Caractérisation des communautés végétales du Miombo en lien avec les paramètres physiques du solpar Pacifique WIMANA KIZILA Université de Lubumbashi - Diplôme d'Etudes Approfondies (DEA) 2012 |

INTRODUCTIONEn Afrique Centro australe, la forêt claire du Miombo occupe une superficie de 3.6 million de Km2 (Giliba et al., 2011). Elle occupe 80 % de la végétation naturelle en Zambie, 11 % pour la république de démocratique du Congo (Banda et al., 2006). Elle est distribuée sur 11 pays : République démocratique du Congo, République Sud-Africaine, Malawi, Burundi, Tanzanie, Angola, Zimbabwe, Zambie, Mozambique, Kenya, Botswana (Campbell et al., 2007). Environ 100 millions des personnes habitant les milieux ruraux et urbains dépendent de cet écosystème (Timberlake & Chidumayo, 2011). Avec des particularités écologique et économique importantes (Frost, 1996). Elle est pourvoyeur de bois d'oeuvre et de bois de chauffe. Elle fournit la matière première pour la production du charbon de bois (Luoga, 2005). Elle est également source de fourrage et de divers produits forestiers non ligneux contribuant à l'alimentation des populations locales (Fanshawe, 1971; Malaisse, 1997). Elle est aussi importante pour des raisons spirituelles et culturelles des populations locales (Gambiza et al., 2000). Cet écosystème a connu au cours du 20e siècle un taux de dégradation élevé sous l'effet de l'accélération des pratiques anthropiques : surexploitation de la ressource bois (bois d'oeuvre et charbon de bois), conversion pour l'agriculture et le pâturage, modifications des régimes de perturbation liés au feu (Campbell, 1996), alors que, le Miombo est une cible importante pour la conservation de la biodiversité et le suivi de changement de la végétation (Panteleo et al., 2011). Face à ces défis, la restauration du Miombo est un enjeu majeur. La restauration écologique doit s'appuyer à la fois sur une connaissance approfondie du fonctionnement écologique des écosystèmes et de leur réponse aux perturbations humaines (Clevel et Aronson, 2010). 2 1. SYNTHESE BIBLIOGRAPHIQUE 1.1. Définition et distribution de la forêt claire Située presqu'entièrement dans la zone tropicale australe, la forêt claire comprend deux bandes : La première s'étend entre la latitude 5°N et 23°N, avec une végétation à prédominance de diverses espèces d'acacias ; et la seconde bande va de 5° S 25° S de latitude. Un massif homogène qui se prolonge jusqu'en Afrique du Sud. Elle est caractérisée par une végétation à dominance de la sous famille de Caesalpinioideae du genre : Brachystegia-Isoberlinia-Julbernardia ; lesquelles espèces sont rarement trouvées en dehors de la forêt claire du Miombo (Strömquist & Backéus, 2010). De par les précipitations et la richesse floristique, la forêt claire a été subdivisée en deux : ? Le Miombo humide, dont les précipitations sont supérieures à 1000 mm de pluies/an. Il est dominé par les espèces telles que : Brachystegia floribumda, B. wangermeeana, Marquesia macroura. ? Le Miombo sec, dont les précipitations sont inférieures à 1000 mm de pluies/an. Il est dominé par les espèces telles que : B. spiciformis, B. boehmii, Julbernardia paniculata. La présente étude s'intéresse à la forêt claire du sud de l'équateur appelée Miombo.
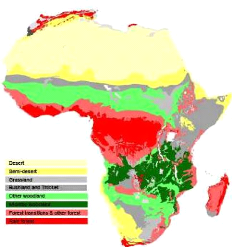
Figure 1: Répartition de la forêt claire et du Miombo en Afrique (WHITE, 1983). 3 1.2. La forêt claire zambézienne au Katanga (Miombo). La forêt claire zambézienne est la formation végétale la plus répandue du haut Katanga, elle couvre 11% du territoire national de la R.D. Congo (Timberlake & Chidumayo, 2011). Cette formation est dominée par les genres Brachystegia-Julbernardia-Isoberlinia, et parsemée par les grandes termitières à originalité floristique très élevée (Schmitz, 1971). 8500 espèces sont recensées à ce jour (Malaisse, 1997). La végétation de la plaine de Lubumbashi a déjà fait l'objet des quelques études : Sys et Schmitz (1959) ; ont démontré qu'il existe dans le Miombo Katangais une corrélation entre la structure, la texture du sol et la végétation en place. Par exemple, un peuplement à Brachystegia boehmii indique la présence dans le sol de cailloux et latérites. Schmitz (1971) a isolé au sein du Miombo Katangais différentes alliances et associations d'espèces végétales. Ces alliances et associations ont été définies en fonction de la végétation, la topographie, la texture et la couleur du sol. Il s'agit de: l'alliance Berlinio-Marquesion, (elle regroupe les types forestiers en liaison entre les forêts semi- caducifoliées et tropophiles) ; l'alliance Mesobrachystegion (ce sont des forêts tropophiles à Brachystegia sur sol argileux/ sablo-argileux et profond avec une bonne économie d'eau), l'alliance Xerobrachystegion (ce sont des latosols bruns/ ocre-jaune, moins riche en éléments, sec et profond). Au sein de ces alliances, on retrouve des sous-associations suivant la nature et les propriétés physiques et chimiques des sols. Malaisse (1997) a tenu compte des critères physionomiques et floristiques tout en ignorant les paramètres du sol. Il ressort de cette analyse que la végétation de la plaine de Lubumbashi est subdivisée en des savanes arbustives à espèces typiques et des forêts claires à Brachystegia spp. La forêt claire du Katanga se situe dans le sous-centre endémisme Katanguien, qui appartient au domaine zambézien et se partage entre un secteur lunda à l'ouest et un secteur bemba à l'est (Malaisse, 1993).Le Haut Katanga appartient au secteur de l'Est (Secteur Bemba). La majeure partie de la réserve minérale est concentrée dans les couches superficielles du sol (Chidumayo, 1997). 