre part ceux de l'exposition Nord soient plus longs que ceux de l'exposition sud. Tableau N°65 : Paramètres statistiques sur la longueur (cm) des cônes par branche en relation avec l'exposition.
Les valeurs du coefficient de variation montrent l'inexistence de grands écarts pour ce caractère. 119 Pour le Belezma, l'intervalle de variation est compris entre 3.57 et 8.61cm, avec une moyenne de5.65cm. DERRIDJ (1990) donne comme intervalle de variation: 5,52 - 6,88 cm ou il note que les stations de l'Atlas saharien ont les longueurs les plus faibles d'une part et que d'autre part les stations situées à des altitudes élevées (cédraies claires) présentent les longueurs les plus grandes. Pour les cédraies françaises, la moyenne trouvée par TOTH (1971) et GEORGE (1980) est comprise entre 5 et 8 cm. 3.3.1.1.2.2 - Le diamètre : Comme pour la longueur, le diamètre moyen des cônes de l'exposition Nord (4.47cm) est plus important que celui de l'exposition Sud (4.37cm) et que pour une même exposition les cônes des branches hautes sont plus gros que ceux des branches basses. Tableau N°66 : Paramètres statistiques sur le diamètre des cônes par branche et par station.
Les coefficients de variation, nous montrent qu'il n'y a pas d'écart entre les cônes, le coéfficient de variation compris entre 8.79 et 12.63%). Comme pour le caractère longueur DERRIDJ (1990) constate que ceux de l'Atlas Saharien présentent les diamètres les plus faibles (4,41 cm), ceux de Tala- Guilef sont plus importants (5,69 cm). L'intervalle de variation donné par GEORGE (1980) est compris entre 4 et 5 cm. Celui de Belezma est compris entre 3,20 et 5,98 cm avec une moyenne de 4,42 cm 3.3.1.2 - Distribution des cônes par classe de poids et de dimensions : Les variations entre les branches, pour le poids, la longueur et le diamètre des cônes selon l'exposition sont portées en annexe VIII et représentées par des histogrammes de classe. Pour chaque classe, nous avons également représenté le nombre moyen des graines fertiles et attaquées. 3.3.1.2.1 - Le poids : Le mode de distribution est consigné dans la figure n°33 1 - L'exposition Nord : 120 La distribution pour ce caractère est la suivante : Les classes 30 - 40 g, 40 - 50 g et 50 - 60 g caractérisent mieux cette exposition avec respectivement 23,33 %, 27,77 % et 17,77 % des cônes pesés. Les classes 60 - 70 g et 20 - 30 g occupent 14,44 et 8,88 %. En outre, ceux qui représentent mieux les branches hautes présentent un mode pour les classes 30 - 40 g, 40 - 50 g, 50 - 60 g et 60 - 70 g. La première classe occupe 26,66 %, la troisième occupe 22,22 %, la deuxième et la quatrième occupent 20 % chacune. Les basses branches sont représentées par quatre classes occupant 80% des cônes : 20 - 30 g (13,33 %), 30 - 40 g (20 %), 40 - 50 g (35,55 %) et 50 - 60 (13,33 %). 2 - L'exposition Sud Le mode le plus représentatif pour l'ensemble des cônes des branches hautes et basses est composé par les classes 20 - 30 g, 30 - 40 g, 40 - 50 g et 50 - 60 g qui occupent respectivement 15,55 %, 26,26 %, 23,33 % et 20 % représentant 85 % des cônes échantillonnés. Pour ceux situés sur lesbranches hautes les classes de poids 20 - 30 g, 30 - 40 g, 40 - 50 g et 50 - 60 g sont les mieux représentées avec par ordre 17,77 %, 24,44 %, 17,77 % et 22,22 %.Les classes de poids 30 - 40 g, 40 - 50 g et 50 - 60 g avec respectivement 28,88 %, 28,88 % et 17,77 % de l'effectif représentent les cônes des branches basses. 3 - Station : Exposition Nord et Sud Les classes les mieux représentées sont comprises entre 30 - 40 g, 40 - 50 g et 50 - 60 g qui occupent respectivement 25 %, 25,55 % et 18,88 % soit 70 % environ de l'ensemble des cônes étudiés. Les classes 20 - 30 g et 60 - 70 g occupent 12,12 % et 11,11 %, alors que celles de faibles dimensions (10 - 20 g) et de fortes dimensions (70 - 80 g) ne s'accaparent que 3 à 4 % environ. Les branches hautes présentent un mode pour les classes 30 - 40 g, 40 - 50 g et 50 - 60 g, qui s'octroient 80 % des cônes échantillonnés. Les classes 20 - 30 g, 30 - 40 g, 40 - 50 g et 50 - 60 g sont les mieux représentées des branches basses avec un total de plus de 85 %. 3.3.1.2.2 - La longueur : La distribution de ce caractère est reportée dans la figure N°34 1 - L'exposition Nord : La distribution des classes de longueur de l'exposition Nord est déterminée par le mode suivant: 40 - 50 cm, 50 - 60 cm, 60 - 70 cm et 70 - 80 cm: ces classes occupent respectivement 25,5 %, 31,11 %, 25,55 % et 14,44 % de l'échantillon. 121
Deuxième partie : Chapitre IV : Etude des cones et des graines Station Sud 5- 0 n n Station Nord Branches hautes 20 - 15- 10- 5- 10_20'20_30'30_40'40_50'50_60'60_70'70_80' 0 n n 30 - 25 - Station Nord et Sud Fig. 33 : Distribution du poids des cônes de cèdre de l'Atlas. Influence de la station et de 122 Branches hautes 20 - 15 - n n 0 10 20.20 30.30 40.40 50.50 60'60 70.70 80' 30 - 25 - 10 - 5- Station Sud 10_20'20_30'30_40'40_50'50_60'60_70'70_80' Station Nord Branches hautes et basses 25 - 20 - 15 - 10 - 5- 10_20 2 _30 30_40 40_50 50_60 6 _ 0 70_80 . 0 n n 30 - 25 - 20 - 15- 10- Branches hautes 30 - Les branches hautes sont caractérisées par 4 classes de longueur : 40 - 50 cm, 50 - 60 cm 60 - 70 cm et 70 - 80 cm. La première et la quatrième occupent chacune 20 % de l'effectif total, alors que la deuxième et la troisième s'accaparent respectivement 31,11 % et 28,88 %. Quant aux branches basses on a le même mode de classe; les fréquences relatives obtenues sont de 31.33 % pour les deux premières classes, la troisième et la quatrième classe n'occupent que 22.22% et 8.88% 2 - L'exposition Sud Cette exposition est représentée par les classes 40-50 cm,50-60 cm et 60-70 cm avec des fréquences relatives de 33,33% ,27,77% et 34.44% pour l'ensemble des cônes étudiés. Pour les branches basses c'est le même mode de classe qui les caractérise : 40-50 cm, 50-60 cm et 60-70 cm. Chaque classe occupe 30% environ de l'effectif. 3 -- Station : Exposition Nord et Sud: Pour l'ensemble de la station, le mode de classement le plus représentatif correspond aux classes 40 - 50 cm, 50 - 60 cm et 60 - 70cm avec 29,44% pour les premières classes et 30 % pour la troisième. Les branches hautes de la station sont caractérisées par les classes 40 - 50 cm, 50 - 60 cm, 60 - 70 cm et 70 - 80 cm. Elles occupent 26,66 %, 28,88 % 31,11 % et 13,33 %. Les classes 40 - 50 cm, 50 - 60 cm et 60 - 70 cm qui représentent chacune 30 % des cônes mesurés appartiennent aux branches basses. 3.3.1.2.3 - Le diamètre : La distribution des classes de diamètre est reportée dans la figure N°35. 1 - L'exposition Nord : La classe de diamètre 40 - 50 cm caractérise la station à exposition Nord avec 77,77 % des cônes mesurés. Les classes 30 - 40 cm et 50 - 60 cm n'occupent que 11,11 % chacune. Les branches hautes et basses sont mieux représentées par classe 40-50cm avec respectivement 82,22 % et 73,33 %. 2 - L'exposition Sud : C'est la classe 40 - 50 cm qui représente l'exposition Sud avec une fréquence relative de 70 %. Les classes 30 - 40 cm et 50 - 60 cm n'occupent que 20 % et 10 % environ. Les branches hautes et basses sont également mieux représentées par la classe 40 - 50 cm avec respectivement 63,82 % et 75,55 %. Les classes 30 - 40 cm et 50 - 60 cm occupent 23 % et 13 % pour les branches hautes et pour les branches basses on a 18% et 7%. 123 Deuxième partie : Chapitre IV : Etude des cones et des graines
-10 -20 -30 -40 Station Nord Branches hautes et basses Station Sud Branches hautes et basses 0 40 - 35 - 30 - 25 - 20 - 15 - 10 - 5- 40 - 35 - 30 - 25 - 20 - 15- 10- 5- 0 ri . i--i . . . . 30 _ 40 40 _ 50 50 _ 60 60 _ 70 70 _ 80 80 _ 90 . Station Nord n 30 _ 40 40 _ 50 50 _ 60 60 _ 70 70 _ 80 80 _ 90 Station Sud
40 Branches basses et hautes 30 20 10
Branches basses et hautes 30 _ 40 40 _ 50 50 _ 60 60 _ 70 70 _ 80 80 _ 90 Station Nord et Sud
124 Branches hautes
100 - 80 - 60 - 40 - 20 - 0
Station Sud 100 -
Station Nord
30 _40 40 _ 50 50 _ 60 11- Station Sud
90 - 80 - 70 - 60 - 50 - 40 - 30 - 20 - 100 80 - 60 - 40 - 20 - 0 -20 - -40 - -60 - -80 - -100 - 80 - 70 - 60 - 50 - 40 - 30 - 20 - 100 30 _40
50 50 _ 60 branches hautes branches hautes et basses 40 _ 50 50 _ 60 Station Nord et Sud
3 -- Station : Exposition Nord et Sud: 125 La distribution par classe de diamètre est représentée par une classe comprise entre 40 - 50 cm qui occupe 73 % de l'effectif. Les branches hautes et basses sont également caractérisées par la même classe, elles occupent 68,47 % et 78,88 % des cônes. La classe 30 - 40 cm est mieux représenté pour les branches hautes (19,56 %) que basses (12,22 %). La classe 50 - 60 cm est plus présente sur les branches hautes (11,95 %) que basse (8,88 %). 3.3.1.2.4- Conclusion La station Nord et les branches hautes représentent les classes dominantes quel que soit le caractère étudié. L'existence une étendue de variation dans le poids ; cette variation s'exprime selon les conditions dans lesquelles évolue le peuplement. L'allure générale des histogrammes représentant les trois caractères poids, longueur et diamètre est celle de GAUSS : une ou plusieurs classes représentant les dimensions les moins fréquentes séparées par les plus fréquentes. 3.3.1.3 - Méthodes statistiques : 3.3.1.3.1 - L'analyse de la variance : Cette analyse a pour objet l'étude des caractères biométriques des cônes mûrs (poids, longueur et diamètre) en fonction des facteurs (station - arbre - branche) et leurs interactions en testant l'hypothèse nulle, c'est - à - dire que les groupes sont issus d'une même et unique population. Le Tableau n°67 montrent que les cônes (N+2) présentent des variations pour les trois caractères biométriques au seuil de 5 %. Tableau N°67:Etude des caractéristiques des cônes de cèdre de l'Atlas en fonction des stations des arbres, des branches et leurs interactions.
3.3.1.3.1.1 - Les critères de classification : 126 Pour ces trois caractères (poids, longueur et diamètre) des différences obtenues sont significatives pour les 3 critères adoptés : Facteurl -Station; Facteur2 - Arbre ; Facteur 3- Branche 3.3.1.3.1.2 - Interaction des critères de classification : La même analyse a donné les résultats suivants : Le poids : Facteur 1 - Facteur 2 : Hautement significative. Facteur 1 - Facteur 3 : Hautement significative. Facteur 2 - Facteur 3 : Non significative. Facteur 1 - Facteur 2 - Facteur 3 : Significative. La longueur : Facteur 1 - Facteur 2 : Hautement significative. Facteur 1 - Facteur 3 : Non significative. Facteur 2 - Facteur 3 : Non significative. Facteur 1 - Facteur 2 - Facteur 3 : Significative Le diamètre : Facteur 1 - Facteur 2 : Hautement significative. Facteur 1 - Facteur 3 : Significative. Facteur 2 - Facteur 3 : Significative. Facteur 1 - Facteur 2 - Facteur 3 : Non significative. 3.3.1.3.2 - La méthode de NEWMAN et KEULS : En complétant l'analyse de la variance par cette méthode, nous avons obtenu des ensembles homogènes au seuil de 5 % pour les trois critères de classification étudiés : station - arbre - branche. 3.3.1.3.2.1 - Les facteurs de classification : 1 - Station Les trois caractères forment des groupes homogènes, qui se discriminent nettement. Les cônes mûrs de l'exposition Nord sont plus dimensionnés que ceux de l'exposition Sud Tableau N°68 : Détermination des groupes homogènes de stations, par le poids, la longueur et le diamètre. Méthode de NEWMAN et KEULS.
2 - Les branches. La même analyse a permis d'obtenir des groupes homogènes pour les trois caractères. Le tableau n° 69, indique que les cônes mûrs de deux années des branches hautes sont plus lourds, plus longs et plus gros que ceux des branches basses. En effet, pour le poids nous enregistrons respectivement une moyenne de 45,65 g et 42,49 g. 127 La longueur obtenue est de 5,75 cm pour les branches hautes et 5,53 cm pour les branches basses ; quant au diamètre, il est respectivement de 4,48 cm et 4,37 cm. Tableau N°69:Détermination des groupes homogènes de branches par le poids, la longueur et le diamètre. Test de NEWMAN et KEULS.
3 - Les arbres: Pour ce critère, nous avons utilisé la moyenne des cônes de la branche haute et de la branche basse. Les arbres de 1 à 15 appartiennent à l'exposition Nord, ceux de l'exposition Sud portent les numéros de 16 à 30. Le test de NEWMAN et KEULS au seuil de 5 % relève 22 groupes pour le poids, 15 pour la longueur et 20 pour le diamètre. 3.3.1.3.2.2 - Interactions des critères de classification : 1 - Facteur station - Facteur branche : Cette interaction n'est significative que pour le poids et le diamètre ; complétée par le test de NEWMAN et KEULS, nous obtenons les résultats suivants : Les deux groupes homogènes dégagés sont formes par 4 interactions.Le premier groupe est composé par l'exposition Nord - branche haute avec 48,58 g pour le poids et 4,57 cm pour le diamètre. Le second groupe et composé de trois interactions : Exposition Nord - branche basse (43.04g - 4.38cm), exposition Sud - branche haute (42.71g -- 4.39cm) et exposition Sud - branche basse (41.94 -- 4.36 cm). Tableau N°70: Détermination des groupes homogènes de l'interaction station - branche par le poids et le diamètre. Test de NEWMAN et KEULS.
128 Sud -- branche basse 4,36 129 Tableau n° 71 : Détermination des groupes homogènes des arbres et / ou l'interaction exposition - arbre par le poids, la longueur et le diamètre des cônes.
Légende : VN : Versant Nord VS : Versant Sud A : Arbre 2 - Arbre - exposition: Pour ce type d'interaction les résultats sont les suivants : poids, diamètre et longueur présente respectivement 22, 20 et 15 groupes homogènes. L'inconvénient de la méthode de NEWMAN et KEULS est qu'elle ne permet pas d'éviter les difficultés d'interprétation, résultants du chevauchement de groupes homogènes. 3.3.2 - Influence de la station sur le poids et la longueur des graines fertiles et attaquées :Les données brutes sont représentées en annexe IX 3.3.2.1 - Caractères biométriques : L'évaluation des principaux caractères biométriques nous a permis de constater que le poids moyen des graines fertiles est deux fois supérieur à celles attaquées. Pour la longueur la différence entre les deux types de semences est minime. 3.3.2.1.1 - Les graines fertiles : Le tableau ci-dessus résume les principaux paramètres des graines fertiles pour les stations Tableau N°72 : Paramètres statistiques sur le poids et la longueur des graines fertiles en fonction de la station.
Il montre que le poids et la longueur moyenne d'une graine fertile produite par l'exposition Nord (0,086 g pour 1,23 cm) sont supérieurs à celle de l'exposition Sud (0,083 g pour 1,14 cm). La moyenne pour les deux expositions est de 0,084 pour 1,18 cm. Il n'y a pas d'écart entre le poids et la longueur des graines fertiles, les valeurs du coefficient de variation sont très faibles. DERRIDJ (1990) rapporte que pour l'année 1987, le poids moyen des graines fertiles varie entre 0,059 g (Tala - Guilef) et un maximum de 0,105 g ( Meurdja).Pour TOTH (1980 b) le poids moyen est compris entre 0,089 g (année de moyenne production) et 0,046 g (année de faible production).La moyenne obtenue au Mont-Ventoux sur 7 années est de 0,068g KROUCHI (1995) en étudiant le poids de graines fertiles selon les arbres a trouvé un poids moyen compris entre 0.061 et 0114g avec une moyenne de 0.083g et un CV de 728%, ce qui montre l'existence de très grands écarts entre les arbres. Ces résultats montrent que le poids des graines fertiles, varie en fonction de la station ; l'année semencière pourrait être également importante. 130 3.3.2.1.2 - Les graines attaquées : Le poids des graines attaquées est compris entre 0,029 g et 0,061 g avec une moyenne de 0,043 g pour l'exposition Nord. Celle de l'exposition Sud oscille entre 0,027 g et 0,054 g avec une moyenne de 0,042 g. Pour la longueur elle est de 1.19 cm pour l'exposition Nord et 1.11cm pour l'exposition Sud. La moyenne pour les deux expositions est de 1,15 cm. Tableau N°73 : Paramètres statistiques sur le poids et la longueur des graines attaquées.
Le coefficient de variation, montre pour ses faibles valeurs qu'il n'existe pas d'écarts pour les deux caractères particulièrement la longueur, pour une même exposition. 3.3.2.1.3-Correlation entre le poids et la longueur. Les liaisons linéaires retenues comme significatives au seuil de 1 %o, sont celles égales ou supérieures à la valeur r = 0,4143 (DDL = 58). Elles montrent l'existence de corrélation entre le poids et la longueur (r = 0,514, pour l'exposition Nord, r = 0,564 pour l'exposition Sud). L'étude des corrélations (Cf. Tableau n°74) indique que pour une même exposition, le poids et la longueur présentent des liaisons linéaires au seuil de 1 %o (r = 0,4143): exposition Nord; r = 0,602 : exposition Sud = 0,668. Tableau N°74:Matrice de corrélation du poids et de la longueur desgraines fertiles et attaquées Influence de l'exposition.
DDL: 58 Légende : P : Poids. L : Longueur. V. N : Versant Nord. G.F : Graines fertiles. G.A : Graines attaquées. V. S : Versant Sud. 131 3.3.2.2 La distribution des classes de poids et de longueur des graines fertiles : Les classes de poids et de longueur sont portées dans le tableau n°75. La représentation graphique de la distribution du poids est portée dans la figure n°36. Tableau N°75:Influence de l'exposition sur la distribution des classes de poids et de longueur des graines fertiles du Cèdre de l'Atlas.
Pour le poids, la classe 0,0700 à 0,0999 g est la plus représentative. Elle occupe 96,66 % des graines fertiles de l'exposition Nord et 86,66 % pour l'exposition Sud. Les autres classes de faibles et de fortes dimensions sont moins présentes.
96,66 10 86,66 5,83 91,66 2,5 3,33 1,66 1,66
Fig. 36 : Distribution ( %) du poids moyen d'une graine fertile selon l'exposition. Quant au caractère longueur la classe 1,20 - 1,30 cm individualise l'exposition Nord avec une fréquence de 63 % environ. L'exposition Sud est représentée par la classe 1,10 - 1,20 avec une fréquence de 58,06 %. Cette distribution montre que pour les deux caractères nous obtenons une classe dominante séparée par une ou plusieurs classes de faibles dimensions. 132 En prenant en considération l'influence du poids sur la germination (Cf. 3ème partie -- chapitre I ) nous pouvons dire que les possibilités de régénération du cèdre de l'Atlas de l'exposition Nord sont meilleures que celles du Sud. 3.3.2.3 - Méthodes statistiques : 3.3.2.3.1 - L'analyse de la variance : Pour comparer le poids et la longueur des graines fertiles et attaquées issues de deux stations nous avons utilisé l'analyse de la variance. Cette dernière nous permet de déterminer d'éventuelles différences entre les graines de même type d'une part et, entre les graines fertiles et attaquées de l'autre part. 3.3.2.3.1.1 - Graines fertiles : Les différences sont significatives pour le caractère longueur et diamètre selon le critère exposition. 3.3.2.3.1.2 - Graines attaquées : Si pour le caractère poids, nous n'obtenons aucune différence significative selon le facteur station (F test < F thé) en revanche nous enregistrons des différences pour le caractère longueur. 3.3.2.3.1.3 - Graines fertiles et graines attaquées : Pour les deux caractères étudiés, les différences obtenues sont significatives entre les deux types de graines aussi bien pour l'exposition Nord que celle Sud. Les résultats de ces quatre analyses sont reportés dans le tableau ci-dessus Tableau N°76 : Etude de l'influence de la station et du type de graines surle poids et lalongueur des graines fertiles et attaquées.
3.3.2.3.2 - Le test de NEWMAN et KEULS : 3.2.2.3.2.1 - Le facteur station : Les résultats obtenus sont reportés sur le tableau n°77 et n°78. 1 - Graines fertiles : 133 Les graines fertiles produites par l'exposition Nord forme le premier groupe homogène avec une moyenne de 0,09 g pour 1.24 cm. Celle de l'exposition Sud forme le second avec 0,08 g pour 1.15 cm par graine. Tableau N°77: Détermination des groupes homogènes de station par le poids et la longueur des graines fertiles.
2 - Graines attaquées : Si le caractère poids, forme un seul groupe (il n'y a pas de différences entre les deux expositions), celui de la longueur a donné deux groupes homogènes Le premier groupe est formé par la station Nord avec une moyenne de 1,19 cm, le second par la station à exposition Sud avec 1,12 cm. Tableau N°78: Détermination des groupes homogènes de station par la longueur des graines attaquées.
3.3.2.3.2.2 - Le facteur type de graines : Pour ce caractère, les groupes homogènes de poids et de longueur se discriminent en fonction de la station. 1 - Station Nord : Les graines fertiles qui forment le premier groupe homogène sont plus lourdes (0,09 g) et plus longues (1,24cm) que les graines attaquées qui constituent le second groupe avec 0,04g pour 1,19 cm. Tableau N°79: Détermination des groupes homogènes de graines fertiles et attaquées par le poids et la longueur selon la station. Méthode de NEWMAN et KEULS.
2 - Station Sud : Comme pour l'exposition Nord, deux groupes se discriminent pour les deux caractères. 134 Le premier groupe est formé par les graines fertiles aussi bien pour le poids que pour la longueur (0,08 g pour 1,15 cm). Le second groupe est représenté par les graines attaquées avec un poids moyen de 0,04g pour une longueur moyenne de 1,12 cm. 3.4 - Etude des corrélations : 3.4.1 - La matrice de corrélation : Cette matrice est composée de dix variables par exposition : le poids (P), la longueur (L), le diamètre des cônes (D) d'une part et le nombre de graines fertiles (GF) et attaquées (GA) de l'autre part selon la position des cônes dans l'arbre. Tableau N°80 : Matrice de corrélation sur les dimensions des cônes et des graines fertiles et attaquées. Influence de la position des cônes sur l'arbre (branche basse et branche haute).
Légende : P : Poids du cône. BB : Branche Basse. GA : Nombre de graines attaquées L : Longueur du cône. BH : Branche Haute. GF : Nombre de graines fertiles. D : Diamètre du cône. Elle étudie par station les relations qui peuvent exister entre : - les dimensions des cônes 135 - les dimensions des cônes et le nombre des deux types de graines Les valeurs de l'étude de corrélation totale retenue comme significative au seuil de 5 %, 1 % et 1 %0 sont respectivement égale ou supérieur à 0,287 ; 0,372 et 0,467. 3.4.2 - Corrélation entre les caractères morphologiques des cônes : Les relations linéaires obtenues sont hautements significatives : - Exposition Nord Les branches basses. Les valeurs obtenues au seuil des 1 %o entre les poids et la longueur, le poids et le diamètre sont respectivement égales à 0,804 et 0,909 ; entre le diamètre et la longueur sont 0,707. Pour les branches hautes les valeurs sont égales à 0,766 ; 0,882 ; 0,617. - Exposition Sud : Les relations pour les branches basses sont respectivement égales à 0,824 entre le poids et la longueur ; 0,879 entre le poids et le diamètre et 0,666 entre la longueur et le diamètre. Celles des branches hautes sont respectivement égales à 0,852 ; 0,895 et 0,732. 3.4.3-Corrélation entre les graines et les caractères morphologiques des cônes : Les corrélations entre le nombre de graines fertiles et les caractères morphologiques des cônes sont consignés dans le tableau n°80, qui montre que ces liaisons sont très significatives à hautements significatives selon la branche et l'exposition. 3.4.3.1 - L'exposition Nord : Le poids, la longueur et le diamètre des cônes sont positivement corrélés à la production des graines fertiles au seuil de 1 %o. Les valeurs obtenues au seuil de 10/00 sont respectivement égales à 0,848 - 0,720 et 0,701 pour les cônes des branches hautes, et 0,688 - 0,651 - 0,540 pour ceux des branches basses. 3.4.3.2 - L'exposition Sud : Les liaisons linéaires obtenues entre le poids et le nombre de graines fertiles sont significatives au seuil de 1 %o pour les cônes des branches hautes (r = 0,666) et de 1 % pour ceux des branches basses (r = 0,432). Quant au caractère longueur, elles sont significatives au seuil de 1 %o aussi bien pour les branches hautes (r = 0,662) que pour les branches basses (r = 0,476). La corrélation entre le diamètre et les graines fertiles obtenue est significative au seuil de 1 % (r = 0,445) pour les cônes des branches hautes, quant aux cônes des branches basses, elle n'est significative qu'au seuil de 5 % (r = 0,355). 3.5-DISCUSSION -- CONCLUSIONAprès la désarticulation manuelle des cônes, les graines obtenues sont de 2 types : les graines fertiles et les graines vaines. Ces dernières sont composées de trois catégories : aplaties, attaquées et en forme de bec. Les graines fertiles et attaquées sont localisées surtout dans la partie centrale du cône. 136 L'étude de la production grainière par cône est représentée par la figure n°37 ; elle montre que l'année de la fructification et la station influencent d'une façon significative la qualité et la quantité des graines : La production totale des graines par cône est très importante quand la fructification est forte, par contre, elle est peu variable entre les stations quand l'année de fructification est faible à moyenne. Celle des graines fertiles est également variable et plus importante sur l'exposition Nord et durant l'année de forte fructification : elles nous renseignent sur l'état de la cédraie Le nombre de graines vaines d'une part et celui des différentes catégories varie également d'un cône à un autre selon l'année et la station. Aussi, quand l'année de production est forte on assiste à une importante production de graines fertiles, ces dernières sont négativement corrélées aux graines tout venant. Ceci nous permet de dire que chez le cèdre de l'Atlas, la majorité des ovules donneront des graines fertiles (la pollinisation et la fécondation atteignent l'optimum) Durant les années de faible à moyenne production, les graines vaines sont composées essentiellement de graines aplaties; ces dernières occupent la quasitotalité des graines totales (tout venant) surtout quand la station est défavorable. Elles ont tendance à diminuer au profit des graines fertiles, au fur et à mesure que la fructification augmente, particulièrement quand les conditions climatiques sont favorables. L'estimation des graines aplaties (graines sans embryon) est très importante pour l'étude de la régénération naturelle du cèdre de l'Atlas. Leur existence est due à l'avortement des ovules (non-pollinisation et /ou non-fécondation) est le résultat de la compétition entre le cycle de reproduction et végétatif, l'état de santé et l'âge de l'arbre ... cette catégorie de graines pourraient nous renseigner sur la qualité de la station et l'état du peuplement. Au Belezma le nombre de ces graines est très important quand la fructification est faible surtout sur l'exposition Sud. Il apparaît l'existence d'un système de compensation entre la production des graines fertiles et aplaties. Autrement dit, le cèdre de l'Atlas, tend à moduler sa production grainière en fonction de la disponibilité des ressources de la station (facteurs extrinsèques) et de l'arbre lui- même (facteurs intrinsèques). Ces facteurs qui caractérisent généralement tous les arbres (forestiers et fruitiers) sont considérés comme étant une adaptation quelles que soient les conditions écologiques : c'est le phénomène de l'alternance entre la fructification et l'accroissement. Cette alternance qui affecte selon COURANJOU (1982) un grand nombre ou la totalité des mécanismes physiologiques qui se traduisent par une diminution de la surface foliaire durant l'année de forte fructification. Ainsi, au cours de nos différentes observations sur l'estimation de la fructification, il n'est pas rare de rencontrer des pieds de cèdre qui portent peu ou pas de cônes mais, sont pourvus d'une importante surface foliaire. Outre, l'année de fructification, la qualité de la station joue également un rôle très important sur la qualité et la quantité des semences. TOTH (1978 a ; 1980 b) rapporte que la production des cônes et la qualité physiologique et biologique des graines de Cedrus atlantica M sont liées aux conditions climatiques et la sylviculture mise en oeuvre durant toute la 137 période du cycle de reproduction ; le nombre de graines dépend du milieu BOUDY (1950) ; il résulte aussi du houppier, du diamètre et de la hauteur de l'arbre : elle est d'autant plus importante quand la cime est plus étalée et éclairée être donc plus espacée (LEE in KROUCHI,1995 ). DERRIDJ,(1990 ) a mis en évidence l'influence de la station sur la production qualitative et quantitative des graines entre les cédraies humides et sèches, ainsi que l'altitude pour une même station. Des études réalisées par KROUCHI (1995) ont montré l'existence de très grandes variabilités dans la production des graines tout venant et des graines fertiles pour une même station. Au Belezma, l'exposition Nord est caractérisée d'une part, par un ensoleillement journalier et saisonnier favorable et d'autre part, d'un froid nocturne et matinal ; ces deux éléments qui influencent fortement l'ensemble des paramètres favorables au développement des graines de cèdre : Floraison -- pollinisation -- maturation -- dissémination et germination. A l'échelle du cône, le nombre de graines attaquées est variable selon la station, l'année de fructification et la position des cônes dans l'arbre. Leur importance est liée aux fluctuations des cônes (hôte) donc à l'alternance de la fructification et aux facteurs climatiques. ROQUES (1983) a montre qu'il existe une étroite corrélation entre la levée des diapauses et l'intensité de la floraison de l'année, cette diapause peut durer 5 années consécutives. Aussi, à travers les résultats obtenus, nous avons noté que plus la production est forte et plus le nombre de graines attaquées est important. Les pullulations des insectes ravageurs de cônes et de graines suivent, selon ROQUES (1983) l'importance de la fructification. L'insecte ravageur des graines s'est adapté au cycle de fructification du Cèdre de l'Atlas : plus le taux de graines attaquées et important, par rapport aux graines fertiles, et plus l'année de fructification est bonne. En outre, le nombre de graines attaquées est relativement plus important sur les branches hautes que les branches basses. Ceci est dû aux facteurs climatiques ou le paramètre lumière, semble jouer le principal rôle ROQUES (1988). L'influence de ces insectes est susceptible de jouer un rôle non négligeable dans la régénération des essences forestières d'altitude. Ainsi, divers auteurs CANTGREL (1988), DEMOLIS (1988) OTT (1988), SKRZYPCZYNSKA (1988) et TRO S SET (1988) ont insisté sur l'importance de l'impact des insectes ravageurs de graines sur la régénération des peuplements forestiers dans les zones tempérés. TOTH (1975a, 1978b 1980c) a souligné l'influence sur la régénération, des ravageurs de graine dans les cedraies artificielles. MOUNA (1993) a répertorié au niveau des cédraies naturelles du Maroc 4 insectes inféodés aux fleurs mâles, aux inflorescences femelles, aux cônes et aux graines. L'étude des caractères biométriques (le poids, la longueur et le diamètre) des cônes en relation avec la station et la position des cônes dans l'arbre est la figure n° 37 synthétise les principaux résultats ; ces derniers nous a permis de constater l'existence d'écarts, particulièrement le poids. Ainsi, ceux de l'exposition Nord sont plus importants que ceux de l'exposition Sud. Par ailleurs, pour une même exposition, les paramètres biométriques des cônes des branches hautes sont les plus dimensionnés, que ceux des branches basses.. L'écart obtenu pour chaque caractère nous permet de constater que le poids apparaît comme étant le meilleur critère pour spécifier les populations. En outre les cônes dont la 138 biomasse est importante possèdent non seulement les dimensions les plus importantes mais sont les plus fertiles (initiation précoce des inflorescences femelles, station favorable forte fructification...). Le poids des cônes murs présentent de grandes variations selon DERRIDJ (1990) les cônes des cédraies de l'Atlas Tellien sont plus lourds que ceux de l'Atlas Saharien. Ces grandes variabilités géographiques ont amené plusieurs auteurs, notamment GAUSSEN (1964) et DERRIDJ (1990) a émettre l'hypothèse selon laquelle le cèdre de l'Atlas Saharien est plus primitif que celui de l'Atlas Tellien. Les tendances évolutives pour les cônes correspondent à une augmentation leur taille (GAUSSEN (1967). Ainsi, nous sommes tentés par l'hypothèse que le cèdre de l'exposition Sud s'est adapté à une sécheresse plus prononcée et constitue peut être un " écotype ". Le poids des graines chez une espèce donnée est considéré selon HARPER et al (in HENDRIX, 1984) comme un caractère constant ; cependant d'autres auteurs ont signalé qu'il peut varier énormément. (SALISBURY BLACK HARPER et al in HENDRIX, 1984). En outre, LALONDE et al (1989) ont noté que les arbres attribueraient un taux de ressources fixe à chacune des graines, au-delà d'un certain seuil l'arbre a tendance à produire une nouvelle graine ;les graines seront donc, approvisionnées de la même manière, il y aurait par conséquent peu de variation dans leurs poids. Il est considéré comme étant le caractère le plus indiqué pour étudier la variabilité géographique des essences forestières. Il est influencé selon BIROT (in DERRIDJ, 1990) par les effets maternels, mais il est davantage sous dépendance climatique. TOTH (1980) note que parmi les facteurs principaux, la pluviométrie et la durée de sécheresse sont les plus importantes. Il a été proposé par BARITEAU et al. (1990) comme critère de sélection chez les cèdres de l'Atlas, TOTH (1978 a) pour étudier les possibilités de régénération naturelle et DERRIDJ (1990) pour discriminer les populations du cèdre en Algérie, KROUCHI (1995) et HARROUCHE (1991) pour analyser le succès reproductif de Cedrus atlantica M. Comme pour les cônes, ce sont les arbres de l'exposition Nord, qui produisent les graines fertiles les plus lourdes et les plus longues. On peut dire, que le poids des graines fertiles est influence par les caractères biométriques des cônes : plus les cônes sont lourds et plus le poids des graines est important. Par ailleurs l'analyse du poids des graines fertiles et attaquées montre que ces derniers sont deux fois moins lourdes que les graines fertiles. DERRIDJ (1990) et HARROUCHE (1991) ont mis en évidence, la variation du poids des graines fertiles entre les cédraies sèches et humides. Ce caractère est lié fortement à l'altitude pour une même station. Nous savons que le cône de cèdre acquiert sa maturité en 32 mois environ, durant lesquels l'arbre subit l'influence des contraintes du milieu, qui aurait pour conséquence la 139 limitation ou non du taux d'approvisionnement sur la biomasse des cônes et donc la production quantitative et qualitative. Cette production est en relation directe avec la biomasse des cônes et dépend de plusieurs facteurs : - Edaphiques, climatiques et microclimatiques. - Nutritionnel en relation avec la position du cône dans l'arbre et celle de la graine dans le fruit. - Phénologique, à savoir l'initiation de l'inflorescence femelle et la synchronisation entre la disponibilité du pollen, les fleurs femelles et l'ouverture de leurs bractées. - Des conditions ayant prévalu durant le cycle de reproduction, surtout l'année N + 1 elles peuvent constituer une contrainte à l'approvisionnement. - Peuplement : densité, traitements sylvicoles, âge, état de santé et caractéristiques dendrométriques (houppier, diamètre et hauteur). - Année de fructification. - Génétique (fertilité de l'arbre). - Altitude pour une même station. - Concurrence entre le cycle reproducteur et végétatif. 140
Introduction : La régénération du cèdre de l'Atlas demeure à l'heure actuelle un problème important pour la pérennité des cédraies Algériennes en général et celle des Aurès en particulier. Elle dépend en grande partie des conditions du milieu. En effet, les paramètres de station particulièrement les températures et les précipitations influencent la fructification, le pouvoir germinatif des graines, la germination, l'installation et le maintien des semis. Ils agissent en même temps et dépendent en grande partie de l'exposition et de l'altitude. D'autres facteurs considérés généralement comme secondaires peuvent devenir des facteurs limitants : sol (physique, chimique et biologique), couvert et strate herbacée. Selon VALETTE (1983) la régénération artificielle en zones méditerranéennes rencontre de sérieuses difficultés en raison des contradictions du milieu souvent difficiles. Au Belezma même si la régénération naturelle du cèdre de l'Atlas est présente dans certains endroits, la situation actuelle, particulièrement sur les expositions défavorables, ne permet pas au cèdre de se régénérer dans les limites acceptables ; les conditions climatiques défavorables et l'absence des travaux sylvicoles associés aux influences néfastes de l'homme ont déséquilibré l'écosystème. Ainsi, la faible densité des peuplements sur les expositions Sud ne permet pas à cette essence de se renouveler normalement, car la présence de vieux individus dépérissants et dispersés limite les possibilités de productions et de dissémination. Les travaux des différents auteurs, ont permis de subdiviser le problème de la régénération en deux sous - problèmes bien distincts : la germination de la graine et le maintien des semis. Ainsi, CARLIER (1988) souligne que parmis les nombreuses causes possibles de l'insuffisance de la régénération des péssières d'altitude pourraient consister en ce que les conditions nécessaires à la germination ou à la conservation des graines ne soient pas habituellement réalisées. CANTEGREL (1988) rapporte que le renouvellement des fôrets pyrénéennes est fonction de la germination. La germination et le maintien des semis en milieu naturel du cèdre de l'Atlas ont été surtout étudiés par LEPOUTRE (1961,1963), LEPOUTRE et PUJOS (1963), MALKI (1992) leurs travaux de recherches montrent que la régénération naturelle est étroitement liée aux facteurs bioclimatiques, qui conditionnent la date de germination, le développement des semis et leur capacité à développer un système racinaire capable d'assurer l'alimentation hydrique durant la période estivale. TOTH (1971, 1973, 1978 a, 1980,1980 c) a mis en évidence l'étroite relation entre le cycle de production des graines, la régénération et les conditions écologiques. Le même auteur a expérimenté également au laboratoire et sur terrain l'influence de différents facteurs (hydratation, température, éclairement, ...etc.) sur la germination des graines de cèdre. Notons également que la germination des graines de Cédrus atlantica M et leur conservation à fait l'objet de plusieurs publications : ZAKI, 1970 ; ABOUROUH, 1983 ; MULLER et al, 1984; DERRIDJ, 1990 ; BARITEAU, 1993 et BARITEAU et al, 1994. La germination pourrait contribuer aux difficultés et aux entraves de la régénération naturelles et assistées. Ces derniers pourraient provenir de : -l'hydratation insuffisante des graines reposant sur le sol nu - la perte de vitalité des graines insuffisamment hydratées principalement quand l'été est précoce - médiocres taux de germination. 141 CHAPITRE I : La germination des graines de cèdre.
