Université Montpellier II
Ecole Doctorale
Systèmes Intégrés en Biologie, Agronomie,
Géosciences,
Hydrosciences et Environnement (Sibaghe).
NOTICE DE TRAVAUX
Présentée pour
l'obtention du diplôme
HABILITATION A DIRIGER DES RECHERCHES
Par
Philippe Lachenaud
Chercheur au
Cirad-Bios
« L'amélioration génétique du
cacaoyer. Des ressources génétiques forestières aux
variétés cultivées »
Soutenue le 3 mars 2010 Membres du jury :
M. CHARRIER André, SupAgro Montpellier et U. Montpellier
2, Président du jury. M. DAVID Jacques, UMR DIAPC, SupAgro
Montpellier
M. LEFEVRE François, Inra-URFM, Avignon
Mme NEEMA Claire, Dpt SVS, AgroParisTech
M. PRAT Daniel, UMR5023, Université Lyon 1
M. SACHE Yvan, UMR Bioger-CPP, Inra AgroParisTech --
Sommaire
I. Curriculum vitae 3
II. Activités d'encadrement et de formation 5
III. Publications et travaux 7
3.1 Publications dans périodiques 7
3.2 Communications orales lors de rencontres internationales
12
3.3 Posters 16
3.4 Divers 17
3.5 Rapports de mission 18
IV. Synthèse des résultats scientifiques 21
4.1 Amélioration génétique et
sélection en Côte d'Ivoire 22
4.1.1 Les variétés hybrides des
générations 1975 et 1979 22
4.1.2 Le programme de sélection récurrente
réciproque 23
4.1.3 L'utilisation des haploïdes doublés 24
4.2 Les cacaoyers spontanés de Guyane 28
4.2.1 Prospections et collecte 28
4.2.2 Sauvegarde 29
4.2.3 Caractérisation et évaluation 29
4.2.4 Diversité génétique et structuration
30
4.2.5 Utilisation dans les programmes d'amélioration
31
4.3 Conclusions 34
4.4 Références citées 34
V. Projet de recherches 38
5.1 Objectifs 38
5.2 Le projet et les résultats attendus 38
5.2.1 La problématique scientifique-Questions de recherche
38
5.2.2 L'état de l'art 39
5.2.3 Le contexte international 40
5.2.4 Les travaux envisagés 40
5.2.5 Les protocoles expérimentaux 41
A) Evaluation de la résistance aux Phytophthora
41
B) Evaluation de la résistance à
Moniliophthora perniciosa 42
C) Evaluation de la résistance à Ceratocystis
fimbriata 43
D) Caractérisation de la dureté des cortex 44
E) Tests à Montpellier (Cirad) 44
F) Prélèvement des endophytes 44
5.2.6 Le calendrier prévisionnel 47
5.3 Perspectives 47
5.4 Références citées 48
I. Curriculum vitae : Philippe LACHENAUD
Nationalité :
Date de naissance :
Française
02 septembre 1952
Adresse professionnelle :
Profession :
UPR Maîtrise des bioagresseurs des cultures pérennes
TA-A31/02
34398 Montpellier Cedex 5, France
Téléphone : +33 4 67 61 56 33
Fax : +33 4 67 61 55 81
Adresse électronique :
philippe.lachenaud@cirad.fr
Chercheur
Fonction :
Généticien cacao
Centre de coopération internationale en recherche
agronomique pour le développement
Département BIOS
Compétences :
Cacao, ressources génétiques,
sélection, phytotechnie
Ressources génétiques cacaoyères (clones,
matériel spontané, descripteurs). Sélection
cacaoyère (modalités, protocoles, dispositifs).
Phytotechnie et physiologie du cacaoyer.
Thèmes de recherche en cours :
- description et caractérisation du matériel
spontané de Guyane,
- étude de la variabilité génétique
du matériel spontané de Guyane. - élaboration du
rendement.
Autres expertises : Régions
d'expérience : Formation :
Facteurs de la Fructification et Remplissage des cabosses.
Déficience en bore sur cacaoyer.
Brésil, Côte d'Ivoire, Equateur, Gabon,
Guyane, Honduras, Saõ Tomé et Principe
Doctorat
Institut National Agronomique (INA-PG) / Paris / France (1991)
Diplôme d'Ingénieur Agronome de l'INA-PG INA-PG /
Paris / France (1977)
Compétences linguistiques :
Expérience professionnelle :
Langue maternelle : Français
Langues de travail : Anglais, Espagnol
Autre langue connue (lue) : Portugais
Depuis 2002 : Cirad-CP, puis Cirad-Bios
Montpellier (France)
Généticien et expert cacao
Ressources génétiques cacaoyères
Sauvegarde et études de la collection de Guyane.
Modélisation de l'élaboration du rendement (Cameroun)
Implications internationales
1997-2002 : CIRAD-CP
CNRA-Bingerville (Côte d'Ivoire)
Généticien Cacao et agronome expert cacao
Responsabilité du programme de Sélection
Récurrente Réciproque + Essai clonal international au CNRA
(projet CFC-IPGRI).
Télédétection des vergers café-cacao
en Côte d'Ivoire (CIRAD-CP/BNETD-CCT).
1996-1997 : CIRAD-CP Montpellier (France)
Généticien et expert cacao
Caractérisation des cacaoyers spontanés de
Guyane.
Expertise cacao-café en Côte d'Ivoire
(télédétection des vergers).
1995-1996 : CIRAD-CP
Guayaquil et Pichilingue (INIAP, Equateur)
Généticien cacao (projet européen
d'amélioration de la qualité du cacao équatorien)
Sélection cacao (résistances aux maladies et
qualité).
1988-1995 : Institut de recherches sur le café et
le cacao (IRCC) - puis CIRADCP
Kourou (Guyane)
Chef de mission IRCC, responsable des programmes cacao et
café
Etude du matériel guyanais spontané (Prospections,
sauvegarde, description, évaluation).
Gestion des collections cacao et café.
1981-1988 : IRCC Divo (Côte d'Ivoire)
Généticien cacao
Ressources génétiques. Sélection
cacaoyère. Phytotechnie cacaoyère.
1980-1981 : Ministère de l'agriculture, Service de
la protection des végétaux Poitiers (France)
Ingénieur d'agronomie contractuel
Adjoint au responsable des avertissements agricoles.
1977-1979 : Institut Français du Café et du
Cacao Divo, Zagné (Côte d'Ivoire)
Volontaire Service National Actif
Implantation des champs semenciers cacao de la SATMACI. Gestion
de la Base de Multiplication et de Vulgarisation de Zagné.
Un des membres fondateurs et membre du comité de
pilotage (Steering Commitee) du réseau CACAONET (« Global Cacao
Genetic Resources Network »), représentant du Cirad.
Membre à deux reprises du comité international
d'organisation des conférences internationales sur la recherche
cacaoyère organisées par la COPAL (en 2006, Costa Rica et en
2009, Bali, Indonésie).
II. Activités d'encadrement et de formation.
2-1 Formations diplômantes
- En 1996-98, j'ai participé à l'encadrement
d'une étudiante en thèse (Universités d'Abidjan et de
Paris V), Mlle Fatoumata Barry. Sa thèse s'intitulait : ((
Extraction des paramètres qualitatifs et quantitatifs pour la
caractérisation des vergers café et cacao de Côte d'Ivoire
par télédétection ». Ma participation a
concerné les aspects agronomiques des cacaoyères (et
caféières).
- Lors de mon dernier séjour de longue durée en
Côte d'Ivoire (1997-2002), j'ai participé à l'encadrement
rapproché d'un thésard et d'un étudiant en DEA de
l'Université de Cocody, pour les aspects (( Génétique
» de leurs travaux. Mathias G. Tahi (thèse soutenue en 2003
: (( Evaluation sur feuille de cacaoyer de la résistance à
Phytophthora palmivora, agent de la pourriture brune des cabosses
») est maintenant chercheur au CNRA, en charge du programme de
Sélection Récurrente et Réciproque du cacaoyer (programme
dont j'étais responsable) et, à ce titre, je continue son ((
encadrement ».
L'étudiant qui suivait un DEA à l'époque,
Désiré N. Pokou, a soutenu une thèse en 2008, avec
un sujet sur la diversité des cacaoyers utilisés en Côte
d'Ivoire ((( Etude de la diversité génétique des
populations de la sélection récurrente réciproque du
cacaoyer (Theobroma cacao L.) et de quelques populations
cultivées en milieu paysan en Côte d'Ivoire"). J'ai
continué à participer à son encadrement lors de son
travail de thèse (j'étais alors en poste à Montpellier).
Son premier article, sur l'évolution de la diversité
génétique entre les deux premiers cycles de SRR (où je
suis co-auteur) a été publié dans la revue Plant
Breeding.
- Je suis membre du comité de thèse d'un
collègue chercheur du Cirad, Patrick Jagoret, dont le sujet
s'intitule : (( Fonctionnement et dynamiques des systèmes de culture
plurispécifiques à base de cacaoyer dans un bassin de production
ancien : le centre Cameroun » (directeur de thèse J. Wery, ED
Sibaghe, soutenance prévue fin 2010).
- En 2007, j'ai participé à la formation d'un
étudiant en licence 3 pro, de l'Université de Montpellier 2,
Renaud Favier, dont le stage s'intitulait (( les cacaoyers de Guyane
française, diversité biologique et chimique ».
- Entre 1981 et 1988 (Station IFCC-IRCC de Divo, Côte
d'Ivoire), j'ai été maître de stage de plusieurs
étudiants de l'ENSA d'Abidjan (devenue ensuite l'ENSA de Yamoussoukro),
où il m'arrivait de donner des cours. Cependant, j'ai laissé sur
place les documents et, bien évidemment puisque cela remonte à 25
ans, j'aurais beaucoup de mal à retrouver les noms et "le devenir" des
stagiaires.
En 1987-88, j'ai participé à l'encadrement
d'Olivier Sounigo (inscrit en thèse à l'Université
d'Orsay) pour un sujet sur (( Le système d'incompatibilité du
cacaoyer et ses implications pour la sélection ». Toutefois, il a
abandonné son projet après le décès de son
directeur de thèse (J. Pernès). Deux articles avaient pourtant
déjà été co-publiés sur ses travaux.
A mentionner que j'ai été également
rapporteur de thèse pour un étudiant de l'University of
Queensland (Australia), Emeras Tade, dont le sujet s'intitulait : ((
Agronomic, physiological and genetic studies on a dwarf mutant of cocoa
(Theobroma cacao L.), soutenance en fin 2008.
2-2 Autres formations
Depuis que je suis en poste à Montpellier (fin 2002),
avec une lettre de mission qui mentionne explicitement mon "appui scientifique"
à plusieurs collègues, mes activités d'encadrement et de
formation ont été les suivantes :
- En 2008, j'ai participé à l'encadrement de
Mathias Tahi (chercheur ivoirien déjà mentionné
plus haut) lors d'un stage de perfectionnement à Montpellier
intitulé : « Sélection récurrente et
réciproque du cacaoyer en Côte d'Ivoire : bilan à
mi-parcours et orientation des recherches ».
- En 2005, j'ai été maître de stage pour
un chercheur camerounais de l'IRAD, Joseph Amang M'Bang. Son sujet
était intitulé : « Approches d'analyse, étude et
modélisation, des données du projet FSP Régional IMP
Cameroun ».
- En 2005, au Honduras, formation de Tito Jimenez
Chacón (et d'une quinzaine de techniciens), ingénieur
responsable en agronomie de l'ONG TechnoServe, à la reconnaissance
phénotypique des divers groupes morpho-génétiques de
cacaoyers présents dans le pays, et en particulier des Criollos.
- En 1996-97, en Côte d'Ivoire, j'ai assuré pendant
plusieurs mois la formation en agronomie cacaoyère d'un technicien
supérieur du BNETD - CCT, Souleymane Bamba.
- Entre 1981 et 1988, en Côte d'Ivoire, j'ai
été responsable de stage pour des cadres chercheurs de l'IRCC, de
l'IRHO, du BDPA, et pour des chercheurs étrangers (stages formels de
plusieurs semaines). Là aussi, il me serait assez difficile de retrouver
les traces. Néanmoins, je peux au moins citer le nom d'une chercheuse
affectée à l'IRCC par son ministère : Jeanne A. K.
N'Goran, qui est devenue plus tard responsable de l'amélioration
génétique cacao au CNRA, puis conseiller du premier ministre pour
les "produits de base" et maintenant responsable de la coopération
internationale au CNRA. Son stage concernait « La sélection
cacaoyère en RCI, aspects théoriques et pratiques ».
Parmi les chercheurs encore au Cirad actuellement, je peux
mentionner Xavier Bonneau et Didier Clément.
III. Publications et travaux
Au 15 octobre 2008, je compte actuellement à mon actif
29 publications avec Impact Factor (ISI), dont 20
en premier auteur, Cf tableau suivant. A mentionner également :
3 chapitres d'ouvrage (en co-auteur) + un en premier auteur, en cours
(Université de Davis, USA).
|
Journal
|
I. F. 2007
(JCR)
|
Nb d'articles (premier auteur)
|
Remarques
|
|
Annals of Forest Sciences
|
1,591
|
1 (1)
|
|
|
Journal of Applied Genetics
|
1,351
|
1
|
(I. F. 2008)
|
|
Scientia Horticulturae
|
0,694
|
1 (1)
|
|
|
Acta Botanica Gallica
|
0,375
|
3 (3)
|
|
|
Genetic Resources and Crop Evolution
|
0,731
|
3 (2)
|
|
|
Euphytica
|
1,050
|
1 (1)
|
|
|
Journal of Horticultural Science
|
0,658
|
3 (3)
|
|
|
Plant Breeding
|
1,092
|
1
|
|
|
Crop Protection
|
1,129
|
1
|
|
|
Canadian Journal of Botany
|
0,985
|
1
|
|
|
Café, Cacao, Thé
|
0,104 (*)
|
12 (9)
|
Fin en 1994
|
|
PLoS ONE
|
|
1
|
Revue récente
|
|
Totaux
|
|
29 (20)
|
|
|
(* : Valeur en 1994)
|
|
|
|
La revue PloS ONE, crée en décembre 2006, est
encore trop récente pour avoir un facteur d'impact. Il est à
prévoir qu'il sera élevé.
La liste de l'ensemble de mes travaux scientifiques figure
ci-dessous (octobre 2008)
- 3-1) Publications dans périodiques (revues avec
Facteur d'Impact en gras)
- premier auteur (37)
- co-auteur (14)
- 3-2) Communications orales lors de rencontres
internationales
- premier auteur (9)
- co-auteur (23)
- 3-3) Communications par posters lors de
congrès (8)
- 3-4) Divers (dont chapitres d'ouvrages)
(17)
- 3-5) Rapports de missions (28)
3.1 Publications dans périodiques
En premier auteur :
- LACHENAUD, Ph., D. ZHANG. 2008.
Genetic diversity and population structure in wild stands of
cacao trees (Theobroma cacao L.) in French Guiana
Annals of Forest Science 65:
310-316.
- LACHENAUD, Ph. 2008.
La collection de cacaoyers (Theobroma cacao) du Cirad en
Guyane française Lettres du Groupe de Ressources
Phytogénétiques d'Aquitaine, 33 : 4-6
- LACHENAUD, Ph., PAULIN, D., DUCAMP, M. & THEVENIN, J.-M.
2007.
Twenty years of agronomic evaluation of wild cocoa trees
(Theobroma cacao L.) from French Guiana.
Scientia Horticulturae 113 (4):
313-321.
- LACHENAUD, Ph. 2007.
Fruit trait variability in wild cocoa trees (Theobroma
cacao L.) from the Camopi and Tanpok basins in French Guiana.
Acta Botanica Gaiica 154 (1),
117-128.
- LACHENAUD, Ph. 2007.
Ortet selection in cacao progenies from the Kérindioutou
river in French Guiana Ingenic Newsletter, 11: 5-7
- LACHENAUD, Ph. 2006.
Les cacaoyers spontanés de Guyane
Les Amis du Muséum National d'Histoire Naturelle,
225, 1-4
- LACHENAUD, Ph., G. OLIVER, Ph. BASTIDE & D. PAULIN.
2006.
Le remplissage des cabosses des cacaoyers spontanés de
Guyane (Theobroma cacao L.)
Acta Botanica Gaiica 153 (1),
105-114
- LACHENAUD, Ph., O. SOUNIGO & B. SALLEE. 2005.
Les cacaoyers spontanés de Guyane française :
état des recherches. Acta Botanica Gaiica 152 (3), 325
- 346
- LACHENAUD, Ph., O. SOUNIGO & D. CLEMENT. 2005. The
compatibility - yield efficiency relationship. Ingenic Newsletter,
10, 13 - 16
- LACHENAUD, Ph. and G. OLIVER. 2005.
Compatibility and duration of pod maturation in Guianan wild
cocoa trees. Preliminary results.
Ingenic Newsletter, 10, 20 -
22.
- LACHENAUD, Ph. and G. OLIVER. 2005.
Variability and selection for morphological bean traits in wild
cacao trees (Theobroma cacao L.) from French Guiana.
Genetic Resources and Crop Evolution 52,
225-231
- LACHENAUD, Ph., O. SOUNIGO and G. OLIVER. 2004.
Genetic structure of Guianan wild cocoa (Theobroma cacao
L.) described using isozyme electrophoresis.
Plant Genetic Resources Newsletter 139 : 24-30.
- LACHENAUD Ph. and J. C. MOTAMAYOR. 2004.
Red pods in progenies from the Euleupousing River in French
Guiana. Ingenic Newsletter , 9, 12-15.
-LACHENAUD, Ph. & C. MONTAGNON. 2002.
Competition effects in cocoa (Theobroma cacao L.) hybrid
trials. Euphytica 128 (1): 97-104
- LACHENAUD, Ph. and G. OLIVER. 2001.
Variability in various agronomic traits of wild cocoa trees
(Theobroma cacao L.) from the Camopi and Tanpok basins (French
Guiana)
Plant Genetic Resources Newsletter 128 : 35-40
- LACHENAUD, Ph.,G. OLIVER and Ph. LETOURMY. 2001.
Agronomic assessment of wild cocoa (Theobroma cacao L.) trees
from the Camopi and Tanpok basins (French Guiana)
Ingenic Newsletter, 6, 24-26.
- LACHENAUD, Ph., G. OLIVER and Ph. LETOURMY. 2000.
Agronomic assessment of wild cocoa trees (Theobroma
cacao L.) from the Camopi and Tanpok basins (French Guiana)
Plant Genetic Resources Newsletter, n° 124 : 1-6
- LACHENAUD, Ph., F. BONNOT, OLIVER, G. 1999.
Use of floral descriptors to study variability in wild cocoa
trees (Theobroma cacao L.) in French Guiana.
