3 Hiérarchisation des paramètres
descriptifs de l'anisotropie élastique du
bois normal
Le chapitre 1 a permis de dresser l'inventaire, via une
étude documentaire pluridisciplinaire, des paramètres
essentiels rendus responsables de la variabilité intra arbre de
l'anisotropie élastique du bois normal. Les valeurs
numériques assignées à chacun de ces mêmes
paramètres, via une technique simple d'optimisation
associant calculs et propositions bibliographiques, ont conduit à
la définition d'un résineux standard virtuel RSV (chapitre 2).
Cette essence fictive a été considérée comme «
médiane » en terme de propriétés mécaniques
élastiques. L'objectif de ce chapitre est de préciser
numériquement, en s'appuyant sur des banques de données
structurelles ou des tendances générales issues de la
bibliographie, l'influence de chacun des paramètres clés,
sur les rapports d'anisotropie élastique du bois massif sans
défaut.
3.1 La porosité, facteur de l'anisotropie
élastique du réseau cellulaire nid d'abeille
Par soucis de simplification, et devant la
disparité des valeurs des rigidités de parois cellulaires
présentées dans la littérature, les études
numériques sont menées dans ce chapitre, sous l'hypothèse
d'une isotropie transverse de la double paroi (ETp = ERp).
Dans la modélisation présentée
précédemment (chapitre 2), la masse volumique (stabilisée
à l'humidité de référence de 12%) est liée
aux paramètres géométriques tissulaires
épaisseurs
(eT, eR) et diamètres (DR, DT) comme à la masse
volumique de la matière ligneuse par les deux expressions (2.12) et
(2.13) données dans le chapitre 2.
A cette première étape de la discussion, les
épaisseurs pariétales, radiale et tangentielle
(posées égales à e), ainsi que les
diamètres radial DR et tangentiel DT, ne seront pas distingués
(DR = DT = D). Le facteur de forme des cellules (supposées
en premier lieu carrées) est alors exprimé par le rapport
adimensionnel e/D.
Les rapports directionnels d'anisotropie élastique L/R et
L/T du tissu ligneux élémentaire sont
alors liés aux rapports d'anisotropie élastique
ELp
ERp
de la double paroi par les relations (3.1),
(3.2) via les coefficients d'anisotropie définis par les
relations (3.3) à (3.5):
(3.1)
E L
E R
e
= 2 1
D
E Lp
=
E Rp
a LR
E Lp
E Rp
E L
e
=1 =e (D 2e)3
E Lp
E Lp
(3.2)
= 2 1
D
E T
1 +
64 D
e3
E Tp
= a LT
E Tp
(3.3)
1 e
=
a F
64 D
(D 2e)3
e3
63
Hiérarchisation des paramètres descriptifs de
l'anisotropie élastique du bois normal
(3.4)
a LR
e
2
= 1
D
e
1 e (D 2e)3
(3.5)
a LT
= 2 1 1 +
=
3
a LR (1 + a F )
D
64 D e
Le rapport aF représente ainsi la
proportion d'anisotropie élastique L/T résultant de la mise
en
flexion de la double paroi cellulaire, tandis que le rapport
aLR fournit une estimation de la proportion d'anisotropie élastique
L/R résultant du renfort radial crée par l'alignement des
parois dans le sens R. Le produit aLT exprime enfin la proportion
d'anisotropie élastique résultant d'un alignement dans le sens
T.
Il convient ici de préciser qu'à cette
étape du travail, la discussion fait état de sens R et de sens T
et non des directions d'orthotropie radiale et tangentielle du matériau
bois, tout ceci afin de caractériser l'influence des
différences disposition en alignement (sens T) ou en quinconces
(sens T) du tissu ligneux sur les caractéristiques du nid d'abeille.
Considérant un échelonnement des masses volumiques
du matériau bois compris entre 0,2 et
1,5 (masse volumique de la matière ligneuse), il est
possible, pour différentes porosités, de déduire les
valeurs du facteur de forme e/D des cellules, supposées, en premier
lieu, carrées
(Tableau 3.1).
