|
DEDICACES
Avec la Grâce du Très Haut sans qui rien
n'aurait été possible aujourd'hui...
A SAVOU, ma mère ; tu n'auras jamais
tort d'avoir toujours cru en moi et d'avoir fait de moi ce que je suis
devenu.
A mon PERE, pour la rigueur qui m'a
façonné...
A mes frères Blandine, Guy Patrick, Diane Olive,
Marie Emma, Yannick Larry, Loïc Jeffrey, je vous serai toujours
reconnaissant de tant d'affection et de sacrifices consentis, quelle chance
ai-je de vous avoir ?!...Et toute ma famille
A mes `'frères'' Hugues et
Ismaël pour tous les moments de joie, de peine et de
galère. Nos dreams deviennent de plus en plus réalité et
le `'HIT'' a sans doute commencé son envol vers les
sommets, soyez prêts !
Sheila, pour ton soutien sans faille et tout
ton amour, trouve ici l'expression de toute ma gratitude ainsi que mon amour
réciproque
A Lamine et Mariétou
Kamara, remerciements infinis pour avoir fait de moi un
privilégié par vos soins affectueux...J'ai des larmes que je ne
peux retenir !
A Lysiane, Marcelle, Ginette, Raïssa,
formidables et spéciales que vous êtes, si vous n'aviez pas
été là, il aurait fallu vous créer. Gros
bisous !
A Bernardin, Jeanne et ses
frères, Stan, Judy, Tidiane,
A Cynthia, , Marlène, Tina, Gali, Alix, Anouchka,
encore merci pour vos leçons de sympathie
A mes amis d'Awendjé...
A tous les membres du
« Kongossa » de la 8 C,
particulièrement Khassim, Lamine `'Niak'', Francis, Guy Blaise
`'Kirikou'' sans oublier mon grand, Simon.
A tous ceux que j'aurais `'oublié'', faute de place,
sachez que je vous porte dans mon
A tous les braves et valeureux agriculteurs qui ont
été victimes du péril acridien, sans oublier tous ceux
(agents, techniciens et volontaires) qui ont contribué à mener la
lutte.
Une pensée très pieuse pour vous tous qui
êtes déjà partis ; une spéciale pour ma
grand-mère Alphonsine `'Kaka'', tu aurais du voir ton Petit papa
aujourd'hui. Je ne t'oublie de là où tu es Taylor, ton vide ne
sera jamais comblé dans la famille. Toi non plus, Michel Wabo, durant
tout mon séjour à l'ENSA, ton souvenir m'est resté
gravé. Que Dieu vous garde !
I. REMERCIEMENTS
A l'issue de ce modeste travail qui couronne notre passage
à l'ENSA, qu'il me soit permis ici d'adresser mes sincères
remerciements à tous ceux là qui de loin ou de près y ont
joué un rôle :
· Pr. Papa Ibra Samb et Dr. Abdoulaye Dramé
respectivement Directeur et Directeur des études de l'ENSA, et à
travers eux, l'ensemble du corps professoral et du personnel de
l'école.
· Messieurs Moussa Fall et Alioune Coly anciens Directeur
et Directeur des études qui m'ont permis d'intégrer cette
école et de réaliser un rêve d'enfance.
· Dr.Saliou Ndiaye, Chef du Département `'PV'', le
grand frère avant d'être l'enseignant, toujours là, avec
les mots qui suffisaient, quand il le fallait.
· La Direction de TROPICA SEM et particulièrement
à Dr. Rémi Nono-Womdim pour m'avoir ouvert les portes de la
station `'Baobab'', sans jamais ménager aucun effort pour m'assurer les
conditions de travail les meilleures. En plus de la rigueur scientifique, de la
disponibilité....Je garde en admiration le charisme du Chef mais surtout
la grande sympathie d'un Aîné.
· Dr. Alain Palloix de l'INRA pour son importante
contribution à ce travail et son action dans la recherche sur les
piments. J'ai été honoré de travailler avec un
spécialiste de renommée mondiale.
· Tous les membres du Jury pour l'honneur que vous me faites
d'évaluer ce travail
· Mamadou Sarr et Fatoumata Coly, mes `'compagnons
techniques'' pour tout leur soutien dans ce travail. Et à travers eux,
l'ensemble du personnel de la station de Keur Ndiaye Lô pour m'avoir
intégré vraiment comme un des leurs.
· Monsieur Ibrahima Diop de la maintenance, messieurs Hann
et Fall, sans oublier madame Faye de la bibliothèque, monsieur Ibrahima
Bâ et tout le personnel de l'ENSA.
· Ma `'Promo'' d'origine, la 19e, d'abord.
Ensuite à ma `'Promo'' d'adoption, la 20e pour tous les
moments communs de dur labeur mais surtout pour l'ambiance formidable. Mention
spéciale à mes confrères `'PV 2004'' ; Sémou,
Seyny, Bassirou et Halimatou
· L'ensemble de la Communauté
étrangère ; Carole, Stella, Noëlla, Djihane, Les
`'Centro'', Ali, Christian, Fanéli, Barry
· A tous mes anciens particulièrement Nicolas
Bassène, Francis Bouba, Richard, Toka, Yves, .. et Mes cadets, Iso
Diatta, Ndiaga, Nabou, Bintou., Modou Marie et tout le reste
II. Résumé
Le piment doux (Capsicum annuum L.) est une plante
maraîchère de grande importance connue et cultivée de par
le monde. Son adaptation à des conditions tropicales chaudes et humides
pose cependant des problèmes en ce sens que la chaleur humide et la
forte pression parasitaire qui y sont relatives affectent grandement la
culture. D'où la nécessité de développer des
variétés dites tolérantes à cet environnement.
Pour ce faire, la sélection d'un matériel
adapté passe par l'intégration de caractères de
résistance et d'adaptation portées par une espèce voisine,
en l'occurrence C. chinense réputée tolérante car
présentant d'intéressantes caractéristiques en saison
pluvieuse.
En conditions d'hivernage, il a été
procédé à l'évaluation et la caractérisation
d'une quarantaine de génotypes, produits de la première
génération d'un rétrocroisement ou backcross de plusieurs
lignées de populations d'origines diverses.
Il ressort de notre analyse que dans l'ensemble (93 %), tous
les génotypes ont démontré un comportement positif
généralisé du point de vue de l'adaptation et donc de la
tolérance à la chaleur humide. Les résultats
suivants :
*Deux niveaux de vigueur ; bonne vigueur (49 %) et
vigueur moyenne (44 %),
*10 % de levée tardive,
*Un rythme de croissance et un niveau d'encombrement moyens
supérieurs à la moyenne des témoins,
*88 % des génotypes présentaient une
précocité de floraison,
*La fructification est normale pour 62 % de l'ensemble du
matériel et le poids moyen des fruits est fort appréciable,
*Du point de vue de leur conformation, les idéotypes
recherchés constituent la grande majorité des fruits obtenus
à raison de 67 % de carrés de type `'Yolo'' et 31 % de demi-longs
de type `'Lamuyo''.
Sur la base de ces indicateurs quelques génotypes ont
été `'choisis'' pour affiner davantage la sélection par
conséquent une meilleure fixation des caractères de
tolérance et d'adaptation aux conditions tropicales chaudes et
humides.
Mots-clés :Capsicum annuum,
Capsicum chinense, adaptation, résistance, hivernage,
backcross
III. Abstract
Sweet pepper (Capsicum annuum L) is an important
fruit vegetable widely known and grown in the world.
However in hot and humid tropics, sweet pepper is susceptible
to all kinds of diseases and the too heavy rainful is harmful to the crops, for
it causes poor fruit set and rotting of the fruits.
For this reason, it's necessary to develop varieties
possessing both adaptation to this kind of environment and resistance to major
diseases.
The appropriate breeding work need the use of resistance genes
of a closely relative species like the hot and aromatic pepper (Capsicum
chinense) in which is reported good adaptation and tolerance to hot and
humid conditions.
During the rainy season, evaluation and characterization of
about forty genotypes resulting from the first generation backcrossing of
various cultivars intercrossed has been performed. The analysis has allowed to
find out significant adaptation traits in the material such as:
*Suitable vigour level; 49 % (good vigour) and 44 % (middle
vigour),
*Acceptable growth rhythm and plant canopy,
*88 % of genotypes showed few days to flowering,
*62 % normal fruit setting,
*Suitable fruit weight and size,
*Fruits types: 67 % `'Yolo'' and 31 % `'Lamuyo''.
Some genotypes have been chosen to confirm and improve the
followed results. The breeding work is likely to deal with fixation of these
significant characters.
Key words: Capsicum annuum, Capsicum
chinense, hot an humid conditions, backcrossing, adaptation
IV. LISTE DES ACRONYMES
AFD : Analyse Factorielle Discriminante
CDH : Centre de Développement de
l'Horticulture
ENSA : Ecole Nationale Supérieure
d'Agriculture
INRA : Institut National de Recherche Agronomique
(France)
ISTA : International Seed Testing
Association
LISTE DES TABLEAUX
|
N°
|
Titre
|
Page
|
|
1
|
Les vitamines du poivron
|
9
|
|
2
|
Les minéraux du poivron
|
9
|
|
3
|
Les composants du poivron
|
10
|
|
4
|
Les noms vernaculaires africains
|
10
|
|
5
|
Les besoins en eau par système d'irrigation
|
13
|
|
6
|
Génétique des caractères
|
22
|
|
7
|
Relevé météorologique de l'hivernage
2004
|
28
|
|
8
|
Liste des parents initiaux
|
29
|
|
9
|
Récapitulatif des croisements
|
30
|
|
10
|
Liste du matériel total testé
|
32
|
|
11
|
Suivi de la fertilisation
|
33
|
|
12
|
Traitements phytosanitaires effectués
|
34
|
|
13
|
Relevé pathologique
|
46
|
|
14
|
Les corrélations totales des variables
|
56
|
|
15
|
Comparatif des principales familles
|
59
|
LISTE DES FIGURES
|
N°
|
Titre
|
Page
|
|
1
|
Les fruits
|
4
|
|
2
|
Coupe longitudinale du fruit
|
5
|
|
3
|
Les types variétaux de poivrons
|
6
|
|
4
|
Coupe d'une fleur
|
8
|
|
5
|
Symptômes de TMV sur fruit
|
15
|
|
6
|
Pucerons sur les feuilles
|
15
|
|
7
|
Symptômes de CMV sur une feuille
|
15
|
|
8
|
Symptômes de CMV sur les fruits
|
15
|
|
9
|
Plante affectée par le flétrissement
|
19
|
|
10
|
Aperçu des fruits récoltés
|
21
|
|
11
|
Situation de la zone des Niayes
|
27
|
|
12
|
Système élémentaire pour goutte à
goutte
|
35
|
|
13
|
Répartition de la levée
|
39
|
|
14
|
Evolution de la levée
|
40
|
|
15
|
Evolution de la croissance moyenne
|
41
|
|
16
|
Evolution de la croissance comparée à mi-cycle et
à la récolte
|
42
|
|
17
|
Evolution du niveau d'encombrement
|
43
|
|
18
|
Evolution des dimensions foliaires
|
44
|
|
19
|
Evolution du niveau de vigueur
|
45
|
|
20
|
Distribution de la floraison
|
47
|
|
21
|
Evolution de la floraison
|
47
|
|
22
|
Distribution de la fructification
|
48
|
|
23
|
Evolution de la fructification
|
49
|
|
24
|
Evolution de la première récolte
|
50
|
|
25
|
Evolution de la seconde récolte
|
50
|
|
26
|
Comparaison des deux récoltes
|
51
|
|
27
|
Evolution du rendement moyen par plante
|
52
|
|
28
|
Evolution du poids moyen des fruits
|
53
|
|
29
|
Evolution du calibre moyen des fruits
|
54
|
|
30
|
Evolution du nombre moyen de graines
|
55
|
|
31
|
Distribution de la couleur des fruits avant maturité
|
57
|
|
32
|
Distribution de la couleur des fruits à
maturité
|
57
|
V. SOMMAIRE
Pages
Dédicaces................................................................................................II
Remerciements........................................................................................III
Résumé...................................................................................................IV
Abstract...................................................................................................V
Liste des
acronymes.................................................................................VI
Liste des
tableaux.....................................................................................VI
Liste des
figures......................................................................................VII
INTRODUCTION
GENERALE.......................................................................1
Première partie : Synthèse
bibliographique
Généralités sur la plante et sa
culture......................................................2
Le genre
Capsicum..........................................................................2
Origine et
distribution..............................................................2
Systématique et botanique : caractères
généraux.......................2
Les espèces cultivées :caractéristiques
principales....................3
L'espèce Capsicum
annuum.............................................................3
Morphologie et types
variétaux.................................................3
Morphologie..................................................................4
Types
variétaux.............................................................5
Développement de la
plante.............................................6
Mode de reproduction et biologie
florale....................................7
mode de
reproduction.....................................................7
Modalités de
l'autogamie.................................................7
Biologie
florale...............................................................7
L'abscission des organes
reproducteurs....................................8
Importance de la
culture................................................................8
Valeur
nutritive........................................................................9
Les divers usages
..................................................................9
Nomenclature
vernaculaire.....................................................10
Agronomie de Capsicum
annuum.........................................................10
Exigences
écologiques..................................................................10
Conduite et pratiques
culturales.....................................................11
Semis et
plantation................................................................11
Fertilisation et
irrigation.........................................................12
Rôle des principaux éléments
minéraux...........................12
Estimation des besoins et modalités pratique de la
fumure12
Les besoins en
eau........................................................12
Entretien
cultural..................................................................12
Lutte contre les mauvaises
herbes..................................12
Paillage ou
mulching......................................................12
Utilisation des filets
d'ombrage.......................................12
Tuteurage.....................................................................13
Les contraintes majeures de la
culture.............................................14
les problèmes
phytosanitaires.................................................14
les maladies
virales.......................................................14
les maladies
fongiques...................................................16
les maladies
bactériennes..............................................18
Les ennemis
animaux.............................................................19
Les accidents
physiologiques.................................................20
Récolte et
conservation..................................................................20
Génétique du piment (Capsicum annuum,
L)..........................................22
Relations
cytogénétiques........................................................22
Génétique des
caractères.......................................................22
Qualité et sélection du
piment..................................................23
Critères de
qualité..................................................................23
Méthodes de
sélection............................................................23
Sélection
massale.........................................................23
Méthode généalogique différée
(ou bulk)..........................23
La filiation unipare (ou
SSD)...........................................24
Le
backcross................................................................24
La sélection
récurrente..................................................24
Facteurs influençant la production de semences
hybrides..........24
La qualité d'une
semence.......................................................24
Les hybrides et la production de
semences..............................24
Expression de
l'hétérosis.......................................................25
Justification de l'utilisation de C.
chinense...............................25
Un point sur la
recherche....................................................................26
Seconde partie : Méthodologie
Cadre de
l'étude..................................................................................27
Localisation..................................................................................27
Agro-écologie du
site.....................................................................27
Matériels et
Méthode...........................................................................29
Matériel
végétal.............................................................................29
Listing des parents
initiaux.....................................................29
Tableau général des
croisements............................................29
Matériel
testé........................................................................31
Méthodologie................................................................................33
Dispositif
expérimental...........................................................33
Fiche technique de
l'essai......................................................33
Observations
réalisées...........................................................36
Pendant la phase
végétative...........................................36
Caractères
quantitatifs..............................................36
Caractères
qualitatifs................................................37
Pendant la phase
générative...........................................37
Caractères
quantitatifs..............................................37
Caractères
qualitatifs................................................38
Traitement statistique des
données..........................................38
Limites
méthodologiques........................................................38
Résultats et
discussion........................................................................39
Evaluation du matériel
végétal.................................................39
En phase
végétative...............................................................39
Caractères
quantitatifs...................................................39
Evolution de la levée des
plantes...............................39
Evolution de la hauteur des
plantes...........................40
La canopée ou
encombrement...................................42
Les dimensions
foliaires.................................44
Caractères
qualitatifs....................................................45
Le
port....................................................................45
La vigueur
générale..................................................45
L'état
sanitaire.........................................................46
En phase
générative...............................................................46
Caractères
quantitatifs...................................................46
La précocité de
floraison...........................................46
Evolution de la
fructification......................................48
Le rendement et ses composantes.............................49
Le rendement génotypique moyen relatif........49
Le rendement génotypique moyen par plante..52
Le poids moyen des fruits............................53
Le calibre moyen des fruits..........................54
Le rendement grainier par génotype..............54
Caractères
qualitatifs....................................................56
La couleur des
fruits.................................................56
Avant
maturité............................................56
A
maturité..................................................57
L'état
sanitaire........................................................58
Caractérisation globale du matériel
végétal..............................58
Conclusion générale et
perspectives........................................................61
Références
bibliographiques....................................................................63
VI. INTRODUCTION GENERALE
Le piment doux (Capsicum annuum L.) plus connu
sous le terme générique de poivron est une plante
maraîchère originaire d'Amérique tropicale, très
appréciée pour ses fruits surtout consommés en tant que
légumes.
C'est une plante qui revêt une grande importance,
tant du point de vue économique, du point de vue alimentaire que du
point de vue de la recherche, en sélection et amélioration
variétales notamment.
Depuis quelques années en effet la production mondiale
de poivrons n'a cessé de s'accroître, passant de 10.769.000 tonnes
en 1991 à 22.168.000 tonnes en 2002 (FAO, 2003), soit du simple au
double en l'espace d'une décennie.
Les poivrons sont réputés être des
sources importantes de vitamines, contribuant ainsi grandement à
l'amélioration de la ration et de l'équilibre alimentaire. Ils
représentent en plus, au même titre que les aubergines et les
courgettes par exemple, d'excellents produits de diversification de la
production agricole.
Cependant, la culture du piment (Capsicum annuum L.)
se heurte à quelques problèmes d'adaptation dans certaines zones
agro-écologiques.
En zone tropicale chaude et humide par exemple, la
quantité et la qualité de la production se trouvent
généralement et essentiellement affectées par la forte
pression parasitaire liée aux conditions climatiques extrêmes.
D'où, des rendements souvent aléatoires liés à
l'adaptation à cet environnement qui mettent en exergue l'épineux
problème de l'étalement de la production maraîchère
sur toute l'année.
