Effets de la fertilisation organique à base de fumier de volailles sur la physiologie et la nutrition du sapin de noël en plantationpar Abbe Jonas ASSAMOI Moncton campus Edmundston - Maitrise es sciences forestières 2006 |

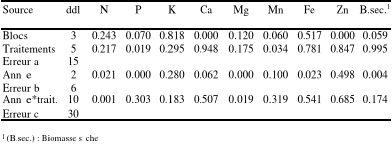
CHAPITRE IIFertilisation organique d'arbres de Noël (Abies balsamea (L.) Mill.) en plantation. Réponses des éléments minéraux et de la coloration foliaire. 2.1 Résumé Un dispositif expérimental a été établi à l'été 2004 dans deux plantations d'arbres de Noël (Abies balsamea (L.) Mill.) du Nouveau-Brunswick afin de réduire les coûts de fertilisation annuelle car le fumier de volailles, en plus de posséder une bonne valeur fertilisante est disponible en grande quantité et à plus faible coût que l'engrais chimique couramment utilisé en plantation d'arbres de Noël. Toutefois, il est nécessaire de démontrer son efficacité avant toute recommandation aux producteurs. Les objectifs spécifiques étaient de déterminer la quantité et la fréquence d'applications du fumier de volailles. Ce dispositif comprend six traitements de fertilisation: le témoin (0 g N arbre-1 an-1); l'engrais chimique de formulation 20-10-10 (20 g N arbre-1 an-1); et quatre niveaux de fertilisation organique avec le fumier de volailles de composition 4-1-2 (20 g N arbre-1 an-1; 40 g Narbre-1 an-1; 40 g N arbre-1 2 ans-1; 80 g N arbre-1 2 ans-1). Des échantillonnages d'aiguilles en vue de la détermination des concentrations d'éléments minéraux dans le feuillage ont été effectués à l'automne des années 2004, 2005 et 2006. L'évaluation des couleurs du feuillage s'est faite à l'automne de ces mêmes années. Les résultats ont indiqué une variation à l'intérieur de limites adéquates des concentrations et contenus foliaires des éléments nutritifs analysés. L'analyse vectorielle des concentrations en azote a indiqué de meilleurs effets des traitements bisannuels au fumier de volailles alors que peu de résultats significatifs ont été obtenus avec le phosphore et le potassium. Les couleurs du feuillage n'ont pas varié significativement entre les traitements au cours des années d'étude. 2.2 Introduction Au Nouveau-Brunswick, comme dans tout l'est canadien, l'industrie des arbres de Noël majoritairement composée du sapin baumier (Abies balsamea (L.) Mill) constitue une activité économique importante. En effet, cette industrie n'a cessé de connaître depuis son installation au Canada vers les années 1945, un essor économique grandissant, générant des revenus importants (environ 5.7 millions de dollars au Nouveau-Brunswick ; Statistique Canada 1996). Malgré cette performance économique, les producteurs d'arbres de Noël sont en proie à des difficultés liées à la mise en place de leurs plantations. De ce fait, des solutions alternatives pourraient leur permettre de réduire les coûts de leurs plantations. Parmi celles-ci, la fertilisation dont l'effet bénéfique pour les arbres est l'augmentation de la quantité d'azote qui est libérée et disponible dans le sol (Meason, Markewitz & Will 2004; Mälkönen 1990; Miller 1981). Par ailleurs, au nord-ouest du Nouveau-Brunswick, les nombreuses entreprises de production de volailles fournissent du fumier, source de matière organique de très bonne valeur fertilisante et à moindre coût que l'engrais chimique couramment utilisé en plantation d'arbres de Noël. Moore, Daniel, Sharpley & Wood (1995) ont montré que presque 90% des déchets de volailles produits étaient utilisés sur des terres agricoles eu égard à leurs faibles coûts comparativement aux fertilisants commerciaux. Actuellement, les producteurs d'arbres de Noël n'optent pas pour cette approche à cause du manque d'informations sur la quantité et le rythme d'application de ce fertilisant. Kingery et al. (1993); Sommerfeldt, Chang & Entz (1988) ont noté une amélioration des propriétés physico-chimiques de sols amendés avec du fumier de volailles comparativement aux sols non amendés; de même Edwards & Daniels (1992) ont noté que dépendamment de la composition individuelle des déchets de volailles, ceux-ci pourraient augmenter la production des cultures via leur capacité à fournir les nutriments et augmenter la qualité du sol. Par conséquent, le remplacement de l'engrais chimique par un engrais organique dans les plantations d'arbres de Noël peut modifier la nutrition des arbres. L'analyse foliaire est la méthode la plus souvent utilisée pour évaluer le statut nutritionnel des arbres de Noël car elle intègre tous les facteurs qui peuvent influer sur la disponibilité et l'absorption des éléments nutritifs. En plus d'indiquer les niveaux critiques des concentrations foliaires en nutriments (Van den Driessche 1974 ; Paarlahti, Reinikainen & Veijalainen 1971), elle indique, le cas échéant, s'il faut modifier le programme de fertilisation. Plusieurs méthodes ont été développées pour évaluer le statut nutritif des plantes incluant les niveaux critiques, les ratios techniques, le système intégré de diagnostic et de recommandation (DRIS) et l'analyse vectorielle. De ces méthodes citées, l'analyse vectorielle est l'outil le plus utilisé dans l'évaluation du statut nutritif des arbres à travers un format graphique intégré (Timmer & Stone 1978). Timmer & Stone (1978) ont utilisé cet outil pour comparer les concentrations foliaires en éléments nutritifs de jeunes plants de sapin baumier fertilisés avec l'azote, le phosphore, le potassium et de la chaux. De même, Imo & Timmer (1997) ont analysé les nutriments de jeunes pousses de Prosopis chilensis avec cet outil. Dans cette étude, l'analyse vectorielle a été utilisée en vue d'examiner les effets de l'application des traitements sur les concentrations et contenus foliaires des éléments N, P, K ainsi que la biomasse sèche des aiguilles échantillonnées. Bruns (1973) a noté que la fertilisation affectait la qualité d'un arbre de Noël sur plusieurs aspects. Les traitements d'azote seul ou combinés au phosphore et au potassium, ont produit un feuillage plus dense, aux teintes plus vert foncé et doté d'aiguilles de plus grandes dimensions. Dans cette étude, la coloration foliaire recherchée par les producteurs correspondait au code 5GY 3/4 fourni par la Charte Munsell. Les objectifs spécifiques de la présente étude sont de déterminer les effets des quantités et fréquences d'applications du fumier de volailles sur (1) les concentrations foliaires en éléments nutritifs et (2) la couleur des aiguilles. 