ABSTRACT
Although the existence of great deal control strategies, the
cowpea, Vigna unguiculata production, still limited by pests.
In this thesis, we study the possibility of Amblyseius swisrkii
(Acari: Phytoseiidae) Athias-Henriot, to reproduce with Megalurothrips
sjostedti Trybom (Thysanoptera: thripidae) larvae instars and pollen of
this western flowering thrips host plants. So our works are conducted to
appreciate the vulnerability at the predation of M. sjostedti larvae
instars. The effect of the pollen of five thrips host plants (Lonchocarpus
sericeus, Vigna unguiculata, Tephrosia candida, Zea
mays and Typha australis) was tested too on the reproduction
function and the survival of the predator. The pollens were extracted from the
flowers of the thrips host plants, collected in different regions of south and
central-Benin. In the laboratory, the vulnerability of thrips was tested on the
first instars of M. sjostedti. (larvae 1 and larvae 2). We used five
(5) larvae for each instars with one (1) predator as densities. The results
show that the predator does not attack the larvae 2 but has a significant
effect on larvae 1 instars of the pest. In addition, phytoseiid nourish and lay
well on all the pollen sources. But the combination of cowpea pollen and larvae
1, reduces the predation rate of the phytoseiid. Nevertheless, A.
swirskii, is an interesting biological agent against M.
sjostedti.
Key words: pollens, diet, life table,
phytoseiid, Amblyseius swirskii, predator, acari, Vigna
unguiculata, Tephrosia candida, Zea mays,
Lonchocarpus sericeus, Typha australis.

INTRODUCTION
Le niébé, Vigna unguiculata (L.)
Walpers, est la légumineuse vivrière la plus importante et la
plus cultivée dans les régions d'Afrique Tropicale (Jakai &
Adalla, 1997). Il peut être cultivé en association avec d'autres
cultures principales tels que le sorgho, le maïs, l'igname ou le mil. Le
niébé se caractérise par sa richesse en protéines
et contribue énormément à la fertilisation des sols
grâce à la fixation symbiotique de l'azote atmosphérique
(Sun et Simbi, 1983). Au plan alimentaire, il occupe une place de choix du fait
qu'il constitue une importante source de protéines et d'énergie
tant pour les hommes que pour les animaux (Rachie, 1985; Anonyme, 2002). Dans
les pays en développement où l'accès aux protéines
d'origine animale est difficile voire impossible pour certaines populations, le
niébé constitue la seule source de protéine la plus
accessible et il est appelé, pour ce fait, «la viande du
pauvre». Ainsi, le niébé est susceptible de combler les
déficits protéiques des pays en développement qui
regorgent les trois quarts (3/4) de la population mondiale, mais qui ne
produisent que seulement le quart de la production mondiale de viande (Delobel
& Tran, cités par Capo-Chichi, 2005). Il constitue alors un aliment
d'appoint non négligeable pour l'amélioration de l'état
nutritionnel des populations des pays pauvres. Malgré cette
utilité du niébé, les rendements et les superficies
emblavées pour sa culture évoluent en dents de scie au
Bénin (INRAB, 1995).
Selon Coulibaly & Lowenber-Deboer (2002), l'Afrique
Occidentale est actuellement loin de couvrir ses besoins en niébé
par sa propre production. Dans cette partie du continent, les rendements moyens
oscillent autour de 230 kg/ha, sans application des pesticides, contre 800 kg/
ha avec application de pesticides (PEDUNE, 1998), alors que le rendement
potentiel du niébé est de 3000 kg/ha (Rusoke & Rubaihayo,
1994). Cette faiblesse du rendement est due au complexe parasitaire
associé à cette culture depuis la levée jusqu'au stockage
(Singh & Allen, 1980; Ahounou, 1990; Atachi, 1998; Agboton, 2004). Parmi
les insectes ravageurs, le thrips floricole Megalurothrips sjostedti
(Trybom), (Thysanoptera: Thripidae) (Syn: Taeniothrips sjostedti)
constituent la contrainte majeure à la production du niébé
dans les régions de l'Afrique au sud du Sahara (Okwakpam, 1967; Singh
& Allen, 1980; Ezueh, 1981; Jakai & Daoust, 1986; Tamo, 1991; Tamo
et al., 1993; Bottenberg et al., 1997). Ce
ravageur est capable d'occasionner des pertes de rendement
atteignant 20% à 80% des récoltes. En cas de fortes infestations,
les pertes de rendements sont totales.
