Dynamique non lineaire et effet de l’instabilite de turing sur les systèmes cellulaires désordonnéspar Boris TAMKO MBOPDA Université de Yaoundé I - Master II of Physique 2019 |

CHAPITRE DEUX
2.3 Rôle des acides dans le cancer 2.3.1 Cause directe du cancer Le lauréat du prix Nobel, le Dr Otto Warburg H, a consacré sa vie à l'étude et à la cause du cancer. Dr Warburg était directeur de l'institut Kaiser Wilhelm (aujourd'hui Institut Max Planck) pour la physiologie cellulaire à Berlin. Otto Heinrich Warburg, né le 8 Octobre 1883 à Fribourg-en-Brisgau (Bade-Wurtemberg), mort à Berlin, était un médecin, physiologiste et biochimiste allemand. Docteur en chimie et docteur en médecine, il fut lauréat du prix Nobel physiologie et médecine pour sa découverte de processus-clés de la respiration cellulaire et de différents systèmes enzymatiques. Ayant démontré que les cellules cancéreuses changent leur métabolisme pour passer à un métabolisme anaérobie il a formulé l'hypothèse, non confirmée, que le cancer ne peut pas se développer dans un milieu riche en oxygène. Parmi ses étudiants, le plus illustre est Hans Adolf Krebs, prix Nobel lui aussi en 1953. Chapitre II : Analyse mathématique et méthodologie 23 Mémoire de MASTER II TAMKO MBOPDA Boris c2019 Le Dr Warburg a été l'un des éminents physiologistes du 20ème siècle, et il a découvert que la cause du cancer est une acidité trop importante dans le corps, signifiant que le pH dans le corps est en dessous du niveau normal de 7,365, qui constitue un état «acide». Warburg a étudié le métabolisme des tumeurs et la respiration des cellules et a découvert que les cellules cancéreuses persistent et prospèrent dans un pH plus faible, à 6.0, à cause de la production d'acide lactique et d'une concentration élevée en CO2. Il croyait fermement qu'il y avait une relation directe entre le pH et l'oxygène. Un pH plus élevé, qui est alcalin, signifie qu'il y a une concentration plus élevée en molécules d'oxygène, tandis qu'un pH plus faible, qui est acide, signifie qu'il y a une concentration plus faible d'oxygène et c'est cette même oxygène qui est nécessaire pour garder des cellules saines. En 1931, il a été lauréat du prix Nobel de physiologie ou médecine pour cette importante découverte sur la cause du cancer. Il a déclaré : «les tissus cancéreux sont acides, alors que les tissus sains sont alcalins. L'eau se divise en ions H+ et OH-,s'il y a un excès d'H+, c'est acide; s'il y a un excès d'OH-, alors c'est alcalin.» Dans son ouvrage le métabolisme des tumeurs, il a prouvé que « toutes les formes de cancer sont caractérisées par deux conditions de base : l'acidose et l'hypoxie. Le manque d'oxygène et l'acidose sont les deux faces d'une même pièce : si vous en avez une, vous avez l'autre.» «Toutes les cellules normales ont un besoin absolu d'oxygène, mais les cellules cancéreuses peuvent vivre sans oxygène, c'est une règle sans exception. Privez une cellule de 35% de son oxygène pendant 48 heures, elle pourra devenir cancéreuse.» Le Dr Warburg a clairement démontré que la première cause du cancer est un manque d'oxygène, ce qui crée un état acide dans le corps humain. Le Dr Warburg a découvert que les cellules cancéreuses sont anaérobies et ne peuvent pas survivre en présence de niveaux élevés d'oxygène, que l'on trouve dans un état alcalin. 2.3.2 L'action du PH sur le développement du cancer D'après les recherches du Dr. Enderlein, une guérison totale de maladies chroniques, ne peut arriver que lorsque le sang est ramené à un pH normal, légèrement alcalin. Etant donné que nous sommes faits à 60-70% d'eau, le niveau du pH a des effets très profonds sur la chimie du corps, la santé et les maladies. Tous les mécanismes régulateurs (respiration, circulation, digestion, production hormonale) ont pour but d'équilibrer le pH, en débarrassant le Chapitre II : Analyse mathématique et méthodologie 24 Mémoire de MASTER II TAMKO MBOPDA Boris c2019 corps et les tissus des résidus acides sans endommager les cellules