2.3 Matériels et méthodes 2.3.1 Les sites d'étude Les sites d'étude sont localisés à l'intérieur de deux plantations de sapin baumier appartenant à deux producteurs d'arbres de Noël du nord-ouest du Nouveau-brunswick. Le site de Saint-Quentin (47o 30' 35" Nord ; 67o 23' 18" Ouest) (Services Nouveau-Brunswick 2002), est situé à plus de 300 m d'altitude au centre d'un plateau dans la chaîne des montagnes des Appalaches sur un sol de type loam à limoneux. Le terrain est plat et couvert en majorité de forêts et de terres agricoles. Le pH est compris entre 5.5 et 6.5. Les températures moyennes pour le mois de janvier sont de -12° C et pour le mois d'août de 17° C (van Groenewoud 1983). La moyenne annuelle des précipitations est d'environ 991 mm. Les conditions météorologiques ayant prévalu durant la période d'étude sont présentées au tableau 1.1 Le site de Connors (47o 13' 0" Nord ; 68o 49' 59" Ouest) (Services Nouveau-Brunswick 2002) est situé à environ 45 km à l'ouest d'Edmundston. Le terrain est vallonné et le territoire est surtout couvert de forêts et de terres agricoles. Le pH est compris entre 5 et 5.5. La moyenne annuelle des précipitations est d'environ 976.4 mm. Les températures moyennes au mois de janvier sont de -7.4° C et au mois d'août de 23° C. Le sol est aussi de type loam à limoneux. Les plantations en étude comportent des arbres âgés de cinq ans et la mise en place du dispositif fut effectuée lorsque ceux-ci étaient âgés de deux ans. La plupart des semis étaient des sauvageons. 2.3.2 Le dispositif expérimental et les traitements de fertilisation Le dispositif expérimental a été établi selon un plan en blocs aléatoires complets et a été installé en juin 2004 dans deux plantations de sapins de Noël. On retrouve dans ce dispositif quatre blocs répétitions. A l'intérieur de chaque bloc, on retrouve treize rangées d'arbres comprenant six rangées pour les six traitements de fertilisation et une rangée d'arbres agissant comme une zone tampon entre deux traitements adjacents. L'utilisation des blocs aléatoires complets se justifie par l'hétérogénéité du matériel expérimental d'une part et d'autre part par les critères de blocking relatifs à des facteurs édapho-climatiques incontrôlés qui pourraient influencer les variables mesurées. L'objectif poursuivi dans le choix de ce plan expérimental est d'augmenter la précision de l'analyse en réduisant la variable aléatoire. La poudre sèche du fumier de volailles a été appliquée au pied des arbres selon les quantités prescrites par le dispositif expérimental. Le fumier utilisé est de la fiente de volailles déshydratée et non compostée, obtenue en vrac par les producteurs d'arbres de Noël auprès de l'entreprise WestCo filiale de ENVIREM Technologies INC. Il a une teneur en matière organique supérieure à 75 % et un pH de 6.3. Les différentes concentrations en éléments minéraux du fumier de volailles utilisées sont présentées dans le tableau 1.2 du chapitre I. L'engrais chimique dénommé McCain Fertilizer est du nitrate d'ammonium de formulation 20-10-10 (N-P-K). La fertilisation a eu lieu au mois de juin des années 2004 à 2006. Les traitements sont : témoin ; 0.5 kg de matière organique (M.O.) arbre-1an-1; 1 kg M.O. arbre-1an-1; 1 kg M.O. arbre-1 2 ans-1; 2 kg M.O. arbre-1 2 ans-1et 115 g de 20-10-10 (NPK) arbre-1an-1. 2.3.3 L'échantillonnage foliaire Les échantillons des aiguilles de la saison de croissance en cours ont été prélevés entre les mois de septembre et octobre des années 2004 à 2006, période où les teneurs en éléments nutritifs des aiguilles sont relativement stables et reflètent le mieux les besoins en éléments nutritifs de l'arbre. Au total 593 arbres ont été échantillonnés dont un arbre sur deux dans chaque unité expérimentale. Par la suite, 250 aiguilles par unité expérimentale ont été prélevées aléatoirement, séchées, pesées et les masses obtenues furent acheminées au Laboratory for Forest Soils and Environmental Quality (Faculty of Forestry and Environmental Management. University of New-Brunswick) pour les analyses foliaires en éléments minéraux. 2.3.4 La coloration foliaire La couleur des aiguilles de l'année courante a été déterminée à l'aide de la Charte Munsell (Munsell Color charts for plant tissues 1977). Les mesures de coloration foliaire ont été effectuées la même journée que l'échantillonnage foliaire, soit à l'automne des années 2004 à 2006 et ont porté sur les 1186 arbres du dispositif expérimental. 2.3.5 L'analyse vectorielle Dans l'analyse vectorielle, le contenu en nutriments est défini comme étant une fonction du poids sec de la plante et de la concentration en nutriments. Il s'obtient par le produit de la concentration et la biomasse sèche. L'analyse vectorielle permet d'examiner la composition en nutriments et la biomasse de la plante dans un diagramme unique où le contenu en nutriments (x), la concentration en nutriments (y) et la biomasse (z) satisfont à la fonction : x = f (y, z) (Haase & Rose 1995). Les valeurs des nutriments et poids sec de référence ou de contrôle sont généralement fixées à 100 afin de faciliter les comparaisons avec un point de départ commun. Les valeurs des autres échantillons sont ensuite placées par rapport à la référence (Haase & Rose 1995). Dans cette étude les valeurs de référence sont celles du traitement chimique de formulation 20-10-10. 2.3.6 Les analyses statistiques Le test d'ANOVA du logiciel de traitement des données SPSS 15.0 for Windows a été utilisé pour évaluer l'effet des traitements sur les concentrations et contenus foliaires en éléments minéraux à un seuil de signification de 10%. Le test de comparaison multiple Tukey du logiciel SPSS a été utilisé pour la comparaison des moyennes des traitements lorsque les tests de l'analyse des variances étaient significatifs. Le test LSD a été utilisé lorsque le test Tukey ne décelait pas de différences statistiques entre les moyennes des effets simples. L'évaluation de la coloration du feuillage a été effectuée avec le test de Khi carré pour l'évaluation des proportions des couleurs suivi d'un test d'analyse de variance en vue d'évaluer l'effet des traitements sur ces proportions de couleur. Le choix du seuil de signification à 10% se justifie d'une part par une variabilité initiale que l'on juge importante par rapport aux variables mesurées et d'autre part pour diminuer les risques d'erreur de type II (â ; ne pas rejeter H0.lorsque H0 est fausse) (Lemelin 2004 ; Scherrer 1984). 2.4 Résultats Les résultats des analyses de variance (ANOVA) sont présentés dans les tableaux 2.1 pour les concentrations foliaires ainsi que les biomasses sèches des aiguilles et 2.2 pour les contenus foliaires des éléments minéraux contenus dans les aiguilles échantillonnées. Tableau 2.1. Sommaire des analyses de variance : degrés de liberté (ddl) et valeurs des tests de F selon l'analyse des variances des concentrations foliaires en éléments nutritifs et les biomasses sèches des aiguilles.