Au nombre des exigences dont il faudrait tenir compte pour
augmenter la production du niébé au Bénin, on peut citer
les traitements phytosanitaires (MDR, 1992). Cette recommandation encourage
l'utilisation des pesticides de synthèse dont les effets nocifs des
résidus, ne sont plus ignorés de nos jours, quel que soit le
respect des doses recommandées. Chose plus grave, l'utilisation des
pesticides induit la résistance chez les ravageurs occasionnant leur
résurgence, et a des effets nocifs sur la santé humaine et sur
l'environnement (Programme Natura/Nectar, 1996). Il en résulte un
important risque d'intoxication difficilement chiffrable (Tissut et
al., 1979). Le faible niveau de revenu des producteurs ne leur permet pas
un accès facile à ces pesticides, du fait de leur coût
très élevé. Il s'avère alors impératif, de
rechercher d'autres méthodes de luttes contre ces ravageurs, dans le
contexte d'une agriculture écologique. On peut citer entre autres la
lutte biologique. En effet, la lutte biologique est une méthode de
protection des cultures dont le but est de réduire la population des
ravageurs en dessous du seuil économique des dégâts. Elle
se base sur l'utilisation des ennemis naturels natifs et ou exotiques pour
lutter contre les ravageurs des cultures. Cette façon de gérer
les ravageurs, permet d'éviter les effets néfastes des pesticides
chimiques sur l'environnement, les producteurs et les consommateurs (Nomikou,
2003). Dans ce contexte, certaines potentialités biologiques de
Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae), relatives
à son régime alimentaire seront évaluées, pour une
lutte biologique efficiente contre le thrips, M. sjostedti. Mais le
succès d'un programme de contrôle biologique doit provenir d'une
connaissance approfondie des paramètres bioécologiques de
l'ennemi naturel (Waag, 1989).
L'objectif global de la présente étude est de
contribuer à l'évaluation des capacités de A.
swirskii à réduire ou à supprimer les populations de
thrips en pullulation
De façon spécifique, il s'agira :
- d'évaluer la prédation de A. swirskii
sur les premier et deuxième stades larvaires de M.
sjostedti;
- de sélectionner les plantes-hôtes dont le pollen
assure au mieux la reproduction de A. swirskii; et
- d'évaluer l'effet de la combinaison du meilleur pollen
et des larves de M. sjostedti, sur le développement, la
reproduction et la survie de A. swirskii.

2.1 Le niébé, Vigna unguiculata
(L.) Walp. (Fabaceae).
Depuis des décennies, le thrips floricole, M.
sjostedti Trybom (Thysanoptera: Thripidae), l'un des principaux ravageurs
cibles du niébé, fait l'objet de nombreux travaux de recherche.
L'attachement des chercheurs à ce domaine, tient des lourdes pertes
qu'occasionnent ce thrips floricole sur la production de niébé
(Okwakpam, 1967; Singh & Allen, 1980; Ezueh, 1981; Jakai & Daoust,
1986; Tamo, 1991; Tamo et al., 1993 ; Bottenberg et al.,
1997). De nos jours, l'approche de lutte intégrée contre les
ravageurs, basée sur la manipulation de l'environnement est de plus en
plus envisagée, pour assurer une méthode de protection saine et
durable qui préserve la santé des producteurs ainsi que celle des
consommateurs (Tissut et al., 1979).
La présente revue de littérature se propose donc
de faire une synthèse des informations nécessaires à une
connaissance générale des facteurs édapho-climatiques,
sociologiques et le complexe parasitaire liés à la production de
niébé.