Si jusqu'à maintenant, de nombreux travaux ont
été menés sur le poivron, ils ont conduit essentiellement
à de pertinents résultats sur les diverses voies de
résistance génétique à la prolifération des
multiples ennemis de la culture. Et à ce titre certaines
variétés présentent d'ailleurs d'intéressantes
résistances aux maladies. Les questions liées à
l'adaptation aux conditions tropicales chaudes et humides demeurent donc
d'actualité et s'associent désormais à des programmes de
recherche privilégiés.
Dans cette dynamique, le présent travail s'inscrit
dans le cadre d'un programme de sélection dont l'objectif est le
développement de variétés de poivrons adaptées aux
conditions intertropicales. A terme, ces variétés devraient
s'adapter à la chaleur humide en cumulant aussi bien des facteurs tant
de résistances, d'adaptation que de tolérance à ces
conditions.
Il s'agira donc pour nous, d'étudier les
caractéristiques du développement végétatif et
génératif vis à vis de la chaleur humide à travers
une évaluation différentielle du comportement de plusieurs
génotypes sélectionnés.
En définitive, nous tenterons de mettre en exergue les
combinaisons qui révéleront les meilleures
propriétés adaptatives et qui seront susceptibles d'être
suivies dans le cadre d'une sélection créatrice ou d'une
innovation variétale.
I. Généralités sur la plante et sa
culture
I.1. Le genre
Capsicum
I.1.1. Origine et distribution
Kalloo (1989) de même que De Witt & Bosland
(1993) rapportent que les premières variétés de
Capsicum, genre créé par Valerius Cordus en 1506 (Chaux
& Foury, 1994), seraient originaires d'une zone localisée entre les
montagnes sud brésiliennes à l'est, la Bolivie à l'ouest,
le Paraguay et le nord Argentine au sud. Ils qualifient cette zone comme `'aire
centrale'' où auraient été représentées
toutes les principales espèces domestiquées du genre. Harlan
(1987) pour sa part, précise deux zones d'origine ; le centre
mésoaméricain pour C. annuum et le centre
d'Amérique du sud pour les autres espèces. Les Capsicum
se seraient alors répandus à travers les Amériques avant
d'être domestiqués indépendamment dans la zone du Mexique
(C. annuum), en Amazonie (C. chinense) et dans les
régions méridionales d'Amérique centrale (C.
frutescens). Les deux autres espèces (C. baccatum et
C. pubescens) sont restées principalement confinées en
Amérique du sud (De Witt & Bosland, 1993).
L'histoire nous enseigne que Christophe Colomb rapporta du
piment en Europe au retour de son premier voyage à la conquête
d'une nouvelle route des épices en 1493.
I.1.2. Systématique et botanique :
caractères généraux
Le piment appartient au genre Capsicum de la
grande famille des Solanaceae comme les aubergines (Solanum
melongena, S. aethiopicum), la tomate ( Lycopersicon
esculentum), la pomme de terre (Solanum tuberosum) ou encore la
tabac ( Nicotiana tabacum).
Dans la sous-classe des Aristidae du groupe des
Dicotylédones évoluées caractérisés par la
gamopétalie (pétales soudés), les Solanaceae
appartiennent à l'ordre des Polémoniales, à port
herbacé et à ovaire supère. (Guignard, 1996)
La classification du genre est assez confuse (Purseglove,
1984). On dénombre une vingtaine de formes spécifiques et de
nombreuses variétés botaniques. De deux (02) (Purseglove, 1984)
à cinq (05) (Eshbaugh, 1977 cité par Chaine-Dogimont, 1993)
espèces ont été domestiquées et sont les plus
fréquemment cultivées suivant les régions du monde;
C.. annuum, C.. baccatum, C.. chinense,
C.. frutescens, C. pubescens.
La saveur piquante de certaines espèces
qualifiées de « piments forts » (par
opposition aux « piments doux ») est liée à
la présence de la Capsaïcine (C18H27NO3), substance irritante du
groupe des vanillyl-amides localisée au niveau du placenta (Anu &
Peter, 2000) et dont la plus forte concentration se rencontre au voisinage des
graines (Messiaen, 1975)
Chez les Capsicum, les graines sont lisses, plus
plates que celles d'aubergine. La germination est identique à celle de
la tomate et de l'aubergine.
Les feuilles sont glabres et lancéolées alors
que les fleurs sont blanches en général et semblables à
celles d'aubergine mais avec des dimensions plus réduites.
Les Capsicum produisent des fruits de forme et de
taille variables ; verts avant leur maturité pour prendre des
colorations jaunes, rouges ou violacées aux stades les plus
avancés .
Les fruits peuvent être allongés, flexueux,
coniques, globuleux à 3 ou 4 loges (lisses ou flexueux),
sphériques ou plats côtelés.
I.1.3. Les espèces
cultivées :caractéristiques principales
On peut distinguer les diverses espèces de
Capsicum cultivées principalement par leur type de
développement ; annuel ou vivace, mais aussi par certains
caractères morphologiques (Messiaen, 1975). Greenleaf (1986) pour sa
part fait état d'une combinaison entre les caractéristiques des
fleurs et des fruits. Ainsi on a :
· Capsicum frutescens
L. : espèce vivace dont la hauteur des
plants dépend du climat, à feuillage fin, à fleurs souvent
en bouquets (Williams et al, 1991) insérées par paires.
Les feuilles sont parfois ovoïdes (De Witt & Bosland, 1993). Le
pédoncule floral est droit à la floraison et les fruits sont
allongés coniques (parfois ronds),très piquants mais peu
parfumés. Ils sont faciles à sécher.
· Capsicum chinense
Jacq. : a un feuillage plus
large et plus gaufré que C. frutescens, mais pousse lui, plus
lentement que C. annuum. Il est plus vivace que C. frutescens
et peut donner au bout de 3 ou 4 ans des arbustes de 1,5 m de haut avec des
troncs de 2 ou 3 cm de diamètre. Ses fruits (2 à 6 par noeud)
sont de forme variée, en lanterne, ou très plats
côtelés, très piquants (jusqu'à 250 000 sur
l'échelle de Scoville) à doux (Grubben & El Tahir, 2004) et
très parfumés. L'espèce est, de plus réputée
pour sa longue période de récolte, ses besoins presque nuls en
pesticides et son coût de production très appréciable. Son
cycle varie entre 80 et 120 jours en moyenne. Les semences sont lentes à
germer. Grubben & El Tahir (2004) ajoutent que l'espèce est
très appréciée en saison des pluies pour sa vigueur et sa
grande résistance à l'anthracnose et aux viroses.
· Capsicum pubescens Ruiz et
Pav., se caractérise par ses feuilles ovoïdes très poilues
(d'où d'ailleurs son nom) et ses fleurs bleues, ses graines noires et
irrégulières. L'espèce est caractéristique des
zones élevées (Chaine-Dogimont, 1993). C'est un grand buisson
pérenne qui peut s'élever jusqu'à 3 m de haut. Son cycle
de croissance est d'au moins 120 jours. Ses fruits se déshydratent et ne
se conservent pas bien. (Eshbaugh, 1977 cité par Chaine-Dogimont,
1993)
· Capsicum baccatum L. :
est une espèce qui se distingue par sa corolle blanche marquée de
chevrons jaunes et ses étamines libres. Elle est surtout utilisée
comme condiment. Elle est caractéristique des Andes, zone dans laquelle
on rencontre également sa forme sauvage (Pickersgill, 1971 citée
par Chaine-Dogimont, 1993). Son cycle est d'environ 120 jours et les plants de
C. baccatum ont du fait de leur persistance, une tendance arbustive.
De Witt & Bosland (1993) ont défini dans ce groupe deux formes
sauvages (baccatum et microcarpum) ainsi qu'une forme
domestiquée (pendulum)
· Capsicum annuum L. que nous
étudierons plus amplement par la suite est une espèce annuelle
comme son nom l'indique. On rencontre dans cette espèce en
général des variétés à fruits doux mais
aussi quelques variétés à fruits piquants.
I.2. L'espèce
Capsicum annuum
I.2.1. Morphologie et types variétaux
I.2.1.1. Morphologie
· C'est une plante herbacée de 0,5 à 1, 5 m
de haut dont le système radiculaire est un pivot assez fort avec des
racines qui ont une tendance à se développer latéralement
dans un rayon de 0,30 à 0,50 cm. La tige se lignifie progressivement
d'où la tendance à un mode pérennant (Chaux & Foury,
1994)
· Les feuilles sont simples, larges, molles,
pétiolées et alternes, très souvent glabres. Elles sont
ovales à elliptiques plus ou moins allongées, à sommet
aigu ; chacune est opposée à la feuille du sympode (= axe+
feuille + fleur).
· Les fleurs sont généralement solitaires,
quelques fois par paires ou en bouquets. Elles sont petites, blanches,
terminales, bisexuées et habituellement pentamériques. La corolle
est composée de cinq (05) pétales soudés qui lui donnent
l'aspect d'un tube. Les étamines alternent avec les lobes des
pétales et le style est unique.
· Le fruit (Fig 1.) est une baie indéhiscente avec
un épais pédoncule qui varie suivant la forme ou la saveur
(piquante ou douce). Le péricarpe est coriace et charnu d'après
la description de Rajput & Parulekar (1998). La baie développe un
ovaire bicarpellaire avec un placenta axial. Ce dernier porte les graines.
(Fig 2.)
· Les graines sont réniformes, plates, à
tégument lisse et de couleur jaune paille. Leur taille est jugée
variable d'après Belleti & Quagliotti (1988) en fonction des
conditions dans lesquelles elles mûrissent ; l'environnement
général de la plante-mère, la position de la baie sur
celle-ci, leur nombre par fruits, le moment de la récolte et celui de
leur extraction du fruit. Un gramme compte environ entre 140 (Purseglove, 1984)
et 150 graines (Chaux & Foury, 1994), et peuvent conserver 50 % de leur
viabilité pendant trois (03) ans lorsqu'elles sont dans un lieu propre,
sec et sombre (Ashworth, 1991)
![]()
Figure 1 : les Fruits
![]() ![]()
(a)
(b)
Figure 2 :
Coupe longitudinale d'un fruit
I.2.1.2. Types variétaux
Il en existe plusieurs qu'on peut regrouper en trois
classes : les carrés, les rectangulaires et les triangulaires
(Fig.3)
· Les carrés : on y distingue les
carrés américains, italiens et hollandais. La forme est
régulière en général, globuleuse chez les
carrés italiens. Les parois sont épaisses et fermes, permettant
ainsi la mécanisation de la récolte et du conditionnement chez
les carrés américains ou type `' blocky''. Les carrés
hollandais ont un calibre plus petit que les autres.
· Les rectangulaires : on distingue dans ce
groupe ; les ½ longs, les ¾ longs et les longs. Les fruits sont
larges, épais, fermes, lisses, réguliers et bien colorés,
en jaune ou rouge essentiellement.
· Les triangulaires : appelés aussi
« cornés » (Mercier, 1999), ils sont proches des
types sauvages d'origine. On rencontre dans ce groupe, des formes piquantes.
Les fruits sont en général lisses et épais. Ils sont
caractérisés par une large gamme de coloris allant du blond
très clair en passant par l'orange, le jaune et le rouge à
maturité.
Il faut enfin ajouter qu'il existe également des formes
condiformes et subsphériques plus aplaties, principalement
utilisées pour l'industrie de transformation.
![]()
(source PHM n° 365-366)
Figure 3 :. les types variétaux de
poivrons
I.2.1.3. Développement de la
plante
Après un développement végétatif
monopodial sur 7 à 9 étages, le message floral est perçu
au niveau du méristème terminal qui donne une à deux
fleurs isolées.
Les bourgeons subterminaux (bourgeons axillaires des
dernières feuilles formées) donnent ensuite un axe qui ne porte
qu'une seule feuille et se termine à son tour par une fleur unique.
D'après Chaux & Foury (1994), la tendance à la
dichotomie serait alors le fait du développement d'une ébauche de
2e feuille pouvant axiller un second bourgeon caulinaire.
I.2.2. Mode de reproduction et biologie florale
I.2.2.1. mode de reproduction
Le piment C. annuum L. est une plante
annuelle préférentiellement autogame (Chaine-Dogimont, 1993) ou
encore autogame facultatif (Pochard et al, 1992 ). Ses fleurs sont
pentamériques et hermaphrodites, et elles sont fréquemment
visitées par les insectes d'où une allogamie résiduelle
qui en résulte.
Poulos (1993) note que chez la plupart des cultivars, les
fleurs diffèrent par la position des stigmates en relation avec les
anthères. Ainsi, les fleurs à longs styles dont les stigmates
s'étendent au-delà des étamines sont plus susceptibles de
pollinisation croisée que les fleurs à styles courts où
l'autopollinisation est surtout de mise.
De Witt & Bosland (1993) ajoutent que la pollinisation
croisée chez le piment C. annuum L. s'accentue avec la
structure de la fleur (Fig.4) mais aussi la présence de nectar qui
attire les insectes.
Diverses méthodes d'autofécondation peuvent
être utilisées : fermeture des fleurs à la colle ou
avec une bande adhésive, isolement de la plante. Les cas de castration
manuelle sont délicats du fait de la fragilité des fleurs et des
sacs polliniques. C'est un procédé très utilisé
pour la production de semences hybrides commerciales, rapportent Pochard et
al (1992)
I.2.2.2. Modalités de
l'autogamie
L'autogamie est une fécondation forcée et exclusive
d'un individu par lui-même. Elle est assurée au plan anatomique
par la structure de la fleur qui facilite l'autofécondation et
empêche l'hybridation. Cependant, 5 conditions biologiques doivent
être réunies à la fois :
Fleurs hermaphrodites, avec étamines et pistil,
Contact ou proximité étamines-stigmates,
Synchronismes des floraisons mâle et femelle,
Absence de systèmes d'auto-incompatibilité et de
stérilité mâle, au moins pendant la période de
réceptivité,
Fleurs closes, ou au moins stigmates protégés
durant leur période de réceptivité
I.2.2.3. Biologie florale
Le nombre de jours requis pour la floraison dépend
principalement de la variété et des conditions environnementales.
Rajput & Parulekar (1998) estiment que pour la plupart des
variétés, la floraison commence au bout de 40 jours après
le repiquage. Les fleurs sont souvent solitaires mais dans certains cas, elles
apparaissent en bouquets. Elles sont bractées, pédicellées
et bisexuées (hermaphrodites).
La réceptivité du stigmate devient effective
à partir du jour de l'anthèse (Kalloo, 1988 ; Rajput &
Parulekar, 1998) et demeure jusqu'à 2 jours après celle-ci
(Rajput & Parulekar, 1998).
Si d'aucuns jugent le moment à partir duquel les
grains de pollen deviennent fertiles, un jour avant l'anthèse (Rajput
& Parulekar, 1998), d'autres le situent le jour de l'anthèse
(Kalloo, 1988).
D'après Kalloo (1988), la durée de
l'anthèse est comprise entre 5 et 6 heures alors que celle de la
déhiscence varie entre 8 et 11 heures.
De nombreux auteurs enfin s'accordent sur le fait que C.
annuum est classiquement une plante à autofécondation qui
connaît cependant des taux variables de pollinisation croisée
naturelle. Kalloo (1988) révèle pour les groupes de vecteurs
suivants ; abeilles et thrips et autres insectes des taux respectivement
compris entre 7 et 37 %, et entre 2 et 78 %.
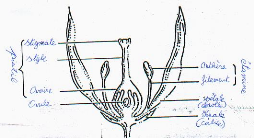
Figure 4 : coupe d'une
fleur
I.2.3. L'abscission des organes reproducteurs
Encore appelée « coulure », c'est
un phénomène majeur caractérisé par la chute des
boutons floraux, des fleurs et des jeunes fruits chez C. annuum dont
il constitue un facteur limitant de la production. Les principales causes de ce
phénomène sont liées à des contraintes
environnementales extrêmes (température, humidité,
luminosité) mais aussi à la pression parasitaire (Wien et
al., 1988).
Les fleurs ouvertes puis les boutons floraux sont les
premiers organes reproducteurs perdus après un ombrage de plus de 90 %.
Pour Chaux & Foury (1994), une profusion de fleurs par rapport à la
surface assimilatrice représente un risque constitutif de coulure. Wien
et al. (1998) imputent un rôle central à l'interaction
entre différents régulateurs de croissance dans le
mécanisme général de la chute des organes reproducteurs
mais aussi végétatifs.
D'après eux, les feuilles ou boutons floraux
fonctionnels et non sénescents produisent une auxine ainsi que d'autres
promoteurs de croissance qui diffusent en dehors de l'organe, le long du
pétiole ou du pédicelle et de cette façon empêchent
toute abscission.
En réalité, en conditions difficiles ou sous
l'influence d'une maladie ou des dommages d'insectes, les feuilles ou boutons
floraux infectés accroissent leur production d'éthylène,
un gaz régulateur de croissance qui va réduire le taux d'auxine
produite par l'organe considéré, diminuer le taux d'auxine
transporté vers le pédicelle ou le pétiole et ainsi
accélérer le phénomène d'abscission.
Autrement dit, selon Wien et al. (1988), c'est
l'équilibre entre les niveaux d'éthylène et d'auxine et
probablement d'autres promoteurs de croissance produits par l'organe en
développement qui détermine ou le maintien ou l'abscission des
boutons floraux et des feuilles.
I.3. Importance de
la culture
I.3.1. Valeur nutritive
Une des principales caractéristiques nutritionnelles
du poivron est sa très grande richesse en vitamines (tabl.1). C'est le
légume frais qui renferme le plus de vitamine C après le persil
(Anonyme, 2004). La provitamine A (carotène) atteint aussi des niveaux
intéressants (jusqu'à 3,5 mg / 100 g chez les poivrons rouges).
On a recensé également des vitamines du groupe B ( de 0,02
à 0,7 mg selon les vitamines) ainsi que de la vitamine E (1,4 mg / 100
g).
On a par ailleurs identifié de nombreux minéraux
(tabl.2) et oligo-éléments dont du potassium, du phosphore,
du magnésium, du calcium. Le fer est présent accompagné du
cuivre, du manganèse et du zinc.
Malgré sa richesse en vitamines la composition
(tabl.3) du poivron fait de lui, un fruit au niveau énergétique
modeste (Anonyme, 2004) ;
Ä Les glucides ( ou sucres) varient entre 2,2 et 4,7 g aux
100 g. Il s'agit surtout du glucose et du fructose, tandis que le saccharose
n'apparaît que sous forme de traces.
Ä Les protides végétaux représentent
environ 0,7 à 1,9 % du total.
