|
_ _
Mémoire
Présenté par
BEN CHEIKH Fatma
Master Sciences et Technologies du Vivant et de
l'Environnement
Mention ERM
Spécialité « De l'Agronomie à
l'Agroécologie »
Recherche de facteurs explicatifs des écarts
à la relation négative rendement - teneur en protéines
chez le blé tendre (Triticum aestivum L.)
Pour l'obtention du diplôme de Master Sciences et
Technologies du Vivant et de l'Environnement
Enseignant responsable du stage : Alexandra
JULLIEN Maître de stage :
David GOUACHE
Soutenu le 28 juin 2012
Remerciements
Je tiens à remercier chaleureusement toutes les
personnes qui ont contribué à l'aboutissement de ce master. Tous
de près ou de loin, vous avez permis que cette belle aventure se passe
dans la joie et la bonne humeur.
Tout d'abord, mes remerciements
s'adressent à madame Alexandra Jullien, qui, sans elle je ne pourrais
jamais finir la rédaction de mon mémoire. Elle n'a pas
arrêté de m'encourager et me poussait à ne pas baisser les
bras jusqu'à la dernière minute.
Ensuite, je remercie monsieur Thierry Doré, notre
responsable de master, un homme que je respecte énormément. Je
n'oublierais jamais les précieux conseils que vous m'avez donnés
et vos encouragements durant toute l'année. Vos mots m'ont
été d'une aide importante, grâce à laquelle j'ai pu
parvenir à achever ce travail. Tout le respect est pour vous monsieur.
Je remercie également mon encadrant de stage
monsieur David Gouache pour son aide à l'établissement de ce
stage.
Un grand merci pour l'équipe d'Arvalis-Orsay. Merci
pour la bonne humeur qui a régné pendant les six mois que j'ai
passé avec vous. Tous mes remerciements s'adressent à vous :
Helena, Elise, Angeline, Guillaume, Delphine, Cécile, Valérie,
Flora, Katia.
Un spécial remerciement à Marion, stagiaire
à Arvalis-Orsay. Merci Marion pour tous les moments passés
ensemble, pour ta joie et la bonne humeur que tu dégages. Tu
étais toujours là pour moi essayant de me remonter le moral quand
rien ne va plus. Merci infiniment Marion : une petite pensée
à notre formation à Boigneville «living young and wild and
free ».
Je ne peux pas écrire ce remerciement sans penser à
mes amis de la maison de la Tunisie, ma nouvelle petite famille à moi.
Sans vous je ne pourrais jamais m'adapter à la vie parisienne aussi
vite. Merci Emira, Zid, Hajer, Kalthoum, Hamra, Amine, Karray, yassine, Tiki,
... les plus fous des fous !
Enfin je remercie chaleureusement mes très chers parents,
ma soeur, mon frère, ma belle-soeur toute ma famille et mes amis (Seima,
Nejla, Amal, Wafa) en Tunisie. Malgré la distance qui nous
séparait vous étiez tous là pour moi quand j'avais besoin
de vous, dans mes pires moments.
Table des matières
LISTE DES FIGURES
3
LISTE DES TABLEAUX
6
LISTE DES ABREVIATIONS
7
Introduction
9
Objectifs
10
Analyse bibliographique
11
1. Fonctionnement
azoté et élaboration de rendement et taux de
protéines
11
1.1. Métabolisme azoté dans la
plante
11
1.2. Effet de
l'azote sur l'élaboration du rendement
11
1.3. Effet de
l'azote sur l'élaboration du taux de protéines
12
2. Relation entre
le rendement et la teneur en protéines
13
2.1. Description de
la relation entre rendement et teneur en protéines
13
2.2. Explication du
paradoxe de la relation entre rendement et teneur en
protéines
14
2.2.1. Facteur
génétique
14
2.2.2.
Métabolisme azoté et métabolisme
carboné
14
2.2.3. Effet de
dilution de l'azote
15
3. Concept de grain
protein deviation « GPD »
15
3.1.
Définition et but du calcul de GPD
15
3.2. Variables
explicatives du GPD
16
3.2.1. Contribution
de la quantité d'azote absorbé en post-floraison dans
l'explication du GPD
3.2.2. Contribution
de la quantité d'azote remobilisée
3
Problématique
18
Matériel et méthodes
19
1. Types de
données manipulées
19
2. Différentes étapes de
traitement et nettoyage de données
20
3. Calcul du
GPD
20
4. Analyse
statistique
21
Résultats et discussion
22
1. Relation entre rendement en grain
et la teneur en protéines
22
2. Calcul des
résidus standardisés (le GPD) de la relation rendement - teneur
en protéines
23
3. Evaluation des
variables explicatives du GPD
24
4. La part de
l'azote absorbé en post-floraison dans l'explication du GPD
25
5. Evaluation des
variables agro-climatiques
28
Discussion
30
1. Relation teneur en protéines -
rendement en grain
30
2. Relation entre le GPD et les
différents métabolismes physiologiques
31
Conclusion
34
Annexe1: Ensemble des variables de la base de
données
35
Références bibliographiques
38
LISTE DES FIGURES
Figure 1:
Relation entre le rendement (t/ha) et quantité d'azote absorbée
(kg N /ha) (les différentes couleurs des triangles expliquent les doses
d'azote) (Barraclough et al., 2010)
3
Figure 2:
Déterminisme du poids des grains et sa composition: A,
métabolisme azoté dans le grain; B: production des assimilats de
carbone; C: accumulation de l'amidon (Triboi et Triboi-Blondel, 2002)
13
Figure 3:
Relation entre le rendement en grain et la teneur en protéines pour 27
variétés dans 27 milieux différents (Bogard, 2011)
14
Figure 4:
Processus physiologique affectant le GPD chez le blé (Bogard, 2011)
16
Figure 5:
Relation entre absorption post-foliaire de l'azote et le GPD (Anonyme 3)
17
Figure 1:
Résumé des différentes étapes de traitement et
nettoyage de données
20
Figure 2:
Relation entre la teneur en protéines et le rendement pour les quatre
variété: Apache, Caphorn, Charger et Soissons
22
Figure 3:
Boxplot montrant les GPDs selon les variétés
23
Figure 4:
Relation entre GPD et l'INN floraison pour chacune des variétés
(la distribution des points est faite selon les années des campagnes
agricoles)
25
Figure 5:
Relation entre l'écart à la relation
PANU-QNremobilisée et le GPD pour les quatre
variétés Apache, Caphorn , Charger et Soissons (la distribution
des points est faite selon les années)
26
Figure 6:
Relation entre le GPD et l'ecart à la relation PANU-QNremobilisée
expliquée par l'INNfloraison pour les quatre variétés.
27
Figure 7:
Relation entre la consommation du carbone pour la production des semences pour
plusieurs espèces et leurs rendements (afin de comparer la performance
de production plusieurs courbes d'iso production ont été
utilisées)
31
Figure 8:
représentation simplifiée de module post-floraison du
modèle Azodyn: accumulation de biomasse et d'azote dans les grains de
blé (Jeuffroy et al., 2000)
32
LISTE DES TABLEAUX
Tableau 1:
Résumé des caractéristiques des sites
3
Tableau 2:
Pentes et coefficients de détermination de la relation rendement -
teneur en protéines pour les quatre variétés
23
Tableau 3:
Coefficients de détermination et Pvalue (á=5%) de la
corrélation entre le GPD et la quantité d'azote accumulée
en floraison (QNfloraison), indice de nutrition azotée
à floraison (INNfloraison), quantité d'azote
absorbée en post-floraison (PANU), quantité d'azote
remobilisée (QNremobiliseé) et Ecart PANU-QNR
24
Tableau
4:Coefficient de determination et Pvalue de la
relation entre l'ecart PANU-QNremobilisée et le Poids mille grain (PMG)
et la biomasse de remplissage (BMremplissage)
28
Tableau 5:
Pvalue des différentes relations entre la quantité d'azote
remobilisée (QNremobilisée), la quantité
d'azote absorbée en post-floraison (PANU), l'ecart à la relation
PANU-QNremobilisée (ecPANU.QNR) et la biomasse de remplissage
(BMremplissage) et les facteurs climatiques.
29
LISTE DES ABREVIATIONS
PANU: quantité d'azote absorbée en
post-floraison
QNremobilisée: quantité d'azote
remobilisée
QNfloraison: quantité d'azote
absorbée en pré-floraison
GPD: Grain Protein Deviation
BMremplissage: biomasse de remplissage
ecPANU-QNremobilisée: écart
à la relation PANU - QNremobilisée
Résumé
Chez le blé tendre (Tritiucum aestivum L.) le
rendement en grain et la teneur en protéines sont les majeurs
critères de la production contribuant à la valeur
économique de cette culture. Producteurs et sélectionneurs
cherchent constamment à concilier entre ces deux critères.
Toutefois, ceci n'est pas toujours évident à cause de la relation
négative entre rendement et teneur en protéines.
Il a été montré que l'évaluation
des résidus de la relation rendement - teneur en protéines (ou ce
qu'on appelle GPD pour Grain Protein Deviation) est un bon moyen aidant
à modifier cette relation négative. De plus on a montré
que ce GPD est lié à la variabilité
génétique de l'absorption d'azote post-floraison, une variable
importante dans l'explication de GPD.
Au cours de ce travail, à partir des analyses de
données, nous avons cherché à expliquer la variation de
GPD dans des conditions pédoclimatique différentes via des
processus de l'absorption azotée qui sont: la quantité d'azote
absorbé en préfloraison(QNfloraison), la
quantité d'azote absorbé en post-floraison (PANU), la
quantité d'azote remobilisée (QNremobilisée).
Au vu de ce travail nous pouvons conclure que le
QNfloraison est la variable qui explique au mieux la variation de
GPD. La QNremobilisée et le PANU ont aussi un effet
significatif sur le GPD mais ils sont influencés par les facteurs
climatiques tels que la température élevée et le
déficit hydrique.
Abstract
In bread wheat (Tritiucum aestivum L.) grain yield (GY) and
grain protein concentration (GPC) are the major criteria of production as they
contribute to the economic value of wheat. Producers and breeders constantly
seek to reconcile these two criteria. However, the negative GPC- GY
relationship makes it hard.
It was shown that the grain protein deviation (GPD) was a good
way to shift this negative relationship. Moreover it was shown that GPD is
linked to the genetic variability of post anthesis nitrogen uptake.
During this work, we tried to explain the variation of GPD in
different environment through the process of nitrogen absorption. These
processes are the amount of nitrogen absorbed at anthesis, of post anthesis
nitrogen uptake (PANU), the amount of nitrogen remobilized.
In view of the work we can conclude that the Nitrogen stored
at anthesis is the variable that explains better the GPD. Nitrogen remobilized
and PANU have a significant effect on GPD but they are influenced by climatic
factors such as high temperature and water deficit.
Introduction
La production de blé tendre « Titicum
aestivum L. » représente la quatrième culture la
plus importante du monde en tonnes après la canne à sucre, le
mais et le riz. La production mondiale en 2010 a atteint les 650.881.002 tonnes
pour une superficie cultivée de l'ordre de 216.974.683 ha (Faostat,
2012).
La nutrition azotée est un facteur clé de
maîtrise de la production de blé. Elle influence le fonctionnement
du couvert dès le tallage herbacé avec des effets dominants
à partir du stade épi 1 cm jusqu'à la floraison. La teneur
en azote régule en effet la croissance foliaire et donc la production de
biomasse. Cela se traduit par des effets visibles sur les composantes du
rendement : en particulier, Nombre d'épi/m², nombre de grain
/épi. De ce fait une déficience en azote aura une incidence
très pénalisante sur le rendement. D'un autre côté,
l'excès d'azote peut provoquer une augmentation du niveau de verse en
diminuant le rapport C/N ou bien augmenter la sensibilité à la
sécheresse surtout dans des conditions de limitation de la
disponibilité d'eau.
La plante absorbe l'azote sous ses deux formes nitrique
(NO3-) et ammoniacale (NH4+). Ces
composants, après assimilation sont utilisés dans la construction
des tissus photosynthétiques contenant des protéines de
synthèse (essentiellement la RUBISCO) et des protéines de
structures. (Pask et al., 2011). L'azote est donc un
élément qui a une influence directe sur la teneur en
protéines. Il entre dans la composition des acides nucléiques
(ADN et ARN), des acides aminés qui composent les protéines (75%)
d'azote est stocké sous forme de protéines) nécessaires
aux mécanismes métaboliques dont dépendent la croissance
et le développement.
