|
UNIVERSITE DE KISANGANI
FACULTE DES
SCIENCES
|
DEPARTEMENT D'ECOLOGIE ET GESTION DES RESSOURCES
VEGETALES
|
|
DYNAMIQUE DE LA REGENERATION DES ESPECES
ABONDANTES
DE LA FORET DE L'ITURI : Cas des
dispositifs permanents de la Reserve de
Faune à Okapis
(Province de l'Ituri, R.D. Congo)
Par
NGONGO MUSELEMU Fiston
Mémoire Présenté en vue de
l'obtention du Grade de Licencié en Sciences
Option : BIOLOGIE
Orientation : BOTANIQUE
Promoteur : Prof. Jean-Remy MAKANA
ANNEE ACADEMIQUE : 2015-2016
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
DEDICACE
i
A Ngongo Likwela et Aishe Ngongo
A Nos parents
A tous ceux qui ont contribué, contribuent et
contribueront à
éveiller nos consciences
A Grace Mave Dhedasi et à son admiration
Fiston NGONGO MUSELEMU
ii
Dynamique de la régénération
des espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
REMERCIEMENTS
Au dénouement de ce travail qui marque la fin de nos
études universitaires, nous tenons à remercier de tout coeur
l'Université de Kisangani en général et la Faculté
des Sciences en particulier pour le cadre confort nous offert. Nous pensons de
ce fait aux autorités académiques et facultaires.
Nous souhaitons ainsi présenter nos remerciements les
plus sincères au Professeur Jean-Remy Makana pour avoir proposé
cette étude et accepté de la conduire en la dirigeant de tout
coeur jusqu'à cette fin, malgré ses multiples fluctuations en
cause de l'éducation et l'encadrement dans le domaine de la gestion de
la biodiversité.
Nous louons le soutien financier du labo LECAFOR, qui nous a
permis non seulement de réaliser cette étude en nous mettant
à possession des données d'inventaires réalisés en
1997, 2001 et 2007 dans les dispositifs permanents de la Réserve de
faune à Okapis à Epulu mais aussi de maquiller ce travail de sa
forme physique.
Notre reconnaissance à l'Eternel Notre Dieu pour le
souffle de vie avec lequel nous sommes en bonne forme. Vraiment merci
Seigneur.
Nous tenons à jeter les fleurs à vous nos
parents Ngongo Litofe et Asina Tunu pour nous avoir donné la vie et fait
de nous ce que nous sommes aujourd'hui.
Nos remerciements s'adressent également aux
Professeurs, Doctorants, Chefs de Travaux et Assistants de la Faculté
des Sciences : Léopold Ndjele, Honorine Ntahobavuka, Hippolyte Nshimba,
Dieudonné Upoki, Jean-Louis Juakaly, Bénoît Dhed'a Djailo,
Réné Oleko, Augustin Banyitiya, Kahindo Muhongya, Tchatchambe Wa
Bandol'an, Pionus Katwala Gatate, Christophe Lomba Bosombo, Faustin Boyemba,
Kamabu Vasombola, Franck Bapeamoni, Felix Bola Mbele (spécialement ici
pour ses précieux conseils et remarques édifiantes), Prosper
Sabongo, John Mabay, Janvier Lisingo, Bijoux Lituka, Jacques Tchatchambe et
tous ceux dont les noms ne sont pas cités ici, pour l'enseignement et
l'encadrement scientifique à l'Université.
A toi notre très chère amie Grace Mave Dhed'asi
pour les encouragements, les énormes sacrifices et privations consentis
pendant les trois dernières années de dur labeur. A toi, nous
présentons une profonde reconnaissance et une remarquable admiration.
Dynamique de la régénération
des espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
A vous nos frères et soeurs : Alphonsine Ngongo, Eric
Ngongo, Tony Ngongo, Bushiri Ngongo, Zuena Ngongo, Vicky Ngongo, Likwela Ngongo
et oncles Dieu Mokuba et Abdallah Nura ; pour votre immense affection.
Que nos compagnons de lutte trouvent à travers ces
lignes nos encouragements ainsi que notre compassion pour eux. Vous êtes
si nombreux pour vous lister dans un tel travail ; mais néanmoins
Mitterrand Banyanga Baruti et Emma Mwinja Ntabona suffisent pour vous
représenter. A tous, nous présentons un grand merci.
Nous ne pouvons pas clore cette section sans pour autant
songer à nos amis fidèles : Steave Ngoy Luhembwe, Georges Abraham
Tchatchambe, Rajac Nyamaifofe, Gloria Oleko, Adriènne Nyere, Eunice
Ntambwe, Heritier Olmedo, Arnaud Kabongo, Cosmos Paluku Kivaya, Jasmin
Mutahinga ; pour leurs encouragements.

Fiston NGONGO MUSELEMU
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
RESUME
Les études de la dynamique de la
régénération dans les milieux forestiers tropicaux
continuent à subsister dans l'insignifiance malgré l'ultime
évolution de la science de nos jours, laissant imprécises et
méconnues les informations tant soient peu exactes sur la
régénération des espèces végétales en
milieu forestier. Ce présent travail a examiné le cas des
forêts de la Réserve de faune à Okapi située dans la
Province de l'Ituri en République Démocratique du Congo. Son
objectif était de réunir des informations fiables sur la
dynamique de la régénération des essences arborescentes
abondantes dans les dispositifs permanents de la Réserve de Faune
à Okapi de la forêt de l'Ituri.
Les résultats obtenus sur deux parcelles de 20 ha
chacune durant deux périodes d'inventaire successif (1996-2001 et
2001-2007) présentent un effectif de 54.267 individus de la
régénération de trente-sept (37) espèces abondantes
de la Réserve de Faune à Okapis ; repartis en 33.752 et 20.515
individus de tempéraments différents, respectivement à
Edoro et Lenda. Les espèces sciaphiles présentent une
densité (abondance) plus élevée dans les deux sites
d'étude et sont suivies des espèces héliophiles et des
espèces tolérantes qui montrent des densités les
faibles.
Les différents taux démographiques
quantifiés révèlent que la mortalité et le
recrutement sont faibles dans les dispositifs permanents de la forêt de
l'Ituri ; ils montrent des taux relativement bas compris entre 0,009 % à
0,42 % de mortalité et 0,006 % à 0,034 % de recrutement. Ces deux
taux sont plus élevés chez les héliophiles que chez les
autres groupes fonctionnels.
Le changement de la population de la
régénération des espèces abondantes de la
Réserve de Faune à Okapi présente une allure stable dans
l'ensemble des périodes d'inventaire. Une petite progression s'est
présentée à la première période à
Edoro.
Enfin, la densité du bois des espèces abondantes
de la Réserve de Faune à Okapis corrèle faiblement avec
les différents paramètres démographiques illustrés
dans ce travail tels que la mortalité et le recrutement diminuent avec
l'augmentation de la densité du bois.
iv
Mots clés : Dynamique,
Régénération, Forêt de l'Ituri, sciaphile,
héliophile, tolérant
v
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
ABSTRACT
The studies of regeneration dynamics in tropical forests
continue to be insignificant despite the progress scientific research, leaving
that area of study little misunderstood. The present work examines forest
regeneration in the Okapi Wildlife Reserve, located in the province of Ituri in
the Democratic Republic of the Congo. Its aim was to gather reliable
information on the dynamics of regeneration of abundant tree species in the
permanent forest plots of the Okapi Reserve in the Ituri Forest.
The results obtained on two plots of 20 hectares each for two
successive periods of inventory (1996-2001 and 2001-2007) yielded 54,267
saplings belonging tothirty-seven (37) species. There were 33,752 and 20,515
individuals in Edoro and Lenda, respectively. Shade-tolerant species had a
higher density (abundance) in both study sites and were followed by
light-demanding species,non-pioneer light-demanding species exhibited the
lowest densities.
Measured demographic parameters revealed that the mortality
and recruitment were low in the permanent plots of the Ituri Forest. Mortality
rates variedfrom0.009% to 0.42% and recruitment rates from 0.006% to 0.034%.
Both rates werehigher among light demanding species than other functional
groups.
Population change of the regeneration of abundant species of
the Reserve Okapi minimal throughout the inventory periods. A small increase
was made in the 1996-2001intervalat theEdoro site.
Finally, the wood density of abundant species of Okapi
Wildlife Reserve has no significant impact on the demographic parameters
analyzed in this work, although mortality and recruitment slightly decreased
with increasing wood density.
Keywords: Dynamics, Regeneration, Ituri
Forest, shade tolerant, light demanding, non-pioneer light demanding
vi
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
LISTE DES FIGURES
Figure 2.1. La carte de la Réserve de faune à Okapi
. 9
Figure 3.1. Abondance (individus/ha) de la
régénération des espèces selon les
tempéraments
dans les deux sites 12
Figure 3.2. Corrélation entre le taux de mortalité
de la régénération et la densité du bois des
espèces dans les deux parcelles permanentes de la Reserve de Faune
à
Okapis . 15
Figure 3.3. Corrélation entre le taux de recrutement de la
régénération et la densité du bois des
espèces dans les deux parcelles permanentes de la Reserve de Faune
à
Okapis 16
Figure 3.4. Corrélation entre le taux de changement de
population de la régénération et la densité du bois
des espèces dans les deux parcelles permanentes de la Reserve de
Faune à Okapis 17
vii
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
LISTE DES TABLEAUX
Tableau 1.1. Tempéraments des espèces prises en
étude 6
Tableau 3.1. Le taux de mortalité de la
régénération des espèces abondantes dans les
parcelles permanentes de la Reserve de Faune à Okapis selon leurs
tempéraments 13
Tableau 3.2. Le taux de recrutement de la
régénération des espèces abondantes dans les
parcelles permanentes de la Reserve de Faune à Okapis selon leurs
tempéraments 14
Tableau 3.3. Le taux de changement de la
régénération des espèces abondantes dans les
parcelles permanentes de la Reserve de Faune à Okapis selon leurs
tempéraments 15
viii
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
TABLE DES MATIERES
DEDICACE i
REMERCIEMENTS ii
RESUME iv
ABSTRACT v
LISTE DES FIGURES vi
LISTE DES TABLEAUX vii
CHAPITRE I: INTRODUCTION GENERALE 1
1.1. Cadre scientifique de l'étude 1
1.1. Problématique 2
1.2. Hypothèses 4
1.3. Objectif 4
1.4. Intérêt du travail 5
1.5. Généralités sur le
tempérament des espèces végétales 5
CHAPITRE II: MATERIEL ET METHODES 8
2.1. Site d'étude 8
2.1. Matériel 9
2.2. Méthodes 9
2.2.1. Description du dispositif expérimental 9
2.2.2. Analyse de la Dynamique 10
2.2.3. Traitement et analyse des données 11
CHAPITRE III : PRESENTATION DES RESULTATS Erreur !
