Introduction
Progressivement, la reconnaissance des
invertébrés dans les processus touchant les
écosystèmes ainsi qu'en tant que composant majeurs de la
biodiversité, a provoquée une augmentation significative de leur
représentation dans les études (Andersen et al, 2002 ; Hites et
al, 2004 ; Rohr et al, 2006 in Groc ,2007). Cependant leur incorporation dans
les études en particulier pour les écosystèmes terrestres,
est plutôt considérée comme onéreuse (Andersen et
al. 2002 ; New, 1996 ; Oliver & Beattie, 1996; Schnell et al. 2003 in Groc,
2007).
En effet, l'identification n'est pas toujours faisable du fait
d'une expertise taxonomique limitée, du temps et/ou des moyens
financiers disponibles, et de la connaissance des caractéristiques
biologiques nécessaires pour identifier les organismes collectés.
Et il en résulte que la majorité de ces études
écologiques concernent plutôt les vertébrés et les
plantes vasculaires, bien que cela tendent à s'équilibrer au fil
du temps. Parmi les invertébrés, les abeilles sauvages
possèdent de nombreux attributs les rendant essentiels dans les
écosystèmes. En effet, de par leur importance écologique
dans la plus part des écosystèmes terrestres, leur
diversité et leur sensibilité aux changements environnementaux,
elles constituent des bioindicatrices pertinentes pour la santé des
écosystèmes (
www.écologie.gouv.fr).
Et en dépit de leur rôle décisif dans le
bien-être des hommes, le grand public et les décideurs ne savent
rien des conséquences de la perte de celles-ci qui sont pourtant d`une
ampleur considérable. Sur ce, elles assurent à elles seules
près de 85% de la pollinisation des plantes sexuées, elles nous
procurent aussi un certain nombre des biens qui possèdent une valeur
économique directe tels que: la nourriture, les médicaments, etc.
(Fao, 2007). Et pourtant aujourd'hui plus de la moitié de 369
espèces d'abeilles sauvages sont menacées. Les causes de ce
phénomène en grande partie sont connues, elles sont toutes dues
aux faits de l'homme, qui par sa présence a modifié
profondément les milieux naturels. En effet ces causes sont parfois
l'introduction de nombreuses espèces de plante à
fleurs horticoles et exotiques, ne fournissant pas de nourriture ou une
nourriture non appropriée à nos abeilles sauvages; les
modifications systématiques de notre territoire, engendrant la perte des
zones de nidification ou la construction de barrières physiques
difficilement franchissables; la présence des parasites causant ainsi
des maladies (varraoses, la fausse teigne ou la teigne de la cire, la loque
américaine); l'utilisation intensive des produits phytosanitaires; des
émissions électromagnétiques; les effets de la pollution
et du changement climatique et l'apparition de nouveaux prédateurs comme
le frelon asiatique(Vereecken et al, 2010 ;
www.abeillesentinelle.net).
Et suite à la méconnaissance prononcée de
la diversité, de la biologie et de la littérature taxonomique des
abeilles sauvages, en dépit de leur importance écologique et
économique au détriment de l'abeille domestique (Apis
mellifera) suscite un attrait justifiant le choix de cette étude
dans les milieux écologiquement contrastés de l'Université
de Kinshasa singulièrement au niveau du jardin expérimental et
dans les abords de la Faculté des Sciences.
L'objet de cette étude est :
> Déterminer la diversité des abeilles sauvages
(farouches) qui s'y trouvent ; > acquérir une meilleure
compréhension du réseau d'interaction plantes
abeilles et la phénologie des abeilles sauvages
(évaluer la succession
d'espèces d'abeilles au cours du temps ;
> discuter la richesse spécifique de ces espèces
au niveau de l'Université de Kinshasa et pour enfin faire une discussion
locale.
Ce travail est subdivisé en trois chapitres
précédés par une introduction. Le chapitre premier
présente les données bibliographiques des abeilles sauvages. Le
chapitre deuxième décrit le milieu de notre étude, les
matériel et méthodes utilisés pour la récolte de
nos données. Le chapitre troisième présente les
résultats obtenus et la discussion de ceux-ci. Enfin une conclusion va
clore notre étude.
Chapitre 1 : Données bibliographiques sur les
abeilles sauvages
1.1. Description systématique et morphologique
Les abeilles sauvages constituent un groupe le plus important
dans le règne animal. Elles appartiennent à l'embranchement des
arthropodes, classe des insectes, sous -classe des ptérygotes, l'ordre
des hyménoptères, le sous-ordre des apocrites, le groupe des
aculéates et dans la superfamille des Apoidea apiformes. Outre les
abeilles, la superfamille des Apoidea inclut également le Sphecidea.
C'est pour cette raison qu'on distingue les Apoidea apiformes et les Apoidea
spheciformes (Ifuta, 2007 ; Alexander, 1992 in Vanderplanck, 2009). Et 85 % de
ces espèces sont réputées solitaires en raison de leurs
moeurs de nidification. Certaines abeilles sauvages creusent leurs nids dans le
sol (Fig.1a), dans des tiges de plante (Fig.1c), dans des bois morts voire
pourris (Fig.1b) ou même sur le grès. D'autres construisent leurs
nids à partir de feuilles ou de mortier (mélange de terre ou de
sable et de salive) dans des cavités appropriées et
habituellement préexistantes telles que des coquilles d'escargot
(Fig.1d) ou des terriers creusés par d'autres insectes xylicoles
(Malyshev, 1968 ; Stephen et al, 1969 ; Iwata, 1976 ; Roubik, 1989 ; O'Toole
& Raw, 1991 ; Pouvreau, 2004 in Vanderplanck, 2009). Les nids comportent
une ou plusieurs petites loges qui forment autant de cellules individuelles
pour le couvain. Ces cellules larvaires peuvent être soit
distribuées d'un bout à l'autre d'une galerie ramifiée
soit disposées les unes à la suite des autres soit
agglomérées toutes ensemble (Wcislo & Cane, 1996 ; Pouvreau,
2004 in Vanderplanck, 2009). Les parois de ces cellules ou parfois même
les pains de pollen sont habituellement tapissés par les femelles
à l'aide d'une sécrétion hydrophobe issue de leur glande
de Dufour (Duffield et al, 1984 ; Hefetz, 1987 in Vanderplanck, 2009).
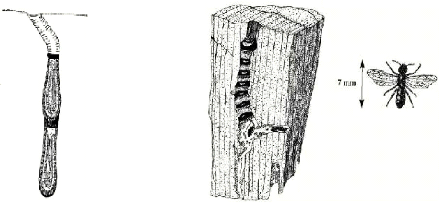
Fig.1a : Nid terricole d'Anthocopa cristatula (Osmiini,
Mégachilidés) (Pouvreau, 2004 in Vanderplanck, 2009)
Figure.1.b: Nid d'Heriades sp. (Mégachilidés) dans
un
morceau de bois (Pouvreau, 2004 in Vanderplanck, 2009)
|
|
|
Fig1c. : Nid rubicole d'Anthocopa ligurica (Osmiini,
Mégachilidés) (d'après Pouvreau,
2004 in Vanderplanck, 2009)).
|
|
Figure.1d : Nid d'osmie (Osmia bicolor,
Mégachilidés) dans une coquille d'escargot dissimulée sous
des brindilles sèches (d'après Pouvreau, 2004 in
Vanderplanck).
|
Halictidae, Melittidae, Meganomidae, Stenotritidae et
Dasypodaidae) et les langues longues (Megachilidae et
Apidae).
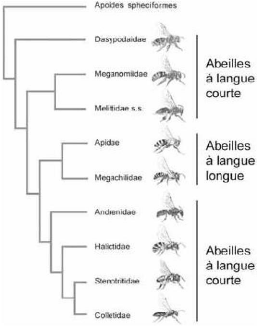
Fig.2 : arbre des Apoïdea distinguant les clades à
langue longue de ceux à langue courte (Danforth et al, 2006 in Michez,
2007).
L'abeille est un insecte, caractérisé par un
corps subdivisé en 3 parties (la tête, le thorax et l'abdomen),
qui sont tous0 couverts d'un squelette externe appelé exosquelette. Ce
dernier confère à l'insecte sa rigidité et permet
l'ancrage des muscles car il constitue une barrière protectrice pour
l'insecte contre les intempéries et des prédateurs. Elles se
caractérisent aussi par la présence des 4 ailes membraneuses.
1.2. Biogéographie des abeilles et centre de
diversité
Bien que les études faunistiques soient limitées
et idiosyncratiques, elles montrent que les abeilles abondent dans la plupart
des habitas de tous les continents, excepté l'Antarctique (Michener,
1979; Roubik, 1989; Neff & Simpson, 1993 in Vanderplanck, 2009) pourvu que
les plantes à fleurs soient présentes et que des
conditions adéquates pour la nidification soient
observées. Actuellement, le nombre d'espèces d'abeilles dans le
monde est estimé à environ 20.000 (dont plus de 16 000
décrites) (
www.discoverlife.org) dont 6
de ces familles sont présentes dans la région subsaharienne
(Urban et al, 2010).
