|

Faculté des Sciences Psychologiques et de
l'Éducation
Influences circadiennes sur le jugement
social
STUHEC SEBASTIEN Mémoire
réalisé sous la direction
de Messieurs les Professeurs Philippe Peigneux et Olivier
Klein en vue de l'obtention du grade de Master en Sciences Psycholojques.
Abstract
Nous avons pour objectif d'approfondir les résultats
d'un article de Bodenhausen, 1990 qui mettait en évidence une variation
circadienne du jugement social. Afin de sélectionner la population de
notre étude, 805 personnes ont répondu à un questionnaire
de chronotype (sujets du matin ou du soir) en ligne. Sur base des scores
à ce questionnaire, 48 sujets ont été testés
à leur domicile, 24 extrêmes du matin (âge moyen : 47 ans)
et 24 extrêmes du soir (âge moyen : 34 ans). Nous avons
testé chaque sujet à deux reprises : une passation au moment
optimal (matin pour les matinaux, soir pour les vespéraux) et une
passation au moment non-optimal (soir pour les matinaux, matin pour les
vespéraux) de leur journée. Dans chaque groupe, la moitié
des sujets a commencé par la passation optimale et l'autre groupe par la
passation non-optimale et il s'est écoulé 7 à 10 jours
entre les deux passations. L'inertie de sommeil a été
contrôlée en faisant passer les sujets 1h30 et 10h30 après
leur réveil. Les sujets ont passé 4 tâches dont l'ordre de
passation a été contre-balancé. Ces tâches mesurent
la vigilance (tâche de vigilance psychomotrice), l'inhibition (GoNoGo),
le jugement social explicite (échelle de préjugés
explicite) et implicite (tâche d'association implicite IAT). Notre
étude met en évidence une variation circadienne de la vigilance
uniquement chez les sujets du soir, ceux-ci étaient plus rapides au
moment optimal de leur journée y compris lorsque l'on fait covarier
l'âge dans les analyses. Nos résultats n'indiquent pas de
différence significative aussi bien pour le jugement social implicite
qu'explicite. Nous n'avons également pas trouvé de
différence significative pour l'inhibition mais nous restons prudents
pour cette mesure en raison d'un problème d'encodage.
Remerciements
3
Remerciements
Il est un certain nombre de personnes sans qui la
réalisation de mes études et de mon mémoire auraient
été fortement compromis et à toutes ces personnes je
souhaite dire merci !
Les nombreuses difficultés que représente ce
travail ont vu un éclairage sans égal grâce à mes
promoteurs Philippe Peigneux et Olivier Klein qui se sont montrés
toujours à la hauteur. Je tiens également à remercier
Philippe Bernard pour les nombreuses heures qu'il m'a consacré et
Nicolas Van Der Linden qui m'a été d'un grand secours.
Je tiens à remercier ma maman qui a toujours cru en moi
et m'a apporté un soutien inconditionnel. Ma famille et belle-famille ne
sont pas en reste et je leur offre toute ma gratitude pour les innombrables
coups de pouce qu'ils m'ont apporté, je pense notamment à ma
chérie Floriane, sa mère Marie et son père Jean-Louis.
J'ai une pensée également en écrivant
cette partie pour le cercle de psychologie, tous ses membres et surtout tous
mes amis. Cette formidable association m'a permis de tenir bon quand le moral
l'était moins et le soutien de tous ceux qui la compose est une
incroyable richesse.
Je remercie également les 805 personnes qui ont pris le
temps de répondre à mon questionnaire de chronotype, à
toutes celles qui ont bien voulu relayer l'information et aux quelques-unes qui
m'ont aidé dans la rencontre avec les bénévoles et dans
mes déplacements.
Il serait malhonnête de clôturer cette partie sans
remercier les 49 bénévoles qui ont accepté que je vienne
à des horaires invraisemblables leur faire passer des tâches qui
avaient un gout plutôt amer pour certains d'entre eux.
A toutes ces personnes et aux nombreux que j'oublie probablement,
merci !
Table des matières
Préface 6
I. Introduction 7
1 Jugement social 7
1.1. Concepts du jugement social 7
1.1.1. Attitudes, croyances et intentions 7
1.1.2. Préjugés, stéréotypes et
discrimination 8
1.1.3. Représentation mentale et formation d'impression
8
1.2. Le stéréotype : historique 9
1.3. Contenu des stéréotypes 10
1.3.1. Modèle par exemplaires et modèle par
prototype 10
1.3.2. Contenu descriptif et contenu explicatif 11
1.4. Caractéristiques et fonctions 12
1.5. Facteurs cognitifs de maintien des stéréotypes
12
1.5.1. Catégoriser la cible 12
1.5.2. Assimiler les attributs de la cible 13
1.5.3. Interpréter le comportement de la cible 13
1.5.4. Expliquer les causes du comportement de la cible 14
1.5.5. Se souvenir de la cible 14
1.5.6. Généraliser la perception de la cible
á sa catégorie sociale 14
1.6. Activation du stéréotype 15
1.6.1. Automaticité 15
1.6.1.1. L'activation des stéréotypes est-elle
vraiment automatique ? 16
1.7. Mesure des stéréotypes 16
1.7.1. Tfiche d'association implicite 17
1.7.2. Les mesures indirectes des préjugés 17
2. Fonctions exécutives 18
2.1. L'inhibition 19
2.1.1. Au niveau cognitif 19
2.1.1.1. Historique 19
2.1.1.2. La relation lobe frontal-inhibition. 20
2.1.1.3. Inhibition contrôlée et automatique. 20
2.1.1.4. Caractéristiques de l'inhibition 21
2.2. Evaluation 22
2.3. Inhibition et stéréotypes 24
3 Les rythmes 25
3.1. Les rythmes biologiques 25
3.2. Régulation homéostatique 25
3.3. Les rythmes circadiens 26
3.3.1. Endogènes 26
3.3.2. Génétiquement déterminés 27
3.3.3. Synchronisés 27
3.4. Interaction entre rythmes circadiens et régulation
homéostatique 28
3.5. Paradigmes d'exploration 29
3.5.1. Désynchronisation forcée 29
3.5.2. Routine constante 30
3.5.3. Chronotype 30
3.5.4. Contraintes méthodologiques 32
3.5.4.1. Difficulté de la tâche 32
3.5.4.2. Différences inter-individuelles 33
3.5.4.3. Inertie de sommeil 33
3.6. Relation avec les fonctions cognitives 33
3.7. Chronotype et inhibition 34
3.8. Chronotype et stéréotypes 35
Questions de recherche et hypothèses 37
Hypothèses 37
II. Méthode 39
Sélection de la population 39
MEQ (Morning Evaluation Questionnaire) 39
Plan expérimental 40
Expérience 41
Tâche Go/No-Go (inhibition) 41
Tâche de vigilance psychomotrice (PVT) (vigilance) 42
Tâche d'association implicite (IAT) (jugement social
implicite) 43
Echelle explicite de préjugés (jugement social
explicite) 44
III. Résultats 46
Tâche de vigilance psychomotrice 46
GoNoGo 48
Tâche d'association implicite 51
Echelle explicite de préjugés 52
Corrélations 54
IV. Discussion 55
Rappel des hypothèses 55
Résultats principaux et interprétation 56
Tâche de vigilance psychomotrice 57
Tâche GoNoGo (mesure de l'inhibition) 58
Echelle explicite 60
Tâche d'association implicite (IAT) 61
Bibliographie 63
Articles 63
Synthèses d'articles 66
Cours 67
Chapitres de livres 67
Livres 68
Ressources multimédia 69
Sites Internet 69
Thèses et mémoires 70
Thèses et mémoires en ligne 70
Annexes 71
Annexe 1 : Questionnaire de chronotype en ligne 71
Annexe 2 : Echelle explicite de préjugés 76
Preface
En 1990, Galen Bodenhausen mettait en évidence que le
jugement social peut être influencé par les rythmes circadiens, un
type de rythme biologique d'une durée d'environ 24 heures. En d'autres
termes, le jugement social serait influencé par le moment de la
journée.
Bodenhausen parvient à ces résultats en
étudiant un premier groupe de sujets qui se lèvent très
tôt le matin dits extrêmes du matin et un second groupe de sujets
qui se lèvent tard dit extrêmes du soir. Il demande à ces
participants d'émettre un jugement social sur un texte. Il constate
alors que lorsque le moment de la journée ne leur est pas favorable
(soir pour les sujets du matin, matin pour les sujets du soir), les sujets sont
plus enclins à porter un jugement social davantage
défavorable.
Quel mécanisme pourrait expliquer cette variation
circadienne du jugement social ?
Une réponse à cette question pourrait être
l'inhibition cognitive. Cette fonction cérébrale est la
capacité à refreiner ses réactions spontanées. En
d'autres termes, l'inhibition permet de s'empêcher de produire certaines
réponses. Dans l'étude de Bodenhausen, nous pouvons nous
interroger si c'est ce dont il est question. Le sujet, dans un moment qui n'est
pas optimal pour lui, ne parvient pas à inhiber un jugement
défavorable. Ce point va donc nous intéresser ici, l'inhibition
cognitive et le jugement social sont-ils liés. Est-ce l'activation de ce
mécanisme qui rend compte de la variation circadienne du jugement social
? Si tel est le cas nous devrions observer tout d'abord une variation
circadienne de l'inhibition et cette variation devrait être similaire
à celle du jugement social. Autrement dit, le moment n'est pas
idéal pour moi, je ne parviens pas à inhiber et mon jugement
social s'en ressent, il est plus défavorable.
I. Introduction
Il est tôt le matin, je suis réveillé par
le bruit assourdissant que ma voisine fait avec son sèchecheveux. Ni une
ni deux, ça trotte dans ma tête, il n'y a qu'une femme pour faire
ça. Et si mon horloge interne n'était pas totalement
étrangère à ma production de stéréotypes
?
Cette question, à première vue anodine, pourrait
n'être pas négligeable dans le milieu des soins ou celui du droit.
En effet, si une infirmière à domicile ou un neuropsychologue ont
des stéréotypes sur les personnes d'origine
étrangère différents le matin du soir, la prise en charge
peut s'en trouver différente.
Pour ce travail nous allons donc nous intéresser à
trois dimensions : le jugement social, les rythmes circadiens et les fonctions
exécutives.
Nous allons commencer par nous interroger sur ce qu'est le
jugement social. 1 Jugement social
1.1. Concepts du jugement social
1.1.1. Attitudes, croyances et intentions
Dimension
conative
Dimension
cognitive