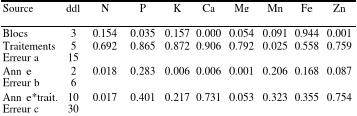
Tableau 2.2. Sommaire des analyses de variance : degrés de liberté (ddl) et valeurs des tests de F selon l'analyse des variances des contenus foliaires en éléments nutritifs.
2.4.1 Concentrations des éléments Azote Entre 2004 et 2006, on a noté une interaction significative (p=0.001 ; Tableau 2.1) entre traitements et années d'étude quant aux concentrations (Figure 2.1) foliaires en azote. Au cours de la saison de croissance de 2004, les concentrations foliaires en azote étaient élevées pour l'ensemble des traitements notamment chez les arbres issus des traitements organiques 1kg/an et 2kg/2ans. En 2005, les concentrations foliaires en azote ont diminué chez les arbres traités au fumier avec les quantités 1kg/2ans et 2kg/2ans. En 2006, une baisse des concentrations a été généralement observée chez tous les arbres traités, sauf pour les traitements au fumier de volailles bisannuels (1kg/2ans et 2 kg/2ans). Les concentrations foliaires en azote ont également varié selon les années (p=0.021 ; Tableau 2.1). Celles-ci étaient en moyenne de 2.34%, 2.24% et 2.17% en 2004, 2005 et 2006 respectivement.
Figure 2.1. Concentrations foliaires en azote en fonction des traitements et des années. p est la probabilité du test de l'interaction entre traitement et année. 2SE=0.09. Phosphore On a noté un effet significatif (p=0.019 ; Tableau 2.1) des traitements sur les concentrations foliaires en phosphore entre 2004 et 2006 (Figure 2.2). Les concentrations foliaires en phosphore ont diminué pour les traitements organique et chimique et sont restées élevées chez le témoin. Cependant ces concentrations des arbres traités au fumier de volailles avec les quantités 1kg/an et 1kg/2ans ne sont statistiquement pas différentes de celles du témoin. Il a également été observé une variation des concentrations foliaires en phosphore selon les années (p=0.000 ; Tableau 2.1). Celles-ci étaient en moyenne de 0.25 %, 0.23 % et 0.20 % en 2004, 2005 et 2006 respectivement.
Figure 2.2. Concentrations foliaires en phosphore en fonction des traitements. Les moyennes affectées de la même lettre ne sont pas statistiquement différentes au seuil á=0.1. p est la probabilité du test de l'effet des traitements sur les concentrations foliaires en phosphore. Potassium Aucun effet des traitements sur les concentrations foliaires en potassium n'a été constaté au cours de cette période (p=0.872; Tableau 2.1). Calcium Les traitements n'ont pas eu d'effet sur les concentrations foliaires en calcium. Toutefois, ces concentrations ont varié selon les années (p=0.062; Tableau 2.1). Celles-ci étaient en moyenne de 0.50%, 0.49% et 0.58% en 2004, 2005 et 2006 respectivement. Magnésium Entre 2004 et 2006, on a noté une interaction significative (p=0.019 ; Tableau 2.1) entre traitements et années (Figure 2.3) quant aux concentrations foliaires en magnésium. Ces concentrations ont généralement augmenté au cours de cette période pour l'ensemble des traitements y compris le témoin. Les concentrations foliaires en magnésium ont également varié selon les années d'étude (p=0.000 ; Tableau 2.1). Celles-ci étaient en moyenne de 0.07%, 0.1 % et 0.1% en 2004, 2005 et 2006 respectivement.
Figure 2.3. Concentrations foliaires en magnésium en fonction des traitements et des années. p est la probabilité du test de l'interaction entre traitement et année. 2SE=0.006 Oligo-éléments Aucun effet des traitements n'a été observé dans les concentrations foliaires en fer et en zinc entre 2004 et 2006. Par contre, on a noté un effet significatif (p=0.034 ; Tableau 2.1) des traitements sur les concentrations foliaires en manganèse entre 2004 et 2006 (Figure 2.4). Les concentrations foliaires en manganèse ont diminué chez le témoin et le traitement au fumier de volailles avec la quantité 0.5kg/an. Les concentrations foliaires en fer ont également varié selon les années d'étude (p=0.023 ; Tableau 2.1). Celles-ci étaient en moyenne de 48 ppm, 67 ppm, et 40 ppm en 2004, 2005 et 2006 respectivement.
Figure 2.4. Concentrations foliaires en manganèse en fonction des traitements. Les moyennes affectées de la même lettre ne sont pas statistiquement différentes au seuil á=0.1. p est la probabilité du test de l'effet des traitements sur les concentrations foliaires en manganèse. 2.4.2 Contenus des éléments Les contenus des aiguilles en azote ont augmenté de 2004 à 2006 (p=0.017 ; Tableau 2.2) chez tous les traitements sauf pour les traitements bisannuels (1kg/2ans et 2kg/2ans) (Figure 2.5). Selon les années, il a été observé des variations dans le contenu des aiguilles pour les éléments minéraux suivants : potassium (p=0.006 ; Tableau 2.2), calcium (p=0.006; Tableau 2.2), magnésium (p=0.001; Tableau 2.2), fer (p=0.168; Tableau 2.2) et zinc (p=0.087, Tableau 2.2). Par contre aucun effet des traitements n'a été constaté dans les contenus foliaires en phosphore (p=0.856), en calcium (p=0.906), en magnésium (p=0.792), en fer (p=0.558) et en zinc (p=0.759). Il a également été observé un effet des traitements (p=0.025 ; Tableau 2.2) dans les contenus foliaires en manganèse (Figure 2.6) ainsi qu'une interaction significative (p=0.053 ; Tableau 2.2) entre les traitements et les années dans les contenus foliaires en magnésium (Figure 2.7).