Genetic Resources and Crop Evolution, 46 :
491-500.
- LACHENAUD, Ph. 1999.
Le cacaoyer et sa culture.
Ingénieurs de la Vie, n° 450,
57.
- LACHENAUD, Ph., D. CLEMENT et G. OLIVER. 1998.
Premiers clones sélectionnés dans les descendants
des cacaoyers (Theobroma cacao) autrefois cultivés en
Guyane.
Plant Genetic Resources Newsletter, n°
113 : 31-34.
- LACHENAUD, Ph., OLIVER, G. 1998.
Influence d'éclaircies sur les rendements d'hybrides de
cacaoyers. Plantations, recherche, développement, vol. 5,
n°1, 34-40.
- LACHENAUD, Ph. , V. MOOLEEDHAR et C. COUTURIER. 1997.
Les cacaoyers spontanés de Guyane. Nouvelles prospections.
Plantations, recherche, développement, vol. 4, n°1,
25-30.
- LACHENAUD, Ph., V. MOOLEEDHAR, Ch. COUTURIER. 1997
Brief report on collection of wild cacao from the Euleupousing
and Yaloupi rivers in French Guiana.
Ingenic Newsletter, 3, 20-21.
- LACHENAUD, Ph. 1997.
Genetic/taxonomic structuring of Theobroma cacao L.
species. Fresh hypotheses Ingenic Newsletter, 3, 10-11.
- LACHENAUD, Ph. and M. DUCAMP. 1996
Characterization and potential utilization of wild cacao from
French Guiana. Ingenic Newsletter, 2, 8-9.
- LACHENAUD, Ph. 1995.
Variations in the number of beans per pod in Theobroma
cacao L. in the Ivory Coast. III. Nutritional factors, cropping effects
and the role of boron.
Journal of Horticultural Science, 70 (1)
7-13.
- LACHENAUD, Ph. 1995.
Variations in the number of beans per pod in Theobroma
cacao L. in the Ivory Coast. II. Pollen germination, fruit setting and
ovule development.
Journal of Horticultural Science, 70 (1)
1-6.
- LACHENAUD, Ph. 1994.
Variations in the number of beans per pod in Theobroma
cacao L. in the Ivory Coast. I. The role of pollen.
Journal of Horticultural Science, 69 (6)
1123-1129.
- LACHENAUD, Ph., D. CLEMENT, B. SALLEE et Ph. BASTIDE. 1994.
Le comportement en Guyane de cacaoyers sélectionnés
en Côte d'Ivoire. Café, Cacao, Thé
(Paris), 38, n°2, 91-102.
- LACHENAUD, Ph. 1994.
Description du remplissage incomplet des cabosses chez
Theobroma cacao L. en Côte d'Ivoire.
Café, Cacao, Thé (Paris), 38,
n°2, 81-89.
- LACHENAUD, Ph. et B. SALLEE. 1993.
Les cacaoyers spontanés de Guyane. Localisation,
écologie et morphologie. Café, Cacao, Thé
(Paris), 37, n°2, 101-114.
- LACHENAUD, Ph. 1991.
Effet de quelques variables saisonnières sur la relation
entre poids de cabosse et poids de fèves fraîches chez le
cacaoyer. Bilan d'observations en Côte d'Ivoire. Café,
Cacao, Thé (Paris), 35, n°2, 113-120.
- LACHENAUD, Ph. et O. SOUNIGO. 1991.
Production de semences de cacaoyer en Côte d'Ivoire.
Influence des proportions relatives des parents d'un champ semencier biclonal
sur la quantité, la qualité et l'origine génétique
des semences.
Café, Cacao, Thé (Paris), 35,
n°2, 105-112.
- LACHENAUD, Ph. 1987.
L'association cacaoyer-bananier plantain. Etude de dispositifs.
Café, Cacao, Thé (Paris), 31,
n°3, 195-202.
- LACHENAUD, Ph. et G. MOSSU. 1985.
Comparative study of the influence of two methods of management
on the production factors of a cocoa farm.
Café, Cacao, Thé (Paris), 29,
n°3, 197-200.
- LACHENAUD, Ph. et G. MOSSU. 1985.
Etude comparative de l'influence de deux modes de conduite sur
les facteurs de rendement d'une cacaoyère.
Café, Cacao, Thé (Paris), 29,
n°1, 21-30.
- LACHENAUD, Ph. 1984.
Une méthode d'évaluation de la production de
fèves fraîches applicable aux essais entièrement
"randomisés".
Café, Cacao, Thé (Paris), 28,
n°2, 83-88.
En co-auteur :
- POKOU N. D., N'GORAN J. A.K., LACHENAUD
Ph., ESKES A.B., MOTAMAYOR, J.C,
SCHNELL R., KOLESNIKOVA-ALLEN M., CLEMENT D., SANGARE A. 2009
Recurrent selection of cocoa populations (Theobroma cacao L.) in
Côte-d'Ivoire: comparative genetic diversity between the first and second
cycles.
Plant Breeding 128 : 514-520
- BASTIDE, Ph., D. PAULIN & Ph.
LACHENAUD. 2008.
Influence de la mortalité des cacaoyers sur la
stabilité de la production dans une plantation industrielle.
Tropicultura 26, 1, 33-38
- MOTAMAYOR, J.C., LACHENAUD, PH., WALLACE DA
SILVA, J., LOOR, G., KUHN, D., BROWN, J. & SCHNELL, R. 2008
Geographic and genetic population differentiation of the
Amazonian chocolate tree. PLoS One, October 2008, Vol. 3, Issue 10, e3311 (8
p.)
- PAULIN, D., M. DUCAMP, PH. LACHENAUD.
2008.
New sources of resistance to Phytophthora megakarya
identified in wild cocoa trees populations of French Guiana.
Crop Protection 27: 1143-1147
- TAHI, G. M., J. A. K. N'GORAN, O. SOUNIGO, P.
LACHENAUD & A. B. ESKES. 2007. Efficacy of simplified methods
to assess pod production in cocoa breeding trials. Ingenic Newsletter,
11, 7-11.
- ASSEMAT, S., PH. LACHENAUD, F. RIBEYRE, F.
DAVRIEUX, J.-L. PRADON & E. CROS. 2005.
Bean quality traits and sensory evaluation of wild Guianan cocoa
populations (Theobroma cacao L.).
Genetic Resources and Crop Evolution, 52 :
911-917
- ESKES, A. B. and PH. LACHENAUD.
2004.
A rootstock-scion experiment with cacao re-analysed for yield
efficiency. Ingenic Newsletter, 9, 43-45.
- SOUNIGO, O., Ph. LACHENAUD, Ph. BASTIDE, C.
CILAS, J. N'GORAN, C. LANAUD. 2003.
Assessment of the value of doubled haploids as progenitors in
cocoa (Theobroma cacao L.) breeding
Journal of Applied Genetics, 44 (3),
339-353.
- N'DOUME, C., Ph. LACHENAUD, A. HUSSARD, H.
NGUYEN V. , A. FLORI. 2000. Etude de faisabilité pour
l'élaboration d'une cartographie statistique d'inventaire des vergers
café et cacao en Côte d'Ivoire par
télédétection satellitale.
Bulletin de la SFPT, 157, 3-10.
- PAULIN, D., G. MOSSU, Ph. LACHENAUD et C.
CILAS. 1993.
La sélection du cacaoyer en Côte d'Ivoire.
Analyse du comportement de soixante-deux hybrides dans quatre
localités. Café, Cacao, Thé (Paris), 37,
n°1, 3-20.
- BEREAU, M., LACHENAUD, Ph. et P.
PLANQUETTE. 1992.
Le cacaoyer en Guyane française.
Cahiers
d'Outre-mer, 45, 179-180.
- LOTODE, R. et Ph. LACHENAUD. 1988.
Méthodologie destinée aux essais de
sélection du cacaoyer. Café, Cacao, Thé
(Paris), 32, n°4, 275-292.
- LANAUD, C., Ph. LACHENAUD et O. SOUNIGO.
1988.
Comportement en croisement des haploïdes doublés de
cacaoyers. Canadian Journal of Botany, 66,
1986-1992.
-LANAUD, C., O. SOUNIGO, Y. K. AMEFIA, D. PAULIN, Ph.
LACHENAUD et D. CLEMENT. 1987.
Nouvelles données sur le fonctionnement du système
d'incompatibilité du cacaoyer et ses conséquences pour la
sélection.
Café, Cacao, Thé (Paris), 31,
n°4, 267-277.
3.2 Communications orales lors de rencontres
internationales.
En premier auteur :
- LACHENAUD, Ph. 2005.
Genetic Effects of Inter-tree Competition in Mixed Cocoa Stands
on Yield, Vigour, and Cropping Efficiency.
Proceedings of the International Workshop on Cocoa Breeding for
Improved Production Systems, 19-21/10/2003, Accra (Ghana), 110 - 117.
- LACHENAUD, Ph., O. SOUNIGO, G. OLIVER. 2005.
Paramètres et structuration génétiques des
cacaoyers spontanés de Guyane française, étudiés
par électrophorèses d'iso-enzymes.
Actes de la 14ème Conférence Internationale sur la
Recherche Cacaoyère, Accra (Ghana), 13-18 /10/2003, vol. 1, 103-110.
- LACHENAUD, Ph. 2005.
Densité évolutive en cacaoculture : la
nécessité des éclaircies.
Actes de la 14ème Conférence Internationale sur la
Recherche Cacaoyère, Accra (Ghana), 13-18 /10/2003, vol. 1, 309-315.
- LACHENAUD, Ph. 2002.
Le dilemme de la densité en cacaoculture, ou la
nécessité des éclaircies. Conférence Internationale
sur l'avenir des cultures pérennes, Yamoussoukro (Côte d'Ivoire),
5-9/11/2001.
CD Rom, ISBN 2-87614-502-2.
- LACHENAUD, Ph, A. ESKES, J. A. K. N'GORAN, D. CLEMENT, I.
KEBE, M. TAHI et C. CILAS. 2001.
Premier cycle de sélection récurrente en Côte
d'Ivoire et choix des géniteurs du second cycle.
in : Actes de la 13ème Conférence Internationale
sur la recherche cacaoyère, Kota Kinabalu (Sabah, Malaysia), 9-14
octobre 2000. Cocoa producer's alliance, Lagos (NGA), 11-22.
- LACHENAUD, Ph. 2000. Manual pollination techniques and
verification of incompatibility. In : Working procedures for cocoa germplasm
evaluation and selection : Proceedings. Rome : IPGRI, p. 98-99. CFC/ICCO/IPGRI
Project Workshop, 1998-02-01/1998-02- 06, Montpellier, France.
- LACHENAUD, Ph. 1994.
Le remplissage incomplet des cabosses en Côte d'Ivoire.
in : Actes de la 11ème Conférence Internationale
sur la recherche cacaoyère, Yamoussoukro (CIV), Cocoa producer's
alliance, Lagos (NGA), p.521-527.
- LACHENAUD, Ph. 1992.
Recent cocoa germplasm conservation initiatives of IRCC in French
Guiana. International Workshop on Conservation, Characterization and
Utilization of Cocoa Genetic Resources, Trinidad, sept.1992, 253-255.
- LACHENAUD, Ph. 1988.
La plantation en haies fruitières : une technique
permettant l'association avec les cultures vivrières et leur
stabilisation.
10ème Conférence Internationale sur la Recherche
Cacaoyère, Santo Domingo, mai 1987, 45-50.
En co-auteur :
- BASTIDE, Ph., Ph. AGUILAR, Ph. LACHENAUD,
D. PAULIN, I. JIMMY, G. BOULETARE. Variabilité du rendement et
mesures de la biomasse sur des cacaoyers adultes au Vanuatu.
15 ème Conférence Internationale sur la
Recherche Cacaoyère, San José, Costa Rica, 9-
14 octobre 2006, vol. I, pp. 291-297.
- PAULIN, D., M. DUCAMP, Ph.
LACHENAUD.
Evaluation des cacaoyers spontanés de Guyane
française pour leur résistance à Phytophthora
megakarya.
15 ème Conférence Internationale sur la
Recherche Cacaoyère, San José, Costa Rica, 9- 14 octobre 2006,
vol. 1, pp. 41-47.
- TAHI, G. M., J. A. K. N'GORAN, Ph.
LACHENAUD, O. SOUNIGO & A. B. ESKES. Mise au point d'une
méthode d'évaluation simplifiée de la productivité
du cacaoyer.
Fifth INGENIC Workshop on Cocoa Breeding for Farmers' Needs, San
José, Costa Rica, 15-17/10/2006 (sous presse)
- N'GUESSAN, F., ESKES, A.B. & LACHENAUD,
Ph. 2005.
Etude de la résistance du cacaoyer aux mirides :
classification des grands groupes de cacaoyers en fonction de leur niveau de
résistance ou sensibilité.
Actes de la Conférence Internationale sur la Recherche
Cacaoyère. 14, 2003-10- 13/2003-10-18, Accra, Ghana, Vol. 1, 559-565
- SOUNIGO, O., J. N'GORAN, D. PAULIN, Ph.
LACHENAUD, D. CLEMENT & A. B. ESKES. 2005.
Comparison of Values as Clones and as Progenitors for Yield,
Vigour and Yield Efficiency: experiences from Côte d'Ivoire.
Proceedings of the International Workshop on Cocoa Breeding for
Improved Production Systems, 19-21/10/2003, Accra (Ghana), 62 - 65.
- N'GORAN, J., Ph. LACHENAUD, I. KEBE, F.
N'GUESSAN, M. TAHI, D. POKOU, O. SOUNIGO, K. N'GORAN, A. ESKES. 2004.
Selection and breeding activities in Côte d'Ivoire.
Workshops on CFC/ICCO/IPGRI Projects. University of Reading, 28
March to 3 April 2004. CD Version.
- N'DOUME, C., Ph. LACHENAUD, A. HUSSARD,
H. NGUYEN V , A. FLORI. 2002. Cartographie statistique d'inventaire des
vergers café et cacao par télédétection
satellitale.
Conférence Internationale sur l'avenir des cultures
pérennes, Yamoussoukro (Côte d'Ivoire), 5-9/11/2001. CD Rom, ISBN
2-87614-502-2.
- N'GORAN, J.A.K., C. MONTAGNON, J.-L. KONAN, Ph.
LACHENAUD, H. LEGNATE. 2002.
Apport du matériel végétal
amélioré sur la durabilité des cultures pérennes.
Conférence Internationale sur l'avenir des cultures pérennes,
Yamoussoukro (Côte d'Ivoire), 5-9/11/2001. CD Rom, ISBN 2-87614-502-2.
- N'DOUME, C., Ph. LACHENAUD, A. HUSSARD, H.
NGUYEN V. , A. FLORI. 2001.
Etude de faisabilité pour l'élaboration d'une
cartographie statistique d'inventaire des vergers café et cacao en
Côte d'Ivoire par télédétection satellitale.
in : Actes de la 13ème Conférence
Internationale sur la recherche cacaoyère, Kota Kinabalu (Sabah,
Malaysia), 9-14 octobre 2000. Cocoa producer's alliance, Lagos (NGA),
1377-1382.
- EFRON Y., N'GORAN J.K.A., LACHENAUD, P.
2000. Evaluation of vigour, yield, pod and
bean traits. In : Working procedures for cocoa germplasm
evaluation and selection : Proceedings. Rome : IPGRI, p. 76-82. CFC/ICCO/IPGRI
Project Workshop, 1998-02- 01/1998-02-06, Montpellier, France.
- N'DOUME, C., PH. LACHENAUD, A. HUSSARD, H.
NGUYEN et A. FLORI. 1999.
Etude de faisabilité pour l'élaboration d'une
cartographie statistique d'inventaire des vergers café et cacao en
Côte d'Ivoire par télédétection satellitale.
Journées géographiques de Daloa, 22-26 mars
1999.
- CLEMENT, D., J. N'GORAN, D. PAULIN, Ph.
LACHENAUD, O. SOUNIGO and A. ESKES. 1999.
Améliorationsgénétique du
cacaoyer en Côte d'Ivoire : nouvelle sortie variétale.
in : Actes 12eme Conférence Internationale sur
la Recherche Cacaoyère, Salvador, Bahia (Brasil), Lagos (NGA) Cocoa
producer's alliance, p. 337-344.
-PAULIN, D., LACHENAUD, Ph. ,LANAUD, C.,
RISTERUCCI, A. M., LAURENT, V., N'GORAN, J. A. K., 1996.
Utilisation des marqueurs morphologiques, isoenzymatiques et
moléculaires pour l'étude de la diversité du cacaoyer
(Theobroma cacao L.)In : CIRAD-CP. Rencontres cacao. Les
différents aspects de la qualité. Actes du séminaire.
Montpellier, 30/06/1995, p. 115.
-PAULIN, D., LAURENT, V., N'GORAN, J. A. K., RISTERUCCI, A.
M., LACHENAUD, Ph., LANAUD, C. 1996.
Use of morphological, isoenzyme and molecular markers in the
study of cocoa (Theobroma cacao) diversity.
EUCARPIA, Meeting on tropical plants, Montpellier, p. 269-270.
- ESKES, A.B., D. PAULIN, D. CLEMENT, J. A. K N'GORAN, O.
SOUNIGO, Ph. LACHENAUD, C. CILAS, D. BERRY and A. YAGMPAM.
1995.
Selection methods applied and genetic knowledge generated in
cacao breeding in Côte d'Ivoire and Cameroon.
International Workshop on cocoa breeding strategies, Kuala
Lumpur, Malaysia, Oct. 1994.
- PAULIN, D., V. LAURENT, J. A. K. N'GORAN, A. M. RISTERUCCI,
Ph. LACHENAUD, C. LANAUD. 1994.
Use of morphological, isoenzyme and molecular markers in the
study of cocoa (Theobroma cacao L.) diversity.
Proceedings of the genetic resources section. Meeting of
Eucarpia, mars 1994, Clermont-Ferrand.
- CLEMENT, D., D. PAULIN, Ph. LACHENAUD et O.
SOUNIGO. 1994.
Etude du comportement d'hybrides issus de clones
hauts-amazoniens, bas-amazoniens et Trinitario, croisés avec des clones
Catongo.
in : Actes 11ème Conférence
Internationale sur la Recherche Cacaoyère, Yamoussoukro (CIV), Lagos
(NGA), Cocoa producer's alliance, p. 393-398.
- SOUNIGO, O., J. N'GORAN, N. COULIBALY, D. CLEMENT et
Ph. LACHENAUD. 1994. Evaluation des clones de cacaoyers
pour la productivité, la résistance aux mirides et la
résistance à la pourriture des cabosses.
in : Actes 11ème Conférence
Internationale sur la Recherche Cacaoyère, Yamoussoukro (CIV), Lagos
(NGA), Cocoa producer's alliance, p. 375-381.
