|
Université de Ouagadougou (UO)
********************
Unité de Formation et de Recherche
en Sciences de la Vie et de la Terre
(UFR/SVT)
*******************
Département de Biologie et
Physiologie
Végétales (BPV)
*******************
Laboratoire Biosciences
N° d'ordre:..........

MEMOIRE
Présenté
En vue de l'obtention
Du
Master Professionnel en Sélection et
Valorisation des Ressources Phyto-Génétiques
(MP/SVRPG)
Par
ALLEIDI Issa
CARACTERISATION MORPHOLOGIQUE
DES ACCESSIONS D'ARACHIDE (Arachis hypogaea L.) POUR LA TENEUR EN
HUILE ET LA TOLERANCE A LA SECHERESSE
THEME :
Soutenu le 12 Mai, 2014 devant le jury composé
de:
Président: ZONGO Jean Didier,
Professeur Titulaire, Université de Ouagadougou, Burkina Faso
Membres: TAMINI Zoumbiessé, Professeur
Titulaire, Université de Ouagadougou, Burkina Faso
TRAORE Ernest R., Assistant, Université de
Ouagadougou, Burkina Faso
Sommaire
Dédicaces
iii
Remerciements
iv
Résumé
vi
Abstract
vii
Liste des figures
viii
Liste des tableaux
ix
Liste des abréviations
x
Introduction
1
CHAPITRE I: REVUE
BIBLIOGRAPHIQUE
3
1.1 Origine et diversification de
l'arachide
4
1.2 Biologie de l'arachide
4
1.2.1 Appareil végétatif
4
1.2.2 Appareil reproductif
5
1.3 Stade de développement de
l'arachide et ses exigences écologiques
5
1.4 Principales contraintes de la
production
6
1.5 La sécheresse
7
1.5.1 Sécheresse & Aridité
7
1.5.2 Sécheresse & stress hydrique
7
1.5.3 Différents types de
sécheresse
8
1.5.4 Manifestations de la sécheresse
9
1.5.5 Effets de la sécheresse
9
1.5.5.1 Sur la surface assimilatrice
9
1.5.5.2 Sur le système racinaire
11
1.5.6 Mécanismes d'adaptation
13
1.5.6.1 Mécanismes phénologiques
14
1.5.6.2 Mécanismes morphologiques
14
1.5.6.3 Mécanismes physiologiques
16
1.5.6.4 Mécanismes moléculaires
18
CHAPITRE II: MATERIEL &
METHODES
20
II.1 Matériel
21
2.1.1 Matériel végétal
21
2.1.2 Site expérimental
21
II.2 Méthodes
22
2.2.1 Essai en conditions naturelles
22
2.2.1.2 Dispositif expérimental.
22
2.2.1.3 Entretien et traitement
22
2.2.1.4 Collecte des données
22
2.2.2 Essai en conditions
contrôlées
25
2.2.2.1 Dispositif expérimental
25
2.2.2.2 Entretien et traitement
26
2.2.2.3 Contrainte hydrique
26
2.2.2.4 Collecte des données
27
2.2.3 Analyse des
données
30
CHAPITRE III: RESULTATS
&DISCUSSION
31
III.1 Résultats
32
3.1.1 En conditions naturelles
32
3.1.1.1 Effet de la conservation sur le taux de
germination
33
3.1.1.2 Caractères quantitatifs
33
3.1.1.3 Caractères qualitatifs
38
3.1.2 En conditions contrôlées
42
3.1.2.1 Classement des accessions selon le
degré de flétrissement
43
3.1.2.2 Caractères quantitatifs
44
3.1.2.3 Caractères Physiologiques
51
3.1.3 Quelques caractères influencés
par l'environnement
59
III.2 Discussion
61
3.2.1 Conditions naturelles (Champ)
61
3.2.2 Conditions contrôlées (pots)
64
CONCLUSION & PERSPECTIVES
72
Conclusion & perspectives
73
Références
bibliographiques
74
Annexe 1: Extraction
d'huile
I
Annexe 2: Calendrier
d'arrosage durant toute l'expérimentation en pots
II
Annexe 3: Vue d'ensemble
des deux expérimentations
III
Annexe 4: Dispositif
expérimental en conditions naturelles
IV
Annexe 5: Dispositif
expérimental en conditions contrôlées
V
Dédicaces
Je Dédie Ce Mémoire
A
Ø Ma mère, décédée le
05/12/2012; que son âme repose en paix
Ø Notre père et ma grande soeur.
Ø Ma grande soeur, la défunte, que son âme
repose en paix.
Ø Toute la famille.
Remerciements
Ce travail est le résultat d'une collaboration
étroite entre plusieurs structures d'une part et d'autre part entre
plusieurs personnalités. Par conséquent, je voudrais exprimer ma
gracieuse reconnaissance à un ensemble de personnes, de près ou
de loin qui ont contribué à l'élaboration de ce document
final. Je tiens à remercier très sincèrement:
- M. ZONGO Jean Didier,
Professeur titulaire de l'Université de Ouagadougou, coordonnateur
du Master en Sélection et Valorisation des Ressources
Phyto-Génétiques (SVRPG) d'avoir accepté la direction de
ce mémoire, malgré ses multiples tâches. C'est un grand
honneur pour moi que vous présidez ce jury.
- M. TAMINI Zoumbiessé,
Professeur titulaire de l'université de Ouagadougou, pour avoir
accepté d'être membre du jury afin d'améliorer la
qualité du document. Je témoigne toute ma gratitude d'avoir
accepté de siéger à ce jury.
- M. TRAORE Ernest Renan,
Assistant à l'UFR/SVT/Université de Ouagadougou, pour les
corrections et suggestions et pour sa participation à ce jury. Merci
encore Docteur.
- M. BAKASSO Yacoubou,
Maître de conférences de l'Université Abdou Moumouni de
Niamey, pour m'avoir mis en contact avec les chercheurs de l'ICRISAT
Sadoré/Niamey et les Doctorants du département de Biologie et
pour l'achat de l'hexane lors de l'extraction d'huile.
- M. HAMIDOU Falalou, Directeur
de recherche à l'ICRISAT Sadoré/Niamey, de m'avoir accepté
dans ce centre, malgré ses multiples occupations. J'ai
bénéficié de ses multiples conseils et des
matériels qu'il a mis à ma disposition pour les travaux de
terrain. Je ne cesserai jamais de lui dire merci.
- M. HASSANE Bissala Y.,
Assistant de recherche, responsable de la banque de gènes d'ICRISAT
Sadoré/ Niamey pour m'avoir autorisé l'accès au
matériel végétal, pour son aide au cours de la
caractérisation des différents types taxonomiques de l'arachide
et pour ses conseils.
- M. HAWEL Maman Sani,
technicien permanent de l'ICRISAT Sadoré/Niamey pour son aide
apportée dans le recrutement permanent de la main d'oeuvre et pour ses
conseils chaleureux qui ne manquaient jamais.
- M. HALILOU Oumarou, Doctorant en
Physiologie végétale, à l'ICRISAT pour ses conseils et son
soutien pour l'installation de l'appareil thermo-hygrométrie au sein de
l'essai.
- M. HASSAN Abdou, technicien
à l'ICRISAT, pour les traitements des plantes qui ne manquaient jamais
dès que le besoin est nécessaire.
- Messieurs les Chercheurs, les
assistants de recherche et le personnel du centre sahélien (ICRISAT) qui
contribuent à sa bonne marche; merci encore.
- Mes remerciements vont également
à:
- M. MARICHATOU Hamani,
Maître de conférences de la faculté d'agronomie/UAM/Niamey,
responsable du laboratoire de DPA, pour m'avoir autorisé à faire
l'extraction d'huile dans son laboratoire; merci encore professeur.
- M. HAOUA Sabo, Docteur en
Biochimie au Département de Chimie de FAST/UAM/Niamey pour ses conseils
qui ne manquaient pas pendant l'extraction d'huile.
- Mme BATIONO/KANDO Pauline,
M. NANEMA Romaric, tous enseignants à l'UFR/SVT et tous
les Doctorants du laboratoire Biosciences pour leurs conseils.
- M. MAMOUDOU Morou, technicien
au laboratoire de Chimie organique, pour son accompagnement au travail et la
recherche des matériels d'extraction d'huile.
- Toute la famille et
les amis pour qui la séparation a été
difficile, mais qui ont su concilier la passion et la raison.
- Tous les professeurs qui m'ont
tenu; merci encore.
- Tous les camarades de la
même section (Master Professionnel /SVRPG) pour leur bonne collaboration
et leur gentillesse lors de ma formation.
Merci à tous.....
Résumé
L'Arachide (Arachis hypogaea L.), est une plante
alimentaire oléagineuse, adaptée aux conditions de
sécheresse. Afin d'identifier les meilleures accessions d'arachide
(teneur en huile adéquate et tolérance à la
sécheresse), vingt accessions appartenant à 5 types taxonomiques
(FST, VUL, PRU, HYR et HYB) et de diverses origines, ont
été évaluées au champ et en pots selon un
dispositif de bloc de Fisher randomisé. Vingt-neuf caractères
agro-morphologiques dont quatorze qualitatifs et quinze quantitatifs ont
été étudiés. Quatorze caractères dont quatre
physiologiques (teneur relative en eau, teneur en chlorophylle, teneur en eau
par rapport à la masse sèche et déficit hydrique) et dix
agro-morphologiques (50 % de floraison, surface foliaire, biomasse
sèche, nombre de gousses par plante, rendement en gousses par plante,
rendement en graines par plante, poids moyen d'une graine, indice de
récolte, indice de tolérance à la sécheresse,
indice de résistance à la sécheresse) ont
été observés sous conditions stressantes dans des pots
contenant 22,896 kg de substrat avec 4 traitements pour la tolérance
à la sécheresse. Quatorze caractères quantitatifs
permettent de discriminer les types taxonomiques d'arachide dont fastigiata
fastigiata Valencia est le meilleur type taxonomique ayant une
bonne teneur en huile (48,525 %). Trois caractères sont
corrélés à l'indice de tolérance à la
sécheresse, tandis que deux caractères sont associés
à l'indice de résistance à la sécheresse.
L'accession A6 est la meilleure accession tolérante à la
sécheresse.
Mots clés: Arachide, Arachis
hypogaea, caractérisation, teneur en huile, tolérance,
sécheresse.
Abstract
The groundnut (Arachis hypogaea L.) is an oilseed
plant food, adapted to drought conditions. To identify the best accessions
(adequate oil content and drought tolerance), twenty accessions belonging to 5
different taxonomic (FST, VUL PRU, HYR, HYB) and diverse origins, have
been evaluated in the field and in pots in a device randomized randomize
complete block design. Twenty-nine characters agro-morphological fourteen
qualitative and fifteen quantitative were studied Fourteen characters with four
physiological (relative water content, chlorophyll content, water content
relative to the dry mass and water deficit) and ten agro-morphological (50 %
flowering, leaf area, dry biomass, number of clove/plant, clove yield/plant,
seed yield/plant, average weight of a seed, harvest, index of drought
tolerance, resistance index drought), were observed under stressful conditions
in pots containing 22.896 kg substrate with four treatment (drought tolerance).
Fourteen relevant quantitative traits allow discrimination types of groundnut,
which hypogeae fastigiata Valencia is the best taxonomic having an oil
content (48,525 %). Three characters are correlated with the index of drought
tolerance, while two characters are associated with the index of drought
resistance. A6 is the best drought tolerance accession.
Keywords: Groundunt, Arachis
hypogaea, characterization oil content, tolerance, drought.
Liste des figures
Figure 1: Photo des plantes
témoins et stressées aux différents traitements
27
Figure 2: Evolution des
paramètres climatiques durant l'expérimentation au champ. Echelle
(110/113).
32
Figure 3: Evolution de la
température journalière durant la période de remplissage
des gousses à la récolte au champ. Echelle (100/112).
33
Figure 4: AFC et classification des
individus. Echelle:(28/82)
40
Figure 5: Dendrogramme issu de la
classification ascendante hiérarchique des accessions d'arachide sur la
base de la distribution de leurs caractères qualitatifs. Echelle
(146/199)
40
Figure 6: Evolution des moyennes de
température et d'humidité relative durant
l'expérimentation.