vivantes. Si le pH dévie, trop acide ou trop alcalin, les cellules s'empoisonnent avec leurs propres déchets toxiques et meurent. Comme les pluies acides sur une forêt et des déchets alcalins dans un lac, un pH déséquilibré peut interrompre les activités et fonctionnements cellulaires, comme les battements de coeur et les étincelles entre les neurones du cerveau. La cause du cancer est liée à deux facteurs qui sont toujours présents lors d'un cancer, le pH acide et le manque d'oxygène. En apprenant à manipuler ces deux facteurs qui seraient la cause du cancer, on peut réduire ou inverser le processus du cancer. Rappelons-nous que le pH est un nombre exponentiel de 10; donc une petite différence dans le pH se traduit en grosse différence dans le nombre d'ions OH-. En d'autres mots, un sang avec un pH de 7.45 contient 64.5% plus d'oxygène qu'un sang donc le pH est 7.3. Le cancer a besoin d'un environnement acide/pauvre en oxygène pour survivre et se développer. Les patients en phase terminale d'un cancer sont environ 1000 fois plus acides qu'ils ne devraient l'être. Cela veut dire un niveau d'oxygène dangereusement critique au niveau cellulaire. En l'absence d'oxygène, le glucose fermente et devient acide lactique. Cela baisse le pH des cellules. Le pH des urines et de la salive des patients en phase terminale de cancer se situe très souvent entre 4.0 et 5.5. Quand le cancer se transforme en métastases, le pH chute davantage. Nos corps ne peuvent pas lutter contre la maladie si le pH n'est pas proprement équilibré. En d'autres termes, c'est «alcalinise ou meurt». 2.4 Modèle utilisé pour le mémoire Le métabolisme énergétique altéré est une caractéristique quasi universelle des tumeurs cancéreuses solides. il est apparu ces dernières années comme une caractéristique phénotypique possible et une avenue pour de nouvelles stratégies de traitement . Le principe du métabolisme altéré dans les tumeurs remonte à l'oeuvre séminale d'Otto Warburg . Il a observé que les carcinomes chez le rat et chez l'homme subissaient un métabolisme du glucose, non pas par phosphorylation oxydative comme dans les cellules normales, mais principalement par glycolyse, voie extra mitochondriale tronquée produisant de l'acide lactique comme sous-produit et généralement réservée aux conditions d'hypoxie. Bien que de nombreuses cellules tumorales soient sujettes à une hypoxie chronique ou transitoire due à un encombrement et à un système vasculaire défectueux, Warburg a noté que les tumeurs maintenaient la voie glycolytique même Chapitre II : Analyse mathématique et méthodologie 25 Mémoire de MASTER II TAMKO MBOPDA Boris c2019 avec suffisamment d'oxygène. Pour la phosphorylation oxydative, indiquant que le passage métabolique de la phosphorylation oxydative à la glycolyse maintenant appelé effet Warburg, ou phénotype glycolytique était un changement fondamental du métabolisme de la tumeur (Warburg 1956). Les décennies qui ont suivi ont confirmé les observations de Warburg. Le phénotype glycolytique est suffisamment répandu, par exemple, pour le succès de l'imagerie par tomographie à émission de positrons au 18 fluoro-2-désoxyglucose (FDG-PET), qui exploite le taux considérablement accru d'absorption du glucose présenté par les tumeurs exprimant le phénotype glycolytique. Les facteurs sous-jacents de l'effet Warburg restent toutefois en grande partie mystérieux, car ils entrainent une baisse significative du rendement absolu en ATP par molécule de glucose par rapport à la voie d'oxydation normale et génèrent une charge toxique en acide lactique. La glycolyse découplée de la respiration peut éventuellement permettre aux cellules tumorales d'optimiser leurs voies cataboliques et anaboliques pour la biosynthèse rapide requise par un programme de croissance intensive. Alternativement, ou peut-être en complément, en acquérant une résistance à l'acidification du microenvironnement, les cellules tumorales exprimant le phénotype glycolytique peuvent acquérir un avantage