En ce qui concerne le centre de diversité des genres
d'Apoïdea Apiformes, il est plus observé dans la région
néotropicale mais le plus grand éventail d'espèces se
rencontre dans les régions xériques et tempérées
(Michener, 1979 in Vanderplanck, 2009). L'abondance d'abeilles solitaires la
plus impressionnante s'observe dans les parts du monde chaud et aride
présentant un caractère semi-désertique; plus
particulièrement dans l'Ouest de l'Amérique du Nord, au Turkestan
ainsi qu'au sein des régions adjacentes, dans le bassin
méditerranéen, dans des parties de l'Afrique du Sud, en Australie
et dans l'Ouest de l'Amérique du Sud.
1.3. Description biologique
L'abeille sauvage, insecte pollinisateur majeur, son
développement est très complexe. En effet, les abeilles sauvages
sont pour la plupart solitaires, c'est-à-dire qu'elles ne forment pas de
société complexe composée d'un couple «royal» et
d'une «cour» d'ouvrières: leur biologie est finalement assez
proche de la plupart des autres insectes, puisqu'on retrouve un mâle, une
femelle et leur descendance. Il existe également d'autres espèces
d'abeilles sauvages«sociales», notamment les bourdons et les
Halitidae.
Le cycle de vie d'une abeille sauvage solitaire est
très simple. Dès l'émergence, les mâles patrouillent
à la recherche des femelles et l'accouplement a lieu dans
l'environnement immédiat de leur naissance. La femelle consacrera
ensuite l'essentiel de sa courte existence (quelques semaines au plus) à
la confection d'un nid au sein duquel elle accumulera pollen, nectar et autres
dérivés floraux qui constitueront l'alimentation de base de sa
progéniture. Et on ne parle pas de famille nombreuse... tout au plus une
dizaine d'oeufs qui seront pondus au fond de leur nid, à l'abri
des prédateurs et autres parasites. Certaines abeilles
solitaires ont troqué leur existence laborieuse pour un mode de vie
strictement parasitaire: on les appelle les abeilles «coucous». Comme
l'oiseau qui squatte le nid d'autres espèces pour y pondre ses oeufs,
ces abeilles sont passées maîtres dans l'infiltration des nids
d'autres espèces d'abeilles sauvages. Une fois au coeur du nid, elles se
débarrassent rapidement des oeufs de l'abeille-hôte et
déposent un oeuf sur les réserves nutritives accumulées
avant de quitter les lieux. A côté des abeilles sauvages
solitaires, il ya aussi les abeilles sauvages sociales. Cependant ces
dernières se caractérisent par une organisation sociale bien
organisée ou structurée, donnant naissance aux castes (couple
royal, ouvrier, soldat et les couvains).
En effet, la cohésion profonde chez les abeilles
sauvages sociales ne passe pas du tout, comme chez les mammifères, par
la sexualité ou la reproduction, puisque l'énorme majorité
des individus de la société est stérile. Curieusement,
c'est la nourriture qui va créer entre les membres de la
communauté des relations extraordinairement étroites, et rendre
chaque individu dépendant de ses congénères. La
communication dans la société peut s'effectuer via les substances
(chimiques ou gazeuses) émises ou les phéromones dans la
reconnaissance des individus conspécificiques, du territoire ou encore
pour déceler les plantes à fleurs déjà
visitées (marquage répulsif) et non visitées (marquage
attractif) (Malekani, 2009;Vereecken et al, 2010 ; Kapyla, 1978 in vereecken et
al, 2009).
Pour ce qui est de la nourriture, les abeilles ont deux
principales sources de nutriment : le nectar, source de glucides (sucres), qui
se transformera en miel, et le pollen des fleurs, unique source de
protéine nutritive. (www.jacheres-apicoles; Latham et al ,2007 ; Fao,
2007).
1.4. Relation plantes -abeilles
Avec 250.000 à 260.000 espèces décrites,
les Angiospermes représentent le groupe de plantes vascularisées
le plus diversifié et le plus étendu sur terre (Soltis &
Soltis, 2004 in Vanderplanck, 2009). Ce succès évolutif trouve
probablement son origine en grande partie au sein des nombreuses interactions
plantes-insectes, clef de voûte des écosystèmes actuels et
aussi à l'élaboration d'un nid à proximité des
ressources alimentaires, étape indispensable à
l'établissement et au maintien des populations d'abeilles sauvages sur
des sites favorables (Malyshev 1932, Cane, 1991 in Vereecken et al, 2006). Ces
interactions peuvent être de plusieurs types: antagonistes dans le cas
d'insectes phytophages ; mutualistes dans le cas d'insectes pollinisateurs.

(a) (b)
Fig.3 : photo de Bombus terrestris (a) et Bombus
pratorum (b) entrain de butiner (Vereecken et al, 2010).
Au cours de leurs visites florales, les d'abeilles
transfèrent des grains de pollens d'une fleur à l'autre
(fécondation croisée) au moyen de sa forte pilosité se
trouvant au niveau de son abdomen en complicité avec les
métatarses de ses pattes postérieures
|
|
|
Fig.4b : Brosse ventrale d'une femelle d'Anthidium sp
(Megachilidae) (Pouvreau, 2004 in Barbouche et al, 2009)
|
Figure 4a : Patte postérieure de la femelle de
Lasioglossum leucozonium (Halictidae) (Pouvreau in Barbouche et
al, 2009)
Latham (2007) révèle une importance
capitale et/ou fondamentale des abeilles dans la santé des
écosystèmes particulièrement au niveau des plantes du
Bas-Congo par le fait que les abeilles sont actives sur ces dernières et
les visitent durant toutes les périodes de l'année(saison
sèche et saison de pluie).Cette attraction périodique des
abeilles sur les plantes, fait que la région du Bas-Congo soit
considérée comme une province prépondérante pour
les revenus économiques du pays et aussi les sièges de nombreuses
études biologique. Cette association (plantes -abeilles) confère
à la province du bas Congo une flore fortement diversifiée qui se
montre par l'existence de beaucoup d'espèces exotiques et
indigènes. Elles pollinisent quasiment toutes les familles
présentes dans la région dont :Manguifera
indica(Anarcadiaceae), Manihot glaziovii (Euphorbiaceae), Maniotes expansa
(Connaraceae), Milletia versicolor (Fabaceae), Musa spp ( Musaceae), Nicotiana
tabacum, Nymphea alba (Nympheaceae), Ocimum graussimun (Lamiaceae), Paropsia
brazzeana, Pentaclethra
etveldeana (Capparaceae),Persea americana (Lauraceae),
Petersianthus
macrocarpus (Lecythidaceae), Phraseolus lunatus
(Fabaceae), Phytolacca dodecandra (Phytolaccaceae), Piliostigma thonnigii
(Caesalpiniaceae), Peptadeniastrum africanum (Mimosaceae),Psidium guajava
(Myrtaceae), Psorosperun febrifigum (Clusiaceae), Rauvolfia vomifora
(Apocynaceae), Sapuim corutum (Euphorbiaceae), Sarcocephallus latifoluis
(Rubiaceae), Sechium edule (cicurbitaceae), Secundaca lonipedunculata
(Polygalaceae), Brassica rappa (Brassicaceae), Brassica juncea (Brassicaceae),
Chaetocarpus africanus (Euphorbiaceae), Cissus rubiginosa (Vitaceae), Citrullus
lanatus (Cucurbitaceae), Cogniauxia podolaena (Curbitaceae), Crescentia cujete
(Bignoniaceae), Crossoptuyx febrifuga (Rubiaceae), Croton sylvaticus
(Euphorbiaceae), Croton mubango (Euphorbiaceae), Cucumeropsis mannu
(Cucurbitaceae), Dichrostachys cinerea (Mimosaceae), Duranta erecte
(Verbenaceae), Elaeis guineensis (Arecaceae), Eucalyptus spp (Myrtaceae),
Gaertnera paniculata (rubiaceae), Grevilla robusta (Proteaceae), Harungana
madagascariensis (Clusiaceae), Helianthus annuus (Asteraceae), Hyptis
suaveolens (Lamiaceae), Lantana camara (Verbenaceae), etc.
Donc pollinisation est un service central à toutes les
fonctions de l'écosystème, la perte de la biodiversité est
inconcevable sans parler de l'effet écologique néfaste qui
suivrait la perte d'un grand nombre des pollinisateurs. (Vereecken et al, 2010
; Fao, 2007).
Cependant des différentes stratégies de butinage
peuvent être observées chez les abeilles. Certaines espèces
arborent de spécialisation dans leur choix de pollen, visitant un nombre
restreint de plantes disponibles dans leur habitat (monolectisme,
oligolectisme) tandis que d'autres butinent un plus large éventail de
plantes hôtes (Tableau 1) (Muller &Kuhlmann, 2008 in Vereecken et al,
2009).