Signet non défini.
3.1. Abondance des espèces dans les sites
d'étude 12
3.2. Paramètres démographiques de la dynamique
de régénération 13
3.2.1. Mortalité 13
3.2.2. Recrutement 14
3.2.3. Changement de population 15
3.3. Densité du bois et paramètres
démographiques de la dynamique de la régénération
15
CHAPITRE IV : DISCUSSION ..19
4.1. Détermination des paramètres
démographiques 19
4.1.1. Mortalité des espèces abondantes en
fonction de tempéraments 19
ix
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
4.1.2. Recrutement et tempéraments des espèces
abondantes 20
4.1.3. Le changement de la population des espèces
abondantes en fonction de
tempéraments ...20
4.2. Déterminisme de la densité du bois sur les
variables démographiques 21
CONCLUSION 22
REFERENCES BIBLIOGRAPHIQUES .23
ANNEXES
26
Dynamique de la régénération
des espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
CHAPITRE I. INTRODUCTION GENERALE
1.1. Cadre scientifique de l'étude
Les équilibres séculaires entre l'homme et la
nature semblent aujourd'hui bouleversés: le changement climatique
s'emballe sous l'influence des quantités critiques des gaz à
effet de serre, la biodiversité diminue à un taux jamais
égal auparavant, les populations des pays du sud ont de plus en plus du
mal à se nourrir et à trouver un cadre de vie adapté. Le
défi qui nous attend demain est de pouvoir garantir à 8 milliards
d'habitants de la planète, à commencer par les plus pauvres, des
conditions de vie décentes, tout en n'hypothéquant pas le
bien-être des générations futures par une exploitation des
ressources naturelles (De Wasseige et al, 2012).
Dans cette perspective, personne ne peut encore douter de
l'importance des forêts tropicales dans la résolution de ces
grandes crises écologiques et économiques. Selon le CIFOR, la
couverture forestière mondiale représente environ 4 milliard
d'hectares, soit à peu près de 30% de la surface terrestre.
Près de 56% de ces forêts sont situées dans les zones
tropicales et subtropicales. Les forêts du bassin du Congo forment le
second bloc continu des forêts tropicales humides de la planète
après le massif Amazonien. Ces forêts sont très
réputées pour leur diversité tant animale que
végétale (De Wasseige et al, 2008).
En République Démocratique du Congo, les
forêts couvrent plus de la moitié de l'ensemble du territoire
national (Forafri, 1999 in Badjoko, 2009). Elles jouent un rôle capital
dans la régulation de l'effet de serre, dans les grands
équilibres climatiques et constituent le plus grand réservoir de
la biodiversité de la planète (Forafri, 1999 in Badjoko,
2009).
Mais ces forêts ne sont pas suffisamment connues et il
n'existe généralement pas de plan de production et
d'aménagement préalable (Lokombe, 2004). D'où, il faut
étudier comment les forêts sont organisées,
c'est-à-dire, comment elles sont construites, quelle est leur
architecture et leur dynamique et quelles sont les structures et le processus
sous-jacents présents dans le mélange apparemment anarchique des
arbres et des espèces (Fournier et Sasson, 1983 in Badjoko, 2004).
2
Dynamique de la régénération
des espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
1.2. Problématique
Toutes les solutions envisagées actuellement pour la
conservation des forêts tropicales humides, comme la mise en place des
réservoirs biologiques, l'exploitation forestière durable, la
restauration des forêts dégradées ou la création des
plantations forestières, nécessitent un approfondissement des
connaissances sur l'écologie et les paramètres de la dynamique de
la régénération des espèces communes dans ces
écosystèmes (Scheil et Van Heist, 2000 ; Picard et Gourlet,
2009).
La régénération naturelle, la base du
renouvellement des peuplements forestiers, est au coeur des
préoccupations des gestionnaires forestiers. Elle se fait par le biais
des graines issues des arbres semenciers présents dans la forêt.
L'un des problèmes majeurs est que cette
régénération peut être bloquée, parfois
pendant plusieurs dizaines d'années (Vanessa, 2012). Ce blocage peut
avoir des causes multiples. En effet, divers facteurs peuvent jouer un
rôle important comme la chute des graines, qui est dépendante du
couvert forestier; la banque de graines dans le sol, qui est petite en
comparaison avec celle des espèces herbacées mais importante pour
les espèces ligneuses pérennes et souvent
représentée par des espèces pionnières, la
prédation, la disponibilité des ressources (lumière,
nutriments) et la compétition entre espèces pour les ressources
(Clark et al., 1999). La disponibilité des ressources et la
compétition entre espèces pour ces dernières paraissent
des facteurs les plus importants dans l'appréciation de la
régénération dans les milieux forestiers.
L'évaluation de l'impact des tempéraments sur la
performance des espèces végétales intéresse
également des études de la dynamique des espèces dans les
forêts tropicales en général et celles de notre
région en particulier (Mbindule, 2014 ; Omvibho, 2014).
Néanmoins, d'autres facteurs comme les
paramètres du milieu et la composition floristique du couvert de sous
étage doivent être pris en considération afin
d'améliorer la gestion forestière ; les paramètres du
milieu apparaissant comme des éléments clés dans la
régénération naturelle de la forêt (Vanessa, 2012 ;
Enzinga, 2013). En effet, d'après Heithecker et Halpern (2006)
cités par Vanessa (2012), la canopée peut influencer la
quantité et la qualité de la lumière disponible, de l'air,
de la température et de l'humidité du sol.
Les conditions d'éclairement évoluent en
fonction de la variation de la canopée de la forêt et la
régénération dépend de la capacité des
jeunes plants de survivre à l'ombre (Messier et al. 1999). Il existe des
espèces ligneuses tolérantes à l'ombre qui peuvent se
régénérer sous le
3
Dynamique de la régénération
des espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
couvert des adultes et des espèces intolérantes
à l'ombre qui ne peuvent se régénérer qu'en
condition lumineuse (trouée, chablis, etc.).
La connaissance en matière de la dynamique des
populations d'arbres est peu développée en RDC, pendant que le
pays vient de s'engager sur la voie de valorisation de ses ressources,
d'où un plan d'aménagement forestier s'avère très
important. La gestion des peuplements exige de pouvoir prédire à
long terme le renouvellement et la qualité du stock exploitable. Pour ce
faire, il est important et d'une grande nécessité de mieux cerner
les processus écologiques qui déterminent la dynamique de la
régénération des espèces (Lubini, 1982 ; Reitsma,
1988 ; Pierlot, 1996 ; Bibani & al., 1998; Jesel, 2005 ).
Les différentes études menées dans les
dispositifs permanents de la Réserve de Faune à Okapi (RFO)
montrent que les espèces appartenant à la famille desFabaceae
sont les plus abondantes et les plus dominantes (Hart, 1985 ; Makana et
Thomas, 2006; Mbusa, 2015). Parmi ces espèces on peut citer
Gilbertiodendron dewevrei, Cynometra alexandri, Julbernardia seretii,
Erythrophleum suaveolens, Albizia gummifera, etc. D'autres espèces
sont abondantes dans la région sont Strombosia pustulata,
Strombosiopsis tetrandra, Zanthoxylum gilletii, Alstonia boonei, Klainedoxa
gabonensis, etc. Parmi ces espèces, certaines sont sciaphiles et
d'autres strictement héliophiles.
Quelles stratégies ces espèces adoptent-elles pour
assurer la croissance de leurs populations?
C'est pour répondre à cette question que cette
étude se propose d'analyser la dynamique de la
régénération des espèces arborescentes dominantes
à l'échelle locale. Les 40 espèces les plus abondantes
dans les parcelles permanentes de la Réserve de Faune à Okapi
seront utilisées dans le cadre de cette étude qui sera
basée sur les données d'inventaires réalisés en
1996 et 2007.
4
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
1.3. Hypothèses
Les hypothèses suivantes ont été
formulées au regard de la question centrale posée ci-dessus:
- Le taux de mortalité de la
régénération est plus élevé pour les
espèces héliophiles que pour les sciaphiles;
- Le taux de recrutement des espèces est plus
élevée pour les espèces héliophiles que pour les
sciaphiles;
- Le taux de changement de population de la
régénération des espèces s'avère plus
élevé pour les espèces sciaphiles que pour les
héliophiles;
- Les paramètres démographiques
(mortalité, de recrutement et changement de population) sont
négativement corrélés avec la densité du bois.
1.4. Objectif
1.4.1. Objectif général
L'objectif global de cette étude est de réunir
des informations fiables sur la dynamique de la
régénération des essences arborescentes dominantes dans
les dispositifs permanents de la Réserve de Faune à Okapi dans la
forêt de l'Ituri.
1.4.2. Objectifs spécifiques
Spécifiquement, ce travail visera à :
- Evaluer et comparer le taux de mortalité de la
régénération des espèces selon leurs
tempéraments (héliophiles, tolérantes et sciaphiles).
- Analyser et comparer le taux de recrutement de la
régénération des espèces selon leurs
tempéraments (héliophiles, tolérantes et sciaphiles).
- Calculer et comparer le taux de changement de population de
la régénération des espèces selon leurs
tempéraments (héliophiles, tolérantes et sciaphiles).
- Déterminer l'impact de la densité du bois sur
les paramètres démographiques de la
régénération des différentes espèces.
5
Dynamique de la régénération
des espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
1.5. Intérêt du travail
L'étude sur la dynamique de la
régénération des espèces dominantes de la Reserve
de faune à Okapis s'inscrit dans le cadre de suivi permanent ou durable
des ressources végétales ligneuses en état de
renouvellement. Ce contexte vise à mieux comprendre les processus
primaires, qualifiés de « base » dans le renouvellement des
peuplements forestiers ; constituant une grande problématique de la
communauté internationale dans l'aménagement et gestion
intégrée pour une bonne conservation des édifices qui
renferment une biodiversité exceptionnelle en l'occurrence des
forêts classées.