1.1 - Définitions :La germination est le passage de la vie latente de la graine à la vie active, sous l'effet de facteurs favorables. Selon EVENARI (in MAZLIAK 1982), c'est un processus physiologique dont les limites sont le début de l'hydratation de la semence et le tout début de la croissance de la radicule. Une semence a germé, lorsque la radicule a percé les enveloppes ou elle est visiblement allongée (COME, 1970). La germination comprend plusieurs phases physiologiques successives, dont la plus importante est appelée germination sensu stricto, qui s'achève juste avant la croissance de la radicule (EVENARI in MAZLIAK, 1982). Le processus de la germination dépend des facteurs intrinsèques (âge et état de la plante, évolution physiologique et morphologique de la graine) et extrinsèques (humidité, température, oxygène) Il comprend trois principales phases successives : - La première phase : c'est la phase d'imbibition de la graine, qui se traduit par une augmentation régulière et importante de l'activité respiratoire. (COME, 1970 ; MAZLIAK, 1982). - La deuxième phase : c'est la germination sensu stricto elle est marquée par un arrêt de l'absorption de l'eau et une activité respiratoire régulière (MAZLIAK, 1982). - La troisième phase : Elle est caractérisée par une reprise de l'absorption de l'eau et une activité respiratoire de plus en plus importante due au développement de la radicule (MAZLIAK, 1982). Selon EVARI (in ANONYME 1992), cette phase se traduit par une activité enzymatique et une augmentation des taux de respiration et d'assimilation qui sont l'indice d'utilisation des éléments nutritifs mis en réserve, et leur transfert vers les zones de croissance. Durant cette phase 3ème, un changement irréversible se produit dans l'embryon, l'arrêt de la germination provoque la mort de l'embryon (TOTH, 1978 a). Les phases de la germination des graines de cèdre de l'Atlas sont facilement observables à l'oeil nu. Elles se traduisent par : - Imbibition par l'eau. - Ouverture du micropyle. - Apparition de la radicule. - Développement et évolution de l'hypocotyle portant les cotylédons, encore encerclées par les enveloppes des graines. - Déploiement des feuilles cotylédonnaires Toutes ces phases sont schématisées dans la figure n°37. 143 1.2- Paramètres caractérisant la germination :144
... 1 D 9 9 Io 5 ii 3 2 1-Imbibition de graines. 2- Ouverture du micropyle. 3-Apparition de la radicule 4-Allongement de la radicule
feuilles cotylédonaires. 1.2.1- Hétérogénéité de la germination :Le pouvoir germinatif des graines dépend en grande partie des conditions dans lesquelles on les place. Les causes de la variabilité des propriétés germinatives sont multiples et mal connues (COME, 1970).Elles dépendent surtout du patrimoine héréditaire. Mais les facteurs de l'environnement peuvent modifier l'expression de ces propriétés d'origine génétique. Cette hétérogénéité est due selon MAZLIAK (1982) à trois catégories de facteurs : -Conditions de développement des semences sur la plante (température, ensoleillement, pluviosité, nature du sol, etc...).L'origine géographique des semences est donc très importante. -Conditions de conservation (méthodes, température humidité...) -Conditions de germination : plus ces conditions sont défavorables, et plus le lot semble être hétérogène. C'est d'ailleurs une méthode très utilisée pour mettre en évidence l'hétérogénéité d'un lot de semences ou des différences d'états physiologiques entre plusieurs lots. Les résultats des essais de germination varient donc selon l'origine des semences, les traitements qu'elles ont subis et les conditions de germination. 1.2.2- Mode d'expression des résultats : Pour exprimer la germination des graines, nous avons utilisé deux paramètres : - La capacité de germination (C.G%) qui exprime le taux de semences capable de germer dans des conditions bien définies. - La vitesse ou temps de germination : c'est le temps moyen de germination (T.M.G. en jours) qui représente l'inverse x 100 du coefficient de vélocité. Le temps moyen de germination, s'exprime de la façon suivante :
N1 est le nombre de graines germées au temps T1, N2 est le nombre de graines qui ont germées entre le temps T1 et T2,... etc. Très souvent, il n'y a pas de lien selon MAZLIAK (1982), entre ces deux paramètres. Il est donc indispensable d'illustrer les résultats par des courbes de germination, qui décrivent complètement l'évolution de la germination des graines placées dans des conditions déterminées, pendant une période donnée (nombre des graines germées en fonction du temps). Pour caractériser le pouvoir germinatif des semences forestières, divers auteurs notamment ABOUROUH (1983), COME (1970), LEBRU (1970), TOTH (1978 a) MULLER (1986) et MULLER et al (1978, 1980 ,1990a, 1990b) et DERRIDJ (1990) ont utilisé ces deux paramètres (T.M.G et C.G) ainsi que la courbe de germination. 2- Méthodologie générale : 145 2.1- Introduction :Les boites de Pétri sont utilisées quand il s'agit de tester de germination sur de très courtes durées : 12h à 48h. Pour la période de germination qui dure au moins 21 jours, comme c'est le cas du cèdre de l'Atlas, l'utilisation des boites de pétri présente divers inconvénients qui risquent de fausser les résultats. En effet, par leur faible capacité de rétention de l'eau, le dessèchement et/ou l'excès d'eau de la couche porteuse de graines (coton ou papier filtre) nécessitent des manipulations onéreuses. Compte tenu, de ces inconvénients, nous avons été amenés à réaliser nous même des germoirs, qui permettent aux graines d'être alimenté régulièrement en eau, sans aucune intervention de notre part, pendant toute la durée de l'expérience. 2.2- Matériel d'étude: 2.2.1- Le germoir : Le germoir utilisé pour les différents tests de germination est constitué d'un flacon, d'un support, d'un papier filtre et d'un papier buvard (Cf. Fig. n° 38 ). Par son volume (215 ml), ce germoir assure une alimentation en eau pendant 6 semaines sans aucune intervention du manipulateur. Notons que le flacon et le support des graines sont en matière plastique (non biodégradable) permettant de mener des tests de germination sans apports d'hydrosolubles, dus aux différents éléments du germoir. 2.2.2- Les graines : Les graines fertiles de cèdre utilisées pour les différents tests de germination sont issues des cônes mûrs de deux années et triées manuellement. Elles sont triées visuellement dans un premier temps, puis par densimétrie (éther de pétrole) ou les graines lourdes (saines) se déposent au fond du conteneur. Un second test visuel est indispensable pour éliminer les graines détériorées (éclatement des poches de résine, blessures...). Selon BARITEAU (1993) le tri à la main est une méthode non traumatisante pour les graines de cèdre ; elle a été préconisée lors du Séminaire International pour le genre Cedrus. Du fait de la dormance primaire, les graines de Cedrus atlantica M subissent un prétraitement par le froid humide pour lever la dormance, après séchage à l'air libre pendant 48h. Les récoltes et les traitements sont utilisés en fonction des buts recherchés. 146
8,60cm8 cm
Fente Boite transparente en plastique
Support en plastique 6cm Papier buvard long. GRAINES GERMOIR Fig. 38:Germoir à alimentation continue en eau employé dans les différents tests de germination. 147 2.3- Mise en germination : 147 Pour les graines de cèdre de l'Atlas, la germination est effectuée selon l'association Internationale d'Essais de Semences (I.S.T.A) à 20 °C ou 20°C - 30°C pendant 21 jours, après au moins 3 semaines de prétraitement sur milieu humide de 3 à 5° C (MULLER et al, 1984 a). Le dispositif expérimental comporte pour chaque traitement 10 germoirs contenant chacun 10 graines fertiles disposées sur du papier filtre, ce dernier est posé directement sur un support dur et transparent. Il présente en son milieu une fente permettant un contact entre le papier filtre, le papier buvard et l'eau. Ainsi l'alimentation en eau des graines est assurée par capillarité. Les tests de germination s'effectuent au laboratoire à température ambiante ; le comptage des graines germées se fait tous les trois jours. Notons que pour ces tests, nous adoptons la définition physiologique de la germination. Cette dernière est contrôlée par divers facteurs, qui agissent tous en même temps. L'analyse d'un facteur, ne doit en aucun cas faire abstraction aux autres. 3- Etude expérimentale de la germination au laboratoire :3.1- Hydratation des graines :3.1.1- But : L'hydratation des graines de cèdre de l'Atlas constitue la première phase de la germination, qui se traduit par une absorption de l'eau sous forme liquide. C'est un phénomène physiologique, qui se termine par le début de la croissance (allongement de la radicule). Cet essai a pour objet de déterminer la quantité d'eau absorbée par les graines fertiles avant la phase de croissance de la radicule : c'est la première phase de la germination. 3.1.2- Matériel et méthode : Trois lots de graines fertiles non dormantes sont séchés à l'air libre jusqu'à stabilisation de leur poids respectif. Chaque lot, est composé de 100 graines ayant pour poids : - l' lot: 7,7321 g.
Ces lots trempés dans de l'eau distillée. Ils seront par la suite pesé après lh, 6h, 12h, 24h, 48h, 72h, 96h et 120h d'immersion ; avant chaque nouvelle pesée les graines sont d'abord séchées avec du papier buvard. 3.1.3- Résultats -- discussion : 148 Les données brutes sont portées en annexe X Les résultats obtenus sont donnés par le tableau n° 81 et représentés graphiquement par la figure n° 39. Tableau N°81 : Evolution du poids en pourcentage des graines fertiles du cèdre immergées dans l'eau distillée pendant 120 heures (5 jours).
Ils nous indiquent que le poids, se stabilise à partir de la 96èmeheure ; les graines n'absorbent pratiquement plus d'eau au-delà de cette période.
Poids de 100 graines 12,9328g 12,3752g 8,0916g Heures 7 9 8 14 - 13 - 12 - 10 - 0 1 6 12 24 48 72 96 123 Fig. 39 : Evolution du poids des graines de cèdre de l'Atlas en fonction du temps de l'hydratation. L aum.nputni uc I cau pai ici gic1,111Gb 1W11,11Gb, pabbt., pai 1Gb vamcb . 149 - Première phase: l'absorption de l'eau est extrêmement rapide au départ. En effet en une heure, les graines enregistrent une augmentation de l'ordre de 20 % en moyenne. Les graines les moins pesantes, n'augmentent que de 14 % environ. - Deuxième phase: elle se traduit par un gain de poids de l'ordre de 5 %. Cette phase d'absorption lente est située entre la 6ème heure et la 12ème heure. - Troisième phase: c'est la deuxième phase d'absorption rapide, elle est comprise entre la 12ème heure et la 24ème heure et se traduit par une augmentation moyenne de l'ordre de 15 %. - Quatrième phase : elle se caractérise par une absorption lente et continue de l'eau, jusqu'à la 96ème heure. Au-delà de cette période, le poids a tendance à se stabiliser. Ces résultats, montrent que l'hydratation des graines, prélude de la germination, passe par 4 phases d'absorption : rapide- lente- rapide- lente. Celle-ci se traduit par une augmentation du poids des graines de l'ordre de 55 %. CARLIER (1988) rapporte qu'aucune germination n'est observée tant que la teneur en eau des graines d'épicéa (Picea abies L) n'atteint pas 51% par rapport au poids des graines anhydres Les graines les plus lourdes, ont tendance à absorber beaucoup plus d'eau que les graines légères et, germent donc rapidement. CHAUSSAT et al (1975) signalent que la courbe d'imbibition est pratiquement la même pour l'ensemble des graines ; elle est rapide pendant les premières heures puis devient lente; quand elle atteint son maximum, la germination survient plus ou moins longtemps (à partir de 72ème heure selon nos essais) Par ses téguments membraneux et perméables, la graine de Cedrus atlantica M, présentent selon TOTH (1978 a) et ABOUROUH (1983) une grande facilité d'hydratation et d'oxygénation. Ceci se traduit par une germination relativement facile. 3.2- Essai de germination : 3.2.1- But : Cet essai a pour but de vérifier si le germoir que nous avons réalisé, permettra aux graines de germer normalement. 3.2.2- Méthodologie : Pour cela, nous avons effectué deux tests de germination, l'un sur le germoir que nous avons réalisé et l'autre sur le germoir de marque BENDER et HOBEIN (ex R.F.A). Ce dernier contrôle automatiquement l'eau et la température. Afin d'éliminer au maximum, l'hétérogénéité de la germination des lots de graines, nous avons récolté 10 cônes (N+2) situés sur la même branche. Après désarticulation et triage les graines fertiles obtenues sont séchées à l'air libre pendant 48 h, puis traitées par le froid humide durant 30 jours. Les essais de germination sont effectués en même temps : le premier sur germoir BENDER et HOBEIN à 20 °C, le second sur boite germoir à température ambiante. Chaque essai a porté sur un lot de 100 graines, à raison de 10 par répétition. 3.2.3- Résultats -- discussion : 150
Jours 3 6 9 12 15 18 21 100 Capacité de germination (%) 90 80 Germoir Bender et Hobein Boite - germoir 70 60 50 40 30 20 10 Troisième partie Chapitre I : La germination de la graine de cèdre de l'Atlas. Les données brutes sont portées en annexe XI Les résultats sont consignés sur le tableau n°82, qui indique la capacité de germination (CG en %) et le temps moyen de germination (T.M.G. en jours). Ils montrent que les deux germoirs utilisés ont donné pratiquement les mêmes résultats. Cependant le germoir BENDER et HOBEIN enregistre un gain de 4 % pour la capacité de germination et 0,4 jours pour le temps moyen de germination. Tableau N°82 : Influence du germoir sur la germination des graines du cèdre de l'Atlas.
La figure n° 40 exprime l'évolution de la germination en fonction du temps.
Elle montre que l'allure des deux courbes est pratiquement la même mais un léger avantage est à signaler pour le germoir BENDER et HOBEÏN. Ceci est dû probablement au facteur température. En effet si l'eau ne pose pas pour les deux types de germoir, le facteur température des boites de germoir dépend des conditions externes. Cet essai, montre que les « boites » germoir, permettent aux graines de germer normalement et peuvent donc être utilisé pour les différents tests de germination. 3.3- Influence de l'exposition, de la position des cônes dans l'arbre et du poids des graines sur la germination : 151 3.3.1- Introduction : L'étude des graines fertiles (Cf. Chapitre IV : Etude des cônes et des graines) a montré que leurs caractères biometriques varient en fonction de l'exposition et de la position des cônes dans l'arbre. Les graines de l'exposition Nord et des branches hautes présentent respectivement des poids plus importants que celles de l'exposition Sud et des branches basses. Il est surtout lié aux réserves accumulées durant le processus de formation particulièrement pendant la phase de la maturité morphologique : il y a élaboration des constituants de la semence, celle-ci s'achève quand l'embryon aura atteint son développement normal. Le poids des graines est surtout influencé par les conditions climatiques. Elles jouent selon TOTH (1978 a) un rôle important sur les aptitudes germinatives des graines. En outre DERRIDJ (1990) a mis en évidence l'influence de l'origine des graines : celles de l'Atlas Tellien (plus dimensionnées) présentent les meilleures capacités de germination par rapport aux graines de l'Atlas Saharien (moins dimensionnées). Compte tenu de la variabilité du poids des graines, nous avons effectué des tests de germination en appliquant les mêmes traitements (durée de stratification) et les mêmes conditions de germination. (Boite - germoir à température ambiante) 3.3.2- Matériel et méthodes : Des graines fertiles provenant de Belezma ont été reparties en fonction de l'exposition, de la branche et de leur poids unitaire. Pour l'exposition, l'échantillonnage a consisté en deux lots de 100 graines, appartenant chacun à une exposition. Quant à la position des cônes dans l'arbre les deux lots de graines proviennent d'un seul pied de cèdre, afin d'éliminer l'hétérogénéité des semences. Le premier lot appartient à la branche la plus haute de l'arbre et le second à celle du plus bas (hauteur d'homme) Pour le poids des graines, ces dernières qui proviennent uniquement de l'exposition Nord sont pesées une par une et classées en lots de 100 graines chacun en fonction de trois catégories (lots) de poids. l' lot : composé de graines fertiles dont le poids unitaire est inférieur ou égal 0,0699g.
Ces graines sont obtenues après désarticulation manuelle, un premier tri visuel, ensuite un tri densimétrique (Ether de pétrole) et enfin un second tri visuel. Elles sont séchées à l'air 152 libre pendant 48 h, ensuite elles subissent un prétraitement par le froid humide pendant 30 jours. La germination est faite dans des boites germoir comme précédemment, la durée est fixée à 21 jours. Le comptage se fait tous les trois jours. 3.3.3- Résultats -- discussion :Les résultats bruts sont données en annexes XII, XIII et XIV Ils sont donnés par les tableaux n°83, n°84 et n°85 et représentés graphiquement dans les figures n°41, n°42 et n°43 - Station : Pour l'ensemble de la station, la capacité et la vitesse de germination obtenues sont de 83,5 % pour 9 jours. Ces deux paramètres montrent que les graines fertiles présentent des aptitudes germinatives satisfaisantes. D'après le tableau n°83 et la figure n°41, nous constatons que : l'exposition Nord a donné les meilleurs résultats (87 % pour 8,52 jours) que l'exposition Sud (80 % pour 9,49 jours) ; la différence des aptitudes germinatives entre les deux expositions est due à la qualité de station. Tableau N°83 : Influence de la station sur la germination.
- Branche: Par ailleurs, pour le paramètre branche, les graines issues des branches hautes présentent des capacités de germination meilleures que celles des branches basses. Par contre les temps moyens de germination sont pratiquement les mêmes (Cf. tableau n°84 et fig. n°42) Tableau N°84 : Influence de la position des cônes dans les branches sur la germination.
- Poids Quant au caractère poids, les résultats (Cf. tableau n° 85 et fig. n°43) montrent que les graines les plus légères donnent les plus faibles résultats (63 % pour 12,28 jours).Les graines les plus lourdes enregistrent les meilleures aptitudes germinatives : C.G= 96 % et T.M.G = 9,44 jours. 153 154 Tableau N°85 : Influence du poids des graines sur la germination.
L'étude de la biométrie des cônes et des graines (Cf. 3ème partie Chapitre IV ) nous a montré que les graines de dimension réduites sont plus fréquentes dans l'exposition Sud et les branches basses que dans les expositions Nord et les branches hautes. Par ailleurs, nous savons également que les graines lourdes germent mieux que les graines légères. En conséquence, nous pensons que le poids des graines à une influence non négligeable sur la régénération naturelle. Il dépend en partie des conditions dans les quelles la graine a évoluée (état sanitaire de l'arbre, son âge, les facteurs climatiques...). 3.4- Influence des différentes parties de la graine sur la germination :3.4.1- Introduction :COME (1970), CHAUSSAT et al (1975), TOTH (1978 a) ainsi que MAZLIAK (1982) signalent que certaines parties de la graine comme le tissu nutritif, les téguments ne sont pas indispensables à la germination, ils peuvent même l'inhiber. 3.4.2- Matériel et méthode : Afin de tester l'influence des différentes parties de la graine, à savoir les téguments et l'aile trois lots de 100 graines sont repartis de la façon suivante : - Un lot représentant le témoin, dont les graines sont sans ailes, mais avec téguments. - Un lot dont les graines sont dépourvues de téguments et d'ailes (graines nues). - Un lot de graines pourvues d'ailes et de téguments. Ces graines ont subi au préalable, un prétraitement par le froid humide et misent à germer dans les conditions précitées. Le comptage se fait tous les 3 jours. Par ailleurs, afin d'éviter d'éventuelles contaminations des graines nues par des champignons ces dernières sont immergées dans de l'eau oxygénée (10 %), avant leurs mises en germination. 3.4.3- Résultats : Les résultats bruts sont données en annexe VX Les résultats de ces tests sont indiqués dans le tableau n° 86, qui donne pour chaque lot la capacité de germination et le temps moyen de germination. 155 Tableau N°86 : Influence des différentes parties de la graine sur ses aptitudes germinatives.
La figure n°44 qui décrit la cinétique de la germination montre qu'il n'y a pas de différences nettes entre les trois lots de semences.
0 0 3 6 9 12 15 18 21
100 - 90 - 80 - 70 - 60 - 50 - 40 - 30 - 20 - 10 - Capacité germination.( %) Jours Fig.44: Influence des différentes parties de la graine de cèdre de l'Atlas sur son pouvoir 3.4.4- Discussion -- conclusion : Les différents tests de germination, montrent que les graines arrivent à germer en l'absence de téguments et en présence d'ailes. Ceci montre que la dormance embryonnaire n'est pas due aux différentes parties de la graine. En effet, le tableau n° 86 montre que le premier lot qui constitue le témoin (graine sans ailes, avec téguments) et le second lot, composé de graines nues (sans ailes et sans téguments) donnent les meilleurs taux de germination (97 %).Les graines nues donnent le meilleur temps moyen de germination avec 7,11 jours. Les téguments semblent agir donc sur la vitesse de germination : les graines nues s'hydratent beaucoup plus vite. Quant aux lots des graines ailées, elles enregistrent les plus mauvais résultats. Notons que les poches résinifères, semblent être indispensable à la germination (MULLER et al, 1984a ; DERRIDJ, 1990) ; d'autres auteurs pensent qu'elles diminuent le pouvoir germinatif des graines (F.A.O, 1992). TOTH (1978 a) et DERRIDJ (1990) ont montré que l'endosperme n'inhibe pas la germination. La dormance primaire est d'origine embryonnaire. 156 D'autre part, selon nos propres expériences, les hydrosolubles des aiguilles vertes des arbres adultes de cèdre, de litière fraîche et ancienne, ainsi que l'humus n'ont aucun effet dépressif sur la germination même à de fortes concentrations (20 %). 3.5- Influence des prétraitements sur la germination : 3.5.1- Introduction : Les conditions nécessaires à la germination sont : l'oxygène, la température, l'humidité (mais sans excès), la lumière (graines de cèdre sont sans photosensibilité) et la maturité physiologique. C'est ce dernier point qui sera traité. Comme pour les diverses graines forestières, celles du cèdre de l'Atlas qui proviennent de cônes murs, et placées dans des conditions favorables, s'imbibent normalement, les micropyles s'ouvrent, mais les radicules n'apparaissent pas. On dit que les graines sont dormantes. Selon MULLER (1986), le phénomène de dormance concerne 60 % des graines forestières. Cette dormance propre à l'embryon (dormance primaire) s'installe selon COME (1970) au cours du développement de la semence sur la plante, avant ou pendant la maturation morphologique. Elle est peut être considérée comme un système régulateur de la germination. Elle est due, selon les différentes théories, à des hormones végétales, à des modifications métaboliques, aux phénomènes de perméabilité à l'eau, à l'oxygène et à la présence de certains composés dans les téguments (COME, 1970 ; MULLER et al 1990). Le degré de maturité des graines de Cedrus atlantica M est lié à l'ouverture des cônes, et à leur aptitude à se désarticuler. TOTH (1978 a) et MULLER (1984a) signalent que cette maturité est caractérisée par la couleur des embryons (couleur verte).Les graines de cèdre à maturité incomplète, donnent des C.G. (%) faibles et de mauvaises aptitudes à la conservation. Les graines du cèdre de l'Atlas, présentent un état de dormance plus ou moins profond. Selon MULLER (1986), elle est hétérogène d'un lot à un autre et même à l'intérieur d'un même lot. Pour germer, la graine de cèdre a besoin de subir pendant quelque temps l'action du froid Ce dernier a deux actions différentes sur les graines : la levée de la dormance primaire et la conservation. Les températures basses agissent sur les graines en provoquant progressivement leur maturation physiologique pendant une période plus ou moins longues (30 à 60 jours selon les auteurs). La levée de dormance réside à faire subir aux graines un froid humide pendant au moins 30 jours. Cette technique appelée stratification consiste à déposer les graines par couches successives dans du sable humide, puis les laisser séjourner dans un réfrigérateur à une température de 4°C durant 30 jours au moins. 157 La combinaison du froid sec puis humide prépare mieux la graine à germer (ZAKI, 1969). La stratification est le prétraitement le plus utilisé pour lever la dormance des graines du cèdre de l'Atlas. (ZAKI, 1969; TOTH, 1978 a, 1978 b; ABOUROUH, 1983; MULLER et al, 1978, 1983; DERRIDJ, 1990). Néanmoins, ce prétraitement est une opération longue et il n'est pas évident d'avoir des chambres froides, pour stoker ce mélange. Pour cela, il existe d'autres méthodes, moins longues et moins coûteuses permettant de lever la dormance embryonnaire par l'immersion des graines dans des solutions aqueuses. En revanche l'utilisation de ces solutions est très délicate, car il faut prendre en considération : La concentration. La durée de trempage. Les graines de Douglas (Pseudostuga menziesii) traitées par l'eau oxygénée est une méthode de lever la dormance plus efficace que la méthode classique par le froid, moins pratique et plus longue (LEBRU, 1970). En outre pour les faînes de Fagus silvatica une pré-réfrigération sans milieu semble meilleure que la stratification classique (tourbe humide), mais elle nécessite le maintien tout au long de ce traitement, d'une teneur en eau des faînes voisine de 30 %. L'installation des faînes dans une solution contenant 150 à 200 g de polyéthylène glycol (P.E.G) par kilogramme, assure ce maintien et évite toute sortie de radicule au cours du passage au froid (MULLER et al, 1983). La levée de dormance par l'eau oxygénée a été utilisée notamment par ABOUROUH (1983) et DERRIDJ (1990). Il s'agit de tremper des graines de cèdre de l'Atlas dans l'eau oxygénée à 35 % pendant une heure; les résultats obtenus sont pratiquement les mêmes que la stratification classique. 3.5.2- But : Il s'agit de comparer l'influence de la durée du froid humide et de la concentration l'eau oxygénée sur les aptitudes germinatives des graines de cèdre. 3.5.3- Matériel et méthodes : Des graines fertiles sont reparties en 6 lots de 100 individus. Trois lots vont subir un prétraitement par le froid humide pendant différentes périodes : les graines sont mélangées avec du sable humide (lavé), où ils subiront une stratification dans un réfrigérateur pendant une période de 15, 30 et 45 jours. Après chaque période, un lot de 100 graines est mis à germer. 158 Deux lots sont traités par l'eau oxygénée. Le premier lot est trempé dans de l'eau oxygénée à 30 %, pendant une durée de 45 mn. Quant au second, son immersion est de 60 mn, à une concentration de 15 %. Le dernier lot qui compose le témoin est mis à germer sans aucun prétraitement En outre l'eau oxygénée (11202) étant peu stable à l'air, les solutions préparées sont utilisées immédiatement. 3.5.4 - Résultats : Les données brutes sont portées en annexe VIX Les résultats de ces tests sont représentés dans le tableau ci - dessus qui indique pour chaque traitement la capacité et le temps moyen de germination. Tableau N°87 : Influence de deux prétraitements sur la capacité et le temps moyen de germination.
La figure n°45 illustre l'évolution de la germination pendant la période de l'essai. 100
o 3 6 9 12 15 18 21
159 3.5.5-Discussion - conclusionLes résultats montrent que le prétraitement des graines améliore leurs aptitudes germinatives Le meilleur résultat est obtenu avec la durée de 45 jours : C.G. est de 96 % pour 6,32 jours. Le froid humide (stratification), méthode très utilisée pour la levée de dormance indique que plus la durée est longue plus les résultats sont meilleurs. La durée de stratification est un paramètre très important, mais il ne doit pas dépasser une certaine limite En effet selon ABOUROUH (1983) et MULLER et al (1984), ce paramètre est de 5 à 6 semaines, car au- delà de cette durée, il n'y a pas d'amélioration du pouvoir germinatif. L'effet de la stratification dépend de la date de récolte (ZAKI, 1970), donc de la maturité des graines. Cette dernière est liée selon MULLER et al (1983) au degré d'ouverture des cônes et à leur aptitude à se désarticuler. Le traitement par l'eau oxygénée a amélioré la germination. Si pour le témoin la C.G. est de 37 % pour un T.M.G de 11,72 jours, les lots traités pendant une heure seulement ont donné de bons résultats. En effet une concentration de 30 % pour 45 mn a donné une C.G. de 70 % et un T.M.G de 9 jours alors que 15 % pour une durée de 60 mn n'a permis d'avoir que 55 % avec T.M.G. de 9,38 jours. Ces résultats montrent que l'utilisation de l'eau oxygénée à 30 % pendant 45 mn, peut remplacer une stratification classique de 15 jours. L'augmentation du temps d'immersion peut accroître le pouvoir germinatif des graines et donc se substitue à une stratification de 45 jours. Une concentration de 30 % a un effet meilleur sur le pouvoir germinatif qu'une concentration de 15%. Cependant la concentration de l'eau oxygénée ne doit pas dépasser 35%. (TRAPPE, in ABOUROUH, 1983). Le traitement des graines par l'eau oxygénée (H202) permet à l'embryon de disposé d'une plus grande quantité d'oxygène (COME, 1970). De plus, c'est un désinfectant, elle nettoie les téguments les rendant ainsi plus perméable à l'eau et à l'oxygène. Signalons, que le séjour des graines de cèdre dans des solutions d'A.I.A (Acide Indole 3 Acide acétique) et l'acide giberellique (G.A.3) pendant 24 h, n'ont pas eu d'effets spectaculaires. Selon DERRIDJ (1990), les concentrations, qui semblent améliorer peu la germination sont faibles (le mg/1), à forte dose (100 mg/1), ces auxines inhibent la germination. A la lumière des recherches actuelles, les prétraitements des graines de cèdre par le froid humide ou l'eau oxygénée sont un préalable à la germination : - Accélération de la germination. - Augmentation du nombre de graines germées. - Diminution de la mortalité. 160 3.6- Influence de la date de récolte et du mode de prétraitement :3.6.1- Introduction :Les pépinières productives de plants sont dépourvues de moyens leur permettant de traiter les graines de Cedrus atlantica M. La méconnaissance de la dormance des graines et donc la non-application des traitements par le froid humide ou sec, se traduit par des rendements médiocres des pépinières et un gaspillage de graines et de plants. En effet, nous avons montré que les graines non traitées ont des capacités de germination les plus faibles et le temps moyen de germination le plus long (3.5 - Influence des prétraitements sur la germination). Par ailleurs, les récoltes des graines au niveau de nos pépinières sont effectuées durant les mois d'Octobre et Novembre. Une fois désarticulées, les graines sont séchées à l'air libre pendant 4 à 5 jours, puis triées grossièrement. Les graines ainsi obtenues, sont utilisées soit immédiatement, soit conservées (le plus souvent) pendant plusieurs mois, voire des années, dans de mauvaises conditions. La conservation des graines par stratification est un préalable à la germination. La période de récolte est un paramètre très important permettant de diminuer au maximum les pertes de graines. TOTH (1978) MULLER et al (1984) notent qu'en France il est préférable d'effectuer la récolte en Octobre - Novembre.En réalité, il faut se rapprocher de la date de désarticulation naturelle. Plus la température est basse avant la récolte, plus la faculté germinative est élevée. Par ailleurs, nous avons trouvé qu'il existe une étroite relation entre le pouvoir germinatif des graines et l'année de leur production : quand l'année de production est mauvaise, la germination est médiocre; année de forte production, la germination est bonne. Ceci est dû à la qualité des graines. 3.6.2- Matériel et méthodes : Le pouvoir germinatif des graines dépend de la date de récolte. Il s'agit donc, de déterminer la meilleure date de récolte, qui permet d'avoir la meilleure capacité de germination, avec des temps moyens de germination les plus bas possible. Pour tester l'influence de ce facteur nous avons travaillé sur une même population : (exposition Nord) à des périodes différentes. Les graines utilisées sont issues de 5 récoltes effectuées le : 2 Septembre - 3 Octobre - 2 Novembre - 5 Décembre - 1 Janvier Durant la première récolte, tous les cônes sont fermés, pour ceux des deux récoltes suivantes, l'ouverture des écailles est entamée. Quant aux cônes du mois de Décembre et Janvier, la désarticulation est entamée, elle est précédée de l'ouverture des écailles qui est le résultat de l'influence de la chaleur et de l'humidité. Après décortication manuelle des cônes, les graines sont traitées, puis séchées à l'air libre. Un lot de 100 graines, est mis à germer immédiatement après le séchage, sans aucun prétraitement. Le second subira une stratification pendant 30 jours. Les observations sont faites tous les 3 jours, pendant 21 jours. 161 3.6.3- Résultats : Les données brutes sont portées en annexe XVII et XVIII. Le tableau n° 88 donne les résultats obtenus pour ces tests. Tableau N°88 : Influence de la date de récolte et du prétraitement par le froid humide sur les capacités et les vitesses de germination.
Les figures n°46 et n°47 représentent l'influence des traitements et des dates de récoltes sur l'évolution du taux de germination durant 21 jours. 3.6.4- Discussion -- conclusion : Les résultats obtenus, montrent que la capacité de germination et la vitesse de germination des graines non traitées, récoltées en Décembre et Janvier (tardivement) sont meilleures que celles récoltées au mois de Septembre, Octobre et Novembre. En effet, le maximum de graines germées a été obtenu pour le mois de Janvier et Décembre avec respectivement 91 % pour 09,46 jours et 70 % pour 09,68 jours. Le minimum est obtenu pour le lot du 2 Septembre (10 % pour 12 jours) et du 3 Octobre (13 % pour 12 jours). Le mois de Novembre a donné des résultats intermédiaires (50 % pour 13.62 jours). Pour les graines traitées par le froid humide hors cônes, les taux de germination obtenus sont comprises entre 74 % et 94 %, avec des temps moyens les plus bas. Néanmoins, ces derniers ne suivent pas les capacités de germination (Décembre et Janvier). Ces tests montrent que les graines récoltées en Septembre et Octobre se caractérisent par un état d'immaturité physiologique prononcée. Cet état de dormance primaire, disparaît après un traitement artificiel par le froid pendant 30 jours, puisqu'on passe d'une capacité de germination de l'ordre de 10 % à plus de 70 %. En outre, en l'absence de prétraitement, nous remarquons que plus la récole est tardive, moins les graines sont dormantes. Pour la récolte de Janvier, il n'y a pratiquement plus d'effet de stratification. La maturation a réduit l'état de dormance. Les graines dormantes enregistrent un taux de germination de 5 % selon TOTH (1978a) ; MULLER et al (1984) obtiennent 12 % pour le mois d'Août, et 19 % pour celui de Septembre. Nous notons également, que plus les écailles sont ouvertes et plus la maturité des graines est meilleure. A partir du mois de Novembre jusqu'à la période de désarticulation totale, les conditions écologiques (températures, humidité) activent la maturation et lèvent la dormance Cette levée semble se faire progressivement au cours du temps (Cf. Figure n°48) 162 163 La durée de stratification dépend de la date de récolte. Une stratification de 30 jours semble insuffisante quand les récoltes sont précoces (Septembre - Octobre) et qu'elle n'est 164 pas indispensable, lorsqu'elle est tardive (Décembre et Janvier).Elle dépend des conditions écologiques précédent la récolte et donc du degré d'ouverture des cônes. Actuellement, la récolte des cônes pour les pépinières productrices de plants de cèdre se fait au mois d'Octobre et Novembre, ce qui se traduit par d'énormes pertes qui peuvent atteindre 90 % selon les résultats que nous avons obtenu : les pépinières ne possèdent pas de chambres froides leur permettant de traiter les graines. Il serait donc plus judicieux de procéder à des récoltes tardives. La période de récolte, serait donc de l'ordre de 5 à 6 mois avant la date de désarticulation naturelle des cônes. Les graines ne présentent pas de trace d'immaturité, à condition de leur faire subir des prétraitements avant leur mise en germination. 3.7- Influence de la conservation des graines sur la germination :3.7.1- Introduction :Le principal objectif de la conservation des graines forestières est de mettre en réserve le plus grand nombre de graines lors des années de forte production en prévision des années déficientes. Nous avons montré que le cycle de reproduction qui dure 29 mois, est caractérisé par une irrégularité de la production quantitative et qualitative des graines. Par ailleurs, l'augmentation des coûts des semences (récoltes, transport, désarticulation...) les mauvaises années de fructification (incendies, sécheresse, insectes...) font de la conservation une opération indispensable pour les pépinières; elle leur permet d'avoir à tout moment des graines saines et fertiles. La conservation des graines de cèdre de l'Atlas est une opération délicate, qui fait appel à divers facteurs: date de récolte - teneur en eau des graines - séchage - température et mode de conservation. Conserver les graines fertiles du cèdre de l'Atlas, consiste à les maintenir en vie de ralentie grâce à une déshydratation (séchage). Les différents travaux sur la conservation montrent que plus la température est basse et plus la teneur en eau des graines doit être faible. Il existe deux méthodes de conservation : - La Conservation traditionnelle : les graines sont conservées dans les cônes. Ces derniers sont étalés dans des endroits bien aérés. Après désarticulation, les graines sont utilisées immédiatement ou bien stockées dans des endroits frais. - La Conservation actuelle : les graines sont conservées hors cônes, dans des récipients hermétiquement clos, leur humidité doit être inférieure à 10 % (ZAKI, 1968; TOTH, 1978 a). MULLER (1986) rapporte que pour les graines dites orthodoxes (Pinea - Abies - Cedrus), la teneur en eau la plus indiqué doit être de l'ordre de 5 à 10% et les températures très basses comprises entre -5 ° et -15 °C. 165 MULLER et al (1984) ont obtenu après 6 années de recherches sur l'amélioration du traitement des graines de Cedrus atlantica M, les résultats suivants : - Le stockage des graines à l'intérieur des cônes à 4 °C, n'est retenu que si la durée de conservation ne dépasse pas 8 mois. Hors cône, l'aptitude germinative est satisfaisante, après 6 années et demi de conservation à condition que l'humidité des graines soit inférieure à 11 %. - La conservation à - 15 °C, s'effectue très mal quand la teneur est supérieur à 11 % ; si les teneurs sont comprises entre 5,5 et 7 % les résultats sont très bons pendant 42 mois Mais à partir de 8 %, la faculté germinative diminue graduellement pour atteindre 30 % en 42 mois de conservation. Les recherches menées par ZAKI (1968), TOTH (1978 a, 1980c) et surtout MULLER et al (1984, 1990) sur la conservation des graines, permettent de déduire qu'on peut opter entre plusieurs méthodes, selon le terme : - Très court terme : 2 à 5 mois; La conservation peut se faire en cônes à une température comprise entre + 4 et + 12 °C. A l'état de graines à + 4°C après séchage (pas de prégermination) ou sans séchage avec risque de prégermination. - Moyen terme : 15 à 18 mois; Les graines doivent être conservés de préférence hors cônes, en adoptant la conservation à long terme. - Long terme : 7 à 5 ans; Après extraction et séchage jusqu'à une teneur en eau comprise entre 5,5 et 7,5%. La conservation est effectuée en récipients hermétiques à une température comprise entre - 5 et -5°C. 3.7.2- Méthodologie d'étude : Afin d'estimer l'importance des pertes, qui peuvent être engendrer par les pépinières productrices de plants de cèdre, nous avons utilisé les mêmes méthodes : dates de récole - décortication des cônes - tri des graines - conservation des graines hors cônes et dans les cônes. Les récoltes ont été effectuées au mois de Novembre. Une partie des graines a été extraite, après immersion des cônes dans de l'eau. Les graines ainsi récupérées, sont conservées dans un sac en toile perforée, qui sera déposé dans un endroit frais et bien aéré. Les cônes restants ont été conservés de la même manière que les graines. Les tests de germination ont été menés comme précédemment à raison de 100 graines par essai. Pour le témoin, les graines sont mises à germer immédiatement après tri et séchage. Quant aux autres essais, ils sont effectués après les différents temps de conservation choisis : 6 - 12 et 24 mois, hors cônes et dans les cônes. 3.7.3- Résultats : Les données brutes sont portées en annexe IXX 166 Les résultats obtenus sont donnés dans le tableau n° 89, montre que les méthodes de stockage utilisées au niveau de nos pépinières causent énormément de pertes. Tableau N°89 : Etude du mode et du temps de conservation sur la capacité de germination et le temps moyen de germination.
La figure n°49 montre que le meilleur mode de conservation est le hors cône.
0 3 6 9 12 15 18 21 Jours --A-- Témoin -- H.C.6mois --)K-- H.C.12mois --.-- H.C.24mois --1-- D.C.6mois -- D.C.12mois --*-- D.C.24mois Fig.49 : Effet du mode de conservation sur la germination des graines de cèdre de l'Atlas. 3.7.4- Discussion -- conclusion :Les graines conservées hors cônes ou dans les cônes pendant 6 mois ont des capacités de germination pratiquement identique : 56 % pour 54 %. La vitesse de germination est en faveur des graines conservées hors cônes. La conservation traditionnelle à court terme (6 mois) a peu d'effets sur les aptitudes germinatives des graines peu dormantes hors cônes ou dans les cônes. Les graines conservées durant une année et plus, nous permettent d'enregistrer des chutes très importantes de leur pouvoir germinatif particulièrement quand elles sont à l'intérieur des cônes. Nous avons également noté, qu'un nombre important de graines a pré-germé au cours de cette conservation, et que les champignons ont causé beaucoup de dégâts. Donc, avant de conserver les graines de cèdre, il faut d'abord les faire sécher. 167 En effet, la vie latente des graines est liée directement à sa teneur en eau (TOTH, 1978a) et que cette dernière, doit être comprise entre 5,5 et 7 % selon MULLER et al (1984). La déshydratation des graines doit se faire progressivement et qu'il ne faut jamais aller au - delà de 5 %, car les macromolécules se dégradent (TOTH, 1980a). Les graines conservées dans les cônes donnent des résultats médiocres, au -delà de 4 à 5 mois (ZAKI, 1968; DEMOVEN, CHAUDEY, MARTIN, in : TOTH, 1978 a ; MULLER et al (1984), MULLER (1986). 4 - Conclusion Ces essais montrent que les graines fertiles de cèdre de l'Atlas, triées par l'éther de pétrole (1 1 = 0,65 kg) germent normalement après prétraitement au froid humide ou à l'eau oxygénée. Par contre le tri densimétrique à l'alcool induit selon BARITEAU (1993) une mortalité non négligeable des graines viables : téguments minces et perméables. Il ressort de cette étude un certain nombre d'information, qu'il faut prendre en considération, afin d'éviter le gaspillage des graines fertiles d'une part et d'augmenter leur faculté germinative de l'autre part. Avant la germination sensu stricto, les graines absorbent au moins 50 % d'eau de leur poids, apres avoir passé par 04 phases d'absorption : rapide -- lente -- rapide - lente. Les graines récoltées sur exposition Nord et sur les branches hautes présentent les meilleures capacités de germination que celles de l'exposition Sud et des branches basses. Nous avons montré dans la 2ème partie, Chapitre W que non seulement l'exposition Nord et les branches hautes sont les plus productives, mais que leur poids est plus important: c'est ce caractère qui explique la différence de la germination entre les stations et la position des cônes dans l'arbre. Les différentes parties de la graine, semblent n'avoir aucun effet sur le taux de germination. En outre le traitement de graines dormantes à l'eau oxygénée est efficace à forte concentration (30 %); cependant la stratification pendant 45 jours (2 à 4° C) constitue le meilleur traitement. La période de récolte serait beaucoup plus étendue, elle est de l'ordre de 5 mois avant la désarticulation totale des cônes, mais avec des récoltes trop précoces, les cônes sont difficiles à décortiquer et les graines plus dormantes. La durée de la germination (temps de latence) est liée à la maturité et à l'état de dormance des graines ; il est de l'ordre de 3 jours. Le temps de trempage nécessaire à la décortication des cônes diminue au fur et à mesure que l'on se rapproche de la désarticulation naturelle. Il est préférable de procéder à des récoltes en Décembre, car les graines ne présentent pas d'immaturité et sont aptes à germer; les graines récoltées précocement, présentent de faibles capacités de germination, à cause de la dormance primaire, cette dernière est éliminer à condition de traiter les graines (eau oxygénée à 30 % ou stratification) Les graines peuvent être conservées pendant 6 ans, sans risque de pré-germination. Ainsi même si la périodicité de la fructification est supérieure a 3 années, il serait possible de 168 stoker les graines à condition que la conservation se fasse hors cônes, et que leur teneur en eau soit comprise entre 5,5 % et 7%. Le but de la conservation est de stoker les graines sans détérioration de leurs embryons. Elle doit être réalisée lors d'une année de bonne production. La conservation des graines dans les cônes, est à proscrire sauf peut être pour de courtes périodes. Actuellement, le désailage (séparation des ailes des graines) «artificielle », entraîne des dommages aux graines fertiles, suite à l'éclatement des poches de résines. D'après TOTH (1980) il faudrait s'efforcer de trouver une méthode rationnelle pour séparer l'aile de sa graine sans endommager cette dernière. Le desailage mécanique semble être dépressif sur la faculté germinative (MULLER et al, 1984). Selon BARITEAU et al (1994) l'éclatement des poches de résine (desailage mécanique, transport) diminue la faculté germinative; le taux de réussite est passé de 65 % à 24 % par cumul de stress. Une étude réalisée en 1992 par BARITEAU (1993) avait mis en évidence la grande fragilité des graines de cèdre et la nécessité des recherches complémentaires sur les techniques de tri ayant pour but l'élimination des parasites (Mégastigmus ssp) 169 CHAPITRE II : Influence des prétraitements des
graines sur le
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
-nU Eau KHC1 |
07,50 |
|
- Granulométrie en % |
|
|
Argile |
61,00 |
|
Limon fin |
18,00 |
|
Limon grossier |
03,50 |
|
Sable fin |
04,50 |
|
Sable grossier |
13,00 |
|
- Matière organique |
|
|
C% |
03,17 |
|
N% |
00,15 |
|
C/N |
21,13 |
|
- Bases échaneeables en meq/100g |
|
|
Ca++ |
24,00 |
|
K+ |
00,76 |
|
Na |
00,70 |
|
Mgr |
15,00 |
|
-12' 2'05 en m |
05,00 |
1.3.2 - Les semis :
Les analyses de semis sont faites à 3 mois après le repiquage. Les résultats bruts sont reportés en annexe
Pour caractériser la croissance et le développement des jeunes plantules diverses auteurs notamment ABOUROUH (1983), DRAPIER (1983), BECKER et al (1985) mesurent les paramètres morphologiques (poids, longueur), le nombre de cotylédons et l'analyse minérale.
Les paramètres morphométriques mesurés
pour chaque plantule sont :
- Poids frais de la partie aérienne et
racinaire (mg).
- Poids sec de la partie aérienne et racinaire (mg).
- Longueur de la partie aérienne et racinaire (pivot) en cm. - Nombre de cotylédons.
Quant à l'analyse minérale (Azote, Phosphore, Potassium, Calcium et Magnésium) elle a été faite sur la partie aérienne et racinaire
1.4 - Techniques d'analyse : 1.4.1 - Analyse du sol :
- pH : pH mètre.
- La granulométrie : pipette de ROBINSON.
- Matière organique : le carbone a été déterminé par la méthode ANNE.
- La teneur en M.0 a été détruite en multipliant le taux de carbone par le coefficient
1,72.
- L'azote total : méthode KJELDHAL.
- Bases échangeables : Ca++ et Mg++ par compléxométrie, Na et K+ par photométrie à flamme.
- Phosphore assimilable : méthode JORET HERBERT.
172
1.4.2 - Analyse du végétal :
Le poids a été déterminé par une balance de précision dont l'erreur est de 1/10 mg. La longueur est obtenue à l'aide de papier millimètre et de pied à coulisse (1/20 mm)
Pour l'analyse minérale des semis nous avons utilisé les mêmes méthodes : compléxométrie, photométrie et JORET HERBERT.