Tableau.1: caractéristiques préférentielles
des abeilles sur les choix floraux.
|
Catégorie
|
Définition
|
|
Monolectisme
|
Récolte de pollen sur une seule espèce de plante
même en présence d'une ou de plusieurs espèces sympatriques
du même genre
|
|
Oligolectisme
|
Récolte de pollen sur deux à plusieurs
espèces appartenant à un genre de plante.
|
|
Mésolectisme
|
Récolte de pollen sur plus de quatre genres de plantes
appartenant à deux ou trois familles de plante.
|
|
Polylectisme
|
Récolte de pollen sur plusieurs familles de plantes.
|
Chapitre 2 : Milieu d'étude, Matériels et
méthodes 2.1. Milieu d'étude
2.1.1. Situation géographique
Notre étude s'est effectuée à l'Unikin
(commune de Lemba). Les sites choisis sont ceux des abords de la Faculté
des sciences et du jardin expérimental de la Faculté des Sciences
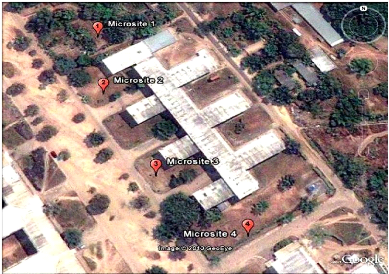
(fig.4). Ils comprennent 4 microsites pour l'étude
d'échantillonnage dont le jardin expérimental (a), les abords du
département de mathématique et de l'informatique (b), les abords
du département de la biologie et de l'environnement (c) et les abords de
l'antenne de l'Ocpt (d). Les coordonnés géographiques
correspondant à chaque microsites sont repris dans le tableau suivant
:
Tableau.2: coordonnés GPS et altitude moyenne de chacun de
microsites échantillonnés.
|
Microsites ou foyers
d'observations
|
Coordonnés GPS
|
Altitude moyenne
|
|
A
|
lat 4°25'3.58» S; long 15°18'29.63» E
|
441m
|
|
B
|
lat 4°25'5.05»S,long 15°18'29.81»E
|
444 m
|
|
C
|
lat 4°25'06.98»S;long 15°18'30.85»E
|
444 m
|
|
D
|
lat 4°25'8.32» S ;long 15°18'32.54»E
|
445 m
|
Fig.5
: Illustration de différents microsites
prospectés.

Jardin expérimental
Les abords du département
des
Mathématiques

Les abords du département de Biologie Les abords
de l'antenne de l'Ocpt
Figure .6 :
: Localisation des microsites sur le site de
l'université de Kinshasa (source googleearth 30 juin 2010).
2.1.2. Climat
L'Université de Kinshasa (Unikin) étant
située dans la province de Kinshasa, elle
- à -
présente un climat du type AW4 c'est dire un
climat tropical humide où la
température moyenne mensuelle le plus froid est
supérieure à 18 °c avec une période
de
sécheres ,2008). Les
se de 4 mois et une période de pluviosité
de 8 mois (Lukoki
valeurs moyennes mensuelles des précipitations
et des températures obtenues à partir de la
station météorologique de BINZA ont permis de tracer le
diagramme ombrothermique de la Fig.6.Les moyennes des
données mensuelles sur la précipitation et la
température sont reprises en annexe 5.

60
40
20
80
0
120
100
Temperature CC)
PrOcipitations (mm)
250
200
150
100
50
0
P récipitation s (m m ) Tem péra tures (°C
)
JAN F E V MAR A V R M A I JU IN JU IL A O U S E P O C T N O V
DEC
M ois
Fig.7: diagramme ombrothermique des moyennes mensuelles des
précipitations et des températures de 2000 à 2009(source
METTELSAT BINZA).
Ce graphe montre la variation des moyennes mensuelles de
pluviosité et des moyennes mensuelles de température de 2000
à 2009. La température moyenne maximale a été
enregistrée en mars (25,5°C) et la température moyenne
minimale a été observée en juillet (22,3°C).
Cependant le mois de novembre se révèle le plus pluvieux (251,7
mm) et le mois le moins pluvieux est le juillet (0,7mm).
2.1.3. Végétation
La végétation est dominée par les plantes
adventices ou colonisatrices, et avec une prédominance des
poacées (Panicum maximum, Panicum repens, Cynodon dactylon,
Eleusine indica, Anthephora cristata, Panicum brevifolium, Eragrostis ciliaris
,Sporobolus puramidalis, Eragrostis tenella, Paspalum notatum etc..) ; des
cyperacées(Mariscus cyperus,Cyperus triangularis, Cyperus rotondus,
Cetaria barbata etc...) et des asteracées (Tithonia
diversifolia, Chromolaena odorata, Tridax procumbers, Synedrella nodiflora ,
Bidens pilosa, Ageratum conyzoides).
En dehors de ces dernières, on note aussi quelques
espèces adventices à l'instar de : Sida acuta, Spermacos
latifolia, Euphorbia hirta, Euphorbia heterophylla , Euphorbia tirrucali,
Croton hirtus, Phyllantus amarus, Desmodium tortiosum,
Asystasia gangetica, Boerhavia diffusa, Oldelendia
corymbosa, Urena lobata, Commelina diffusa auxquelles s'ajoutent quelques
arbustes et arbres : Caesalpinia pulcherina, Treculia africana, Ficus elastica,
Lagestroema speciosum, Moringa sp, Averrhoa
caramboa,etc...(Dibaluka,2009).
2.2. Matériels et Méthodes 2.2.1. Collectes
des spécimens
Les collectes des données se sont effectuées
à l'Unikin particulièrement au niveau du jardin
expérimental et sur les abords de la faculté des sciences.
A cet effet, nous avons effectué 23 descentes sur
terrain au rythme de deux descentes par mois durant 12 mois (17 mai 2009 au 30
mai 2010). Les techniques des récoltes utilisées sont le
piégeage aux coupelles et celui de filet.
2.2.2. Piégeage aux coupelles ou piégeage
passif
Les coupelles sont de petits bols colorés qui attirent
les insectes et les capturent dans l'eau (Potts et al, 2007). Ces
dernières constituent une méthode indirecte ou non tributaire du
récolteur, car la prise des abeilles est déléguée
par les pièges autonomes (piège à coupelle) (Dufrêne
et Carré, 2009) et aussi de référence pour des nombreuses
études sur l'apidofaune. L'avantage qu'a cette méthode, est que
l'échantillonnage est plus prolongé et se fait
simultanément dans tous les sites possibles retenus pour les
échantillonnages (
www.alarmproject.net ;
http://online.sfsu.edu/-beeplot/).
Des nombreux insectes floricoles, parmi lesquels les abeilles
sauvages sont irrésistiblement attirées par certaines couleurs
(Kirk, 1984 ; Dafni et al, 1990 ; Thorp 1999 in Potts et al ,2007).En
général, la couleur jaune est la plus attractive pour les
hyménoptères (abeilles, guêpes et fourmis) et les
diptères (mouches) ; le blanc attire les mouches et les insectes
phytophages, mais peut également repousser certains
hyménoptères ; le rouge est attractif pour certains
coléoptères.
Comme précisé ci-dessus, différents types
d'insectes sont attirés par différentes couleurs, ce qui importe
d'utiliser un ensemble de coupelles de différentes couleurs, parmi
lesquelles le bleu, le jaune et le blanc sont souvent les plus efficaces.
2.2.2.1. Principe
Le principe était simple du fait que nos coupelles
colorées remplies d'eau savonnée sont perçues comme une
grande fleur. Les abeilles s'en approchent, pensant pouvoir y récolter
le pollen et/ou nectar, elles s'y noient dès qu'elles entrent en contact
avec la surface de l'eau savonnée. Sans savon dans l'eau les
pièges sont inefficaces (Potts et al, 2007).
2.2.2.2. Description de l'équipement
D'après cette méthode, les coupelles doivent
respecter un diamètre de 10 à 20 cm et une profondeur de 7
à 15 cm. Ces dimensions correspondent à une grande partie des
bols en plastique et des récipients communément
utilisés.
Les coupelles étaient traitées à
l'intérieure et à l'extérieure à l'aide de peinture
colorée fluorescente sous rayon ultra-violet (U-V).Cette étape
est indispensable puisque les insectes sont beaucoup plus sensibles que l'homme
aux U-V, ces derniers attirent davantage les insectes que les couleurs non
fluorescentes sous rayon U-V (
http://online.sfsu.edu/~beeplot/;Dufrêne
et Carré, 2009).
Deux petits trous de 2 à 3 mm de diamètre sur
les cotés de la coupelle ,à environ 1cm en dessous du bord
supérieur ont été réalisés .Ces trous
permettront d'évacuer l'excès d'eau accumulée dans les
coupelles au cours de forte pluie ,tout en conservant les insectes
capturés dans le piège .Une seconde paire de trous aux
caractéristiques semblables était également
réalisée afin de permettre de fixer les coupelles à une
ficelle dans l'éventualité où le piège devrait
être installé en hauteur, dans un arbre ou dans un buisson.
2.2.2.3. Installation des pièges à
coupelles
L'installation des pièges à coupelles
était fonction de la structure aérienne de la flore du milieu
visité par les abeilles et aussi de la zone où la
probabilité de trouver les abeilles est importante.