4 Dans ce secteur, la diversité d'espèces endémiques est élevée dans différentes familles et genres, tels que les genres Crotalaria, espèces et variétés de Brachystegia, Uapaca, Cassia, Protea, Thesium, Polygonum, ... (Schmitz, 1971). 1.2.1 Conditions environnementales La forêt claire du Katanga est sous un climat à « saisonnalité » prononcée. Celle-ci est sous dépendance non seulement des précipitations, mais également, de la température (Schmitz, 1971). Cette saisonnalité, couplée au passage du feu anthropique, influence le fonctionnement de cet écosystème (Malaisse, 1979). Au Haut Katanga, la moyenne des précipitations annuelles est de 1225 l/m2. Avec une moyenne de température de 20.0°C. La saison sèche a une durée moyenne de 5 mois (Mai-Septembre); Et la saison pluvieuse dure 4 mois (Novembre-Mars). Cela soumet la végétation à un contraste bien marqué (Schmitz, 1971). Ce pourrait être un facteur majeur qui détermine la limite sud de la distribution du Miombo dans la région zambézienne (Werger & Cootzee, 1978). La géologie du Katanga est complexe. Elle tire son origine à partir des invasions marines par accumulation de sédiments, des mouvements tectoniques violents comme la dépression « Kamalondo ». Il s'en est suivi la formation des pénéplaines et des hauts plateaux qui sont une conséquence des grands effondrements et des différentes phases d'érosions. Ces processus ont débouchés à la formation des Hauts plateaux à sable blanc (Kalahari), des collines à affleurements rocheux ou non, des plaines, et des ravins (Schmitz, 1971). Au Katanga les sols sont ferralitiques, profonds, à horizons peu contrastés et de transition graduelle. Les sols les plus fréquents sont du type ferralitique de couleur rouge et jaune, souvent associés à des lithosols sur cuirasses ferrugineuses, ou sur des sédiments meubles sableux (Duvigneaud, 1958). 5 Le Miombo des environs de Lubumbashi évolue sur des latosols zonaux subdivisés en trois séries suivant la teneur en oxyde de Fer (Schmitz, 1971) : ? Les sols rouges sur roches carbonatées et dolomitiques. ? Les sols ocres-rouges sur roches argileuses ou siliceuses. ? Les sols jaunes sur roches argileuses ou siliceuses, nappe peu profonde. 1.2.2. Dégradation et déforestation du Miombo Katangais La dégradation est perçue comme une atteinte à la qualité des milieux forestiers (Kabulu et al., 2008). La dégradation et la déforestation sont des processus très liés. Le premier terme a trait aux aspects qualitatifs, alors que le second a trait aux aspects quantitatifs. Elles sont associées à l'érosion de la diversité floristique et faunistique. Dans le Miombo, 191 espèces d'arbres sont en danger dû à la production de charbon de bois, au feu et à l'agriculture. 140.000 ha de Miombo sont perdus par année, soit un taux de 4-6% (Timberlake, 2000). Au Haut Katanga, avant les années 1910, la périphérie de la ville de Lubumbashi était couverte par le Miombo parsemé des tâches de forêts denses sèches et de forêts galeries. Les études de Keay et al. (1988), cité par Malaisse (1997), montrent qu'environ 80 % de ces forêts ont disparu ; ce qui représente environ 1000 km2 autour de la ville de Lubumbashi. L'OSFAC (Observatoire Satellital des Forêts d'Afrique Centrale), a estimé la perte du couvert à 2.3 %. Soit un taux annuel de 0.323 % entre 2000 et 2005. Plus récemment, une étude de la dynamique spatio-temporelle des forêts claires au tour de Lubumbashi a montré que la vitesse de déforestation entre 1958-2009 est de 2.4% par an (Munyemba, 2010). Les causes de cette déforestation sont multiples. Elle a été corrélée avec les densités de populations et la croissance des activités minières ( Ernst & Verhegghen, 2010) . A cela s'ajoute une urbanisation anarchique due à l'expansion démographique , l'agriculture qui entraine une savanisation du Miombo, le prélèvement abusif des essences forestières pour la fabrication de charbon de bois, le braconnage qui s'est accentué après la chute de la Gécamines (principale entreprise minière de la région) à la fin des années 1980 (Faucon, 2009) et les feux de brousse. Leur fréquence et intensité, sont deux facteurs qui influencent la Les objectifs majeurs de la restauration peuvent être appréhendés à deux niveaux (S.E.R.I.S.P.W, 2004): 6 survie des espèces et la formation de graines (Nangendo et al., 2005). Le feu a une grande influence sur le cycle de vie des diverses espèces. Lors de son passage, le feu affecte même les jeunes pousses, ce qui les rend vulnérables (Campbell, 1996). La Province s'est industrialisée à partir de 1911 avec l'expansion des activités minières au Katanga, et a connu une croissance exponentielle ces 15 dernières années. L'exploitation minière est directement liée à la déforestation. Des grandes superficies de forêts claires disparaissent chaque jour pour permettre l'accès aux gisements, et même pour l'extraction des minerais. Ce qui facilite aussi l'accès aux charbonniers à des forêts autrefois inaccessibles ! En plus, la plupart des entreprises minières du Katanga utilisent encore des techniques archaïques et non respectueuses de l'environnement. Il convient de signaler que, depuis quelques temps, les exploitants miniers tentent d'étendre leurs activités d'exploitation aux forêts faisant partie du domaine public de l'Etat (forêts classées). On place dans cette catégorie, les parcs nationaux (réserves intégrales), les domaines de chasse, les forêts urbaines, les jardins botaniques et les périmètres de reboisement de l'Etat ou des entités décentralisés (Code forestier, 2002). Le processus de restauration et de conservation de la biodiversité doit passer par une étude approfondie de la question. Seule une gestion raisonnée des écosystèmes pourra limiter le nombre d'extinction des espèces sauvages (Olivieri & Vitelis, 2001). 