Après repiquage nous avons arrêté l'expérimentation au bout de 90 jours. Durant cette période la graine de cèdre n'utilise en principe que ces réserves (ABOUROUH, 1983).
Nous avons observé des mortalités ainsi que des différences morphologiques des semis selon les traitements tant sur les parties aériennes que souterraines.
Les données brutes obtenues après analyse sont reportées dans le tableau en annexe XXII. 2.1 -- Repiquage :
Le nombre et le taux de reprise des graines repiquées après germination dans des « boites germoirs » sont portés dans le tableau ci-dessous.
Tableau N°91: Reprise des graines germées après repiquage dans un sol forestier (Sous-cédraie).
|
Traitements |
Nombre de graines repiquées |
Reprise des graines repiquées |
|
|
Nombre |
Taux (%) |
||
|
Témoin |
30 |
17 |
56,66 |
|
Froid humide |
30 |
30 |
100 |
|
Eau oxygénée |
30 |
28 |
93,33 |
Le tableau n°91, montre que le taux de reprise après repiquage des graines traitées par le froid humide est de 100 %, celui de l'eau oxygénée a donné un taux de 93,33 % alors que pour les graines non traitées les résultats enregistrés sont les plus faibles (56,66 %).
Cette expérience, qui mérite d'être poursuivie est d'un intérêt pratique certain pour les pépiniéristes. En effet, le traitement des graines de cèdre de l'Atlas pour lever la dormance primaire permet d'avoir les meilleurs taux de germination et de reprise.
2.2 - Caractéristiques morphologiques des semis :
Afin de mettre en évidence l'influence des prétraitements des graines sur le développement de semis, nous avons adopté la méthode suivante :
- Analyse des principales caractéristiques morphologiques des semis par des paramètres statistiques.
- Etude du poids, de la longueur et du nombre de cotylédons. - Analyse de la variance pour chaque caractère.
173
2.2.1 - Paramètres statistiques :
Le tableau n°92 qui exprime les caractères morphologiques des semis de cèdre de l'Atlas, élevés sous serres pendant 3 mois, montre que les graines dormantes ayant subies des prétraitements donnent les meilleurs résultats aussi bien pour la partie aérienne que racinaire
La figure n°50 représente la morphologie générale des plants de 3 mois.
Tableau N°92: Paramètres statistiques sur les caractères morphométriques des semis de cèdre de l'Atlas de trois mois. Influence des prétraitements
|
Valeur minimale |
Valeur maximale |
Moyenne |
Coefficient de variation % |
|||||||||||
|
T |
11202 |
Fil |
T |
11202 |
Fil |
T |
11202 |
Fil |
T |
11202 |
Fil |
|||
|
Poids |
Frais |
Tige |
121.70 |
200.6 |
165.2 |
305.20 |
572.6 |
550.2 |
207.45 |
368.54 |
344.01 |
29.41 |
28.46 |
28.39 |
|
Racine |
23.50 |
31.80 |
40.5 |
130.30 |
241 |
288.2 |
42.84 |
139.88 |
124.02 |
54.78 |
43.62 |
48.91 |
||
|
Total |
162.90 |
251.7 |
206.07 |
435.50 |
799.8 |
789.8 |
250.30 |
507.14 |
468.61 |
29.43 |
31.41 |
31.72 |
||
|
Sec |
Tige |
31.31 |
50.2 |
20.6 |
180.60 |
192.1 |
171.3 |
64.95 |
105.46 |
87.95 |
53.79 |
36.38 |
48.12 |
|
|
Racine |
10.54 |
12.07 |
7.09 |
40.10 |
66.30 |
90.0 |
19.46 |
39.15 |
36.15 |
38.15 |
47.90 |
59.63 |
||
|
Total |
50.62 |
63.9 |
41.6 |
200.10 |
225.3 |
250.8 |
84.44 |
143.54 |
124.20 |
44.38 |
37.92 |
42.75 |
||
|
Longueur |
Tige |
4.84 |
5.26 |
4.26 |
6.65 |
6.96 |
7.16 |
5.62 |
6.02 |
5.66 |
9.48 |
9.94 |
12.23 |
|
|
Racine |
4.30 |
5.47 |
5.03 |
11.72 |
24.30 |
22.34 |
6.78 |
14.99 |
14.00 |
23.75 |
36.33 |
34.29 |
||
|
Totale |
9.61 |
10.76 |
9.94 |
17.29 |
31.26 |
28.7 |
12.40 |
21.02 |
19.66 |
14.30 |
27.19 |
27.06 |
||
|
Nombre de cotylédons |
27 |
27 |
23 |
39 |
53 |
51 |
31.11 |
38 |
37 |
12.36 |
18.06 |
17.83 |
||
Nombre de variables 10. Nombre d'observations : 18 pour le témom (T). 28 pour l'eau oxygénée (1202). 30 pour le froid humide (Fil)
2.2.1.1 - Le poids : 2.2.1.1.1 - Le poids frais :
L'examen des données montre que le poids frais des semis de 3 mois de la partie aérienne est compris entre 121,70 pour le témoin et 572.70 mg pour les semis dont les graines ont été traitées par l'eau oxygénée. La plus forte moyenne est de 365.01 mg (H202).Quant à la partie racinaire les chiffres obtenus indiquent que ce poids varie entre 23.50 mg pour le témoin et 288.20 mg pour les semis dont les graines ont été traitées par le froid humide. La meilleure moyenne est obtenue par le prétraitement à l'eau oxygénée de 138.91 mg (11202).
Les valeurs du coefficient de variation, montrent que l'écart entre la partie aérienne des semis est peu important (C.V < 30.00 %), celui de la biomasse racinaire est importante particulièrement pour le Témoin (C.V = 54.98 %).
2.2.1.1.2 - Le poids sec :
Les données obtenues, indiquent que le poids sec des semis de la partie aérienne est compris entre un minimum de 20.06 mg (Froid) et un maximum de 192.10 mg (11202), avec une moyenne de 105.46mg (11202). Celui de la partie racinaire varie entre 7.09 mg (11202) et 90 mg (Froid) La moyenne est de 39.15 mg (11202).
Le poids sec total en mg est compris entre 41.60 (Froid) et 255.30 mg (11202) la moyenne la plus importante est obtenue pour les semis dont les graines ont subi l'influence de l'eau oxygénée (144.62).
Pour cet indice, le coefficient de variation obtenu est supérieur à 36 % quel que soit le traitement. Ceci montre que la biomasse des semis exprime une variabilité selon les traitements ; c'est le traitement par le froid qui donne des semis dont le développement de la partie racinaire présente les différences les plus significatives (C.V. = 59.73 %).
174
|
Fig. 50 : Morphologie de jeunes semis de cèdre de l'Atlas âgés de 3 trois. Influence du froid humide et de l'eau oxygénée. |
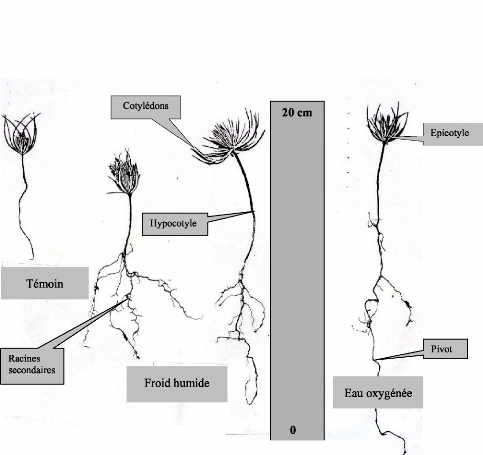
Froid humide
Eau oxygénée
20 cm
0
2.2.1.2 - La longueur :
La longueur de la partie aérienne des semis dont les graines ont été traitées au froid, varie entre 4.26 cm et 7.16 cm. La plus forte moyenne est de 6.02 cm, elle est enregistrée par l'H2O2 ; en effet le développement de la partie aérienne des jeunes plants de cèdre de l'Atlas dont les graines n'ont pas été conditionnées est sensiblement le même. Celui dont les graines ont subi des prétraitements au froid humide et à l'eau oxygénée présentent des différences significatives (C.V = 44.60 %).L'écart de la longueur selon les différents traitements n'est pas significatif.
Quant à celle du pivot, elle est comprise entre 4.30 (Témoin) et 24.30 cm (11202). La plus forte moyenne obtenue est de 14.99 cm (11202). La valeur des coefficients de variation indique que le développement du système racinaire présente de grands écarts entre les semis pour un même traitement.
175
Le nombre de cotylédons est compris entre 23 (Froid) et 53 individus (11202). Le coefficient de variation montre qu'il n'y pas a de différences significatives entre le nombre de cotylédons pour chaque traitement. Le nombre de cotylédons le plus important est enregistré pour le traitement à l'eau oxygénée, (37,44 individus) puis le froid humide (37,27) et enfin le témoin (32,27).
2.2.1.4 - Conclusion
Afin de mieux apprécier le développement des jeunes plantules, nous avons représenté dans le graphe ci-dessous les moyennes de la longueur de la partie aérienne et racinaire, le nombre de cotylédons et le poids sec des deux parties ; ceci montre que le traitement par le froid humide ou l'eau oxygénée a permit non seulement d'accroître le pouvoir germinatif des graines mais active la croissance et le développement des plantules.
q Témoin Froid humide El Eau oxygénée
105,46
87,9E
64,95
31,11
37 38

5,62 5,66 6,02
I I I.

19,46
6,78 14 14,99
36'1539,15
|
Longueur Racine Longueur Tige |
Nombre de |
Poids Sec Racine Poids Sec Tige |
Fig. 51 : Représentation de plant moyen âgé de 3 mois selon les traitements.
L'étude des différentes corrélations a pour objet de déterminer d'éventuelles relations entre les différents caractères morphologiques des semis. La matrice est composée de 10 variables (poids frais total, poids frais de la partie aérienne, poids frais de la parie racinaire, poids sec total, poids sec de la partie aérienne, poids sec de la partie racinaire, la longueur totale, la longueur de la partie aérienne, le pivot et le nombre de cotylédons) à 10 observations pour le témoin, 28 pour l'eau oxygénée et 30 pour le froid humide.
176
Tableau N° 93 : Matrice de corrélation des principaux caractères morphométriques des plantules de cèdre de l'Atlas de 3 mois.
|
TEMOIN |
|||||||||||||||||||
|
Poids frais |
Poids sec |
Longueur |
Nombre de |
||||||||||||||||
|
Total |
Aérien |
Racinai |
Total |
Aérien |
Racinai |
Totale |
Aérienne |
Racinaire |
|||||||||||
|
Poids frais |
total |
1.000 |
|||||||||||||||||
|
aérien |
0.957 |
1.000 |
|||||||||||||||||
|
racinaire |
0.652 |
0.402 |
1.000 |
||||||||||||||||
|
Poids sec |
total |
0.631 |
0.750 |
0.030 |
1.000 |
||||||||||||||
|
aérien |
0.604 |
0.710 |
0.052 |
0.981 |
1.000 |
||||||||||||||
|
racinaire |
0.336 |
0.441 |
-0.092 |
0.428 |
0.244 |
1.000 |
|||||||||||||
|
Longueur |
totale |
0.681 |
0.521 |
0.785 |
0.069 |
0.003 |
0.332 |
1.000 |
|||||||||||
|
aérien |
0.471 |
0.554 |
0.038 |
0.307 |
0.201 |
0.602 |
0.441 |
1.000 |
|||||||||||
|
racinaire |
0.595 |
0.390 |
0.852 |
-0.026 |
-0.063 |
0.167 |
0.955 |
0.155 |
1.000 |
||||||||||
|
Nombre de cotylédons |
0.830 |
0.808 |
0.504 |
0.653 |
0.611 |
0.416 |
0.465 |
0.312 |
0.408 |
1.000 |
|||||||||
|
FROID HUMIDE |
|||||||||||||||||||
|
Poids frais |
Poids sec |
Longueur |
Nombre de |
||||||||||||||||
|
Total |
Aérien |
Racinaire |
Total |
Aérien |
Racinaire |
Totale |
Aérienne |
Racinaire |
|||||||||||
|
Poids frais |
total |
1.000 |
|||||||||||||||||
|
aérien |
0.966 |
1.000 |
|||||||||||||||||
|
racinaire |
0.908 |
0.768 |
1.000 |
||||||||||||||||
|
Poids sec |
total |
0.890 |
0.927 |
0.697 |
1.000 |
||||||||||||||
|
aérien |
0.763 |
0.835 |
0.531 |
0.923 |
1.000 |
||||||||||||||
|
racinaire |
0.701 |
0.652 |
0.679 |
0.654 |
0.314 |
1.000 |
|||||||||||||
|
Longueur |
tntale |
0.887 |
0.865 |
0.794 |
0.796 |
0.621 |
0.748 |
1.000 |
|||||||||||
|
aérien |
0.591 |
0.533 |
0.601 |
0.403 |
0.181 |
0.642 |
0.780 |
1.000 |
|||||||||||
|
racinaire |
0.897 |
0.882 |
0.793 |
0.824 |
0.662 |
0.736 |
0.996 |
0.720 |
1.000 |
||||||||||
|
Nombre de cotylédons |
0.924 |
0.886 |
0.851 |
0.833 |
0.655 |
0.772 |
0.856 |
0.638 |
0.859 |
1.000 |
|||||||||
|
EAU O X Y G É N É E |
|||||||||||||||||||
|
Poids frais |
Poids sec |
Longueur |
Nombre de |
||||||||||||||||
|
Total |
Aérien |
Racinaire |
Total |
Aérien |
Racinaire |
Totale |
Aérienne |
Racinaire |
Cotylédons |
||||||||||
|
Poids frais |
total |
1.000 |
|||||||||||||||||
|
aérien |
0.983 |
1.000 |
|||||||||||||||||
|
racinaire |
0.939 |
0.861 |
1.000 |
||||||||||||||||
|
Poids sec |
total |
0.879 |
0.917 |
0.724 |
1.000 |
||||||||||||||
|
aérien |
0.895 |
0.930 |
0.743 |
0.982 |
1.000 |
||||||||||||||
|
racinaire |
0.824 |
0.870 |
0.662 |
0.978 |
0.935 |
1.000 |
|||||||||||||
|
Longueur |
tntale |
0.805 |
0.759 |
0.821 |
0.610 |
0.629 |
0.552 |
1.000 |
|||||||||||
|
aérien |
0.662 |
0.684 |
0.575 |
0.617 |
0.611 |
0.564 |
0.664 |
1.000 |
|||||||||||
|
racinaire |
0.794 |
0.744 |
0.817 |
0.593 |
0.613 |
0.536 |
0.998 |
0.620 |
1.000 |
||||||||||
|
Nombre de cotylédons |
0.902 |
0.919 |
0.769 |
0.864 |
0.897 |
0.797 |
0.729 |
0.647 |
0.716 |
1.000 |
|||||||||
Nombre de variables : 10.
Nombre d'observations :18 pour le témoin ( T ).28 pour l'eau oxygénée ( 11202).30 pour le froid humide (F.H
|
Test de signification : |
0.05 |
0.01 |
0.001 |
|
Témoin : |
0.468 |
0.561 |
0.673 |
|
H202 |
0.373 |
0.478 |
0.588 |
|
Froid : |
0.361 |
0.462 |
0.570 |
Parmi les différents paramètres morphométriques considérés, on peut constater que les différentes liaisons linéaires sont les plus significatives quand les graines sont traitées.
Ainsi, en prenant par exemple :
-Le poids sec des différentes parties du plant qui semblent mieux rendre compte de l'importance de la production de la matière sèche et donc de celui du développement et de la croissance les jeunes plantules sont hautement significatives.
-Le système racinaire, paramètre très important dont dépend la survie des plantules, est également très significatif rendant compte de son accroissement et donc de l'efficacité de l'absorption hydro-minérale.
-Le nombre de cotylédons permet d'apprécier l'activité photosynthétique, les corrélations obtenues avec la longueur du système racinaire mettent en évidence l'amplification du développement.
177
2.2.3 - Les variations :
L'analyse de la variance a été réalisée sur 17 observations par traitement. Les résultats sont représentés dans le tableau n°94.
Tableau N°94 :Analyse de variance sur les caractères morphologiques des semis de 3mois en fonction du prétraitement des graines par l'eau oxygénée et le froid humide.
|
Source |
Poids Frais |
Poids Sec |
D |
F. ". |
|||||||||||||||||
|
Total |
Partie |
Partie |
Total |
Partie aérienne |
Partie racinaire |
||||||||||||||||
|
C.M |
F.test |
C.M |
F. test |
C.M |
F.test |
C.M |
F. test |
C.M |
F.test |
C.M |
F.test |
||||||||||
|
Traitement |
322534.5 |
13.7 |
121896.58 |
11.74 |
45505.31 |
12.74 |
17239.59 |
5.75 |
7486.59 |
4.18 |
2286.33 |
6.14 |
02 |
3.19 |
|||||||
|
Résiduelle |
23544.60 |
/ |
10383.96 |
/ |
3573.01 |
/ |
2998.71 |
/ |
1793.22 |
/ |
372.10 |
/ |
48 |
||||||||
|
Total |
35503.69 |
/ |
14844.47 |
/ |
5250.31 |
/ |
3568.34 |
/ |
2020.96 |
/ |
448.67 |
/ |
50 |
||||||||
|
Source |
Longueur |
Nombre de |
D |
F |
|||||||||||||||||
|
Totale |
Partie aérienne |
Partie racinaire |
|||||||||||||||||||
|
C.M |
F. test |
C.M |
F. test |
C.M |
F. test |
C.M |
F. test |
||||||||||||||
|
Traitement |
360.10 |
12.86 |
0.77 |
2.23 |
336.59 |
13.88 |
229.23 |
4.64 |
2 |
3.19 |
|||||||||||
|
Résiduelle |
27.99 |
/ |
0.34 |
/ |
3.74 |
/ |
49.36 |
/ |
48 |
||||||||||||
|
Total |
41.28 |
/ |
0.36 |
/ |
36.74 |
/ |
56.55 |
/ |
50 |
||||||||||||
Pour le facteur traitement, cette analyse fait apparaître au seuil de 5% des différences significatives pour les caractères suivants :
- Poids frais et sec : Total, partie aérienne et racinaire
- Longueur : Total et pivot.
- Nombre de cotylédons.
Le test de NEWMAN et KEULS a donné les groupes suivants : 2.2.3.1-Etude pondérale
2.2.3.1.1 - Le poids frais :
Deux groupes homogènes se discriminent pour le poids frais (Cf. Tableau n°95). - Total
- De la partie aérienne
- De la racinaire
Pour ces trois caractères :
Le premier groupe est toujours représenté par le prétraitement à l'eau oxygénée et au froid humide
Le second par le témoin (aucun prétraitement).
Tableau N°95 :Détermination
des groupes homogènes de traitement par le poids frais (total,
partie
aérienne et racinaire) Test de NEWMAN et KEULS.
|
Traitements |
Poids frais total |
Poids frais de la |
Poids frais de la |
Groupes |
|
Eau oxygénée |
503.44 |
360.71 |
136.32 |
A |
|
Froid humide |
471 |
346.51 |
128.02 |
|
|
Témoin |
250.31 |
207.46 |
42.85 |
B |
178
2.2.3.1.2- Le poids sec :
Nous avons obtenu deux groupes homogènes : le poids sec total et celui de la partie racinaire ; le premier est représenté par les prétraitements et le second par le témoin.
Tableau N°96 : Détermination des groupes homogènes de traitement par le poids sec total et de la partie racinaire) Test de NEWMAN et KEULS.
|
Traitements |
Poids sec total |
Poids sec de la
partie |
Groupes homogènes |
|
Eau oxygénée |
144.57 |
39.56 |
A |
|
Froid humide |
129.63 |
38.48 |
|
|
Témoin |
83.49 |
18.96 |
B |
Quant au poids sec de la partie aérienne nous obtenons 3 groupes homogènes : le premier est formé par l'eau oxygénée avec 106.10 mg, le second par le froid humide (groupe intermédiaire) avec 90.25 mg et le témoin constitue le troisième groupe avec seulement 64.62.
Tableau N°97:Détermination des groupes homogènes de traitement par le poids sec de la partie aérienne. Test de NEWMAN et KEULS.
|
Traitements |
Poids sec de la partie aérienne |
Groupes homogènes |
||
|
Eau oxygénée |
106.10 |
A |
||
|
Froid humide |
90.25 |
A B |
||
|
Témoin |
64.52 |
B |
2.2.3.2 - La longueur : 2.2.3.2.1- La longueur totale :
La longueur totale des semis (la tige et pivot) se discriminent en deux groupes homogènes: Le premier est représenté par le traitement de l'eau oxygénée et du froid avec respectivement 20,82 et 19,84 cm. Le second groupe constitue le témoin est formé par les plantules dont les graines non traitées enregistrent une moyenne de 12,41cm (Cf. Tableau n°98).
Tableau N°98 : Détermination de groupes homogènes de traitement par la longueur. Méthode de NEWMAN et KEULS.
|
Traitements |
Longueur totale |
Longueur du pivot |
Groupes homogènes |
|
Eau oxygénée |
20,82 |
14,80 |
A |
|
Froid humide |
19,84 |
14.14 |
|
|
Témoin |
12,41 |
6.79 |
B |
2.2.3.2.2-Le pivot :
Quant à la longueur du pivot (Cf. Tableau n°98) les deux groupes homogènes se séparent. Le premier groupe formé par les plantules dont les graines ont été traitées par l'eau oxygénée et le froid humide avec dans l'ordre 14,80 cm et 14.14 cm. Le second groupe se compose du témoin donne la plus faible longueur avec seulement 6.79 cm.
179
2.2.3.2.3 - La longueur aérienne :
Le tableau ci-dessus montre qu'il n'y a pas de différences significatives entre les moyennes pour les trois traitements. C'est le témoin qui donne les lus mauvais résultats.
Tableau N°99:Détermination des groupes homogènes de traitement par la longueur de la partie aérienne. Test de NEWMAN et KEULS.
|
Traitements |
La longueur aérienne |
Groupes homogènes |
|
Eau oxygénée |
6.02 |
A |
|
Froid humide |
5.70 |
|
|
Témoin |
5.62 |
2.2.3.3 -Les cotylédons :
Pour ce caractère nous obtenons deux groupes homogènes qui se séparent : le premier est formé par les semis dont les graines ont été traitées par l'eau oxygénée et le froid humide avec respectivement les moyennes de 37.67 et 37.29 cotylédons le second constitue le témoin donne 31.12.
Tableau N°100 : Détermination des groupes homogènes de traitement par le nombre de Cotylédons. Test de NEWMAN et KEULS.
|
Traitements |
Cotylédons |
Groupes homogènes |
|
Eau oxygénée |
37.67 |
A |
|
Froid humide |
37.29 |
|
|
Témoin |
31.12 |
B |
Le rôle physiologique des éléments minéraux au sein du végétal est primordial dans les différentes réactions du complexe métabolique.
Parmi les éléments biogènes majeurs qui rentrent dans la composition des différentes substances organiques on peut citer l'azote (N) et le phosphore (P). Les autres éléments (K, Ca, Mg) catalysent les différentes réactions de synthèses et énergétiques d'une part et interviennent dans les mécanismes de construction ainsi que dans les phénomènes de régulation de l'autre part (MAZLIAK, 1981).
Les résultats analytiques de ces éléments biogènes, dans la partie aérienne et racinaire des semis de cèdre de l'Atlas, dont les graines ont été traitées par le froid humide et l'eau oxygénée sont reportés dans le tableau ci-dessous.
Tableau N°101:Composition chimique de la partie aérienne et racinaire (en %) des semis de Cedrus atlantica M âgés de 3 mois
|
Traitements |
Semis |
N |
P |
K |
Ca |
Mg |
|
Témoin |
Aérienne |
0.97 |
0.13 |
0.67 |
0.50 |
0.14 |
|
Racinaire |
0.95 |
0.10 |
0.41 |
0.38 |
019 |
|
|
Froid humide |
Aérienne |
1.92 |
0.20 |
0.86 |
0.61 |
0.16 |
|
Racinaire |
1.86 |
0.17 |
0.47 |
0.42 |
0.19 |
|
|
Eau oxygénée |
Aérienne |
1.95 |
0.21 |
0.93 |
0.66 |
0.16 |
|
Racinaire |
1.90 |
0.19 |
0.51 |
0.43 |
0.21 |
Qui montre que les semis dont les graines ont été traitées donnent les meilleurs résultats. En outre, la composition chimique ayant des graines traitées par l'eau oxygénée est la plus élevée aussi bien pour la partie aérienne que souterraine.
180
Les plantules de Cedrus atlantica M obtenue à partir des graines traitées se sont révélées très intéressante pour étudier :
- Le pouvoir de reprise des graines germées.
- Les caractéristiques morphologiques et minérales des plantules obtenues après repiquage.
Ainsi, notre étude a montré que le nombre de graines germées ayant repris leur développement après repiquage est beaucoup plus intéressant pour les graines traitées. La croissance et le développement des semis sont également stimulés par le froid humide et l'eau oxygénée.
En effet, après traitements des graines, les semis obtenus présentent des caractéristiques morphologiques et minérales largement significatives par rapport à ceux dont les graines n'ont subi aucun prétraitement pour lever la dormance. Cette levée a permis non seulement d'augmenter le pouvoir germinatif et la reprise après repiquage des graines mais active le développement et l'accroissement des jeunes plantules.
Les graines traitées par le froid humide et l'eau oxygénée ont donné des plantules dont les caractéristiques morphologiques sont significatives par rapport au témoin. En effet, les valeurs obtenues par l'étude des variables (le poids frais, le poids sec, la longueur et le nombre de cotylédons) ont montré que le développement et l'accroissement des semis sont activés par les deux prétraitements (Cf. figure n°51).
Les données morphologiques obtenues par ABOUROUH (1983) sur des semis de Cedrus atlantica M. âgés de 4 mois élevés sur sol forestier et celles des semis élevés par nos soins sont reportées sur le tableau ci-dessus.
Tableau N°102 : Données morphologiques du cèdre de l'Atlas
|
Semis de 4 mois |
S Semis de 3 mois |
||||
|
Témoin |
Eau oxygénée |
Froid humide |
|||
|
Longueur. |
Tige |
11,60 |
5.62 |
6.02 |
5.66 |
|
Pivot |
31,80 |
6,78 |
14.99 |
13.87 |
|
|
Poids sec. |
Aérienne ( P.A |
0,538 |
0,083 |
0,129 |
0,144 |
|
Racinaire ( P.R ) |
0,211 |
0,019 |
0,038 |
0,039 |
|
|
Total ( PT) |
0.749 |
0.102 |
0.167 |
0.183 |
|
|
Rapport |
P.A. / P.R. |
2,54 |
4,40 |
3,36 |
3,65 |
|
PR / PT |
0.281 |
0.186 |
0.227 |
0.213 |
|
Les rapports PA/PR et PR/PT ont été utilisés respectivement par ABOUROUH (1983) et DRAPIER (1983); ils traduisent le déséquilibre entre la partie aérienne et souterraine des semis.
En prenant en considération ces rapports nous constatons que les semis issues des prétraitements ont des rapports qui se rapprochent de ceux obtenus par ABOUROUH (1983) nettement plus faibles particulièrement PR/ PT.
181
182
Les semis dont les graines n'ont pas subi de lever de dormance, ont un développement racinaire plus réduit que ceux dont les graines ont été traitées.
Le faible développement du système racinaire rend les semis moins adaptés à l'absorption des éléments minéraux, ceci est peut être l'une des causes de l'absence des semis singulièrement dans les expositions sud du Belezma. PAGES (in PAGES ; 1991) a noté que l'accomplissement des fonctions d'encrage et d'absorption hydro - minérale du système racinaire sont étroitement liées à son architecture c'est à dire à sa forme.
En outre, les semis originaires des graines traitées par l'eau oxygénée favorisent mieux l'assimilation des éléments biogènes.
Notons que pour un même traitement, la partie racinaire a une teneur en éléments majeurs plus faible que la partie aérienne, exception faite au magnésium. Ainsi les semi-issus de graines traitées à l'eau oxygénée sont plus riches en éléments minéraux que ceux du froid humide ; le témoin présente les plus faibles moyennes.
La figure n° 52 et le tableau n° 101 qui représentent les teneurs des éléments minéraux en pourcentage de celui du témoin montre que :
L'azote qui constitue un élément plastique et rentre dans la composition des différentes substances organiques présente la teneur la plus importante par rapport aux autres éléments..
Les semi-issues des graines traitées à l'eau oxygénée présentent les meilleurs teneurs en N avec pour la partie aérienne 201 % et la partie racinaire 201 % par rapport au témoin. Ceux dont les graines sont traitées au froid humide présentent respectivement les valeurs suivantes: 198 % et 196 %.
Ceci montre que les semis des graines traitées assurent leurs fonctions d'assimilation chlorophyllienne d'une manière plus active. Un taux d'azote suffisant assure un bon développement des pousses végétatives.
Quant aux autres éléments nutritifs (P, K, Ca, Mg) qui traduisent le degré de croissance et de développement des semis leur taux est beaucoup plus faible que l'azote, particulièrement le phosphore (0,10 à 0,21 %) et le magnésium (0,14 à 0,21 %).
En effet, le phosphore est un élément plastique (constitution), qui contribue à la division cellulaire et le processus de respiration et de photosynthèse.
Le potassium qui favorise l'assimilation du CO2, est surtout abondant dans les organes à croissance rapide.
Le calcium est surtout nécessaire au déroulement des mitoses, alors que le magnésium qui est un constituant de la chlorophylle est indispensable au déroulement de la photosynthèse.
L'importance donc de ces éléments majeurs (P, K, Ca et Mg) traduit le degré de la croissance et du développement des semis.
Les analyses minérales des semis constituent la meilleure méthode pour l'appréciation de statut nutritionnel.
183
WALKERT et al (in ABOUROUH, 1983) ont donné la concentration en éléments nutritifs des aiguilles du cèdre rouge de l'ouest des U.S.A ; elle est estimée en pourcentage du poids sec :
|
Eléments |
Concentration en % |
|
|
N |
1,5 |
|
|
P |
0.12 |
|
|
K |
0,39 à |
0,78 |
|
Ca |
0,10 à |
0,20 |
|
Mg |
0,12 à |
0,32 |
En prenant en considération ces données analytiques, nous pourrions dire que la concentration en éléments nutritifs pour les semis dont les graines sont traitées, dépasse la concentration foliaire déficitaire.
La biogenèse est fortement influencée par la levée de dormance.
184

185
Les différents travaux sur le cèdre de l'Atlas dans son aire d'origine, montrent que sa régénération naturelle est étroitement liée aux facteurs climatiques : températures et précipitations en particulier ; ces dernières sont très liées à l'exposition et à l'altitude ; ces deux facteurs conditionnent la production des graines (floraison, pollinisation, développement et maturation des cônes) la désarticulation des cônes, la dissémination des graines, la date de germination, le développement des semis et leur capacité de développer un système racinaire, capable d'assurer une alimentation en eau durant la saison estivale. Cette dernière provoque la disparition de la quasi-totalité des jeunes plantules.
BOUDY (1952), MARION (1955), LEPOUTRE (1961), TOTH (1978 a et 1987 b), NEDJAHI (1988), DERRIDJ (1990), LAHMADI et al (1993, EZZAHIRI et al (1994), signalent l'importance de l'implantation du système racinaire des semis de cèdre de l'Atlas, pour échapper à la sécheresse, surtout estivale. Lors de la réunion de travail organisée à Chambéry en 1988 sur les phénomènes de la régénération des forêts d'altitude, divers chercheurs notamment DEMOLIS, PIUSSI, OTT et OSSET insistèrent sur l'importance du travail du sol pour l'amélioration de la régénération.
L'étude climatique a montré que le Belezma est caractérisé par l'irrégularité et l'insuffisance des précipitations et une sécheresse estivale prononcée qui dure plus 6 mois. L'amélioration des conditions édaphiques comme le travail du sol, permet d'améliorer la germination (lit de semence) le développement du système racinaire (facilité de pénétration) et le maintien des semis, en limitant les dégâts causés par le déficit pluviométrique et les hautes altitudes.
Nos différentes observations sur le terrain nous en permis de constater que l'exposition et l'altitude constituent les facteurs les plus influents ; néanmoins, le nombre de semis est extrêmement faible. Afm d'améliorer la régénération, nous avons essayé de créer des conditions édaphiques favorables, grâce à de simples travaux de crochetage du sol ; pour cela, nous avons expérimenter 3 modalités : témoin, travail superficiel et profond du sol.
Avant de présenter la méthodologie d'étude nous permettant d'évaluer l'aptitude à la régénération naturelle au Belezma, il convient d'abord de préciser les conditions des différentes observations :
- La cédraie n'a subi aucun traitement sylvicole ; ces derniers jouent un rôle très important dans la production grainière.
- L'état sanitaire et l'âge sont très variables selon les stations ; ils jouent également un rôle important dans la régénération.
Il n'a été possible de tenir de prendre en considération ces deux points d'où une perte d'information.
Notons que les observations ont été réalisé durant l'année de forte production.
Etant donné que le rayon de la dissémination des graines est de l'ordre de 20 à 50 m autour des semenciers, nous avons été amenés à limiter nos observations à l'intérieur (Sous couvert) et au voisinage des arbres (A découvert).
Afin de suivre l'évolution des semis issus de la régénération au-delà de la saison estivale nous avons installé un dispositif expérimental pendant l'année de forte production, sur deux stations à situation écologique différente : Station à exposition Nord et Station à exposition Sud en prenant en considération l'altitude, le couvert et le degré du travail de sol.
186
Le tableau ci-dessous synthétise la méthode
utilisée pour l'étude de la
régénération.
Tableau N°103:Etude de la
régénération naturelle du cèdre de l'Atlas :
méthodologie générale
|
Exposition |
Altitude (m) |
Couvert |
Nombre de placeaux |
Modalités |
|
Versant [180 placeaux |
< 1700m 10 placettes [90 placeaux ] |
Couvert [45 |
Sous 9 placeaux de 1 m2 chacun par placette: |
x 5 placeaux : Témoin |
|
3 x 5 placeaux : Travail |
||||
|
3 x 5 placeaux : Travail |
||||
|
A [45 |
9 placeaux de 1 m 2 chacun par placette: Soit 15 placeaux par modalité |
3 x 5 placeaux : Témoin |
||
|
3 x 5 placeaux : Travail |
||||
|
3 x 5 placeaux : Travail |
||||
|
1700 - 1800m |
couvert [45 |
Sous 9 placeaux de 1 m2 chacun par placette: |
x 5 placeaux : Témoin |
|
|
3 x 5 placeaux : Travail |
||||
|
3 x 5 placeaux : Travail |
||||
|
A [45 |
9 placeaux de 1 m2 chacun par |
3 x 5 placeaux : Témoin |
||
|
3 x 5 placeaux : Travail |
||||
|
3 x 5 placeaux : Travail |
||||
|
Versant [180 placeaux del m2 chacun] |
< 1700m |
couvert [45 |
Sous 9 placeaux de 1 m2 chacun par placette: |
x 5 placeaux : Témoin |
|
3 x 5 placeaux : Travail |
||||
|
3 x 5 placeaux : Travail |
||||
|
A [45 |
9 placeaux de 1 m2 chacun par |
3 x 5 placeaux : Témoin |
||
|
3 x 5 placeaux : Travail |
||||
|
3 x 5 placeaux : Travail |
||||
|
1700 - 1800m |
couvert [45 |
Sous 9 placeaux de 1 m2 chacun par placette: |
x 5 placeaux : Témoin |
|
|
3 x 5 placeaux : Travail |
||||
|
3 x 5 placeaux : Travail |
||||
|
A [45 |
9 placeaux de 1 m2 chacun par |
3 x 5 placeaux : Témoin |
||
|
3 x 5 placeaux : Travail |
||||
|
3 x 5 placeaux : Travail |
187
Pour chaque station, nous avons choisi deux tranches altitudinales : la première est inférieure à 1.700 m et la seconde est comprise entre 1.700 m et 1.800 m.
Dans chaque tranche nous avons installé 10 placettes de forme carrée ayant chacune 100m2, 5 sous couverts et 5 à découvert ; chaque placette est divisée en placeaux de 1 m2. On a choisi 9 placeaux par placette. Pour chaque placette, nous avons retenu trois modalités pour trois placeaux: travail profond du sol (T.P.S)-travail superficiel du sol (T.S.S)-sol intact (Témoin: T)
Ainsi, le nombre total de placeaux de 1m2 sur lequel sont faites toutes les observations est 360, soit 180 par exposition, ce qui représente 90 par tranche altitudinale, 45 sous couvert et 45 à découvert (15 pour le témoin, 15 pour le travail superficiel du sol et 15 pour le travail profond du sol)
Les premiers travaux de préparation du sol ont été effectués au mois d'Octobre, avant la désarticulation des cônes. Durant ce mois, le sol est relativement humide ce qui facilite son crochetage. Un second travail a été réalisé en Novembre pour mieux ameublir et affiner le crochetage, afin de permettre aux eaux de pluie de s'infiltrer.
Pour faciliter les observations, nous avons utilisé un gabarit en fer forgé ayant la même forme et la même superficie que les plateaux (1m2).
Pour éviter toutes confusions entre les placeaux, le premier et le second placeau ont été matérialisés par des piquets métalliques peints respectivement en rouge et en blanc. Un carnet de terrain, nous permettra de situer l'ensemble des plateaux et le report de toutes les observations.
Les résultats bruts obtenus sont consignés en annexe XX et XXI
Afin de mettre en évidence l'influence de l'altitude, du couvert et du travail du sol sur la germination, le développement et la survie des semis de l'année au - delà de la sécheresse estivale, nous avons adopté après la récolte des données la démarche suivante :
- Nous avons dans un premier temps illustré nos résultats par des courbes afin de suivre l'évolution mensuelle du nombre de semis.
- Ensuite nous avons étudié statistiquement les deux stations séparément en même temps en considérant les observations mensuelles comme étant des répétitions ; puis, une autre analyse pour chaque mois, en utilisant l'analyse de la variance.
- Enfin, une analyse en composantes principales (A.C.P) qui étudie l'évolution de la germination de l'ensemble des placettes en fonction des mois d'observations. Elle nous permettra d'identifier les catégories de placettes les plus convenables au développement des semis, selon leurs particularités,
Les représentations graphiques, figure n°54, montrent que le nombre de semis diminue progressivement en fonction du temps. En effet, à partir d'Avril pour l'exposition Sud et Mai, mois qui correspondent au nombre le plus élevé de plantules, nous assistons à une diminution graduelle des semis ; les reliquats obtenus à la fin de la saison estivale sont très faibles à nuls.
En effet, la figure n°53, le tableau n°104 et celui en annexe XXII résument la situation des semis après la saison estivale : - Le nombre de plantules, les reliquats ainsi que les semis disparus ou en voie de l'être (jaunissement) et sains - Le taux de réussite
188
189
190
Tableau N°104 : Evolution du nombre de semis de l'année en fonction du degré du travail du sol et du couvert selon l'exposition.
|
Altitude inférieure à 1700 m |
Altitude comprise entre 1700 - 1800 |
||||||||||||
|
Sous Couvert |
I A Découvert |
Sous Couvert |
I A Découvert |
||||||||||
|
T |
T.S.S |
T.S.P |
T |
T.S.S |
T.S.P |
T |
T.S.S |
T.S.P |
T |
T.S.S |
T.S.P |
||
|
Versant Nord |
Avril |
49 |
79 |
112 |
45 |
88 |
134 |
62 |
79 |
129 |
57 |
120 |
223 |
|
Décembre |
15 |
21 |
46 |
16 |
29 |
54 |
23 |
28 |
51 |
18 |
42 |
87 |
|
|
Taux de mortalité en % |
69 |
73 |
59 |
64 |
67 |
60 |
63 |
64 |
60 |
68 |
65 |
61 |
|
|
Versant Sud |
Avril |
10 |
17 |
31 |
09 |
19 |
25 |
28 |
29 |
54 |
13 |
23 |
49 |
|
Décembre |
00 |
02 |
09 |
00 |
00 |
00 |
02 |
08 |
12 |
00 |
00 |
03 |
|
|
Taux de mortalité en % |
100 |
88 |
71 |
100 |
100 |
100 |
93 |
72 |
78 |
100 |
100 |
94 |
|
Léeende : T : Témoin - T.S.S : Travail superficiel du sol - T.S.P : Travail du sol profond
Contrairement à l'exposition Sud, l'exposition Nord est plus favorable à la survie des semis de l'année. En effet, selon les traitements, le taux de mortalité est compris entre 59 et 73 % pour l'exposition Nord, celui de l'exposition Sud varie entre 71 et 100 %. Le découvert a plutôt une action défavorable.
En vue de comparer l'influence de 4 stations différentes sur l'évolution des semis naturels depuis leurs apparitions jusqu'à la dissémination des graines de l'année d'après, nous avons procédé à l'analyse de la variance de manière à mettre en évidence l'existence d'éventuelles différences entre les semis et d 'éventuelles interactions.
Tableau N°105: Etude de l'influence de l'Exposition, l'Altitude, du Couvert et du degré de travail du sol sur le nombre moyen des semis - naturels par l'analyse de la variance
|
Source de variation |
Carré moyen |
DDL |
F test |
F théorique |
|
|
Facteur 1 : Exposition |
180511.42 |
1 |
364.35 |
3.888 |
|
|
Facteur 2 : Altitude |
9652.16 |
1 |
19.48 |
||
|
Facteur 3 : Couvert |
1804.16 |
1 |
3.64 |
||
|
Facteur 4 : travail du sol |
32771.59 |
2 |
66.15 |
3.038 |
|
|
Interactions |
F 1 -F2 |
3792.03 |
1 |
7.65 |
3.888 |
|
F 1 -F3 |
7326.05 |
1 |
14.79 |
||
|
F 1 -F4 |
17157.88 |
2 |
34.63 |
3.038 |
|
|
F2-F3 |
951.84 |
1 |
1.92 |
3.888 |
|
|
F2-F4 |
1355.42 |
2 |
2.74 |
3.038 |
|
|
F3-F4 |
1772.22 |
2 |
3.58 |
||
|
F 1 -F2-F3 |
2343.91 |
1 |
4.73 |
3.888 |
|
|
F 1-F2-F4 |
1119.30 |
2 |
2.26 |
3.038 |
|
|
F 1 -F3-F4 |
2581.49 |
2 |
5.21 |
||
|
F2-F3-F4 |
1120.92 |
2 |
2.26 |
||
|
F 1 -F2-F3-F4 |
599.33 |
2 |
1.21 |
||
|
Résiduelle |
495.43 |
216 |
|||
|
Totale |
1800.63 |
239 |
|||
3.2.1- Les facteurs :
Il fait apparaître pour les critères de classification étudiés des différences significatives au seuil de 5 % pour : l'exposition, l'altitude et degré de travail du sol. Quant au couvert, les différences ne sont pas significatives
3.2.1.1- Facteur 1 : Exposition :
Les deux expositions se différencient en deux groupes homogènes. Le tableau n°106 montre que la station à exposition Nord (plus humide et plus fraîche) est la plus favorable à la régénération naturelle.
191
Tableau N°106:Caractérisation des groupes homogènes de l'exposition par le nombre moyen de semis. Test de NEWMAN et KEULS au seuil de 5%.
|
Stations |
Moyennes |
Groupes homogènes |
|
Exposition Nord |
65.87 |
A |
|
Exposition Sud |
11.02 |
B |
3.2.1.2 - Facteur 2 : L'Altitude :
Les deux tranches altitudinales se différencient en deux groupes homogènes : le premier est formé par les hautes altitudes avec des moyennes intéressantes.