42
Figure 7: Evolution
des moyennes de température (T) et d'humidité relative (HR)
durant le 1èe stress hydrique appliqué au stade 50 %
de floraison. Echelle (100/111)
42
Figure 8: Evolution des moyennes de
température (T) et d'humidité relative (HR) durant le
1èrstress hydrique appliqué au stade fructification.
Echelle (107/128).
43
Figure 9: Evolution
des moyennes de température (T) et d'humidité relative (HR)
durant le 1èe stress hydrique appliqué au stade de
remplissage de gousses. Echelle (100/100).
43
Figure 10: Variation de poids moyen
d'une graine des plantes témoins (T0), stressées aux
stades 50% de floraison (T1), de fructification (T2) et
de remplissage des gousses (T3). Echelle
(100/100).
47
Figure 11: Dendrogramme
d'identification des accessions tolérantes à la sécheresse
sur la base des caractères corrélés à l'indice de
tolérance à la sécheresse. Echelle (157/194).
58
Figure 12: Photo de Graines d'A6
(accessions tolérante) aux différents traitements(T1, stress
appliqué au stade de 50 % F;T2, stress appliqué au stade de
fructification; T3,stress appliqué au stade de remplissage de
gousses
58
Figure 13: Evolution des
températures minimale (T Min) et maximale (T Max) en conditions
naturelles (N) et contrôlées (C) du semis au stade 50% de
floraison.
59
Liste
des tableaux
Tableau I: Taxon, origine, donneur et iso
donneur du matériel végétal utilisé
21
Tableau II: Liste des paramètres
étudiés
25
Tableau III: Listes de caractères
physiologiques et des indices
29
Tableau IV: Effet de la conservation sur le
taux germination
33
Tableau V: Performance de l'ensemble des
accessions en conditions naturelles.
35
Tableau VI: Performance des accessions
d'arachide pour les 15 caractères quantitatifs
36
Tableau VII: Matrice de
corrélation
37
Tableau VIII: Performance moyennes des
groupes taxonomiques
38
Tableau IX: Caractères qualitatifs
des 20 accessions d'arachide (Arachis hypogaea L.).
41
Tableau X: Regroupement des accessions en
fonction date de 50 % F et SF
45
Tableau XI: Analyse de variance des
caractères quantitatifs
48
Tableau XII: Regroupement des accessions
d'arachide en fonction de leurs BS, RGP et RgP
49
Tableau XIII: Regroupement des accessions
d'arachide en fonction des indices Ir, IRS, ITR
50
Tableau XIV: Regroupement des
accessions d'arachide en fonction de la T.Ch
52
Tableau XV: Analyse de variance des
caractères physiologiques
54
Tableau XVI: Regroupement des accessions
d'arachide en fonction de TRE, TE/MS et DH aux différents
traitements
55
Tableau XVII: Matrice de corrélation
en conditions contrôlées.
57
Tableau XVIII: Comparaison de DF et 50 % F
dans les deux conditions de culture
60
Liste
des abréviations
50%F: Cinquante pour cent de floraison
ANOVA: Analyse de Variance
CFL: Couleur de la Fleur
Cg: Couleur de la graine
COG: Compacité de la gousse
D: Dure
DC: Durée du cycle
DFL: Début de Floraison
DH: Déficit Hydrique
FFO: Forme du Foliole
FPT: Fleur Présente sur la tige
principale
HPL: Hauteur de la plante
IBPGR: International Board for plant Genetic
Resources (Now Biodiversity international)
ICG: ICRISAT Centre Groundnut
ICRISAT: International Crops Research of
Institute for the Semi-Arid Tropical
J.Org: Jaune orangé
JAS: Jours Après Semis
L: Léger
LAI: Indice de surface foliaire
M: Modéré
Mi: Mince
Mo: Moyen
NGP: Nombre de Gousses par Plante
Nmg/G: Nombre maximum de graines par
gousse
NRA: Niveau de ramification
NRG: Nombre des réticulations par
gousse
P: Prononcé
PF: Poids frais des folioles de la
troisième feuille de la tige principale
PGT: Pigmentation de la tige
PGY: pigmentation de Gynophores
PMg: Poids Moyen d'une graine
PPL: Port de la plante
Pr: Proéminente
pr: profond
PS: Poids Sec des folioles de la
troisième feuille de la tige principale
PT: Poids en pleine turgescence des folioles
de la troisième feuille de la tige principale
RGP: Rendement en gousses par Plante
RgP: Rendement en graines par Plante
SAM: Sensibilité aux maladies
Sb: Sans bec
Se: Sans étranglement
SF: Surface Foliaire
Sr: Sans réticulation
T0: Plantes
«témoins»
T1: Plantes
«stressées» au stade 50 % de floraison suivis des stress
intermittents
T2: Plantes
«stressées» au stade fructification suivis des stress
intermittents
T3: Plantes
«stressées» au stade remplissage des gousses suivis des stress
intermittents
T.Ch: Teneur en chlorophylle de la
troisième feuille de la tige principale
T: Taux de levée
Tr: Très
TAG: Taux de germination
TBG: Type de bec de la gousse
TEG: Type d'Etranglement de la gousse
TE/MS: Teneur en eau par rapport à la
Masse sèche
TEH: Teneur en huile
TIF: Type d'Inflorescence
TRE: Teneur Relative en eau de la
troisième de la tige principale
TRG: Type de Réticulation de la
gousse
Introduction
La production des graines oléagineuses s'avère
nécessaire dans l'atteinte d'une alimentation riche et
équilibrée. Les plantes oléagineuses, les plus
utilisées dans le monde sont: les amandes, le colza, le coton, l'olive,
le palmier, le soja, le tournesol, le sésame et l'arachide. Cette
dernière spéculation est la plus cultivée en Afrique et en
particulier en Afrique subsaharienne.
De 2009 à 2012, la production d'arachide en Afrique
était de 9 603 820 tonnes avec une perte enregistrée de 617 010
tonnes. La production et la perte se repartissent de la façon suivante:
Afrique de l'Ouest (6 132 589 tonnes de production et 345 456 tonnes de perte
estimée), Afrique de l'Est (1 252 542 tonnes de production contre 79 928
tonnes de perte estimée), Afrique du Nord (1 220 759 tonnes de
production et 113 820 tonnes de perte de production), Afrique du sud (104 249
tonnes de production, 4 500 tonnes de perte) et Afrique centrale (893 681
tonnes de production contre 73 305 tonnes de perte estimée ) selon
FAOSTAT, (2013).
La réduction de la production d'arachide non
décortiquée en Afrique occidentale est due à la faible
production des pays membres de cette Afrique occidentale. Selon FAOSTAT,
(2013), la perte de la production d'arachide au Niger ne cesse d'augmenter (2
654 tonnes de perte estimée en 1979 contre 7 605 tonnes en 2012).
Cette espèce oléagineuse qui est l'une des
rares légumineuses à fructification souterraine à
être utilisée dans le monde (MAYEUX, 1999) possède des
atouts. Elle est riche, en huile (44 et 56 %), en protéines (22 à
30 %), en potassium, en phosphate et en magnésium (CONKERTONE et
al., 1989). Elle nécessite moins des intrants fertilisants.
L'arachide est parfois utilisée dans les rotations culturales (contre
les maladies), la restauration des sols pauvres (Engrais vert). Elle est
utilisée dans l'alimentation humaine et animale. L'arachide est surtout
cultivée dans les pays sahéliens à cause de son
adaptabilité à ces conditions climatiques sahéliennes.
Le Niger est un pays sahélien, situé en Afrique
occidentale, entre 11°40' et 23°30' N et 0°10' et 16° E. Il
est un pays enclavé ou totalement continental avec un climat aride
à semi-aride. L'arachide y tient une part essentielle, puisqu'elle est
une oléagineuse adaptée aux zones aride et semi-aride. L'arachide
est la première culture de rente introduite au moment de la colonisation
(BAINA et al., 2007). Elle joue un rôle important dans
l'économie du pays (direct dans la commercialisation en huile, ses sous
produits pour exportation et soit indirect dans l'alimentation des
bétails destinés à la commercialisation vers les pays
côtiers).
L'arachide, plante économe en intrants est
adaptée aux situations de fortes chaleurs, de faible
pluviométrie, aux sols sableux, sablo-limoneux, limoneux et
argilo-limoneux. L'arachide doit être caractérisée afin de
sélectionner des variétés les plus productives et ayant
une teneur en huile adéquate.
Les changements climatiques qui se manifestent par
l'augmentation de la température, l'aggravation du déficit
hydrique affectent la productivité d'arachide. Il convient donc de
réfléchir aux voies et moyens pour améliorer sa
productivité dans les zones où elle est cultivée.
En effet, l'objectif général de cette
étude est de caractériser des accessions d'arachide locales et
améliorées de la banque de gènes d'ICRISAT pour
connaître leur teneur en huile et leurs réponses à la
sécheresse. Les objectifs spécifiques sont de:
Ø Evaluer l'effet de la conservation «ex
situ» sur la viabilité des semences conservées
ex-situ;
Ø Identifier des accessions riches en huile afin
d'identifier le type taxonomique ayant une teneur en huile adéquate;
Ø Identifier des accessions tolérantes aux
différentes poches de sécheresse qui interviennent aux stades
cinquante pour cent de floraison, fructification et remplissage de gousses
suivis des poches de sécheresse intermittentes;
Ce présent mémoire de master s'articule en
quatre parties que sont la revue bibliographique, le matériel et les
méthodes, les résultats et discussion et enfin la conclusion
à partir de laquelle des perspectives ont été
dégagées.
CHAPITRE I: REVUE
BIBLIOGRAPHIQUE
1.1 Origine et diversification de l'arachide
L'arachide, (Arachis hypogaea L.) est une
espèce tétraploïde (SINGH et al., 1998; HERSELMAN,
2003) avec une garniture chromosomique de 2n= 40. Elle est originaire
d'Amérique du Sud. Elle a pour centre primaire la Bolivie, l'Argentine
et le Paraguay (KRAPOVICKAS, 1969; HAMMONS, 1973 et KHALFAOUI, 1988).
L'arachide a pour centre secondaire Guaranis au Sud-Est du Brésil et
Uruguay (KRAPOVICKAS, 1969; GREGORY, 1976). L'arachide a été
introduite en Afrique, en Inde et l'Extrême Orient à partir du
Brésil, par les Portugais. Puis dans le pacifique, en Indonésie
et en Chine à partir de la côte Ouest de l'Amérique du Sud
par les Espagnols. A partir du XVème siècle.
L'arachide s'est ensuite distribuée dans le reste du monde notamment
Amérique du Nord. C'est l'une des rares légumineuses à
fructification souterraine à être utilisée à travers
le monde (MAYEUX, 1999).
1.2 Biologie de l'arachide
L'arachide est une espèce appartenant à la
famille des Légumineuses, sous famille des Papilionacées
(Fabacée), tribu des Arachidinées (GILLIER et al., 1969)
et sous tribu des Stylosanthinées (CHEVALIER, 1934; KHALFAOUI, 1988).
Elle comprend plus de soixante espèces annuelles et pérennes,
dont les trente deux décrites se repartiessent en sept sections:
Arachis, Erectodées, Caulorhizae, Rhizomatosae, Extranervosae,
Ambinervosae et Triseminalae (GREGORY et al., 1976; et
KRAPOVICKAS, 1969). La section Arachis comprend trois
séries: annuelle, pérenne et amphiplode. Cette
dernière série compte deux espèces (Arachis monticola
et Arachis hypogaea). L'espèce hypogaea a aussi
deux sous-espèces à savoir: Arachis hypogaea hypogaea et
Arachis hypogaea fastigiata. Chacune de ces
sous-espèces compte au moins deux variétés: les
variétés hypogaea et hirsuta appartiennent
à la sous espèce hypogaea; les variétés
fastigiata, vulgaris, aequatoriana et
peruviana appartiennent à la sous-espèce
fastigiata. Les types cultivés sont Virginia bunch
et runner, Valencia peruviana, Spanish et Peruvian hirsuta,
Eaquatorian. L'arachide est une plante fortement autogame.