2.1.1- Ecologie, distribution et
production
Le niébé se développe dans les conditions
de chaleur et de luminosité intense. C'est une plante thermophile qui
requiert tout au long de sa croissance, une température oscillant entre
25 et 28°C et une pluviométrie de 750 à 1000 mm (Anochili,
1978). Il vit bien dans les sols profonds et bien drainés et est
tolérant à la sécheresse ainsi qu'à la
salinité du sol. Il peut se développer sous des conditions
environnementales variées et sur des sols pauvres, sans addition
d'engrais azotés. Il peut être semé en culture pure ou en
association avec d'autres cultures. Dans les associations de cultures, les
paramètres tels que la densité de plantation, les arrangements
spatiaux, la date de semis, le nombre de cultures associées varient
d'une région à une autre. Ces paramètres tiennent compte
de la fertilité des sols, de la disponibilité des semences et des
besoins alimentaires de la famille. Parmi les principales légumineuses
vivrières, le niébé et l'arachide occupent une place
importante en superficie et en production. Parfois les superficies du
niébé dépassent celles de l'arachide. En 2003, le
niébé a pris le dessus sur toutes les légumineuses avec
une contribution de 7 1,5% des emblavures, suivi de voandzou 10,6% (INRAB,
1995). Cela montre que le niébé est aussi une source de revenu
non
négligeable pour les producteurs. Mais l'essentiel de
la production est assuré par les petits paysans qui cultivent le
niébé avec des méthodes encore extensives.
Le niébé est cultivé au Bénin sur
toute l'étendue du territoire national. Les zones de prédilection
se rencontrent par ordre d'importance dans l'Ouémé, le Mono et le
Zou (MAEP, 2004). Les statistiques sur les douze dernières années
montrent que les superficies de production et les rendements évoluent en
dents de scie. Les rendements sont restés faibles et ne dépassent
pas 600 kg/ha (Agboton, 2004). Cette faiblesse est due au fait que le
niébé est toujours fortement parasité au champ et le
manque de traitements phytosanitaires adéquats affecte le rendement. Au
nombre des exigences dont il faudra tenir compte pour augmenter la production
du niébé, le Ministère du Développement Rural (MDR,
1992) préconise d'assurer le traitement phytosanitaire comme le font
déjà si bien les petits producteurs du coton. Ce travail doit se
faire en collaboration avec les structures d'encadrement technique
opérant sur le terrain.
2.1.2- Importance et utilisation du
niébé
Le niébé constitue la plus importante
légumineuse à graine cultivée en Afrique Tropicale. Il se
caractérise surtout par sa richesse en protéines (24-28%) et
autres constituants comme l'eau 11%, les hydrates de carbone 56,8%, les lipides
(1,3%), les fibres (3,9%), les cendres (3,6%), la vitamine A (32,42UI /100g),
la vitamine D (26- 78,02ug/100g) et la vitamine E (3,07-5,07 mg/100g). les
grains de niébé contiennent aussi du calcium (90mg/100g) du fer
(6-7mg/100g), de l'acide nicotinique (2mg/100g), de la thiamine (0,9mg/100g).
Mais il est pauvre en lysine (Platt, 1962; Oyenuga, 1968; Ogounmodedo &
Oyenuga, 1968; IITA, 1975).
Le niébé fournit une valeur
énergétique de 342 calories par 100g de graines (Oyenuga, 1968).
Sa capacité à fixer de l'azote atmosphérique lui
confère le rôle important de précédent cultural et
du maintien de la fertilité des sols (Ahounou, 1990). En dehors de ces
graines, les feuilles du niébé sont consommées comme
légume par les hommes, et comme fourrage par le bétail (INRAB,
1995). Le niébé sert aussi à la fabrication des colorants
verts (Bezpaly, 1984).
2.1.3- Complexe parasitaire du
niébé
Un vaste éventail de ravageurs s'attaque au
niébé depuis la levée jusqu'à la maturation
complète, occasionnant ainsi d'énormes dégâts
(Ahounou, 1990). Ces ravageurs ont été catégorisés
en trois groupes par Singh et al. (1979) :
+ Les insectes de pré-floraison
Il s'agit des coléoptères Chrysomelidae,
Ootheca mutabilis (Sahlb) et Paraluperodes quaternus
(Fairmare); des hémiptères Aphididae, Aphis craccivora
(Koch), des thysanoptères Thripidae du feuillage, Sericothrips
occipitalis (Hood), et les familles Jassidae, Empoasca dolichi
(Paoli).
+ Les insectes floricoles du
niébé:
Ce sont:
- les thysanoptères parasites des fleurs,
Megalurothrips sjostedti (Tryb.).
- les lépidoptères foreurs de gousses,
Maruca vitrata (Fabricius) et Cydia ptychora (Meyrick)
- les hétéroptères suceurs des gousses
Anoplocnemis curvipes (F.), Riptortus dentipes (F.),
Acanthomia tomentosicollis (Stäl), A. horrida (Germ) et
Nezara viridula (L.)