Intention
Je vais acheter
le nouveau disque
de Richard
Séguin
Attitude
J'aime
le nouveau disque
de Richard Séguin
Sources de la croyance Croyance
Expérience personnelle
J'ai écouté le nouveau disque de Richard
Séguin Interaction sociale
Mon meilleur ami aime le nouveau disque de Richard
Séguin.
J'ai lu une critique élogieuse du nouveau disque de
Richard Séguin
Inférence
Les disques de Richard Séguin sont bons en
général : celui-ci doit donc l'être aussi
Le nouveau disque
de Richard Séguin
est une oeuvre
de
qualité.
Figure 1 : attitude, croyance et intention (Bédard,
Déziel & Lamarche, 2006)
Comme l'illustre la figure 1, on peut distinguer 3 dimensions
de la personne. L'attitude est un état affectif favorable ou
défavorable que l'on ressent envers tout ce qui peut être
représenté, incluant les personnes. L'attitude tient du domaine
affectif et transparait dans les croyances et intentions de la personne.
Une croyance est une conviction se rapportant à tout objet
représentable. Elle trouve trois origines possibles :
l'expérience personnelle, l'interaction sociale et
l'inférence.
Les intentions, elles, relèvent du domaine des actions qui
sont projetées envers un objet/sujet concret, abstrait réel ou
imaginaire (Bédard, Déziel & Lamarche, 2006).
1.1.2. Préjugés, stéréotypes et
discrimination
Le stéréotype est une croyance répartie sur
les traits d'individus se rapportant à un groupe. Le
stéréotype est donc une conviction.
Le préjugé est une attitude, générale
dépréciative envers les individus d'un groupe. Le
préjugé est donc un état affectif.
La discrimination est un comportement dévalorisant
vis-à-vis de personnes d'un groupe. Un comportement est la partie d'une
action qui est visible à un observateur. La discrimination relève
donc de l'action (Bédard et al., 2006).
1.1.3. Représentation mentale et formation
d'impression
On distingue 3 formes de représentations : les
représentations imagées, les représentations
propositionnelles et les représentations liées à
l'action.
- Les représentations imagées permettent de coder
les renseignements visuels : agencement dans l'espace, structure, couleur,
etc.
- Les représentations propositionnelles sont fortement
reliées au langage et reflètent notre connaissance sur les
caractères des objets de notre environnement et sur les rapports que ces
objets entretiennent.
- Les représentations liées à l'action
sont relatives à l'exécution et à l'aspect
sémantique,
elles sont une connaissance implicite sur le développement
et la régulation de l'action.
Les représentations mentales nous permettent de stocker
l'information en ceci que leur contenu sur les objets, situations ou
événements est conservé en mémoire. Ensuite, comme
vu plus haut elles nous renseignent sur le monde, l'explicitent. Qu'il s'agisse
des représentations transitoires, conservées en mémoire de
travail ou des représentations permanentes, stabilisées dans la
mémoire à long terme, les informations doivent être
rapidement accessibles. Enfin, elles permettent de guider l'action des
individus en ceci qu'elles lui décrivent l'environnement (Meunier,
2009).
Les processus impliqués dans les attributions causales
et les représentations mentales des autres sont au coeur de la
perception interpersonnelle. Les impressions que nous formons des autres
guident nos attentes, nos perceptions et interprétations des
comportements des autres mais aussi nos réponses cognitives, affectives
et comportementales envers les autres (Mrug & Hoza, 2007).
1.2. Le stéréotype : historique
Nous pouvons facilement percevoir la présence de
stéréotypes néanmoins ce concept n'est pas si
évident à saisir. On trouve ce terme dès le
18ème siècle, il trouve sa racine étymologique
dans les mots grecs stereos (solide) et tùpos
(empreinte, caractère), il correspond à cette époque
à un procédé d'impression qui permet de « convertir
en planches solides un seul bloc des pages préalablement
composées en caractères mobiles " (Mortier, 1963; in
Légal, 2008). Les stéréotypes offraient un
mécanisme typographique rapide, peu onéreux mais de peu de
valeur.
Un siècle plus tard, le concept de
stéréotypie est appliqué par les psychiatres pour
dénommer une attitude sous un angle temporel et topographique (Ashmore
& Del Boca, 1981 ; in Légal, 2008). On peut dès lors
remarquer que dès sa conception, le stéréotype se
réfère à quelque chose de figé, constant, rigide et
répété (Légal, 2008).
On doit le terme de stéréotype en sciences
sociales à Lippman qui en 1922 perçoit cette notion comme un
mécanisme simplificateur qui permet de gérer «
l'environnement réel, qui est à la fois trop grand, trop complexe
et trop évanescent pour une connaissance directe " (Lippman, 1922 ; in
Bosche, 2007).
En 1943, Jean Stoetzel, apporte des précisions sur la
notion de stéréotype. Il les décrit comme « porteurs
de signification " (Pichon, 2009). Le stéréotype est parfois
associé à un raisonnement syllogistique (Stening & Everett,
1979 ; in Bosche, 2007). Par exemple : les français font souvent la
grève, M. Dupont est français donc M. Dupont fait souvent la
grève. En 1987, Fisher (in Pichon, 2009) va plus loin dans la
définition du stéréotype et lui attribue la notion de
catégorisation fonctionnelle, le stéréotype se
définit comme « une catégorisation descriptive
simplifiée par laquelle nous cherchons à situer autrui ou des
groupes d'individus ".
1.3. Contenu des stéréotypes
Un stéréotype est formé de l'assortiment
de divers éléments de connaissances. Il est question d'un
système cognitif comprenant des connaissances et des
représentations mentales, apposés à une catégorie
ou un groupe, et qui est conservé en mémoire. L'appartenance
à une catégorie est déterminée selon Bruner,
Goodnow et Austin, 1956 (in Légal, 2008) par un nombre fini de traits
sachant que chacun d'entre eux est essentiel. Cette vision a rapidement
été délaissée au bénéficie des
paradigmes de la psychologie cognitive (Légal, 2008). Rips, Shoben et
Smith, 1973 (in Johnson, 2001) nuancent cette vision, pour eux certains objets
d'une catégorie comme une bouteille ou une plante sont plus typiques
d'une catégorie que d'autres objets.
Rosch et Mervis, 1975 (in Johnson, 2001) suggèrent que
l'inscription dans une catégorie se fait selon un certain niveau. Une
catégorie serait représentée par un élément
prototypique qui est le plus caractéristique de celle-ci.
1.3.1. Modèle par exemplaires et modèle par
prototype
Deux types de modèles se sont développés
pour expliquer la façon dont ces structures cognitives sont acquises :
le modèle par exemplaires et le modèle par prototypes. L'approche
associationniste est basée sur les principes d'un apprentissage
associatif et met l'accent sur l'histoire de l'apprentissage l'individu. Les
bons exemples de la catégorie sont ceux qui ont fréquemment
été associés à la catégorie dans le
passé. La mémoire se limite alors à retenir peu à
peu les informations données. Alternativement, la comparaison
basée sur la similarité implique un processus de
détermination du degré avec lequel un exemple partage les traits
ou propriétés commun(e)s avec les exemples stockés du
concept. Dans ce cadre théorique, les bons exemples de la
catégorie sont ceux qui sont similaires au prototype de la
catégorie. C'est donc le prototype et non l'information sur l'objet ou
l'individu, qui va permettre de décider l'inscription ou non aux membres
de la catégorie (Johnson, 2001).
Recentrons notre question : Qu'est-ce qu'on rencontre au sein
du stéréotype ? Il comprend un ensemble de connaissances à
propos des traits d'une catégorie, l'aspect physique, le comportement ou
la personnalité de ses membres. On y repère des exemplaires
typiques de la catégorie (Légal, 2008). Par exemple, examinons
quelques traits souvent reliés aux personnes
d'origine nord-africaine. Quand vous songez à cette
catégorie d'individus, des traits vous viennent en tête. Au sein
de ceux-ci, vous avez peut-être estimé qu'une personne de cette
catégorie parle fort et en arabe (comportements), que ses cheveux sont
bruns, crépus et touffus (caractéristiques physiques), qu'elle a
une réflexion limitée (capacités), est avare et agressive
(personnalité). Les stéréotypes que vous exprimez ne sont
pas nécessairement négatifs et vous pouvez également
penser qu'une personne d'origine nord-africaine est chaleureuse et extravertie
(comportements), que son teint est doré et son regard expressif
(caractéristiques physiques), qu'elle a de bonnes capacités
à courir (capacités), qu'elle est accueillante et
généreuse (personnalité). Vous avez éventuellement
imaginé le visage de votre voisin, de Tariq Ramadan ou Zinedine Zidane
(exemplaire typique).
On trouvera des variations conséquentes concernant le
degré de définition et de contenu des stéréotypes.
De ce fait, des stéréotypes sont emplis de contenu et
amène vers des connaissances s'appliquant à de nombreuses
caractéristiques et comportements de la catégorie visée
tandis que d'autres seront maigres et seront restreints à un nombre
d'informations très faible. Ces variations trouvent leur origine dans
différents éléments : culture, enseignement,
considération individuelle du stéréotype etc. Le contenu
des stéréotypes d'une personne avec un niveau de
préjugés élevé sera probablement composé
majoritairement d'éléments négatifs (les maghrébins
sont paresseux, virulents, etc.) alors que celui des personnes avec un bas
niveau de préjugés contiendra non seulement des
éléments négatifs mais aussi des éléments
positifs (les maghrébins sont virulents mais ils sont bons
marathoniens). Les stéréotypes sont donc différents d'une
personne à l'autre.
Par définition, les stéréotypes sont des
croyances, parfois erronées mais contenant fréquemment des
éléments de vérité. Quoi qu'il en soit, les
stéréotypes sont des schématisations de la
réalité et reflètent une inclination à
l'exagération des ressemblances au sein du groupe
stéréotypé (Légal, 2008).
1.3.2. Contenu descriptif et contenu explicatif
Le contenu descriptif des stéréotypes est
étudié afin de mettre en évidence la perception
homogène des autres groupes. Dans ces investigations, une mesure porte
sur l'endogroupe et l'exogroupe. Cette mesure peut par exemple être
effectuée par questionnaire où les sujets doivent évaluer
si les traits proposés s'appliquent ou non à un groupe
communautaire (ex : les japonais sont travailleurs). Ce type de mesure permet
de mettre en évidence ce que le sujet
exprime explicitement sur une population mais présente
l'inconvénient d'être peu sensible notamment à cause du
biais de désirabilité sociale.
Le contenu explicatif des stéréotypes concerne
la fonction du stéréotype, ses dimensions cognitive, affective et
émotionnelle. Les études portent ici sur le « sens commun
», c'est-àdire l'explication que tout-un-chacun tente de donner
à ses cognitions. Ainsi, lorsque le sujet est placé dans un cas
de dissonance cognitive (par exemple : « je suis militant
écologiste» or « je prends un bain tous les jours »), on
pourra généralement mettre en évidence chez le sujet un
travail de rationalisation afin de rétablir la consonance (ex : «
je suis en train d'installer un bac de récupération d'eau de
pluie pour mon potager biologique, ce qui compense l'eau de mon bain
».)
Ainsi, le stéréotype sert à simplifier
l'environnement mais pas uniquement en réduisant l'information, il est
aussi pour fonction de remplir un besoin d'explication de la
réalité (SalesWuillemin, 2006).
1.4. Caractéristiques et fonctions
L'accessibilité d'un stéréotype
correspond à la probabilité qu'un groupe social soit
activé. Plus une catégorie est accessible, plus il est
vraisemblable que celle-ci soit stimulée par une entité et donc
employée pour catégoriser cet élément (Bry, 2008 ;
in Pichon, 2009). L'accessibilité serait dépendante de la
fréquence et de la récence d'activation. Elle serait essentielle
à l'activation d'une catégorie mentale.
L'utilisation majeure des stéréotypes est
pratique, ils offrent la possibilité de simplifier la
réalité. Ils nous servent dès lors à gérer
la quantité d'informations sur les objets et les individus qui nous
entourent et guident nos comportements (Klein, 2008-2009).
1.5. Facteurs cognitifs de maintien des
stéréotypes
1.5.1. Catégoriser la cible
Les personnes et les objets sont classifiés en
catégories. L'appartenance à une catégorie répond
de divers facteurs tels que les catégories qui sont souvent
activées, la catégorie qui exprime au mieux le comportement de la
cible ou encore les buts du moment.
La catégorisation va permettre de simplifier, de mobiliser
les acquisitions passées, d'agencer l'environnement et de guider les
comportements.
La catégorisation va orienter l'attention et
résulter en une accentuation de la similarité des membres d'une
catégorie et l'exagération des différences entre
catégories (Waroquier & Klein, 2007).
1.5.2. Assimiler les attributs de la cible
Lorsque nous catégorisons un individu comme appartenant
à une catégorie sociale particulière (ex : Alexandre est
paresseux), pour que le stéréotype que nous portons sur cet
individu soit modifié il est nécessaire que de l'information
contre-stéréotypique (ex : ma voisine m'a dit qu'il travaille 8
heures par jour dans une association) soit traitée. Néanmoins,
cette information est généralement ignorée lors de la
formation d'impression et ce pour deux raisons.
Comme vu ci-dessus, les stéréotypes orientent le
processus attentionnel (Fiske, 1998 ; in Waroquier & Klein, 2006). Nous
recherchons en l'autre ce que nous prévoyons de visualiser. Par exemple,
convié chez un ami d'origine maghrébine, notre choix de cadeau se
fera peutêtre davantage dans une confiserie de loukoum que dans une
fromagerie.
Mais encore, l'information qui est cohérente avec le
stéréotype serait plus facilement intégrée (ex:
Bodenhausen & Macrae, 1998 ; in Waroquier & Klein, 2006). Il semble
plus simple d'assimiler l'information suivant laquelle une femme au foyer fait
régulièrement le ménage (comportement
stéréotypique) plutôt que d'assimiler qu'elle lit des
livres scientifiques (comportement neutre) ou qu'elle consacre ses week-end
à la mécanique automobile (comportement
contre-stéréotypique). En mémoire, le groupe social et
l'attribut seraient unis par des liens de causalité (une femme au foyer
doit s'occuper de sa maison et doit donc faire le ménage, etc.),
permettant l'assimilation de l'information concordante avec le
stéréotype.
1.5.3. Interpréter le comportement de la cible
Les comportements de la cible, neutres ou mêmes
contre-stéréotypiques font fréquemment l'objet d'une
interprétation biaisée qui renforce le
stéréotype.
En 2007, Correll, Park, Judd et Wittenbrink utilisent un jeu
vidéo pour simuler des rencontres avec des cibles potentiellement
hostiles. Leur étude s'intéresse au biais racial dans une
décision tirer/ne pas tirer. L'expérience révèle
notamment que la tendance des participants à commettre une erreur (tirer
sur un noir non armé ou ne pas tirer sur un blanc armé) est
fortement augmentée lorsque ceux-ci ont précédemment lu
l'histoire d'un criminel noir.
1.5.4. Expliquer les causes du comportement de la cible
En 1977, Ross (in Riggio & Garcia, 2009)
définissait l'erreur d'attribution fondamentale comme étant la
tendance à surestimer les facteurs situationnels pour les comportements
des autres.
Cette erreur d'attribution fondamentale trouve une place de
choix dans les stéréotypes. En effet, l'erreur d'attribution
fondamentale est une double tendance, celle d'octroyer les comportements
concordants avec un stéréotype à des facteurs internes et
la tendance à interpréter les comportements
contre-stéréotypiques comme relevant de l'environnement. Ainsi,
Jacques apprend l'histoire d'un jeune, Brahim, qui a aidé une femme
blessée dans le métro. Jacques explique ce comportement par une
attribution extérieure à Brahim (Brahim est étudiant
infirmier et ses amis l'ont incité à aider la femme) plutôt
qu'à une disposition interne (Brahim est altruiste).
1.5.5. Se souvenir de la cible
Doosje, Spears, De Redelijkheid et Van Onna, 2007 ont
étudié le rôle de la consistance sur la mémoire des
groupes de hauts et bas statuts. Leurs résultats indiquent une
mémoire supérieure pour l'information consistante avec le
stéréotype en comparaison avec une information inconsistante.
Cependant, leur seconde étude indique que certains sujets («
high-identifiers ») mobilisent davantage de ressources cognitives pour
retenir l'information inconsistante et s'en souviennent par conséquent
mieux que les autres sujets (« low identifiers »). Cette étude
va contre l'idée que l'identification intra-groupe est reliée
à l'information favorable à propos de ce groupe. Apparemment, la
motivation à résoudre des inconsistances cognitives
amènerait à une mémoire supérieure pour les
informations nonfavorables comparativement à la motivation à
défendre une image positive en retenant uniquement les informations
favorables.
1.5.6. Généraliser la perception de la cible
à sa catégorie sociale
Même lorsque les gens sont confrontés à
des personnes qui remettent en question leur stéréotype sur
l'exogroupe, ils admettent des exceptions individuelles en re-cloisonnant le
stéréotype (Allport, 1954 & 1979 ; in Dovidio, Glick &
Rudman, 2005). Les personnes créent de petits sous-types qui contiennent
les exceptions, protégeant ainsi leur catégorie globale.
Les sous-types sont pratiques parce qu'ils permettent aux gens
de retenir leurs catégories qui sont confortables. Les gens trouvent
pratique de retenir des catégories générales parce
qu'elles nécessitent plus d'effort et l'effort est
désagréable (Allport, 1954 &1979 ; in Dovidio et al., 2005).
Mais cet aspect pratique a ses limites et le sous-type peut alors devenir
sous-groupe. Alors que les sous-types maintiennent le stéréotype,
les sous-groupes augmentent la perception de variabilité au sein de la
catégorie et limitent la perception rigide d'une catégorie
(Dovidio et al., 2005).
1.6. Activation du stéréotype
Quand les gens sont motivés à
déprécier un individu, ils sont susceptibles d'activer des
stéréotypes négatifs qui autrement seraient restés
dormants. De manière similaire, quand ils sont motivés à
penser fortement à un individu, ils sont plus enclins à activer
des stéréotypes positifs qui autrement n'auraient pas
été activés. Ces idées peuvent paraitre
controversées parce que certaines études ont indiqué que
les stéréotypes sont toujours activés de manière
automatique lorsque l'individu est exposé aux membres de groupe
stéréotypé (cf. Devine, 1989 ; in Kunda & Sinclair,
1999).
Cependant des recherches plus récentes indiquent que
l'activation de pensées stéréotypiques n'est pas
automatique pour tout le monde ou dans toutes les situations (Radvansky,
Copeland & Von Hippel, 2010)
1.6.1. Automaticité
Les premières applications de la distinction entre
processus automatiques et contrôlés dans l'étude des
préjugés, déclaraient que l'activation des
stéréotypes était automatique. Par exemple, Devine en 1989
(in Devine & Monteith, 1999) indiqua que l'activation des
stéréotypes ne nécessitait ni intention ni attention ou
capacité, ainsi lorsqu'un indice approprié est
présenté (ex : genre, niveau social) l'activation du
stéréotype suit inévitablement. Devine a
interprété les résultats de son étude comme
indiquant que l'activation des stéréotypes est automatique aussi
bien pour les individus avec un niveau haut que bas de
préjugés.
Des études utilisant des procédures alternatives
ont également suggéré que les stéréotypes de
genre et d'age peuvent être activés automatique. Par exemple,
Purdue et Gurtman (in Devine & Monteith, 1999) ont utilisé une
procédure de masquage de l'amorçage avant que les sujets n'en
soient conscients. L'évaluation des traits négatifs était
plus rapide quand ils étaient
précédés d'une amorce «
âgé » qu'une amorce « jeune ». Purdue et Gurtman
interprètent ces résultats comme un « âgisme »
automatique. Selon Devine et selon Purdue et Gurtman, si les participants ne
sont pas conscients de l'amorce, leurs réactions ne peut pas
refléter un processus contrôlé (Devine & Monteith,
1999).
Banaji et Hardin, 1996 étudient les
stéréotypes de genre en utilisant un temps de présentation
entre l'amorce et la cible relativement court, empéchant
l'exécution de processus contrôlés. Les jugements pour des
paires amorce-cibles consistent sont plus rapides que les jugements pour les
paires inconsistantes indiquant une activation automatique des
stéréotypes concernant le genre.
1.6.1.1. L'activation des stéréotypes
est-elle vraiment automatique ?
L'automaticité regroupe au moins 4 traits qui peuvent
être définis en relation avec les objectifs : (non) intentionnel,
(in)contrôlé/ (in)contrôlable, autonome, dirigé vers
un but, (in)dépendant d'un but (Moors & De Houwer, 2006)
De ces caractéristiques, Bargh, 1989 (in Devine &
Monteith, 1999) distingue 3 types d'automaticité :
- L'automaticité pré-consciente : processus
automatiques qui se produisent avant la prise de conscience et ne
nécessitant que la présence d'un élément
déclencheur pour être initiés.
- L'automaticité post-consciente : elle requière
certains types de traitement conscient
des stimuli pertinents mais les résultats de ce traitement
sont involontaires
- L'automaticité dépendante du but : elle
requière un traitement intentionnel, dirigé
vers un but dont les résultats peuvent être
volontaires ou non.
1.7. Mesure des stéréotypes
Pour évaluer l'application de stéréotypes
et préjugés, de multiples procédés ont
été élaborés. Les techniques ont
évolué depuis les mesures classiques relativement globales et
extrêmement directes, en passant par les mesures d'accessibilité
et les mesures classiques et indirectes des préjugés plus fines.
Toutes ces approches montrent cependant des bénéfices et des
désavantages.
Au fil du temps, la mesure des préjugés a
évolué jusqu'à des mesures indirectes moins
réactives et sensibles aux normes sociales.
1.7.1. Tâche d'association implicite
Une tâche d'association implicite (IAT) mesure la
différence d'association entre 2 concepts cible et un attribut. Les 2
concepts apparaissent dans une tâche à deux choix (ex :
insectes/fleurs) et l'attribut dans une seconde tâche (ex :
plaisant/déplaisant). Quand les instructions obligent à
répondre dans le sens d'une haute association entre catégories
(ex : insectes + déplaisant), la performance est plus rapide que lorsque
l'association est faible (ex : insectes + plaisant) (Greenwald, McGhee &
Schwartz, 2008).
1.7.2. Les mesures indirectes des préjugés
Ces mesures se font sous forme de questionnaires ou
d'échelles. Elles essaient de minimiser les biais inhérents aux
normes sociales (stratégies d'autoreprésentation,
désirabilité sociale) en utilisant des chemins
détournés, sans parvenir cependant à les éliminer
totalement.
De cette manière, l'échelle de racisme moderne
(McConahay, 1986 ; in Légal, 2008), dans son énonciation, adopte
des énoncés plus abstraits, reliés de manière plus
détournée aux conduites racistes. L'Echelle moderne de sexisme
(Swim, Aikin, Hall & Hunter, 1995 ; in Légal, 2008), est
passée de mesures requérant un degré d'accord (de «
pas du tout" à « tout à fait") avec des formulations du type
« Les femmes ne sont généralement pas aussi intelligentes
que les hommes " à des mesures plus sophistiquées telles
que« La discrimination envers les femmes n'est plus un problème de
nos jours " (Légal, 2008).
Comme nous l'avons vu, les stéréotypes ont un
but heuristique, ils nous renseignent sur le monde, nous permettent de s'y
adapter. Les fonctions exécutives sont les mécanismes
quinous permettent de nous adapter à notre monde. Avant de
nous intéresser au lien entre ces deux concepts, penchons notre
attention sur les fonctions exécutives.
2. Fonctions exécutives
Les fonctions exécutives englobent une série de
processus de haut niveau dont la fonction principale est de faciliter
l'adaptation à des situations nouvelles ou complexes (Collette, Hogge,
Salmon, Van der Linden, 2006 ; in Schmidt, 2009)
Pour réaliser des tâches routinières, les
mécanismes mis en place sont automatiques, ne nécessitant pas de
réflexion et très peu d'attention. Les situations nouvelles,
elles, nécessitent la sélection d'une action appropriée,
demandant attention et réflexion (Degiorgio, Fery, Polus & Watelet,
s.d.).
La mise en évidence des mécanismes
désignés sous l'appellation « fonctionnement exécutif
» provient d'études neuropsychologiques de patients
cérébro-lésés et d'études d'imagerie
cérébrale fonctionnelle de sujets sains. Ces études
mettent en exergue une distinction entre les processus étudiés
mais pas une totale indépendance (Meulemans, Collette & Van der
Linden, 2004).
Ces fonctions qui permettent de s'adapter à des situations
nouvelles, qui permettent à l'individu de s'adapter, sont les suivantes
:
- L'inhibition (capacité de s'empêcher de produire
une réponse automatique) - La mise à jour (rafraîchir le
contenu de sa mémoire de travail)
- La flexibilité mentale (passer d'un comportement
à un autre)
- La récupération active d'informations en
mémoire
- L'attention divisée (être attentif à deux
activités en méme temps)
- La planification (organiser une série d'actions)
Le fonctionnement exécutif a initialement
été associé à l'activité dans les
régions cérébrales frontales (Schmidt, 2009). A ce sujet,
des tâches relevant du cortex pré-frontal relativement complexes
(Tour de Londres, Wisconsin Card Sorting Test, tâche de raisonnement
logique) se sont révélées sensibles aux effets du moment
de la journée et à la déprivation de sommeil (Drumond,
Brown, Salamat, Gillin, 2004; Jones, Harrison, 2001; in Blatter & Cajochen,
2006). Un nombre grandissant d'études récentes démontre
qu'une partie du fonctionnement exécutif dépend de régions
cérébrales postérieures (essentiellement
pariétales) (Harrison, Jones & Waterhouse, 2007 ; in Schmidt,
2009).
2.1. L'inhibition
L'inhibition est la capacité à s'empêcher
de produire une réponse automatique, à arréter la
production d'une réponse en cours et écarter les stimulations non
pertinentes pour l'activité en cours (Degiorgio et al., s.d.). Cette
fonction cognitive trouve donc sa place dans une étude sur les
stéréotypes et les rythmes circadiens. En effet, si un sujet ne
parvient pas à s'empêcher de produire une réponse
automatique et qu'à ce moment de la journée sa réponse est
davantage stéréotypique, un pont peut être jeté
entre les deux notions.
2.1.1. Au niveau cognitif
Quelle est la différence entre focaliser son attention
sur quelque chose et inhiber le reste ? Dans une tâche Stroop, il est
difficile de déterminer si le phénomène basique - le
ralentissement qui se manifeste lors d'essais incongruents - est dû
à une activation plus importante de la couleur de l'encre ou à
l'inhibition du nom des mots, ou les deux. Il en est de même pour de
nombreuses tâches exécutives-attentionnelles. Mais il y a un cas
-- L'inhibition de la réponse - dans lequel l'inhibition, plus que
l'attention, est clairement la clé. L'inhibition de la réponse
est la suppression de la réponse partiellement préparée
(Smith & Kosslyn, 2009)
2.1.1.1. Historique
Le premier cas de changement comportemental rapporté
dans de la littérature neuroscientifique est celui de Phineas Gage
(Kotowicz, 2007), ayant subi une perforation frontale par une barre de
métal. Pour décrire les troubles comportementaux de Phineas Gage,
Harlow, 1968 (in Meulemans et al., 2004) utilise déjà le concept
d'inhibition. Ce concept sera utilisé dans des domaines divers tel que
la psychopathologie, la psychologie développementale ou encore
neuropsychologie (Meulemans et al., 2004).
En neuropsychologie, l'inhibition est habituellement
décrite comme un processus inverse, qui freine un processus actif plus
important que l'on nomme Activation (Boujon, 2002)
Trois fonctions d'inhibition en mémoire de travail ont
été identifiées (Hasher al., 1999 ; in Meulemans et al.,
2004) : entraver le passage d'informations non appropriés,
empêcher l'exécution d'une réponse prédominante et
empécher le passage d'informations devenues non
pertinentes. Les mécanismes inhibiteurs seraient
fonctionnellement indépendants (Connely & Hasher, 1993 ; in
Meulemans et al., 2004).
Pour des raisons historiques (Bianchi, 1895; Harlow, 1968;
Luria, 1966; Shallice, 1982 ; in Andrés, 2003), un lien
spécifique entre inhibition et cortex frontal a été
fortement suggéré en neuropsychologie (Andrés, 2003).
2.1.1.2. La relation lobe frontal-inhibition.
Les lésions des lobes frontaux affectent un
système qui est fortement impliqué dans l'aptitude à
inhiber ou supprimer les stimuli ou des associations qui ne sont pas
pertinentes pour la tâche à accomplir (Dempster, 1991).
Des données en PET scan indiquent une
sensibilité à l'interférence accrue dans le cortex
préfrontal (Jonides et al., 1998 ; in Thompson-Schill, Jonides,
Marshuetz, Smith, D'Esposito, Kan, Knight, & Swick., 2002 ).
Des études indiquent la présence de troubles
(par exemple pour l'imitation, Brass et al., 2003 ; in Van Leeuwen, Van Baaren,
Martin, Dijksterhuis & Bekkering, 2009) chez des patients frontaux mettant
en avant un déficit d'inhibition. Mais encore, les lésions
frontales ont été reliées à un fonctionnement
inhibiteur réduit (Passingham, 1993 ; Smith & Jonides, 1999 ; in Van
Leeuwen et al., 2009).
Ainsi, lorsqu'il demande au patient d'imprimer à sa
main tour à tour trois dispositions (main fermée, paume sur le
bureau et main tendue placée sur le côté), Luria (1978 ; in
Boujon, 2002) constate que les patients avec lésion frontale peuvent
reproduire de façon répétitive un méme
élément de la série. Quand il demande à ces
patients de faire un dessin d'un cercle et d'une croix à tour de
rôle, ceux-ci parviennent à recopier le premier dessin de
manière itérative, sans changement entre les dessins. Luria
déduit de cette inclination à s'obstiner la présence d'un
problème dans l'inhibition des routines d'actions automatisées
(Meulemans et al., 2004).
2.1.1.3. Inhibition contrôlée et automatique.
John Hughling Jackson est à l'origine, à la fin
du 19ème siècle, d'une théorie novatrice sur
les déficits liés à un dommage cérébral.
D'après cet auteur, les déficits liés à une
lésion se focalisent sur les aspects les plus complexes alors que les
plus simples et donc les plus automatiques sont relativement
préservés (Gagnon & Goulet, 1992 ; in Siéroff, 2003).
Par la
suite, il a été exposé à diverses
reprises que les atteintes cérébrales perturbent peu les
caractéristiques les plus simples d'une fonction (par exemple,
dénommer la couleur de l'encre dans une tâche Stroop) alors que
les plus élevés semblent plus affectés (Siéroff,
2003).
Il existe donc une distinction pour certains auteurs
(Arbuthnott, 1995 ; Popp & Kipp, 1998 ; in Meulemans et al., 2004) entre
mécanismes intentionnels ou contrôlés et involontaires ou
automatiques. Une tâche d'inhibition contrôlée se
caractérise par l'aptitude à inhiber volontairement des
réponses prépondérantes ou spontanées lorsque c'est
requis. Ce type d'inhibition, de nature exécutive, est réduit
à la suppression consciente de réponses non appropriées.
Une tâche d'inhibition automatique nécessite des mécanismes
inhibiteurs se manifestent en dehors de la conscience. Ce type d'inhibition est
une réaction résiduelle automatique de traitement de
l'information appropriée. On la nomme généralement
inhibition réactionnelle (Meulemans et al., 2004).
2.1.1.4. Caractéristiques de l'inhibition
En 1975, Posner & Snyder mettaient en avant que
l'inhibition est un mécanisme actif de suppression. Ils ont mis en
évidence que pour qu'une action ou une information pertinente soit
sélectionnée, il est nécessaire que les actions ou
informations inappropriées soient inhibées. L'inhibition dans la
sélection s'exécuterait en fonction des caractères des
facteurs distracteurs et ceci pour permettre l'efficience des
procédés de traitement de la cible. Cette vision n'a pas toujours
été unanime, en effet Gathercole et Broadbent, 1987 (in Boujon,
2002) ont présagé que la baisse des interférences dues aux
distracteurs serait liée à un affaiblissement passif dans le
temps d'activation de leurs représentations.
L'inhibition est un phénomène adaptatif, son
effet sur le temps de réaction est lié, comme l'activation,
à la pertinence de l'information véhiculée par le stimulus
qui permet la réalisation de l'action (nommé indice ou amorce).
Quand l'indiçage apporte une contribution notable pour la
réalisation de l'action (fréquence concordante dans 80% des cas),
l'effet commun d'inhibition et de facilitation survient. Selon Posner et
Snyder, 1975 lorsque la fréquence de validité de l'indice baisse,
l'effet d'inhibition se dissipe. Ceci indique que l'effet d'inhibition est
adaptatif, volontaire et actif.
L'inhibition est un mécanisme central de suppression qui
intervient dans de nombreuses
situations expérimentales, aussi bien
la dénomination d'objets que la décision lexicale ou
encore
des épreuves de classification (Neill & Westberry, 1987). En effet,
elle peut se
manifester y compris lors de changements physiques ou
sémantiques du stimulus ou de la réponse.
L'initialisation est lente mais durable, elle prend entre 50
et 100ms, ce qui est lent comparativement à l'activation mais ce
phénomène dure quelques secondes (Neill & Westberry, 1987)
Ungerleider et Mishkin, 1992 (in Smith & Kosslyn, 2009)
ont démontré en induisant des lésions
cérébrales chez le singe qu'il y a 2 phénomènes qui
interviennent dans la sélection : le phénomène « what
» qui consiste à identifier la cible et le phénomène
« where » qui consiste à la localiser. La localisation serait
encodée précocement, laisse libre cours au processus
d'identification (Boujon, 2002).
Une variation quelconque de direction du mouvement au cours de
cette période entraîne un retard pour identifier la cible, c'est
ce qu'on appelle le phénomène d'inhibition de retour (IOR)
indiquant que l'inhibition se centrerait sur l'objet.
Robertson et al., 2004 (in Zhang & Zhang, 2006) ont
suggéré cependant que cet effet pourrait être
réinterprété en tant que processus spatiaux de
réorientation. Les résultats de Zhang et Zhang, 2006 vont dans ce
sens et indiquent que l'IOR basée sur l'objet dans les affichages
dynamiques résulterait davantage de la mise à jour de
l'inhibition spatiale plutôt que l'inhibition d'objet.
L'étape sur laquelle agirait l'inhibition
relèverait du but fixé par la tâche. Peu importe la
modalité perceptive des stimuli ou des réponses, l'effet
d'inhibition se maintient et reste similaire lors du maintien d'une
modalité ou du passage d'une à une autre. Lorsque la consigne est
définie selon un acte à effectuer au sein de la tâche, les
changements n'interviennent plus sur l'effet inhibiteur (Banich, 2004).
2.2. Evaluation
- L'amorçage négatif
On présente des paires de lettres successives
écrites en couleurs différentes. Le sujet doit lire à voix
haut une des deux lettres, toujours la lettre de la même couleur (par
exemple, celle écrite en rouge). Dans la condition non congruente, la
lettre que le sujet devait ignorer est maintenant la cible, la lettre qu'il
doit dénommer. On constate alors une augmentation du temps de
réaction (Le Rouzo, 2008).
- Le Stroop