sélectif par rapport aux cellules saines voisines sensibles à l'acide, qui leur permet d'envahir le microenvironnement. L'hypothèse d'une invasion à médiation acide, est motivée par le fait de considérer une tumeur comme une espèce invasive, soumise à des pressions sélectives de plus en plus sévères et à une évolution somatique au sein du microenvironnement d'une population en bonne santé; par conséquent , il est sujet à une représentation mathématique en tant que système de réaction-diffusion à l'échelle du tissu. Alors que de nombreux modèles mathématiques ont été développé pour explorer les relations entre invasion tumorale, tissu-acide, métabolisme cellulaire et exigences énergétiques, Gatenby et Gawlinski ont été les premiers émettre hypothèse d'invasion à médiation acide dans un cadre réaction-diffusion . En partant du principe que les tumeurs considérées exprimaient déjà l'effet Warburg, les auteurs ont isolé les conséquences du phénotype glycolytique sur l'invasion à l'interface entre la tumeur en croissance et le tissu sain environnant. Le système d'équations aux dérivées partielles non linéaires qui en résulte est décrit en détail dans Gatenby et Gawlinski (1996). Toutefois, par souci de commodité, nous reproduisons les équations ici, U(x,T) indiquant la densité de cellules saines (fonction de la variable spatiale x en unités de centimètres et le temps T, mesuré en secondes) V(x,T) la densité des cellules tumorales et W(x,T) la concentration d'acide Chapitre II : Analyse mathématique et méthodologie 26 Mémoire de MASTER II TAMKO MBOPDA Boris c2019 lactique extracellulaire dépassant les concentrations normales d'acide tissulaire. Les équations de Gatenby-Gawlinski (Gatenby-Gawlinski, 1996 ) sont les suivantes :
L'indice «1» représente ici l'association des paramètres avec les cellules saines, «2» avec les cellules tumorales et «3» avec un excès d'acide lactique extracellulaire. Les constantes K1,2 représentent les capacités porteuses des tissus, ä1 le taux de mort cellulaire saine induite par l'acide, ä3 le taux de clairance de l'acide en excès par tamponnement et évacuation vasculaire, ñ1,2 les taux de prolifération cellulaire, ñ3 le taux de production d'acide en excès par les cellules tumorales, Ä2 le coefficient de diffusion en espace libre des cellules tumorales et Ä3 le coefficient de diffusion chimique de l'excès d'acide. Une caractéristique clé de ce modèle est le terme de diffusion tumorale à densité limitée dans l'équation. (2), l'idée étant qu'un tissu sain fonctionnant à sa pleine capacité portante agira patiemment contre nous- mêmes, sans qu'il soit réduit, dans Gatenby et Gawlinski (1996), solution numérique des équations (1) à (3) du système 2.2 ont capturé deux types de comportement : l'invasion par une tumeur hétérogène composée de cellules saines et malignes, et l'invasion par une tumeur homogène tuant toutes les cellules saines derrière son bord d'attaque. En outre les tumeurs suffisamment agressives, les auteurs ont prédit l'ouverture d'un écart interstitiel entre les cellules malignes en progression et les cellules saines en régression, comportement qui n'avait pas été signalé auparavant, avant de détecter des lacunes dans les échantillons de tête et de cou fixés et congelés instantanément de carcinomes à cellules squameuses humaines. cette prédiction et la détection subséquente de l'écart interstitiel ont été reconnues comme une illustration élégante des avantages d'une approche hybride mathématique et expérimentale de l'étude du cancer [20]. cet écart pourrait peut-être avoir un potentiel diagnostique potentiel tumeurs agréssives. Le modèle de Gatenby-Gawlinski est l'un des nombreus études suggérant que l'acidité pourrait jouer un rôle important dans la progression tumorale [21]. si cela est vrai, cela pourrait conduire à de nouvelles stratégies thérapeutiques. L'un de ces traitement en cours de développement, la thérapie tampon, était motivé par les conclusions de la modélisation de Gatenby et Gawlinski [22] et visait à priver une tumeur en développement de son avantage sélectif en neutralisant le Chapitre II : Analyse mathématique et méthodologie 27 pH du microenvironnement F23]. 