Néanmoins son taux d'allogamie est de 0,2 % pour le type
Virginia et 6,6 % pour ceux de Spanish (SAUGER, 1949) et
Valencia (BOUFFIL, 1951) dans les zones où l'activité
des abeilles est intense (NIGAM et al., 1983).
1.2.1 Appareil végétatif
L'arachide est une plante oléagineuse des zones
semi-arides et arides dont le système radiculaire est puissant. Ce
système radiculaire est constitué par une racine primaire
pivotante qui s'enfonce verticalement dans le sol à une certaine
profondeur. L'absorption de l'eau et des éléments minéraux
est assurée par le parenchyme cortical des radicelles. Son
système racinaire présente des nodules lui permettant de fixer
l'azote atmosphérique.
La tige principale de l'arachide est toujours
érigée, avec de nombreuses variantes de ports: rampant 1, rampant
2 et rampant 3, semi érigé et érigé (IBPGR et
al., 1992).
Sur les tiges de l'arachide les noeuds peuvent être soit
végétatifs, reproductifs ou stériles. L'arachide est une
plante à fort développement végétatif. Les branches
végétative et reproductive de l'arachide peuvent être
alternées, série, irrégulière avec fleurs ou
irrégulière sans fleurs.
Les feuilles de l'arachide sont paripennées,
possèdent généralement 4 folioles. Néanmoins,
certaines variétés possèdent au moins une feuille
possédant 5 à 6 folioles soit 3 folioles en haut et 2 en bas ou
vice versa. Ce caractère est le plus souvent observé à la
quatrième feuille de la tige principale. Les feuilles de l'arachide sont
munies à leur base de deux stipules engainantes.
Les folioles sont de formes variées en allant de
cunéiforme au linéaire-lancéolé (IBPGR et
al., 1992). Elles sont opposées par paire et de couleurs
variées allant du jaune au vert bleuté. Les feuilles ont une
position diurne (le jour, les feuilles sont dressées et les folioles
largement ouvertes) et nocturne (la nuit, les pétioles se courbent vers
le sol et les folioles se rapprochent deux à deux.
1.2.2 Appareil reproductif
L'inflorescence est de deux types, soit simple ou
composée. Les fleurs sont presque sessiles et apparaissent à
l'aisselle des pétioles. La fleur est de type papilionacé (cinq
sépales et cinq pétales), de couleurs variées (blanche
à l'orange grena). Elle a 12 étamines (10 sont soudées en
tube et 2 sont libres), et un ovaire infère unicarpellaire contenant 2
à 6 ovules. L'ovaire est inséré sur un support, le
gynophore. Après fécondation, l'ovaire est porté en terre
par le développement de gynophore qui s'allonge et s'enfonce dans le sol
par géotropisme positif.
Le fruit mûrit à une profondeur de 3 à
5 cm et contient 1 à 6 graines par gousse suivant les
variétés. Les gousses peuvent avoir différents types
d'étranglement, bec, réticulations selon les types taxonomiques.
Les graines sont couvertes d'un tégument
séminal rose ou saumon. Parfois, Ce tégument présente
plusieurs couleurs très variées en allant de blanc au marron
(IBPGR et al., 1992). L'amande est composée de deux
cotylédons gorgée des matières grasses et d'un embryon
facilement distingué.
1.3 Stade de développement de l'arachide et ses
exigences écologiques
Arachis hypogaea est une plante ayant 5 stades de
développement en allant de la phase de germination à la phase de
maturation:
- Phase de germination: Cette phase correspond à
l'imbibition de la graine (24 à 48 h), après la mise en terre de
la graine jusqu'à l'apparition de la radicule (5 à 6 jours
après semis).
La germination de l'arachide est de type hypogé. Trois
semaines après la germination, les nodosités apparaissent sur les
racines.
- Phase de croissance: Elle correspond à
l'accroissement de la tige principale et l'apparition des rameaux
cotylédonaires. Ces cotylédons persistent très longtemps
et présentent comme deux petits moignons ridés. Un peu plus tard
deux autres rameaux apparaissent par rapport aux précédents.
- Phase de floraison: Généralement, elle
commence 20 à 40 jours après semis. Elle peut durer plusieurs
mois selon les variétés et les conditions de culture.
- Phase de fructification: Elle commence une semaine
après fécondation. Cette phase de fructification souterraine
requiert un sol léger, bien drainé, de pH variant de 5 à
8,5; de températures journalières oscillant entre 27 °C et
33° C et 500 mm de pluies bien distribuées durant tout au long du
cycle de son développement.
Par exemple, Valencia à port
érigé, cycle court (90 jours) dont la majorité des gousses
possède 3 à 4 loges soit 3 à 4 graines par gousse.
Spanish à port érigé, cycle court ou hâtif
(90-110 jours) dont les gousses ont généralement deux loges; soit
deux graines par gousse. Virginia soit à port rampant ou
érigé, de cycle moyen à long (100-150 jours), dont les
gousses ont une ou deux loges. L'arachide est une plante qui répond aux
engrais azotés, aux ajouts de matières organiques, de calcium, de
phosphate, de soufre et de cobalt.
- Phase de maturation: Ce stade varie selon les
variétés; il y a des variétés de 75, 85, 90, 95,
110 jours voire même 120 jours.
1.4 Principales contraintes de la production
L'arachide est une légumineuse largement produite
dans le monde pour sa forte teneur en huile (CELIA et al., 2005). Elle
est utilisée dans l'alimentation humaine (huile pour la consommation, le
beurre très populaire aux Etats-Unis, la farine en biscuiterie, arachide
bouillie, grillée ou salée); dans l'alimentation animale (fanes
utilisées comme fourrage, tourteau pour le bétail dans certain
pays; industrie (huile pour savonnerie) et dans l'agriculture (engrais vert,
rotation culturale). La teneur en huile d'arachide est variable selon les types
taxonomiques.
En effet, la sous espèce Arachis hypogaea
hypogaea a une teneur en huile qui varie entre 36 et 47 % d'huile, tandis
que celle de la sous espèce Arachis hypogaea fastigiata varie
entre 45 et 50 % (CELIA et al., 2005). Selon WIKIPEDIA (23 h 10,
26/01/2014), 100 g d'huile d'arachide contient 39 g d'acides gras
mono-insaturés, 38 g d'acides gras essentiels oméga-6, 23 g
d'arachide gras saturés (palmitique, stérique,
béhénique, arachidique), et moins de 0,1 g d'arachide gras
essentiel oméga-3. Contrairement aux autres huiles (olive, colza,
tournesol, soja), l'huile d'arachide est très riche en acides gras
saturés. Elle résiste à l'oxydation et aux fortes
températures (CELIA et al., 2005), donc peut être
conservée en conditions climatiques de zones sahéliennes.
C'est une huile qui contient 9 cal/g et ne dégage pas
de fumée quand elle est chauffée. Malgré son importance
alimentaire, industrielle, cosmétique; l'arachide est confrontée
à un certain nombre des contraintes notamment les contraintes biotiques
telles que les maladies fongiques, bactéricides, les viroses et les
ennemis (insectes). Elle est principalement confrontée aux contraintes
abiotiques telles que la sécheresse.
1.5 La sécheresse
La notion de «sécheresse» est
difficile à définir de façon précise puisqu'elle
est relative aux zones et à la manière dont les ressources en eau
sont utilisées par les plantes. Elle peut être
considérée comme un phénomène rare ou
fréquent, normal ou catastrophique. Elle peut se définir comme,
une période étendue de diminution des pluies par rapport à
un régime normal autour duquel l'environnement local et
l'activité humaine se sont stabilisés. C'est une pénurie
et/ou une petite quantité d'eau donnée, face à des besoins
précis dans le temps et dans l'espace, un manque d'eau sur une
période significativement longue ayant des impacts sur les cinq
règnes (Végétal, Animal, Champignons, Protiste,
Monère). Elle est une contrainte abiotique et risque d'être l'une
des conséquences majeures du changement climatique.
Du point de vue agronomique, la notion de la
sécheresse exprime un «déficit hydrique». Ce
déficit hydrique peut être défini comme une
réduction de la teneur en eau, du potentiel hydrique ou du potentiel de
turgescence. La sécheresse peut être épisodiquement naturel
(modification de la température de la surface de l'air et de
l'humidité du sol) ou anthropique (émission de gaz à effet
de serre, déforestation, désertification), fait suite à un
déficit pluviométrique, sur des longues périodes durant
lesquelles les précipitations sont anormalement faibles ou insuffisantes
pour maintenir l'humidité du sol et l'hygrométrie normale de
l'air afin d'assurer une meilleure production agricole.
1.5.1 Sécheresse & Aridité
Le terme «sécheresse» recouvre des
notions différentes. Elle peut être définie comme un manque
d'eau ponctuel, un déficit de pluviométrie non
systématique et imprévisible pouvant être
caractérisée par: l'intensité de sa déviation par
rapport aux valeurs moyennes ou normales de pluviométrie, la
durée, la période d'occurrence, l'extension géographique,
la dynamique de mise en place et l'époque d'apparition par rapport au
cycle cultural pour les plantes. La «sécheresse»
diffère de «l'aridité» par le fait que,
l'aridité est un déficit systématique de
pluviométrie qui se manifeste dans des zones peu propices aux
activités agricoles.
1.5.2 Sécheresse & stress hydrique
Un «stress hydrique» peut être
défini non seulement comme un facteur abiotique ayant un impact sur le
fonctionnement normal des organismes vivants (LEVITT, 1980). Il peut être
aussi une mauvaise quantité d'eau disponible à la plante.
Par exemple, une plante cultivée sur un sol
permanemment humide, inondé aurait un développement anormal,
voire même sa croissance inhibé. L'excès d'eau peut
être donc considéré comme un «stress
hydrique». Ce type de stress s'explique par la réduction
d'oxygène dans le sol (problème d'aération des racines),
la diminution d'absorption des éléments nutritifs qui sont
indispensables au développement de la plante (HOPKINS et al.,
2003). En bref, le «stress» peut être thermique,
salin, oxydatif et hydrique. Le «stress hydrique» est le
plus fréquent en Afrique subsaharienne où la pluviométrie
devient de plus en plus faible à telle enseigne que l'expression de ce
terme «stress» est considérée comme un
«stress hydrique». Ce stress hydrique peut être
considéré comme une sécheresse, lorsqu'il est dû au
manque d'eau, donc un déficit hydrique. Par contre le stress hydrique
diffère de la sécheresse, lorsqu'il est dû à
l'accès d'eau (plantes cultivées dans des zones permanemment
humides). En Afrique subsaharienne où les cultures seraient les plus
exposées au déficit hydrique. Les deux termes (stress hydrique et
sécheresse) sont communs, à telle enseigne qu'il est difficile de
distinguer «stress hydrique» et «sécheresse». Le
stress hydrique ou sécheresse est la première contrainte
abiotique qui entraine des différences significatives entre les
rendements d'une culture (SORRELLS et al., 2000; IJAZ RASOOL et
al., 2012).
1.5.3 Différents types de sécheresse
La sécheresse peut détruire les
récoltes partiellement ou totalement. Elle constitue l'une de
principales contraintes de réduction de la production agricole. La
sécheresse devient alors un facteur de famine régionale, exode,
souvent accompagnée des troubles sociaux puis des conflits armés,
notamment dans les régions où il y aurait peu des ressources
génétiques. Elle est un facteur complexe selon
WIKIPEDIA/sécheresse (
www.secheresse).
Du point de vue des domaines d'application ou
d'utilisation, trois types de sécheresses sont distingués:
-la première est la sécheresse
météorologique qui survient lorsqu'il existe une période
prolongée de précipitations en dessous de la moyenne;
-la seconde est la sécheresse hydrologique qui survient
lorsque les réserves d'eau disponibles dans les nappes aquifères,
lacs et réservoirs descendent en dessous de la moyenne;
-la troisième est la sécheresse agricole ou
«sécheresse-rhizosphèrique», causée par une
insuffisance de l'eau dans le réservoir superficiel du sol pendant la
saison culturale. Elle se manifeste lorsqu'il n'existe pas une humidité
assez suffisante pour le développement des cultures. Ce type de
sécheresse agricole peut se réaliser même si les
précipitations sont normales, non seulement à cause de la nature
ou état du sol, de la mauvaise itinéraire technique, mais aussi
lorsqu'il y a une faible humidité du sol associée aux fortes
températures du sol.