1.3. La restauration des groupements végétaux 1.3.1. Définition La restauration écologique est un processus qui assiste l'autoréparation d'un écosystème qui a été dégradé, endommagé ou détruit (Society for Ecological Restoration International Science & Policy Working Group., 2004). Si les écosystèmes des pays du Sud sont surexploités et dégradés, il faut retenir que toute politique d'aménagement doit avoir à l'esprit que la population locale demeure la force motrice dans les écosystèmes naturels (Aronson et al., 1995). La distribution du Miombo est bien connue. Cependant, les groupements végétaux, les différents modèles d'associations des espèces et les facteurs influençant la distribution des 7 ? Au niveau des écosystèmes : Maintenir ou accroître la productivité primaire ou secondaire et améliorer la biodiversité. ? Au niveau des paysages : Faciliter la réintégration au cas où, le paysage a été fortement fragmentés. Il est difficile de reconstituer exactement le fonctionnement des écosystèmes historiques, mais il est possible de ré-établir les espèces des groupements végétaux connues (Aronson et al, 1995). 1.3.2. Etapes de la restauration des groupements végétaux Les gestionnaires de l'environnement sont confrontés à des difficultés dans la mise en place des pratiques de restauration écologiques des écosystèmes (Bioret, 2006). La confrontation des données paléo écologiques, génétiques, associées aux caractéristiques biologiques des espèces charismatiques des groupements végétaux, conduit à améliorer les connaissances sur la réactivité future des groupements végétaux (Whattaker et al., 2005). La connaissance des conditions historiques est un point de départ important dans la détermination de la zone de référence (état dans lequel se trouvait l'habitat avant qu'il soit modifié) et de la zone à réhabiliter (Society for Ecological Restoration International Science & Policy Working Group, 2004). Les sources des données qui seront utilisées dans la délimitation de la zone de référence/zone à réhabiliter sont notamment: Les descriptions écologiques, les photographies aériennes de la végétation (anciennes et récentes), les vestiges du site qui restent à restaurer (indiquant les conditions physiques et le biotope antérieurs), les descriptions écologiques (d'écosystèmes similaires intacts), les spécimens des herbiers. Ceci permet de représenter sur un graphique la trajectoire des groupements végétaux à restaurer, à partir de données écologiques et de modèles de prédiction (Society for Ecological Restoration International Science & Policy Working Group, 2004). 8 espèces sont mal expliqués. Alors que, la gestion durable des écosystèmes du Miombo doit passer par leur compréhension. 2. OBJECTIFS L'objectif général de cette étude est de caractériser les communautés végétales de Mikembo en lien avec les paramètres édaphiques. Pour atteindre cet objectif, trois objectifs spécifiques:
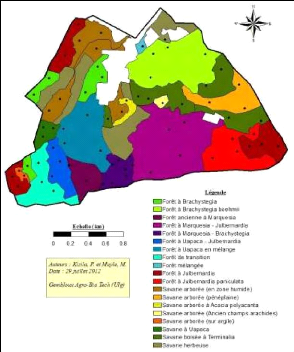
16 A cela s'ajoute trois paramètres pour la dynamique de reproduction : ? Moyenne des tiges régénérées par graine (Pgr). ? Moyenne des tiges régénérées sur souche (Pso). ? Moyenne des tiges régénérées sur tronc (Ptr). Les graphiques des distributions des tiges par classes de diamètre ont été établis pour chaque communauté végétale. Ensuite des graphiques pour les espèces qui dominent la canopée des différentes communautés végétales de Mikembo en particulier et du Miombo en général. Ce sont les espèces : Brachystegia boehmii, Julbernardia paniculata, Julbernardia globiflora, Uapaca nitida et Marquesia macroura. Ces graphiques ont permis de distinguer les espèces ayant un bon profil dans la dynamique de régénération du Miombo. Enfin, l'effet groupe a été testé sur les différents paramètres au niveau du peuplement par une analyse de la variance suivi d'une structuration de moyenne dans Minitab 16. 3.5.4. Paramètres du sol. Des relations ont été établies entre d'une part, les abondances et la couleur, la profondeur du sol; Et d'autre part, les espèces indicatrices et la couleur, la profondeur du sol et de la charge. En enfin, un test de khi carré suivi de la structuration de moyenne a été effectuée dans Minitab 16. Pour voir si la répartition de placettes dans différentes communautés est fonction des paramètres édaphiques mesurés. 17 4. RESULTATS. 4.1. Carte de végétation et inventaires. La Figure 5, illustre les 19 formations végétales, qui occupent 413 ha sur 442 ha. Les points noirs sont les 69 placettes projetées aléatoirement.
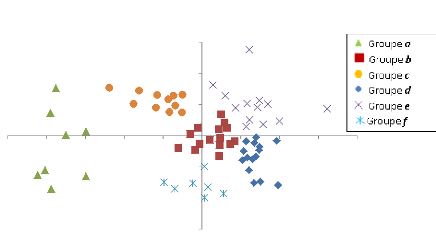
Figure 5: La carte de végétation préliminaire de Mikembo. 4.2. Caractérisation des communautés. La Figure 6, montre la première ordination qui a révélé 6 groupes de placettes. Les savanes arborées sont reprises dans le groupe f, les zones humides dans le groupe c et les peuplements à Brachystegia boehmii dans le groupe e. Les groupes a, b et d sont mélangés.