Figure 2.5. Contenus foliaires en azote en fonction des traitements et des années. p est la probabilité du test de l'interaction entre traitement et année. 2 SE= 0.3.
Figure 2.6. Contenus foliaires en manganèse en fonction des traitements. Les moyennes affectées de la même lettre ne sont pas statistiquement différentes au seuil á=0.1. p est la probabilité du test de l'effet des traitements sur les contenus foliaires en manganèse.
Figure 2.7. Contenus foliaires en magnésium en fonction des traitements et des années. p est la probabilité du test de l'interaction entre année et traitement. 2SE=0.012. 2.4.3 Biomasses sèches des aiguilles Aucun effet des traitements sur les biomasses sèches des aiguilles n'a été observé (p=0.995 ; Tableau 1.3). Cependant elles ont augmenté avec les années (Figure 2.8) ayant des valeurs plus importantes en 2006.
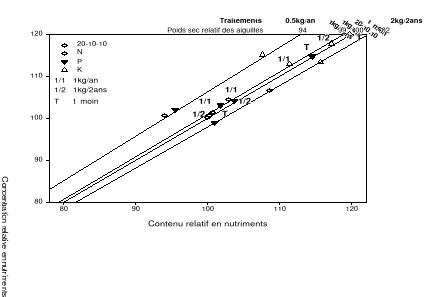
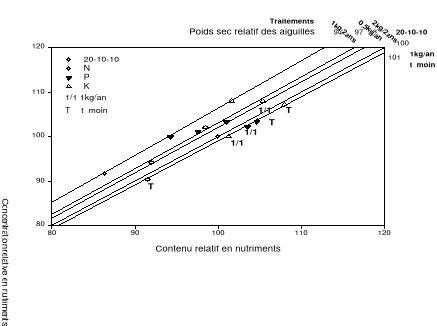
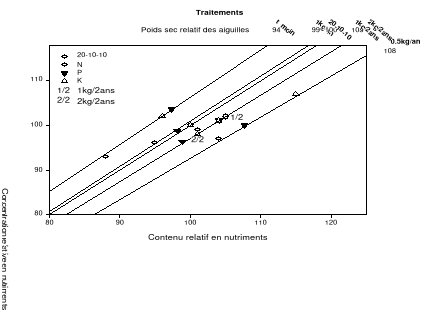
Figure 2.8. Valeurs moyennes de la biomasse sèche en fonction des années. Les moyennes affectées des lettres différentes sont statistiquement différentes au seuil á=0.1. p est la probabilité du test de l'effet des traitements sur la biomasse sèche des aiguilles. 2.4.4 L'analyse vectorielle Azote L'analyse vectorielle des aiguilles échantillonnées en 2004 indique une augmentation des concentrations et contenus foliaires en azote sans changement notable de la biomasse sèche pour tous les traitements à l'exception du traitement avec 0.5 kg/an de fumier de volailles par rapport à la référence (traitement chimique). Ces résultats observés au cours de cette année indiquent une consommation de luxe en azote pour tous les traitements à l'exception du traitement avec 0.5 kg/an de fumier de volailles par rapport à la référence (traitement chimique). À noter que les concentrations, contenus et biomasses sèches les plus importants furent observés chez les arbres traités avec 2kg/2ans de fumier de volailles (Figure 2.9a). En 2005, l'analyse vectorielle indique un effet d'antagonisme chez les traitements bisannuels avec le fumier de volailles de même que ceux n'ayant reçu aucune application de fertilisant comparativement à l'engrais chimique (Figure 2.9b). En 2006, Les arbres traités avec 1kg/2ans de fumier de volailles ont eu les concentrations et contenus foliaires en azote les plus élevés (Figure 2.9c). En général, les arbres témoins ont connu une baisse des concentrations et contenus foliaires de même que la biomasse sèche au cours des années 2004 à 2006. Phosphore L'analyse vectorielle des aiguilles échantillonnées en 2004 indique une augmentation des concentrations et contenus foliaires en phosphore sans changement de la biomasse sèche pour tous les traitements à l'exception du traitement avec 0.5 kg/an de fumier de volailles par rapport au traitement de référence. (Figure 2.9a). Ce résultat indique que le phosphore est en consommation de luxe comme observé chez l'azote au cours de cette année. En 2005, les traitements au fumier de volailles avec les quantités 1kg/2ans et 0.5kg/an ont induit une diminution des concentrations et contenus foliaires en phosphore sans changement de la biomasse. Ce résultat indique que le phosphore apporté en 2005 par la fertilisation avec ces quantités est en excès entraînant un effet d'antagonisme (Figure 2.9b). En 2006, aucun résultat significatif ne fut noté bien qu'on ait observé une hausse non significative de la biomasse sèche chez les arbres traités au fumier de volailles avec la quantité 0.5kg/an. Potassium En 2004 et 2005 tous les traitements y compris le témoin ont induit une augmentation en concentration, contenu sans changement de la biomasse sèche comparativement au traitement chimique (point de référence). Ce résultat indique une consommation de luxe en potassium chez ces traitements. En 2006, aucun résultat significatif ne fut noté bien qu'on ait observé une hausse non significative de la biomasse sèche chez les arbres traités au fumier de volailles avec la quantité 0.5kg/an. a) 2004
b) 2005
c) 2006
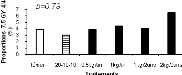
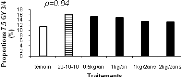
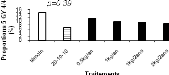
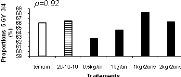
Figure 2.9. Réponses en nutriments (azote, phosphore et potassium) et poids sec des aiguilles du sapin baumier selon les traitements en 2004 (a), 2005 (b) et 2006 (c). 2.4.5 Couleurs du feuillage Les résultats du test de Khi carré n'étaient pas conformes aux conditions d'application de ce test puisque 80% des classes ne possédaient pas une fréquence théorique supérieure ou égale à 5 (Scherrer 1984). Pour y remédier, une seconde analyse a été réalisée en sélectionnant dans la base de données les couleurs les plus représentées (effectifs plus importants) et considérées optimales de par les meilleures couleurs observées. Cette analyse a porté sur les couleurs 7.5 GY 3/4; 7.5 GY 4/4 ; 7.5 GY 3/4 ; 5 GY 4/4 et 5GY 3/4. Les résultats obtenus ont satisfait aux conditions d'application du test de Khi Carré puisque aucune classe n'avait un effectif théorique inférieur à 5. La signification asymptotique (bilatérale) n'était pas significative (p=0.631). On en a déduit que la distribution des couleurs n'était statistiquement pas différente entre les traitements. Les proportions des cinq couleurs représentatives par bloc et par traitement ci-dessus énumérées ont été ensuite analysées à l'aide de l'analyse de variance (ANOVA). Aucun traitement n'a modifié la coloration foliaire entre 2004 et 2006. Les seuils de signification obtenus pour chacun des codes de couleur étaient : 7.5 GY 3/4 (p=0.68), 7.5 GY 4/4 (p=0.79), 7.5 GY 3/4 (p=0.84), 5 GY 4/4 (p=0.39), 5 GY 3/4 (p=0.92). a) b)
c) d)
e)
Figure 2.10. Effets des traitements sur les proportions des couleurs 7.5 GY 3/4 (a), 7.5 GY 4/4 (b), 7.5 GY 3/4 (c), 5 GY 4/4 (d) et 5 GY 3/4 (e).