Figure 3 : Le Stroop lecture
Figure 4 : Le Stroop interférence

Figure 2 : Le Stroop dénomination
Cette tâche se déroule en 3 parties. La
première partie est la dénomination : on présente une
feuille sur laquelle se trouvent plusieurs lignes de rectangles en couleur
(bleu, vert, rouge et jaune) dont le sujet doit donner la couleur (figure
2).
A la seconde étape, on présente des noms de
couleurs écrits en noir (bleu, vert, rouge et jaune) que le sujet doit
lire (figure 3).
La troisième étape est celle de
l'interférence. On présente des noms de couleurs écrits
(bleu, vert, rouge et jaune) dont la couleur d'impression ne correspond pas
à la couleur écrite. Le sujet doit donner la couleur dans
laquelle les mots sont écrits (figure 4). Il doit donc inhiber la
lecture du mot.
- Les réponses contrariées
Il s'agit ici de tester l'aptitude à instaurer et
entretenir un plan d'exécution pour un déplacement non
automatisé. L'examinateur effectue un geste, le sujet doit produire
l'inverse. Par exemple, l'examinateur tape sur la table une fois, le sujet doit
taper deux fois. En revanche si l'examinateur tape sur la table deux fois, le
sujet doit taper une fois.
- Le Hayling
Le Hayling vise à tester la capacité du sujet
à s'empêcher de donner une réponse dominante. (Peigneux,
2009-2010).
On lui présente des phrases dans lesquelles il manque le
dernier mot.
La première phase de la tâche pour le sujet est de
compléter par un mot congruent. Par exemple : « les arbres perdent
leurs... » on attend que le sujet réponde « feuille ».
Dans la seconde phase de la tâche, le sujet doit
compléter la phrase par un mot non lié sémantiquement.
Par exemple : « les chats ne font pas des... " le sujet
peut répondre « télévision " mais il ne peut pas
répondre « chien " qui est la réponse attendue ou «
éléphant " une réponse sémantiquement
liée.
- Les épreuves de type Go/No-Go
Diverses observations indiquent que des patients frontaux
éprouvent une difficulté à contrôler les mouvements
volontaires. Ces observations montrent que des patients lésés au
niveau préfrontal ne parviennent pas à inhiber leur
réponse motrice à un stimulus alors qu'il leur est demandé
de ne pas répondre pour ce stimulus. La tâche utilisée pour
ce type d'observation se nomme « Go-No Go ", elle a été
développée expérimentalement par Drewe en 1975. Ce dernier
a rapporté que des patients frontaux, plus précisément
avec lésion médiane, avaient significativement plus de
difficultés pour inhiber leur réponse à un ou deux stimuli
que des sujets ayant une lésion localisée ailleurs au sein du
cerveau. (Boujon, 2002).
2.3. Inhibition et stéréotypes
Seulement une poignée d'études ont pointé
la possibilité d'inhiber l'activation des stéréotypes et
plusieurs d'entre elles ont été discutées (Kunda &
Sinclair, 1999). Ce qu'il est intéressant de remarquer dans le cadre de
ce mémoire, c'est que l'inhibition semble bien avoir un impact sur les
stéréotypes et que si l'inhibition ne peut pas accomplir son
action, les stéréotypes paraissent s'exprimer librement. Ainsi,
des études sur la motivation indiquent qu'il serait possible dans
certains cas d'inhiber notre comportement stéréotypique (Macrae,
Bodenhausen & Milne, 1995 ; in Kunda et al., 1999).
L'inhibition cognitive et l'inhibition des
stéréotypes sont-ils un seul et même mécanisme ? La
question ne semble pas avoir passionné les chercheurs et la
littérature sur le sujet est inexistante.
Pour faire le parallèle entre ces deux notions que ce
sont l'inhibition cognitive et l'inhibition des stéréotypes, il
est intéressant de voir si elles suivent la même variation au
cours de la journée. A cette fin, nous allons nous intéresser
désormais aux rythmes.
3 Les rythmes
Les activités des êtres vivants se
déroulent de manière périodique, suivent un rythme et ont
lieu à des fréquences très variables. Ces rythmes
correspondent à une succession régulière ou
périodique de mouvements, de faits et de phénomènes (Bear,
Connors & Paradiso, 2007).
3.1. Les rythmes biologiques
Les rythmes dont nous parlons sont des rythmes biologiques qui
ne sont autres qu'une variation cyclique d'un phénomène
biologique (physiologie, croissance, comportement des animaux, etc.) dont la
période s'ajuste sur celle de phénomènes externes (Bear et
al., 2007). Ils vont de la milliseconde à plusieurs années. En
chronobiologie, on distingue entre les rythmes circadiens, infradiens,
ultradiens et circannuels.
Les rythmes nycthéméraux suivent la
révolution de la terre sur elle-même et permettent l'adaptation de
l'organisme aux changements quotidiens de luminosité. En l'absence
d'information environnementale, certains vont persister, présentant une
période proche de 24h (entre 20 et 28h), ces rythmes sont dits
circadiens (Achard, 2003).
Les auteurs qualifient d'ultradiens tous les rythmes d'une
période inférieure à 20 heures (fréquence
cardiaque, stades de sommeil..) et d'infradiens (cycle menstruel...) tous ceux
d'une période supérieure à 28 heures (Beugnet-Lambert,
1988). Les rythmes circannuels ont une période d'une année (cycle
migratoire des oiseaux, hibernation...) (Gwinner, 1977).
Quatre paramètres sont à prendre en compte pour
caractériser un rythme biologique : la période, l'acrophase,
l'amplitude et le niveau moyen. La période est la durée totale
d'un cycle complet (en secondes, minutes, heures...). L'acrophase, dont
l'opposé est la batyphase, correspond au pic de la variable
mesurée au cours du temps. L'amplitude représente la
variabilité de la valeur biologique mesurée sur une
période considérée. Le niveau moyen (ou MESOR) n'est autre
que la moyenne des mesures de la variable (Estruch, 2003).
3.2. Régulation homéostatique
L'homéostasie a été définie
dès 1932 par Cannon comme le processus de coordination physiologique qui
maintient l'état de veille dans l'organisme. Le processus
homéostatique a fréquemment été traduit comme une
fatigue cellulaire ou métabolique (Kong et al., 2002 ; in Mongrain,
2006). Ce mécanisme augmente la propension au sommeil sur base du temps
passé éveillé et réduit la propension à
l'endormissement en réponse à un sommeil prolongé
(Borbely & Achermann, 2005 ; in Schmidt, 2009). Le
processus homéostatique augmente exponentiellement au cours de
l'éveil et diminue de la méme façon durant le sommeil : il
reflète le besoin de dormir et l'intensité du sommeil (Daan et
al., 1984 ; in Mongrain, 2006).
3.3. Les rythmes circadiens
Les premiers écrits sur les rythmes circadiens
remontent à l'Antiquité avec le scribe Androsthènes qui
décrivit le comportement nycthéméral des feuilles du
tamarinier, celles-ci s'ouvraient le jour et se refermaient la nuit.
Au XVIème siècle le physiologiste et
médecin italien Santorio constata une variation journalière du
poids corporel. Il mit ainsi en évidence pour la première fois la
présence d'un rythme circadien chez l'Homme (Levett & Agarwal,
1979).
3.3.1. Endogènes
En 1729, Ortous de Mairan découvre que le mimosa,
méme lorsqu'il est placé dans l'obscurité permanente,
continue à ouvrir et fermer ses feuilles. Il montra ainsi que les
rythmes circadiens sont endogènes puisqu'ils perdurent en l'absence
d'indices environnementaux.
En 1832, Augustin de Candolle découvrit que les
feuilles s'ouvraient une à deux heures plus tôt chaque jour et
acquéraient par conséquent une périodicité de 22
à 23 heures. Il en conclut qu'il existe une tendance inhérente
aux plantes à montrer un mouvement périodique. Il avait ainsi mis
en évidence l'existence d'une période endogène en cours
libre ("free running period") proche, mais pas forcément
égale, à la période du cycle jour/nuit.
Aschoff et Wever (1962) enfermèrent des volontaires
dans une cave en absence d'indices environnementaux. Siffre (1964), lui,
vécut seul dans une caverne souterraine pendant deux mois. Ces deux
expériences ont révélé l'apparition
spontanée d'une période en cours libre du cycle
activité/repos d'environ 25 h, indiquant que les humains, comme les
plantes et les animaux, possédaient un système circadien
endogène (Gonze, s.d.).
Aujourd'hui, on considère par définition que les
rythmes circadiens sont endogènes. Dans des conditions constantes, ils
se déroulent librement avec une période qui dévie
légèrement du cycle lumière-obscurité de 24h
auxquels ils sont synchronisés naturellement (Schmidt, 2009).
3.3.2. Génétiquement déterminés
En 1935, Erwin Bunning démontre que la période
en cours libre du rythme circadien des plantes et des insectes est
héritée génétiquement, en croisant des parents de
périodes endogènes différentes.
Depuis une dizaine d'années, plusieurs gènes
impliqués dans l'horloge circadienne interne ont pu être
caractérisés. Le premier organisme génétiquement
modifié présentant un rythme circadien d'activité
altéré est la mouche (Konopka & Benzer, 1971). Les premiers
gènes horloge ("clock genes") ont été
détectés chez la mouche (Drosophilia) et le champignon
(Neurospora), leur étude a permis de mettre en évidence
les régulations qui confèrent à ces gènes une
expression circadienne. Par la suite, de nombreux "clock genes" ont
été trouvés chez les cyanobactéries, les plantes
(Arabidopsis) et les mammifères, y compris l'Homme (Gonze,
s.d.).
Des travaux de Linkowski, 1999 sur des jumeaux
suggèrent que certains composants du sommeil humain pourraient
être génétiquement déterminés. Ils ont
notamment trouvé un effet génétique fortement significatif
concernant les stades 2 et 4 de sommeil ainsi qu'en sommeil delta. Selon les
mêmes auteurs certains aspects de la rythmicité circadienne,
notamment l'expression endogène de cortisol, semblent très
similaires chez des jumeaux monozygotes mais pas chez des dizygotes,
suggérant que dans la plupart des organismes, les facteurs
génétiques contrôlent la génération des
rythmes circadiens (Linkowski, 1993 ; in Schmidt, 2009).
3.3.3. Synchronisés
Les rythmes circadiens sont certes endogènes mais ils
sont également resynchronisés en fonction de facteurs externes
appelés Zeitgebers ou synchroniseurs. Les Zeitgebers sont des agents ou
évènements environnementaux qui donnent des indices sur l'heure
biologique. Chez l'humain, le synchroniseur le plus important est le cycle
jour-nuit mais il en existe d'autres comme les obligations sociales (heure de
coucher, heure du repas...). La synchronisation peut être
considérée comme un état dans lequel deux oscillateurs ou
plus ont la même fréquence à cause de leurs influences
mutuelles ou unilatérales.
Pour que la synchronisation s'effectue, elle nécessite
un processus d'entraînement (adaptation
du rythme biologique à
un synchroniseur) qui consiste au couplage d'un rythme endogène
à
un cycle externe (ou zeitgeber), comme le cycle jour-nuit ou le
cycle de température, avec le résultat que ces deux oscillations
ont la même fréquence (Schmidt, 2009).
3.4. Interaction entre rythmes circadiens et
régulation homéostatique
Il existe deux mécanismes qui sous-tendent la
régulation du cycle veille-sommeil. Ces deux processus sont
conceptualisés dans le modèle de Borbely et al., 1982 (Borbely et
al., 1982 ; in Achermann, 2004) (figure 5).
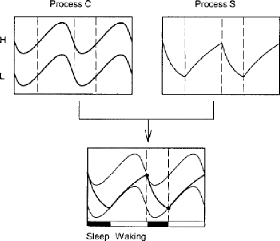
Figure 5 : Représentation schématique du
modèle deux processus de régulation du sommeil de Borbely et al.,
1982 (Achermann, 2004)
Ce modèle se base sur l'interaction entre deux
processus : le processus circadien (C) et le processus homéostatique (S)
qui agissent en synchronie ou en opposition le long du cycle de 24h.
Le processus homéostatique (S) est défini comme
le processus de promotion du sommeil. Au cours du processus S les performances
cognitives et l'alerte diminuent et on constate un accroissement de la fatigue.
Durant le sommeil, le processus S diminue faisant retomber la pression de
sommeil (Schmidt, Colette, Cajochen, & Peigneux, 2007).
Le processus circadien (C) correspond aux variations
oscillatoires pendant 24h de la propension au sommeil. La propension au sommeil
basée sur le processus circadien est à son niveau le plus bas
durant les premières heures du soir et atteint son maximum au petit
matin, quand la pression homéostatique est basse (Van Dongen &
Dinges, 2003 ; in Schmidt et al., 2007). Ce processus est dépendant du
temps, indépendant du sommeil ou de l'éveil, module
deux seuils (H et L) qui déterminent le début et la
fin des épisodes de sommeil, respectivement (Achermann, 2004).
3.5. Paradigmes d'exploration
Il existe actuellement plusieurs procédés pour
étudier les variations des rythmes circadiens. On trouve parmi ceux-ci
le paradigme de désynchronisation forcée, le paradigme de routine
constante et enfin une investigation basée sur le chronotype. Ces
paradigmes diffèrent considérablement dans la façon dont
ils contrôlent les paramètres circadiens et
homéostatiques.
3.5.1. Désynchronisation forcée
Il existe un paradigme qui permet de rompre le synchronisme
entre les mécanismes C et S afin de déterminer l'impact de chacun
de ces deux processus. Le paradigme de désynchronisation forcée
consiste à isoler des sujets des indices temporels et de les soumettre
à des durées de journées modifiées. Un exemple de
cette technique est celui de Nathaniel Kleitman et Bruce Richardson en 1938 qui
ont passé 32 jours enfermés dans une grotte du Kentucky (figure
6).
Figure 6 : Kleitman et Richardson enfermés 32 jours dans
une grotte
Le processus circadien ne reçoit plus d'indices
temporels (variations de luminosité,
activitéextérieure), il ne peut dès lors plus
suivre le rythme journalier et suit son propre rythme
(Schmidt et al., 2007).
Cette technique très difficile à mettre en place
notamment pour des raisons de temps, de
moyens et de précautions
éthiques n'est pas appropriée dans le cadre de ce travail et ne
sera
1
http://www.archiveofsleep.com/post/4494298875/cave-experiment-by-nathaniel-kleitman-and-bruce
dès lors pas utilisée. Le paradigme que nous
allons utiliser (expliqué plus loin) ne permet pas de
différencier entre le processus homéostatique et circadien chez
nos sujets. Cependant, bien que les bases neuro-anatomiques et fonctionnelles
de ces deux processus semblent différentes, ils participent tous deux
à la régulation du sommeil et à la performance en
éveil (Schmidt, 2009).
3.5.2. Routine constante
Le paradigme de routine constante permet de démasquer
les rythmes circadiens endogènes, normalement incorporés dans le
cycle veille-sommeil. Les activités quotidiennes, qu'elles soient
d'origine interne ou externe ont le potentiel de masquer les « vrais
» rythmes endogènes.
Initialement, le paradigme de routine constante est
défini par des conditions environnementales constantes. L'idée
qui sous-tend ce paradigme est que la suppression des cycles périodiques
extérieurs (ex : variation de la lumière) suffit pour
découvrir la contribution du processus circadien. Ainsi, les
études à long terme sur les animaux se font avec une
lumière constante ou encore dans la pénombre de manière
constante.
Les études sur les humains ont
révélé que le contrôle des cycles périodiques
extérieurs ne suffit pas à révéler la contribution
directe de l'horloge circadienne. Les protocoles incluent désormais le
contrôle du comportement. Ainsi, on prive les sujets de sommeil, on
contrôle la position, la prise de nourriture, les activités. Pour
observer un cycle circadien complet, une routine constante doit durer
suffisamment longtemps pour que les changements résiduels se dissipent.
Les routines constantes durent donc plus de 24h (Duffy & Dijk, 2002).
La limitation majeure de cette approche est qu'elle ne permet
pas de séparer la composante circadienne du cycle veille-sommeil
(Schmidt et al., 2007).
3.5.3. Chronotype
Chez les humains, les rythmes circadiens ont été
mis en évidence depuis 1939 (Kleitman, 1963 ; in Mongrain, 2006). Il y a
des personnes dites du matin (matinales), qui vont se coucher relativement
tôt et qui ont davantage tendance à être actif le matin,
d'autres personnes sont dites du soir (vespérales), elles se
lèvent relativement plus tard et privilégient donc une
activité plus tard dans la journée.
La phase circadienne se manifeste environ deux heures plus
tôt chez les sujets ayant un chronotype du matin par rapport à des
sujets vespéraux aussi bien au niveau physiologique que
comportemental.
La régulation homéostatique réagirait
également au chronotype. Les sujets matinaux dissipent plus vite la
pression homéostatique que les sujets du soir au cours du sommeil. A
l'éveil au contraire ils accumulent la pression homéostatique
plus rapidement que les vespéraux.
Ces types circadiens peuvent être identifiés par
questionnaires et fournir des informations sur les caractéristiques
circadienne et homéostatique du cycle veille/sommeil d'un sujet
(Taillard, 2009).
Dans l'état actuel des choses, deux questionnaires
prédominent la littérature dans l'étude du chronotype
(Schmidt et al., 2007), le plus populaire est le Morningness-Eveningness
Questionnaire (MEQ, Horne & Östberg, 1976), l'autre est le Munich
Chronotype Questionnaire (MCTQ, Roenneberg, Wirz-Justice, & Merrow,
2003).
Dans une perspective neuropsychologique, une limitation
majeure de l'utilisation des protocoles de désynchronisation
forcée et de routine constante réside dans la mise en place de
tâches cognitives qui doivent être administrées à
intervalle régulier. Dans cette optique, il semble que l'approche
basée sur le chronotype soit davantage appropriée à
l'étude de l'influence de l'horloge biologique sur les variations de
performance dans les fonctions cognitives de haut-niveau (Schmidt et al.,
2007).
Le questionnaire de Horne et Östberg est le plus
répandu pour mesurer le chronotype chez l'adulte. Le sujet est
interrogé sur son état de fatigue et ses moments
préférés de la journée pour la réalisation
d'activités, pour les repas ou encore son cycle/veille sommeil au