La sécheresse agricole freine aussi la croissance
végétative des plantes, diminue les composantes du rendement, met
en danger le bétail (LAURE et al., 2010). Elle est l'un des
critères ou paramètres, les plus importants, dans un programme de
sélection et d'amélioration variétale dans les zones
à faible pluviométrie; afin d'assurer une production agricole
adéquate. La sécheresse se manifeste par plusieurs
façons.
1.5.4 Manifestations de la sécheresse
La sécheresse provoque un stress hydrique. Elle est
la plus frappante dans les zones agricoles et l'un des principaux facteurs
limitant la production agricole mondiale (TURNER, 1986; PASSIOURA et
al., 1996). La sécheresse est un phénomène
naturel qui se manifeste aussi bien par un déficit pluviométrique
entrainant un déficit de production (BARAKAT et al., 1997) que
par une diminution importante de la longueur de la saison des pluies, la
fréquence accrue des arrêts des pluies «poches de
sécheresse» plus ou moins longues pendant la période
culturale (ZAGRE et al., 1998). Elle peut être exprimée
aussi d'une part, par la restriction de la disponibilité en eau du sol,
d'autre part pat l'augmentation d'évaporation et
d'évapotranspiration accrue par l'élévation des facteurs
climatiques tels que la température l'ensoleillement, la vitesse du
vent. En Afrique, où la culture en saisons pluvieuses est très
largement majoritaire, la sécheresse est une contrainte permanente de la
production agricole (CECCARELLI et al., 1996). Elle affecte la
croissance des plantes.
1.5.5 Effets de la sécheresse
Les effets de la sécheresse se font ressentir sur les
parties végétatives avant d'affecter les parties reproductives.
Par exemple chez le riz pluvial, les parties végétatives les plus
affectées par la sécheresse sont la hauteur de la plante, le
tallage, la surface foliaire, la matière sèche et le
diamètre des tiges. Pour les parties reproductives: la longueur des
panicules par exemple. La durée du cycle et de l'épiaison sont
affectées par la sécheresse. Les variétés ne se
comportent pas de la même façon vis-à-vis des
périodes sur lesquelles la sécheresse intervient. Par exemple une
variété peut être plus affectée par la
sécheresse à un stade de son développement et être
moins affectée à un autre stade de développement (REYNIER
et al., 1978). Les possibilités de criblage des
variétés vis-à-vis de la sécheresse posent
énormément des problèmes. Car non seulement, les
comparaisons variétales n'apportent peu d'information sur la
tolérance variétale. Selon le stade de développement, le
classement des variétés peut s'inverser. En plus, les
variétés ayant l'enracinement le plus profond ne se
révèlent pas forcement les plus résistantes.
1.5.5.1 Sur la surface
assimilatrice
La croissance végétative des plantes en
condition de sécheresse est fortement perturbée. Par exemple chez
le tournesol, la croissance foliaire soumise à un déficit
hydrique est arrêtée très rapidement lorsque le potentiel
hydrique foliaire est de -0,4 Mpa (BOYER, 1968).
La sécheresse a un effet négatif sur: la hauteur
de la plante (IJAZRASOOL, 2012), la matière sèche (KRAMER, 1983;
CHOWDHRY et al., 1999; HASSAN, 2003; NOORKA et al., 2009a,b;
MOHARRAM et al., 2011) et la surface foliaire. Cette diminution de la
biomasse aérienne, provient de la sénescence
prématuré ou diminution de l'expansion foliaire. Au niveau
cellulaire, la diminution de la croissance végétative est due
à l'inextensibilité de la paroi cellulaire et les
réactions biochimiques impliquées dans le processus de relaxation
de cette paroi (MATTHEWS et al., 1984; COSGROVE, 1993).
L'extensibilité assure la turgescence cellulaire. Par exemple lorsque la
valeur de la turgescence est inferieure à la valeur seuil, la surface
foliaire totale diminue en passant par la sénescence foliaire.
§ Sur la photosynthèse
La photosynthèse se déroule dans les
chloroplastes des organes chlorophylliens. Ces organes sont les plus souvent
les feuilles, mais parfois des tiges (Leptadenia hastata). Elle est un
ensemble des processus aux cours desquels, en présence de la
lumière et des éléments minéraux, les
végétaux verts synthétisent des molécules
organiques (glucose, lipides et protides). C'est une réaction
nécessitant de la fixation de CO2 sur le Rudi-P (Rubilose
bi-phosphate), libération de l'oxygène O2 dans
l'atmosphère, en présence de la lumière et d'une
alimentation purement minérale. Au cours d'une période de
sécheresse, la réduction de la photosynthèse est
liée à la diminution du potentiel hydrique foliaire. Elle est
aussi liée à la fermeture des stomates, proportionnelle à
la baisse du potentiel hydrique (BOIS et al., 1983) et à la
limitation des métabolismes carbonique et enzymatique (ZINSELMEYER et
al., 1999; KIM et al., 2000). La réduction de la
fixation du CO2 limite la régénération du
Rudi-P (Ribulose bi-phosphate) substrat du cycle de Calvin (GIMENEZ et
al., 1992). Lors d'un abaissement du potentiel hydrique, la
conductance stomatique diminue. Chaque plante, a son potentiel hydrique moyen
du sol, auquel la plante a une alimentation en eau suffisante. Par exemple chez
le tournesol, lorsque le potentiel hydrique se situe à -2 Mpa, la
fixation de CO2 diminue (MOJAYAD, 1993). Chez le riz le potentiel normal
d'absorption est de -200 mbar selon (BOIS et al., (1983). Lorsque ce
potentiel hydrique chute rapidement, la fixation du CO2 est
limitée, donc la photosynthèse se trouve perturbée. Le
contrôle de la régulation stomatique est lié à la
turgescence cellulaire, l'Acide Abscissique (ZHANG et al., 1989; DAVIS
et al., 1994). Cet Acide Abscissique (ABA) est probablement en forte
concentration dans les feuilles lors d'un déficit hydrique (TARDIEU et
al., 1993a).
§ Sur la Respiration
La respiration se réalise dans les mitochondries
qui sont des organites présents dans toutes les cellules respiratoires.
La respiration est une réaction de dégradation
totale des composés organiques aboutissant à la formation des
corps minéraux, responsable de l'oxydation des composés
organiques (consommation d'O2, libération de CO2).
La respiration est affectée par la réduction d'oxygène et
l'absorption des éléments nutritifs dissous dans l'eau (HOPKINS
et al., 2003).
1.5.5.2 Sur le
système racinaire
Le stress hydrique ou sécheresse entraine une
réduction du diamètre de la racine. Par exemple chez le riz et le
tournesol, il peut avoir une réduction pouvant attendre 40 % de
diamètre de la racine (CRUZIAT, 1974 ; BOIS et al., 1983). La
sécheresse provoque une perte du potentiel de turgescence. Par exemple
lorsqu'une plante subit un déficit hydrique, les cellules perdent la
turgescence. Cette perte de turgescence entraine une modification de la
structure, un ralentissement de la croissance de la plante. Selon les
variétés, la croissance racinaire peut être maintenue par
un ajustement osmotique qui limite la baisse du potentiel de turgescence
(TURNER, 1988). Selon TURNER et al., (2001), une sélection
s'opérant sur la base du système racinaire présente deux
inconvénients que sont l'impraticabilité du criblage au champ
pour cette caractéristique à grande échelle et la
difficulté de corréler des observations au champ à celles
qui sont faites en pots.
1.5.5.2a Sur la pression de turgescence
L'eau, est le constituant fondamental de la cellule et
représente 50 à 90 % de la matière fraîche. Elle
assure la turgescence cellulaire. Celle-ci, intervient dans l'ouverture des
stomates, le maintien du volume du chloroplaste (GUPTA et al.,
1987).
1.5.5.2b Sur la croissance de la plante
La croissance est une augmentation de la masse de la
matière fraîche. Elle se traduit par une augmentation de la taille
et de la masse corporelle. Elle est assurée par des zones
privilégiées (méristèmes). Ces
méristèmes sont des petits massifs des cellules
indifférenciées, gardant chaque organe les
propriétés des cellules embryonnaires. La croissance en longueur
est assurée par les méristèmes primaires tandis que celle
en épaisseur est assurée par les méristèmes
secondaires. Ces deux méristèmes sont à l'origine de
l'anatomie des plantes.
La sécheresse affecte considérablement cette
croissance. Elle est l'un des facteurs majeurs qui limite la production
végétale en conditions naturelles (GABRIEL et al.,
2008). Par exemple une poche de sécheresse intervenue durant une saison
culturale entraine une forte diminution de la réserve en eau du sol et
limite la croissance de la plante (GABRIEL. et al., 1996). Lorsque
l'humidité du sol diminue, le potentiel hydrique foliaire
nécessaire (provoquant le mouvement d'eau depuis le sol jusqu'aux
feuilles) est plus faible (LACAZE., 2006).
1.5.5.2c Sur le rendement et ses composantes
Le rendement d'une culture est un caractère
quantitatif, complexe et faiblement héritable. Le rendement est
habituellement et négativement corrélé à la
sénescence foliaire précoce (MERRIEN al., 1981). Il est
surtout influencé par les pressions parasitaires et d'adventices, la
densité de semis, la date de semis et les facteurs climatiques et
édaphiques. Le rendement est significativement et positivement
corrélé à certains caractères. Par exemple chez
l'arachide le rendement est associé à la hauteur de la plante, au
nombre des gousses par pied, au poids moyen d'une gousse et au taux de
maturité, (ZAGRE, 2004). Le rendement est aussi lié à
l'ajustement osmotique (AO) et à l'efficacité d'utilisation de
l'eau par la plante (REKIKA, 1997; TEULAT, 1997a; TEULAT et al.,
1997b; MERAH, 1999; TEULAT. et al., 2001) dont la plupart des
études ont montré que la disponibilité en eau intervient
dans l'élaboration du rendement. Par exemple chez le tournesol,
l'élaboration du rendement est liée à la consommation en
eau par son impact: sur la surface assimilatrice, la durée du
fonctionnement de la surface foliaire et le taux d'assimilation du carbone par
les feuilles (MERRIEN et al., 1981a). Selon IJAZRASOOL et
al., (2012), la hauteur de la plante, surface foliaire du drapeau et
le rendement en graines sont des caractères liés au rendement.
Des observations faites sur des caractères agro-morphologiques
intervenant dans les composantes de rendement montrent qu'il y a une
variabilité importante dans les formes architecturales des plantes
(FELICITY et al., 2010). Cette variabilité doit être
largement exploitée par les sélectionneurs afin de mettre des
variétés (tolérantes à la sécheresse)
à la disposition des populations. L'attente de cet objectif
s'élabore tout au long du cycle de développement de la plante.
C'est ainsi que, les premiers travaux ont démontré l'importance
de la période de remplissage des graines d'une culture (TRIBOI et
al., 2002). Les études ont mis en évidence le rôle
de l'activité photosynthétique en post floraison dans
l'élaboration de rendement. Cette activité est ralentie par le
maintien de la surface foliaire et son activité métabolique lors
d'une sécheresse (DEBAEKE et al., 2004; TRIBOI-BLONDE et
al., 2004). Au regard de tout ce qui précède, tout
facteur abiotique ou biotique affectant l'un des composantes de rendement a des
répercussions sur le rendement.
Par ailleurs, tout déficit hydrique qui agit sur le
tallage, la surface foliaire; entraine une baisse de rendement (LARBI et
al., 1997). Par exemple chez le sésame, des études ont
montré qu'un déficit hydrique entraine une réduction du
rendement en passant par la réduction des composantes du rendement
(nombre de capsules, poids de capsule).
