|
UNIVERSITE DE KISANGANI
FACULTE DES SCIENCES
AGRONOMIQUES
Option « Eaux et Forêts »

BP 2012 KISANGANI
Activité cambiale et croissance de
Pericopsis elata (Harms) Van Meeuwen:
inventaire forestier et
analyse diachronique d'échantillons de la zone
cambiale dans les
Réserves de Yoko et de Yangambi
(Province Orientale/RDC)
Ir. ONOTAMBA KONDJO Patrick
Mémoire
Présenté en vue de l'obtention du titre de
Diplômé d'Etude Supéieure en Aménagement durable des
forêts
Promoteur : Professeur Honorine NTAHOBAVUKA
(Université de Kisangani)
Co-promoteur : Docteur Ir. Hans BEECKMAN
(Musée Royal de l'Afrique Centrale)
Co-promoteur : Professeur Charles De CANNIERE
(Université Libre de Bruxelles)
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Table des matières
Table des matières i
Liste des tableaux iv
Liste des figures v
Liste des Annexes vi
Liste des abréviations vi
Dédicace vii
Remerciements vii
Résumé ix
Abstract x
|
1.
|
CONTEXTE ET INTRODUCTION
1.1. Problématique
1.2. Généralités sur Pericopsis
elata (Harms) Van Meeuwen
|
1
1
2
|
|
1.2.1.
|
Description botanique
|
2
|
|
1.2.2.
|
Aire de distribution
|
2
|
|
1.2.3.
|
Ecologie de l'espèce
|
3
|
|
2.
|
HYPOTHESE ET OBJECTIFS
|
4
|
|
2.1. Hypothèse
|
4
|
|
2.2. Objectifs
|
4
|
|
2.3. Intérêt du travail
|
4
|
|
3.
|
MATERIELS ET METHODES
|
5
|
|
|
3.1. Sites
|
5
|
|
3.1.1.
|
La Réserve Forestière de la Yoko
|
5
|
|
3.1.2.
|
La Réserve de Biosphère de Yangambi
|
7
|
|
3.1.2.1.
|
Le Bloc d'aménagement
|
9
|
|
3.1.2.2.
|
Le Bloc Ole
|
10
|
|
3.2. Matériels
|
10
|
|
3.2.1.
|
Matériels techniques
|
10
|
|
3.2.1.1.
|
Matériels pour l'étude de la phytodiversité
|
10
|
|
3.2.1.2.
|
Matériels pour l'étude de l'activité
cambiale
|
11
|
|
3.2.1.3.
|
Matériels pour l'étude de la dendrochronologie
|
11
|

i
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
|
3.2.2. Matériels biologiques
3.3. Méthodes
|
12
13
|
|
3.3.1.
|
Caractérisation du peuplement de Pericopsis elata
|
13
|
|
3.3.1.1.
|
Détermination botanique
|
13
|
|
3.3.1.2.
|
Mesure des diamètres de référence
|
13
|
|
3.3.1.3.
|
Positionnement des arbres
|
13
|
|
3.3.1.4.
|
Traitement des données
|
13
|
|
3.3.2.
|
Etude de la croissance de Pericopsis elata
|
15
|
|
3.3.2.1.
|
Activité cambiale
|
15
|
|
3.3.2.2.
|
Dendrochronologie
|
17
|
|
3.3.2.3.
|
Influence des essences accompagnatrice sur la croissance de
Pericopsis
|
|
|
elata
|
20
|
|
|
4.
|
RESULTATS
|
22
|
|
4.1. Caractérisation du peuplement de Pericopsis
elata
|
22
|
|
4.1.1.
|
Phytodiversité du Bloc d'aménagement
|
22
|
|
4.1.1.1.
|
Richesse spécifique aréale
|
22
|
|
4.1.1.2.
|
Abondance relative
|
22
|
|
4.1.1.3.
|
Dominance relative
|
23
|
|
4.1.1.4.
|
Structure diamétrique spécifique de Pericopsis
elata
|
23
|
|
4.1.2.
|
Distribution spatiale de Pericopsis elata
|
24
|
|
4.2. Etude de la croissance de Pericopsis elata
|
27
|
|
4.2.1.
|
Activité cambiale
|
27
|
|
4.2.2.
|
Dendrochronologie
|
31
|
|
4.2.2.1.
|
Le cross-dating
|
31
|
|
4.2.2.2.
|
Mesure de l'épaisseur des cernes de croissance
|
32
|
|
4.2.2.3.
|
Estimation de la productivité ligneuse
|
32
|
|
4.2.2.4.
|
Corrélation avec le climat
|
33
|
|
4.2.3.
|
Influence des grandes essences sur la croissance de
Pericopsis elata
|
35
|
|
5.
|
DISCUSSIONS
|
36
|
|
5.1. Caractérisation du peuplement de Pericopsis
elata
|
36
|
ii
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
5.1.1. Phytodiversité du bloc d'aménagement 36
5.1.1.1. Richesse spécifique aréale 36
5.1.1.2. Abondance et dominance relative 36
5.1.2. Distribution spatiale de Pericopsis elata 37
5.2. Etude de la croissance de Pericopsis elata 37
5.2.1. Activité cambiale 37
5.2.2. Dendrochronologie 37
5.2.2.1. Largeur des cernes de croissance 37
5.2.2.2. Estimation de la productivité ligneuse 38
5.2.2.3. Corrélation avec le climat 38
5.2.3. Influence des grandes essences sur la croissance
Pericopsis elata 38
CONCLUSIONS ET SUGGESSIONS 40
REFERENCES BIBLIOGRAPHIQUES 42
ANNEXES 49
iii
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Liste des tableaux
Tableau 1: Superficies des layons inventoriés dans le Bloc
d'aménagement 13
Tableau 2: Nombre d'échantillons prélevés
par arbres et par récolte pour l'analyse de l'activité cambiale
16
Tableau 3: Description des quatre échantillons pour
l'analyse dendrochronologique 17
Tableau 4: Structure verticale du Pericopsis elata dans
le bloc d'aménagement 24
Tableau 5: Paramètres des calculs de distance au plus
proche voisin 26
Tableau 6: Corrélation entre les paramètres
climatiques et le nombre moyen de cellules dans la zone cambiale 30
Tableau 7: Cross-dating intra-arbres 31
Tableau 8: Cross-dating inter-arbres 31
Tableau 9: Paramètres climatiques contribuant le plus
à largeur de cernes 33
Tableau 10: Indice d'influence des grandes essences sur la
croissance du Pericopsis elata 35
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Liste des figures
Figure 1: Aire de distribution de Pericopsis elata 3
Figure 2: Localisation de la zone d'étude de la
Réserve Forestière de la Yoko 6
Figure 3: Diagramme Ombrothermique de la RFY de 1951 à
2006 7
Figure 4: Diagramme Ombrothermique de la RBY de 2000 à
2009 8
Figure 5: Nombre de jours de pluies de 2000 à 2009 9
Figure 6: Localisation de la zone d'étude de la
Réserve de Biosphère de Yangambi 10
Figure 8: Vue macroscopique montrant les limites de cernes de
croissance de Pericopsis elata
12
Figure 7: Vue microscopique du bois de Pericopsis elata
12
Figure 9: Coupe des écorces-bois en cube de 1 cm3
16
Figure 10 : Microtome portatif 17
Figure 11: Rondelles avec 11 rayons de comptage 18
Figure 12: Mesurage de l'épaisseur des cernes à
l'aide du programme TSAP Win 19
Figure 13: Adondance relative des espèces du Bloc
d'Aménagement 22
Figure 14: Dominance relative des espèces du Bloc
d'aménagement 23
Figure 15: Structure diamètrique de Pericopsis elata
23
Figure 16: Représentation spatiale des pieds de
Pericopsis elata dans le Bloc d'aménagement
de RBY 25
Figure 17: Vue microscopique de la zone cambiale de
Pericopsis elata 27
Figure 18: nmc dans la zone cambiale par récolte pour le
premier arbre 27
Figure 19: nmc dans la zone cambiale par récolte pour le
deuxième arbre 27
Figure 22: nmc dans la zone cambiale de quatre arbres 28
Figure 20: nmc dans la zone cambiale par récolte pour le
quatrième arbre 28
Figure 21: nmc dans la zone cambiale par récolte pour le
troisième arbre 28
Figure 23: Moyenne mensuelle des cellules produites dans la zone
cambiale 29
Figure 24: Corrélation entre le nombre de cellules dans la
zone cambiale et la précipitation 29
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Figure 25: Corrélation entre le nombre de cellules dans la
zone cambiale et la température 30
Figure 26: Evolution des largeurs de cernes de croissance au
cours de temps 32
Figure 27: Accroissement courant et moyen annuel (cm) 32
Figure 28: Corrélation entre la largeur des cernes et la
température 34
Figure 29: Corrélation entre la largeur de cernes de
crissance et la précipitation 34
Liste des Annexes
Annexe 1: Fonds floristique du bloc d'aménagement 49
Annexe 2: Nombre des cellules non différenciées par
file radiale et par récolte 59
Annexe 3: Largeur moyenne de cernes de croissance en mm pour les
quatre arbres 59
Liste des abréviations
ACA : Accroissement Courant Annuel
AMA : Accroissement Moyen Annuel
CHP : Circonférence à Hauteur de la Poitrine
CITES :Convention on International Trade in Endangered Species
COMIFAC : Commission des Foréts d'Afrique Centrale
DER : Densité Relative
DHP : Diamètre à Hauteur de la Poitrine
DME : Diamètre Minimum d'Exploitabilité
DOR : Dominance Relative
IFA : Institut Facultaire des sciences Agronomique
INERA : Institut National pour l'Etude et la Recherche
Agronomique
LAREF : Laboratoire des Recherches Forestières de Luki /
Bas -- Congo
MECNEF : Ministère de l'Environnement, de la Conservation
de la Nature, des
Eaux et Forêts
MRAC : Musée Royal de l'Afrique Centrale
RBY : Réserve de Biosphère de Yangambi
RDC : République Démocratique du Congo
RFY : Réserve Forestière de Yoko
vi
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Dédicace
Combien je suis si fier de dédier ce travail de dur
labeur à toi ma très chère fille Patricia
KONDJO
MANUNGA, car ta présence a su me donner de la force et de la
détermination
d'amener à bon port ce fruit d'intellectualisme
et de sagesse.
Merci mon bébé chéri ...
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Remerciements
Ce travail a été un grand parcours. Pour oser
l'entreprendre et surtout, le mener à terme, il nous a fallu beaucoup de
soutien, de l'aide et de la collaboration de nombreuses personnalités.
Mais nous ne saurons cependant, dans les quelques paragraphes ci-après
auxquels les exigences académiques nous astreignent, exprimer toute
notre gratitude à tous ceux qui n'ont cessé de nous encourager,
de nous soutenir et de nous faciliter la tâche.
Notre marque de reconnaissance va plus particulièrement
au Dr. Hans BEECKMAN, qui a accepté de diriger ce travail et qui est
à l'origine de l'idée novatrice de cette étude. Nous
mesurons la confiance qu'il nous a accordée en nous proposant ce sujet,
malgré nos connaissances plutôt modestes en anatomie du bois et en
dendrochronologie. Il a eu un regard scientifique à l'évolution
de ce travail par ses remarques et conseils pertinents.
Nous adressons nos remerciements aux Professeurs Honorine
NTAHOBAVUKA et Charles DECANNIERE respectivement Promoteur et co-promoteur pour
l'encadrement et surtout pour leur façon si gentille de nous faire
comprendre nos erreurs afin de perfectionner notre travail.
Qu'il nous soit permis d'adresser nos remerciements de
manière sincère à Mr Benjamin TOIRAMBE BAMONINGA qui,
malgré ses multiples occupations, a accepté d'orienter et
d'encadrer ce travail. Nous avons trouvé en lui, dès le
début les qualités d'un bon maître, lucide et patient,
rigoureux dans la lecture, remarques et critiques mais aussi et surtout un
homme très attentif à notre désir de savoir. Qu'il trouve
ici la marque de notre reconnaissance.
Nous remercions le Doctorant Prosper SABONGO, Dr. Agathe DIE,
Dr. Claire DELVAUX, Dr. Camille COURALET et Dr. Corneille EWANGO pour leur
encadrement tant sur le plan scientifique que morale.
Notre signe de reconnaissance va également aux
coordonateurs du projet AFORCO, Professeurs Jan BOGAERT, Jean Pierre MATE MWERU
et le doctorant Léon IYONGO WAYAMONGO, respectivement coordonateur,
coordonateur local et coordonateur local adjoint pour leur dévouement
à ce projet qui a concouru à la qualité de notre
formation.
A l'Ir Jean Maron MALOTI responsable du laboratoire des
recherches des écosystèmes forestiers de Luki qui nous a
accueillis et aidés pendant le déroulement de l'analyse de nos
échantillons.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
La collecte des données sur terrain exige toujours un
travail en équipe. Sur ce, nous remercions les techniciens BONDELE
KIBINDA, et Justin YAOLI pour leur aide et disponibilité.
Nous tenons à remercier avec beaucoup d'amour et
d'estime nos parents André KONDJO SHOKO et Annie MANUNGA LOMAME, qui
nous ont été d'un secours incommensurable sur tous les plans.
Vous aviez su nous remonter le moral quand nous étions faibles. Nous
sommes si fiers de vous avoir comme parents et que le Bon Dieu vous rende au
centuple tous ces bienfaits.
A Helena NDINGA KIMPEMBE pour tous les bonheurs et peines
partagés ensemble pendant toute cette formation, pour sa contribution
dans la réflexion de notre mémoire et pour ce beau cadeau que tu
m'as offert, symbole de notre existence en la personne de notre fille Patricia
KONDJO MANUNGA.
A mes collègues du projet AFORCO avec qui nous avions
passé des moments de peines et de joies, nous citons : Jean de Dieu
MALONGOLA, Jean Fiston MIKWA, John TSHIBAMBA, Joseph ADHEKA, Joseph OMATOKO,
Josias BONYOMA, Mathie NKUMA, Nathalie NGALYA, Nellas BAUMA, Prosper DECHUVI,
le feu Valentin LOBELA et particulièrement ceux avec qui nous
étions si proche et avec qui nous avions partagé des souvenirs
inoubliables : nous pensons à Helena NDINGA, Jean-Pierre Pitchou MENIKO
TO HULU, Albert ANGBONGA, Aimé MOTONDO et Maurice NGEMALE.
A toute la famille KONDJO, pour votre marque d'affection, par vos
appels téléphoniques, vos prières et conseils. Que ce
travail soit pour vous un model d'excellence.
Nous ne pouvons clore ces mots de remerciement sans penser
à la personne qui nous est très intime et qui nous a surtout
soutenu par la prière et par des réconforts moraux sans lesquels
nos efforts seraient vains, Sandra MALENGA KILOZO affectueusement appelé
Cindy. Qu'elle s'estime heureuse d'avoir marqué nos esprits.
Patrick ONOTAMBA KONDJO
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Résumé
Pericopsis elata est une essence de forte valeur
commerciale recherché pour son bois. Il est aujourd'hui
considéré comme espèce en danger en RDC oü des
stratégies d'assistance à la régénération
naturelle et au reboisement spécifique sont peu
développées. La connaissance de sa vitesse de croissance et de la
structure de sa population permettraient de fixer de manière
précise le DME (Diamètre minimum d'exploitabilité) et
favoriseraient la planification de l'exploitation de cette essence sans risque
de compromettre sa pérennité.
L'objectif principal du présent travail est
d'évaluer la dynamique d'une population de Pericopsis elata
dans les conditions climatiques de la zone équatoriale. Deux sites ont
été choisis : la Réserve Forestière de la Yoko et
la Réserve de Biosphère de Yangambi. L'étude avait deux
volets : premièrement une étude de la caractérisation du
peuplement de Pericopsis elata où la phytodiversité a
été réalisée grâce à un inventaire
floristique de tous les individus à dhp = 10 cm présent dans le
bloc d'aménagement de Réserve de Biosphère de Yangambi.
Deuxièmement, sur la croissance du Pericopsis elata par
l'analyse de l'activité cambiale sur des échantillons
écorce-bois prélevées bimensuellement du 5 mai au 20
septembre 2010 dans la plantation (bloc Ole) de la Réserve de
Biosphère de Yangambi, par l'analyse des profils des cernes de
croissance de quatre rondelles prélevées dans une plantation de
la Réserve Forestière d la Yoko (dendrochronologie) et ainsi que
par l'influence des essences accompagnatrices sur la croissance de cette
espèce.
Sur les 10.956 individus répartis dans 229
espèces et 35 familles, Pericopsis elata compte 429 individus
sur une superficie de 21 ha. La famille de Fabaceae est la plus
abondante. Cette espèce présente une distribution
agrégée. La production maximale de cellules cambiales s'observe
en septembre, un ralentissement de la division cellulaire dans la zone cambiale
est observé au mois de juillet L'accroissement moyen annuel en surface
terrière est de 0,06 m2/an soit un accroissement moyen annuel
en diamètre de 0,27 cm/an. Les cernes de croissance sont synchrones. La
croissance de cette espèce est plus liée à la
présence des grandes essences qu'aux paramètres climatiques.
Mots clés : Pericopsis elata, inventaire
floristique, activité cambiale, dendrochronologie, cernes de croissance,
accroissement moyen annuel, Réserve de Biosphère de Yangambi,
Réserve forestière de la Yoko.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Abstract
Pericopsis elata is a species of high commercial
value sought for timber. It is now considered endangered species in the DRC
where strategies to support natural regeneration and reforestation are specific
underdeveloped. Knowledge of its growth rate and structure of its population
would determine accurately the DME (minimum exploitable diameter) and advanced
planning for the operation of the fuel without jeopardizing its survival. The
main objective of this study was to assess the dynamics of a population of
Pericopsis elata in climatic conditions in the equatorial
zone. Two sites were chosen: the Forest Reserve Yoko and Yangambi Biosphere
Reserve. The study was twofold: first, a study of the characterization of the
settlement of Pericopsis elata where plant diversity has been achieved
through a floristic inventory of all individuals = 10 cm dbh present in the
block management of Biosphere Reserve of Yangambi. Second, the growth of
Pericopsis elata by analysis of cambial activity on wood-bark samples
collected bimonthly from May 5 to September 20, 2010 in the plantation (block
Ole) of Yangambi Biosphere Reserve by the profile analysis of growth rings of
four slices taken from a plantation in the Forest Reserve of the Yoko
(dendrochronology) and as well as the influence of species accompanying the
growth of this species.
Of the 10,956 individuals in 229 species and 35 families,
Pericopsis elata has 429 people on an area of 21 ha. The family
Fabaceae is the most abundant. This species has an aggregate
distribution. The maximum production of cambial cells is observed in September,
a slowdown of cell division in the cambial zone is observed in July The annual
average increase in basal area is 0.06 square meter per year or an average
annual increase in diameter of 0.27 cm / year. Growth rings are synchronous.
The growth of this species is more related to the presence of large species
than climatic parameters.
Keywords: Pericopsis elata floristic inventory, cambial
activity, dendrochronology, tree ring growth, average annual growth, Yangambi
Biosphere Reserve, Forest Reserve Yoko.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
1. CONTEXTE ET INTRODUCTION
1.1. Problématique
Les forêts tropicales disparaissent à une vitesse
alarmante et il est important de développer des techniques pour
s'assurer de la conservation de leur écosystème. La gestion ou la
conservation de ces foréts dépende d'une multitude de
critères biologiques. Beaucoup de ces critères découlent
de l'estimation de la biodiversité et impliquent la connaissance de la
composition d'espèces. Il s'agit d'un réel challenge en
forét tropicale humide, car cette dernière à une
biodiversité très riche et une forte densité (Quentin,
2010).
Cependant ces forêts subissent des pressions qui sont de
plus en plus forte et se caractérisent par une diminution de ces
formations forestières. En RDC, ses forêts naturelles font l'objet
d'une exploitation du type sélectif qui consiste à
prélever uniquement les arbres de quelques essences dont la valeur
commerciale justifie leur recherche et elle ne s'accompagne d'aucune mesure en
faveur du renouvellement des effectifs exploités. Pourtant, la
valorisation de ces essences précieuses est essentielle car elle
constitue la meilleure garantie pour le maintien de la capacité de
charge des populations pour une exploitation forestière durable, voire
un maintien à long terme de grands massifs forestiers. Pericopsis
elata est l'une de ces essences de forte valeur commerciale
recherché pour son bois. Il est aujourd'hui considéré
comme espèce en danger en RDC oü des stratégies d'assistance
à la régénération naturelle et au reboisement
spécifique sont peu développées. La connaissance de sa
vitesse de croissance et la structure de sa population permettraient de fixer
de manière précise le DME (Diamètre minimum
d'exploitabilité) et favoriseraient la planification de l'exploitation
de cette essence sans risque de compromettre sa pérennité.
En outre, les changements planétaires (changements
climatiques, augmentation du taux de CO2 atmosphérique et augmentation
des dépôts azotés) sont susceptibles d'avoir un effet sur
la production des écosystèmes forestiers. La dendrochronologie
qui est une science utilisant les fluctuations de la croissance des arbres dans
le temps (Magnin, 2001) et une des méthodes de datation permettant de
déterminer la durée de vie d'un arbre et de préciser
l'année et la saison de son abattage, offre un moyen de vérifier
cette hypothèse en milieu naturel, car les données de croissance
radiale peuvent être interprétées comme des indicateurs de
la productivité des écosystèmes forestiers.
1

Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
1.2. Généralités sur Pericopsis
elata (Harms) Van Meeuwen
1.2.1. Description botanique
Les descriptions botaniques du Pericopsis elata sont les
suivantes :
(i) Famille : Fabaceae sous famille de Faboideae
(ii) arbre de taille moyenne pouvant atteindre 45 à 50 m de haut
(iii) fût dépourvu de branches jusqu'à une hauteur de 30
à 35 m, rectiligne et cylindrique, parfois tortueux, jusqu'à 130
à 150 cm de diamètre, souvent avec des contreforts bas et obtus ;
surface de l'écorce crème ou grisâtre, s'écaillant
en fins morceaux laissant des taches rouge-brun, écorce interne
jaunâtre, fonçant à l'orange sale (iv) cime aplatie, a
branches massives, étalées (v) rameaux pendants, glabres (vi)
Feuilles alternes, composées imparipennées a 5-7-9 parfois 11
folioles ; stipules linéaires, d'environ 7 mm de long, caduques ;
pétiole et rachis mesurant ensemble 7 a 20 cm de long,
légèrement sillonnés sur la face supérieure ;
stipelles filiformes, de 2 a 5 mm de long, persistantes ; pétiolules de
3-7 mm de long ; folioles alternes, elliptiques a ovales-elliptiques, de 3-8 cm
× 1,5-3,5 cm, la terminale plus grande, légèrement
cunéiformes a arrondies a la base, courtement acuminées à
l'apex, glabres, pennatinervées a 5 a 11 paires de fines nervures (vii)
Inflorescence : panicule mince, retombante, atteignant 12 cm de long à
l'extrémité d'une pousse, a pubescence clairsemée ;
bractées jusqu'à 4 mm de long. Fleurs bisexuées,
papilionacées, blanches a crème ou verdâtres ;
pédicelle de 5 a 9 mm de long, mince ; calice campanulé, de 6 a
12 mm de long, a grands lobes triangulaires, finement poilus ; corolle a
étendard presque circulaire de 11 a 13 mm de diamètre, a onglet,
pétales des ailes et de la carène d'environ 15 mm de long ;
étamines 10, libres, de 9 a 19 mm de long, glabres ; ovaire
supère, aplati, de 5 a 6 mm de long, pubescent, style mince,
recourbé. (viii) Fruit : gousse oblongue-linéaire, aplatie de 7 a
17 cm × 2,5-3 cm, brièvement stipitée, vaguement
ailée sur les bords, brune, lisse, a nervures réticulées,
indéhiscente, contenant 1 a 4 graines (ix) Graines discoïdes, de 12
a 15 mm de diamètre, brunes (x) Plantule a germination
épigée ; hypocotyle de 1 a 2 cm de long, épicotyle de 4 a
5 cm de long ; cotylédons ovales, d'environ 12 mm de long,
légèrement charnus ; 2 premières feuilles opposées,
simples, les suivantes alternes et 3 foliolées.
1.2.2. Aire de distribution
D'après Fouarge et Louis (1943) l'espèce fut
découverte par Milbread, en 1911 dans la forêt équatoriale
primitive au sud du Cameroun. En suite, on l'a reconnue au Nigeria et en Cote
d'ivoire (Minef, 2002 ; Kabala et al. ,2008). Pericopsis elata est
répertorié dans 6 pays

Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
africains à savoir : le Cameroun, la RCA, la
République du Congo, la Côte d'Ivoire, le Ghana, le Nigeria et la
RDC (figure 1).
Mais c'est en RDC oü on possède les derniers
grands peuplements de Pericopsis elata (sous l'appellation
d'Afrormosia, Mogoya ou Olé). Sa distribution est disjointe,
limitée, mais bien dispersée ; il est concentré dans les
forêts semi-sempervirentes de Kisangani - Banalia -- Yangambi et de
Lisala -- Bongandanga.
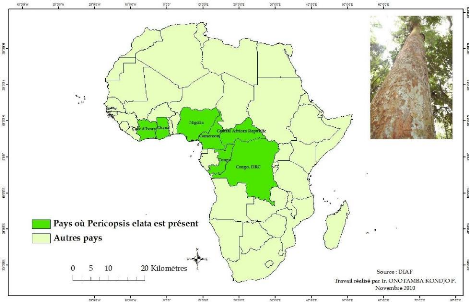
Figure 1: Aire de distribution de Pericopsis
elata
1.2.3. Ecologie de l'espèce
Pericopsis elata est caractéristique de la
forêt semi-décidue, en particulier dans les endroits
marécageux, plats et perturbés. Il est présent dans des
régions où la pluviométrie annuelle est de 1000-1500 mm.
Espèce héliophile, caractéristique de l'alliance
oxystigmoscorodophloeion (Lebrun et Gilbert, 1954). Les tiges de
Pericopsis elata croissent en bouquet ou en tache de peuplement
monospécifique quasiment purs et elles dominent suivant leur habitat,
tout en acceptant les autres espèces dans son développement
(Lebrun et Gilbert, 1954). En forêt naturelle, la densité des
tiges de Pericopsis elata diminue au fur et à mesure que le
peuplement vieillit. En raison de la densité de la forét, de
l'étagement des strates arborées et de la présence
permanente des feuilles ; la lumière y pénètre
difficilement (Boyemba, 2006 ; Bourland ,2008).

Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
2. HYPOTHESE ET OBJECTIFS
2.1. Hypothèse
L'hypothèse fondamentale découlant de cette
problématique est la suivante :
La croissance en diamètre du Pericopsis elata en
zone équatoriale dépendrait de :
a. Facteurs climatiques : précipitation et
température ;
b. L'influence des essences accompagnatrices.
2.2. Objectifs
L'objectif principal du présent travail
est d'évaluer la dynamique d'une population de Pericopsis elata
dans les conditions climatiques de la zone équatoriale.
Pour y parvenir, les objectifs spécifiques
ci-après ont été fixés :
a. connaître la phytodiversité du bloc
d'aménagement par la connaissance de la richesse floristique,
l'abondance et la dominance relative des espèces et des familles, la
surface terrière, l'indice d'importance relative et la structure
diamétrique spécifique de Pericopsis elata ;
b. Etudier la distribution spatiale de Pericopsis elata
dans le bloc d'aménagement de la RBY pour arriver à :

regrouper les Codominants et les dominés autour des
émergents et les Dominants ;
c. 
Etudier la périodicité de croissance du
Pericopsis elata par l'évaluation de : son activité
cambiale (nombre des cellules produites au cours de temps),
la fréquence (saisonnalité) de la formation des
cellules cambiales;
d. Etudier la dendrochronologie par l'analyse des profils des
cernes ;
e. Etudier l'influence des grandes essences telles que
Entandrophragma, Erythrophleum, Petersianthus macrocarpus etc. sur la
croissance de Pericopsis elata.
2.3. Intérêt du travail
L'intérêt que revét ce travail est de
fournir les informations nécessaires sur la vitesse de croissance et la
structure de la population de Pericopsis elata. Cela permettrait de
mettre sur pied une politique de gestion et de conservation durable.

Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
3. MATERIELS ET METHODES
3.1. Sites
Le choix des sites retenu pour cette étude est lié
aux objectifs et aux contraintes administratives :



L'impact des résultats sur la phytodiversité et
la distribution spatiale sont mieux exprimés dans une forêt
naturelle que dans une plantation ; de ce fait elles ont été
étudiées dans le Bloc d'aménagement de la RBY ;
La dendrochronologie n'a pas pu être
étudiée dans la RBY indépendamment de notre bonne
volonté. En effet l'abattage des pieds de Pericopsis elata est
strictement interdit dans cette Réserve. Nous nous sommes donc rabattu
dans une plantation de Pericopsis elata de la RFY après
négociation.
Il nous a semblé important d'étudier
l'activité cambiale dans une plantation (Bloc olé/RBY) que dans
la forêt naturelle pour la simple raison que Pericopsis elata
est aujourd'hui listé dans l'annexe II de la CITES ; promouvoir les
études de cette espèce dans une plantation fournirait des
informations très capitales dans la pérennisation et la prise de
décision par les autorités politico-administratives.
3.1.1. La Réserve Forestière de la Yoko
La Réserve Forestière de la Yoko est
délimitée au Nord par la ville de Kisangani et une bande des
foréts fortement dégradées, au Sud et à l'Est par
la rivière Biaro qui forme une demi-boucle en suivant cette direction,
à l'Ouest par la voie ferrée et la route le long de laquelle elle
se prolonge des points kilométriques 21 à 38 (Lomba & Ndjele,
1998).
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Elle est située dans le district de la Tshopo, dans le
territoire d'Ubundu et dans la collectivité Bakumu-Mangongo. Elle est
baignée par la rivière Yoko qui la subdivise en deux parties dont
la réserve nord avec 3.370 ha et la réserve sud avec 3.605 ha,
soit une superficie globale de 6.975 ha. Elle a comme coordonnées
géographiques : longitude Nord : 00° 29' 40,2», latitude Est -
Quest : 25° 28' 90,6» et une altitude moyenne de : 430 m (Figure
4).
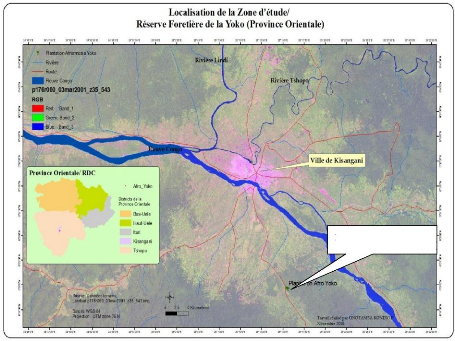
Plantation YOKO
Figure 2: Localisation de la zone d'étude de la
Réserve Forestière de la Yoko
6

Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
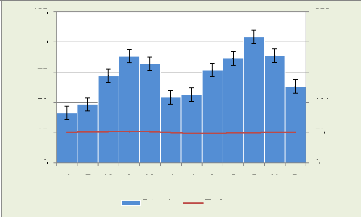
Les conditions climatiques de la Réserve Forestière
de la Yoko sont consignées dans la figure 5 ci-dessous :
T °C moy
125
100
25
75
50
0
J F M A M J JASON D
P (mm) T° C moy
250
200
0
50
150
100
P moy (mm)
Figure 3: Diagramme Ombrothermique de la RFY de 1951
à 2006
En tenant compte des irrégularités dans le
prélèvement des données climatiques de la réserve
et en suivant sa situation à la périphérie de Kisangani,
la Réserve de Yoko bénéficie globalement du climat
régional de la ville de Kisangani du type Af, de la classification de
KÖPPEN (Ifuta, 1993).
3.1.2. La Réserve de Biosphère de
Yangambi
La Réserve de Biosphère de Yangambi est
située administrativement dans les territoires d'Isangi et de Banalia,
dans le district de la Tshopo, à l'Ouest de la ville de Kisangani, dans
la province Orientale, au Nord-Est de la RDC et correspond à une
étendue comprise entre 24°16' et 25°08' de longitudes Est et
0°38' et 1°10' latitudes Nord, avec des altitudes variables entre 400
et 500m. Elle est traversée par un réseau hydrographique
important dont la rivière Lobilo forme le principal bassin avec ses
multiples affluents qui, ensemble, se jettent dans le fleuve Congo.
Situation climatique
La RBY se situe dans la zone influencée par le climat du
type Af de Köppen (Bultot, 1977) oü il pleut pratiquement toute
l'année.

Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
L'analyse des données météorologiques de
la station d'agroclimatologie de l'INERA Yangambi (figure 6 et 7) pour la
période de 2000 à 2009 montre que la moyenne maximale annuelle
des températures est de 31,1°C, la minimale de 20°C et la
moyenne de 25,7°C. Les trois gammes de températures restent presque
constantes toute l'année, ne montrant que de très faibles
amplitudes. Leur tendance générale est à la baisse pendant
le second semestre de l'année ; ce qui confirme que la grande saison
sèche se manifeste vers fin et le début de l'année
(décembre, janvier et février), alors que la petite saison
sèche est à peine esquissée au second semestre (juin et
juillet).
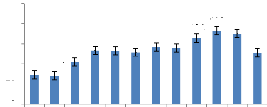
Les totaux des moyennes mensuelles des précipitations
et du nombre de jours de pluies sont respectivement de 1776 mm et de 180 jours.
Globalement il pleut en moyenne une fois tous les deux jours dans la RBY,
excepté les mois de janvier et de février. Les deux moyennes
augmentent pendant le deuxième semestre de l'année, ce qui
indique que la petite saison des pluies se manifeste au premier semestre (avril
et mai) et la grande saison au deuxième (août à novembre).
Cette répartition est confirmée par la tendance à la
baisse des températures notées pendant le deuxième
semestre de l'année. Les plus faibles pluies sont observées en
janvier, février et juin de chaque année ; et les plus abondantes
pluies en octobre.

T°C moy ; Bares d'erreur
125
100
25
75
50
0
J F M A M J J A S O N D
P (mm) T° C moy
250
200
300
50
0
150
100
P (mm)
Figure 4: Diagramme Ombrothermique de la RBY de 2000
à 2009
Source : Station agrométéorologique de
l'INERA Yangambi
Il en va presque de même en ce qui concerne le nombre de
jours de pluies : il pleut pendant moins de jours en janvier et février
et plus de jours en octobre et novembre. Cette

Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
manifestation des pluies en deux périodes de
l'année pourrait influencer profondément la phénologie et
la croissance de la végétation.
250
164
133 132 129 142 140
104
72 69,7
182 175
127
0
200
150
100
50
|
Janvier
Ryder
|
Mars
Avril
Mai
Jui
n
Juillet
Aoirt
Septembre
Octobre
Novembre
Decembre
|
Figure 5: Nombre de jours de pluies de 2000 a
2009
Source : Station agrométéorologique de l'INERA
Yangambi
L'étude sur la phytodiversité et l'analyse de
l'activité cambiale se fait respectivement dans les blocs
ci-après :
3.1.2.1. Le Bloc d'aménagement
La forét d'aménagement se trouve sur le plateau
Yangambi. Ce dernier est situé à une distance d'environ 100 Km de
Kisangani, au Nord-Est, à quelques 8 Km de L'Institut Facultaire des
Sciences Agronomiques de Yangambi (I.F.A.). Son altitude se rapproche de 500 m
au-dessus du niveau de la mer. Ses coordonnés géographiques sont
les suivantes longitude 24°28'36,4» et latitude
0°48'13,0».
Cette forêt fait partie de la réserve
forestière du Groupe forestier de l'INERA. Sur le plan hydrographique,
cette forêt est située dans le bassin de la rivière
Lusambila et de la rivière Boombo. Elle couvre une superficie de 255 ha
; cette superficie a été calculée à l'aide du
logiciel Arc GIS 9.2 sur base des données récolté sur
terrain (figure 8). Elle se trouve à environs 30 m à l'Ouest de
la route qui mène au point kilométrique 5.
C'est une forét primaire dominée par
Scorodophleus zenkeri Harms, à l'intérieur duquel 20
layons ont été ouverts.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
3.1.2.2. Le Bloc Ole
Il couvre 24,20 ha. C'est une parcelle qui a été
soumis d'un enrichissement par layon depuis 1940 (figure 8).
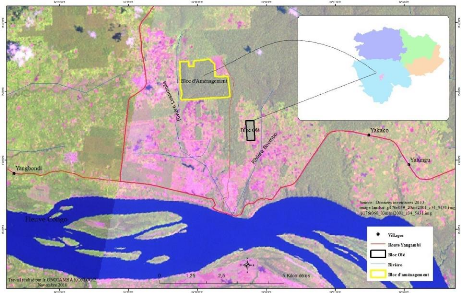
Figure 6: Localisation de la zone d'étude de la
Réserve de Biosphère de Yangambi
3.2. Matériels
3.2.1. Matériels techniques
3.2.1.1. Matériels pour l'étude de la
phytodiversité
Pour la récolte de données relative à la
phytodiversité, les équipements ci-après ont
été utilisés :
Un appareil photographique pour prendre des images, une paire
de jumelles pour viser les organes végétaux des arbres
supérieurs aux fins d'identification, un GPS de marque etrex Vista HCx
pour prélever les différents waypoints, cinq machettes pour le
layonnage, deux mètres ruban de 50 m pour chaîner les longueurs et
les largeurs des layons et de 10m pour mesurer les circonférence
à hauteur de poitrine (Chp), les blocs notes et les stylos à
bille/crayons pour noter de données floristiques, deux paquets des
craies marqueurs forestiers, trois bombes à marquage forestier, une lime
pour aiguiser les machettes, une boussole pour orienter les machetteurs,
etc.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
3.2.1.2. Matériels pour l'étude de
l'activité cambiale
Les matériels techniques pour l'étude de
l'activité cambiale sont constitués de blocs écorces-bois,
quatre ciseaux menuisiers et deux marteaux pour prélever les
échantillons, un scalpel, un pinceau, deux pinces, une scie à
bois pour la coupe des échantillons de ces derniers en cube de
1cm3, des lames et lamelles pour fixer les échantillons en
vue de faire des coupes permanentes, une bouteille du baume de canada, une tige
en verre pour déposer le baume de canada sur la coupe, des boites en
plastiques, un système de chauffage (briquet), une camera montée
sur un microscope pour la prise des photos, un microscope.
3.2.1.3. Matériels pour l'étude de la
dendrochronologie
Les matériels ci-après ont servi pour atteindre
les objectifs dendrochronologiques : une tronçonneuse pour la coupe des
arbres, une ponceuse pour polir la surface transversale des rondelles, de
disques de ponçage de rugosités différentes en vue d'une
bonne visibilité des cernes de croissance, une table de mesure
couplée à un microscope (LINTAB) pour le comptage de cernes, un
crayon, une latte et une gomme pour le traçage des rayons de comptage et
une loupe de grossissement 10 x pour visualiser les limites de cernes.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
3.2.2. Matériels biologiques
Quatre specimens de Pericopsis elata
récoltés dans une plantation de 1954 à la RFY dont la
provenance et l'âge ne sont pas connus et 36 échantillons
d'écorce-bois de Pericopsis elata récolté dans
une vieille plantation de 1940 ont constitués nos matériels
biologiques.
En effet, Pericopsis elata est une espèce semi
poreuse c'est-à-dire qu'il y a réduction de taille des vaisseaux
du bois initial au bois final, en coupe transversale, les gommes semblent
envahir complètement le lumen de certains vaisseaux, ces derniers sont
disposés en une association de courtes files radiales de 2 ou 3
vaisseaux associées à des vaisseaux isolés. Les fibres
sont présentes. Les parenchymes axials sont fusiformes et en files
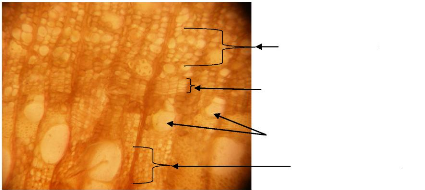
(figure 2). Les limites des cernes de Pericopsis elata sont distinctes
comme le présente la figure 3.
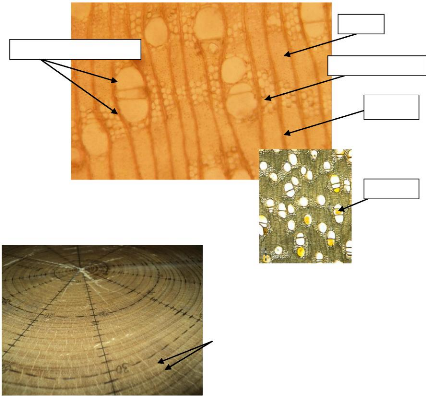
Gomme
Figure 7: Vue microscopique du bois de Pericopsis
elata
Fibres
Vaisseaux accolés (2 ou 3)
Parenchyme
Rayons
Limite d'un cerne de croissance
Figure 8: Vue macroscopique montrant les limites de
cernes de croissance de Pericopsis elata
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
3.3. Méthodes
3.3.1. Caractérisation du peuplement de Pericopsis
elata
Les données récoltées pour la
caractérisation du peuplement de Pericopsis elata portent sur
l'identification botanique, la mesure du diamètre à hauteur de
poitrine de tous les arbres de dhp = 10 cm ainsi que la localisation des pieds
de Pericopsis elata.
3.3.1.1. Détermination botanique
La détermination botanique se fait de façon fine
pour tous les arbres présents, avec prélèvement
d'échantillons pour chaque espèce non identifiée sur
terrain. Cette détermination s'est faite en s'appuyant sur les
connaissances des indicateurs de l'INERA Yangambi.
3.3.1.2. Mesure des diamètres de
référence
Le diamètre minimum d'inventaire a été
fixé à 10 cm conformément aux exigences fixé par la
COMIFAC. Le diamètre de référence des arbres vivants de
plus de 10 cm de dhp a été mesuré, à l'aide d'un
ruban souple métallique.
Un trait de peinture a été marqué à
l'emplacement exact oil le mètre-ruban a été
positionné sur le tronc lors de la prise des mesures.
3.3.1.3. Positionnement des arbres
Deux méthodes peuvent titre utilisées pour
localiser les arbres au sein des placeaux : la méthode classique,
utilisant la boussole et les décamètres ou chaînes
d'arpenteur et la méthode par triangulation, utilisant un
télémètre laser. C'est la première méthode
qui a été utilisée dans le cadre de ce mémoire.
3.3.1.4. Traitement des données
(a) Phytodiversité
La récolte de données sur la phytodiversité
a reposé sur quatre layons dont les écartements et les
superficies sont présentés dans le tableau 1
Tableau 1: Superficies des layons inventoriés
dans le Bloc d'aménagement
Layons Dimension Superficie
(ha)
Layon 1 800 m x 50 m 4
Layon 5 800 m x 50 m 4
Layon 10 1300 m x 50 m 6,5
Layon 15 1300 m x 50 m 6,5
Les paramètres d'analyse présentés dans le
présent mémoire sont essentiellement ceux caractérisant la
structure d'un peuplement forestier à savoir la richesse
spécifique,
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
l'abondance relative, la dominance relative, l'indice
d'importance relative et la structure diamétrique.
(i) Richesse spécifique aréale
La richesse aréale indique le nombre d'espèces
par unité de surface. C'est l'une des mesures les plus communes de la
biodiversité. Elle indique le nombre d'espèces recensées
par unité de surface (Monot 1955, Walker 1992 et 1995).
(ii) Fréquence relative des taxons
La fréquence d'un taxon (FRR) est exprimée en
considérant le nombre de relevés où le taxon sur la somme
de relevé de tous les taxons présents. La formule permettant de
calculer la fréquence relative d'un taxon est la suivante :
FRR
FRR = FRR × 100
(iii) Abondance des taxons
Elle tient compte du nombre d'individus dans une
espèce ou dans une famille, sans tenir compte de sa taille. L'abondance
d'un taxon est donnée par la place que celui-ci occupe sur la liste
décroissante de densité relative de tous les taxons.
(iv) Dominance des taxons
La dominance des taxons permet de visualiser le mieux la
situation de la forêt car elle met en évidence les espèces
qui occupent le plus de place dans la forêt. Elle tient compte de la
taille des individus par la prise en compte de leur surface terrière.
La surface terrière (basal area) est la surface
occupée par les troncs à hauteur de poitrine. Elle est
exprimée en m2/ha. La surface terrière individuelle
est notée g et est donnée
C2
par la formule ci-après : g = 4ð Les arbres
étant considérés comme cylindriques et
leurcirconférence étant notée C.
La dominance relative d'un taxon exprime la proportion de la
surface terrière du taxon (espèce ou famille) par rapport
à la surface terrière totale.
Surface terrière du taxon × 100
Dominance relative du taxon =
Surface terrière totale
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
(v) Densité relative
La densité est un descripteur de base de l'état
de la parcelle et un indice simple de la compétition moyenne dans le
peuplement. Elle est définie comme le nombre total de tiges par
unité de surface.
La formule permettant de calculer la densité relative
d'un taxon est la suivante :
Nombre d'individusdu taxon
Densité relative d'un taxon = × 100
Nombre total d individusdans l' echantillon
(vi) Importance relative des taxons
L'importance relative a été calculée par la
formule : IVI =FRR+DER+DOR avec FRR comme Fréquence relative DER
Densité relative et DOR Dominance relative.
(vii) La structure diamétrique
spécifique
C'est la répartition des tiges par classes de
diamètre d'une espèce. Pour des raisons pratiques, des classes de
taille constante de largeur de 10 cm ont été choisi.
(b) Distribution spatiale
Pour ce paramètre, nous avons tenu compte de tous les
pieds de Pericopsis elata dans le 255 ha.
(i) Localisation des pieds
Grâce aux coordonnées cartésiennes (X, Y)
prélevées sur terrain, une carte de l'emplacement de chaque pied
a pu etre élaborée.
(ii) Analyse de la distribution spatiale
Nous avons utilisé la méthode de Clark et Evans oil
l'on calcul la distance du plus proche voisin. Cette méthode est
décrite par Nshimba (2008).
3.3.2. Etude de la croissance de Pericopsis elata
3.3.2.1. Activité cambiale
L'évaluation de l'activité cambiale de
Pericopsis elata, a nécessité le prélèvement
des blocs écorce-bois contenant la zone cambiale. Au total 36
échantillons ont été prélevés dans quatre
arbre en raison d'un bloc écorce-bois pour chaque arbre et à
chaque récolte. Le prélèvement de ces échantillons
s'est fait entre 1 à 1,30m au-dessus du sol et pendant une
période allant du 5 mai au 20 septembre 2010 (tableau 2). La dimension
de l'échantillon a été
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
suffisamment grandes (8 cm de long, 4 cm de large et 4 cm
d'épaisseur) pour éviter que l'écorce ne se détache
du bois (ce qui ferait perdre la zone cambiale).
Tableau 2: Nombre d'échantillons
prélevés par arbres et par récolte pour l'analyse
de
l'activité cambiale
|
Arbre N°1
(dhp en cm)
|
Arbre N°2
(dhp en cm)
|
Arbre N°3
(dhp en cm)
|
Arbre N°4
(dhp en cm)
|
|
Date de récolte
|
34
|
46
|
|
55
|
72
|
|
5 Mai 2010
|
1
|
1
|
|
1
|
1
|
|
5 juin 2010
|
1
|
1
|
|
1
|
1
|
|
20 juin 2010
|
1
|
1
|
|
1
|
1
|
|
5 juillet 2010
|
1
|
1
|
|
1
|
1
|
|
20 juillet 2010
|
1
|
1
|
|
1
|
1
|
|
5 Aaoût 2010
|
1
|
1
|
|
1
|
1
|
|
20 Août 2010
|
1
|
1
|
|
1
|
1
|
|
5 septembre 2010
|
1
|
1
|
|
1
|
1
|
|
20 septembre 2010
|
1
|
1
|
|
1
|
1
|
|
Total
|
|
|
36
|
|
|
Les échantillons prélevés ont
été immergés dans une solution de boisson traditionnelle
alcoolisée de forte concentration (Lotoko) faute de FAA
(Formaldéhyde-Acide acétique-Alcool) et conservés dans un
bocal hermétiquement fermé. Les coupes microscopiques de 15
à 20 um sont faites suivant les étapes ci-après :
1. la coupe des écorces-bois pour en faire des cubes de 1
cm3 (Figure 9)

Scie
Fixateur
Bloc écorce-bois
Cambium
N° Echantillon
Ecorce
Bois
Figure 9: Coupe des écorces-bois en cube de 1
cm3
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
2. pour chaque cube, faire des coupes de 15-20 um à
l'aide d'un microtome portatif (Figure 10)

Figure 10 : Microtome portatif
3. Fixer les coupes sur la lame et lamelle pour faire des coupes
permanentes.
L'analyse de ces échantillons a été faite au
Laboratoire des Recherches Forestières de Luki / Bas -- Congo, LAREF en
sigle.
(a) Comptage des cellules cambiales par file
radiale
Le nombre de cellules cambiales a été
déterminé sur 30 files radiales pour chaque arbre et pour chaque
récolte. Le comptage s'est fait dans le sens moelle-écorce
(au-dessus du xylème secondaire jusqu'en dessous du phloème.
3.3.2.2. Dendrochronologie
(a) Prélèvement des rondelles et analyse
des profils des cernes
Les rondelles de Pericopsis elata ont
été récoltées dans la Réserve
Forestière de la Yoko. A l'aide d'une tronçonneuse, on effectue
des coupes transversales dans le tronc d'un pied (spécimen) pour obtenir
des rondelles. Par cette méthode, nous avons prélevé 4
rondelles sur 4 arbres c'est-à-dire une rondelle par arbre, à 60
cm du sol (tableau 3).
Tableau 3: Description des quatre échantillons
pour l'analyse dendrochronologique
|
Date d'abattage
|
Année de la
plantation
|
Code
échantillon
|
Diamètre moyen (cm)
|
Hauteur fût
(m)
|
Coordonnées géographiques.
|
|
24-mai-2010
|
1954
|
Y1
|
38,28
|
19,25
|
N 00° 19' 37,2" E 025°15'28,7"
|
|
24-mai-2010
|
1954
|
Y2
|
39,01
|
21,78
|
N 00° 19' 39,5" E 025°15'27,2"
|
|
24-mai-2010
|
1954
|
Y3
|
63,42
|
22
|
N 00° 19' 35,3" E 025°15'35,1"
|
|
24-mai-2010
|
1954
|
Y4
|
46,32
|
10,79
|
N 00° 19' 35,6" E 025°15'36,3"
|
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
(b) Préparation de la surface de la rondelle
à observer
Pour préparer la surface de la rondelle à
observer, cette dernière a été immobilisé à
l'aide d'un support sur une table ordinaire qu'on polit une des 2 faces avec le
disque de plus grande rugosité (50), jusqu'au nivelage complet de
celle-ci.
Des disques de rugosités décroissantes (80 ;
120; 220 ; 320 ; 500 et 800) sont successivement utilisés pour obtenir
l'absence totale de dénivellation sur cette face afin de rendre plus
visibles les cernes de croissance sur la rondelle. La qualité de cette
préparation est importante car c'est d'elle que dépendra la
visibilité des limites des zones d'accroissements parfois très
discrètes chez certaines essences (Detienne et al., 1998
cité par Die, 2004).
(c) Analyse de la surface
préparée

Afin de réduire les effets des
irrégularités diamétriques, plusieurs rayons de comptage
ont été tracés sur la face ainsi poncée (Figure
11). Néanmoins, seuls 3 à 4 ont été
considérés compte tenu de la forme plus ou moins
régulière des rondelles. Chaque rayon est marqué par le
symbole R suivi d'une lettre soit Ra, Rb, Rc et Rd de la moelle vers
l'écorce.
Figure 11: Rondelles avec 11 rayons de
comptage
(d) Observation microscopique de la surface de la
rondelle
L'observation a été faite grâce à une
loupe binoculaire pour détecter et analyser avec une certitude et
précision la nature des limites de cernes.
(e) Mesures de l'épaisseur des cernes et
synchronisation des profils de cernes de
croissance
Les tests statistiques pour une comparaison des séries
des largeurs de cernes sont effectués grâce au programme TSAP-Win.
Ce dernier permet de faire des mesures des largeurs de cernes, la
standardisation et l'interdatation statistique et visuelle des séries
(Figure 12). Les facteurs analysés dans le cadre de notre travail sont :
GIK (Gleichlaüfigkeit) et TVBP (Student's t-value d'après la
standardisation de Baillie & Pilcher).
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)


GIK indique si la synchronisation entre deux séries au
sein d'un méme arbre est bonne, réussie. Il s'agit d'un test de
Gleichlaüfigkeit (stat G). Le seuil critique pour le GIK est d'environ 60%
(Trouet & al., 2009).
TV-BP : il s'agit d'un test de Student t d'après la
standardisation de Baillie & Pilcher. Ce test est utilisé pour
tester la synchronisation des séries de largeur de cernes de croissance
entre plusieurs arbres pris deux à deux. Le seuil de TV-BP égal
à 2 est accepté pour les espèces tropicales ((Trouet &
al., 2009).
Figure 12: Mesurage de l'épaisseur des cernes
à l'aide du programme TSAP Win (f) Estimation de la productivité
ligneuse en surface terrière
Pour effectuer des études d'analyse de cernes de
croissance, il est préférable de prélever les rondelles
à un niveau qui se rapproche le plus de la base du tronc en vue de
déterminer l'âge de l'arbre. Ce procédé permet
d'éviter les effets (resserrement prononcé des cernes, nombre de
cernes insuffisant anormalement) dus à la décroissance
diamétrique.
Les calculs des accroissements sont été fait sur
base de la surface terrière. Le fait de prendre au hasard un seul
diamètre pour calculer la surface terrière ne permet pas de tenir
compte des réalités dans les autres directions de croissance sur
la même surface. Cela peut ainsi conduire à des
imprécisions dans les résultats (Rondeux, 1999). Par
conséquent, deux diamètres ont été optés
compte tenu des formes plus ou moins régulières. Mais quelle que
soit la forme de la rondelle, la surface terrière (g) se calcule suivant
la formule (Rondeux, 1999): g : surface terrière
irdm 2
g =
4
dm 2 : Diamètre moyen par arbre
Le diamètre moyen (dm) est obtenu de la manière
suivante :
dm = >ICm× 2
lcm : largeur de cernes moyen
dm : diamètre moyen
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Dans le cadre de ce mémoire, l'accroissement moyen
annuel est la moyenne arithmétique des accroissements courants annuels
cumulés en surfaces terrières par année. Il se calcul par
la formule suivante :
gmayt cu.
AMA = avec t : âge en année
L'accroissement courant annuel est la moyenne de surface
terrière de chaque cerne de croissance.
ACA = gmoy de chaque cerne
(g) Corrélation avec le climat
Nous avons utilisé des tests statistiques de la
corrélation simple et la régression linéaire. La
corrélation simple permet d'établir le signe de
l'évolution simultanée de la croissance par rapport aux facteurs
climatiques et la régression linéaire permet d'isoler le facteur
qui a le plus d'influence sur l'épaisseur des cernes de croissance
(activité cambiale).