Ä Les lipides (ou graisses) ne dépassent pas 0,2
à 0,4 g aux 100 g
Ä Les fibres sont relativement abondantes (2 g aux 100 g)
Tableau1 : les vitamines du
poivron
I.4. Vitamines
|
|
Vitamine C (acide ascorbique)
|
|
Provitamine A (carotène)
|
|
Vitamine B1 (thiamine)
|
|
Vitamine B2 (riboflavine)
|
|
Vitamine B3 ou PP (nicotinamide)
|
|
Vitamine B5 (acide panothénique)
|
|
Vitamine B6 (pyridoxine)
|
|
Vitamine B9 (acide folique)
|
|
Vitamine E ( tocophérols)
|
II. III. Minéraux
|
(mg)
|
|
Potassium
|
170.0
|
|
Phosphore
|
26.00
|
|
Calcium
|
9.000
|
|
Magnesium
|
13.00
|
|
Sodium
|
2.000
|
|
Chlore
|
18.00
|
|
Fer
|
0.400
|
|
Cuivre
|
0.100
|
|
Zinc
|
0.200
|
|
Manganèse
|
0.100
|
Tableau
2 : les minéraux du poivron
IV. Composants
|
(g)
|
|
Glucides
|
3.50
|
|
Protides
|
1.10
|
|
Lipides
|
0.30
|
|
Eau
|
91.0
|
|
Fibres alimentaires
|
2.00
|
Tableau 3 :
les composants du poivron
IV.1.1. Les divers
usages
Le piment doux est cultivé pour de multiples
usages. On lui reconnaît principalement un usage alimentaire en tant que
légume ; consommé cru ou non, mur ou non. Les fruits sont
parfois mis en conserves ; entiers pelés ou coupés en
lanières, appertisés au vinaigre, surtout dans les pays à
haut niveau de vie, rapportent Pochard et al. (1992). Les fruits sont
aussi dans certains cas transformés en poudre, séchés ou
fermentés.
Palloix & Phaly (1996) évoquent aussi un usage
religieux chez les Incas du Pérou et un usage médicinal
comme stimulant histaminique.
C'est enfin selon Dupriez & De Leener (1987) un excellent
laxatif et analgésique.
IV.1.2.
Nomenclature vernaculaire
Du fait de sa grande distribution, le piment Capsicum annuum
L. est bien connu de par le monde et sous des appellations
différentes suivant les zones géographiques ; Deghi
mirch en Hindi, Pilipili hoho en Swahili,
Sipen ngonpo en Tibétain. Dupriez & De Leener
(1987) ont rapporté les autres noms suivants de la zone tropicale
africaine :
Tableau 4 : Les noms vernaculaires
africains
|
Langues
|
C. annuum, L
|
V. Langues
|
C. annuum, L
|
|
Dendi
|
VI. Touka
|
Maka
|
Bage anini
|
|
Bobo
|
Kepanano sa
|
Mifi
|
Sog ndog
|
|
Dagara
|
Simaa
|
Fufuldé Foulbé
|
Atangou
|
|
Dioula
|
Foronto bani
|
Ashanti
|
Mako
|
|
Mooré
|
Foronto
|
Bambara
|
Furondo kumba
|
|
Banganté
|
Sôg nekale
|
Yoruba
|
Tatase, atarodo
|
|
Bassa
|
Tomato hilôba
|
Mandingue Socé
|
Kano
|
|
Douala
|
Ndongo mindene
|
Malinké
|
Foronto
|
|
Akposso
|
Djakali
|
Mina
|
Gboyébéssé
|
|
Ewondo
|
Ondongo eprwebe
|
Wolof
|
Kani
|
VII. Agronomie de
Capsicum annuum
VII.1. Exigences écologiques
Le piment doux est une des plantes maraîchères
les plus thermophiles. Son développement optimal s'observe dans la
fourchette des températures variant entre 16 et 26° C (Messiaen,
1975).
Même si la plante n'est pas très sensible au
photopériodisme, pour des amplitudes nycto-diurnes faibles (ordre de
5° C) le comportement peut varier selon le niveau de la moyenne
journalière (Chaux & Foury, 1994). Elle requiert d'après
Messiaen (1975) des éclairements inférieurs à 50% du
rayonnement solaire naturel, surtout pour les jeunes plants et son zéro
végétatif se situe à 14° C.
Messiaen (1975) recommande la culture de Capsicum annuum
L. durant les mois frais et dans la mesure du possible, sous ombrage
naturel (papayers, jeunes bananiers ou haies d'arbustes) ou artificiel
(grillages artificiels).
Le piment s'adapte bien à la saison sèche des
climats sahéliens ou `'Sud chinois'' c'est à dire dans les
régions localisées entre 25 à 30 ° de latitude. La
culture se développe de plus en plus en savane durant la saison
sèche.
La plante requiert enfin des sols souples, profonds,
à humidité circulante (Laumonnier, 1979), mais s'adapte assez
bien à une large gamme de sols tant qu'ils sont bien drainés.
Williams et al. (1991) estiment que C. annuum peut avoir une
bonne croissance dans les plaines sableuses. Le facteur humus est d'importance
pour cette culture exigeante par rapport à d'autres Solanacées
telles les aubergines ou la tomate. Le PH convenable semble se situer entre 6.5
et 7.
VII.2. Conduite et pratiques
culturales
VII.2.1. Semis et plantation
En conditions tropicales, le choix variétal se
restreint encore à quelques variétés présentant une
forte rusticité, toutefois il convient de rechercher des
variétés résistantes et susceptibles de forte
productivité.
Les conditions de culture restent les mêmes
d'après Desai et al (1997) quelle qu'en soit la
finalité de la production ; fruits frais pour le marché ou
production grainière.
Les semences provenant de fruits à maturité
complète seront semées en pépinière composée
de terre légère et fraîche (Kroll, 1994), les poivrons se
prêtant mal au semis direct car moins compétitifs vis à vis
des mauvaises herbes. De plus, l'importante pression parasitaire dont ils sont
susceptibles fait que le choix du terrain participe déjà à
la lutte phytosanitaire préventive intégrée contre les
nuisibles telluriques.
Le repiquage en place définitive sera effectué
entre 25 et 40 jours après semis, en doubles lignes (voire triples)
espacées de 1 m avec des écartements variant entre 0,4 et 0,8 m
en tous sens, selon le type variétal utilisé.
Laumonnier (1979) préconise une plantation pas
très profonde, le collet légèrement au-dessus du sol, car
la plante est très sensible à un certain nombre de maladies du
collet.
Il sera nécessaire de prendre en compte les
précédents favorables tels que les Alliacées, les
céréales, la canne à sucre, l'amarante, le chou au
détriment des Solanacées notamment les aubergines et dans une
moindre mesure les Astéracées et les Légumineuses.
VII.2.2. Fertilisation et irrigation
VII.2.2.1. Rôle des principaux
éléments minéraux
L'azote N :
C'est un élément de croissance dont
l'excès doit être évité notamment en phase de
floraison et de fructification. Toutefois sa carence est à l'origine de
branches courtes, rabougries et peu nombreuses avec des petites feuilles
déformées (Mitra, 1990). La couleur de ces dernières
évolue progressivement du vert clair à un vert plus ou moins
jaunâtre et elles se détachent prématurément. De
plus les fruits sont petits, maigres et chlorosés.
Le phosphore P :
C'est un élément de stabilisation de la
plante (croissance radiculaire) et de fructification. Il doit être
optimisé en phase de développement (floraison -
fructification).
En cas de carence de cet élément, Mitra (1990)
dans sa description des symptômes trouve que les feuilles sont petites,
resserrées et incurvées de l'intérieur. Les vieilles
feuilles jaunissent avec des bords roses. Les fruits sont menus et
déformés. D'où, une nutrition correcte de cet
élément influence positivement la résistance de la plante
à certaines maladies.
Le potassium K :
C'est aussi un élément de croissance et de
fructification dont la disponibilité peut-être influencée
par de fortes teneurs en sodium du sol (cas des sols salés).
Sa carence perturbe la croissance de la plante. Le nombre de
feuilles est très réduit, leur taille petite et leur couleur
jaunâtre.
Mitra (1990) rapporte que des petites lésions
nécrotiques peuvent se développer le long des nervures avant
d'entraîner une défoliation.
VII.2.2.2. Estimation des besoins et modalités
pratique de la fumure
Le bilan suivant 117 (N) - 87 (P) - 177 (K) correspondant
à un équilibre 1-0,7-1,5, constitue une moyenne basée sur
trois (03) estimations rapportées, avec un apport moyen de fumier de 15
à 20 t/ha (Tropicasem, 2004). Le CDH préconise dans les
conditions du Sénégal, 90-90-180 qui peut-être couvert
à base de 10-10-20.
Les conditions optimales des apports seront :
· Azote : fractionnement tout au long du cycle
· Phosphore : apport en fond (60 % environ),
disponibilité en début de phase reproductive
· Potassium : disponibilité de la floraison
à la première récolte (15 % en fond, fractionnement)
· Calcium et Magnésium : apport en fond
VII.2.2.3. Les besoins en eau
Ils peuvent se répartir en besoins intrinsèques et
en besoins en termes d'irrigation ; les premiers correspondant à
la quantité d'eau consommée par le couple plante-sol, c'est
à dire à l'évapotranspiration de la culture (ET culture)
qui a été calculée à partir de
l'évapotranspiration de référence (ET0). Ils sont
d'environ 4,5 mm/j soit 4,5l/m²/jour (Tropicasem, 2001). Les seconds en
fonction du mode d'apport (irrigation de surface, aspersion ou
micro-irrigation), du climat, du type de sols et des éventuelles pertes
inhérentes peuvent être synthétisés comme suit
en fonction des stades phénologiques de la culture:
Tableau 5 : Les besoins en eau par système
d'irrigation
|
Système d'irrigation
|
Phase 1(semis-levée ; 20j)
(l/m²/j)
|
Phase 2 (jeune plante ; 50j)
(l/m²/j)
|
Phase 3 (plante adulte ; 230j)
(l/m²/j)
|
Apports totaux (m3/ha)
|
|
Irrigation de surface
|
4
|
6.5
|
11.2
|
30000
|
|
Aspersion
|
3
|
4.5
|
7.8
|
20800
|
|
Micro-irrigation
|
3
|
3
|
5.34
|
14200
|
(Tropiculture n°54 - mars 2001)
VII.2.3. Entretien cultural
VII.2.3.1. Lutte contre les mauvaises
herbes
Le contrôle des mauvaises herbes dont la croissance rapide
affecte l'ensoleillement (De Witt & Bosland, 1993) est une des plus
importantes opérations dans la réussite d'une culture de
poivrons qu'ils compétissent en plus pour les nutriments et
l'humidité du sol. Il peut se faire soit manuellement soit
mécaniquement. Mais l'utilisation d'herbicides devient de plus en plus
fréquente.
Aux stades jeunes, un binage peut s'avérer utile pour
minimiser les mauvaises herbes et ameublir le sol pour en améliorer
l'aération. Plusieurs désherbages seront nécessaires.
VII.2.3.2. Paillage ou mulching
C'est une technique qui s'est avérée efficace en
ce sens que l'application d'un matériel à la surface du sol
retarde la croissance des adventices, conserve l'humidité en maintenant
uniforme la température du sol. La culture a un meilleur aspect et les
rendements s'en trouvent ainsi améliorés
VII.2.3.3. Utilisation des filets
d'ombrage
Ils ont un effet positif sur le bon développement des
plantes. En ce sens, El-Aidy et al (1989) ont démontré
leur nette influence ; les filets d'ombrage améliorent sensiblement
la hauteur des plants, la surface foliaire, les poids frais et secs des plants
ainsi que la qualité des fruits et le rendement total.
VII.2.3.4. Tuteurage
Au fur et à mesure de leur croissance, les plants seront
régulièrement tuteurer afin d'éviter toute rupture des
ramifications qui affectent le rendement.
VII.3. Les contraintes majeures de la
culture
Le poivron est d'autant plus sensible à une grande
variété de maladies que sa culture a lieu en saison humide
où les dégâts sont les plus importants. Il est de plus
passible de nombreuses attaques d'insectes et de parasites, et d'accidents
physiologiques.
VII.3.1. les problèmes phytosanitaires
VII.3.1.1. les maladies virales
D'après Poulos (1987), une trentaine de virus
différents sont supposés attaquer les Capsicum et les
maladies qui s'en suivent sont en général sous les tropiques, les
plus importantes en terme de gravité des dégâts. Elles
surviennent souvent en complexe dans une même culture et dans une
même plante.
· La mosaïque du tabac : elle est
provoquée par le TMV (Tobacco Mosaic Virus) appartenant au type des
tobamovirus. C'est un virus qui se transmet par contact. La maladie est
caractérisée par une mosaïque du feuillage, vert clair
à blanc (Gébré-Sélassié et al.,
1994) et l'apparition de plages mal colorées sur les fruits, ou des
réactions nécrotiques plus ou moins graves selon les stades
végétatifs et les variétés sur les feuilles et les
tiges (Messiaen et al, 1991). La croissance des plantes est ralentie,
les fruits sont marbrés, de maturation irrégulière avec un
aspect souvent bosselé (fig.5)
(Gébré-Sélassié et al., 1994).
La lutte passe par l'utilisation de semences garanties sans
virus, c'est à dire avec une observation rigoureuse des règles
prophylactiques ; désinfection des semences, trempage des graines
contaminées dans une solution à 10 % de phosphate trisodique
pendant 30 minutes. Mais aussi l'utilisation de variétés
résistantes qui reste à l'heure actuelle le moyen le plus
efficace et le plus économique.
· La mosaïque du concombre : c'est une maladie
causée par le CMV (Cucumber MosaicVirus) qui se transmet par
différentes espèces de pucerons (fig.6) sous le mode non
persistant, c'est à dire qu'il est transmissible immédiatement
après la prise de nourriture du vecteur. Et la capacité de
transmettre est de courte durée, quelques minutes selon
Gébré-Sélassié et al. (1994). Elle se
manifeste par des symptômes de ring-spots nécrotiques sur les
feuilles (fig.7) adultes au moment de l'infection, puis une mosaïque
chlorotique et déformante du feuillage ultérieur (on parle de
forme filiforme). Les fruits déjà formés au moment de
l'infection présentent des dessins creux en forme d'anneaux et des
lignes sinueuses qui les déprécient et les rendent sensibles aux
coups de soleil (fig.8). Par contre la fructification ultérieure est
annulée (Messiaen et al., 1991).
Une lutte attentive sera nécessaire contre les pucerons
vecteurs de cette maladie, ceci dès le stade de la production des
plants. (Laumonnier, 1979)
 Figure 5 : Symptômes du TMV sur
fruit Figure 5 : Symptômes du TMV sur
fruit
 Figure 6 : Pucerons sur les
feuilles Figure 6 : Pucerons sur les
feuilles
 Figure 7 : Symptômes de CMV sur une
feuille Figure 7 : Symptômes de CMV sur une
feuille
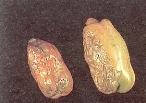 Figure 8 : Symptômes de CMV sur les
fruits Figure 8 : Symptômes de CMV sur les
fruits
D'autres types de virus adaptés suivant les climats et
les continents ont été également
évoqués :
§ Le virus Y de la pomme de terre (PVY) des
régions tempérées, méditerranéennes et
subtropicales de l'Ancien et du Nouveau Monde.
Transmis selon le mode non persistant par de nombreuses
espèces de pucerons. Il se manifeste soit par une mosaïque verte le
long des nervures (`'vein banding'' en anglais), soit par une nécrose
commencant par les nervures et gagnant parfois les pétioles, les tiges
et les fruits. Les feuilles, les fleurs et les fruits les plus atteints tombent
(Gébré-Sélassié et al., 1994).
§ Le Pepper veinal mottle virus (PVMV) africain
Il est encore appelé virus de la panachure du poivron.
Il cause de très sévères déformations du
feuillage ; les feuilles sont très petites, cloquées, avec
une mosaïque verte sur cloques et nervures. Les plants sont rabougris
(Déclert, 1990).
§ Le Tobacco etch virus (TEV) plus spécialement
Nord américain,
§ Le Pepper mottle virus (PMV) du sud des Etats-Unis,
§ Le Chili veinal mottle virus (CVMV) qui sévit
dans le sud-est asiatique,
§ Le virus du flétrissement de la fève ou
Brood bean wilt virus (BBWV) au sud de l'Italie et du Maroc,
§ La mosaïque de la luzerne ou Alfalfa mosaic virus
(AMV)
Dans tous les cas, la plupart des symptômes
d'infestations virales sont des tâches, des marbrures, des
déformations et décolorations des feuilles, la production de
feuilles anormales ainsi qu'une croissance générale rabougrie
(Agromisa, Agrodok n°13 )
VII.3.1.2. les maladies fongiques
· Oïdium : son agent causal Leveillula
taurica a une évolution très rapide marquée par un
feutrage blanc à la face inférieure des feuilles qui
coïncide avec une nécrose en `'point de tapisserie'' (Messiaen et
al, 1991) aboutissant à leur dessèchement et à
une défoliation des plantes (Déclert, 1990). La croissance des
fruits ralentit et la floraison s'arrête. C'est une maladie très
développée par temps chaud en l'absence de pluies. Mais elle est
cependant favorisée par une humidité assez élevée
(70 à 80%) surtout la nuit. Dans les conditions du Sénégal
où elle est considérée comme la principale maladie (CDH,
1986), elle est très redoutée en saison sèche. La
détermination biologique de cette maladie peut se faire par
détection et identification des conidies à la loupe binoculaire
ou par examen microscopique de ruban adhésif appliqué sur la face
inférieure des taches. Les aubergines et la tomate en sont des plantes
hôtes.
· L'anthracnose des fruits : son agent causal est
soit Colletotrichum capsici, soit Colletotrichum nigrum et
elle a des dégâts importants par temps de pluie.
Les symptômes consistent selon Déclert (1990) en
de larges plages de pourritures humides en général sur
l'extrémité ou les flancs des fruits, colorées en brun et
progressivement déprimées. Selon les cas, elles se couvrent d'une
mince gelée rose orangée, masse de conidies d'acervules (cas de
C. nigrum) ou de ponctuations noires en disposition circulaire et
concentrique (cas de C. capsici). Les fruits tombés sur le sol
constituent une source importante de contamination.