- PAULIN, D., G. MOSSU, Ph. LACHENAUD, A. B.
ESKES. 1994.
Genetic analysis of a factoriel crossing scheme with cacao
hybrids tested in four locations in Ivory Coast.
Proceedings International Cocoa Conference, Kuala Lumpur
(Malaysia), sept. 1991, 73.
- ESKES, A. B., C. CILAS, D. PAULIN, D. CLEMENT, O. SOUNIGO,
Ph. LACHENAUD and J. A. K. N'GORAN. 1992.
CIRAD-IRCC Involvement in evaluation and utilization of cocoa
germplasm. International Workshop on Conservation, Characterization and
Utilization of Cocoa Genetic Resources, Trinidad, sept.1992, 389-397.
- N'GORAN, J. A. K., Ph. LACHENAUD, Ph.
BASTIDE and D. PAULIN. 1992. Cocoa germplasm conservation initiatives in
Côte d'Ivoire.
International Workshop on Conservation, Characterization and
Utilization of Cocoa Genetic Resources, Trinidad, sept.1992, 221-227.
- PAULIN, D., LACHENAUD, Ph., MOSSU, G.,
ESKES, A. B. 1992.
Genetic analysis of a North Carolina design II crossing scheme
with cacao (Theobroma cacao) hybrids tested in four locations in
Côte d'Ivoire.
13ème congrès EUCARPIA, Angers, juillet
1992, 567-568.
-LANAUD, C., O. SOUNIGO, D. PAULIN, Ph.
LACHENAUD et D. CLEMENT. 1988. Nouvelles données sur le
fonctionnement du système d'incompatibilité du cacaoyer et ses
conséquences pour la sélection.
10ème Conférence Internationale sur la
Recherche Cacaoyère, Santo Domingo, mai 1987, 573-580.
3.3 Posters
- LACHENAUD, Ph., D. PAULIN, M. DUCAMP & J.-M.
THEVENIN.
Les clones spontanés GU de Guyane française :
synthèse des résultats acquis et aide au choix des
géniteurs.
15 ème Conférence Internationale sur la
Recherche Cacaoyère, San José, Costa Rica, 9-
14 octobre 2006, Vol. I, pp. 539-547
- ZHANG, D. and PH. LACHENAUD.
Genetic variation and population structure in the wild cocoa
germplasm from French Guiana.
15 ème Conférence Internationale sur la
Recherche Cacaoyère, San José, Costa Rica, 9-
14 octobre 2006 (sous presse)
POKOU, N. D., N'GORAN J. A. K., KOLESHNIKOVA-ALLEN, M., ESKES,
A., LACHENAUD, Ph., SANGARE, A.
Diversité génétique des populations de
cacaoyers (Theobroma cacao L.) obtenues après le premier cycle
de sélection récurrente.
15 ème Conférence Internationale sur la
Recherche Cacaoyère, San José, Costa Rica, 9- 14 octobre 2006
(sous presse)
- LACHENAUD P., OLIVER G., LETOURMY P. 2001. Agronomic
evaluation of spontaneous cocoa-trees (Theobroma cacao L.) in the
Camopi and Tanpok basins (French Guiana). In : Actes de la
13ème Conférence Internationale sur la recherche
cacaoyère, Kota Kinabalu (Sabah, Malaysia), 9-14 octobre 2000. Cocoa
producer's alliance, Lagos (NGA),169.
- N'DOUME, C., Ph. LACHENAUD, A. HUSSARD, H. NGUYEN V. , A.
FLORI. 2000.
A feasibility study for the implementation of statistical
inventory mapping of coffee and cocoa plantations in Côte d'Ivoire by
satellite remote sensing.
XIXème Congrès de la
Société Internationale de Photogrammétrie et de
Télédétection (ISPRS), AMSTERDAM, 16-23/7/2000.
- SOUNIGO, O., C. LANAUD, Ph. LACHENAUD et Ph. BASTIDE. 1994.
Etude du comportement en croisement de quatorze haploïdes
doubles de cacaoyers (Theobroma cacao L.) en Côte d'Ivoire.
11ème Conférence Internationale sur la
Recherche Cacaoyère, Yamoussoukro, Côte d'Ivoire, Juillet 1993.
- VINCENT, A., D. CLEMENT, Ph. BASTIDE et Ph. LACHENAUD. 1994.
Etude de la répartition de la production et de son
incidence sur la pourriture brune (P. palmivora) dans une parcelle
clonale de Hauts-amazoniens.
11ème Conférence Internationale sur la
Recherche Cacaoyère, Yamoussoukro, Côte d'Ivoire, Juillet 1993.
- LACHENAUD, Ph. and O. SOUNIGO. 1994.
Influence of several densities of pollinator trees in a biclonal
seed garden in Côte d'Ivoire, on the quantity, quality and genetic origin
of seeds.
Proceedings International Cocoa Conference, Kuala Lumpur,
Malaysia, sept. 1991, 575.
3.4 Divers
(dont chapitres d'ouvrages, en gras)
- N'GORAN, J. A. K., Ph. LACHENAUD, I. B. KEBE, K. F.
N. N'GUESSAN, G. M. TAHI, D. POKOU, O. SOUNIGO, K. N'GORAN & A. B. ESKES.
2007.
Population breeding approaches applied in cocoa selection
in Côte d'Ivoire.
In: Global Approaches to Cocoa Germplasm Utilization and
Conservation, A. B. Eskes and Y. Efron, Eds., pp. 35-40. CFC
Amsterdam
- N'GUESSAN, K. F., A. B. ESKES & Ph. LACHENAUD.
2007.
Mirid resistance studies in Côte d'Ivoire: field
observations on recent and cumulative damage.
In: Global Approaches to Cocoa Germplasm Utilization and
Conservation, A. B. Eskes and Y. Efron, Eds., pp. 170-176. CFC
Amsterdam
- NGUYEN, H., Ph. LACHENAUD et A. FLORI, CIRAD-CP. 1997.
Cartographie analytique et statistique des vergers cacao et
café de deux zones pilotes DALOA et SOUBRE de Côte d'Ivoire.
CP SIC n° 859, octobre 1997 (60 p. + cartes).
- FLORI, A., Ph. LACHENAUD, H. NGUYEN. 1997.
Analyse statistique de l'enquête CIRAD-CCT dans les zones
pilotes de DALOA et SOUBRE. RCI, janvier-février 1997.
Doc n° CP-806, juin 1997 (12 p. + annexes).
- LACHENAUD, Ph. 1993.
Les cacaoyers spontanés de Guyane.
INFOS, Recherche et Technologie (Crestig, Cayenne), 26, Janvier
1993.
- LACHENAUD, Ph. 1991.
Facteurs de la fructification chez le cacaoyer (Theobroma
cacao L.). Influence sur le nombre de graines par fruit.
Thèse de Doctorat, Institut National Agronomique
PARIS-GRIGNON, 188 p.
- LACHENAUD, Ph., D. LOUETTE, F. RUF. 1990. Bahia face
à la crise.
CIRAD-DSA, 63 p.
- RUF, F., D. LOUETTE, Ph. LACHENAUD 1990.
Le Brésil.
In : la compétitivité du cacao
Africain.
Ministère de la Coopération-CIRAD (PARIS).
p 238-257.
- LACHENAUD, Ph. 1988.
Les essais en haies fruitières en Côte d'Ivoire.
In : Agronomie du cacaoyer. Paris, France, CIRAD-IRCC, 4 p.
Journées Agronomiques de l'IRCC, 1988/09/06-09, Montpellier, France.
- LACHENAUD Ph. 1988.
La greffe - buts et techniques.
In : Journées d'étude sur le cacaoyer :
amélioration du cacaoyer. Montpellier, France, CIRAD-IRCC, n.p. (2
p.). Journées d'Etude sur le Cacaoyer, 1985/07/09-11, Montpellier,
France.
- LACHENAUD, Ph. 1988.
Modes de culture pour l'expression des potentialités du
matériel amélioré.
In : Journées d études sur le cacaoyer :
amélioration du cacaoyer. Montpellier, France, CIRAD-IRCC, n.p. (2
p.). Journées d'Etude sur le Cacaoyer, 1985/07/09-11, Montpellier,
France.
- LANAUD, C., D. PAULIN et Ph. LACHENAUD. 1985.
Note sur l'amélioration génétique cacaoyer
en Côte d'Ivoire. Document interne IRCC, Direction de Côte d'Ivoire
(14 p.).
- LACHENAUD, Ph., J. BIGNON. 1985.
Contribution à l'étude de la pollinisation du
cacaoyer. Le cacaoyer est-il insuffisamment pollinisé ? Document interne
IRCC Côte d'Ivoire, 37 p.
- LACHENAUD, Ph. 1983.
Problèmes posés par l'ombrage du cacaoyer.
Evolution des techniques culturales traditionnelles.
Institut de Recherches du Café et du Cacao, 25ème
anniversaire, 1958-1982, 79-80.
- LACHENAUD, Ph. 1983.
La culture du cacaoyer en haies fruitières. Premiers
résultats (Divo, Côte d'Ivoire)
Institut de Recherches du
Café et du Cacao, 25ème anniversaire, 1958-1982, 81.
- LACHENAUD, Ph. 1983.
Production de semences hybrides de cacaoyers. Les champs
semenciers.
Institut de Recherches du Café et du Cacao, 25ème
anniversaire, 1958-1982, 89-90.
- LACHENAUD, Ph. 1982.
Comportement de jeunes cacaoyères en grande saison
sèche (Divo). Gerdat-IFCC (Divo, 5 p.)
3.5 Rapports de mission
- LACHENAUD, Ph. 2009.
Une filière cacao en Martinique. Rapport de mission (7-11
juin 2009). Cirad-BIOS, UPR 31, N° A-31 2009/04, juin 2009 (15 p.).
- LACHENAUD, Ph. 2008.
Rapport de mission en Guyane. Projet FEDER « DICACAO »
24 juin - 5 juillet 2008-09-01
CIRAD - BIOS, UPR 31, n° 2199, juillet 2008.
- LACHENAUD, Ph. 2008.
Evaluation et sauvegarde de la collection de cacaoyers de Guyane.
Rapport de mission. 10-26 janvier 2008.
CIRAD-BIOS, UPR 31, n° 2159, Février 2008.
- DUCAMP, M. & LACHENAUD, Ph. 2007.
Evaluation et sauvegarde de la collection de cacaoyers de Guyane
Rapport de mission (3-18 juin 2007), CIRAD-BIOS, n° 2100-07
- LACHENAUD, Ph. 2007.
Sauvegarde de la collection de cacaoyers de Guyane. Rapport de
mission. 10-20 décembre 2006.
Cirad-UPR 31, n° 2035-07.
- LACHENAUD, Ph. 2006.
Sauvegarde de la collection de cacaoyers de Guyane. Rapport de
mission. 7-14 décembre 2005.
CP SIC n° 1896-2006. Cirad-Cultures pérennes.
- MONTAGNON, C., E. SOMARRIBA, Ph. LACHENAUD. 2006.
Lineamientos para el Plan Estratégico de Cacao, Nicaragua
Cirad-CATIE, Managua, 14 febrero 2006 (9 p.)
- LACHENAUD, Ph. 2005.
Project on "Revitalization of the Honduran cocoa sector"
TechnoServe Honduras. Mission Report (4-17 September 2005)
CP SIC N° 1872-2005. Cirad Tree Crops.
- LACHENAUD, Ph. 2005.
Sauvegarde de la collection de cacaoyers de Guyane. Rapport de
mission. 4-18 mars 2005.
CP SIC n° 1813. Cirad-Cultures pérennes.
- LACHENAUD, Ph. 2004.
Sauvegarde de la collection de cacaoyers de Guyane.
Rapport de mission. 26 février-29 mars 2004.
CP n° 1709. Programme cacao. Cirad-Département
Cultures pérennes.
- LACHENAUD, Ph. 2003.
14ème Conférence Internationale sur la
Recherche Cacaoyère. Accra (Ghana) 13-18 octobre 2003.
4ème Atelier International (INGENIC) &
4ème Séminaire International (INCOPED). Accra (Ghana)
19-21 octobre 2003. Rapport collectif de participation. Doc n° CP-1659,
décembre 2003.
- LACHENAUD, Ph. 2003.
Sauvegarde de la collection de cacaoyers de Guyane. Rapport de
mission (II) (5-18 février 2003)
Doc n° CP-1591, mars 2003.
- LACHENAUD, Ph. 2002.
Rapport de la mission conjointe INRA-CIRAD au Cameroun (23-30
novembre 2002) Doc n° CP-1570, déc. 2002.
- LACHENAUD, Ph. 2002.
Sauvegarde de la collection de cacaoyers de Guyane.
Rapport de mission (17-29 avril 2002), Doc n°
CP-1490, mai 2002.
- LACHENAUD, Ph. 2001.
Situation du programme cacao en Guyane et état de son
dispositif de recherche. Rapport de mission (20-27 février 2001)
Doc n° CP-1489, mars 2001.
- LACHENAUD, Ph. 1998.
Le programme cacao en Guyane. Rapport de mission 22 novembre au 2
décembre 1998.
Doc n° CP-1061, 12-1998.
- LACHENAUD, Ph. 1997.
Rapport de mission en Côte d'Ivoire, du 13/1 au 23/2/1997.
Doc n° CP-746, mars 1997 (13 p. + annexes).
- NGUYEN, H., Ph. LACHENAUD. 1996.
Rapport de mission en Côte d'Ivoire (du 21 au 31 octobre
1996) Doc n° Cp-692, Nov. 1996 (12 p. + annexes).
- ESKES, A., O. SOUNIGO, Ph. LACHENAUD. 1992.
Compte-rendu de mission : atelier de travail sur les
Ressources Genetiques du cacao, Trinidad (13-18 septembre 1992). Visite au
Venezuela (18-22 septembre 1992) (23 p.) Document IRCC-CIRAD.
- LACHENAUD, Ph. 1989.
Rapport de mission au Gabon (3/12/89 - 7/12/89). Document
IRCC-SOCA 2 (8 p.).
- LACHENAUD, Ph. 1989.
Rapport de mission en République Démocratique de
São Tomé et Principe. (7- 11/12/89). Document IRCC-SOCA 2. (7
p.).
- LACHENAUD, Ph. 1989.
Rapport de mission en République Démocratique de
São Tomé et Principe. (31/3/89 au 4/4/89). Document IRCC-SOCA 2
(11 p.).
- LACHENAUD, Ph. 1989.
Rapport de mission en Equateur et au Pérou (21-31 janvier
1989). CIRAD-IRCC (15 p.)
- LACHENAUD, Ph. 1988.
Rapport de mission en République Démocratique de
São Tomé et Principe, du 28 avril au 2 mai 1988.Document
IRCC-SOCA 2 (7 p.).
- LACHENAUD, Ph. 1988.
Rapport de mission au Gabon auprès de la SOCAGAB, du 22 au
28 avril 1988. Document IRCC-SOCA 2 (9 p.).
- LACHENAUD, Ph. 1987.
Rapport de mission en République Démocratique de
São Tomé et Principe, du 19 au 26 juin 1987.
Document IRCC-SOCA 2 (28 p.).
- LACHENAUD, Ph. 1987.
Rapport de mission au Gabon Document IRCC-SOCA 2 (10 p.).
- LACHENAUD, Ph. 1986.
Rapport de mission à São Tomé.
Document interne IRCC (confidentiel, 11 p.).
IV Synthèse des résultats
scientifiques
L'amélioration génétique du
cacaoyer. Des ressources génétiques forestières aux
variétés cultivées.
Introduction
Le cacaoyer (Theobroma cacao, Linné 1753) est
un arbre de la famille des Malvacées, originaire des forêts
pluviales néotropicales, depuis le sud du Mexique (où il ne
pourrait être que féral) jusqu'au nord de la Bolivie. Deux
sous-espèces géographiques sont reconnues, T. cacao
subsp. cacao et T. cacao subsp. sphaerocarpum
(Chevalier) Cuatrecasas 1964, qui correspondent aux deux principaux
groupes cultivés, appelés Criollo et Forastero (le
troisième, Trinitario, est un hybride entre les deux autres). La
connaissance de la structuration génétique de l'espèce a
été affinée depuis (Lanaud et al., 1999 ;
Motamayor et al., 2002, 2008) : l'existence des deux
sous-espèces est maintenant contestée, au profit de celle de
plusieurs groupes morpho-géographiques. L'un de ces groupes est
constitué par les cacaoyers spontanés de Guyane
française.
Le cacaoyer, qui peut atteindre à l'état sauvage
un développement important (25 m de hauteur et 20 m de diamètre
de frondaison), est une plante cauliflore : les inflorescences, et donc les
fruits, sont localisés sur le tronc et les branches. Sur le tronc et les
branches principales, les zones d'apparition des inflorescences évoluent
en massifs appelés "coussinets floraux", qui produisent des fleurs
pendant toute la vie de l'arbre. L'inflorescence est une cyme bipare à
courtes ramifications (1 mm environ). La fleur, petite, est
régulière, complète et pentamère. Le fruit, la
"cabosse", est exploité pour ses graines. Une cabosse normale contient
de 30 à 60 graines entourées d'un abondant mucilage sucré
qui, fermentées et séchées (appelées alors
«fèves»), constituent le cacao marchand, matière
première du chocolat. Le cacaoyer, domestiqué en Amérique
centrale par les Mayas et révélé aux Européens en
1502, fut ensuite répandu par ces derniers dans les zones favorables de
tous les continents.
J'ai commencé à travailler en
amélioration du cacaoyer en 1977, à l'IFCC (Institut
Français du Café et du Cacao), comme VSNA1, en
Côte d'Ivoire. J'ai été embauché ensuite comme
ingénieur-chercheur en 1981. Depuis cette époque, mes travaux sur
le cacaoyer ont concerné deux aspects principaux :
- L'amélioration génétique et la
sélection des variétés en Côte d'Ivoire ; - Les
ressources génétiques sauvages de Guyane.
En parallèle, et fréquemment en liaison avec ces
deux domaines principaux, j'ai également travaillé en :
- Phytotechnie (problématiques du greffage, de la
densité, des concurrences) ;
- Physiologie de la production et élaboration du
rendement (cf ma thèse : Lachenaud,
1991);
- Télédétection des cacaoyères.
Dans cette synthèse, seuls seront abordés les deux
premiers points.
1 Volontaire du Service National Actif
J'ai effectué trois séjours de longue
durée en RCI : de novembre 1977 à avril 1979, de mars 1981
à juin 1988 et de septembre 1997 à juillet 2002, soit environ 13
ans et demi en tout. J'ai pu m'impliquer dans les domaines suivants :
4-1-1 Les variétés hybrides des
générations 1975 et 1979.