On calcule les valeurs du coefficient
d'anisotropie géométrique aLR correspondant à
l'arrangement cellulaire d'alignement sens R, à l'aide de (3.4),
ainsi que les valeurs du coefficient d'anisotropie
géométrique, aLT correspondant à l'arrangement cellulaire
d'alignement sens T (3.5). On accède enfin au coefficient sans dimension
aF à l'aide de (3.3), représentant la proportion d'anisotropie
résultant de la mise en flexion de certains éléments de
la paroi cellulaire. Les valeurs numériques
correspondantes sont reportées dans le Tableau
3.1.
Un premier examen du Tableau 3.1 montre que la porosité Po
varie dans un large intervalle de
86% à 1%, lorsque les masses volumiques ñ de la
structure considérée sont comprises dans l'intervalle 0,2
à 1,5 g/cm3. A une porosité quasi nulle correspond
bien une masse volumique proche de la valeur admise pour la matière
ligneuse (ñm= 1,51 g/cm3).
Les résultats affichés dans le Tableau 3.1,
permettent de conclure que, dans le cadre de l'étude
(i.e. cellules carrées), la porosité
intervient, via le rapport de forme e/D, dans l'anisotropie
élastique apparente du matériau bois assimilé à une
structure nid d'abeille.
Dans le cas des arrangements en alignement suivant la
direction R, la porosité Po intervient directement (Po = (aLR
-1)2) par l'intermédiaire du coefficient multiplicatif aLR
dont la valeur numérique reste comprise dans l'intervalle de 1 à
2.
Dans le cas des arrangements en quinconces faisant
intervenir un comportement en
« flexion » de certains éléments de la
paroi cellulaire (portion BCE soumise à une flexion 3 points, Figure 2.3
du chapitre 2), on constate (Tableau 3.1) d'une part que le coefficient aLT
varie dans une plus large gamme allant de 23 à 1, et d'autre
part que des anisotropies classiques L/T, exprimées par une valeur
du facteur aLT de l'ordre de 20, (valeurs courantes
64
Hiérarchisation des paramètres descriptifs de
l'anisotropie élastique du bois normal
dans le bois initial et final, Bodig et al, 1982, comme
dans le bois massif, Guitard et al 1987),
ne peuvent être atteintes que pour des masses volumiques
inférieures ñ à 0,25 g/cm3 (Figure
3.1). Cette dernière valeur de masse volumique
apparaît bien restrictive en regard de l'intervalle des variations
de cette même grandeur classiquement observable sur la majorité
des essences résineuses (Venet et al, 1986, Keller, 1999).
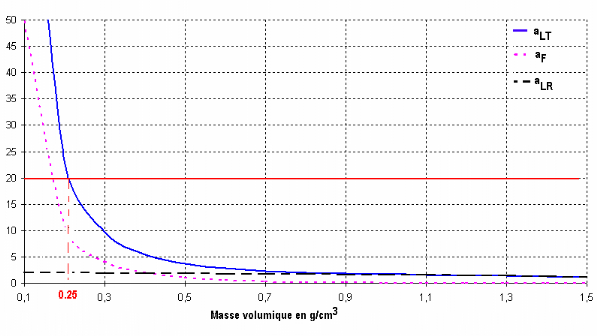
Figure 3.1 Evolutions des rapports
géométriques avec la masse volumique ñ
|
(g/cm3)