1.6. Généralités sur le
tempérament des espèces végétales
Le tempérament des espèces
végétales peut être défini comme « un ensemble
des réactions de croissance et de développement
présentées par un arbre dans son environnement durant son cycle
de vie » (Oldeman et Van Dijk, 1991). Ces réactions sont
basées sur leur stratégie d'exploitation des ressources et
spécialement en lumière solaire ainsi que de leurs spectres
liés à la dispersion ou dissémination (Swaine et Whitemore
1988 ; Doucet, 2003).
En forêt tropicale humide, deux groupes (parfois trois)
sont classiquement reconnus depuis Aubréville (1938) et Van Steenis
(1956) selon la place et le rôle des espèces dans les
différentes phases de la dynamique forestière [phases
d'ouverture, de maturation et de fermeture définies par Swaine et
Whitmore (op.cit)].
Les espèces pionnières
sont présentes dans les ouvertures (de taille suffisante)
du couvert forestier d'origine naturelle ou anthropique. Ce sont les
premières espèces à coloniser les zones perturbées.
Elles peuvent être facilement reconnues dans les forêts tropicales
d'Amérique (genre Cecropia) comme dans les forêts d'Asie
(genre Macaranga) et dans les forêts d'Afrique centrale (genres
Musanga, Macaranga, Alstonia, etc.), notamment celles dont les
semences se conservent dans la banque de graines du sol (Prévost,
1983).
Un ensemble de traits biologiques relatifs à la
biologie de la reproduction, à l'installation et à la croissance
de juvéniles et d'arbres, à la physiologie et à la
morphologie de différents stades
6
Dynamique de la régénération
des espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
de développement sont associés aux
espèces pionnières (Bazzaz et Pickett, 1980 ; Turner, 2001).
Ces espèces pionnières ont une durée de
vie limitée (quelques décennies), une fructification
précoce, continue et abondante. Les graines peuvent garder leur pouvoir
germinatif pendant plusieurs mois, voire plusieurs années et constituent
une réserve abondante dans le sol. Tout le cycle de développement
d'un individu (de la graine à l'arbre mature) se déroule en
pleine lumière ; d'où l'appellation
«héliophiles» leur est attribuées. Les arbres sont
caractérisés par une croissance très rapide (Lilian et
al., 2003).
Les espèces non-pionnières
(appelées aussi
«climaciques») se définissent par
opposition aux espèces précédentes. Elles se retrouvent
dans toutes les strates de hauteur (des espèces de sous-bois aux
espèces émergentes). Une caractéristique
générale est la tolérance à l'absence de
lumière par les plantules et les jeunes arbres ; d'où
l'appellation d'espèces sciaphiles leur
attribuée (Lilian et al. Op.cit).
Un troisième groupe d'espèces est parfois
distingué. Ce dernier groupe intermédiaire est défini pour
désigner les espèces nomades à vie longue (Van Steenis,
1956), les secondaires tardives (Budowski, 1965) ou encore les géantes
anémochores. Ces espèces présentent avec les
pionnières des caractéristiques communes mais ont une
durée de vie plus longue, une croissance moins rapide et atteignent une
plus grande taille (Lilian et al. op.cit). Elles peuvent être
désignées par les termes «héliophiles
non-pionnières » ou « espèces
tolérantes».
Le tableau 1.1. ci-dessous présente la liste des
espèces l'étude, associées à leurs
tempéraments.
Tableau 1.1. : Tempéraments des espèces
considérées pour cette étude
|
No
|
Espèces
|
Tempéraments
|
|
1
|
Albizia gummifera
|
Héliophile
|
|
2
|
Angylocalyx pynaertii
|
Sciaphile
|
|
3
|
Anonidium mannii
|
Tolérante
|
|
4
|
Anthonotha macrophylla
|
Tolérante
|
|
5
|
Blighia welwitschii
|
Tolérante
|
|
6
|
Celtis mildbraedii
|
Héliophile
|
|
7
|
Chrysophyllum boukokoense
|
Tolérante
|
|
8
|
Chrysophyllum lacourtianum
|
Tolérante
|
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
7
|
9
|
Cleistanthus michelsonii
|
Tolérante
|
|
10
|
Cola lateritia
|
Tolérante
|
|
11
|
Cynometra alexandri
|
Sciaphile
|
|
12
|
Dialium corbisieri
|
Sciaphile
|
|
13
|
Dialium excelsum
|
Sciaphile
|
|
14
|
Dialium pentandrum
|
Sciaphile
|
|
15
|
Erythrophleum suaveolens
|
Tolérante
|
|
16
|
Gilbertiodendron dewevrei
|
Sciaphile
|
|
17
|
Greenwayodendron suaveolens
|
Sciaphile
|
|
18
|
Grewia obligoneura
|
Héliophile
|
|
19
|
Guarea cedrata
|
Tolérante
|
|
20
|
Hallea stipulosa
|
Héliophile
|
|
21
|
Irvingia excelsa
|
Tolérante
|
|
22
|
Irvingia grandifolia
|
Tolérante
|
|
23
|
Julbernardia seretii
|
Sciaphile
|
|
24
|
Klainedoxa gabonensis
|
Tolérante
|
|
25
|
Macaranga schweinfurthii
|
Héliophile
|
|
26
|
Manilkara zenkeri
|
Sciaphile
|
|
27
|
Parinari excelsa
|
Tolérante
|
|
28
|
Pseudospondias microcarpa
|
Tolérante
|
|
29
|
Pycnanthus angolensis
|
Héliophile
|
|
30
|
Sarcocephalus pobeguinii
|
Héliophile
|
|
31
|
Staudtia kamerunensis
|
Sciaphile
|
|
32
|
Strombosia pustulata
|
Sciaphile
|
|
33
|
Strombosiopsis tetrandra
|
Sciaphile
|
|
34
|
Treculia africana
|
Héliophile
|
|
35
|
Uapaca corbisieri
|
Héliophile
|
|
36
|
Uapaca guineensis
|
Héliophile
|
|
37
|
Zanthoxylum gilletii
|
Héliophile
|
8
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
CHAPITRE II. MATERIEL ET METHODES
2.1. Site d'étude
Les dispositifs qui ont servi de base à
l'exécution de ce travail sont ceux installés dans la
Réserve de Faune à Okapis (RFO) par le Centre de Formation et de
Recherche en Conservation Forestière (CEFRECOF) en 1994. Cette
réserve est localisée dans le Nord-est de la R.D. Congo dans
l'actuelle Province de l'Ituri située entre 1° et 2° 29' N, et
28° et 29° 4' E (Hart et Carrick, 1996 ; Makana et Thomas, 2005). La
topographie de la région est douce avec des collines à roulement
occasionnel. L'altitude varie de 600 m à l'Ouest à plus de 1000 m
à l'Est (Hart et al., 1996). La pluviométrie moyenne annuelle est
de 1700 mm, avec une saison sèche de décembre à
février au cours de laquelle les précipitations mensuelles sont
de l'ordre de < 100 mm (Hart et Carrick 1996, Makana et Thomas, 2005).
La réserve est caractérisée par deux
principaux types forestiers. Le site de Lenda, situé à 1°18'
N, 28°39' au sud du village d'Epulu est localisé dans une zone de
forêt monodominante à Gilbertiodendron dewevrei. Celui
d'Edoro est situé entre 1°34'N et 28°32'E et sa
végétation est du type forêt mixte dominée
généralement par des espèces de canopée de la
famille de Fabacées, dont Cynometra alexandri et
Julbernardia seretii.
D'autres types de végétation présentes
dans le bassin de la rivière Ituri comprennent les forêts
marécageuses le long des cours d'eau et la végétation
xérophile sur les sommets des rochers, collines, mais aussi les deux
sont beaucoup moins rependus que les deux principaux types de forêts
mentionnés (Hart et al, 1996).
9
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
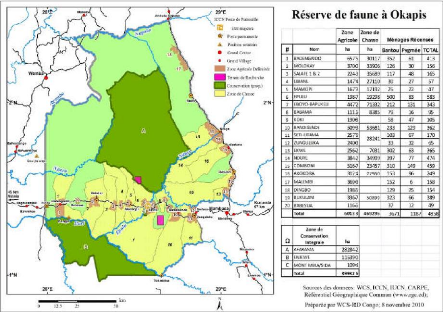
Figure 2.1. : La carte de la Réserve de
faune à Okapi
(
http://www.memoireonline.com/08/11/4776/Rapport-de-stage--la-Reserve-de-Faune--Okapis.html)
2.2. Matériel
Le matériel biologique du présent
travail est constitué des individus jeunes (plantules
et juvéniles) de 37 espèces arborescentes les
plus abondantes dans les parcelles d'étude et
inventoriées successivement en 1996, 2001 et 2007. Les plantules
sont tous individus à 1 cm = dhp < 4 cm, tandis que
les juvéniles sont des individus à 4 cm = dhp
< 10 cm.
2.3. Méthodes
2.3.1. Description du dispositif expérimental
La récolte des données a
été réalisée dans quatre parcelles rectangulaires
de 200 x 500 m de 10 ha chacune établies depuis 1996 par l'appui
technique de WCS et du center for tropical
Forest Science (CTFS) de Smithsonian
Tropical Research Institute (ST RI) (Makana et
al,
2004).
10
Dynamique de la régénération
des espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
Ces parcelles, divisées en 250 placettes de 20 m ×
20 m et subdivisées chacune en 16 sous placettes de 5 m × 5 m ;
étaient installées dans deux sites différents à
savoir Edoro et Lenda ; celles de Lenda sont orientées sur un axe
Nord-sud et distantes de 500 m chacune, par contre celles d'Edoro sont
orientées le long de la direction Ouest-est (Makana,
1999).
Un inventaire initial a été
réalisé sur la période allant de 1994-1996 et les
recensements ultérieurs étaient accomplis en 2001 et 2007.
Pendant ces recensements, les arbres du recensement précèdent ont
été remesurés, soient enregistrés comme morts. Un
arbre a été considéré comme mort si toutes ses
tiges et branches étaient soient morts ou visiblement absents (Makana,
op.cit).
Les tiges nouvellement recrutées étaient celles
qui avaient atteint le diamètre minimal (1 cm dhp) au moment du nouveau
recensement et ces tiges ont été ajoutées dans
la liste de base des données.