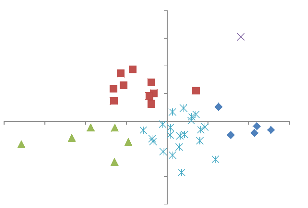
18 0,5 0 -2,5 -2 -1,5 -1 -0,5 0 0,5 1 1,5 2 NMDS1 -0,5 -1 -1,5 1,5 1 NMDS2 Figure 6: Les six groupes issus de la première ordination réalisée sur 69 placettes. Le tableau 3, reprend les placettes dans différents groupes définit par la première ordination. Toutes les savanes boisées sont reprises dans le groupe f, les savanes arborées dans le groupe c et les peuplements à Brachystegia boehmii dans le groupe e. Dans le groupe d, on trouve des nombreuses formations à Julbernardia, des Marquesia et Brachystegia, Les groupes a, b et d sont moins différenciés. Tableau 3: La répartition des placettes dans différents groupes après la première ordination. Groupe a Groupe b Groupe c Groupe d Groupe e Groupe f BR3 BCA1 PN1 JCA1 BBC1 AP1 JN2 BN1 PN2 JCA2 BB CA1 JSE3 BN2 PN3 JN1 BBC3 CA2 MA2 BR2 TL1 JP1 BBC4 FM1 MB4 MA1 ZHL1 JP2 BR1 TB1 MJ1 MA3 ZHR1 JP3 TN1 TB2 MJ2 MA4 ZHR2 JP4 TN2 TB3 MJ3 SU1 ZHR3 JSE1 TBBC1 UJ1 SU2 JSE2 TMB1 UJ2 SU3 MB1 TS1 UM3 TS2 MB2 ZHN1 UM4 UJ3 MB3 ZT1 UM1 MJ4 ZT2 UM2 ZT3 ZHN2 Une deuxième ordination a été effectuée avec les placettes des groupes qui ce sont révélés très hétérogène (groupes a, b et d). Soit un total de 43 placettes. Le résultat de cette deuxième analyse présente une distribution des placettes en 4 groupes et une placette isolée. 19 Le groupe 1 comprend les placettes des forêts de transition, le groupe 2, reprend les peuplements à Julbernardia spp., alors que le groupe 3, regroupe toutes les placettes des zones matures à gros fûts. Le groupe 4, est constitué du reste des placettes forestières. Le groupe 5 est constitué d'une placette isolée qui a été supprimée.
2 NMDS2 1,5 1 0,5 Groupe 1 Groupe 2 Groupe 3 Groupe 4 Placette isolée 0 -2 -1,5 -1 -0,5 0 NMDS1 0,5 1 1,5 -0,5 -1 -1,5 Figure 7: Les 5 groupes issus de la deuxième classification effectuée sur les groupes a; b ; d (Nombre de placettes : 43). Les groupes a, e et i de la première classification, et le groupe 4 de la seconde classification ont été remis, pour une troisième ordination. Les résultats de cette analyse ont montrés que, mise à part le groupe e (zones humides) de la première analyse, séparer ces placettes étaient inutile parce qu'ils reflètent la réalité de terrain. La distribution définitive des placettes dans différents groupes est reprise dans le tableau 4. Le groupe F est constitué des placettes qui se situe dans la savane, le groupe G, est constitué des placettes en zone humide, le groupe I, a été exclu parce que c'est une placette isolée des autres groupes. 20 Tableau 4: La distribution des placettes dans différents groupes après la troisième ordination Groupe Groupe Groupe Groupe Groupe Groupe Groupe Groupe Groupe A B C D E F G H I BR3 JCA1 BR2 AP1 BCA1 SU1 PN1 ZHL1 BBC1 JN1 UM4 JCA2 MA3 CA1 BN1 SU2 PN2 ZHR1 BB ZT1 JP1 MB1 CA2 BN2 SU3 PN3 ZHR2 BBC3 ZT2 JP2 MB2 FM1 JN2 TS2 TL1 ZHR3 BBC4 ZT3 JP3 MB3 TB1 JSE3 UJ1 BR1 JP4 UM1 TB2 MA1 UJ2 TN1 JSE1 TB3 MA2 UJ3 TN2 JSE2 TBBC1 MA4 UM2 JN1 MJ3 TMB1 MB4 UM3 MJ4 TS1 MJ1 ZHN2 ZHN1 MJ2 Pour les groupes restant, les résultats sur l'abondance et les espèces indicatrices dans différentes strates (arbustes-arborée dominé-arborée dominante) ainsi que la régénération est repris dans les tableaux 5 et 6. Le tableau 3, reprend les abondances des espèces à différentes strates et dans la régénération dans les 8 communautés végétales. Au niveau du peuplement : On remarque que U. nitida est plus abondante dans le groupe 1, avec 39.6%. Dans les groupe 2 c'est J. paniculata (43.1%) ; P. tinctorius (13.5%) dans le groupe 3 ; C. adenogonium (36.9%) dans le groupe 4 ; J. globiflora (26.2%) dans le groupe 5 ; Acacia spp. (36.9) dans le groupe 6 ; A. polyacantha (89.6%) dans le groupe 7 ; et B. boehmii (62.5%) dans le groupe 8. Au niveau des strates : Les deux premiers groupes sont dominés par U.nitida, le groupe 3 par P. tinctorius ; le groupe 4 : C. adenogonium, groupe 5 : J.globiflora ; groupe 6 : Acacia sp. Le groupe 7 : A polyacantha. Et le groupe 8 : B.boehmii. Au niveau de la régénération les espèces les plus dominantes par ordre de numéro des groupes : U.nitida, J. globiflora, D. boehmii, D. boehmii, J. globiflora, A. venosum, A. polyacantha, B.boehmii. 21 Tableau 5: Les abondances des espèces par strates, par peuplement et dans la régénération pour les 8 communautés végétales (Abod : Abondance).