travers de 19 questions. Les scores du questionnaire vont de 16 à 86, le
sujet est considéré comme extrême du soir si son score est
en deçà de 31 et comme extrême du matin si son score est
audelà de 69.
Un nombre conséquent d'études ont
suggéré que la performance circadienne à des tests
cognitifs est largement dépendant de paramètres
spécifiques de la tâche, incluant notamment la durée et la
difficulté, la méthode d'administration et les variables
mesurées (Bonnet, 2005 ; in Schmidt, 2009). La performance cognitive ne
serait pas uniquement déterminée par l'activation des
systèmes de régulation sous-jacents mais elle serait
également modulée par des mécanismes compensatoires tels
que des facteurs motivationnels ou les attentes vis-à-vis de
l'expérience (Schmidt, 2009).
L'utilisation d'un protocole standardisé et hautement
contrôlé (routines constantes et désynchronisation
forcée) ainsi qu'une tâche de vigilance (PVT) ont permis de
quantifier l'homéostasie éveil-sommeil et les effets
dépendant des rythmes circadiens sur la cognition humaine (Blatter &
Cajochen, 2006).
3.5.4. Contraintes méthodologiques
La performance cognitive englobe les réponses
comportementales à des tâches de complexité
différente, aussi bien au niveau de la réaction psychomotrice
qu'au niveau de réponses nécessitant des fonctions cognitives de
haut niveau (par exemple mémoire, langage, fonctions exécutives).
Ces tâches sont clairement différenciées de l'humeur, de la
fatigue et du sentiment d'endormissement bien qu'elles y soient
corrélées. D'un point de vue méthodologique, mesurer les
rythmes circadiens dans la performance humaine est plus complexe que de mesurer
les rythmes circadiens de température ou la sécrétion
pinéale de mélatonine.
Deux problèmes principaux apparaissent dans la mesure
de la performance cognitive circadienne : le type de tâches
utilisé et les différences inter-individuelles de performance
à une tâche (Blatter & Cajochen, 2006)
3.5.4.1. Difficulté de la tâche
A première vue, il semblerait que la complexité
de tâche ne joue pas un rôle crucial dans la recherche circadienne.
La PVT (ou tâche de vigilance psychomotrice), très sensible aux
baisses de performances dues au manque de sommeil et à la variation
circadienne peut difficilement être considérée comme une
tâche complexe. Des tâches plus complexes testant les fonctions
exécutives sont susceptibles également d'être sensibles
à la perte de sommeil et à la phase circadienne.
Cependant, les résultats d'un des tests
neuropsychologiques les plus utilisés, notamment dans les processus
inhibiteurs, le test Stroop couleur de mots, montre des effets inconsistants
d'une étude à l'autre, indiquant un effet du mot de la
journée dans certaines études et pas dans d'autres. Une
explication pour ces résultats divergents est que le contrôle
exécutif n'est pas un processus unitaire mais plutôt des processus
indépendants qui sont reliés. La déprivation de sommeil et
le moment de la journée n'affecteraient dès lors que certaines
composantes du système exécutif (Blatter & Cajochen,
2006).
3.5.4.2. Différences inter-individuelles
On dénote des différences inter-individuelles
marquées dans plusieurs aspects circadiens et reliés au sommeil
de la physiologie tel que la longueur de la période circadienne (de
23,9h à 24,5h), dans le chronotype (matinal, vespéral), la
durée de sommeil ou encore la perte de sommeil. La plupart de ces
aspects sont liés à l'âge, aux traits de
personnalité.
Des études ont révélé que les
individus diffèrent dans leur propension au sommeil ainsi que dans la
baisse de la performance cognitive observée pendant une privation de
sommeil.
3.5.4.3. Inertie de sommeil
L'inertie de sommeil est une période suivant le
réveil suivant laquelle l'attention et la performance des sujets sont
affaiblies de telle sorte que le niveau d'éveil normal n'est pas
rencontré (Balkin & Badia, 1988 ; in Silva & Duffy, 2008).
Silva et Duffy, 2008 testent des adultes âgés
(plus ou moins 65 ans) au travers d'une tâche DSST au travers d'un
paradigme de désynchronisation (13,33h d'éveil et 6,67h de
sommeil par « jour »). La tâche DSST comprend des paires de
nombres-symboles (ex: 1/+, 2/ Ë) suivi d'une liste de chiffres dans
laquelle le sujet doit compléter par le symbole correspondant. Ils
constatent que la performance des sujets s'améliore au fur et à
mesure de leur éveil.
3.6. Relation avec les fonctions cognitives
Les oscillations circadiennes semblent induire de nombreuses
variations dans les performances cognitives, c'est le cas par exemple de la
vigilance (Adam, Rétey, Khatami & Landolt, 2006), l'alerte (Van
Dongen & Dinges, 2000), la mémoire de travail (Folkard et al., 1994;
in Schmidt, 2009) ou encore la mémoire déclarative (Cajochen et
al., 1999 ; in Schmidt, 2009).
L'évaluation de l'effet du moment de la journée
sur les performances cognitives a commencé par des mesures
psychophysiologiques. Nathaniel Kleitman, pionnier dans la recherche sur les
rythmes circadiens constata une variation diurne dans la vitesse et
l'exactitude de la performance cognitive, la meilleure performance étant
l'après-midi et la plus mauvaise tôt le matin ou tard dans la nuit
(Kleitman, 1933 ; in Blatter & Cajochen, 2006). Il mit également
en évidence que ces variations sont associées au
rythme diurne de la température corporelle et qu'un changement
spontané ou induit de celle-ci engendrait un ralentissement du temps de
réaction.
Quarante ans plus tard, en 1979, Aschoff et Wever
utilisèrent le paradigme de désynchronisation forcée. Ce
fut la première mesure de la performance circadienne et la
première évidence que les rythmes circadiens et le cycle
veille-sommeil contribuent tous deux à la variation de la performance
cognitive (Blatter & Cajochen, 2006).
Une étude de Taillard, Philip, Claustrat, Capelli,
Coste, Chaumet et Sagaspe (2011) met en évidence que les sujets du soir
parviennent à maintenir une alerte optimale tout au long de la nuit
alors que les sujets du matin n'y parviennent pas. Pour les deux chronotypes,
la performance circadienne est corrélée à la somnolence
subjective.
Des variations circadiennes ont été
observées également sur la mémoire de travail. Rouch,
Wild, Ansiau et Marquié (2005) constatent que les travailleurs en
équipe qui ont des horaires de travail fluctuants ont de moins bonnes
performances dans le rappel immédiat que les travailleurs ayant des
horaires fixes. Leurs résultats suggèrent que cette baisse de
performance n'est pas tant le fruit de troubles de sommeil mais davantage d'une
désynchronisation des rythmes circadiens. Rowe, Hasher et Turcotte
(2009) comparent des sujets matinaux âgés à des sujets
vespéraux âgés dans une tâche de mémoire de
travail visuo-spatiale (empan de Corsi). La tâche leur est
présentée soit en ordre ascendant (sets de plus en plus grands)
ou descendant (sets les plus longs en premier). Dans la condition descendante,
ils observent que les matinaux âgés ont une performance bien
meilleure lorsqu'ils sont au pic de leur journée.
3.7. Chronotype et inhibition
Habituellement, on considère que des sujets
testés au moment optimal de leur journée ont de meilleures
performances comparés à des sujets au moment non optimal, c'est
l'effet de synchronie (May & Hasher, 1998 ; in Schmidt, 2009). Des
études suggèrent que la synchronie entre les périodes
d'éveil circadien importe vraiment dans des tâches cognitives
contrôlées, principalement dans des tâches inhibitrices
telles que stop-signal ou go/no-go (Schmidt, 2009).
Hasher et ses collègues (Hasher, Lustig & Zachs
2008) se sont intéressés aux processus inhibiteurs et leurs
résultats mettent en avant que la nature des fonctions inhibitrices
varie au fur et à mesure de la journée en fonction du
modèle circadien individuel. Ils se sont intéressés
à 3 processus de la fonction inhibitrice : l'accès, la
délétion et la répression.
Le processus d'accès de la fonction inhibitrice permet
de déterminer parmi les représentations activées laquelle
est au centre de l'attention. Une étude de May, 1990 (in Hasher, Lustig
& Zachs, 2008) propose une tâche dans laquelle les sujets doivent
générer un mot (ex : fromage) à partir de 3 mots non
reliés (ex : rat, bleu et chalet). Les sujets sont soumis à une
distraction au cours de l'expérience. La performance des sujets du matin
est affectée le matin et l'aprèsmidi par le distracteur mais
davantage l'après-midi. Les sujets du soir ont une bonne performance
optimale le soir mais leur performance est affectée le matin.
Le processus de délétion est critique pour
supprimer les informations non pertinentes du focus attentionnel. On
présente une phrase avec un mot manquant prédictible (ex : blanc
bonnet et bonnet ...) qui est complété par le participant puis
remplacé quelques secondes plus tard par un mot moins concordant (ex :
miteux). Le mot utilisé par le participant devient non-pertinent. Les
expérimentateurs mesurent ensuite le temps d'accès au mot
non-pertinent. Chez les sujets du soir, l'après-midi, le mot
non-pertinent est supprimé de sorte que son utilisation de nouveau est
au-dessous du niveau de base. Le matin en revanche, il est au-dessus du niveau
de base. Les sujets du matin montrent également une variation
circadienne avec une performance plus mauvaise l'après-midi.
Le processus de répression est le mécanisme de
contrôle des réponses. C'est le mécanisme actuellement le
plus utilisé selon Miyake et al., 2000 (in Hasher et al., 2008) et que
l'on réfère directement à l'inhibition. Il est
étudié à travers des tâches comme le Stroop ou le
Go-NoGo. Le nombre d'erreurs commis par les sujets dans une tâche de type
Go-NoGo apparait plus important au moment non-optimal de la journée
aussi bien pour les sujets du matin que du soir.
3.8. Chronotype et stéréotypes
Bodenhausen, 1990 fait l'hypothèse que les variations
circadiennes du niveau d'éveil seraient liées à la
tendance à développer des stéréotypes sociaux en
vertu des effets sur la motivation et la capacité de traitement. Son
hypothèse a été confirmée puisque les sujets ont
montré davantage de biais stéréotypiques dans leur
jugement au moment non optimal de la journée (le matin pour les sujets
vespéraux, le soir pour les sujets du matin). Ces résultats se
retrouvent
dans un jugement de probabilités concernant les
caractéristiques d'une personne et dans la perception de la
culpabilité suite à des allégations de mauvaise conduite
chez des étudiants. Ces résultats suggèrent que les
processus biologiques doivent être considérés dans les
tentatives de conceptualiser les déterminants de
stéréotypes.
|
Questions de recherche et hypothèses
|
37
|
Questions de recherche
Nous allons nous intéresser dans un premier temps
à la réplicabilité des résultats de Bodenhausen,
1990 aussi bien au niveau explicite qu'implicite en contrôlant l'inertie
de sommeil. Rappelons que celui-ci a mis en évidence que l'activation
des stéréotypes semble influencée par les rythmes
circadiens au moins de manière explicite.
Nous tenterons également de vérifier s'il existe
des variations circadiennes dans une tâche d'inhibition cognitive telle
que le GoNoGo. Cette tâche semble en effet faire l'objet de variations
circadiennes (Schmidt, 2009).
Nous nous interrogerons si les variations obtenues sont la
conséquence d'une variation de la vigilance.
Enfin, nous étudierons s'il existe une corrélation
entre les mesures portant sur le jugement social et l'inhibition cognitive.
Hypothèses
H1 : Les stéréotypes varient en fonction du moment
de la journée.
Bodenhausen, 1990 constate que les variations circadiennes du
niveau d'éveil seraient reliées à la propension à
activer les stéréotypes sociaux. Notre hypothèse est qu'au
niveau explicite, en contrôlant l'inertie de sommeil, on ne constate pas
d'effet en raison d'un biais de désirabilité sociale. En
revanche, au niveau implicite, y compris en contrôlant l'effet d'inertie
de sommeil, nous nous attendons à obtenir le même type de
résultats que Bodenhausen.
H2 : L'inhibition varie en fonction du moment de la
journée
Des études précédentes (Hasher et al.,
2008) indiquent que les variations circadiennes ont une influence sur les
performances inhibitrices. Notre hypothèse est que les performances en
inhibition, et plus particulièrement dans une tâche de type GoNoGo
(cf. Schmidt, 2009), sont moins bonnes lorsque la passation est
effectuée au moment non-optimal de la journée (matin pour les
vespéraux, soir pour les matinaux).
Si l'inhibition ne peut jouer son rôle alors
l'expression des stéréotypes semble s'exprimer pleinement (Kunda
et al., 1999). L'hypothèse que nous formulons ici est simple, plus la
performance en tâche GoNoGo est mauvaise, plus l'expression des
stéréotypes sera forte et inversement.
II. Méthode
Sélection de la population
MEQ (Morning Evaluation Questionnaire)
Ce questionnaire (annexe 1) a été administré
afin de sélectionner la population.
Celui-ci investigue les caractéristiques de sommeil
personnelles, si le sujet est du matin ou du soir. En fonction du score obtenu
au questionnaire le sujet est classé vespéral ou matinal.
La procédure est la suivante :
1. Le questionnaire est posté en ligne
2. Des affiches sont posées et une large diffusion est
effectuée sur les réseaux personnels.
3. Le sujet est classé dans le groupe vespéral ou
matinal selon le score obtenu au questionnaire.
4. On propose deux dates de passation et un horaire au sujet.
Graphique 1 : répartition du chronotype dans
l'échantillon
805 personnes ont répondu au questionnaire et le
chronotype se répartit de manière gaussienne (Graphique 1). L'age
moyen de la population ayant répondu à ce questionnaire est de
36,83 ans et le score moyen est de 52, 94, ce qui équivaut à un
score neutre, légèrement du matin. Les sujets extrêmes du
matin, ayant un score supérieur ou égal à 70,
représentent 3,97% de la population testée. Les sujets
extrêmes du soir, ayant un score inférieur ou égal à
31, représentent 4,46% de la population testée.
Sur base de ce questionnaire, 2 groupes ont été
formés : un groupe composé de 24 sujets extremes du matin et un
groupe composé de 24 sujets extremes du soir. En raison d'une
difficulté pratique de recrutement, le score seuil pour les sujets
extrêmes du soir a été augmenté à 32.
L'age moyen des sujets extremes du matin qui ont
été testés est de 47,08 ans contre 33,75 ans pour les
sujets extremes du matin. Cette différence d'age entre les deux groupes
est significative (t(1,46)=-2.99 ; p=0,04) et n'a pas pu être
contrôlée de manière expérimentale. Ce facteur sera
donc intégré dans les analyses statistiques.
Plan expérimental
Comme dit dans la section précédente, il existe
deux groupes de participants : les extrêmes du matin ou « matinaux
» et les extrêmes du soir ou « vespéraux ».
Chaque sujet a été testé à 2
reprises : une fois au moment non-optimal (10h30 après le réveil
pour les matinaux, 1h30 après le réveil pour les
vespéraux), une fois au moment optimal (1h30 après le
réveil pour les matinaux, 10h30 après le réveil pour les
vespéraux).
Les sujets sont testés 1h30 et 10h30 après leur
lever afin de contrôler l'inertie de sommeil (Schmidt et al., 2009).
Ces passations ont été contre-balancées,
ainsi la moitié des sujets a commencé au moment optimal tandis
que l'autre moitié a commencé par la passation non-optimale.
La passation 1h30 après le lever a été
commencée en moyenne 1h27 après le lever et la condition 10h30
post-réveil a été commencée en moyenne 10h29
après le lever.
L'ordre de passation des tâches a été
contrôlé afin de s'assurer que l'ordre dans lequel le sujet
effectue les tâches n'a pas d'impact sur sa performance.
|
Ordre de passation
|
|
C1
|
PVT-IAT-GoNoGo
|
|
PVT-GoNoGo-IAT
|
|
C3
|
IAT-PVT-GoNoGo
|
|
C4
|
IAT-GoNoGo-PVT
|
|
C5
|
GoNoGo-IAT-PVT
|
|
C6
|
GoNoGo-PVT-IAT
|
Tableau 1 : Contre-balancement de l'ordre de passation
Ainsi chaque groupe a été réparti en 6
conditions, chaque condition correspondant à l'ordre dans lequel les
sujets passent les tâches (tableau 1).
En résumé, voici le plan expérimental
utilisé (tableau 2).
|
24 sujets matinaux
|
24 sujets vespéraux
|
|
C1
|
|
C3
|
C4
|
C5
|
C6
|
C1
|
|
C3
|
C4
|
C5
|
C6
|
|
Ordre de passation 1h30 ap lever puis 10h30 ap lever
|
1,2
|
3,4
|
5,6
|
7,8
|
9, 10
|
11,12
|
25,26
|
27,28
|
29, 30
|
31,32
|
33,34
|
35,36
|
|
Ordre de passation 10h30 ap lever puis 1h30 ap lever
|
13,14
|
15,16
|
17,18
|
19, 20
|
21,22
|
23,24
|
37,38
|
39, 40
|
41,42
|
43,44
|
45,46
|
47,48
|
Numéros de sujets Tableau 2 : résumé du plan
expérimental
Enfin, afin d'éviter un effet test-retest, il s'est
écoulé 7 à 10 jours (8,3 jours en moyenne) entre les 2
passations pour chaque sujet.
Expérience
Le sujet effectue 4 tâches : une tâche de type
Go/No-Go (inhibition), une tâche PVT (vigilance), une tâche IAT
(jugement social implicite) et une échelle explicite de
préjugés (jugement social explicite). Les 3 premières
tâches sont contrebalancées comme expliqué dans la partie
précédente et la 4ème tâche (échelle
explicite) est toujours présentée en dernier. Le temps de
réalisation de l'ensemble de ces tâches lors d'une passation
avoisine les 45 minutes.
L'ensemble des tâches informatisées a
été réalisé sur un PC portable Dell Inspiron 6000
cadencé à 1,6GHz avec 1 Go RAM.
Tâche Go/No-Go (inhibition)
La tâche Go/No-Go utilisée est une tâche
informatisée dans laquelle le sujet doit appuyer quand il voit n'importe
quelle lettre à l'exception d'une (dans notre cas la lettre X) (figure
7). Cette tâche va permettre de mesurer la capacité du sujet
à inhiber une réponse prépotente. La tâche dure
approximativement 20 minutes.
Condition go Condition no-go
(appuyer lorsqu'une lettre apparait) (ne pas appuyer lorsque la
lettre X apparait)