2.3.2. Analyse de la Dynamique ? Mortalité et
recrutement
Selon Madelaine (2009), on définit par N0 l'effectif
initial du peuplement, NS et Nt respectivement le nombre de
survivants et le nombre de recrutés après une période
d'observation t et Nf = NS+ Nt, le nombre total d'individus observés
à la fin de la période t. On pose aussi Nm= N0 -
Ns, le nombre de morts durant la période t. Sous
l'hypothèse que les processus de mortalité et de
recrutement sont continus et constants sur la période d'observation, les
taux de mortalité, X, et de recrutement, r, sont dérivés
du modèle exponentiel (Phillips et Gentry, 1994 ; Condit et al., 1995 ;
Sheil et al, 1995) tel que :
ë (%)= ln
(N0)-ln(NS)
t
r%)_ ln (NS)-ln(Nt)
(%)--
t
11
Dynamique de la régénération
des espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
? Changement de population
Taux de changement annuel (%) =
in(N5)-in(N0)
t
Ou N0 et Ns sont les nombres d'individus au
début et à la fin de la période de mensuration (Condit et
al., 1996 cités par Makana, 2004).
2.4. Traitement et analyse des données
La base de données utilisée dans la
présente étude a été conçue à partir
du logiciel Excel après les différentes campagnes de mensuration
des arbres dans les parcelles d'étude. Ce logiciel a aussi servi
à la réalisation de différentes manipulations et analyses
des données.
L'ANOVA a été utilisée pour comparer des
moyennes de taux de mortalité, de recrutement et de changement de
population des individus des 37 espèces et le test t de student a servi
à comparer d'une part les moyennes de ces composantes dans les deux
parcelles d'étude et d'autre part les densités ou abondance des
espèces dans les sites d'études. La corrélation entre la
densité du bois et les composantes de la dynamique a été
évaluée par la technique de régression.
12
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
CHAPITRE III. PRESENTATION DES RESULTATS
Le présent chapitre présente les
différents résultats obtenus au cours de cette étude qui
porte sur l'évaluation des paramètres de la dynamique de la
régénération des espèces les plus abondantes des
dispositifs permanents de la Réserve de Faune à Okapis. Il se
subdivise en trois principales sections. La première présentera
l'abondance des espèces étudiées en fonction de leurs
tempéraments ; la deuxième, constituant le coeur du travail, va
étaler les observations des composantes de la dynamique de la
régénération des espèces et la dernière
section montrera les différentes corrélations entre la
densité de bois de ces espèces et les différentes
composantes de la dynamique examinées.
3.1. Tempérament et abondance des
espèces
Les résultats d'inventaire montrent que les
espèces sciaphiles poussent abondamment dans les deux parcelles formant
les dispositifs permanents de la Réserve de faune à Okapis. Elles
présentent dans les deux parcelles d'Edoro une abondance moyenne de 100
et 92 individus par hectare respectivement dans la première (1996-2001)
et la seconde (2001-2007) période de recensement. Cette densité
diminue sensiblement dans le site de Lenda où ces espèces
présentent une abondance de 53 et 55 individus par hectare
respectivement dans la première et seconde campagne d'inventaire. Les
autres groupes d'espèces ont de plus faibles abondances dans les deux
sites (Figures 3.1)
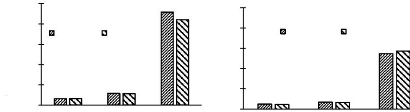
Lenda 2001 Lenda 2007
B
Edoro 2001 Edoro 2007
A
0
Abondance (indiv.ha-1)
100
80
60
40
20
100
80
60
40
20
0
Heliophile Tolerant Sciaphile Heliophile Tolerant
Sciaphile
Temperament Temperament
Figure 3.1. Abondance (individus/ha) de la
régénération des espèces selon les
tempéraments dans les deux sites (A pour Edoro et B pour Lenda)
La figure 3.1 montre la répartition des valeurs des
moyennes de l'abondance de la régénération des
espèces dans les parcelles permanentes de la Réserve de Faune
à Okapis.
13
Dynamique de la régénération
des espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
Les espèces sciaphiles sont les plus abondantes tant
à Edoro qu'à Lenda et pour la première comme pour la
seconde période d'inventaire. Les analyses montrent qu'il y a une
différence significative entre les tempéraments à Edoro le
second intervalle (F = 3,37et p = 0,045) et à Lenda les deux intervalles
(F = 3,44 ; p = 0,043 et F = 3,26 ; p = 0,049, respectivement).
3.2. Paramètres démographiques de la
dynamique de régénération
Les divers constituants de la dynamique de la
régénération des espèces de la Réserve de
faune à Okapi sont présentés dans cette sous-section. Les
détails pour chaque espèce sont mis dans les annexes.
3.2.1. Mortalité
Le tableau 3.1 présente les taux de mortalité de
la régénération des espèces selon leurs
tempéraments dans les dispositifs permanents de la Réserve de
Faune à Okapis.
Tableau 3.1. Taux de mortalité de la
régénération des espèces dans les parcelles
permanentes de la Réserve de Faune à Okapis selon leurs
tempéraments
La mortalité élevée se manifeste chez les
jeunes héliophiles dans les deux sites. Les espèces
tolérantes viennent en deuxième position et les sciaphiles
connaissent une mortalité faible dans ces deux sites. En appliquant le
test statistique, ces valeurs montrent une différence significative
entre les tempéraments pour le site de Lenda (F = 6,071 et p = 0,013
pour la première période et F = 4,581 et p = 0,0278 pour la
seconde période), tandis que pour Edoro la différence est
marginale (F = 2,764 et p = 0,095 pour la première période et F =
3,573 et p = 0,055 pour la seconde période)
Par contre, le taux de mortalité n'a pas
été significativement différent entre les deux sites au
cours de deux intervalles de temps de mensuration (t = -0,515 ; p = 0,63 pour
la première période et t = -0,23 p = 0,83pour la seconde
période)
14
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
3.2.2. Recrutement
Le tableau qui suit renseigne sur les taux de recrutement des
espèces arborescentes communes en fonction de leurs tempéraments
dans les deux sites de la Réserve de Faune à Okapi.
Tableau 3.2. Le taux de recrutement de la
régénération des espèces abondantes dans les
parcelles permanentes de la Réserve de Faune à Okapis selon leurs
tempéraments
Sites et Périodes
|
Tempéraments
|
Edoro
1996-2001 2001-2007
|
Lenda
1996-2001 2001-2007
|
|
Héliophiles
|
0,025 #177; 0,021
|
0,034 #177; 0,031
|
0,019 #177; 0,013
|
0,029 #177; 0,032
|
|
Sciaphiles
|
0,011 #177; 0,008
|
0,015 #177; 0,010
|
0,010 #177; 0,006
|
0,012 #177; 0,008
|
|
Tolérantes
|
0,027 #177; 0,024
|
0,026 #177; 0,018
|
0,006 #177; 0,006
|
0,014 #177; 0,013
|
|
F
|
3,907
|
3,455
|
4,493
|
1,932
|
|
P-value
|
0,044
|
0,057
|
0,025
|
0,178
|
On note des variations importantes dans les taux de
recrutement entre les différents tempéraments (Tableau 3.2.). La
différence est significative à la première période
dans tous les sites (0,044 et 0,025 respectivement à Edoro et Lenda).
Cependant, il n'y a pas de différence de recrutement entre les deux
sites pendant les deux intervalles d'inventaires (t = 1,47 ; p-value = 0,21
pour la première période et t = 0,87 p-value = 0,43 pour la
seconde)
15
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
3.2.3. Changement de population
Le tableau suivant renseigne sur les taux de changement de
population des espèces arborescentes abondantes en fonction de leurs
tempéraments dans les deux sites de la Réserve de Faune à
Okapi.
Tableau 3.3. Taux de changement de population de la
régénération des espèces les plus communes dans les
parcelles permanentes de la Réserve de Faune à Okapis
Sites et Périodes
|
Tempéraments
|
Edoro
1996-2001 2001-2007
|
Lenda
1996-2001 2001-2007
|
|
Héliophiles
|
0,001
|
#177; 0,026
|
-0,004 #177; 0,032
|
-0,015 #177; 0,023
|
-0,015
|
#177; 0,049
|
|
Sciaphiles
|
0,001
|
#177; 0,013
|
0,001 #177; 0,014
|
0,001 #177; 0,008
|
-0,002
|
#177; 0,010
|
|
Tolérantes
|
0,013
|
#177; 0,028
|
0,006 #177; 0,023
|
-0,012 #177; 0,0015-0,009
|
#177; 0,017
|
|
F
|
|
0,66
|
0,354
|
5,072
|
|
1,095
|
|
P-value
|
|
0,53
|
0,707
|
0,02
|
|
0,36
|
L'analyse du tableau ci-dessus montre de différences
entre les taux de changement de population des espèces pour les
différents tempéraments. Cependant, cette différence n'est
significative que pour le premier intervalle de mensuration (p = 0,02) dans le
site de Lenda.
3.3. Densité du bois et paramètres
démographique de la dynamique de la
régénération
Les figures suivantes présentent les rapports entre les
différents taux des composantes de la dynamique de la
régénération et la densité du bois des
espèces étudiées dans la Reserve de Faune à
Okapis.
16
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
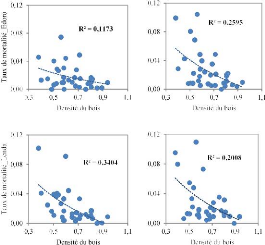
A
A
B
B
Figure 3.2. Corrélation entre le taux de
mortalité de la régénération et la densité
du bois des espèces dans les deux parcelles permanentes de la Reserve de
Faune à Okapis (A : 1996-2001 et B : 2001-2007)
On observe d'après la figure précédente
que la densité du bois a une influence sur la mortalité de
l'ensemble de la régénération dans les deux sites
d'étude. Il y a une relation inverse entre la densité de bois et
la mortalité ; la mortalité diminue avec l'augmentation de la
densité de bois.
La figure 3.3 montre que le taux de recrutement diminue aussi
avec l'augmentation de la densité de bois. Cette relation est
relativement plus forte à Lenda qu'à Edoro car les coefficients
de détermination sont plus élevés pour le site de
Lenda.