Au niveau du jardin expérimental, nous avons
installé 15 pièges à coupelles rassemblés en 5
groupes de 3 coupelles (1bleu ; 1jaune et 1 blanche).Un groupe était
placé au centre et les 4 autres dans les 4 extrémités du
jardin la contrée. Chaque groupe des coupelles étaient
accrochés sur une branche d'arbre car il s'agissait des buissons
(fig.8a).

Figure 8a: l'emplacement des Figure 8b :l'emplacement des
pièges à coupelles dans les pièges à
coupelles au ras du sol
buissons au jardin expérimental. sur les abords de la
faculté des
sciences.
Dans les abords de la faculté, les coupelles
étaient placées individuellement au ras du sol afin que les
pièges soient bien vus par les abeilles par ce que ledit milieu
était dominé par les plantes herbacées (fig.8b).
2.2.2.4. Relevé des pièges à
coupelles
Une fois placée, les pièges à coupelles
sont récoltés après 30 heures de séjours qui
couvrent toute la période d'activité des abeilles. Après
récolte, Chaque échantillon était placé dans des
fioles portant information du site, de la date et de l'heure de la capture,
dans lesquelles on a ajouté l'alcool dénaturé à 70%
ce qui permettra de conserver les spécimens récoltés
jusqu'à leur préparation (Westphal et al, 2009 ;
http://online.sfsu.edu/~beeplot).
2.2.3. Piégeage au filet ou piégeage
actif
Cette méthode directe est tributaire du
récolteur (Dufrêne et Carré, 2009),elle exige à un
récolteur à parcourir le terrain d'étude muni de son filet
pendant une durée de temps bien définie (7 heures du temps) de
manière à couvrir toutes les périodes d'activités
des abeilles et de capturer certaines espèces d'abeilles susceptibles de
ne pas visiter les coupelles colorées (p.ex. les espèces visitant
des fleurs qui ne sont pas à proximité des coupelles) ou qui sont
capables de s'en échapper (p.ex. les espèces de gros gabarit)
(Potts et al, 2007). A cet effet la capture des spécimens d'abeilles a
été faite au moyen de filet entomologique du type manuel.
Nous avons subdivisé les sites en 4 sections ou 4
foyers d'observation par ce que les activités des abeilles
diffèrent selon le temps. D'après nos observations, des
nombreuses espèces abeilles butinent très tôt matin (6
à 7h30'), et une majorité butine entre 10 et 11 heures (Potts,
2007), et c'est dans cet esprit que nous avons délimité nos sites
en foyers d'observation ou en section sus évoqué. Et les abeilles
actives les aprèsmidi.
En ce qui concerne le piégeage au filet, nous avons
pratiqué deux types de méthodes: la capture des abeilles en plein
vol et la capture des abeilles sur les inflorescences visitées pour la
récolte de pollen et/ou de nectar.
2.2.4. Préparation des abeilles pour leur
conservation et identification
La préparation des spécimens capturés
consistait à faciliter l'identification ultérieure. Pour ce
faire, nous avons étalés les spécimens capturés sur
une frigolite, puis épinglés au niveau du thorax par une
épingle de la marque Entomophinx n°1. Ensuite, les membres (ailes,
pattes, antennes) ont été bien écartés, car ces
parties sont importantes pour l'identification (fig. 9). Après
l'épinglage, nos échantillons ont été
laissés à l'air libre pendant 3 jours pour assurer leur
séchage; nous les avons ensuite placés dans la boite
entomologique (Fig. 10).

Fig.9: épinglage d'une abeille (Potts et al, 2007)

Fig.10: illustration des abeilles préparées dans
la boite entomologique
En ce qui concerne l'identification de nos spécimens
d'abeilles, elle a été faite possible par Dr Alain Pauly de
l'institut royal des Sciences naturelles de Belgique du département
d'entomologie, spécialiste des Abeilles africaines.
2.3. Analyse statistique
Les méthodes d'analyse statistique sont distinctes et
variés proposées par plusieurs auteurs dont DAGET (1976) et
SOUTHWOOD (1978) in Noudjoud(2006). Dans ce contexte nous avons
évalué la biodiversité au niveau á ou
diversité locale (composition et structure des espèces)
d'après whittaker (1960) in Marcon (2010).
2.3.1. Evaluation de la biodiversité au niveau
á 2.3.1.1. Les indices écologiques de composition
Pour exploiter les résultats, plusieurs indices
écologiques de composition sont utilisés tels que la richesse
totale, la richesse moyenne et l'abondance rélative.
2.3.1.1.1. Richesse totale
Selon MULLER(1985) in Noudjoud(2006), la richesse totale
représente l'un de paramètres fondamentaux
caractéristiques d'un peuplement. La richesse totale S est le nombre
total des espèces que comporte un peuplement considéré
dans un écosystème donné (RAMADE, 1984 in Louadi et al,
2010).
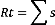
Où :
> Rt : richesse totale des espèces d'un peuplement.
> s : nombre d'espèce
2.3.1.1.2. Richesse moyenne
La richesse moyenne correspond au nombre moyen des espèces
contactées à chaque relevé (Blondel, 1979 in Riad, 1992).
La richesse moyenne est représentée par la formule suivante :
Si
S= Nr
Où :
> S : richesse moyenne d'un peuplement donné.
> Si : le nombre moyen observé à chacun des
relevés. > Nr : le nombre de relevés.
2.3.1.1.3. Abondance relative
L'abondance relative est le rapport du nombre des individus de
l'espèce prise en considération au nombre total des individus de
toutes espèces confondus (ZAIME et GAUTIER, 1989 in Noudjoud ,2006).
Elle est représentée par la formule suivante :
A.R= N/X100
N
Où :
> A.R. (°/°) : l'abondance relative ou
fréquence centésimale.
> Ni : le nombre des individus de l'espèce prise en
considération. > N : le nombre total des individus de toutes
espèces confondues.
2.4.1.1.4. Fréquence d'occurrence et constance
La fréquence d'occurrence d'une espèce est le
rapport exprimé en pourcentage entre le nombre total de
prélèvement où cette espèce est notée et le
nombre total de tous les prélèvements effectués.
Pi*100
F =
P
Où :
> F : fréquence d'occurrence ;
> Pi: nombre total de prélèvements
contenant l'espèce prise en considération ; > P : nombre total
de prélèvement effectués.
Selon Dajoz (1985) in Noudjoud (2006), on distingue :
> Les espèces omniprésentes (F:100%) ; >
constantes (75=F=100) ;
> les espèces régulières (50=F=75) ; >
les accessoires (25 =F =50) ;
> et les espèces accidentelles (F=25).
2.3.1.2. Les indices écologiques de structure
Parmi les indices synthétiques ou
non-paramétriques, basés sur les abondances relatives des
espèces, l'indice de SHANNON et WEAVER(1963) in Marcon (2010) sont les
plus fréquemment utilisés. Ces indices tendent à
synthétisent à la fois le nombre d'espèces à la
fois le nombre d'espèces et l'équilibre de leur
répartition dans le milieu (DUFRENE, 1992 in khalaf et al, 2009).
Les coefficients calculés reliés aux
caractéristiques des espèces sont les suivants :
H'=-- ? Pi * log2Pi
Où :
> H' :l'indice de diversité ;
> Pi=ni/N (abondance relative des espèces)
;
> ni : le nombre d'individus dans le premier groupe
taxonomique ; > N : le nombre d'individus dans la station ;
Cet indice permet d'effectuer une mesure de la composition en
espèces d'un écosystème, en termes du nombre
d'espèces et de leurs abondances relatives. Cependant, des peuplements
à physionomie très différente peuvent avoir la même
diversité : H' est maximal quand toutes les espèces sont
également représentées dans l'échantillon.
Il convient donc de calculer parallèlement aux indices
de diversité H' et l'équitabilité E, en rapportant la
diversité observée à la diversité théorique
maximale par équirepartition des effectifs entre les espèces
présentes :
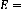
H'
log2 S
L'équitabilité varie de 0 et 1 : elle tend vers
0 quand la quasi-totalité des effectifs est concentrée sur une
même espèce, ce qui signifie les caractères d'un milieu
relativement peu diversifié soumis à de facteurs contraignants
(BARTLETT ET HIORNS, 1973 in Colignon et al, 2010), elle est de l'ordre de 1
lorsque toutes les espèces ont une même abondance.
L'espérance d'Hulrbert représente le nombre d'espèces
espérées dans un tirage aléatoire de 100 spécimens
et ce pour le site d'échantillonnage .Il s'exprime en espèce/100
spécimens. L'indice de l'espérance de Hulrbert (Es) évalue
la diversité d'un site. La formule simplifiée par Hulrbert(1971)
in Louadi et al (2010) est :
Es = [1 - (N - ni/N)100]
Où
> ni : nombre des spécimens de l'espèce
;
> N : nombre total des spécimens de la station ;
> Es : le nombre d'espèces espéré dans
une prise aléatoire de 100 spécimens ; unité :
espèce/100 spécimens.