|
0,2
|
0,3
|
0,4
|
0,5
|
0,6
|
0,7
|
0,8
|
0,9
|
1
|
1,1
|
1,2
|
1,3
|
1,4
|
1,5
|
|
Po (%)
|
0,86
|
0,80
|
0,73
|
0,66
|
0,60
|
0,53
|
0,47
|
0,40
|
0,33
|
0,27
|
0,20
|
0,13
|
0,07
|
0,01
|
|
e/D
|
0,03
|
0,05
|
0,07
|
0,09
|
0,11
|
0,13
|
0,16
|
0,18
|
0,21
|
0,24
|
0,27
|
0,31
|
0,37
|
0,46
|
|
aF
|
10,73
|
4,07
|
1,93
|
1,03
|
0,58
|
0,34
|
0,20
|
0,12
|
0,06
|
0,03
|
0,01
|
0,01
|
0,00
|
0,00
|
|
aLR
|
1,93
|
1,89
|
1,85
|
1,81
|
1,77
|
1,73
|
1,68
|
1,63
|
1,58
|
1,52
|
1,45
|
1,37
|
1,26
|
1,08
|
|
aLT
|
22,67
|
9,62
|
5,45
|
3,69
|
2,81
|
2,32
|
2,02
|
1,83
|
1,69
|
1,57
|
1,48
|
1,38
|
1,27
|
1,08
|
Tableau 3-1 Comparaison de l'évolution des
coefficients d'anisotropie avec la porosité Po
L'étendue de l'intervalle de variations du coefficient
d'anisotropie aLT pourrait laisser penser que le fonctionnement en flexion
des parois cellulaires est une des raisons de la très forte
anisotropie L/T observée expérimentalement sur le bois initial
comme sur le bois final.
Pourtant, d'après la Figure 3.1, construite à
partir des données présentées au Tableau 3.1, la prise en
compte de la flexion des parois, traduite par la valeur du facteur aF, est peu
justifiée
dés que la masse volumique est supérieure
à 0,6 g/cm3. En effet, pour une telle valeur du
paramètre, au facteur de forme des cellules e/D = 0,58, correspond un
facteur de forme de l'élément sollicité en flexion
très faible L/h = (D-e) /2e = 3,5. Pour des cellules assimilables
à des cellules de bois final (parois épaisses et diamètres
cellulaires assez faibles), une partie de l'anisotropie L/T, est donc
directement imputable à un effet « quantité de
matière ».
65
Hiérarchisation des paramètres descriptifs de
l'anisotropie élastique du bois normal
Pour évaluer le degré d'anisotropie induit dans le
plan transverse du nid d'abeille par la seule
différence des arrangements en alignement radial et en
quinconces, les variations du rapport aRT d'anisotropie transverse
tissulaire ER / ET exprimé en (3.6), en fonction de la masse
volumique, sont reportées dans le Tableau 3.2 et la Figure 3.2.
E 1
e (D 2e)3 E E E
(3.6)
R
= 1 +
Rp
F
= (1 + a
) Rp = a Rp
E T
64 D
e3
E Tp
E Tp
RT
E
Tp
L'examen simultané de la Figure 3.2 et du Tableau 3.2
permet de constater qu'une anisotropie

transverse macroscopique R/T classique (Guitard et al, 1987)
supérieure ou égale à 2 ne peut être attribué
à la seule différence d'arrangement en quinconce ou en alignement
que pour des masses volumiques inférieures à 0,5
g/cm3.