1.5.6 Mécanismes
d'adaptation
La sécheresse est un stress abiotique affectant le
rendement d'une plante cultivée et la stabilité de son rendement
suivant les années. L'impact de la sécheresse sur le rendement
dépend à la fois du génotype de la plante, de
l'espèce considérée et des techniques culturales (LAURE et
al., 2010). La sécheresse se manifeste par la combinaison d'une
part, de la restriction de la disponibilité en eau du sol et d'autre
part, de l'augmentation d'une demande évaporatrice.
Pour palier cette contrainte abiotique, les plantes ont
élaboré des «stratégies d'adaptation»
permettant d'assurer leurs physiologies normales. Cette stratégie
d'adaptation est la capacité d'une plante de maintenir ses fonctions
physiologiques, de croître et de maintenir le rendement dans des
conditions stressantes (TURNER, 1997; JONES et al., 1992). Elle est
considérée comme une dynamique dont la résultante est la
«tolérance».
Tolérance: La notion de
«tolérance» reste à définir de
manière consensuelle, dans la mesure où l'appellation des termes
résistance et tolérance à la sécheresse
dépend des auteurs et des domaines d'application. Par exemple les
écologues définissent la tolérance comme la
capacité d'une plante à survivre dans une zone à faible
apport d'eau (LEVITT et al., 1980).
Les agronomes expliquent que la tolérance à la
sécheresse est la capacité d'une plante à croître et
à produire des rendements satisfaisants dans des zones sujettes à
des déficits hydriques épisodiques (TURNER, 1997).
Quant aux physiologistes, la tolérance est une
stratégie permettant à une plante d'assurer ses fonctions
physiologiques malgré la dégradation de son état hydrique.
Par exemple, le maintien de la turgescence lors d'un déficit hydrique
permet de retarder la fermeture des stomates (MOJAYAD et al., 1994),
de maintenir le volume chloroplastique (GUPTA et al., 1987), de
réduire le flétrissement foliaire (JONES et al., 1980)
et de maintenir les fonctions cellulaires indispensables à la survie.
Ces aptitudes permettent aux plantes de tolérer la sécheresse
(LUDLOW et al., 1983).
Du point de vue génétique, la tolérance
est la capacité du potentiel génétique de la plante de
s'exprimer son expression phénotypique dans un environnement stressant.
L'expression de gènes aboutissant à l'accumulation des osmolytes
sont considérés comme les principaux mécanismes de
tolérance au déficit hydrique (RAMANJULU et al., 2002).
Par exemple chez le tournesol, les capacités osmotiques dépendent
de son génotype (CHIMENTI et al., 2002). Selon JONES (1992), la
tolérance peut être classée en deux catégories en se
basant sur le potentiel chimique de la plante: Il s'agit de
l'«évitement», qui correspond à une
augmentation ou le maintien du potentiel hydrique du tissu et la
«tolérance» proprement dite qui est le potentiel
chimique des tissus diminue.
La tolérance est le résultat des
mécanismes physiologiques, biochimiques et moléculaires complexes
permettant aux plantes tolérantes de produire un rendement
satisfaisant.
Ce processus complexe permet de sélectionner des
plantes tolérantes à la sécheresse en fonction de la
disponibilité des moyens des sélectionneurs.
1.5.6.1 Mécanismes phénologiques
Esquive: L'esquive est la première
stratégie des plantes d'éviter la sécheresse. C'est l'un
des types d'adaptation à l'environnement qui permet aux plantes
d'éviter les périodes critiques pour pouvoir produire.
L'esquive permet à la plante d'atténuer les
effets des contraintes hydriques par accomplissement de son cycle de
développement avant l'installation de la sécheresse (BEN NACEUR
et al., 1999; AMIGUES et al., 2006). Par ce système,
de nombreuses variétés ont été créées
pour assurer une production adéquate dans les zones où de poches
de sécheresse interviennent en fin de saison (SUBBARAO, 1995; FUKAI et
al., 1995; TURNER et al., 2001). Les plantes qui esquivent ne
subissent jamais la sécheresse. Dans un programme de sélection et
d'amélioration, l'esquive ne devrait pas être
considérée comme un critère de sélection dans les
zones où il aurait des poches de sécheresses qui interviennent en
début ou en pleine saison culturale. L'esquive ne peut se raisonner
qu'à l'échelle d'exploitation agricole et des systèmes de
culture en utilisant des variétés plus précoces ou
semi-précoces selon les zones agro écologiques.
1.5.6.2 Mécanismes morphologiques
Les plantes sont capables de mettre en place toute une
série de réponses physiologiques leur permettant de
tolérer la sécheresse afin de s'adapter aux conditions
environnementales. Elles peuvent atténuer les effets du stress sur leur
métabolisme afin d'assurer leur survie et leur reproduction. Le
mécanisme le plus connu est appelé «évitement».
Evitement: L'évitement est le
deuxième type d'adaptation des plantes. Il est la capacité d'une
plante de maintenir son état hydrique satisfaisant, permettant de
limiter les effets du stress (grâce à des adaptations comme le
flétrissement, transformation des feuilles en épines ou en
écailles, orientation des feuilles parallèlement à la
direction du rayonnement solaire, l'enroulement des feuilles, présence
des cires à la surface des feuilles, réduction de la surface
foliaire). Cette stratégie permet aux plantes de survivre tout en
assurant une productivité satisfaisante. Cette dernière est
liée à la fois à la réduction de la transpiration
et à une optimisation d'eau par les racines. L'évitement se
manifeste par des modifications des parties aériennes et souterraines
(SLAMA et al., 2005).
Après l'installation de la sécheresse, une
diminution de la croissance et une modification des mouvements d'eau sont
associées à des réorientations du métabolisme et
à une fermeture des stomates. Cette fermeture des stomates assure une
bonne conservation de l'état hydrique de la plante. Par exemple lorsque
la sécheresse se réalise, il y a un ralentissement de la
croissance végétative et reproductive, en d'autres termes, une
réduction de la biomasse aérienne et souterraine.
Ce ralentissement résulte d'une diminution de la
vitesse de division cellulaire (GRANIER et al. 2000) et d'une
modification des propriétés physico-chimiques des parois
pecto-cellulosiques (plus rigides). Ceci empêche la croissance et le
développement de la plante. Ces processus limitent les échanges
gazeux. Des études récentes ont montré que ce
phénomène n'est pas une conséquence passive du manque
d'eau dans les cellules, mais plutôt une réponse
contrôlée par la plante, dont le résultat est
d'éviter le stress hydrique.
Si la croissance des parties aériennes est
altérée en cas de stress hydrique, celle des racines est moins
touchée vis-à-vis de la réduction du système
racinaire et favorise une augmentation de l'absorption de l'eau et des
éléments nutritifs via l'enracinement, la colonisation rapide du
sol, l'augmentation du diamètre des vaisseaux (WU et al.,
2000).
1.5.6.2a Surface foliaire
L'eau est une ressource naturelle inégalement
répartie dans la plante selon le stade de développement de la
plante. Elle doit être en quantité suffisante pour couvrir les
besoins des plantes en eau et préserver les écosystèmes
(BLANCHON, 2009). La réduction de la surface foliaire et la diminution
de la conductance stomatique jouent un rôle déterminant dans les
mécanismes permettant de réduire la transpiration. Cette
réduction de la surface foliaire est un moyen de réduction de
perte d'eau par transpiration (PERRIER, 1975). Lors d'une sécheresse
précoce, la réduction de la surface foliaire associée
à une diminution de l'expansion foliaire s'accompagne d'une baisse de
rendement lorsque l'indice de surface foliaire (LAI) est en deçà
de la valeur seuil. Par exemple chez le tournesol, lorsque LAI chute en dessous
de 2,5 à la floraison, le rendement baisse (MERRIEN et al.,
1990). Le rendement est un caractère complexe, corrélé
à la durée de la vie de la surface foliaire après
floraison, fortement et négativement affecté par la
sénescence prématurée due à un déficit
hydrique. La régulation de conductance stomatique dépend du
potentiel hydrique foliaire et de l'humidité de l'air au champ (TURNER,
1997). Le potentiel hydrique est fonction du génotype de la plante
(MOJAYAD, 1993) et de son stade de développement (MORIZET et
al., 1990). Cependant, les génotypes à faible
conductance sont plus sensibles au déficit de vapeur et à la
baisse du potentiel hydrique foliaire que les génotypes à forte
conductance. Donc la faible conductance est un critère de
sélection pour chercher des variétés adaptées
à la sécheresse. A l'échelle de la plante, l'eau est
acheminé des racines aux feuilles par les vaisseaux conducteurs en
particulier le xylème; tandis qu'à l'échelle des tissus,
l'eau circule de cellule en cellule par la voie apoplastique (passage libre
à la surface des parois entourant les cellules végétales)
et voie symplastique (passage au travers des membranes). Des études
récentes ont montré que, les échanges d'eau
transmembranaires sont réalisés au travers des pores
transmembranaires appelées aquaporines (LUU et al., 2005).
Les aquaporines sont des protéines incluses dans les
membranes plasmiques, composants significatifs dans le transport cellulaire de
l'eau (MAUREL et al., 2001; TEYRMAN et al. 2002). Elles
assurent la conductivité hydrique et augmentent la
perméabilité de l'eau dans la cellule en allant de 10 à 20
fois (MAUREL et al., 2001). Lorsqu'il y a un stress hydrique,
l'ouverture et la répartition des aquaporines dans les membranes sont
modifiées. Ce qui permet de moduler le trajet de l'eau au sein des
tissus ou organes (ZHU et al., 2006). L'expression du gène
d'aquaporine est corrélée à la teneur relative en eau
(RWC).
Les gènes d'aquaporines assurent la
perméabilité de l'eau et l'augmentation du flux de l'eau
(YAMAGUCHI et al., 1992; YAMADA et al., 1997). La
répression de gènes d'aquaporines diminue la
perméabilité de l'eau dans la cellule pendant les périodes
de sécheresse (SMART et al., 2001).
1.5.6.2b Stomates et Cuticule
En cas de stress hydrique, un mécanisme
signalétique permet à la plante de fermer ses stomates. Il en
résulte une réduction des pertes d'eau par transpiration et le
maintien de l'état hydrique des cellules. Quant à la cuticule,
des études ont montré que certaines plantes réduisent la
perte d'eau par une augmentation de l'épaisseur de la cuticule lors un
déficit hydrique prolongé.
1.5.6.2c Système racinaire
L'optimisation de l'absorption d'eau est liée
à un ensemble des caractères quantitatifs tels que la masse, le
volume et la profondeur racinaire (RAMANJULU et al., 2002). Chez le
blé, la longueur des racines primaires et la finesse racinaire
paraissent les plus reliées à la tolérante à la
sécheresse (DAALOUL al., 2013). De nombreuses études ont
montré que les plantes adaptées aux zones arides sont
tolérantes à la sécheresse et possèdent un
enracinement profond et extensif capable d'extraire l'eau du sol. Selon BEN
SALM et al., (1991), un système radiculaire profond permet au
blé de résister aux conditions stressantes. L'aptitude des
racines à extraire l'eau du sol, est une réponse pour
l'élaboration de la production en graines (PASSIOURA, 1994; SUBBARAO,
1995). La croissance racinaire en condition de stress hydrique peut être
maintenue par l'ajustement osmotique qui limite la baisse du potentiel de
turgescence (TURNER, 1986).
1.5.6.3 Mécanismes physiologiques
1.5.6.3aTeneur relative en eau (RWC)
La tolérance à la sécheresse est le
résultat de nombreux caractères anatomique, morphologique,
biochimique, physiologique constitutifs ou inductifs qui interagissent pour
permettre le maintien du processus de la croissance et du développement
de la plante dans les conditions stressantes. La sécheresse diminue la
croissance et le développement de la plante donc la productivité,
en affectant la turgescence cellulaire qui est liée à
l'altération de l'état hydrique cellulaire.