· La cercosporiose : elle est causée par
Cercospora capsici en saison de pluies. Sur les feuilles jeunes, on
observe de petites taches grises, arrondies, qui mesurent 2 à 5 mm de
diamètre et deviennent irrégulières,
délimitées par une marge fine, brun foncé et
prolongées par un halo jaune clair. Les tissus nécrosés se
percent par petites plaques. Les feuilles jaunissent, se dessèchent et
tombent. Les répercussions sur la production sont
généralement faibles. (Déclert, 1990)
· La corynesporiose : provoquée par
Corynespora cassiicola en saison des pluies et sous hygrométrie
et température élevées, elle se distingue par des petites
taches foliaires brunes, arrondies à irrégulières, de 4
à 6 mm de diamètre se rapportant à Corynespora.
Elles se caractérisent par leur marge épaisse, constituée
de plis très rapprochés lui donnant un certain relief. Le centre
de la tache se crevasse, mais la trouaison n'est pas totale, les tissus
nécrosés restant en général attachés au
pourtour de la tache. Limitées aux feuilles âgées, les
attaques ne sont pas importantes. (Déclert, 1990)
· Le dépérissement des rameaux : son
agent causal est Choanephora cucurbitarum par temps pluvieux. En
dessous des extrémités, les rameaux jeunes sont atteints de
pourriture brune ; ils se recouvrent d'un duvet clairsemé hyalin
abondant, surmonté de pulvérulence noire (fructification du
champignon parasite, aisément identifiable à la loupe).
Les rameaux atteints s'affaissent, pendent et se
dessèchent. La production des fruits est sévèrement
réduite.
Amaranthus spp, Solanum melongena, le gombo,
le haricot, la laitue, la pastèque sont des plantes hôtes du
pathogène. La lutte devrait consister en la pulvérisation de
bouillies de Captane ou de Thirame (30-40 g/dal) (Déclert, 1990)
· Le dépérissement à
sclérotes : son agent est Sclerotium rolfsii. Les feuilles
jaunissent, se flétrissent et tombent. On observe souvent, au voisinage
immédiat du collet, la présence de petites mèches blanches
soyeuses, plus ou moins agrégées aux particules de terre, et de
petits sclérotes blancs à beiges (brunissant avec le temps).
Les racines sont l'objet d'une pourriture sèche
très importante. Selon le degré d'infestation du sol en
sclérotes (cultures précédentes), les plantations peuvent
être fortement décimées. (Déclert, 1990)
· Le mildiou des fruits : causée par
Phytophthora capsici en saison des pluies. Il se distingue par une
pourriture blême se développant sur la partie apicale du fruit, et
qui apparaît moins déprimée et moins ridée que pour
l'anthracnose.
Les fruits atteints tombent sur le sol, où ils
achèvent leur décomposition.
L'identification de Phytphthora peut-être
rapidement obtenue avec la technique des pastilles de pétunia.
La protection des fruits est assurée par des
pulvérisations de bouillies de Captafol (30-40 g/dal), de Dichlofluanide
(10-20 g/dal) ou d'émulsions de Triforine (10-20 ml/dal).
(Déclert, 1990)
· La moisissure des feuilles : elle est
provoquée par Cercospora unamunoi sous hygrométrie
élevée. La moisissure se manifeste par des lésions
chlorotiques diffuses, subcirculaires à allongées, ayant pour
dimension 3 à 12 mm. Elles se distinguent de celles de l'oïdium par
l'examen de la face inférieure des feuilles, où se trouvent des
taches nécrosées, brun grisâtre, circulaires à
arrondies, mesurant 2 à 10 mm et entourées d'un halo vert clair.
Une plage finement pulvérulente brun olivâtre les recouvre. Les
feuilles atteintes s'enroulent, se dessèchent et se détachent.
Les fruits sont petits et difformes. (Déclert, 1990)
· La stemphyliose : causée par
Stemphylium sp. en saison sèche, cette affection se
caractérise par la présence de petites taches foliaires
arrondies, de 2 à 3 mm de diamètre, grises et annelées de
noir, avec une marge en bourrelet épais ; le centre se fend et se
troue ultérieurement. Les dégâts sont peu importants.
(Déclert, 1990)
· Les taches foliaires blanches : causées par
Mycosphaerella sp, cette affection se caractérise sur les
jeunes feuilles au sommet de la plante par des petites taches, blanches
à grisâtres, mesurant 2 à 4 mm, de contour
irrégulier à anguleux, entourées d'une marge brune, et en
général, localisées au centre de la feuille, le long de la
nervure principale.
Les parenchymes nécrosés se détachent,
créant une trouaison du feuillage. Les dégâts ne sont pas
importants.
Dans le cadre de la lutte, la pulvérisation de
bouillies de Mancozèbe ou de Zirame peut-être recommandée.
(Déclert, 1990)
· Les taches noires duveteuses des fruits : elles
ont pour agent causal est Curvularia sp sous hygrométrie
élevée. On observe une tache noire veloutée sur le flanc
des fruits, recouvrant une plage de pourriture à évolution lente.
Les dégâts ne sont ni importants, ni fréquents.
(Déclert, 1990)
· La fonte de semis : causée par Pythium
aphanidermatum sur sol contaminé ou détrempé.
Déclert (1990) décrit un dépérissement des
plantules, débutant par le flétrissement
généralisé du feuillage. Le plus souvent le collet
apparaît anormalement aminci. Rapidement les collets brunissent, les
plantules versent et disparaissent en quelques jours sous l'effet d'une
macération totale. Arrachées, les racines apparaissent
anormalement courtes et effilochées.
VII.3.1.3. les maladies
bactériennes
· Le flétrissement bactérien : c'est
une maladie importante causée par Ralstonia solanacearum. Ses
symptômes sont un jaunissement et un flétrissement des plus
vieilles feuilles basales. La caractéristique de cette maladie est le
brusque et rapide dessèchement par temps ensoleillé suite
à une période pluvieuse (fig.9).
On peu diagnostiquer la maladie en coupant les racines ou les
tiges. Les plantes affectées produisent une sève sombre et les
tissus radiculaires internes sont colorés en brun.
Les moyens de lutte sont une meilleure prise en compte de
`'l'effet précédent''...
· La moucheture bactérienne : elle est
causée par Xanthomonas axonopodis p.v vesicatoria
décrit en 1994 par Bouzar et al. (Bassim et al.,
2004) , ses symptômes consistent en l'apparition de petites
formations véreuses sur les feuilles et les fruits, que d'autres
pathogènes peuvent utiliser pour infecter les fruits. Les saisons
très pluvieuses favorisent l'infestation qui réduit
considérablement la fructification et la qualité des fruits.
Les rotations culturales et le traitement de semences à
la chaleur (25 minutes à 50° C) sont de bons moyens de lutte
préventive.

Figure 9 : Plante affectée par le
flétrissement
VII.3.2. Les ennemis animaux
Ce sont principalement :
· Les vers des fruits
o Heliothis armigera encore appelé
Helicoverpa armigera (Noctuelle de la tomate) dont les chenilles vert
clair à brun noir (3,5 - 4 cm) trouent et évident les fruits en
les pourrissant.
o Cryptophlebia leucotreta dont les chenilles
rosâtres (1,5 cm) creusent des galeries dans la chair du fruit qui
pourrit.
· La mouche méditerranéenne des
fruits : Ceratis capitata dont la femelle pond ses oeufs sous la
peau des fruits et les asticots se nourrissent de la chair en y creusant des
galeries.
· Le puceron vert : Myzus persicae dont les
importantes colonies sous les feuilles entraînent la déformation
de celles-ci et des pousses terminales arrêtant ainsi la croissance.
· Les acariens, essentiellement la femelle de
Polyphagotarsonemus latus (0,125 à 0,2 mm et invisible à
l'oeil nu) qui, en suçant la sève depuis la face
inférieure des feuilles cause une nécrose.
· Les mouches blanches
Bemisia tabaci dont les adultes tiennent sur la face
inférieure des feuilles et aspirent la sève de la plante
Aleurodicus dispersus qui suce la sève de la
plante depuis la face inférieure des feuilles
VII.3.3. Les accidents physiologiques
Ils sont causés par des carences en nutriments ou des
conditions climatiques extrêmes telles que la sécheresse ou la
température.
· Fissures ou craquelures des fruits suite à un
gonflement trop important du fruit, qui se développent sous l'effet
de trop fortes températures et des variations d'humidité.
Le paillage est un excellent moyen de prévention.
· Coups de soleil : ils sont
caractérisés par des lésions nécrotiques grises
à la surface du fruit dont les parties exposées jaunissent en
premier.
Dans ce cas, des haies vives, sources d'ombrage peuvent
s'avérer utiles
· Les taches noires ou tâches amères
(stip) : causées par la carence en calcium ou par des
concentrations trop élevées de cations comme NH4+, K+, Mg?+ dans
le sol. Dans ce cas la salinité du sol peut être réduite
par un abondant arrosage qui filtrera les sels.
· Absence de graines dans les fruits ; sous l'effet
de température ou d'une luminosité trop faible, la
fertilité des fleurs et par voie de conséquence le nombre de
graines dans les fruits est réduit.
· Nécrose apicale (blossom-end rot)
consécutive à un déficit hydrique important est
favorisée par le manque de calcium. Les jeunes fruits sont plus
sensibles que les fruits à plein grossissement ; la partie
terminale des fruits devient beige.
VII.4. Récolte et
conservation
Selon les variétés et le but de la
production, la période de récolte peut varier. Les poivrons sont
généralement récoltés verts (fig.10), manuellement,
avec leur pédoncule lorsque les fruits n'ont pas encore atteint la
maturité complète car étant d'un meilleur rapport
(Beniest, 1987) quoique nutritionnellement meilleurs à l'état
mûr (jaunes ou rouges selon les variétés. Il est
recommandé de prévoir la récolte entre 50 et 55 jours
après la floraison pour les fruits verts (Laumonnier, 1979) ou 60
à 80 jours repiquage (Beniest, 1987). Dans ce cas, les risques de
pourriture, d'attaques d'insectes ou de coups de soleil sont limités.
La récolte peut durer trois (03) mois ou plus tant que
les niveaux de maladies sont contrôlables ou alors que la production
devient insignifiante.
Les rendements en fruits frais sont de l'ordre de 10 à
20 tonnes à l'hectare mais théoriquement des rendements de 40
tonnes sont possibles alors que les rendements en grains sont de l'ordre de 120
kg/ha.
Pour Chaux & Foury (1994), le piment C. annuum
peut être conservé plusieurs semaines en chambre froide à
7-10° C sous H.R. de 90-95 %. Un film perforé donne satisfaction
pour le maintien de la qualité du produit.
Dans la pratique, la durée d'entreposage peut atteindre
le mois.

Figure
10 : Aperçu des fruits récoltés
VIII. Génétique du piment (Capsicum annuum
L.)
VIII.1. Relations
cytogénétiques
Tous les Capsicum ont un nombre de chromosomes
identique : 2n = 2x = 24.
De nombreuses études ( morphologiques, résultats
d'intercroisements, zymogrammes) ont permis de s'accorder sur l'existence de
deux groupes distincts : l'un à fleurs violettes et l'autre
à fleurs blanches ( Chaux & Foury, 1994). C'est pourquoi Pochard et
al (1992) ont indiqué que C. annuum ne peut se
croiser avec le groupe d'espèces à fleurs violettes. Par contre,
il se croise facilement avec C. chinense et difficilement avec C.
baccatum.
Ces mêmes auteurs évoquent aussi les
différences de translocations entre espèces, C. annuum
différant de C. chinense et de C. baccatum
respectivement par une et trois translocations.
Les travaux de Greenleaf (1986) sur les possibilités de
croisements interspécifiques et la fertilité des hybrides issus
de ces croisements et les caryotypes de quelques espèces nous
renseignent davantage sur les possibilités ou non de certains
croisements interspécifiques.
VIII.2.
Génétique des caractères
Elle pourrait se résumer aux caractères
recensés dans le tableau suivant :
Tableau 6 : Génétique des
caractères
|
Caractère
|
Nombre de gènes
|
Types d'action du gène
|
|
Orientation du fruit
|
Monogénique (up+ up+)
|
Etat pendant domine érigé
|
|
Couleur du fruit
|
Monogénique
|
Rouge domine orange
|
|
Longueur du pédicelle
|
Monogénique
|
Court domine long
|
|
Saveur piquante
|
Monogénique
|
Piquante domine douce
|
|
Forme du fruit
|
Monogénique
|
Raccourcie ou arrondie domine forme anguleuse
|
|
Apex du fruit
|
Monogénique
|
Pointe incomplète domine carrée
|
|
Texture de la chair
|
Monogénique
|
Douce,molle,fruit facilement séparé du calice
domine fruit fortement attaché au calice
|
|
Tenure de la plante
|
Monogénique
|
Erigé,grand,élevé domine
écrasé
|
|
Envergure de la plante
|
Monogénique
|
Déployé domine érigé
|
|
Lobes des fruits
|
Monogénique
|
Multilobé domine bilobé
|
|
Production des grains
|
Monogénique
|
« graineux » domine sans grains
|
|
Stérilité complète
|
|
Fertilité complète domine
stérilité
|
|
Stérilité femelle
|
|
Fertilité femelle domine stérilité
|
Source : Génétique des caractères
(Peter K. V., 1998)
VIII.3.
Qualité et sélection du piment
VIII.3.1. Critères de
qualité
Ils sont nombreux et variables suivant les habitudes de
consommation et d'usages des fruits de piment (Capsicum annuum L.).
Ils déterminent ainsi les objectifs de sélection. Et
au-delà de la recherche d'idéotypes adaptés aux conditions
culturales des zones tropicales chaudes et humides qui nous intéressent,
nous pouvons retenir à la suite de Hennart (1996), quoique la notion de
qualité soit difficile à cerner, que les fruits de qualité
présentent les caractéristiques suivantes :
Ä régularité du point de vue de la forme et
du calibre qui dépend de la disposition sur la plante ; les fruits
se gênant souvent les uns les autres lorsqu'ils sont volumineux, surtout
lors de la première nouaison,
Ä la paroi doit être généralement
épaisse pour faciliter la manutention et la conservation,
Ä la coloration doit être la plus foncée
possible, en vert ou à maturité,
Ä l'existence d'une extrémité à 4
loges est également un critère de qualité.
Par ailleurs Chaux & Foury (1994) signalent en plus que
les critères de fermeté, d'intégrité
représentent des normes à la commercialisation. Et selon les
normes françaises, pour un fruit de première
catégorie la largeur ne doit pas être inférieure
à :
o poivron long pointu : 30 mm
o forme carrée, épointée : 50 mm
o forme carrée, pointue : 40 mm
o forme aplatie (« tomate ») : 55
mm
VIII.3.2. Méthodes de
sélection
Selon Poulos (1993), les méthodes
de sélection les plus appropriées sont la sélection
massale, la sélection de lignées pures, le backcross ou
rétrocroisement, la méthode pedigree et la sélection
généalogique par filiation monograine ou unipare (ou Single Seed
Descent, SSD). Leur choix est fonction d'après Moreno-Gonzalez &
Cubero (1993) de certains facteurs tels le système de reproduction,
l'hétérosis, la structure cytogénétique, la
différenciation entre les caractères qualitatifs et quantitatifs,
l'action des gènes, l'héritabilité et la combinaison entre
méthodes classiques et spéciales
VIII.3.2.1. Sélection
massale
Les individus reproducteurs sont choisis dans la masse des
plantes cultivées et leur descendance est elle-même
cultivée en masse, sans séparation. Conséquences :
· il y a un seul cycle de reproduction par an,
· l'efficacité de la sélection est
influencée par la précocité d'expression des
caractères
· la valeur de la sélection faite dépend de
l'héritabilité des caractères
VIII.3.2.2. Méthode
généalogique différée (ou bulk)
Elle est une suite de la précédente qui a pour
effet la mise en dose double des caractères récessifs, donc leur
apparition, ce qui se révèle un avantage dans la mesure où
ces caractères récessifs sont intéressants.
VIII.3.2.3. La filiation unipare (ou
SSD)
Méthode utilisée surtout pour les plantes
à cycle court, donnant plusieurs générations par an.
Pendant 6 à 7 générations, on prélève une
graine par plante et on la ressème. On garde ainsi un exemplaire au
moins de chaque plante de la population de départ, sans en
éliminer.
Ensuite on pratique une sélection
généalogique sur ces plantes devenues homozygotes suite
aux multiplications successives.
VIII.3.2.4. Le backcross
Il est utilisé surtout pour introduire un
caractère déficient chez certaines lignées qui manquent
d'une qualité même si l'ensemble des autres est satisfaisant. On
l'a principalement utilisée pour introduire des gènes de
résistances aux maladies ou modifier un caractère biochimique ou
alors pour l'introduction de stérilité mâle dans le but de
créer des variétés hybrides
VIII.3.2.5. La sélection
récurrente
On peut évoquer la création de
« populations sources » par sélection
récurrente qui serviront par d'autres, à la sélection de
variétés fixées, homogènes et stables.
La méthode est dite récurrente car elle
procède par cycles successifs ; la population
améliorée produite à la fin de chaque cycle sert
aussitôt de matériel de départ pour le cycle suivant. Le
nombre de cycles étant très variable, des plantes de ces
populations peuvent à tout moment être extraites pour la
création variétale.
VIII.4. Facteurs
influençant la production de semences hybrides
VIII.4.1. La qualité d'une
semence
De façon générale, elle peut être
appréciée à travers divers critères quantitatifs
définis par les normes internationales (ISTA) dont les plus importants
sont :
o Le pouvoir initial de germination,
o La vigueur germinative,
o La pureté spécifique et variétale,
o L'état sanitaire,
o Le taux d'humidité,
o Le poids de 1000 graines en rapport avec le nombre moyen de
graines au gramme
VIII.4.2. Les hybrides et la production
de semences
Les hybrides ne peuvent être reproduits par leurs
utilisateurs car ils entreraient en disjonction génétique,
altérant l'homogénéité et le niveau de
productivité de la variété.
La production de semences hybrides est réalisée
dans des conditions pratiques qui visent un double objectif :
o Maximiser le rendement en semences hybrides à travers
un dispositif qui assure un taux maximum de plantes femelles car elles
présentent le meilleur rendement en garantissant une meilleure
qualité,
o Eviter les autofécondations ou les hybridations
indésirables car l'homogénéité et la pureté
génétique des hybrides exigent d'éviter les pollinisations
parasites du parent femelle provoquées par du pollen étranger ou
par une autofécondation
VIII.4.3. Expression de
l'hétérosis
L'expression de l'hétérosis qui désigne
la supériorité des hybrides de première
génération sur les parents impliqués dans le croisement,
c'est à dire l'augmentation de la vigueur de la descendance, est
influencée par les facteurs environnementaux.