Figure 7 : illustration de la tâche GoNoGo
La tâche utilisée est une traduction
française de celle utilisée dans l'article de Hinshaw, Greenhill,
Shafritz, Vitolo, Kotler, Jarrett et Glover, 2007 disponible sur le site
internet de l'institut Sackler2. La tâche a été
exécutée grâce au logiciel e-prime 2.0.8.89. Elle est
composée de 5 blocs comprenant chacun 129 items dont 25 items NoGo et
104 items Go.
Tâche de vigilance psychomotrice (PVT)
(vigilance)
Cette tâche créée par Dinges, 1985 (in
Schmidt, 2009) est également une tâche informatisée dans
laquelle le sujet doit répondre le plus vite possible lorsqu'il voit un
compteur défiler, sans pour autant anticiper sa réponse. Les
mesures prises en compte sont le temps de réaction (TR), le nombre
d'erreurs d'omission (TR de plus de 500 ms), le nombre de commissions (le sujet
appuie alors qu'il ne devrait pas, le temps de réaction est
inférieur à 100 ms) et la variabilité intra-sujet
(différence entre les temps de réaction les 10% plus rapides et
les 10% moins rapides). Ces mesures vont nous permettre de vérifier si
le sujet maintient sa vigilance lors des tâches proposées. Elle va
nous permettre également de vérifier également si le
maintien de la vigilance est équivalent au moment optimal et non
optimal. Cette tâche dure 10 minutes.
L'intervalle entre les stimuli est aléatoire entre 2 et
10s. Le sujet voit ainsi apparaitre approximativement 70 stimuli pendant la
réalisation de la tâche.
2
http://www.sacklerinstitute.org/cornell/assays_and_tools/
Tâche d'association implicite (IAT) (jugement social
implicite)
La tâche a été adaptée de la version
Racism IAT sur le site de Millisecond3. La tâche est
exécutée via le logiciel Inquisit 3.0.4.0.
La tâche est composée de 8 items cibles á
savoir 8 prénoms masculins, 4 prénoms á consonance
francophone (Pascal, Grégory, Damien, Ferdinand) et 4 prénoms
á consonance maghrébine (Mohamed, Ahmed, Ali, Youssef). Les
prénoms francophones et maghrébins ont été
associés sur base de leur fréquence dans la population
belge4. Les deux types d'attributs (Bon/Mauvais) comportent chacun 8
items (Bon : merveilleux, superbe, plaisir, beau, plaisant, glorieux, charmant,
magnifique / Mauvais : tragique, horrible, agonie, douloureux, terrible,
épouvantable, humiliant, méchant).
Lors de cette tâche, le sujet appuie chaque fois sur la
touche E pour associer la catégorie de gauche. Il appuie sur I pour la
catégorie de droite. Lorsqu'il commet une erreur, une croix rouge
apparait en dessous de l'item pour l'inviter à corriger.