De nombreuses études ont montré une relation
négative qui existe entre le rendement et la teneur en protéines.
Ces deux variables résultent de l'intégration de processus en
rapport à la fois avec métabolisme du carbone et de l'azote. Le
rendement pour sa part dépend des facteurs déterminants
l'assimilation photosynthétique, le stockage et la remobilisation des
hydrates de carbone. La teneur en protéines d'autre part dépend
de la quantité d'azote absorbée et remobilisée vers le
grain. Diriger la production pour répondre à ces deux facteurs
demeure une tâche délicate à cause de cette relation
négative.
De nombreux travaux ont eu pour but d'améliorer
conjointement le rendement et la teneur en protéines. A cette fin, on a
deux leviers. Un levier agronomique jouant sur la conduite de la fertilisation
azotée en optimisant l'efficience de l'utilisation des engrais
azotés et un levier génétique par l'amélioration
des plantes et l'étude de la variabilité génétique
existante. La plupart des études qui portaient sur le levier agronomique
montrent l'importance de l'absorption post-floraison et l'importance de sa
valorisation. D'une autre part, peu de travaux ont porté sur
l'étude du levier génétique (Oury et Godin, 2007 ;
Bogard, 2011 ; Bogard et al., 2010 ; LE Gouis et
al., ; Monaghan et al., 2001). Ces travaux ont conduit
à l'utilisation du « GPD » pour Grain Protein
Deviation. C'est l'écart à la relation teneur en protéines
- rendement en grain. Le GPD est calculé comme les résidus de la
régression entre la teneur en protéines et le rendement. Le GPD a
été étudié dans le but d'identifier les
variétés qui ont des teneurs en protéines
supérieures à celle prédite par la relation. Le GPD est
affecté par quelques facteurs physiologiques tels que la remobilisation
de l'azote accumulé pendant la floraison (Barbottin et al.,
2004) et l'absorption post-floraison (Bogard, 2011). Toutefois le rôle
que peut jouer le pédoclimat sur le GPD via ces processus n'a pas
été étudié en tant que tel. En effet le climat agit
sur tous les paramètres qui conditionnent la relation teneur en
protéines - rendement. La température et la disponibilité
en eau affectent beaucoup les conditions de la disponibilité et
d'absorption d'azote.
Le but de ce travail sera de pouvoir comprendre les effets
pédoclimatiques sur la relation teneur en protéines - rendement
en grain et voir le fonctionnement des paramètres physiologiques :
absorption d'azote en floraison, absorption d'azote en post-floraison,
remobilisation de l'azote, qui sont en relation avec celle-ci.
Objectifs
ï Calcul du GPD pour caractériser la
variabilité interannuelle du climat et multilocale affectant la
relation rendement - teneur en protéines
ï A l'instar du travail fait par Mattieu Bogard (2011)
dans le cadre d'une thèse qui porte sur l'explication du GPD via la
variabilité génétique, vérifier s'il est possible
d'expliquer les variations interannuelles et multilocales de GPD par un surplus
d'absorption d'azote en post-floraison.
ï Rechercher les variables agroclimatiques
corrélées au GPD et/ou au surplus d'absorption post-floraison.
Analyse bibliographique
1. Fonctionnement
azoté et élaboration de rendement et taux de
protéines
1.1. Métabolisme azotée dans la plante
L'azote est un élément majeur de la nutrition
chez le blé. L'introduction des variétés
« semi-naines » a permis d'accroître les
quantités d'azote valorisables par un couvert de blé (Barraclough
et al., 2010). L'azote agit sur la croissance de l'appareil
végétatif, en contrôlant la taille des feuilles et en
jouant sur le devenir des talles. Il influence également fortement le
nombre d'épillets fertiles. Lorsque le statut azoté de la culture
passe au-dessous du seuil défini comme l'état permettant la
croissance maximale de la culture, celle -ci est en carence. Cette carence
azotée agit sur les fonctions de croissance en réduisant
l'accumulation de biomasse, l'indice foliaire, son expansion ou en
accélérant le processus de sénescence. La contrainte
azotée peut également affecter l'efficience de conversion de
l'énergie lumineuse (Jeuffroy et Recous, 1999).
L'azote est absorbé par les racines sous ses deux
formes ; nitrique (NO3-) et ammoniacale
(NH4+). L'absorption se fait grâce à des
transporteurs au niveau de la racine (NRT: nitrate transporter et AMT: ammonium
transporter). Dans la plante l'ion nitrate prend deux chemins Il subit une
réduction dans les feuilles pour enfin aboutir à l'ion
NH4+. Cependant, dans le cas de faible besoin en azote il
est stocké dans les vacuoles pour constituer un pool de réserve.
(Gate, 1995). De l'autre coté l'ammonium est assimilé dans les
racines. Les produits de l'assimilation de l'azote sont des acides organiques
et acides aminés. (Morot-Gaudry, 1997) L'assimilation de l'azote se fait
à l'aide de deux enzymes le GS (glutamine synthétase) et la GOGAT
(glutamate synthase). Ces deux enzymes intègrent l'ammonium dans une
molécule organique qui sera à l'origine de la formation de deux
acides aminés: la glutamine et le glutamate. Des protéines seront
par la suite formées grâce à des acides aminés
dérivant du glutamine et le glutamate (Masclaux et al., 2001
cité par Ben Slimane, 2010).
1.2. Effet de l'azote sur
l'élaboration du rendement
L'élaboration du rendement dépend de
l'absorption azotée. Cette absorption se fait selon la
disponibilité de l'élément dans le sol et est fonction des
besoins de la plante. En effet, les besoins de la culture varient selon les
stades de croissance. On a montré que les plus forts besoins sont entre
la phase d'épi 1 cm et la floraison. Un déficit azoté
pendant cette période affecte le rendement. Au cours de la montaison,
l'azote agit sur la montée à épis des talles
herbacées, la croissance des feuilles et l'élongation des
entre-noeuds. De plus, une carence en azote dans la phase de montaison
altère le nombre de grains (Jeuffroy et Recous, 1999). L'accumulation de
biomasse dans les grains est le résultat de la confrontation entre la
demande des grains en biomasse et l'offre en assimilats carbonés au
niveau de la culture. La quantité de biomasse disponible pour le
remplissage des grains est dépendante de la production de biomasse par
la culture après la floraison (source majoritaire des assimilats
carbonés pour les grains) et d'une fraction fixe de la biomasse à
floraison, correspondant à la remobilisation des sucres solubles
stockés dans les tiges avant ce stade (O'Leary et al., 1985
cité par Barbottin, 2004).
La nutrition azotée intervient donc dès le
tallage avec des effets dominants à partir du stade épi 1 cm
jusqu'à la floraison sur les composantes du rendement : nombre
d'épis/m² et nombre de grain (Gate, 1995).
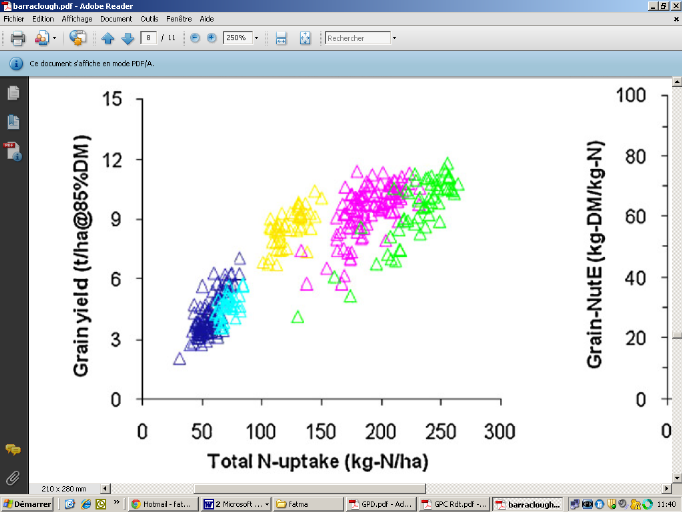
Figure 1: Relation entre le rendement (t/ha)
et quantité d'azote absorbée (kg N /ha) (les différentes
couleurs des triangles expliquent les doses d'azote) (Barraclough et
al., 2010)
1.3. Effet de l'azote sur
l'élaboration du taux de protéines
L'azote assimilé disponible au niveau des grains est
essentiellement utilisé pour la synthèse des protéines qui
seront accumulées et stockées dans l'endosperme. (Triboi et
Triboi-Blondel, 2002) L'azote est stocké dans le grain sous forme de
protéines. Le stockage se fait essentiellement au cours de la
période post-floraison grâce à la remobilisation de l'azote
des parties végétatives en cours de sénescence. (Shewry,
2007). Il faut noter que l'accumulation d'azote dépend de la
disponibilité en azote au niveau de la plante. (Barbottin, 2004) On a
montré que même un faible apport en azote dans la phase
post-floraison affecte positivement la teneur en protéine (Gate, 1995).
Trois modalités de fertilisation azotée ont une
influence sur la teneur en protéines :
- la dose apportée qui avec laquelle
la teneur en protéine augmente.
- la forme d'azote utilisée où
on constate que la forme nitrate permet une augmentation du
taux de protéines comparée aux formes
ammoniaco-uréiques.
- le fractionnement des apports, surtout en
appliquant un apport tardif qui permet d'augmenter la teneur en
protéines les apports tardifs grâces à une
meilleure utilisation de l'azote par le blé, permettent d'augmenter la
teneur en protéines (Anonyme 1, 2011).
La figure (2) est un diagramme séparant les deux
mécanismes azoté et carboné pour mieux comprendre leur
interaction et leur impact sur l'établissement de rendement et la teneur
en protéines.
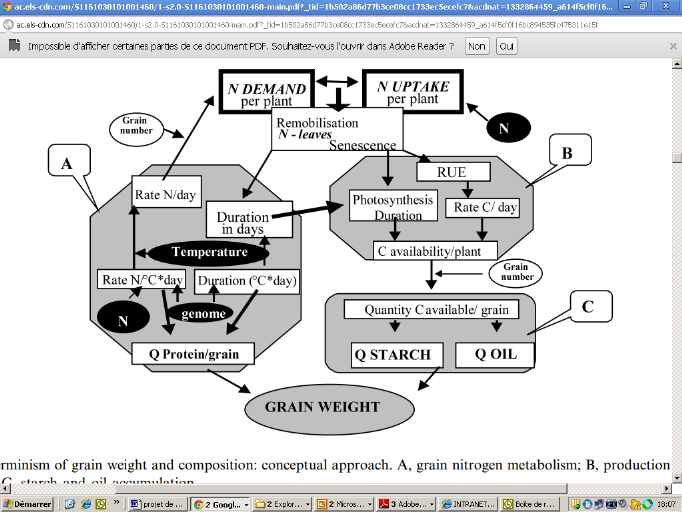
Figure 2: Déterminisme du poids des
grains et sa composition: A, métabolisme azoté dans le grain; B:
production des assimilats de carbone; C: accumulation de l'amidon (Triboi et
Triboi-Blondel, 2002)
2. Relation entre le
rendement et la teneur en protéines
2.1. Description de la
relation entre rendement et teneur en protéines
Les deux critères : rendement en grain et la
teneur en protéines étaient l'objet de plusieurs recherches.