22 Le Tableau 6, montre les espèces indicatrices dans le peuplement, dans différentes strates et dans la régénération. Au niveau du peuplement, le groupe A, a quatre espèces indicatrices, le groupe B : J. paniculata, dans les groupes C et F : il n'y a pas des espèces indicatrices. Le groupe D : C. adenogonium et Terminalia sp. Le groupe E : J. globiflora. Le groupe G : A. polyacantha. Le groupe H : B. boehmii et B. longifolia. Au niveau de différentes strates ce sont les genres : Uapaca, Monotes, Julbernardia, Annona, Combretum, piliostigma, Acacia, Brachystegia. Alors que dans la régénération on a les genres : Monotes, Uapaca, Parinari, Julbernardia, Dalbergia, Combretum, Piliostigma, Brachystegia, Pseudolachnostylis, Plyllocosmus. Tableau 6: Les espèces indicatrices par strate, par peuplement et dans la régénération pour les 8 communautés végétales.
23 Suite à cette première analyse, le Tableau 7 illustre la nouvelle classification des groupes: 8 communautés ont été créées sur base de la structure et de la composition floristique. On a trois forêts hétérogènes, deux savanes arborées, une savane boisée, forêt de régénération, forêt ancienne. Tableau 7: Reclassification des communautés végétales tenant compte de la structure et de la composition floristique.
4.3. Structure des communautés. Les Tableau 8 et 9, montre qu'il existe des différences significatives entre communautés pour différents paramètres dendrométriques. Les valeurs de p sont inférieures à 0,05. Alors que pour la hauteur dominante il n'y a pas des différences significatives p=0.077. Il existe des différences significatives entre communautés pour tous ces paramètres (p<0.05), sauf le pourcentage de régénération par tige (p=0.43). Lorsqu'on observe le Tableau 8, les forêts hétérogènes (B. boehmii, J. globiflora, J. paniculata), et la forêt ancienne, enregistrent des surfaces terière entre 13.5 m2/ha et 16.1 m2/ha. Cette gamme de valeurs relève de la forêt claire. Alors que dans les savanes, deux gammes de valeurs peuvent être distinguées: 3.2 m2/ha pour les deux savanes à Acacia sp. et 5.3 m2/ha, pour la savane à Combretum et Terminalia. 24 Tableau 8: Différents paramètres de structure dans différentes communautés végétales. La valeur sous parenthèse représente l'erreur standard, les lettres représentent les groupes.
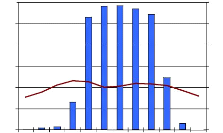
25 4.4. La distribution de tiges par catégorie de diamètre dans différentes communautés végétales. La figure 8, montre la distribution de fréquence dans la forêt de régénération à Uapaca. La distribution est asymétrique. La classe 5, a moins d'individus. La classe 10 enregistre plus d'individus que toutes les autres. 3500 3000 2500 NHA 2000 1500 1000 500 0
5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 Classe de diamètre Figure 8: La distribution des tiges par classe de diamètre dans la forêt de régénération à Uapaca sp. La figure 9, montre la distribution de fréquence d'Uapaca nitida dans la forêt de régénération à Uapaca sp. qui présente une distribution asymétrique. Il n y a pas d'individus dans les classes 25, 30, 35.
Classe de diamètre Figure 9: La distribution de tiges par classe de diamètre d'U. nitida dans la forêt de régénération à Uapaca sp. 26 La figure 10, montre la distribution de tiges par classe de diamètre dans la forêt hétérogène à J. paniculata, environs 6000 tiges/ ha, alors que la classe de recrutement n'a qu'environ 4900 tiges/ha. 7000 6000 5000
2000 1000 0
5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 Classe de diamètre Figure 10: La distribution de fréquence dans la forêt hétérogène à J. paniculata. La figure 11, illustre la distribution de tiges par catégorie de grosseur de J. paniculata dans la forêt hétérogène à J. paniculata. La classe 5 enregistre environs 1700 tiges/ha. La classe 10 a environs 2800 tiges/ ha. 3000 2500 2000 NHA 1500 1000 500 0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 Classe de diamètre Figure 11: La structure de J. paniculata dans la forêt hétérogène à J. paniculata. 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 27 La figure 12, montre la distribution de tiges par catégorie de diamètre dans la forêt ancienne à P.tinctorius. La structure est inéquiènne avec des ondulations.
2000 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 Classe de diamètre Figure 12: La structure de la forêt ancienne à P. tinctorius. La figure 13. Montre la distribution de tiges par catégorie de grosseur dans la savane boisée à C. adenogonium. Dans cette communauté, le nombre de tiges dans la classe 10 est plus élevé. Par contre le nombre de tiges dans la classe 5 semble relativement faible. Entre la classe 30 et la classe 55, il y a un vide.
Classe de diamètre Figure 13: La structure de la savane boisée à C. adenogonium. La figure 14, représente la distribution de fréquence de la forêt hétérogène à J. globiflora qui est inéquiènne comme toute structure naturelle. Mais il convient de signaler que dans les classes 60 ; 65 ; 85 et 95, il y a absence d'individus.
28 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 Classe de diamètre Figure 14: La structure de la Forêt hétérogène à J.globiflora. La figure 15, illustre la distribution de tiges de M.macroura par catégorie de grosseur dans la forêt hétérogène à J. globiflora. M. macroura présente une distribution dont plusieurs classes manquent d'individus. Cela s'observe surtout à partir de la classe 40 jusqu'à la classe 100. Donc les individus âgés. Par contre, on observe une régularité d'individus entre les classes 5-35.