Les composantes climatiques utilisées dans ce travail sont
présentées comme suit : La précipitation moyenne mensuelle
(1954 -- 2006) ;
La température moyenne mensuelle (1954 -- 2006)
3.3.2.3. Influence des essences accompagnatrice sur la
croissance de Pericopsis elata
Pour ce paramètre, nous n'avons tenu compte que des
espèces accompagnatrice de Pericopsis elata aux endroits
où ce dernier est fort concentré. Ce paramètre à
été étudié dans tout le bloc d'aménagement
c'est-à-dire 255ha. Les coordonnées géographiques ont
été prises pour chaque arbre inventorié. En vue de mettre
en évidence l'influence des essences accompagnatrices sur la croissance
de Pericopsis elata, un indice a été crée mais
qui doit être pris avec beaucoup de réserve, car il n'a pas fait
objet des plusieurs expérimentations et que le seuil d'influence n'est
pas encore déterminé. Les paramètres que nous avions pris
en compte pour évaluer l'influence sont les suivants :
|
La densité relative ;
La dominance relative ;
La surface de la couronne ;
La distance moyenne minimale.
|
Nous avons estimé que prendre les paramètres de
compétition pour corréler à la croissance de
Pericopsis elata pourrait apporter les éléments de
lumière pour évaluer cette influence.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
(a) Conversion des coordonnées
géographiques en coordonnées cartésiennes
Afin de convertir les coordonnées géographiques
(Longitude, latitude) en coordonnées cartésiennes (X, Y), nous
avons procédé de la manière suivante :
a. Appeler le shapefile sur l'interface Arc Map ;
b. Cliquer sur l'outil « Xtools Pro » du logiciel
Arc GIS 9.2 puis « Table operation » puis « X, Y, Z cordinates
» ensuite spécifier le système des coordonnées. Il
faudrait se rassurer que le système soit en système des
coordonnées projetées pour permettre les calculs de distance
métrique.
(b) Calcul des distances
Une fois convertie, les coordonnées cartésiennes de
chaque individu sont utilisées pour déterminer leur distance par
rapport à chaque semencier de Pericopsis elata
inventoriés grâce à la formule :
Distance AB = (XA -- XB)2 + (YA -- YB)2 +
(ZA -- ZB)2
A = Espèces présentes dans le site Y = latitude en
mètre
B = Espèce cible Z = Altitude en mètre
X= longitude en mètre
(c) Moyenne des distances minimales des
essences
Pour déterminer la distance minimale moyenne de chaque
espèce, nous avons procédé par le calcul de la distance
minimale de chaque individu par rapport à chaque semencier de
Pericopsis elata puis calculé la distance moyenne minimale de
chaque individu pour chaque espèce.
(d) Calcul de l'indice d'influence
Afin de mettre en évidence l'influence des essences
accompagnatrices, un indice d'influence a été créé.
Ce dernier tient compte de la densité relative, de la dominance
relative, de la surface de la couronne et de la distance moyenne minimale de
l'essence par rapport à l'espèce cible, en l'occurrence ici
Pericopsis elata.
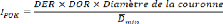
Où IPOK est l'indice d'influence,
DER : densité relative, DOR : dominance relative et
Dmin : Distance minimale moyenne
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
4. RESULTATS
4.1. Caractérisation du peuplement de Pericopsis
elata
4.1.1. Phytodiversité du Bloc
d'aménagement
4.1.1.1. Richesse spécifique aréale
Sur l'ensemble de 4 layons qui couvre 21 ha, 10.596 arbres
à dbh =10cm ont été inventoriés, ce qui correspond
à une densité de 504,6 pieds/ha. Ces arbres appartiennent
à 229 espèces reparties dans 35 familles (annexe 1).
4.1.1.2. Abondance relative
L'abondance relative sur base de données d'inventaire
réalisé dans le Bloc d'aménagement est consignée
dans la figure 13

Guarea thompsonii.
332
3%
Carapa procera 294
3%
Pancovia
harmsiana
350
3%
Cola griseiflora
366
3%
Petersianthus
macrocarpu374
3%
Tridesmostemon
claessensii 385
4%
Autres esp.
5997
57%
Pericopsis elata
429
4%
Staudtia stipitata
694
7%
Olax gambecola
542
5%
Scorodophloeus
zenkeri Harms
833
8%
Figure 13: Abondance relative des espèces du
Bloc d'Aménagement
Sur base de données d'inventaire, il sied de signaler
que par rapport aux familles, c'est la famille de Fabaceae qui est la
plus abondante avec 2053 individus soit 19%, suivi de Meliaceae 1376
soit 13%, de Myristicaceae avec 1050 soit 10% d'Olacaceae
avec 859 individus soit 8% (Annexe 1). Au sein de la famille Fabaceae
Pericopsis elata vient en deuxième position après
Scorodophleus zenkeri avec 429 tiges correspondant à 20% et en
quatrième position avec 4% après Scorodophleus zenkeri,
staudtia stipitata et Olax gambecola pour l'ensemble des
familles.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
4.1.1.3. Dominance relative
La proportion de la surface terrière qu'occupe
Pericopsis elata par rapport aux autres espèces sur l'ensemble
des layons inventoriés est représentée dans la figure 14
cidessous.
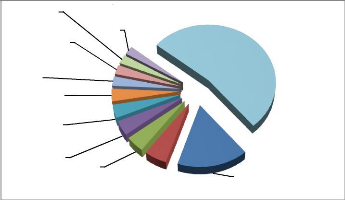
Drypetes
gossweileri
0,93
Pericopsis elata
3%
1,22
4%
Anonidium mannii
Staudtia stipitata.
1,28
4%
Olax gambecola
0,85
2%
Panda oleosa
0,91
3%
1,41
4%
Petersianthus
macrocarpus
1,52
4%
Guarea thompsonii
0,80
2%
Tridesmostemon
claessensii
1,81
5%
Autres esp.
18,62
53%
Scorodophloeus
zenkeri 5,68
16%
Figure 14: Dominance relative des espèces du Bloc
d'aménagement
Pericposis elata fait parti des espèces les
dominantes du bloc d'aménagement occupant la sixième position
avec une surface terrière de 1,22%.
4.1.1.4. Structure diamétrique spécifique
de Pericopsis elata
La répartition des tiges par classe de diamètre de
Pericopsis elata est consignée dans la figure 15.
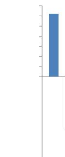
15 9 4 4 9 3 6 2 3 1
63
20 - 30
310
350
300
250
200
150
100
50
Nbre d'ind.
0
10 - 20
|
30 - 40
|
40 - 50
|
50 - 60
|
60 - 70
|
70 - 80
|
80 - 90
|
90 - 100
|
100 -110
|
110 -120
|
120 -130
|
|
|
|
|
|
|
|
|
|
|
Pericopsis elata (Harms) Van Meeuwen
Classe de dhp (cm)
Figure 15: Structure diamètrique de
Pericopsis elata
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
La figure 15 montre la structure diamètrique
spécifique de Pericopsis elata qui est reflétée
par l'abondance relative. Celle-ci est une exponentielle décroissante
typique des forêts denses humides (Rollet, 1974 ; Forni, 1997,
cités par Doucet, 2003) avec une abondance des individus dans les deux
premières classe de dhp.
4.1.2. Distribution spatiale de Pericopsis elata
Un total de 2327 pieds de Pericopsis elata (tableau
4) de chp = 10 cm a été inventorié sur une superficie de
255 ha. Tous les pieds sont présentés spatialement dans la figure
16. Les calculs de la distribution spatiale sont consignés dans le
tableau 5.
Tableau 4: Structure verticale du Pericopsis
elata dans le bloc d'aménagement
|
Structure verticale
|
Nombre de tiges
|
Circonférence à hauteur
de la poitrine
moyenne
(cm)
|
Moyenne de hauteur
fût (m)
|
|
Dominés
|
1379
|
29,6
|
|
|
Co-dominants
|
635
|
59,3
|
13,2
|
|
Emergents
|
188
|
275,8
|
19,2
|
|
Dominants
|
125
|
136,8
|
13,6
|
|
Total général
|
2327
|
|
|
Il ressort du tableau 4 que les dominés sont les plus
abondants avec 1379 tiges, suivi de co-dominants, des émergents et enfin
des dominants.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
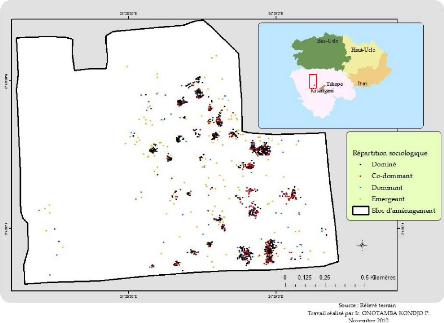
Figure 16: Représentation spatiale des pieds de
Pericopsis elata dans le Bloc
d'aménagement de
RBY
La figure 16, montre comment sont répartie
spatialemenent les individus de Pericopsis elata dans le bloc
d'aménagement. Nous constatons que les dominés en jaune sont plus
abondant et forment des agrégats isolés mais qui doivent
être testé statistiquement par la méthode de Clark et Evans
(Tableau 5).
Pour des calculs statistiques, les arbres dont le dhp est = 10 cm
ont été rajoutés dans la liste (de 2.327 à 2.331
pieds de Pericopsis elatata).
a. La densité de la population
(p)
Nombre d'individus dans la zone
d'étude 2331
|
P=
|
|
2550000 m2 = 0,0009141 ind./m2
|
|
surface de la zone
d'étude
|
|
r o=
|
14994,29
|
= 6,432557 m
|
|
2331
|
b. La distance moyenne observée jusqu'au voisin le
plus proche
--
ro = riavec ri Distance jusqu'au voisin le
plus proche pour l'individu i
n
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
c.
|
La distance attendue jusqu'au voisin le plus proche
(rE)
|
|
1
|
1
|
|
|
|
|
|
=
|
16,5516643 m
|
|
rE =
2A ??
|
--rE =
2A 0,0009141
|
|
|
|
d.
|
La mesure d'éloignement de la distance
observée de la distribution aléatoire
(R)
|
6,432557
16,5516643
-- R=
= 0,38863506
ro
rE