La teneur relative en eau (RWC ou TRE), le potentiel
hydrique (Øw) et ses composantes, le
potentiel de turgescence (Øt) et le potentiel osmotique
(Øs), sont les paramètres les plus
étudiés pour évaluer l'état hydrique des plantes
(TEZAR et al., 2002). La diminution du potentiel hydrique foliaire et
de la teneur relative en eau observée au cours d'une réponse du
déficit hydrique aboutit à une diminution de la
photosynthèse chez le tournesol (TEZARA et al., 2002). De
nombreuses études ont démontré que l'ajustement osmotique
est associé à la tolérance à la sécheresse
(CLARK et al.,1982; MORGAN, 1983, 1984; SCHONFELD et al.,
1988; JONGDEE, 2002; CHIMENTI et al., 2002; OBER al., 2005,
CHIMENTI et al., 2006).
En cas de déficit hydrique dans le sol ou lorsque le
potentiel hydrique foliaire est réduit, les tissus de la plante peuvent
perdre de l'eau. Cependant, certaines plantes sont capables de maintenir leur
turgescence cellulaire (Øt
=Øw-Øs) par l'accumulation
contrôlée des osmoprotectants (acide aminés, sucres,
polyols, proline, les composés à ammonium quaternaire et les
composés tertiaires de sulfonium et les ions (TURNER et al.,
1980; ZHANG et al., 1999; ABEBE et al., 2003). Ce
phénomène est appelé ajustement osmotique, permettant de
maintenir le volume cellulaire correct, malgré les pertes d'eau
engendrées par ce déficit hydrique. L'ajustement osmotique (AO)
est l'accumulation active des solutés dans la cellule. Il est sans doute
le mécanisme le plus important pour maintenir la turgescence ainsi que
l'activité physiologique lorsque le potentiel hydrique diminue (MORGAN,
2005; CHIMENTI et al., 2005). Chez le tournesol, l'AO contribue
à maintenir le rendement du tournesol dans les conditions de
sécheresse (CHIMENTI et al., 2002). Il
se calcule de la façon suivante: AO= Øs FT(non
stressé)- Øs FT( stressée),
avec Øs FT = Potentiel osmotique à la
pleine turgescence. Cet ajustement osmotique permet à la plante
d'assurer le durcissement de ses feuilles et tiges. Lorsqu'il y a perte de
turgescence, il peut y avoir une tolérance à la
déshydratation des tissus (TURNER, 1986, BRAY, 2000). De nombreux
travaux ont montré qu'une modification de l'expression de gènes
impliqués dans les voies de biosynthèse d'osmolytes est en
réponse avec le déficit hydrique. Il
paraît que le mauvais remplissage des épis en osmolyte, en
particulier le sucres, et notamment les saccharoses est la cause de
l'arrêt de développement des grains (BOYLE et al., 1991;
ZINZELMEIR et al.,1995a). Cependant, le processus de remplissage des
graines d'arachide et le signale cellulaire déclencheur de la carence en
lipides et l'avortement restent à identifier. Par exemple, l'expression
simultanée du gène de la P-5-C synthéase (Acide 5
carboxylique de la proline) et la répression du gène de la
proline déshydrogénase (ProDH) conduit à une accumulation
de la proline pendant un déficit hydrique (YOSHIBA et al.,
1997). Un transporteur (ProT1) de la proline a été induit lors
d'un stress hydrique (SCHWACKE et al., 1999). Le mécanisme de
la protection fourni par ces osmolytes reste à être
expliqué clairement (RAMANJULU et al., 2002). Quant aux sucres
solubles, il a été montré que les sucres jouent un
rôle déterminant dans l'ajustement osmotique et la stabilisation
des certaines protéines (CARPENTER et al., 1990).
1.5.6.4 Mécanismes moléculaires
Une plante soumise à un stress hydrique aboutit
à la mise en place de nombreuses réponses cellulaires ayant des
conséquences importantes sur sa croissance et son
développement.
La plante doit se protéger contre les effets du stress
en modifiant son métabolisme, tout en essayant de maintenir ses
capacités de production. Les réponses sont multiples et reposent
sur des mécanismes cellulaires impliquant de nombreuses
protéines, enzymes, hormones, donc de nombreux gènes. Il est
important d'identifier des gènes candidats pour une éventuelle
amélioration génétique des variétés
tolérante à la sécheresse et de teneur en huile
adéquate.
1.5.6.4a Les gènes
Les protéines, les enzymes et les hormones sont
codées par des gènes. L'expression de ces gènes peut
être modifiée en fonction des conditions environnementales telles
que les conditions stressantes (HARBI et al., 2010). La
détermination de l'expression des gènes potentiellement
impliqués dans les réponses cellulaires d'une plante lors d'un
déficit hydrique a été mise en évidence par
plusieurs techniques, telles que les techniques transcriptomique et
protéomique (SEKI et al., 2002; SHIMOZAKI et al.,
2003). Les gènes dont l'expression est régulée par ces
deux voies de signalisation codent des protéines impliquées dans
la réponse au stress hydrique (SHINOZAKI et al., 2007).
Grâce au séquençage, analyse génétique et de
logiciel de cartegène, il est possible de localiser ou identifier des
QTL (Quantitatives Trait Locus). La détection des QTL est un moyen
rapide et efficace pour l'amélioration des plantes tolérantes
à la sécheresse et ayant une bonne teneur en huile. En effet,
certaines études ont montré qu'il y a des QTL qui sont
liés à la tolérance de la sécheresse. Par exemple
chez le maïs, un QTL de la teneur en ABA associé au
développement des racines a été identifié (TUBERO
et al., 1998). Chez Arabidopsis thaliana, un QTL lié
à une forte accumulation d'ABA dans les feuilles est associé
à une augmentation de rendement en conditions stressantes, a
été identifié (LANDI et al., 2001). Un QTL
favorisant le maintien de la croissance foliaire en cas de stress hydrique a
été aussi identifié (REYMOND et al., 2003).
L'identification de QTL favorable pour un trait donné est celle de
gènes impliqués dans les réponses d'adaptation à la
sécheresse sont des outils biotechnologiques permettant aux
sélectionneurs de cribler les individus afin d'accélérer
le processus de sélection pour l'amélioration
génétique.
1.5.6.4b Acide abscissique
L'Acide Abscissique (ABA) est une hormone
végétale, responsable de la sénescence des feuilles et
produite en cas de stress hydrique ou du vieillissement des feuilles dans des
conditions normales. Cette hormone envoie des signaux impliquant, des facteurs
de transcription nommés AREB (ABA Responsitive Element Binding) (ABE et
al., 1997; UNO et al., 2000).
Selon YAMAGUSHI-SHINOZAKI et al., (2005), il y a
aussi les facteurs DREB (Drought Responsive Element Binding) qui sont
impliqués dans la réponse à la sécheresse. L'ABA
est un effecteur de fermeture des stomates ou de synthèse
d'osmoprotectants (TARDIEU et al., 1993b).
1.5.6.4c Les protéines et enzymes
De nombreuses études ont montré l'implication
des protéines et des enzymes dans la réponse des plantes en
conditions stressantes. En effet, les protéines du groupe LEA
(Late-Embryogenesis-Abundant protéines) sont des protéines qui
s'accumulent au cours des dernières étapes de
l'embryogenèse et dans des conditions stressantes (RAMANJULU et
al., 2002).
Les déshydrines sont immunologiquement distinctes des
protéines, connues sous le nom de LEA D11. Ces LEA D11 sont de sous
groupe des LEA (DURE et al., 1989) et elles ont été
décrites chez de nombreuses espèces végétales (BRAY
et al., 1997). Par exemple chez le tournesol un ADNc de ces
protéines, Ha Dhn1, induit par le stress hydrique a été
isolé (OUVRARD et al., 1996). Les enzymes qui sont
impliquées en cas de stress hydrique lors d'un processus photochimique
sont principalement: la superoxide dismutase (SOD), l'ascorbate peroxydase
(APX), catalase (CAT), la glutahion-S-transférase (GST) et la glutathion
peroxydase (GPX). Selon REDDY et al., (2004), ces enzymes sont des
antioxydants lors d'un stress oxydatif.
CHAPITRE II: MATERIEL & METHODES
II.1 Matériel
2.1.1 Matériel
végétal
Le matériel utilisé est constitué de 20
accessions d'arachide locales et améliorées de la banque de
gènes de l'ICRISAT Sadoré (Niamey/Niger). Cinq types taxonomiques
(fastigiata vulgaris Spanish (VUL), fastigiata fastigiata Valancia
(FST), fastigiata peruviana Valencia peruviana (PRU) hypogaea runner Virginia
runner (HYR), hypogaea bunch Virginia bunch (HYB)), de diverses origines
(Mali, Sénégal etc.) et de deux statuts biologiques (9
améliorées et 11 locales) ont été
sélectionnés pour les expérimentations en conditions
naturelles et contrôlées.
Tableau I: Taxon, origine,
donneur et iso donneur du matériel végétal
utilisé
|
Code
|
Génotypes
|
Année
de-régénération
|
Taxonomique
|
BST
|
Sources
|
Donneur
|
Iso-donneur
|
|
A1
|
ICG 10005
|
2009/2010
|
PRU
|
LR
|
5PER
|
IAC
|
IND
|
|
A2
|
ICG 8352
|
2008/2009
|
HYB
|
BL
|
9TWN
|
IAC
|
IND
|
|
A3
|
ICG 8385
|
2009/2010
|
HYB
|
BL
|
9USA
|
IAC
|
IND
|
|
A4
|
ICG 8398
|
1998/2009
|
HYB
|
BL
|
9USA
|
IAC
|
IND
|
|
A5
|
ICG 8429
|
2009/2010
|
HYR
|
LR
|
2MWI
|
IAC
|
IND
|
|
A6
|
ICG 8431
|
1999/2010
|
VUL
|
LR
|
2MWI
|
IAC
|
IND
|
|
A7
|
ICG 8439
|
2009/2010
|
HYR
|
LR
|
5MWI
|
IAC
|
IND
|
|
A8
|
ICG 8443
|
2005/2008
|
VUL
|
LR
|
9ZAF
|
IAC
|
IND
|
|
A9
|
ICG 8454
|
1996/1999
|
VUL
|
BL
|
9ZWE
|
IAC
|
IND
|
|
A10
|
ICG 8460
|
1999/2007
|
VUL
|
LR
|
9ZWE
|
IAC
|
IND
|
|
A11
|
ICG 8550
|
1996/1999
|
FST
|
LR
|
9MDG
|
IAC
|
IND
|
|
A12
|
ICG 8594
|
1998/2008
|
PRU
|
LR
|
9ZAR
|
IAC
|
IND
|
|
A13
|
ICG 8606
|
2008/2010
|
FST
|
BL
|
9WZE
|
IAC
|
IND
|
|
A14
|
ICG 8724
|
1999/2009
|
HYB
|
BL
|
9MLI
|
IAC
|
IND
|
|
A15
|
ICG 8746
|
1997/2007
|
HYR
|
BL
|
9SEN
|
IAC
|
IND
|
|
A16
|
ICG 8781
|
1999/2009
|
HYR
|
LR
|
2ZMB
|
IAC
|
IND
|
|
A17
|
ICG 8848
|
2001/2005
|
FST
|
BL
|
9ZWE
|
IAC
|
IND
|
|
A18
|
ICG 8863
|
2001/2007
|
FST
|
BL
|
9ZWE
|
IAC
|
IND
|
|
A19
|
ICG 9989
|
2009/2010
|
PRU
|
LR
|
9PER
|
IAC
|
IND
|
|
A20
|
ICG 9991
|
2009/2010
|
PRU
|
LR
|
5PER
|
IAC
|
IND
|
A: Accession
Génotypes: ICG, Icrisat
Centre Groundnut (Arachide). .
Taxonomique: VUL,
Vulgaris; FST, Fastigiata; PRU, Perivian; HYR, Hirsuta; HYB, Bunch.
BST: BL, breeder's line; LR,
Landrace.
Sources: MLI, Mali; MWI, Malawi;
MDG, Madagascar; PER, Pérou; SEN, Sénégal; TWN, Taiwan;
USA, Etat Unies d'Amérique; ZAF, Afrique du Sud; ZAR, Zaïre; ZWE,
Zimbabwe; ZMB, Zombie.