5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 Classe de diamètre Figure 15: La distribution de tiges par classe de diamètre pour M. macroura. La figure 16, montre la distribution de tiges par catégorie de grosseur de J. globiflora dans la forêt hétérogène à J globiflora. 29
NHA 4500 4000 5000 3500 3000 2500 2000 1500 1000 500 0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 Classe de diamètre Figure 16: La structure de J. globiflora. La figure 17, illustre la distribution de fréquence de la forêt hétérogène à B.boehmii. La structure est asymétrique à gauche. La classe 10 contient environs 3500 tiges/par. Alors que la classe 5 étant considéré comme la classe de recrutement n'a qu'environs 2300 tiges/ ha. 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 4000 3500 3000 2500 NHA 2000 1500 1000 500 0 Classe de diamètre Figure 17: La structure de la Forêt hétérogène à B. boehmii. La figure 18, illustre la distribution de tiges par catégorie de grosseur de l'espèce B.boehmii dans la forêt hétérogène à Brachystegia. La structure reste asymétrique comme pour la forêt hétérogène à B. boehmii. 30
NHA 2500 2000 1500 1000 500 0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 Classe de diamètre Figure 18: La structure de l'espèce B. boehmii dans la Forêt hétérogène à B. boehmii. La figure 19, montre la distribution de tiges par catégorie de diamètre dans la savane arborée à Acacia spp. La classe de recrutement enregistre peu de tiges. La classe 10 renferme environs 220 tiges.
5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 Classe de diamètre Figure 19: La structure de la savane arborée à Acacia spp. Le Nombre de tiges /ha des savanes arborées à Acacia spp.est faible, le maximum de tiges se concentre dans la classe 10, suivi de la classe 15. Par contre la classe 5 qui est considérée comme la classe de recrutement enregistre peu de tiges. La figure 20, illustre la distribution de tiges par classe de diamètre en savane arborée à A. polyacantha. 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 31
Le NHA est relativement faible, environs 190 tiges/ ha pour la classe 15, suivi de la classe 10 qui enregistre environs 160 tiges/ ha. Il convient de signaler que la classe 40 n'a pas d'individus. Environs 20 tiges dans la classe 45.
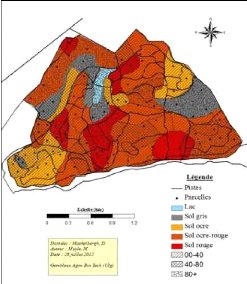
Classe de diamètre Figure 20: La structure de la savane arborée à A. polyacantha. 4.5. Facteurs édaphiques Le Tableau 10, montre la répartition des placettes suivant la couleur du sol. Le test Khi carré a montré que la couleur du sol, influence moins la répartition des placettes dans différentes communautés végétales (Khi carré: 64,789; dl: 24; p=0.07). La valeur de p (p-value) est une approximation qui n'est probablement pas valide. Parce qu'il y a beaucoup de vides. Les forêts hétérogènes à B. boehmii, à J. globiflora et paniculata, ainsi que la forêt de régénération se situent sur sol ocre, ocre-rouge, et rouge. 100% des savanes sont sur sol Gris. Par contre, la forêt à P.tinctorius et La savane boisée à C. adenogonium semblent n'est pas avoir des préférences. La forêt de régénération à Uapaca spp. semble s'établir sur sol ocre et ocre-rouge. Les trois forêts hétérogènes sur sol ocre-ocre rouge et rouge. Tableau 10: Répartition des placettes en fonction des couleurs du sol.
32 Le Tableau 11, illustre la répartition des placettes par groupe selon les différentes profondeurs de sol. Les sols les plus profonds hébergent 100 % des placettes de la savane arborée à Acacia spp. Le teste Khi carré a montré qu'il existe une agrégation des placettes en fonction de différentes profondeurs du sol (Khi carré: 27,681; dl: 16; p : 0,035). La savane arborée à Acacia polyacantha est absente sur sol moins profond, alors que la forêt de régénération et celle à B. boehmii sont aussi manquants sur sol plus profond. Les autres n'ont pas une spécificité. Tableau 11: Influence de la profondeur du sol sur la répartition de placettes.
4.6. Influence de la charge caillouteuse Le Tableau 12, montre les espèces indicatrices de la charge caillouteuse dans différentes strates ainsi que dans la régénération. La présence de la charge caillouteuse a peu d'influence sur la répartition des placettes. Presque toutes les espèces sont indicatrices des sols avec charge. Par contre ; seule M. katangensis a présenté une valeur indicatrices de 27% pour les sols sans charge. M. macroura et J. paniculata sont des espèces indicatrices de la charge dans la strate arborée dominante et arborée dominée. M. macroura est absente dans la régénération. Alors que J. paniculata est présente. B.boehmii dans les strates arborée dominée et arbustive. Uapaca spp. est présent dans la régénération et dans la strate arbustive. 33 Tableau 12: Espèces indicatrices en fonction de la présence ou absence de la charge caillouteuse
34 Le Tableau 13, montre que M. macroura est un indicateur des sols peu profond. Cette espèce a une valeur indicatrice de 34.6% dans le peuplement, et 34.1%, dans la strate dominante, 28.7% la strate arborée, ainsi que dans la régénération 40.5%. Pour les sols profonds (40-80cm), C. adenogonium apparait comme l'espèce indicatrice, parce qu'elle se retrouve dans différentes strates. Mais absente dans la régénération. Les sols plus profonds (80 cm et plus), enregistrent le genre Uapaca au niveau du peuplement (U. sensibarica, U. nitida, U. pilosa), dans différentes strates, et même dans la régénération (U. nitida). Les sols plus profonds semblent ne pas avoir une spécificité en espèce. Tableau 13: Les espèces indicatrices suivant différentes profondeur du sol.