Après une première phase de sélection
massale dans le matériel végétal utilisé localement
(Forastero « Amelonado ouest-Africain »), l'amélioration du
cacaoyer en Côte d'Ivoire s'est orientée vers la création
d'hybrides de clones. Ainsi, depuis la fin des années 50, des Forasteros
Haut-Amazoniens (d'abord de niveau G2, puis ensuite G0 et G1, y compris des
haploïdes doublés ; Lanaud et al., 1988) ont
été croisés avec des Forasteros BasAmazoniens Amelonados
«locaux» ou des Trinitarios. Des centaines d'hybrides ont ainsi
été créés et comparés dans des essais en
paires simples («single pairs»). Les meilleurs croisements ont
été distribués aux planteurs à partir de 1974 (une
douzaine de familles). Initialement, les critères de sélection
étaient avant tout la productivité et la taille des fèves,
et secondairement, les résistances aux divers aléas, dont la
pourriture brune (causée par Phytophthora palmivora) et les
punaises mirides. Les descendances hybrides vulgarisées, avec des
rendements pouvant atteindre ou dépasser 2500 kg/ha de cacao marchand en
station de recherche (Besse, 1977), ont nettement surclassé le vieux
matériel Amelonado pour ce critère, mais également pour la
vigueur juvénile (facilitant l'implantation) et la tolérance aux
mirides, fléau de la cacaoculture ivoirienne. Utilisés tels que
distribués (une douzaine seulement de familles F1) ou plus souvent sous
forme de descendances produites chez les planteurs lors de croisements naturels
entre individus de diverses F1, les hybrides (au sens large) ont
été globalement très appréciés, et dans
certaines régions, les Amelonados sont en voie de disparition.
En règle générale, pour la
productivité, les hybrides à mâle Amelonado se sont
montrés supérieurs à ceux comportant un mâle
Trinitario. Toutefois, le minimum commercial de 1 g de moyenne pour le poids
moyen des fèves a parfois imposé de garder des géniteurs
Trinitarios (à grosses fèves). En plus d'une taille de
fève plus grande, les hybrides vulgarisés présentent
certains avantages technologiques par rapport à l'Amelonado, par exemple
un taux de matière grasse plus élevé (Besse, 1977).
Toutefois, la base génétique de ce matériel alors
utilisé par les planteurs était véritablement
étroite.
Lors de mon premier séjour, j'ai pu participer à
la plantation des champs semenciers de la Satmaci (Société de
développement), représentant environ 150 Ha, en trois sites. Il
s'agissait des formules hybrides de la première génération
(Besse, 1977). J'ai été impliqué dans toutes les
opérations (multiplication des clones parents par bouturage, gestion des
parcs-à-bois, préparation des parcelles, plantation, suivi). Ces
champs semenciers ont permis à la Côte d'Ivoire de devenir le
premier producteur mondial.
A partir de 1979, de nouvelles combinaisons hybrides ont
été créés par Mossu (Paulin et al., 1993)
et testées dans un réseau multilocal d'essais. Ces hybrides
étaient nettement plus variés que les premiers.
Etant revenu en RCI, sur la station de Divo, en mars 1981,
j'ai pu suivre, pratiquement depuis les premières observations (vigueur
juvénile), l'essai A19/1 (un des cinq du réseau initial,
planté en juin 1979 à Divo) et planter un autre essai (A18/1), en
mai 1981, soit 95 descendances en tout, en « randomisation totale »,
avec des effectifs familiaux respectivement de 20 et 40. Après plus de
10 ans de suivi, les résultats du réseau d'essais
ont été co-publiés en 1993 (Paulin et
al., 1993). En 1988, après une analyse de l'ensemble des essais de
sélection cacaoyère plantés à Divo, dont les
protocoles étaient souvent trop différents, avec des
témoins parfois absents, j'ai pu planter un essai synthétique,
D15/2, regroupant les meilleures combinaisons hybrides repérées
sur la station. Les résultats de l'essai D15/2 furent co-publiés
en 1999 (Clément et al., 1999) : ils représentent la
première sortie variétale en Côte d'Ivoire depuis la
génération d'hybrides de 1975. Il s'agit de 7 familles
proposées en mélange, présentant une meilleure
résistance à la pourriture brune que les hybrides
précédents, pour un gain de productivité de 18 % par
rapport à la moyenne des témoins de la génération
précédente. Les clones femelles impliqués sont : PA 150,
P7, NA 32, T79/501, T60/887, T85/799 et UPA 409, tous Forasteros
hauts-Amazoniens, et les mâles IFC 2, IFC 5 (Amelonados (( locaux »)
et POR (Trinitario du Vénézuela)
Entre 1981 et 1988, j'ai pu également travailler sur la
méthodologie des essais de sélection en (( randomisation totale
» (Lachenaud, 1984 ; Lotodé et Lachenaud, 1988 ; Lachenaud et
Sounigo, 1991), faire diverses introductions de matériel
végétal à partir des stations de quarantaine (et en
particulier du matériel végétal originaire de Guyane, via
Montpellier) et planter, en 1987 à Divo, avant que les résultats
définitifs du réseau d'essais 1979 ne soient connus, quatre
champs semenciers (un par famille hybride) qui sont encore utilisés
actuellement. En poste en Guyane, j'ai pu étudier dans un essai
comparatif d'hybrides planté en 1984 (en (( randomisation »
totale), le comportement des hybrides ivoiriens de la génération
précédente (Besse, 1977), et en publier les résultats
(Lachenaud et al., 1994). J'ai donc pu acquérir une très
bonne connaissance des hybrides vulgarisés en Côte d'Ivoire.
4-1-2 Le programme de sélection récurrente
réciproque (SRR)
Depuis 1990, le programme d'amélioration
génétique du cacaoyer en Côte d'Ivoire s'est orienté
vers la création de variétés certes toujours productives
(le niveau atteint dès 1975 est considéré comme
acceptable, et on vient de voir que la sortie variétale de 1999
présentait déjà un progrès de 18 %), mais
présentant une meilleure adaptation au milieu, particulièrement
quant à la tolérance aux pourritures (surtout celles
causées par P. palmivora) et aux insectes (Paulin et Eskes,
1995 ; Sounigo et al., 1999). Il était donc nécessaire
de prendre en compte de nouveaux critères de sélection,
d'élargir la base génétique du matériel
distribué et d'adopter des méthodes d'amélioration plus
efficaces. Un schéma de sélection, basé sur
l'amélioration récurrente de populations complémentaires
à bases génétiques élargies, a donc
été envisagé (Clément et al., 1994). Il
devait assurer, sur plusieurs (( cycles », un cumul de gènes
favorables (dont ceux de résistances), tout en maintenant une
variabilité génétique importante. Bien que la sortie
variétale (descendances ou clones) soit possible à tout moment du
programme, y compris donc pendant les deux premiers cycles de brassage
intra-populations, elle n'est envisagée à dessein que lors des
croisements entre populations améliorées à l'issue des
deux premiers cycles. Etant donné l'expérience acquise avec les
premiers hybrides, le choix de deux populations de base complémentaires
s'est donc porté sur les Forasteros Haut-Amazoniens (HA) d'une part et
les Forasteros Bas-Amazoniens et Trinitarios d'autre part (BA+T). La seconde
population est donc une population composite, à créer lors du
premier cycle de brassage (Fig. 1).
Le choix des clones constitutifs du premier cycle a
reposé sur leurs valeurs propres ou en croisement (connues localement ou
par la littérature), l'information disponible à l'époque
sur la structuration génétique de l'espèce (Lanaud, 1987)
et leurs origines géographiques.
Les parcelles ont été plantées en 1991-1992,
à Divo, et suivies jusqu'en 1998-99.
J'ai été affecté de nouveau en RCI en
septembre 1997, pour prendre la responsabilité du programme de SRR,
c'est-à-dire, en pratique, clôturer les essais du premier cycle,
analyser les résultats, les publier et mettre en place le second cycle.
Ces activités s'inscrivaient dans le cadre d'un projet CFC /ICCO/IPGRI
intitulé (( Cocoa Germplasm utilization and conservation: a global
approach ».
La sélection des géniteurs du second cycle de
brassage est du type «individu-famille». Les meilleures familles sont
identifiées sur la base de leurs performances en test, mais aussi sur
l'AGC des parents. Les critères de sélection retenus pour la
sélection des familles ont été les suivants :
- la mortalité ;
- la productivité (Lachenaud, 1984, 1991);
- le rapport Production-vigueur (« Cropping, ou Yield,
Efficiency »), c'est-à-dire le rapport de la production
cumulée à la section en cm2 à 50 cm du sol
à la clôture de l'essai (Lotodé et Lachenaud, 1988);
- le pourcentage de cabosses pourries ;
- le poids moyen d'une cabosse saine, surtout utile dans la
population (BA+T), car bien corrélé avec le poids moyen d'une
fève ;
- le classement des géniteurs pour leur réaction
à P. palmivora, évaluée par le
«testfeuille» (Tahi et al., 2000).
Les résultats du premier cycle (cf Tableaux 1 et 2) et
la sélection des géniteurs du second ont été
publiés (Lachenaud, in Eskes, 1999 ; Lachenaud et al.,
2001). En 2000 et 2001, j'ai pu planter 5 parcelles (4 à Divo, une
à Abengourou, soit 4,7 Ha en tout) liées au deuxième cycle
du programme de SRR, comprenant environ 190 croisements, plus les
témoins.
Le chercheur ivoirien actuellement en charge du programme (M.
Tahi) est venu à Montpellier en avril-mai 2008 pour analyser les
données disponibles issues de ces parcelles : les premiers
résultats sont très encourageants (Tab. 3). Cependant, seuls pour
l'instant les résultats concernant l'évolution de la
diversité entre les deux premiers cycle sont en cours de publication
(Pokou et al., 2008). Les autres résultats devraient être
publiés vers 2011-12.
Egalement, à l'issue du premier cycle, 7 familles ont
pu être sélectionnées pour essais de confirmation en sortie
variétale. Elles présentent un progrès considérable
par rapport aux précédents hybrides (y compris en matière
de productivité) et ont été incluses (pour 6 d'entre
elles), en 2001, dans un essai planté à Abengourou (Tab. 4).
4-1-3 L'utilisation des haploïdes doublés
A partir de 1982, j'ai pu planter et suivre à Divo deux
essais de C. Lanaud et un de G. Mossu comprenant des géniteurs
haploïdes doublés (D7, E2/1, E2/2, pour un total de 84 familles, y
compris les témoins), parallèlement à la recherche
d'haploïdes. Certains résultats ont été
copubliés (Lanaud et al, 1988 ; Sounigo et al., 2003).
Ils ont montré qu'il était possible de créer des familles
intéressantes en croisant deux parents haploïdes doublés
(moyennant un certain choix des géniteurs). Ces familles
n'étaient toutefois pas systématiquement plus homogènes
(pour les critères de sélection) que celles avec un (ou deux)
géniteurs diploïdes « normaux ». Les résultats ont
également montré qu'obtenir des haploïdes doublés
à partir de géniteurs prometteurs (à partir d'un seul
cycle d'autofécondation) était un bon moyen de les
améliorer. Dans le programme de SRR figure un clone haploïde
doublé (H 1258).
HA3
BAT3
collection de travail
sélection
sélection
HA0
BAT0
sortie variétale
sortie variétale
BA x BA BA x T
HA x HA
BAT!
BAT1 x BAT1
HA!
HA1 x HA1
sortie variétale
sortie variétale
BAT2
BAT2 x BAT2
HA2
HA2 x HA2
sélection
sortie variétale
BAT2 x HA2
sélection
sélection
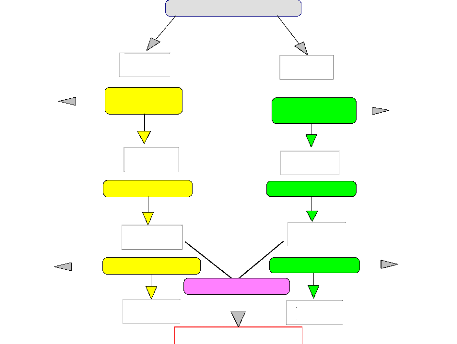
Figure 1 : le schéma du programme de
Sélection Récurrente et Réciproque du cacaoyer en
Côte d'Ivoire (d'après Clément et al., 1994). Deux
cycles de brassage intra-groupes précèdent la première
confrontation inter-groupes (BAT2 x HA2). Le cycle 3 pourra être suivi
d'autres cycles.
Tableau 1 : synthèse des analyses de la variance
(type H3 SS) réalisées sur l'ensemble des croisements HA x HA du
premier cycle de SR en RCI, pour 4 critères de sélection (TS =
très significatif, HS = hautement significatif, THS = très
hautement significatif, NS = non significatif), d'après Lachenaud et
al. (2001)
|
Critère
|
Signification des effets :
& % & x %
|
Meilleurs géniteurs
|
|
Production
|
TS
|
THS
|
NS
|
SCA 6, MO 81, IMC 57 AMAZ 15-15, PA 150 NA 32,
|
|
Production / vigueur
|
THS
|
THS
|
TS
|
SCA 6, AMAZ 15-15 T 60/887, PA 150, MO 81, T 79/501
|
|
Pertes par pourritures
|
NS
|
HS
|
NS
|
IMC 67, MO 98, IMC 57, PA 4, T 60/887, PA 150 IMC 78, SCA 6
|
|
Poids moyen de cabosse
|
THS
|
THS
|
NS
|
IMC 78, MO 81,
AMAZ 15-15, IMC 6, UPA 134, T 60/887
|
Tableau 2 : synthèse des analyses de la variance
(type H3 SS) réalisées sur l'ensemble des croisements (BA+T) x
(BA+T) du premier cycle de SR en RCI, pour 4 critères de
sélection (HS = hautement significatif, THS = très hautement
significatif, NS = non significatif), d'après Lachenaud et al.
(2001)
|
Critère
|
Signification des effets :
& % &x %
|
Meilleurs géniteurs
|
|
Production
|
THS
|
THS
|
THS
|
IFC 303, W41, CC 10 ICS 84, IFC 304, WA 40 IFC 1, IFC 371
|
|
Production / vigueur
|
THS
|
THS
|
THS
|
IFC 303, CC 10, WA 40 ICS 84, MAT 1-6,
IFC 304
|
|
Pertes par pourritures
|
THS
|
NS
|
THS
|
IFC 6, GS 29, VENC4-11 ACU 85, IFC 14, SNK 12, N 38, ICS 89
|
|
Poids moyen de cabosse
|
THS
|
HS
|
NS
|
SNK 12, W 41, GS 29 CC 10, VENC4 -11, ICS 95
|
Tableau 3 : Moyennes des divers hybrides et groupes
(ajustées par rapport aux témoins, pour les variables PP
(production potentielle), CP (cabosses pourries), RPVIG (rapport production /
vigueur), NCab (nombre total de cabosse par arbre), PMCab (poids moyen d'une
cabosse) et Sec (section), d'après M.Tahi (rapport de stage 2008 - comm.
pers.)
|
Groupes
|
Nombre
de
familles
|
PP
(kg de
cab.)
|
CP
(%)
|
RPVIG
(kg/cm2)
|
NCab
|
PMCab
(kg de
cab.)
|
Sec
(cm2)
|
|
Familles du test
intergroupes
|
6
|
103,9 a
|
14,7 ab
|
0,68 a
|
198,8 b
|
0,52 a
|
162,8 a
|
|
Hybrides 1979
|
4
|
59,5 b
|
13,4 ab
|
0,42 b
|
100,1 cd
|
0,49 a
|
149,5 ab
|
|
Hybrides avec
|
|
|
|
|
|
|
|
|
Guyanais
|
10
|
110,9 a
|
9,4 a
|
0,78 a
|
249,2 a
|
0,46 ab
|
155,4 ab
|
|
HA2
|
75
|
64,0 b
|
7,9 a
|
0,47 b
|
134,5 c
|
0,44 ab
|
140,9 abc
|
|
(BA+T) 2
|
70
|
25,4 c
|
12,9 ab
|
0,22 c
|
54,7 de
|
0,42 ab
|
111,9 ed
|
|
HA1
|
78
|
60,8 b
|
15,7 ab
|
0,46 b
|
128,8 c
|
0,41 ab
|
132,6 bcd
|
|
Hybrides 1975
(Besse)
|
10
|
53,9 b
|
18,5 b
|
0,46 b
|
96,25 cd
|
0,38 b
|
118,7 cd
|
|
(BA+T) 1
|
116
|
24,4 c
|
13,8 ab
|
0,25 c
|
30,42 e
|
0,36 b
|
92,4 e
|
Les moyennes suivies des mêmes lettres sont statistiquement
identiques (Test de Student Newman et Keuls au seuil de 5 %)
Tableau 4 : familles
pré-sélectionnées pour la sortie variétale,
à l'issue du premier cycle de brassage, avec * : moyenne, ** : bonne,
*** : très bonne performance, (n) : classement dans l'essai. (le
témoin est un hybride de la génération 1979).
D'après Lachenaud et al., (2001)
|
Croisements
|
Production
(en % du témoin)
|
Production/vigueur
|
Pourritures
|
|
MOQ 413 x SCA 6
|
236
|
*** (1)
|
**
|
|
POR x T60/887
|
209
|
*** (3)
|
**
|
|
PA 150 x POR
|
133
|
**
|
**
|
|
SCA 6 x LF 1
|
125
|
***
|
**
|
|
IFC 303 x IFC 1
|
168
|
***(3)
|
*
|
|
IFC 29 x IFC 303
|
166
|
*** (4)
|
*
|
|
N 38 x GU 133/1
|
156
|
*** (1)
|
***
|
4-2 Les cacaoyers spontanés de Guyane
En Guyane française, l'histoire de la cacaoculture se
déroule de 1734 aux débuts du XXème
siècle, et connaît son apogée vers 1840, où quelques
cinquante tonnes d'un cacao réputé de qualité
supérieure étaient produites annuellement. La provenance du
matériel végétal alors utilisé dans les plantations
reste inconnue (Lachenaud & Sallée, 1993). L'utilisation des
cacaoyers spontanés locaux, si elle ne peut être totalement
exclue, est sans aucun doute restée marginale (Lanaud, 1987).
Depuis 1729 l'existence de "forêts" de cacaoyers
sauvages est mentionnée par les explorateurs du sud de la Guyane. Les
localisations indiquées, assez imprécises, restreignent toutefois
leur présence aux berges des rivières Kérindioutou (nom de
l'Oyapok dans son cours supérieur), Camopi, Yaloupi et Euleupousing,
toutes affluents de l'Oyapok, frontière actuelle entre la Guyane et le
Brésil. Également mentionnés, les cours supérieurs
de la rivière Tanpok et de ses affluents, bien qu'appartenant au bassin
du Maroni, se situent cependant dans cette même région. Aucun
témoignage consistant, passé ou actuel, ne fait état de
l'existence de véritables cacaoyers spontanés ailleurs en Guyane
(dans ses limites actuelles).