|
(g/cm3)
|
Po (%)
|
e/D
|
aRT
|
|
0,20
|
0,86
|
0,03
|
15,42
|
|
0,30
|
0,80
|
0,05
|
5,56
|
|
0,40
|
0,73
|
0,07
|
3,03
|
|
0,50
|
0,66
|
0,09
|
2,06
|
|
0,60
|
0,60
|
0,11
|
1,61
|
|
0,70
|
0,53
|
0,13
|
1,37
|
|
0,80
|
0,47
|
0,16
|
1,19
|
|
0,90
|
0,40
|
0,18
|
1,13
|
|
1,00
|
0,33
|
0,21
|
1,07
|
|
1,10
|
0,27
|
0,24
|
1,04
|
|
1,20
|
0,20
|
0,27
|
1,02
|
|
1,30
|
0,13
|
0,31
|
1,01
|
|
1,40
|
0,07
|
0,37
|
1,00
|
|
1,50
|
0,01
|
0,46
|
1,00
|
Tableau 3-2 Variations des rapports de forme
e/D
et du rapport d'anisotropie aRT avec la
masse volumique
Figure 3.2 Evolutions du facteur aRT
avec la masse volumique
3.1.1 Comparaison au modèle de Gibson et al et
influence de l'élément de paroi fléchie
Afin d'élucider l'influence de la morphologie
cellulaire sur les rapports d'anisotropie
élastique du tissu ligneux, la modélisation de
Guitard et al (2002), retenue ici, est confrontée
à celle formulée par Gibson et al
(1988) (et reprise par Koponen et al en 1991), décrivant
le matériau bois comme une structure nid d'abeille
hexagonale non régulière. Cette confrontation est d'autant
plus aisée que les dispositions alignées et en quinconces
évoquées dans le modèle de Guitard et al
(2002) représentent deux cas « limites » de la
géométrie tissulaire décrite et admise par Gibson et
al (1988).
Ainsi, si la configuration des cellules en alignement (sens R)
envisagée ici est à rapprocher
(en terme de réponse mécanique) de la
configuration en carré, proposée par ces derniers auteurs,
la disposition en quinconce (sens T, Figure 1.8b du chapitre 1),
correspond à la structure nid d'abeille hexagonale (sens 2),
rappelée sur la Figure 1.8a du chapitre 1.
66
Hiérarchisation des paramètres descriptifs de
l'anisotropie élastique du bois normal
Les particularités géométriques du
modèle (b), proposé par Guitard et al, (Figure 1.8b)
sont
les suivantes: la valeur du paramètre angulaire è
est prise nulle (è = 0), tandis que les valeurs numériques
assignées aux paramètres géométriques
(paramètres de forme) l et h, caractérisant
la cellule rectangulaire (b) sont DR /2 et h = DT.
La différence essentielle entre le modèle (a) et
le modèle (b), réside ainsi dans le choix de la longueur utile
des éléments de paroi sollicités en flexion. Selon
Gibson et al (1988), la longueur des éléments
fléchis, notée l, n'est pas liée à
l'épaisseur de la paroi. Dans le cas du modèle (b), la
longueur utile de l'élément fléchi, noté (DR
- 2eT), est, de façon évidente, fonction décroissante
de cette même épaisseur de paroi. En conséquence, pour un
diamètre
DR de cellule fixé, le modèle (b) sera plus
rigide que le modèle (a) au fur et à mesure que l'on
considère des parois plus épaisses, c'est à dire
des masses volumiques plus élevées. Ceci rejoint la remarque
faite au paragraphe précédent (paragraphe 3.1).
L'estimation du module d'Young du tissu ligneux, soumis à
une sollicitation dans la direction
2, proposée par Gibson et al (1988) est
donnée par la relation suivante (3.7):
E t 3 (h
+ sinè)
(3.7)
2
E 2paroi
=
l
l
cos3è
Cette relation appliquée à la configuration
géométrique (b), envisagée notamment par Guitard
et al (2002) et présentée sur la
Figure 1.8 b, en utilisant les notations propres à la cellule
rectangulaire devient (3.8):
3
(3.8)
ET = 128 DT
eR
ERp
DR DR
Cette dernière expression est à comparer
à la relation (3.9), résultant de l'inversion de la
relation (2.19) du chapitre 2, qui peut se mettre sous la forme
suivante:
(3.9)
E T
= E Tp eT 1
2
E E D 1
e D
2e
3
Rp Rp
R
1 +
E Tp T
R T
64
E Rp
D T e R
L'expression (3.9) diffère de (3.8) par
l'intermédiaire de deux termes. Le "1", premier terme à
l'intérieur du crochet au dénominateur,
représente la contribution des éléments de paroi
sollicités en traction compression à la rigidité du nid
d'abeille. Ce terme est négligé a priori par les auteurs du
modèle (a). Le second terme du crochet au dénominateur
est lié à la contribution des éléments de paroi
sollicités en flexion (Figure 2.3, chapitre 2).