Donneurs: IAC, Collection
international d'Arachide
Iso donneurs: IND, Inde
2.1.2 Site expérimental
Les essais ont été conduits au sein de la
station de recherche de l'ICRISAT Sadoré/Niamey/Niger. Le centre
s'étend sur 500 ha et est situé à 40 km au Sud-Est de
Niamey capitale du Niger, à 22 km Nord-Ouest de Say et 7 km à
l'Ouest du fleuve Niger. Il se situe entre latitude 13°15'N et longitude
2°18'E.
II.2 Méthodes
2.2.1 Essai en conditions
naturelles
2.2.1.2 Dispositif
expérimental.
Le dispositif utilisé est un bloc de Fisher
randomisé (Annexe 4). Les graines des 20 accessions ont
été mises en culture le 26 Août 2013, selon le dispositif
de blocs de Fisher Randomisés avec trois (3) répétitions
afin d'étudier leurs caractéristiques agro-morphologiques. Les
graines ont été semées après une pluie de 18,3 mm
(25 Août 2013) sur un sol sableux, une application de fumure de fond
(minéral et organique) et un labour. Trois (3) graines ont
été semées par poquet dans les conditions suivantes:
température maximale (31,1 °C), température minimale (24
°C); températures du sol à 10 cm de profondeur: 24,2 °C
et 32,7° C à 7 h et 13 h respectivement soit une température
moyenne journalière du sol de 28,45° C; températures du sol
à 5 cm: 26,1° C et 35° C à 7 h et 13 h respectivement
soit une température moyenne du sol de 30,5° C; humidité
relative maximale et minimale sont 96 et 69 % respectivement soit 82,5 %
d'humidité relative moyenne; la vitesse du vent était de 1,4
km/h. Chaque répétition est constituée de 20 parcelles
élémentaires de dimension de 4800 cm2 (60 cm x 80 cm).
Les écartements entre les répétitions d'une part, et
d'autre part entre les parcelles est de 1 m. Chaque parcelle dispose de deux
lignes de semis écartées de 60 cm. Chaque ligne comportait cinq
(5) poquets écartés de 20 cm
2.2.1.3 Entretien et
traitement
Le démariage a été effectué le
lendemain du premier sarclage (17ème JAS), à raison
d'une plante par poquet dans le but d'éliminer les plantes
blessées lors du premier sarclage. Le deuxième sarclage a eu lieu
au 30ème JAS. Au stade début floraison, il y a eu une
application de phosphate.
La pluviométrie enregistrée durant toute
l'expérimentation était de 54,8 mm repartie de la façon
suivante: le mois d'Août, 34,7 mm repartie en deux jours; le mois de
Septembre, 17, 6 mm en six jours et enfin le mois d'Octobre, 2,5 mm en deux
jours. Le déficit hydrique a été récompensé
par un système d'irrigation (rampe frontale) avec une irrigation de10 mm
d'eau, deux fois par semaine.
2.2.1.4 Collecte des
données
La culture a fait l'objet d'observations du stade semis
jusqu'à la date de récolte afin de noter les vingt-neuf
caractères étudiés dont 15 quantitatifs et 14 qualitatifs.
Par ailleurs toutes les mesures ont faites sur l'ensemble des plantes de chaque
parcelle élémentaire, excepté la surface foliaire. Quant
à l'effet de la conservation sur le taux de germination des semences
d'arachide, le taux moyen de germination des semences conservées a
été déterminé en calculant les taux moyens de
l'ensemble des semences conservées pendant 3; 4; 5; 6; 8 et 14 ans. Les
données climatiques ont été prises auprès du
service météo de la station de recherche.
2.2.1.4a Caractères quantitatifs
Toutes les variables quantitatives ont été
mesurées au champ à l'exception des variables post
récoltes et la teneur en huile (quantifiée au laboratoire par
extraction).
Le taux de germination (TAG) a été
calculé au 10ème JAS; ce pourcentage a consisté
à calculer le rapport entre le nombre de graines germées et le
nombre total des graines semées multiplier par cent.
Le taux de levée (T) a été calculé
au 15ème JAS; il correspond au rapport entre les plantules
viables et le nombre total de plantes.
Le début de floraison (DFL) correspond au nombre de
jours passés du semis jusqu'à l'apparition de la première
fleur de chaque plante.
La date de cinquante pour cent de floraison (50% F) correspond
à la date à laquelle la moitié des plantes par accessions
a fleurit.
La surface foliaire (SF) a été mesurée
à l'aide d'un appareil Area Meter LI 3100. Sa détermination a
consisté à prélever la troisième feuille de la tige
principale de trois plantes par accession dans une répétition,
puis la scannée avec cet appareil.
La hauteur de la plante (HAP) a été
mesurée au 70 ème après levée, c'est
à dire 80ème JAS, avec un traçoir gradué
et 40 cm de longueur. Elle a été mesurée depuis le collet
jusqu'au bourgeon terminal de la tige principale de la plante de toutes les
plantes de la parcelle.
L'indice de surface foliaire (LAI) a été
déterminé au 70ème JAS avec l'appareil LAI
-2000PCA.
La durée du cycle (DC) a été notée
du 70ème au 100ème JAS; elle correspond le
nombre de jours passés du semis jusqu'à la maturité des
gousses.
Le nombre des gousses par plante (NGP) a été
compté au moment de la récolte (107 au 108ème
JAS soit deux jours de récolte), il correspond le rapport entre le
nombre de gousses récoltées et le nombre total de plantes
retenues.
Le rendement en gousses par plante (RGP) a été
déterminé, il correspond au rapport entre le poids des gousses et
le nombre total de plantes donatrices de ces gousses, après 14 jours de
séchage sous la température ambiante et à
l'éclairement solaire (23/12/2013).
Le nombre de réticulation par gousse (NRG) a
été noté après la détermination de RGP; il a
consisté à compter le nombre de réticulations par
gousse.
Le nombre maximum de graines par gousse (Nmg/G) a
été noté au moment du comptage de gousses et
confirmé au moment du décorticage.
Le rendement en graines par plante (RgP) a
été calculé après 5 jours de décorticage des
gousses; il correspond au rapport entre le poids de graines et le nombre total
de plantes.
Le poids moyen d'une graine (PMg) a été
déterminé après décorticage des gousses et
élimination des graines défectueuses (graines germées,
attaquées par les termites, triquées, trouées et
pourries). Sa détermination a consisté à prendre une
poignée de graines à la main au hasard, ensuite à peser
avec une balance de précision (d=0,0001) et compter le nombre de graines
de cette poignée. Puis le rapport a été calculé
entre le poids de cet échantillon et le nombre de graines retenues.
La teneur en huile (TEH) a été quantifiée
par l'extraction d'huile. L'huile a été extraite à partir
de l'hexane dans un appareil de Soxhlet pendant 3 heures (Annexe 1).
2.2.1.4b Caractères qualitatifs
La détermination des caractères qualitatifs
s'est effectuée au champ et au laboratoire.
La forme de la foliole (FFO) a été
enregistrée au stade de développement complet de la
troisième feuille de la tige principale (IBPGR et al.,
1992).
Le port de la plante (PPL) a été observé
au 80ème JAS. La sensibilité aux maladies (SAM) a
été observée tout au long de l'expérimentation.
La couleur de la fleur (CFL), le type d'inflorescence (TIF) et
la présence de fleurs sur la tige principale (FPT) ont été
enregistrés du stade floraison au stade gynophorisation.
La pigmentation des gynophores (PGY), des tiges (PGT) et le
niveau de ramification (NRA) ont été notées au moment de
la récolte.
Les types de bec de la gousse (TBG), d'étranglement de
la gousse (TEG), de réticulation de la gousse (TRG), la compacité
de la gousse (COG), la couleur de la graine (Cg) ont été
notés au moment de décorticage.
Tableau II:
Liste des paramètres étudiés
|
Caractères quantitatifs
|
Caractères qualitatifs
|
|
Paramètres
|
Abréviations
|
Paramètres
|
Abréviations
|
|
1. Taux de germination
|
TAG (%)
|
1. Forme de la foliole
|
FFO
|
|
2. Levée
|
T (%)
|
2. Port de la plante
|
PPL
|
|
3. Début de floraison
|
DFL (jours)
|
3. Sensibilité aux maladies
|
SAM
|
|
4. Cinquante pour cent de floraison
|
50 % F (jours)
|
4. Couleur de la fleur
|
CFL
|
|
5. Surface foliaire
|
SF (cm2)
|
5. Type d'inflorescence
|
TIF
|
|
6. Hauteur de la tige principale de la plante
|
HPL (cm)
|
6. Fleur présente sur la tige principale
|
FPT
|
|
7. Indice de surface foliaire
|
LAI
|
7. Pigmentation du gynophore
|
PGY
|
|
8. Durée du cycle
|
DC (jours)
|
8. Pigmentation de la tige
|
PGT
|
|
9. Nombre de gousses par plante
|
NGP
|
9. Niveau de ramification
|
NRA
|
|
10. Rendement en gousses par plante
|
RGP (g/plante)
|
10. Type de bec
de la gousse
|
TBG
|
|
11. Nombre de réticulations par gousse
|
NRG
|
11. Type 'd'étranglement de la gousse
|
TEG
|
|
12. Nombre maximum de graines par gousse
|
Nmg/G
|
12. Type de réticulation de la gousse
|
TRG
|
|
13. Poids moyen d'une graine
|
PMg (g)
|
13. Compacité de la gousse
|
COG
|
|
14. Rendement en graines par plante
|
RgP (g/plante)
|
14. Couleur de la graine
|
Cg
|
|
15. Teneur en huile
|
TEH (%)
|
|
2.2.2 Essai en conditions contrôlées
2.2.2.1 Dispositif
expérimental
L'essai en contions contrôlées a
été mené au sein de la même station de recherche
dans des pots selon un dispositif de bloc de Fisher randomisé avec trois
(3) répétitions et quatre (4) traitements (T0,
T1, T2, T3) (Annexe 5). Les mêmes
accessions d'arachide ont été reconduites en conditions
contrôlées afin d'évaluer leur performance vis à vis
de la sécheresse. Chaque accession est présentée par un
seul pot dans un traitement. Ce qui donne un total de 240 pots (20
variétés x 1 pot/variété x 4 variantes de
traitement x 3 répétitions) pour cet essai. Le semis a
été fait le 18/09/2013, en raison de trois graines par pot (25 cm
de profondeur, 95 cm de diamètre supérieur et 93 cm de
diamètre inférieur).
Le fond de chaque pot est préalablement perforé
(3 trous). Chaque pot contient un substrat composé de 4,33 kg de
gravions (première couche inférieure) pour permettre
l'aération des racines lors de la formation des gousses et la
détermination de la quantité d'eau nécessaire par pot et
une couche supérieure (50 % du sable lessivé, 25 % argile et 25 %
fumure organique). La quantité du substrat allouée par pot
était de 22,896 kg (18,06 kg du sable et 4,33 kg des gravions).
2.2.2.2 Entretien et
traitement
Lors de cette expérimentation, deux démariages
ont été effectués: un premier au 12ème
JAS et un deuxième au 17ème JAS afin de choisir les
plantules vigoureuses parmi les deux restantes lors du premier
démariage. Trois scarifiages ont été effectués aux
10, 15 et 30ème JAS respectivement dans le but de casser la
croûte du substrat causé par l'arrosage. Lors de cet essai, des
traitements d'insecticide (Décis) et fongicide (0,825 g furadan/pot) ont
été faits. 1 g DAP (Di-ammonium-phosphate) par pot et 1,5 g
phosphate par pot ont été appliqués aux
20ème et 37ème JAS respectivement.
2.2.2.3 Contrainte
hydrique
Toutes les plantes ont été arrosées
régulièrement, en fonction de leur besoin en eau jusqu'à
ce que 14 accessions d'arachide ayant atteint leur stade 50 % de floraison:
c'est à dire lorsque 70 % (14/20*100) d'accessions ayant atteint le
stade 50 % de floraison. Par ailleurs, les autres plantes n'ayant pas atteint
leur stade 50 % F, avaient au moins chacun deux plantes ayant porté des
fleurs. C'est à ce stade que le premier stress hydrique (T1)
a été appliqué. Ce stress hydrique consistait à
interrompre l'arrosage des plantes tandis que l'arrosage était continu
pour les autres plantes selon leur besoin en eau.
