3. Méthode et
Analyses effectuées
? Méthode
Nous avons utilisé la méthode NPPEN et la
méthode d'homogénéisation, lesquelles sont décrites
dans les sections précédentes.
? Analyse 1
La première analyse consistait à comparer les
résultats du modèle NPPEN utilisé avec trois descripteurs
environnementaux (SST, bathymétrie, SSS,) à ceux obtenus avec le
modèle utilisé avec trois descripteurs environnementaux et un
descripteur biotique (SST, bathymétrie, SSS, présence de
proie C. finmarchicus) (Fig. V.2). L'objet de cette analyse
était de mesurer les différences liées à
l'utilisation du paramètre biotique C. finmarchicus.
? Analyse 2
Dans cette deuxième analyse, le NPPEN a
été utilisé avec les paramètres SST,
bathymétrie, SSS, et avec ou sans le descripteur biotique
C. finmarchicus (Figs. V.3 à V.5). La probabilité
de présence moyenne de la morue de l'Atlantique a d'abord
été modélisée pour les périodes 1960-1969,
2000-2006 et 2090-2099 scénario B2 (Fig. V.3). Puis, l'évolution
annuelle moyenne des probabilités de présence de la morue a
été calculée pour chacune des trois zones de
l'étude (zone « Terre-Neuve », zone CIEM Va et zone
CIME IV), de 1960 à 2006 (Figs. V.4a et V.5a) et pour
la période de 2006 à 2100 (Figs. V.4b, c et d et
V.5b, c et d) en fonction des différents scénarios
d'évolution du climat.
? Analyse 3
La probabilité moyenne de présence de la morue
de l'Atlantique a été calculée pour les trois zones
prospectées (zone « Terre-Neuve », zone CIEM Va et
zone CIEM IV), pour chaque décennie entre 1960 et 2100 et pour chaque
scénario d'évolution du climat. Les probabilités moyennes
de présence calculées ont été distribuées en
dix classes (de 0 à 1). Les distributions en classe ont
été reportées sur les figures V.4 et V.5. Seule
l'évolution future des probabilités basées sur le
scénario B2 a été illustrée et la distribution en
classe des probabilités de présence pour la décennie
1960-1969 a été conservée dans chaque graphique à
des fins de comparaisons (histogrammes bleus).
4. Principaux
résultats obtenus
? Analyse 1
Les résultats obtenus à l'aide du modèle
NPPEN basé sur quatre paramètres (SST, bathymétrie SSS et
présence de C. finmarchicus) différent des
résultats obtenus sans la présence du paramètre biotique
« copépode » (Figs. V.2 et V.3). Par rapport au
modèle utilisé sans paramètre biotique, on note une
estimation inférieure de la probabilité de présence de la
morue de l'Atlantique en mer de Norvège, mer de Barents, mer de
Groenland, dans les eaux islandaises et aux alentours immédiats de
Terre-Neuve (Fig. V.2). Par contre, au sud de la mer Nord et en mer du
Labrador, les valeurs des probabilités de présence de la morue,
calculées par le modèle utilisant la présence de proie,
sont supérieures à celles qui sont produites sans le
paramètre biotique (Fig. V.2).
? Analyse 2
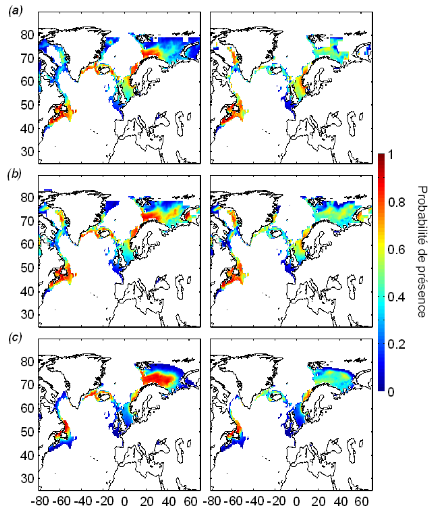
La cartographie de l'évolution des probabilités
de présence de la morue en Atlantique Nord montre un mouvement
biogéographique de cette espèce dirigé vers le Nord, entre
les périodes 1960-1969 et 2000-2006 (Fig. V.3a, b). Le
mouvement biogéographique observé est plus prononcé
côté Est de l'océan Atlantique (la mer du Nord et la mer de
Barents). À l'avenir, ce déplacement latitudinal risque de se
poursuivre, et son intensité dépend du type de scénario
d'évolution du climat choisi. L'intégration du facteur biotique
C. finmarchicus, dans la modélisation de la
présence de la morue de l'Atlantique, fournit des probabilités
généralement inférieures à celles
modélisées sans ce paramètre (Fig. V.3). L'amplitude des
changements biogéographiques est tempérée par
l'utilisation du descripteur biotique.