17
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
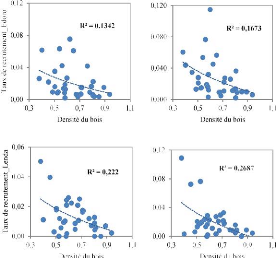
A
A
B
B
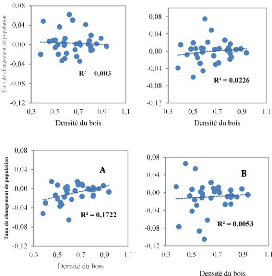
Figure 3.4. Corrélation entre le taux de
recrutement de la régénération et la densité du
bois des espèces dans les deux parcelles permanentes de la Reserve de
Faune à Okapis (A : 1996-2001 et B : 2001-2007)
A
B
Figure 3.5. Corrélation entre le taux de changement de
population de la régénération et la densité du bois
(A : 1996-2001 et B : 2001-2007)
18
Dynamique de la régénération
des espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
Contrairement aux taux de mortalité et de recrutement,
le taux de changement de population montre une relation positive avec la
densité de bois. Les espèces de haute densité de bois
montrent de populations de juvéniles ayant une croissance plus
élevée que celles à faible densité de bois. Mais
cette relation est très faible.
19
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
CHAPITE IV. DISCUSSION
Le présent chapitre se focalise sur la confrontation
des observations faites au cours de ce travail et celles des autres auteurs
ayant travaillé sur cet aspect dans la Reserve de Faune à Okapis
et dans les autres sites.
Les données d'inventaire indiquent que dans l'ensemble
le site d'Edoro a plus de régénération que celui de Lenda
(33.752 individus contre 20.515). Ces résultats sont conformes à
ceux d'autres analyses faites dans l'Ituri et qui indiquent que les
forêts mixtes d'Edoro ont une densité d'arbres plus
élevée que la foret monodominante de Lenda (Makana, 1999 ; Makana
et al., 2004).
4.1. Détermination des paramètres
démographiques
4.1.1. Mortalité des espèces abondantes en
fonction de tempéraments
Sur l'ensemble de la période de suivi, le taux de
mortalité de tous les individus de toutes espèces confondues,
varie de 0,009% à 0,042%. Ce taux est plus élevé chez les
héliophiles dans tous les deux sites avec 0,024 #177; 0,021% et 0,038
#177; 0,032% à Edoro respectivement en première et seconde
période et 0,034 #177; 0,029% et 0,044 #177; 0,042% à Lenda
respectivement en premier et second épisode. Le taux le plus bas est
observé chez les espèces sciaphiles pour la période
2001-2007.La première hypothèse de ce travail selon laquelle le
taux de mortalité est plus élevé pour les espèces
héliophiles que pour les sciaphiles a donc été
confirmée, bien que cette différence n'est pas
nécessairement statistiquement significative.
Les valeurs observées dans cette étude sont
relativement plus faibles que celles dégagées par Omvibho (2014)
dans le dispositif d'Edoro (moyenne de 1,24#177;0,18%). Le taux de
mortalité de la deuxième période est toujours
estimé supérieur à celui de la première ; cette
observation est autant faite dans la forêt sempervirente des Ghâts
occidentaux de l'Inde (dispositif d'étude permanent d'Uppangala)
(Madeleine, 2009). Madeleine trouve un taux de mortalité moyen de 0,82%
; variant de 0,65% à 0,93% pour une période 18 ans. Cette
particularité des forêts naturelles des Ghâts a
déjà été appuyée par Pélissier
(1995). Sogbossi (2012) présente un taux annuel de mortalité des
arbres dans la forêt de la Lama au Bénin se situant entre 0,75% et
1,15%, soit une moyenne d'environ 1% pour l'ensemble de la forêt dense.
Cette mortalité est trop élevée chez les jeunes essences
héliophiles qui souffrent d'un manque de lumière
20
Dynamique de la régénération
des espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
nécessaire à leur croissance et à leur
développement, donc de ce fait plus sujettes à la
mortalité naturelle.
Des taux de mortalité relativement
écartés de ceux du présent travail (0,98%) sont
observés en Guyane Française (Delcamp 2007, Madelaine et al.
2007) mais ces taux sont inférieurs à ceux estimés
à 1,48% à Pasoh en Malaisie et 2,03% à Barro Colorado
Island au Panama (Condit et al. 1999) ; 2,03% à La Selva au Costa Rica
(Lieberman et al. 1985) et 1,77% à Kade au Ghana (Swaine et al., 1987).
Condit et al. (2006) trouvent des taux de mortalité compris entre 1,04%
(Pasoh, Malaisie) et 17,06% (Mudumalai, forêt sèche du sud de
l'Inde) pour les juvéniles de moins de 10 cm de dbh.
4.1.2. Recrutement et tempéraments des
espèces abondantes
Le taux de recrutement varie fortement de 0,006% chez les
espèces sciaphiles à 0,034% aux héliophiles. Ainsi, la
deuxième hypothèse selon laquelle le recrutement est
élevé chez les espèces héliophiles que les autres
groupes, est positivement vérifiée.
Le taux de recrutement le plus élevé est
observé pour la première période de suivi (0,034 #177;
0,031% à Edoro et 0,029 #177; 0,032% à Lenda) et est
significativement différent des taux de recrutement estimés pour
la seconde période.
Le taux de recrutement estimé à la Reserve de
Faune à Okapi est loin du taux estimé à la Piste de
Saint-Elie en Guyane Française, soit 0,81% (Madelaine et al. 2007), mais
inférieur aux taux estimés à 2,83 à Pasoh en
Malaisie et 2,62 à Barro Colorado Island au Panama (Condit et al. 1999).
Ces estimations placent la forêt d'Epulu parmi les peuplements les moins
dynamiques, caractérisés à la fois par une faible
mortalité et un faible recrutement.
4.1.3. Le changement de la population des espèces
abondantes en fonction de tempéraments
Le changement de population de la
régénération des espèces les plus abondantes de la
Reserve de faune à Okapis varie selon le tempérament, bien que
ces variations ne soient pas statistiquement significatives. La même
chose a été observée dans le site d'Edoro et Lenda, au
cours de deux études menées séparément à
Epulu (Mbindule, 2014 et Omvibho, 2014). Ces analyses vont de pair avec les
résultats de l'étude de Poorter et Werger (1999).
21
Dynamique de la régénération
des espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
Nonobstant cet aspect global, la différence remarquable
de taux de changement se signale chez les espèces de la
régénération du dispositif de Lenda durant la
période de 1996-2001 (p-value < 0,05) et généralement
les espèces tolérantes ont un taux élevé de
changement de sa population, suivi des sciaphiles durant les deux intervalles
différents d'inventaire des plantes dans la Reserve de Faune à
Okapi. Ce constant nous pousse à accepter notre troisième
hypothèse stipulant que la régénération les
espèces sciaphiles présente un taux élevé de
changement de population que les héliophiles.
4.2. Déterminisme de la densité du bois
sur les variables démographiques
L'un des critères d'une grande importance et
très essentiel dans les études de la dynamique des peuplements
forestiers est la densité du bois (Makana, 2004; Madeleine, 2009). Elle
renvoie le plus souvent à des modèles des corrélations
étroites aux caractéristiques fonctionnels des espèces
végétales; telle que la texture et aux variations de l'abondance
des espèces (
www.inra.fr).
Les différentes corrélations entre de la
densité du bois et les différentes variables
démographiques ont prouvé une relation faible mais constante
entre elles. Globalement les espèces à bois léger
(généralement les héliophiles) ont des taux de
mortalité et de recrutement plus élevés que celles
à bois plus lourd (Makana, 2004).On peut donc dire que la
quatrième hypothèse selon laquelle les taux de mortalité
et de recrutement sont négativement associés à la
densité de bois a aussi été confirmée.
Contrairement aux constats faits sur ce travail, Omvibho
(2014) a démontré une corrélation positive (R2 > 0,5)
pour les espèces confondues d'arbres dans le dispositif permanent
d'Edoro et Kavali (2013) a trouvé que la densité du bois
influence significativement la relation hauteur-diamètre, les
espèces à bois dense étant plus capables d'atteindre des
hauteurs plus importantes que celles à bois moins dense
22
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
CONCLUSION
Ce travail a porté sur la dynamique de la
régénération des espèces arborescentes les plus
abondantes des dispositifs permanents de la Reserve de Faune à Okapis.
Un effectif de 54.267 individus de la régénération de 36
espèces a été dénombré, dont 33.752
individus à Edoro et 20.515 individus à Lenda.
Le taux de mortalité est nettement plus
élevé chez les espèces héliophiles que chez les
autres groupes fonctionnels. Le taux de recrutement est beaucoup plus faible
pour les sciaphiles que pour les deux autres groupes. Le taux de changement de
population montre une plus faible variation entre les tempéraments, sauf
pour les sciaphiles dans le site de Lenda où il est constamment plus
élevé que chez les autres groupes.
De façon générale, les espèces de
la forêt de l'Ituri ont montré moins de dynamisme que celles des
autres sites tropicaux, exhibant des taux de mortalité et de recrutement
plus faibles que dans plusieurs autres sites.
La densité du bois a montré une relation
négative avec les variables démographiques, de sorte que tant le
taux de mortalité que celui de recrutement ont diminué avec
l'augmentation de la densité de bois.
Le présent travail, comme toute autre étude
scientifique, présente des limites et d'autres études
évaluant d'autres aspects de la dynamique de la
régénération, associées aux études
écologiques poussées sont à envisager dans ces dispositifs
de suivi permanent des essences botaniques, en vue cerner au mieux l'importance
et la mise en valeur des ressources végétales de la Reserve de
Faune à Okapis.
26
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
REFERENCES BIBLIOGRAPHIQUES
Aubréville A. 1938. La Forêt coloniale : les
forêts de l'Afrique occidentale française. Annales des Sciences
coloniales, pp. 1-9.
Badjoko, D.H., 2009. Etudes de la structure des
émergents et dominants dans le bloc sud du dispositif de la
réserve forestière de Yoko. Mémoire inédit. Fac.
Sc. UNIKIS, 58p.
Bazzaz F., Pickett S. 1980. Physiological ecology of tropical
succession: a comparative review. Annual Review of Ecology and Systematics,
vol. 11, pp. 287-310.
Bibani, M. R., Jonkers, W.B.J. et Essama, E. J., 1998.
Phénologie de 86essences productrices de bois d'oeuvre de la forêt
dense humide sempervirente du sud-Cameroun. Résultats
préliminaires, Séminaire Forafri, Libreville-Gabon, p16.