J'ai été affecté en Guyane d'octobre 1988
à juin 1995, puis ai continué à superviser les
activités locales jusqu'à aujourd'hui.
Depuis 1978, le Cirad (à l'époque, IFCC, puis en
1981, IRCC) s'est fixé comme objectif l'étude des cacaoyers de
Guyane, d'abord ceux anciennement cultivés (Clément, 1986), que
l'on pensait issus des cacaoyers sauvages locaux, puis les cacaoyers
spontanés (prospection, collecte, sauvegarde, caractérisation,
évaluation), dont les populations, d'accès
particulièrement difficile, sont situées en zone indienne
protégée (extrême sud-est du département). Le but de
ces études est bien entendu d'utiliser, si possible, ces cacaoyers
"inconnus" dans les programmes d'amélioration génétique de
cette plante. J'ai été personnellement impliqué dans tous
les aspects liés à cet objectif, depuis la collecte en
forêt primaire, la sauvegarde ex situ, la caractérisation
et l'évaluation, l'étude de la diversité et de la
structuration génétique, la sélection de géniteurs,
la mise à disposition des sélectionneurs et l'utilisation dans
les programmes d'amélioration (Lachenaud et al., 2005).
4-2-1 Prospection et collecte
J'ai pu organiser (et participer à) deux prospections en
forêt primaire : - en 1990 : rivières Kérindioutou et
Oyapok
Cette prospection (29 juin - 17 juillet 1990) a
concerné le cours supérieur de l'Oyapok, appelé
Kérindioutou en amont du village amérindien de Trois-Sauts. Trois
populations furent identifiées et collectées : Borne 7, vers les
sources de la Kérindioutou, Ker, sur le cours aval de la
Kérindioutou et Pina, sur l'Oyapok proprement dit (Lachenaud et
Sallée, 1993).
- en 1995 : rivières Oyapok, Euleupousing et Yaloupi
Du 13 au 26 avril 1995, dans le cadre d'un projet F.I.C.
(Fonds Interministériel Caraïbes, France) associant le Cirad et le
Cocoa Research Unit (C. R. U.) de l'University of the West Indies (Trinidad et
Tobago), une seconde prospection sur le haut-Oyapok et ses affluents fut
entreprise. Deux nouvelles zones à cacaoyers furent découvertes
sur les berges des criques Euleupousing et Yaloupi, (Elp et Yal) et quelques
prélèvements furent effectués le long du fleuve Oyapok
(Lachenaud et al., 1997).
4-2-2 Sauvegarde
La sauvegarde ex situ des représentants des
pieds-mères sauvages (il s'agit alors soit de semenceaux issus de
descendances libres, soit plus rarement de clones des pieds-mères)
s'effectue en parcelles de collection, sur la station de Paracou-Combi (commune
de Sinnamary). Il en va de même des ortets sélectionnés sur
place après caractérisation et évaluation. La sauvegarde
peut aussi être effectuée en partie en station de quarantaine. Le
germplasme GU, principalement prospecté en 1987 (7 pieds-mères
furent collectés en 1985) est toujours bien représenté
plus de vingt ans après sa collecte : 94 % des pieds-mères ont
encore des représentants en collections, en Guyane ou ailleurs, ce qui
est un résultat assez exceptionnel. En effet, Bartley (2005) fait
remarquer que le Brazilian Genetic Resources Program a perdu plus de 50 % des
pieds-mères collectés depuis 1976. Pour les prospections de 1990
et 1995 (Clones Borne 7, Ker, Pina, Elp, Yal etc..), 81 % des piedsmères
collectés sont toujours représentés en collection.
Les bons résultats du programme du Cirad peuvent
être expliqués, en partie, par le fait que pratiquement seules des
cabosses ont été prélevées sur les
pieds-mères (le taux de réussite des greffes est toujours plus
faible) et que, très rapidement, ce matériel a été
distribué dans plusieurs pays. Il est actuellement présent dans
dix-sept pays producteurs (Lachenaud et al., 2007). En 2004-2005, une
parcelle sous ombrière artificielle, regroupant l'ensemble des clones
intéressants et décrits (« core collection » d'environ
190 clones spontanés), a été installée à
Paracou-Combi (parcelle S5).
4-2-3 Caractérisation et évaluation
Les principales activités de recherche menées en
Guyane ont concerné la caractérisation et l'évaluation du
matériel génétique spontané (aspects morphologiques
et biochimiques, comportement agronomique) dans le but de fournir des
indications pratiques aux chercheurs pour l'utilisation des cacaoyers
guyanais.
Une partie des travaux de caractérisation (et la
prospection de 1995) furent financés par le FIC, pour un projet qui
associait le Cirad et le CRU de Trinidad (1994-1996).
Les arbres issus des prospections 1987 et 1990 ont
été observés individuellement pendant dix ans pour les
descripteurs agronomiques suivants, qui sont tous des critères de
sélection :
- la croissance juvénile (accroissement de section au
collet entre un et deux ans au champ) ; - la vigueur adulte (section du tronc
à dix ans à 50 cm du sol) ;
- la précocité de production (= production
cumulée à cinq ans) ;
- la production (cumulée à dix ans) ;
- le rapport production / vigueur à dix ans (= "Cropping
Efficiency") ;
- le pourcentage de cabosses pourries (cumul à dix ans)
;
- le poids moyen d'une cabosse.
Pour le même matériel végétal,
certains descripteurs de fleurs, de fruits et de fèves (qu'ils soient ou
non des critères de sélection, qualitatifs ou quantitatifs) ont
été étudiés sur des individus
pré-sélectionnés (pour leur croissance juvénile et
leur précocité) ou sur les clones des pieds-mères
sauvages. Les effectifs utilisés, par clone ou arbre
étudié, sont généralement ceux
préconisés par l'IPGRI2 (Anon. 1981). La
compatibilité et la durée de maturation ont également
été étudiées.
2 International Plant Genetic Resources Institute
(Rome), maintenant Bioversity
L'évaluation comprend également l'étude
de la valeur en croisement du matériel spontané de Guyane, pour
divers critères de sélection agronomiques, comme la vigueur, la
productivité, le rapport production / vigueur, les résistances.
Un essai factoriel incomplet 5 x 8 fut planté à Paracou-Combi en
1997-98 (7 clones guyanais spontanés et 6 clones d'autres groupes,
Forasteros et Trinitarios). Suite à divers problèmes logistiques,
cet essai fut arrêté en 2001 et seules les données de
vigueur juvénile sont disponibles (Lachenaud et al., 2007). Les
résultats acquis pour ce critère nécessitent une
confirmation pour d'autres critères, mais, tout comme les
résultats obtenus en Côte d'Ivoire, ils semblent indiquer les
mérites considérables des clones GU en hybridation.
Les résultats de toutes ces études menées
en Guyane ont été publiés : Lachenaud et al.,
1999, 2000, 2006 ; Lachenaud et Oliver, 2001, 2005a, 2005b ; Lachenaud, 2007
(voir par ex., Tab. 5). Dans cette rubrique, on peut également citer les
études concernant les qualités technologiques et sensorielles des
chocolats élaborés à partir des cacaos guyanais, dont une
synthèse a été publiée par Assemat et al.,
(2005). Les origines (populations, descendances, clones) à
préférer, suivant leurs performances par rapport aux divers buts
des sélectionneurs, ont pu être identifiées (Lachenaud
et al., 2007). Il reste toutefois de nombreuses données
à analyser et publier, en particulier concernant le matériel de
la dernière prospection, en 1995.
En plus de la publication des résultats, les
données sont, depuis le début, transmises
régulièrement à l'Université de Reading (UK) qui
les intègre dans la banque de données ICGD (International Cocoa
Germplasm Database).
En dehors de la Guyane, c'est surtout le comportement
vis-à-vis des bioagresseurs qui a été
étudié, sur des effectifs de clones variables, souvent faibles
cependant. Une revue des résultats acquis a été
présentée par Lachenaud et al., (2007). Paulin et
al., (2006, 2008) ont publié les résultats des
tests-feuilles menés au Cirad à Montpellier vis-à-vis de
la résistance aux P. palmivora et P. megakarya,
agents de la maladie de la pourriture brune (ex. Tab. 7). Au
Brésil, J.L. Pires a confirmé l'intérêt de certains
génotypes guyanais, et du groupe dans son ensemble, comme nouvelle
source de résistance à la maladie du balai-de-sorcière,
causée par Moniliophthora perniciosa (Pires, 2003). En
Côte d'Ivoire, N'guessan et al. ont montré que le
comportement des clones guyanais vis-à-vis des punaises mirides est
globalement excellent et supérieur ou égal à celui des
meilleures origines (N'guessan et al., 2007).
4-2-4 Diversité génétique et
structuration
Les descripteurs cités précédemment sont
sensibles à l'action du milieu, même si certains,
présentant une forte héritabilité (au sens large), ont une
très bonne valeur discriminante, comme les descripteurs floraux
(Lachenaud et al., 1999). Les marqueurs biochimiques et
moléculaires ne présentent pas cet inconvénient, car ils
permettent la caractérisation du génotype en certains points
(locus). L'électrophorèse d'iso-enzymes sur gel d'amidon a
été la méthode retenue en 1994, pour sa fiabilité
et sa simplicité. Six systèmes enzymatiques furent
utilisés dans les études menées à Kourou de 1995
à 1998 (PGM, PGI, MDH, PAC, ADH et IDH). Certains de ces systèmes
impliquant plusieurs gènes, le nombre total de locus
considéré fut de neuf. 138 clones représentant dix-sept
populations furent ainsi étudiés (Lachenaud et al.,
2004).
L'ensemble des données de caractérisation
(descripteurs biochimiques, floraux,
agronomiques) a donc permis
d'étudier la variabilité rencontrée dans le
matériel spontané de
Guyane, ainsi que la structuration de
cette variabilité. Dès 2004, les diverses populations
spontanées de cacaoyers ont pu être
regroupées en deux ensembles : un ensemble (( amont » et un (( aval
» (Tab. 5).
Plus récemment, une étude quasi exhaustive (189
clones représentant 18 demes) a été menée en
collaboration avec l'USDA, à l'aide de 15 marqueurs micro-satellites ((
standardisés » pour le cacaoyer, et a donné des
résultats importants sur la diversité génétique des
cacaoyers sauvages de Guyane et la structuration de cette diversité
(Lachenaud et Zhang, 2008). Ils confirment, en ce qui concerne les
paramètres de génétique des populations (He, Ho, Fst), les
résultats obtenus à l'aide de marqueurs iso-enzymatiques
(Lachenaud et al., 2004). La structuration en populations confirme
également les résultats précédents et montre que
les (( populations » identifiées en forêt primaire par les
collecteurs sont bien une réalité. Une importante structuration
existe dans la métapopulation de cacaoyers spontanés guyanais (cf
Tab. 6), avec un Fst de 0,20, qui pourrait s'expliquer par les
caractéristiques biologiques du cacaoyer et les événements
climatiques ayant affecté les forêts guyanaises au Quaternaire.
4-2-5 Utilisation dans les programmes d'amélioration
Les essais de géniteurs guyanais spontanés,
encore peu nombreux et généralement récents, n'ont produit
que peu de résultats publiés. En Côte d'Ivoire, les
résultats acquis semblent montrer une très bonne transmission aux
descendants de la résistance au P. palmivora des
géniteurs spontanés guyanais : ainsi le clone GU 123-B,
classé très résistant au test-feuille, voit ses
descendants (avec les géniteurs P7, IFC 1 et IFC 11) également
classés très résistants (Anon. 2004). Dans ce même
pays, une descendance avec GU 133-1 (classé résistant) a
été pré-sélectionnée pour essais multilocaux
avant vulgarisation, pour sa productivité élevée, son
excellent "Cropping efficiency" et son très faible % de pertes par
pourritures (cf Tab. 4, page 27 ; Lachenaud et al., 2001).
J'ai pu planter dans ce pays, en 2001, un essai factoriel
impliquant trois clones guyanais GU (originaires de la rivière Camopi),
demi-frères de clones déjà connus comme résistants
au P. palmivora au test-feuille. Les résultats disponibles
(Anon. 2006 ; Tahi, comm. pers., 2008) montrent des performances excellentes
pour le rendement et la résistance au champ à la pourriture brune
des familles comportant un géniteur guyanais.
Des clones guyanais GU sont également présents
dans quelques croisements à Trinidad (3 clones), au Ghana (14 clones),
en Amérique (un clone dans un essai multilocal implanté dans 5
pays). Suite à la publication d'une revue sur les performances du
germplasme GU (Lachenaud et al., 2007), de nombreux clones ont pu
être conseillés comme géniteurs, suivant les
critères de sélection choisis. Trois clones ((
élites», c'est-à-dire excellents ou très bons pour
l'ensemble des critères, ont même été
identifiés et livrés aux partenaires en RCI et au Costa Rica. Ils
ont également été proposés, parmi d'autres, au
Cameroun et au Nigeria.
Tableau 5: Average values observed for four bean
traits, in the main five populations and the two groups, with ABW = average
bean weight, L = average bean length, W = average bean width, and %FB =
percentage of flat beans (in Lachenaud and Oliver, 2005 a).
Only means followed by a different letter are significantly
different (P < 0.05)
|
Germplasm
|
Number
|
ABW
|
L
|
W
|
%FB
|
|
of trees
|
(g)
|
(mm)
|
(mm)
|
|
|
Cam 1
|
21
|
3.05
|
22.60
|
11.97
|
4.44 c
|
|
Cam 3
|
9
|
2.91
|
22.92
|
12.34
|
5.81 bc
|
|
Cam 7
|
14
|
2.99
|
22.92
|
12.21
|
3.40 c
|
|
Cam 9
|
32
|
2.92
|
22.40
|
11.50
|
8.58 ab
|
|
Cam 13
|
9
|
2.92
|
22.82
|
12.03
|
11.50 a
|
|
"Upstream group» (1)
|
39
|
3.05
|
22.75
|
12.09 a
|
4.33 b
|
|
"Downstream group» (2)
|
54
|
2.92
|
22.56
|
11.74 b
|
8.79 a
|
Tableau 6. Inter-population variation measured by Nei's
distances (1972, under the diagonal) and pairwise FST (above the
diagonal), with Cam = Camopi and Elp = Euleupousing
|
Population
|
Cam 1
|
Cam 3
|
Cam 7
|
Cam 9
|
Cam 12
|
Cam 13
|
Elp
|
"Primitive"
|
|
Cam 1
|
-
|
0.136
|
0.037
|
0.081
|
0.148
|
0.078
|
0.050
|
-
|
|
Cam 3
|
0.138
|
-
|
0.150
|
0.040
|
0.050
|
0.050
|
0.147
|
-
|
|
Cam 7
|
0.036
|
0.201
|
-
|
0.098
|
0.168
|
0.074
|
0.060
|
-
|
|
Cam 9
|
0.090
|
0.032
|
0.141
|
-
|
0.053
|
0.026
|
0.109
|
-
|
|
Cam 12
|
0.164
|
0.042
|
0.237
|
0.046
|
-
|
0.087
|
0.166
|
-
|
|
Cam 13
|
0.085
|
0.035
|
0.101
|
0.029
|
0.078
|
-
|
0.099
|
-
|
|
Elp
|
0.056
|
0.187
|
0.076
|
0.145
|
0.224
|
0.122
|
-
|
-
|
|
"Primitive"
|
0.811
|
0.867
|
0.718
|
0.776
|
0.805
|
0.772
|
0.545
|
-
|
|
"Parinari"
|
1.202
|
1.202
|
1.005
|
1.054
|
1.126
|
1.092
|
0.931
|
0.511
|
Populations differentiations (FST) are all
significant, except for Cam 1 - Cam 7 and Cam 9 -
Cam 13. « Primitive
» and « Parinari » are control groups (in Lachenaud and Zhang,
2008)
Tableau 7: General classification of the Guianan clones,
with adjusted mean values (in bold, control clones) : 0 < HR<1< R<
2< MR < 2.5 < S < 3.5 < HS <5. (in Paulin et al.,
2008)
Clone Adjusted Degree of
mean resistance
GU 123-V 0.16 HR
GU 226-V 0.59 HR
KER 1-L 0.60 HR
GU 195-V 0.67 HR
GU 125-C 0.71 HR
B7-B6 0.84 HR
ELP 20-A 0.89 HR
GU 269-V 1.03 R
ELP 16-A 1.06 R
GU 255-V 1.11 R
GU 241-V 1.13 R
ELP 22-A 1.18 R
B7-B5 1.19 R
IMC47 1.27 R
GU 301-A 1.34 R
GU 134-B 1.34 R
GU 263-V 1.35 R
GU 254-A 1.37 R
GU 285-B 1.43 R
B7-B3 1.43 R
GU 143-B 1.47 R
GU-353-V 1.50 R
GU 183-G 1.53 R
GU 134-C 1.55 R
GU 143-A 1.58 R
GU 156-B 1.60 R
GU 139-A 1.65 R
GU 285-A 1.76 R
GU 175-V 1.79 R
GU 268-A 1.80 R
GU 303-B 1.80 R
GU 230-B 1.88 R
GU 307-V 1.91 R
GU 147-P 1.94 R
GU 230-C 1.96 R
GU 140-S 1.99 R
GU 237-V 2.00 R
GU 257-E 2.08 MR
B7-A2 2.18 MR
GU 213-V 2.21 MR
GU 233-P 2.28 MR
GU 185-G 2.37 MR
KER 11.1-A 2.54 S
KER 6 2.57 S
PINA 2.62 S
GU 134-A 2.63 S
YAL 3 2.64 S
GU 129-B 2.66 S
GU 329-V 2.67 S
ELP 28-A 2.71 S
ELP 2-B 2.81 S
ELP 37-A 3.04 S
KER 3 3.08 S
OC77 3.10 S
KER 8-R 3.16 S
MXC67 3.16 S
GU 312-V 3.17 S
GU 138-A 3.26 S
GF23 3.28 S
ICS1 3.36 S
GU 265-V 3.40 S
EQX3360 3.56 HS
ELP 40-B 3.61 HS
KER 9 3.66 HS
OYA 2-B 3.73 HS
4-3 Conclusions
Les travaux présentés dans cette courte
synthèse s'inscrivent dans une démarche d'amélioration des
variétés cultivées de cacaoyer, dans une optique de
développement (mandat du Cirad). Depuis mon premier contact avec la
recherche cacaoyère (en novembre 1977), j'ai pu m'impliquer dans presque
toutes les disciplines et domaines concernant la plante et les peuplements
cultivés, depuis la collecte des ressources génétiques
sauvages en forêt primaire jusqu`à la
télédétection des vergers, en passant par la
caractérisation et l'évaluation des ressources
génétiques (phénotypage), l'amélioration des
variétés, la sélection, la pollinisation, la physiologie
de la production et les composantes du rendement, la phytotechnie, etc...