Figure 8 : 2 concepts cibles

2 concepts cibles apparaissent dans une tâche á
deux choix (Maghrébin/Autre) (figure 8). Dans l'exemple ici, le sujet
voit apparaitre le prénom « Pascal » au centre de
l'écran, il appuie sur la touche correspondant á la
catégorie « Autre » (la touche E sur le clavier).
Le sujet continue á répondre avec les
mêmes touches á une seconde tâche qui concerne l'attribut
(Bon/Mauvais) (figure 9). Dans l'exemple, le sujet appuie sur la touche
correspondant á la catégorie « Mauvais » (touche I).
Figure 9 : attribut
|
Ensuite, l'attribut et les concepts cibles sont
mélangés. Ainsi, le sujet voit apparaitre une valeur de
l'attribut d'un côté (ex : Bon) ainsi qu'un concept cible (ex :
Maghrébin). Et de l'autre côté il voit apparaitre la
seconde valeur de l'attribut (ex : Mauvais) et le second concept cible (ex :
Mauvais). Au milieu de l'écran apparait soit un prénom
|
|
|
Figure 10.1 : conditions mélangées.
Prénom.
|
|
|
|
|
(figure 10.1) soit un mot (figure 10.2). Dans l'exemple de la
figure 10.1, le sujet appuie sur la touche correspondant à « Autre
» (touche I). Dans l'exemple de la figure 10.2, il appuie sur la touche
correspondant à « Mauvais » (touche I également).
|
|
|
Figure 10.2 : conditions mélangées. Mot.
|
La tâche est contre-balancée afin qu'elle ne
commence pas toujours par la méme association concept-attribut.
Pour l'analyse des résultats, nous porterons notre
intérét sur le score D. Il s'agit d'une mesure qui divise la
différence entre la moyenne des blocs tests par l'écart-type de
l'ensemble des temps de réaction des deux blocs. Les résultats de
Greenwald, Nosek et Banaji, 2003 indiquent que l'algorithme conventionnel de
l'IAT est dépassé par la mesure D pour (a) la magnitude de la
corrélation implicite-explicite, (b) la résistance à la
contamination par des réponses de vitesse différente, (c) la
résistance à l'effet de réduction du score IAT, (d) la
sensibilité aux effets connus sur les mesures IAT et (e) la latence
implicite explicite dans l'analyse factorielle confirmatoire.
Cette mesure D étant plus fine et plus sensible, c'est
elle que nous retiendrons pour l'analyse des résultats de l'IAT.
Nous nous attendons ici à ce que le score D soit de
manière générale positif, ce qui indiquerait une
préférence de l'échantillon pour la population
francophone. Mais surtout nous nous attendons à ce qu'au moment
non-optimal de leur journée les sujets aient un score D plus positif
qu'au moment non optimal. Cela indiquerait que la préférence pour
la population endogène (francophone) est plus marquée au moment
non-optimal de la journée.
Echelle explicite de préjugés (jugement social
explicite)
Dans cette échelle (annexe 2), on présente au sujet
des phrases ambiguës, une par une. A chacune de ses phrases, le sujet doit
répondre de « pas du tout d'accord » à « tout
à fait d'accord ».
Par exemple :
Les personnes d'origine maghrébine travaillent aussi dur
pour aller de l'avant que la plupart des autres Belges.
1. Tout a fait d'accord
2. Plutôt d'accord
3. Plutôt pas d'accord
4. Pas du tout d'accord
Gette tâche va permettre de déterminer si le
sujet a une propension plus importante au préjugé de
manière explicite au moment non-optimal de sa journée. Gette
tâche dure environ 5 minutes.
La méthodologie utilisée pour l'analyse de
l'échelle explicite repose sur l'article de Laïla Benraiss (2004).
L'échelle est composée de 6 questions construites sous forme
d'échelles de Likert à 4 positions. Il s'agit donc d'une
échelle de type ordinal où chaque item ne peut prendre que 4
valeurs. Nous effectuons une étude exploratoire afin de purifier
l'instrument de mesure. Le test de Bartlett teste l'hypothèse nulle de
corrélation des variables. Le seuil de risque est proche de 0 (p=0,000),
l'hypothèse des corrélations nulles peut être
rejetée. Ici, les données sont ainsi corrélées et
donc factorisables. L'indice KMO donné par le test de Kaiser, Meyer et
Olkin est de 0,575. Get indice et faible et peu acceptable.
L'alpha de Cronnbach permet de mesurer la fiabilité des
items à mesurer ce qu'ils sont censés mesurer. Lorsque l'alpha
des questions approche 1, la cohérence interne de l'échelle est
bonne, lorsqu'elle approche de 0, la cohérence est faible. Un alpha
acceptable est compris entre 0,6 et 0,7 (Nunnally, 1978 ; in Benraiss, 2004).
L'alpha de Gronnbach ici est de 0,522, ce qui est également faible.
Nous décidons dès lors de supprimer la question
5, ce qui porte l'indice KMO à 0,606 et l'alpha de Cronnbach à
0,650 qui sont des données acceptables. Un score composite est
calculé sur base des items retenus. Ge score correspond à la
moyenne des scores de chaque question.
III. Résultats
Tâche de vigilance psychomotrice (mesure de la
vigilance)
Pour la tâche de vigilance psychomotrice, des analyses
séparées ont porté sur le TR moyen pour les TRs > 100
ms et < 500 ms, le nombre d'omissions (TRs > 500ms), le nombre de
commissions (TRs < 100 ms) et la variabilité des TRs
(différence de TR moyen entre les 10% les plus rapides et les 10% les
plus lents).
Sur l'ensemble des sujets, le temps de réaction (TR)
moyen est de 333.24 ms (s.d. 38,83), le nombre d'omissions moyen est de 3.95
(s.d. 5,27), le nombre de commissions moyen est de 0.53 (s.d. 2,11) et la
variabilité moyenne est de 161.08 ms (s.d 20,32). Le sujet 18 a
été écarté des analyses parce que son TR est
au-delà de 2 écart-types.
Le test de Kolmogorov-Smirnoff nous indique que le TR moyen et
la variabilité sont distribuées normalement (p>0.20). Ce n'est
en revanche pas le cas des omissions et commissions (p<0.01).
Nous avons effectué une analyse de la variance à
mesures répétées pour le TR moyen et la variabilité
avec un facteur intra-groupe Optimalité (moment de test optimal vs.
non-optimal) et deux facteurs inter-groupes Chronotype (matinal vs.
vespéral) et ordre de Passation de la condition optimale (lors de la
1ère vs. lors de la 2nde passation), avec
l'âge en covariée.
âche de vigilance psychomotrice
Graphique 2 : effet significatif d'optimalité sur le temps
de réaction chez les matinaux et vespéraux
Les résultats de l'ANOVA sur les temps de
réaction moyen révèlent un effet d'interaction
Optimalité*Chronotype (F(1,43)=9,0639;p=0,004) (graphique 2). Tous les
autres effets sont non-significatifs (ps > .13). Les tests post-hoc
Tukey HSD montrent que le TR moyen est plus rapide au moment optimal que
non-optimal dans le groupe Vespéral (TR optimal 310 ms (s.d. 30) vs TR
non-optimal 327 ms (s.d. 36) ; p = .003), ce qui n'est pas le cas dans le
groupe Matinal (TR optimal 345 ms (s.d. 37) vs TR non-optimal 343 ms (s.d.
39,49) ; p = 0.97). Par ailleurs le TR moyen au moment optimal est plus rapide
dans le groupe Vespéral que dans le groupe Matinal (p = .0024). Les
autres effets ne sont pas significatifs (ps>0,20).
Aucun effet significatif n'a été observé
pour une anova similaire portant sur la variabilité du TR
(ps>0,24).
Les analyses effectuées pour les omissions et
commissions sont des tests non-paramétriques de type Wilcoxon pour
échantillons pairés. On constate une tendance à commettre
davantage de commissions au moment non-optimal chez les vespéraux
(Commissions optimal 0 (s.d. 0) vs Commissions non-optimal 0,21 (s.d. 0,51) ;
W=0,00 ; p=0,07). Tous les autres effets sont
Vespéraux
non-significatifs (ps>0,12).
Les moyennes et écart-types des mesures de la PVT sont
reprises dans le tableau 3 ci-dessous.
|
Matinaux
|
Vespéraux
|
|
Moyenne
|
Ecart-type
|
Moyenne
|
Ecart-type
|
|
TR Moyen Optimal
|
345,40
|
37,00
|
310,11
|
30,11
|
|
TR Moyen Non Optimal
|
343,29
|
39,50
|
326,65
|
36,06
|
|
Omissions Optimal
|
4,83
|
3,88
|
1,92
|
2,24
|
|
Omissions Non Optimal
|
3,83
|
2,90
|
2,79
|
3,89
|
|
Commissions Optimal
|
0,61
|
1,75
|
0,00
|
0,00
|
|
Commissions Non Optimal
|
1,39
|
4,52
|
0,21
|
0,51
|
|
Variabilité Optimal
|
167,76
|
30,21
|
156,38
|
22,60
|
|
Variabilité Non Optimal
|
158,63
|
23,37
|
163,42
|
25,15
|
Tableau 3 : Moyennes et écart-types dans les mesures de
la tâche de vigilance psychomotrice chez les matinaux et
vespéraux
GoNoGo (mesure de l'inhibition)
Malheureusement, une erreur lors de l'enregistrement des
données s'est produite pour cette tâche. Seules les données
de 13 sujets du matin ont été effectivement enregistrées
lors des deux passations ainsi que celles de 9 sujets du soir.
Pour la tâche GoNoGo, des analyses
séparées ont été effectuées sur le temps de
réaction (TR) moyen, le pourcentage d'appuis intempestifs, le
coefficient de variation et le TR moyen post-X. Une analyse portant sur la
comparaison TR moyen et TR moyen post-X a également été
réalisée. Les sujets 16,20 et 35 ont été
écartés des analyses de TR moyen et de TR moyen post-X (leur
performance étant au-delà de 2 écart-types).
Sur l'ensemble des sujets, le TR moyen est de 396,26 ms avec
un écart-type de 61,24ms le TR moyen post-X est de 385,93ms avec un
écart-type de 65,03, le pourcentage d'appuis intempestifs moyen est de
4,34%, et le coefficient de variation moyen est de 4,34. Les sujets 13 et 14
sont exclus de l'analyse pour les appuis intempestifs.
Le test de Kolmogorov-Smirnoff nous indique que le TR
médian, le nombre d'appuis intempestifs et le TR médian post-X
sont distribuées normalement (p>0.15). Ce n'est en revanche pas le
cas du coefficient de variation (p<0.01).
Nous avons effectué une analyse de la variance à
mesures répétées pour le TR moyen, le pourcentage d'appuis
intempestifs et le TR moyen post-X avec un facteur intra-groupe
Optimalité (moment de test optimal vs. non-optimal) et deux facteurs
intergroupes Chronotype (matinal vs. vespéral) et ordre de Passation de
la condition optimale (lors de la 1ère vs. lors de la
2nde passation), avec l'âge en covariée.
GoN
Graphique 3 : Effet non significatif d'optimalité sur le
TR moyen de la tftche GoNoGo chez les matinaux et vespéraux lors de la
première et seconde passation.
Les résultats de l'ANOVA sur le TR moyen
relèvent un effet significatif de l'interaction
Optimalité*Chronotype*Passation (F(1,15)=14,06;p=0,002) (graphique 3)
mais cet effet n'est pas confirmé lors des analyses à posteriori
(ps>0,09). On constate également un effet d'Optimalité
marginalement significatif pour le TR moyen post-X (F(1,17)=4,27 ; p=0,056) et
un effet d'interaction Optimalité*Chronotype*Passation (F(1,17)=11,62 ;
p=0,004) également non confirmés par les analyses à
posteriori (p>0,23). Tous les autres effets sont nonsignificatifs
(ps>0,11). Les analyses mettent en évidence un effet
d'Optimalité marginalement significatif pour le nombre d'appuis
intempestifs (F(1,17)=4,34 ; p=0,05) non confirmé par les analyses
à posteriori (p=0,09).
Les analyses effectuées pour le coefficient de variation
sont des tests non-paramétriques de type Wilcoxon pour
échantillons pairés. On ne constate aucun effet significatif
(ps>0,69).
p o op pa p ma
Nous avons effectué une analyse de la variance à
mesures répétées pour le TR moyen avec un
Mainaux
facteurs inter-groupes Chronotype (matinal vs. vespéral)
et ordre de Passation de la condition optimale (lors de la
1ère vs. lors de la 2nde passation), avec
l'âge en covariée.
Lorsque les sujets sont testés au moment non-optimal,
leur TR moyen post-X est significativement plus rapide que lors de l'ensemble
de la tâche (TR moyen post-X non-optimal= 370,72 vs TR moyen tâche
non-optimal=381,37 ; F(1,19)=12,19 ; p=0,003), confirmé par les analyses
à posteriori (p=0,0002). Au moment optimal, on constate un effet
significatif Groupe*Passation (F(1,16)=4,59 ; p=0,047) non confirmé par
les analyses à posteriori (ps>0,25). Tous les autres effets sont
non-significatifs (p>0,25).
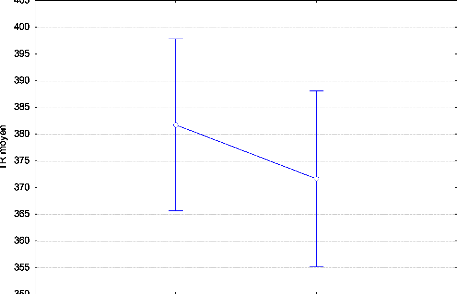
Graphique 4 : Effet significatif du temps de réaction
moyen lors de la tftche ou après un X pour l'ensemble des sujets.
Lorsque les sujets sont testés au moment non-optimal,
leur TR moyen post-X est significativement plus rapide que lors de l'ensemble
de la tâche (TR moyen post-X non-optimal= 370,72 vs TR moyen tâche
non-optimal=381,37 ; F(1,19)=12,19 ; p=0,003) (graphique 4), confirmé
par les analyses à posteriori (p=0,002). Au moment optimal, on constate
un effet significatif Groupe*Passation (F(1,16)=4,59 ; p=0,047) non
confirmé par les analyses à posteriori (ps>0,25). Tous les
autres effets sont non-significatifs (p>0,25).
Le tableau 4 ci-dessous reprend les moyennes et
écart-types des sujets matinaux et vespéraux pour les
différentes mesures de la tâche GoNoGo.
|
Matinaux
|
Vespéraux
|
|
Moyenne
|
Ecart-type
|
Moyenne
|
Ecart-type
|
|
TR Moyen Optimal
|
388,14
|
49,22
|
379,70
|
41,44
|
|
TR Moyen Non Optimal
|
384,52
|
42,94
|
376,96
|
39,13
|
|
TR Moyen Post-X Optimal
|
375,46
|
53,85
|
366,89
|
42,13
|
|
TR Moyen Post-X Non Optimal
|
373,31
|
45,38
|
367,11
|
40,24
|
|
Appuis intempestifs Optimal
|
4,51
|
3,46
|
4,24
|
2,66
|
|
Appuis intempestifs Non Optimal
|
5,44
|
3,38
|
4,55
|
2,85
|
|
Coefficient de variation Optimal
|
0,45
|
0,03
|
0,43
|
0,011
|
|
Coefficient de variation Non Optimal
|
0,42
|
0,12
|
0,43
|
0,018
|
Tableau 4 : Moyennes et écarts-types des mesures de la
tâche GoNoGo
71610101WWo6iOioOimSfi6iii (mesure
implicite des stéréotypes)
Pour la Tfiche d'Association Implicite (IAT), des analyses sont
effectuées sur le score D.
Sur l'ensemble des sujets, le score D moyen est de 0,65 (s.d.
0,41), ce qui indique une attitude plus positive envers la population
francophone. Le sujet 21 est exclu de l'analyse (score audelà de 2
écart-types).
Nous avons effectué une analyse de la variance à
mesures répétées pour le score D avec un facteur
intra-groupe Optimalité (moment de test optimal vs. non-optimal), deux
facteurs intergroupes Chronotype (matinal vs. vespéral) et ordre de
Passation de la condition optimale (lors de la 1ère vs. lors
de la 2nde passation), avec l'age en covariée.
p
Graphique 5 : effet d'optimalité non significatif sur le
score D de l'IAT chez les matinaux et vespéraux.
L'effet d'interaction Optimalité*Chronotype est
non-significatif (F(1,42)=0,73 ; p=0,40) (graphique 5). Tous les autres effets
sont également non-significatifs (ps>0,11).
Les moyennes et écarts-type de l'IAT chez les matinaux et
vespéraux sont reprises dans le tableau 5 ci-dessous.
|
Matinaux
|
Vespéraux
|
|
Moyenne
|
Ecart-type
|
Moyenne
|
Ecart-type
|
|
D-Score Optimal
|
0,62
|
0,39
|
0,47
|
0,57
|
|
D-Score Non Optimal
|
0,72
|
0,46
|
0,71
|
0,41
|
Tableau 5 : moyennes et écarts-types du D-score.
Echelle explicite de
préjugés
Pour l'échelle explicite de préjugés, des
analyses séparées sont réalisées pour le score
composite moyen.
Sur l'ensemble des sujets, le score moyen obtenu est de 2,08
(s.d. 0,44) ce qui correspond à
Vespéraux
p
un score légèrement favorable aux personnes
maghrébines (le score allant de 1 à 4, un score
de 2,5 correspondant à une attitude ni favorable ni
défavorable). Aucun sujet n'a été exclu de l'analyse.
Nous avons effectué une analyse de la variance à
mesures répétées pour le score D avec un facteur
intra-groupe Optimalité (moment de test optimal vs. non-optimal), deux
facteurs intergroupes Chronotype (matinal vs. vespéral) et ordre de
Passation de la condition optimale (lors de la 1ère vs. lors
de la 2nde passation), avec l'âge en covariée.
|
|
|
Graphique 6 : effet d'optimalité non-significatif sur le
score à l'échelle explicite chez les matinaux et
vespéraux.
|
L'effet d'interaction Optimalité*Chronotype est
non-significatif (F(1,43)=0,49 ; p=0,49). On constate un effet
d'optimalité (F(1,43)=4,95 ; p=0,03) non confirmé par les
analyses à posteriori (p=0,12). Tous les autres effets sont
également non-significatifs (ps>0,22).
Les moyennes et écart-types sont repris dans le tableau 6
ci-dessous.
|
Matinaux
|
Vespéraux
|
|
Moyenne
|
Ecart-type
|
Moyenne
|
Ecart-type
|
|
Score Optimal
|
2,00
|
0,42
|
2,13
|
0,47
|
|
Score Non Optimal
|
2,06
|
0,47
|
2,15
|
0,44
|
Tableau 6 : Score moyen à l'échelle explicite
Correlations
Nous avons porté notre attention sur le lien entre la
vigilance et les capacités inhibitrices. Les conclusions de ces analyses
devront toutefois être tempérées par le fait que des
données sont manquantes pour la tâche d'inhibition suite à
un problème technique. Le TR moyen et les omissions de la tâche de
vigilance psychomotrice sont significativement corrélés à
la moyenne de la tâche GoNoGo et la moyenne post-X (0,56<r<0,59
optimal; 0,47<r<0.69 non-optimal ; p<0,05). Le nombre de commissions
est corrélé significativement au coefficient de variation de la
tâche GoNoGo au moment non-optimal (r=0,49 ; p<0,05).
Nous avons aussi focalisé notre regard sur la relation
entre la vigilance et l'activation de stéréotypes. Au niveau
implicite, le nombre de commissions est corrélé significativement
au score D de la tâche d'association implicite lors de la passation
optimale (r=0,56 ; p<0,05). Au niveau explicite, les corrélations ne
sont pas significatives (0<r<0,24).
Nous nous sommes également intéressés
à la relation existant entre les capacités d'inhibition telles
qu'évaluées dans la tâche Go-NoGo et l'activation de
stéréotypes. Au niveau implicite, la corrélation entre les
indices d'inhibition et le score D de l'IAT ne sont pas significatives
(0,03<r<0,09). Au niveau explicite, la moyenne de la tâche GoNoGo
est significativement corrélée au score explicite dans la
condition optimale (r=0,45;p<0,05).
Enfin, nous avons porté notre intérét sur
le lien entre l'activation implicite et explicite des
stéréotypes. Les corrélations entre l'échelle
explicite et le score D de l'IAT ne sont pas corrélées
significativement (0,07<r<0,42).
IV. Discussion
Rappel des hypothèses
L'objectif principal de ce travail a été
d'étudier le lien entre les rythmes circadiens et le jugement social.
Nous avons tenté de lier ces deux aspects au travers de l'inhibition
cognitive. Autrement dit, nous avons procédé en 3 temps : 1. Voir
s'il existe un lien entre la performance en inhibition et le rythme circadien.
2. Voir si la performance inhibitrice est liée au jugement social 3.Voir
si ces trois notions sont liées entre elles (ex : au moment non optimal
de ma journée, j'ai plus de difficultés à inhiber mes
réponses et j'active davantage mes stéréotypes).
A cette fin, nous avons recruté deux groupes de
participants sur base d'un questionnaire de chronotype : les sujets
extrêmes du soir et les sujets extrêmes du matin. Ces sujets ont
été vu au moment optimal (le matin pour les matinaux, le soir
pour les vespéraux) et non-optimal de leur journée (le soir pour
les matinaux, le matin pour les vespéraux). L'ordre de passation
(optimal puis non-optimal vs non-optimal puis optimal) a été
contrebalancé et il s'est écoulé 7 à 10 jours entre
les deux passations.
Les sujets ont été testés au travers de
quatre tâches contre-balancées, l'une d'entre elles est la
tâche de vigilance psychomotrice qui permet de s'assurer du maintien de
leur attention durant l'expérience. La tâche de GoNoGo est une
seconde tâche à laquelle les sujets ont été
confrontés, elle mesure la capacité des sujets à inhiber
la réponse à un élément dominant. La
troisième tâche est l'IAT ou tâche d'association implicite
au cours de laquelle on mesure la préférence implicite des sujets
pour une population (maghrébin/francophone). Enfin, ils ont
été testés au travers d'une échelle explicite de
préjugés afin de voir s'il existait une différence dans la
réponse exprimée au niveau implicite et explicite par les sujets
sur les mêmes populations.
Nous nous attendions dans un premier temps à ce que nos
résultats sur la tâche de vigilance indiquent un temps de
réaction plus rapide lors de la condition optimale uniquement chez les
sujets du soir. Nous nous attendions également pour cette tâche
à obtenir des temps de réaction plus rapides pour les sujets du
soir que du matin.
Là où nous nous attendions à ce que la
vigilance réagisse au facteur circadien, notre attente se
portait
également sur des variations circadiennes dans les autres tâches.
Ainsi, la première de
nos hypothèses est qu'au moment
non-optimal de leur journée, tous les sujets (matinaux et
vespéraux) éprouvent davantage de
difficultés à inhiber leurs stéréotypes au niveau
implicite mais non au niveau explicite. Nous nous attendons donc à ce
que nos sujets révèlent une préférence plus
importante pour la population francophone (endogroupe) que pour la population
maghrébine (exogroupe) au moment non-optimal de leur journée.
Cette préférence devrait se marquer par un score D plus important
dans la tâche d'association implicite au moment non-optimal. En revanche,
nous nous attendons à ne constater aucune différence sur
l'échelle explicite en raison d'un biais de désirabilité
sociale.
La seconde hypothèse est que l'inhibition cognitive
subirait le méme sort et que l'inhibition serait moins bonne au moment
non-optimal de la journée. Nous nous attendons ici à ce que nos
sujets appuient plus souvent lorsque cela ne leur est pas demandé (appui
intempestif) dans la condition non-optimale. Cela se traduirait par un
pourcentage d'appuis intempestifs plus importants dans la tâche
GoNoGo.
La troisième et dernière hypothèse
rejoint les deux premières et postule que l'inhibition des
stéréotypes serait influencée par l'inhibition cognitive.
Notre attente ici est que les sujets aient des performances fortement
corrélées aussi bien au moment optimal qu'au moment non-optimal
de leur journée entre la tâche GoNoGo et la tâche
d'association implicite. Ceci se traduirait par un nombre d'appuis intempestifs
et un score D congruents (moment non-optimal : nombreux appuis intempestifs,
score D élevé / moment optimal : peu d'appuis intempestifs, score
D faible).
Résultats principaux et interprétation
Désormais, nous allons présenter les
différents résultats de cette étude et soumettre des
interprétations en lien avec la partie théorique de cette
recherche.
Les données seront présentées successivement
puis nous nous intéresserons aux liens existant entre ces
données.
Tâche de vigilance psychomotrice
Nous allons tout d'abord nous intéresser à la
mesure contrôle de cette étude, la vigilance, au travers des
différentes conditions à savoir le groupe d'appartenance (matinal
vs vespéral) et le moment de la passation (optimal vs non-optimal).
Concernant la variabilité (différence entre les temps de
réaction les 10% les plus rapides et les 10% les moins rapides), les
omissions (>500ms) et les commissions (<100ms) nous ne constatons aucune
différence significative de l'effet du moment (optimal vs non-optimal)
aussi bien pour les sujets du matin que pour les sujets du soir. En revanche,
là où l'on ne constate pas de différence significative
concernant le temps de réaction chez les sujets du matin, les sujets du
soir sont significativement plus rapides le soir.
Ensuite, concernant les différences de groupe, nous ne
constatons aucune différence entre les sujets matinaux pour la
variabilité, les omissions et les commissions. En revanche, les sujets
du soir sont significativement plus rapides que les sujets du matin. Bien que
l'on constate un effet significatif de l'age, ces effets se maintiennent
lorsque l'age est contrôlé.
En réalisant les contrastes locaux pour les
différentes mesures, voilà ce que nous obtenons :
(1) Pas d'effet pour les mesures de variabilité,
omissions et commissions.
(2) Pas d'effet de moment (optimal vs non-optimal) pour les
sujets matinaux
(3) Effet significatif de moment (optimal vs non-optimal) sur le
temps de réaction pour les sujets vespéraux
(4) Effet significatif de groupe (matinal vs vespéral)
sur le temps de réaction
Le maintien du nombre d'omissions, commissions et de la
variabilité nous indiquent que les sujets focalisent leur attention avec
autant de consistance au moment optimal que non-optimal. Cette absence d'effet