Comme évoqué dans le début de ce paragraphe,
la différence entre les deux expressions porte
sur la correction en (2eT) de la longueur des
éléments fléchis (DR-2 eT). En négligeant chacun
de ces deux effets, c'est à dire en annulant chacun des
deux termes, (3.9) se réduit strictement
à (3.8).
67
Hiérarchisation des paramètres descriptifs de
l'anisotropie élastique du bois normal
Pour apprécier l'incidence de la correction de
la longueur des éléments fléchis sur
l'anisotropie tissulaire L/T, il convient de comparer
numériquement les valeurs du coefficient aLT défini en (3.5)
à son équivalent aG (3.10) déduit de (3.8).
1 E Tp e
(D 2e
)3
e D e
(3.5)
a LT
= 1 + T
R T 1 2 R +
R R
64
E Rp D T
e R 3
D T D T eT
1 E
e D
3
e D e
(3.10)
= = Tp T R
1 2 R +
R R
a
G 64
E Rp
D T e R
D T D T
eT
Remarque : les expressions précédentes
(3.6) à (3.10), faisant intervenir le rapport
d'anisotropie transverse bipariétal
E Rp
, ont été laissées
en l'état pour « montrer »
E Tp
l'influence des propriétés mécaniques
de la double cloison sur les différents coefficients
d'anisotropie tissulaires. Comme précisé en début de ce
chapitre, les applications numériques sont néanmoins
menées, faute d'information sur le degré d'anisotropie transverse
de la paroi, sous l'hypothèse d'une isotropie transverse de la double
cloison.
Sous les conditions précisées, une application
numérique est présentée dans le Tableau 3.3.

Figure 3.3 Comparaison de l'évolution des
coefficients géométriques d'anisotropie
(aLT) du modèle de Guitard et al
et
du coefficient équivalent
(aG) du modèle proposé par Gibson
et al
|
ñ (g/cm3)
|
0,2
|
0,3
|
0,4
|
0,5
|
0,7
|
1
|
1,1
|
1,2
|
1,3
|
1,4
|
1,5
|
|
aLT
|
22,67
|
9,62
|
5,45
|
3,69
|
2,32
|
1,69
|
1,57
|
1,48
|
1,38
|
1,27
|
1,08
|
|
aG
|
25,67
|
10,78
|
5,71
|
3,42
|
1,51
|
0,56
|
0,41
|
0,30
|
0,22
|
0,15
|
0,08
|
Tableau 3-3 Evolution des valeurs numériques
des coefficients d'anisotropie (aLT) et (aG) avec
la masse volumique
68
Hiérarchisation des paramètres descriptifs de
l'anisotropie élastique du bois normal
La Figure 3.3 représente, en fonction de la masse
volumique ñ, les valeurs du coefficient
géométrique d'anisotropie aLT du modèle
(b) de Guitard et al et du coefficient équivalent aG
correspondant au modèle (a) de Gibson et al. On constate que
pour des masses volumiques faibles, inférieures à 0,5
g/cm3, les deux schématisations sont quasi
équivalentes et prévoient
des anisotropies L/T d'autant plus élevées que la
valeur de la masse volumique est faible.
Pour des structures de densité supérieure
à 0,5, le modèle de Guitard et al (2002) (sens T)
indique une anisotropie géométrique tissulaire L/T
supérieure à celle prévue par le modèle
proposé par Gibson et al (1988). Néanmoins, pour
de telles valeurs de densité, le niveau d'anisotropie
prévue par le modèle (b) à cellules rectangulaires est peu
supérieur à 3 et tout à fait insuffisant pour rendre
compte des rapports L/T observés expérimentalement.
|
|