Le coeur du travail a cependant toujours été
l'utilisation des ressources génétiques pour
l'amélioration des variétés cultivées, avec, depuis
quelques années, les résistances aux bioagresseurs comme
critères de sélection prépondérants. Les travaux
ont toujours été réalisés dans le cadre de
collaborations (en expatriation ou non) avec les pays producteurs dont
principalement le premier d'entre eux, la Côte d'Ivoire, et dans
ce cas avec une composante (( formation » primordiale, mais aussi avec des
institutions de pays du nord, comme l'USDA par exemple. Les résultats
pratiques les plus marquants sont la création de très bonnes
variétés en Côte d'Ivoire (productives, résistantes,
sélectionnées sur leur (( cropping efficiency », produisant
un cacao de bonne qualité) et la sélection de clones
spontanés guyanais intéressants pour de nombreux critères,
en particulier les résistances aux maladies (cf Tab. 7, pour la
résistance au P. megakarya).
4-4 Références citées
Anon., 1981. Genetic Resources of Cocoa. Report "IBPGR Working
Group on Genetic Resources of cocoa", IBPGR Secretariat, Roma, 25 p.
Anon., 2004. CFC/ICCO/IPGRI Project on: Cocoa Germplasm
Utilization and Conservation: a Global Approach. Final Individual Reports April
1998-September 2003, Volumes 1 & 2.
Anon., 2006. CFC/ICCO/IPGRI Project on: Cocoa Productivity and
Quality Improvement, a Participatory Approach. Appendix to the General Progress
Report Year 2. Individual Institute Reports. 1, June 2005-31 May 2006.
Assemat, S., Lachenaud, Ph., Ribeyre, F., Davrieux, F., Pradon,
J.-L., Cros, E., 2005. Bean quality traits and sensory evaluation of wild
Guianan cocoa populations (Theobroma cacao L.). Genetic Resources
and Crop Evolution 52, 911-917.
Bartley, B. G. D., 2005. The Genetic Diversity of Cacao and its
Utilization. CABI Publishing (UK and USA), 341 p.
Besse, J. 1977. La sélection générative du
cacaoyer en Côte d'Ivoire : bilan et orientation des recherches.
5ème Conférence Internationale sur la Recherche Cacaoyère,
Ibadan (Nigéria), 95-103.
Clément D., 1986. Cacaoyers de Guyane. Prospections.
Café Cacao Thé (Paris), 30 (1), 11- 36.
Clément, D., A. Eskes, O. Sounigo et J. N'goran. 1994.
Amélioration génétique du cacaoyer en Côte d'Ivoire.
Présentation d'un nouveau schéma de sélection.
11ème Conférence Internationale sur la Recherche
Cacaoyère, Yamoussoukro (Côte d'Ivoire), 451-455.
Clément, D., J. N'goran, D. Paulin, Ph. Lachenaud, O.
Sounigo, A. B. Eskes. 1999. Amélioration génétique du
cacaoyer en Côte d'Ivoire. Nouvelle sortie variétale.
12ème Conférence Internationale sur la Recherche
Cacaoyère, Salvador, Bahia (Brésil) 1996, 337-344.
Cuatrecasas J., 1964. Cacao and its allies. A taxonomic
revision of the genus Theobroma. Contributions from the United States
National Herbarium, 35 (6). Smithsonian Institution. Washington, D. C.
Eskes, A. B. 1999. CFC/ICCO/IPGRI project on Cocoa Germplasm
Utilization and conservation - a global approach. Appendix to general progress
report of year 1, Individual Institutes Reports, IPGRI, June 1999.
Lachenaud, Ph. 1984. Une méthode d'évaluation de la
production de fèves fraîches applicable aux essais
entièrement «randomisés».
Café, Cacao, Thé (Paris), 28 (2),
83-87.
Lachenaud, Ph. 1991. Effet de quelques variables
saisonnières sur la relation entre poids de cabosse et poids de
fèves fraîches chez le cacaoyer. Bilan d'observations en
Côte d'Ivoire. Café, Cacao, Thé (Paris), 35 (2),
113-120.
Lachenaud, Ph. 1991. Facteurs de la fructification chez le
cacaoyer (Theobroma cacao L.). Influence sur le nombre de graines par
fruit. Thèse, Institut National Agronomique ParisGrignon, 188 p.
Lachenaud, Ph. 2007. Fruit trait variability in wild cocoa trees
(Theobroma cacao L.) from the Camopi and Tanpok basins in French
Guiana. Acta Botanica Gallica 154 (1), 117-128.
Lachenaud, Ph., Sallée, B., 1993. Les cacaoyers
spontanés de Guyane. Localisation, écologie, morphologie.
Café, Cacao, Thé (Paris) 37, 101-114.
Lachenaud, Ph., D. Clément, B. Sallée, Ph. Bastide.
1994.
Le comportement en Guyane de cacaoyers sélectionnés
en Côte d'Ivoire. Café, Cacao, Thé (Paris), 38
(2), 91-101.
Lachenaud, Ph., Mooleedhar, V., Couturier, C., 1997. Wild cocoa
trees in French Guiana. New surveys. Plantations Recherche
Développement 4, 25-32.
Lachenaud, Ph., Bonnot, F., Oliver, G., 1999. Use of floral
descriptors to study variability in wild cocoa trees (Theobroma cacao
L.) in French Guiana. Genetic Resources and Crop Evolution 46,
491-501.
Lachenaud, Ph., Oliver, G., Letourmy, Ph., 2000. Agronomic
assessment of wild cocoa trees (Theobroma cacao L.) from the Camopi
and Tanpok basins (French Guiana). PGR Newsletter 124, 1-6.
Lachenaud, Ph., Oliver, G., 2001. Variability in various
agronomic traits of wild cocoa trees (Theobroma cacao L.) from the
Camopi and Tanpok basins (French Guiana). PGR Newsletter 128,
35-40.
Lachenaud, Ph, A. Eskes, J. A. K. N'Goran, D. Clément, I.
Kebe, M. Tahi et C. Cilas. 2001. Premier cycle de sélection
récurrente en Côte d'Ivoire et choix des géniteurs du
second cycle.
in : Actes de la 13ème Conférence Internationale
sur la recherche cacaoyère, Kota Kinabalu (Sabah, Malaysia), 9-14
octobre 2000. Cocoa producer's alliance, Lagos (NGA), 11-22.
Lachenaud, Ph., Sounigo, O., Oliver, G., 2004. Genetic structure
of Guianan wild cocoa (Theobroma cacao L.) described using isozyme
electrophoresis. PGR Newsletter 139, 24-30.
Lachenaud Ph, Sounigo O, Sallée B. 2005. Les cacaoyers
spontanés de Guyane française : état des recherches.
Acta Botanica Gallica 152 (3): 325 - 346
Lachenaud, Ph., Oliver, G., 2005a. Variability and selection
for morphological bean traits in wild cocoa trees (Theobroma cacao L.)
from French Guiana. Genetic Resources and Crop Evolution 52,
225-231.
Lachenaud, Ph., Oliver, G., 2005b. Compatibility and Duration of
Pod Maturation in Guianan Wild Cocoa Trees - Preliminary results. Ingenic
Newsletter 10, 20-22.
Lachenaud, Ph., Oliver, G., Bastide, Ph., Paulin, D., 2006. Le
remplissage des cabosses des cacaoyers spontanés de Guyane
(Theobroma cacao L.). Acta Botanica Gallica 153, 105- 114.
Lachenaud Ph., Paulin D., Ducamp M., Thevenin J.-M., 2007. Twenty
years of agronomic evaluation of wild cocoa trees (Theobroma cacao L.)
from French Guiana. Sciencia Horticulturae 113: 313-321.
Lachenaud, Ph., D. Zhang. 2008. Genetic diversity and
population structure in wild stands of
cacao trees (Theobroma cacao
L.) in French Guiana. Annals of Forest Science 65: 310-316.
Lanaud, 1987. Nouvelles données sur la biologie du
cacaoyer (Theobroma cacao L.). Thèse de doctorat d'état,
Université de Paris-sud, 106 p (+ annexes).
Lanaud, C., Ph. Lachenaud, O. Sounigo. 1988. Comportement en
croisement des haploïdes doublés de cacaoyers (Theobroma
cacao). Canadian Journal of Botany, 66, 1986-1992.
Lanaud C., J.C. Motamayor & O. Sounigo, 1999. Le cacaoyer.
In: Diversité génétique des plantes tropicales
cultivées. P. Hamon et al., Eds. Collection
Repères, Cirad, 387 p.
Lotodé, R., Ph. Lachenaud. 1988. Méthodologie
destinée aux essais de sélection du cacaoyer. Café,
Cacao, Thé (Paris), 32 (4), 275-292.
Motamayor J. C., A.-M. Risterucci, P. A. Lopez, C.F. Ortiz, A.
Moreno & C. Lanaud, 2002. Cacao domestication I : the origin of the cacao
cultivated by the Mayas. Heredity 89, 380- 386.
Motamayor, J.C., Lachenaud, Ph., Wallace Da Silva E Mota, J.,
Loor, G., Kuhn, D., Brown, J. & Schnell, R. 2008.
Geographic and genetic population differentiation of the
Amazonian chocolate tree. PLoS One, October 2008, Vol. 3, Issue 10, e3311 (8
p.)
N'guessan, K.F., Eskes, A.B., Lachenaud, Ph., 2007. Mirid
resistance studies in Côte d'Ivoire: field observations on recent and
cumulative damage. In: Eskes, A.B., Efron, Y. (Eds.), Global Approaches to
Cocoa Germplasm Utilization and Conservation, CFC Technical Paper N° 50,
pp. 170-181.
Paulin, D., G. Mossu, Ph. Lachenaud, C. Cilas. 1993. La
sélection du cacaoyer en Côte d'Ivoire. Analyse du comportement de
soixante-deux hybrides dans quatre localités. Café, Cacao,
Thé (Paris), 37 (1), 3-19.
Paulin, D., A. B. Eskes. 1995. Le cacaoyer : stratégies de
sélection. Plantations Recherche Développement, 2 (6),
5-13.
Paulin, D., Ducamp, M., Lachenaud, Ph., 2006. Evaluation des
cacaoyers spontanés de Guyane française pour leur
résistance à Phytophthora megakarya. Proceedings of the
15th International Conference on Cocoa Research, San José
(Costa Rica), 9-14/10/2006 (sous presse).
Paulin, D., M. Ducamp, P. Lachenaud. 2008. New sources of
resistance to Phytophthora megakarya identified in wild cocoa tree
populations of French Guiana. Crop Protection 27: 1143-1147.
Pires, J.L., 2003. Avaliação quantitativo e
molecular de germoplasma para o melhoramento do cacaueiro com enfase na
produtividade dos frutos e rêsistencia. Ph D Thesis, Universidad federal
de Viçosa, MG, Brazil (242 p.).
Pokou N. D., N'goran J. A.K., Lachenaud Ph., Eskes A.B.,
Kolesnikova-Allen M., Clement D., Sounigo O., Sangare A. 2008. Recurrent
selection of cocoa populations (Theobroma cacao L.) in
Côte-d'Ivoire: comparative genetic diversity between the first and second
cycles. Plant Breeding (sous presse)
Sounigo, O., Coulibaly, N., Brun L. A., N'Goran J.A.K., Cilas C.,
Eskes A.B. 1999. Etude de la sensibilité aux mirides (Distantiella
theobromae et Sahlbergella
singularis) dans des essais de descendances de cacaoyers
de Côte d'Ivoire.
Proceedings 12ème Conférence Internationale sur la
recherche Cacaoyère, 1996, Salvador de Bahia (Brésil),
331-336.
Sounigo, O., Ph. Lachenaud, Ph. Bastide, C. Cilas, J. N'Goran, C.
Lanaud. 2003. Assessment of the value of doubled haploids as progenitors in
cocoa (Theobroma cacao L.) breeding. Journal of Applied
Genetics, 44 (3), 339-353.
Tahi, M., I. Kébé, A. B. Eskes, S. Ouattara, A.
Sangaré and F. Mondeil. 2000.
Rapid screening of cacao genotypes for field resistance to
Phytophthora palmivora using leaves, twigs and roots. European
Journal of Plant Pathology, 106, 87-94
V Le projet de recherches
Diversité des cacaoyers spontanés de
Guyane. Utilisation dans la lutte contre les bioagresseurs.
5.1 - Objectifs
Le projet a pour objectif principal d'enrichir les
connaissances sur la diversité d'une plante pérenne tropicale de
grande importance économique, le cacaoyer (Theobroma cacao L.)
dans le but de mieux lutter contre les maladies. Il vise à
obtenir des clones de cacaoyers spontanés de Guyane utilisables dans la
création de variétés durablement résistantes. Le
matériel génétique spontané guyanais présent
dans la collection du Cirad à Sinnamary en Guyane sera
évalué exhaustivement pour sa résistance à trois
des principales maladies du cacaoyer (les pourritures à Phytophthora
spp., le balai-de-sorcière dû à Moniliophthora
perniciosa et le "mal de machete", dû à Ceratocystis
fimbriata 3), domaine où des résultats
prometteurs ont déjà été acquis. L'ensemble des
clones sera également caractérisé pour la
résistance du cortex des cabosses à la pénétration,
facteur de la résistance au (( Cocoa Pod Borer » (Conopomorpha
cramerella), important ravageur des cabosses du sud - est asiatique qui
étend actuellement son aire de dispersion.
A l'aide des résultats obtenus, et de ceux
déjà acquis en caractérisation morphologique et en
évaluation agronomique, des clones pourront être proposés
aux sélectionneurs et chercheurs en amélioration du cacaoyer,
afin qu'ils puissent les intégrer dans leurs programmes
d'amélioration nationaux, pour lutter contre les bioagresseurs
présents chez eux. Ils pourront aussi intégrer dans leurs futures
variétés des gènes de résistance pour une ((
amélioration préventive » (cas de la
résistance aux maladies américaines du cacaoyer,
balai-de-sorcière et moniliose, à intégrer dans les
variétés cultivées en Afrique ou en Asie, par exemple).
D'autre part, l'intérêt des cacaoyers
spontanés de Guyane ne se limite pas à être des sources de
résistance génétique à divers bioagresseurs,
utilisables en lutte génétique. Ils pourraient
également héberger des communautés de champignons
endophytes, comme c'est le cas dans d'autres zones d'origine, utiles en
lutte biologique. Il s'agit là d'un domaine nouveau, totalement
inconnu en ce qui concerne ces cacaoyers. Ces associations d'endophytes,
perdues ( ?) avec la domestication, pourraient, après restauration en
culture, participer à la protection du cacaoyer vis-à-vis des
maladies citées.
5.2 Le projet et les résultats attendus
5.2.1 La problématique scientifique
Les cacaoyers spontanés du sud-est de la Guyane
française ont fait l'objet de plusieurs expéditions de
prospections et de collecte entre 1985 et 1995 (Lachenaud & Sallée,
1993 ; Lachenaud et al., 1997). Les nombreuses études
menées sur ce nouveau matériel génétique (par
exemple Sounigo et al., 1996, 1999, 2001 ; Lachenaud et al.,
2000 , 2001, 2004, 2008) en ont montré la grande originalité. La
caractérisation et l'évaluation de ce matériel ont pour
but de fournir des indications pratiques aux sélectionneurs pour son
utilisation en amélioration génétique (Lachenaud et
al., 2007) ; ainsi la variabilité (diversité) pour les
critères agronomiques (vigueur, précocité, production,
"cropping-efficiency", taille des cabosses, résistance au champ
vis-à-vis des pourritures ; Lachenaud et al. 2000, 2001), la
3 En synonymie avec Ceratocystis cacaofunesta
(Engelbrecht & Harrington, 2005)
granulométrie et les caractères morphologiques
des fèves (Lachenaud & Oliver, 2005) et des fruits (Lachenaud,
2007), les caractéristiques organoleptiques (Assemat et al.,
2005) des importantes populations des bassins des rivières Camopi et
Tanpok ont été publiées. Il en est de même de
certains résultats sur la structuration génétique du
groupe dans son ensemble et des paramètres de génétique
des populations (Lachenaud et al., 1999, 2004 ; Lachenaud et Zhang,
2008). Parallèlement, l'intérêt potentiel de ce
matériel se précise dans la lutte contre les pourritures brunes,
avec la mise en évidence de clones très résistants (Paulin
et al., 2005, 2006, 2008 ; Lachenaud et al., 2007), dans la
lutte contre la maladie du balai-de-sorcière (Pires, 2003 ; Lachenaud
et al., 2007) et dans la lutte contre les mirides (N'guessan et
al., 2005, 2007). Ces organismes sont responsables de la relative
stagnation de la production cacaoyère dans toutes les zones productrices
de cacao. Il s'agit donc d'un matériel assez exceptionnel et il est
important, d'une part de poursuivre les prospections de matériel sauvage
(car il reste en Guyane des zones non prospectées) et, d'autre part, de
caractériser et évaluer l'ensemble des clones disponibles
pour leur comportement vis-à-vis de certains bioagresseurs.
Les champignons endophytes sont un groupe ancien qui a
co-évolué en symbiose avec les plantes supérieures. Il
semble que lorsque les plantes sont extraites des forêts de leurs centres
d'origine ou de diversification, ces endophytes soient perdus (Taylor et
al., 1999; Evans et al., 2003). En effet, des
échantillonnages dans des plantations de cacaoyers situées en
dehors des zones d'origine (à la fois en Afrique et Amérique)
révèlent des groupements d'endophytes très pauvres, ce qui
pourrait expliquer les nombreux problèmes sanitaires rencontrés.
En Equateur et au Panamá, les endophytes trouvés en forêt
sur des cacaoyers sauvages et naturalisés (Arnold & Herre, 2003 ;
Arnold et al., 2003) ont montré qu'ils peuvent conférer
aux cacaoyers cultivés une résistance vis-à-vis des
pathogènes (Clay, 2004). Le "centre d'origine" du cacaoyer se situe en
zone amazonienne et, par exemple, des données préliminaires de
haute-Amazonie montrent que les groupements d'endophytes du cacaoyer y
comprennent des espèces de groupes non connus comme habituellement
endophytes. Les hypothèses à tester dans le projet seront donc
que les cacaoyers des populations naturelles de Guyane hébergent des
groupements d'endophytes différents de ceux d'autres zones et que ces
endophytes pourraient être exploités dans une stratégie de
protection durable des cultures.
Cette partie du travail, à effectuer en
coopération avec le laboratoire "Systematic Botany and Mycology" de
l'USDA (Beltsville, Maryland, USA), pourrait constituer un autre projet (de
thèse), à part entière, si les premiers résultats
se révèlent encourageants.