2.5 Discussion 2.5.1 Analyses foliaires Les traitements organiques annuels n'ont pas modifié significativement les concentrations des éléments minéraux par rapport au traitement chimique contrairement aux traitements organiques bisannuels. De plus, les variations observées ont toujours été dans des proportions adéquates pour ces éléments indiquant l'absence d'effet toxique ou d'une déficience. Les traitements appliqués ont modifié les concentrations foliaires en azote au cours des années 2004 à 2006. Des résultats similaires furent observés par Leaf, Berglund & Léonard (1970) dans une expérience sur le feuillage de 40 espèces de pin rouge (Pinus resinosa, Ait.) au cours de laquelle, ils ont observé des différences significatives des concentrations foliaires en N, P, K, Ca et Mg sur une période de six années. Dans un essai de l'application d'un fertilisant sur l'épinette de Norvège, Tamm (1968) a montré des variations substantielles des concentrations en nutriments dans les parcelles témoins sur une période de six années comme observées par Leaf et al. (1970). En 2005 les concentrations des traitements annuels demeurent élevées par contre celles du témoin et des traitements bisannuels ont diminué fort probablement parce qu'aucun traitement n'a été appliqué au cours de cette année dans ces unités expérimentales. En ce qui concerne les concentrations observées chez le témoin en 2004, il est plausible de croire que l'azote ait été transloqué vers les aiguilles de l'année courante afin de maintenir une concentration comparable aux autres traitements. Toutefois, dès 2005, la concentration en azote a diminué chez le témoin. En 2006, la concentration foliaire en azote diminue chez tous les traitements. Cette diminution pourrait s'expliquer par la forte pluviométrie enregistrée au cours du mois de juin 2006 (Tableau 1.1; Chapitre I), période d'application des fertilisants qui aurait occasionné un lessivage de cet élément. Cependant, les concentrations en azote foliaire obtenues sont dans des proportions adéquates (entre 1.8 et 2% ; Hatch 1987). Les concentrations foliaires en phosphore chez les arbres traités sont en baisse comparativement au traitement témoin bien que les concentrations obtenues soient dans les limites adéquates (0.2 à 0.5% ; Heller et al. 1998). Cette baisse des concentrations chez les arbres fertilisés pourrait s'expliquer par une insuffisance du phosphore disponible pour le maintien de la biomasse végétale produite. Cette insuffisance serait attribuable à la fixation des ions phosphoriques aux hydroxydes de fer et d'aluminium, aux colloïdes minéraux et organiques (Blanchet 1959), les rendant de ce fait moins assimilables par les arbres. L'effet des concentrations en calcium observé durant les années 2004 à 2005 pourrait s'expliquer par un manque ou un excès d'humidité dans le sol qui interfère avec l'absorption du calcium (Kirkby & Pilbeam 1984). Aussi, Marschner (1986) attribue une baisse des concentrations en calcium à la compétition de cet élément avec d'autres cations tels que NH4+ (ammonium), K+ (potassium), Mg++ (magnésium) et le Na+ (sodium) pour son absorption par les racines. Les résultats indiquent des proportions adéquates en magnésium dans les tissus foliaires et une augmentation des concentrations et des contenus en fonction des traitements et des années dénotant ainsi de sa parfaite assimilation pour l'ensemble des traitements. En général, les concentrations et contenus en manganèse dans les tissus foliaires ne diffèrent pas de manière significative chez les arbres traités à l'engrais chimique et au fumier de volailles, laissant supposer une absorption de cet élément dans les mêmes quantités. L'effet des concentrations en fer et en zinc observé au cours des années pourrait être lié à des phénomènes d'antagonisme ou de synergie et même à des facteurs climatiques intervenant en milieu incontrôlé. 2.5.2 Analyses vectorielles Les analyses vectorielles n'ont pas montré de problème de déficience en azote, phosphore ou en potassium (Figure 2.9). Toutefois, un antagonisme a été noté chez l'azote au niveau des traitements bisannuels au fumier de volailles et les arbres témoins en 2005. De plus ces analyses ont montré que les traitements bisannuels au fumier de volailles ont produit les meilleurs effets sur les concentrations et contenus foliaires en azote des aiguilles échantillonnées en 2004 et 2006. Ce fait étant lié à une plus importante disponibilité des éléments minéraux associée à ces quantités et fréquences d'applications ainsi qu'une meilleure absorption de ceux-ci dans la solution du sol. La baisse des concentrations en phosphore chez les arbres traités peut être attribuable à un effet d'antagonisme de l'azote sur l'absorption du phosphore comme rapporté par Bekele, Hudnall & Tiarks (1999) lors d'une expérience dans une plantation juvénile de Pinus taeda suite à une fertilisation azotée. Aussi Olykan, Adams, Normeyer & McLaren (1995) ont observé une baisse significative des concentrations foliaires en phosphore dans une plantation de Pinus radiata âgée de 4 ans une année après une fertilisation azotée. Les variations de biomasse sèche observées dans l'analyse vectorielle reflètent bien les traitements administrés. La biomasse sèche des aiguilles chez le témoin diminue de 2004 à 2006 alors que celles des traitements organiques bisannuels fluctuent en réponse à la fertilisation appliquée en 2004 et 2006. 2.5.3 La coloration foliaire Les tests de coloration foliaire ont montré que la fertilisation aussi bien chimique qu'organique n'a pas influencé la couleur des aiguilles au cours de ces trois années d'étude. On peut supposer que les éléments minéraux majeurs tels que l'azote et le magnésium, constituants essentiels de la chlorophylle étant dans des proportions adéquates, la couleur des aiguilles ne pouvaient pas varier. Aussi est-il important de mentionner que la coloration recherchée par les producteurs correspondant au code 5GY3/4 a dominé dans toutes les unités expérimentales du dispositif étudié.