R=
Critères :
· R = 1 Distribution aléatoire
· R est proche de 0 la distribution est
agrégée
· R est #177; 2,15 la distribution est uniforme
Tableau 5: Paramètres des calculs de distance au
plus proche voisin
Paramètres calculés Tous les individus
n 2331
p (nbre Ind./m2) 0, OOO9141
r i min(m) 0,05
r i max(m) 164,24
r o(m) 6,43
r E(m) 16,54
R 0,39
Type de distribution Agrégée
L'indice d'agrégation R indique que la distribution
spatiale des individus inventoriés est agrégée car son R =
0,39, largement inférieur à 1 pour le type aléatoire et
à 2,15 pour le type uniforme.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
4.2. Etude de la croissance de Pericopsis elata
4.2.1. Activité cambiale
Tous les paramètres étudiés dans le domaine
de l'activité cambiale fait référence aux qutre arbres de
la plantation (Bloc Ole) de la RBY.
La figure 17 montre la disposition des cellules non
différenciées de la zone cambiales
par file.
Cellules du phloème
#177;4 Cellules non différenciées par file
dans
zone cambiale
Cellules en différentiation
Cellules du xylème secondaire
Figure 17: Vue microscopique de la zone cambiale de
Pericopsis elata
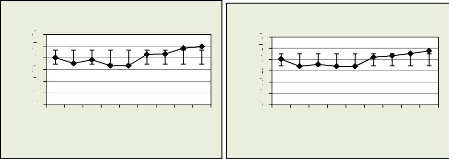
Les figures 18, 19, 20, 21 illustrent le nombre des cellules dans
la zone cambiale par mois
M ai1
Jui n1
Jui n_2 Jui I I et1 Jui I I et 2 Ao0t1 AoOt_2 SQpt_1 SQpt_2
Mai 1 Jui n1 Jui n_2 Jui I I et1 Jui I I et2 AoCit1 AoCit_2
SQpt_1 SQpt_2
A1
A2
6
5
4
3
2
1
0
nmc par file radiale
6
5
4
3
2
1
0
nmc par file radiale
Figure 18: nmc dans la zone cambiale par Figure 19: nmc
dans la zone cambiale
récolte pour le premier arbre par récolte
pour le deuxième arbre
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
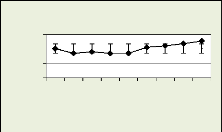
nmc par file radiale
4
2
6
0
Mai _l Juinl Juin_2 Juilletl Juillet2 Aofit_l Aofit2 Sept_1
Sept_2
A3
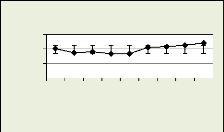
nmc par file radiale
4
2
6
0
Mai _l Juinl Juin_2 Juilletl Juillet2 Aofit_l Aofit2 Sept_1
Sept_2
A4
Figure 21: nmc dans la zone cambiale par récolte
pour le troisième arbre figurs
Figure 20: nmc dans la zone cambiale
par
récolte pour le quatrième arbre
Il se dégage pour les figures 18, 19, 20 et 21 que le
nombre de cellules non différenciées produitent par file radiale
atteint son maximum au mois de septembre avec un ralentissement au mois de
juillet et une petite reprise au mois de mai.
La moyenne des cellules non différenciées dans la
zone cambiale de ces quatre arbres par récolte est consignée dans
la figure 22.
nmc de 4 arbres
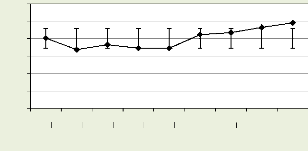
Juin 1
Mai 1
Juin2
Juillet2
Sept_1
Sept_2
Aofit_l
Juilletl
Aofit2
6
5
4
3
2
1
nmc de tous les arbres
0
Figure 22: nmc dans la zone cambiale de quatre
arbres
La tendance de la courbe pour cette figure est identique
à celle des figures 18, 19, 20
et 21.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Pour mettre en évidence la corrélation entre les
données climatiques et le nombre de cellules prélevé
bimensuellement, une moyenne mensuelle de nombre de cellules a
été calculée et consignée dans la figure 23.
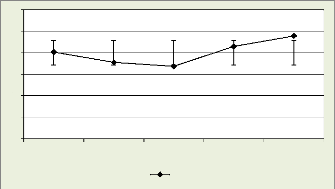
Mai Juin Juillet Août Sept
nmc
6
5
4
3
2
1
0
Figure 23: Moyenne mensuelle des cellules produites dans
la zone cambiale
Les figures 24 et 25 montrent la corrélation entre les
paramètres climatiques et le nombre des cellules produites dans la zone
cambiale pour chaque mois.
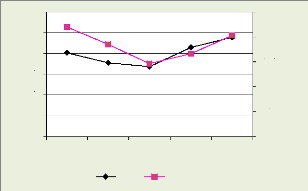
nmc
4
6
3
2
0
5
1
Mai Juin Juillet Août Sept
nmc Précip (mm)
250
200
50
0
150
100
Precip (mm)
Figure 24: Corrélation entre le nombre de
cellules dans la zone cambiale et la
précipitation
Il apparait dans cette figure que la tendance de cellules
produites dans la zone cambiale suit la même tendance que la courbe de
précipitation. Ces deux paramètres sont moyennement
corrélés avec R2= 0,515.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
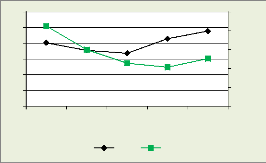
4
0
6
5
3
2
1
Mai Juin Juillet Août Sept
nmc T°C
26,5
26,0
25,5
25,0
24,5
24,0
Figure 25: Corrélation entre le nombre de
cellules dans la zone cambiale et la
température
La figure 25 montre que ces deux paramètres ne sont pas
corrélés car R2= -0,029.
Tableau 6: Corrélation entre les
paramètres climatiques et le nombre moyen de cellules dans la
zone
cambiale
|
nmc
|
Précip
|
T°C
|
|
nmc Corrélation de Pearson
|
1
|
0,513
|
-0,029
|
|
Sig. (bilatérale)
|
|
0,377
|
0,963
|
|
N
|
5
|
5
|
5
|
|
Précip Corrélation de Pearson
|
0,513
|
1
|
0,824
|
|
Sig. (bilatérale)
|
0,377
|
|
0,086
|
|
N
|
5
|
5
|
5
|
|
T°C Corrélation de Pearson
|
-0,029
|
0,824
|
1
|
|
Sig. (bilatérale)
|
0,963
|
0,086
|
|
|
N
|
5
|
5
|
5
|
Il ressort dans ce tableau que la précipitation a plus
de l'influence sur la production des cellules que la température et le
seuil de probabilité se rapproche plus de 0,05 pour la
précipitation (0,377) que celui de la température (0,963).
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
4.2.2. Dendrochronologie
Tous les paramètres étudiés dans le domaine
de la dendrochronologie sont prélevés dans quatre arbres de la
plantation de la RFY.
4.2.2.1. Le cross-dating
Le cross-dating permet d'établir la concordance entre
les largeurs de cernes de plusieurs échantillons et permet aussi
d'attribuer à chaque cerne l'année exacte de sa formation. Le
tableau 7 montre la synchronisation des moyennes des rayons à
l'intérieur de chaque arbre et le tableau 8 illustre la synchronisation
entre les différents arbres.
|
Tableau 7: Cross-dating intra-arbres
|
Tableau 8: Cross-dating inter-arbres
|
|
Arbre-moyenne
|
GIK
|
Arbre
|
TV-BP
|
|
Yoko1-m
|
66
|
Yoko1-m
|
2
|
|
Yoko2-m
|
60
|
Yoko2-m
|
2
|
|
Yoko3-m
|
71
|
Yoko3-m
|
2
|
|
Yoko4-m
|
65
|
Yoko4-m
|
3
|
|
Yoko-m
|
66
|
Yoko-m
|
2
|
Au regard du tableau 7 le test GIK montre que la moyenne
trouvée de 66 est supérieure au seuil critique,
c'est-à-dire que la synchronisation entre les séries est
très bonne d'oü il y a concordance entre les cernes des rayons du
même arbre. Le tableau 8 ci-dessus montre que le TV-BP est égal au
seuil critique accepté pour les espèces tropicales. Cela signifie
que la synchronisation entre les arbres est bonne et la concordance entre les
cernes de croissance est correcte. Cela permet de dater les arbres avec plus de
certitude possible à 60 ans.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
4.2.2.2. Mesure de l'épaisseur des cernes de
croissance
Les largeurs moyennes de cernes de croissance (annexe 3)
après synchronisation sont consignées dans la figure 26.
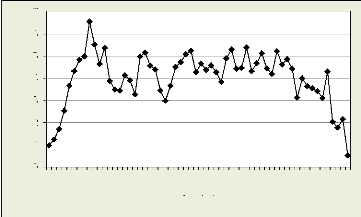
I argeur cerne (mm)
4
2
7
6
5
3
0
1
1
4
7 10 13 16 19 22 25 28 31 34 37 40 43 46 49 52 55 58
Age (an)
Figure 26: Evolution des largeurs de cernes de croissance
au cours de temps
La figure 26 ci-dessus, montre qu'il y a une croissance
exponentielle dans les l0 premières années avec un pic à
la 9ème année avec 6,56 mm. La courbe se stabilise
à partir de la 28ème année puis décroit
brutalement à la 55ème année.
4.2.2.3. Estimation de la productivité
ligneuse
La figure 27 ci-dessous montre l'allure de l'ACA et de l'AMA
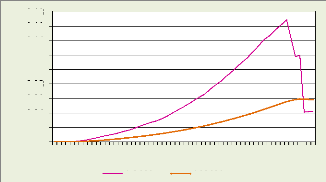
Accroissement (m2/an)
0,18
0,16
0,14
0,12
0,08
0,06
0,04
0,02
0,1
0
1 4 7 10 13 16 19 22 25 28 31 34 37 40 43 46 49 52 55 58
ACAYm AMAYm
Age
Figure 27: Accroissement courant et moyen annuel
(cm)
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Il ressort de la figure 27 que l'accroissement courant annuel
en surface terrière est presque nul dans les 5 premières
années et s'en suit d'une croissance exponentielle qui atteint son pic
à la 54ème année avec 0,16 m2 par an
et décroit pour s'intercepter à la courbe de l'accroissement
moyen à la 58ème année. Cette interception
traduit l'âge théorique pour une optimalisation d'une exploitation
de bois. L'accroissement moyen annuel en surface terrière est de 0,06
m2/an soit un accroissement moyen annuel en diamètre de 0,27
cm/an pour une période de 60 ans.
4.2.2.4. Corrélation avec le climat
Le tableau 8 ci-dessous montre le paramètre climatique qui
influent le plus sur la largeur de cernes.
Tableau 9: Paramètres climatiques contribuant le
plus à largeur de cernes
|
|
Largeur de
cernes
|
Précip.
Totale
|
T°C
moyen
|
|
Largeur de
cernes
|
1,000
|
-0,003
|
0,320
|
|
Corrélation de
Pearson
|
Précip. Totale
|
-0,003
|
1,000
|
0,102
|
|
T°C moyen
|
0,320
|
0,102
|
1,000
|
A la lumière de ce tableau, il se dégage que la
contribution de la précipitation et de la température sur la
largeur des cernes est faible, mais néanmoins la température
contribue en grande partie par rapport à la précipitation car son
R2 = 0,320 ce qui est compris entre 0,2 et 0,5.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Les figures 23 et 24 illustrent l'allure des courbes de
précipitation à celle de largeur des cernes de croissance et de
celle de la température.
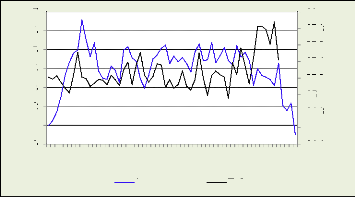
Largeur carne
4
2
7
6
5
3
0
1
1
4
7 10 13 16 19 22 25 28 31 34 37 40 43 46 49 52 55 58
Largeur cernes T°C
25,8
25,6
25,4
25,2
24,8
24,6
24,4
26
25
T°C moy.
Figure 28: Corrélation entre la largeur des
cernes et la température
La figure 28 ci-dessus montre que les largeurs de cernes de
croissances sont moyennement corrélées à celle de
données de la température R2=0,320.
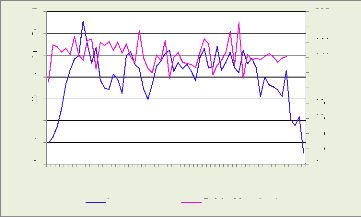
Largeur came
7
4
2
0
6
5
3
1
1
4
7 10 13 16 19 22 25 28 31 34 37 40 43 46 49 52 55 58
Largeur cernes Précip. Moy. (mm)
40
200
20
0
80
60
180
160
140
120
100
Precip. Moy. (mm)
Figure 29: Corrélation entre la largeur de cernes
de crissance et la précipitation
La figure 29 ci-haut montre la tendance des courbes de la largeur
des cernes de croissance qui est faiblement corrélée à
celle de la précipitation R2= 0,003.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
4.2.3. Influence des grandes essences sur la croissance de
Pericopsis elata
Le degré d'influence dépend de la dominance
relative, de la densité relative et du diamètre de la couronne
d'une essence sur la distance minimale qui la sépare avec
l'espèce cible. Le tableau 9 montre le degré d'influence des
grandes essences sur Pericopsis elata.
Tableau 10: Indice d'influence des grandes essences
sur la croissance du Pericopsis elata (Distance Moy.min (m)=
distance moyenne minimale en mètre, Dhp moy. (cm)=Dhp moyen en cm, Nbre
d'ind.=Nombre d'individu, DER=densité relative, DOR=Dominance relative,
IPOK=Indice d'Influence).
|
N° Nom scientifique
|
Distance
Moy.min.
(m)
|
Dhp moy.
(cm)
|
Nbre
d'ind.
|
Diamètre
moy.
couronne (m)
|
DER
|
DOR
|
IPOK
|
|
1.
|
Entandrophragma spp
|
96,22
|
88,5
|
44
|
13,920
|
30,8
|
0,294
|
1,307
|
|
2.
|
Petersianthus macrocarpus
|
90,23
|
87,7
|
18
|
13,616
|
12,6
|
0,616
|
1,170
|
|
3.
|
Erythrophleum suaveolens
|
131,19
|
90,0
|
21
|
14,087
|
14,7
|
0,345
|
1,135
|
|
4.
|
Trilepisium
|
|
|
|
|
|
|
|
|
madagascariensis
|
29,19
|
72,9
|
6
|
12,000
|
4,2
|
0,246
|
0,424
|
|
5.
|
Prioria balsamifera
|
47,10
|
90,4
|
7
|
11,500
|
4,9
|
0,322
|
0,385
|
|
6.
|
Prioria oxyphylla
|
118,90
|
69,5
|
12
|
11,790
|
8,4
|
0,299
|
0,249
|
|
7.
|
Piptadeniastrum africanum
|
112,79
|
88,7
|
7
|
14,625
|
4,9
|
0,275
|
0,174
|
|
8.
|
Pterocarpus soyauxii
|
54,70
|
102,2
|
5
|
12,881
|
3,5
|
0,203
|
0,167
|
|
9.
|
Canarium schweifurthii
|
239,40
|
76,7
|
4
|
22,432
|
2,8
|
0,068
|
0,018
|
|
10.
|
Pericopsis elata
|
|
263,30
|
138
|
13,870
|
|
0,285
|
|
Il ressort de ce tableau que les espèces du genre
Entandrophrama ont plus d'influence positive que négative sur
la croissance du Pericopsis elata avec un indice d'influence de 1,307,
suivi de Petersianthus macrocarpus avec 1,170 puis Erythrophleum
suaveolens 1,135 il est suivi de Trilepisium madagascariensis
0,424 etc.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
5. DISCUSSIONS
5.1. Caractérisation du peuplement de Pericopsis
elata
5.1.1. Phytodiversité du bloc
d'aménagement
5.1.1.1. Richesse spécifique aréale
La richesse spécifique aréale trouvée
dans ce bloc de 11 espèces par hectare soit 229 espèces pour 21
hectares peut paraître faible dans la perspective d'une comparaison
intercontinentale et continentale. A titre d'exemple, Prance et al.
(1976) et Wong & Whitmore (1970), cités par Puig (2001), estiment
des richesses spécifiques pouvant atteindre 179 espèces par
hectare en Manaus au Brésil (Amérique du Sud) et 328
espèces sur 4ha en Pasoh/Malaisie (Asie tropicale), Gartlan et
al., 1986 et Newberry et al., 1986, cités par Puig, (2001)
estiment que des richesses spécifiques pouvant atteindre 39
espèces sur 0,64ha Douala-Edea et de 75 espèces sur 0,64ha au
Parc de Korup au Cameroun. Toutefois, elle demeure remarquable dans certaines
zones forestières de la RDC Comparativement à la Réserve
de biosphère de Luki où 118 espèces ligneuses (à
dbh =10cm) inventoriées sur une superficie estimée à
7550ha (Muembo & Nsenga, 2006), cette richesse spécifique de la RBY
paraît très élevée et très
diversifiée. Cet écart considérable se justifierait
surtout par le régime pluviométrique qui arrose abondamment les
forêts de la région de Yangambi et ses environs (plus 1800mm/an).
Cette richesse est aussi très élevée en considérant
des valeurs obtenues des autres sites du pays (exemple des résultats des
inventaires d'aménagement de concessions 27/03 Bikoro dans la Province
de l'Equateur et 36/04 Ubundu dans la Province Orientale donnant respectivement
195 espèces sur 768,5ha et 103 espèces sur 67,467ha (FRM, 2007 et
2008), indiquant que les forêts de la RBY sont dans l'ensemble
relativement denses.
5.1.1.2. Abondance et dominance relative
L'inventaire floristique a permis de recenser au total 10.956
individus repartis dans 229 espèces ligneuses regroupées en 35
familles. La famille de Fabaceae est la plus abondante et
l'espèce Scorodophleus zenkeri Harms est la plus abondante car
il compte à lui seul 833 individus soit 8% et présente un indice
d'importance, une densité et une surface terrière très
élevé respectivement de IVI = 24,525, DER = 7,861 % et 5,686.
Pericopsis elata est la deuxième espèce au sein de la
famille de Fabaceae et la quatrième sur l'ensemble des
espèces. La dominance de Scorodophloeus zenkeri, semble
être dictée par les facteurs sol et topographie. En effet selon
Vande weghe (2004) et Lebrun et Gilbert (1954) Scorodophloeus zenkeri
domine généralement les plateaux sabloneux et forme avec
Julbernadia seretii un
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
noyau d'espèces dominantes dans l'alliance
Oxystigmo-Scorodophloeion (Lebrun et Gilbert 1954) appelée
aussi « forêts semi-caducifoliés à Scorodophloeus
zenkeri ».
5.1.2. Distribution spatiale de Pericopsis elata
La distribution agrégée de Pericopsis elata
trouvée dans ce travail, est confirmée par Lebrun et
Gilbert, 1954, Boyemba 2006 et Begaa, 2009.
5.2. Etude de la croissance de Pericopsis elata
5.2.1. Activité cambiale
Les résultats trouvés dans ce mémoire ont
montré que le nombre le plus élevé des cellules par file
radiale dans la zone cambiale s'est observé au mois de d'Août et
au mois de septembre avec une moyenne de cinq cellules, un ralentissement au
mois de juillet, mois pendant lequel il a été observé le
plus de cellules en différentiation. Ces nombres de cellules non
différenciées dans la zone cambiale sont faiblement
corrélés avec les données climatiques mises en ensemble
c'est-à-dire la température et la précipitation
R2= 0,222. Contrairement à l'étude menée par
Karumanchi S. et Kishore S., 2001 pour l'espèce Azadirachta indica
oü les cycles saisonniers dans différents types
d'écosystèmes forestiers de l'état de Gujarat ont
été comparés entre des arbres provenant de forêts
humide décidue (MDF), sèche décidue (DDF) et de savane
arborée (SF), ils ont donné des résultats suivants : La
division des cellules cambiales commence en février et juin pour les
MDF, la croissance radiale maximale s'est observée en
août-septembre, au moment des fortes pluies, et un arrêt en janvier
et mai pendant les périodes les plus sèches de l'année.
Dans les DDF, le premier démarrage de croissance radiale commence en
janvier, avec un maximum en avril et un arrêt en mai. Une seconde
période d'activité commence en juin avec l'arrivée des
pluies, atteint son maximum en octobre et s'arrête en décembre.
Dans les SF le cambium reste actif toute l'année avec trois pics
d'activité en février, juillet et octobre. Lorsque les feuilles
arrivent à maturité complète en novembre, la vitesse de
division cellulaire du cambium est relativement faible dans les MDF et le SF et
nulle dans les DDF. Dans le cambium, les rayons présentent des espaces
intercellulaires importants pendant les mois secs pour les trois types de
forêts.
5.2.2. Dendrochronologie
5.2.2.1. Largeur des cernes de croissance
Les résultats sur la largeur de cernes de croissance
ont montré qu'il y a un pic au cours de la vie de l'arbre, à la
neuvième année avec une largeur de 6,56 mm. Ce pic pourrait
s'expliquer par la croissance rapide de l'arbre à son jeune age et la
décroissance brusque
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
pourrait être expliqué par une forte
compétition des essences accompagnatrice de Pericopsis
elata.
5.2.2.2. Estimation de la productivité
ligneuse
L'AMA en surface terrière dans la plantation de Yoko
pour une période de 60 ans de 0,06 m2/an soit un AMA en
diamètre de 0,27 cm/an pour Pericopsis elata paraît
très faible dans la perspective d'une comparaison continentale et
locale. Au Ghana, Adler (1989) a trouvé pour la même espèce
un accroissement diamétrique de 1,3 cm/an pour une période de 55
ans. En Côte d'ivoire, CITES, 2003 donne une valeur atteignant de 0, 85
à 1cm/an dans une plantation de 65 ans, Doucet, 2007 a trouvé
0,35 cm/an à 60 ans pour la même espèce au cameroun.
D'après Assumani, 2009 l'accroissement annuel moyen actuel de
Pericopsis elata dans les plantations forestières de
l'I.N.E.RA. -Yangambi varie entre 0,64 et 0,69 cm/an ce qui est
supérieur à celui trouvé dans ce mémoire. Ce faible
accroissement s'explique par le non suivi de traitements sylvicoles, car cette
plantation date de 1954 ; il s'en suit d'une forte compétition inter
espèce et la méthodologie utilisée est différente
de celle utilisée par les autres auteurs.
5.2.2.3. Corrélation avec le climat
Les résultats trouvés, montrent que les
paramètres climatiques, on une faible influence sur l'épaisseur
de cernes de croissance, bien que la température influe plus que la
précipitation sur la croissance de cette espèce, mais ensemble le
pourcentage est faible avec R2= 0,158. Ceci confirme que la
croissance du Pericopsis elata dépend des autres facteurs et en
l'occurrence les essences accompagnatrices par la recherche de la
lumière.
5.2.3. Influence des grandes essences sur la croissance
Pericopsis elata
Les résultats trouvés dans le cadre de ce
mémoire montrent que Entadrophragma spp, Erythrophleum
suaveolens et Petersianthus macrocarpus (espèces qui
présentent une densité et une dominance relative
élevées) contribuent positivement que négativement sur la
croissance de Pericopsis elata. La raison pour laquelle, nous
attribuons la croissance du Pericopsis elata à ce facteur
d'influence est la suivante :
|
Positivement Pericopsis elata est une essence de
lumière. En effet les grandes essences citées ci-haut permettent
le développement des chenilles qui font objet d'une récolte
intensive par la population riveraine ; leur pénétration en
forét perturbe l'écosystème et crée des bonnes
conditions pour son développement en laissant pénétrer la
lumière. Cette dernière est un facteur prépondérant
pour la croissance de cette espèce.
|
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)


La défoliation des essences accompagnatrice tels que
les espèces du genre Entandrophragma qui perdent le deux tiers
de leurs feuilles favorise la disponibilité des éléments
minéraux dans le sol. De ce fait, Pericopsis elata
bénéficie de ces éléments mineraux pour sa
croissance.
Négativement, il y a une forte compétition
verticale et horizontale entre les essences accompagnatrices et Pericopsis
elata.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
CONCLUSIONS ET SUGGESSIONS
Au terme de cette étude porté sur la dynamique
de Pericopsis elata (Harms) Van Meeuwen qui avait pour objectif de
connaître la phytodiversité de la zone d'étude,
d'étudier la distribution spatiale, la périodicité de
croissance, la dendrochronologie de Pericopsis elata et
d'étudier l'influence des grandes essences sur la croissance de cette
espèce.
10.596 tiges de dhp = 10cm ont été
dénombrées sur 21 ha reparties dans 229 espèces et 35
familles. La famille de Fabaceae présente une importance
relative élevée avec 66,223 au sein de la quelle Pericopsis
elata vient en deuxième position après Scorodophleus
zenkeri avec 429 tiges correspondant à 20%. La structure
diamétrique est en forme exponentielle décroissante typique des
forêts denses humides avec une abondance des individus dans les deux
premières classes de dhp.
Cette espèce présente une distribution
agrégée. La croissance en termes de division des cellules dans la
zone cambiale pour la période de récolte ont montré
Pericopsis elata produit plus de cellules au mois d'août et
septembre, avec une réduction de cellules au mois de juillet. Les
données climatiques ont une faible influence sur la division des
cellules de la zone cambiale.
Les profiles de cernes de croissance sont synchrone que
ça soit entre les rayons d'un même arbre et entre les cernes de
différents arbres. L'accroissement moyen annuel en surface
terrière est de 0,06 m2/an soit un accroissement moyen annuel
en diamètre de 0,27 cm/an pour une période de 60 ans. La
croissance de cette espèce est plus liée à la
présence des grandes essences qu'aux paramètres climatiques
(température et précipitation) car la corrélation a
montré que ces paramètres influents à un faible
pourcentage.
Eu égards aux résultats trouvés dans ce
mémoire nous suggérons ce qui suit :