nous invite donc à penser que les sujets ne sont pas plus distraits au
moment optimal ou non-optimal de leur journée.
Nous aurions pu nous attendre à ce que les sujets du
matin, tout comme les sujets du soir, aient une performance significativement
plus lente au moment non-optimal de leur journée à savoir le
soir. Une explication quelque peu intuitive de ce phénomène
réside dans le profil des sujets extremes du matin. En effet, ce type de
profil est généralement la résultante d'un emploi qui
impose des horaires matinaux. Ces sujets ont ainsi des journées
régulières, qui sont « normalisées » depuis de
nombreuses années pour certains. Ces sujets, lorsqu'ils sont
éveillés depuis 10h30 sont testés en
milieu d'après-midi, moment où dans leur vie quotidienne, ils
sont autant susceptibles d'être aussi actifs qu'à leur lever. Les
sujets vespéraux sont généralement inscrits dans un mode
de vie qui ne les contraint pas à être actifs dès leur
lever, qu'ils soient jeunes et joueurs de jeux vidéo ou qu'ils
travaillent de nuit. Leur performance le matin s'en trouve dès lors
ralentie. Un point de vue inverse, lui aussi intuitif, serait d'envisager que
le moment optimal des matinaux est en réalité un moment
contraint. De par leur emploi, ils ont été obligés de
devenir productifs le matin. Avec le temps, leur performance le matin s'est
améliorée pour atteindre la performance moyenne de leur
journée. Schmidt et al., 2009 ont testé des sujets au travers
d'une tâche de vigilance psychomotrice et obtiennent des résultats
similaires à ceux obtenus dans notre étude. En effet, les sujets
du soir sont plus rapides le soir mais on ne constate pas de différence
chez les sujets du matin. Dans cette études, les auteurs mettent en
évidence une activation plus importante chez les vespéraux
notamment dans le locus coerulus et le noyau suprachiasmatique (aires
importantes dans les rythmes circadiennes). Cette activation
cérébrale plus importante chez les vespéraux pourrait
être à l'origine de leur meilleure performance le soir.
L'explication la plus cohérente que nous pouvons
apporter concernant la stabilité de la performance chez les matinaux
réside dans l'atténuation circadienne chez les plus
âgés. Aussi bien dans notre étude que de manière
générale, les sujets extrêmes du matin sont
généralement plus âgés que les sujets extrêmes
du soir. Or, on sait que l'age joue un rôle sur le fonctionnement
circadien et qu'avec l'age celui-ci tend à s'aplanir. Une étude
d'Adam et al.,2006 met notamment en évidence que lors d'une privation de
sommeil, la performance des âgés tend à rester beaucoup
plus stable que celle des plus jeunes qui s'effondre.
Tâche GoNoGo (mesure de l'inhibition)
A nouveau, nous avons comparé la performance des sujets
matinaux et vespéraux au moment optimal et non optimal de leur
journée. Les résultats que nous avons obtenus sont contraires
à notre hypothèse : quel que soit le groupe testé, le
temps de réaction et le nombre d'appuis intempestifs ne varient pas. En
effet, nous n'avons pas mis en évidence d'effet significatif du moment
(optimal vs non-optimal) aussi bien pour les matinaux que les vespéraux.
Ces résultats vont à l'encontre de la littérature (May
& Hasher, 1998 ; in Hasher et al., 2008). En effet, May et Hasher
constatent que les sujets du matin produisent globalement plus d'erreurs
et que tous les sujets (matinaux ou vespéraux) effectuent
davantage d'erreurs au moment non-optimal.
Nous restons prudents quant aux données obtenues dans
cette tâche car plusieurs biais ont contaminé la tâche. Le
premier d'entre eux et probablement le plus important est que de nombreuses
données n'ont pas été enregistrées par le
programme, ce qui réduit de manière importante le nombre de
sujets et dès lors la probabilité d'obtenir un effet. De
surcroit, cela n'a pas affecté de manière égale le nombre
de sujets matinaux et vespéraux créant une
inégalité entre les groupes. Ensuite, le contre-balancement de
l'ordre de passation des tâches a lui aussi été
affecté par ce nombre de sujets manquants.
Ensuite, la longueur de la tâche n'était
probablement pas adaptée au testing. En effet, la tâche durait 20
à 25 minutes, ce qui s'est avéré particulièrement
long pour de nombreux sujets qui ont manifesté leur
désintérêt. Ce biais a probablement créé une
sorte d'effet plancher avec une performance dans l'ensemble des conditions
davantage régie par un désintérét de la tâche
que par un effet d'optimalité. Blatter et Cajochen, 2006 mettent en
garde contre cet effet de durée et recommandent l'utilisation de
tâches de vigilance courtes (5 à 10 minutes) lors d'une
déprivation de sommeil, le sujet devenant trop distractible par la
suite.
La corrélation significative entre le temps de
réaction moyen de la PVT et celui du GoNoGo nous invite cependant
à considérer que la performance obtenue par les sujets dont le
score est enregistré est interprétable. L'inhibition est une
fonction cognitive de haut niveau, plus exigeante qu'une tâche de
vigilance. On peut imaginer que de par l'effort cognitif nécessaire, les
sujets sont davantage affectés par la difficulté de la
tâche que par un éventuel effet du moment de la journée.
Ces résultats ne vont cependant pas dans le sens de la
littérature. Manly, Lewis, Robertson, Watson et Datta, 2002 (in Schmidt,
2007) constatent un important effet du moment de la journée, les sujets
étant moins efficients tôt le matin qu'en début
d'après-midi et le soir.
Nous comparons désormais le temps de réaction
à l'ensemble de la tâche à celui obtenu après un X
(élément pour lequel il ne faut pas appuyer). Nous aurions eu
tendance à penser que le sujet devant inhiber sa réponse, il se
prépare à la réponse suivante, à l'inhiber de
nouveau et son temps de réaction s'en trouve ralenti. Pourtant au moment
optimal de la passation, il n'y a pas d'effet significatif et le sujet se
prépare de la même manière à un item suivant un X ou
non. Là où nous obtenons un résultat quelque peu
surprenant, c'est qu'au moment non-optimal, les sujets répondent plus
rapidement après avoir vu un X. Une réponse qui pourrait
être apportée à ce résultat est que
l'attention du sujet étant captée par un élément
inhabituel, il focalise davantage son attention et son potentiel de
préparation en est alors accru. Sa vigilance étant plus grande,
il met moins de temps à répondre à l'élément
qui suit un X. (Je viens de voir un X, je me suis fait/j'aurais pu me faire
piéger, je reste bien attentif !). Buschman et Miller, 2007 ont mis en
évidence une activation cérébrale lors de la
présentation d'un distracteur qui serait dû à une attention
de type bottom-up. Dans notre expérience, nous pouvons penser que le
sujet traite l'ensemble de la tâche de manière top-down et le
stimulus X lui apparait comme un « distracteur » qui vient capter son
attention sur un mode bottom-up. Le sujet, sous l'influence de ce distracteur
focalise alors son attention au cas où un nouveau distracteur
apparaitrait. Ce phénomène ne se produit qu'au moment non-optimal
de la journée, le sujet étant plus à même de
réagir à un « distracteur ».
Echelle explicite
Pour cette tâche, nous avons également
comparé la performance des sujets matinaux et vespéraux à
leur moment optimal et non-optimal. Les résultats que nous avons obtenu
confirment notre hypothèse : quel que soit le groupe testé et le
moment, les sujets ne manifestent pas de variations dans la mesure des
préjugés au niveau explicite. Nous n'avons pas mis en avant
d'effet significatif du moment (optimal vs non-optimal) aussi bien pour les
matinaux que les vespéraux. Ces résultats vont à
l'encontre de la littérature (Bodenhausen, 1990). En effet, les
résultats de Bodenhausen indiquent que les sujets émettent un
jugement social plus défavorable au moment non-optimal de leur
journée.
La première explication que nous pourrions accorder
à ce résultat viendrait d'un effet de récupération
des items de l'échelle. L'échelle est composée de 5 items
et nous pourrions penser que les sujets se souviennent de leur réponse
d'une passation à l'autre, effaçant ainsi l'effet du moment de la
passation. Cependant, en espaçant les passations de 7 à 10 jours
nous pouvons considérer que ce facteur de mémorisation soit
supprimé puisque les sujets n'ont normalement pas effectué un
encodage en mémoire à long terme.
La seconde explication viendrait de la sensibilité de
l'échelle. Les sujets sont susceptibles de manière socialement
désirable et une échelle à 4 niveaux (tout à fait
d'accord à pas du tout d'accord) n'est pas suffisamment sensible pour
détecter un changement de réponse. Les deux premières
explications indiquent dès lors qu'il aurait été
probablement préférable d'utiliser
une tâche plus complexe comme celle de Bodenhausen, 1990
qui utilise une tâche de jugement sur un texte.
Une autre vision de ce résultat viendrait du fait que
les résultats obtenus par Bodenhausen sont liés à
l'inertie de sommeil et la pression homéostatique. En effet, tous les
sujets testés par cet auteur sont testés à 9h du matin et
à 20h. Les sujets extrêmes du soir ne sont normalement pas encore
réveillés à 9h du matin et cela entraine chez eux un effet
de fatigue qui pourrait expliquer leur performance plus mauvaise. Les sujets
extrêmes du matin quant à eux, sont pour certains levés
depuis 15h lorsqu'ils passent la tâche à 20h, ils sont donc soumis
à une pression de sommeil importante qui elle aussi nuance leurs
résultats.
Un compromis entre ces explications résiderait donc
dans l'utilisation d'une tâche moins sujette à l'effet de
désirabilité sociale que les sujets passent 1h30 et 10h30
après le lever. L'analyse de la prochaine tâche, la tâche
d'association implicite devrait donc nous renseigner davantage à ce
sujet.
Tâche d'association implicite (IAT)
Nous avons testé ici si les sujets manifestent une
préférence pour une population (francophone/maghrébine)
plus importante en fonction du moment auquel ils sont testés. Tout
d'abord l'échantillon testé présente de manière
générale une légère préférence pour
la population francophone. Cette préférence, comme nous pouvions
nous y attendre, ne diffère pas significativement entre les sujets du
matin et du soir.
En revanche, notre hypothèse n'est pas
confirmée. Là où nous pensions qu'au moment non-optimal la
préférence serait plus marquée au niveau implicite pour la
population francophone nous ne constatons aucune différence
significative aussi bien pour les sujets du matin que du soir. Si les
résultats obtenus par Bodenhausen, 1990 n'étaient pas liés
à l'inertie et la pression de sommeil, nous nous attendrions à
obtenir le même type de résultats.
Nous pourrions suspecter un manque de sensibilité du
score D mais les résultats utilisant l'algorithme classique de l'IAT
vont dans le méme sens. Nous pourrions également inférer
la présence de biais méthodologique tels que les bruits
environnants au domicile mais cela devrait également influencer la
performance dans la tâche de vigilance.
Nos résultats semblent dès lors indiquer que les
résultats obtenus par Bodenhausen sont
davantage le reflet de
l'inertie et la pression de sommeil que d'un effet circadien. Afin de s'en
assurer, il serait nécessaire de faire passer les
mêmes tâches que Bodenhausen en contrôlant cette fois-ci
l'inertie et la pression de sommeil.
L'activation des stéréotypes ne semble pas
reliée à la vigilance. En effet, là où nos sujets
du soir sont plus performants le soir, l'activation de leurs
stéréotypes, elle, ne semble pas varier aussi bien au niveau
implicite qu'explicite. Les résultats obtenus concernant l'inhibition
nous invitent à être prudents mais en se penchant sur la
littérature (Hasher et al., 2008), la performance des sujets du matin
est moins bonne le soir. On ne constate cependant pas dans notre étude
de variation dans l'activation des stéréotypes. On pourrait
dès lors penser que ces notions ne sont pas directement reliées.
Nous restons cependant prudents car les auteurs ne s'accordent pas quant
à la présence d'effets circadiens dans la tâche GoNoGo
(Blatter & Cajochen, 2006).
Dans la continuité de cette étude, il serait
pertinent de tester les sujets dès leur réveil au travers de la
méme tâche d'association implicite (IAT). Si des résultats
apparaissaient lors de cette expérience, cela nous inviterait alors
à repenser les résultats de Bodenhausen en mettant en avant que
des résultats se produisent au lever en raison d'une inertie de sommeil
mais que lorsque ce facteur est contrôlé ces résultats
tendent à disparaitre.
Bibliographie
Articles
Achermann, P. (2004). The two-process Model of Sleep Regulation
Revisited. Aviat Space Environ Med, 75(3, Suppl.) , A37-43.
Adam, M., Rétey, V.J., Khatami, R. & Landolt, H.P
(2006). Age-Related Changes in the Time Course of Vigilant Attention During 40
Hours Without Sleep in Men. Sleep, 29(1), 55- 57.
Banaji, M. & Hardin, C.D. (1996) Automatic stereotyping.
Psychological Science, 7(3)136- 141.
Benraiss, L. (2004). Méthologie de construction d'une
échelle de mesure: application du paradigm de Churchill. Communication
dans le cadre du XVème Congrès Annuel de l'Association
Francophone de la Gestion des Ressources Humaines (AGRH), du 1er au 4
septembre, Montréal (Canada).
Blatter, K. & Cajochen, C. (2006). Circadian rhythms in
cognitive performance: Methodological constraints, protocols, theoretical
underpinnings. Physiology & Behavior, 90(2-3), 196-208.
Bodenhausen, G.V. (1990). Stereotypes as judgmental heuristics:
evidence of circadian variations in discrimination. Psychological Science
1(5), 319-322.
Buschman, T.J. & Miller, E.K. (2007). Top-Down Versus
Bottom-Up Control of Attention in the Prefrontal and Posterior Parietal
Cortices. Science 315 (5820), 1860-1862.
Correll, J., Park, B., Judd, C.M. & Wittenbrink, B. (2007).
The influence of stereotypes on decisions to shoot. European Journal of
Social Psychology, 37, 1102-1117.
Devine, P. & Monteith, M. J. (1999). Automaticity and Control
in Stereotyping. In Dual-process theories in social psychology. éd
Chaiken, Shelly, 339-360.
Doosje, B., Spears, R., De Redelijkheid, H. & Van Onna, J.
(2007). Memory for stereotype (in)consistent information: The role of in-group
identification. British Journal of Social Psychology , 46, 115-128.
Duffy, J.F. & Dijk, D.J. (2002). Getting through to circadian
oscillators: why use constant routines? Journal of Biological Rythms
17(1), 4-13.
Greenwald, A., Nosek, A.B. & Banaji, M.R.
(2003).Understanding and Using the Implicit Association Test: I. An Improved
Scoring Algorythm. Journal of Personality and Social Psychology 85(2),
197-216.
Gwinner, E. (1977). Circannual rhythms in bird migration. Annual
Review of Ecology and Systematics, 12, 381-405.
Hinshaw, S.P., Greenhill, L.L, Shafritz, K.M., Vitolo, A.,
Kotler, L.A., Jarrett, M.A. & Glover, G., 2007. Frontostriatal Connectivity
and Its Role in Cognitive
Control in Parent-Child Dyads With ADHD. American Journal of
Psychiatry 164, 1729-1736.
Koch, U. & Magnusson, A.K. (2009). Unconventional GABA
release: mechanisms and function. Current Opinion in Neurobiology, 19,
305-310.
Konopka, R.J & Benzer, S. (1971). Clock Mutants of
Drosophilia Melanogaster. Proceedings of the National Academy of Sciences
of the United States of America, 68 (9), 2112-2116.
Horne, J.A & Östberg, O. (1976). A self-assessment
questionnaire to determine morningesseveningness in human circadian
rhythms. International Journal of Chronobiology, 4, 97-110
Kotowicz, Z. (2007). The strange case of Phineas Gage.
History of the human sciences, 20(1), 115-131.
Kunda, Z. & Sinclair, L. (1999). Motivated Reasoning With
Stereotypes: Activation, Application, and Inhibition. Psychology Inquiry
10(1), 12-22.
Levett, J. & Agarwal, G. (1979). The first man/machine
interaction in medicine: the pulsilogium of Sanctorius. Medical
Instrumentation 13(1), 61-63.
Linkowski, P. (1999). Sleep Patterns in twins. J Sleep
Res, 8 Suppl 1, 11-13.
Moors, A. & De Houwer, J. (2006). Automaticity: A Theoretical
and Conceptual Analysis. Psychological bulletin, 132(2), 297-326.
Mrug, S., Hoza, B. (2007) Impression formation and modifiability:
Testing a theoretical model. Merrill-Palmer Quarterly: Journal of
Developmental Psychology, 53(4), 631-659.
Neill & Westberry (1987). Selective Attention and the
Suppression of Cognitive Noise. Journal of Experimental Psychology 13
(2), 327-334.
Riggio, H.R. & Garcia, A.L. (2009). The Power of Situations:
Jonestown and the Fundamental Attribution Error. Teaching of Psychology,
36, 108-112.
Rouchy, I., Wildy, P., Ansiau, D. & Marquié, J.C.
(2005). Shiftwork experience, age and cognitive performance. Ergonomics
48 (10), 1282 - 1293.
Rowe,G., Hasher, L. & Turcotte,J. (2009).Age and synchrony
effects in visuospatial working memory. The Quartely Journal of
Experimental Psychology 62 (10), 1873-1880.
Roenneberg, T., Wirz-Justice, A. & Merrow, M. (2003). Life
between Clocks: Daily Temporal Patterns of Humans Chronotypes. Journal of
Biological Rhythms, 18 (1), 80-90.
Schmidt,C., Colette,F., Leclercq,Y., Sterpenich, V.,
Vandewalle,G., Berthomier,P., Berthomier,C.,
Philipps,C.,Tinguely,G.,Darsaud,A.,Gais,S.,Schabus,M.,Desseilles,M.,DangVu,T.T.,Salmon,E.,Balteau,E.,Degueldre,D.,Luxen,A.,Maquet,P.,Cajochen,C.&
Peigneux,P. (2009).Homeostatic Sleep Pressure and Responses to Sustained
Attention in the Suprachiasmatic Area. Science 324, 516-518.
Silva, E.J. & Duffy, J.F. (2008). Sleep Inertia Varies With
Circadian Phase and Sleep Stage in Older Adults. Behavorial Neuroscience
122 (4), 928-935.
Taillard, J. (2009). L'évaluation du chronotype en
clinique du sommeil. Médecine du sommeil, 6, 31-34.
Thompson-Schill, S.L., Jonides, J., Marshuetz, C., Smith,
E.E., D'Esposito, M., Kan, I.P., Knight, R. T & Swick, D. (2002). Effects
of frontal lobe damage on interference effects in working memory.
Cognitive, Affective, & Behavioral Neuroscience, 2 (2), 109-120
Van Dongen, H.P.A. & Donge, D. (2000). Circadian Rhythms
in Fatigue, Alertness and Performance. In: Kryger, M.H., Roth, T. & Dement,
W.D.C. (2000), Principles and Practice of Sleep Medicine (3rd ed.),
pp. 391--399.
Van Leeuwen, M.L, Van Baaren, R.B., Martin, D, Dijksterhuis,
A. & Bekkering, H. (2009). Executive functioning and imitation: Increasing
working memory load facilitates behavioural imitation. Neuropsychologia,
47, 3265-3270
Synthèses d'ILticlDs
Andrés, P. (2003). Frontal cortex as the central
executive of working memory: Time to revise our view. Cortex: A Journal
Devoted to the Study of the Nervous System and Behavior, 39(4- 5),
871-895.
Taillard, J., Philip, P., Claustrat, B., Capelli, A., Coste, O.,
Chaumet, G. & Sagaspe, P. (2011). Time Course of Neurobehavioral Alertness
During Extended Wakefulness in Morning- and Evening-Type Healthy Sleepers.
Chronobiology International 28 (6), 520-527.
Zhang, M. & Zhang, Y. (2006). Object Based-Inhibition of
Return in Dynamic Displays: Object Inhibition or Updating of Spatial
Inhibition? Acta Psychologica Sinica, 38(6), 798- 804.
Cours
Estruch, X. (2003). Applications de la chronobiologie à la
planification des cours et aux rythmes scolaires. Cours de Maîtrise
STAPS Université du Littoral-Côte d'Opale, Dunkerque.
Klein, O. (2008-2009). Cognition sociale. Cours de
3ème bachelier faculté des sciences psychologiques et
de l'éducation Université Libre de Bruxelles, Bruxelles.
Peigneux, P. (2009-2010). Neuropsychologie clinique. Cours de
Master 1 Neuropsychologie et développement cognitif Université
Libre de Bruxelles, Bruxelles.
Chapitres de livres
Banich, M.T. (2004). Chapter 11 Executive Function. In Cognitive
Neuroscience and Neuropsychology second edition. Boston: Houghton-Mifflin,
365-392.
Greenwald, A.G., McGhee, D.E. & Schwartz, J.L.K (2008).
Measuring Individual Differences in Implicit Cognition: The Implicit
Association Test. In Fazio, H.R. & Petty, R.E. Atttitudes Key Readings.
Their Structure, Function, and Consequences. Psychology Press, 109-
131.
Hasher, L., Lustig, C. & Zacks, R. (2008). Inhibitory
Mechanisms and the Control of Attention. In Conway A.R.A. Variation in working
memory. Oxford University Press.
Meunier, J.M. (2009). Chapitre 2 Les représentations. In
Mémoires, représentations et traitements. Dunod,
61-110.
Posner & Snyder (1975). Facilitation and Inhibition in the
Processing of Signals. In Rabbit, P.M.A & Dornic, S. Attention and
Performance V. Academic Press Inc, 669-682.
Siéroff, E. (2003). L'inhibition en neuropsychologie. In
L'inhibition. Ed S. Moutier, Paris, 37- 59.
Waroquier, L. & Klein, O. (2006). De la difficulté
de se débarrasser de ses habitudes mentales : les mécanismes
cognitifs impliqués dans la persistance des stéréotypes.
In Klein, O. & Pohls, S. Psychologie des stéréotypes et des
préjugés, Labor, Bruxelles.
Livres
Bear, M.F, Connors, B. & Paradiso, M.A (2007). Neurosciences,
à la découverte du cerveau. 3ème éd.,
éditions Pradel.
Bédard, L., Déziel, J. & Lamarche, L. (2006).
Introduction à la psychologie sociale vivre, penser et agir avec les
autres. 2ème édition. ERPI Sciences
humaines.
Beugnet-Lambert, C. Lancry, A., Leconte, P. (1988).
Chronopsychologie rythmes et activités humaines. Presses
Universitaires de Lille.
Bosche, M. (2007) Anthropologie Interculturelle : Immersion au
coeur d'un monde métissé. eBooks/OpenAnthropology.
Boujon, C. (2002). L'inhibition au carrefour des neurosciences et
des sciences de la cognition, fonctionnement normal et pathologique.
Solal.
Cannon, W.B. (1932). The wisdom of the body. W.W. Norton
& Company.
Légal, J-B. (2008). Stéréotypes,
préjugés et discrimination. éd. Dunod.
Meulemans, T., Collette, F. & Van der Linden, M. (2004).
Neuropsychologie des fonctions exécutives. éd. Solal.
Pradat-Diehl, P., Azouvi, P. & Brun, V. (2006). Fonctions
exécutives et rééducation. éd. Masson.
Smith, E. & Kosslyn, S.M. (2009). Cognitive psychology, Mind
and brain. Pearson Education.
Ressources multimédia
Sites Internet
Degiorgio C, Fery P, Polus B, Watelet A. CRFNA. Comprendre les
fonctions exécutives
http://www.crfna.be/Portals/0/fonctions
exécutives.pdf
Didier Gonze. Unité de Chronobiologie Théorique.
http://student.ulb.ac.be/~dgonze/CLOCKS/history.html
Gouvernement fédéral belge
http://economie.fgov.be/fr/modules/publications/statistiques/population/prenoms_de_la_popul
ation_totale.jsp
Millisecond.
http://www.millisecond.com/download/samples/v3/IAT/
Sackler Institute.
http://www.sacklerinstitute.org/cornell/assays_and_tools/
Thèses et mémoires
Pichon C.L (2009). L'amorçage automatique de
comportements sociaux: mythe ou réalité. Mémoire
réalisé sous la direction de messieurs les professeurs Axel
Cleeremans et Olivier Klein en collaboration avec monsieur Stéphane
Doyen, doctorant, en vue de l'obtention du grade de Master en Sciences
Psychologiques
Schmidt, C. (2009). Chronotype and time-of-day influences on
cognition in young and elderly subjects: a behavioral and functional
neuroimaging approach. Thèse soutenue à l'Université
de Liège en vue de l'obtention du garde de docteur en sciences
psychologiques et de l'éducation.
Thèses et mémoires en
ligne
Achard, V. (2003). Influence de l'obésité sur le
rythme nycthéméral du PAI-1 plasmatique et de ses facteurs de
régulation transcriptionnelle. Mémoire présenté
à l'Ecole Nationale Supérieure Agronomique de Montpellier en vue
de l'obtention du diplôme d'études approfondies en endocrinologie
cellulaire et moléculaire.
Mongrain, V. (2006). Rythmes circadiens et mécanismes
homéostatiques de
récupération chez des personnes de type matinal
ou vespéral Th~se présentée à
l'Université de Montréal dans la Faculté des études
supérieures en vue de l'obtention du grade de Philosophiae Doctor
(Ph.D.) en Sciences Neurologiques