- Les coléoptères dévoreurs de fleurs
Mylabris farquharsoni
+ Les insectes de stock:
Les coléoptères Bruchidae Callosobruchus
maculatus (Fabricius), et Bruhidius atrolineatus (Pic). Nombre de
ces ravageurs sont des vecteurs des maladies virales, très
préjudiciables au niébé, en termes de qualité du
produit et du rendement. La mosaïque jaune du niébé est la
maladie virale la plus fréquemment transmise par les insectes floricoles
comme S. occipitalis, M. sjostedti, les
coléoptères P. quaternus, Nematocerus acerbus
Wilke ( Curculionidae) et Zonocerus variegatus F (Acridoidea:
Pygomophidae). (Whitney et al., cité par Sourokou, 1985).
En Afrique tropicale, en dehors des dégâts
spécifiques à chaque ravageur, le thrips M. sjostedti
peut occasionner jusqu'à 60% voire 100% de perte de rendement (Singh
& Allen, 1980; Ezueh, 1981); les larves causant plus de dégâts
que les adultes (Salifu, 1986).
2.2 Megalurothrips sjostedti (Thysanoptera:
Thripidae) Trybom
Le thrips floricole, M. sjostedti a
bénéficié de nombreuses études relatives à
son origine, sa distribution, sa taxonomie, sa morphologie et sa
bioécologie. Aussi des méthodes de lutte ont-elles
été développées pour la gestion de ce ravageur
redoutable au niébé.
2.2.1- Origine et distribution
:
L'espèce Megalurothrips sjostedti a
été rapportée pour la première fois sur le
continent africain, précisément en Afrique de l'Est, au
début du vingtième siècle (1905). Non seulement sa
répartition est restreinte au continent africain, mais il constitue la
seule espèce du genre retrouvée en Afrique (Palmer, 1987).
L'abondance en Asie de l'Est de certaines espèces de ce genre telles que
M. typicus (Bagnall), M. usitatus (Bagnall), laisse
suggérer l'Asie tropicale comme origine du genre Megalurothrips
(Palmer, 1987). Selon Salifu (1986), M. sjostedti est répandu
dans les agro-écosystèmes des pays comme l'Afrique du Sud, le
Bénin, le Cameroun, le Congo Démocratique, la Côte
d'Ivoire, le Gabon, la Gambie, le Ghana, la Guinée Equatoriale,
l'Ouganda et la Tanzanie. Autrefois appelé Taeniothrips
sjostedti, M. sjostedti est la seule espèce ayant le
statut de ravageur des cultures (Palmer, 1987). En Asie de l'Est, cette
espèce n'est pas nuisible aux cultures, puisque ses populations sont
maintenues en dessous du seuil économique des dégâts, par
des ennemis naturels. Cet argument renforce l'hypothèse selon laquelle
M. sjostedti ne soit pas d'origine africaine (Tamo et al.,
1997).
2.2.2- Taxonomie
Megalurothrips sjostedti appartient au super-ordre
des Thysanoptéroides, à l'ordre des Thysanoptères, au
sous-ordre des Terebrantia, à la famille des Thripidae, et à la
sous- famille des Thripinae (Lewis, 1997). Selon Palmer (1987), le genre
Megalurothrips regroupe des espèces facilement identifiables
par leur grande taille. Au stade adulte, ils sont de couleur noire alors que
les stades larvaires sont de couleur variable: blanchâtre, orange ou
rouge en fonction du régime alimentaire. Les mâles de M.
sjostedti sont caractérisés par un segment abdominal de
forme conique, alors que la femelle possède un petit ovipositeur
externe.
2.2.3- Morphologie, Biologie et Ecologie
Les thrips floricoles sont de petits insectes de couleur
noirâtre de 1 à 2 mm de long, pourvus d'ailes étroites,
allongées et frangées de longs cils. On observe une segmentation
bien distincte des antennes, des pattes et de l'abdomen. De petite taille et
d'un noir pas trop foncé, les mâles sont estimés à
5% de la population, tandis que les femelles très noires, sont plus
grandes et plus abondantes (Tamo, 1991). La photo 1 montre les adultes
mâles et femelles de M. sjostedti.