5.2.2 L'état de l'art
Dans le cadre du projet CFC/ICCO/IPGRI (1998-2003) "Cocoa
Germplasm Utilization and Conservation : a global approach", des clones de
cacaoyers spontanés de Guyane, essentiellement issus de la prospection
1987 (clones GU, en nombre limité) ont été testés
vis-à-vis de plusieurs espèces de Phytophthora, par le
test précoce sur disques de feuilles. Les résultats acquis au
Cirad à Montpellier, au CNRA (RCI), au CRIG (Ghana) montrent que des
clones très résistants, souvent supérieurs aux
références, existent dans le matériel guyanais (IPGRI,
2001 ; Paulin et al. 2005, 2006, 2008 ; Lachenaud et al.,
2007). Au Brésil, J. Pires (Cepec) a confirmé
l'intérêt du groupe dans son ensemble comme nouvelle source de
résistance au balai-de-sorcière (Pires, 2003). Toutefois, la plus
grande partie du matériel génétique guyanais n'a pas
été testée, et en particulier les populations
prospectées en 1990 et 1995 (dont certaines se révèlent
les plus variées, Sounigo et al. 2001; Lachenaud et
al. 2004). De nouvelles et précieuses sources de résistance
aux maladies citées seront donc probablement découvertes chez ces
cacaoyers. Vis-à-vis de la résistance aux insectes, les
résultats obtenus en RCI contre les mirides (N'guessan et al.,
2007) sont très encourageants. La forte proportion de cortex des
cabosses des clones guyanais (Lachenaud, 2007) pourrait constituer un avantage
dans la lutte contre le « Pod Borer ».
5.2.3 Le contexte international
La collection guyanaise du Cirad, qui est la collection de
référence pour le matériel spontané de Guyane, est
une collection importante (plus de 550 "accessions" au total) où sont
représentés environ 200 pieds-mères issus de vingt
populations sauvages originaires de cinq bassins. Seule une partie
limitée du matériel génétique sauvage guyanais a
été transférée, soit au Cirad à Montpellier,
soit à l'Université de Reading (station de quarantaine), soit
(par l'intermédiaire des quarantaines) dans une quinzaine de pays
producteurs de cacao (dont Brésil, Côte d'Ivoire, Cameroun,
Equateur, etc...). Ce matériel a essentiellement été
étudié par le Cirad en Guyane (caractérisation
morpho-agronomique, structuration de la variabilité). Les tests de
résistance aux bioagresseurs sont relativement récents, et ont
toujours été réalisés sur des effectifs
limités. Une (( core collection >, d'environ 200 clones,
établie en Guyane en 2004-05, sous ombrière artificielle,
présente les conditions idéales
d'homogénéité pour tester localement (à l'aide du
test-feuille) l'ensemble des génotypes contre les pourritures brunes
causées par P. palmivora et P. capsici, les deux
espèces présentes en Guyane, ainsi que pour tester (sur
branchettes) la résistance au (( Mal de machete >,.
5.2.4 Les travaux envisagés
Concernant la caractérisation et l'évaluation des
cacaoyers de Guyane, les activités suivantes seront menées dans
le cadre du projet :
- Identification (sur la base de critères
morphologiques) de souches locales de Phytophthora pathogènes
du cacaoyer (P. palmivora et P. capsici). Isolement et mise en culture
des souches pour leur caractérisation moléculaire.
Stratégie identique pour M. perniciosa et C.
fimbriata. (à réaliser en Guyane par un étudiant de
niveau M1 de l'Université Antilles-Guyane).
- Evaluation (au laboratoire) d'environ 190 génotypes
guyanais pour leur résistance aux Phytophthora, sur disques de
feuilles (et sur cabosses détachées pour un échantillon de
clones résistants). Tests en pépinière sur les mêmes
génotypes pour la résistance à M. perniciosa.
(par dépôt d'une goutte gélosée d'inoculum).
Test sur baguettes des mêmes génotypes pour l'évaluation de
la résistance à C. fimbriata. Test au
pénétromètre pour évaluer la dureté des
cabosses (à réaliser en Guyane).
- Transfert du matériel végétal
résistant et/ou intéressant pour d'autres critères au
Cirad à Montpellier pour l'évaluation de son niveau de
résistance vis-à-vis de P. megakarya (espèce
très dangereuse mais non présente en Amérique), et
également de souches plus agressives de M. perniciosa
originaires d'autres pays d'Amérique, voire de souches de
Lasiodiplodia sp. (à réaliser à Montpellier, en
partie par un étudiant M1).
5.2.5 Les protocoles expérimentaux
Tous les protocoles devront faire l'objet d'une adaptation locale
; voici, dans l'état actuel des connaissances, ce que l'on peut proposer
:
A) Evaluation de la résistance aux Phytophthora
- Clones de cacaoyers à tester
L'étude concernera la parcelle S5 de la station de
Combi, installée sous ombrière artificielle. 191 objets seront
testés, dont 186 clones de cacaoyers spontanés guyanais. Les
autres origines sont représentées par deux témoins de
résistance (SCA 6, répété deux fois, et T 60/887)
et GF 24, un clone guyanais "Primitif" (cloné dans les
variétés anciennement cultivées) déjà inclus
dans un essai multilocal CFC/ICCO/IPGRI. Les témoins de
sensibilité sont deux clones spontanés guyanais,
déjà testés à Montpellier.
- Témoins Résistants : SCA 6 et T 60/887
- Témoins Sensibles : ELP 40-B
(répété deux fois) et OYA 2-B
- Matériel fongique : souches de Phytophthora
spp.
On utilisera des souches locales (testées pour leur
virulence) de P. palmivora et de P. capsici.
- Culture des souches : la culture des souches se
réalise sur milieu gélosé PDA ou V8 en boîtes de
Petri ou tubes de verre. La température de stockage doit être
comprise entre 20 et 25°C, à l'obscurité.
- Maintien du pouvoir pathogène : l'alternance, tous
les 6 mois, entre culture en boîtes de Petri et en tubes de verre permet
généralement de maintenir l'agressivité des souches.
Toutefois, si une perte d'agressivité d'une souche était
constatée, il faudrait alors inoculer cette souche sur des cabosses
matures vertes, puis l'isoler à nouveau sur milieu spécifique, en
boîtes ou en tubes, à 25°C et à l'obscurité.
- préparation de l'inoculum : 10 jours avant la date
prévue d'inoculation, les souches à inoculer (3 ou 4) sont
repiquées sur milieu V8 + betasitostérol gélosé en
boites de Petri (diamètre : 10 cm). Les boites sont placées
pendant 3 jours à 25°C et à l'obscurité. Puis elles
sont placées à 25°C et à la lumière
(photopériode 12h/12h) afin d'obtenir la formation des sporocystes qui
seront à l'origine de la production des zoospores.
- Inoculation, incubation, notation des
symptômes
Le jour de l'inoculation, 4 à 5 ml d'eau
distillée stérile seront rajoutés dans chaque boîte.
Les boîtes seront alors placées dans un
réfrigérateur à 4°C pendant 30 mn. Le choc froid
favorise la libération de zoospores contenues dans les sporocystes. Les
boîtes sont ensuite placées à 20°C pendant 2 heures
pour que les zoospores puissent être relâchées dans l'eau.
Puis, pour chaque souche, la solution contenant les zoospores est
récupérée dans un bécher. Un comptage des zoospores
est fait grâce à une cellule de Malassez. La suspension de
zoospores sera diluée afin d'obtenir une concentration de 300 000
zoospores / ml.
L'inoculation avec la suspension calibrée doit se faire le
plus rapidement possible afin d'éviter que les zoospores ne perdent leur
capacité à nager dans l'eau.
L'évaluation de base comprendra 5 à 6
répétitions, séparées par au moins
un mois. Il s'agira en fait de séries, car il est probable que certains
clones manqueront à chaque série.
Lors de chaque répétition, on utilisera :
- Une feuille par clone (on s'efforcera de prendre tous les
clones lors de chaque répétition), avec à chaque fois les
témoins de résistance et de sensibilité) au stade 3,
c'est-à-dire des feuilles de 50 à 60 jours, cueillies le matin
(6h30- 9h00), stockées en glacière et rapportées au
laboratoire le plus rapidement possible.
- 20 disques par feuille (10 pour P. palmivora, 10
pour P. capsici), déposés face inférieure vers le
haut. Dans chaque bac, chaque clone sera représenté par deux
disques (un pour P. palmivora et 1 pour P. capsici). Lors de
chaque répétition, la disposition des disques sera la même
dans chaque bac (d'où la nécessité de faire un plan lors
du placement des disques).
- 1 gouttelette de 10 uL par disque, déposée avec
une micro-pipette à répétition. - Durée
d'incubation : 7 jours à 25°C et à l'obscurité
- Lecture des symptômes suivant une échelle de 0
à 5 :
0 : absence de symptômes
1 : minuscules points nécrotiques (= points de
pénétration)
2 : points nécrotiques nombreux (points en
réseau)
3 : taches se rejoignant (taches en réseau)
4 : grandes taches uniformes (marbrées)
5 : très grandes taches nécrotiques,
dépassant parfois les limites de la goutte d'inoculation (taches
vraies).
Dans un deuxième temps (ou à Montpellier),
quelques répétitions de confirmation seront effectuées
avec une concentration élevée (>1.0 million de zoospores /ml)
pour les clones classés "Très résistants" après les
tests de base.
B) Evaluation de la résistance à
Moniliophthora perniciosa
- Clones de cacaoyers à
tester
Il s'agira des mêmes clones que ceux utilisés lors
des tests de résistance à Phytophthora spp. Les
témoins seront les clones SCA 6 (résistant) et ICS 46
(sensible).
5 greffes seront nécessaires par clone. Les plants devront
être âgés de 6 à 8 mois lors des inoculations.
- Matériel fongique : souches de M. perniciosa
- En début de petite saison des pluies (novembre),
récolter des balais-de-sorcière secs non
détériorés sur Theobroma cacao et T.
grandiflorum.
- Désinfecter les balais en les trempant dans une solution
d'hypochlorite de sodium (30 minutes, 0,5 % de chlore).
- Pendre les balais dans une unité de production de
basidiocarpes et faire alterner des périodes humides et des
périodes sèches pour stimuler la production de basidiocarpes.
- Les basidiocarpes seront récoltés
délicatement et placés au dessus d'un bécher contenant une
solution de glycérol 16 % / MES4 0,01M / Tween 0,01% à
pH 6,1. La libération des spores a lieu de nuit.
- Le lendemain matin, évaluer la concentration en
spores à l'aide d'une cellule de comptage et vérifier le taux de
germination sur milieu de culture "eau gélosée" (4 heures
après ensemencement). Calibrer la suspension pour une utilisation
immédiate ; sinon, la récupérer dans des cryotubes (1,8 -
2 ml) qui seront stockés dans l'azote liquide jusqu'à
l'utilisation ultérieure.
- Inoculation, incubation, notation des
symptômes
La méthode d'inoculation proposée est une
adaptation de celle développée par SurujdeoMaharaj et
al. (2003).
4 MES = acide
2-morpholinoéthanesulfonique-1-hydrate ; PM = 195,24
- Environ 2 semaines avant la date d'inoculation prévue,
tailler les plants afin d'induire la formation de pousses
végétatives synchrones.
- Sortir les cryotubes de l'azote liquide et les faire
dégeler au bain-marie (30°C) quelques minutes.
- Inoculer chaque bourgeon à l'aide d'une goutte
gélosée d'inoculum, au stade "Flush 2", tel que décrit
dans Greathouse et al. (1971), à une concentration à
préciser localement.
- Incuber en atmosphère saturée en humidité
(chambre d'incubation) à une température d'environ 24-26°C
pendant 48 heures.
- Après la période d'incubation, les plants seront
remis en pépinière, sous ombrière.
- La première variable analysée sera la "date
d'apparition du premier symptôme" : pour cela, chaque bourgeon sera
observé quotidiennement pendant les quatre premières semaines
afin de détecter l'émergence des symptômes. Ensuite, pour
les plants résistants, les observations seront faites tous les 15
jours.
- La deuxième variable analysée sera le
"diamètre maximal du balai à sa base" : pour cela, la base du
balai sera mesurée au pied-à-coulisse une fois par semaine
jusqu'à obtention du diamètre maximal.
- Le pourcentage de bourgeons présentant des
symptômes (gonflement, nécrose, balai) ainsi que le pourcentage de
bourgeons malades ayant développé un balai seront aussi
évalués.
Chaque plant constitue une répétition. Il n'est
prévu qu'une seule série d'inoculation (sur 3 ou 4 jours).
Toutefois, une seconde série sera possible, un an après, sur les
mêmes plants, après plusieurs tailles importantes.
C) Evaluation de la résistance à
Ceratocystis fimbriata
- Clones de cacaoyers à
tester
Il s'agira des mêmes clones que ceux utilisés
lors des tests de résistance à Phytophthora spp.
(2-5-1). Lors des tailles des arbres en parcelle S5, deux branches (ou
branchettes) de 30 cm, aoûtées, seront
récupérées pour chaque clone à tester. Cinq clones
témoins seront utilisés (IMC67, EET 400, résistants ou
moyennement résistants, CCN 51, ICS 95, sensibles et PA 120, très
sensible ; Delgado et Echandi, 1965 ; Delgado et Suarez, 2005 ; Silva et
al, 2004 ; Delgado, 2007).
- Matériel fongique : souches de C. fimbriata
- Isolement de souches sur de jeunes plantules ou arbres
attaqués par les Xyleborus, coléoptères scolytes
qui forent des galeries au niveau du collet des plantules et qui introduisent
ainsi le pathogène dans la plante (Grosclaude et al., 1988 ;
Vigouroux, 1979 ).
- Les souches seront multipliées et conservées en
collection au laboratoire à Kourou, sur milieu gélosé PDA,
avec régénération périodique.
- Une première série de tests
préliminaires sur les 5 clones témoins permettra de
sélectionner la souche la plus agressive, qui sera ensuite
utilisée au cours des tests complets d'évaluation.
- Inoculation, incubation, notation des
symptômes
On s'inspirera des travaux de Delgado et Echandi (1965),
Dominguez (1971, 1976), Przybyl (1988), Delgado et Suarez (2005) et Delgado
(2007) :
- Chaque branchette sera découpée en trois morceaux
de 10 cm de long. Au total, 6 morceaux seront inoculés par clone et par
date d'inoculation.
- Sur chaque morceau, deux perforations (1 mm de
diamètre) seront effectuées à l'aide
d'une
mini-perceuse : une à 1 cm d'une extrémité (témoin,
sans inoculation), l'autre à 3 cm
(inoculation avec du mycélium ou une suspension de spores,
suivant les résultats des essais préliminaires).
- Les morceaux seront placés dans des plateaux en
conditions humides, à 25°C et avec un éclairage 12h /12h.
- Les notations des dimensions de la nécrose se feront au
bout de 3 et 7 jours. On clivera la moitié des branchettes à 3
jours et l'autre à 7.
Par date d'inoculation, 50 clones peuvent être
testés (les clones témoins seront inoculés pour toutes les
dates). Chaque clone sera inoculé 5 fois (donc 5 x 6 = 30 morceaux de
branchettes inoculées par clone). Les clones résistants seront
ensuite confirmés au champ, par inoculations artificielles (Silva et
al., 2007)
D) Caractérisation de la dureté des
cortex.
Ces travaux, assez novateurs, s'appliqueront aux mêmes
clones que ceux des tests précédents, mais, nécessitant
des cabosses, pourront prendre beaucoup plus de temps. Ils seront menés
parallèlement à des tests identiques se déroulant en Asie
du sud-est (sur des clones différents). Ils reposent sur l'utilisation
d'un pénétromètre pour fruits. Le protocole sera à
adapter localement, les bases étant les suivantes (Bekele et Butler,
2000 ; Bekele et al., 1997, 2001) :
- Utilisation de cabosses normales (non
déformées) présentant les premiers signes de
maturité (environ trois semaines avant la pleine maturité),
testées le jour même de la récolte ;
- Les mesures seront effectuées en zig-zag, dans la zone
équatoriale de la cabosse (sur une bande de 2 cm de large) ;
- 15 mesures seront effectuées pour chaque cabosse : 5
dans chacun des sillons principaux et secondaires, et 5 dans les côtes
;
- 5 cabosses seront utilisées (au moins) par clone ;
- Des témoins seront choisis en fonction des tests
réalisés en Asie du sud-est, afin de pouvoir faire le lien avec
nos données. Les témoins de dureté du cortex seront les
clones ICS 84, ICS 1 et ICS 6 (Bekele et al., 2001).
Les cacaoyers guyanais ayant les cortex particulièrement
épais, ils pourraient être intéressants dans la lutte
contre le Cocoa Pod Borer.
E) Tests à Montpellier (Cirad)
Parmi les clones qui se révèleront "Très
résistants" ou "Résistants" au P. palmivora et à
M. perniciosa en Guyane, certains seront expédiés
à Montpellier pour y être testés contre P.
megakarya (et également, contre des souches très agressives
de M. perniciosa).
Les baguettes des clones retenus seront
expédiées à Montpellier dès la conclusion des tests
en Guyane. Dix plants de chaque clone seront nécessaires (5 pour les
tests M. perniciosa et 5 pour les tests-feuille vis-à-vis de
P. megakarya). Des semences seront donc expédiées de
Guyane six mois auparavant, afin de disposer en temps voulu de porte-greffes au
bon stade. Les baguettes des clones seront expédiées en deux
fois, à 2 mois d'intervalle, en juin et août de l'année 2.
Les tests à Montpellier commenceront lorsque les plants greffés
auront 6 mois.
F) Prélèvements des endophytes
Les souches endophytes des champignons Trichoderma
sont capables de former des associations endophytes dans les troncs des
arbres. Certains de ces endophytes montrent un bon potentiel pour le
contrôle des maladies du cacaoyer (Evans et al., 2003 ; Holmes
et al., 2004 ; Crozier et al. , 2006 ; Samuels et al.,
2006 ).
Les cultures de Trichoderma et/ou autres seront
obtenues à partir de deux sources : les endophytes des troncs et des
feuilles de cacao (et d'autres espèces ligneuses), d'une part, et les
endophytes issus du sol, d'autre part.
- Collecte d'endophytes des troncs
La diversité des champignons endophytes sera
étudiée dans des populations naturelles de cacaoyers. Les
prospecteurs rechercheront des cacaoyers sauvages et y échantillonneront
les endophytes des feuilles et des troncs. On recherchera en particulier des
cacaoyers apparemment résistant à la maladie du
balai-de-sorcière.
Echantiionnage
Le nombre d'arbres à échantillonner
dépendra du nombre de cacaoyers sauvages trouvés et sera donc
déterminé au moment de la prospection, l'objectif étant
d'échantillonner au moins 6 arbres sur chaque site. Les données
GPS de chaque arbre seront enregistrées.