La qualité de quelques hybrides peut varier au fil des
ans en fonction des conditions qui ont prévalu au cours de
l'année de production des semences. Kalloo (1989) révèle
que les meilleures conditions pour une production de semences de qualité
et en quantité sont : une température moyenne,
l'intensité lumineuse, l'humidité et une bonne fertilité
du sol.
Chez Capsicum annuum, Kalloo (1989) tout comme Doshi
& Shukla (2000) rapportent des taux d'hétérosis
élevés pour le rendement en fruits frais, le nombre de fruits par
plante, leur longueur, leur épaisseur et leur taille.
VIII.5.
Justification de l'utilisation de C. chinense
Parce qu'il n'existe pas toujours à
l'intérieur de l'espèce travaillée les
caractéristiques que l'on désire introduire dans la
variété à créer, on fait appel à des plantes
d'espèces voisines ; formes spontanées, résistantes
(Demarly & Sibi, 1989). Pour Palloix et al (1995), la recherche de
gènes de résistance doit être menée dans des
variétés ou des espèces très différentes du
type variétal souhaité. Ainsi, la plupart des résistances
ont été découvertes dans des piments cultivés ou
sauvages dont les plantes parfois pérennes en zone tropicale produisent
des petits fruits « brûlants ». Sous ce rapport, de
nombreux auteurs ont évoqué plusieurs raisons liées aux
caractéristiques de C. chinense pour justifier son utilisation
comme source pour le transfert de gènes de résistance sur C.
annuum. Parmi eux :
Ö Saccardo & Ramulu (1977) ont
révélé que C .chinense était une
importante source de résistances aux maladies et présentait des
taux de fructification ainsi qu'un rendement en grains par plante
intéressants chez les produits d'un back-cross d'hybrides F1,
principalement lorsque C. chinense est femelle ;
Ö Kounowsky et al (1985) ont affiné les
recherches en précisant que C .chinense était porteur de
résistances au TMV, au TEV, et à Verticillium entre
autres ;
Ö Les travaux de Cheng (1988) ont
révélé enfin d'intéressantes
caractéristiques comme un péricarpe doux, vert foncé de
certaines variétés de C. chinense semblables aux C
.annuum bien que les fruits soient plus petits et les formes
différentes. Les C. chinense croissent en conditions humides
comme pérennes deux (02) à quatre (04) années exempts de
maladies ou d'attaques d'insectes. Leur période de production est longue
et les fruits sont croustillants, doux et sans aigreur.
Ce sont là, autant de raisons valables qui pourraient
justifier et servir à l'étude du potentiel de fructification
sous conditions difficiles, dans notre cas, humides, et d'augmenter la taille
des fruits et l'épaisseur du péricarpe chez C.annuum.
IX. Un point sur la recherche...
Depuis la création des premiers types de poivrons
hybrides ( comme le « Lamuyo » créé en 1973
par l'INRA), la recherche sur piment n'a cessé d'évoluer et les
approches sont désormais aussi nombreuses que variées en
s'orientant par exemple vers « la résistance
génétique aux bioagresseurs » (Palloix et al.,
2001). La recherche comprend toutefois plusieurs axes, depuis la cartographie
du génome jusqu'aux outils pour la sélection et l'innovation
variétale en passant par la construction de résistances,
l'analyse de résistances aux parasites ou encore l'estimation d'indices
de distances génétiques entre les variétés.
I. Cadre de l'étude
I.1. Localisation
L'essai a été mené à la station
`'Baobab'' de Tropica Sem sise à Keur Ndiaye Lô,
localité située dans le département de Rufisque, c'est
à dire dans la zone climatique subcanarienne qui caractérise la
bande côtière du Sénégal : les Niayes
(fig.11).
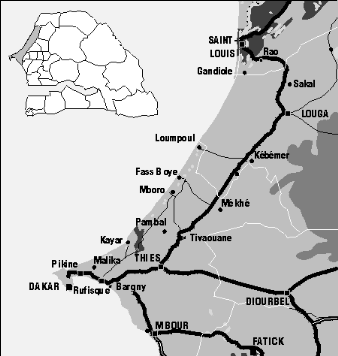
Figure
11 : Situation de la zone des Niayes
I.2. Agro-écologie du site
Ce milieu assez original est constitué des dunes et
dépressions souvent inondées par l'affleurement de la nappe
phréatique. La zone est caractérisée par la circulation
des alizés maritimes soufflés par les courants froids des
Açores. On y distingue une saison humide concentrée sur trois
mois (juillet, août et septembre) et une saison sèche qui dure les
autres neuf mois (Fall et al., 2001).
La température mensuelle la plus chaude oscille autour
de 27.5 °C et survient en juillet et août. Elle peut
légèrement s'élever selon Fall et al. (2001) du
fait de la présence de l'harmattan. Un important taux d'humidité
relative est lié à la proximité de la mer.
Les précipitations sont dictées par la
présence de la mousson en provenance du sud issue de l'anticyclone de
Sainte-Hélène durant l'hivernage. (Fall et al., 2001)
Les sols de type minéraux à pseudo gley sont
argilo-sableux, légèrement calcaires, assez lourds, riches en
matières organiques, avec une forte capacité de
rétention.
La végétation est constituée d'une
savane arborée clairsemée où les espèces dominantes
sont Adansonia digitata, Acacia raddiana ou encore
Zizyphus mauritiana. Du fait de la présence dans la zone de
nombreuses exploitations, des espèces nouvelles telles
Leuceana.., Eucalyptus, Camadulensis
utilisées comme brise-vents sont de plus en plus abondantes. Le tapis
herbacé est essentiellement composé de Graminées
saisonnières et d'Euphorbiacées.
Durant la période couvrant la durée de notre
essai, les données consignées dans le tableau X suivant ont
été enregistrées par les services
météorologiques.
Tableau 7 : Relevé
météorologique de l'hivernage 2004
|
Juillet
|
Août
|
Septembre
|
Octobre
|
Novembre
|
Décembre
|
|
Température moyenne (°C)
|
27
|
27,8
|
28,4
|
27
|
25,4
|
18,6
|
|
Température maximum (°C)
|
30
|
31,5
|
33,5
|
37,6
|
33,6
|
28,2
|
|
Température minimum (°C)
|
23
|
21,5
|
24
|
21
|
17
|
14
|
|
Humidité relative (%)
|
87,5
71,6
|
88,4
69,5
|
91,7
70
|
87
53
|
86,4
41,2
|
88,6
47,6
|
|
Total des précipitations (mm)
|
92
|
118
|
80
|
-
|
-
|
-
|
( Source : Service météorologie Dakar-Yoff,
2004)
II. Matériels et
Méthode
II.1. Matériel
végétal
A l'origine des lignées de piments issues de
populations de petits fruits de type chinense (Chi 59 et Chi 45),
brûlants, choisis pour leur résistance et leur adaptation à
l'hivernage mais aussi pour leur forte productivité, ont
été croisées à des variétés de
piments doux à gros fruits d'origines géographiques diverses
(poivrons tropicaux et poivrons tempérés)
sélectionnés ou pour leur adaptation en saison fraîche et
sèche ou pour leurs caractéristiques horticoles.
La première génération des produits de
ces croisements qui a subi un premier backcross, a donné le
matériel d'étude dont il est question d'en faire une
évaluation.
II.1.1. Listing des parents initiaux
Tableau 8 : Liste des parents
initiaux
|
Code génotype
|
Caractéristique
|
|
010239
|
Poivron doux tropical 1
|
|
010240
|
Poivron doux tropical 2
|
|
010241
|
Poivron doux tropical 3
|
|
012042
|
Poivron doux tropical 4
|
|
010249
|
F1 (Vania x Chi 8)
|
|
010250
|
Vania, poivron tempéré résistant à
CMV et TMV
|
|
010255
|
F1 (Vania x Chi 45)
|
|
010258
|
(F1010239 x Chi 59)
|
|
010259
|
I1(F1010239 x Chi 59)
|
|
010260
|
BC1[(F1010239 x Chi 59)1
|
|
010172
|
Se 100
|
Obtentions INRA, résistantes au potyvirus
|
|
010173
|
Se 101
|
|
010174
|
Se 102
|
|
010175
|
Se 103
|
|
010176
|
Se 104
|
|
001225
|
Piment résistant
|
II.1.2. Tableau général des
croisements
Dans ce tableau, tous les croisements recensés ont
abouti à la constitution du matériel de départ de notre
étude.
Tableau 9 : Récapitulatif des
croisements (Cf. page suivante)
|
010172
|
010173
|
010174
|
010175
|
010176
|
010258
|
010259
|
010250
|
001225
|
010242
|
010260
|
010239
|
|
Se 100
|
Se 101
|
Se 102
|
Se 103
|
Se 104
|
(F1010239 x Chi 59)
|
I1(F1010239 x Chi 59)
|
Vania
|
????
|
|
BC1[(F1010239 x Chi 59)1
|
|
|
010239
|
IX.1. X
|
IX.2. X
|
x
|
x
|
x
|
x
|
x
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
IX.3. 010259
|
|
|
|
|
|
|
|
IX.4. x
|
|
|
|
x
|
|
I1(F1010239 x Chi 59)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
010250
|
|
|
|
|
|
|
|
|
IX.5. x
|
x
|
x
|
|
|
Vania
|
|
|
|
|
|
|
|
|
|
|
|
|
|
010249
|
|
|
|
|
|
|
|
IX.6. x
|
|
|
|
|
|
F1 Vania x Chi 8
|
|
|
|
|
|
|
|
|
|
|
|
|
|
010241
|
IX.7. X
|
X
|
X
|
X
|
X
|
|
|
|
IX.8. IX.9. IX.10. IX.11. X
|
|
|
IX.12. IX.13. X
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
010242
|
IX.14. X
|
IX.15. X
|
X
|
X
|
|
|
|
|
IX.16. IX.17. IX.18. IX.19. X
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
010240
|
x
|
x
|
x
|
x
|
x
|
|
|
|
IX.20. IX.21. IX.22. IX.23. X
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
010255
|
|
|
|
|
|
|
|
|
|
|
IX.24. x
|
|
|
F1 (Vania x Chi 45)
|
|
|
|
|
|
|
|
|
|
|
|
|
II.1.3. Matériel
testé
Des génotypes issus du matériel obtenu par les
précédents croisements, rétrocroisés, ont
constitué notre matériel listé dans le tableau ci-dessous,
auxquels ont été associés deux témoins de
référence ; 001237 pour la qualité
et le type de fruits et 040133 pour la résistance aux
maladies et l'adaptation en saison fraîche et sèche.
Tableau 10 : Liste du matériel
total testé ( Cf. page suivante)
X. CodeGénotypes
|
Génération
|
Fournisseur
|
Parents
|
Ancêtre
|
|
030187
|
BC
|
TRS
|
010239x010172
|
|
|
030188
|
BC
|
TRS
|
010239x010173
|
|
|
030189
|
BC
|
TRS
|
010239x010174
|
|
|
030190
|
BC
|
TRS
|
010239x010175
|
|
|
030191
|
BC
|
TRS
|
010239x010176
|
|
|
030192
|
BC
|
TRS
|
010239x010258
|
|
|
030193
|
BC
|
TRS
|
010239x010259
|
|
|
030194
|
BC
|
TRS
|
010240x010172
|
|
|
030195
|
BC
|
TRS
|
010240x010173
|
|
|
030196
|
BC
|
TRS
|
010240x010174
|
|
|
030197
|
BC
|
TRS
|
010240x010175
|
|
|
030198
|
BC
|
TRS
|
010240x010176
|
|
|
030199
|
BC
|
TRS
|
010241x010172
|
|
|
030200
|
BC
|
TRS
|
010241x010173
|
|
|
030201
|
BC
|
TRS
|
010241x010174
|
|
|
030202
|
BC
|
TRS
|
010241x010175
|
|
|
030203
|
BC
|
TRS
|
010241x010176
|
|
|
030204
|
BC
|
TRS
|
010242x010172
|
|
|
030205
|
BC
|
TRS
|
010242 x G10010173
|
|
|
030206
|
BC
|
TRS
|
010242 x 010174
|
|
|
030207
|
BC
|
TRS
|
010242 x 010175
|
|
|
030208
|
BC
|
TRS
|
010249 x 010250
|
|
|
030209
|
BC
|
TRS
|
010250 x 001225
|
|
|
030210
|
BC
|
TRS
|
010250 x 010242
|
|
|
030211
|
BC
|
TRS
|
010250 x 010260
|
|
|
030212
|
BC
|
TRS
|
010255 x 010250
|
|
|
030213
|
BC
|
TRS
|
010259 x 010239
|
|
|
030214
|
BC
|
TRS
|
010259 x 010250
|
|
|
040199
|
BC1
|
TRS
|
|
030187
|
|
040200
|
BC1
|
TRS
|
|
030187
|
|
040201
|
BC1
|
TRS
|
|
030187
|
|
040202
|
BC1
|
TRS
|
|
030187
|
|
040203
|
BC1
|
TRS
|
|
030188
|
|
040204
|
BC1
|
TRS
|
|
030188
|
|
040205
|
BC1
|
TRS
|
|
030188
|
|
040206
|
BC1
|
TRS
|
|
030189
|
|
040207
|
BC1
|
TRS
|
|
030189
|
|
040208
|
BC1
|
TRS
|
|
030189
|
|
040209
|
BC1
|
TRS
|
|
030190
|
|
040210
|
BC1
|
TRS
|
|
030190
|
|
040211
|
BC1
|
TRS
|
|
030190
|
|
040212
|
BC1
|
TRS
|
|
030190
|
|
040213
|
BC1
|
TRS
|
|
030191
|
|
040214
|
BC1
|
TRS
|
|
030191
|
|
040215
|
BC1
|
TRS
|
|
030191
|
|
040216
|
BC1
|
TRS
|
|
030191
|
II.2. Méthodologie
II.2.1. Dispositif
expérimental
Pour cette étude qui se devait essentiellement de
caractériser nos différents génotypes de façon
à mieux appréhender et cerner leurs comportements
héréditaires respectifs, nous avons adopté un dispositif
en blocs, sans répétition, avec des lignes simples.
II.2.2. Fiche technique de l'essai
Ö Le semis a été
effectué le 27 juillet 2004 sur plaques alvéolaires avec du
terreau professionnel. Pour chaque génotype, nous avons semé 1
à 2 graines par alvéole, selon la disponibilité en
semences. Les plaques ont ensuite été placées sous
ombrière dans le tunnel réservé aux
pépinières.
Ö La plantation en place définitive
a eu lieu le 08 septembre 2004 sur la parcelle C1 d'une superficie de 2500
m². Un mois avant, le sol a subi un labour profond et une quinzaine de
jours avant plantation, un hersage et un passage au vibroculteur ont
été effectués.
Ö Les écartements
pratiqués : 0,40 cm sur les lignes et 0,60 cm entre les lignes
Ö La protection phytosanitaire a
consisté au cours du cycle en une série de traitements tant
préventifs que curatifs dont les détails sont consignés
dans le tableau 12
Ö L'irrigation était assurée
par un dispositif élémentaire de « goutte à
goutte » (fig.12) à 2 bars pendant 45 mn à 1 heure
chaque jour.
Ö Entretien des cultures :
Ä Du semis à la plantation,les plants ont
été régulièrement arrosés et traités
au Rovral contre les agents de fonte de semis
Ä Des sarclo-binages ont été effectués
à la demande afin de supprimer les adventices
Ä Un tuteurage a permis de maintenir les plantes bien
dressées face à l'action du vent et sous le poids des fruits.
Ö La fertilisation a consisté en
l'apport d'éléments fertilisants à P + 40, P + 60 et P +
75 jours comme présenté sur le tableau :
Ö La récolte a eu lieu à la
maturité physiologique, au stade vert, sur les plantes utiles.
Tableau
11 : Suivi de la fertilisation
|
Période
|
Eléments
|
Quantité
|
|
P+40
|
18-46-00
|
7kg
|
|
K2SO4
|
4 kg
|
|
Urée
|
4 kg
|
|
P+60
|
Urée
|
6 kg
|
|
Sulfate de potasse
|
18 kg
|
|
P+75
|
K2SO4
|
10 kg
|
|
Nitard mix
|
2 kg
|
|
9-23-30
|
10 kg
|
|
Urée
|
10 kg
|
Tableau 12 : Traitements phytosanitaires
effectués
|
Date
|
Produit utilisé
|
Ravageurs contrôlés
|
|
Nom
(formulation)
|
Matière active
|
Dose appliquée
|
|
17/09/04
|
dequinèbe
|
|
|
|
|
|
|
|
|
|
06/10/04
|
Charamba
Vélizia
|
|
|
|
|
11/10/04
|
Acaron
Vélizia
|
|
|
|
|
12/10/04
|
Rovral (P.M)
|
Iprodione
|
15g /10L
|
|
|
22/10/04
|
Spiclopride
Vélizia
|
|
|
|
|
26/10/04
|
Vélizia
|
|
|
|
|
28/10/04
|
Orthène 50
Anvil
Vélizia
|
Acéphate
|
7,5g / 10L
|
Insectes
|
|
10/11/04
|
Décis
Vélizia
Néolon
|
Deltaméthrine
|
12,5g/ha
|
Aleurodes,thrips
|
|
12/11/04
|
Vélizia
|
|
|
|
|
22/11/04
|
Acaron
Vélizia
|
|
|
|
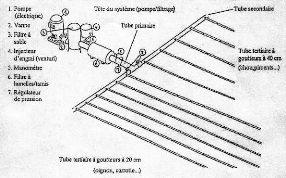
Figure 12 : Système
élémentaire pour `'goutte à goutte''
II.2.3. Observations
réalisées
Elles ont porté sur les caractéristiques du
développement végétatif, de la production et de la
tolérance aux maladies et ravageurs.
II.2.3.1. Pendant la phase
végétative
II.2.3.1.1. Caractères
quantitatifs
â Croissance végétative
Le rythme de croissance de plantes a pu être
apprécié à travers d'une part, l'évolution de la
levée des différents génotypes semés et d'autre
part à travers la hauteur des plantes et le niveau d'encombrement
foliaire associé par conséquent aux dimensions foliaires.