Ainsi la relation entre ces deux variables était bien
étudiée. La majorité des études montrent qu'entre
le rendement et la teneur en protéines la relation est plutôt
négative. (Monaghan et al, 2001, Oury et Godin, 2007, LeGouis et
al.,2010). Ces résultats ont été confirmés
récemment par l'étude de Bogard (2011). Il a trouvé une
relation négative très significative entre les deux
critères (
Figure 3)
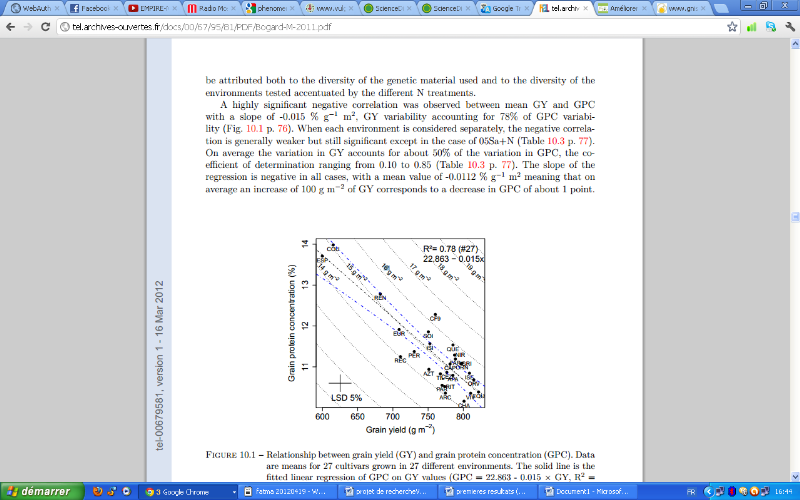
Figure 3:
Relation entre le rendement en grain et la teneur en protéines
pour 27 variétés dans 27 milieux différents (Bogard,
2011)
2.2. Explication du
paradoxe de la relation entre rendement et teneur en
protéines
2.2.1. Facteur
génétique
Une première étude était faite dans le
cadre d'expliquer la relation entre le rendement et la teneur en
protéines par rapport aux variétés. La comparaison
était faite entre des variétés anciennes et des
variétés récentes dites
« performantes ». Les résultats étaient que
les variétés anciennes ont un taux de protéines
élevé mais à faible rendement. Tandis que les
variétés récentes affichent un bon rendement contre une
teneur faible en protéines (Barraclough et al., 2010). Ceci met
en évidence la relation négative qui existe entre les deux
variables.
Dans le but d'expliquer le paradoxe de la relation, certains
auteurs Pepe et Robert, 1975 ; Miezen et al., 1977, McNeal et Berg, 1978
(cité par Bogard, 2011) ont trouvé que c'est le facteur
génétique est responsable de cette relation. Ils ont
proposé que la nature de la relation soit due essentiellement aux
facteurs génétiques par la présence des gènes
à effet pléiotropes affectant les deux caractères ou bien
par la présence des gènes à effet antagoniste fortement
liés génétiquement.
2.2.2. Métabolisme
azoté et métabolisme carboné
A côté du facteur génétique, une
forte hypothèse concernant la compétition entre les
métabolismes azotés et carbonés peut expliquer cette
relation négative (bhatia et rabson, 1976 cité par Bogard 2011).
On a montré qu'il existe une étroite interaction
entre ces deux métabolismes. L'azote est remobilisé des organes
végétatifs vers les grains via le processus de
sénescence : par conséquent, la perte d'azote dans les
organes végétatifs contribue à réduire l'indice
foliaire et l'efficience de conversion du rayonnement en biomasse. Le
remplissage des grains en biomasse, qui dépend de l'interception
lumineuse pilotée par l'indice foliaire et de son efficience de
conversion, est donc directement impacté par cette remobilisation
(Jeuffroy et al., 2000 ).
La nature de la relation entre le rendement et la teneur en
protéines résulte essentiellement des interconnexions entre
métabolismes carbonés et azotés (Bogard, 2011). Ces deux
variables résultent de l'intégration de processus en rapport
à la fois avec les métabolismes du carbone et de l'azote. Le
rapport C/N du grain étant largement en faveur du carbone, le rendement
en grains dépend en premier lieu des facteurs déterminant
l'assimilation photosynthétique, le stockage et la remobilisation des
hydrates de carbone. Pour la teneur en protéines, cela dépend
fortement de la quantité d'azote absorbée et remobilisée
vers le grain (Bogard, 2011).
La teneur en protéines des grains dépend de la
richesse de la plante en acides aminés durant la période
post-floraison. Les acides aminés, pendant la méiose, permettent
la formation des protéines de structure et métabolique, qui, en
phase de remplissage deviennent des protéines de réserve. (Martre
et al., 2003 cité par Bertheloot, (2009). Le
développement des grains nécessite la réalisation d'un
mécanisme dont le processus permet le recyclage des protéines des
organes végétatifs via leur dégradation en acides
aminés et leur transfert vers les organes en croissance (les grains en
post-floraison): c'est la remobilisation (Hirel et al., 2007
cité par Ben Slimane, 2010). Un nombre élevé de grain
accélère la remobilisation de l'azote engendrant une
accélération de la sénescence pendant le remplissage et
qui par conséquent affecte la photosynthèse. (Bogard et
al., 2011).
2.2.3. Effet de dilution
de l'azote
Selon Przulj et Momcilovic (2001) la concentration en
protéines dans le grain ne dépend pas seulement de la
quantité d'azote dans le grain mais aussi de la concentration en
carbohydrates.
On a montré que cet effet antagoniste entre les deux
caractères de la production est essentiellement dû à un
phénomène de dilution de l'azote absorbé au profit du
carbone
Slafer et al. 1990 cité par Monaghan et
al (2001) ont montré que l'augmentation de la teneur en
protéines, peut se faire par une accumulation plus importante de l'azote
absorbé en préfloraison combinée avec une forte
remobilisation de l'azote des parties végétatives vers les
grains.
3. Concept de grain protein
deviation « GPD »
3.1. Définition et but
du calcul de GPD
Le GPD a été étudié pour la
première fois par Mongham et al (2001) en Angleterre dans le but de
mesurer la capacité des variétés de blé tendre
d'atteindre un taux de protéines meilleur que prévu par rapport
un rendement donné.
Pour évaluer la relation négative reliant le
rendement et la teneur en protéine, on a proposé d'utiliser la
déviation à partir de la régression linéaire entre
le rendement et la teneur en protéine ; c'est ce qu'on appelle
« la déviation grain protéine » :
« Grain Protein deviation » (Anonyme2, Monaghan et al.,
2001, Oury et Goudin, 2007, LeGouis et al, 2010, ...).
Le GPD étant définie comme étant le
« résidus standardisés » de la
régression linéaire du rendement en grain (t/ha) et la teneur en
protéines (%) (Oury et Godin, 2007 ; Monghan et
al., 2001 ).
Oury et Goudin (2007) ont proposé d'utiliser une sorte
d'algorithme d'ajustement dans le but d'éliminer les outliers et
évaluer la vraie position de la régression linéaire pour
un ensemble de données. L'algorithme fonctionne en éliminant tous
les points dont les résidus sont supérieurs à |1.96|.
L'itération est stoppée quand on a plus de valeur en dessus de la
valeur seuil
Dans leur étude Oury et Godin (2007) ont utilisé
la formule (1) pour calculer les résidus de la
régression :
GPD=Sd= ri / ^s*v (1-hi) (1);
^s est la déviation résiduelle
standard ; ^s=v((1/n-p)*?ri²)
Avec : ri= résidus des
observations, p=2, n=nombre des observations,
hi=la valeur de la diagonale de la matrice.
En effet, étant donné que les GPD sont des
résidus standardisés donc, par définition, dans la
condition d'une distribution normale, la moyenne de GPD est nulle et 95% des
valeurs de GPD standardisées sont comprises entre -1.96 et +1.96. Cette
dernière valeur correspond au quantile d'ordre 2.5% pour P=0.975.
L'une des méthodes utilisées pour obtenir le GPD
est d'utiliser un algorithme pour éliminer tous les points qui ont des
résidus supérieur à |1.96| l'itération est
stoppée quand on a plus de valeur en dessus de la valeur seuil. Le but
de cette méthode est de neutraliser l'effet des données
aberrantes. On aura donc une régression linéaire précise
et qui représente tous les points (Oury et Godin, 2007).
3.2. Variables explicatives
du GPD
Afin de décomposer le GPD et d'identifier les
caractères associés, la méthode d'analyse a
consisté à rechercher les corrélations permettant
d'établir un lien entre ce caractère et d'autres variables. Pour
cela, les coefficients de corrélation des indicateurs statistiques
jugeant de la force de l'association entre deux variables ont été
analysés (Bogard et al., 2011).
Monaghan et al. (2001) ont utilisé la méthode de
« step-wise multiple régression » qui consiste
à ajouter des variables pour voir leur contribution dans l'explication
du GPD. Ils ont constaté que l'accumulation de l'azote post-floraison et
la quantité d'azote remobilisée sont les variables les plus
importantes qui ont une influence sur le GPD.
3.2.1. Contribution de la
quantité d'azote absorbé en post-floraison dans l'explication du
GPD
Le Gouis et al ont montré qu'il y a une
corrélation entre la quantité d'azote remobilisé et la
quantité d'azote absorbé post-floraison avec l'écart
à la relation rendement teneur en protéines. Cependant, la
corrélation n'est pas significative entre la quantité d'azote
remobilisée et le GPD. La corrélation est par contre
significative avec la quantité d'azote absorbé en post-floraison
qui explique environ 50 % du GPD (Le Gouis et al., 2010).
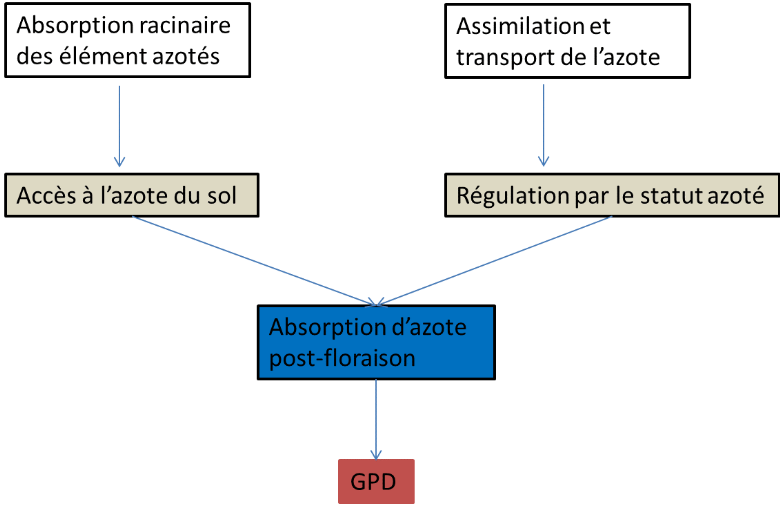
Figure 4: Processus physiologique affectant
le GPD chez le blé (Bogard, 2011)
Bogard (2011) en étudiant la relation entre GPD et
l'azote absorbé pendant la période post-floraison a trouvé
que ces deux variables sont très bien corrélés
indépendamment de la date de floraison (figure 6). Monaghan et
al (2001) ont trouvé que l'explication de l'importance du PANU
dans la détermination du GPD est liée à la
différence de la répartition de l'azote accumulé en
floraison et après floraison
L'absorption post- floraison, bien que relativement faible,
est une source significative d'azote pendant la phase de remplissage (Le Gouis
et al., 2010) mais elle est très variable. Elle dépend
du site, du type de sol notamment de sa réserve d'azote disponible
à la plante ainsi que de la réserve utile.
Sur plantes attaquées par une maladie foliaire on peut
observer l'importance de l'absorption azotée post-foliaire. Bancal
et al., (2008) ont observé que l'accumulation d'azote dans les
parties aériennes s'arrête plus tard dans des parcelles
traitées aux fongicides que dans celles où les plantes sont
attaquées par une maladie fongique. De façon assez
générale, le bilan d'azote entre floraison et maturité
montre que l'absorption de l'azote post-floraison est significativement
réduite chez les plantes malades avec un effet significatif de
l'année, donc de la sévérité des maladies et
souvent aussi de la fertilisation azotée. La diminution de l'absorption
semble donc fonction de la précocité et de la
sévérité de l'infection. Leitch et Jenkings (1995) ont
observé une réduction de 11% de l'absorption post-floraison dans
le cas d'une infestation par la Septoriose foliaire, mais sans effet de la
fertilisation azotée (Ben Slimane, 2010)
D'autre part l'étude des variétés
« stay-green » de sorgho met en évidence
l'importance de l'absorption azotée post-floraison. Les
« stay-green » sont des variétés qui ont une
activité photosynthétique plus longue même sous des
conditions hydriques limitantes. Ces variétés sont
caractérisées par une concentration d'azote dans les
différents compartiments toujours élevée. En effet, cette
expression de « stay-green » est la conséquence de
l'équilibre entre la demande en azote des grains et la réserve de
l'azote durant le remplissage (Borell et Hammer, 2000). Borell et Hammer
(2000), pour une étude sur le sorgho ont montré que la
concentration de l'azote dans les différents compartiments de la plante
est toujours plus élevée que chez les autres
variétés.