Budowski, G. 1965. Distribution of tropical american rain
forest species in the light of successional processes. Turrialba, vol. 15, pp.
40-42.
Clark J. S. Beckage B. Camill P. Cleveland B. Hillerislambert
J. Lichetr J. Mclachlan J. Mohan J. Wyckoff P. (1999) Interpreting
recruitment limitation in forests. American Journal of Botany 86 (1):
1-16.
Condit, R., S. P. Hubbell, and R. B. Foster. 1995. Mortality
rates of 205 neotropical tree and shrub species and the impact of a severe
drought. Ecological Monographs 65:419-439.
Condit, R., P. S. Ashton, N. Manokaran, J. V. La Frankie, S.
P. Hubbell, and R. B. Foster.1999. Dynamics of the forest communities at Pasoh
and Barro Colorado: comparing two 50-ha plots. Philosophical transactions -
Royal Society of London. Biological Sciences. 354:1739-1748.
Condit, R., P. S. Ashton, S. Bunyavejchewin, H. S. Dattaraja,
S. Davies, S. Esufali, C. Ewango, R. Foster, I. A. U. N. Gunatilleke, C. V. S.
Gunatilleke, P. Hall, K. E. Harms, T. Hart, C. Hernandez, S. P. Hubbell, A.
Itoh, S. Kiratiprayoon, J. LaFrankie, S. L. de Lao, J.-R. Makana, M. Nur
Supardi Noor, A. Rahman Kassim, S. Russo, R. Sukumar, C. Samper, H. S. Suresh,
S. Tan, S. Thomas,
24
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
R. Valencia, M. Vallejo, G. Villa, and Z. T. 2006. The importance
of demographic niches to tree diversity. Science 313:98-101.
De wasseige C., de Marcken P., Bayol N., Hiol Hiol F. et
Mayaux Ph., Desclée B., Nasi R., Billan A., Defourny P. et Eba'a Atyi
R., 2012. Les forêts du Bassin du Congo. Etat des forêts. 2010.
Delcamp, M. 2007. Groupes "fonctionnels" d'espèces et
prédiction de la dynamique des peuplements d'arbres après
perturbation en forêt dense tropical humide : exemple en Guyane
française. Ph. D. Montpellier II.
Dupuy, B., L. Durrieu de Madron, Y. Petrucci. 1998.
Sylviculture des peuplements naturels en forêt dense humide africaine:
acquis et recommandations. Bois et Forêts des Tropiques 257 : 5-21.
Enzinga, Y., 2013. Structure et Biomasse de deux types
forestiers en Province Orientale (R.D. Congo). Mémoire inédit,
Fac. Sc., UNIKIS, 30p.
Ewango, C- E.N., 2010. The liana assemblage of a Congolian
rainforest. Diversity, Structure and Dynamics. Thesis, 161p.
Hart, T.B., 1985. The ecology of asingle-species-dominant
forest and of a mixed forest in Zaire, Africa. Ph.D. Thesis. Michigan State
University, USA.
Hart, J. A. and P. Carrick. 1996. Climate of the
Réserve de Faune à Okapis: Rainfall and Temperature in the Epulu
Sector, 1986-1995. CEFRECOF Working Paper n02.
Hart, T. B., J. A. Hart, R. Dechamps, M. Fournier and M.
Ataholo. 1996. Changes inforest composition over the last 4000 years in the
lturi basin, Zaire. ln: van der Maesen L.1.G. et al. (eds), The Diversity of
African Plants. Kluwer AcademicPublishers, the Netherlands, pp. 545-563.
Heithecker TD, Halpern CB (2006) Variation in microclimate
associated with dispersed-retention harvests in coniferous forests of western
Washington. Forest Ecology and Management 226:60-71.
25
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
Kavali, T. , 2013. Analyse du rapport hauteur-diamètre
des essences forestières dans la partie orientale du Bassin du Congo
(TFE) Cas des forêts de l'Ituri et des environs de Kisangani,
Mémoire, inédit, Fac. Sc , 38p.
Lieberman, D., M. Lieberman, R. Peralta, and G. S. Hartshorn.
1985. Mortality patterns and stand turnover rates in a wet tropical forest in
Costa Rica. Journal of Ecology 73:915- 924.
Lilian, B., Olivier, F., Molino, JF., Gourley-Fleury et
Sabatier. 2003. Diversité spécifique et regroupement
d'espèces arborescentes en forêt Guyanaise. Revu. For. Fr. LV.
131-146
Lokombe D., 2004. Caractéristiques
dendrométriques et stratégies d'aménagement de la
forêt dense humide à Gilbertiodendron dewevrei en
région de Bengamisa, Thèse inédite, IFA, 223p
Lubini, A. 1982. Végétation messicole et
postculturale des Sous-Régions de Kisangani et de la Tshopo
(Haut-Zaïre). Thèse de doctorat Université de Kisangani,
489p.
Madelaine, A-C., 2009. Dynamique des peuplements forestiers
tropicaux hétérogènes : variabilité inter et
intraspécifique de la croissance des arbres et trajectoires de
développement en forêt dense humide sempervirente, dans les
Ghâts occidentaux de l'Inde, thèse de doctorat, Université
Montpellier 2 - Montpellier SUPAGRO, école doctorale SIBAGHE
(Systèmes Intégrés en Biologie, Agronomie,
Géosciences, Hydrosciences, Environnement) ,202p.
Madelaine, C., R. Pélissier, G. Vincent, J.-F. Molino,
D. Sabatier, M.-F. Prévost, and C. de Namur. 2007. Mortality and
recruitment in a lowland tropical rain forest of French Guiana: effects of soil
type and species guild. Journal of Tropical Ecology 23:277- 287.
Makana, J.-R.,1999; Forest structure, Species Diversity and
Spatial Patterns of Trees in Monodominant and Mixed Stands in the Ituri Forest,
Democratic Republic of Congo. 128pages.
Makana, J.-R. 2004. Ecology and sustainable of African
mahoganies and selected other timber species in northeastern Congo Basin,
Democratic Republic of Congo.
26
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
Makana, J.-R. & S.C. Thomas 2005. Effects of light-gaps
and litter removal on the seedling performance of six African timber species.
Biotropica 37(2): 227-237.
Makana, J.R., and Thomas, S.C. 2006. Impacts of selective
logging and agricultural clearing on forest structure, floristic composition
and diversity, and timber tree regeneration in the Ituri Forest, Democratic
Republic of Congo. Biodiversity and Conservation 15(4): 1375-1397.
Mbindule, M. 2014. Dynamique des populations des
espèces arborescentes (arbres et arbustives de la forêt primaire
de l'Ituri (Province Orientale, R.D. Congo). Mémoire inédit. Fac.
Sc. UNIKIS. 30 p
Messier C. Doucet R. Ruel J C. Claveau Y. Kelly C. Lechowicz M
J. (1999) Functional ecology of advance regeneration in relation to light
in boreal forests. Can. J. For. Res. 29: 812-823.
Mbusa, M., 2015. Analyse de la composition floristique et de
la diversité des forêts monodominante et mixte de l'Ituri. Cas des
parcelles permanentes de la Reserve de Faune à Okapis (Province
Orientale, RD Congo). Mémoire inédit. Fac Sc. Unikis. 28 p.
Oldeman R.A.A., Van Dijk J. 1991. Diagnosis of the temperament of
tropical rain forest trees. pp. 21-65.
Omvibho, A. 2014. Dynamique des populations des espèces
arborescentes (Arbres et arbustives de la forêt primaire de l'Ituri
(Province Orientale, R.D. Congo). Mémoire inédit. Fac. Sc.
UNIKIS. 28 p
Pélissier, R. 1995. Relations entre
l'hétérogénéité spatiale et la dynamique de
renouvellement d'une forêt dense humide sempervirente (Forêt
d'Uppangala - Ghâts occidentaux de l'Inde.). Ph.D. Thesis. University of
Lyon, France.
Phillips, O. L., and A. H. Gentry. 1994. Increasing turnover
through time in tropical forests. Science 263:954-958.
Picard N. & Gourlet-Fleury S., 2008. Manuel de
référence pour l'installation de dispositifs permanents en
forêt de production dans le Bassin du Congo. CIRAD,
27
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
Département Environnements et Sociétés,
UPR Dynamique des forêts naturelles, Montpellier, France, 265 p.
Pierlot., R. 1966. Structure et composition des forêts
denses d'Afrique centrale, spécialement celles du Kivu. Ac. Roy. Sc.
Outre-Mer, CI. Sc. Nat. & Méd., 16 : 120-130.
Poorter, L., and M. J. A. Werger. 1999. Light environment,
sapling architecture, and leaf display in six rain forest tree species.
American Journal of Botany 86:14641473.
Prévost, M. F., and H. Puig. 1981. Accroissement
diamétral des arbres en Guyane : observation sur quelques arbres de
forêt primaire et de forêt secondaire. Bulletin du Muséum
National d'Histoire Naturelle 3:147-171.
Prevost, M.-F., 1983, Les fruits et les graines des
espèces végétales pionnières de Guyane
française, Revue d'écologie - La Terre et La Vie, 38(2) :
82-92p
Reitsma, J.M. 1988. Végétation forestière du
Gabon. Tropenbos Technical Series 1, Wageningen, The Netherlands, 142
p.
Sara Bastien-Henri, 2008. Biomasse aérienne en
plantations d'arbres tropicaux, Mémoire de maîtrise,
Université du Québec à Montréal, 51p
Sebastien, J., 2005. Écologie et dynamique de la
régénération de Dicorynia guianensis
(Caesalpiniaceae) dans une forêt guyanaise. Thèse de
doctorat. Institut national agronomique de Paris-Grignon288p.
Sheil, D., Burslem, D. F. R. and Aider, D. 1995. The
interpretation and misinterpretation of mortality rate measures. Journal of
Ecology 83: 331-333.
Swaine, M.D., D. Lieberman, and F.E. Putz. 1987. The dynamics
of tree populations in tropical forest: a review. Journal of Tropical Ecology;
Page 359-361.
Sogbossi CM, 2012. Structure et dynamique des peuplements
forestiers naturels du Sud-Benin. Résumé de Mémoire soumis
au 12è congrès forestier mondial. Québec city.
Canada. 14 p
Turner I.M., 2001. The ecology of trees in the tropical rain
forest. Cambridge : Cambridge University Press, 298 p.