Les mesures de la hauteur des plantes et du niveau
d'encombrement ont été effectuées respectivement à
P + 34 j (c'est à dire 34 jours après plantation) et à P +
57 j (57 jours après plantation) sur 10 plantes choisies au hasard, mais
assez représentatives de l'ensemble des plants, pour chaque
génotype.
Dans le cas des dimensions foliaires, elles ont
été mesurées sur 5 plantes pour chaque génotype,
alors que l'évolution de la levée chez les différents
génotypes de notre étude a consisté en un suivi de la
vitesse de germination.
Remarque : pour les
génotypes dont le nombre de plantes après semis était
inférieur à 10, les moyennes ont été faites sur un
nombre de plantes moindre pour ce qui est de la mesure de la hauteur ou du
niveau d'encombrement.
â Suivi de l'évolution de la
levée :
L'opération a consisté à établir
sur la base du nombre de jours entre la date de semis et la date de germination
de la graine, la durée de la levée. L'apparition effective d'un
bourgeon de germination a été le critère pris en compte
pour chaque génotype.
â La hauteur des plantes :
Sur chaque plante retenue, la plus grande hauteur a
été mesurée à l'aide d'un décamètre.
Puis la moyenne de toutes les hauteurs a été calculée
pour chaque génotype.
â L'encombrement foliaire :
Son évaluation a consisté à mesurer de
façon croisée, les deux plus grands diamètres de la
frondaison de chaque plante retenue, d'en calculer une `'surface
d'encombrement'' puis d'en faire la moyenne sur l'ensemble des plantes retenues
pour chaque génotype.
â Les dimensions foliaires :
Une plante assez représentative de l'ensemble a
été choisie dans chaque génotype en fonction de sa bonne
tenue. Dix (10) feuilles prélevées sur les étages
inférieurs (05) et sur les étages supérieurs (05) ont
été mesurées suivant la plus grande longueur et la plus
grande largeur.
II.2.3.1.2. Caractères
qualitatifs
Les observations ont porté essentiellement sur le port
des plantes, la vigueur générale ainsi que l'état
sanitaire général.
Le port :
Il s'agissait d'établir une distinction entre les types
de port des plantes de nos différents génotypes.
La vigueur générale :
Sur la base d'une appréciation visuelle de l'ensemble
des plantes, chaque génotype a été classé
suivant 6 classes caractérisant 6 niveaux d'observations.
L'appréciation des niveaux se faisant de façon croissante.
(niveau 1 = manque total de vigueur, plante presque tombante, niveau 2 = faible
vigueur, niveau 3 = vigueur moyenne, niveau 4 = assez bonne vigueur, niveau 5 =
bonne vigueur, niveau 6 = excellente vigueur ou plante vigoureuse)
L'état sanitaire général :
Il a été observé sur chaque
génotype en établissant un listing des symptômes
pathologiques notés sur l'ensemble des plantes.
II.2.3.2. Pendant la phase
générative
II.2.3.2.1. Caractères
quantitatifs
Les observations ont porté essentiellement
sur :
La précocité de la floraison :
Elle a été estimée à travers le
nombre de jours entre la plantation et l'apparition des boutons floraux sur au
moins une plante pour chaque génotype. En même temps, une
distinction a été établie en listant les numéros
sur lesquels l'anthèse avait déjà eu lieu.
Le nombre de fruits par plante ou encore niveau de
nouaison :
Il a été mesuré en comptant sur... pieds
par numéros, le nombre de fruits noués
Le rendement et ses composantes :
o Rendement génotypique
o Poids moyen des fruits
o Calibre / dimensions des fruits
o Rendement moyen grainier génotypique
II.2.3.2.2. Caractères
qualitatifs
La couleur des fruits immatures : suivant
l'intensité, la couleur des fruits à été
classée sur une échelle de 1 à 6.
La couleur des fruits à maturité : les
fruits ont été classés suivant qu'ils soient Rouges ou
Jaunes.
L'état sanitaire
II.2.4. Traitement statistique des
données
Ici, il s'agit pour nous d'essayer d'avoir une meilleure
connaissance de notre matériel végétal à
travers:
Une caractérisation des différents
génotypes sur la base de leurs caractères
végétatifs et génératifs,
Une évaluation de leurs rendements respectifs ainsi
que des composantes de ceux-ci.
Pour ce faire, nous avons eu recours à l'utilisation du
tableur Microsoft EXCEL et du logiciel SPSS avec lequel certaines
données ont fait l'objet d'une analyse factorielle discriminante.
II.2.5. Limites
méthodologiques
Les résultats de cette étude auraient pu
être encore plus pertinents si nous n'avions pas rencontré
quelques entraves au bon déroulement de nos travaux comme :
o Un démarrage quelque peu tardif lié à
des problèmes de calendrier académique
o Un défaut logistique lié à l'absence de
carmin acétique qui nous aurait permis de déterminer l'index de
fertilité pollinique des plantes
III. Résultats et
discussion
III.1. Evaluation du matériel
végétal
III.1.1. En phase
végétative
III.1.1.1. Caractères
quantitatifs
III.1.1.1.1. Evolution de la levée des
plantes
La levée pour nos différents génotypes a
eu lieu entre 7 et 20 jours après semis et le diagramme qui suit
représente pour chaque date, la proportion des génotypes
relatifs.
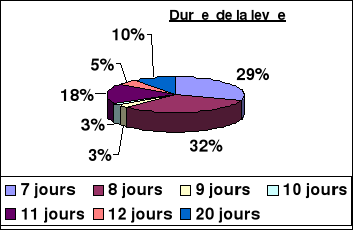
Figure 13 : Répartition de la
levée
Nous voyons que 10% de nos génotypes sont quelque peu
en marge avec des levées au bout de 20 jours. Mais dans l'ensemble, 67%
des génotypes ont levé entre 7 et 10 jours qui correspondent
respectivement à la duré de la levée chez les
témoins 001237 et 040133.
En considérant que la levée chez C.
annuum intervient entre 3 et 7 d'après Chaux & Foury (1994), on
peut dire que tous les génotypes ayant levé au-delà de 7
jours sont un peu tardifs et au-delà de 12 jours on peut parler de
levée tardive.
Toutefois, la distribution pour les différents
génotypes se présente comme suit :
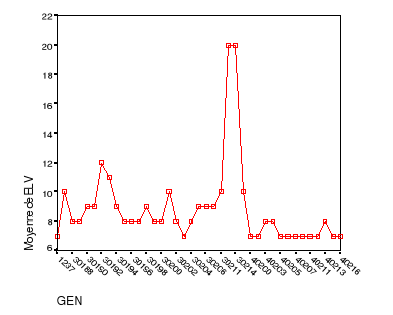
Figure 14 : Evolution de la
levée
Pour les principales familles, on note que :
030187 a levé au bout de 10 jours
alors que 040201 et 040202 ont levé
au bout de 7 jours.
030188 a levé au bout de 8 jours comme
040203, 040204 alors que
040205 a levé au bout de 7 jours.
030189 a levé en 8 jours contre 7 pour
040206, 040207 et 040208
030190 a levé en 9 jours contre 7
jours pour 040211 et 040212
030191 a levé en 9 jours alors que
040213, 040214 et 040215 ont
levé au bout de 7 jours
Dans tous les cas,on remarque que 10 % de nos génotypes
sont très largement en marge avec une levée au bout de 20 jours
(030214, 030213)
III.1.1.1.2. Evolution de la hauteur des
plantes
Elle a été appréciée dans un
premier temps à travers la hauteur moyenne des plantes à mi-cycle
dans chaque génotype et le graphique qui suit nous montre
l'évolution moyenne de la hauteur des plantes :
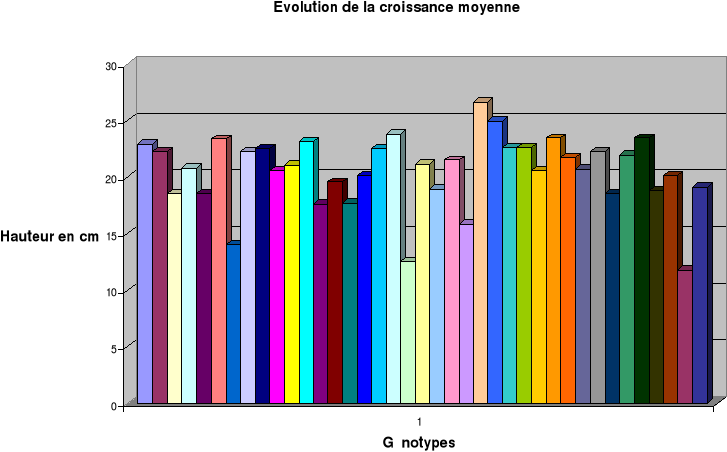
Figure 15 : Evolution de la croissance
moyenne
Ä Seuls 3 génotypes ont une hauteur moyenne en
dessous de 15 cm (030214, 040412, 030213)
Ä 1 seul génotype a une hauteur moyenne de plus de
25 cm (030191)
Ä L'essentiel des génotypes ayant une hauteur
moyenne comprise entre 15 et 25 cm
Ä Nos témoins 9 (001237) et 13 (040133) culminent
respectivement à 22.5 cm et à 17.6 cm d'où leur moyenne de
20,5 cm qui semble être celle d'une grande partie de nos
génotypes.
Pour ce qui est des principales familles, on note
essentiellement une nette différence entre 030188 et
040205 (respectivement 19.5 cm contre 13.1 cm) et une
légère entre 030189 (17.7 cm) et
040206 (20.11 cm), 040207 (22.5 cm) et
040208 (23.8 cm).
030190 supérieur à
040211 et 040212
Ensuite, la hauteur des plantes a été
appréciée à travers une comparaison entre la hauteur
moyenne à mi-cycle et la hauteur moyenne des plantes au moment de la
première récolte, comme le montre le graphique suivant pour
voir la progression de la croissance:
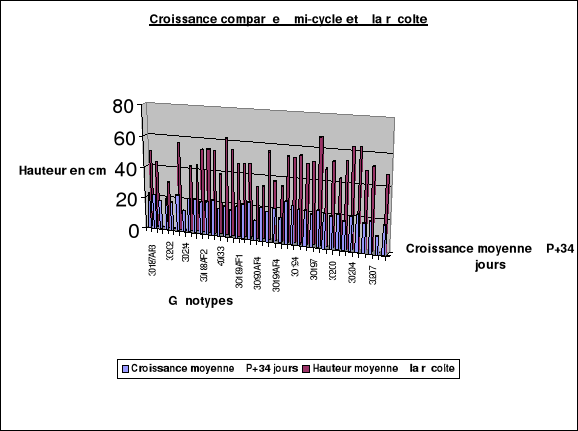
Figure 16 : Evolution de la croissance
comparée à mi-cycle et à la
récolte
Dans ce cas, les plus fortes progressions ont
été notées pour 030188,
030198, 030204, 030205. Par
contre 030202 et 040212 présentaient
les progressions les plus faibles entre le niveau de croissance à
mi-cycle et la hauteur des plantes à la récolte.
Le témoin 001237 présente une
légère régression alors que 040133
présente lui une faible progression.
III.1.1.1.3. La canopée ou
encombrement
On a pu l'apprécier en comparant pour chaque
génotype, le niveau moyen d'encombrement comme représenté
sur le graphique suivant. On voit que les niveaux moyens d'encombrement sont
les plus importants pour 030214, 030190,
01237, 030192 et 030189.
Dans l'ensemble, la plupart des génotypes ont des
niveaux d'encombrement relativement proches sauf pour 040213,
040212 et 030196 qui présentent les
niveaux d'encombrement les plus bas.
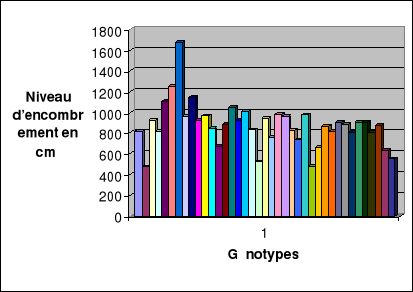
Figure 17 :Evolution du niveau
d'encombrement
III.1.1.1.4. Les dimensions
foliaires
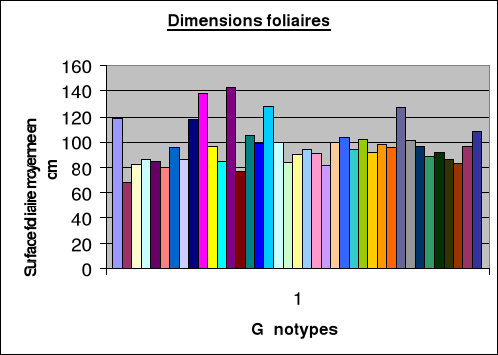
Figure 18 : Evolution des dimensions
foliaires
Pour l'essentiel des génotypes, la surface foliaire
moyenne est comprise entre 80 et 100 cm². Quatre génotypes
présentent des dimensions foliaires supérieures à 120
cm² ( le témoin 040133, 040204,
040207 et 030200) alors que 6
génotypes ont des dimensions foliaires comprises entre 100 et 120
cm². Un seul génotype a des dimensions foliaires inférieures
à 80 cm².
L'analyse factorielle discriminante que nous avons
effectuée à l'aide du logiciel SPSS sur les variables qui
précèdent nous a permis de voir que le nombre de jours à
partir duquel avait lieu la levée des plantes de nos différents
génotypes n'avait pas de lien direct avec la hauteur moyenne, les
dimensions foliaires ou encore le niveau d'encombrement. Ce, du fait qu'entre
ces variables, il existe une corrélation négative (cf.
Tableau des corrélations totales). Autrement dit, la levée des
plants n'influe pas sur la croissance générale et le
développement végétatif. Cependant, la croissance
était bien corrélée aux dimensions foliaires et au niveau
d'encombrement tout comme, les dimensions foliaires et le niveau d'encombrement
étaient corrélés entre eux.
La lenteur de la germination pourrait s'expliquer par
l'héritabilité de ce caractère propre à
C. chinense dont les semences sont
réputées lentes à germer
III.1.1.2. Caractères
qualitatifs
Pour ceux-ci, nous avons obtenu les résultats
suivants :
III.1.1.2.1. Le port
Nous avons pu simplement constater que la
quasi-totalité de nos génotypes présentaient des plantes
à port dressé ou érigé. Pour deux d'entre eux,on a
tout de même noté la présence d'une seule plante dans
chacun des cas, à port évasé :
030202 et 030213. On a bien la confirmation
que le port dressé, érigé domine le port
écrasé, évasé.
III.1.1.2.2. La vigueur
générale
Selon les classes établies, les niveaux de vigueur
sont représentés par le graphique suivant :
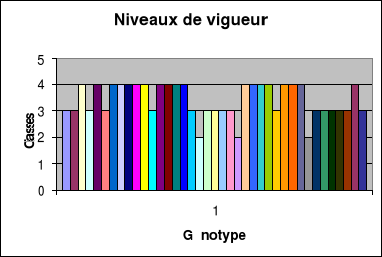
Figure 19 : Evolution du niveau de
vigueur
Trois classes de vigueur se distinguent nettement et on
remarque que dans l'ensemble,nos génotypes se sont bien comportés
vis à vis des conditions climatiques auxquelles ils ont
été confrontés. Toutefois, trois génotypes ont un
niveau de vigueur relativement bas car en dessous du niveau moyen
040208, 030211 et 030201.
Cela nous montre que le critère de vigueur a bel et
bien été déjà fixé ce niveau de
sélection correspondant à la première
génération de backcross. Mais les caractéristiques de la
lignée pure recherchée seront progressivement fixés suite
à plusieurs croisements avec cette lignée qu'on qualifiera de
`'récurrente''.
III.1.1.2.3. L'état
sanitaire
En utilisant certaines clés de détermination
phytopathologiques, nous sommes parvenus à diagnostiquer dans notre
matériel, quelques pathologies sur la base de l'inventaire de leurs
symptômes. Et les pathologies suivantes ont été
notés :
1- Cercosporiose due à Cercospora
capsici :
2- Moisissure des feuilles due à Cercospora
unamonoi ;
3- Corynesporiose due à Corynespora
cassiicola ;
4- Dépérissement des rameaux du à
Choanephora cucurbitarum ;
5- Dépérissement à sclérotes du
à Sclérotium rolfsii
Tableau 13 : Relevé
pathologique
|
Pathologie (soupçonnée)
|
Génotypes affectés
|
|
Cercosporiose
|
040203,040202,040206,030200,030201,030206,040211,030197,
|
|
Moisissure des feuilles
|
030189,030199,030206,040201,040207
|
|
Corynesporiose
|
030196,030203
|
|
Dépérissement des rameaux
|
040202,030199,030196,030200,030206
|
|
Dépérissement à
sclérotes
|
34 génotypes
|
Les dégâts les plus importants et les plus
signifiants ont été notés dans les cas de
dépérissement à sclérotes dans la mesure où
cette pathologie se manifeste sur les racines et le collet des plantes. Elle
entraîne de ce fait une pourriture sèche très importante
qui décime les plantes qui après jaunissement et
flétrissement du feuillage meurent.
Quelques cas de chute de boutons floraux ou de fleurs
caractéristiques de la coulure ont été observés
mais la fréquence des pieds affectés et la quantité des
organes perdus ne permettaient pas d'en faire des statistiques importantes.
III.1.2. En phase
générative
III.1.2.1. Caractères
quantitatifs
III.1.2.1.1. La précocité de
floraison
On a pu établir 7 classes correspondant au nombre de
jours à partir desquels, la floraison a commencé, pour chaque
génotype. Le diagramme suivant nous montre la répartition des
proportions de nos génotypes en fonction du nombre de jours.
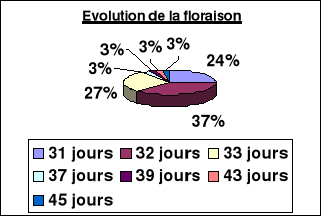
Figure 20 : Distribution de la
floraison
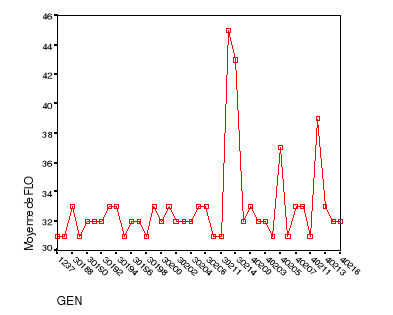
Figure 21 : Evolution de la
floraison
L'essentiel des génotypes ayant fleuri entre 31 et 33
jours.