2.4.2. Distribution d'abondance
C'est la distribution des fréquences obtenues en
classant les espèces par ordre de fréquences relatives ou
absolues décroissantes. Elle permet de comprendre la distribution
spatiale des espèces et la structure de la population d'abeilles. Cette
étude est menée par la méthode de Log-linéaire
(modèle de MOTOMURA). Les calculs et l'établissement des graphes
des droites pour cette méthode sont rendus possibles par le logiciel de
statistique Origin 6.1.
Chapitre 3 : Résultats
3.1. Faune des abeilles sauvages
L'étude de la biodiversité des Abeilles sauvages
a mis en évidence 17 espèces recensées à
l'Université de Kinshasa plus précisément au niveau du
jardin expérimental et sur les abords de la Faculté des Sciences
sur un total de 59 individus ou spécimens récoltés,
regroupés dans 8 genres et repartis dans 4 familles (tab.3). Les images
des espèces d'abeilles récoltées sont reprises en annexe
6.
Tableau 3: Espèces d'abeilles sauvages inventoriées
à l'université de Kinshasa.
|
Famille
|
Sous-famille
|
Genre
|
Sous-genre
|
Espèce
|
|
Apidae
|
Apinae
|
Crocisapidia
|
|
chandleri
|
|
Amegilla
|
|
sp
|
|
Thyreus
|
|
sp
|
|
Xylocopinae
|
Xylocopa
|
Koptortosoma
|
imitator
|
|
nigrita
|
|
albiceps
|
|
calens
|
|
Mesotrichia
|
combusta
|
|
Halictidae
|
Halictinae
|
Halictus
|
Seladonia
|
jucundus
|
|
|
|
|
|
|
Megachilidae
|
Megachilinae
|
Megachile
|
Chalicodoma
|
rufipennis
|
|
sp
|
|
bituberculata
|
|
sp2
|
|
eurymera
|
|
Gronoceras
|
cincta
|
|
Ceolioxys
|
|
sp
|
|
Euaspis
|
|
erythros
|
3.2. Phénologie
Les abeilles sont des espèces dont leurs
activités sont fonction de plusieurs facteurs à l'occurrence: la
disponibilité de nourriture (présence des fleurs pour le
butinage), le changement des saisons (pluie ou sèche), la
présence ou l'absence du soleil, etc. Le tableau 4 indique la
répartition de nos récoltes pendant la durée de
l'étude.
Tableau.4 : Nombre des spécimens listés par famille
durant l'année d'étude (2009- 2010).
|
Année
|
2009
|
2010
|
|
Mois
|
Mai
|
Jn
|
Jt
|
Ao
|
Sept
|
Oct
|
Nov
|
Dec
|
Jan
|
Fév
|
Mars
|
Avr
|
Mai
|
|
Apidae
|
16
|
1
|
2
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
2
|
4
|
6
|
|
Halictidae
|
0
|
0
|
1
|
2
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
|
Megachilidae
|
3
|
4
|
6
|
6
|
0
|
0
|
0
|
0
|
0
|
1
|
0
|
3
|
1
|
Les abeilles sauvages sont actives selon les facteurs
climatiques et la disponibilité de ressources florales. Les effectifs de
ces derniers varient selon les mois, et les résultats notés dans
le site de l'Unikin montrent bien que la plupart des abeilles sauvages sont
mieux représentées. Leurs représentations coïncident
avec la floraison d'un maximum de plantes et avec l'installation de conditions
clementes. Tels que ce graphe nous montre, on note que les Apidae sont
nombreuses au mois de mai (premier pic) et la population disparaît
à partir du mois d'août jusqu'au mois de janvier, et elles
reprennent leurs activités au mois de février pour être
maximale au mois de mai.
En ce qui concerne les Megachilidae, elles apparaissent
à partir du mois Mai pour être maximales au mois de
juillet-août. Elles disparaissent également à partir du
mois de septembre jusqu'au mois de janvier. Elles reprennent leur vol au mois
de février jusqu'au mois de mai. Les Halictidae n'ont été
seulement récoltées entre le mois de juillet et août. Nous
ne les avons pas récoltées dans les restes de la période
de l'étude.
3. 3.Evaluation de biodiversité par les indices
écologiques de Composition 3.3.1. Abondance relative
Les résultats sur l'abondance relative de chaque
espèce sont consignés sur le tableau ci-après :
Tableau .5: Abondance relative des abeilles sauvages au niveau de
l'Université de Kinshasa.
|
Espèces
|
Nombre d'individus
|
Abondance relative%
|
|
Amegilla sp
|
8
|
13,6
|
|
Crocisapidia chandleri
|
2
|
3,4
|
|
Thyreus sp
|
1
|
1,7
|
|
Xylocopa albiceps
|
1
|
1,7
|
|
Xylocopa combusta
|
8
|
13,6
|
|
Xylocopa imitator
|
4
|
6,7
|
|
Xylocopa nigrita
|
5
|
8,5
|
|
Xyolocopa callens
|
4
|
6,8
|
|
Total Apidae
|
33
|
55,9
|
|
Coelioxys sp
|
1
|
1,7
|
|
Euaspis erythros
|
1
|
1,7
|
|
Megachile rufipennis
|
10
|
16,9
|
|
Megachile sp2
|
1
|
1,7
|
|
Megachile bituberculata
|
3
|
5,08
|
|
Megachile cincta
|
1
|
1,7
|
|
Megachile eurymera
|
3
|
5,08
|
|
Megachile sp
|
3
|
5,08
|
|
Total Megachilidae
|
23
|
39
|
|
Halictus jacundus
|
3
|
5,08
|
|
Total Halictidae
|
3
|
5,08
|
Les résultats du tableau ci-haut montrent les
différentes valeurs d'abondances relatives trouvées pour chaque
taxon. On note que le nombre des individus en 2010 est plus faible que celui de
2009. Cependant pour l'année 2009, on remarque la présence de
trois familles (Apidae, Halictidae et Megachilidae) tandis qu'en 2010 on
observe la présence des Apidae, des Megachilidae et avec une absence
remarquable des Halictidae.
En ce qui concerne l'abondance des espèces, la famille
des Apidae et celle des Megachilidae sont les plus abondantes avec une
proportion égale de 47,7%, suivie des Halictidae avec 5,08.
Apidae Megachilidae Halictidae
47.1%
47.1%
5.88%
Fig.11 : Répartition du nombre des espèces par
familles.
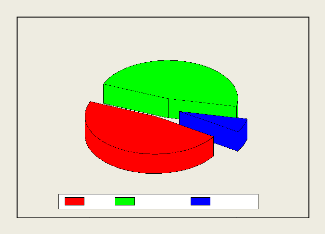
Cependant pour ce qui est de l'abondance des individus ou des
spécimens, la famille des Apidae est plus abondante, représentant
à elle seule 55,9 % des spécimens observés, suivie des
Megachilidae avec 39 % d'abondance et vient ensuite la famille des Halictidae
avec 5,08 %.
5.08%
39%
55.9%
Apidea Halictidea Megachilidea
Fig. 12: Abondance des spécimens par familles.
3.3.2. Richesse spécifique (totale) et
moyenne.
La valeur de la richesse spécifique totale(S) et celle
de la richesse spécifique moyenne(s) telles que reprises dans le tableau
suivant, nous montrent que l'Université de Kinshasa est plus
diversifiée en termes des abeilles sauvages.
Tableau.6 : Les valeurs de la richesse spécifique
totale(S) et moyenne(s).
|
Richesse spécifique totale (S)
|
Richesse spécifique moyenne(s)
|
|
17
|
1,95
|
3.3.3. Indice d'occurrence et fréquence
Tableau.7 : Fréquence d'occurrence et constante des
abeilles sauvages au niveau de l'université de Kinshasa.
|
Espèces
|
Occurrence
|
Fréquence d'occurrence
|
Statut écologique
|
|
Amegilla sp
|
8
|
34,78
|
Accidentelle
|
|
Coelioxys sp
|
1
|
4,34
|
Accidentelle
|
|
Crocisapidia chandleri
|
2
|
8,69
|
Accidentelle
|
|
Euaspis erythros
|
1
|
4,34
|
Accidentelle
|
|
Halictus jucundus
|
2
|
8,69
|
Accidentelle
|
|
Megachile bituberculata
|
2
|
8,69
|
Accidentelle
|
|
Megachile cincta
|
1
|
4,34
|
Accidentelle
|
|
Megachile eurymera
|
1
|
4,34
|
Accidentelle
|
|
Megachile rufipennis
|
5
|
21,73
|
Accidentelle
|
|
Megachile sp
|
2
|
8,69
|
Accidentelle
|
|
Megachile sp2
|
1
|
4,34
|
Accidentelle
|
|
Thyreus sp
|
1
|
4,34
|
Accidentelle
|
|
Xylocopa albiceps
|
1
|
4,34
|
Accidentelle
|
|
Xylocopa combusta
|
3
|
13,04
|
Accidentelle
|
|
Xylocopa calens
|
3
|
13,04
|
Accidentelle
|
|
Xylocopa imitator
|
3
|
13,04
|
Accidentelle
|
|
Xylocopa nigrita
|
3
|
13,04
|
Accidentelle
|
D'après ce tableau, nous avons enregistré une seule
espèce accessoire, il s'agit d'Amegilla sp. D'autres
espèces sont accidentelles.