Tableau N°107: Caractérisation des groupes homogènes d'altitude par le nombre moyen de semis. Test de NEWMAN et KEULS au seuil de 5%.
|
Altitudes |
Moyennes |
Groupes homogènes |
|
Altitude comprise entre 1.700 -1800 m |
44.78 |
A |
|
Altitude inférieure à 1.700 m |
32.10 |
B |
En effet, le tableau n°107 montre que les altitudes situées entre 1700-1800 m sont les plus favorables à la régénération naturelle. Ces dernières sont plus arrosées et plus fraîches.
3.2.1.3- Facteur 3 : Le Couvert :
Le facteur couvert n'a aucune influence sur la régénération. (Cf. Tableau n°108). En effet, l'analyse n'a donné qu'un seul groupe homogène ; toutefois les stations à découvert permettent aux semis de mieux se développer.
Tableau N°108 : Caractérisation des groupes homogènes du couvert par le nombre moyen de semis. Test de NEWMAN et KEULS au seuil de 5%.
|
Couvert |
Moyennes |
Groupes homogènes |
|
A Découvert |
41.18 |
A |
|
A Couvert |
35.78 |
L'influence modérée du couvert, peut s'expliquer par la densité des peuplements. 3.2.1.4 - Facteur 3 : Le Degré de travail du sol :
Le tableau n°109 montre que le degré de travail du sol se discrimine en trois groupes.
Tableau N°109: Détermination des groupes homogènes du degré de travail du sol par le nombre moyen de plantules. Test de NEWMAN et KEULS.
|
Degré de travail du sol |
Nombre moyen de semis |
Groupes homogènes |
|
|
Travail profond du sol (T.P.S) |
60.20 |
A |
|
|
Travail superficiel du sol (T.S.S) |
34.95 |
B |
|
|
Témoin (T) |
20.17 |
C |
Le premier groupe avec 60.20 plantules correspond au travail profond du sol (T.P.S), le second groupe composé par le travail superficiel du sol (T.S.S) avec 34.95 plantules et le dernier groupe sans travail du sol (Témoin) n'a enregistré qu'une moyenne de 20.17 plantules.
Ce critère, dévoile que le cèdre se régénère d'une manière plus réussie quand le substrat est travaillé et ameubli profondément.
192
3.1.2-Etude des interactions :
Etant donné que la régénération naturelle du cèdre de l'Atlas est incertaine, l'étude des différentes interactions devient très délicate, car elle dépend de l'ampleur de l'incidence du paramètre le plus significatif.
Les résultats de ces interactions sont significatifs pour celles formées par : [Exposition -
Altitude] ; [ Exposition -- Couvert ] ; [Exposition -- Travail du sol] ;[Altitude-Couvert ] ; [Altitude -- Travail du sol ] ; [Couvert - Travail du sol] ; [Exposition - Altitude -- Couvert ] ; [Exposition - Altitude -- Travail de sol] ; [Exposition - Couvert - Travail du sol] ; [Altitude - Couvert - Travail du sol ] [ Exposition - Altitude - Couvert -- Travail du sol]
Le test de NEWMAN et KEULS au seuil de 5 % a permis de mettre en évidence des groupes homogènes selon les paramètres étudiés.
3.1.2.1 - Exposition - Altitude :
L'influence synchronique de l'Exposition et de l'Altitude montre que l'exposition est le paramètre le plus influent. Ainsi les plus fortes moyennes sont dénombrées sur l'exposition Nord quelle que soit l'altitude.
Tableau N°110 : Caractérisation des groupes homogènes de l'exposition et de l'altitude par le nombre moyen de semis. Test de NEWMAN et KEULS au seuil de 5%.
|
Interaction : Exposition - Altitude |
Moyennes |
Groupes homogènes |
|
|
Exposition Nord --Altitude 1700-1800m |
76.18 |
A |
|
|
Exposition Nord --Altitude <1700m |
55.55 |
B |
|
|
Exposition Sud --Altitude 1700-1800m |
13.38 |
C |
|
|
Exposition Sud --Altitude <1700m |
8.65 |
En effet, la lecture du tableau n° 110 montre que la station à exposition Nord à haute altitude est de loin celle qui a permis une meilleure régénération, elle forme le premier groupe avec une moyenne de 76,18 plantules ; le second groupe avec 55.55 semis se compose de l'interaction Exposition Nord --Altitude <1700m.Le troisième groupe est formé par deux interactions Exposition Sud --Altitude 1700-1800m et Exposition Sud --Altitude <1700m avec les plus médiocres résultats, donne 13,38 et 8.65 plantules.
Les hautes altitudes associées aux expositions favorables assurent les meilleures conditions favorables à la régénération.
3.1.2.2 - Exposition - Couvert :
Cette interaction formée par l'Exposition et le Couvert et dont les résultats sont portés sur le tableau n°111, a permis d'avoir 3 groupes homogènes qui se discriminent nettement.
Tableau N°111: Caractérisation des groupes homogènes de l'exposition et du couvert par le nombre moyen de semis. Test de NEWMAN et KEULS au seuil de 5%.
|
Exposition - Couvert |
Moyennes |
Groupes homogènes |
||
|
Exposition Nord- A découvert |
74.13 |
A |
||
|
Exposition Nord -- A couvert |
57.60 |
B |
||
|
Exposition Sud -- A découvert |
13.80 |
C |
||
|
Exposition Sud -- A couvert |
8.23 |
Cette interaction est semblable à celle de l'Exposition - Altitude. En effet, nous obtenons le même nombre de groupes homogènes et c'est l'exposition qui semble être le facteur déterminant.
193
Points vus
Points cachés
Les résultats de cette interaction, qui sont portés sur le tableau n° 112, ont permis d'avoir 4 groupes homogènes :
Le premier est composé de l'interaction Exposition Nord -- Travail profond du sol avec une moyenne de 103.45 plantules.
Le second est formé par l'interaction Exposition Nord - Travail superficiel du sol avec 59.63 semis
Le troisième groupe est constitué par l'interaction Exposition Nord -- Témoin avec seulement 34.53 semis
Le quatrième et dernier est composé de 3 interactions de l'exposition Sud avec 16.95, 10.27 et 5.82 plantules selon le degré de travail du substrat.
Tableau N°112:Caractérisation des groupes homogènes de l'exposition et de degré de travail du sol par le nombre moyen de plantules. Test de NEWMAN et KEULS.
|
Exposition - Degré de travail du sol |
Moyennes |
Groupes homogènes |
||
|
Exposition Nord -- Travail profond du sol |
103.45 |
A |
||
|
Exposition Nord - Travail superficiel du sol |
59.63 |
B |
||
|
Exposition Nord -- Témoin |
34.53 |
C |
||
|
Exposition Sud -- Travail profond du sol |
16.95 |
D |
||
|
Exposition Sud -- Travail superficiel du sol |
10.27 |
|||
|
Exposition Sud - Témoin |
5.82 |
|||
Cette interaction montre que l'influence de deux facteurs favorables (l'exposition Nord et le travail profond du sol) engendre des circonstances bénéfiques à la régénération. L'exposition constitue le paramètre principal.
3.1.2.4 - Altitude - Couvert :
L'action simultanée du Couvert et de l'Altitude prise en considération n'est pas significative: un seul groupe est obtenu. Toutefois, Les hautes altitudes associées au découvert semblent être les meilleures stations ; à couvert la régénération est moindre à cause probablement de l'ensoleillement (Cf. Tableau n '113).
Tableau N°113 : Caractérisation des groupes homogènes de l'altitude et du couvert par le nombre moyen de semis. Test de NEWMAN et KEULS au seuil de 5%.
|
Altitude - Couvert |
Moyennes |
Groupes homogènes |
|
Altitude comprise entre 1.700 -1800 m- A découvert |
46.52 |
A |
|
Altitude comprise entre 1.700 -1800 m- Sous - Couvert |
40.05 |
|
|
Altitude comprise < 1.700 m - A découvert |
32.85 |
|
|
Altitude comprise < 1.700 m- Sous - Couvert |
31.35 |
En effet, les hautes altitudes donnent les résultats compris entre 46.52 et 40.05, par contre les basses altitudes n'ont enregistré que 31.35 et 32.85 selon le couvert.
3.1.2.5 - Altitude -- Travail du sol
Le tableau N°114 montre que l'action simultanée de l'Altitude et du Travail du sol n'est pas significative pour l'ensemble des placettes : un seul groupe est obtenu.
194
Tableau N°114 : Caractérisation des groupes homogènes de l'altitude et du couvert par le nombre moyen de semis. Test de NEWMAN et KEULS au seuil de 5%.
|
Altitude -- Degré de travail du sol |
Moyennes |
Groupes homogènes |
|
Altitude comprise entre 1.700 -1800 m- Travail profond du sol |
71.28 |
A |
|
Altitude comprise < 1.700 m - Travail profond du sol |
49.13 |
|
|
Altitude comprise entre 1.700 -1800 m- Travail superficiel du sol |
39.30 |
|
|
Altitude comprise < 1.700 m- Travail superficiel du sol |
30.60 |
|
|
Altitude comprise entre 1.700 -1800 m- Témoin |
23.77 |
|
|
Altitude comprise < 1.700 m- Témoin |
16.58 |
Néanmoins, les tranches altitudinales les plus hautes associées à l'importance du degré du travail du sol assurent les meilleures conditions favorables à la régénération.
3.1.2.6 -Couvert - Degré de travail du sol :
Le tableau n°115 montre que cette interaction a donné 4 groupes qui se discriminent distinctement.
Le premier groupe avec 67.88 semis est composé par le travail profond du sol et le découvert.
Le second avec 52.53 est formé par les stations A Couvert -- Travail profond du sol.
Le troisième groupe est constitué de 2 interactions : A Découvert -- Travail superficiel
du sol A Couvert -- Travail superficiel du sol avec respectivement 37.20 et 32.70 semis.
Le dernier groupe avec les plus faibles moyennes est composé par les placettes A
Découvert et A Couvert sans le travail du sol.
Tableau N°115:Caractérisation des groupes homogènes de l'exposition et du degré de travail du sol par le nombre moyen de semis. Test de NEWMAN et KEULS.
|
Exposition -- Degré de travail du sol |
Moyennes |
Groupes homogènes |
|||
|
A Découvert - Travail profond du sol |
67.88 |
A |
|||
|
Sous Couvert -- Travail nrofond du sol |
52.53 |
B |
|||
|
A Découvert -- Travail superficiel du sol |
37.20 |
C |
|||
|
Sous Couvert -- Travail superficiel du sol |
32.70 |
||||
|
Sous Couvert - Témoin |
21.88 |
D |
|||
|
A Découvert - Témoin |
18.48 |
||||
Cette interaction montre que le degré de travail du sol est le paramètre le plus déterminant.
3.1.2.7 -Exposition - Altitude - Couvert :
La distribution des groupes homogènes obtenue pour cette interaction (ExpositionAltitude-Couvert) a donné 3 groupes homogènes :
Le premier groupe est constitué par l'interaction Exposition Nord-Altitude 17001800m-- A Découvert avec semis avec une moyenne de 89.57 semis.
Le deuxième groupe est formé par 3 interactions à savoir l'Exposition Nord - Altitude 1700-1800m-Sous Couvert, l'Exposition Nord--Altitude < 1700m-A Découvert et l'Exposition Nord -- Altitude < 1700 m -- Sous Couvert avec respectivement 62.80, 58.70 et 52.40 plantules.
Le troisième groupe est composé par l'exposition Sud avec les quatre niveaux étudiés.
195
Tableau N°116:Caractérisation des groupes homogènes de l'interaction Exposition-Altitude et Couvert par le nombre moyen de semis. Test de NEWMAN et KEULS.
|
Exposition - Altitude - Couvert : |
Moyennes |
Groupes homogènes |
|
Exposition Nord - Altitude 1700-1800m -- A Découvert |
89.57 |
A |
|
Exposition Nord - Altitude 1700-1800m -- Sous Couvert |
62.80 |
B |
|
Exposition Nord -- Altitude < 1700 m -- A Découvert |
58.70 |
|
|
Exposition Nord -- Altitude < 1700 m -- Sous Couvert |
52.40 |
|
|
Exposition Sud - Altitude 1700-1800m -- Sous Couvert |
17.30 |
C |
|
Exposition Sud -- Altitude < 1700 m -- Sous Couvert |
10.30 |
|
|
Exposition Sud - Altitude 1700-1800m -- A Découvert |
9.47 |
|
|
Exposition Sud -- Altitude < 1700 m-- A Découvert |
7.00 |
Cette interaction entre les 3 facteurs, montre que les plus favorables (Exposition Nord, les hautes altitudes et le découvert) permettent aux jeunes semis de trouver des conditions propices à leur croissance.
Les résultats de cette interaction sont portés sur le tableau N° 117 qui montre que l'influence simultanée de l'Exposition - Altitude -- Travail de sol n'a donné qu'un seul groupe homogène.
Tableau N°117 : Caractérisation des groupes homogènes de l'exposition de l'altitude et du travail du sol par le nombre moyen de semis. Test de NEWMAN et KEULS.
|
Exposition - Altitude -- Degré de travail du sol. |
Moyennes |
Groupes homogènes |
|
Exposition Nord - Altitude 1700-1800m- Travail profond |
122.80 |
A |
|
Exposition Nord - Altitude <1700 m- Travail profond |
84.10 |
|
|
Exposition Nord - Altitude 1700-1800 m- Travail superficiel |
66.15 |
|
|
Exposition Nord - Altitude <1700 m- Travail superficiel |
53.10 |
|
|
Exposition Nord - Altitude 1700-1800m- Témoin |
39.60 |
|
|
Exposition Nord - Altitude <1700 - Témoin |
29.45 |
|
|
Exposition Sud - Altitude 1700-1800m - Travail profond |
19.75 |
|
|
Exposition Sud - Altitude <1700 m - Travail profond |
14.15 |
|
|
Exposition Sud - Altitude 1700-1800m - Travail superficiel |
12.45 |
|
|
Exposition Sud - Altitude <1700 m - Travail superficiel |
8.10 |
|
|
Exposition Sud - Altitude 1700-1800m - Témoin |
7.95 |
|
|
Exposition Sud - Altitude <1700 m - Témoin |
3.70 |
Cependant, l'influence de facteurs favorables à savoir l'Exposition Nord- l'Altitude 1700-1800m -- Travail profond du sol facilite la régénération ; par contre celle de facteurs défavorables comme l'Exposition Sud - l'Altitude <1700m -- Sol non travaillé sont néfaste.
Notons que, ce sont les placettes situées sur l'exposition Nord qui permettent au cèdre de l'Atlas de se développer convenablement.
3.1.2.9- Exposition - Couvert - Travail du sol :
Cette interaction montre que l'influence simultanée des facteurs favorables donne les meilleurs résultats. Les groupes homogènes obtenus sont au nombre de 6 :
Le premier groupe est formé par l'interaction exposition Nord et le découvert avec 122.80 plants. Le second groupe est composé par l'interaction exposition Nord -Sous couvert avec 84.10 plants. Le troisième groupe est formé par deux interactions Exposition Nord - A découvert--
Travail superficiel du sol et Exposition Nord - Sous Couvert-- Travail superficiel du sol avec
respectivement 66.30 et 52.95 semis.
196
Tableau N°118: Caractérisation des groupes homogènes de l'exposition, du couvert et du travail du sol par le nombre moyen de semis par le test de NEWMAN et KEULS.
|
Exposition - Couvert - Degré de travail du sol. |
Moyennes |
Groupes homogènes |
|
Exposition Nord - A découvert- Travail profond du sol |
122.80 |
A |
|
Exposition Nord- Sous Couvert- Travail profond du sol |
84.10 |
B |
|
Exposition Nord - A découvert-- Travail superficiel du sol |
66.30 |
C |
|
Exposition Nord - Sous Couvert-- Travail superficiel du sol |
52.95 |
|
|
Exposition Nord - Sous Couvert -Témoin |
37.75 |
D |
|
Exposition Nord- A découvert-Témoin |
33.30 |
|
|
Exposition Sud - Sous Couvert - Travail profond du sol |
20.95 |
DE |
|
Exposition Sud - A découvert - Travail profond du sol |
12.95 |
E |
|
Exposition Sud - Sous Couvert - Travail superficiel du sol |
12.45 |
|
|
Exposition Sud - A découvert - Travail superficiel du sol |
8.10 |
|
|
Exposition Sud - Sous Couvert-Témoin |
8.00 |
|
|
Exposition Sud - A découvert-Témoin |
3.65 |
Le quatrième groupe est formé également par deux interactions : Exposition Nord - Sous Couvert --Témoin avec 37.75 plantules et Exposition Nord- A découvert-Témoin avec 33.30 plantules.
Le cinquième groupe constitué par l'interaction Exposition Sud - Sous Couvert - Travail profond du sol avec 20.95 plantules ; c'est un groupe intermédiaire entre le quatrième et le sixième.
Le sixième et dernier groupe est formé par 5 interactions de l'exposition Sud.
Pour cette interaction nous pouvons tirer les mêmes conclusions que l'interaction Exposition - Altitude -- Travail de sol : c'est l'exposition qui forme le facteur déterminant.
3.1.2.10 -Altitude - Couvert - Degré de travail du sol
L'action simultanée de l'Altitude, du Couvert et le Degré de travail du sol n'est pas significative: un seul groupe est obtenu. Cependant, la sommation des facteurs propice au développement des jeunes plantules donne les meilleures moyennes.
Tableau N°119 : Caractérisation des groupes homogènes de l'Altitude et du Couvert et du Travail du sol par le nombre moyen de semis. Test de NEWMAN et KEULS.
|
Altitude -- Couvert -- Degré de travail de sol : |
Moyennes |
Groupes homogènes |
|
Altitude 1700-1800m - A Découvert - Travail profond |
85.00 |
A |
|
Altitude 1700-1800m - Sous Couvert - Travail profond |
57.55 |
|
|
Altitude <1700 m - A Découvert - Travail profond |
50.75 |
|
|
Altitude <1700 m - Sous Couvert - Travail profond |
47.50 |
|
|
Altitude 1700-1800m - A Découvert - Travail superficiel |
42.80 |
|
|
Altitude 1700-1800m - Sous Couvert - Travail superficiel |
35.80 |
|
|
Altitude <1700 m - A Découvert - Travail superficiel |
31.60 |
|
|
Altitude 1700-1800m - Sous Couvert - Travail superficiel |
29.60 |
|
|
Altitude 1700-1800m - Sous Couvert - Témoin |
26.80 |
|
|
Altitude 1700-1800m - A Découvert - Témoin |
20.75 |
|
|
Altitude <1700 m - Sous Couvert - Témoin |
16.95 |
|
|
Altitude <1700 m - A Découvert - Témoin |
16.20 |
Ainsi les placettes ayant les caractéristiques suivantes Altitude 1700-1800m-A Découvert - Travail profond, ont donné 85.00 plantules, par contre celles situées à basse altitude à couvert ou à découvert sans aucun travail de sol n'ont donné moins de 16 plantules.
3.1.2.11 - Exposition - Altitude - Couvert -- Degré de travail du sol :
Cette interaction montre que l'influence «Exposition-Altitude-Couvert-Travail » du sol n'est pas significative ; en effet nous obtenons un seul groupe homogène (Cf. tableau n°120)
197
Tableau N°120 : Caractérisation des groupes homogènes de l'exposition de l'altitude, du couvert et du travail du sol d'altitude par le nombre moyen de semis.
|
Exposition - Altitude - Couvert - Degré de travail du sol |
Moyenne |
C V (%) |
G.H |
||
|
Exposition Nord-Altitude 1700-1800m- A Découvert -Travail profond |
154.20 |
65. 85 |
36.04 |
67.02 |
A |
|
Exposition Nord-Altitude 1700-1800m- Sous Couvert -Travail profond |
91.40 |
35.50 |
|||
|
Exposition Nord-Altitude <1700m- A Découvert -Travail profond |
91.40 |
36.49 |
|||
|
Exposition Nord-Altitude 1700-1800m- A Découvert -Travail |
76.90 |
41.27 |
|||
|
Exposition Nord-Altitude <1700m- Sous Couvert -Travail profond |
76.68 |
35.66 |
|||
|
Exposition Nord-Altitude <1700m- A Découvert -Travail superficiel |
55.70 |
44.28 |
|||
|
Exposition Nord-Altitude 1700-1800m- Sous Couvert -Travail |
55.40 |
39.96 |
|||
|
Exposition Nord-Altitude <1700m- Sous Couvert - Travail superficiel |
50.50 |
48.61 |
|||
|
Exposition Nord-Altitude 1700-1800m- Sous Couvert -Témoin |
41.60 |
37.87 |
|||
|
Exposition Nord-Altitude 1700-1800m- A Découvert -Témoin |
37.87 |
41.60 |
|||
|
Exposition Nord-Altitude <1700m- Sous Couvert -Témoin |
29.90 |
45.59 |
|||
|
Exposition Nord-Altitude <1700m- A Découvert -Témoin |
29.00 |
41.26 |
|||
|
Exposition Sud-Altitude 1700-1800m- Sous Couvert -Travail nrofond |
23.70 |
11. 02 |
61.63 |
104.77 |
|
|
Exposition Sud-Altitude <1700 m - Sous Couvert -Travail profond |
18.20 |
52.21 |
|||
|
Exposition Sud-Altitude 1700-1800m - Sous Couvert -Travail |
16.20 |
56.62 |
|||
|
Exposition Sud-Altitude 1700-1800m - A Découvert -Travail profond |
15.80 |
100.34 |
|||
|
Exposition Sud-Altitude 1700-1800m - Sous Couvert -Témoin |
12.00 |
93.31 |
|||
|
Exposition Sud-Altitude <1700 m - A Découvert -Travail profond |
10.10 |
112.14 |
|||
|
Exposition Sud - Altitude 1700 -1800m - A Découvert - Travail |
08.70 |
104.34 |
|||
|
Exposition Sud-Altitude <1700 m - Sous Couvert -Travail superficiel |
08.70 |
68.78 |
|||
|
Exposition Sud-Altitude <1700m - A Découvert -Travail superficiel |
07.50 |
117.79 |
|||
|
Exposition Sud-Altitude <1700m - Sous Couvert -Témoin |
04.00 |
114.01 |
|||
|
Exposition Sud-Altitude 1700-1800m - A Découvert -Témoin |
03.90 |
128.43 |
|||
|
Exposition Sud-Altitude <1700m - A Découvert -Témoin |
03.40 |
123.52 |
|||
Cependant le facteur Exposition semble être le plus sigmficatif par rapport aux autres. En effet, les interactions composées de l'exposition Nord sont les plus propices ; nous observons aussi que la régénération des différentes placeaux est extrêmement variable, particulièrement celle du Sud
Les résultats de cette analyse montrent que, la régénération de la cédraie de Belezma est régie facteurs dont leur influence est variable ; ainsi, selon l'importance de l'un ou de l'autre nous pouvons les classer en 3 types :
Le premier facteur, que l'on peut qualifier de limitant, est capital: C'est l'exposition. En effet quelle que soit l'interaction, c'est toujours ce paramètre qui détermine l'importance de la régénération.
Le second, dont les effets sur les semis se font sentir, quand le paramètre limitant est franchement défavorable : c'est le facteur altitudinal.
Le troisième facteur à savoir le couvert, qui n'a pas pratiquement aucun effet, quand le premier ou le second facteur sont défavorables.
Le travail du sol, facteur améliorant, pourrait être classé comme secondaire.
La sommation de deux ou plusieurs facteurs positifs comme l'exposition Nord, les hautes altitudes et le travail profond du sol amplifie la régénération ; alors que s'ils sont négatifs comme l'exposition sud, les basses altitudes et le découvert, les semis n'ont aucune chance de s'installer.
3.2.2 - Analyse par mois :
Cette étude qui analyse l'évolution mensuelle des semis pour chaque mois en fonction de l'exposition, de l'altitude, du couvert, du degré de travail du sol et de leurs interactions sont reportés dans le tableau n°121. Elle fait ressortir au seuil de 5% des différences significatives. Afm d'obtenir des discriminations entre les moyennes selon les caractéristiques des stations, l'analyse de la variance est complétée par le test de NEWMAN et KEULS.
198
199
3.2.2.1 - Facteur 1 : L'exposition :
Pour chaque mois les deux types de stations se discriminent en deux groupes homogènes (Cf. Tableau n°122). La station Nord qui est la plus favorable forme le premier groupe et donne toujours les meilleurs résultats depuis l'apparition des semis (Mars) jusqu'au mois de Décembre.
Tableau N°122:Détermination des groupes homogènes de l'exposition par le nombre de semis naturels. Test de NEWMAN et KEULS.
|
Mois |
Expositions |
Nombre de semis |
Groupes Homogènes |
|
Mars |
Versant Nord |
93.42 |
A |
|
Versant Sud |
22,58 |
B |
|
|
Avril |
Versant Nord |
98.08 |
A |
|
Versant Sud |
25,58 |
B |
|
|
Mai |
Versant Nord |
97.83 |
A |
|
Versant Sud |
20,75 |
B |
|
|
Juin |
Versant Nord |
88.83 |
A |
|
Versant Sud |
17,92 |
B |
|
|
Juillet |
Versant Nord |
7250 |
A |
|
Versant Sud |
07,17 |
B |
|
|
Août |
Versant Nord |
54.00 |
A |
|
Versant Sud |
03,83 |
B |
|
|
Septembre |
Versant Nord |
44,08 |
A |
|
Versant Sud |
03,42 |
B |
|
|
Octobre |
Versant Nord |
3758 |
A |
|
Versant Sud |
03,08 |
B |
|
|
Novembre |
Versant Nord |
36.50 |
A |
|
Versant Sud |
03.00 |
B |
|
|
Décembre |
Versant Nord |
35.83 |
A |
|
Versant Sud |
03,00 |
B |
Notons une décroissance progressive du nombre de plantules en fonction du temps. Ainsi, pour l'exposition favorable nous passons de 98.08 à 35.83 plantules, soit une perte 63.44% ; quant à l'exposition Sud, le nombre qui été de 25,58 atteint après la période estivale et automnale, 3 plantules, soit une perte de 22.58 plantules ce qui correspond à 86.71%.
L'exposition est le facteur déterminant, elle influence non seulement la germination mais également la survie des semis quelle que soit la période. Ainsi, la configuration des monts de Belezma, crée des conditions assez particulières qui font que les stations situées sur les expositions nord sont plus arrosées et plus froides que celles localisées sur les expositions sud
3.2.2.2 - Facteur 2 : L'altitude :
Le tableau n°123 montre que les deux altitudes se discriminent pour les mois humides : Mars Avril et Mai.
Pour les mois secs (Juin et Juillet) il n'y a pas de différences entre les deux altitudes, celui d'Août, les deux altitudes se discriminent, celle comprise à 1.700 - 1800 m enregistre une survie de 34,50 plantules, l'altitude inférieure à 1.700 m a permis d'avoir que 23,33 plantules. Les mois de Septembre, Octobre, Novembre et Décembre n'enregistrent aucune différence significative.
L'altitude comprise à 1.700 - 1800 m a permis la meilleure survie que l'altitude inférieure à 1.700 m et que nous enregistrons une mortalité progressive des semis en relation avec le temps.
200
Tableau N°123: Détermination des groupes homogènes de l'altitude par le nombre de semis.
|
Mois |
Altitudes |
Nombre de semis |
Grounes Homogènes |
||
|
Mars |
Altitude 1700 -1800 m |
66.25 |
A |
||
|
Altitude inférieure à 1700 m |
49,75 |
B |
|||
|
Avril |
Altitude 1700 -1800 m |
72.17 |
A |
||
|
Altitude inférieure à 1700 m |
51,50 |
B |
|||
|
Mai |
Altitude 1700 -1800 m |
68.58 |
A |
||
|
Altitude inférieure à 1700 m |
50,00 |
B |
|||
|
Juin |
Altitude 1700 -1800 m |
61.91 |
A |
||
|
Altitude inférieure à 1700 m |
43.93 |
||||
|
Juillet |
Altitude 1700 -1800 m |
45.10 |
A |
||
|
Altitude inférieure à 1700 m |
34,50 |
||||
|
Août |
Altitude 1700 -1800 m |
34.50 |
A |
||
|
Altitude inférieure à 1700 m |
23,33 |
B |
|||
|
Septembre |
Altitude 1700 -1800 m |
28.50 |
A |
||
|
Altitude inférieure à 1700 m |
19,00 |
||||
|
Octobre |
Altitude 1700 -1800 m |
24.41 |
A |
||
|
Altitude inférieure à 1700 m |
16,25 |
||||
|
Novembre |
Altitude 1700 -1800 m |
23.50 |
A |
||
|
Altitude inférieure à 1700 m |
16,00 |
||||
|
Décembre |
Altitude 1700 -1800 m |
22.83 |
A |
||
|
Altitude inférieure à 1700 m |
16.00 |
L'analyse de ce facteur, montre que durant la saison printaniere, période humide, les hautes altitudes sont propices a la régénération ; mais dès que les chaleurs estivales arrivent, la tranche altitudinale étudiée n'arrive pas à compenser l'effet néfaste de ces températures.
3.2.2.3 - Facteur 3: le couvert :
L'influence de ce facteur n'est pas significative. Lesmoyennes sont portées dans le tableau n° 124. Tableau N°124 : Détermination des groupes homogènes du couvert par le nombre de semis
|
Mois |
Couvert |
Nombre de semis par |
Groupes Homogènes |
|
Mars |
A Découvert |
63.33 |
A |
|
Sous couvert |
52.67 |
||
|
Avril |
A Découvert |
67.08 |
A |
|
Sous couvert |
56.58 |
||
|
Mai |
A Découvert |
64.08 |
A |
|
Sous couvert |
54.50 |
||
|
Juin |
A Découvert |
56.08 |
A |
|
Sous couvert |
50.66 |
||
|
Juillet |
A Découvert |
42.16 |
A |
|
Sous couvert |
37.49 |
||
|
Août |
A Découvert |
30.50 |
A |
|
Sous couvert |
27.33 |
||
|
Septembre |
A Découvert |
25.83 |
A |
|
Sous couvert |
21.25 |
||
|
Octobre |
A Découvert |
21.41 |
A |
|
Sous couvert |
19.25 |
||
|
Novembre |
A Découvert |
21.00 |
A |
|
Sous couvert |
18.75 |
||
|
Décembre |
A Découvert |
20.75 |
A |
|
Sous couvert |
18.08 |
Si le couvert n'a pas beaucoup d'influence, à cause probablement de la faible densité, il faut noter que la moyenne de la germination, de lacroissance et du développement sont supérieurs à découvert que sous couvert; ceci peut s'expliquer par le fait que sur l'exposition Nord, le nombre de plants est de loin très important à découvert (l'ensoleillement n'est pas très important et l'humidité est persistante).
201
Le test de NEWMAN et KEULS au seuil de 5 %, a permis de discriminer le degré de travail du sol en deux ou trois groupes homogènes selon les mois (Cf. Tableau n°125).
Tableau N°125: Détermination des groupes homogènes de degré du travail du sol par le nombre de naturels. Test de NEWMAN et KEULS,
|
Mois |
Degré du travail du sol |
Nombre de semis |
Groupes |
Homogènes |
|
|
Mars |
Travail profond |
86,50 |
A |
||
|
Travail superficiel |
54,63 |
B |
|||
|
Aucun (Témoin.) |
32,88 |
C |
|||
|
Avril |
Travail profond |
94,63 |
A |
||
|
Travail superficiel |
65,75 |
B |
|||
|
Aucun (Témoin.) |
34,13 |
C |
|||
|
Mai |
Travail profond |
89,63 |
A |
||
|
Travail superficiel |
55,63 |
B |
|||
|
Aucun (Témoin.) |
35,63 |
C |
|||
|
Juin |
Travail profond |
82,38 |
A |
||
|
Travail superficiel |
49,00 |
B |
|||
|
Aucun (Témoin.) |
28,75 |
||||
|
Juillet |
Travail profond |
62,63 |
A |
||
|
Travail superficiel |
38,00 |
B |
|||
|
Aucun (Témoin.) |
18,88 |
C |
|||
|
Août |
Travail profond |
47,50 |
A |
||
|
Travail superficiel |
25,25 |
B |
|||
|
Aucun (Témoin.) |
14,00 |
||||
|
Septembre |
Travail profond |
38,88 |
A |
||
|
Travail superficiel |
20,75 |
AB |
|||
|
Aucun (Témoin.) |
11,63 |
B |
|||
|
Octobre |
Travail profond |
33,88 |
A |
||
|
Travail superficiel |
20,75 |
B |
|||
|
Aucun (Témoin.) |
09,88 |
||||
|
Novembre |
Travail profond |
33,25 |
A |
||
|
Travail superficiel |
16,25 |
B |
|||
|
Aucun (Témoin.) |
9,75 |
||||
|
Décembre |
Travail profond |
32,75 |
A |
||
|
Travail superficiel |
16,25 |
B |
|||
|
Aucun (Témoin.) |
09,25 |
||||
Les trois premiers mois, à savoir Mars, Avril et Mai se discriminent en 3 groupes homogènes: le premier, formé par le travail profond du sol a permis d'avoir les meilleurs résultats, le second groupe qui a donné des résultats intermédiaires, se compose par le travail superficiel du sol. Le troisième et dernier groupe avec les plus faibles résultats forme le témoin (sol non travaillé).
Le quatrième mois (Juin) est formé par deux groupes homogènes: le premier est représenté respectivement par le travail profond du sol et le second est constitué par le sol travaillé superficiellement et non travaillé.
Le mois de juillet comme les trois premiers mois (Mars, Avril et Mai) se compose de trois groupes homogènes : le travail profond du sol le travail superficiel du sol et enfin le témoin.
Le sixième mois (Août) est analogue au mois de Juin avec deux groupes homogènes.
Le mois de Septembre est composé de trois groupes: le premier est constitué par le travail profond du sol (T.P.S) avec 38,88 plantules. Le dernier groupe (témoin) où les plantules arrivent difficilement à survivre (11,63 plantules). Le deuxième groupe est formé par le travail superficiel du sol présente des résultats intermédiaires entre le premier et le troisième groupe avec 20.75 semis.
202
Les trois derniers mois (Octobre, Novembre et Décembre) forment comme le mois d'Août deux groupes homogènes qui se discriminent nettement : le premier groupe est formé par le travail profond du sol favorisant avec une survie importante de plantules. Le second groupe avec de faibles résultats, se compose du travail superficiel du sol et du témoin.
Le travail du sol, que nous avons classé comme facteur secondaire, est déterminant pendant les premiers mois humides; mais, dés que la sécheresse s'installe, c'est la qualité du travail du sol qui devient décisive, en effet le travail profond est concluant ; en revanche, le travail superficiel n'a pratiquement plus d 'effet.
3.2.3.5 - Les interactions :
3.2.3.5.1 - Exposition - Couvert (F1 -F3) :
Les résultats de cette interaction sont portés dans le tableau n '126.
Tableau N°126 : Détermination des groupes homogènes de l'interaction exposition-couvert par le nombre de semis de la régénération naturelle.Test de NEWMAN et KEULS
|
Mois |
Exposition - Couvert |
Moyennes |
Groupes homogènes |
|
|
Mars |
Versant Nord |
A découvert |
106.67 |
A |
|
Sous couvert. |
80.17 |
B |
||
|
Versant Sud |
Sous couvert. |
25.17 |
C |
|
|
A découvert |
20.00 |
|||
|
Avril |
Versant Nord |
A découvert |
111.17 |
A |
|
Sous couvert. |
85.00 |
|||
|
Versant Sud |
Sous couvert. |
28.17 |
||
|
A découvert |
23.00 |
|||
|
Mai |
Versant Nord |
A découvert |
110.17 |
A |
|
Sous couvert. |
85.50 |
|||
|
Versant Sud |
Sous couvert. |
23.50 |
||
|
A découvert |
18.00 |
|||
|
Juin |
Versant Nord |
A découvert |
97.83 |
A |
|
Sous couvert. |
79.83 |
|||
|
Versant Sud |
Cnne rnnvert |
11 Sn |
||
|
A découvert |
14.33 |
|||
|
Juillet |
Versant Nord |
A découvert |
80.83 |
A |
|
Sous couvert. |
64.16 |
|||
|
Versant Sud |
Sous couvert. |
10.83 |
||
|
A découvert |
03.50 |
|||
|
Août |
Versant Nord |
A découvert |
59.83 |
A |
|
Sous couvert. |
48.17 |
|||
|
Versant Sud |
Sous couvert. |
06.50 |
||
|
A découvert |
01.17 |
|||
|
Septembre |
Versant Nord |
A découvert |
50.00 |
A |
|
Sous couvert. |
36.50 |
|||
|
Versant Sud |
Sous couvert. |
06.00 |
||
|
A découvert |
00.83 |
|||
|
Octobre |
Versant Nord |
A découvert |
42.33 |
A |
|
Sous couvert. |
32.83 |
|||
|
Versant Sud |
Sous couvert. |
05.67 |
||
|
A découvert |
00.50 |
|||
|
Novembre |
Versant Nord |
A découvert |
41.50 |
A |
|
Sous couvert. |
32.00 |
|||
|
Versant Sud |
Sous couvert. |
05.50 |
||
|
A découvert |
00.50 |
|||
|
Décembre |
Versant Nord |
A découvert |
41.00 |
A |
|
Sous couvert. |
30.67 |
|||
|
Versant Sud |
Sous couvert. |
05.50 |
||
|
A découvert |
00.50 |
|||
203
Cette interaction a permis d'avoir trois groupes homogènes uniquement pour le mois de Mars. Les autres mois (Avril, Mai, Juin, Juillet, Août, Septembre, Octobre, Novembre et Décembre) ne présentent aucune différence.
En effet, le mois de Mars est constitué par les interactions suivantes:
-Station à exposition Nord -- découvert : moyenne de plantules est de 106,67. -Station à exposition Nord -- couvert : moyenne de 80,17 semis
-Station à exposition Sud - couvert et Station à exposition Sud découvert avec respectivement des moyennes de 25,17 et 20 plantules.
Celles d'Avril, Mai, Juin, Juillet, Août, Septembre Octobre Novembre et Décembre constitue un seul groupe pour les deux ; cependant le maintien des semis est meilleur sur la station à exposition Nord.
En examinant le tableau n°127, nous constatons que : les six premiers mois, à savoir Mars, Avril, Mai, Juin, Juillet et Août, 3 groupes homogènes se discriminent : le premier groupe est représenté par le versant Nord - travail profond du sol avec la meilleure moyenne, le second groupe est formé par le versant Nord - travail superficiel du sol avec une moyenne intermédiaire et le troisième est constitué par 4 interactions : exposition Nord - témoin (sol non travaillé), exposition Sud - travail profond du sol, exposition Sud - travail superficiel du sol et exposition sud - sol non travaillé.
Quant au mois de Septembre, il ne constitue qu'un seul groupe, il n'y a aucune différence significative entre les 6 interactions.
Quant aux trois derniers mois, Octobre Novembre et Décembre, l'interaction est représentée par deux groupes homogènes qui se fractionnent nettement, le premier est formé par le versant Nord - travail profond du sol, le second groupe est constitué par 5 interactions a savoir versant Nord - travail superficiel du sol, versant Nord - sol non travaillé et enfin l'exposition Sud -3 degrés de travail du sol (T.P.S, T.S.S et T).
Cette interaction montre que l'association de la station à exposition Nord (station favorable) associée au travail profond du sol, donne les meilleures moyennes quelque soit la période des observations. Par contre, quand le travail est superficiel son effet devient de plus en plus insignifiant au fur et à mesure que les températures augmentent et que les pluies se raréfient. A partir du mois d'Octobre, période des premières chutes pluies automnales et la diminution des températures, nous assistons à une reprise du développement et de l'accroissement des semis.
3.2.3 - Conclusion :
Cette étude montre que la régénération du cèdre de l'Atlas est extrêmement complexe; en effet, non seulement tous les facteurs agissent simultanément, mais l'ampleur de leurs actions diffère selon la station ; de plus, un facteur peut dans certains cas, favorise ou défavorise l'effet d'un ou plusieurs paramètres. Nous avons aussi révélé que, de part leur fragilité les semis, (Cf. fig.51) éprouvent de grandes difficultés pour passer l'année estivale ; il faut donc trouver des solutions pour permettre aux semis de s'installer ("irrigation", fertilisation, mycorhization...)
Aussi, l'utilité de cette de cette analyse, est d'avoir répertorier par ordre d'importance l'ampleur de tous les facteurs analysés.
204
Tableau N°127: Détermination des groupes homogènes de l'interaction exposition-travail du sol par le nombre de semis.
|
Mois |
Exposition -Travail du sol |
Nombre de semis |
Groupes homogènes |
|
|
Mars |
Versant Nord |
Travail profond du sol |
141.25 |
A |
|
Travail superficiel du |
87.75 |
B |
||
|
Témoin |
51.25 |
C |
||
|
Versant Sud |
Travail profond du sol |
31.75 |
||
|
Travail superficiel du |
21.50 |
|||
|
Témoin |
14.50 |
|||
|
Avril |
Versant Nord |
Travail profond du sol |
149.50 |
A |
|
Travail superficiel du |
91.50 |
B |
||
|
Témoin |
53.25 |
C |
||
|
Versant Sud |
Travail profond du sol |
39.75 |
||
|
Travail superficiel du |
22.00 |
|||
|
Témoin |
15.00 |
|||
|
Mai |
Versant Nord |
Travail profond du sol |
149.75 |
A |
|
Travail superficiel du |
91.50 |
B |
||
|
Témoin |
52.25 |
C |
||
|
Versant Sud |
Travail profond du sol |
29.50 |
||
|
Travail superficiel du |
19.75 |
|||
|
Témoin |
13.00 |
|||
|
Juin |
Versant Nord |
Travail profond du sol |
138.50 |
A |
|
Travail superficiel du |
81.00 |
B |
||
|
Témoin |
47.50 |
C |
||
|
Versant Sud |
Travail profond du sol |
26.25 |
||
|
Travail superficiel du |
17.00 |
|||
|
Témoin |
10.50 |
|||
|
Juillet |
Versant Nord |
Travail profond du sol |
113.75 |
A |
|
Travail superficiel du |
68.75 |
B |
||
|
Témoin |
35.00 |
|||
|
Versant Sud |
Travail profond du sol |
11.50 |
||
|
Travail superficiel du |
07.25 |
|||
|
Témoin |
02.75 |
|||
|
Août |
Versant Nord |
Travail profond du sol |
88.25 |
A |
|
Travail superficiel du |
46.25 |
B |
||
|
Témoin |
27.50 |
C |
||
|
Versant Sud |
Travail profond du sol |
06.75 |
||
|
Travail superficiel du |
04.25 |
|||
|
Témoin |
00.25 |
|||
|
Septembre |
Versant Nord |
Travail profond du sol |
71.75 |
A |
|
Travail superficiel du |
37.75 |
|||
|
Témoin |
22.75 |
|||
|
Versant Sud |
Travail profond du sol |
06.25 |
||
|
Travail superficiel du |
04.25 |
|||
|
Témoin |
00.50 |
|||
|
Octobre |
Versant Nord |
Travail profond du sol |
61.75 |
A |
|
Travail superficiel du |
31.75 |
B |
||
|
Témoin |
19.25 |
|||
|
Versant Sud |
Travail profond du sol |
06.00 |
||
|
Travail superficiel du |
02.75 |
|||
|
Témoin |
00.50 |
|||
|
Novembre |
Versant Nord |
Travail nrnfnnr1 tin en] |
Afi Sn |
A |
|
Travail ennerfiriel tin en] |
ln fifi |
B |
||
|
Témoin |
19.00 |
|||
|
Versant Sud |
Travail nrnfnnr1 tin en] |
fi4 fifi |
||
|
Travail superficiel du sol |
02.50 |
|||
|
Témoin |
00.50 |
|||
|
Décembre |
Versant Nord |
Travail nrnfnnr1 tin en] |
SQ Sfi |
A |
|
Travail ennerfiriel tin en] |
ln fifi |
B |
||
|
Témoin |
18.00 |
|||
|
Versant Sud |
Travail nmfoncl (ln sol |
fi6 fill |
||
|
Travail superficiel du sol |
02.50 |
|||
|
Témoin |
00.50 |
|||
205
Etant donné que la méthode statistique utilisée précédemment à savoir le test de NEW et KEULS qui analysait une seule caractéristique à la fois, nous avons jugé utile de traiter l'ensemble des variables par une analyse en composantes principales (A.C.P) afin d'individualiser les différentes placettes par l'ensemble des variables étudiées.