1,5 mm

Photo 1: Mâle (à gauche) et femelle
(à droite) des thrips floricoles
Source: GOERGEN (communication personnelle)
IITA. Insect Museum, Cotonou Bénin
Selon Salifu (1986), le développement de l'oeuf
à l'adulte dure 19 jours dans les conditions de température de
26,8 #177; 1,4°C et de 65 #177; 10,4% d'humidité relative. Quatre
stades de développement sont identifiés: deux stades larvaires
(larve stade 1 et larve stade 2), un stade de pré nymphe et un stade de
nymphe. Les larves de stades 1 et 2 sont de très petite taille ; leurs
yeux sont rouges net brillants, avec des antennes, des pattes et l'abdomen
courts. Les nymphes se forment dans le sol. En l'absence du
niébé, les thrips se nourrissent et se reproduisent sur plusieurs
plantes hôtes. On y rencontre majoritairement des Fabaceae, quelques
Mimosaceae et Cesalpiniaceae, et une plante non légumineuse,
Cochlospermum planchoni Hook (Bisaceae) (Tamo et al., 1993a).
La photo 2 montre les différents stades de développement de
M. sjostedti.
b
a
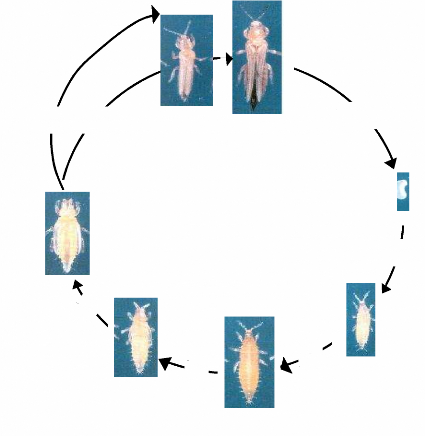
1 jour
3 - 4 jours
d
3 - 4 jours
c
g
3 - 4
jours
3 - 4
jours
f
e
2-3
jours
a) Adulte mâle b) Adulte femelle c) oeuf d) Larve
de stade 1 e) Larve de stade 2 f) Pré nymphe g) Nymphe
Photo 2 : cycle de développement M.
sjostedti, à 25°C
Source: Adapté de Tamo (1991).
2.2.4- Méthode de lutte
contre les thrips
La connaissance incomplète des caractères
biologiques de M. sjostedti restreint la gamme des mesures de lutte
contre cet insecte. Toutefois, plusieurs méthodes de lutte ont
donné des résultats appréciables (Sourokou, 1985).
+ Lutte chimique
Les pyréthrinoïdes synthétiques en
formulation simple ou combinée avec d'autres insecticides
contrôlent bien la population des thrips (IITA 1982). Le monocrotophos,
appliqué sur niébé s'est révélé plus
efficace que le fenvalérate et le DDT
(dichlodiphényltrichloroéthane) au sud du Nigéria entre
1973 et 1975. Le cymbush et le mélange cymbush (cypermétthrine) +
diméthoate sont significativement plus efficaces que atellic et
diméthoate. Un test de comparaisons d'insecticides, a conduit à
la conclusion que azodrine assure significativement un meilleur contrôle
des thrips et permet d'avoir de hauts rendements par rapport au DDT et au
sevimol. Le stade critique de menace des ravageurs est atteint à partir
du 37ème jour après semis. Singh (1980) suggéra
des interventions chimiques entre le 35ème et le
40ème jour; cependant, le début de floraison du
niébé dépend de son cycle et de sa sensibilité au
photopériodisme. Malgré leur efficacité, les effets
pervers de ces insecticides font accorder une attention particulière aux
méthodes de lutte plus écologiques (Tissut et al.,
1979).
+ Lutte culturale
Il est préférable que dans un système
d'association maïs-niébé, le maïs soit semé
avant le niébé, de façon que le maïs entre en
floraison avant celle du niébé. Cette pratique contribue au bon
développement du niébé et permet aussi de réduire
les dégâts occasionnés par les thrips. Lors de la
déhiscence des panicules, les méloïdes sont attirés
par les pollens du niébé (IITA, 1982). Par contre Rathore et
al., (1984) rapportent qu'en culture intercalaire, il n'y a pas de
différence significative entre la densité de population des
thrips sur le niébé en culture pure et celle associée au
maïs (IITA/SAFGRAD, 1984).
