I.1.4 / Caractère du cycle oestrien
Chez tous les mammifères, à l'exception des
primates, la femelle n'accepte l'accouplement que pendant la période des
chaleurs ou oestrus. C'est à la fin de cette période que se fait
l'ovulation.
Chez les bovins, le cycle sexuel dont la durée varie, peut
être divisé en quatre phases correspondant à
différentes étapes de l'activité ovarienne :
v Proestrus (avant l'oestrus) : c'est la phase
de croissance et de maturation folliculaire ;
v Oestrus ou chaleurs : cette phase est
caractérisée par la ponte ovulaire. Pendant cette phase, la
femelle recherche, attire, accepte le mâle (chevauche et se laisse
chevaucher par ses congénères) ;
v Metoestrus ou post oestrus : c'est la phase
anabolique du corps jaune. Le devenir de celui-ci est conditionné
à celui de l'ovule. Si ce dernier est fécondé, le corps
jaune reste actif empêchant la maturation de nouveaux follicules. Mais si
la fécondation n'a pas eu lieu, le corps jaune
dégénère : c'est le phase lutéale ;
v Diostrus ou anoestrus : elle correspond
à la période de lutéolyse, ou période de repos
sexuel.
I.2 / Les composantes du cycle sexuel chez la
femelle
On distingue trois composantes du cycle sexuel chez la vache : v
Les événements cellulaires ovariens ;
v Les événements hormonaux ;
v Le comportement sexuel.
I.2.1 / Les événement cellulaires
ovariens
Cette composante est celle du clinicien car les modifications
qui la caractérisent se situent essentiellement sur la
périphérie de l'ovaire : par exploration transrectale, il a donc
la possibilité de les apprécier. Les événements
cellulaires ovariens se produisent de façon régulière,
cyclique et permettent de distinguer deux phases pendant la durée du
cycle sexuel :
v une phase folliculaire qui se définit
comme une phase de croissance brutale,
explosive et terminale d'un ou plusieurs follicules à
antrum aboutissant à l'ovulation.
Le diamètre de ces follicules passe de 2 mm à 20 mm
en quelques 2 à 4 jours.
Le début de cette phase ne peut que se définir
conventionnellement ; la fin se situe lors
de l'expulsion de l'ovocyte. Cette phase folliculaire est le
période terminale de la
folliculogénèse amorcée dès la
moitié de la vie foetale.
Ce processus folliculogénétique échappe
donc en quasi totalité à l'activité cyclique ; il ne lui
est relié qu'épisodiquement chez la vache adulte non gestante,
tous les 21 jours pendant 3 à 4 jours.
V' Une phase lutéale ;
immédiatement après l'ovulation. A partir du caillot sanguin
consécutif à l'ovulation le remaniement histologique et
biochimique aboutit à l'existence de cellules lutéales et
à la formation du corps jaune. Cette transformation initiale demande 24
à 48 heures. D'abord de couleur rouge rosée puis jaune
rosée. Le corps jaune devient, parallèlement à son
développement, franchement jaune. La phase lutéale, dont la
durée est de 16 à 18 jours peut se résumer en trois
périodes :
> Première période : croissance d'une
durée de 5 à 8 jours passant de la taille de
quelques mm à 20 mm ;
> Deuxième période : maintien du
développement pendant 8 à 10 jours ;
> Troisième période : régression
(lutéolyse) brutale en 12 / 24 heures
essentiellement d'origine vasculaire, à la suite de la
quelle sa taille diminue
progressivement jusqu'à disparaître.
La dégradation du corps jaune provoquant la chute
rapide de la sécrétion de la progestérone est due à
un facteur lutéolytique appelé prostaglandine F2 alpha :
(PGF2a).On peut estimer que cette prostaglandine secrétée par
l'utérus, est acheminée à l'ovaire par la veine
utéroovarienne grâce à un mécanisme à contre
courant vasculaire.
La sécrétion de la prostaglandine F2 alpha
(PGF2a) se fait sous forme de bombardements intensifs conduisant à une
lutéolyse irréversible et à la chute définitive du
taux de progestérone. Cette composante laisse apparaître une
inégalité importante entre la phase folliculaire (2-4 jours) et
la phase lutéale (16- 18 jours).
I.2.1.1 / La folliculogénèse (Croissance
folliculaire)
La folliculogénèse est l'ensemble des
phénomènes qui participent à la croissance et la
maturation des follicules. La croissance résulte de trois
phénomènes qui ont successivement un rôle essentiel :
V' augmentation de la taille de l'ovocyte ;
V' multiplication des cellules de la granulosa ;
V' augmentation de la taille de l'antrum.
Quand un follicule s'échappe de la réserve des
follicules primordiaux et commence sa croissance, celle-ci continuera
jusqu'à ce que le follicule subisse l'atrésie ou ovule.
D'après (Thibault et Levasseur, 1979) Il n'y a de
réserve de follicules en croissance ou de follicules à antrum.
I.2.1.1.1 / Initiation de la croissance
folliculaire
Le nombre de follicules quittant la réserve chaque jour
est proportionnel à sa taille, il diminue donc en fonction de l'age
(Pedersen, 1972) cite par Thibault et Levasseur, 1979 . Chez la plupart des
mammifères, les follicules primordiaux évoluent très
lentement vers le stade intermédiaire et primaire.