Cette méthode qui est principalement descriptive, traite donc l'évolution du nombre de semis inventoriés de chaque mois en fonction des caractéristiques de chaque placette, à savoir l'exposition, l'altitude, le couvert et le travail de sol. Selon PHILIPEAU (1985) cette méthode consiste à présenter sous forme de graphique les informations d'un tableau formé de lignes et colonnes. Dans le cas de notre étude, les lignes sont composées plateaux selon leurs caractéristiques sur lesquels sont portées en colonnes les variables (nombre de semis)
3.3.2 -Variables à analyser :
L'analyse a porté sur l'évolution du nombre de semis depuis leurs apparitions jusqu'à la dissémination des graines de l'année suivante, de l'ensemble des 360 plateaux de 1m2 réparties sur 40 placettes à raison de 9 placeaux par placette : 3 pour le témoin, 3 pour le travail superficiel du sol et 3 pour le travail profond du sol.
3.3.3 - Présentation des résultats et discussion : 3.3.3.1-Matrice de corrélations :
La matrice de corrélation fait apparaître (Cf. tableau n '128) des relations linaires positives et négatives entre le nombre de semis observés par mois et certaines stations selon leurs caractéristiques. Ainsi, l'exposition exprime mieux une étroite liaison négative avec la régénération naturelle et /ou assistée quelle que soit la période ; par contre la manifestation du travail du sol est de moindre importance. Le couvert et l'altitude (1700-1800m) ne présentent aucune liaison particulière.
Tableau n° 128 : Matrice de corrélation.
|
Exp. |
Alt. |
Couv |
Trav |
Mars |
Avril |
Mai |
Juin |
Juil |
Août |
Sept |
Oct |
Nov |
Déc |
||
|
Exp. |
1.000 |
||||||||||||||
|
Alt. |
0.000 |
1.000 |
|||||||||||||
|
Couve. |
0.000 |
0.000 |
1.000 |
||||||||||||
|
Trav. sol |
0.000 |
0.000 |
0000 |
1.000 |
|||||||||||
|
P E I O D E |
Mars |
-0.642 |
0.084 |
0.111 |
0.456 |
1.000 |
|||||||||
|
Avril |
-0.717 |
0.204 |
0.104 |
0.489 |
0.997 |
1.000 |
|||||||||
|
Mai |
-0.741 |
0.179 |
0.092 |
0.447 |
0.998 |
0.994 |
1.000 |
||||||||
|
Juin |
-0.740 |
0.178 |
0.057 |
0.457 |
0.995 |
0.992 |
0.998 |
1.000 |
|||||||
|
Juil |
-0.751 |
0.174 |
0.058 |
0.428 |
0.992 |
0.984 |
0.993 |
0.994 |
1.000 |
||||||
|
Août |
-0.760 |
0.169 |
0.048 |
0.414 |
0.988 |
0.981 |
0.991 |
0.995 |
0.992 |
1.000 |
|||||
|
Sept |
-0.750 |
0.175 |
0.061 |
0.410 |
0.991 |
0.985 |
0.993 |
0.995 |
0.991 |
0.997 |
1.000 |
||||
|
Oct |
-0.745 |
0.172 |
0.053 |
0.426 |
0.989 |
0.986 |
0.992 |
0.994 |
0.987 |
0.994 |
0.997 |
1.000 |
|||
|
Nov |
-0.754 |
0.173 |
0.050 |
0.429 |
0.987 |
0.984 |
0.991 |
0.992 |
0.986 |
0.993 |
0.995 |
0.999 |
1.000 |
||
|
Déc |
-0.746 |
0.155 |
0.061 |
0.436 |
0.989 |
0.986 |
0.991 |
0.993 |
0.987 |
0.992 |
0.996 |
0.999 |
0.999 |
1.00 |
|
significatif à 5% : 0.406
significatif à 1% : 0.517
significatif à 1 %o : 0.630
206
3.3.3.2-Détermination des composantes principales
La caractérisation des composantes principales est réalisée essentiellement sur la part de l'information exprimée par la variance maximale sur les principaux axes, c'est à dire le pourcentage de la variance expliquée par les axes en valeurs propres.
Tableau N°129: Contribution à la variance totale (pourcentage expliqué par les 5 premiers axes)
|
Axe n° |
1 |
2 |
3 |
4 |
5 |
|
Pourcentage de chaque axe |
76.7 |
7.2 |
7.2 |
7.2 |
1.4 |
|
Pourcentage cumulé |
76.7 |
83.9 |
91.3 |
98.3 |
99.7 |
La proportion de variance occupée par les deux premiers axes (axe 1 et axe 2) est de 83.9%. Donc, nous nous tenons à l'interprétation de ces deux composantes qui sont explicatives.
3.3.3.3-Etude des deux premières composantes :
Le sens de chaque axe est expliqué par la corrélation du nombre de semis, en relation avec les composantes principales.
Ainsi l'étude de cette matrice, représentée par le tableau n°130, entre les deux axes (F1 et F2) et les variables, nous autorise à déterminer l'organisation de ces variables sur le plan 1-2 du cercle de corrélation.
Le premier et le second axe sont corrélés négativement avec l'exposition ; le travail du sol est négativement lié à l'axe 2, par contre la liaison est faible mais positive à l'axe 1; par contre le couvert et l'altitude ne présentent aucune liaison par rapport a l'axe 1.
Tableau n°130: Matrice de corrélations entre les variables et les 5 principaux axes.
|
Variables |
N° des composantes (axes) |
|||||
|
1 |
2 |
3 |
4 |
5 |
||
|
Exposition |
-0.7633 |
-0.4667 |
0.0840 |
0.2511 |
0.3597 |
|
|
Altitude |
0.1740 |
-0.4022 |
0.8134 |
-0.3735 |
-0.0815 |
|
|
Couvert |
0.0711 |
-0.5624 |
-0.5694 |
-0.5945 |
-0.0306 |
|
|
Travail de sol |
0.4492 |
-0.5514 |
-0.0836 |
0.6663 |
-0.2075 |
|
|
Période. |
Mars |
0.9959 |
-0.0381 |
-0.0233 |
-0.0120 |
0.0275 |
|
Avril |
0.9928 |
-0.0752 |
0.0064 |
0.0070 |
0.0078 |
|
|
Mai |
0.9973 |
-0.0235 |
-0.0064 |
-0.0090 |
0.0326 |
|
|
Juin |
0.9979 |
-0.0086 |
0.0130 |
0.0180 |
0.0335 |
|
|
Juillet |
0.9943 |
0.0343 |
-0.0331 |
0.0097 |
0.0183 |
|
|
Août |
0.9951 |
0.0335 |
0.0125 |
-0.0069 |
0.0456 |
|
|
Septembre |
0.9957 |
0.0208 |
0.0110 |
-0.0174 |
0.0663 |
|
|
Octobre |
0.9958 |
0.0159 |
0.0129 |
0.0009 |
0.0607 |
|
|
Novembre |
0.9955 |
0.0197 |
0.0140 |
0.0013 |
0.0402 |
|
|
Décembre |
0.9957 |
0.0132 |
-0.0064 |
0.0087 |
0.0533 |
|
Mars
Mai - Juin
Juillet
Août - Septembre - Octobre - Novembre - Décembre
207
Les périodes sont situées sur le cercle de corrélation et donc, fortement corrélées à l'axe F1 ; elles sont groupées en trois nuages :
Mars, Mai, Juin
Avril
Juillet, Août, Septembre, Octobre, Novembre et Décembre.
Le mois de Mars, période d'apparition des jeunes plantules de cèdre révèle leurs importances selon la qualité de la station.
Avril est caractérisé par la germination tardive.
Mai, Juin nous renseignent sur le prélude de mortalité
Juillet, Août, Septembre, Octobre, Novembre et Décembre nous informent sur mortalité graduelle des plantules
La représentation et l'organisation de ces variables, sont illustrées par la figure ci- dessous.
|
F2 |
|||
|
Fl |
|||
|
Fig. 56 : Cercle des corrélations. |
|||
Une variable est d'autant mieux représentée sur le plan qu'elle est plus proche du cercle. 3.3.4. -Etude de l'organisation des stations
La figure 57 qui représente la projection des points correspondants aux différentes placettes dans le premier plan principal : F1 -- F2 ; il explique 83.9 % de l'inertie totale et, reflète donc la quasi-totalité de l'information.
L'axe F1 explique à lui seul 76.3 % de l'inertie totale et indique l'exposition : c'est l'axe de l'évolution des semis.
La seconde composante qui n'explique que 7.2% de la variance totale ; elle caractérise l'altitude et le couvert.
Les différentes placettes sont groupées en deux grands ensembles groupes A et B, dont leur distribution avec une augmentation du nombre de plantules progresse de la gauche vers la droite.
208
L'ensemble A
Il est formé par les placettes de l'exposition Nord ; ce sont les plus favorables à la régénération, il est composé de 3 nuages : Al, A2 et A3
Le premier (Al):
Il est caractérisé par le plus grand nombre de plantules, particulièrement durant la période estivale ; il est formé par les placettes dont les caractéristiques sont :
Altitude 1700 /1800m--Travail profond du sol-A découvert (parcelle12: 10.28 semis par m2)
En effet ce groupe est fortement corrélé aux mois d'Août et Septembre ; ceci montre clairement l'importance du travail profond du sol sur la survie des semis, particulièrement quand les autres facteurs ne s'éloignent pas de l'optimum.
Le second nuage (A2)
Il est déterminé par des régénérations acceptables comprises entre 91.40 et 76.70 semis; il est formé par 4 placettes ayant les caractéristiques suivantes :
Altitude1700/1800m --Travail profond du sol - Sous couvert (parcelles 9 : 6.09 semis/m2) Altitude<1700 m --Travail profond du sol - A découvert (parcelles 6 : 6.09 semis/m2) Altitude 1700/1800 m --Travail superficiel du sol - A découvert (parcelles 11: 5.12 semis/m2) Altitude < 1700m -- Travail profond du sol - Sous couvert (parcelles 3 : 5.11 plantules/m2)
Le nuage (A3)
Il forme les placettes dont la régénération est relativement faible mais convenable ; il comprend également 7 placettes :
Altitude<1700 m -- Travail superficiel du sol - A découvert (parcelles 5 : 3.71 semis/m2) Altitude 1700 / 1800m --Travail superficiel du sol- Sous couvert (parcelles 8 : 3.69 sentis/m2) Altitude< 1700 m-- Travail superficiel du sol - Sous couvert (parcelles 2 : 3.36 plantules/m2) Altitude 1700 / 1800m -- Témoin- Sous couvert (parcelles 7 : 2.77setnis/m2)
Altitude 1700 / 1800m -- Témoin- A découvert (parcelles 10 : 2.50semis/m2) Altitude <1700 m -- Témoin- A découvert (parcelles 4 : 1.93 semishn2)
Altitude <1700 m-- Témoin- Sous couvert (parcelles 1:1.99 semis/m2)
L'ensemble B :
Il est composé l'ensemble des placettes de l'exposition Sud (parcelles 13 à 24); toutes ces stations sont caractérisées par de très faibles densités de plantules (entre 0.26 à 1.58 plantules par m2). Il est indiqué par une survie extrêmement médiocre de semis ; toutes les placettes sont situées à l'extrémité gauche de l'axe Fl.
3.3.5-Conclusion :
L'avantage de cette analyse, nous a permis de réunir dans un ensemble ou sous- ensemble, les placettes qui expriment les mêmes affinités. Ainsi, les placettes localisées dans l'exposition Nord, les hautes altitudes, le découvert et ayant subies un travail profond du sol (placettes n°12) présentent les conditions optimales a la régénération naturelle : germination et maintien des plantules après la saison sèche.
Le travail du sol est globalement positif et améliore nettement les conditions du milieu quand les autres facteurs sont convenables (placettes n°003, 006 et 009); mais dés que ces derniers deviennent défavorables, particulièrement durant la période allant du printemps jusqu'à la fin de l'automne, son effet est insignifiant.
Les tranches altitudinales sont peu influentes, à cause probablement de l'intervalle pris en considération d'une part, d'autre part la faible densité à l'hectare, expliquent éventuellement, son inefficacité quant à la protection des jeunes semis, particulièrement durant la période des hautes chaleurs.
209
210
Après la dissémination des graines (décembre - janvier) et suite à l'élévation de la température, celles fertiles ayant absorbées au moins 50% d'eau par rapport à leur propre poids s'ouvrent au printemps en libérant au mois de Mars et Avril une plantule formée d'un hypocotyle portant une rosette de cotylédons disposés en spirale dont le nombre varie entre 3 et 9 selon les individus; il augmentera pour atteindre 20 à 50 individus selon les semis au bout de 3 mois.
Contrairement aux semis élevés en pépinière qui atteignent des dimensions appréciables (15 à 20 cm) en une année, ceux de la régénération naturelle ne mesurent que 5-8 cm de longueur, après déploiement des feuilles cotylédonaires ; ils sont de ce fait confrontés au stress hydrique puis thermique durant la période estivale, ensuite celui du froid pendant la période hivernale.(Cf. figure n° 58)
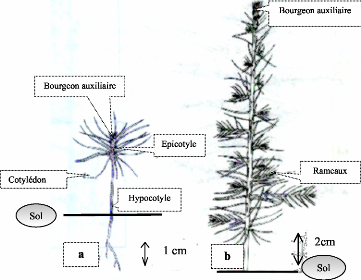
Hypocotyle
î 1 cm
Cotylédon
Epicotyle
....
. . Bourgeon auxiliaire
Rameaux
......
..........
2cm
a
Bourgeon auxiliaire
Fig. 58 : Plante d'une année issue d'une régénération naturelle (a) ou de pépinière (b)
Le traitement des résultats a montré que les différences obtenues sont significatives entre les différentes placettes pour une même exposition et pour les deux expositions.
En effet, l'étude de la station Nord, de la station Sud ensuite les deux en même temps a montré que :
- L'exposition Nord est la plus favorable à la régénération naturelle.
- L'altitude comprise entre 1700-1800 m donne les meilleurs résultats.
- Le travail profond du sol, influence positivement la germination et le développement particulièrement
- Le couvert joue un rôle différent selon l'exposition : il est favorable sinon indispensable sur l'exposition Sud ; sur l'exposition Nord s'il est néfaste pendant la période froide et humide, il devient propice durant la période estivale.
211
Soulignons que toutes les placettes se distinguent par une mortalité graduelle des semis, qui peut conduire dans certaines conditions à leur élimination totale.
Le suivi mensuel de ces plantules est représenté par la figure n°55 et le tableau n° 117 résume l'état des placettes après une année d'observation.
Tableau N°117:Evolution du nombre de semis en fonction de la station et du travail du sol.
|
Exposition Nord |
Exposition Sud |
Station |
||||||||
|
Avr |
Dec |
Taux de |
Avr |
Dec |
Taux de |
Avr |
Dec |
Taux de |
||
|
Altitude |
<1.700 |
507 |
181 |
35.70 |
111 |
11 |
9.90 |
703 |
192 |
27.31 |
|
1.700 -1.800 |
670 |
249 |
37.16 |
196 |
25 |
12.75 |
781 |
274 |
35.08 |
|
|
Couvert |
Sous couvert |
510 |
184 |
36.08 |
169 |
33 |
19.53 |
679 |
217 |
31.96 |
|
A découvert |
667 |
246 |
36.88 |
138 |
03 |
2.17 |
805 |
249 |
31.93 |
|
|
Travail |
Témoin |
213 |
072 |
33.80 |
60 |
02 |
3.33 |
273 |
074 |
27.10 |
|
Superficiel |
366 |
120 |
32.79 |
88 |
10 |
11.36 |
454 |
130 |
28.63 |
|
|
Profond |
598 |
238 |
39.80 |
159 |
24 |
15.09 |
757 |
262 |
34.61 |
|
|
Nombre total de semis |
1177 |
430 |
36.03 |
307 |
36 |
11.73 |
1484 |
466 |
30.95 |
|
La régénération est étroitement liée aux différents facteurs stationnels et, chacun peut avoir une importance plus ou moins grande ; pris un à un l'influence de ces facteurs est facilement décelables comme le montre la figure n° 54 ; mais, en réalité l'action combinée de l'ensemble de tous les facteurs qui avant ou lors de l'ensemencement permettent de dégager des conditions favorables à la germination, à l'installation et à la survie des semis.
La date d'apparition des semis n'est pas déterminée selon LEPOUTRE (1963) par la hauteur des pluies, mais étroitement dépendante de la valeur et la durée des températures maximales (M) . Cette température doit atteindre 9 à 10 °C pendant au moins 9 à 10 jours, pour des températures minimales (m) elle doit être de l'ordre -- 5 ° C.
Néanmoins, si durant la germination, les précipitations se font rare comme au mois de Avril (1981) ou Mars (1982), on assistera à un ralentissement de l'hydratation des graines, entraînant ainsi, une perte de leur vitalité qui se traduit par un arrêt de la germination.
EZZAHIRI et al (1994) ont mis en évidence la dépendance de la régénération naturelle du cèdre vis avis de certains paramètres liés à la station et au peuplement : paramètres climatiques (humidité, températures...), facteurs édaphiques (nature du substrat, profondeur et texture), et la structure de la végétation.
LEPOUTRE (1964) en étudiant la régénération naturelle, estime que l'interaction des facteurs climatiques peut provoquer une très grande mortalité des jeunes semis de cèdre: gelées précoces en hautes altitudes et/ou une sécheresse prononcée en basse altitude.
Au Belezma, les conditions du milieu sont souvent défavorables ; elles se traduisent par :
Une sécheresse estivale très prononcée. Irrégularités des précipitations.
Fréquences des vents desséchants. Sol compact (argileux).
Absence totale de traitements sylvicoles.
212
Aussi, l'analyse de l'évolution des semis dévoile que les mois qui suivent la germination et l'apparition des aiguilles (Mai à Octobre) sur l'exposition Sud et à découvert constituent la première phase critique qui est déterminante pour la survie des plantules de l'année. Pour les stations sous couvert, surtout celles de l'exposition Nord, cette phase qui relativement tardive débute à partir de Juin jusqu'à Août - Septembre. Quant à la période hivernale qui se rapporte aux semis âgés de 7 à 8 mois, qui forme la deuxième phase critique, commence à partir de Décembre et se termine au mois de février, elle est plus longue sur les hautes altitudes et plus rigoureuse sous couvert.
On peut dire que les semis du cèdre évoluent dans des conditions clémentes pendant 2 à 3 mois seulement sur une année ; les placettes bénéficiant d'une bonne régénération naturelles sont les mieux protégés par«l'effet du microclimat».
Dans notre station, les deux paramètres essentiels à la régénération sont l'altitude et de l'exposition. En effet l'apparition (avril) et le maintien des semis au-delà de la saison estivale est plus intéressante sur l'exposition Nord et à haute altitude.
Les conditions thermiques expliquent en grande partie la présence des plages de semis naturels ; mais, l'arrivée précoce des hautes températures estivales associées à la rareté de l'eau les fait disparaître ; cependant, ceux protégés par un couvert arrivent dans bien des cas à se maintenir durant la période estivale.
111,66
ri Altitude<1700m
El Altitude1700 -1800m
84,5
41,5
30,16
32,66
18,5 1,83 4,16
|
Avril |
Décembre |
Avril |
Décembre |
Fig . 59: Effets de l'exposition et de l'altitude sur la germination et le maintien des semis.
Les autres facteurs comme le couvert, la strate herbacée, les conditions du sol ... peuvent jouer un rôle non négligeable dès qu'on s'écarte des conditions favorables à la régénération naturelle.
Au Belezma, la régénération se comporte différemment selon le couvert et l'exposition (Cf. figure ci-dessous). Sur l'exposition Sud, les semis s'installent mieux sous abri, mais également autour des souches, des arbres morts, sur des micro - accidents du relief (replats, bord des pistes...), ceci confère de l'ombre au semis; sur celle du Nord ils ont tendance à se régénérer relativement mieux à découvert que sous couvert .
213
Dans les cédraies humides, la régénération s'installe également mieux dans les zones relativement protégées un abri comme les cépées de chêne vert est favorable à l'installation lente des semis de cèdre. (MARION, 1955 ; VLASSEV, 1973).
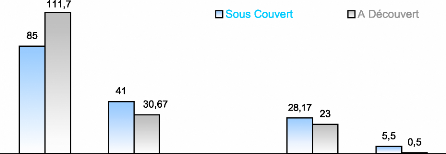
28,17 23
1
Sous Couvert
D A Découvert
85
5'5 0,5
41
111,7
30,67
Avril Décembre Avril Décembre
Fig . 60: Effets du couvert selon l'exposition sur la germination et le maintien des semis.
Cette installation est due au tempérament du cèdre de l'Atlas; il a tendance à s'implanter dans des conditions bien précises qui parfois semblent être opposées. C'est une essence d'ombre à basse altitude et de lumière à haute altitude (LEPOUTRE, 1964). Ce n'est pas une essence typique de la lumière (AUSSENAC, 1984).
De point de vue climatique le cèdre se renouvelle selon EZZAHIRI et al (1994) dans les micro - stations qui bénéficient d'apport de précipitations ce qui améliore le bilan hydrique et atténue la continentalité. Ces micro- accidents du relief, jouent également un rôle dans la protection des semis en diminuant la vitesse de ruissellement des eaux. Au Belezma , les précipitations printanières et automnales par leur agressivité causent de graves dommages aux semis de l'année.
TAN(1987 et 1992) note que les paramètres comme l'enneigement, la violence des averses, les températures, la rugosité sol, l'humidité de l'air et du sol, sont liées à la régénération du sapin ; il a également évoqué l'érosion des semis ainsi que les facteurs qui s'y opposent : replats, base du tronc, souches et surtout la couverture muscinale. Ainsi, les facteurs microclimatiques et pédologiques agissent sur les premiers stades de développement et d'implantation des semis du sapin (BECKER et al 1984 et 1985).
La dynamique de la régénération de cette espèce est extrêmement complexe, elle est en relation directe avec l'humidité du sol et de l'air ainsi que les températures ; dans ce cas on peut considérer que le couvert est un facteur important : il agit nom seulement sur l'humidité édaphique et l'humidité relative qui constituent un "facteur limitant" pour les cédraies naturelles, mais protége les jeunes semis contre l'ensoleillement lors de la saison estivale qui peut durer plus de 6 mois au cours de l'année (mai à octobre).
Elle est liée à tous les facteurs écologiques qui régissent la disponibilité de l'eau pour la plante : les discriminations entre les expositions du nombre de plantules sont dues surtout à la persistante de l'humidité
214
Sur exposition Sud, les placettes sous couvert sont mieux protégées que celles à découvert : les pertes en eau par évaporation sont relativement peu importantes, il y a plus de survie. Sur exposition Nord, les températures sont plus clémentes, l'humidité de l'air et du sol sont plus importantes, qui favorisent outre la production grainière, la germination, l'installation et le maintien des semis les semis en tendance à mieux s'installer, particulièrement sur les placettes à couvert clair, mais également autour des souches, des arbres morts. En effet, sur cette exposition le nombre de semis ayant dépassé la période estivale est de 430 ce qui correspond à un taux de mortalité de l'ordre de 64 %, celui de l'exposition Sud atteint 36 semis soit un taux de mortalité de l'ordre de 90 %.
Ainsi, AUSSENAC (1984), a indiqué que c'est surtout la sécheresse estivale, qui constitue le facteur limitant. EMBERGER (in QUEZEL, 1957) a noté qu'au Maroc, le cèdre de l'Atlas manque sur presque tout le Haut Atlas, sans doute à cause du nombre trop élevé de mois secs.
Ainsi les jeunes plantules, ont peu de chance de survivre à la sécheresse : le développement du système racinaire n'a pas eut le temps nécessaire d'atteindre les profondeurs, leurs permettant d'échapper à la dessiccation des horizons superficiels. La précocité et l'importance de la sécheresse conditionnent la survie des semis et dépendent des facteurs de la station. En outre, malgré la capacité du cèdre à résister à la sécheresse, les semis d'une saison de végétation sont selon AUSSENAC (1984), très sensibles à la sécheresse et beaucoup plus sensibles que ceux de deux années. Nos résultats montrent que le taux de réussite sur la station Nord est de 36%, alors que pour celle du Sud, on enregistre que 11.73%.
Les semis sont très sensibles à l'ensoleillement direct; DUCREY (1993) rapporte que le cèdre de l'Atlas, ne contrôle pas très efficacement ses pertes en eau et son adaptation réside dans sa bonne croissance racinaire et sa capacité à prospérer le sol en profondeur, et que les vagues de croissance aérienne successives alternent avec les vagues de croissance racinaire. Le milieu a une action quantitative et non qualitative sur le développement du système racinaire (ATGER, 1991). LEPOUTRE (1963 -- 1964) signale que le système racinaire peut pousser toute l'année par vagues successives de 2 à 4 semaines et que si les conditions sont favorables, la croissance peut atteindre 40 cm en 4 mois.
En prenant en considération l'action simultanée de deux facteurs stationnels, à savoir le découvert et les basses altitudes l'installation des semis particulièrement sur les expositions Sud devient impossible. Si ces deux facteurs sont dans un premier temps favorables à la germination (hausse précoce de la température), ils deviennent par la suite extrêmement défavorables au maintien des semis, on assistera alors à la disparition progressive mais totale des plantules au cours de la saison estivale. Sous couvert, la germination est inhibée par les basses températures au printemps, surtout à haute altitude ; par la suite le maintien des semis est favorisé par le couvert.
Aussi, les cédraies riches en litière rendent selon LEPOUTRE (1961 et 1963) les conditions du milieu édaphique défavorables à la germination et à la survie des semis. Elles sont défavorables à partir de 1850 -- 1900 m (plus l'altitude diminue et plus le couvert devient favorable), l'altitude de 1550 m représente les conditions optimum
Cependant au Belezma, les stations riches en litière sont peux importantes ; elles sont localisées sur les expositions Nord (peuplement fermé), la régénération est défavorisée dans un premier temps par la présence de la matière organique mal décomposée, qui agit négativement sur la germination, elle ne l'est plus quand les hautes chaleurs arrivent, la litière devient plutôt favorable grâce à sa capacité de rétention de l'humidité. Ainsi, DRAPIER (1985) a mis en évidence le type de station (végétation et humus) et l'aptitude à la régénération.
215
La fermeture des stomates chez le cèdre de l'Atlas est tardive (GUEUL, 1983) et qu'il a encore selon AUSSENAC et al (1983) une photosynthèse au cours d'une sécheresse, bien après que la croissance en hauteur ait été arrêtée et que c'est surtout la sécheresse atmosphérique qui influence la croissance des semis. L'humidité de l'air joue un rôle fondamental dans les premiers stades du développement du sapin (TAN et al 1992). BARITAU (1993), CRIEU et al (in DUCREY, 1993) notent qu'on assiste à une diminution très rapide de la croissance dès qu'il y a sécheresse atmosphérique ; or il faut signaler que le Belezma se caractérise par une faible humidité relative.
La croissance racinaire et la photosynthèse du cèdre de l'Atlas sont interdépendantes, elles évoluent selon l'état hydrique du plant et donc celui du sol ; la croissance des racines varie en fonction des conditions nutritionnelles de la station ; les champignons ectomycorhiziens jouent un rôle très important dans la résistance des semis à la sécheresse et dans la stimulation dans leur croissance radiculaire ; la vitesse de croissance initiale de la racine doit être considérée comme le facteur fondamental pour la survie des semis. Cependant le cèdre à l'état jeune est difficile à se mycorhizer même après des inoculations artificielles. (LEPOUTRE, 1963 et 1964 ; ABOUROUH, 1983 et 1993 ; LAHMADI et al in LAHMADI et al, 1993).
Les travaux réalisés par les chercheurs, ont montré que le sol par ses conditions physiques et chimiques accentuera ou amortira les effets du climat. Si ce dernier est favorable les conditions édaphiques jouent un rôle mineur, mais dès qu'on s'écarte de l'optimum climatique, elles deviennent défavorables (litière mal décomposée, argile).
LEPOUTRE (1964 et 1966) insiste sur le fait qu'il n'y a pas de sol permettant ou interdisant la régénération du cèdre, mais il y a simplement des types de sol qui la facilitent plus ou moins. Les sols sableux facilitent l'installation des semis mais leur dessiccation (rapide) rende le maintien des semis extrêmement difficile alors que les sols profonds sont favorables. Sur sol argileux ou marneux, le développement du cèdre de l'Atlas est très difficile sinon impossible (problèmes de réserve utile).
La fertilité de la station est conditionnée par les fissures de la roche mère RIPERT et al (1993). Le cèdre de l'Atlas, exploite bien les fissures de la roche mère, il appréciera également les sols terreux et profonds (alluvions) à conditions que ceux-ci ne soient pas compacts et asphyxiants (facilités de pénétration des racines). Cette compacité qui pourrait être éliminée par un travail du sol, permettra d'améliorer les propriétés physiques et donc d'assurer un meilleur taux de survie des jeunes semis.
BOUDY (1952), LEPOUTRE (1961et1963), TOTH (1978) et DERRIDJ (1990) signalent que les sols meubles et profonds permettent une bonne régénération du cèdre de 1 'Atlas : rétention de l'eau et pénétration facile des racines. GHEHL et al (1989) notent que les caractéristiques photosynthétiques (surface foliaire x taux d'assimilation de CO2) des plants de Cedrus atlantica M. élevés en conteneurs sont fortement modulées par le type de substrat.
DUCHAUFFOUR (1988) a soulevé également les difficultés de la régénération du sapin (Ables alba MILL) dû aux problèmes de l'enracinement.
Des travaux réalisés par EZZAHIRI et BELGHAZI (2000) ont montré que la régénération du cèdre est tributaire de certains éléments du sol comme l'argile, le magnésium et le sodium. Les meilleures régénérations sont obtenues sur des sols caractérisés par des taux de carbone de 1%, en magnésium de 16 meq /100g, en sodium de 0.70meq /100g et une teneur de l'ordre de 22% d'argile. PUTOD (1979) signale l'influence particulièrement néfaste de la texture argileuse. Or, si les sols de Belezma sont bien pourvus en éléments minéraux, l'argile est véritable handicap pour la régénération, il est de 60 % (Cf. Tableau n°90).
216
La concurrence herbacée dans les cédraies naturelles a été signalée par BOUDY (1950) LEPOUTRE (1963) et NEDJAHI (1988), elle se traduit par la diminution de l'humidité édaphique mise à disposition des plantules et sur la réception des graines par le sol. Selon FROCHOT et al (1986), la strate herbacée représente un sérieux obstacle aux régénérations naturelles : gène physique, compétition pour la lumière, l'eau et les éléments nutritifs ; émission de substances toxiques pour les jeunes semis. PIUSSI (1988) a observé l'influence nocive du tapis végétal à l'égard des semis : pour les péssières, les plants issus de la régénération naturelle peuvent survivre si les arbres et les arbustes sont éliminés.
Ainsi quand les conditions climatiques sont relativement favorables (sous couvert sur les expositions Sud -- à découvert sur celle du Nord -- altitude moyenne) le travail du sol qui améliore la structure des horizons, facilite la décomposition de la matière organique, l'infiltration des eaux réduit la concurrence herbacée et permet également de créer un lit de semence pour les graines.
DEMOLIS, OTT et TROSSET rapportèrent lors de la réunion de travail en 1988 sur la régénération des forêts d'altitude que si le sol est dégagé et le premier horizon remue (le décapage des couches organiques est à déconseiller) de bonnes conditions sont crées pour la régénération des forêts d'altitude des zones tempérées. En outre, PELLISSIER et al (1992) ont mis en évidence le blocage de la germination due à l'humus ; l'influence de ce dernier persiste les années suivant la levée des semis et se concrétise par une disparition considérable des jeunes plants de sapin.
TOTH (1978a et 1982), note que si les conditions climatiques, surtout la pluviométrie, compensent les conditions édaphiques, ces dernières peuvent être améliorées par le travail mécanique du sol. Ce dernier a permis selon les données de TOTH (1978) et DERRIDJ (1990), d'avoir les meilleurs résultats aussi bien pour la germination que le maintien des semis du cèdre de l'Atlas. Il semble donc intéressant de préparer le sol pour favoriser la germination lors des années de grainée.
Nos résultats illustre l'intérêt du travail du sol nom seulement sur la germination mais sur le maintien des semis.
149,5
|
|
|
||||||
|
||||||||
|
||||||||
|
||||||||
|
||||||||
q Travail.Superficiel.Sol ri Travail.Profond.Sol

39,75
2
152
1
0,5 1,66 6
Avril Décembre
Exposition Nord Exposition Sud
Fig.61 : Effets du couvert selon l'exposition sur la germination et le maintien des semis.
217
Le travail du sol est très apprécié par les semis du cèdre de l'Atlas : le système racinaire profite de la moindre humidité, pour s'enfoncer dans les couches profondes ; l'élimination de végétation concurrentielle facilitera la pénétration des racines.
Pour obtenir les meilleurs résultats, il faut que le travail du sol atteigne 40 -- 60 cm et même parfois d'avantage. En effet, nos résultats ont mis en évidence l'importance du travail sol particulièrement au-delà de 40 cm où, non seulement le nombre de graines germées est important (2.11 semis par m2) mais également leur maintien. Ce travail permet de diminuer le taux d'argile, ce dernier constitue un véritable obstacle à la régénération (germination, développement et maintien)
Des études dendrochronologiques du cèdre menées par TILL (1985), CHBOUKI, MEKO et STOCKTOM (in LAMHAMEDI et al 1993) ont révélé que le climat ou évolue les cédraies naturelles, est caractérisé par une alternance d'une période favorable humide et une période défavorable sèche ; la dynamique de la cédraie, notamment en matière de régénération dans le temps ne pourrait être due qu'au facteur limitant qui est l'eau. De ce fait, il est actuellement admis avec une grande certitude que la régénération du cèdre de l'Atlas est intimement liée à de longues périodes de sécheresse.
218
Climat : Méso et microclimat.
- Précipitations - Humidité
- Températures

Floraison
Dissémination
Année de fructification.
Position des cônes
dans l'arbre
Germination
Installations des plantules
A
Travail du sol
Qualité de la station
- Facteurs orographiques
* Altitude
* Exposition
*
pente
-Facteurs édaphiques
--
1REGENERATION
- Composition
- Peuplement
- Structure
- Age
A l'issue de ce travail qui a pour objet, l'étude de la régénération du cèdre de l'Atlas (Cedrus Atlantica M), il est important de présenter brièvement les principaux résultats obtenus.
Cette régénération comporte deux cycles :
- Le cycle de production des graines : Floraison - Fructification
- Le cycle d'ensemencement : Dissémination - Germination - Installation des plantules.
Le schéma ci-dessous servant à montrer les phases essentielles de ces deux cycles, qui sont étroitement liés entre-eux et dépendent :
- Des facteurs climatiques, dont les plus déterminants sont les précipitations, l'humidité atmosphérique et les températures ; elles agissent simultanément et conditionnent le fonctionnement de l'écosystème "cédraie " par leurs actions directes et indirectes.
- De la station : exposition, altitude, couvert ...
- De l'état du peuplement : composition, âge, structure...
|
Fig.62 : Etude des principaux paramètres affectant le processus de la régénération du cèdre de l'Atlas (Cedrus atlantica M) |
220
I-Le cycle de production.
1- Le cycle de reproduction.
L'étude phénologique de l'appareil de reproduction nous a permis de préciser les dates d'apparitions des inflorescences femelles (Fin Août) et mâles (Fin juin) ainsi que les principales phénophases. La maturité de la fleur mâle se traduit par la production des grains de pollen et dure 3 à 4 mois ; celle de l'inflorescence femelle dure un mois au maximum et se caractérise par l'ouverture des bractées et devient le récepteur du pollen. Au printemps de l'année suivante, on assiste à une augmentation lente du poids et des dimensions ; après fécondation, qui aura lieu vers la fin du moi de Mai, l'inflorescence femelle devient cônelet, sa croissance devient exponentielle et atteint sa maturité morphologique vers le mois d'Août --mi-Septembre ; un mois après, sa couleur qui était d'un vert clair devient marron, le cône atteint sa maturité physiologique. A partir du mois d'Août jusqu'à fin Décembre, parfois à la fin de la première semaine de janvier, on assiste à la désarticulation des cônes et à la dissémination des graines, qui peut aller.
L'évolution des principaux caractères biométriques pendant un cycle complet se résume ainsi : si le poids, la longueur et le diamètre augmentent peu durant la première année (N), l'année d'après (N+1) se caractérise par de grandes transformations morphologiques dues à la pollinisation, fécondation..., les dimensions s'accroissent considérablement alors que, la 3ème année (N+2) se caractérise surtout par des transformations physiologiques qui se traduisent par le changement de couleur et l'achèvement d'élaboration des graines .
L'échelle proposée, nous permettant de suivre chronologiquement les principales phénophases durant un cycle complet de reproduction :
Floraison - Fructification - Désarticulation et dissémination. 2-L'estimation de la fructification
Pour cela, nous avons utilisé la méthode de TOTH, qui a été réalisée pour les cédraies humides (précipitations sont de l'ordre de 700 à 1.100 mm), dans un premier temps, puis nous l'avons adaptée à la cédraie de Belezma (précipitations sont comprises entre 500 à 700 mm) en déterminant le nombre des inflorescences femelles et des cônes pour l'estimation de la fructification et donc les possibilités de récoltes. Ceci nous a permis de montrer que pour estimer la production grainière et évaluer les possibilités de récolte le facteur climat est primordial, car de lui dépende la qualité et la quantité de graines.
3- La variabilité de la production des organes de reproduction.
Cette variabilité est en relation avec les facteurs intrinsèques et extrinsèques. Elle se traduit par l'alternance de la fructification. Cette dernière est due généralement à un épuisement important des réserves dans l'arbre (Facteurs intrinsèques).
Le nombre des inflorescences femelles et des cônes est plus important sur les branches hautes quelle que soit l'exposition. (Facteurs extrinsèques)
Quant au facteur exposition, si la floraison est meilleure sur l'exposition Sud, à cause de l'ensoleillement, la fructification est meilleure sur les expositions Nord. Les pertes des inflorescences en enregistrées sont plus importantes sur l'exposition Sud.
221
4- L'alternance de la fructification.
Les résultats ont montré que le cèdre de l'Atlas ne peut pas assurer une forte production de graines chaque année. Généralement une bonne année de fructification est suivie de deux ou de trois années moyennes à mauvaises.
5 - Les cônes et des graines.
La désarticulation artificielle des cônes.
Les deux facteurs déterminants sont l'humidité et la chaleur: immersion des cônes durant une période de 24 à 36 h, suivie d'un séchage à l'air libre pendant 6 heures au moins ; ensuite une autre imbibition de 2h au moins. Ceci permettra de récupérer la quasi-totalité des graines; ces dernières sont composées de graines fertiles et vaines (aplaties, attaquées et en forme de bec)
Répartition des graines dans le cône.
La décortication manuelle des cônes a montré que les graines fertiles, aplaties et attaquées sont surtout localisées au milieu du cône (partie fertile), celles en forme de bec se trouvent aussi bien dans la partie haute que la partie basse.
La production des différents types de graines.
Elle est fonction surtout de l'année de la fructification (l'alternance), de la station et de la position des cônes dans l'arbre.
En effet, nous avons constaté l'existence de relations hautement significatives entre la qualité des graines et l'année de fructification pour une même station et également entre les stations : une mauvaise année de production de cônes correspond à une qualité médiocre de graines, particulièrement sur l'exposition sud et inversement à une forte production de cônes correspond une bonne qualité de graines.
La production qualitative et quantitative des graines de l'exposition Nord (Station favorable) est meilleure que celle de l'exposition Sud (Station défavorable).
Les cônes initiés sur les branches hautes de l'arbre, c'est à dire les plus éclairées sont les plus fertiles.
Biométrie des cônes et des graines pendant l'année de forte production.
Les dimensions des cônes jouent également un rôle très important dans la régénération naturelle. En effet, on a constaté que les cônes les plus dimensionnées produisent non seulement le plus grand nombre de graines fertiles mais également les plus lourdes, c'est à dire celles qui présentent les meilleurs taux de germination et les temps moyens de germination les plus bas et donc capable de donner des semis les plus développés.
Pour une même exposition le poids, la longueur et le diamètre des cônes des branches hautes sont supérieurs à ceux des branches basses, en outre l'exposition Nord produit les cônes les plus dimensionnés, c'est à dire les plus fertiles, que celle du Sud.
222
H - La régénération 1- La germination -Le germoir.
La réalisation par nos soins, d'un germoir à alimentation d'eau continue par capillarité grâce à un papier buvard nous a permis de mener des tests de germination depuis l'apparition de la radicule jusqu'au stade cotylédonaire, soit une duré de 21 jours.
-Mise en germination.
En conditions artificielles, il est facile de levée la dormance grâce à des prétraitements par un froid humide (- 1 à + 4 °C) durant une période d'un mois ou bien par un traitement à l'eau oxygénée à 30 % pendant une durée de 45 mn.
Les graines fertiles extraites à partir de cônes mûrs, présentent une dormance plus ou moins profonde, celle-ci est susceptible d'être levée si des traitements au froid humide ou à l'eau oxygénée lui sont appliqués ; ces graines sont incapables de germer si elles ne s'imbibent pas d'eau en absorbant au moins 50 % par rapport à leur propre poids; cette hydratation passe par 4 phases: rapide- lente -rapide- lente. Après l'imbibition qui dure 72 heures (temps de latence) la germination est entamée.
-Influence de quelques facteurs sur la germination.
L'exposition, la position des cônes dans les branches et le poids des graines jouent un rôle très important dans la variabilité du pouvoir germinatif et donc dans la régénération. Les graines initiées sur les expositions favorables et les branches hautes se caractérisent par des facultés germinatives les plus élevées et donc la possibilité de développer des systèmes racinaires puissants, susceptibles de résister à l'aridité climatique estivale.
Les différents tests ont montré que les parties de la graine (la membrane, l'endosperme et l'aile) qu'elles n'ont pratiquement aucun effet sur le pouvoir germinatif.
Par contre, les paramètres comme la date de récolte et le mode de traitement déterminent dans une large mesure le pouvoir germinatif. Ainsi les graines qui seront récoltes à la fin de l'automne (Novembre) ou bien au début de l'hiver (Décembre) présentent sans prétraitement des taux de germination comprises entre 50 -70%, quand le prétraitement leur est appliqué, ces taux seront supérieurs ou égaux à 90 %;
La période de récolte des graines est extrêmement intéressante pour les pépinières productrices de plants.
Ces deux paramètres sont très importants dans la variabilité du pouvoir germinatif ; ce sont les températures basses qui déterminent les possibilités de germination (levée de dormance) ; il faut donc procéder à des récoltes tardives (fin de l'automne) afm d'éviter tout prétraitement « artificielle », car les graines ont déjà subi l'influence du froid humide avant leur dissémination.
223
2-Influence des prétraitements sur le développement des plantules :
Cette étude a mis en évidence une nette relation entre le prétraitement des graines et l'aptitude à la régénération.