Des échantillons issus de cacaoyers introduits
(collection de Paracou-Combi) et de la végétation dominante
environnante seront également récoltés, afin de comparer
les différentes espèces d'endophytes.
Isolement
On suivra le protocole d' Evans et al (2003).
Une surface d'écorce d'environ 8 x 6 cm sera
prélevée sur chaque tronc à hauteur d'épaule (1,5
m) à l'aide d'une machette désinfectée (alcool + flamme).
La partie exposée sera nettoyée avec un scalpel (lame n° 22)
et la couche sous-jacente avec une lame n° 10a. Dix tranches triangulaires
d'aubier (0,8 x 0,5 cm) seront prélevées et
transférées individuellement (avec des pinces stériles)
dans des boîtes de Petri (3 cm de diamètre) contenant deux types
de milieux : 5 à base de 20 % de PDA (Potato Dextrose Agar) et d'une
solution de 10 mg/l de pénicilline-streptomycine, et 5 de MEA (Malt
Extract Agar) + chloramphenicol (0,05 g/l). Les boîtes de Petri seront
scellées avec du ruban adhésif et conservées en
boîtes plastiques. Les fenêtres ouvertes dans les troncs seront
obturées immédiatement avec un mastic horticole pour blessures.
Tous les instruments seront désinfectés à l'alcool (+
flamme). Les boîtes de Petri seront envoyées au laboratoire de
l'USDA à Beltsville (USA) avec un permis APHIS (Animal and Plant Health
Inspection Service).
Les cultures seront examinées pendant une période
de 8 semaines, en incubation à 25°C. Du mycélium et des
spores seront isolés dès leur apparition.
- Collecte d'endophytes des feuilles
Echantiionnage
Sur les mêmes arbres, on prélèvera des
feuilles. De même, des échantillons de feuilles issus de la
végétation dominante seront également
récoltés, ainsi que des cacaoyers introduits, afin de comparer
les différentes espèces d'endophytes.
Isolement
Six feuilles seront récoltées sur chaque arbre.
Les feuilles seront celles de la dernière poussée foliaire et
devront être exemptes de dommages d'insectes ou de traces de maladies. 16
segments de la lamelle moyenne de chaque feuille (2 mm2 chacun)
seront prélevés, à égale distance de la nervure
médiane et de la bordure, et stérilisés dans des
tubes Sterlin de 8 ml contenant une solution d'hypochlorite de
sodium (à 5 %) pendant 5 mn. Ensuite, les segments seront rincés
successivement dans 3 tubes Sterlin contenant de l'eau distillée
stérile. Les segments seront ensuite transférés (avec des
pinces stérilisées à l'alcool) dans des boîtes de
Petri contenant de l'agar (comme dans le cas des échantillons de
troncs). L'isolement sera réalisé autant que possible dans les 4
heures suivant la collecte des feuilles, avec une cellule portable.
- Echantillonnage des Trichoderma des sols.
Des Trichoderma seront isolés du sol autour
des arbres sélectionnés, ce qui donnera une indication possible
de l'origine des endophytes des troncs. Au maximum 5 échantillons de
sols seront collectés dans un rayon de deux mètres autour du
tronc de chaque cacaoyer échantillonné. Les Trichoderma
peuvent être isolés à l'aide d'un milieu spécifique
sélectionné (Papavizas & Lumsden 1982). Des
échantillons de quelques grammes de sol seront prélevés
sous la litière, stockés dans des sacs en papier et isolés
jusqu'au retour au laboratoire.
- Essais in vitro du potentiel de contrôle microbien
Une évaluation in vitro du mycoparasitisme et
de la production d'antibiotiques des cultures sera réalisée, si
nécessaire. Une des hypothèses à tester est que le genre
Trichoderma offre le meilleur potentiel de contrôle microbien
parce qu'il ne comprend pas de pathogène de la plante et parce qu'il a
déjà été montré que des Trichoderma
sont efficaces dans le contrôle des maladies pour beaucoup de
cultures.
Aucun essai in vitro n'est fiable à 100%, mais
il est essentiel d'avoir une méthode d'estimation du potentiel du
contrôle microbien. Pour les essais sur le mycoparasitisme potentiel des
espèces de Moniliophthora, ce projet utilisera le test de
pré-inoculation modifié comme décrit par Evans et
al (2003).
Une boîte de Petri (9 cm) est inoculée d'un
côté avec le pathogène-cible. Le pathogène se
développe et remplit la boîte ; une bande d'agar (env 15 x 3 mm)
contenant le mycopathogène proposé est déposée au
côté opposé, puis la boîte est mise à incuber
(une semaine à 25 °C). Des prélèvements dans l'agar
(de 3 mm de diamètre) sont effectués le long d'une ligne allant
du point d'inoculation du pathogène à celui de l'antagoniste. Les
divers prélèvements sont placés séparément
dans des boîtes de Petri contenant du milieu PDA 20% et incubées
à 25°C. Les boîtes sont ensuite examinées pour
déterminer à quelle distance (du point de
prélèvement au point d'inoculation du myco-pathogène) on
peut retrouver le myco-pathogène.
Cette procédure est répétée chaque
semaine pendant trois semaines. Le fait de retrouver le myco-pathogène
testé dans tous les prélèvements indique un
mycoparasitisme efficace.
5.2.6 Le calendrier prévisionnel
|
1.1
|
1.2
|
1.3
|
|
1.4
|
|
2.1
|
|
2.2
|
2.3
|
2.4
|
3.1
|
3.2
|
3.3
|
3.4
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Année.Trimestre
1
2
3
4
5
6
7
8
1 : Recherche, identification et caractérisation de
souches des trois pathogènes (Guyane + Montpellier).
2 : Tests-feuille Phytophthora en Guyane
3 : Tests-feuille P. megakarya à Montpellier
4 : Tests M. perniciosa en Guyane
5 : Tests M. perniciosa à Montpellier
6 : Tests C. fimbriata en Guyane
7 : Tests de résistance des cortex de cabosses
(Guyane)
8 : Analyses statistiques et interprétation de l'ensemble
des résultats.
5.3 Perspectives
A l'issue des trois années prévues pour la
durée du projet, on peut espérer les résultats suivants
:
- La sélection de nombreux clones résistants
à chacune des maladies ; - La sélection de certains clones
résistants aux trois maladies ;
- Un catalogue des espèces de champignons endophytes du
cacaoyer présentes en Guyane ;
- Les premiers résultats de tests biologiques.
Dans la mesure où ces résultats seront acquis
(même seulement en partie), il sera possible d'envisager ensuite les
travaux suivants :
- « Pré-breeding » en croisant les clones
sélectionnés à l'issue du projet et/ou déjà
décrits (descripteurs agro-morphologiques), afin de cumuler les
gènes intéressants dans de nouveaux géniteurs, utilisables
en tant que clones ou comme parents pour la création et la
sélection de variétés résistantes (y compris en
amélioration préventive).
- Tester « in planta » les
potentialités en lutte biologique des espèces ou souches
d'endophytes s'étant révélées efficaces à
l'issue des tests biologiques de laboratoire, et éventuellement tester
des formulations (associations).
- Vérifier dans le groupe des cacaoyers
spontanés de Guyane, la pertinence des marqueurs microsatellites
proposés par Santos et al. (2007) pour identifier les
génotypes résistants au balai-de-sorcière.
5.4 Références citées
- Arnold, A.E., Herre, E.A. 2003. Canopy cover and leaf age
affect colonization by tropical fungal endophytes: ecological pattern and
process in Theobroma cacao (Malvaceae). Mycologia 95:
388-398.
- Arnold, A.E., L.C. Meija, D. Kyllo, E.I. Rojas, Z. Maynard, N.
Robbins & E.A. Herre. 2003. Fungal endophytes limit pathogen damage in a
tropical tree. PNAS 100 (n°26) : 15649- 15654.
- Assemat S., Ph. Lachenaud, F. Ribeyre, F. Davrieux, J.-L.
Pradon & E. Cros, 2005. Bean quality traits and sensory evaluation of wild
Guianan cocoa populations (Theobroma cacao L.). Genetic Resources
and Crop Evolution 52: 911-917
- Bekele, F. and D. R. Butler. 2000. Proposed short list of cocoa
descriptors for characterization. In : Working procedures for cocoa germplasm
evaluation and selection. A. B. Eskes, J.M.M. Engels & R. A. Lass, edit.
2000, IPGRI (Rome, Italy), pp. 41-48.
- Bekele, F., Bidaisee, G. and Rampat, R. 1997. A preliminary
study of pod husk hardness of accessions in the International cocoa Genebank,
Trinidad. In : Annual report 1996, CRU, The University of the west Indies, St
Augustine, Trinidad and Tobago, 67-76.
- Bekele, F. L., G.G. Bidaisee and W. M. Mollineau. 2001.
Morphological characterization of germplasm from the ICG, T: highlights of
recent findings. In : Annual report 2000, CRU, The University of the west
Indies, St Augustine, Trinidad and Tobago, 16-21.
- Clay, K. 2004. Fungi and the food of the gods. Nature
(London) 427: 401-402
- Crozier J., Thomas S.E., Aime M.C., Evans H.C. and Holmes K.A.
2006. Molecular characterization of fungal endophytic morphospecies isolated
from stems and pods of Theobroma cacao. Plant Pathology, 55,
783-791.
- Delgado, J. C. & E. Echandi. 1965. Evaluación de la
resistancia de especies y clones de cacao al mal del machete provocado por
Ceratocystis fimbriata. Turrialba 15 (4): 286-289
- Delgado, R. 2007. Assessment of resistance of Nacional cocoa
accessions to Ceratocystis fimbriata in Ecuador. Ingenic
Newsletter 11 : 29-30.
- Delgado, R. A. & C. Suárez Capello. 2005.
Evaluación de la resistancia al Mal del Machete en clones
internacionales de cacao en Ecuador.
14th International Cocoa Research Conference, Accra,
Ghana, 13-18 October 2003, 1273- 1277.
- Dominguez, P. F. 1971. Avance del trabajo sobre selection de
plantas de cacao por resistencia al hongo Ceratocystis fimbriata. Rev. Fac.
Agron. VI (2), 5-29.
- Dominguez, P. F. 1976. Relacion entre vigor de las plantas y
resistencia al hongo Ceratocystis fimbriata en cacao. Rev. Fac.
Agron. Maracay IX (1) : 5-21.
- Evans, H.C., Holmes, K.A. & Thomas, S.E. 2003. Endophytes
and mycoparasites associated with an indigenous forest tree, Theobroma
gileri, in Ecuador. Mycological Progress 2: 149-160.
- Greathouse, D.C., Laetsch, W.M., Phinney, B.O. 1971. The
shoot-growth rhythm of a tropical tree, Theobroma cacao. American
Journal of Botany 58, 281-286.
- Grosclaude, G., Olivier, R, Pizzuto, J-C, Romiti, C &
Madec, S. 1988. Détection par piégeage du Ceratocystis
fimbriata f. platani. Application à l'étude de la
persistance du parasite dans du bois infecté. Forest Pathology
18 (7), 385-390.
- Holmes K.A., Schroers H.J., Thomas S.E., Evans H.C. and Samuels
G.J. 2004. Taxonomy and biocontrol potential of a new species of
Trichoderma from the Amazon basin of South America. Mycological
Progress, 3 (2), 199-210.
- IPGRI (Eskes, A. B). 2001. CFC/ICCO/IPGRI project on: Cocoa
Germplasm Utilization and Conservation : a Global Approch. Appendix to
Six-Month General Progress Report (Year 4). IPGRI, december 2001.
- Lachenaud, Ph. 2007. Fruit trait variability in wild cocoa
trees (Theobroma cacao L.) from the Camopi and Tanpok basins in French
Guiana. Acta Botanica Gallica 154 (1), 117-128.
- Lachenaud Ph. & B. Sallée, 1993. Les cacaoyers
spontanés de Guyane. Localisation, écologie et morphologie.
Café, Cacao, Thé 37 (2), 101-114.
- Lachenaud, Ph., V. Mooleedhar & C. Couturier, 1997. Wild
cocoa trees in French Guiana. New surveys. Plantations, Recherche,
Développement 4 (1), 25-32.
- Lachenaud Ph., F. Bonnot & G. Oliver, 1999. Use of floral
descriptors to study variability in wild cocoa trees (Theobroma cacao
L.) in French Guiana. Genetic Resources and Crop Evolution 46,
491-500.
- Lachenaud Ph., G. Oliver & Ph. Letourmy, 2000. Agronomic
assessment of wild cocoa trees (Theobroma cacao L.) from the Camopi
and Tanpok basins (French Guiana). Plant Genetic Resources Newsletter
124, 1-6.
- Lachenaud Ph. & G. Oliver, 2001. Variability in various
agronomic traits of wild cocoa trees (Theobroma cacao L.) from the
Camopi and Tanpok basins (French Guiana). Plant Genetic Resources
Newsletter 128, 35-40.
- Lachenaud Ph., O. Sounigo & G. Oliver, 2004. Description of
genetic structure in Guianan wild cocoa trees (Theobroma cacao L.)
using Isozyme Electrophoresis. Plant Genetic Resources Newsletter 139,
24-30.
- Lachenaud Ph. & G. Oliver, 2005. Variability and
selection for morphological bean traits in wild cacao trees (Theobroma
cacao L.) from French Guiana. Genetic Resources and Crop
Evolution 52: 225-231.
- Lachenaud Ph., D. Paulin, M. Ducamp, J.-M. Thevenin. 2007.
Twenty years of agronomic evaluatin of wild cocoa trees (Theobroma
cacao L.) from French Guiana. Scientia Horticulturae 113:
313-321.
- Lachenaud, Ph., D. Zhang. 2008. Genetic diversity and
population structure in wild stands of cacao trees (Theobroma cacao
L.) in French Guiana. Annals of Forest Science 65: 310- 316.
- N'guessan, F., A. B. Eskes, Ph. Lachenaud. 2005. Study on the
resistance of cocoa trees to mirids in Côte d'Ivoire: classification of
major groups by their level of resistance /
sensitivity. Conférence Internationale sur la Recherche
Cacaoyère. 14, 2003-10-13/2003-10- 18, Accra, Ghana, vol 1, 559-565
-N'guessan, K.F., Eskes, A.B., Lachenaud, Ph. 2007. Mirid
resistance studies in Côte d'Ivoire: field observations on recent and
cumulative damage. In: Eskes, A.B., Efron, Y. (Eds), Global Approaches to Cocoa
Germplasm Utilization and Conservation. CFC Technical Paper 50, pp. 170-181
- Papavizas, GC, Lumsden, RD. 1982. Improved medium for isolation
of Trichoderma spp. from soil. Plant Disease 66 :
1019-1020
- Paulin D., M. Ducamp, K. Vezian-Bonnemayre & A.B. Eskes,
2005.- Evaluation par le test d'inoculation sur feuille du niveau de
résistance de trente clones vis-à-vis de plusieurs espèces
de Phytophthora. Conférence Internationale sur la Recherche
Cacaoyère. 14, 2003- 10-13/2003-10-18, Accra, Ghana, vol. 1, 85-93.
- Paulin, D., Ducamp, M., Lachenaud, Ph., 2006. Evaluation des
cacaoyers spontanés de Guyane française pour leur
résistance à Phytophthora megakarya. Proceedings of the
15th International Conference on Cocoa Research, San José
(Costa Rica), 9-14/10/2006 (sous presse).
- Paulin, D., M. Ducamp, P. Lachenaud. 2008. New sources of
resistance to Phytophthora megakarya identified in wild cocoa tree
populations of French Guiana. Crop Protection 27: 1143-1147.
- Pires, J. L. 2003. Avaliação quantitativo e
molecular de germoplasma para o melhoramento do cacaueiro com enfase na
produtividade dos frutos e rêsistencia (Capitulo 3 : Rêsistencia
à Vassoura-De-Bruxa e Podriadão). Ph D Thesis, Universidad
federal de Viçosa, MG,
Brasil.
- Przybyl, K. 1988. The response of Populus "nE42" to
infection by Ceratocystis fimbriata f. platani from cacao tree and
plane. Forest pathology 18 (1), 8-12.
- Samuels, G.J., Suarez, C., Solis, K., Holmes, K.A., Thomas,
S.E., Ismaiel, A., Evans, H.C. 2006. Trichoderma theobromicola and
T. paucisporum: two new species from South America. Mycological
Research 110: 381-392.
- Santos, R.M.F., U.V. Lopes, R. C. Bahia, R.C.B. Machado, D.
Ahnert & R.X. Corrêa. 2007. Marcadores microssatélites
relacionados com a resistência à vassoura-de-bruxa do cacaueiro.
Pesq. Agropec. Bras., Brasilia, v42, n°8, 1137-42.
- Silva, S. D. V. M., M. C. Paim & W. M. Castro. 2004. Cacau
"Jaca" resistente a Ceratocystis fimbriata na região cacaueira da Bahia,
Brasil. Fitopatolol. Bras. 29 (5) : 538- 540.
- Silva, S. D. V. M., E. P. Mandarino, V. O. Damaceno & L. P.
Santos Filho. 2007. Reação de genótipos de cacaueiros a
isolados de Ceratocystis cacaofunesta. Fitopatol. Bras. 32
(6) : 505-506.
- Sounigo O., Y. Christopher & R. Umaharan, 1996. Genetic
diversity assessment of Theobroma cacao L. using iso-enzyme and RAPD
analyses. Report for 1996, 35-51. Cocoa Research Unit, The University of the
West Indies, St. Augustine, Trinidad and Tobago.
- Sounigo O., S. Ramdahin, R. Umaharan, Y. Christopher, 1999.-
Assessing cacao genetic diversity using IE and RAPD techniques. Annual Report
1998, 25-28. Cocoa Research Unit, The University of the West Indies, St.
Augustine, Trinidad and Tobago.
- Sounigo O., S. Ramdahin, Y. Christopher, 2001.- Etude de la
diversité génétique du cacaoyer à l'aide des RAPD.
13th International Conference on Cocoa Research, Kota Kinabalu (9-14/10/2000),
Sabah, Malaysia, 119-126.
- Surujdeo-Maharaj, S., Umaharan, P., Butler, D.R., Sreenivasan,
T.N. 2003. An optimized screening method for identifying levels of resistance
to Crinipellis perniciosa in cocoa (Theobroma cacao). Plant
Pathology 52:464-475.
-Taylor, J.E., Hyde, K.D. & Jones, E.B.G. 1999. Endophytic
fungi associated with the temperate palm, Trachycarpus fortunei,
within and outside its natural range. New Phytologist 142:
335-346.
- Vigouroux, A. 1979. Une méthode simple de recherche de
Ceratocystis fimbriata f. platanisur arbre en place.
Forest pathology 9 (5), 316-320.