GPD
251657728
Absorption post-floraison N (g/m²)
251656704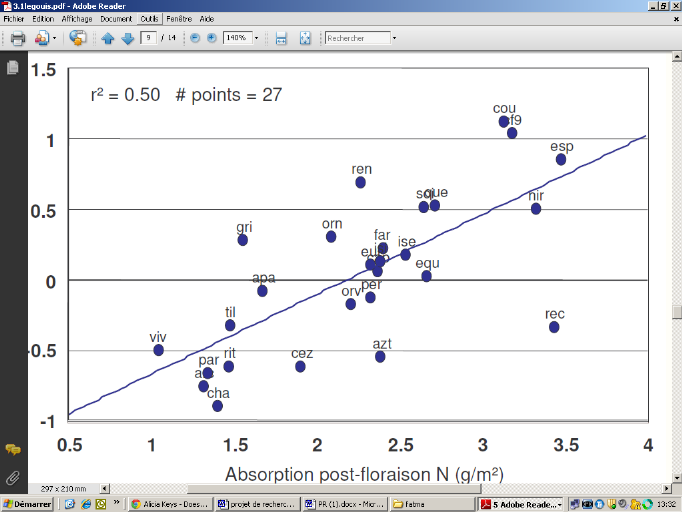
Figure 5: Relation entre absorption
post-foliaire de l'azote et le GPD (Anonyme 3)
3.2.2. Contribution de la
quantité d'azote remobilisée
L'azote remobilisé est définie comme
étant la quantité d'azote mesurée en floraison et qui n'a
pas été récupérée dans les pailles à
la récolte Pask et al (2011). L'étude de cette portion
d'azote est intéressante puisque elle constitue la majeure source de la
présence de l'azote dans le grain lors du remplissage. Elle
représente 60 à 90% de l'azote des grains (Pask et al.,
2011 ; Barbottin, 2005).
La remobilisation de l'azote dépend fortement de la
quantité de l'azote absorbé par la culture pendant la phase de
floraison. En effet ; la quantité d'azote remobilisée est
d'autant plus importante que la quantité d'azote absorbé en
floraison est importante. Toutefois, la remobilisation de l'azote des parties
végétatives vers les grains dépend de plusieurs facteurs
environnementaux (stress hydrique et stress thermique) au cours de la
période de remplissage Elle dépend aussi de l'état
sanitaire des plantes : en cas de maladies foliaires, la quantité
d'azote remobilisée sera fortement affectée. De plus on a
indiqué que la remobilisation de l'azote est indépendante du
facteur génétique dans le cas où il n'y a aucun facteur
environnemental limitant (Barbottin et al., 2005).
Une étude sur l'orge a montré les mêmes
résultats obtenus sur le blé tendre et on a montré le
rôle que joue la remobilisation de l'azote sur le rendement. En effet, le
rendement est d'autant plus élevée que la quantité d'azote
remobilisée est importante. (Przulj et Momcilovic, 2001).
Ces études mettent en avant l'importance de la
remobilisation de l'azote comme un caractère pouvant expliquer le GPD.
Problématique
Le GPD exprime la déviation à la relation teneur
en protéines et le rendement. Ces deux variables dépendent
énormément de l'élément azote dans la plante. Parmi
les majeures sources d'accumulation d'azote dans le grain, on trouve la
remobilisation de l'azote accumulé au cours de la phase
préfloraison et l'absorption de l'azote en post-floraison. Il est donc
intéressant de voir les effets des facteurs pédoclimatiques qui
agissent sur ces processus. Pask et al (2011) ont montré que
parmi les facteurs qui ont une influence sur l'absorption post-floraison, on
trouve : le type de sol, la disponibilité de l'azote minéral
et la réserve en eau du sol.
De plus, l'absorption post-floraison dépend de la
quantité d'azote fournie par le sol pendant le remplissage. Les
fournitures du sol correspondent soit au « reste du dernier apport»
soit au produit de la minéralisation. Concernant la première
hypothèse, mis à part des situations exceptionnelles de carence
induite et de sécheresse ou d'accidents ayant fortement
pénalisé la croissance, la demande de la plante est telle qu'on
peut supposer que les quantités restantes sont
généralement négligeables. La minéralisation
dépend quant à elle à la fois du climat et des
caractéristiques du sol. Les caractéristiques du sol fixent la
vitesse potentielle de minéralisation. Celle-ci est ensuite
affectée par le climat, qui est plus ou moins favorable à la
réalisation de cette vitesse maximale (Comifer, 2011). L'effet du climat
se calcule via la prise en compte les conditions de température et
d'humidité de l'horizon minéralisant : c'est le concept de
jours normalisés.
D'un autre côté, la capacité d'absorption
est liée au maintien des surfaces vertes. En effet, une rapide
sénescence causée par une forte remobilisation de l'azote peut
limiter la capacité d'absorption d'azote pendant le remplissage (Mi et
al., 2000). La cinétique de la sénescence est fortement
dépendante des conditions environnementales telles que la
disponibilité de l'eau, la température ou bien une carence
azotée précoce.
Dans sa thèse, Bogard (2011) a montré que la
variabilité génétique du GPD suit une variabilité
génétique de l'absorption en post-floraison. Si on se met dans
différents contextes agro-climatiques, cette variable serait-elle
toujours aussi importante dans l'explication de GPD ? D'autre part,
faut-il prendre en considération d'autres variables du processus
azoté telles que l'azote absorbé en préfloraison et
l'azote remobilisé des parties végétatives vers les grains
pour expliquer le GPD ?
Dans le cas où ces variables ont une influence sur la
variation de GPD, il serait alors intéressant d'étudier leur
variation selon les variations pédoclimatiques. Dans quelles conditions
pédoclimatiques les variables explicatives du GPD sont
valorisées ?
Le travail va donc se dérouler en plusieurs
étapes. La première consiste à calculer le GPD. La seconde
demande de vérifier quelle proportion des variations
pédoclimatiques du GPD est expliquée par l'absorption
post-floraison ou par la quantité d'azote remobilisée. Enfin, on
cherchera, par analyse de corrélations, à relier les variations
de GPD ou d'azote absorbé post-floraison ou d'azote
remobilisé à des indicateurs agro-climatiques
représentant les effets du climat sur les processus de fournitures
d'azote ou de capacité d'absorption de la plante. Les indicateurs
agro-climatiques sont en l'occurrence : le déficit hydrique
(ETR/ETM), somme de température > 25°, somme de rayonnement
global, ...
Matériel et
méthodes
1. Types de données
manipulées
Les données utilisées au cours de ce stage sont
des données tirées des essais en micro-parcelles. Il s'agit d'un
large réseau, représentant donc une grande diversité de
pédo-climats, d'essais qui servent à l'équipe
« Physiologie » comme observatoire de la croissance et de
l'élaboration du rendement et de la qualité. Y sont ainsi
mesurées des biomasses et des teneurs en azote, permettant de calculer
des quantités d'azote absorbées. Par contre, il n'existe pas de
mesure systématique des variables du sol qui permettraient
d'accéder directement ou même par calcul aux variables de
fournitures d'azote par le sol. Ces essais sont généralement
conduits proches de l'optimum technique recommandé régionalement.
Sont également intégrés dans ce jeu de données des
essais sur la thématique de la fertilisation azotée, mettant en
jeu des variations de pratiques de fertilisation parfois très
importantes. Le jeu de données comporte 159 variables et 373
observations. Dans ce jeu de données on a testé quatre
variétés de blé tendre cultivées dans des parcelles
réparties sur 16 régions et ayant 7 types de sol
différents (
Tableau 1).
Tableau 1: Résumé des
caractéristiques des sites
|
Régions
|
Type de sol
|
Variété
|
|
Auvergne
|
2
|
Argile
|
4
|
Apache
|
76
|
|
Bourgogne
|
26
|
Argilo-calcaire profond
|
40
|
Caphorn
|
85
|
|
Bretagne
|
2
|
Calcaire superficiel
|
34
|
Charger
|
50
|
|
Centre
|
23
|
craie de champagne
|
1
|
Soissons
|
162
|
|
Centre BP
|
90
|
Graviers
|
14
|
|
|
|
Centre Est
|
40
|
Limon
|
155
|
|
|
|
Ile de France
|
31
|
Limoneux-Argileux
|
125
|
|
|
|
Midi-pyrenées
|
2
|
|
|
|
|
|
Nord
|
28
|
|
|
|
|
|
Nord Est
|
4
|
|
|
|
|
|
Normandie
|
10
|
|
|
|
|
|
Ouest
|
54
|
|
|
|
|
|
Pays de la Loire
|
10
|
|
|
|
|
|
Poitou-Charentes
|
11
|
|
|
|
|
|
Région Ouest
|
8
|
|
|
|
|
|
Rhone Alpes
|
16
|
|
|
|
|
La base de données contient différents types de
données concernant la date d'apparition de chaque stade (date de semis,
date de floraison, date d'épiaison, date de récolte...) ainsi que
les données qui sont en relation avec les composantes de rendement et de
la teneur en protéines (nombre de grain, nombre de tiges, PMG, %
azote...). Deux autres variables ont été calculées et
ajoutées à ce jeu de données qui sont la
quantité d'azote absorbé en post-floraison (PANU), et la
quantité d'azote remobilisé (Annexe 1).
§ 251658752Quantité d'azote absorbé
en post-floraison= Quantité d'azote accumulée à
la récolte - quantité d'azote accumulée en floraison
§ Quantité d'azote
remobilisé = quantité d'azote absorbé en
floraison - quantité d'azote dans les pailles à la récolte
(Kg/ha)
2. Différentes étapes de
traitement et nettoyage de données
Afin d'aboutir à évaluer les
corrélations et les relations entre les variables, on a dû faire
un travail sur la base de données. La 1ère
étape du travail consiste à extraire les données de la
base de données Panoramix, les mettre en forme et les valider
(cohérence des unités, des mesures entre elles, etc...)...
Ensuite, on a calculé le GPD et analyser les corrélations entre
GPD et les variables explicative de sa variation. La
Figure 6 résume ce travail qui est fait sur
les données.
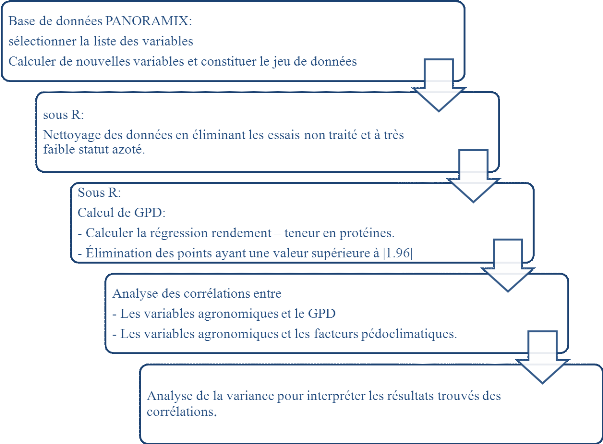
Figure 6:
Résumé des différentes étapes de
traitement et nettoyage de données
3. Calcul du
GPD
Tout d'abord on a commencé par trier les données
pour être « dans le domaine de validité » du
concept de GPD. En effet, la relation négative rendement -
protéines correspond à des conduites azotées proches de
l'optimum. Dans des essais fertilisation, on observe pour les niveaux de
fertilisation les plus faibles, une augmentation conjointe du rendement et de
la protéine avec l'accroissement des doses apportées. Le tri est
fait en filtrant les données et en ne laissant que les points qui
représentent un bon état de la culture. (Le filtrage est fait par
rapport à l'état azoté à floraison (donc
l'INNfloraison > 0, 8), l'état de santé
des cultures, et les modalités de fertilisations). Au-delà de
cette 1ère précaution, Oury et Goudin (2003) ont mis
en avant le fait que le calcul du GPD est extrêmement dépendant du
jeu de données utilisé car il dépend de la droite de
régression qui est calculée. Pour stabiliser son calcul, il
propose d'établir la droite de régression de manière
itérative, en ôtant les points dont les résidus sont trop
éloignés de la droite de régression. L'itération
est stoppée quand il n'y aura plus de valeur au-dessus de la valeur
seuil de résidus standardisés de +/- 1.96.Une fois la
régression finale établie, on calcule le GPD pour tous les points
de la base.