3.4. Evaluation de la biodiversité par les indices
écologiques de structure
3.4.1. Indices de Shannon -Weaver, indice de
l'espérance de Hurlbert et indice d'équitabilité de
Pielou.
L'indice de diversité de SHANNON-WEAVER (H') et
équitabilité des espèces récoltées sont
repris dans le tableau 10.les calculs de l'indice de diversité de
SHANNON-WEAVER (H') et de l'équitabilité sont repris en annexes3
et en annexe 4 pour l'espérance de Hulrbert.
Tableau .8: Les différentes valeurs calculées par
l'indice de Shannon-Weaver, de l'espérance Hulrbert et
d'équitabilité.
|
Indices
|
Valeurs
|
|
Indice de Shannon-Weaver(H')
|
3,71 bits
|
|
Indice de diversité maximale(Hmax)
|
4,09
|
|
Equitabilité
|
0,91
|
|
Indice de l'espérance d'Hulrbert(Es)
|
16 ,14 espèce/100 spécimens
|
L'indice de diversité de Shannon (H') qui est de 3,71
bits, se rapproche de la diversité maximale (Hmax) dont la valeur est de
4,09 ce qui veut dire que le peuplement des abeilles sauvages au niveau de
l'Unikin est diversifié et que la richesse spécifique est
importante. L'équitabilité est élevée de 0,91.
Cette Valeur indique la population est en équilibre entre elle. La
diversité spécifique est élevée d'après
Hulrbert avec 16,14 espèces par 100 spécimens, ce qui veut dire
l'espérance de tirer aléatoirement le nombre des espèces
dans 100 individus est 16.
3.4.2. Distribution d'abondance
Les espèces récoltées durant la
période d'étude sont classées par la fréquence
absolue et relative décroissante. Nous avons en abscisse le rang des
espèces ou le nombre des espèces, et en ordonné le nombre
des spécimens.
La courbe logarithmique d'abondance des espèces
exprimées en effectifs et classées par rangs décroissants
révèle que les espèces listées dans le site
obéit à la loi de log normale de Motomura (progression
géométrique), qui stipule que chaque taxon utilise des ressources
auxquelles elle a accès selon sa position hiérarchique. La valeur
du coefficient de corrélation calculée (r =0,96547) tend vers 1,
c'est-à-dire la distribution des espèces au sein de l'Unikin
obéit la loi de log normale ou géométrique de Motomura.
Donc les partages des ressources du milieu, ainsi que l'occupation du milieu se
fait selon une hiérarchie bien établie (du plus grand au plus
petit). Les calculs
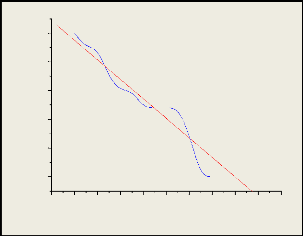
Nombre des specimens(logni)
0,8
0,6
0,4
0,2
0,0
1,0
-2 0 2 4 6 8 10 12 14 16 18
Nombre des espèces
y=-0,06792x+0,9498 r=0,96547
de la distribution d'abondance pour les espèces de
l'Unikin sont repris en annexe 1 et 2.
10
8
6
4
Nombre des specimens(Ni)
2
0 2 4 6 8 10 12 14 16 18
Nombre des espèces
0





Fig.13 : représentation graphique de la distribution des
espèces d'abeilles sauvages.
3.2. Discussion
L'étude de la biodiversité des abeilles sauvages
au niveau de notre site d'étude a révélé la
présence de 3 familles (Apidae, Halictidae et Megachilidae) sur 6
familles (Halictidae, Andrenidae, Collectidae, Megachilidae, Apidae et
Mellitidae) durant une année échelonnée du 17 mai 2009 au
30 mai. Cette constatation corrobore avec les résultats de symphonie
naturelle et celui de Kisantu (Bukaka, 2010 communication personnelle; Kasidi,
2010 communication personnelle).
En ce qui concerne l'abondance des familles en terme des
spécimens au niveau du site de l'Unikin, les Apidae sont abondantes
(55,07%), suivie des Megachilidae (47%) et les Halictidae en dernière
position (5,08%).Par ailleurs, l'inventaire des abeilles sauvages établi
au niveau de Symphonie naturelle par Bukaka(communication personnelle),
révèle la famille des Megachilidae comme la plus abondante (49%)
suivie les Halictidae en exéco avec les Apidae(25,5%). Par contre au
niveau du jardin botanique de Kisantu, l'abondance des abeilles sauvages
s'observent en forte proportion chez les Apidae (43,7%), puis les Halictidae
(34,4%) et les Megachilidae (21,9%) (Communication personnelle). Cette faible
proportion des Halictidae par rapport à la Symphonie naturelle et au
jardin botanique de Kisantu peut s'expliquer par la manque des ressources
florales spécifiques recherchées par les abeilles par rapport aux
autres sites (Symphonie naturelle et Jardin botanique de Kisantu), car la
Faculté des sciences et ses environs se situent en plein milieu urbain.
Ce qui n'est pas le cas avec les autres sites.
Pour ce qui est de la diversité des familles, au niveau
de l'Unikin la famille des Apidae et celle des Megachilidae sont les plus
diversifiées avec une proportion égale soit de 47,5% et afin la
famille des Halictidae avec 5%.De ce fait la présente étude ne
montre pas les résultats similaires avec ceux du jardin botanique et de
la symphonie naturelle car l'abondance notable d'espèce au niveau du
jardin botanique de Kisantu est plus enregistrée dans la famille des
Halictidae(44,6%),suivie des Apidae(33,9%) et les
Megachilidae(21,4%)(Kasidi,2010 communication personnelle). Et quant à
la forêt
de symphonie naturelle, les résultats
enregistrés contrarient les résultats de l'Unikin, car la
symphonie naturelle présente 43,3% pour les Apidae, 30% pour les
Halictidae puis 26,7% pour les Megachilidae (Bukaka, 2010).De ce fait, ces
résultats peuvent s'expliquer par le statut écologique
particulier des espèces au sein de la famille et aussi par les habitats
exploités.
La richesse spécifique totale enregistrée
à l'Unikin est faible (17 espèces) par rapport à la
symphonie naturelle (30 espèces) et du jardin botanique de Kisantu (54
espèces).Cette faible diversité spécifique observée
dans notre étude sur le site l'Unikin résulterait d'une forte
activité anthropique caractérisée par le
défrichement des arbres et l'installation de nouvelle infrastructure
académique et aussi sur le suréchantillonnage au niveau du jardin
botanique de Kisantu .En revanche, l'activité des abeilles et la liste
des espèces sont réduites.
En considérant les différents résultats
obtenus par la fréquence d'occurrence des espèces d'abeilles
sauvages sur le site de l'Unikin, les espèces accidentelles sont
majoritaires contre une espèce accessoire (Amegilla sp). Par
ailleurs les résultats obtenus sur la fréquence d'occurrence au
niveau de la symphonie naturelle (Bukaka, 2010) relève bien que les
espèces accidentelles sont majoritaires par rapport aux espèces
accessoires. De ce fait, la justification de ces différents
résultats serait plus due à la raréfaction de plantes
utiles pour le butinage et à la destruction récurrente des
habitats.
La diversité assez impressionnante
dégagée par l'indice de Shannon-Weaver (3,71 bits), traduit une
forte diversité sur le site de l'Unikin. Cette information est aussi
confirmée par la valeur de l'espérance de Hulrbert
calculée toujours dans ledit site où on espère tirer
aléatoirement 16 espèces pour 100 individus. Ce qui signifie
l'apidofaune est encore riche dans notre site, dominé par les
activités anthropiques.
individus que pour la disponibilité des ressources
florale. Et quant à la distribution d'abondance, les espèces de
l'Unikin obéit à la loi de log linéaire ou de la
progression géométrique de Motomura.
Conclusion et perspectives
L'objectif de ce travail a été de
déterminer la diversité des abeilles sauvages au niveau de
l'Université de Kinshasa plus précisément sur les abords
de la Faculté des sciences, ainsi qu'au jardin expérimental. A
l'issu de 23 prélèvements pour cette présente
étude, axée sur la connaissance de la biodiversité des
abeilles sauvages du 17 mai 2009 au 30 mai 2010 au niveau de
l'Université de Kinshasa, nous avons inventorié 17 espèces
sur un total de 59 individus, regroupés dans 8 genres puis repartis dans
3 familles (Apidae Halictidae et Megachilidae), ce qui représente une
proportion de 1/3 sur l'ensemble des familles connues. Cependant la famille des
Apidae est la plus représentée et celle des Halictidae est la
moins représentée.