Les différents traitements ont fait varier les concentrations foliaires en éléments nutritifs à l'intérieur des proportions adéquates et n'ont pas modifié la coloration des aiguilles. De plus, l'analyse vectorielle a montré que les traitements bisannuels au fumier de volailles ont produit les meilleurs effets sur les concentrations et contenus foliaires en azote des aiguilles échantillonnées sans avoir varié la biomasse de celles-ci. Cette accumulation d'éléments nutritifs notamment l'azote dans les aiguilles sans allocation vers leur croissance suscite de l'interrogation sur les applications de ce fertilisant organique. Les questions qui se posent sont de savoir si ces applications résulteront à long terme à un effet toxique ou en un effet bénéfique pour la croissance des arbres. Aussi, les observations chez les arbres témoins laissent suggérer l'avantage d'une expérimentation sur des sites préalablement exempts d'application de fertilisants pour mener à bien une meilleure discussion des variables mesurées suite aux traitements. Eu égard aux résultats de l'analyse vectorielle chez les arbres traités avec les quantités bisannuelles de fumier de volailles en réponse à la fertilisation en 2004 et en 2006, le traitement 1kg/2ans de fumier de volailles pourrait être recommandée aux producteurs. L'application de cette quantité leur permettrait de réduire les coûts liés à l'achat des fertilisants et de la manutention en plantation d'arbres de Noël. Cependant, il conviendrait d'encourager la poursuite de cette étude et réaliser un suivi des nutriments dans le sol. CONCLUSION GÉNÉRALE Nos résultats indiquent que le traitement au fumier de volailles est aussi efficace que l'engrais chimique couramment utilisé en plantation d'arbres de Noël et que les quantité et fréquence d'applications du fumier de volailles ont influencé positivement (1) la capacité photosynthétique maximale (Amax), (2) la surface foliaire spécifique, (3) la respiration du sol, et (4) la croissance en diamètre au collet des arbres. La hauteur des arbres n'a pas été influencée par les traitements à cause de la taille de la flèche terminale par les producteurs. Les différents traitements de fumier de volailles ont également varié les concentrations foliaires en éléments nutritifs à l'intérieur des proportions adéquates et n'ont pas modifié la coloration des aiguilles. De plus, l'analyse vectorielle a montré que les traitements bisannuels au fumier de volailles ont produit les meilleurs effets sur les concentrations et contenus foliaires en azote des aiguilles échantillonnées. L'absence de différences très significatives entre le témoin et les traitements organiques pour la Amax, la surface foliaire spécifique et le taux relatif de croissance en diamètre observée dans le chapitre 1, de même que les concentrations toujours dans des proportions adéquates dans le chapitre 2 suscitent des interrogations sur l'opportunité de fertiliser les plants ou non à ce stade de croissance. De plus, la comparaison entre le témoin et le traitement chimique semble nous indiquer que la fertilisation chimique pourrait ne pas être optimale pour la croissance des arbres. Selon les résultats obtenus et les coûts associés à l'achat et à la manutention des fertilisants en plantation d'arbres de Noël, nous recommandons aux producteurs d'appliquer 1kg de fumier de volailles par arbre tous les deux ans. Il faut tout de même rappeler aux producteurs d'évaluer leur coût en fonction du prix de la tonne métrique et de la quantité appliquée en engrais chimique ou en engrais organique. Dans notre étude, nous appliquions 5 fois plus de matière organique (M.O.) que d'engrais chimique pour obtenir la même quantité d'azote (0,5 kg M.O. par arbre vs 0,1 kg 20-10-10 par arbre). Donc, le prix revient au même si la tonne d'engrais chimique de formulation 20-10-10 vaut 400$ et que la tonne de M.O. équivaut à 80$. Toutefois, la fertilisation organique améliore les propriétés physico-chimiques du sol contrairement à l'engrais chimique. Nous recommandons également d'évaluer la période de temps nécessaire pour l'apparition de carences nutritives chez le témoin et ainsi confirmer la nécessité de fertiliser ou non les arbres. Un suivi des nutriments dans la solution du sol pourrait aussi répondre à la question du lessivage des nutriments et suggérer des quantités d'application inférieures, voire 0,5 kg tous les deux ou trois ans. De plus, nous suggérons de surveiller les concentrations en oligo-éléments tels le cuivre et le zinc qui sont généralement abondants dans le fumier de volailles et qui pourraient s'accumuler dans le sol et causer éventuellement des effets toxiques chez les arbres. REFERENCES Albaugh, T.J., Allen, H.L., Dougherty, P.M., Kress, L.W. & King, J.S. (1998). Leaf area and above and belowground growth responses of loblolly pine to nutrient and water additions. Forest Science , 44: 328. Bauhus, J. & Khanna, K. (1999). The significance of microbial biomass and activity in forest soils. 77-110 in N. Rastin and J. Bauhus,(eds). Going underground: ecological studies in forest soils. Research Signpost, Trivandrum, India. Bekele, A., Hudnall, W.H. & Tiarks, A.E. (1999). Vector analysis identify loblolly pine (Pinus taeda L.) phosphorus deficiency on a Beauregard soil. General Technical Report-Southern Research Station, USDA Forest service, 30, 474-479. Blanchet, R. (1959). C.R. Acad. Agric., 45, 247-252. Bonin, P. (1971). Normes de reboisement des terres privées. Service de la restauration forestière. Ministère des terres et forêts, Québec. 