Après avoir obtenu les GPD, des régressions
linéaires entre le GPD, la quantité d'azote absorbé en
préfloraison, l'indice de nutrition azotée en floraison, l'azote
absorbé en post-floraison et l'azote remobilisé pendant la phase
de remplissage sont étudiées afin de trouver des variables
explicatives du GPD.
Le GPD nous indique si on a fait plus ou moins de
protéines ou de rendement qu'attendu. Le décomposer en un effet
absorption d'azote en pré-floraison, absorption d'azote en
post-floraison et azote remobilisé permet d'affiner le diagnostic en
évaluant si l'écart à la droite est plutôt
lié à l'élaboration du rendement (pré- ou
post-floraison) ou à l'élaboration de la teneur en
protéines.
4. Analyse
statistique
La régression linéaire de la relation entre le
rendement et la teneur en protéines a été calculée
pour les différents points du jeu de données. Ensuite les GPD ont
été définis comme les résidus standardisés
de cette relation. L'ensemble des analyses statistiques et les figures a
été réalisé avec le logiciel statistique R.
L'évaluation des corrélations entre les variables a
été faite par l'analyse de variance en utilisant le test de
Fisher.
Résultats et
discussion
Les résultats qui seront exposés à la
suite de ce document concernent un sous jeu de données extrait de la
base de données panoramix. En fait, ces données sont le
résultat d'une multitude d'essais qui ont été fait sur
plusieurs années dans différents endroits et pour
différents buts.
Nous avons effectué un nettoyage en éliminant
les lignes dans lesquels des données manquaient. Puis nous avons
ajouté de nouvelles variables qui sont la quantité d'azote
absorbé en pos-floraison, la quantité d'azote remobilisée,
la biomasse de remplissage en éliminant les valeurs insensées.
Ces valeurs éliminées concernaient surtout la quantité
d'azote absorbée en post-floraison vu qu'on a eu des valeurs très
étranges. Nous avons donc conservé un nouveau jeu de
données sur lequel nous avons travaillé.
1. Relation entre rendement en grain et la
teneur en protéines
Nous avons établi la relation entre le rendement en
grains (q/ha) et la teneur en protéines des grains (%). Nous avons
obtenu une corrélation significativement négative avec une pente
de l'ordre de -0.034 et un coefficient de corrélation
égale à 0.35 dans l'absence du facteur
variété (
Erreur ! Source du renvoi
introuvable.).
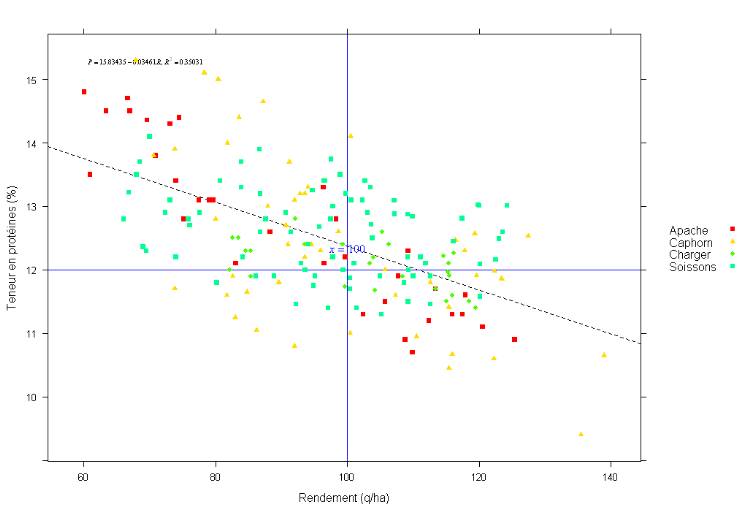
Figure 7: Relation entre la teneur en
protéines et le rendement pour les quatre variétés:
Apache, Caphorn, Charger et Soissons
Le
Erreur ! Source du renvoi
introuvable. résume les valeurs de la pente et des
coefficients de détermination obtenus suite à la
corrélation faite pour chaque variété.
Tableau 2: Pentes
et coefficients de détermination de la relation rendement - teneur en
protéines pour les quatre variétés
|
Variété
|
Pente
|
Coefficients de détermination (R²)
|
|
Charger
|
-0.015
|
0.30
|
|
Soissons
|
-0.013
|
0.1
|
|
Apache
|
-0.05
|
0.83
|
|
Caphorn
|
-0.044
|
0.34
|
Les valeurs des coefficients de corrélation
diffèrent d'une variété à une autre. En effet, pour
la variété Apache la relation est très bien
expliquée avec R² de l'ordre de 0.83. Cependant,
ce coefficient est très faible pour la variété Soissons.
L'analyse de variance de la relation rendement - teneur en protéines a
montré qu'il n'y a pas effet significatif de l'interaction rendement x
variété. Cependant, il existe un effet significatif de la
variété au seuil de á=5% avec un P-value de l'ordre de
0.022.
2. Calcul des
résidus standardisés (le GPD) de la relation rendement - teneur
en protéines
Le GPD est par définition les résidus de la
régression qui existe entre le rendement et la teneur en
protéines. Dans le but d'obtenir une droite de référence
pour les GPD, nous avons effectué une itération éliminant
les points qui sont supérieur à |1.96| (selon la loi de la
distribution normale des résidus). Les valeurs obtenues sont comprises
entre -1.88% et 1.82%.
Dans un second temps, nous avons calculé les GPDs pour
tout le jeu de données. Il est égal à la différence
entre la valeur de la teneur en protéines mesurée et sa valeur
prédite par le modèle (régression entre teneur en
protéines et rendement). Le résultat est comme montré dans
la
Erreur ! Source du renvoi
introuvable.. Les valeurs de GPD obtenues pour les quatre
variétés sont comprises entre - 9.13% et 3.46%.
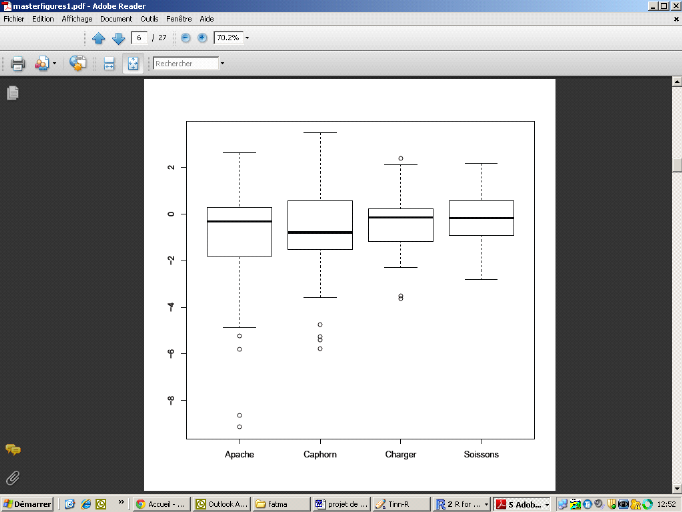
Figure 8: Boxplot montrant les GPDs selon les
variétés
3. Evaluation des
variables explicatives du GPD
Différentes corrélations ont été
évaluées entre des variables explicatives du processus
azotés (absorption d'azote en floraison, absorption d'azote en
post-floraison et la remobilisation d'azote) et le GPD. Le but était de
trouver quelle est parmi ces variables physiologiques celle qui explique au
mieux la déviation à la relation rendement - teneur en
protéine. Le
Erreur ! Source du renvoi
introuvable. résume le résultat trouvé pour
les différentes relations. Nous pouvons remarquer que parmi les quatre
variables, l'indice de nutrition azotée et la quantité d'azote
absorbée en floraison expliquent bien la variation du GPD. Cependant,
ceci n'exclue pas l'importance de l'absorption azotée en post-floraison
et la part d'azote remobilisé dans la détermination du GPD. En
effet l'analyse de variance a montré qu'il y a un effet significatif de
PANU1(*) et de
QNremobilisée2(*) sur le GPD (Pvalue égale à
1.013e-8 et 2.2e-16 respectivement).
Tableau 3:
Coefficients de détermination et Pvalue (á=5%) de la
corrélation entre le GPD et la quantité d'azote accumulée
en floraison (QNfloraison), indice de nutrition azotée
à floraison (INNfloraison), quantité d'azote
absorbée en post-floraison (PANU), quantité d'azote
remobilisée (QNremobiliseé) et Ecart PANU-QNR
|
GPD
|
|
Pvalue (á=5%)
|
R²
|
|
Apache
|
Caphorn
|
Charger
|
Soissons
|
Apache
|
Caphorn
|
Charger
|
Soissons
|
|
QNfloraison
|
<2.2e-16
|
0,53
|
0,60
|
0,37
|
0,04
|
|
INNfloraison
|
<2e-16
|
0,58
|
0,68
|
0,52
|
0,05
|
|
PANU
|
1.013e-8
|
0,12
|
0,05
|
0,1
|
0,1
|
|
QNremobiliseé
|
2.2e-16
|
0,44
|
0,43
|
0,18
|
0,017
|
|
Ecart PANU-QNR
|
<2.2e-16
|
0.66
|
0.55
|
0.49
|
0.17
|
Suite à l'analyse de variance des variables, on a
remarqué que, pour le modèle contenant le GPD et la PANU, il y a
un effet additif de la variété sur le PANU sans qu'il y soit un
effet significatif d'interaction. D'une part, l'effet de l'interaction entre
variété et l'INN, le QNfloraison et la
QNremobilisée est significative avec un P-value de l'ordre de
<2e-16 ; 1.32e-15 et 3.92e-11
respectivement.
Ainsi, comme premier résultat, nous pouvons tirer
l'importance du statut azoté de la plante pendant la floraison dans la
détermination de la relation qui existe entre la teneur en
protéine et le rendement. Ce résultat est confirmé par la
Erreur ! Source du renvoi
introuvable. qui montre que lorsque les points sont au-dessous
d'un état de nutrition azotée optimal (inférieur à
1) nous avons dans la majorité des cas des valeurs négatives de
GPD (excepté la variété Soissons). Ceci reflète un
résultat agronomique classique qui, pour un INN égale à 1
permet une conduite optimale du blé en rendement comme en teneur en
protéines. Toutefois, d'après le graphique nous pouvons voir que
même pour une valeur suboptimale d'INN (INN=0 ,8), nous avons des
valeurs de GPD variables. Ceci laisse une marge d'action en post-floraison sur
le pilotage d'un troisième apport. Dans le cas contraire où
l'indice de nutrition azotée est inférieur à 0,8 toutes
les valeurs de GPD sont négatives. Cependant la variété
« Soissons » présente des résultats
différents par rapport aux trois autres présentant un coefficient
de corrélation très faible mais montrant un effet d'interaction
sur la relation significatif pour un P-value(test de Student) de l'ordre de 2.8
e -16 .