En effet, le repiquage des graines prégermées après traitement au froid humide et à l'eau oxygénée dans des pots, sur substrat récolté sous cédraie, a donné les plus forts taux de reprise; en outre, les jeunes plantules obtenues présentent par rapport au témoin, les meilleures caractéristiques morphologiques et ne présentent aucun déficit de point de vue de leur composition minérale. Aussi, les graines prégermées dotées de radicule de l'ordre de 1 à 2 mm continuent à se développer normalement une fois repiquées, en donnant un enracinement puissant et ramifié ; cependant, si la longueur des radicules de ces graines est assez longue, plus de 5mm, la reprise est nulle.
3- Installation des plantules.
Cette étude, a pour objet de traiter l'influence du degré de travail du sol sur la régénération en prenant en considération les facteurs de station à savoir l'exposition, l'altitude et le couvert. Elle a été réalisée durant l'année de forte fructification ; globalement la régénération obtenue est satisfaisante sur l'exposition Nord, malgré les conditions climatiques souvent difficiles. Après la germination et l'apparition des cotylédons, on a assisté à une diminution progressive du nombre de plantules jusqu'à leur disparition totale dans certaines conditions, comme le découvert sur l'exposition Sud.
Le renouvellement de la cédraie est déterminé par l'importance de la sécheresse qui est en relation avec l'altitude, l'exposition et le couvert.
La comparaison de l'installation des semis a montré que:
- Si les plantules arrivent à s'installer et à se développer sur l'exposition Nord, elles éprouvent de grosses difficultés à se maintenir sur celle du Sud.
- Sur l'exposition Sud, l'apparition des plantules est précoce et atteint son maximum au mois d'Avril; il n'y a pas de germination tardive. Sur celle du Nord, si nous avons enregistré des germinations tardives (Mai) sur des altitudes comprises entre 1.700-1.800 m leur nombre reste faible.
- Sur les hautes altitudes, l'installation des semis est relativement facile quelle que soit l'exposition; ces plantules semblent apprécier le découvert sur l'exposition Nord à cause des basses températures ; sur celle du Sud l'abri devient indispensable à leur maintien: l'humidité persistante associée à des températures clémentes favorisent l'installation.
-Le travail du sol est extrêmement favorable à la germination et au maintien des semis de cèdre de l'Atlas : après la dissémination, la graine trouve l'humidité nécessaire à sa germination ; au stade cotylédonaire, le système racinaire bénéficie non seulement d'une humidité favorable pour son accroissement mais n'éprouve aucune difficulté pour s'enfoncer dans les horizons meubles et profonds du sol.
L'influence simultanée de plusieurs facteurs jouent un rôle déterminant dans la germination, le développent et le maintien des semis au-delà de la période estivale ; période qui se caractérise par la rareté des précipitations et les hautes températures. Ainsi, sur l'exposition Nord, la combinaison du travail profond du sol et le découvert est favorable à la germination et à la survie des plantules : lumière, humidité suffisante et une protection contre l'ensoleillement direct. Aussi, dans les stations favorables, l'action des hautes altitudes (supérieures à 1.700m) et du découvert facilite la régénération ; celles défavorables et sous couvert, le maintien des semis est globalement intéressant uniquement pour les placettes ayant subi un travail profond du sol.
224
RésuméLa régénération du cèdre de l'Atlas (Cedrus atlantica M) a été étudiée dans deux stations : Station à exposition Nord et Station à exposition Sud. Cette étude comprend essentiellement deux parties : le cycle de production et le cycle d'ensemencement. La phénologie de l'appareil de reproduction a été abordée pendant un cycle complet par des mesures biométriques au laboratoire durant les principales phénophases déterminées sur terrain. L'estimation des inflorescences femelles, des cônes d'une année et deux années nous ont permis de prévenir l'importance de la fructification une à deux années à l'avance et de relever que la variabilité de la production de ces organes est en relation avec la station, les branches et surtout l'année de la fructification. Ces paramètres sont déterminants pour le poids et les dimensions des cônes et donc sur la production qualitative et quantitative des graines : plus la production des cônes est forte et plus celle des graines est bonne. L'étude de l'amplitude de la fructification a montré que le cèdre de l'Atlas ne peut pas assurer chaque année une forte fructification. La régénération a été traitée au laboratoire et sur terrain. Au laboratoire nous avons étudié le pouvoir germinatif de graine en relation avec divers paramètres. Le poids des graines a une influence sur leur pouvoir germinatif, il est influencé par l'année de fructification, par la station, la position du cône dans l'arbre et la grosseur de cône. La date de récolte et le mode de conservation des graines déterminent la capacité de germination. La dormance des graines peut être levée après traitement par l'eau oxygénée ou au froid humide ; les plantules issues de ces graines présentent un bon développement, ce qui leur permet d'avoir de meilleures chances d'échapper à la sécheresse. Sur terrain, le travail profond du sol, l'exposition Nord et le couvert donnent de meilleures chances aux semis de survivre, pendant la période estivale. Mots-clés : Biométrie, Cedrus atlantica, cône, cônelet, fructification, inflorescences femelles, phénologie, phénophase, plantule, régénération. |
SummumThe regeneration of the cedar of the Atlas (Cedrus atlantica M) was studied in two stations: A Station with a North exhibition and a Station with a South exhibition. This study understands essentiality two parts: the cycle of production and the cycle of sowing. The reproduction phenology was approached during a complete cycle by biometric measures in the laboratory during the main clauses phénophases determined on ground. The estimation of female inflorescences, cones of one year and two years allowed us to fore cast the importance of the fruiting one in two years in advance. The variability of the production of these organs is in connection with the station, the branches and especially the year of the fruiting. These parameters are determining for the weight and the dimensions of cones and so on the qualitative and quantitative production of seeds: the more the production of cones is high and the more that of the seeds is good. The study of the amplitude of the fruiting showed that the cedar of the Atlas can not provide every year a good fruiting. The regeneration was studied in the laboratory out doors. In the laboratory we studied the germinal power of seed in connection with different parameters. The weight of seeds has an influence on their germinal power; it is influenced by the year of fruiting by the station, the position of the cone in the tree and the cone thickness. The date of harvest and the mode of conservation of seeds determine the capacity of germination. The dormance of seeds can be raised after treatment by the hydrogen peroxide or in the wet cold, plantules stemming from this show presents a good development, this allows them to have the best chances to aroid aridity. Out doors, the deep work of the soil, the North exhibition and the covert give better chances to the sowing to survive during summery periods Keywords: Biometry, Cedrus atlantica, cone, conelet, fruiting, seed, female inflorescences, phenology, phenophase, plantule, regeneration. |
BARITEAU, M. (1993) :
L'amélioration génétique des cèdres en France.
Silva Mediterranea -- Actes du Séminaire International sur le cèdre de l'Atlas. Ifrane (Maroc) ; 7 --11 juin 1993 : 464 -- 472.
BARITEAU, M. et FERRANDES, P ; (1990) :
L'amélioration des cèdres en France. Symposium International sur le cèdre. Antalya -- Turquie : 366 -- 379.
BARITEAU, M. ALPTEKIN, C.U. et BRAHIC, P. (1994) :
La faculté germinative des organes de cèdre (Cedrus atlantica M.) : influence des traitements densimétriques à l'alcool, du desailage et de la stratification. Ann. Rech. For. Maroc, Vol. 2 : 500 -- 509.
BECKER, M. et DRAPIER, G. (1984) :
Rôles de l'allélopathie dans les difficultés de régénération du Sapin (Abies_alba Mill). I -- Propriétés phytotoxiques des hydrosolubles d'aiguilles de sapin.
Acta OEecologia. OEecol. Plant., 5 (13), 4 : 347 -- 356.
BECKER, M. (1985) :
Les difficultés de régénération naturelle du sapin (Ables alba Mill) dans les Vosges. Etude écologique. R.F.F. XXXVII -- 1-- 45 -- 55.
BECKER, M. (1985) :
Démarche méthodologique préconisée pour la typologie des stations forestières. Colloques phytosociologiques XIV -- Phytosociologie et foresterie.
I.N.R.A -- C.N.R.F. Laboratoire de phyto -- écologie forestière. Champenoux 54280 Seichamps : 299 -- 311.
BECKER, M. et DRAPIER, G. (1985) :
Rôles de l'allélopathie dans les difficultés de régénération du sapin (Ables alba Mill).
II -- Etudes des lessivats naturelles, de feuillages, de litières et d'humus. Acta OEcologia OEcol. Plant., 6 (20), 1 : 31 -- 40.
BENABID, A. (1993) :
Biogéographie phytosociologie et phytodynamique des cédraies de l'Atlas Cedrus atlantica (Manetti).
Silva Mediterranea Actes du Séminaire International sur le cèdre de l'Atlas Ifrane (Maroc) ; 7 --11 juin 1993 : 62 -- 69.
BENTOUATI, A. (1993) :
Première approche d'étude de la croissance et de la production du cèdre de l'Atlas dans le massif de Belezma.
Thè. Mag. Inst. d'Agro. de Batna : 63 p.
BIDAUT, M. (1971) :
Variation et spéciation chez les végétaux supérieurs.
Notions fondamentales et systématique moderne. Doin Editeurs, Paris : 147 p.
B.N.E.F . (1986) :
Etude d'aménagement du Parc National de Belezma. Phase II : Inventaire et analyse économique. Bureau National des Etudes Forestières ;
Blida : 1-- 29.
BONNET- MASSIMBERT (1984) :
Biologie florale et cycle de reproduction de quelques arbres. Douglas, Pin Sylvestre, Chêne. I.N.R.A. Paris: 219-242
BOUDY, P. (1950) :
Economie forestière Nord -- Africaine.
Monographie et traitement des essences forestières. Ed. Larose, T 2 : 529 -- 619.
227
BOUDY, P. (1952) :
Considération sur la forêt algérienne et sur la forêt tunisienne.
Comptes rendus hebdomadaires des séances de l'Académie d'Agriculture de
France. T 39, séances des 8 et 15 Octobre 1952 : 529 -- 543.
BYOUNG --GUIENG, Y. (1976) :
Croissance du cèdre de l'Atlas (Cedrus atlantica Manetti) en relation avec quelques variables du milieu en Languedoc -- Roussillon (France).
Thè. Doc. Ing. Université des Sciences Techniques du Languedoc : 1 -- 10.
CANTEGREL,R.(1988):
Note sur le dépérissement et le renouvellement des forets pyrénéennes de pin à crochets (Pinus uncinata ,RAMOND ) .
Régénération des forets d'altitude.
Université de Savoie.Office National des forêts. 35-42
CARBAYE, J. ; LEROY, Ph. ; LE TACON, F. et LEVY, G. (1970) :
Réflexions sur une méthode d'études des relations entre facteurs écologiques et caractéristiques des peuplements.
Ann. Sci. For., 27 (3) : 303 -- 321.
CARLIER,G. (1988):
Observations sur les conditions de germination et de la longévité des graines d'épicéa ( Picéa abiès.(L.) KARSTEN. Régénération des forets d'altitude. Université de Savoie.Office National des forêts. 65-70
CHAUSSAT, F. et LEDAUNFF, Y. (1975) :
La germination des semences
Ed. Gauthier -- Villard, Paris : 11-- 36.
CHRISTOPHE, C. (1980) :
Variabilité intraspécifique de l'épicéa : stabilité et homogénéité des provenances et régions de provenances allemandes.
Ann. Sci. Forest. Vol. 37.2: 109 -- 133.
CLAUZEL, L. (1976) :
La plantation à racines nues du cèdre de l'Atlas. R.F.F. XXVIII -- 3 : 202 -- 204.
COME, D ; (1970) :
Les obstacles à la germination des graines. Ed. Masson et Cie., Paris : 162 p.
COURANJOU, T. (1982) :
Les résultats de quinze années de recherches sur les aspects génétiques de
l'alternance, posent de nombreux problèmes sur le physiologie.
Hème colloque sur les recherches forestières. Bordeaux : 107 -- 123.
COURANJOU, T. (1983) :
Facteurs variétaux de l'alternance des arbres fruitiers.
Résultats de quinze années de recherches chez le prunier domestique. Fruits Oct. 1983. Vol. 38. 10: 706 -728.
DAGNELIE, P. (1975) :
Théorie et méthodes statistiques, 2.
Ed. agronomiques de Gembloux : 463 p.
DEMOLIS,Ch . (1988):
La régénération de l'épicéa dans le Jura.
Régénération des forets d'altitude.
Université de Savoie.Office National des forêts. 61-64
228
DERRIDJ, A. (1990) :
Etude des populations de Cedrus atlantica M. en Algérie. Thè. Doc. Uni. Paul Sebastier, Toulouse : 288 p.
DRAPIER, J. (1983) :
Les difficultés de régénération des sapinières vosgiennes : importance de l'humus et rôle de l'allélopathie.
Thè. Doc. Uni. NANCY I -- INRA. Champenoux : 109 p.
DRAPIER, J. (1985) :
Les difficultés de régénération naturelle du sapin (Ables alba Mill) dans les Vosges. Etude écologique. RFF. XXXVII --1 : 45 -- 55.
DUCHAUFOUR, Ph.(1988):
Les péssières d'altitude :Physiologie de la nutrition et problèmes de régénération. Régénération des forets d'altitude.
Université de Savoie.Office National des forêts. 43-47.
DUCREY, M. (1981) :
Action des basses températures hivernales sur la photosynthèse du cèdre et du douglas. Ann. Sei. Forest., 38 (3) : 317 -- 329.
DUCREY, M. (1993) :
Adaptation du cèdre de l'Atlas (Cedrus atlantica Manetti) au climat méditerranéen. Aspect écophysiologique de sa réaction à la sécheresse.
Silva Mediterranea Actes du Séminaire International sur le cèdre de l'Atlas. Ifran (Maroc) ; 7 --11 juin 1993: 140 --150.
EMBERGER, L. (1971) :
Travaux de botanique et d'écologie. MASSON et Cie., Editeurs -- Paris : 520 p.
EZZAHIRI, M. ; BELGHAZI, B. et BEHMAD, M ; (1994) :
Bilan régénération naturelle de la cédraie dans les parcelles clôturées du Moyen Atlas, Maroc.
Ann. Rech. For. Maroc, 27 (Spécial), Vol. 1, I. XXXVI : 259 --267.
EZZAHIRI, M. ; BELGHAZI, B. (2000):
Synthèse de quelques résultats sur la régénération naturelle du cèdre de l'Atlas au Moyen Atlas (Maroc).
Science et changements planétaires / Sécheresse.vol 11.n°2 :79--89
F.A.O (1992) :
Guide de manipulation des semences forestières.
Organisation des Nations Unies pour l'Alimentation et l'Agriculture. Etude F.A.O. Forêt. Rome : 27 -- 55.
FAUREL, L. et LAFITTE, R. (1949) :
Facteurs de répartition des cédraies dans les massifs de l'Aurès et de Belezma. Extrait du Bull Soc. Hist. Nat. de l'Afrique du Nord. Tome 40 : 178 -- 186.
FERRANDES,H. (1986):
Amélioration génétique des arbres forestiers R.F.F. N°Spécial .139-142
FROCHOT, H. ; PICARD, J.F. et DREFEUS, Ph. (1986) :
La végétation herbacée obstacle aux plantations. R.F.F. XXXVIII. N° 3.
GAUSSEN, H. (1964) :
Les gymnospermes actuelles et fossiles.
Trav. Lab. For. Toulouse, T 2 (1), Fasc. VII : 273 -- 480.
229
GAUSSEN, H. (1967) :
Les gymnospermes actuelles et fossiles.
Trav. Lab. Toulouse, vol.I, Fasc. IV, Chap. XI :294-301
G.E.M.A.R.E.F. (1983) :
Les cèdres
Groupe d'Aix - En -- Provence. Division I.F.M : 1-- 7.
GENSAC, P. (1988) :
Régénération des forêts d'altitude.
Réunion de travail organisée à
Chambéry du 20/ 22 septembre 1988.
Uni. De Savoie -- Office National
des Forêts (Savoie) France : 168 p
GEORGE,J . (1980):
La récolte des graines de cèdre.
Rev. For . France. XXXII. 6. 544-446
GODET, J.P. (1974) :
Les comparaisons des moyennes et des variances. Application à l'agronomie. Bureau d'études Statistiques de l'I.T.C.F. Paris : 55 p.
GODET, M. et PHILIPPEAU,G .( 1992):
Comment interpréter les résultats d'un analyse de la variance. Collection S.T.A.T-I.T.C.F. Paris : 50p
GOUNOT, M. (1969) :
Méthodes d'étude quantitative de la végétation.
MASSON et Cie., Editeurs 120, Boulevard saint -- Germain. Paris : 314 p.
GUEHL, J.M. ; FALCONNET, G. et CRUEZ, J. (1989) :
Caractéristiques physiologiques et survie après plantation de plants de Cedrus atlantica élevés en conteneurs sur différents types de substrats de culture.
Ann. Sci. For. 46: 1-- 14.
HALIMI, A. (1980) :
L'Atlas Blideen, Climat et étage végétaux. Ed. O.P.U. Alger: 523 p.
HARROUCHE, A. (1991):
Etude comparative de quelques provenances algériennes de Cedrus atlantica Manetti. I.N.R.F. Ann; Rech. Forest : 61 -- 83.
HENDRIX, S D. (1984):
Variation in seed weight and its effects on germination in Pistanica sativa L Amer.J. Bot. 76(6): 790-805
HERTER,F ; BALANDIER,P. ; MAUGET, J.C. ;RAGEAU,R et BONHOMME, M. (1991) : Conséquences des conditions climatiques durant la croissance estivale et la période de repos sur la capacité de croissance des bourgeons chez deux espèces fruitières tempérées : le Pommier et le Pécher.
I.N.R.A : l'ARBRE, biologie et développement -- C. Edelined -- Naturalia Monspeliensia. HS: 417 -- 430.
KROUCHI, F. (1995):
Contribution à l'étude de l'organisation reproductive du cèdre de l'Atlas (Cedrus
atlantica Manetti) à Tal -- Guilef (Djurdjura Nord Occidental).
Thè. Mag. INA Alger : 105 p. 6 Annexes.
LACAZE, J.F (1984):
Régénération des forets par les semis I.N.R.A. Paris. 209-217
230
LAHMADI, M. et CHBOUKI, N. (1994) :
Les principaux facteurs influençant la régénération naturelle du cèdre de l'Atlas (Cedrus adantica M.).
Ann. Rech. For. Maroc. 27 Vol 2 : 244 -253.
LAÏMOUCHI, A. (1983) :
Contribution à la production du cèdre de l'atlas (Cedrus adantica. M) à Chréa (W.Blida Thè .Ing . Agr . I.N.A. El- Harrach . Alger. 96p.
LAÏMOUCHI, A. (1996) :
Contribution à l'étude écophysiologique de Cedrus atlantica Manetti au Parc National de Chréa (Wilaya de Blida).
Thè. Mag. INA. Alger : 83 p III Annexes
LALONDE, R.G. ROITBERG, B D. (1989) :
Ressources limitation and offspring size and number of trade offs in Cirsium arvence (Asteraceae) .
Amer.J.Bot. 76 (8) : 1107-1113.
LAMOTTE, M. (1971) :
Initiation aux méthodes statistiques en biologie ; Ed. MASSON et Cie. Paris : 144 p.
LANIER, L. (1993) :
Les champignons des cédraies en Algérie.(Etude comparative.) Ann. Rech. For Maroc. 27 Vol 2 :554-561.
LAPIE, G. (1909) :
Etude phytogéographique de la Kabylie du Djurdjura. Thèse es Sciences, Université Paris : 156 p.
LEBRU, C. (1970) :
Prétraitement des graines de Douglas à l'eau oxygénée. R.F.F. XXII -- 4 : 473 -- 476.
LEDANT, JP. (1975):
Essences forestières algériennes (Indigènes et exotiques ) Notes de dendrologie. Dépt de forêt.
INA El-Harrach .Alger :133p
LEPOUTRE, B. (1961) :
Recherches sur les conditions édaphiques de la régénération des cédraies marocaines. Ann. Rech. For. Au Maroc, T 6, Fasc. 2, S.R.F. Rabat : 211 p.
LEPOUTRE, B. (1963) :
Observations sur la régénération du cèdre par taches. Ann. Rech. Maroc. Fasc. 1, S.R.F Rabat : 17 p.
LEPOUTRE, B. (1964) :
Premier essai de synthèse sur le mécanisme de régénération du cèdre de l'Atlas dans le Moyen Atlas marocain.
Ann. Rech. For. Maroc. T 7, Rabat : 57 -- 163.
LEPOUTRE, B. et PUJOS (1963) :
Facteurs climatiques déterminant les conditions de germination et d'installation des plantules de cèdre.
Ann. Rech. For. Maroc. T7. 23-54
LE TACON, F. et OSWALD. H (1977) :
Influence de la fertilisation minérale sur la fructification du hêtre ( Fagus silvatica L) Ann. Sci For. , 34 (2).80-108
231
LETREUCH -- BELAROUCI, N. (1991):
Les reboisements en Algérie et leurs perspectives d'avenir.
Vol. 1 ; Office des Publications Universitaires. Alger : 294 p
MALKI, H. (1992) :
Contribution à l'étude de l'influence du climat et des facteurs physiques sur la régénération du cèdre de l'Atlas (Cedrus atlantica M.) dans les monts de Belezma (Algérie). Thè. Doc. Uni. de Paris -- Sorbonne : 187 p + 37 Annexes.
MARION, J. (1955) :
La régénération naturelle d cèdre de l'Atlas dans les cédraies du rebord septentrional du Moyen Atlas Occidental calcaire.
Ann. Rech. For. au Maroc.
Rapport annuels 1953 -- 1954 -- 1955, Fasc. 1:33 -- 149.
MAZLIAK, P. (1982) :
Physiologie végétale II, croissance et développement. Collection Méthodes des Herman, Paris : 465 p.
MEDDOUR, H. (1992) :
Contribution à l'étude dendrochronologique de deux provenances de pin d'Alep (Pinus halepensis Mill) en Algérie. Cas d'une station littorale et d'une autre continentale.
Thè. Ing. Agro. NA. 177 p.
M'HIRIT, 0. (1982) :
Etude écologique et forestière des cédraies du Rif Marocain : Essai sur une approche multidimensionnelle de la phytoécologie et de la production du cèdre de l'Atlas
Ann. Rech. For. Maroc. Vol.2 : 499p
M'HIRIT, 0. (1993) :
Le cèdre de l'Atlas (Cedrus atlantica. Manetti )
Présentation générale et état des connaissances à travers le Réseau.Méditerranéen " Le CEDRE "
Ann. Rech. For. Au Maroc. T 22 : 4-15
MONET, L. (1964) :
Manuel de Paléontologie végétale.
Masson et Cie, 3ème édition. Librairie de l'Académie de Médecine. Paris. 168 -- 169.
MOUNA, M. (1993) :
Entomofaune du cèdre de l'Atlas au Maroc.
Silva Mediterranea Actes du Séminaire International sur le cèdre de l'Atlas. Ifrane (Maroc) ; 7 --11 juin 1993 : 515 -- 522.
MULLER, C. et BONNET -- MASSIMBERT, M. (1983) :
Amélioration de la germination des faînes (Fagus silvatica) par traitement en présence de polyéthylène glycol.
Station d'amélioration des arbres fruitiers Champénoux. INRA .
Ann. Sci. For. 40 (2) : 157 -- 164
MULLER, C. (1986) :
Le point sur la conservation des semences forestières et la levée de dormance. R.F.F.XXXVIII-3- 200 - 204
MULLER, C. ; LAROPPE, E. et BONNET -- MASSIMBERT, M. (1984) :
Nouvelles voies dans le traitement des graines dormantes de certains feuillus: Frêne, Merisier .Rev. For. Fr. XLII- 3: 329-345.
MULLER, C. et BONNET -- MASSIMBERT, M. (1990 ) :
Levée de dormance des faines avant leur conservation : résultats préliminaires INRA. Station d'amélioration des arbres forestiers. Champénoux : 386 -- 394.
232
MULLER, C. et BONNET -- MASSIMBERT, M. et LAROPPE,F (1990 ) :
Nouvelles voies dans le traitement des graines dormantes de certains feuillus : Hêtre Frêne , Merisier
Rev. For. XLII --3. 329-345
MULTON, J.L. (1982):
Conservation et stockage des graines et des grains et produits dérives. Céréales oléagineux, protéagineux, aliments pour animaux.
Sciences et techniques Agro -- Alimentaire. Tech. et doc. Lavoisier. Paris : 586 p.
MUNAUT, A.V. (1982) :
Les cernes de croissances des arbres (la dendrochrotypologie). Typologie des sources du moyen âge occidental.
Edit. BREPOLS TURNHOT-BELGIUM. 52 p.
NEDJAHI, A. (1988) :
La cédraie du Chréa (Atlas Blideen).
Phénologie, productivité, régénération.
Thè. Doc. 3éme cycle. Uni. NANCY I : 184 p.
NEDJAHI, A. (1993) :
Etude de la croissance radiale des provenances du cèdre de l'Atlas (Cedrus adantica Manetti) en Algérie.
Silva Mediterranea Actes du Séminaire International sur le cèdre de l'Atlas. Ifrane (Maroc) ; 7 --11 juin 1993: 452 -- 461.
OSWALD, H.(1984)
Floraison, pollinisation et fructification chez le Hêtre ( Fagus silvatica L.) I.N.R.A. Paris .243-258.
OSSET,E .(1988):
Problèmes de régénération dans les forets résineuses subalpines riches en mégaphorbiais . Régénérations des forets d'altitude.
Université de Savoie.Office National des Foret : 49-56.
OTT, E.(1988):
Problèmes de régénération dans les forets résineuses riches en mégaphorbiais . Régénérations des forets d'altitude. Université de Savoie.Office National des Foret : 42-56
PAGES, L. ; KERVELLA, J. et CHADOEUF, J. (1991) :
Mise en place du système racinaire du pécher (Pinus persitu L. BATSCH) ; I.N.R.A. L'ARBRE Biologie et développement -C. Edelin et Naturalia Monspeliensia.H.S:646- 647.
PELLISSIER, F. et TROSSET, L. (1992) :
Les difficultés de régénération naturelle des péssières subalpines : prédation des graines au sol et blocages dus à l'humus.
INRA. Ann. Sci. For. 49 : 383 -- 388.
PEYERIMHOFF, P. (1941) :
Carte forestière de l'Algérie et de la Tunisie.
1 Vol. Imp. Pape Baconnier Frères : 70 p., 1 carte.
PHILIPPEAU, G. (1985) :
Théorie des plans d'expérience, Application à l'Agronomie. Service des Etudes Statistiques de l'I.T.C.F. Paris : 55 -- 75.
PIUSSI,P.(1988):
Accroissement et survie des jeunes épicéas dans une trouées en coupe. Régénérations des forets d'altitude.
Université de Savoie.Office National des Foret : 57-60
PRADAL, F. (1979) :
Variabilité génétique et écophysiologique du cèdre.
Mémoire de fin d'études à l'E.N.I.T.E.F -- INRA. Bordeaux et de Ruscas : 1 - 6.
233
POLUNIN, N. (1967) :
Eléments de géographie botanique.
GAUTHIER -- VILLARS, Paris : 195 -- 198.
PUJOS, A. (1964) :
Les milieux de la cédraie marocaine.
Etude d'une classification des cédraies du Moyen Atlas et du Rif en fonction du sol et du climat de la régénération actuelle dans les peuplements.
Ann. Rech. For. Maroc . 8: Rabat 283
PUTOD,R .(1979) :
Le cèdre de l'Atlas en France. Bull . Vulg : 75 p
QUEZEL, P. (1957) :
Le peuplement végétal des hautes montagnes d'Afrique du Nord. Ed. Le Chevalier, Paris : 15-30
RIPERT, C. et BOISSEAU, B. (1993) :
Ecologie et croissance de cèdre de l'Atlas en Provence.
Silva Mediterranea Actes du Séminaire International sur le cèdre de l'Atlas. Ifrane (Maroc) ; 7 --11 juin 1993: 156 --164.
ROQUES, C. (1983) :
Les insectes ravageurs de cônes et graines des conifères en France. Station Zoologique Biocenotique. Forest. INRA.
Centre de recherche d'Orléans. Paris:71-73.
ROQUES, C. (1988) :
Impact des insectes ravageurs des cônes et graines sur les potentialités de régénération naturelle des principales essences constituant les forets d'altitude du Brianconnais.Régénérations des forets d'altitude.
Université de Savoie.Office National des Foret : 17-28
SABATIER, S. et BARTHELEMY, D. (1993) :
Premières observations sur l'architecture du cèdre de l'Atlas, Cedrus atlantica (ENDL.) MANETTIEX CARRIERE (PINCEAE).
Silva Mediterranea Actes du Séminaire International sur le cèdre de l'Atlas. Ifrane (Maroc) ; 7 --11 juin 1993 : 364 -- 377.
SARI, Dj. (1977) :
L'homme et l'érosion dans l'Ouarsenis (Algérie). SNED. Alger. 63 -- 204.
SCHOENBERGER, A. (1970) :
Inventaire des massifs de S'GAG et Chelia. FAO -- projet Algérie 15 : 14 p.
SKRZYPCZYNSKA,M.(1988)
Megastigmus suspectus BORR ./ HYM ., TORYMIDAE / et Resseliella piceae SEITN./DIPT., CECIDOMY , IIDAE / dans la zone du sapin Ables alba Mill. en Pologne. Régénérations des forets d'altitude.
Université de Savoie.Office National des Forêts : 29-33
STEWART,P ( 1969) :
Quotient pluviométrique et dégradation de la biosphère Bull.Soc.Hist.Nat.Afr.du Nord ;Alger,59 :14
STEWART,P ( 1974) :
Cours de sylviculture.
Institut National Agronomique.
Dpt de foresterie. El-Harrach -Alger :101p
234
TAN, B.S ; (1987) :
Etude de l'influence de quelques facteurs écologiques sur les premiers stades de la régénération du sapin (Ables alba) à l'étage montagnard du Jura.
Rev. Ecol. Biol. Sol. 24 (4) : 623 -- 635.
TAN, B.S ; (1987) :
Effets des facteurs physiques sur les premiers stades de la régénération naturelle Sapin pectiné (Ables alba M) dans quelques stations du Jura.
Ann.Sci.For.49.337-350
TILL, C. (1985) :
Recherches dendrochronologiques sur le cèdre de l'Allas (C,edrus atlantica Endl. Carrière) au Maroc. Thè. Doc. Uni. Catholique de Louvain Fac des Sciences. Louvain - la Neuve. 231 p.
TOTH, J. (1970) :
Plus que centenaire et plein d'avenir : le cèdre en France. R.F.F. XXII. 3 : 355 -- 364
TOTH, J. (1971) :
Le cèdre (Cedrus atlantica) en France.
Bull. de vulgarisation forestière n° 4 : 5 -- 19.
TOTH, J. (1973 a) :
Contribution à l'étude de la dissémination des graines de cèdre (Cedrus_atlantica Manetti) sur la face du Mont -- Ventoux (Vaucluse).
Inst. Nat. de Rech. Agro. C.N.R.F. Doc n° 6. Avignon Dist. Lim. : 1-- 28.
TOTH, J. (1973 b) :
Première approche de la production potentielle du cèdre de l'Atlas dans le Sud de la France. R.F.F XXV. 5 : 381 -- 389.
TOTH, J. (1975) :
Etude sur la dissémination des graines et de la
régénération naturelle du cèdre au
Mont-Ventoux.
Dispositif expérimental n°84-7. Protocole d'installation -
Avignon : 1- 7.
TOTH, J. (1978 a) :
Contribution à l'étude de la fructification et de la régénération du cèdre de l'Atlas (Cedrus atlantica M.) dans le Sud de la France.
Thè. Doc. Ing. Fac. St. Jérôme -- Marseille, France : 136 p.
TOTH, J. (1978 b) :
Prévision de production de cônes de cèdre ;
Estimation qualitative et quantitative.
Protocole n° 11.
Ministère de l'Agriculture. I.N.R.A. Stat. Sylv. Medit. Avignon:1- 3
TOTH, J. (1980 a):
Le cèdre dans quelques pays du pourtour méditerranéen et dans deux autres pays à grande importance forestière. Forêt Méditerranéenne. T. II. N° 1 : 23 -- 30.
TOTH, J. (1980 b) :
Le cèdre II. La graine :Dissémination ; extraction, qualité, germination, conservation. La forêt privée. Rev. For. Europ. n° 131 : 78 --84.
TOTH, J. (1980 c) :
Le cèdre III. La graine des plants en pépinière, reboisement, régénération naturelle. La forêt privée. Rev. For ; Europe. n° 132 : 41 -- 47.
TOTH, J. (1981) :
Contribution à l'étude monographique du cèdre méditerranéen. INRA. Station d'Avignon : 25 p.
TOTH, J. (1982) :
Analyse de la croissance juvénile sur trois essences résineuses -- cèdre -- pin noir et pin de salzmann dans le reboisement de Belbezet (Gara).
Foret méditerranéenne. T. IV n° 2 : 143 -- 146.
235
TOTH, J. (1982 -- 1984) :
Quelques éléments nouveaux pour mieux situer et caractériser le cèdre de l'Himalaya en France vis-à-vis du cèdre de l'Atlas et du cèdre du Liban en France méridionale.
INRA. Centre de recherche forestière d'Avignon. Bull. Soc et Sci. Nat. Vaucluse : 41- 49.
TOTH, J. (1983) :
La prévision et l'estimation de récoltes de cônes de cèdre de l'Atlas Doc. Int. Stat. Sylv. I.N.R.A. Avignon : 10 p
TOTH, J. (1984) :
La prévision des possibilités de récoltes de cônes de cèdre de l'Atlas(Cedrus atlantica Manetti). Bull. Tech. n° 15 O.N.F : 39 -- 51.
TOTH, J. (1987 a) :
Installation et développement des semis naturels « pin noir d'Autriche, pin sylvestre et cèdre de l'Atlas » après incendie sur le Mont -- Ventoux (Vaucluse).
Forêt méditerranéenne. T. IX. n° 1 : 27 -- 36.
TOTH, J. (1987 b) :
Effets des facteurs d'environnement sur l'accroissement du cèdre de l'Atlas. Bull. Soc. Sci. Nat. Vaucluse : 71 -- 77.
TOTH, J. (1988 a) :
Le Mont -- Ventoux forestier.
Doc. interne n°7.C.N.R.F.Station de sylviculture méditerranéenne. Avignon : 28p.
TOTH, J. (1988 b) :
Croissance juvénile de trois provenances françaises de cèdre de l'Atlas étudiées dans une mono -- station à Citon.
Doc. interne n° LINRA-CNRF-Station de sylviculture méditerranéenne.Avignon : 80 p.
TOTH, J. (1990 a) :
Le cèdre III. Intérêt paysager.
Cédraie touristique. Forêt privée n° 195 : 8 p.
TOTH, J. (1990 b) :
Croissance, Sylviculture et production du cèdre de l'Atlas (Cedrus adantica_Manetti) implanté dans une zone naturelle du chêne vert (Quercus ilex L.) en région méditerranéenne Française.
Exposé présenté au Symposium International du cèdre à Antalya (Turquie) : 1-- 7.
TROSSET, L.( 1988) :
La régénération des péssières d'altitude :comportement des semis expérimentaux soumis à différents traitements
Régénérations des forets d'altitude.
Université de Savoie.Office National des Foret : 99-112
VALETTE,J.Ch.(1983) :
Influence du mode de plantation de cèdres 1+OG ( Sacs de polyéthylène) sur leur reprise et leur croissance initiale.Etude menée en pépinière.
R.F.XXXV-4 :289-233
VLASSEV, V. (1973) :
Croissance en groupes du cèdre dans la montagne des Babors.
Ann. de l'Inst. Nat. Agro. Alger, II. 2: 115 --141.
ZAKI, A. (1969) :
Premières études sur les phénomènes de dormance de la graine de cèdre de l'Atlas et sur l'influence de différents facteurs à l'égard de sa germination. Ann. Rech. For. Maroc, T 2. Ed. Tech. Nord Africaine : 23 -- 52.