36 p. Brix, H. (1983). Effects of thinning and nitrogen fertilization on growth of Douglas-fir: relative contribution of foliage quantity and efficiency. Can. J. For. Res., 13, 67-175. Brouwer, R. (1962a). Nutritive influences on the distribution of dry matter in the plant. Neth. J. Agric. Sci., 10, 399-408. Brouwer, R. (1962b). Distribution of dry matter in the plant. Neth. J. Agric. Sci., 10, 361-376. Brouwer, R. (1983). Functional equilibrium: sense or nonsense? Neth. J. Agric. Sci., 31, 399-408. Bruns, P.E. (1973). Cultural practices, fertilizing and foliar analysis of balsam fir Christmas trees. New Hampshire Agricultural Experiment Station, Durham. Bull. 501. p. Burton, D. & Beauchamps, E. (1994). Profile nitrous oxide and carbon dioxide concentrations in a soil subject to freezing. Soil Sci. Soc. Am. J., 58, p. 115-122. Carlyle, J.C., Carlyle & Than, U.B. (1988). Abiotic controls of soil respiration beneath an eighteen-year-old Pinus radiata stand in South-eastern Australia, J. Ecol., 76, pp. 654-662. Chabalier, P.F., Kerchove, V., Macary, H. (2006). Guide de la fertilisation organique à la Réunion, CIRAD, Chambre d'Agriculture de la Réunion, 304 p. Clark, J. (1961). Photosynthesis and Respiration. State University College of forestry at Syracuse University. Syracuse, New-york. Cornelissen, J.H.C. (1999). A triangular relationship between leaf size and seed size among woodyspecies: Allometry, ontogeny, ecology and taxonomy. Oecologia, 118, 248-255. Diaz-Raviña, M., Acea, M. J., & Carballas, T. (1993). Microbial biomass and its contribution to nutrient concentrations in forest soils. Soil Biol. Biochem. , 25, 25-31. Edwards, D.R. & Daniels, T.C. (1992). Environmental impacts of non-farm poultry waste disposal: a review. Bioresource Technology , 41, 9-33. Elliott, Lynch, L.F., Elliott & J.M., Lynch (1994). Biodiversity and soil resilience in D.J. Greenland and I. Szabolc, Editors, Soil Resilience and Sustainable Land Use, CAB International, Wallingford, UK, pp. 353-364. Environnement Canada (2007). Données climatiques normales et moyennes climatiques [En ligne]. Disponible : http://www.climate.weatheroffice.ec.gc.ca/climateData/monthlydata_f.html. Page consultée le 09 septembre 2007. Ericsson T. (1995). Growth and shoot: root ratio of seedlings in relation to nutrient availability. Plant and Soil, 169, 205-214. Farrar, J.F. & Jones, D.L. (2000). The control of carbon acquisition by roots. New. Phytol., 147, 43-53. Field, C., Merino, J. M. & Mooney, H.A. (1983). Compromises between water-use efficiency and nitrogen-use efficiency in five species of California evergreens. Oecologia, 60, 384-389. Girvan, M.S., Bullimore, A.S., Ball, J.N., Pretty & Osborn, A.M. (2004). Responses of active bacterial and fungal communities in soils under winter wheat to different fertilizer and pesticide regimens. Applied and Environmental Microbiology, 70, pp. 2692-2701. Haase, D.L., Rose, R. (1995). Vector analysis and its use for interpreting plant nutrient shifts in response to silvicultural treatments. Forest Science, 41, 51-66. Hatch, D.K. (1987). Soils and foliage analysis; tool for the Christmas tree grower. Note 1987: 06. 1987.09.04. Heller, R., Esnault, R. & Lance, C. (1998). Physiologie végétale-1. Nutrition. Edit Dunold, pp : 30-167. Imo, M., Timmer, V. R. (1997). Vector diagnosis of nutrient dynamics in Mesquites seedlings. Forest Science, Vol. 43, No. 2, 268-273. Janssens, I.A., Lankreijer, H., Matteucci, G. (2001).
Productivity overshadows Karin, E., Bertilsson, S., Cederlund, H., Stenström, J. & Hallin, S. (2006). Long-term impact of fertilization on activity and composition of bacterial communities and metabolic guilds in agricultural soil. Soil Biology and Biochemistry. Volume 39, Issue 1, January 2006, Pages 106-115. Keller, T. & W., Koch. (1962). Der Einfluss der Mineralstoffernäfrung auf CO2-Gaswechsel und Blattpigmentgehalf der Papel. I. Mitt. Schweiz. Anst. forstl. Versuchsw. 38(2): 253-282. Killham, K. (1994). Soil Ecology. Cambridge: Cambridge University Press, 260 p. Kingery, W.L., Wood, C.W., Delaney, D.P., Williams, J.C., Mullins, G.L., Van Santen, E. (1993). Implications of long-term land application of poultry litter on tall fescue pastures. J. Prod. Agric., 6, 390- 395. Kirkby, E.A. & D.J. Pilbeam (1984). Calcium as a plant nutrient. Plant Cell Environ., 7, 397-405. Körner, C.H. (1994). Biomass fractionation in plants: a reconsideration of definitions based on plant functions. In. Roy J, Garnier, éd. A whole plant perspective on carbon-nitrogen interactions, SPB, the Hague, 173-185. Kozlowski, T.T., P.J., Kramer & S.G., Pallardy (1991). The Physiological Ecology of Woody Plants, San Diego, California, Academic Press, inc. 657 p. Kramer, P.J. & T.T., Kozlowski (1960). Physiology of trees. McGraw-Hill Book co., Inc., 642 pp. Lambers, H. & H. Poorter (1992). Inherent variation in growth rate between higher plants : a search for physiological causes & ecological consequences. Advances in Ecological Research , 23, 187-261. Lambers, H., Chapin, F.S. III & Pons, T. (1998). Plant Physiological Ecology. Springer Verlag, Berlin. Leaf, A.L., J.V., Berglund & R.E., Léonard (1970). Annual variation in foliage of fertilized and/or irrigated red pine plantations. Proc. Soil Sci Soc. Am., 34, 677-682. Lemelin, A. (2004). Méthodes quantitatives. [En ligne]. Disponible : http://www.ucs.inrs.ca/cours/lemelin/eur8213/2_3.pdf Page consultée le 15 août 2007. Liu, Ristaino, B., Liu & J.B., Ristaino (2003). Microbial community structure in soils from organic and conventional agroecosystems. Phytopathology, 96 , p. S53. Maier, C.A., K. H., Johnsen, J. Butnor, L. Kress, P. Anderson (2002). Effects of nutrients and CO2 amendments on branch growth, phenology and gas exchange in 13 year-old loblolly pine (Pinus taeda) trees. Tree Phys: (in press). Mälkönen, E. (1990). Estimation