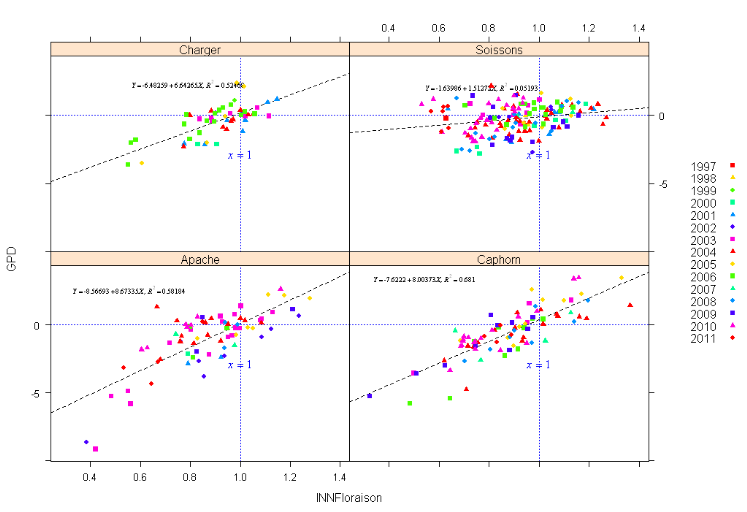 Figure
9: Relation entre GPD et l'INN floraison pour chacune des
variétés (la distribution des points est faite selon les
années des campagnes agricoles) Figure
9: Relation entre GPD et l'INN floraison pour chacune des
variétés (la distribution des points est faite selon les
années des campagnes agricoles)
4. La part de l'azote
absorbé en post-floraison dans l'explication du GPD
L'absorption d'azote en préfloraison et la
quantité d'azote absorbé en post-floraison sont deux processus
reliés entre eux. Obtenir une corrélation relativement faible
entre le GPD et le PANU3(*),
peut être dans une part, expliqué par cette
«dépendance» entre le QNfloraison4(*) et le PANU. Autrement dit, dans
la relation entre GPD - PANU, il existe une part de PANU négligé
qui n'est pas expliquée par QNfloraison ou
QNremobilisée. En effet, pour tenir compte de l'absorption
post-floraison sur le GPD nous avons étudié l'écart
à la relation PANU-QNfloraison et
PANU-QNremobilisée.
L'établissement d'une relation entre le GPD et les deux
variables a montré que l'écart à la relation
PANU-QNremobilisée explique mieux le GPD. De plus, nous avons
trouvé une étroite relation entre la quantité d'azote
absorbé en préfloraison et la quantité d'azote
remobilisée avec un R² compris entre 0.7 et 0.9. De ce fait la part
d'azote absorbé en post-floraison dans l'explication de GPD sera
expliquée à travers l'étude de la relation
GPD-écart PANU-QNremobilisée. La
Erreur ! Source du renvoi
introuvable. montre cette relation. L'écart entre les deux
variables en question explique de 50% à 60% la variation de GPD pour les
variétés Apache, Charger et Caphorn et seulement 18% pour la
variété Soissons. Cette dernière, comme dans le cas
d'étude de la relation entre GPD et INN, a montré un faible
coefficient de corrélation tout en ayant un effet significatif
d'interaction avec l'écart PANU-QNR sur la relation.
En outre, les deux variables INN et l'écart à la
relation PANU-QNremobilisée semble être
dépendants. Il existe une relation similaire qui relie ces deux facteurs
au GPD. L'analyse de variance du modèle intégrant à la
fois le GPD, l'écart à la relation PANU-
QNremobilisée et l'INNfloraison montre un effet
significatif de l'interaction entre les deux variables avec un Pvalue de
l'ordre de 3.76e-16. La
Figure 11 explique la relation GPD - écart
PANU-QNremobilisée en tenant compte de l'indice de la
nutrition azotée. La figure explique les
résultats trouvés auparavant. En effet, pour un INN optimale
(entre 0,8 et 1) les valeurs de GPD sont positives. La biomasse est alors
suffisante pour assurer un bon nombre de grain et une demande en azote forte
pour générer de l'absorption post-floraison. De plus les valeurs
de l'écart PANU-QNremobilisée sont plutôt
positives. Dans le cas contraire où les valeurs de l'INN à
floraison sont faibles (< 0.8) tous les points du GPD affichent des valeurs
négatives tout comme les valeurs de l'écart
PANU-QNremobilisée.
Ainsi, il nous a paru important d'expliquer cet écart
à partir des variables agronomiques qui déterminent le rendement
ou bien la teneur en protéines pour voir le poids relatif entre le
pré et le post floraison.
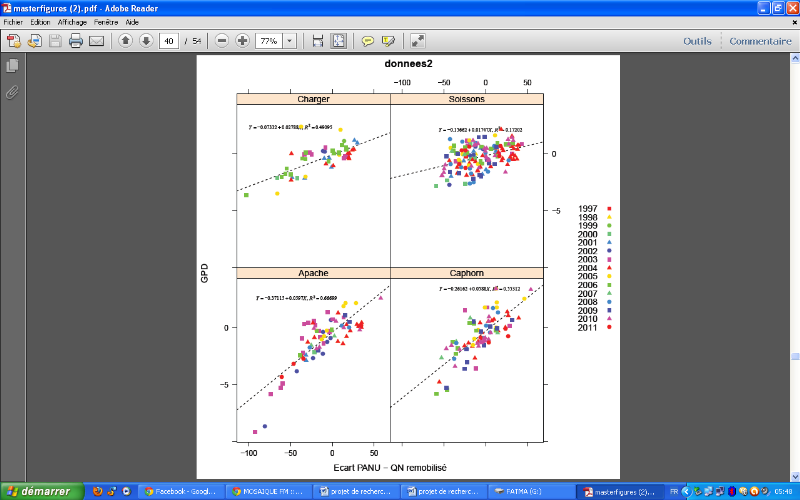
Figure 10: Relation entre l'écart
à la relation PANU-QNremobilisée et le GPD pour les
quatre variétés Apache, Caphorn , Charger et Soissons (la
distribution des points est faite selon les années)
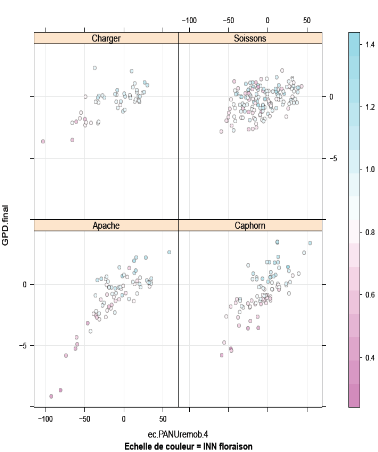
Figure 11:
Relation entre le GPD et l'écart à la relation
PANU-QNremobilisée expliquée par l'INNfloraison
pour les quatre variétés.
Le choix était misé sur le PMG (poids mille
grains) et la biomasse aérienne de remplissage. Le
Erreur ! Source du renvoi
introuvable. résume le résultat de l'analyse de
variance pour les deux variables agronomiques. La biomasse aérienne
pendant le remplissage explique bien cet écart entre la PANU et la
QNremobilisée. En fait, la biomasse de remplissage est la
différence entre la biomasse aérienne totale à la
récolte et la biomasse aérienne en floraison, donc c'est celle
produite pendant la phase post-floraison à partir de l'azote
remobilisé ou bien absorbé par les racines pendant cette phase.
De ce fait nous pouvons percevoir le rôle de l'absorption azotée
tardive sur le GPD.
Tableau
4:Coefficient de détermination et Pvalue de la relation entre
l'ecart PANU-QNremobilisée et le Poids mille grain (PMG) et la biomasse
de remplissage (BMremplissage)
|
Ecart PANU-QNremobilisée
|
PMG5(*)
|
|
Pvalue (á=0.5)
|
R²
|
|
Apache
|
Caphorn
|
Charger
|
Soissons
|
Apache
|
Caphorn
|
Charger
|
Soissons
|
|
1.189e-10
|
0,16
|
0,033
|
0,0033
|
0,28
|
|
BMremplissage6(*)
|
|
Pvalue
|
R²
|
|
Apache
|
Caphorn
|
Charger
|
Soissons
|
Apache
|
Caphorn
|
Charger
|
Soissons
|
|
2.2e-16
|
0,3
|
0,17
|
0,4
|
0,17
|
5. Evaluation des
variables agro-climatiques
Le contexte agro-climatique est susceptible de modifier le
poids des déterminants du GPD. Nous avons bien remarqué que les
variables PANU et QNremobilisé explique d'une manière ou d'une
autre la variation de GPD. Ces variables dépendent
énormément des conditions pédo-climatiques du site.
Après avoir connu les variables agronomiques explicatives de GPD, on a
étudié la relation qui existe entre les variables agronomiques et
les différents facteurs climatiques susceptibles de les modifier. Les
facteurs climatiques sont:
- Somme du rayonnement (phase floraison jusqu'à
maturité) : sRg
- Nombre des jours ou le ETR/ETM est <1 :
ETR/ETM<1
- Somme de T>25°C (phase floraison jusqu'à
maturité): somme T>25°C
- Nombre de jours pour T>25°C (phase floraison
jusqu'à maturité): NjoursT>25°C
- Somme de T<15°C (phase floraison jusqu'à
maturité): sT<15°C
En effectuant les corrélations entre les
différentes variables, on a obtenu des coefficients de
détermination très faible (
Tableau 6). On a eu recours à l'analyse de
variance pour voir l'effet de chaque facteur climatique sur les variables
agronomique. Le
Erreur ! Source du renvoi
introuvable. résume les résultats trouvés.
Une température élevée et un
déficit hydrique pendant la phase de remplissage ont un effet
significatif sur la QNremobilisée et le PANU. En effet, une
succession de jours avec une température dépassant 25°C ou
bien le maintien d'une température élevée pendant la phase
post-floraison diminue la quantité de l'azote remobilisé des
parties végétatives vers les grains ainsi que la capacité
de la plante à absorber de l'azote pendant cette phase. Ce même
effet est observé lorsqu'un déficit hydrique a lieu.
Tableau 5: Pvalue
(á=5%) des différentes relations entre la quantité d'azote
remobilisée (QNremobilisée), la quantité
d'azote absorbée en post-floraison (PANU), l'ecart à la relation
PANU-QNremobilisée (ecPANU.QNR) et la biomasse de remplissage
(BMremplissage) et les facteurs climatiques.
|
|
NjoursT>25°C
|
somme T>25°C
|
ETR/ETM<1
|
Spluie
|
Setr
|
sT<15°C
|
sRg
|
|
QNremobilisée
|
0.0015
|
0.0035
|
0.011
|
0.00069
|
0.01
|
ns
|
ns
|
|
PANU
|
1.25e-8
|
ns
|
0.00424
|
0.019
|
ns
|
ns
|
ns
|
|
ecPANU.QNR
|
7.033e-11
|
ns
|
ns
|
ns
|
ns
|
ns
|
ns
|
|
BMremplissage
|
9.914e-9
|
ns
|
ns
|
0.009
|
0.036
|
0.042
|
0.016
|
Tableau 6:
Coefficients de corrélation des differentes corrélations entre la
quantité d'azote remobilisée (QNR), la quantité d'azote
absorbée en post-floraison (PANU), l'ecart à la relation
PANU-QNremobilisée (ecPANU.QNR) et la biomasse de remplissage (BMremp)
et les facteurs climatiques pour les variétés Apache (A), Caphorn
(C), Charger (Ch), Soissons (S)
|
NjoursT>25°C
|
somme T>25°C
|
ETR/ETM<1
|
Spluie
|
Setr
|
|
|
A
|
C
|
Ch
|
S
|
A
|
C
|
Ch
|
S
|
A
|
C
|
Ch
|
S
|
A
|
C
|
Ch
|
S
|
A
|
C
|
Ch
|
S
|
|
|
QNR
|
0.007
|
0.0723
|
0.009
|
0.032
|
0.00015
|
0.03
|
0.07
|
0.0026
|
0.0376
|
0.0037
|
0.0090
|
0.0102
|
0.0130
|
0
|
0.0065
|
0.0517
|
0.0607
|
0.0144
|
0.0361
|
|
|
|
PANU
|
0.09
|
0.0002
|
0.2
|
0.096
|
0.09
|
0.0013
|
0.12
|
0.1138
|
0.0564
|
0.0004
|
0.0166
|
0.0685
|
4e-05
|
4e-05
|
1e-05
|
0.0470
|
0.0609
|
0.0047
|
0.0049
|
0.0011
|
|
|
ecPANU.QNRemoilisée
|
0.09936
|
0.04604
|
0.12
|
0.2
|
0.06
|
0.0253
|
0.0102
|
0.1618
|
0.0023
|
0.0006
|
0.0016
|
0.0503
|
0.0067
|
2e-05
|
0.0060
|
0.0093
|
0.0002
|
0.0004
|
0.0115
|
0.0015
|
|
|
BMremp
|
0.08373
|
0.034
|
0.13
|
0.085
|
0.044
|
0.037
|
0.0281
|
0.0736
|
4e-04
|
0.0005
|
0.0030
|
0.0147
|
0.0351
|
0.0004
|
0.00097
|
0.0065
|
0.0088
|
0.0003
|
0.0021
|
0.0038
|
|
Discussion
Le travail a pour but d'étudier la relation qui existe
entre le rendement et la teneur en protéines sous un. Ce travail fait
suite au travail de Bogard (2011) qui a évalué cette relation
pour analyser la variabilité génétique des
variétés. il a utilisé pour ça 27
variétés différentes. Dans notre cas on a utilisé
quatre variétés cultivées dans des conditions
environnementales différentes et l'objectif est plutôt d'analyser
la variabilité environnementale autour de la relation.