236
|
Mois |
Station |
Versant Nord |
Versant Sud |
||||||||
|
Altitude en m |
|||||||||||
|
1400 |
1600 |
1700 |
1800 |
2000 |
1400 |
1600 |
1700 |
1800 |
2000 |
||
|
Janvier |
45.36 |
50.21 |
58.19 |
62.18 |
66.18 |
74.11 |
47.79 |
51.77 |
53.77 |
55.77 |
59.75 |
|
Février |
48.20 |
53.35 |
61.84 |
66.08 |
70.32 |
78.75 |
50.78 |
55.02 |
57.14 |
59.26 |
63.49 |
|
Mars |
53.36 |
59.06 |
68.46 |
73.15 |
77.85 |
87.19 |
56.22 |
60.91 |
63.25 |
65.65 |
70.29 |
|
Avril |
44.22 |
48.95 |
56.73 |
60.62 |
64.51 |
72.25 |
46.59 |
50.47 |
52.42 |
54.36 |
58.25 |
|
Mai |
49.58 |
54.88 |
63.61 |
67.67 |
72.33 |
81.01 |
52.23 |
56.59 |
58.77 |
60.95 |
65.31 |
|
Juin |
17.67 |
19.56 |
22.67 |
24.22 |
25.78 |
28.87 |
18.61 |
20.17 |
20.94 |
21.72 |
23.27 |
|
Juillet |
03.37 |
3.73 |
4.32 |
4.62 |
4.91 |
5.50 |
3.55 |
3.84 |
3.39 |
4.14 |
4.43 |
|
Août |
13.11 |
14.51 |
16.82 |
17.97 |
19.12 |
21.42 |
13.81 |
14.96 |
15.54 |
16.11 |
17.27 |
|
Septembre |
33.07 |
36.60 |
42.42 |
45.33 |
48.24 |
54.03 |
34.84 |
37.74 |
39.20 |
40.64 |
43.56 |
|
Octobre |
40.48 |
44.81 |
51.93 |
55.49 |
59.06 |
66.14 |
42.64 |
46.20 |
47.98 |
49.77 |
53.32 |
|
Novembre |
54.34 |
60.15 |
69.71 |
74.50 |
79.28 |
88.79 |
57.27 |
62.02 |
64.42 |
66.81 |
71.58 |
|
Décembre |
52.11 |
57.68 |
66.85 |
71.44 |
76.03 |
85.14 |
54.90 |
59.48 |
61.77 |
64.06 |
68.64 |
|
Moyenne |
37.90 |
41.97 |
48.63 |
51.97 |
55.30 |
61.97 |
39.93 |
43.27 |
44.93 |
46.60 |
49.93 |
|
Années |
Précipitations |
Précipitations saisonnières (mm) |
Régime |
|||
|
Hiver (H) |
Printemps (P) |
Eté (E) |
Automne (A) |
|||
|
1975 |
361 |
150 |
127 |
04 |
80 |
HPAE |
|
1976 |
588 |
181 |
179 |
81 |
198 |
A1-IPE |
|
1977 |
338 |
114 |
143 |
23 |
60 |
PHAE |
|
1978 |
319 |
60 |
142 |
25 |
92 |
PARE |
|
1979 |
403 |
98 |
144 |
48 |
112 |
PARE |
|
1980 |
601 |
206 |
243 |
11 |
141 |
PHAE |
|
1981 |
248 |
70 |
68 |
55 |
56 |
HPAE |
|
1982 |
709 |
184 |
154 |
25 |
345 |
AHPE |
|
1983 |
202 |
57 |
57 |
11 |
77 |
AHPE |
|
1984 |
454 |
265 |
46 |
33 |
109 |
HAPE |
|
1985 |
583 |
242 |
193 |
00 |
148 |
HPAE |
|
1986 |
495 |
155 |
144 |
06 |
189 |
AHPE |
|
1987 |
423 |
175 |
95 |
64 |
89 |
HPAE |
|
1988 |
296 |
131 |
65 |
42 |
57 |
HPAE |
|
1989 |
439 |
90 |
134 |
147 |
67 |
EPHA |
|
1990 |
591 |
195 |
243 |
54 |
98 |
PHAE |
|
1991 |
566 |
96 |
271 |
28 |
172 |
PARE |
|
1992 |
691 |
175 |
268 |
21 |
228 |
PARE |
|
1993 |
395 |
127 |
117 |
23 |
129 |
AHPE |
|
Moy |
456 |
146 |
147 |
34 |
129 |
PHAE |
238
|
Années |
Mois |
Versant Nord |
Versant Sud |
||||||
|
Poids |
Longueur |
Poids |
Longueur |
||||||
|
g |
% / cônes |
g |
% / cônes |
g |
% / cônes |
g |
% / cônes |
||
|
N |
10 Sept |
0.0695 |
0.0705 |
1.08 |
13.70 |
0.0783 |
0.0855 |
1.01 |
14.06 |
|
05 Oct |
0.0987 |
0.1000 |
1.21 |
15.35 |
0.0941 |
0.102 |
1.20 |
16.71 |
|
|
15 Oct |
0.114 |
0.115 |
1.28 |
16.24 |
0.0978 |
0.106 |
1.21 |
16.85 |
|
|
30 Nov |
0.115 |
0.116 |
1.28 |
16.24 |
0.0978 |
0.106 |
1.21 |
16.85 |
|
|
N + 1 |
30 Mars |
0.201 |
0.204 |
1.51 |
19.16 |
0.178 |
0.194 |
1.49 |
20.75 |
|
15 Avr |
0.335 |
0.340 |
1.90 |
24.11 |
0.196 |
0.214 |
1.51 |
21.03 |
|
|
01 Mai |
1.301 |
1.320 |
2.30 |
29.18 |
0.370 |
0.404 |
1.57 |
21.86 |
|
|
20 Mai |
3.028 |
3.073 |
2.63 |
33.27 |
1.780 |
1.944 |
2.07 |
28.83 |
|
|
02 Jui |
15.199 |
15.427 |
4.33 |
54.94 |
6.718 |
7.337 |
2.98 |
41.50 |
|
|
11 Jui |
30.215 |
30.668 |
4.35 |
55.20 |
27.432 |
29.960 |
4.24 |
59.05 |
|
|
09 Juil |
80.951 |
82.116 |
6.72 |
85.27 |
78.840 |
86.106 |
6.74 |
93.87 |
|
|
30Nov |
91.542 |
92.916 |
6.75 |
85.65 |
88.936 |
97.133 |
6.76 |
94.15 |
|
|
N + 2 |
04 Fev |
81.236 |
82.455 |
6.51 |
82.61 |
76.312 |
83.345 |
6.73 |
93.73 |
|
06Juil |
80.556 |
81.765 |
6.38 |
80.96 |
74.111 |
80.941 |
5.97 |
83.14 |
|
|
23Dec |
89.651 |
90.996 |
7.33 |
93.02 |
85.102 |
92.945 |
6.50 |
90.52 |
|
|
N+ 3 |
13 Jan |
98.521 |
100 |
7.88 |
100 |
91.561 |
100 |
7.18 |
100 |
|
Années de |
Nombres d'inflorescences |
Nombre de |
Nombre de cônes |
Année de |
||
|
Sept N |
Oct N |
Mai N+1 |
Août N+1 |
Sept N+2 |
Cônes N+2 |
|
|
1974 |
824 |
504 |
368 |
104 |
104 |
1976 |
|
1975 |
146 |
116 |
48 |
2 |
2 |
1977 |
|
1976 |
250 |
186 |
56 |
68 |
68 |
1978 |
|
1977 |
1204 |
466 |
162 |
284 |
284 |
1979 |
|
1978 |
43 |
14 |
2 |
0 |
0 |
1980 |
|
1979 |
283 |
183 |
68 |
11 |
11 |
1981 |
|
1980 |
1142 |
900 |
315 |
151 |
151 |
1982 |
|
1981 |
3 |
2 |
0 |
104 |
104 |
1983 |
|
1982 |
42 |
14 |
11 |
2 |
2 |
1984 |
|
1983 |
1125 |
316 |
185 |
68 |
68 |
1985 |
|
Moyenne |
392 |
270 |
151 |
79 |
79 |
--- |
TOTH (1984)
239
Sud : Faible uroduction
|
Arbre- échantillons N° |
07 |
08 |
09 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
Total |
|
|
Inflorescences |
Branches basses |
45 |
18 |
91 |
17 |
46 |
28 |
36 |
21 |
101 |
32 |
19 |
54 |
37 |
61 |
53 |
659 |
|
Branches hautes |
83 |
57 |
101 |
68 |
36 |
17 |
39 |
18 |
83 |
76 |
93 |
68 |
44 |
26 |
35 |
874 |
|
|
Somme des deux branches |
128 |
75 |
192 |
85 |
82 |
45 |
75 |
39 |
184 |
108 |
112 |
122 |
81 |
87 |
88 |
1503 |
|
|
cônes N+1 |
Branches basses |
1 |
1 |
2 |
1 |
2 |
2 |
2 |
1 |
1 |
2 |
3 |
2 |
1 |
2 |
1 |
24 |
|
Branches hautes |
4 |
3 |
2 |
3 |
4 |
2 |
3 |
3 |
2 |
2 |
5 |
2 |
5 |
1 |
4 |
45 |
|
|
Somme des deux branches |
5 |
4 |
4 |
4 |
6 |
4 |
5 |
4 |
3 |
4 |
8 |
4 |
6 |
3 |
5 |
69 |
|
|
cônes N+2 |
Branches basses |
1 |
0 |
2 |
1 |
0 |
1 |
1 |
0 |
1 |
0 |
0 |
1 |
0 |
1 |
0 |
9 |
|
Branches hautes |
2 |
1 |
1 |
1 |
1 |
2 |
0 |
1 |
0 |
0 |
1 |
0 |
1 |
1 |
0 |
12 |
|
|
Somme des deux branches |
3 |
1 |
3 |
2 |
1 |
3 |
1 |
1 |
1 |
0 |
1 |
1 |
1 |
2 |
0 |
21 |
|
Sud : Moyenne uroduction
|
Arbre- échantillons N° |
07 |
08 |
09 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
Total |
|
|
Inflorescences |
Branches basses |
14 |
12 |
31 |
8 |
19 |
25 |
8 |
5 |
13 |
63 |
10 |
37 |
61 |
60 |
28 |
394 |
|
Branches hautes |
23 |
12 |
17 |
32 |
9 |
11 |
65 |
4 |
5 |
49 |
43 |
26 |
71 |
44 |
49 |
460 |
|
|
Somme des deux branches |
37 |
24 |
48 |
40 |
28 |
36 |
73 |
9 |
18 |
112 |
53 |
63 |
132 |
104 |
77 |
854 |
|
|
cônes N+1 |
Branches basses |
21 |
5 |
9 |
3 |
7 |
13 |
21 |
9 |
31 |
11 |
8 |
6 |
4 |
12 |
8 |
168 |
|
Branches hautes |
10 |
11 |
6 |
17 |
6 |
17 |
17 |
5 |
16 |
6 |
41 |
18 |
19 |
6 |
10 |
205 |
|
|
Somme des deux branches |
31 |
16 |
15 |
20 |
13 |
30 |
38 |
14 |
47 |
17 |
49 |
24 |
23 |
18 |
18 |
373 |
|
|
cônes N+2 |
Branches basses |
1 |
1 |
2 |
1 |
2 |
2 |
2 |
1 |
1 |
2 |
3 |
2 |
1 |
2 |
1 |
24 |
|
Branches hautes |
4 |
3 |
2 |
3 |
4 |
2 |
3 |
3 |
2 |
2 |
5 |
2 |
5 |
1 |
4 |
45 |
|
|
Somme des deux branches |
5 |
4 |
4 |
4 |
6 |
4 |
5 |
4 |
3 |
4 |
8 |
4 |
6 |
3 |
5 |
69 |
|
Sud : Forte uroduction
|
Arbre- échantillons N° |
07 |
08 |
09 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
Total |
|
|
Inflorescences |
Branches basses |
0 |
2 |
1 |
3 |
0 |
0 |
1 |
15 |
5 |
1 |
5 |
1 |
1 |
1 |
2 |
38 |
|
Branches hautes |
1 |
9 |
0 |
4 |
1 |
6 |
0 |
9 |
12 |
1 |
2 |
0 |
5 |
0 |
6 |
56 |
|
|
Somme des deux branches |
1 |
11 |
1 |
7 |
1 |
6 |
1 |
24 |
17 |
2 |
7 |
1 |
6 |
1 |
8 |
94 |
|
|
cônes N+1 |
Branches basses |
12 |
10 |
2 |
2 |
3 |
4 |
2 |
2 |
4 |
3 |
8 |
4 |
32 |
6 |
2 |
96 |
|
Branches hautes |
19 |
12 |
9 |
2 |
5 |
6 |
31 |
3 |
2 |
7 |
21 |
5 |
19 |
4 |
12 |
157 |
|
|
Somme des deux branches |
31 |
22 |
11 |
4 |
8 |
10 |
33 |
5 |
6 |
10 |
29 |
9 |
51 |
10 |
14 |
253 |
|
|
cônes N+2 |
Branches basses |
21 |
5 |
9 |
3 |
7 |
13 |
21 |
9 |
31 |
11 |
8 |
6 |
4 |
12 |
8 |
168 |
|
Branches hautes |
10 |
11 |
6 |
17 |
6 |
17 |
17 |
5 |
16 |
6 |
41 |
18 |
19 |
6 |
10 |
205 |
|
|
Somme des deux branches |
31 |
16 |
15 |
20 |
13 |
30 |
38 |
14 |
47 |
17 |
49 |
24 |
23 |
18 |
18 |
373 |
|
Nord : Faible uroduction
|
Arbre- échantillons N° |
07 |
08 |
09 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
Total |
|
|
Inflorescences |
Branches basses |
57 |
11 |
23 |
65 |
43 |
32 |
32 |
48 |
22 |
23 |
11 |
26 |
22 |
28 |
24 |
467 |
|
Branches hautes |
72 |
49 |
76 |
54 |
48 |
58 |
58 |
66 |
26 |
14 |
21 |
15 |
55 |
51 |
53 |
716 |
|
|
Somme des deux branches |
129 |
60 |
99 |
119 |
91 |
90 |
90 |
114 |
48 |
37 |
32 |
41 |
77 |
79 |
64 |
1183 |
|
|
cônes N+1 |
Branches basses |
2 |
4 |
2 |
1 |
11 |
1 |
1 |
8 |
1 |
1 |
2 |
1 |
4 |
1 |
1 |
42 |
|
Branches hautes |
4 |
5 |
4 |
3 |
5 |
3 |
3 |
7 |
2 |
2 |
2 |
7 |
8 |
1 |
2 |
56 |
|
|
Somme des deux branches |
6 |
9 |
6 |
4 |
16 |
4 |
4 |
15 |
3 |
3 |
4 |
8 |
12 |
2 |
3 |
98 |
|
|
cônes N+2 |
Branches basses |
1 |
0 |
3 |
0 |
2 |
1 |
1 |
1 |
0 |
0 |
1 |
0 |
1 |
0 |
0 |
11 |
|
Branches hautes |
3 |
1 |
2 |
2 |
1 |
3 |
3 |
2 |
2 |
1 |
1 |
2 |
1 |
2 |
1 |
27 |
|
|
Somme des deux branches |
4 |
1 |
5 |
2 |
3 |
4 |
4 |
3 |
2 |
1 |
2 |
2 |
2 |
2 |
1 |
38 |
|
Nord : Moyenne uroduction
|
Arbre- échantillons N° |
07 |
08 |
09 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
Total |
|
|
Inflorescences |
Branches basses |
15 |
20 |
8 |
10 |
11 |
10 |
112 |
25 |
25 |
5 |
25 |
4 |
3 |
1 |
6 |
280 |
|
Branches hautes |
89 |
31 |
15 |
6 |
32 |
36 |
40 |
28 |
40 |
10 |
18 |
4 |
75 |
0 |
26 |
450 |
|
|
Somme des detuc branches |
104 |
51 |
23 |
16 |
43 |
46 |
152 |
53 |
65 |
15 |
43 |
8 |
78 |
1 |
32 |
730 |
|
|
cônes N+1 |
Branches basses |
32 |
6 |
8 |
5 |
40 |
34 |
16 |
36 |
15 |
12 |
5 |
17 |
11 |
12 |
10 |
259 |
|
Branches hautes |
37 |
32 |
32 |
4 |
44 |
31 |
34 |
31 |
10 |
6 |
2 |
5 |
31 |
32 |
40 |
370 |
|
|
Somme des deux branches |
69 |
38 |
40 |
9 |
84 |
65 |
50 |
67 |
25 |
18 |
7 |
22 |
41 |
44 |
50 |
629 |
|
|
cônes N+2 |
Branches basses |
2 |
4 |
2 |
1 |
11 |
1 |
1 |
8 |
1 |
1 |
2 |
1 |
4 |
1 |
1 |
42 |
|
Branches hautes |
4 |
5 |
4 |
3 |
5 |
3 |
3 |
7 |
2 |
2 |
2 |
7 |
8 |
1 |
2 |
56 |
|
|
Somme des deux branches |
6 |
9 |
6 |
4 |
16 |
4 |
4 |
15 |
3 |
3 |
4 |
8 |
12 |
2 |
3 |
98 |
|
Nord : Forte uroduction
|
Arbre- échantillons N° |
07 |
08 |
09 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
Total |
|
|
Inflorescences |
Branches basses |
1 |
0 |
0 |
3 |
10 |
0 |
0 |
1 |
0 |
6 |
3 |
0 |
0 |
6 |
0 |
30 |
|
Branches hautes |
0 |
2 |
0 |
4 |
11 |
0 |
0 |
1 |
0 |
8 |
2 |
0 |
0 |
12 |
0 |
40 |
|
|
Somme des deux branches |
1 |
2 |
0 |
7 |
21 |
0 |
0 |
2 |
0 |
14 |
5 |
0 |
0 |
18 |
0 |
70 |
|
|
cônes N+1 |
Branches basses |
12 |
0 |
3 |
1 |
10 |
6 |
47 |
1 |
5 |
2 |
11 |
2 |
1 |
1 |
3 |
105 |
|
Branches hautes |
23 |
4 |
5 |
6 |
28 |
5 |
19 |
7 |
1 |
8 |
17 |
3 |
33 |
0 |
21 |
180 |
|
|
Somme des deux branches |
35 |
4 |
8 |
7 |
38 |
11 |
66 |
8 |
6 |
10 |
28 |
5 |
34 |
1 |
24 |
285 |
|
|
cônes N+2 |
Branches basses |
32 |
6 |
8 |
5 |
40 |
34 |
16 |
36 |
15 |
12 |
5 |
17 |
11 |
12 |
10 |
259 |
|
Branches hautes |
37 |
32 |
32 |
4 |
44 |
31 |
34 |
31 |
10 |
6 |
2 |
5 |
30 |
32 |
40 |
370 |
|
|
Somme des detuc branches |
69 |
38 |
40 |
9 |
84 |
65 |
50 |
67 |
25 |
18 |
7 |
22 |
41 |
44 |
50 |
629 |
|
240
Exposition Nord
Année de faible production
|
Canes N° |
1 |
2 |
1 |
4 |
5 |
6 |
7 |
2 |
9 |
10 |
11 |
12 |
11 |
14 |
15 |
16 |
17 |
12 |
19 |
20 |
21 |
22 |
21 |
24 |
25 |
26 |
27 |
22 |
29 |
10 |
|
|
GRAINES FERTILES |
67 |
69 |
66 |
80 |
16 |
29 |
8 |
3 |
26 |
95 |
2 |
44 |
9 |
3 |
72 |
48 |
61 |
36 |
68 |
7 |
42 |
68 |
63 |
38 |
15 |
39 |
6 |
50 |
21 |
28 |
|
|
GRAINES |
ATTAQUEES |
5 |
3 |
2 |
4 |
1 |
7 |
0 |
1 |
4 |
2 |
0 |
5 |
3 |
3 |
4 |
5 |
2 |
0 |
1 |
2 |
3 |
2 |
1 |
3 |
0 |
4 |
0 |
1 |
3 |
2 |
|
APLATIES |
53 |
46 |
50 |
38 |
98 |
77 |
109 |
100 |
83 |
9 |
108 |
82 |
106 |
111 |
29 |
51 |
59 |
58 |
56 |
103 |
67 |
51 |
43 |
59 |
94 |
57 |
120 |
55 |
87 |
56 |
|
|
AIGUILLES |
9 |
10 |
9 |
14 |
32 |
11 |
10 |
14 |
13 |
12 |
36 |
9 |
10 |
3 |
11 |
7 |
10 |
15 |
13 |
18 |
20 |
16 |
14 |
16 |
23 |
8 |
13 |
16 |
12 |
12 |
|
|
GRAINES VAINES |
67 |
59 |
61 |
56 |
131 |
95 |
119 |
115 |
100 |
23 |
154 |
96 |
119 |
126 |
44 |
63 |
71 |
73 |
70 |
123 |
90 |
69 |
58 |
78 |
117 |
69 |
133 |
72 |
102 |
70 |
|
|
GRAINES TOTALES PAR CÔNE |
134 |
128 |
127 |
136 |
147 |
124 |
127 |
118 |
126 |
118 |
156 |
140 |
128 |
129 |
116 |
111 |
132 |
109 |
138 |
130 |
132 |
137 |
121 |
116 |
132 |
108 |
139 |
122 |
123 |
98 |
|
Année de moyenne production
|
Canes N° |
1 |
2 |
1 |
4 |
5 |
6 |
7 |
2 |
9 |
10 |
11 |
12 |
11 |
14 |
15 |
16 |
17 |
12 |
19 |
20 |
21 |
22 |
21 |
24 |
25 |
26 |
27 |
22 |
29 |
10 |
|
|
GRAINES FERTILES |
61 |
32 |
48 |
77 |
23 |
51 |
47 |
31 |
38 |
18 |
71 |
23 |
28 |
61 |
25 |
68 |
63 |
7 |
46 |
55 |
69 |
36 |
43 |
58 |
66 |
64 |
53 |
40 |
49 |
13 |
|
|
GRAINES |
ATTAQUEES |
1 |
7 |
3 |
8 |
5 |
3 |
4 |
7 |
4 |
6 |
11 |
6 |
8 |
8 |
11 |
4 |
11 |
5 |
3 |
7 |
16 |
4 |
8 |
11 |
8 |
1 |
2 |
6 |
4 |
6 |
|
APLATIES |
78 |
57 |
46 |
26 |
83 |
45 |
83 |
62 |
42 |
61 |
49 |
83 |
70 |
61 |
58 |
62 |
45 |
60 |
42 |
58 |
70 |
63 |
72 |
83 |
52 |
83 |
68 |
78 |
82 |
83 |
|
|
AIGUILLES |
6 |
4 |
27 |
14 |
18 |
16 |
18 |
12 |
21 |
9 |
7 |
9 |
22 |
9 |
16 |
3 |
6 |
13 |
29 |
19 |
2 |
6 |
4 |
21 |
2 |
3 |
4 |
9 |
17 |
4 |
|
|
GRAINES VAINES |
85 |
68 |
76 |
48 |
129 |
64 |
105 |
81 |
67 |
76 |
67 |
98 |
100 |
78 |
85 |
68 |
62 |
78 |
74 |
85 |
88 |
73 |
84 |
115 |
62 |
87 |
74 |
95 |
103 |
93 |
|
|
GRAINES TOTALES PAR CÔNE |
146 |
100 |
124 |
125 |
152 |
115 |
152 |
112 |
105 |
94 |
138 |
121 |
128 |
139 |
110 |
137 |
125 |
85 |
120 |
139 |
157 |
109 |
127 |
173 |
128 |
151 |
127 |
135 |
152 |
106 |
|
Année de forte production
|
Canes N° |
1 |
2 |
1 |
4 |
5 |
6 |
7 |
2 |
9 |
10 |
11 |
12 |
11 |
14 |
15 |
16 |
17 |
12 |
19 |
20 |
21 |
22 |
21 |
24 |
25 |
26 |
27 |
22 |
29 |
10 |
|
|
GRAINES FERTILES |
65 |
74 |
47 |
95 |
82 |
81 |
122 |
91 |
65 |
66 |
71 |
26 |
117 |
91 |
75 |
72 |
77 |
123 |
63 |
64 |
61 |
44 |
58 |
41 |
61 |
49 |
69 |
119 |
85 |
128 |
|
|
GRAINES |
ATTAQUEES |
33 |
2 |
20 |
28 |
17 |
19 |
5 |
31 |
30 |
35 |
13 |
2 |
19 |
5 |
31 |
15 |
14 |
12 |
0 |
6 |
24 |
8 |
3 |
74 |
3 |
13 |
2 |
12 |
10 |
7 |
|
APLATIES |
27 |
30 |
31 |
26 |
26 |
10 |
27 |
22 |
21 |
23 |
32 |
87 |
10 |
50 |
23 |
38 |
25 |
33 |
135 |
50 |
23 |
111 |
55 |
31 |
54 |
80 |
55 |
28 |
36 |
25 |
|
|
AIGUILLES |
29 |
9 |
28 |
18 |
40 |
43 |
6 |
19 |
20 |
27 |
12 |
19 |
15 |
6 |
38 |
12 |
32 |
10 |
8 |
8 |
59 |
14 |
58 |
10 |
20 |
17 |
10 |
17 |
22 |
22 |
|
|
GRAINES VAINES |
89 |
41 |
79 |
72 |
83 |
70 |
38 |
72 |
71 |
85 |
57 |
108 |
44 |
61 |
92 |
65 |
71 |
55 |
143 |
64 |
106 |
133 |
116 |
115 |
77 |
110 |
62 |
57 |
68 |
54 |
|
|
GRAINES TOTALES PAR CÔNE |
154 |
115 |
126 |
167 |
165 |
153 |
160 |
163 |
136 |
151 |
128 |
134 |
161 |
152 |
167 |
137 |
148 |
178 |
206 |
128 |
167 |
177 |
174 |
156 |
138 |
159 |
131 |
176 |
153 |
182 |
|
Ex position Sud
Année de faible production
|
Cônes N° |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
15 |
16 |
17 |
18 |
19 |
20 |
21 |
22 |
23 |
24 |
25 |
26 |
27 |
28 |
29 |
30 |
|
|
GRAINES FERTILES |
3 |
11 |
5 |
18 |
3 |
7 |
6 |
2 |
5 |
2 |
3 |
4 |
6 |
6 |
7 |
1 |
3 |
4 |
4 |
17 |
3 |
1 |
6 |
4 |
0 |
5 |
3 |
1 |
5 |
4 |
|
|
GRAINES |
ATTAQUEES |
1 |
0 |
0 |
4 |
2 |
3 |
1 |
0 |
0 |
1 |
0 |
0 |
2 |
1 |
1 |
0 |
3 |
0 |
0 |
10 |
1 |
0 |
1 |
1 |
0 |
0 |
0 |
1 |
0 |
0 |
|
APLATIES |
115 |
137 |
121 |
108 |
123 |
109 |
124 |
125 |
110 |
118 |
100 |
112 |
112 |
109 |
123 |
115 |
116 |
119 |
109 |
96 |
111 |
115 |
101 |
115 |
107 |
120 |
113 |
128 |
112 |
114 |
|
|
AIGUILLES |
22 |
17 |
25 |
18 |
23 |
12 |
17 |
18 |
22 |
17 |
18 |
18 |
18 |
13 |
18 |
26 |
5 |
10 |
17 |
21 |
17 |
15 |
19 |
20 |
18 |
12 |
18 |
35 |
18 |
21 |
|
|
GRAINES VAINES |
138 |
154 |
146 |
130 |
148 |
124 |
142 |
143 |
132 |
136 |
118 |
130 |
132 |
123 |
142 |
141 |
124 |
129 |
126 |
118 |
129 |
130 |
121 |
136 |
125 |
132 |
131 |
164 |
130 |
135 |
|
|
GRAINES TOTALES PAR CÔNE |
141 |
165 |
151 |
148 |
151 |
131 |
148 |
145 |
137 |
138 |
121 |
134 |
138 |
129 |
149 |
142 |
127 |
133 |
130 |
135 |
132 |
131 |
127 |
140 |
125 |
137 |
134 |
165 |
135 |
139 |
|
Année de moyenne production
|
Canes N° |
1 |
2 |
1 |
4 |
5 |
6 |
7 |
2 |
9 |
10 |
11 |
12 |
11 |
14 |
15 |
16 |
17 |
12 |
19 |
20 |
21 |
22 |
21 |
24 |
25 |
26 |
27 |
22 |
29 |
10 |
|
|
GRAINES FERTILES |
27 |
25 |
29 |
38 |
24 |
4 |
51 |
20 |
24 |
6 |
27 |
12 |
52 |
25 |
47 |
10 |
33 |
41 |
53 |
38 |
10 |
43 |
12 |
17 |
20 |
47 |
15 |
10 |
9 |
11 |
|
|
GRAINES |
ATTAQUEES |
13 |
12 |
18 |
19 |
8 |
16 |
9 |
3 |
6 |
14 |
8 |
3 |
10 |
9 |
15 |
6 |
3 |
5 |
5 |
19 |
5 |
13 |
3 |
5 |
11 |
15 |
7 |
3 |
8 |
6 |
|
APLATIES |
57 |
49 |
80 |
26 |
34 |
55 |
25 |
79 |
97 |
58 |
98 |
92 |
38 |
30 |
16 |
97 |
54 |
41 |
27 |
26 |
17 |
53 |
92 |
54 |
21 |
16 |
19 |
68 |
28 |
66 |
|
|
AIGUILLES |
16 |
21 |
17 |
18 |
14 |
58 |
2 |
4 |
2 |
54 |
2 |
2 |
23 |
5 |
12 |
2 |
20 |
6 |
13 |
18 |
4 |
22 |
2 |
14 |
12 |
12 |
15 |
9 |
27 |
21 |
|
|
GRAINES VAINES |
83 |
82 |
115 |
63 |
56 |
129 |
36 |
86 |
105 |
126 |
108 |
97 |
71 |
54 |
43 |
105 |
77 |
52 |
45 |
63 |
26 |
88 |
97 |
73 |
44 |
43 |
44 |
80 |
63 |
93 |
|
|
GRAINES TOTALES PAR CÔNE |
110 |
107 |
144 |
101 |
80 |
133 |
87 |
106 |
129 |
132 |
135 |
109 |
123 |
79 |
90 |
115 |
110 |
93 |
98 |
101 |
36 |
131 |
109 |
90 |
64 |
90 |
56 |
90 |
72 |
104 |
|
Année de forte production
|
Canes N° |
1 |
2 |
1 |
4 |
5 |
6 |
7 |
2 |
9 |
10 |
11 |
12 |
11 |
14 |
15 |
16 |
17 |
12 |
19 |
20 |
21 |
22 |
21 |
24 |
25 |
26 |
27 |
22 |
29 |
10 |
|
|
GRAINES FERTILES |
60 |
105 |
70 |
74 |
108 |
29 |
71 |
33 |
53 |
114 |
81 |
50 |
55 |
26 |
67 |
36 |
103 |
36 |
2 |
101 |
100 |
88 |
99 |
62 |
71 |
40 |
99 |
106 |
54 |
67 |
|
|
GRAINES |
ATTAQUEES |
23 |
25 |
7 |
9 |
2 |
7 |
13 |
26 |
18 |
9 |
14 |
61 |
13 |
10 |
14 |
18 |
7 |
10 |
70 |
10 |
7 |
7 |
15 |
10 |
8 |
10 |
9 |
14 |
20 |
21 |
|
APLATIES |
29 |
24 |
42 |
50 |
31 |
110 |
37 |
51 |
43 |
45 |
39 |
42 |
38 |
97 |
5 |
79 |
37 |
30 |
8 |
14 |
43 |
64 |
31 |
24 |
69 |
44 |
38 |
23 |
75 |
76 |
|
|
AIGUILLES |
26 |
12 |
18 |
8 |
14 |
15 |
28 |
30 |
9 |
7 |
20 |
61 |
22 |
17 |
56 |
4 |
16 |
21 |
32 |
25 |
15 |
11 |
14 |
25 |
112 |
26 |
18 |
22 |
23 |
16 |
|
|
GRAINES VAINES |
78 |
61 |
67 |
67 |
47 |
132 |
78 |
107 |
70 |
61 |
73 |
164 |
73 |
124 |
80 |
101 |
60 |
61 |
110 |
49 |
65 |
82 |
60 |
59 |
88 |
80 |
65 |
59 |
118 |
||
|
GRAINES TOTALES PAR CÔNE |
138 |
166 |
137 |
141 |
155 |
161 |
149 |
140 |
123 |
175 |
154 |
214 |
128 |
150 |
147 |
137 |
163 |
97 |
112 |
150 |
165 |
170 |
159 |
121 |
159 |
120 |
164 |
165 |
172 |
180 |
|
Exuosition Nord
|
ARBRE N° 1 |
ARBRE N° 2 |
ARBRE N°3 |
ARBRE N°4 |
ARBRE N°5 |
|||||||||||||||||||||||||||
|
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
||||||||||||||||||||||
|
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
||
|
Poids .g |
33.89 |
42.23 |
42.35 |
52.02 |
46.80 |
53.01 |
54.69 |
37.52 |
40.97 |
60.03 |
55.10 |
57.06 |
38.36 |
22.05 |
41.55 |
37.25 |
37.45 |
37.60 |
36.04 |
17.04 |
28.83 |
17.70 |
23.23 |
22.26 |
43.33 |
44.73 |
50.33 |
68.46 |
59.63 |
62.42 |
|
|
Longueur cm |
4.83 |
4.93 |
5.12 |
5.83 |
5.75 |
5.86 |
6.35 |
5.44 |
8.61 |
6.45 |
6.24 |
6.38 |
4.90 |
4.20 |
5.42 |
5.24 |
5.40 |
5.47 |
5.39 |
3.61 |
4.85 |
4.24 |
4.69 |
4.35 |
5.72 |
5.82 |
6.44 |
7.22 |
6.83 |
6.91 |
|
|
Diamètre cm |
4.11 |
4.08 |
4.50 |
4.59 |
4.40 |
4.61 |
4.74 |
4.27 |
4.29 |
4.98 |
4.85 |
4.88 |
4.41 |
3.58 |
4.42 |
4.34 |
4.14 |
4.17 |
4.05 |
3.39 |
4.12 |
3.36 |
3.72 |
4.59 |
4.47 |
4.43 |
4.64 |
5.01 |
4.82 |
4.76 |
|
|
Graines |
Fertiles |
27 |
25 |
43 |
50 |
58 |
52 |
50 |
33 |
56 |
81 |
75 |
77 |
56 |
29 |
58 |
47 |
64 |
38 |
24 |
10 |
10 |
20 |
4 |
4 |
41 |
38 |
54 |
86 |
51 |
94 |
|
Attaquées |
13 |
12 |
13 |
8 |
6 |
8 |
3 |
3 |
3 |
11 |
6 |
9 |
8 |
18 |
10 |
9 |
9 |
19 |
8 |
3 |
5 |
11 |
16 |
18 |
5 |
4 |
5 |
3 |
9 |
10 |
|
|
A R B R E N° 6 |
A R B R E N° 7 |
A R B R E N°8 |
A R B R E N°9 |
A R B R E N°10 |
|||||||||||||||||||||||||||
|
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
||||||||||||||||||||||
|
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
||
|
Poids .g |
29.85 |
37.48 |
41.46 |
37.53 |
32.54 |
34.66 |
77.85 |
52.95 |
74.85 |
35.32 |
52.08 |
79.87 |
65.63 |
56.37 |
54.78 |
49.12 |
55.94 |
56.72 |
32.72 |
49.38 |
45.26 |
65.34 |
46.82 |
48.75 |
68.65 |
66.89 |
63.32 |
65.47 |
65.45 |
72.48 |
|
|
Longueur cm |
4.21 |
4.81 |
5.45 |
4.20 |
4.97 |
4.15 |
7.30 |
5.98 |
7.08 |
5.45 |
6.28 |
7.64 |
7.12 |
6.58 |
6.40 |
6.05 |
6.49 |
6.52 |
6.02 |
6.85 |
6.11 |
7.8 |
7.11 |
7.51 |
7.90 |
6.82 |
6.09 |
7.19 |
7.24 |
7.33 |
|
|
Diamètre cm |
4.12 |
4.12 |
4.35 |
4.39 |
4.13 |
4.15 |
5.60 |
5.13 |
5.62 |
4.45 |
5.10 |
5.56 |
4.98 |
4.82 |
4.71 |
4.55 |
4.79 |
4.77 |
3.35 |
4.35 |
4.31 |
4.65 |
4.3 |
4.38 |
5.04 |
5.03 |
4.99 |
4.92 |
4.90 |
5.02 |
|
|
Graines |
Fertiles |
20 |
10 |
24 |
6 |
27 |
12 |
51 |
48 |
73 |
61 |
78 |
83 |
83 |
52 |
92 |
79 |
92 |
89 |
53 |
71 |
35 |
86 |
57 |
47 |
94 |
56 |
76 |
123 |
100 |
116 |
|
Attaquées |
3 |
6 |
6 |
5 |
8 |
3 |
16 |
17 |
12 |
15 |
19 |
21 |
22 |
12 |
11 |
7 |
14 |
17 |
5 |
4 |
9 |
15 |
12 |
15 |
5 |
3 |
4 |
7 |
n |
3 |
|
|
ARBRE N° 11 |
ARBRE N° 12 |
ARBRE N°13 |
ARBRE N°14 |
ARBRE N°15 |
|||||||||||||||||||||||||||
|
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
||||||||||||||||||||||
|
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
||
|
Poids .g |
41.14 |
40.06 |
50.43 |
39.67 |
63.62 |
60.10 |
36.21 |
22.08 |
23.77 |
42.01 |
40.36 |
31.69 |
42.63 |
39.88 |
44.36 |
45.53 |
66.98 |
55.32 |
41.02 |
40.30 |
36.10 |
55.22 |
36.60 |
42.88 |
28.29 |
40.49 |
18.59 |
37.63 |
41.98 |
38.13 |
|
|
Longueur cm |
4.81 |
4.95 |
6.46 |
4.98 |
7.03 |
6.83 |
4.61 |
4.59 |
4.49 |
5.21 |
5.28 |
5.31 |
5.71 |
5.68 |
5.72 |
4.78 |
6.08 |
6.01 |
5.69 |
5.58 |
4.82 |
6.24 |
4.90 |
5.76 |
4.79 |
5.39 |
3.57 |
5.38 |
5.61 |
5.50 |
|
|
Diamètre cm |
4.01 |
4.11 |
4.60 |
4.40 |
4.86 |
4.68 |
3.81 |
3.98 |
3.76 |
4.51 |
4.49 |
4.36 |
4.44 |
4.41 |
4.47 |
4.44 |
4.94 |
4.96 |
4.49 |
4.45 |
4.38 |
5.02 |
4.39 |
4.48 |
4.36 |
4.62 |
3.21 |
4.64 |
4.71 |
4.65 |
|
|
Graines |
Fertiles |
56 |
53 |
47 |
42 |
82 |
58 |
22 |
10 |
12 |
42 |
31 |
26 |
40 |
36 |
51 |
43 |
82 |
58 |
48 |
40 |
29 |
52 |
31 |
59 |
22 |
23 |
18 |
26 |
31 |
18 |
|
Attaquées |
1 |
1 |
2 |
6 |
8 |
10 |
1 |
1 |
0 |
1 |
2 |
3 |
1 |
2 |
1 |
3 |
4 |
6 |
4 |
2 |
1 |
8 |
12 |
16 |
6 |
4 |
3 |
12 |
16 |
13 |
|
Exuosition Sud
|
ARBRE N° 1 |
ARBRE N° 2 |
ARBRE N°3 |
ARBRE N°4 |
ARBRE N°5 |
|||||||||||||||||||||||||||
|
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
||||||||||||||||||||||
|
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
||
|
Poids .g |
38.85 |
44.98 |
38.86 |
50.50 |
53.78 |
51.33 |
34.00 |
32.49 |
49.39 |
38.34 |
40.54 |
42.06 |
49.98 |
49.94 |
35.65 |
46.39 |
46.07 |
49.05 |
57.51 |
67.74 |
59.83 |
70.19 |
63.31 |
58.83 |
65.48 |
58.81 |
59.48 |
63.52 |
44.84 |
71.88 |
|
|
Longueur cm |
5.32 |
5.93 |
5.47 |
6.24 |
6.36 |
6.35 |
4.54 |
4.49 |
4.75 |
5.14 |
5.08 |
4.76 |
6.04 |
6.05 |
5.27 |
6.30 |
6.01 |
6.27 |
6.21 |
6.05 |
6.23 |
6.60 |
6.53 |
5.79 |
6.31 |
6.18 |
6.11 |
5.84 |
5.80 |
6.62 |
|
|
Diamètre cm |
4.08 |
4.48 |
4.32 |
4.60 |
4.65 |
4.50 |
4.32 |
4.20 |
4.52 |
4.69 |
4.54 |
4.66 |
4.75 |
4.70 |
4.10 |
4.58 |
4.47 |
4.44 |
4.89 |
5.22 |
4.60 |
5.10 |
5.17 |
5 |
5.11 |
4.88 |
5.98 |
5.02 |
4.55 |
5.29 |
|
|
Graines |
Fertiles |
7 |
25 |
9 |
43 |
29 |
31 |
9 |
3 |
8 |
4 |
9 |
13 |
56 |
87 |
77 |
26 |
33 |
18 |
36 |
33 |
48 |
86 |
56 |
10 |
16 |
53 |
62 |
37 |
17 |
64 |
|
Attaquées |
2 |
3 |
4 |
5 |
3 |
11 |
6 |
2 |
3 |
3 |
3 |
7 |
25 |
22 |
39 |
36 |
32 |
34 |
11 |
6 |
5 |
15 |
7 |
5 |
9 |
16 |
9 |
6 |
6 |
10 |
|
|
A R B R E N° 6 |
A R B R E N° 7 |
A R B R E N°8 |
A R B R E N°9 |
B RARE N°10 |
|||||||||||||||||||||||||||
|
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
||||||||||||||||||||||
|
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
||
|
Poids .g |
37.44 |
33.64 |
30.63 |
33.49 |
36.91 |
27.81 |
63.10 |
50.94 |
42.26 |
55.64 |
42.43 |
56.37 |
19.78 |
22.57 |
23.22 |
21.21 |
26.01 |
33.85 |
55.44 |
58.29 |
53.31 |
57.72 |
63.17 |
67.17 |
45.72 |
43.80 |
41.11 |
47.45 |
54.85 |
51.96 |
|
|
Longueur cm |
5.52 |
4.49 |
4.87 |
5.43 |
5.20 |
5.06 |
7.15 |
6.27 |
6.20 |
6.51 |
6.18 |
6.55 |
4.21 |
4.22 |
4.81 |
4.61 |
4.78 |
5.06 |
6.38 |
6.60 |
6.22 |
7.91 |
6.57 |
6.36 |
6.46 |
6.40 |
6.22 |
6.65 |
7.47 |
7.34 |
|
|
Diamètre cm |
4.34 |
4.29 |
4.07 |
4.15 |
4.02 |
3.84 |
4.94 |
4.10 |
4.52 |
4.83 |
4.90 |
4.70 |
3.44 |
3.55 |
3.20 |
3.38 |
3.68 |
4.02 |
4.73 |
4.78 |
4.68 |
5.52 |
4.83 |
4.98 |
4.44 |
4.31 |
4.36 |
438 |
4.56 |
4.40 |
|
|
Graines |
Fertiles |
31 |
25 |
25 |
50 |
33 |
15 |
61 |
44 |
30 |
53 |
64 |
41 |
5 |
19 |
4 |
18 |
28 |
36 |
30 |
6 |
56 |
3 |
14 |
90 |
69 |
57 |
65 |
65 |
65 |
69 |
|
Attaquées |
17 |
4 |
10 |
8 |
7 |
11 |
13 |
19 |
16 |
17 |
16 |
19 |
5 |
4 |
0 |
3 |
12 |
13 |
6 |
12 |
9 |
2 |
2 |
13 |
15 |
12 |
12 |
24 |
24 |
14 |
|
|
ARBRE N° 11 |
ARBRE N° 12 |
ARBRE N°13 |
ARBRE N°14 |
ARBRE N° 15 |
|||||||||||||||||||||||||||
|
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
Branche Basse |
Branche Haute |
||||||||||||||||||||||
|
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
1 |
2 |
3 |
||
|
Poids .g |
38.34 |
38.07 |
34.02 |
32.38 |
34.20 |
31.18 |
17.81 |
21.61 |
22.04 |
25.03 |
23.17 |
18.81 |
41.26 |
40.32 |
42.21 |
35.59 |
32.56 |
29.51 |
27.19 |
28.43 |
36.91 |
17.86 |
22.33 |
23.71 |
46.73 |
48.36 |
37.21 |
37.21 |
38.63 |
43.1 |
|
|
Longueur cm |
5.53 |
5.52 |
5.47 |
4.62 |
4.64 |
4.58 |
4.11 |
4.46 |
4.79 |
4.38 |
4.80 |
4.25 |
5.02 |
4.76 |
4.98 |
4.56 |
4.47 |
4.36 |
4.16 |
4.18 |
5.12 |
4.11 |
4.22 |
4.21 |
5.89 |
6.01 |
5.21 |
5.01 |
5.36 |
5.26 |
|
|
Diamètre cm |
4.09 |
4.11 |
4.01 |
4.00 |
4.02 |
3.98 |
3.38 |
3.42 |
3.41 |
3.55 |
3.32 |
3.28 |
4.61 |
4.62 |
4.67 |
4.33 |
4.22 |
3.88 |
3.76 |
3.73 |
4.04 |
3.42 |
4.02 |
3.96 |
4.72 |
4.83 |
4.90 |
4.66 |
4.73 |
4.86 |
|
|
Graines |
Fertiles |
17 |
13 |
21 |
4 |
9 |
8 |
6 |
4 |
2 |
7 |
11 |
3 |
27 |
26 |
22 |
11 |
8 |
9 |
9 |
21 |
17 |
3 |
1 |
6 |
21 |
15 |
16 |
11 |
16 |
18 |
|
Attaquées |
2 |
2 |
3 |
1 |
2 |
1 |
2 |
5 |
1 |
6 |
3 |
9 |
11 |
8 |
2 |
21 |
14 |
6 |
3 |
1 |
2 |
16 |
9 |
7 |
11 |
3 |
9 |
8 |
2 |
6 |
|
|
EXPOSITION |
STATION |
||||||||
|
NORD |
SUD |
MOYENNE |
|||||||
|
Branches |
Branches |
Mo |
Branches |
Branches |
Mo |
Branches |
Branches |
Mo y |
|
|
10-20 |
0.22 |
04.44 |
03.33 |
0.4.44 |
04.44 |
04.44 |
03.33 |
04.44 |
03.88 |
|
20-30 |
04.44 |
13.33 |
08.88 |
17.77 |
06.66 |
15.55 |
11.11 |
13.33 |
12.22 |
|
30-40 |
26.66 |
20.00 |
23.33 |
24.44 |
28.88 |
26.66 |
25.55 |
24.44 |
25.00 |
|
40-50 |
20.00 |
35.55 |
27.77 |
17.77 |
28.28 |
23.33 |
18.88 |
32.22 |
25.55 |
|
50-60 |
22.22 |
13.33 |
17.77 |
22.22 |
17.77 |
20.00 |
22.22 |
15.55 |
18.88 |
|
60-70 |
20.00 |
08.80 |
14.44 |
04.44 |
06.66 |
07.77 |
14.44 |
07.77 |
11.11 |
|
70-80 |
04.44 |
02.22 |
04.44 |
02.22 |
00.00 |
02.22 |
04.44 |
02.22 |
03.33 |
Distribution des cônes par classe de longueur.
|
EXPOSITION |
STATION |
||||||||
|
NORD |
SUD |
MOYENNE |
|||||||
|
Branches |
Branches |
Mo |
Branches |
Branches |
Mo |
Branches |
Branches |
Mo y |
|
|
30-40 |
00.00 |
04.44 |
02.22 |
00.00 |
00.00 |
00.00 |
00.00 |
02.22 |
01.11 |
|
40-50 |
20.00 |
31.11 |
25.55 |
33.33 |
33.33 |
33.33 |
26.26 |
32.22 |
29.44 |
|
50-60 |
31.11 |
31.11 |
31.31 |
26.26 |
28.28 |
27.77 |
28.88 |
30.00 |
29.44 |
|
60-70 |
28.88 |
22.22 |
25.55 |
33.33 |
35.55 |
34.44 |
31.31 |
28.88 |
30.00 |
|
70-80 |
20.00 |
08.88 |
14.44 |
06.66 |
02.22 |
04.44 |
13.33 |
05.55 |
09.44 |
|
80-90 |
00.00 |
02.22 |
01.11 |
00.00 |
00.00 |
00.00 |
00.00 |
01.00 |
00.55 |
Distribution des cônes par classe de diamètre.