En ce qui concerne la fréquentation des espèces,
l'espèce Amegilla sp est la plus remarquable et les moins
remarquables sont Euaspis erythros, Coelioxys sp, Megachile cincta,
Megachile sp2, Xylocopa albiceps, Xylocopa chandleri, Xylocopa imitator,
Thyreus sp.
Pour ce qui est de la phénologie, le mois de mai est le
plus fréquenté par les abeilles sauvages tandis que du septembre
au janvier les abeilles disparaissent.
Et quant à l'écologie, le peuplement des
abeilles sauvages au niveau de l'Unikin est en équilibre avec une
diversité élevée.
Ce travail constitue un point du départ de nos
recherches futures. Au vu de l'originalité des abeilles sauvages, il
serait opportun de continuer à établir des inventaires plus
exhaustifs à travers toute l'étendue de l'Université de
Kinshasa et à tous les coins de la ville de Kinshasa de manière
à mieux maitriser les connaissances des vecteurs de pollen. Leur
protection efficiente contre la dégradation de leurs habitats sera utile
ou d'une importance capitale de gestion optimale des écosystèmes
et de l'environnement général.
Références bibliographiques
1. Barbouche N. & Chouchène M., 2009
: Inventaire des abeilles sauvages en Tunisie et impact des pesticides
sur leur biodiversité. Institut national agronomique.6p.
2. Colignon P., Hastir P., Gaspar C. & Francis,
2010 : Effets de l'environnement proche sur la biodiversité
entomologique en cultures maraîchères de plein champ. Parasitica,
2000,56(2-3) :59-70.
3. Dibaluka S., 2009 : Notes des travaux
pratiques de l'écologie végétale à l'intention des
étudiants de première licence biologie (écologie).
4. Dufrêne E. et Carré G.,
2009 : Relevés faunistiques et les méthodes
d'échantillonnage des populations d'abeilles.41p. CNRS, INRA.
5. Fao, 2007 : Plan d'action de l'initiative
africaine sur les pollinisateurs(IAP) ,42p. ISBN 978-92-5-205893-9.
6. Groc S, 2007: Structure taxonomique et
application de la notion de minimalisme taxonomique à la
mymécofaune des Nouragues, Guyane Française. Mémoire de
Master 2 SEP (systématique, Evolution et paléontologie, parcours
systématique et évolution).Université Pierre et Marie
Curie-Paris VI, 32 p.
7. Ifuta, 2007:cours de zoologie destiné
aux étudiants de deuxième graduat biologie, Tome 1,116 p,
Université de Kinshasa.
8. khalaf G., Slim K., Abi-ghanem C., Nakhle K.,
Fakhri M., 2009: caractérisation et corrélation des
paramètres biotiques et abiotiques des eaux du Nahr El
Bared.19p.Lebanese science journal, vol.10, n°1,2009.
9. Latham. P &Konda K M, 2007: les plantes
utiles du Bas-Congo, 256 p. ISBN n°9780955420818.
10. Louadi K., Bendifallah L & Doumandji SE., 2010
: Apoidea et leur diversité au nord d'Algérie.
Publication scientifique, 18(1) :85-102.
11. Malekani JM, 2009 : cours
de l'éthologie destiné aux étudiants de première
licence biologie (écologie), Université de Kinshasa,60 page.
12. Marcon E, 2010: Mesure de la
biodiversité, écologie des forêts, 58p. CNRS, INRA.
13. Michez D, 2007: La nouvelle
classification des abeilles (Hyménoptère, Apoidea, Apiformes) ou
la chute de l'abeille mellifère (Apis mellifera L) de son
piédestal.4p.Osmia n°1-Hiver 2006-2007.
14. Noudjoud M. ,2006 : contribution
à la connaissance des abeilles sauvages (Hyménoptère,
Apoidea) dans les milieux naturels et cultivés de la région de
khenchela, Mémoire de master, Université de Constantine, 139p.
15. Potts S.G., Roberts S., Vereecken N.J et pauly
A., 2007 : Estimation de la biodiversité en abeilles: manuel de
méthodologie. 21p. Darwin initiative Project/15/021, Manuel
méthodologique.
16. Punga K.J, 2009 : cours de
l'entomologie destiné aux étudiants de première licence
biologie, Université de Kinshasa.
regions.Ecological Monographs,78(4),208,pp.653-671.
17. Riad E.B, 1992:Densité de l'avifaune
dans deux subéraies du Maroc à l'aide de quadrat et
I.K.A.Bull.inst.Sci., Rabat, 1992, n°16, pp.144-151.
18. Urban E, Pauly A. & Kuhlmann .M,
2010: the bees of subsaharian Africa.145p.volume 7(2010), ISSN
1784-1283(Hard copy)-ISSN 1784-1291(on line pdf), D/2010/0339/2.
19. Vanderplanck M. 2009: Métabolisme
stérolique de deux espèces solitaires spécialistes sur
saule, Mémoire d'étude approfondie, Université de
Mons-Hainaut, l78p.
20. Vereecken N,Michez D,Colomb & wollast, 2010
:Connaitre et aider nos abeilles sauvages.acceuillir nos abeilles
sauvages,4 pages. L'homme et l'oiseau 3/2010.
21. Vereecken NJ,Toffin E& Michez,
2006:Observation à la biologie et à la nidification de
quelques abeilles sauvages psammophiles d'interêt wallonie,10 pages.Revue
trimestrielle de conservation de la nature et gestion durable
d'ardenne et gaume. 4ième trimestre 2006.Parcs
et réserves.Volume 61- fascicule 4.
22. Vereecken NJ & Michez D. & 2010 :
les abeilles sauvages, une biodiversité insoupçonnée, 4p.
Université de mons, laboratoire de zoologie.
23. Vereecken NJ., Schewenninger H., Gogala A.,
Stuart PM R., 2009: mise à jour de la distribution de l'abeille
du lierre,Collectes hederae schmidt et westrich.2p.Osmia n°3.
24. Vereecken NJ & Barbier E, 2009 :
Premières données sur la présence de l'abeille asiatique
Megachile (Callomegachile) sculpuralis SMITH
(Hymenoptera, Megachilidae) en europe. Osmia n°3-2009 ; 3 pages.
25. Westphal C.,Ricardo B., Gabriel C., Ellen L.,
Morison N., Petanidou T., Potts S.G., Roberts S., Szentgyorgyi H., Tsheulin
T,Vissiere B., Woyciechowski M.,Jacobus C., Biersmeijer, William E., Settele,
2009: Measuring bee diversity in different european habitats and
biogeographical.
Sites internet
1.
www.alarmproject.net/efficacité
des pièges colorés sur l'attraction des abeilles/consulté
le 3/08/2010.
2.
www.jacheres-apicoles.fr/importance
des abeilles sur la santé des écosystèmes/consulté
le 12/11/2009.
3.
www.abeillesentinelle.net/rôle
des abeilles dans l'agriculture/ consulté le 12/11/2009.
4.
www.écologie.gouv.fr/rôle
des abeilles dans la santé des écosystèmes/consulté
le 16/03/2010.
5.
www.
discoverlife.org/diversité des abeilles à
l'échelle mondiale /consulté le 03/08/2010.
6.
http://online.sfsu.edu/~beeplot/efficacité
des coupelles sur l'attraction des abeilles /consulté le 03/08/2010.
7.
http://Zoologie.umh.ac.be/hymenoptera/
les espèces d'abeilles sauvages /consulté le 12/07/2011
43
Table des matières
Introduction 1
Chapitre 1 : Données bibliographiques sur les abeilles
sauvages 3
1.1. Description systématique et morphologique 3
1.2. Biogéographie des abeilles et centre de
diversité 5
1.3. Description biologique 6
1.4. Relation plantes -abeilles 8
Chapitre 2 : Milieu d'étude, Matériels et
méthodes 12
2.1. Milieu d'étude 12
2.1.1. Situation géographique 12
2.1.2. Climat 14
2.1.3. Végétation 15
2.2. Matériels et Méthodes 16
2.2.1. Collectes des spécimens 16
2.2.2. Piégeage aux coupelles ou piégeage passif
16
2.2.2.1. Principe 17
2.2.2.2. Description de l'équipement 17
2.2.2.3. Installation des pièges à coupelles 18
2.2.2.4. Relevé des pièges à coupelles 19
2.2.3. Piégeage au filet ou piégeage actif 19
2.2.4. Préparation des abeilles pour leur conservation et
identification 20
2.3. Analyse statistique 21
2.3.1. Evaluation de la biodiversité au niveau á
21
2.3.1.1. Les indices écologiques de composition 21
2.3.1.1.1. Richesse totale 21
2.3.1.1.2. Richesse moyenne 22
2.3.1.1.3. Abondance relative 22
2.4.1.1.4. Fréquence d'occurrence et constance 22
2.3.1.2. Les indices écologiques de structure 23
2.4.2. Distribution d'abondance 25
Chapitre 3 : Résultats 26
3.1. Faune des abeilles sauvages 26
3.2. Phénologie 27
3. 3.Evaluation de biodiversité par les indices
écologiques de Composition 29
3.3.1. Abondance relative 29
3.3.2. Richesse spécifique (totale) et moyenne. 31
3.3.3. Indice d'occurrence et fréquence 31
3.4. Evaluation de la biodiversité par les indices
écologiques de structure 32
3.4.1. Indices de Shannon -Weaver, indice de l'espérance
de Hurlbert et indice d'équitabilité de
Pielou. 32
3.4.2. Distribution d'abondance 33
3.2. Discussion 35
Conclusion et perspectives 38
Références bibliographiques 39
Annexes 45
Annexes
Annexe 1 : Distribution d'abondance de 2009 à
2010 par ordre décroissant.