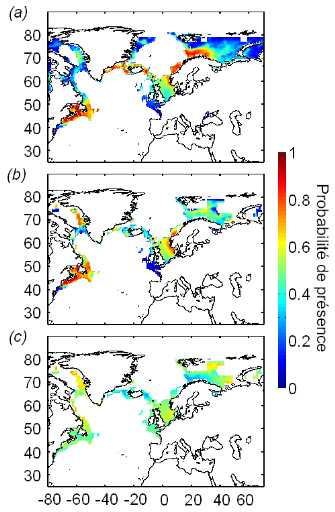
Figure V.2 : Distribution potentielle
(probabilité de présence) de la morue de l'Atlantique pour la
décennie 1960-1969 modélisée avec le NPPEN à partir
de (a) la SST, la bathymétrie et la SSS,
(b) la SST, la bathymétrie, la SSS et la
présence modélisée de C. finmarchicus, et
(c) différence des probabilités de
présence (a) - (b).
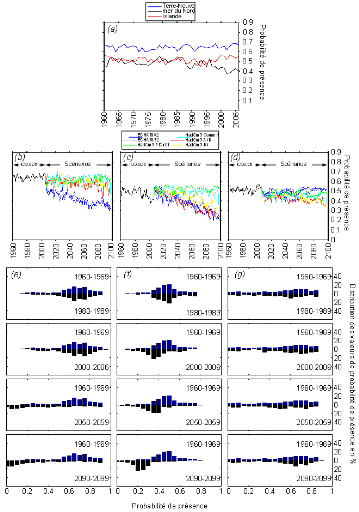
De 1960 à 2006, l'évolution annuelle des
probabilités de présence de la morue en Atlantique nord,
modélisée par le NPPEN (Figs. V.4a et V.5a),
montre trois types de situations différents. Dans la zone
« Terre-Neuve », l'évolution à long terme des
probabilités de présence est relativement stable, bien qu'avec
une variabilité interannuelle prononcée ; dans les deux cas
(présence ou non du facteur biotique dans l'analyse) les valeurs de ces
probabilités oscillent entre 0,6 et 0,7. En mer du Nord, les valeurs de
probabilités sont comprises entre 0,4 et 0,6 et sont globalement plus
faibles quand elles sont calculées sans le facteur biotique ; dans
les deux cas (présence ou non du facteur biotique dans l'analyse), une
nette tendance à la diminution dans cette zone est observée
à partir des années 1980. Dans les eaux islandaises, entre 1960
et 2006, si le facteur biotique n'est pas utilisé, les
probabilités de présence se situent entre 0,45 et 0,6, elles
semblent avoir déclinées très légèrement
jusque dans les années 1990, années à partir desquelles
elles ont entamé une hausse ; si le facteur biotique est pris en
compte, les probabilités de présence calculées sont
inférieures à 0,35 et aucune tendance à l'augmentation ou
la diminution n'est observée de façon nette.
De 2006 à 2100, l'évolution annuelle des
probabilités potentielles de présence de la morue de l'Atlantique
(Figs. V.4 et V.5), montre qu'en zone
« Terre-Neuve » (Figs. V.4b et V.5b),
seuls les scénarios SRES A2 et B2 (lignes bleues) provoquent une chute
importante et rapide des probabilités de présence de la morue de
l'Atlantique. Avec les scénarios A1B et B1, les probabilités de
présence se maintiennent à un niveau stable jusqu'en 2070 ;
avec les autres scénarios, COMMIT et PICNTRL, les probabilités de
présence n'évoluent pas. L'utilisation ou non du paramètre
biotique n'a pas d'influence sensible sur l'évolution des
probabilités en zone « Terre-Neuve ». En Mer du Nord
(Figs. V.4c et V.5c), seuls les scénarios COMMIT et
PICNTRL (lignes verte et turquoise) prévoient le maintient d'une
présence équivalente à celle actuelle. Tous les autres
scénarios prédisent une chute progressive et irréversible
de la présence de la morue; l'utilisation ou non du paramètre
biotique n'a pas d'influence sensible sur l'évolution des
probabilités. Enfin, en Islande (Figs. V.4d et
V.5d), l'analyse pratiquée, sans le facteur biotique, montre
une stabilisation des probabilités à un niveau plus faible
qu'avant 2006 quelque soit le scénario utilisé. Une hausse
très faible des valeurs des probabilités de présence
annuelles est attendue, dans le cas où le facteur
« présence de proie » est considéré
(Fig. V.5d).