étendre l'étude aux autres plantations et aux
foréts naturelles sur le territoire national, en vue de disposer de
données plus fiables sur la croissance et les variations climatiques
intervenues ;
parcourir tous les cycles saisonniers pour déterminer
avec exactitude la période pendant laquelle il y a vraiment arrêt
de production de cellules cambiales.
Dans le cadre de la contribution pour les études de la
dynamique forestière
en zone tropicale, cet indice qui a
été crée en vue d'évaluer le
degréd'influence des autres espèces sur la croissance
de l'espèce du Pericopsis
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
elata doit faire objet de beaucoup
d'expérimentation afin de dégager les arguments
écologiques solides permettant d'expliquer la faible croissance de
Pericospsis elata. Dans la méme file d'idée faire des
études plus approfondie sur les essences accompagnatrice en
corrélation avec les facteurs environnementaux (lumière,
matière organique, etc.).
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
REFERENCES BIBLIOGRAPHIQUES
AETFAT, Missouri Botanic Garden, June 1985: Missouri Botanic
Garden, Missouri.
AKAGOU ZEDONG H.C., 2008. Gestion durable de Pericopsis elata
au Cameroun. Rapport de l'Organe de Gestion CITES présenté
à l'atelier OIBT/CITES dans le cadre du projet : s'assurer que le
commerce international du bois de Pericopsis elata (Afrormosia ou Assamela)
n'est pas préjudiciable à sa conservation, 2-4 avril 2008, Kribi,
Cameroun, 6 p
AKE ASSI, L., 1988 : Espèces rares et en voie
d'extinction de la flore de la Côte d'Ivoire. In: Goldblatt, P. and
Lowry, P.P. (eds.) Modern systematic studies in African Botany. Proc Eleventh
Plenary Meeting, AETFAT, Missouri Botanic Garden, June 1985. Missouri Botanic
Garden, Missouri.
ALDER D., 1989. Natural forest increment, growth and yield.
In: Wong, J.L.G. (ed.) Forest inventory project, seminar proceedings, Accra.
Overseas Developpement Admistration, UK and Ghana Forestry Department. and
Conservation, 5 : 447- 459.
ANON, 1979: Tropical legumes: Resources for the future.
Washington, DC: National Academy of Sciences.
ASSUMANI, Dieu Merci, 2009 : Bilan dendrométrique de
plantations expérimentales de Pericopsis elata (Harms) Van Meeuwen et
Millettia laurentii De Wild. installées à Yangambi (R.D.Congo)
entre 1938 et 1942, 108 p.
ATIBT, 2002: Technical reports on Pericopsis elata. Unpublished
document, 72 p.
AUZEL, P., FETEKE, F., FOMETE, T. NGUIFFO, A. and DJEUKAM, R.,
2001 : Impact de l'exploitation forestière sur las fiscalité, sur
l'aménagement et sur le développement local. Forests Monitor,
IUCN, DFID.
BAILLIE MGL & Pilcher JR, 1973: A simple crossdating
program for tree-ring research. Tree-Ring Bulletin, 33: 7-14.
BEDEL F., DURRIEU de MADRON L., DUPUY B. ; 1997 : Dynamique de
croissance dans des peuplements exploités et éclaircis de
forêt dense africaine : le dispositif de
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
M'Baïki en République Centrafricaine (1982 - 1995).
Projet FORAFRI 60 p. + annexes.
BEECKMAN, H., 2007: Collection MRAC. BOIS.
BEGAA YENDJOGI S., 2009 : Analyse des conditions de
régénération de Pericopsis elata (Harms) Van Meeuwen dans
la forêt naturelle aménagée de Yangambi (RDC)
BOURLAND N., 2008. Quelques éléments de
synthèse sur l'écologie de Pericopsis elata au sud-Est
du Cameroun. Travail présenté à l'atelier OIBT/CITES dans
le cadre du projet : s'assurer que le commerce international du bois de
Pericopsis elata (Afrormosia ou Assamela) n'est pas
préjudiciable à sa conservation. 6 p
BOYEMBA F., 2006. Diversité et
régénération des essences forestières
exploitées dans les forêts des environs de Kisangani.
Mémoire de DEA. ULB. Fac des Sciences, Laboratoire de Botanique et
Phytotaxonomie, 101 p.
BROWN, S., GILLEPSIE A. & A. LUGO. 1989. Biomass
estimation methods for tropical forests with applications to forest inventory
data. Forest Science, 35: 881-902.
BULTOT F., 1977. Atlas climatique du bassin zaïrois. IVe:
pression atmosphérique, vent en surface et en altitude,
température et humidité de l'air en altitude,
nébulosité et visibilité, classification climatique,
propriétés chimiques de l'air et des précipitations.
Bruxelles: Publ. INEAC., hors série. 344 cartes, 11 figures et 35
tableaux.
CAP SCIENCE, 2005. : Dossier enseignant. « Du coeur à
l'ouvrage ».
CARMEN Regina Marcati, VERONICA Angyalossy and RAY Franklin
Evert, IAWA Journal, Vol. 27 (2), 2006: 199-211: Seasonal variation in wood
formation of cedrela fissilis (meliaceae)
CEFDHAC, 1999 Bases pour la mise en cohérence des
politiques et lois forestières des pays d'Afrique Centrale. IUCN.
CHAMBERS, J.Q., J. DOS SANTOS, R.J. RIBEIRO & N. HIGUCHI.
2001. Tree damage, allometric relationships, and aboveground net primary
production in central Amazon forest. Forest Ecology and Management, 152:
73-84.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
CHAVE, J. R. CONDIT, S. LAO, J. P. CASPERSEN, R. B. FOSTER
& S. P. HUBBELL. 2003. Spatial and temporal variation of biomass in a
tropical forest: results from a large census plot in Panama. Journal of
Ecology, 91: 240-252.
CHAVE, J., ANDALO, C., BROWN, S., CAIRNS, M.A., CHAMBERS,
J.Q., EAMUS, D., FÖLSTER, H., FROMARD, F., HIGUCHI, N., KIRA, T., LESCURE,
J.P., NELSON, B.W., OGAWA, H., PUIG, H., RIÉRA, B., YAMAKURA, T., 2005:
Tree allometry and improved estimation of carbon stocks and balance in tropical
forests. Oecologia 145: 87-99. "
CITES, 2003: Review of Significant Trade: Pericopsis
elata, PC 14 Doc. 9.2.2. DEMARQUEZ B., 1997 : Etude de l'accroissement en
diamètre de plusieurs essences.
DETIENNE P. et MARIAUX A. 1977 : Nature et
périodicité des cernes dans les bois rouges des
Méliacées africaines. Bois et Forêts des tropiques
175:52-61.
DETIENNE P., 1989: Appearance and periodicity of growth rings in
some tropical woods.
DETIENNE P., OYONO F., DURRIEU de MADRON L., DEMARQUEZ B.,
Robert NASI, 1998 : L'analyse de cernes : applications aux études de
croissance de quelques essences en peuplements naturels de forêt dense
africaine. Projet Forafri, document n°15, 40p
DIE, Agathe, 2004 : contribution à l'étude de la
croissance en épaisseur du Teck de plantation : Tectona grandis L.F.
(Verbenaneae), à Gognoa (centre - Ouest de la Côte d'Ivoire), par
l'analyse des cernes de croissance et par observations anatomiques.
DUNN, R.M and OTU, D.O., 1994: Reconnaissance inventory of
high forest and swamp forest areas in Cross River State, Nigeria.
FAO Forestry Department, 1986: Databook on endangered tree and
shrub species and their provenances.
FONGNZOSSIE Fedoung Evariste, TSABANG Nolé, NKONGMENECK
Bernard-Aloys, NGUENANG Guy Merlin, AUZEL Philippe, CHRISTINA Ellis, KAMOU
Edouard, BALOUMA Jean Marie, APALO Paul, MATHIEU Halford,
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
VALBUENA Martha et VALÈRE M, Vol. 1 (3) : 204-221, 2008 :
Les peuplements d'arbres du Sanctuaire à gorilles de Mengamé au
sud Cameroun.
FORESTS MONITOR, 2001: Sold down the river. The need to control
transnational forestry corporations: a European case study. Forests Monitor
Ltd, Cambridge, UK.
FORNI, E., 1997 : Types de foréts dans l'est du
Cameroun et étude de la structure diamétrique de quelques
essences. Memoir for the Diploma in Agronomic Science and Biology.
Faculté Universitaire des Sciences Agronomiques de Gembloux.
FOUARGE et LOUIS 1943. Essence forestière et bois du Congo
.Fasc . Afrorrmosia elata. Publ. INEAC, Bruxelles, 22p.
FREDERIC BEDEL, Luc DURRIEU de MADRON, Bernard DUPUY, Vincent
FAVRICHON, Henri Felix MAITRE, Avner BAR HEN, Philippe NARBONI, 1998: Dynamique
de croissance dans des peuplements exploités et éclaircis de
forêt dense africaine. Le dispositif de M'Baïki en République
Centrafricaine (1982 - 1995).
FRM., 2007. Projet d'aménagement des concessions des
sociétés SODEFOR, SOFORMA
HAWTHORNE, W.D., 1995: Ecological profiles of Ghanaian forest
trees. Oxford Forestry Institute. 345 p.
HOWLAND P., 1979: Pericopsis elata (Afrormosia). Commonwealth
Forestry Institute Occasional Papers 9,
IAWA Bulletin n.s., Vol 10 (2):123-132
IFUTA, N.B.1993. Paramètres écologiques et
hormonaux durant la croissance et La
reproduction d'Epomops franqueti (Mammalia : Chiroptera) de la
forêt Ombrophile équatoriale de Masako (Kisangani-Zaïre).
Thèse inédite, KUL, 142 p.
KABALA F., Kutelama-a-Seleko D., Toirambe B., Massudi
Mayan'Kenda C., 2008. Utilisation durable de l'Afrormosia en Afrique - cas de
la République Démocratique du Congo : situation au cours des
trois dernières années (2005, 2006, 2007). Rapport national
soumis par le Ministère de L'Environnement, Conservation de la Nature et
Tourisme (MINECNT), PROJET OIBT/CITES :
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
s'assurer que le commerce international du bois de Pericopsis
elata (Afrormosia ou Assamela) n'est pas préjudiciable à sa
conservation. 19 p.
KARUMANCHI S. Raoa,* and Kishore S. Rajputb 20 April 2001:
India Relationship between seasonal cambial activity, development of xylem and
phenology in Azadirachta indica growing in different forests of Gujarat
State
KOMBOZI Lono P., 2009 : Etude de la structure des
émergents et des dominants dans la forêt de Yangambi (Cas de la
forét d'aménagement de l'INERA-Yangambi) 70 p.
LEBRUN J. et GILBERT G., 1954. Une classification
écologique des forêts du Congo. Bruxelles : Publ. INEAC.,
série scient. n° 63, 89p.
LOMBA, B.L. et NDJELE, M-B. 1998 Utilisation de la
méthode de transect en vue de l' étude de la
phytodiversité dans la Réserve de Yoko (Ubundu, R.D. Congo).
Annales(11), Fac. Sci. UNIKIS, 35-46 p.
MAGNIN, C., 2001. : La Dendrochronologie. Laboratoire d'Analyses
et d'Expertise (L.A.E.), Bordeaux.
MARIAUX A., 1967 : Les cernes dans les bois tropicaux africains.
Nature et périodicité. Bois et Forêts des Tropiques 113:
3-14 & 114: 23-37.
MARIAUX A., 1969 : La périodicité des cernes dans
le bois de Limba. Bois et Forêts de
MARIAUX A., 1970 : La périodicité de formation des
cernes dans le bois de l'Okoumé. Bois et Forêt des Tropiques
131:37-50.
MEEFCP/ Projet d'aménagement forestier pilote de la Sangha
M'baéré.14p.
MINEF, 2002. Ministry of Environnement and Forestry. Note
technique sur le Pericopsis elata (Assamela/ Afromosia).Cameroun
86p.
MINEF, 2004. Rapport Technique sur Pericopsis elata (Assamela),
Ministère de l'Environnement et des Foréts, 8 p. + annexes
MONOT, T. 1955. Sur un cas excéptionnel de richesse
aréale : les sables de Mauritanie orientale. Recueil trav. Labo. Bot.
Géol. Et Zool. Fac. Sc. Montpellier, t7 : 63-67.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
NSHIMBA, S.M., 2008. Etude floristique, écologique et
phytosociologique des foréts de l'île Mbiye à Kisangani, RD
Congo. Thèse de Docteur en Sciences. ULB, 389 p.
OLA-ADAMS, B. (1977) Conservation of the genetic resources of
indigenous forest trees in Nigeria: Possibilities and limitations. Forest
Genetic Resources Information 7:1-9.
p.
PUIG H., 2001: la forêt tropicale humide. Ed. Berlin.
Paris, France. 448p.
QUENTIN Reusens, 2010 : Apport de la
télédétection à très haute résolution
pour la caractérisation d'un peuplement de forét tropicale
humide, etude de cas. 68p.
RONDEUX J., 1999.- La mesure des arbres et des peuplements
forestiers. Les presses Agronomiques de Gembloux (Belgique). 17pp
SAYER, J.A., Harcourt, C.S. and Collins, N.M., 1992: The
Conservation Atlas of Tropical Forests. Africa.
SCHWEINGRUBER F. H., 1987. : Tree Rings: Basics and Applications
of Dendrochronology. D. Reidel Publishing Company, Netherlands, 255-261 p
SCHWEINGRUBER F. H, 1988: Tree Rings. Reidel, Dordrecht,
Netherlands.
SGS Trade Assurance Services, 2002: Forest law assessment in
selected African countries. Final Draft. World Bank/WWF Alliance.
SOKI, K. 1994. Biologie et écologie des termites
(Isoptère) des forêts ombrophiles du Nord
TROUET, V., J. ESPER, N. E. GRAHAM, A. BAKER, J. D. SCOURSE,
and D.C. FRANK. 2009: Persistent positive North Atlantic Oscillation mode
dominated the Medieval Climate Anomaly. Science 324: 78 - 80.
VANDENPUT R., 1981. Les principales cultures en Afrique centrale.
Tournai: édit. Lesaffre, 458
VITTOZ Pascal, 2002. : Dendrochronologie et dynamique
forestière au pays des Kiwis. Walker, B.H. 1992. Biodiversity and
écological redundancy. Conservation Biology, 6: 18-22.
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Walker, B.H. 1995. Conservising Biological diversity throught
ecosystem resilience Conservation Biology, 9: 747-752.
WCMC, 1998: Contribution to an evaluation of tree species
using the new CITES Listing Criteria prepared by the UNEP World Conservation
Monitoring Centre (UNEPWCMC) on behalf of the CITES Management Authority of the
Netherlands.


Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la zone cambiale dans les Réserves de Yoko et
de Yangambi (Province
Orientale/RDC)
ANNEXES
Annexe 1: Fonds floristique du bloc
d'aménagement
(N= Nombre d'individus de chaque espèce ; FR= Nombre
d'unités d'échantillonnage contenant l'espèce ; G= Surface
terrière ; DOR= Dominance relative ; DER= Densité relative ; FRR=
Fréquence relative ; IVI= Indice de l'importance relative de
l'espèce)
|
Famille
|
Espèces
|
N
|
FR
|
G
(m2/ha)
|
DER
(%)
|
DOR
(%)
|
FRR
(%)
|
IVI
|
|
Anacardiaceae
|
Antrocaryon nannanii De Wild.
|
3
|
2
|
0,066
|
0,028
|
0,186
|
0,326
|
0,541
|
|
Total Anacardiaceae
|
3
|
2
|
0,066
|
0,028
|
0,186
|
0,326
|
0,541
|
|
Annonaceae
|
Anonidium mannii (Oliv.) Engl. & Diels
|
273
|
4
|
1,443
|
2,576
|
4,064
|
0,653
|
7,293
|
|
Cleistopholis glauca Pierre ex Engl. & Diels
|
2
|
2
|
0,014
|
0,019
|
0,039
|
0,326
|
0,385
|
|
Greenwayodendron suaveolens
|
76
|
1
|
0,233
|
0,717
|
0,657
|
0,163
|
1,538
|
|
Isolona hexaloba (Pierre) Engl. & Diels
|
14
|
2
|
0,064
|
0,132
|
0,179
|
0,326
|
0,637
|
|
Isolona thonneri (De Wild. & T. Durand) Engl. &
Diels
|
56
|
4
|
0,075
|
0,529
|
0,211
|
0,653
|
1,392
|
|
Monodora myristica (Gaertn.) Dunal
|
60
|
2
|
0,126
|
0,566
|
0,356
|
0,326
|
1,249
|
|
Polyalthia suaveolens Engl. & Diels
|
224
|
3
|
0,742
|
2,114
|
2,089
|
0,489
|
4,693
|
|
Xylopia aethiopica (Dunal) A.Rich.
|
7
|
3
|
0,048
|
0,066
|
0,136
|
0,489
|
0,691
|
|
Xylopia africana (Benth.) Oliv.
|
4
|
2
|
0,020
|
0,038
|
0,057
|
0,326
|
0,421
|
|
Xylopia parviflora (A. Rich.) Benth.
|
1
|
1
|
0,006
|
0,009
|
0,018
|
0,163
|
0,190
|
|
Total Annonaceae
|
717
|
24
|
2,772
|
6,767
|
7,807
|
3,915
|
18,489
|
|
Apocynaceae
|
Alstonia boonei De Wild.
|
3
|
2
|
0,017
|
0,028
|
0,049
|
0,326
|
0,404
|
|
Funtumia africana (Benth.) Stapf
|
4
|
3
|
0,004
|
0,038
|
0,012
|
0,489
|
0,539
|
|
Funtumia elastica (Preuss) Stapf
|
14
|
4
|
0,063
|
0,132
|
0,176
|
0,653
|
0,961
|
|
Pleiocarpa pycnantha (K. Schum.) Stapf
|
16
|
3
|
0,018
|
0,151
|
0,050
|
0,489
|
0,690
|
|
Pleiocarpa pycnantha (K. Schum.) Stapf
|
30
|
2
|
0,023
|
0,283
|
0,064
|
0,326
|
0,674
|
|
Tabernaemontana durissima Stapf
|
106
|
4
|
0,132
|
1,000
|
0,371
|
0,653
|
2,024
|
|
Voacanga chalotiana
|
2
|
1
|
0,022
|
0,019
|
0,061
|
0,163
|
0,243
|




Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la zone cambiale dans les Réserves de Yoko et
de Yangambi (Province
Orientale/RDC)
|
Famille
|
Espèces
|
N
|
FR
|
G
(m2/ha)
|
DER
(%)
|
DOR
(%)
|
FRR
(%)
|
IVI
|
|
Total Apocynaceae
|
175
|
19
|
0,278
|
1,652
|
0,783
|
3,100
|
5,534
|
|
Bignoniaceae
|
Fernandoa adolfi-frederici (Gilg & Mildbr.) Heine
|
3
|
1
|
0,028
|
0,028
|
0,077
|
0,163
|
0,269
|
|
Total Bignoniaceae
|
3
|
1
|
0,028
|
0,028
|
0,077
|
0,163
|
0,269
|
|
Burseraceae
|
Canarium schweinfurthii Engl.
|
7
|
3
|
0,032
|
0,066
|
0,089
|
0,489
|
0,644
|
|
Dacryodes edulis (G. Don) H.J. Lam
|
17
|
4
|
0,034
|
0,160
|
0,097
|
0,653
|
0,910
|
|
Dacryodes yangambiensis Louis & Troupin
|
10
|
4
|
0,022
|
0,094
|
0,062
|
0,653
|
0,809
|
|
Total Burseraceae
|
34
|
11
|
0,088
|
0,321
|
0,248
|
1,794
|
2,363
|
|
Chrysobalanaceae
|
Magnistipula butayei
|
2
|
1
|
0,003
|
0,019
|
0,009
|
0,163
|
0,191
|
|
Maranthes glabra (Oliv.) Prance
|
5
|
2
|
0,016
|
0,047
|
0,046
|
0,326
|
0,420
|
|
Parinari excelsa Sabine
|
4
|
1
|
0,007
|
0,038
|
0,018
|
0,163
|
0,219
|
|
Total Chrysobalanaceae
|
11
|
4
|
0,026
|
0,104
|
0,073
|
0,653
|
0,830
|
|
Clusiaceae
|
Allanblackia floribunda Oliv.
|
38
|
4
|
0,091
|
0,359
|
0,256
|
0,653
|
1,267
|
|
Garcinia cf. letestui Pellegrin
|
1
|
1
|
0,000
|
0,009
|
0,001
|
0,163
|
0,174
|
|
Garcinia epunctata Stapf
|
39
|
4
|
0,041
|
0,368
|
0,115
|
0,653
|
1,136
|
|
Garcinia kola Heckel
|
2
|
2
|
0,003
|
0,019
|
0,009
|
0,326
|
0,354
|
|
Garcinia ovalifolia Oliv.
|
1
|
1
|
0,001
|
0,009
|
0,003
|
0,163
|
0,175
|
|
Garcinia punctata Oliv.
|
190
|
4
|
0,282
|
1,793
|
0,793
|
0,653
|
3,239
|
|
Mammea africana Sabine
|
1
|
1
|
0,002
|
0,009
|
0,006
|
0,163
|
0,178
|
|
Symphonia globulifera L.
|
36
|
4
|
0,202
|
0,340
|
0,569
|
0,653
|
1,561
|
|
Total Clusiaceae
|
308
|
21
|
0,622
|
2,907
|
1,753
|
3,426
|
8,085
|
|
Combretaceae
|
Combretum lokele Liben
|
26
|
4
|
0,187
|
0,245
|
0,527
|
0,653
|
1,425
|
|
Combretum zeyheri Sond.
|
10
|
4
|
0,010
|
0,094
|
0,028
|
0,653
|
0,775
|
|
Pteleopsis hylodendron Mildbr.
|
8
|
4
|
0,053
|
0,076
|
0,150
|
0,653
|
0,878
|
|
Total Combretaceae
|
44
|
12
|
0,250
|
0,415
|
0,705
|
1,958
|
3,078
|
|
Dichapetalaceae
|
Dichapetalum sp.
|
1
|
1
|
0,001
|
0,009
|
0,002
|
0,163
|
0,175
|
|
Total Dichapetalaceae
|
1
|
1
|
0,001
|
0,009
|
0,002
|
0,163
|
0,175
|
|
Ebenaceae
|
Diospyros boala De Wild.
|
83
|
4
|
0,206
|
0,783
|
0,581
|
0,653
|
2,017
|
|
Diospyros crassiflora Hiern
|
64
|
4
|
0,151
|
0,604
|
0,426
|
0,653
|
1,683
|
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la zone cambiale dans les Réserves de Yoko et
de Yangambi (Province
Orientale/RDC)
|
Famille
|
Espèces
|
N
|
FR
|
G
(m2/ha)
|
DER
(%)
|
DOR
(%)
|
FRR
(%)
|
IVI
|
|
Diospyros deltoidea F. White
|
6
|
2
|
0,006
|
0,057
|
0,018
|
0,326
|
0,401
|
|
Diospyros ebenum Koenig ex Retz.
|
43
|
4
|
0,111
|
0,406
|
0,312
|
0,653
|
1,370
|
|
Diospyros hoyleana F. White
|
1
|
1
|
0,001
|
0,009
|
0,004
|
0,163
|
0,177
|
|
Diospyros lancifolia Roxb.
|
2
|
2
|
0,002
|
0,019
|
0,005
|
0,326
|
0,350
|
|
Maba laurentii De Wild.
|
5
|
2
|
0,008
|
0,047
|
0,022
|
0,326
|
0,396
|
|
Total Ebenaceae
|
204
|
19
|
0,486
|
1,925
|
1,369
|
3,100
|
6,394
|
|
Euphorbiaceae
|
Antidesma laciniatum Müll. Arg.
|
2
|
1
|
0,003
|
0,019
|
0,007
|
0,163
|
0,189
|
|
Antidesma membranaceum
|
1
|
1
|
0,000
|
0,009
|
0,001
|
0,163
|
0,174
|
|
Cleistanthus polystachyus Hook.f. ex Planch.
|
14
|
2
|
0,055
|
0,132
|
0,154
|
0,326
|
0,612
|
|
Cleistanthus ripicola J. Léonard
|
1
|
1
|
0,016
|
0,009
|
0,044
|
0,163
|
0,216
|
|
Cleistanthus sp.
|
6
|
2
|
0,012
|
0,057
|
0,034
|
0,326
|
0,417
|
|
Discoglypremna caloneura (Pax) Prain
|
6
|
2
|
0,018
|
0,057
|
0,051
|
0,326
|
0,434
|
|
Drypetes cf. trunci spinosa
|
3
|
3
|
0,006
|
0,028
|
0,016
|
0,489
|
0,534
|
|
Drypetes gossweileri S. Moore
|
177
|
4
|
0,937
|
1,670
|
2,638
|
0,653
|
4,961
|
|
Drypetes leonensis Pax
|
59
|
5
|
0,129
|
0,557
|
0,362
|
0,816
|
1,735
|
|
Drypetes likwa J. Léonard
|
56
|
5
|
0,125
|
0,529
|
0,353
|
0,816
|
1,697
|
|
Drypetes lisolinoli J. Leonard
|
19
|
3
|
0,037
|
0,179
|
0,105
|
0,489
|
0,773
|
|
Drypetes louisii J. Leonard.
|
39
|
4
|
0,114
|
0,368
|
0,321
|
0,653
|
1,342
|
|
Drypetes sp.
|
1
|
1
|
0,002
|
0,009
|
0,005
|
0,163
|
0,178
|
|
Drypetes spinosodentata (Pax) Hutch.
|
7
|
4
|
0,021
|
0,066
|
0,059
|
0,653
|
0,777
|
|
Hymenocardia ulmoides Oliv.
|
9
|
2
|
0,007
|
0,085
|
0,020
|
0,326
|
0,432
|
|
Macaranga capensis (Baill.) Benth. ex Sim
|
1
|
1
|
0,007
|
0,009
|
0,018
|
0,163
|
0,191
|
|
Macaranga lancifolia Pax
|
13
|
4
|
0,022
|
0,123
|
0,062
|
0,653
|
0,837
|
|
Macaranga laurentii De Wild.
|
2
|
1
|
0,002
|
0,019
|
0,006
|
0,163
|
0,188
|
|
Macaranga spinosa Müll.Arg.
|
3
|
3
|
0,003
|
0,028
|
0,007
|
0,489
|
0,525
|
|
Macaranga zenkeri Pax
|
4
|
2
|
0,007
|
0,038
|
0,020
|
0,326
|
0,384
|
|
Microdesmis puberula Hook. f. ex Planch.
|
4
|
3
|
0,002
|
0,038
|
0,005
|
0,489
|
0,533
|
|
Phyllanthus discoideus (Baill.) Müll.Arg.
|
1
|
1
|
0,006
|
0,009
|
0,016
|
0,163
|
0,188
|




Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la zone cambiale dans les Réserves de Yoko et
de Yangambi (Province
Orientale/RDC)
|
Famille
|
Espèces
|
N
|
FR
|
G
(m2/ha)
|
DER
(%)
|
DOR
(%)
|
FRR
(%)
|
IVI
|
|
Ricinodendron heudelotii (Baill.) Pierre ex Heckel
|
6
|
3
|
0,095
|
0,057
|
0,268
|
0,489
|
0,814
|
|
Tetrorchidium didymostemon (Baill.) Pax & K.Hoffm
|
5
|
2
|
0,009
|
0,047
|
0,026
|
0,326
|
0,399
|
|
Uapaca guineensis Müll. Arg.
|
15
|
3
|
0,077
|
0,142
|
0,216
|
0,489
|
0,847
|
|
Uapaca heudelotii Baill.
|
2
|
2
|
0,004
|
0,019
|
0,012
|
0,326
|
0,357
|
|
Total Euphorbiaceae
|
456
|
65
|
1,714
|
4,304
|
4,827
|
10,604
|
19,734
|
|
Fabaceae
|
Afzelia bella Harms
|
3
|
1
|
0,002
|
0,028
|
0,006
|
0,163
|
0,197
|
|
Albizia adianthifolia (Schumach.) W.Wight
|
6
|
4
|
0,020
|
0,057
|
0,056
|
0,653
|
0,765
|
|
Albizia gummifera (J.F.Gmel.) C.A.Sm.
|
8
|
4
|
0,107
|
0,076
|
0,301
|
0,653
|
1,029
|
|
Albizia laurentii De Wild.
|
13
|
4
|
0,030
|
0,123
|
0,083
|
0,653
|
0,859
|
|
Angylocalyx pynaertii De Wild.
|
2
|
1
|
0,002
|
0,019
|
0,005
|
0,163
|
0,187
|
|
Anthonotha fragrans (Baker f.) Exell & Hellcoat
|
2
|
2
|
0,001
|
0,019
|
0,003
|
0,326
|
0,349
|
|
Anthonotha macrophylla P.Beauv.
|
18
|
4
|
0,015
|
0,170
|
0,041
|
0,653
|
0,864
|
|
Baikiaea insignis Benth.
|
4
|
2
|
0,008
|
0,038
|
0,021
|
0,326
|
0,385
|
|
Baphia laurentii De Wild.
|
1
|
1
|
0,001
|
0,009
|
0,002
|
0,163
|
0,175
|
|
Baphia laurifolia Baill.
|
13
|
4
|
0,017
|
0,123
|
0,048
|
0,653
|
0,824
|
|
Berlinia acuminata
|
1
|
1
|
0,001
|
0,009
|
0,003
|
0,163
|
0,175
|
|
Cynometra alexandri C.H. Wright
|
1
|
1
|
0,000
|
0,009
|
0,001
|
0,163
|
0,174
|
|
Cynometra hankei Harms
|
34
|
3
|
0,326
|
0,321
|
0,919
|
0,489
|
1,729
|
|
Dialium congolensis De Wild. & T.Durand
|
1
|
1
|
0,000
|
0,009
|
0,001
|
0,163
|
0,174
|
|
Dialium excelsium J. Louis ex Stey.
|
8
|
2
|
0,094
|
0,076
|
0,265
|
0,326
|
0,667
|
|
Dialium leonensis Pax
|
1
|
1
|
0,001
|
0,009
|
0,002
|
0,163
|
0,175
|
|
Dialium pachyphyllum Harms
|
100
|
4
|
0,148
|
0,944
|
0,417
|
0,653
|
2,013
|
|
Dialium polyanthum Harms
|
40
|
4
|
0,040
|
0,378
|
0,112
|
0,653
|
1,142
|
|
Dialium sp.
|
4
|
2
|
0,004
|
0,038
|
0,011
|
0,326
|
0,375
|
|
Erythrophleum lasianthum Corbishley
|
23
|
4
|
0,323
|
0,217
|
0,909
|
0,653
|
1,779
|
|
Fillaeopsis discophora Harms
|
1
|
1
|
0,013
|
0,009
|
0,036
|
0,163
|
0,208
|
|
Gilbertiodendron dewevrei (De Wild.) J.
Léonard
|
52
|
3
|
0,188
|
0,491
|
0,530
|
0,489
|
1,510
|
|
Gilletiodendron mildbraedii (Harms) Vermoesen
|
5
|
3
|
0,009
|
0,047
|
0,024
|
0,489
|
0,561
|
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la zone cambiale dans les Réserves de Yoko et
de Yangambi (Province
Orientale/RDC)
|
Famille
|
Espèces
|
N
|
FR
|
G
(m2/ha)
|
DER
(%)
|
DOR
(%)
|
FRR
(%)
|
IVI
|
|
Guibourtia demeusei (Harms) J. Léonard
|
1
|
1
|
0,000
|
0,009
|
0,001
|
0,163
|
0,174
|
|
Millettia drastica Welw.
|
7
|
3
|
0,019
|
0,066
|
0,054
|
0,489
|
0,609
|
|
Millettia dubia De Wild.
|
3
|
2
|
0,003
|
0,028
|
0,008
|
0,326
|
0,363
|
|
Millettia sp.
|
1
|
1
|
0,001
|
0,009
|
0,002
|
0,163
|
0,174
|
|
Paramacrolobium coeruleum (Taub.) J. Léonard
|
26
|
4
|
0,042
|
0,245
|
0,120
|
0,653
|
1,017
|
|
Parkia bicolor A. Chev.
|
1
|
1
|
0,001
|
0,009
|
0,002
|
0,163
|
0,174
|
|
Parkia filicoidea Welw. ex Oliv.
|
3
|
2
|
0,007
|
0,028
|
0,020
|
0,326
|
0,374
|
|
Pentaclethra macrophylla Benth.
|
56
|
4
|
0,194
|
0,529
|
0,547
|
0,653
|
1,728
|
|
Pericopsis elata (Harms) Van Meeuwen
|
429
|
4
|
1,222
|
4,049
|
3,731
|
0,653
|
8,432
|
|
Piptadeniastrum africanum (Hook.f.) Brenan
|
10
|
4
|
0,179
|
0,094
|
0,503
|
0,653
|
1,250
|
|
Prioria balsamifera (Vermoesen) Breteler
|
47
|
4
|
0,257
|
0,444
|
0,724
|
0,653
|
1,820
|
|
Prioria oxyphylla (Harms) Breteler
|
63
|
4
|
0,450
|
0,595
|
1,268
|
0,653
|
2,515
|
|
Pterocarpus soyauxii Taub.
|
65
|
4
|
0,425
|
0,613
|
1,196
|
0,653
|
2,462
|
|
Schotia romii De Wild.
|
137
|
4
|
0,307
|
1,293
|
0,865
|
0,653
|
2,810
|
|
Scorodophloeus zenkeri Harms
|
833
|
4
|
5,686
|
7,861
|
16,011
|
0,653
|
24,525
|
|
Sp2 non identifiée
|
17
|
1
|
0,042
|
0,160
|
0,118
|
0,163
|
0,442
|
|
Tessmania africana Harms
|
3
|
1
|
0,012
|
0,028
|
0,032
|
0,163
|
0,224
|
|
Tetrapleura tetraptera (Schumach. & Thonn.) Taub.
|
10
|
4
|
0,024
|
0,094
|
0,068
|
0,653
|
0,814
|
|
Total Fabaceae
|
2053
|
109
|
10,322
|
19,375
|
29,066
|
17,781
|
66,223
|
|
Flacourtiaceae
|
Barteria fistulosa Mast.
|
29
|
3
|
0,022
|
0,274
|
0,063
|
0,489
|
0,826
|
|
Barteria nigritiana Hook.f.
|
18
|
4
|
0,019
|
0,170
|
0,053
|
0,653
|
0,875
|
|
Homalium laurentii De Wild.
|
12
|
4
|
0,017
|
0,113
|
0,049
|
0,653
|
0,815
|
|
Homalium sp.
|
17
|
4
|
0,013
|
0,160
|
0,036
|
0,653
|
0,849
|
|
Oncoba glauca (P. Beauv.) Gilg.
|
11
|
3
|
0,010
|
0,104
|
0,029
|
0,489
|
0,622
|
|
Oncoba welwitschii Oliv.
|
9
|
2
|
0,013
|
0,085
|
0,037
|
0,326
|
0,448
|
|
Total Flacourtiaceae
|
96
|
20
|
0,095
|
0,906
|
0,266
|
3,263
|
4,435
|
|
Huaceae
|
Afrostyrax lepidophyllus Mildbr.
|
11
|
4
|
0,035
|
0,104
|
0,098
|
0,653
|
0,854
|
|
Hua gabonii Pierre ex De Wild.
|
6
|
2
|
0,003
|
0,057
|
0,009
|
0,326
|
0,392
|




Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la zone cambiale dans les Réserves de Yoko et
de Yangambi (Province
Orientale/RDC)
|
Famille
|
Espèces
|
N
|
FR
|
G
(m2/ha)
|
DER
(%)
|
DOR
(%)
|
FRR
(%)
|
IVI
|
|
Total Huaceae
|
17
|
6
|
0,038
|
0,160
|
0,107
|
0,979
|
1,246
|
|
Irvingiaceae
|
Irvingia gabonensis (Aubry-Lecomte ex O'Rorke) Baill.
|
2
|
2
|
0,032
|
0,019
|
0,090
|
0,326
|
0,435
|
|
Irvingia grandifolia (Engl.) Engl.
|
21
|
4
|
0,185
|
0,198
|
0,522
|
0,653
|
1,373
|
|
Klainedoxa gabonensis Pierre ex Engl.
|
34
|
4
|
0,264
|
0,321
|
0,743
|
0,653
|
1,716
|
|
Total Irvingiaceae
|
57
|
10
|
0,481
|
0,538
|
1,354
|
1,631
|
3,524
|
|
Lauraceae
|
Beilschmiedia congolana Robyns & Wilczek
|
5
|
2
|
0,018
|
0,047
|
0,050
|
0,326
|
0,424
|
|
Beilschmiedia corbisieri (Robyns) Robyns &
Wilczek
|
13
|
3
|
0,023
|
0,123
|
0,066
|
0,489
|
0,678
|
|
Beilschmiedia euryneura (Stapf) Robyns & Wilczek
|
6
|
1
|
0,004
|
0,057
|
0,012
|
0,163
|
0,231
|
|
Beilschmiedia gilbertii Robyns & Wilczek
|
1
|
1
|
0,005
|
0,009
|
0,013
|
0,163
|
0,185
|
|
Beilschmiedia yangambiensis Robyns & R.Wilczek
|
1
|
1
|
0,047
|
0,009
|
0,133
|
0,163
|
0,306
|
|
Total Lauraceae
|
26
|
8
|
0,097
|
0,245
|
0,274
|
1,305
|
1,824
|
|
Lecythidaceae
|
Napoleona imperialis P. Beauv.
|
8
|
2
|
0,005
|
0,076
|
0,014
|
0,326
|
0,416
|
|
Petersianthus macrocarpus (P. Beauv.) Liben
|
374
|
4
|
1,579
|
3,530
|
4,445
|
0,653
|
8,627
|
|
Total Lecythidaceae
|
382
|
6
|
1,584
|
3,605
|
4,460
|
0,979
|
9,044
|
|
Linaceae
|
Ochthocosmus africanus Hook. f.
|
5
|
3
|
0,010
|
0,047
|
0,029
|
0,489
|
0,566
|
|
Total Linaceae
|
5
|
3
|
0,010
|
0,047
|
0,029
|
0,489
|
0,566
|
|
Malvaceae/Sterculioideae
|
Cola acuminata (P. Beauv.) Schott. & Endl.
|
15
|
3
|
0,016
|
0,142
|
0,046
|
0,489
|
0,677
|
|
Cola altissima Engl.
|
27
|
4
|
0,029
|
0,255
|
0,081
|
0,653
|
0,988
|
|
Cola griseiflora De Wild.
|
366
|
4
|
0,408
|
3,454
|
1,148
|
0,653
|
5,254
|
|
Cola lateritia K.Schum.
|
3
|
2
|
0,009
|
0,028
|
0,025
|
0,326
|
0,380
|
|
Leptonychia batangensis (C. H. Wright) Burret
|
3
|
1
|
0,002
|
0,028
|
0,007
|
0,163
|
0,198
|
|
Pterygota bequaertii De Wild.
|
6
|
3
|
0,058
|
0,057
|
0,164
|
0,489
|
0,710
|
|
Scaphopetalum thonnerii De Wild. & T. Durand
|
4
|
3
|
0,002
|
0,038
|
0,006
|
0,489
|
0,533
|
|
Sterculia bequaertii De Wild.
|
7
|
3
|
0,052
|
0,066
|
0,147
|
0,489
|
0,702
|
|
Total Malvaceae/Sterculioideae
|
431
|
23
|
0,577
|
4,068
|
1,624
|
3,752
|
9,443
|
|
Melastomataceae
|
Bellucia axinanthera Triana
|
35
|
3
|
0,058
|
0,330
|
0,164
|
0,489
|
0,984
|
|
Memecylon cyaneum De Wild.
|
1
|
1
|
0,001
|
0,009
|
0,002
|
0,163
|
0,175
|
|
Total Melastomataceae
|
36
|
4
|
0,059
|
0,340
|
0,166
|
0,653
|
1,159
|
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la zone cambiale dans les Réserves de Yoko et
de Yangambi (Province
Orientale/RDC)
|
Famille
|
Espèces
|
N
|
FR
|
G
(m2/ha)
|
DER
(%)
|
DOR
(%)
|
FRR
(%)
|
IVI
|
|
Meliaceae
|
Carapa procera DC.
|
294
|
4
|
0,274
|
2,775
|
0,772
|
0,653
|
4,200
|
|
Entandrophragma angolense (Welw.) C.DC.
|
57
|
4
|
0,636
|
0,538
|
1,791
|
0,653
|
2,982
|
|
Entandrophragma candollei Harms
|
12
|
4
|
0,112
|
0,113
|
0,316
|
0,653
|
1,082
|
|
Entandrophragma cylindricum (Sprague) Sprague
|
20
|
4
|
0,241
|
0,189
|
0,679
|
0,653
|
1,521
|
|
Entandrophragma utile (Dawe & Sprague) Sprague
|
1
|
1
|
0,006
|
0,009
|
0,016
|
0,163
|
0,188
|
|
Guarea cedrata (A.Chev.) Pellegr.
|
173
|
6
|
0,708
|
1,633
|
1,994
|
0,979
|
4,605
|
|
Guarea laurentii De Wild.
|
176
|
2
|
0,521
|
1,661
|
1,468
|
0,326
|
3,455
|
|
Guarea thompsonii Sprague & Hutch.
|
332
|
4
|
0,799
|
3,133
|
2,250
|
0,653
|
6,035
|
|
Khaya anthotheca (Welw.) C. DC.
|
1
|
1
|
0,001
|
0,009
|
0,001
|
0,163
|
0,174
|
|
Lovoa trichiloides Harms
|
10
|
3
|
0,075
|
0,094
|
0,210
|
0,489
|
0,794
|
|
Trichilia gilgiana Harms
|
142
|
4
|
0,239
|
1,340
|
0,672
|
0,653
|
2,665
|
|
Trichilia laurentii De Wild.
|
13
|
2
|
0,026
|
0,123
|
0,074
|
0,326
|
0,523
|
|
Trichilia monadelpha (Thonn.) J. J. de Wilde
|
46
|
2
|
0,149
|
0,434
|
0,421
|
0,326
|
1,181
|
|
Trichilia rubescens Oliv.
|
37
|
4
|
0,097
|
0,349
|
0,273
|
0,653
|
1,275
|
|
Trichilia tessmannii Harms
|
28
|
1
|
0,081
|
0,264
|
0,229
|
0,163
|
0,657
|
|
Trichilia welwitschii C. DC.
|
34
|
3
|
0,062
|
0,321
|
0,175
|
0,489
|
0,985
|
|
Total Meliaceae
|
1376
|
49
|
4,028
|
12,986
|
11,343
|
7,993
|
32,322
|
|
Moraceae
|
Antiaris welwitschii Engl.
|
9
|
4
|
0,043
|
0,085
|
0,121
|
0,653
|
0,858
|
|
Milicia excelsa (Welw.) C. C. Berg.
|
1
|
1
|
0,010
|
0,009
|
0,029
|
0,163
|
0,202
|
|
Musanga cecropioides R. Br.
|
28
|
4
|
0,116
|
0,264
|
0,326
|
0,653
|
1,242
|
|
Myrianthus arboreus P. Beauv.
|
20
|
4
|
0,033
|
0,189
|
0,094
|
0,653
|
0,935
|
|
Treculia africana Desc.
|
13
|
4
|
0,021
|
0,123
|
0,060
|
0,653
|
0,835
|
|
Trilepisium madagascariense DC.
|
139
|
4
|
0,622
|
1,312
|
1,751
|
0,653
|
3,716
|
|
Total Moraceae
|
210
|
21
|
0,845
|
1,982
|
2,381
|
3,426
|
7,788
|
|
Myristicaceae
|
Coelocaryon botryoides Vermoesen
|
45
|
2
|
0,079
|
0,425
|
0,223
|
0,326
|
0,974
|
|
Coelocaryon nannanii De Wild.
|
1
|
1
|
0,002
|
0,009
|
0,007
|
0,163
|
0,179
|
|
Coelocaryon preussii Warb.
|
222
|
4
|
0,229
|
2,095
|
0,644
|
0,653
|
3,391
|
|
Pycnanthus angolensis (Welw.) Warb.
|
88
|
4
|
0,121
|
0,831
|
0,341
|
0,653
|
1,824
|




Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la zone cambiale dans les Réserves de Yoko et
de Yangambi (Province
Orientale/RDC)
|
Famille
|
Espèces
|
N
|
FR
|
G
(m2/ha)
|
DER
(%)
|
DOR
(%)
|
FRR
(%)
|
IVI
|
|
Staudtia stipitata Warb.
|
694
|
4
|
1,296
|
6,550
|
3,650
|
0,653
|
10,852
|
|
Total Myristicaceae
|
1050
|
15
|
1,727
|
9,909
|
4,864
|
2,447
|
17,221
|
|
Olacaceae
|
Aplandra zenkeri Engl.
|
3
|
2
|
0,003
|
0,028
|
0,009
|
0,326
|
0,364
|
|
Heisteria parvifolia Sm.
|
2
|
2
|
0,001
|
0,019
|
0,002
|
0,326
|
0,347
|
|
Olax gambecola Baill.
|
542
|
4
|
0,851
|
5,115
|
2,396
|
0,653
|
8,163
|
|
Olax latifolia Engl.
|
1
|
1
|
0,001
|
0,009
|
0,004
|
0,163
|
0,176
|
|
Ongokea gore (Hua) Pierre
|
14
|
4
|
0,158
|
0,132
|
0,444
|
0,653
|
1,229
|
|
Strombosia glaucescens Engl.
|
87
|
4
|
0,403
|
0,821
|
1,136
|
0,653
|
2,609
|
|
Strombosia grandifolia Hook. f. ex Benth.
|
119
|
4
|
0,481
|
1,123
|
1,356
|
0,653
|
3,131
|
|
Strombosia nigropunctata J. Louis & J.
Léonard
|
53
|
3
|
0,323
|
0,500
|
0,910
|
0,489
|
1,899
|
|
Strombosia pustulata Oliv.
|
1
|
1
|
0,006
|
0,009
|
0,018
|
0,163
|
0,190
|
|
Strombosia sp.
|
1
|
1
|
0,011
|
0,009
|
0,032
|
0,163
|
0,205
|
|
Strombosiopsis tetrandra Engl.
|
36
|
4
|
0,457
|
0,340
|
1,288
|
0,653
|
2,280
|
|
Total Olacaceae
|
859
|
30
|
2,697
|
8,107
|
7,594
|
4,894
|
20,595
|
|
Pandaceae
|
Panda oleosa Pierre
|
199
|
4
|
0,912
|
1,878
|
2,569
|
0,653
|
5,100
|
|
Total Pandaceae
|
199
|
4
|
0,912
|
1,878
|
2,569
|
0,653
|
5,100
|
|
Rhamnaceae
|
Maesopsis eminii Engl.
|
4
|
3
|
0,017
|
0,038
|
0,047
|
0,489
|
0,574
|
|
Total Rhamnaceae
|
4
|
3
|
0,017
|
0,038
|
0,047
|
0,489
|
0,574
|
|
Rubiaceae
|
Aidia micrantha (K. Schum.) Bullock ex F. White
|
10
|
3
|
0,006
|
0,094
|
0,017
|
0,489
|
0,601
|
|
Bertiera dewevrei De Wild. & T.Durand
|
1
|
1
|
0,000
|
0,009
|
0,001
|
0,163
|
0,174
|
|
Bertiera parviflora Spruce ex K.Schum.
|
1
|
1
|
0,001
|
0,009
|
0,002
|
0,163
|
0,174
|
|
Colletoecema dewevrei (De Wild.) E.M.A.Petit
|
4
|
2
|
0,005
|
0,038
|
0,015
|
0,326
|
0,379
|
|
Craterispermum cerinantum Hiern
|
5
|
2
|
0,008
|
0,047
|
0,024
|
0,326
|
0,397
|
|
Dictyandra arborescens Welw. ex Hook.f.
|
1
|
1
|
0,001
|
0,009
|
0,002
|
0,163
|
0,174
|
|
Gardenia tchibangensis Pellegr.
|
42
|
5
|
0,042
|
0,396
|
0,117
|
0,816
|
1,330
|
|
Leptactina angolensis (Hutch.) Bullock ex I.Nogueira
|
1
|
1
|
0,000
|
0,009
|
0,001
|
0,163
|
0,174
|
|
Massularia acuminata (G. Don) Bullock
|
15
|
3
|
0,008
|
0,142
|
0,022
|
0,489
|
0,653
|
|
Morinda lucida Benth.
|
1
|
1
|
0,001
|
0,009
|
0,004
|
0,163
|
0,176
|
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la zone cambiale dans les Réserves de Yoko et
de Yangambi (Province
Orientale/RDC)
|
Famille
|
Espèces
|
N
|
FR
|
G
(m2/ha)
|
DER
(%)
|
DOR
(%)
|
FRR
(%)
|
IVI
|
|
Pausinystalia pynaertii De Wild.
|
1
|
1
|
0,001
|
0,009
|
0,003
|
0,163
|
0,175
|
|
Psydrax anordianum (De Wild. & T. Durand) Bridson
|
2
|
2
|
0,002
|
0,019
|
0,005
|
0,326
|
0,350
|
|
Psydrax palma (K.Schum.) Bridson
|
3
|
2
|
0,011
|
0,028
|
0,032
|
0,326
|
0,387
|
|
Randia africana G. Den
|
22
|
4
|
0,011
|
0,208
|
0,032
|
0,653
|
0,892
|
|
Randia congolana De Wild. & T. Durand
|
8
|
4
|
0,004
|
0,076
|
0,012
|
0,653
|
0,740
|
|
Randia sp.
|
2
|
1
|
0,001
|
0,019
|
0,004
|
0,163
|
0,186
|
|
Sarcocephalus diderrichii De Wild. & T. Durand
|
13
|
4
|
0,094
|
0,123
|
0,264
|
0,653
|
1,039
|
|
Sp1 non identifiée
|
2
|
1
|
0,008
|
0,019
|
0,024
|
0,163
|
0,206
|
|
Total Rubiaceae
|
134
|
39
|
0,206
|
1,265
|
0,579
|
6,362
|
8,206
|
|
Rutaceae
|
Zanthoxylum lemaierei (De Wild.) Waterm.
|
3
|
1
|
0,007
|
0,028
|
0,020
|
0,163
|
0,211
|
|
Zanthoxylum lemaierei (De Wild.) Waterm.
|
2
|
1
|
0,008
|
0,019
|
0,021
|
0,163
|
0,203
|
|
Zanthoxylum gilletii (De Wild.) Waterm.
|
1
|
1
|
0,000
|
0,009
|
0,001
|
0,163
|
0,174
|
|
Total Rutaceae
|
6
|
3
|
0,015
|
0,057
|
0,042
|
0,489
|
0,588
|
|
Sapindaceae
|
Allophylus africanus P. Beauv.
|
7
|
3
|
0,007
|
0,066
|
0,020
|
0,489
|
0,575
|
|
Blighia unijugata Baker
|
1
|
1
|
0,002
|
0,009
|
0,007
|
0,163
|
0,179
|
|
Blighia welwitschii (Hiern) Radlk.
|
25
|
4
|
0,256
|
0,236
|
0,722
|
0,653
|
1,611
|
|
Chytranthus macrobotrys (Gilg) Exell &
Mendonça
|
3
|
1
|
0,003
|
0,028
|
0,010
|
0,163
|
0,201
|
|
Pancovia harmsiana Gilg.
|
350
|
4
|
0,367
|
3,303
|
1,034
|
0,653
|
4,990
|
|
Pancovia laurentii (De Wild.) Gilg. ex De Wild.
|
80
|
4
|
0,119
|
0,755
|
0,334
|
0,653
|
1,741
|
|
Total Sapindaceae
|
466
|
17
|
0,755
|
4,398
|
2,126
|
2,773
|
9,297
|
|
Sapotaceae
|
Autranella congolensis (De Wild.) A.Chev.
|
2
|
2
|
0,001
|
0,019
|
0,004
|
0,326
|
0,349
|
|
Chrysophyllum africanum A.DC.
|
140
|
4
|
0,444
|
1,321
|
1,249
|
0,653
|
3,223
|
|
Chrysophyllum beguei Aubrév. & Pellegr.
|
3
|
1
|
0,004
|
0,028
|
0,010
|
0,163
|
0,202
|
|
Chrysophyllum lacourtianum De Wild.
|
103
|
4
|
0,617
|
0,972
|
1,737
|
0,653
|
3,361
|
|
Chrysophyllum perpulchrum Mildbr.
|
1
|
1
|
0,003
|
0,009
|
0,009
|
0,163
|
0,181
|
|
Chrysophyllum pruniforme Pierre ex Engl.
|
7
|
3
|
0,039
|
0,066
|
0,110
|
0,489
|
0,665
|
|
Synsepalum subcordatum De Wild.
|
16
|
1
|
0,064
|
0,151
|
0,181
|
0,163
|
0,495
|
|
Synsepalum stipulatum (Radlk.) Engl.
|
24
|
4
|
0,035
|
0,227
|
0,099
|
0,653
|
0,978
|
Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la zone cambiale dans les Réserves de Yoko et
de Yangambi (Province
Orientale/RDC)
|
Famille
|
Espèces
|
N
|
FR
|
G
(m2/ha)
|
DER
(%)
|
DOR
(%)
|
FRR
(%)
|
IVI
|
|
Synsepalum subcordatum De Wild.
|
58
|
4
|
0,220
|
0,547
|
0,620
|
0,653
|
1,820
|
|
Tridesmostemon claessensii De Wild.
|
385
|
4
|
1,867
|
3,633
|
5,256
|
0,653
|
9,542
|
|
Total Sapotaceae
|
739
|
28
|
3,294
|
6,974
|
9,275
|
4,568
|
20,817
|
|
Simaroubaceae
|
Hannoa klaineana Pierre & Engl.
|
130
|
4
|
0,189
|
1,227
|
0,532
|
0,653
|
2,412
|
|
Total Simaroubaceae
|
130
|
4
|
0,189
|
1,227
|
0,532
|
0,653
|
2,412
|
|
Tiliaceae
|
Desplatsia dewevrei (De Wild. & T.Durand) Burret
|
8
|
4
|
0,014
|
0,076
|
0,040
|
0,653
|
0,768
|
|
Glyphaea brevis (Spreng.) Monach.
|
4
|
2
|
0,003
|
0,038
|
0,008
|
0,326
|
0,372
|
|
Grewia pinnatifida Mast.
|
25
|
3
|
0,042
|
0,236
|
0,119
|
0,489
|
0,844
|
|
Grewia sp.
|
5
|
3
|
0,008
|
0,047
|
0,022
|
0,489
|
0,559
|
|
Total Tiliaceae
|
42
|
12
|
0,067
|
0,396
|
0,189
|
1,958
|
2,543
|
|
Ulmaceae
|
Celtis mildbraedii Engl.
|
132
|
3
|
0,571
|
1,246
|
1,608
|
0,489
|
3,343
|
|
Celtis soyauxii Engl.
|
31
|
3
|
0,093
|
0,293
|
0,261
|
0,489
|
1,043
|
|
Celtis tessmannii Rendle
|
118
|
4
|
0,388
|
1,114
|
1,092
|
0,653
|
2,859
|
|
Total Ulmaceae
|
281
|
10
|
1,052
|
2,652
|
2,962
|
1,631
|
7,245
|
|
Verbenaceae
|
Vitex congolensis De Wild. & T. Durand
|
7
|
3
|
0,023
|
0,066
|
0,065
|
0,489
|
0,621
|
|
Vitex mombassae Vatke
|
3
|
2
|
0,003
|
0,028
|
0,009
|
0,326
|
0,363
|
|
Vitex welwitschii Gürke
|
29
|
4
|
0,086
|
0,274
|
0,242
|
0,653
|
1,168
|
|
Total Verbenaceae
|
39
|
9
|
0,112
|
0,368
|
0,316
|
1,468
|
2,152
|
|
Violaceae
|
Rinorea aruwimiensis
|
2
|
1
|
0,001
|
0,019
|
0,004
|
0,163
|
0,186
|
|
Total Violaceae
|
2
|
1
|
0,001
|
0,019
|
0,004
|
0,163
|
0,186
|
|
Total général
|
10596
|
613
|
35,513
|
100,000
|
100,000
|
100,000
|
300,000
|


Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Annexe 2: Nombre des cellules non
différenciées par file radiale et par récolte
|
A1
|
A2
|
A3
|
A4
|
nmc
|
|
Mai_1
|
4,02
|
4,03
|
4,05
|
4,03
|
4,03
|
|
Juin_1
|
3,53
|
3,40
|
3,40
|
3,47
|
3,37
|
|
Juin_2
|
3,83
|
3,57
|
3,60
|
3,60
|
3,65
|
|
Juillet_1
|
3,33
|
3,40
|
3,40
|
3,33
|
3,45
|
|
Juillet_2
|
3,33
|
3,40
|
3,40
|
3,33
|
3,45
|
|
Août_1
|
4,30
|
4,20
|
4,23
|
4,20
|
4,23
|
|
Août_2
|
4,33
|
4,33
|
4,43
|
4,30
|
4,35
|
|
Sept_1
|
4,83
|
4,53
|
4,73
|
4,50
|
4,65
|
|
Sept_2
|
4,97
|
4,77
|
5,10
|
4,80
|
4,91
|
Annexe 3: Largeur moyenne de cernes de croissance en
mm
pour les quatre arbres
Rayon moy Rayon moy Rayon moy Rayon moy Cernes Y1 Y2 Y3
Y4
|
1
|
0,89
|
1,31
|
0,98
|
0,69
|
|
2
|
1,14
|
1,30
|
1,79
|
0,73
|
|
3
|
1,49
|
2,61
|
1,74
|
1,03
|
|
4
|
5,10
|
2,90
|
1,50
|
0,64
|
|
5
|
5,27
|
5,32
|
2,50
|
1,54
|
|
6
|
3,36
|
8,18
|
2,37
|
3,38
|
|
7
|
5,61
|
8,56
|
3,18
|
1,96
|
|
8
|
4,76
|
8,88
|
3,88
|
2,43
|
|
9
|
4,09
|
10,64
|
5,90
|
5,60
|
|
10
|
2,47
|
8,55
|
5,82
|
5,24
|
|
11
|
5,01
|
7,34
|
3,77
|
2,45
|
|
12
|
4,35
|
7,53
|
4,73
|
4,86
|
|
13
|
3,58
|
5,19
|
2,12
|
4,59
|
|
14
|
1,81
|
5,77
|
2,74
|
3,62
|
|
15
|
0,86
|
5,44
|
4,55
|
2,89
|
|
16
|
2,53
|
6,77
|
2,44
|
4,79
|
|
17
|
2,72
|
5,22
|
3,28
|
4,36
|
|
18
|
3,08
|
2,78
|
3,68
|
3,50
|
|
19
|
3,67
|
4,72
|
4,64
|
6,87
|
|
20
|
4,53
|
4,40
|
5,57
|
6,12
|
|
21
|
4,04
|
2,73
|
5,31
|
6,21
|
|
22
|
3,31
|
1,69
|
6,02
|
6,54
|
|
23
|
3,83
|
2,22
|
5,37
|
2,36
|
|
24
|
2,58
|
2,32
|
4,98
|
2,03
|
|
25
|
2,38
|
1,35
|
6,43
|
4,48
|
|
26
|
3,32
|
2,55
|
7,29
|
4,86
|
|
27
|
4,13
|
3,79
|
6,66
|
4,34
|
|
28
|
2,95
|
2,93
|
7,75
|
6,69
|
|
29
|
3,65
|
1,69
|
8,16
|
7,46
|
|
30
|
3,92
|
0,75
|
6,52
|
5,89
|
|
31
|
3,08
|
1,68
|
6,73
|
7,16
|
|
32
|
3,83
|
2,08
|
5,34
|
6,25
|
|
33
|
3,37
|
1,39
|
7,22
|
6,34
|
|
34
|
3,13
|
1,90
|
6,40
|
5,65
|
|
35
|
4,30
|
1,16
|
4,89
|
4,98
|
|
36
|
4,66
|
2,72
|
6,49
|
5,73
|

Activité cambiale et croissance de Pericopsis elata
(Harms) Van Meeuwen: inventaire forestier, analyse diachronique
d'échantillons de la
zone cambiale dans les Réserves de Yoko
et de Yangambi (Province Orientale/RDC)
Rayon moy Rayon moy Rayon moy Rayon moy Cernes Y1 Y2 Y3
Y4
|
37
|
3,78
|
2,78
|
9,55
|
5,07
|
|
38
|
2,45
|
2,74
|
8,37
|
4,13
|
|
39
|
5,98
|
3,14
|
4,80
|
3,94
|
|
40
|
4,74
|
3,32
|
9,11
|
4,37
|
|
41
|
5,20
|
2,60
|
5,18
|
4,28
|
|
42
|
5,81
|
2,71
|
6,45
|
3,75
|
|
43
|
4,90
|
2,44
|
8,98
|
4,18
|
|
44
|
4,38
|
2,52
|
6,74
|
4,11
|
|
45
|
4,67
|
3,46
|
4,87
|
3,78
|
|
46
|
5,15
|
2,58
|
9,59
|
3,57
|
|
47
|
4,18
|
2,18
|
8,90
|
3,22
|
|
48
|
3,50
|
1,35
|
10,52
|
4,06
|
|
49
|
3,73
|
2,30
|
9,56
|
2,12
|
|
50
|
2,72
|
1,51
|
5,11
|
3,13
|
|
51
|
2,38
|
1,78
|
8,34
|
3,48
|
|
52
|
4,24
|
2,10
|
5,70
|
2,53
|
|
53
|
4,02
|
1,31
|
6,27
|
2,58
|
|
54
|
0,39
|
1,83
|
8,71
|
2,72
|
|
55
|
39,01
|
2,44
|
4,29
|
2,57
|
|
56
|
|
38,28
|
6,41
|
2,17
|
|
57
|
|
|
0,90
|
3,19
|
|
58
|
|
|
63,42
|
1,78
|
|
59
|
|
|
|
2,16
|
|
60
|
|
|
|
0,52
|

|
|