La croissance et l'atrésie des plus petits follicules
sont peu dépendantes des gonadotropines et de leurs variations
cycliques, mais les hormones modulent probablement les capacités de
synthèse et de maturation des cellules de la granulosa.
I.2.1.1.2 / Développement folliculaire
terminal
Les follicules en fin de croissance sont dépendants des
gonadotropines, la taille folliculaire où apparaît la
dépendance est de 2 mm chez la vache. La folliculogénèse
terminale débute dès ce stade et s'achève avec
l'ovulation. L'intervalle de temps nécessaire à son
déroulement est de 4 à 5 jours chez la vache.
Les événements de la
folliculogénèse terminale
Au cours de la folliculogénèse terminale, la
croissance des follicules susceptibles d'ovuler se déroule de
façon synchrone et coordonnée.
La folliculogénèse terminale est activée
dès que le corps jaune régresse. Tous les follicules
gonado-dépendants présents sur les ovaires entrent alors en
croissance terminale (recrutement). Ils forment alors une
cohorte qui comprend plusieurs follicules de taille et de sensibilité
différente aux gonadotropines. A mi-phase folliculaire, une
sélection se produit et la taille de la cohorte est
réduite au nombre d'ovulations caractéristiques de la race ou
l'espèce. Le (ou les) follicule destiné à ovuler,
reconnaissable par la taille, est appelé « follicule
dominant » (Driancourt et al., 1991).
Pendant la période de dominance sont observés :
V' La croissance et la maturation terminale du (ou des) follicule
pré ovulatoire ; V' La régression par atrésie des autres
follicules de la cohorte ;
V' Le blocage du recrutement de nouveaux follicules.
Le recrutement coïncide avec l'apparition d'une
activité aromatase dans la granulosa, la sélection coïncide
avec l'apparition des récepteurs de LH sur la granulosa, une forte
réduction des quantités de certaines protéines de liaison
dans le liquide folliculaire et une forte production d'inhibine (Driancourt et
al., 1991).
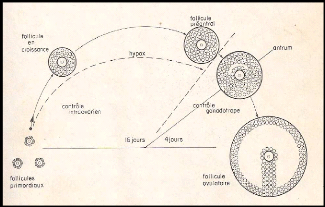
Figure 1 : Régulation de la croissance
folliculaire.
De la réserve de follicules primordiaux
s'échappent régulièrement des follicules qui commencent
leur croissance. Celles-ci se déroule même en absence de
gonadotropes (hypophysectomie= hypox) les gonadotropines (FSH puis FSH et LH)
deviennent indispensable peu avant la formation de l 'antrum
I.2.1.1.3 / Régulation des mécanismes de la
folliculogénèse terminale
Deux niveaux de régulations sont
généralement évoqués : les régulations
endocrines (FSH et LH) et les régulations locales qui affinent les
messages endocrines. Au sein de chaque follicule, des régulations de
nature paracrine (entre thèque et granulosa, entre granulosa et ovocyte)
et autocrine renforcent ou inhibent les effets des gonadotropines sur le
développement folliculaire.
Contrôle du recrutement :
Chez les primates comme chez les mammifères
domestiques, FSH constitue le signal endocrine impliqué dans le
recrutement : (a) dans presque toute les espèces étudiées,
il existe un synchronisme presque parfait entre niveaux de FSH
élevés et recrutement, (b) l'administration de liquide
folliculaire riche en inhibine, au moment du recrutement déprime les
niveaux de FSH et bloque le recrutement, (c) chez des femmes hypogonadotropes,
des
singes ou brebis rendus hypogonadotropes par administration
d'un antagoniste de la GnRH, l'administration de FSH (pure ou recombinante) est
capable d'initier une croissance folliculaire terminal.
Chaque individu présente un niveau seuil de FSH en
dessous duquel le recrutement n'est pas induit. Des variations limités
de FSH (30-50 %) autour de ce niveau seuil déclenchent ou nom le
recrutement. Il est également probable que pour un individu
donné, tous les follicules ne présentent pas les mêmes
seuils.
Les conséquences de cette
hétérogénéité fonctionnelle sur le devenir
de chaque follicule (dominant ou atrétique) sont évidentes ; le
potentiel de survie et le développement d'un follicule ayant des besoins
limités en FSH est très supérieur à celui d'un
follicule beaucoup moins sensible.
Contrôle de la sélection
Deux hypothèses sont généralement
avancés pour expliquer la sélection : l'une purement gonadotrope
et l'autre prenant en compte un facteur local. L'hypothèse gonadotrope
fait intervenir la sensibilité individuelle des follicules de la cohorte
à la FSH et le rétrocontrôle négatif sur le niveau
de FSH, exercé par l'oestradiol et l'inhibine, dont la
sécrétion augmente avec la croissance folliculaire. La chute de
niveau de FSH bloque la croissance / maturation des follicules de la cohorte
qui avait les besoins les plus élevés en FSH (presque toujours
les plus petits).