35 5. DISCUSSION. 5.1. Discussion de la méthodologie. La technique des relevés temporaires aléatoire a permis la collecte des données sur la flore du sanctuaire Mikembo. 69 placettes de 18 m de rayon ont été distribuées sur une superficie de 442 ha. Soit un taux de sondage de 1.7%. Dans le souci d'augmenter la précision, l'idéale aurait été de diminuer la taille de la placette, mais, augmenter leur nombre pour ce même taux de sondage (Rondeux, 1999). Pour améliorer la qualité de la carte de végétation, il était important d'utiliser les relevés systématiques (Rondeux, 1999). Mais cependant, les relevés aléatoires se prêtent mieux dans les analyses statistiques parce qu'ils attribuent à tous les individus la même probabilité de faire partir de l'inventaire. La taille du relevé est variable et fonction de la diversité en espèce suivant la région d'étude. Dans la région zambézienne cette taille est de 1000 m2 (Jayakumar et al., 2011 ) et le diamètre de pré-comptage est généralement de 10 cm, pour la strate arborée (Sokpon et al., 2006). On pourrait aller en dessous surtout s'il s'agit des études de la dynamique de régénération comme l'a proposé Hakizimana (2012). Ce pourquoi dans le cas présent, le diamètre de pré-comptage a été fixé à 2 cm. De plus, vus les objectifs poursuivis, ce diamètre de pré-comptage a facilité l'inventaire de la flore globale de la zone d'étude comme le confirme certains auteurs dans la région zambézienne (Banda et al., 2006 ; Schwartz & Caro, 2003). Pour des études de la dynamique des communautés végétales, il serait intéressant de faire un suivi temporel de placettes permanentes, ce qui n'a pas été possible par manque de temps. Ce pourquoi, les résultats de cette étude ce sont basés sur une approche indirecte prenant en compte les comparaisons de composition floristique entres différents stades au sein des groupements végétaux, tout en considérant uniquement les ressemblances floristiques entre groupements végétaux. Les analyses sur les relations sol-communautés végétales, seront discutées avec prudence. Vu que la carte de sol utilisée dans cette étude reste encore provisoire. Elle résulte d'une extrapolation de 53 points de sondage réalisé sur une superficie de 442 ha. La concession de Mikembo renferme une hétérogénéité de la végétation sur une petite zone. Les résultats de l'inventaire floristique sur l'ensemble de 442 ha ont révélés 80 espèces 5.2. Hétérogénéité de la végétation. 36 (Tableau 17 en annexe). Ces résultats avoisinent la richesse en espèce aperçue par Backeus et al., (2005) : 86 espèces ; Luoga (2002) : 79 espèces ; dans d'autres réserves forestières du Miombo de superficie similaire. Après analyse par ordination, 8 communautés végétales ont été définies sur base de critères objectifs. Il s'agit de : La forêt hétérogène à B.boehmii, à J. paniculata, à J. globiflora, la forêt de régénération à Uapaca sp., la forêt ancienne à P. tinctorius, la savane à Acacia sp., la savane à A. polyacantha et la savane boisée à C. adenogonium (Tableau 7). La forêt ancienne à Pterocarpus a comme espèces indicatrice : Diplorhynchus condilocarpon dans la strate arborée et Dalbergia boehmii dans la régénération. La forêt hétérogène à B.boehmii, renferme B. boehmii, comme espèce abondante et indicatrice, et quelques Uapaca dans la régénération. Alors que la forêt à J. paniculata a comme espèce indicatrice : J. paniculata et Diplorhynchus condilocarpon. Ces communautés appartiennent à l'ordre de Julbernardio-Brachystegietalia spiciformis dans l'alliance Mesobrachystegion en accord avec la description des groupements végétaux décrits par Schmitz (1971). Trois communautés végétales ont des surfaces terrières entre 13m2/ha et 16 m2/ha. Pour la savane boisée, 5.3 m2/ha, les savanes arborées, 3.2 m2/ha (Tableau 8). Ces valeurs coïncident avec celles déterminées par François Malaisse (communications personnelles F. Malaisse). 5.3. Caractérisation des communautés végétales et la dynamique de régénération. La plus part des espèces les plus abondantes au niveau du peuplement et même dans différentes strates ont été trouvées aussi dans la régénération (tableau 5). La densité de la régénération (dhp < 2 cm) varie entre 27 tiges/hectare (Savane arborée à A. polyacantha) et 2105 tiges/hectare (Forêt hétérogène à B. boehmii). Ceci montre une grande variabilité en termes de paramètres dendrométriques. Comme on a peu d'information sur l'histoire récente des communautés végétales à Mikembo, on pourrait se servir des paramètres dendrométriques mesurés et de la structure des peuplements pour tenter d'expliquer la dynamique de la régénération, le degré de perturbation, ainsi que l'évolution de différentes communautés végétales. (Hakizimana, 2012). L'allure générale de la distribution de fréquence dans différentes communautés végétales dans la concession de Mikembo reste une courbe en « J » inversé comme dans tout peuplement naturel. Mais il convient de signaler quelques anomalies sur la plus part des 37 structures des communautés végétales. La forêt hétérogène à B.boehmii (Fig. 17), montre une structure dont la classe de recrutement (5 cm) a 2300 tiges, alors que la classe de 10 cm enregistre 3400 tiges/ha. Et lorsqu'on observe le tableau 9. Environ 50 tiges se développent sur souche et 20 sur tige. Comme le sol de cette communauté est parsemé des affleurements rocheux, il est donc difficile de cultiver dans cette zone ; par contre, ceci serait expliqué par une exploitation de bois dans le passé comme le confirme aussi Schmitz (1971). Pour la communauté à J. globiflora, on remarque quelques troués dans les classes supérieures (Fig.14). La composition floristique de la communauté renseigne que, M. macroura présente les arbres les plus gros dans cette communauté parce que son bois est difficile à exploiter (Smith & Allen, 2004). En effet, les plus gros individus de M. macroura résistent mal aux orages (observations personnelle). Malgré cela, sa régénération est difficile dans un peuplement fermé (Schmitz, 1971). La structure de M. macroura dans cette communauté à P. tinctorius confirme cette hypothèse (Fig. 15). On pourrait dire que la chute régulière des gros individus de M. macroura serait à la base de ces vides qui, par contre favoriserait l'installation des jeunes individus de J. globiflora