2.5 Description de l'instabilité de Turing Est-il possible de trouver une règle générale pouvant expliquer la diversité des motifs observés dans la nature? pourquoi les Zèbres ont-ils des rayures et les lépards des taches? pour un mathématicien, il est très intéressant de pouvoir regrouper, sous un seul cocept, plusieurs problèmes qui nous semblent, à première vue, totalement sans lien. En 1952, Alan Turing publia son célèbre article «The chemical Basis of Morphogenesis» dans lequel il avance une idée orignale : la morphogenèse serait causée par la perte de stabilité, due à la diffusion, d'une position d'équilibre de réactants chimiques. En d'autres termes, des équations de réaction-diffusion pouraient modéliser un pelage tacheté, rayé ou bien plus complexe encore. Bien que les biologistes n'aient actuellement pas la tecnologie nécessaire pour confirmer l'hypothèse de Alan Turing, certains motifs obtenus grâce aux modélisations sont si proches de la réalité que cette hypothèse semble être une explications les plus prometteuses.
FIGURE 2.1 - differents motifs que nous pouvons retrouver dans la nature [24] 2.5.1 Problématique Bien que nous puissions étendre la notion de formation de motifs sur un espace en 2D, comme les taches sur la peau d'un léopard, ou en 3D, comme la forme de la tête d'une echinacea, seule l'équation de réation-diffusion en 1D sera présentée afin de rendre les explications plus intuitives. Mais dans quel contexte biologique une simple «ligne» espace en 1D. Peut-elle engendrer des motifs intéressants? la réponse réside dans le développement de la coquille d'un mollusque! Mémoire de MASTER II TAMKO MBOPDA Boris c2019 Chapitre II : Analyse mathématique et méthodologie 28 Mémoire de MASTER II TAMKO MBOPDA Boris c2019 Aussi surprenant que cela puisse paraître, Hans Meinhardt, chercheur à l'Institut Max Planck pour le développement en biologie, a entièrement consacré à ces petits invertébrés son livre The Algorithmic Beauty of Sea Shells [25] dans lequel il s'intéresse à la diversité fulgurante des motifs engendrés par ceux-ci. Lorsque le mollusque agrandit sa coquille, une rangée de cellules ajoute une fine ligne de matière au rebord du coquillage et, au même moment, dépose des pigments à certains endroits sur cette nouvelle ligne de matière. L'hypothèse est que le taux de concentration d'une substance chimique quelconque, présente dans cette rangée de cellules, déterminerait la quantité ou l'intensité des pigments déposés sur la bordure de la coquille. En d'autres termes, si la concentration de la subtance chimique n'est pas uniforme tout le long de la ligne de la diffusion de celle-ci à travers les parois cellulaires. Donc, en étudiant les variations des concentrations des subtanceces responsables de la pigmentation des coquillages dans l'espace et dans le temps, il nous serait possible de reproduire les motifs observés sur les coquillages. c'est ce que Hans Meinhardt a entrepris de décrire grâce aux équations de réaction-diffusion. Intuitivement, nous savons maintenant que la concentration d'une subtance chimique u(x,t) dépend de deux choses : de la positon x, et du temps t. (Si nous avion été dans une espace en 2D ou 3D, x aurait été un vecteur, mais en 1D x est un scalaire). Il est donc naturel que l'équation de réaction-diffusion associée contienne des dérivées partielles représentant la variation des concentrations dans l'espace (x) et dans le temps (t). Les équations de réaction-diffusion sont de la forme:
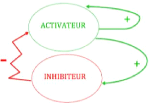