Annexes
Annexe 1 : Questionnaire de chronotype en ligne
Le questionnaire est disponible à l'adresse
chronotype.eu.ma
QUESTIONNAIRE DE TYPOLOGIE CIRCADIENNE
Adapté de: Horne, J. A. and O. Ostberg (1976). "A
self-assessment questionnaire to determine morningness-eveningness in human
circadian rhythms." Int J Chronobiol 4(2): 97-110.
1. Si vous viviez à votre rythme (celui qui vous
plaît le plus), à quelle heure vous lèveriezvous,
étant entièrement libre d'organiser votre journée ?

entre 5h00 et 6h30 du matin entre 6h30 et 7h45 du matin entre
7h45 et 9h45 du matin entre 9h45 et 11h00 du matin entre 11h du matin et
midi
2. Si vous viviez à votre rythme (celui qui vous
plaît le plus), à quelle heure vous mettriezvous au lit,
étant entièrement libre d'organiser votre journée ?

entre 20h00 et 21h00
entre 21h00 et 22h15
entre 22h15 et 0h30
entre 0h30 et 1h45 du matin
entre 1h45 et 3h00 du matin
3. Si vous devez vous lever à une heure précise
(plutôt tôt), le réveil vous est-il indispensable ?
beaucoup assez
peu
pas du tout
Dans des conditions adéquates (environnement favorable,
sans contraintes particulières...), à quel point vous est-il
facile de vous lever le matin ?

pas facile du tout pas très facile

assez facile
très facile
5. Comment vous sentez-vous durant la demi-heure qui suit votre
réveil du matin?

pas du tout éveillé(e) peu
éveillé(e)
relativement éveillé(e) très
éveillé(e)
6. Quel est votre appétit durant la demi-heure qui suit
votre réveil du matin ?

pas bon du tout
pas bon
assez bon
très bon
7. Comment vous sentez-vous durant la demi-heure qui suit votre
réveil du matin ?

très fatigué(e)
relativement fatigué(e) relativement en forme
très en forme
8. Quand vous n'avez pas d'obligations le lendemain, à
quelle heure vous couchez-vous par rapport à votre heure habituelle de
coucher ?

Plus de 2 heures plus tard 1 à 2 heures plus tard
moins d'1 heure plus tard rarement ou jamais plus tard
9. Vous avez décidé de faire du sport. Un(e)
ami(e) vous propose de faire des séances d'1 heure et ce 2 fois par
semaine. le meilleur moment pour lui est de 7 h à 8 h du matin. Ne
considérant que le rythme qui vous convient le mieux, dans quelle forme
penseriez-vous être?

vous trouvez cela très difficile
vous trouvez cela difficile
forme raisonnable
bonne forme
10.

A quel moment de la soiréée vous sentez-vous
fatigué(e) au point de vous endormir ? entre 2h00 et 3h00 du matin

entre 0h45 et 2h00 du matin
entre 22h15 et 0h45 entre 21h00 et 22h15 entre 20h00 et 21h00
11. Vous souhaitez être au mieux de votre forme pour un
examen qui vous demande un effort intellectuel intense durant deux heures. Vous
êtes entièrement libre de le passer quand vous le souhaitez.
Quelle heure choisiriez-vous ?

de 19 heures à 21 heures de 15 heures à 17
heures de 11 heures à 13 heures de 8 heures à 10 heures
12. Si vous alliez au lit à 23 heures, à quel
niveau de fatigue seriez-vous ?

pas du tout fatigué(e) un peu fatigué(e)
relativement fatigué(e) très fatigué(e)
13. Pour une raison quelconque, vous vous couchez quelques
heures plus tard que d'habitude, mais vous n'êtes pas obligé(e) de
vous lever à une heure précise le lendemain. Laquelle des
propositions suivantes choisiriez-vous ?

vous vous réveillez plus tard que d'habitude
vous vous réveillez comme d'habitude mais vous vous
rendormez
vous vous levez comme d'habitude mais vous vous recouchez par la
suite vous vous réveillez comme d'habitude et vous ne vous rendormez
plus
14. Pour effectuer une garde de nuit, vous êtes
obligé d'être éveillé entre 4 h et 6 h du matin.
Vous n'avez pas d'obligations le lendemain. Laquelle des propositions suivantes
vous convient le mieux?

vous n'irez au lit qu'une fois la garde terminée
vous faites une sieste avant et vous vous couchez après
la garde vous vous couchez avant et faites une sieste après la garde
vous dormez ce qu'il vous faut avant d'effectuer la garde et ne
vous recouchez pas après
15. Vous devez faire deux heures de travail physique intense,
mais vous êtes entièrement libre d'organiser votre journée.
Laquelle des périodes suivantes choisiriez-vous?

de 19 heures à 21 heures

de 15 heures à 17 heures de 11 heures à 13
heures de 8 heures à 10 heures
16. Vous avez décidé de faire du sport. Un ami
vous propose une séance d'entraînement d'1 heure et ce 2 fois par
semaine. le meilleur moment pour lui est de 22 h à 23 h. Ne
considérant que le rythme qui vous convient le mieux, dans quelle forme
penseriez-vous être?

bonne forme
forme raisonnable
vous trouvez cela difficile vous trouvez cela très
difficile
17. Supposez que vous pouvez choisir vos horaires de travail.
Admettons que vous travaillez 5 heures par jour et que votre travail est
intéressant et bien payé. A quelle heure commenceriez-vous ces 5
heures de travail consécutives?

entre minuit et 2h entre 2h et 4h
entre 4h et 5h45 entre 5h45 et 7h30 entre 7h30 et 9h entre 9h et
10h
entre 10h et 12h entre 12h et 14h30 entre 14h30 et 17h entre
17h et 19h30 entre 19h30 et 22h entre 22h et minuit
18. A quelle heure de la journée vous sentez-vous dans
votre meilleure forme ?

entre 22h et minuit entre 20h et 22h entre 18h30 et 20h entre 17h
et 18h30 entre 14h30 et 17h entre 12h et 14h30 entre 10h et 12h entre 8h et
10h

entre 6h30 et 8h
entre 5h et 6h30
entre 2h et 5h du matin entre minuit et 2h du matin
19. On dit parfois que quelqu'un est un sujet du matin ou un
sujet du soir. Vous considérezvous comme étant du matin ou du
soir?

tout à fait un sujet du soir plutôt un sujet du
soir
plutôt un sujet du matin tout à fait un sujet du
matin Commentaires éventuels

Votre score est :
>70 Vous êtes tout à fait du matin 59-70 Vous
êtes modérément du matin 42-59 Vous êtes neutre
31-42 Vous êtes modérément du soir <31
Vous êtes tout à fait du soir
Annexe 2 : Echelle explicite de préjugés
1. Les personnes d'origine maghrébine travaillent aussi
dur pour aller de l'avant que la plupart des autres Belges.
1 2 3 4
Tout a fait d'accord Plutôt d'accord Plutôt pas
d'accord Pas du tout d'accord
2. Quelle est la part de responsabilité qui incombe
aux personnes d'origine maghrébine dans
les tensions raciales qui
existent en Belgique actuellement ?
|
1
|
2
|
3
|
4
|
|
Toute la
|
Une grande part de
|
Une certaine part de
|
Aucune part de
|
|
responsabilité leur
|
responsabilité leur
|
responsabilité leur
|
responsabilité ne leur
|
|
incombe
|
incombe
|
incombe
|
incombe
|
3. En général, les personnes d'origine
maghrébine ne se plaignent pas autant qu'elles
devraient le faire
à propos de leur situation dans la société belge.
1 2 3 4
Tout a fait d'accord Plutôt d'accord Plutôt pas
d'accord Pas du tout d'accord
4. La discrimination des personnes
d'origine maghrébine n'est plus un problème en Belgique.
1 2 3 4
Tout a fait d'accord Plutôt d'accord Plutôt pas
d'accord Pas du tout d'accord
5. Y a-t-il eu énormément de changements
réels dans la situation des personnes d'origine
maghrébine
lors des dernières années, beaucoup, seulement un peu ou pas du
tout ?
1 2 3 4
Enormément Beaucoup Seulement un peu Pas du tout
6. Lors des dernières années, les personnes
d'origine maghrébine ont reçu moins qu'elles ne
le
méritent.
1 2 3 4
| 