dont la structure est dominée par les tiges de petit diamètre dans la même communauté(Fig. 16). La structure de la savane boisée à C. adenogonium est inéquiènne. Toutefois, cette communauté enregistre 77.5% de tiges sur souches (Tableau 9). Avec un faible nombre des tiges par ha dans la régénération (Tableau 8). On pourrait supposer que ces zones furent occupées par des anciens villages grâce à sa proximité au cours d'eau, et que les espèces se régénèrent mal. La structure de la savane arborée à Acacia spp. et à A. polyacantha montre que la classe de recrutement à peu de tiges/ ha (Fig. 19 et 20). Mais ces deux conditions semblent avoir différentes origines. Dans la savane à Acacia spp., en moyenne 25 tiges/ha se développe à partir des graines contre 75 tiges sur souche. Alors que dans la savane à Acacia polyacantha, 80 tiges/ha proviennent des graines, contre 18 tiges/ha régénère par souche. Pour A. polyacantha ce blocage serait dû à la difficulté que les graines ont pour germer, étant donné qu'il se développe sur sol engorgé d'eau de façon permanente; Alors que pour Acacia spp., il semble que cette partie était occupée par des anciens champs qui pour leur installation, des quantités énormes de bois ont été coupées tout en laissant des souches. A ce jour, cette communauté serait en train de se régénérer à partir des souches abandonnées sous terre. La 38 régénération par graine constitue environ 25% seulement (Tableau 9). Ce sont donc des peuplements sans avenir parce que la classe de recrutement est faiblement approvisionnée par germination de graine. La structure de la forêt claire de régénération à Uapaca sp., semble asymétrique gauche. Malgré cela, la communauté semble inéquiènne bien que la classe 5 a moins d'individus que la classe 10 (Fig. 8). Par contre l'espèce U. nitida enregistre très peu d'individus dans la classe de recrutement et un vide anormal dans les classes 25-30-35 (Fig. 9). La moitié de tiges ce sont développées sur souche et une autre moitié par graine (Tableau 9); Bien que la communauté ait présentée des signes de dégradation dans le passé (présence des billons). Ceci s'expliquerait par le fait que, les Uapaca, ont une capacité de se régénérer par rejet des souches et se développent bien en condition naturelle grâce aux associations avec les champignons (Ramanankierana, 2006). 5.4. Influence des paramètres du sol sur la répartition des placettes dans différentes communautés végétales. Le test khi carré a montré que la couleur du sol a moins d'influence sur la répartition des placettes (Tableau 10). Cette répartition serait contrôlée par d'autres facteurs. Mais cependant, le même test a montré une différence significative entre la répartition des placettes dans différentes communautés en fonction de la profondeur du sol (Khi carré : 27.681; dl: 16; p= 0.035) (Tableau 11). Alors que, des nombreuses études ont montrée des relations positives entre la couleur, la profondeur du sol et la végétation en place (Schmitz, 1971; Lawton 1978; Smith & Allen, 2004). Néanmoins, à partir des Tableaux 10 et 11, on pourrait tirer des informations intéressantes sur les communautés végétales qui ont la totalité de leurs placettes sur un même type de sol. Telle que la savane à Acacia polyacantha, et à Acacia sp. Il semble que A. polyacantha et acacia sp., ont des préférences sur sols de couleur gris (Tableau 10). Acacia polyacantha étant une espèce qui colonise les zones alluvionnaires périodiquement inondées (Luoga et al., 2004), la présence de l'eau presqu'en permanence dans le sol de cette communauté pourrait justifier le choix de cette espèce au détriment des autres. La présence de la charge caillouteuse a révélé des espèces telles que : J. paniculata, dans la strate dominante; Dalbergia boehmii dans la strate dominée; Ekebergia bengwelensis, dans la 39 strate arbustive. Et dans la régénération on retrouve, Phyllocosmus lemaireanus et Pseudolachnostylis maprouneifolia, M. macroura. Alors que l'absence de la charge n'a révélé qu'une espèce : Monotes katangensis (Tableau 12). Toutes les espèces indicatrices de la charge se retrouvent dans la communauté à P. tinctorius. 6. CONCLUSION ET PERSPECTIVE Cette étude s'inscrit dans le cadre de la contribution à la gestion durable des écosystèmes forestiers (Miombo) au Katanga. La croissance démographique enregistrée dans cette province au cours de la dernière décennie, avec comme conséquence, augmentation de l'exploitation des ressources forestières, principalement le bois, a causé la perte et la fragmentation des habitats naturels. Cette première étude de la végétation de Mikembo a identifié huit communautés végétales sur base des critères objectifs. Il existe des relations positives entre, la profondeur du sol et les communautés en place. Par contre la couleur semble indépendante des communautés végétales dans le cas présent. Les trois facteurs édaphiques choisis pour caractériser le sol s'avèrent approximatifs pour un domaine aussi vaste et complexe comme le sol. Cette étude devra être répétée dans une zone plus étendue et pendant deux périodes de l'année tout en intégrant les aspects chimiques du sol, pour comprendre au mieux les relations entre facteurs édaphiques et communautés végétales en place. Les analyses chimiques proposées sont : Les bases échangeables, la teneur en Fer mobile, l'Azote, le Phosphore. L'écologie de la régénération des espèces du Miombo en condition naturelle est mal connue. Ce qui rend difficile l'adoption d'une stratégie de gestion pour des zones plus ou moins intactes ou la restauration des zones fortement fragmentées. Parmi les 5 espèces emblématiques du Miombo retenues dans cette étude, J. paniculata et J. globiflora, semble avoir un bon profil de régénération vu leur structure diamétrique (en « j » inversé) en condition naturelle. Alors que, la régénération de Marquesia macroura, Uapaca nitida et Brachystegia boehmii, semble difficile. Il serait important d'effectuer des tests de germination de ces espèces en condition contrôlée et in situ. Et comme ce sont en majorité des 40 espèces ectomicorhizées, des études d'identifications d'inoculum spécifiques devraient être menée en marge de cette étude. En suite comparer l'efficacité de ces deux conditions. 41 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||