où Du est la constante de diffusion. Certains d'entre vous auront peut-être reconnu la forme de l'équation de la diffusion de la chaleur. Or, avant de nous attarder sur les aspects mathématiques des termes de réaction et de diffusion, nous resterons dans les explications intuitives de la théorie d'Alan Turing. Cela nous permettra ainsi de renforcer notre compréhension de base sur l'importance du rôle de la diffusion dans la formation des motifs. Mais quel est le lien entre les équation de réaction-diffusion et la formation de motifs? Dans son article «The Chemical Basis of Morphogenesis » publié en 1952, Alan Turing proposa une idée brillante. Selon lui, la formation d'un motif hétérogène et stable dans le temps peut se produire si, en l'absence de diffusion, la concentration des substances chimiques est stable, mais qu'à l'ajout du phénomène de la diffusion, il y a perte d'équilibre de cette position de stabilité. C'est ce que nous appelons l'instabilité de Turing. Imaginons un désert initialement complètement Chapitre II : Analyse mathématique et méthodologie 29 Mémoire de MASTER II TAMKO MBOPDA Boris c2019 plat, à un état stable, lorsque le vent se met à souffler, les petits grains de sable «diffuseront» et s'aggloméreront là où il y a de petites irrégularités jusqu'à la formation de dunes. Un motif est alors créé. Cette idée est bien intrigante puisque le concept de diffusion est normalement pour nous un synonyme d'homogénéité. En effet, lorsque vous submergez votre sachet de thé dans l'eau bouillante, vous n'obtiendrez pas des taches de thé partout dans votre tasse, mais bien un mélange homogène après un certain temps. Alors comment est-il possible d'obtenir la formation d'un motif? Regardons ensemble le comportement d'un système typique d'équations de réaction diffusion comprenant un activateur et un inhibiteur. Il sera donc question de deux substances qui interagissent et influencent mutuellement leur production. Mise en situation : L'idée derrière les systèmes d'activation-inhibition est que l'activateur est auto-catalytique, c'est-à-dire qu'il encourage sa propre production, et permet aussi la production d'un inhibiteur. Or, le rôle principal de l'inhibiteur est de ralentir la production de l'activa-teur (voir figure). Parmi toutes mes lectures, c'est probablement l'image donnée par J.D. Murray dans son livre Mathematical Biology [26] qui m'a le plus aidée à comprendre le phénomène. Imaginons un champ où la végétation est très sèche et qui abrite d'innombrables sauterelles. Supposons que ces insectes produisent énormément d'humidité en transpirant lorsqu'elles ont très chaud. Si une personne allume un feu dans le champ, les flammes vont évidemment se propager. Ici, les sauterelles joueront le rôle de l'inhibiteur et les flammes représenteront l'ac-tivateur. S'il n'y avait pas d'humidité, les flammes se propageraient sur toute la surface et il n'en résulterait rien d'autre qu'un champ totalement brûlé. Par contre, supposons que les
FIGURE 2.2 - Interactions entre un activateur et un inhibiteur [24] flammes réchauffent les sauterelles qui, à leur tour, produisent suffisamment d'humidité pour protéger la végétation des brûlure. Un motif peut alors être créé de la façon suivante. Le feu commence à se répandre avec un coefficient de diffusion Df. Lorsque les sauterelles sentent le Chapitre II : Analyse mathématique et méthodologie 30 Mémoire de MASTER II TAMKO MBOPDA Boris c2019 front de flammes approcher, elles s'enfuient avec un coefficient de diffusion Ds. Evidemment, les sauterelles doivent s'enfuir plus rapidement que les flammes ne se propagent (Ds»Df). Les insectes se mettent alors à transpirer et l'humidité qui en résulte empêche le feu de brûler la végétation imbibée d'eau. Les flammes sont alors confinées dans un domaine fermé qui dépend des coefficients de diffusion de l'activateur (le feu) et de l'inhibiteur (les insectes). Supposons maintenant qu'il n'y ait pas seulement un point de départ pour notre feu, mais bien plusieurs points choisis de façon aléatoire. En répétant l'idée précédente, nous pouvons facilement imaginer que les parties brûlées du champ formeront un motif tacheté (non homogène) puisque les sauterelles produiront de l'humidité entre les différents feux. |
| ||||||||||||||||||||