Nous avons pu confirmer que la relation qui existe entre la
teneur en protéines et le rendement en grains est négative et que
l'amélioration de ces deux critères dans la production de
blé tendre demeure un peu difficile à cause de cet antagonisme.
Le calcul de la déviation à la régression
entre teneur en protéines et rendement permet d'évaluer et
d'améliorer cette relation. Le GPD est ensuite corrélé
avec différents mécanismes physiologiques. On a pu montrer suite
aux différentes corrélations que parmi les mécanismes
physiologiques l'absorption azotée en pré-floraison est une
composante très importante de la variation de GPD. De plus,
malgré la faible corrélation qu'on a obtenu entre le GPD d'un
côté et la quantité d'azote remobilisé et la
quantité d'azote absorbé en post-floraison d'un autre
côté, l'analyse de variance a montré que ces deux
mécanismes physiologiques ont un effet significatif sur le GPD. Entre
ces deux processus de l'absorption azotée, la remobilisation de l'azote
était mieux corrélée avec le GPD que l'absorption
azotée post-floraison.
Au cours de cette étude nous sommes passés de
l'étude des résidus de la relation teneur en protéines -
rendement en grains à celle des résidus entre l'absorption
post-floraison et la quantité d'azote remobilisée. Le but
était de mieux comprendre la variation de GPD à travers un effet
sur l'absorption d'azote post-floraison.
Concernant l'évaluation des facteurs
pédoclimatiques, les corrélations obtenues entre les variables
explicatives de GPD et les facteurs climatiques étaient faibles. Ce
résultat peut être la conséquence de
l'hétérogénéité des données
utilisées. Cependant, on a pu montrer l'effet de quelques facteurs
environnementaux sur les variables agronomiques. Il y a un effet significatif
de la température et de déficit hydrique sur l'azote
absorbé en post-floraison et l'azote remobilisé.
1. Relation teneur en protéines -
rendement en grain
Le phénomène de dilution peut expliquer
l'antagonisme entre le rendement et la teneur en protéines puisque les
deux variables dépendent beaucoup des métabolismes
carbonés et azotés au sein de la culture.
Les résultats ont montré qu'entre le rendement
et la teneur en protéines, la relation est négative. Seulement,
on peut remarquer que les valeurs de la pente sont relativement faibles par
rapport à ce qui a été montré dans la thèse
de Bogard (2011). L'utilisation d'un nombre assez grand de points (373
observations) sans recours à l'utilisation des moyennes peut affecter
les résultats. En effet, l'utilisation des moyennes aide à
neutraliser l'interaction G X E dont le rendement et la teneur en
protéines sont très dépendants et par conséquent
permet une meilleure estimation de la relation teneur en protéines -
rendement (Oury et Goudin, 2007 ; Bogard, 2011). De plus, le faible
coefficient de corrélation obtenu chez la variété Soissons
peut être la conséquence du bruit introduit par les facteurs
environnementaux vu que le nombre d'observation chez la variété
en question est supérieur aux autres variétés.
Il est certain que la teneur en protéines est le
premier critère de qualité chez les producteurs de blé
tendre (Jeuffroy et al., 2000). Toutefois, le grain contient d'autres
composants contribuant à sa qualité nutritionnelle. Une
hypothèse a été étudiée par Munier-Jolain et
Salon (2005) avait pour but de voir s'il y a un antagonisme entre un rendement
élevé et une bonne qualité nutritionnelle du grain. On a
pu montrer que pour toutes les espèces étudiées il existe
une relation négative entre les composants du rendement et ceux de la
qualité nutritionnelle (
Erreur ! Source du renvoi
introuvable.) (Munier-Jolain et Salon, 2005). Une telle
étude peur être intéressante pour mieux évaluer la
relation entre le rendement en grain et les critères qualitatifs du
grain.
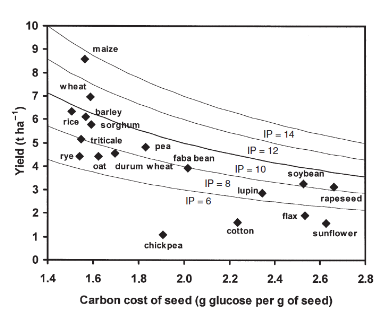
Figure 12: Relation entre la consommation du
carbone pour la production des semences pour plusieurs espèces et leurs
rendements (afin de comparer la performance de production plusieurs courbes
d'iso production ont été utilisées)
2. Relation entre le GPD et
les différents métabolismes physiologiques
Plusieurs corrélations ont été
étudiées entre le GPD et les variables du processus azoté.
Le statut azoté de la plante en floraison
exprimé par l'INNfloraison a été
déterminant dans l'explication de GPD. La fertilisation azotée
avant la floraison est par conséquent déterminante pour la
production de blé. Jeuffroy et al. (1999) et Jeuffroy et
al. (2000) ont montré l'importance de cette phase dans la
détermination du rendement et de la teneur en protéines à
partir du modèle Azodyn qui simule le fonctionnement de la culture de
blé en conditions de nutrition azotée variées (
Erreur ! Source du renvoi
introuvable.). La teneur en protéines et le rendement sont
optimisés grâce à un ensemble de stratégie de
fertilisation azotée effectué dans la plupart des cas au cours de
la période préfloraison (Jeuffroy et Oury, 2012).
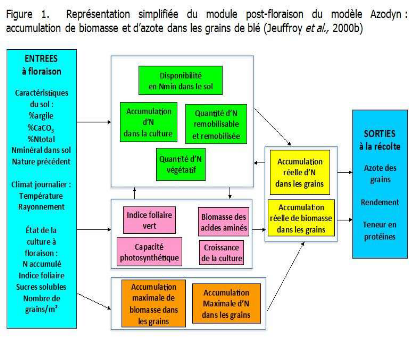
Figure 13: représentation
simplifiée de module post-floraison du modèle Azodyn:
accumulation de biomasse et d'azote dans les grains de blé (Jeuffroy et
al., 2000)
Mis à part l'importance de l'azote absorbé en
préfloraison, les deux mécanismes physiologiques, en
l'occurrence, l'absorption post-floraison et l'azote remobilisé ont
montré un effet significatif sur l'écart à la relation
teneur en protéines - rendement.
Concernant la quantité d'azote remobilisée, on a
pu obtenir une corrélation avec le GPD (R² égale à
0.44 pour la variété Apache) montrant l'importance de cette
composante dans l'explication du GPD. Il a été montré que
la remobilisation de l'azote vers le grain est la source la plus importante
dans l'accumulation de l'azote dans le grain. Il est le principal processus
responsable de variations de rendement (Bancal et al., 2008). Par
conséquent, en ciblant l'amélioration variétale sur la
quantité d'azote absorbée en préfloraison et en maintenant
une forte efficacité de remobilisation de l'azote, on peut
améliorer la teneur en protéines sans réduire le rendement
et donc améliorer la relation qui existe entre les deux variables
(Slafer et al., 1989; Monaghan, 2001). Cependant, plusieurs auteurs
dans leurs études n'ont pas trouvé de relation significative
entre l'azote remobilisé et le GPD (Le Gouis et al., 2010;
Bogard, 2011).
La variabilité génétique de GPD
était bien expliqué par la variabilité
génétique de l'absorption de l'azote en phase post-floraison
indépendamment de la quantité d'azote absorbée à
floraison (Bogard, 2011; LeGouis et al., 2010; Oury et Goudin, 2007).
Le Gouis et al. (2010) ont montré que le GPD est
expliqué à 50 % par la quantité d'azote absorbé en
post-floraison (PANU). Ces résultats ne correspondaient pas à ce
qu'on a trouvé dans l'étude de l'absorption azoté en
post-floraison. En effet dans des conditions multi-environnementales, le PANU
expliquait mal le GPD (R² < = 0,1). Ceci peut être
expliqué par la grande variabilité et
hétérogénéité des données. Le PANU
est très sensible aux conditions climatiques pendant la phase de
remplissage (Pask et al., 2011) et un déficit hydrique, par
exemple, provoque la diminution de la contribution du PANU dans
l'amélioration de la teneur en protéines (Monaghan et
al., 2001).
L'explication du GPD de point de vue physiologique est un peu
complexe. En effet cette variable dépend à la fois de trois
processus azotée: l'azote accumulé en floraison, l'azote
remobilisé et l'azote absorbé en post-floraison, mais à
des degrés de dépendances différents.
On propose d'optimiser la fertilisation tardive dans le but
améliorer la relation teneur en protéines - rendement (Monaghan
et al., 2001). De nouvelles stratégies de fertilisation
azotée sont de plus en plus conseillées. Il s'agit de supprimer
le premier apport et de décaler le deuxième et le
troisième apport vers des dates plus tardives. D'autre part, dans la
perspective de diminuer la quantité d'intrants (plan ecophyto 2018)
associer des légumineuses aux cultures céréalières
aide à augmenter la teneur en protéines tout en réduisant
le recours à la fertilisation azotée (Jeuffroy et Goudin, 2012).
Ainsi, avoir un bon équilibre entre rendement et
qualité de production dépend des facteurs environnementaux,
génétiques et l'interaction entre les deux. Pour
l'amélioration de cette relation, le GPD est considéré
comme un bon moyen.
Conclusion
Chez le blé (Titicum aestivum L.) mis à
part le rendement, la teneur en protéines représente un
critère très important déterminant la valeur
économique de blé. L'objectif d'une bonne production de
blé a pour but de concilier entre rendement et teneur en
protéines. Toutefois, ceci reste difficile à réaliser
à cause de la relation négative qui existe entre les deux
critères.
Une des façons de comprendre la relation qui existe
entre rendement et teneur en protéines est de calculer l'écart
à cette relation (ce qu'on appelle GPD pour Grain Protein Deviation). Le
GPD a pour but d'identifier les génotypes qui présentent une
teneur en protéines supérieure à celle prédite par
la relation teneur en protéines - rendement. L'efficacité de cet
outil a été démontrée dans plusieurs études
précédentes. En effet, on a montré que le GPD est
partiellement sous le contrôle génétique, on l'utilise pour
l'évaluation des variétés de blé en cours
d'inscription au catalogue officiel français.
Les processus azotés tels que l'absorption de l'azote
en pré et post-floraison ou encore la remobilisation d'azote
accumulé pendant la floraison des parties végétatives vers
les grains sont les majeurs sources d'azote et de matières sèches
dans le grain. Afin de mettre en évidence les caractères
affectant séparément le rendement et la teneur en
protéines on a proposé de corréler ces processus avec le
GPD. L'absorption azotée en post-floraison explique bien le GPD. En
effet, la variabilité génétique pour le GPD est
reliée à la variabilité génétique de
l'absorption post-floraison.
Au cours de ce travail, l'objectif était de voir la
possibilité d'expliquer les variations interannuelles et multilocales de
GPD via les mêmes processus azotés qu'on a déjà
étudiés pour expliquer la variabilité
génétique.
Ainsi on a pu montrer l'importance de l'absorption
azotée en préfloraison comme processus expliquant la variation de
GPD. Le coefficient de déterminant de la corrélation entre le
QNfloraison et le GPD était de l'ordre de 0,8 (pour la
variété Apache). Ce résultat est logique vu l'importance
de la phase préfloraison dans la détermination du nombre de
grains formés ainsi que les quantités d'azote et de carbone
disponibles pour la croissance des grains après la floraison. Toutefois,
la remobilisation de l'azote et l'absorption post-floraison de l'azote avaient
un effet significatif sur le GPD.