Pour les principales familles, on note que :
030187 a fleuri au bout de 31 jours alors que
040201 et 040202 ont fleuri respectivement
au bout de 33 et 32 jours.
030188 a fleuri au bout de 33 jours contre 32
jours pour 040203, 31 jours pour 040204 et
37 jours pour 040205.
030189 a fleuri après 31 jours comme
040206, contre 33 jours pour 040207 et
040208
030190 a fleuri au bout de32 jours contre 31
jours pour 040211 et 39 jours pour 040212
030191 a fleuri au bout de32 jours comme
040215 alors que 040213 a fleuri au bout de
33 jours.
Comparativement C. chinense pousse plus lentement
que C. annuum. A ce niveau on ne peut pas savoir dans quel sens
évolue la fixation de ce caractère pour expliquer la lenteur ou
la relative vitesse de floraison. Cependant, en considérant la moyenne
de jours requis pour la floraison de Rajput & Parulekar (1998) de 40 jours
chez C. annuum, nous pouvons dire que les 88 % de génotypes de
notre matériel ayant fleuri au bout de 33 jours (moyenne des
extrêmes) présentent une précocité du point de vue
de la floraison.
III.1.2.1.2. Evolution de la
fructification
L'évolution de la fructification a permis de
constaté qu'entre 2 et 3 jours après le début de la
floraison, c'est à dire entre 33 et 35 jours après plantation
plus de la moitié de nos génotypes (62 %) avait commencé
à fructifier, comme le montre le diagramme suivant :
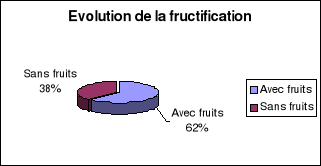
Figure 22 : Distribution de la
fructification
Les premières plantes à avoir fleuri ont
été évidemment les premières à avoir
fructifié et pour les génotypes n'ayant pas encore
fructifié, elle s'est ensuite progressivement déroulée
sauf pour les génotypes que nous n'avons pas récoltés
comme 030192, 030193, 030213
et 030214.
Une comparaison du taux de fructification et du taux de
floraison confirme bien la corrélation négative qui existe entre
les variables Fructification et floraison.
Par la suite, le nombre de fruit moyen par plante par
génotype a été représenté sur le graphique
suivant :
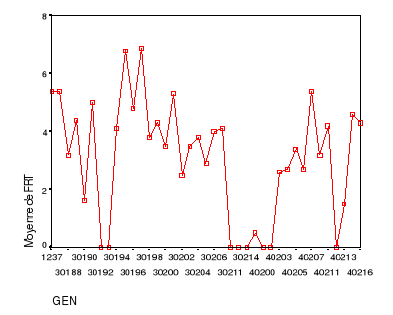
Figure 23 : Evolution de la
fructification
Le graphique de la figure 23 ci-dessus nous montre que 8
génotypes ne présentent aucun fruit sur l'ensemble des plantes
(040201, 030193, 030192,
030214, 040212, 030211,
030213, 040202). Deux génotypes se distinguent
avec des productions moyennes par plante supérieures à 6 fruits
(030195 et 030197) pendant que trois autres
se distinguent avec des productions moyennes par plantes inférieures
à 2 fruits (040213, 030190 et
040133)
III.1.2.1.3. Le rendement et ses
composantes
III.1.2.1.3.1. Le rendement génotypique
moyen relatif
Dans un premier temps, nous avons établi, en
considérant les deux premières récoltes
effectuées, le rendement génotypique moyen relatif
représenté par les graphiques ci-dessous :
-la première récolte
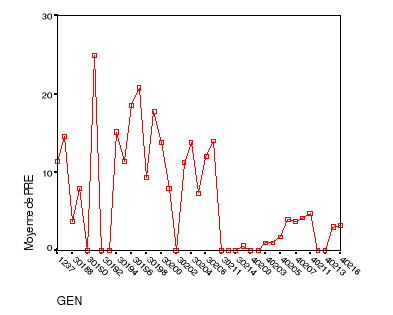
Figure 24 : Evolution de la première
récolte
-la seconde récolte
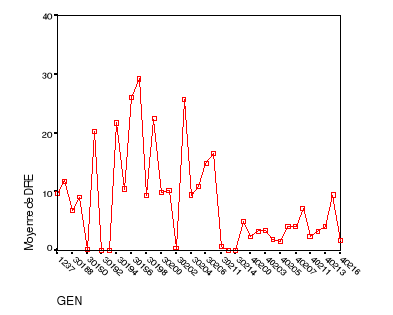
Figure 25 : Evolution de la seconde
récolte
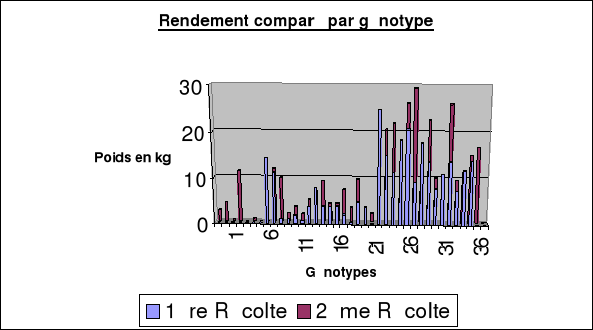
Figure 26 : Comparaison des deux
récoltes
On note qu'à partir du génotype 24
(030191), les rendements sont plus importants et plus souvent
supérieurs à 10 kg avec des pics à 25 kg lors de la
première récolte pour 030191 et 20.8 kg pour
030197. Lors de la deuxième récolte, on a 29.8
kg pour 030197, 26.12 kg pour 030196 ou
encore 25.84 kg pour 030203.
Nos principales familles présentent des rendements
élevés par rapport aux descendants issus de backcross
(matériel homozygote) sauf pour 030190.Dans l'ensemble,
nous notons une nette différence entre les rendements des individus et
ceux de leurs descendants issus du premier backcross.
III.1.2.1.3.2. Le rendement génotypique moyen
par plante
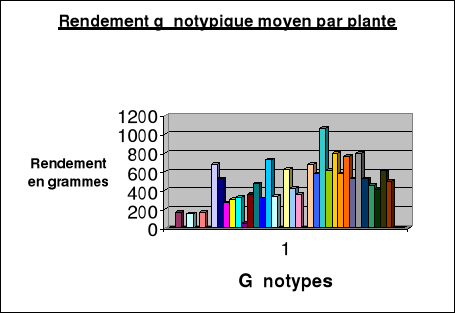
Figure 27 : Evolution du rendement moyen
par plante
Le génotype 030195 on le voit
présente de loin le rendement moyen par plante, le plus
élevé (1059,644 g) alors que les témoins ont
respectivement 526,284 g et 47,39 g pour 001237 et
040133.
III.1.2.1.3.3. Le poids moyen des
fruits
Pour chacun des nos différents génotypes, le
poids moyen d'un fruit a été estimé et le graphique de la
figure 28 qui suit, nous en donne un aperçu.
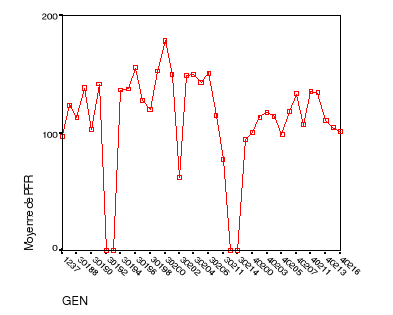
Figure 28 :Evolution du poids moyen des
fruits
Nous pouvons classer l'ensemble de nos génotypes en
deux groupes, en ce qui concerne le poids moyen des fruits dans la mesure
où le graphique ci-dessus nous montre deux niveaux ; un premier
où le poids moyen des fruits est inférieur à 100grammes et
un second où le poids moyen des fruits est supérieur à 100
grammes. Nos témoins 001237 et 040133,
appartiennent à ce groupe avec respectivement 97.46 et 94,78 grammes.
030200 a les fruits dont le poids moyen est
le plus élevé alors que 030202 a les fruits
dont le poids moyen est le plus faible.
III.1.2.1.3.4. Le calibre moyen des
fruits
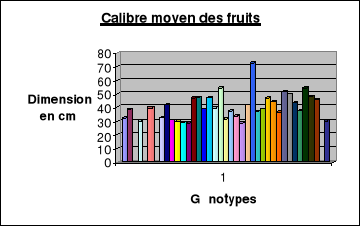
Figure 29 : Evolution du calibre moyen des
fruits
Les fruits issus de différents génotypes de
notre étude sont de calibre relativement moyen, cependant quelques
numéros présentent des fruits d'assez gros calibre comme
030194 (72.8 cm²) qui a possèdent les fruits de
plus gros calibre, 040212 (54.06 cm²) ou encore
030205 (54.02 cm²) et 030200 (51.8
cm²). Les témoins 001237 et
040133 ont respectivement comme calibre moyen des
fruits ; 42.16 et 28.6 cm².
III.1.2.1.3.5. Le rendement grainier par
génotype
Pour évaluer le rendement de nos différents
génotypes en graines, le graphique qui va suivre nous donne la
répartition dans chacun du nombre de graines par fruit pour chacun des
génotypes.
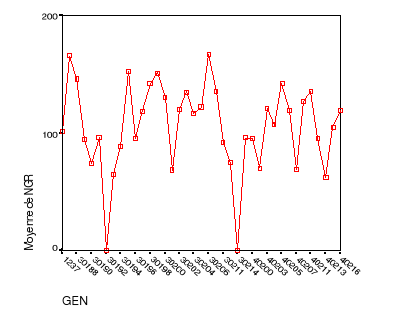
Figure 30 : Evolution du nombre moyen de
graines
Le nombre de graines par fruit est très variable mais
dans l'ensemble, il est voisin de 100 graines. Dans trois cas, il excède
150 (030195, 030206,
030199). Les témoins présentent des fruits
à 101,8 graines pour 001237 contre 96 graines pour
040133.
Suivants les résultats de notre analyse factorielle
discriminante, la première récolte est :
très fortement corrélée de façon
positive à la seconde récolte,
bien corrélée positivement au poids moyen des
fruits et au calibre moyen des fruits,
faiblement corrélée positivement au nombre
moyen de graines par fruit
La seconde récolte est bien corrélée
positivement au poids moyen des fruits et au calibre moyen des fruits et
nettement moins au nombre moyen de graines.
Le poids moyen des fruits est très fortement
corrélé de façon positive au calibre moyen des fruits et
nettement moins au nombre moyen de graines par fruit.
De façon générale, le tableau de
corrélations totales nous montre que toutes les composantes du rendement
sont liées entre elles.
Tableau 14 : Les corrélations totales
des variables
ELV CRS DIF CAN FLO FRT PRE DRE
PFR CAM NGR
ELV 1.000
CRS -0.457 1.000
DIF -0.037 0.040 1.000
CAN -0.299 0.469 0.127 1.000
FLO 0.736 -0.376 -0.048 -0.299 1.000
FRT -0.299 0.391 0.014 0.465 -0.313 1.000
PRE -0.192 0.392 -0.014 0.177 -0.306 0.522 1.000
DRE -0.238 0.358 -0.023 0.120 -0.270 0.456 0.890 1.000
PFR -0.605 0.471 0.044 0.276 -0.437 0.394 0.520 0.515
1.000
CAM -0.552 0.614 0.028 0.404 -0.352 0.464 0.537 0.565
0.723 1.000
NGR -0.363 0.290 -0.065 0.190 -0.265 0.248 0.297 0.266
0.389 0.364 1.000
Signification des variables :
ELV: évolution de la levée en nombre de jours
après semis
CRS: croissance en nombre de jours après semis
DIF: dimensions foliaires
CAN: canopée ou niveau d'encombrement
FLO: floraison en nombre de jours après semis
FRT: fructification (nombre de fruits par plante)
PRE: première récolte
DRE: deuxième récolte
PFR: poids moyen des fruits
CAM: calibre moyen des fruits
NGR: nombre moyen de graines par fruit
III.1.2.2. Caractères
qualitatifs
III.1.2.2.1. La couleur des fruits
III.1.2.2.1.1. Avant maturité
Les différents degrés de vert sont
répartis en classe suivant que la couleur soit très
foncée, claire, brillante,etc. Et trois classes ont été
établies ; la classe III avec une coloration vert `'normal'', la
classe IV avec une coloration vert foncé et la classe V avec une
coloration vert très foncé et brillant.
Et le diagramme nous montre que plus de la moitié des
génotypes de notre étude ont des fruits à
immaturité dont la couleur appartient à la classe IV
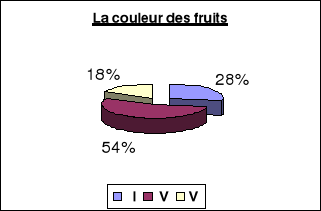
Figure 31 : Distribution de la couleur des
fruits avant maturité
III.1.2.2.1.2. A maturité
A maturité, les fruits ont été
classés selon leur couleur et le digramme suivant donne les proportions
des différentes couleurs obtenues dans nos génotypes.
85 % des fruits sont rouges à maturité, 10 %
sont jaunes et 5 % présentent des fruits rouges et jaunes
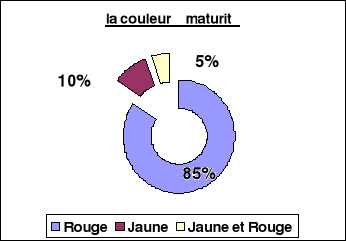
Figure 32 : Distribution de la couleur des
fruits à maturité
III.1.2.2.2. L'état sanitaire
Pendant la phase reproductive, les plantes ont
été principalement attaquées par la mouche
méditerranéenne des fruits ; Ceratis capitata dont
les piqûres sur les fruits ont ouvert la voie à plusieurs
pathologies identifiées à l'aide de clés de
détermination, parmi lesquelles on peut citer:
les taches noires duveteuses sur les fruits dues à
Curvularia sp. (1)
le mildiou des fruits du à Phytophthora capsici
(2)
l'anthracnose due à Colletotrichum nigrum
(3)
Du fait des dégâts très importants de la
mouche méditerranéenne, l'estimation des dégâts des
maladies citées plus haut s`est quelque peu biaisée.
Toutefois, pour l'ensemble on a noté (1) :
très peu ; (2) : peu et (3) : importante.
III.2. Caractérisation globale du
matériel végétal
D'après notre analyse factorielle discriminante, il
serait prématuré de tirer des conclusions générales
sur la base de ce seul essai, toutefois, les résultats obtenus nous ont
permis de mettre en évidence quelques faits intéressants. Nous
avons obtenu une classification en 8 groupes basée principalement sur
la forme des fruits d'où nous parvenons nettement à distinguer
les `'carrés'', les `'rectangulaires'' et les `'triangulaires'' dans les
proportions suivantes :
|
Groupes
|
Type variétal
|
Effectif en %
|
|
1 , 2 , 3 , 4
|
Carré
|
67.69
|
|
5, 6
|
Rectangulaire
|
31.28
|
|
7, 8
|
Triangulaire
|
1.03
|
L'analyse nous donne un taux de 67.2 % d'individus bien
classés par rapport aux différents groupes comme pour confirmer
la tendance `'carrée'' de fruits de la plupart des génotypes
constituant notre matériel végétal.
L'analyse pourrait encore être affinée pour nos
différents groupes dans la mesure où les différences
provenant des types variétaux ne sont pas importantes.
En effet dans le type `'carré'' regroupant les groupes
1, 2, 3 et 4, on peut évoquer l'existence de carrés
américains (`'blocky''), hollandais et même italiens (§
I.2.1.2).
Dans le type `'rectangulaire'', il faut évoquer les
types demi-longs ou trois-quarts longs. Le type `'demi-long'' est
également un type très apprécié constituant par
conséquent avec les `'carrés'' les types variétaux les
plus intéressants, car étant le type désirable.
Cependant, tous nos groupes sont assez proches les uns des
autres dans la mesure où généalogiquement,tous les
individus sont proches ; selon les cas, juste un des parents mâles
ou femelle diffère.
De plus, notre matériel étant en
ségrégation, nous avons des fruits à 3 ou 4 loges.
Les principales familles :
030187 : les fruits qui
présentent des bonnes caractéristiques horticoles sont de type
`'carré'' appartenant aux quatre groupes alors que pour ce qui est
de :
040201 et 040202, on
retrouve respectivement une forte proportion de carrés du groupe 1
(60%) et une forte proportion de carrés du groupe 2 (60%).
030188 : les fruits sont de type
carré des groupes 1 et 2 essentiellement alors que pour 040203
et 040204 on retrouve des carrés de tous les
groupes (1, 2, 3 et 4) et pour 040205 on a une forte
proportion de carrés du groupe 1 (60%).
030189 : les fruits sont à 50%,
des carrés du groupe 4. On retrouve les mêmes proportions pour
040206 et 040207 alors que pour
040208 on a une forte proportion de carrés du groupe 1
(60%).
030190 : on retrouve une proportion
importante de rectangulaires du groupe 6 (70%) alors que pour
040211 on a 70% de carrés des groupes 1, 2, 3 et 4 et
pour 040212, on a 70% de rectangulaires du groupe 5.
030191 : on a 70% de carrés
alors que pour 040213 on retrouve 80% de rectangulaires, 70%
de carrés pour 040215 et 70% de rectangulaires du
groupe 5 pour 040216.