+ Lutte biologique
En relation avec les questions écologiques, doivent
être abordés, les problèmes des êtres vivants
auxiliaires de l'agriculture, des pollinisateurs et des prédateurs
divers
s'attaquant aux nuisibles. Dans le processus de contrôle
des thrips, des prédateurs et des parasitoïdes ont
été identifiés. Orius amnesius, O.
albidipennis (Hémiptère: Anthocoridae); Cheilomenes
sulphurea (Coléoptère: Coccinellidae), Iphyseius
sp. (Acari : Phytoseiidae) (Tamo et al., 1997) sont des
prédateurs identifiés pour une lutte biologique contre M.
sjostedti. Au nombre des parasitoïdes identifiés pour la
réduction naturelle des populations de M. sjostedti, on peut
citer: Ceranisus menes (Gahan) (Hyménoptère: Eulophidae)
(Diop, 1999), Ceranisus femoratus (Gahan) (Hyménoptère:
Eulophidae) (Agboton, 2004) et des genres de la famille Trichogrammatidae que
sont Megaphragma sp et Oligosita sp (Tamo et al.,
1997). Le tableau 1 présente la liste de quelques prédateurs de
thrips classés par ordre, famille, genre et espèce. Toutefois, la
question de l'efficacité de ces différents auxiliaires contre les
thrips se pose. Alors, des méthodes de lutte culturale et des
variétales sont aussi utilisées.
Tableau 1: Liste des prédateurs de
thrips
|
Ordre
|
Famille
|
Genre
|
Espèces
|
Proies (Thrips)
|
|
Orthoptère
|
Gryllidae
|
Oecanthus
|
Longicauda Turanicus
|
Thrips tabaci
Thrips tabaci
|
|
Névroptère
|
Chrysopidae
|
Chrysopa
|
Alobana Carnea Vulgaris Plorabula
|
Selenothrips rubrocinctus Selenothrips rubrocinctus
Odontothrips intermedius Caliothrips faciatus
|
|
Leucochrysa
|
Marquesi Submacula
|
Selenothrips rubrocinctus
Selenothrips
rubrocinctus
|
|
Hemerobiidae (brown lacewings)
|
Hemero bi us
|
Maliformicus Pacificus
|
Taeniothrips inconsequens
Taeniothrips
inconsequens
|
|
Diptère
|
Cecidomyiidae
|
Adelgimyza Artrocnodax
|
Thripiperda Occidentalis
|
Liothrips olae T. palmi
|
|
Asilidae
|
Machinu
|
Annuleps
|
Haplothrips sp.
|
|
Dolichopodidae
|
Condilostylus
|
Pacifus
|
Taeniothrips inconsequens
|
|
Syrohidae
|
Baccha Mesograpta
|
Livida Marginata
|
Gynaikothrips ficorum Thrips tabaci
|
|
Chloropididae
|
Lioscinella
|
Sabrosky
|
Teuchothrips sp.
|
|
Hyménoptère
|
Vespidae
|
Polistes
|
Hebraeus
|
Rhipiphorothrips cruentatus
|
|
Azteca Pheidole
|
Chartifox Megacephla
|
-
Liothrips urichi
|
|
Formicidae
|
Wasmannia
|
Auropunctata
|
Selenothrips rubrocinctus
|
|
Mesostigmates
|
Phytoseiidae
|
Amblyseius
|
Addoensis
Andersoni Stipulatus Swirskii
|
Heliothrips syvanus
Frankliniella occidentalis, Thrips tabaci
Scirtothrips citri
Thrips tabaci, Retrithrips syriacus
|
|
Typhlodromus
|
Pyri
Athiase
Occidentalis Waltersi
|
Drepanothrips reuteri Retrithrips syriacus Retrithrips
syriacus Frankliniella occidentalis
|
Source: Van Rijn (2002) et Agboton (2004)
·. Résistance
variétale
Des travaux d'amélioration génétique
développés par l'IITA et INRAB ont permis d'identifier du
matériel résistant aux thrips et d'autres ravageurs (IITA, 1979;
INRAB, 1995). Le tableau 2 présente quelques variétés
résistantes de niébé. IITA (op cite) a
développé des cultivars tels TVu 1509, 2870, 6507, 7133,
remarquables quant à leur degré de résistance. IITA (1982)
suggéra que cette résistance est due à une antibiose
contenue dans le niébé. Les cultivars à floraison
très précoce sont utilisés dans la lutte contre les thrips
parce qu'ils échappent en partie aux attaquent de ces thrips.
|
|