Au vu de cette synthèse des résultats, nous
pouvons conclure que le GPD est un outil intéressant dans la
sélection des variétés ayant à la fois un bon
rendement et une teneur en protéines élevée. Ce GPD est
lié aux métabolismes azotés de la plante.
Il sera intéressant par ailleurs de réaliser un
travail de caractérisation et de typologie des environnements
étudiés qui permettent d'identifier et de quantifier les facteurs
limitants dans chaque environnement. On pourra alors d'intégrer des
aspects génétiques et physiologiques expliquant le GPD.
Annexe1: Ensemble des
variables de la base de données
|
Site.Type de sol
|
|
Site.Sol.Réserve utile
|
|
Site.Sol.Réserve facilement utilisable
|
|
Site.Station locale
|
|
Site.Agriculteur
|
|
Site.Code postal
|
|
Site.Libellé
|
|
Site.Région
|
|
Site.Région administrative
|
|
Site.Département
|
|
Campagne Agricole.Année
|
|
Essai
|
|
Code culture précédent
|
|
Code labour
|
|
Semis.Date
|
|
Protocole
|
|
Espèce
|
|
Variété
|
|
Complément
|
|
Identifiant
|
|
Parcelle élémentaire
|
|
Sans traitement fongicide
|
|
Commentaire
|
|
Semis.Densité
|
|
Levée.Date observée
|
|
Levée.Date mesurée
|
|
Levée.Densité.Mesure
|
|
Epi 1cm.Date observée
|
|
Epi 1cm.Date mesurée
|
|
Epi 1cm.Densité.Mesure
|
|
Epi 1cm.% Tiges + 3f
|
|
Epi 1cm.Tiges + 3f.Mesure
|
|
Epi 1cm.Biomasse aérienne.Mesure
|
|
Epi 1cm.Teneur azote aérien.Mesure
|
|
Epi 1cm.INN
|
|
Deux noeuds.Date observée
|
|
Deux noeuds.Biomasse aérienne.Mesure
|
|
Deux noeuds.Teneur azote aérien.Mesure
|
|
Deux noeuds.Azote absorbé aérien
|
|
Deux noeuds.INN
|
|
Deux noeuds.Indice absorption N
|
|
Dernière feuille.Date mesurée
|
|
Dernière feuille.Biomasse aérienne.Mesure
|
|
Dernière feuille.Teneur azote aérien.Mesure
|
|
Dernière feuille.Azote absorbé aérien
|
|
Dernière feuille.INN
|
|
Epiaison.Date observée
|
|
Floraison.Date observée
|
|
Floraison.Date mesurée
|
|
Floraison.Epis par m²
|
|
Floraison.Epillets fertiles par épi
|
|
Floraison.Biomasse aérienne.Mesure
|
|
Floraison.Teneur azote aérien.Mesure
|
|
Floraison.Azote absorbé aérien.Mesure
|
|
Floraison.INN
|
|
Récolte.Date observée
|
|
Récolte.Rendement.Mesure
|
|
Récolte.PMG.Mesure
|
|
Récolte.Teneur protéines.Mesure
|
|
Récolte.Azote absorbé grain.Mesure
|
|
Récolte.Biomasse aérienne.Mesure
|
|
Récolte.Grains par épi.Mesure
|
|
Récolte.Grains par m².Mesure
|
|
Qualité.Poids Spécifique.Valeur
|
|
Qualité.Grain sur Paille.Valeur
|
|
Qualité.Teneur Azote Paille.Valeur
|
|
Qualité.Teneur Azote Grain.Valeur
|
|
Azote.Date 1
|
|
Azote.Dose 1
|
|
Azote.Produit 1
|
|
Azote.Commentaire 1
|
|
Azote.Date 2
|
|
Azote.Dose 2
|
|
Azote.Produit 2
|
|
Azote.Commentaire 2
|
|
Azote.Date 3
|
|
Azote.Dose 3
|
|
Azote.Produit 3
|
|
Azote.Commentaire 3
|
|
Azote.Date 4
|
|
Azote.Dose 4
|
|
Azote.Produit 4
|
|
Azote.Commentaire 4
|
|
Azote.Date 5
|
|
Azote.Dose 5
|
|
Azote.Produit 5
|
|
Azote.Commentaire 5
|
|
Azote.Dose totale
|
|
Azote.Nombre
|
|
Irrigation.Date 1
|
|
Irrigation.Dose 1
|
|
Irrigation.Produit 1
|
|
Irrigation.Commentaire 1
|
|
Irrigation.Date 2
|
|
Irrigation.Dose 2
|
|
Irrigation.Produit 2
|
|
Irrigation.Commentaire 2
|
|
Irrigation.Date 3
|
|
Irrigation.Dose 3
|
|
Irrigation.Produit 3
|
|
Irrigation.Commentaire 3
|
|
Irrigation.Date 4
|
|
Irrigation.Dose 4
|
|
Irrigation.Produit 4
|
|
Irrigation.Commentaire 4
|
|
Irrigation.Date 5
|
|
Irrigation.Dose 5
|
|
Irrigation.Produit 5
|
|
Irrigation.Commentaire 5
|
|
Irrigation.Date 6
|
|
Irrigation.Dose 6
|
|
Irrigation.Produit 6
|
|
Irrigation.Commentaire 6
|
|
Irrigation.Date 7
|
|
Irrigation.Dose 7
|
|
Irrigation.Produit 7
|
|
Irrigation.Commentaire 7
|
|
Irrigation.Dose totale
|
|
Irrigation.Nombre
|
Références
bibliographiques
Anonyme1: Principes et lois de fertilisation.
Available from internet < URL «
http://www.yara.fr/fertilizer/fertilizer_facts/fertilisation_laws/nitrogen_and_quality.aspx»
Anonyme 2: Bogard M, Moreau D, Allard V, Martre P, Le
Gouis J. Genetic analysis of grain protein deviation in wheat
Asseng, S., & Milroy, S. P. (2006). Simulation of
environmental and genetic effects on grain protein concentration in wheat.
European Journal of Agronomy, 25(2), 119-128.
Atilio, JB, 2006. Physiology and biochemistry of
source-regulated protein accumulation in the wheat grain. Journal of plant
physiology, 164: 581-590
Bancal MO, Roche R, Bancal P, 2008. In wheat
crops, late foliar diseases decreased nitrogen yiel through N uptake rather
than through N remobilization variations. Annals of botany.
Barbottin, A. (2004). Utilisation d'un
modèle de culture pour évaluer le comportement des
génotypes?: Pertinence de l'utilisation d'Azodyn pour analyser la
variabilité du rendement et de la teneur en protéines du
blé tendre. Agronomie, Paris. Thesis.
Barbottin, A., Lecomte, C., Bouchard, C., & Jeuffroy,
M.-H. (2005). Nitrogen Remobilization during Grain Filling in Wheat.
Crop Science, 45(3), 1141-1150.
Barraclough, P. B., Howarth, J. R., Jones, J.,
Lopez-Bellido, R., Parmar, S., Shepherd, C. E., & Hawkesford, M.
J. (2010). Nitrogen efficiency of wheat: Genotypic and environmental
variation and prospects for improvement. European Journal of Agronomy,
33(1), 1-11.
Bertheloot, J. (2009). Distribution de
l'azote chez le blé (Triticum aestivum L.) après la floraison?:
un modèle dynamique fondé sur une approche
structure-fonction. Thèse, Agroparitech, France : 132
« p. »
Ben Slimane R, 2010. Effets de la septoriose
foliaire sur la sénescence et les flux d'azote pendant le remplissage
des grains chez le blé tendre.Thèse,AgroParisTech,France:186
« p. »
Bogard M, 2011.Analyse
génétique et écophysiologique de l'écart à
la relation teneur en protéines - rendement en grains chez le blé
tendre (Triticum aestivum L.).These, Université Blaise Pascal,
France : 193 « p. »
Bogard, M., Moreau, D., Allard, V., Martre, P., & J,
L. G. (2011). Genetic analysis of grain protein deviation in wheat,
1-3.
Bogard M, Jourdan M, Allard V, artre P, Perretant MR,
Ravel C, Heumez E, Orford S, Snape J, Griffiths S, Gaju O, Foulks J, LE Gouis
J, 2011. Anthesis date mainely explained correlation between
post-anthesis leaf senescence, grain yiel, and grain protein concentration in a
winter wheat population segregating for flowering time QTLs. Journal of
experimental botany. 1 - 16.
Borrell A et Hammer GL, 2000. Nitrogen Dynamics
and tje physiological Basis of Stay-Green in Sorghum. Crop Science.40: 1295 -
1307
Casals M-L, 1998. Demande RMCT, Thème:
intégration d'un module azote dans les modèles agroclimatiques de
l'ITCF. ITCF : service technique de production.
Comifer, (2011). Calcul de la fertilisation
azotée, Guide méthodologique pour
l'établissement des prescriptions locales.
Gate P, 1995. Ecophysiologie du
blé.In : Technique et Documentation - Lavoisier eds.429p.
Jeuffroy, M.-hélène, Gate, P., Machet,
J.-M., & Recous, S. (2011). Gestion de l'azote en agriculture?:
enjeux environnementaux et perspectives agronomiques.
Jeuffroy M.H., Recous S, 1999. Azodyn: a simple
model simulating the date of nitrogen
deficiency for decision support in wheat fertilization. European
Journal of Agronomy 10 : 129-144.
Jeuffroy, M.H., M.L. Girard, et C. Barré.
2000. Blé tendre : comprendre et prévoir la
teneur en protéines. Perspectives Agricoles 261 : 24 - 31.
Jeuffroy M.H et Oury F.X,2012. Impact des
nouvelles techniques de production, impliquant de faibles niveaux d'intrants,
sur la quantité de protéines du
blé.CIAG
Le Gouis Allard V, Joseph V, Cormier D, Messaoud J,
Bonnemoy B, Lemaire P, Rougeol S, Taulemesse , 2010. .
Déterminismes génétique et
écophysiologiques de la dynamique d'absorption de l'azote
post-floraison
Monaghan JM., Snape JW., CHojecki A.JanS. &
Kettlewell PS. (2001). The use of grain deviation for identifying
wheat cultivars with high grain protein concentration and yield.
Euphytica, 122, 309-317
Morot-Gaudry J-F,1997.assimilation de l'azote
chez les plantes: aspects physiologique, biochimique et moléculaires.
.INRA,PARIS
Mi, G., Tang, L., Zhang, F., & Zhang, J.
(2000). Is nitrogen uptake after anthesis in wheat regulated by sink size?
Field Crops Research, 68(3), 183-190.
Munier-Jolain, N. G., & Salon, C. (2005).
Are the carbon costs of seed production related to the quantitative and
qualitative performance? An appraisal for legumes and other crops. Plant,
Cell and Environment, 28(11), 1388-1395.
Oury, F.-X., & Godin, C. (2007). Yield and grain protein
concentration in bread wheat: how to use the negative relationship between the
two characters to identify favourable genotypes? Euphytica,
157(1-2), 45-57.
Pask A.J.D, Bradley R.S, Jamieson P.D
& Foulkes M.J, (2011). Quantifying how winter
wheat crops accumulate and use nitrogen reserves during growth. Field Crop
Research,126, 104 - 118
Triboi, E., & Anne-Marie Triboi-Blondel.
(2002). Productivity and grain or seed composition?: a new approach to an old
problem -- invited paper. European Journal of Agronomy, 16,
163-186.
Semenov M.A, Jamieson P.D, Martre P, 2007.
Deconvoluting nitrogen use efficiency in wheat: A simulation study. European
Journal of Agronomy, 26 : 283-294
Shewry P-R,2007. Improving the protein content
and composition of cereal grain. Journal of Cereal Science, 46 : 239-250
Slafer A., G., & H.Andrade, F. (1989). Genetic I m p r o v e
m e n t in Bread Wheat (Triticum aestivum ) Yield in Argentina. Field Crops
Research, 21, 289-296.
* 1 Quantité d'azote
absorbée en post-floraison
* 2 Quantité d'azote
remobilisée
* 3 Azote absorbé en
post-floraison.
* 4 Quantité d'azote
absorbé en floraison.
* 5 Poids mille grains
* 6 Biomasse pendant le
remplissage
| 