28
Dynamique de la régénération des
espèces abondantes des forêts de l'Ituri : Cas des dispositifs
permanents de
la Réserve de faune à
Okapi
(c)Fiston Ngongo
Van Steenis C.G.J., 1956. Rejuvenation as a factor for judging
the status of vegetation types - the biological nomad theory. pp. 212-215. In :
Symposium on humid tropics vegetation / UNESCO Ed. Paris : UNESCO,
Vanessa, V., 2012. Dynamique de la flore et de la
régénération ligneuse après contrôle de la
végétation en forêt vosgienne. Rapport de stage.
Département d'Ecologie, Biodiversité et Evolution. EBE. Paris. 55
p
29
Annexes
ANNEXES
Annexe 1 : Les différents paramètres de
la dynamique de la régénération des espèces
abondantes dans la parcelle d'Edoro
|
Espèce
|
Tempérament
|
1996 - 2001
|
2001 - 2007
|
1996 - 2001
|
2001-2007
|
|
N0
|
Nt
|
N1
|
t
|
N0
|
Nt
|
N1
|
t
|
Mortalité
|
Recrutement
|
Chang_Po
|
Mortalité
|
Recrutement
|
Chang_Po
|
|
ALBIGU
|
Héliophile
|
119
|
92
|
115
|
6,37
|
115
|
74
|
83
|
5,47
|
0,040
|
0,035
|
-0,005
|
0,081
|
0,021
|
-0,060
|
|
ANTHMA
|
Héliophile
|
769
|
696
|
723
|
6,04
|
723
|
651
|
684
|
5,33
|
0,017
|
0,006
|
-0,010
|
0,020
|
0,009
|
-0,010
|
|
CELTMI
|
Héliophile
|
99
|
99
|
107
|
6,10
|
107
|
104
|
111
|
5,43
|
0,000
|
0,013
|
0,013
|
0,005
|
0,012
|
0,007
|
|
COLALA
|
Héliophile
|
353
|
340
|
375
|
6,15
|
375
|
358
|
417
|
5,44
|
0,006
|
0,016
|
0,010
|
0,009
|
0,028
|
0,020
|
|
ERYTSU
|
Héliophile
|
93
|
77
|
78
|
6,20
|
78
|
60
|
61
|
5,42
|
0,030
|
0,002
|
-0,028
|
0,048
|
0,003
|
-0,045
|
|
GREWOL
|
Héliophile
|
1039
|
937
|
1051
|
6,16
|
1051
|
903
|
1074
|
5,44
|
0,017
|
0,019
|
0,002
|
0,028
|
0,032
|
0,004
|
|
HALLST
|
Héliophile
|
24
|
22
|
25
|
5,87
|
25
|
20
|
24
|
5,26
|
0,015
|
0,022
|
0,007
|
0,042
|
0,035
|
-0,008
|
|
KLAIGA
|
Héliophile
|
160
|
151
|
157
|
6,14
|
157
|
149
|
154
|
5,43
|
0,009
|
0,006
|
-0,003
|
0,010
|
0,006
|
-0,004
|
|
MACASC
|
Héliophile
|
60
|
45
|
53
|
6,28
|
53
|
31
|
43
|
5,41
|
0,046
|
0,026
|
-0,020
|
0,099
|
0,061
|
-0,039
|
|
PYCNAN
|
Héliophile
|
25
|
23
|
34
|
6,35
|
34
|
29
|
37
|
5,55
|
0,013
|
0,062
|
0,048
|
0,029
|
0,044
|
0,015
|
|
SARCPO
|
Héliophile
|
60
|
50
|
53
|
6,31
|
53
|
47
|
51
|
5,42
|
0,029
|
0,009
|
-0,020
|
0,022
|
0,015
|
-0,007
|
|
TRECAF
|
Héliophile
|
66
|
66
|
70
|
6,32
|
70
|
67
|
72
|
5,47
|
0,000
|
0,009
|
0,009
|
0,008
|
0,013
|
0,005
|
|
UAPACO
|
Héliophile
|
39
|
36
|
57
|
6,11
|
57
|
50
|
64
|
5,46
|
0,013
|
0,075
|
0,062
|
0,024
|
0,045
|
0,021
|
|
UAPAGU
|
Héliophile
|
232
|
145
|
189
|
6,30
|
189
|
106
|
162
|
5,53
|
0,075
|
0,042
|
-0,033
|
0,104
|
0,077
|
-0,028
|
|
ZANTGI
|
Héliophile
|
29
|
22
|
26
|
6,24
|
26
|
21
|
39
|
5,39
|
0,044
|
0,027
|
-0,017
|
0,040
|
0,115
|
0,075
|
|
ANGYPY
|
Sciaphile
|
642
|
624
|
648
|
6,12
|
648
|
631
|
663
|
5,40
|
0,005
|
0,006
|
0,002
|
0,005
|
0,009
|
0,004
|
|
ANONMA
|
Sciaphile
|
545
|
535
|
637
|
6,15
|
637
|
617
|
722
|
5,47
|
0,003
|
0,028
|
0,025
|
0,006
|
0,029
|
0,023
|
|
CYNOAL
|
Sciaphile
|
3302
|
3020
|
3092
|
6,19
|
3092
|
2742
|
2879
|
5,46
|
0,014
|
0,004
|
-0,011
|
0,022
|
0,009
|
-0,013
|
|
DIALCO
|
Sciaphile
|
338
|
311
|
323
|
6,14
|
323
|
293
|
315
|
5,41
|
0,014
|
0,006
|
-0,007
|
0,018
|
0,013
|
-0,005
|
|
DIALEX
|
Sciaphile
|
123
|
116
|
120
|
6,15
|
120
|
113
|
121
|
5,42
|
0,010
|
0,006
|
-0,004
|
0,011
|
0,013
|
0,002
|
|
DIALPE
|
Sciaphile
|
616
|
556
|
573
|
6,17
|
573
|
513
|
519
|
5,44
|
0,017
|
0,005
|
-0,012
|
0,020
|
0,002
|
-0,018
|
30
Dynamique de la
régénération des espèces abondantes des
forêts de l'Ituri : Cas des dispositifs permanents de la Réserve
de faune à Okapi
(c)Fiston Ngongo
|
GILBDE
|
Sciaphile
|
736
|
721
|
755
|
6,85
|
755
|
733
|
779
|
5,60
|
0,003
|
0,007
|
0,004
|
0,005
|
0,011
|
0,006
|
|
GREESU
|
Sciaphile
|
2832
|
2674
|
2908
|
6,10
|
2908
|
2731
|
3026
|
5,41
|
0,009
|
0,014
|
0,004
|
0,012
|
0,019
|
0,007
|
|
JULBSE
|
Sciaphile
|
15307
|
12905
|
13181
|
6,15
|
13181
|
10907
|
11183
|
5,41
|
0,028
|
0,003
|
-0,024
|
0,035
|
0,005
|
-0,030
|
|
MANIZE
|
Sciaphile
|
265
|
262
|
287
|
6,06
|
287
|
281
|
296
|
5,33
|
0,002
|
0,015
|
0,013
|
0,004
|
0,010
|
0,006
|
|
STAUKA
|
Sciaphile
|
53
|
52
|
60
|
6,42
|
60
|
54
|
66
|
5,52
|
0,003
|
0,022
|
0,019
|
0,019
|
0,036
|
0,017
|
|
STROPU
|
Sciaphile
|
842
|
803
|
849
|
6,11
|
849
|
810
|
861
|
5,41
|
0,008
|
0,009
|
0,001
|
0,009
|
0,011
|
0,003
|
|
STROTE
|
Sciaphile
|
454
|
413
|
453
|
6,21
|
453
|
404
|
469
|
5,45
|
0,015
|
0,015
|
0,000
|
0,021
|
0,027
|
0,006
|
|
BLIGWE
|
Tolérant
|
20
|
20
|
26
|
6,29
|
26
|
26
|
30
|
5,51
|
0,000
|
0,042
|
0,042
|
0,000
|
0,026
|
0,026
|
|
CHRYBO
|
Tolérant
|
198
|
186
|
272
|
6,25
|
272
|
261
|
356
|
5,50
|
0,010
|
0,061
|
0,051
|
0,008
|
0,056
|
0,049
|
|
CLEIMI
|
Tolérant
|
613
|
574
|
607
|
6,73
|
607
|
549
|
600
|
5,61
|
0,010
|
0,008
|
-0,001
|
0,018
|
0,016
|
-0,002
|
|
GUARCE
|
Tolérant
|
338
|
321
|
338
|
6,21
|
338
|
323
|
350
|
5,48
|
0,008
|
0,008
|
0,000
|
0,008
|
0,015
|
0,006
|
|
IRVIEX
|
Tolérant
|
92
|
91
|
93
|
6,23
|
93
|
90
|
95
|
5,48
|
0,002
|
0,003
|
0,002
|
0,006
|
0,010
|
0,004
|
|
IRVIGR
|
Tolérant
|
39
|
37
|
41
|
6,28
|
41
|
40
|
43
|
5,54
|
0,008
|
0,016
|
0,008
|
0,004
|
0,013
|
0,009
|
|
PARIEX
|
Tolérant
|
138
|
102
|
112
|
6,17
|
112
|
86
|
97
|
5,43
|
0,049
|
0,015
|
-0,034
|
0,049
|
0,022
|
-0,026
|
|
PSEUMI
|
Tolérant
|
21
|
18
|
26
|
5,82
|
26
|
18
|
24
|
5,36
|
0,026
|
0,063
|
0,037
|
0,069
|
0,054
|
-0,015
|
Annexe 2 : Les différents paramètres de
la dynamique de la régénération des espèces
abondantes dans la parcelle de Lenda
|
Espèce
|
Tempérament
|
1996 - 2001
|
2001 - 2007
|
1996 - 2001
|
2001-2007
|
|
N0
|
Nt
|
N1
|
t
|
N0
|
Nt
|
N1
|
t
|
Mortalité
|
Recrutement
|
Chang_Po
|
Mortalité
|
Recrutement
|
Chang_Po
|
|
ALBIGU
|
Héliophile
|
128
|
102
|
113
|
5,77
|
113
|
80
|
87
|
6,32
|
0,039
|
0,018
|
-0,022
|
0,055
|
0,013