L'autre hypothèse allie l'effet de la chute de FSH
à l'action d'un facteur local (intra-ovarien) produit par le plus gros
follicule de la cohorte et inhibant la prolifération ou
différenciation cellulaire (aromatase, apparition des récepteurs
de LH sur la granulosa) dans les autres follicules de la cohorte.
Contrôle de la dominance
Bien que les concentrations de FSH circulant soient
réduites, le (ou les) follicules dominant poursuit sa croissance et sa
maturation car ses besoins en FSH sont réduits. Les niveaux de FSH
nécessaire à la survie du follicule dominant sont d'environ 50 %
inférieur à ceux qui sont nécessaires au recrutement.
Trois propriétés du follicule dominant permettent
d'expliquer son aptitude à survivre dans un environnement appauvri en
FSH :
V' L'acquisition de récepteurs de LH sur la granulosa ;
V' L'amplification de la réponse folliculaire à FSH
et LH par des régulateurs paracrines ou autocrines ;
V' Une vascularisation sélectivement amplifiée
pourrait assurer une diffusion facile de FSH et de LH.
I.2.1.2 / Ovulation
L'ovulation est la libération d'un ou plusieurs
gamètes femelles, au stade ovocyte II,aptes à être
fécondés, après rupture d'un ou plusieurs follicules
préovulatoires.
I.2.1.2.1 / Mécanisme de l'ovulation
Arrivé au terme de sa croissance, en réponse
à une forte élévation des gonadotrophines, la
décharge ovulante, le follicule éclate, non pas par suite d'une
augmentation de la pression interne mais en raison de la fragilisation de la
paroi du follicule, et libère l'ovocyte II.
Les changements morphologiques et cytologiques au cours de
l'ovulation se déroulent comme suit :
V' les cellules de la granulosa se dissocient ;
V' le cumulus est libéré dans le liquide
folliculaire ;
V' la couronne radiée reste en place et l'ensemble ovocyte
couronne radiée s'entoure de glycoprotéines ;
V' les fibres collagènes des thèques sont
dissociées sous l'action d'un système enzymatique complexe puis
digérées par un enzyme, la cathepsine ;
V' l'épithélium ovarien se desquame ; une zone sans
vascularisation, le stigma, apparaît, au niveau de laquelle va se
produire la déchirure du follicule.
Aussitôt avant l'ovulation, il y a reprise de la
méiose et l'obtention d'un ovocyte II.
I.2.1.3 / Le corps jaune
C'est une structure qui apparaît immédiatement
après l'ovulation par suite d'une transformation morphologique et
fonctionnelle du follicule après libération de l'ovocyte.
On distingue trois phases dans l'évolution du corps jaune
:
V' la phase de croissance ou
lutéogénèse : après l'ovulation, la
cavité folliculaire se remplit d'un caillot de sang. les cellules de la
granulosa encerclent le caillot, s'hypertrophient ; leur noyau devient
polyploïde tandis que le tissu formé se vascularise abondamment
;
V' la phase de maintien ou lutéotrophie :
c'est la période pendant laquelle le corps jaune maintient son
développement et son activité endocrine ;
V' la phase de régression ou
lutéolyse : le corps jaune régresse rapidement mais
reste cependant présent pendant plusieurs semaines sous la forme d'un
organite de petite
taille. Parallèlement, le taux de progestérone
diminue brutalement.
S'il y a gestation, la lutéolyse n'a pas lieu ; le corps
jaune évolue en corps jaune de gestation. La cyclicité est
arrêtée par un signal provenant de l'utérus et indiquant la
présence d'un embryon ; cette information est donnée entre le
15ème et 17ème jour du cycle chez la
vache.
Si l'ovocyte libéré après ovulation n'a pas
été fécondé, le corps jaune régresse et
permet l'apparition d'un nouveau cycle ovulatoire.
I.2.2 / Les événements
hormonaux
On peut distinguer trois niveaux de contrôle hormonal de
l'activité cyclique: niveau ovarien, hypophysaire et hypothalamique.
I.2.2.1 / Hormones ovariennes
L'ovaire exerce une fonction hormonale par
l'intermédiaire des glandes endocrines qu'il héberge à son
sein. Les hormones sont de nature stéroïdiennes et dérives
du cholestérol. Leur structure est très bien connu et leur
synthèse très facile.
Au plan hormonal, le cycle sexuel de la vache comprend deux
phases successives: la phase oestrogénique courte (3 jours environ) et
la phase progestéronique (beaucoup plus longue: 17- 18 jours). La
première est concomitante de la croissance folliculaire terminale
précédent l'ovulation. Elle se caractérise par des niveaux
croissants d'oestrogènes essentiellement d'oestradiol 17 â dont le
maximum est de l'ordre de 10-20 pg / ml chez la vache dans le plasma
périphérique.
La phase lutéale se caractérise par une
élévation progressive de la progestérone dont la
concentration moyenne atteint un plateau vers le 8 ème jour, qui
persiste jusque vers le 16- 17ème jour après l'oestrus, puis
s'abaisse brutalement et ses concentrations devient très faibles au
cours des jours entourant l'ovulation.
| 