Figure V.3 : Distribution potentielle
(probabilités de présence) de la morue de l'Atlantique pour les
périodes temporelles 1960-1969 (a), 2000-2006
(b) et 2090-2099 scénario B2(c),
modélisée avec le NPPEN. Les paramètres pris en compte
sont : la SST, la bathymétrie, la SSS (colonne de gauche et droite)
et la présence modélisée par le NPPEN de C.
finmarchicus (colonne de droite).
? Analyse 3
Les trois régions marines constituent des habitats aux
potentiels d'accueil de la morue différents et fluctuants dans le temps
(Figs. V.4e, f et g et V.5e, f et g).
L'étude de l'évolution des probabilités
de présence, entre les périodes 1960-1969 et 1980-1989, montre
peu de changements et ce quelque soit la zone considérée. Entre
les périodes 1960-1969 et 2000-2006, des changements sont observables en
mer du Nord uniquement, représentés par un affaiblissement dans
leur ensemble des probabilités de présence de la morue (Figs.
V.4f et V.5f). Après 2050, seule la zone CIEM Va
islandaise conserve une distribution de probabilités similaire à
celle des années 1960-1969. En mer du Nord, le glissement de la
majorité des probabilités vers des valeurs plus faibles
s'accentue. Du côté de Terre-Neuve, le même
phénomène est prédit à partir de la période
2050-2059.
La comparaison de la décennie 1960-1969 entre les
figures V.4 et V.5 montre que, de façon général,
l'utilisation du paramètre biotique ne modifie pas la distribution des
probabilités de présence en zone
« Terre-Neuve ». En mer du Nord, l'utilisation du
paramètre biotique estime les probabilités supérieures
(entre 0,6 et 0,8) à celles produites sans ce paramètre (entre
0,4 et 0,6). Dans les eaux islandaises, l'utilisation du paramètre
biotique ne permet pas aux probabilités de présence de
dépasser la valeur 0,7. Sans le paramètre biotique, le
modèle évalue que les probabilités de présence de
la morue peuvent atteindre la valeur de 0,9.
L'évolution des probabilités de présence
de la morue de l'Atlantique, décennie par décennie,
diffère sensiblement quand est pris en compte le facteur biotique proie
dans le modèle. Dans la zone « Terre-Neuve »,
l'utilisation du facteur biotique concentre un peu plus les probabilités
de présence vers des valeurs très faibles proche de 0,1. En mer
du Nord, le glissement vers des valeurs plus faibles de la distribution des
probabilités s'opère entre les périodes 1960-1969 et
2000-2006, mais il est moins intense quand est inclut le facteur biotique proie
dans la modélisation. Autour de l'Islande, l'évolution des
probabilités de présence ne semble pas être affectée
par l'utilisation ou non du paramètre biotique proie.
Figure V.4 : Évolutions annuelles
de la probabilité de présence de la morue calculée, sans
C. finmarchicus, à Terre-Neuve (en bleu), en mer du Nord (en
noire) et en Islande (en rouge) modélisées, par le NPPEN de 1960
à 2006 (a) et en fonction des scénarios
d'évolution du GIEC Terre-Neuve (b) en mer du Nord
(c) et en Islande (d). Comparaison des
distributions des probabilités de présence de la morue, entre les
périodes 1960-1969/1980-1989, 1960-1969/2000-2006, 1960-1969/2050-2059
et 1960-1969/2090-2099, à Terre-Neuve (e), en mer du
Nord (f) et en Islande (g).
| 