|
Espèces
|
Nombre d'individus
|
Position
|
|
Megachile rufipennis
|
10
|
1
|
|
Amegilla sp
|
8
|
2
|
|
Xylocopa combusta
|
8
|
3
|
|
Xylocopa nigrita
|
5
|
4
|
|
Xylocopa calens
|
4
|
5
|
|
Xylocopa imitator
|
4
|
6
|
|
Halictus jucundus
|
3
|
7
|
|
Megachile bituberculata
|
3
|
8
|
|
Megachile eurymera
|
3
|
9
|
|
Megachile sp
|
3
|
10
|
|
Crocisapidia chandleri
|
2
|
11
|
|
Coelioxys sp
|
1
|
12
|
|
Euaspis erythros
|
1
|
13
|
|
Megachile cincta
|
1
|
14
|
|
Megachile sp2
|
1
|
15
|
|
Thyreus sp
|
1
|
16
|
|
Xylocopa albipes
|
1
|
17
|
Annexe 2 : Distribution d'abondance des abeilles sauvages
ajustée au modèle de Motomura de 2009-2010.
|
Espèces
|
Logni
|
position
|
|
Megachile rufipennis
|
1
|
0
|
|
Amegilla sp
|
0,90
|
1
|
|
Xylocopa combusta
|
0,90
|
2
|
|
Xylocopa nigrita
|
0,69
|
3
|
|
Xylocopa calens
|
0,6
|
4
|
|
Xylocopa imitator
|
0,6
|
5
|
|
Halictus jucundus
|
0,48
|
6
|
|
Megachile bituberculata
|
0,48
|
7
|
|
Megachile eurymera
|
0,48
|
8
|
|
Megachile sp
|
0,48
|
9
|
|
Crocisapidia chandleri
|
0,30
|
10
|
|
Coelioxys sp
|
0
|
11
|
|
Euaspis erythros
|
0
|
12
|
|
Megachile cincta
|
0
|
13
|
|
Megachile sp2
|
0
|
14
|
|
Thyreus sp
|
0
|
15
|
|
Xylocopa albipes
|
0
|
16
|
Annexe 3 : Indices de Shannon-Weaver (H') et
Equitabiité (E) basés sur les nombre des spécimens de
2009-2010.
|
Espèces
|
N ind.
|
Pi
|
ln pi
|
Log2pi
|
Pi*logPi
|
|
Amegilla sp
|
8
|
0,14
|
-1,96
|
-2,84
|
-0,40
|
|
Coelioxys sp
|
1
|
0,02
|
-3,91
|
-0,56
|
-0,11
|
|
Crocisapidia chandleri
|
2
|
0,03
|
-3,50
|
-5,05
|
-0,15
|
|
Euaspis erythros
|
1
|
0,02
|
-3,91
|
-0,56
|
-0,11
|
|
Halictus jacundus
|
3
|
0,05
|
-3,00
|
-0,43
|
- 0,22
|
|
Megachile bituberculata
|
3
|
0,05
|
-3,00
|
-0,43
|
- 0,22
|
|
Megachile cincta
|
1
|
0,02
|
-3,91
|
-0,56
|
-0,11
|
|
Megachile eurymera
|
3
|
0,05
|
-3,00
|
-0,43
|
- 0,22
|
|
Megachile rufipennis
|
10
|
0,17
|
-1,17
|
-2,56
|
-0,43
|
|
Megachile sp
|
3
|
0,05
|
-3,00
|
-0,43
|
- 0,22
|
|
Megachile sp2
|
1
|
0,02
|
-3,91
|
-0,56
|
-0,11
|
|
Thyreus sp
|
1
|
0,02
|
-3,91
|
-0,56
|
-0,11
|
|
Xylocopa albipes
|
1
|
0,02
|
-3,91
|
-0,56
|
-0,11
|
|
Xylocopa combusta
|
8
|
0,14
|
-1,96
|
-2,84
|
-0,40
|
|
Xylocopa callens
|
4
|
0,07
|
-2,66
|
-3,84
|
-0,27
|
|
Xylocopa imitator
|
4
|
0,07
|
-2,66
|
3,84
|
-0,27
|
|
Xylocopa nigrita
|
5
|
0,08
|
-2 ,53
|
-3,64
|
-0,29
|
|
Total
|
59
|
|
- pt Iog2pt
|
-3,71
|
|
Indice de shannon- weaver(H')
|
H'=3,71
|
|
|
|
|
Hmax
|
4,09
|
|
|
|
|
Equitabilité
|
0,91
|
|
|
|
Annexe 4 : indice de l'espérance d'Hulrbert
|
Espèces
|
Ni
|
N-ni
|
N-ni/N
|
(N-ni/N)^100
|
1-(N-ni/N)^100
|
|
Amegilla sp
|
8
|
51
|
0,86
|
2,82E-07
|
1,00E+00
|
|
Coelioxys sp
|
1
|
58
|
0,98
|
0,13
|
0,87
|
|
Crocisapidia chandleri
|
2
|
57
|
0 ,97
|
0,05
|
|
|
Euaspis erythros
|
1
|
58
|
0,98
|
0,13
|
0,87
|
|
Halictus jacundus
|
3
|
56
|
0,95
|
0,01
|
0,99
|
|
Megachile bituberculata
|
3
|
56
|
0,95
|
0,01
|
0,99
|
|
Megachile cincta
|
1
|
58
|
0,98
|
0,13
|
0,87
|
|
Megachile eurymera
|
4
|
55
|
0,93
|
0,00
|
1
|
|
Megachile rufipennis
|
10
|
49
|
0,83
|
8,09E-09
|
1,00E+00
|
|
Megachile sp
|
3
|
56
|
0,95
|
0,01
|
0,99
|
|
Megachile sp2
|
1
|
58
|
0,98
|
0,13
|
0,87
|
|
Thyreus sp
|
1
|
58
|
0,98
|
0,13
|
0,87
|
|
Xylocopa albicepes
|
1
|
58
|
0,98
|
0,13
|
0,87
|
|
Xylocopa combusta
|
8
|
51
|
0,86
|
2,82E-07
|
1,00E+00
|
|
Xylocopa callens
|
4
|
55
|
0,93
|
0,00
|
1
|
|
Xylocopa imitator
|
4
|
55
|
0,93
|
0,00
|
1
|
|
Xylocopa nigrita
|
4
|
55
|
0,93
|
0,00
|
1
|
Annexe 5 : Donnees climatiques de la station
meteorologique Metelsat de Binza (2000-2010).
|
Mois
|
Précipitations (mm)
|
Température (°C)
|
|
Janvier
|
170,5
|
24,9
|
|
Février
|
182,7
|
25,4
|
|
Mars
|
172,4
|
25,8
|
|
Avril
|
205,6
|
25,6
|
|
Mai
|
152,2
|
25,1
|
|
Juin
|
5,6
|
23,0
|
|
Juillet
|
0,7
|
22,3
|
|
Août
|
8,0
|
22,8
|
|
Septembre
|
53,3
|
24,4
|
|
Octobre
|
180
|
24,8
|
|
Novembre
|
251,7
|
24,7
|
|
Décembre
|
196,1
|
24,8
|
Annexe 6. Quelques abeilles inventoriees dans la station
d'etude en 2009-2010. (httv://
Zoologie.uinh.ac.be/hvinenovtera).
|
|
|
|
|
Amegilla sp.
|
Coelioxys sp.
|
Crocisapidia
chandleri
|
Halictus jucundus
|
|
|
|
|
|
Megachile
bituberculatau
|
Megachile sp.
|
Thyreus sp.
|
Xylocopa albiceps
|
|
|
|
|
Xylocopa calens
|
Xylocopa imitator
|
Annexe 7 : Listes des plantes sur lesquelles les abeilles
sauvages ont ete capturees.
|
Plantes
|
Familles
|
|
Asystasia gangetica
|
Acanthaceae
|
|
Boerhavia diffusa
|
|
|
Caesalpinia pulcherrina
|
Fabaceae
|
|
Chromolaena odorata
|
Asteraceae
|
|
Cyperus triangularus
|
Cyperaceae
|
|
Eleusina indica
|
Poaceae
|
|
Panicum maximum
|
Poaceae
|
|
Tithonia diversifolia
|
Asteraceae
|
|
Tridax procumbers
|
Asteraceae
|
|
Turnera ulmifolia
|
|
|
Urena lobata
|
Malvaceae
|