|
-0,041
|
|
ANTHMA
|
Héliophile
|
173
|
167
|
188
|
5,67
|
188
|
175
|
200
|
6,33
|
0,006
|
0,021
|
0,015
|
0,011
|
0,021
|
0,010
|
|
CELTMI
|
Héliophile
|
258
|
252
|
255
|
6,20
|
255
|
242
|
245
|
6,34
|
0,004
|
0,002
|
-0,002
|
0,008
|
0,002
|
-0,006
|
|
COLALA
|
Héliophile
|
319
|
299
|
334
|
5,93
|
334
|
308
|
351
|
6,33
|
0,011
|
0,019
|
0,008
|
0,013
|
0,021
|
0,008
|
|
ERYTSU
|
Héliophile
|
41
|
34
|
39
|
5,63
|
39
|
35
|
42
|
6,33
|
0,033
|
0,024
|
-0,009
|
0,017
|
0,029
|
0,012
|
|
GREWOL
|
Héliophile
|
403
|
366
|
413
|
5,76
|
413
|
367
|
408
|
6,34
|
0,017
|
0,021
|
0,004
|
0,019
|
0,017
|
-0,002
|
31
Dynamique de la
régénération des espèces abondantes des
forêts de l'Ituri : Cas des dispositifs permanents de la Réserve
de faune à Okapi
(c)Fiston Ngongo
|
HALLST
|
Héliophile
|
23
|
20
|
25
|
5,62
|
25
|
24
|
38
|
6,34
|
0,025
|
0,040
|
0,015
|
0,006
|
0,072
|
0,066
|
|
KLAIGA
|
Héliophile
|
75
|
71
|
72
|
6,20
|
72
|
68
|
70
|
6,36
|
0,009
|
0,002
|
-0,007
|
0,009
|
0,005
|
-0,004
|
|
MACASC
|
Héliophile
|
103
|
58
|
77
|
5,63
|
77
|
42
|
84
|
6,37
|
0,102
|
0,050
|
-0,052
|
0,095
|
0,109
|
0,014
|
|
PYCNAN
|
Héliophile
|
96
|
76
|
81
|
5,71
|
81
|
49
|
50
|
6,31
|
0,041
|
0,011
|
-0,030
|
0,080
|
0,003
|
-0,076
|
|
SARCPO
|
Héliophile
|
54
|
43
|
46
|
5,61
|
46
|
40
|
65
|
6,36
|
0,041
|
0,012
|
-0,029
|
0,022
|
0,076
|
0,054
|
|
TRECAF
|
Héliophile
|
177
|
160
|
163
|
6,21
|
163
|
150
|
156
|
6,35
|
0,016
|
0,003
|
-0,013
|
0,013
|
0,006
|
-0,007
|
|
UAPACO
|
Héliophile
|
74
|
57
|
65
|
5,86
|
65
|
41
|
44
|
6,31
|
0,045
|
0,022
|
-0,022
|
0,073
|
0,011
|
-0,062
|
|
UAPAGU
|
Héliophile
|
42
|
36
|
38
|
5,75
|
38
|
19
|
22
|
6,33
|
0,027
|
0,009
|
-0,017
|
0,110
|
0,023
|
-0,086
|
|
ZANTGI
|
Héliophile
|
42
|
25
|
29
|
5,70
|
29
|
13
|
15
|
6,30
|
0,091
|
0,026
|
-0,065
|
0,127
|
0,023
|
-0,105
|
|
ANGYPY
|
Sciaphile
|
408
|
402
|
428
|
5,84
|
428
|
413
|
435
|
6,33
|
0,003
|
0,011
|
0,008
|
0,006
|
0,008
|
0,003
|
|
ANONMA
|
Sciaphile
|
370
|
353
|
372
|
6,20
|
372
|
354
|
364
|
6,34
|
0,008
|
0,008
|
0,001
|
0,008
|
0,004
|
-0,003
|
|
CYNOAL
|
Sciaphile
|
218
|
209
|
217
|
6,36
|
217
|
197
|
208
|
6,35
|
0,007
|
0,006
|
-0,001
|
0,015
|
0,009
|
-0,007
|
|
DIALCO
|
Sciaphile
|
408
|
384
|
414
|
5,91
|
414
|
388
|
429
|
6,33
|
0,010
|
0,013
|
0,002
|
0,010
|
0,016
|
0,006
|
|
DIALEX
|
Sciaphile
|
56
|
51
|
51
|
5,91
|
51
|
47
|
49
|
6,34
|
0,016
|
0,000
|
-0,016
|
0,013
|
0,007
|
-0,006
|
|
DIALPE
|
Sciaphile
|
373
|
353
|
372
|
5,89
|
372
|
339
|
349
|
6,33
|
0,009
|
0,009
|
0,000
|
0,015
|
0,005
|
-0,010
|
|
GILBDE
|
Sciaphile
|
6688
|
6442
|
7077
|
5,78
|
7077
|
6687
|
7673
|
6,33
|
0,006
|
0,016
|
0,010
|
0,009
|
0,022
|
0,013
|
|
GREESU
|
Sciaphile
|
1355
|
1289
|
1370
|
5,95
|
1370
|
1275
|
1388
|
6,34
|
0,008
|
0,010
|
0,002
|
0,011
|
0,013
|
0,002
|
|
JULBSE
|
Sciaphile
|
2032
|
1867
|
2160
|
5,81
|
2160
|
1890
|
2278
|
6,32
|
0,015
|
0,025
|
0,011
|
0,021
|
0,030
|
0,008
|
|
MANIZE
|
Sciaphile
|
588
|
586
|
610
|
5,73
|
610
|
610
|
645
|
6,31
|
0,001
|
0,007
|
0,006
|
0,000
|
0,009
|
0,009
|
|
STAUKA
|
Sciaphile
|
649
|
586
|
601
|
5,92
|
601
|
492
|
511
|
6,34
|
0,017
|
0,004
|
-0,013
|
0,032
|
0,006
|
-0,026
|
|
STROPU
|
Sciaphile
|
169
|
160
|
167
|
5,90
|
167
|
150
|
158
|
6,33
|
0,009
|
0,007
|
-0,002
|
0,017
|
0,008
|
-0,009
|
|
STROTE
|
Sciaphile
|
395
|
372
|
411
|
5,79
|
411
|
367
|
411
|
6,31
|
0,010
|
0,017
|
0,007
|
0,018
|
0,018
|
0,000
|
|
BLIGWE
|
Tolérant
|
339
|
316
|
339
|
5,89
|
339
|
271
|
287
|
6,33
|
0,012
|
0,012
|
0,000
|
0,035
|
0,009
|
-0,026
|
|
CHRYBO
|
Tolérant
|
638
|
608
|
625
|
5,84
|
625
|
567
|
579
|
6,33
|
0,008
|
0,005
|
-0,004
|
0,015
|
0,003
|
-0,012
|
|
CHRYLA
|
Tolérant
|
39
|
36
|
38
|
6,20
|
38
|
37
|
44
|
6,33
|
0,013
|
0,009
|
-0,004
|
0,004
|
0,027
|
0,023
|
|
CLEIMI
|
Tolérant
|
44
|
43
|
43
|
5,56
|
43
|
40
|
40
|
6,35
|
0,004
|
0,000
|
-0,004
|
0,011
|
0,000
|
-0,011
|
|
GUARCE
|
Tolérant
|
26
|
21
|
21
|
5,88
|
21
|
17
|
19
|
6,35
|
0,036
|
0,000
|
-0,036
|
0,033
|
0,018
|
-0,016
|
Dynamique de la
régénération des espèces abondantes des
forêts de l'Ituri : Cas des dispositifs permanents de la Réserve
de faune à Okapi
(c)Fiston Ngongo
|
IRVIEX
|
Tolérant
|
34
|
34
|
34
|
6,10
|
34
|
32
|
32
|
6,35
|
0,000
|
0,000
|
0,000
|
0,010
|
0,000
|
-0,010
|
|
PARIEX
|
Tolérant
|
34
|
28
|
31
|
5,83
|
31
|
26
|
32
|
6,31
|
0,033
|
0,017
|
-0,016
|
0,028
|
0,033
|
0,005
|
|
PSEUMI
|
Tolérant
|
74
|
59
|
61
|
5,87
|
61
|
45
|
51
|
6,34
|
0,039
|
0,006
|
-0,033
|
0,048
|
0,020
|
-0,028
|
Annexe 3 : Le test t-student fait pour comparer
les densités (abondances) et les taux démographiques entre les
sites
|
First period
|
Second period
|
|
TMP
|
Edoro_Density
|
Lenda_Density
|
Edoro_Density
|
Lenda_Density
|
|
Helio
|
10,377
|
6,460
|
10,557
|
6,693
|
|
Shade Tol
|
91,869
|
54,808
|
100,212
|
52,727
|
|
Toler
|
9,469
|
7,450
|
9,119
|
7,675
|
|
t-student
|
t=0,45 p-value=0,67
|
t=0,52 p-value=0,62
|
|
First period
|
Second period
|
|
TMP
|
Edoro_Mortality
|
Lenda_Mortality
|
Edoro_Mortality
|
Lenda_Mortality
|
|
Helio
|
0,024
|
0,034
|
0,038
|
0,044
|
|
Shade Tol
|
0,01
|
0,009
|
0,014
|
0,013
|
|
Toler
|
0,014
|
0,018
|
0,02
|
0,023
|
|
t-student
|
t=-0,515 p-value=0,63
|
t=-0,23 p-value=0,83
|
32
|
First period
|
Second period First period Second period
|
|
TMP
|
Edoro_Recruitment
|
Lenda_Recruitment
|
Edoro_Recruitment
|
Lenda_Recruitment TMP Edoro_Pop change
|
Lenda_Pop change Edoro_Pop change
|
Lenda_Pop change
|
|
Helio
|
0,025
|
0,019
|
0,034
|
0,029 Helio 0,001
|
-0,015 -0,004
|
-0,015
|
|
Shade Tol
|
0,011
|
0,01
|
0,015
|
0,012 Shade Tol 0,001
|
0,001 0,001
|
-0,002
|
|
Toler
|
0,027
|
0,006
|
0,026
|
0,014 Toler 0,013
|
-0,012 0,006
|
-0,009
|
|
t-student
|
t=1,47 p-value=0,21
|
t=0,87 p-value=0,43 t-student t=2,16
p-value=0,097 t=2,04 p-valu=0,11
|