|
EXPOSITION |
STATION |
||||||||
|
NORD |
SUD |
MOYENNE |
|||||||
|
Branches |
Branches |
Mo |
Branches |
Branches |
Mo |
Branches |
Branches |
Mo y |
|
|
30-40 |
06.66 |
15.55 |
11.11 |
23.40 |
17.77 |
20.65 |
12.21 |
16.66 |
14.43 |
|
40-50 |
82.22 |
73.33 |
77.77 |
63.82 |
75.55 |
69.56 |
68.47 |
78.88 |
73.62 |
|
50-60 |
11.11 |
11.11 |
11.11 |
12.76 |
06.66 |
09.78 |
11.95 |
08.88 |
10.43 |
243
|
Versant Nord |
Versant Sud |
|||||||
|
Graines fertiles |
Graines attaquées |
Graines fertiles |
Graines attaquées |
|||||
|
Poids. g |
Long. cm |
Poids. g |
Long. cm |
Poids. g |
Long. cm |
Poids. g |
Long. cm |
|
|
0.0891 |
1.27 |
0.0402 |
1.15 |
0.0684 |
1.14 |
0.0367 |
1.11 |
|
|
0.0939 |
1.22 |
0.0438 |
1.17 |
0.0923 |
1.20 |
0.0537 |
1.20 |
|
|
0.0818 |
1.31 |
0.0403 |
1.14 |
0.0745 |
1.21 |
0.0540 |
1.14 |
|
|
0.0998 |
1.34 |
0.0464 |
1.21 |
0.0931 |
1.16 |
0.0376 |
1.15 |
|
|
0.0891 |
1.22 |
0.0372 |
1.09 |
0.0844 |
1.15 |
0.0443 |
1.14 |
|
|
0.0892 |
1.27 |
0.0490 |
1.24 |
0.0930 |
1.14 |
0.0374 |
1.15 |
|
|
0.0825 |
1.30 |
0.0463 |
1.10 |
0.0668 |
1.15 |
0.0402 |
1.10 |
|
|
0.0745 |
1.24 |
0.0580 |
1.26 |
0.0599 |
1.03 |
0.0451 |
1.13 |
|
|
0.1085 |
1.22 |
0.0480 |
1.02 |
0.0729 |
1.07 |
0.0370 |
1.12 |
|
|
0.0920 |
1.21 |
0.0475 |
1.30 |
0.0850 |
1.23 |
0.0458 |
1.11 |
|
|
0.0951 |
1.21 |
0.0432 |
1.25 |
0.0915 |
1.24 |
0.0540 |
1.13 |
|
|
0.0869 |
1.15 |
0.0431 |
1.24 |
0.0879 |
1.11 |
0.0458 |
1.12 |
|
|
0.0861 |
1.25 |
0.0453 |
1.20 |
0.0830 |
1.12 |
0.0457 |
1.14 |
|
|
0.0958 |
1.28 |
0.0435 |
1.22 |
0.0877 |
1.18 |
0.0405 |
1.11 |
|
|
0.0911 |
1.28 |
0.0392 |
1.13 |
0.0692 |
1.09 |
0.0376 |
1.01 |
|
|
0.0999 |
1.34 |
0.0460 |
1.20 |
0.0953 |
1.16 |
0.0404 |
1.10 |
|
|
0.0782 |
1.23 |
0.0295 |
1.12 |
0.0930 |
1.15 |
0.0271 |
0.92 |
|
|
0.0807 |
1.24 |
0.0491 |
1.20 |
0.0758 |
1.11 |
0.0465 |
1.12 |
|
|
0.0960 |
1.12 |
0.0428 |
1.29 |
0.0891 |
1.11 |
0.0378 |
1.11 |
|
|
0.0809 |
1.22 |
0.0434 |
1.15 |
0.0866 |
1.15 |
0.0396 |
1.08 |
|
|
0.0952 |
1.29 |
0.0398 |
1.16 |
0.0751 |
1.21 |
0.0427 |
1.09 |
|
|
0.0875 |
1.14 |
0.0381 |
1.10 |
0.0860 |
1.10 |
0.0377 |
1.11 |
|
|
0.0731 |
1.13 |
0.0393 |
1.12 |
0.0822 |
1.02 |
0.0373 |
1.13 |
|
|
0.0870 |
1.32 |
0.0478 |
1.13 |
0.0574 |
1.03 |
0.0418 |
1.14 |
|
|
0.0942 |
1.32 |
0.0484 |
1.25 |
0.0926 |
1.18 |
0.0465 |
1.15 |
|
|
0.0923 |
1.26 |
0.0478 |
1.26 |
0.0804 |
1.14 |
0.0514 |
1.22 |
|
|
0.0765 |
1.13 |
0.0429 |
1.24 |
0.0735 |
1.18 |
0.0376 |
1.01 |
|
|
0.0814 |
1.25 |
0.0429 |
1.20 |
0.0804 |
1.12 |
0.0465 |
1.15 |
|
|
0.0886 |
1.24 |
0.0436 |
1.23 |
0.0728 |
1.07 |
0.0425 |
1.16 |
|
|
0.0941 |
1.18 |
0.0501 |
1.23 |
0.0669 |
1.06 |
0.0418 |
1.19 |
|
|
0.0998 |
1.31 |
0.0475 |
1.18 |
0.0950 |
1.32 |
0.0417 |
1.20 |
|
|
0.0731 |
1.22 |
0.0470 |
1.25 |
0.0903 |
1.25 |
0.0391 |
1.06 |
|
|
0.0943 |
1.24 |
0.0402 |
1.15 |
0.0986 |
1.24 |
0.0409 |
1.10 |
|
|
0.0889 |
1.26 |
0.0389 |
1.08 |
0.0875 |
1.21 |
0.0376 |
1.01 |
|
|
0.0887 |
1.30 |
0.0439 |
1.13 |
0.0735 |
1.18 |
0.0540 |
1.21 |
|
|
0.0930 |
1.28 |
0.0452 |
1.26 |
0.1085 |
1.22 |
0.0396 |
1.05 |
|
|
0.0836 |
1.31 |
0.0390 |
1.13 |
0.0836 |
1.11 |
0.0426 |
1.06 |
|
|
0.0836 |
1.34 |
0.0329 |
1.11 |
0.0838 |
1.10 |
0.0477 |
1.16 |
|
|
0.0880 |
1.23 |
0.0424 |
1.31 |
0.0801 |
1.11 |
0.0376 |
1.04 |
|
|
0.0694 |
1.12 |
0.0377 |
1.26 |
0.0732 |
1.08 |
0.0427 |
1.06 |
|
|
0.0769 |
1.21 |
0.0432 |
1.12 |
0.0745 |
1.13 |
0.0470 |
1.19 |
|
|
0.0853 |
1.28 |
0.0358 |
1.12 |
0.1020 |
1.21 |
0.0541 |
1.20 |
|
|
0.0906 |
1.23 |
0.0392 |
1.13 |
0.0783 |
1.13 |
0.0412 |
1.13 |
|
|
0.0905 |
1.25 |
0.0545 |
1.38 |
0.0924 |
1.17 |
0.0465 |
1.11 |
|
|
0.0859 |
1.25 |
0.0591 |
1.28 |
0.0876 |
1.21 |
0.0472 |
1.16 |
|
|
0.0726 |
1.15 |
0.0491 |
1.26 |
0.0924 |
1.15 |
0.0476 |
1.18 |
|
|
0.0756 |
1.11 |
0.0439 |
1.24 |
0.0833 |
1.12 |
0.0395 |
1.09 |
|
|
0.0828 |
1.18 |
0.0377 |
1.26 |
0.0784 |
1.03 |
0.0372 |
1.14 |
|
|
0.0906 |
1.28 |
0.0416 |
1.21 |
0.0916 |
1.15 |
0.0378 |
1.06 |
|
|
0.0864 |
1.30 |
0.0611 |
1.35 |
0.0854 |
1.14 |
0.0429 |
1.04 |
|
|
0.0628 |
1.12 |
0.0406 |
1.12 |
0.0890 |
1.15 |
0.0458 |
1.16 |
|
|
0.0838 |
1.19 |
0.0401 |
1.12 |
0.0892 |
1.18 |
0.0377 |
1.11 |
|
|
0.0930 |
1.29 |
0.0474 |
1.13 |
0.0831 |
1.04 |
0.0476 |
1.11 |
|
|
0.0874 |
1.24 |
0.0436 |
1.12 |
0.0877 |
1.17 |
0.0421 |
1.13 |
|
|
0.0950 |
1.21 |
0.0441 |
1.21 |
0.0905 |
1.11 |
0.0468 |
1.15 |
|
|
0.0773 |
1.11 |
0.0440 |
1.22 |
0.0800 |
1.13 |
0.0456 |
1.12 |
|
|
0.0898 |
1.24 |
0.0405 |
1.17 |
0.0858 |
1.26 |
0.0412 |
1.12 |
|
|
0.0941 |
1.26 |
0.0473 |
1.19 |
0.0750 |
1.18 |
0.0464 |
1.14 |
|
|
0.0908 |
1.28 |
0.0416 |
1.28 |
0.0866 |
1.14 |
0.0459 |
1.13 |
|
|
0.0857 |
1.23 |
0.0332 |
1.11 |
0.0833 |
1.08 |
0.0469 |
1.16 |
|
244
|
Temps |
Poids de 100 graines ( g) |
Moyenne |
||
|
Lot 1 |
Lot 2 |
Lot 3 |
||
|
0 |
7.7321 |
7.9136 |
8.0916 |
7.9124 |
|
1 |
8.8316 |
9.8476 |
9.9178 |
9.5323 |
|
6 |
9.1973 |
10.0494 |
10.1759 |
9.8075 |
|
12 |
9.8213 |
11.0829 |
10.5182 |
10.4741 |
|
24 |
10.8333 |
11.1755 |
11.4682 |
11.1590 |
|
48 |
11.2393 |
11.5807 |
11.9140 |
11.5780 |
|
72 |
11.5115 |
12.1196 |
12.6358 |
12.0889 |
|
96 |
11.8463 |
12.3650 |
12.9279 |
12.3797 |
|
120 |
11.8525 |
12.3752 |
12.9328 |
12.3868 |
Annexe XI: Influence du type de germoir sur la germination des graines de cèdre.
|
Jours |
Germoir BENDER et HOBEIN. |
Boîtes germoir. |
||
|
C.G. (%) |
T.M.G. (Jours) |
C.G. (%) |
T.M.G. (Jours) |
|
|
0 |
0 |
- |
0 |
- |
|
3 |
14 |
- |
11 |
- |
|
6 |
23 |
- |
20 |
- |
|
9 |
47 |
- |
43 |
- |
|
12 |
62 |
- |
57 |
- |
|
15 |
73 |
- |
66 |
- |
|
18 |
84 |
- |
81 |
- |
|
21 |
86 |
10.43 |
82 |
10.82 |
|
Jours |
Versant Nord |
Versant Sud |
||
|
C.G. (%) |
T.M.G. (Jours) |
C.G. (%) |
T.M.G. (Jours) |
|
|
0 |
0 |
- |
0 |
- |
|
3 |
23 |
- |
17 |
- |
|
6 |
40 |
- |
27 |
- |
|
9 |
63 |
- |
43 |
- |
|
12 |
73 |
- |
70 |
- |
|
15 |
80 |
- |
73 |
- |
|
18 |
83 |
- |
77 |
- |
|
21 |
87 |
08.52 |
80 |
09.49 |
|
Jours |
Branche haute |
Branche basse |
||
|
C.G. (%) |
T.M.G. (Jours) |
C.G. (%) |
T.M.G. (Jours) |
|
|
0 |
0 |
- |
0 |
- |
|
3 |
28 |
- |
39 |
- |
|
6 |
44 |
- |
48 |
- |
|
9 |
52 |
- |
61 |
- |
|
12 |
59 |
- |
68 |
- |
|
15 |
68 |
- |
75 |
- |
|
18 |
74 |
- |
81 |
- |
|
21 |
80 |
08.81 |
92 |
08.87 |
245
|
Jours |
Classes de poids |
|||||
|
0.04 - 0.0699 |
0.0700 - 0.0999 |
Supérieure à 0.1000 |
||||
|
C.G. (%) |
T.M.G. (Jours) |
C.G. (%) |
T.M.G. (Jours) |
C.G. (%) |
T.M.G. (Jours) |
|
|
0 |
0 |
- |
0 |
- |
0 |
- |
|
3 |
0 |
- |
13 |
- |
27 |
- |
|
6 |
10 |
- |
23 |
- |
36 |
- |
|
9 |
33 |
- |
40 |
- |
53 |
- |
|
12 |
40 |
- |
63 |
- |
70 |
- |
|
15 |
47 |
- |
73 |
- |
83 |
- |
|
18 |
53 |
- |
80 |
- |
93 |
- |
|
21 |
63 |
12.28 |
83 |
10.44 |
96 |
9.68 |
|
Jours |
Témoin : Graines sans ailes avec téguments. |
Graines sans ailes et sans téguments. |
Graines avec ailes et téguments. |
|||
|
C.G.(%) |
T.M.G. (Jours) |
C.G.(%) |
T.M.G. (Jours) |
C.G.(%) |
T.M.G. (Jours) |
|
|
0 |
0 |
- |
0 |
- |
0 |
- |
|
3 |
27 |
- |
30 |
- |
17 |
- |
|
6 |
63 |
- |
70 |
- |
60 |
- |
|
9 |
73 |
- |
76 |
- |
73 |
- |
|
12 |
80 |
- |
83 |
- |
80 |
- |
|
15 |
90 |
- |
93 |
- |
87 |
- |
|
18 |
97 |
- |
97 |
- |
90 |
- |
|
21 |
97 |
07.70 |
97 |
07.11 |
93 |
07.87 |
|
Jours |
Témoin |
Eau oxygénée |
Durée du prétraitement (stratification). |
|||||||||||
|
15 % - 45 mn. |
30% - 45 mn |
15 |
ours |
30 |
jours |
45 |
jours |
|||||||
|
C.G. |
T.M.G. |
C.G. |
T.M.G |
C.G. |
T.M.G |
C.G. |
T.M.G |
C.G. |
T.M.G. |
C.G. |
T.M.G. |
|||
|
0 |
0 |
0 |
0 |
0 |
0 |
- |
0 |
- |
||||||
|
3 |
0 |
- |
20 |
- |
12 |
- |
23 |
- |
47 |
- |
53 |
- |
||
|
6 |
7 |
- |
36 |
- |
28 |
- |
42 |
- |
63 |
- |
76 |
- |
||
|
9 |
13 |
- |
50 |
- |
31 |
- |
50 |
- |
70 |
- |
76 |
- |
||
|
12 |
27 |
- |
53 |
- |
40 |
- |
57 |
- |
76 |
- |
80 |
- |
||
|
15 |
30 |
- |
57 |
- |
48 |
- |
60 |
- |
80 |
- |
90 |
- |
||
|
18 |
36 |
- |
63 |
- |
54 |
- |
63 |
- |
87 |
- |
96 |
- |
||
|
21 |
37 |
11.84 |
70 |
9,40 |
55 |
9.38 |
73 |
08.87 |
87 |
06.41 |
96 |
06.56 |
||
|
Jours |
02Septembre |
03Octobre |
02Novembre |
05Décembre |
01Janvier |
|||||
|
C.G. |
T.M.G. |
C.G. |
T.M.G. |
C.G. (%). |
T.M.G. |
C.G. |
T.M.G. |
C.G. |
T.M.G. |
|
|
0 |
0 |
- |
0 |
- |
0 |
0 |
0 |
- |
||
|
3 |
16 |
- |
20 |
- |
13 |
- |
23 |
- |
20 |
- |
|
6 |
34 |
- |
30 |
- |
40 |
- |
53 |
- |
37 |
- |
|
9 |
53 |
- |
47 |
- |
53 |
- |
57 |
- |
63 |
- |
|
12 |
67 |
- |
60 |
- |
70 |
- |
80 |
- |
77 |
- |
|
15 |
70 |
- |
73 |
- |
80 |
- |
83 |
- |
83 |
- |
|
18 |
73 |
- |
77 |
- |
83 |
- |
87 |
- |
90 |
- |
|
21 |
74 |
08.31 |
80 |
09.48 |
86 |
09.17 |
90 |
08.23 |
94 |
09.19 |
|
Jours |
02Septembre |
03Octobre |
02Novembre |
05Décembre |
01Janvier |
|||||
|
C.G. |
T.M.G. |
C.G. |
T.M.G. |
C.G. |
T.M.G. |
C.G |
T.M.G. |
C.G. |
T.M.G. |
|
|
0 |
0 |
- |
0 |
- |
0 |
- |
0 |
- |
0 |
- |
|
3 |
0 |
- |
0 |
- |
3 |
- |
13 |
- |
23 |
- |
|
6 |
0 |
- |
3 |
- |
7 |
- |
20 |
- |
34 |
- |
|
9 |
3 |
- |
3 |
- |
13 |
- |
43 |
- |
60 |
- |
|
12 |
7 |
- |
7 |
- |
23 |
- |
57 |
- |
73 |
- |
|
15 |
10 |
- |
13 |
- |
34 |
- |
64 |
- |
77 |
- |
|
18 |
10 |
- |
13 |
- |
43 |
- |
67 |
- |
83 |
- |
|
21 |
10 |
12.00 |
13 |
12.00 |
50 |
13.62 |
70 |
09.68 |
91 |
09.46 |
|
Jours |
Témoin |
Hors cône. |
Dans les cônes. |
|||||||||||
|
06 mois |
12 mois |
24 mois |
06 mois |
12 mois |
24 mois |
|||||||||
|
C.G. |
T.M.G. |
C.G. |
T.M.G |
C.G. |
T.M.G |
C.G. |
T.M.G. |
C.G. |
T.M.G |
C.G. |
T.M.G |
C.G. |
T.M.G. |
|
|
0 |
0 |
- |
0 |
- |
0 |
0 |
0 |
- |
0 |
- |
0 |
- |
||
|
3 |
20 |
- |
10 |
- |
0 |
- |
0 |
- |
10 |
- |
0 |
- |
0 |
- |
|
6 |
33 |
- |
27 |
- |
3 |
- |
0 |
- |
23 |
- |
0 |
- |
0 |
- |
|
9 |
40 |
- |
37 |
- |
7 |
- |
3 |
- |
32 |
- |
0 |
- |
0 |
- |
|
12 |
44 |
- |
43 |
- |
13 |
- |
6 |
- |
40 |
- |
4 |
- |
0 |
- |
|
15 |
57 |
- |
52 |
- |
20 |
- |
7 |
- |
47 |
- |
4 |
- |
0 |
- |
|
18 |
60 |
- |
56 |
- |
23 |
- |
10 |
- |
50 |
- |
6 |
- |
3 |
- |
|
21 |
60 |
08.30 |
56 |
08.94 |
23 |
12.39 |
10 |
13.20 |
54 |
09.77 |
7 |
15.00 |
4 |
18.75 |
246
Exposition Nord
|
MOIS |
A LTITUDE<1.700m |
ALTITUDE 1.700-1800m |
||||||||||
|
Sous Couvert |
A Découvert |
Sous Couvert |
A Découvert |
|||||||||
|
Témoin |
Travail du sol |
Témoin |
Travail du sol |
Témoin |
Travail du sol |
Témoin |
Travail du sol |
|||||
|
Superficiel |
Profond |
Superficiel |
Profond |
Superficiel |
Profond |
Superficiel |
Profond |
|||||
|
Mars |
46 |
76 |
104 |
45 |
87 |
129 |
58 |
77 |
120 |
56 |
111 |
212 |
|
Avril |
49 |
79 |
112 |
45 |
88 |
134 |
62 |
79 |
129 |
57 |
120 |
223 |
|
Mai |
46 |
78 |
110 |
43 |
84 |
132 |
63 |
83 |
133 |
57 |
121 |
224 |
|
Juin |
41 |
72 |
102 |
38 |
72 |
121 |
58 |
80 |
126 |
51 |
100 |
205 |
|
Juillet |
31 |
66 |
88 |
29 |
66 |
98 |
37 |
62 |
101 |
43 |
81 |
168 |
|
Août |
22 |
41 |
61 |
23 |
39 |
77 |
34 |
48 |
83 |
31 |
57 |
132 |
|
Septembre |
18 |
29 |
53 |
19 |
34 |
61 |
29 |
39 |
61 |
25 |
49 |
112 |
|
Octobre |
16 |
22 |
46 |
16 |
29 |
54 |
26 |
29 |
55 |
19 |
44 |
92 |
|
Novembre |
15 |
21 |
46 |
16 |
29 |
54 |
26 |
29 |
55 |
19 |
44 |
87 |
|
Décembre |
15 |
21 |
46 |
16 |
29 |
54 |
23 |
28 |
51 |
18 |
42 |
87 |
Exposition Sud
|
MOIS |
A LTITUDE<1.700m |
ALTITUDE 1.700-1800m |
||||||||||
|
Sous Couvert |
A Découvert |
Sous Couvert |
A Découvert |
|||||||||
|
Témoin |
Travail du sol |
Témoin |
Travail du sol |
Témoin |
Travail du sol |
Témoin |
Travail du sol |
|||||
|
Superficiel |
Profond |
Superficiel |
Profond |
Superficiel |
Profond |
Superficiel |
Profond |
|||||
|
Mars |
10 |
17 |
31 |
9 |
19 |
24 |
28 |
28 |
37 |
11 |
22 |
35 |
|
Avril |
10 |
17 |
31 |
9 |
19 |
25 |
28 |
29 |
54 |
13 |
23 |
49 |
|
Mai |
10 |
15 |
29 |
9 |
19 |
25 |
25 |
26 |
36 |
8 |
19 |
28 |
|
Juin |
8 |
13 |
27 |
7 |
16 |
21 |
20 |
26 |
35 |
7 |
13 |
22 |
|
Juillet |
2 |
9 |
17 |
0 |
2 |
6 |
9 |
13 |
15 |
0 |
5 |
8 |
|
Août |
0 |
6 |
11 |
0 |
0 |
0 |
2 |
8 |
12 |
0 |
3 |
4 |
|
Septembre |
0 |
5 |
9 |
0 |
0 |
0 |
2 |
8 |
12 |
0 |
2 |
3 |
|
Octobre |
0 |
3 |
9 |
0 |
0 |
0 |
2 |
8 |
12 |
0 |
0 |
3 |
|
Novembre |
0 |
2 |
9 |
0 |
0 |
0 |
2 |
8 |
12 |
0 |
0 |
3 |
|
Décembre |
0 |
2 |
9 |
0 |
0 |
0 |
2 |
8 |
12 |
0 |
0 |
3 |
Exposition Nord
|
Mois |
A L T I T UDE<1.700m |
ALTITUDE 1.700-1800m |
|||||||||||
|
Sous Couvert |
A Découvert |
Sous Couvert |
A Découvert |
||||||||||
|
Témoin |
Travail du sol |
Témoin |
Travail du sol |
Témoin |
Travail du sol |
Témoin |
Travail du sol |
||||||
|
Superficiel |
Profond |
Superficiel |
Profond |
Superficiel |
Profond |
Superficie 1 |
Profond |
||||||
|
Mars |
46 |
76 |
104 |
45 |
87 |
129 |
58 |
77 |
120 |
56 |
111 |
212 |
|
|
Avril |
49 |
79 |
112 |
45 |
88 |
134 |
62 |
79 |
129 |
57 |
120 |
223 |
|
|
Sept |
18 |
29 |
53 |
19 |
34 |
61 |
29 |
39 |
61 |
25 |
49 |
112 |
|
|
Dispams |
9 |
13 |
10 |
7 |
22 |
3 |
24 |
21 |
34 |
26 |
39 |
55 |
|
|
Mort |
22 |
37 |
49 |
15 |
28 |
54 |
9 |
23 |
38 |
12 |
33 |
57 |
|
|
Jaunissement |
3 |
8 |
7 |
7 |
9 |
23 |
7 |
11 |
10 |
1 |
7 |
25 |
|
|
Sains |
15 |
21 |
46 |
16 |
29 |
54 |
23 |
28 |
51 |
18 |
42 |
87 |
|
Exposition Sud
|
Mois |
A L T I T UDE<1.700m |
ALTITUDE 1.700-1800m |
|||||||||||
|
Sous Couvert |
A Découvert |
Sous Couvert |
A Découvert |
||||||||||
|
Témoin |
Travail du sol |
Témoin |
Travail du sol |
Témoin |
Travail du sol |
Témoin |
Travail du sol |
||||||
|
Superficiel |
Profond |
Superficiel |
Profond |
Superficiel |
Profond |
Superficiel |
Profond |
||||||
|
Mars |
10 |
17 |
31 |
9 |
19 |
24 |
28 |
28 |
37 |
11 |
22 |
35 |
|
|
Avril |
10 |
17 |
31 |
9 |
19 |
25 |
28 |
29 |
54 |
13 |
23 |
49 |
|
|
Sept |
0 |
5 |
9 |
0 |
0 |
0 |
2 |
8 |
12 |
0 |
2 |
3 |
|
|
Dispams |
0 |
9 |
8 |
6 |
15 |
7 |
21 |
10 |
21 |
0 |
8 |
29 |
|
|
Mort |
0 |
6 |
12 |
2 |
3 |
16 |
3 |
9 |
16 |
11 |
15 |
15 |
|
|
Jaunissement |
0 |
0 |
2 |
1 |
1 |
2 |
2 |
2 |
5 |
2 |
0 |
2 |
|
|
Sains |
0 |
2 |
9 |
0 |
0 |
0 |
2 |
8 |
12 |
0 |
0 |
3 |
|
247
Annexe XXII : Caractères morphologiques des semis de cèdre de l'Atlas.
Froid humide
|
Plants |
Poids en mg |
Longueur en cm |
Nombre de |
|||||||
|
Frais |
Sec |
|||||||||
|
Total |
Aérienne |
Racinaire |
Total |
Aérienne |
Racinaire |
Total |
Aérienne |
Racinaire |
||
|
1 |
643.5 |
451.70 |
191.8 |
145.7 |
101.3 |
44.40 |
25.49 |
6.14 |
19.35 |
44 |
|
2 |
206.7 |
165.20 |
41.5 |
41.6 |
27.7 |
13.90 |
11.61 |
5.08 |
06.53 |
23 |
|
3 |
593.1 |
431.50 |
161.6 |
173.3 |
162.9 |
10.40 |
23.30 |
6.11 |
17.19 |
39 |
|
4 |
461.7 |
391.10 |
70.6 |
132.0 |
93.7 |
38.30 |
21.36 |
5.24 |
16.12 |
33 |
|
5 |
789.8 |
501.60 |
288.2 |
195.4 |
131.6 |
63.80 |
23.85 |
6.27 |
17.58 |
48 |
|
6 |
299.0 |
250.90 |
48.1 |
64.8 |
55.3 |
9.50 |
10.31 |
4.34 |
05.97 |
31 |
|
7 |
480.7 |
370.10 |
110.6 |
172.0 |
115.1 |
56.90 |
23.73 |
6.12 |
17.61 |
43 |
|
8 |
242.6 |
202.10 |
40.5 |
60.6 |
48.9 |
11.70 |
9.94 |
4.91 |
05.03 |
30 |
|
9 |
270.1 |
226.70 |
43.4 |
69.8 |
59.6 |
10.20 |
10.83 |
4.86 |
05.97 |
27 |
|
10 |
469.6 |
321.10 |
148.5 |
132.3 |
81.9 |
50.40 |
24.42 |
6.13 |
18.29 |
37 |
|
11 |
331.2 |
280.30 |
50.9 |
101.9 |
57.6 |
44.30 |
17.26 |
5.95 |
11.31 |
29 |
|
12 |
641.6 |
480.61 |
161.0 |
190.6 |
137.3 |
53.30 |
26.10 |
6.22 |
19.88 |
43 |
|
13 |
761.3 |
550.90 |
210.4 |
250.8 |
171.3 |
79.50 |
27.42 |
6.23 |
21.19 |
51 |
|
14 |
519.9 |
401.30 |
118.6 |
183.7 |
134.6 |
49.10 |
21.91 |
5.44 |
16.47 |
45 |
|
15 |
491.5 |
330.60 |
160.9 |
130.8 |
95.0 |
35.80 |
18.26 |
5.16 |
13.10 |
35 |
|
16 |
293.6 |
183.20 |
110.4 |
46.99 |
39.9 |
07.09 |
16.96 |
5.53 |
11.43 |
31 |
|
17 |
561.1 |
341.70 |
219.4 |
110.6 |
20.6 |
90.00 |
24.51 |
7.16 |
17.35 |
45 |
|
18 |
442.3 |
330.10 |
112.2 |
69.9 |
40.1 |
29.80 |
20.11 |
5.90 |
14.21 |
37 |
|
19 |
481.1 |
349.50 |
131.6 |
120.2 |
97.3 |
22.90 |
20.13 |
5.77 |
14.36 |
35 |
|
20 |
496.8 |
381.20 |
115.6 |
130.7 |
93.5 |
37.20 |
18.50 |
5.28 |
13.22 |
37 |
|
21 |
331.1 |
266.30 |
64.8 |
100.8 |
74.5 |
26.30 |
14.83 |
5.29 |
09.54 |
33 |
|
22 |
480.8 |
360.70 |
120.1 |
141.6 |
92.2 |
49.40 |
22.59 |
6.10 |
16.49 |
36 |
|
23 |
590.9 |
427.60 |
163.3 |
188.4 |
133.6 |
54.80 |
22.55 |
5.61 |
16.94 |
44 |
|
24 |
435.6 |
288.30 |
147.3 |
76.1 |
33.5 |
42.60 |
20.52 |
6.19 |
14.33 |
38 |
|
25 |
583.1 |
422.50 |
160.6 |
165.6 |
153.4 |
12.20 |
22.40 |
5.83 |
16.57 |
40 |
|
26 |
300.2 |
252.50 |
47.70 |
65.1 |
54.5 |
10.60 |
11.11 |
4.64 |
06.47 |
31 |
|
27 |
275.6 |
231.40 |
44.20 |
70.1 |
59.6 |
10.50 |
11.60 |
4.85 |
06.75 |
28 |
|
28 |
645.7 |
483.60 |
162.1 |
192.3 |
137.0 |
55.30 |
28.70 |
6.36 |
22.34 |
43 |
|
29 |
498.5 |
331.90 |
166.6 |
133.6 |
98.0 |
35.60 |
18.59 |
4.26 |
14.33 |
36 |
|
30 |
440.3 |
332.10 |
108.2 |
68.9 |
40.0 |
28.90 |
21.20 |
6.84 |
14.36 |
38 |
Eau oxveénée
|
Plants |
Poids en mg |
Longueur en cm |
Nombre de |
|||||||
|
Frais |
Sec |
|||||||||
|
Total |
Aérienne |
Racinaire |
Total |
Aérienne |
Racinaire |
Total |
Aérienne |
Racinaire |
||
|
1 |
315.6 |
253.8 |
61.8 |
64.19 |
52.12 |
12.07 |
19.53 |
5.26 |
14.27 |
31 |
|
2 |
453.3 |
321.6 |
131.7 |
74.48 |
61.19 |
13.29 |
24.78 |
5.97 |
18.81 |
37 |
|
3 |
362.1 |
279.6 |
82.5 |
103.0 |
80.97 |
22.03 |
15.73 |
6.10 |
9.63 |
37 |
|
4 |
591.3 |
430.7 |
160.6 |
181.6 |
127.3 |
54.3 |
27.41 |
6.21 |
21.20 |
41 |
|
5 |
725.6 |
501.2 |
224.4 |
209.3 |
148.5 |
60.8 |
25.38 |
6.23 |
19.15 |
49 |
|
6 |
552.6 |
401.6 |
151.0 |
188.5 |
137.20 |
51.30 |
21.63 |
6.21 |
15.42 |
45 |
|
7 |
591.8 |
432.3 |
159.5 |
191.7 |
125.8 |
65.9 |
24.14 |
6.27 |
17.87 |
41 |
|
8 |
799.8 |
572.6 |
227.2 |
255.3 |
192.1 |
63.2 |
27.92 |
6.19 |
21.73 |
53 |
|
9 |
623.1 |
441.2 |
181.9 |
198.6 |
150.1 |
48.50 |
31.26 |
6.96 |
24.30 |
45 |
|
10 |
641.7 |
483.6 |
158.1 |
191.9 |
136.6 |
55.3 |
19.18 |
6.38 |
12.80 |
43 |
|
11 |
482.4 |
314.1 |
168.3 |
133.2 |
96.41 |
36.79 |
15.74 |
5.27 |
10.47 |
31 |
|
12 |
654.2 |
440.2 |
214.0 |
156.7 |
113.9 |
42.8 |
29.56 |
6.54 |
23.02 |
34 |
|
13 |
251.7 |
200.6 |
51.10 |
106.8 |
77.40 |
29.40 |
12.08 |
5.29 |
6.79 |
29 |
|
14 |
262.3 |
230.5 |
31.8 |
63.9 |
50.2 |
13.7 |
12.54 |
6.16 |
6.38 |
27 |
|
15 |
726.1 |
511.3 |
214.8 |
200.2 |
145.60 |
54.60 |
24.11 |
5.96 |
18.15 |
44 |
|
16 |
268.2 |
217.1 |
51.10 |
67.9 |
53.90 |
14.00 |
12.25 |
6.07 |
6,18 |
30 |
|
17 |
256.7 |
209.1 |
47.60 |
70.5 |
54.40 |
16.1 |
10.76 |
5.29 |
5.47 |
27 |
|
18 |
384.6 |
287.7 |
96.9 |
69.3 |
56.65 |
12.65 |
22.15 |
5.61 |
16.54 |
34 |
|
19 |
559.6 |
403.8 |
155.8 |
164.6 |
118.9 |
45.70 |
22.84 |
6.18 |
16.66 |
41 |
|
20 |
648.00 |
468.83 |
179.17 |
211.8 |
151.70 |
60.10 |
24.56 |
6.22 |
18.34 |
46 |
|
21 |
582.4 |
412.9 |
169.5 |
174.56 |
127.70 |
46.86 |
20.06 |
6.20 |
13.86 |
40 |
|
22 |
389.4 |
290.4 |
99 |
109.00 |
80.50 |
28.50 |
18.56 |
5.99 |
12.57 |
30 |
|
23 |
417 |
312.5 |
104.5 |
112.86 |
84.63 |
28.23 |
15.70 |
5.77 |
9.93 |
34 |
|
24 |
554.6 |
324.3 |
230.3 |
75.55 |
62.21 |
13.34 |
25.81 |
6.03 |
19.78 |
38 |
|
25 |
592.4 |
431.7 |
160.7 |
183.2 |
128.90 |
54.3 |
27.63 |
6.36 |
21.27 |
42 |
|
26 |
593.6 |
432.3 |
161.3 |
191.10 |
124.8 |
66.3 |
24.94 |
6.31 |
18.63 |
42 |
|
27 |
643.2 |
481.90 |
161.3 |
192.3 |
135.7 |
56.60 |
19.22 |
6.39 |
12.83 |
43 |
|
28 |
276.7 |
222.9 |
53.8 |
107.4 |
77.7 |
29.7 |
13.10 |
5.31 |
7.79 |
30 |
Temoin
|
Poids en mg |
Longueur en cm |
Nombre de |
||||||||
|
Plants |
Frais |
Sec |
||||||||
|
Total |
Aérienne |
Racinaire |
Total |
Aérienne |
Racinaire |
Total |
Aérienne |
Racinaire |
||
|
1 |
231.2 |
181.1 |
50.1 |
64.23 |
52.21 |
12.02 |
11.81 |
5.41 |
6.40 |
32 |
|
2 |
435.5 |
305.2 |
130.3 |
74.27 |
61.56 |
12.71 |
17.29 |
5.57 |
11.72 |
38 |
|
3 |
324.3 |
281.9 |
42.40 |
119.6 |
79.10 |
40.10 |
13.04 |
6.00 |
7.04 |
39 |
|
4 |
325.4 |
280.9 |
44.50 |
200.1 |
180.6 |
19.50 |
10.67 |
5.21 |
5.46 |
37 |
|
5 |
176.6 |
151.5 |
25.10 |
54.81 |
44.00 |
10.81 |
9.61 |
5.31 |
4.30 |
27 |
|
6 |
162.9 |
121.7 |
41.20 |
50.62 |
31.31 |
19.31 |
12.71 |
5.23 |
7.48 |
30 |
|
7 |
233.2 |
191.5 |
41.70 |
78.76 |
59.19 |
19.57 |
12.63 |
6.07 |
6.56 |
27 |
|
8 |
192.1 |
142.3 |
49.80 |
63.21 |
44.25 |
18.96 |
13.19 |
5.20 |
7.99 |
27 |
|
9 |
225.4 |
201.6 |
23.80 |
75.38 |
61.21 |
14.17 |
11.63 |
4.84 |
6.79 |
29 |
|
10 |
342.7 |
302.1 |
40.60 |
122.40 |
90.32 |
32.08 |
14.74 |
6.61 |
8.13 |
31 |
|
11 |
236.3 |
204.2 |
32.10 |
82.72 |
69.71 |
13.01 |
10.75 |
5.03 |
5.72 |
29 |
|
12 |
231.5 |
187.3 |
44.20 |
70.86 |
49.89 |
20.97 |
12.88 |
6.11 |
6.77 |
30 |
|
13 |
334.2 |
296.6 |
37.60 |
130.71 |
108.68 |
22.03 |
13.79 |
6.65 |
7.14 |
36 |
|
14 |
167.6 |
124.4 |
43.20 |
53.18 |
32.71 |
20.47 |
12.51 |
5.09 |
7.42 |
28 |
|
15 |
223.7 |
198.6 |
25.10 |
62.81 |
41.21 |
21.60 |
11.93 |
5.78 |
6.15 |
31 |
|
16 |
178.2 |
154.7 |
23.50 |
54.36 |
43.82 |
10.54 |
10.03 |
5.34 |
4.69 |
28 |
|
17 |
234.4 |
201.2 |
33.20 |
77.53 |
54.54 |
22.99 |
11.68 |
6.09 |
5.59 |
30 |
248
|
NGFf |
NGAtf |
NGApf |
NGBf |
NGVf |
NGTVf |
NGFm |
NGAtm |
NGApm |
NGBm |
NGVm |
NGTVm |
NGFF |
NGatF |
NGapF |
NGBF |
NGVF |
NGTVF |
|||||||||||||||||
|
NGFf |
1.000 |
|||||||||||||||||||||||||||||||||
|
NGAtf |
0.280 |
1.000 |
||||||||||||||||||||||||||||||||
|
NGApf |
-0.935 |
-0.284 |
1.000 |
|||||||||||||||||||||||||||||||
|
NGBf |
-0.286 |
-0.483 |
0.266 |
1.000 |
||||||||||||||||||||||||||||||
|
NGVf |
-0.922 |
-0.316 |
0.976 |
0.452 |
1.000 |
|||||||||||||||||||||||||||||
|
NGTVf |
-0.131 |
-0.185 |
0.423 |
0.518 |
0.505 |
1.000 |
||||||||||||||||||||||||||||
|
NGFm |
-0.085 |
0.158 |
0.168 |
0.072 |
0.201 |
0.323 |
1.000 |
|||||||||||||||||||||||||||
|
NGAtm |
-0.018 |
-0.116 |
-0.030 |
0.337 |
0.060 |
0.115 |
0.163 |
1.000 |
||||||||||||||||||||||||||
|
NGApm |
-0.251 |
-0.064 |
0.115 |
-0.038 |
0.177 |
-0.261 |
-0.285 |
-0.115 |
1.000 |
|||||||||||||||||||||||||
|
NGBm |
-0.056 |
0.013 |
0.133 |
-0.028 |
0.087 |
0.069 |
-0.188 |
-0.163 |
-0.222 |
1.000 |
||||||||||||||||||||||||
|
NGVm |
-0.296 |
-0.177 |
0.231 |
0.147 |
0.222 |
-0.091 |
-0.366 |
0.005 |
0.839 |
0.257 |
1.000 |
|||||||||||||||||||||||
|
NGTVm |
-0.328 |
0.049 |
0.349 |
0.188 |
0.371 |
0.219 |
0.613 |
0.153 |
0.450 |
0.042 |
0.510 |
1.000 |
||||||||||||||||||||||
|
NGFF |
-0 997 |
-0 970 |
0 176 |
0 045 |
0 117 |
-0 110 |
-0 997 |
-0 009 |
0 070 |
0 OM |
0 lOR |
-0 1R9 |
1 000 |
|||||||||||||||||||||
|
NGatF |
0.194 |
0.300 |
-0.295 |
0.031 |
-0.253 |
-0.215 |
0.068 |
0.270 |
0.021 |
0.262 |
0.189 |
0.225 |
-0.161 |
1.000 |
||||||||||||||||||||
|
NGapF |
0.168 |
-0.143 |
-0.040 |
-0.060 |
-0.057 |
0.226 |
0.051 |
-0.303 |
0.010 |
-0.008 |
-0.076 |
-0.021 |
-0.482 |
-0.436 |
1.000 |
|||||||||||||||||||
|
NGBF |
0.282 |
0.222 |
-0.306 |
0.153 |
-0.249 |
-0.010 |
-0.010 |
0.317 |
0.043 |
-0.175 |
0.081 |
0.059 |
-0.179 |
0.138 |
-0.282 |
1.000 |
||||||||||||||||||
|
NGVF |
0.450 |
0.137 |
-0.386 |
0.037 |
-0.349 |
0.109 |
0.085 |
0.006 |
0.047 |
0.052 |
0.071 |
0.139 |
-0.704 |
0.176 |
0.678 |
0.308 |
1.000 |
|||||||||||||||||
|
NGTVF |
0.211 |
-0.156 |
-0.283 |
-0.010 |
-0.283 |
-0.253 |
-0.268 |
0.002 |
0.147 |
0.154 |
0.228 |
-0.052 |
0.353 |
0.030 |
0.273 |
0.182 |
0.415 |
1.000 |
||||||||||||||||
|
SGFf |
SGAtf |
SGApf |
SGBf |
SGVf |
SGTVf |
SGFm |
SGAtm |
SGApm |
SGBm |
SGVm |
SGTVm |
SGFF |
SGatF |
SGapF |
SGBF |
SGVF |
SGTVF |
|||||||||||||||||
|
SGFf |
1.000 |
|||||||||||||||||||||||||||||||||
|
SGAtf |
0.699 |
1.000 |
||||||||||||||||||||||||||||||||
|
SGApf |
0.216 |
-0.405 |
1.000 |
|||||||||||||||||||||||||||||||
|
SGBf |
-0.092 |
-0.032 |
0.136 |
1.000 |
||||||||||||||||||||||||||||||
|
SGVf |
-0.178 |
-0.289 |
0.870 |
0.597 |
1.000 |
|||||||||||||||||||||||||||||
|
SGTVf |
0.209 |
-0.018 |
0.781 |
0.558 |
0.925 |
1.000 |
||||||||||||||||||||||||||||
|
SGFm |
0 29R |
0 161 |
0 061 |
-0 100 |
-0 091 |
0 099 |
1 000 |
|||||||||||||||||||||||||||
|
SGAtm |
0.572 |
0.444 |
-0.072 |
-0.049 |
-0.005 |
0.164 |
0.304 |
1.000 |
||||||||||||||||||||||||||
|
SGApm |
-0.281 |
-0.262 |
-0.062 |
0.334 |
0.096 |
-0.013 |
-0.421 |
-0.354 |
1.000 |
|||||||||||||||||||||||||
|
SGBm |
0.125 |
0.240 |
0.029 |
-0.219 |
-0.043 |
-0.003 |
-0.214 |
0.490 |
-0.171 |
1.000 |
||||||||||||||||||||||||
|
SGVm |
-0.108 |
-0.062 |
-0.069 |
0.198 |
0.045 |
0.003 |
-0.466 |
0.068 |
0.882 |
0.397 |
1.000 |
|||||||||||||||||||||||
|
SGTVm |
0.063 |
0.032 |
-0.038 |
0.040 |
-0.005 |
0.019 |
0.091 |
0.263 |
0.568 |
0.313 |
0.839 |
1.000 |
||||||||||||||||||||||
|
SGFF |
0 OR1 |
0.275 |
0.110 |
0.201 |
0.242 |
0.242 |
-0 91R |
0.084 |
-0.032 |
0.123 |
0.011 |
-0.102 |
1.000 |
|||||||||||||||||||||
|
SGatF |
-0.058 |
-0.251 |
-0.023 |
0.060 |
-0.024 |
-0.046 |
0.080 |
-0.347 |
0.203 |
-0.208 |
0.027 |
0.081 |
-0.469 |
1.000 |
||||||||||||||||||||
|
SGapF |
-0.163 |
-0.155 |
-0.194 |
-0.192 |
-0.248 |
-0.309 |
-0.446 |
0.076 |
0.177 |
0.314 |
0.309 |
0.075 |
-0.328 |
-0.227 |
1.000 |
|||||||||||||||||||
|
SGBF |
-0.192 |
-0.116 |
-0.037 |
-0.031 |
-0.100 |
-0.183 |
0.090 |
-0.003 |
0.237 |
-0.187 |
-0.331 |
-0.314 |
-0.155 |
0.233 |
-0.072 |
1.000 |
||||||||||||||||||
|
SGVF |
-0.162 |
-0.289 |
-0.132 |
-0.156 |
0.205 |
-0.267 |
-0.227 |
-0.159 |
0.163 |
0.053 |
0.174 |
0.059 |
-0.667 |
0.575 |
0.577 |
0.282 |
1.000 |
|||||||||||||||||
|
SGTVF |
-0.090 |
0.006 |
-0.017 |
0.072 |
0.024 |
0.011 |
-0.544 |
-0.83 |
0.153 |
0.219 |
0.245 |
-0.060 |
0.472 |
0.038 |
0.269 |
0.138 |
0.341 |
1.000 |
||||||||||||||||
Légende N :Versant Nord - S : Versant Sud - f faible - m : moyenne F : forte - GF fertile - GV : vaine - at : attaquées - ap : aplaties - B : bec - TV : tout venant