Nous pouvons donc voir que pour les principales familles, il
existe encore une hétérogénéité liée
à la disjonction des caractères que nous évoquions
déjà plus haut. De plus, les performances relativement faibles
dans certains cas, pourraient être attribuées soit à un
défaut de sélection soit à l'effet des gènes
récessifs
Tableau 15 : Comparatif des principales
familles
|
Génotype
|
CRS (cm)
|
FLO (jours)
|
FRT
|
RDT(kg)
|
NGR
|
|
030187
|
22.25
|
31
|
5.4
|
26.46
|
166
|
|
040201
|
22.9
|
33
|
0
|
2.38
|
95.3
|
|
040202
|
19.1
|
32
|
0
|
3.4
|
70.6
|
|
030188
|
19.5
|
33
|
3.2
|
10.58
|
149.5
|
|
040203
|
21
|
32
|
2.6
|
4.44
|
121.1
|
|
040204
|
20.6
|
31
|
2.4
|
2.98
|
107.4
|
|
040205
|
13.1
|
37
|
3.4
|
3.38
|
96
|
|
030189
|
17.7
|
31
|
4.4
|
17.16
|
98.7
|
|
040206
|
20.11
|
31
|
2.7
|
8.06
|
119.7
|
|
040207
|
22.5
|
33
|
5.4
|
7.98
|
69
|
|
040208
|
23.8
|
33
|
3.2
|
11.38
|
127.4
|
|
030190
|
23.33
|
32
|
1.6
|
0.6
|
74.1
|
|
040211
|
18.9
|
31
|
4.2
|
7.3
|
135.6
|
|
040212
|
12.5
|
39
|
0
|
3.22
|
95.5
|
|
030191
|
26.6
|
32
|
5
|
45.32
|
96.8
|
|
040213
|
22.2
|
33
|
1.5
|
4.18
|
62.5
|
|
040215
|
21.1
|
32
|
4.6
|
12.5
|
105.3
|
|
040216
|
21.5
|
32
|
4.6
|
4.98
|
119.3
|
Le tableau qui précède nous permet de dire que
les caractères ici répertoriés ; le niveau de
croissance, la précocité de floraison, le nombre de fruits par
plante, le rendement en poids de fruits et le rendement grainier sont assez
significatifs pour porter un choix sur le 030187 et le
030191 dans la mesure où ces numéros
présentent un bon niveau pour l'ensemble des caractères choisis
même si pour le rendement en poids de fruits, on note une grande
différence (26.46 kg contre 45.32 kg) qui pourrait s'expliquer par le
fait que dans le cas de 030191, les fruits comprennent aussi
bien des formes rectangulaires que carrées. Alors que pour le
030187,on ne retrouve que des formes carrées.
Mais en comparaison, la descendance des backcross est moins
intéressante (car inférieure) du point de vue des
caractères choisis sauf dans le cas de 030190 où
les descendants sont supérieurs.
Par ailleurs, il faut noter que les descendants homozygotes de
backcross présentent un rendement grainier satisfaisant même si de
façon générale leur production fruitière reste
faible. Sans doute du fait de la disjonction des caractères.
D'un point de vue global, nous pouvons retenir que dans
l'ensemble nos génotypes présentent des valeurs de
caractères acceptables car le plus souvent voisines ou
supérieures à celles de nos témoins 01237
et 040133. Certains génotypes se sont très bien
comportés pour un ou deux caractères parfois avec des valeurs
extrêmes. D'autres avec des valeurs de caractères de
caractères moyennes ou voisines de celles des témoins
méritent un plus grande attention en ce sens qu'ils représentent
pour les critères choisis,les meilleurs génotypes.
Dans ce groupe, en fonction de l'ensemble des
caractères étudiés, on peut citer une dizaine de
génotypes :
030187 qui présente une bonne
adaptation comme en témoignent sa bonne vigueur (classe 4), sa bonne
nouaison (5,4 fruits), sont rendement génotypique moyen par plante
(670,734 g), ses fruits carrés et son rendement moyen en graines.
030189
030191
030194
030195
030196
030200
030197
030201
030207
Sur le plan de la généalogie, il faut souligner
que les génotypes choisis ci-dessus ont pour parent femelle
010239 (030187, 030189 et
030191) ou les poivrons tropicaux n°
2 ; 010240 (030194,
030195, 030196 et 030197),
n° 3 ; 010241 (030200 et
030201) et n° 4 ; 010242
(030207).
X.1. Conclusions et
perspectives
Proposer des voies et moyens stratégiques pour la
sélection de piments (Capsicum annum L.) adaptés
à la chaleur humide, tel était l'objectif majeur de notre
étude qui se devait en outre de « déceler »
parmi les génotypes de notre matériel d'étude, ceux qui
répondent au critère de conformation de fruits.
En effet, si l'objectif était de
sélectionner, en conditions hivernales, les meilleurs génotypes
adaptés, la saveur douce et la forme des fruits ; des gros
carrés de type `'Yolo'' et des demi-longs de type `'Lamuyo'' devaient
s'adjoindre à cette adaptation.
Dans notre essai, il ressort que pour la plus grande partie
des génotypes constituant notre matériel végétal,
un comportement positif généralisé a été
noté pour ce qui est de l'adaptation à des conditions de
températures et d'humidité défavorables à la
culture de poivrons.
Un taux important (49 %) de génotypes
présente un niveau de vigueur, indice indéniable et fondamental
d'une bonne adaptation, semblable à celui des témoins.
Quelques-uns (44 %), quoique bien adaptés également
présentent cependant un niveau de vigueur moyen. Ce qui démontre
aussi que les résistances aux divers bio-agresseurs liés à
cet environnement ont été bien héritées de la part
des parents initiaux à travers les cycles de sélection
précédents.
Cette tendance se confirme avec l'appréciation d'autres
critères tels que le rythme de croissance, la canopée ou niveau
d'encombrement des plantes et même les dimensions foliaires. Donc, du
point de vue du développement végétatif,
Un autre aspect relatif au caractère aléatoire
de la production du fait d'une pression parasitaire et virale très
importante est également pris en compte dans ce cycle de
sélection.
On note des taux moyens de productivité en fruits de
l'ordre de 7 à 8 fruits qui sont fort appréciables lorsqu'on sait
qu'en moyenne, une plante produit une bonne quinzaine de fruits.
Ce sont donc là, des caractères qui permettent
de caractériser une bonne adaptation certes mais également des
voies qu'il convient de poursuivre et d'améliorer pour affiner la
sélection adaptative des piments à l'hivernage.
Ainsi, la combinaison du plus grand nombre de
critères nous a permis d'élire dans l'ensemble de notre
matériel, des génotypes qui en plus de répondre
positivement à l'adaptation aux conditions climatiques ont pu produire
des fruits de qualité recherchée à savoir des formes
carrées de type `'Yolo'' et des formes demi-longues de type `'Lamuyo''.
On peur citer 030187, 030189, 030191, 030194, 030195, 030196, 030197,
030200, 030201, 030207.
Comme on le voit, cette étude, avec son double
objectif s'est avérée concluante dans l'ensemble. Toutefois, il
convient de relativiser les résultats obtenus dans la mesure où
le travail de sélection est un travail de longue haleine dont
l'accumulation des confirmations de résultats positifs seule, est une
gageure de succès. Un questionnement demeure cependant à propos
de la `'chute des performances'' des homozygotes dans la mesure où le
lien entre l'effet dépressif des performances et la parfaite homozygotie
reste à établir.
C'est pourquoi dans la poursuite de ce travail
sélectif, nous préconisons de :
â Poursuivre le travail entrepris en décalant le
cycle de la plante pour qu'il soit en plus grande partie calé entre les
mois de juillet et d'août pour que les plantes soient plus
confrontées à la rudesse de l'hivernage. Cela permettra d'avoir
des résultats encore plus significatifs.
â Essayer de réaliser des rétrocroisements
sur le matériel d'origine généalogique poivrons tropicaux
n° 2, n° 3 et n° 4 pour avoir une idée de leurs
comportements respectifs à comparer avec les ancêtres.
â Introduire dans les prochains cycles de
sélection un témoin de type Chinense à observer
pour mieux caractériser l'adaptabilité du matériel au
moyen d'une comparaison directe.
â De tester encore les produits homozygotes issus de
rétrocroisements des principales familles pour permettre de trouver une
réponse à la baisse des performances de ces derniers par rapport
aux parents issus de la sélection généalogique
classique.
AGROMISA, [nd]. Capsicum growing. AGRODOK
n° 13, Agromisa, Wageningen, Pays-Bas, 22 p
ANONYME, 2004.
www.aprifel.com:
Fiches nutritionnelles pour produits ; le poivron.
ANU A. & PETER K. V., 2000. The chemistry of
paprika. Capsicum & Eggplant Newsletter n° 19, juin 2000.P Belletti,
Ed. University of Turin (DIVAPRA-Agricultural Genetics) ,Italy,
pp 19-22
ASHWORTH S., 1991. Seed to seed: seed saving
techniques for the vegetable gardener. Seed Saver Publications, Decorah, Iowa /
USA, 222 p
BASSIM H., BASSIM E., JONES J. B., MINSAVAGE G. V.,
DICKSTEIN E. R., 2004. Bacterial spot of tomato and pepper caused by
Xanthomonas axonopodis p.v vesicatoria in the Western
Mediterranean Region of Turkey. Plant Disease, Vol. 88, Number 1, January 2004,
p 85.
BELLETI P., QUAGLIOTTI L., 1988. Problems of
seed production and storage of pepper. In: AVRDC, 1989. Tomato and Pepper
production in the Tropics; Proceedings of the International Symposium on
Integrated Management Practices. Tainan, Taiwan. 21-26 march 1988, pp 28-41
BENIEST J. , 1987. Guide pratique du
maraîchage au Sénégal, CDH. Dakar, 144 p
CDH, 1986. Les cultures
maraîchères au Sénégal : bilan des
activités de 1972 à 1985. ISRA-CDH, Dakar /
Sénégal, 265 p
CHAINE-DOGIMONT C., 1993. Etude
génétique de trois systèmes de résistance par
hypersensibilité ou séquestration aux trois virus principaux
infectant le piment (Capsicum annuum L.). Thèse de
docteur, INA-PG. Paris, 194 p
CHENG S.S., 1988. The use of Capsicum
chinense as sweet pepper cultivars and sources for gene transfer. In:
AVRDC, 1989. Tomato and Pepper production in the Tropics; Proceedings of the
International Symposium on Integrated Management Practices. Tainan, Taiwan.
21-26 march 1988, pp 55-62
CHAUX C. & FOURY C., 1994. Productions
Légumières. Tome 3 : Légumineuses potagères -
Légumes fruits. Coll. « AGRICULTURE D'AUJOURD'HUI :
Sciences, Techniques, Applications ».Tec & Doc. Lavoisier, Paris,
France. 563 p
DECLERT C., 1990. Manuel de phytopathologie
maraîchère tropicale : cultures de Côte d'Ivoire. Coll.
« Didactiques ». Editions de l'ORSTOM, Paris, 333 p
DEMARLY Y. & SIBI M., 1989.
Améliorations des plantes et biotechnologies. Universités
Francophones « Sciences en marche ».Editions John Libbey
Eurotext, Paris, France. 152 p
DESAI B.B., KOTECHA P.M., SALUNKHE D.K.,
1997. Seeds handbook : biology, production, processing and
storage. Marcel Dekker, Inc., New York, USA. 627 p
DE WITT D., BOSLAND P. W., 1993.The pepper
garden. Ten Speed Press. Berkeley, California, USA. 240 p
DOSHI K. M. & SHUKLA P. T., 2000. Expression
of heterosis in chilli (Capsicum annuum L.). In: Capsicum &
Eggplant Newsletter n° 19, juin 2000. P. Belletti, Ed. University of
Turin (DIVAPRA-Agricultural Genetics) ,Italy, pp 66-69
DUPRIEZ H., LEENER Ph. de, 1987. Jardins et
vergers d'Afrique. Terres et Vie / CTA. Editions L'harmattan. Nivelles,
Belgique, 354 p
EL-AIDY F., EL-AFRY M., IBRAHIEM F., 1988.
The influence of shade nets on the growth and yield of sweet pepper. In: AVRDC,
1989. Tomato and Pepper production in the Tropics; Proceedings of the
International Symposium on Integrated Management Practices. Tainan, Taiwan.
21-26 march 1988, pp 345 - 348
FALL A. S., FALL S. T., CISSE I., BADIANE A. N., DIAO
M. B., FALL C. A., 2001. Caractéristiques de la zone des
Niayes. Dans: FALL S. T. & FALL A. S., 2001.Cités horticoles en
sursis ? L'agriculture dans les grandes Niayes au Sénégal.
Editions CRDI, Ottawa (Ontario), Canada, 120 p.
FAO, 2003. Annuaire de la production 2002,
vol. n° 56. Collection FAO Statistiques n° 176. Rome, 261 p
GEBRE - SELASSIE K., GOGNALONS P., MARCHOUX G.,
1994. ; Poivron : les viroses sous abris et plein champ.
FRUITS & LEGUMES, n°120 - juin 94, pp 52 - 55
GUIGNARD J-L., 1996. Botanique.
10e édition révisée. Collection
« ABREGES ». Masson, Paris, 278 p
GREENLEAF W. C., 1986. Pepper Breeding. In:
Mark J-Basset, Ed., 1986. Breeding Vegetables Crops, Part I. AVI Publishing. Pp
67-134
GRUBEN G. J. H & EL TAHIR I. M., 2004.
Capsicum annuum L. In: GRUBEN G. J. H & DENTON O. A. (Editors).
Plant Resources Of Tropical Africa 2. Vegetables. PROTA Foundation, Wageningen,
Netherlands / Backhuys Publishers, Leiden, Netherlands / CTA, Wageningen,
Netherlands. Pp 154 - 163.
HARLAN J.R., 1987. Les plantes
cultivées et l'homme. ACCT/ CILF, collection « Techniques
Vivantes ». Presses Universitaires de France, 414 p
HENNART J.W., 1995. Qualité et
sélection du piment. PHM - Revue Horticole, décembre 1995-janvier
1996, n°365-366, pp 52 - 55
KALLOO, 1988. Vegetable breeding, Vol. I. CRC
Press, Inc. Boca Raton / Florida, USA, 239 p
KOUNOWSKY J.S., TODOROVA J.J., STOIMENOVA E.S.,
1985. Capsicum chinense source of resistance to Leveillula
solanacearum F. Sp capsici Gol. And Tobacco Mosaic Virus. CAPSICUM NEWSLETTER
n°4. pp....
KROLL R. , 1994. Les cultures
maraîchères. Coll. « Le technicien d'agriculture
tropicale ». Maisonneuve et Larose. ACCT / CTA, Paris, France, 219
p
LAUMONNIER R. , 1979. Les cultures
légumières et maraîchères, tome III. 3e
édition. Collection « Encyclopédie Agricole »
Editions J-B. Baillière, Paris, France, 276 p
MERCIER J.F., 1995. La culture du piment. PHM
- Revue Horticole, décembre 1995-janvier 1996, n°365-366, pp 44 -
51
MESSIAEN C.M. , 1975.
Le potager tropical, tome 2 : cultures spéciales. Collection
« Techniques vivantes ». Presses Universitaires de France,
197 p
MESSIAEN C.M., BLANCARD D., ROUXEL. F., LAFON
R., 1991.Les maladies des plantes
maraîchères. Ed. INRA, Paris, France, 552 p
MITRA S. K., 1990. Chilli. In : MITRA S.
K., SADHU M. K., BOSE T. K., 1990. Nutrition of vegetable crops. Ed. Naya
Prokash. Calcutta, India, pp 101 - 105
MORENO-GONZALEZ J. & CUBERO J. I., 1993.
Selection strategies and choice of breeding methods. In: HAYWARD M. D.,
BOSEMARK N. O., ROMAGOSA I., 1993. Plant Breeding: principles and prospects.
Plant breeding series 1. Chapman & Hall, London, pp 281-313
PALLOIX A., PHALY T., 1995. Histoire du
piment : de la plante sauvage aux variétés modernes. PHM -
Revue Horticole, décembre 1995-janvier 1996, n°365-366, pp 41-
43
PALLOIX A., DAUBEZE A.M., NEMOUCHI G., RUFFINATO A.,
1995. Maladies et ravageurs, les solutions apportées par la
lutte génétique. PHM - Revue Horticole, décembre
1995-janvier 1996, n°365-366, pp 56 - 61
PALLOIX A., LEFEBVRE V., CARANTA C., DAUBEZE A.M.,
MOURY B., PFLIEGER S., AHMED E.A., THALUIS A., RUFFEL S. & CHENG C.M.,
2001. Recherches sur le piment. Rapport d'activités 1997-2000
de l'Unité de Génétique et d'Amélioration des
Fruits et Légumes. INRA Avignon - Montfavet. France, pp
PETER K.V., 1998. Genetics and Breeding of
vegetables. Indian Council of Agricultural Research, New Delhi. India, 333 p
POCHARD E., PALLOIX A., DAUBEZE A.M.,
1992. Le piment. Dans : GALLAIS A. et
BANNEROT H., éditeurs., 1992. Amélioration des espèces
végétales cultivées : objectifs et critères de
sélection. Ed. INRA, Paris, pp 420 - 434
POULOS J. M., 1993. Pepper breeding.
In : CHADHA M. L., AMZAD HOSSAIN A. K. M., MONOWAR HOSSAIN S. M., 1993.
Breeding of Solanaceous and cole crops : A compilation of lecture
materials of training course held in Bari, Joydebpur, Gazipur, Bangladesh 29
september-3 october 1991. AVRDC, Taipei. Pp 85-121
PURSEGLOVE J.W., 1984.
Tropical Crops : Dycotiledons. Ed. Longman Group Ltd, Singapore, 719 p
RAJPUT J. C., PARULEKAR Y. R., 1998.
Capsicum. In: SALUNKHE D. K., KADDAM S. S., 1998. Handbook of vegetable science
and technology; production, composition, storage and processing. Marcel Dekker,
Inc. New York , USA, pp 203 - 224
SACCARDO F. & K. SREE RAMULU., 1977.
Cytological investigations in the genus Capsicum. « CAPSICUM 77 »
Comptes rendus du 3e congrès EUCARPIA sur la génétique et
la sélection du piment, 5-8 juillet 1977. Station d'Amélioration
des Plantes maraîchères, INRA, Avignon - Montfavet, France, pp
51-66
TROPICASEM, 2001. En savoir plus sur le
piment : gestion de l'eau et irrigation en culture intensive. Tropiculture
n° 54, mars 2001. Edition Tropicasem, Dakar / Sénégal, pp
4-5
TROPICASEM, 2004. Mieux réussir la
fertigation du piment. Tropiculture n° 92, mai 2004. Edition Tropicasem,
Dakar / Sénégal, pp 1-2
WIEN H. C., TRIPP K. E., HERNANDEZ-ARMENTA R. &
TURNER A. D., 1988. Abscission of reproductive structures in pepper:
causes, mechanisms and control. In: AVRDC, 1989. Tomato and Pepper production
in the Tropics; Proceedings of the International Symposium on Integrated
Management Practices. Tainan, Taiwan. 21-26 march 1988, pp 150-165
WILLIAMS C.N., UZO J.O., PEREGRINE W.T.H.,
1991. Vegetable Production in the Tropics. «Intermediate
Tropical Agriculture series». Ed. Longman Scientifical & Technical.
Malaysia, 179 p
   
| 