of nitrogen saturation on the basis of long term-term fertilization experiments. Plant and Soil, 128, 75-82. Margolis, H. A., & Brand, D. G. (1990). An ecophysiological basis for understanding plantation establishment. Can. J. For. Res. 20: 375-390. Marschner, H. (1986). Mineral nutrition of higher plants. Chapter 8: functions of mineral nutrients macronutrients. Academic Press, 195-267. Meason, D.F., Markewitz, D. & Will, R.E. (2004). Annual fertilization and interspecific competition control: effects on in situ forest floor nitrogen fluxes of different-aged Pinus taeda stands in southeast Georgia, USA. Can. J. For. Res., 34, 1802-1818. Miller, H.G. (1981). Forest fertilization: Some Guiding Concept. Forestry 54(2), 157-167. Mitchell, A.K., Hinckley, T.M. (1993). Effects of foliar nitrogen concentration on photosynthesis and water use efficiency in Douglas-fir. Tree Physiology, 12, 403-410. Moore, P.A., Daniel, T.C., Sharpley, A.N. & Wood, C.W. (1995). Poultry manure management: Environmental sound options. Journal of soil and Water Conservation, 50, 621-327. Munsell Color (1977). Munsell Color Charts for Plant Tissues. Minsell Color. Baltimore. Nissen, Wander, T.M., Nissen & M.W., Wander (2003). Management and soil-quality effects on fertilizer-use efficiency and leaching. Soil Science Society of America Journal, 67, pp. 1524-1532. Olykan, S.T., Adams, J.A., Normeyer, A.H., McLaren, R.G. (1995). Micronutrient and macronutrient uptake by Pinus radiata and soil boron fraction as affected by added nitrogen and boron. New Zealand Journal of Forestry Science, 25, 61-72. Paarlahti, K., Paarlahti, A., Reinikainen & H. Veijalainen (1971). Nutritional diagnosis of Scots pine stands by needle and peat analysis, Commun. Inst. Forest. Fenn., 74, (5.), p. 58. Perrin, D., Laitat, E., Yernaux, M. & Aubinet, M. (2004). Modélisation de la réponse des flux de respiration d'un sol forestier selon les principales variables climatiques. Biotechnol. Agron. Soc. Environ. 8 (1), 15-25. Pirson, A. (1958). Mineralstoffe und Photosynthese. In Encyclopedia Plant Physiology (W. Ruhland, ed.). IV: 355-381. Spring-Verlag, Berlin. Raich, J.W., Potter CS. (1995). Global patterns of carbon dioxide emissions from soils. Global Biogeochem.Cycles, 9, p. 23-26. Richardson, B.W., Dickens, E.D. & McElvany, B.C. (2004). Old-field thinned loblolly pine plantation fertilization with diammonium phosphate plus urea and poultry litter 4 year growth and product class distribution results. General Technical Report Southern Research Station, USDA Forest Service. N°.SRS-71, pp. 395-397. Services Nouveau-Brunswick (2002). Atlas du Nouveau-Brunswick, deuxième édition révisée, 2002. Gouvernement du Nouveau-Brunswick, Fredericton. Sheriff, D.W. (1996). Responses of carbon gain and growth of Pinus radiata stands to thinning and fertilizing. Tree Physiol., 16, 527-536. Scherrer, B. (1984). Biostatistique. Gaétan Morin. Boucherville Smolander, H., Oker-Blom, P. (1989). The effect of nitrogen content on the photosynthesis of Scots pine needles and shoots. Ann. Sc.For., 46, 473s-475s. Sommerfeldt, T.G., Chang, C., Entz, T. (1988). Long-term annual manure applications increase soil organic matter and nitrogen and decrease carbon to nitrogen ratio. Soil Sci. Soc. Am. J., 52, 1668-1672. SPSS. 2002. SPSS version 15.0, SPSS Inc., Chicago, IL. Statistique Canada (1996). Programme national de données sur les forêts. Ministère des Ressources Naturelles du Canada. Service canadien des forêts, Ottawa, Canada. Swift, M. J., Heal, O. W. & Anderson, J. M. (1979). Decomposition in Terrestrial Ecosystems. in UoC Press, ed. Studies in ecology, Berkeley. Tamm (1968). An attempt to assess the optimum nitrogen level in Norway spruce under field conditions. Studia Forestall Suercia 61.pp. 67. Tejada, M., Hernandez, M.T. & Garcia, C. (2006). Application of two organic amendments on soil restoration. Effects on the soil biological properties. Journal of Environmental Quality, 35 (4): 1010-1017, Jul-Aug 2006. Teskey R.O., Gholz H.L. & Cropper W.P. (1994). Influence of climate and fertilization on net photosynthesis of mature slash pine. Tree Physiology, 14, 1215-1227. Timmer, V.R., Stone, E.L. (1978). Comparative foliar analysis of young balsam fir fertilized with nitrogen, phosphorus, potassium and lime. Soil Sci. Soc. Am. J., 42, 125-130. Van, Groenwoud H. (1983). Summary of climatic data pertaining to
the Climatic Van den Driessche, R. (1974). Prediction of mineral nutrient status of trees by foliar analysis, The Bot. Rev. 40 (3), pp. 347-393. Vose, J.M. & Allen, H.L. (1988). Leaf area, stem wood growth, and nutrition relationships in loblolly pine. Forest science , 34 , 547-563. Waring, R.H. & W.H., Schlesinger (1985). Forest Ecosystems: Concepts and Management, Academic Press, Orlando, Florida, 340p. Westerman, Tucker, R.L., Westerman & T.C., Tucker (1979). In situ transformations of nitrogen-15 labelled materials in Sonora Desert soils, Soil Science Society of America Journal , 43 , pp. 95-100. Westoby, M. (1998). A leaf-height-seed (LHS) plant ecology strategy scheme. Plant and soil, 199, 213-227. Wilhoit, J., Stokes, T., Samuelson, L., Johnson, J. (1999). Influence of poultry litter fertilization 18-year-old loblolly pine stand. Communications in Soil Science and Plant Analysis, Vol.30, N° ¾, pp. 509-518, 24. Wilson, B.J., Peters, N.C.B., Wright, K.J. & Atkins, H.A. (1988). The influence of crop competition on the seed production of Lamium purpureum, Viola arvensis and Papaver rhoeas in winter wheat aspects. Appl. Biol.,18, 71-80. Xu, M. & Qi, Y. (2001). Soil surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California. Glob. Chang. Biol., 7, pp. 667-677. |
|