Le premier stress hydrique (T1) a
été appliqué du 26ème au
35ème JAS, soit une durée de 10 jours de stress
hydrique.
Après la levée du premier stress hydrique, ces
plantes «stressées» ont subi régulièrement des
stress intermittents (période d'arrosage suivie de période de
stress hydrique). La durée du stress est variable suivant la
température, la vitesse du vent et l'ensoleillement du site
d'expérimentation.
Le traitement T2 a été imposé
du 44ème au 50ème JAS soit une durée
de 7 jours de stress hydrique. Enfin le traitement T3 a été
imposé du 65ème au 70ème JAS soit
une durée de 6 jours de stress hydrique (figure 1).
La durée totale du stress hydrique durant cette
expérimentation varie selon les traitements. En effet, les plantes
«stressées» au stade 50 % F avaient une durée de
contrainte hydrique de 47 jours de stress intermittents. Celles
«stressées» aux stades de fructification et de remplissage des
gousses avaient des durées de 31 et 24 jours de stress intermittents
respectivement (Annexe 2).
Plante témoin
Figure 1: Photo des plantes
témoins et stressées aux différents traitements
Figure Photo 1: Plantes témoins et stressées aux
différents traitements
Plante stressée remplissage de gousses
Plante stressée 50% F
Plante stressée au stade fructification
   
2.2.2.4 Collecte des
données
2.2.2.4 a Paramètres climatiques
Afin de connaître les conditions climatiques dans
lesquelles les essais ont été conduits, un
thermo-hygromètre a été installé sur le site
d'expérimentation. Cet appareil enregistre la température et
l'humidité relative à chaque 30 mn. Ces paramètres
climatiques permettent de localiser la période à laquelle la
contrainte hydrique devient sévère pour la croissance et le
développement de la plante. La moyenne journalière de ces
paramètres (température et humidité relative) a
été calculée. Elle consistait à calculer la moyenne
de 48 valeurs enregistrées par jour. L'intervalle d'enregistrement de
cet appareil est compris entre 0 °C et 98°C pour la
température entre 25 % et 96 % pour l'humidité relative (HR).
2.2.2.4b Caractères Morphologiques
Lors de cette expérimentation en pots, des
observations ont été faites afin d'appliquer les
différents traitements et de mesurer les caractères. En plus des
observations matinales, toutes les 13 heures, des observations
supplémentaires ont été aussi faites durant toute
l'expérimentation afin de noter le degré de flétrissement
des plantes pendant le stress hydrique. Les caractères
étudiés sont les suivants:
2.2.2.4b 1 Surface foliaire (SF)
La détermination de la surface foliaire a
été faite aussi bien pour les plantes «témoins»
que pour les plantes «stressées» uniquement au stade 50 % de
floraison. En effet, pour les plantes «témoins», elle a
porté sur les mêmes feuilles qui ont été
utilisées pour la détermination de la teneur en eau par rapport
à la masse sèche (TE/MS). En plus de cela, un autre
prélèvement des feuilles a été effectué sur
les plantes «stressées». Ce prélèvement a
concerné la troisième feuille de la tige principale des plantes
«stressées», juste au niveau de l'intersection
stipule-pétiole, neuf jours après la levée de la
deuxième série de la contrainte hydrique appliquée au
stade 50 % F (60ème JAS); période correspondant
à la reprise de la croissance et du développement des plantes
«stressées».
La détermination de la surface foliaire a
été effectuée par le même appareil qui a servi
à la mesure de la surface foliaire des accessions d'arachide en
conditions naturelles.
2.2.2.4b.2 Biomasse sèche (BS)
Après avoir enlevé les gousses, les fanes
fraîches ont été exposées deux heures à l'air
libre puis mises dans des enveloppes (après avoir secoué ces
fanes afin d'éliminer le sable qui adhère aux racines). Ensuite
ces enveloppes ont été exposées à
l'éclairement solaire pendant trois jours. Avant de les
transférer à l'étuve à 100°C pendant 24
heures, les enveloppes ont été pesées. La biomasse
sèche à été déterminée par la formule
suivante: BS= Pf-Pv avec BS: biomasse sèche, Pf: poids d'enveloppe
contenant les fanes, Pv: poids d'enveloppe vide; cette biomasse exprime en
gramme par plante.
2.2.2.4b.3 Rendement et ses composantes
Le rendement d'une culture est un caractère complexe
à faible héritabilité. Il est le plus souvent
déterminé à partir de ses composantes. Ses composantes
sont variables selon les spéculations. Chez l'arachide, les composantes
du rendement peuvent être le nombre des gynophores fertiles, l'envergure,
le nombre des gousses, le nombre de graines par gousse, le poids moyen d'une
graine et le taux de maturité. Le rendement peut être
évalué par plante ou par superficie. Il peut être
déterminé à partir de poids des gousses, poids moyen d'une
graine ou encore poids des graines par plante ou par superficie. Dans cette
étude le rendement a été déterminé par
plante dans le but de déterminer l'indice de tolérance à
la sécheresse (ITR).
Les composantes qui ont été mises en
évidence dans cette expérimentation en pots sont le rendement en
graines par plante (RgP), le poids moyen d'une graine (PMg) et le nombre de
gousses par plante (NGP). Ce NGP a été déterminé au
moment de la récolte (101ème JAS) afin de s'assurer du
nombre exact de gousses par plante. Les plantes ont été
arrosées au 100ème JAS afin de faciliter la
récolte. La récolte a été faite par
répétition et par traitement.
2.2.2.4c Détermination des indices
Afin d'identifier les accessions de meilleur rendement en
graines, à caractère fourrager, résistante et
tolérante à la sécheresse, des indices ont
été calculés. L'indice de récolte (Ir) a
été calculé par le rapport entre le rendement en graines
par plante (RgP) et la biomasse sèche (BS) de la plante.
L'indice de résistance à la sécheresse
(IRS) a été calculé par la formule de WILSON et
al., (1978). Il est égale au rapport entre la masse de la
matière sèche des plantes «stressées» (MMSS) et
la masse de la matière sèche des plantes
«témoins» (MMST).
L'indice de tolérance à la sécheresse a
été calculé par la formule utilisée par SEIFALLAH
(2007). Il correspond au rapport entre le rendement en graines des plantes
«stressées» (RgPs) et celui des plantes
«témoins» (RgPt).
2.2.2.4d Caractères Physiologiques
Dans le but de connaître les réponses des
accessions d'arachide vis-à-vis de la sécheresse, 4
paramètres physiologiques ont été fixés. Le
déficit hydrique (DH) a été calculé selon la
formule de GABRIEL CORNIC (2008). Il a concerné sur les mêmes
feuilles utilisées pour la détermination de laT.Ch et de la TRE.
La teneur en eau par rapport à la masse sèche a été
déterminée au stade 50% de floraison.
La teneur en chlorophylle (T.Ch) a été
déterminée juste avant la levée de chaque première
contrainte hydrique imposée aux différents stades de
développement de la plante (50 % F, fructification et remplissage des
gousses) au niveau de la troisième feuille de la tige principale des
plantes «témoin» et «stressées». Cette
détermination de la teneur de la chlorophylle a été faite
sur toutes les folioles de la troisième feuille de la tige principale
à l'aide d'un lecteur de chlorophylle (SPAD) au moment de
l'ensoleillement.
La teneur relative en eau des feuilles (TRE) a
été calculée selon la formule de CLARK et al.,
(1982). Cette formule a été modifiée à trois
niveaux que sont:
l'utilisation des folioles au lieu des disques foliaires;
la température d'imbibition: 2° C pendant 24 h au
lieu de 4°C pendant 24 h;
la durée de séchage: 75 °C pendant 48 h au
lieu de 70 °C pendant 48 h.
La détermination de cette teneur relative en eau (TRE)
a été réalisée sur les mêmes feuilles
utilisées lors de la mesure de la teneur en chlorophylle.
Tableau III: Listes de
caractères physiologiques et des indices
|
Paramètres
|
Abréviations
|
Formules
|
|
1. Déficit hydrique
|
DH
|
DH (%) =100*(Ms-Ma)/MS
|
|
2. Teneur en eau par rapport à la masse sèche
|
TE/MS
|
TE/MS (%) = (MF-MS)*100)/MS
|
|
3. Teneur relative en eau des feuilles
|
TRE
|
TRE (%)=100* (PF-PS)/(PT-PS)
|
|
4. Teneur en chlorophylle
|
T.Ch
|
T.Ch (mg/g Mf)
|
|
5. Teneur relative en eau
|
TRE
|
TRE(%)=100* (PF-PS/PT-PS)
|
|
1. Indice de récolte
|
Ir
|
Ir= RgP/BS
|
|
2. Indice de résistance à la
sécheresse
|
IRS
|
IRS= MMSS/MMST
|
|
|
3. Indice de tolérance à la sécheresse
|
ITS
|
ITS= RgPs/RgPt
|
2.2.3 Analyse des
données
Afin d'obtenir des données statistiques, les
données collectées (données brutes) ont été
traitées par le logiciel Excel 2010. Ce même logiciel a
servi aussi d'établir les courbes des paramètres climatiques sous
lesquels les accessions ont été évaluées.
Les analyses de variance des différents
paramètres ont été faites par le logiciel XLSTAT
version 7.1 avec un intervalle de confiance de 95 % soit un risque
alpha de 5 %. Afin de regrouper les accessions suivant leurs
performances et en fonction des objectifs attendus, trois types de test ont
été retenus à savoir le test de ppds (plus petite
différence significative protégée), le test de Duncan et
le test de Tukey. Ce même logiciel a servi pour l'AFD (Analyse
Factorielle Discriminant) et l'AFC (Analyse Factorielle de Correspondances)
alors que la CAH (Classification Ascendant Hiérarchisée) a
été réalisée par le logiciel Genstat
10ème version dont la distance euclidienne et la liaison
simple ont été fixées lors de l'analyse.
CHAPITRE III: RESULTATS
&DISCUSSION
III.1 Résultats
3.1.1 En conditions naturelles
Le développement des plantes s'est effectué
sous les paramètres climatiques de la station de recherche de l'ICRISAT.
La température moyenne journalière et la radiation solaire
évoluent ensemble. Plus la radiation solaire est élevée,
plus la température est importante. Ces deux paramètres
augmentent du 26 Août au 5 Septembre et se stabilisent relativement vers
la période froide. Quant à l'humidité relative, elle
était plus importante pendant la saison pluvieuse. Elle diminue
progressivement vers la fin de cette saison et ensuite croît faiblement
à nouveau à partir du début de la période froide.
Cette humidité relative évolue de façon aléatoire
avec la température. Ces deux paramètres avaient presque la
même valeur à la date du 04 Novembre. Cette humidité
relative évolue en antagonisme avec la vitesse du vent, c'est à
dire plus elle augmente, plus la vitesse du vent diminue. Cette vitesse du vent
augmente l'évaporation du bac (figure 2). Les sommes de ces
paramètres climatiques durant l'essai sont: Température
(2939,1° C); pluviométrie moyenne (54,8 mm/j); humidité
relative (5318 %); évaporation du bac (648,3 mm/j); radiation solaire
(2182,3 MJ/m2) et enfin la vitesse du vent a été
estimée à 310,4 m/s. La figure 3 montre l'évolution de la
température journalière au stade remplissage des gousses à
la récolte. Cette température a fluctué entre 27,6° C
et 30,7° C avec une somme de température journalière
(229,3° C).
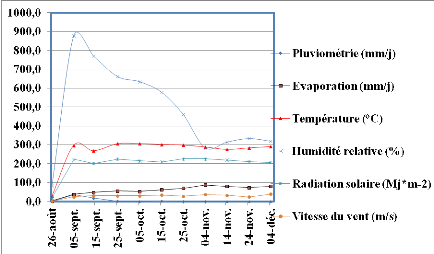 Figure 2: Evolution des
paramètres climatiques durant l'expérimentation au champ. Echelle
(110/113). Figure 2: Evolution des
paramètres climatiques durant l'expérimentation au champ. Echelle
(110/113).
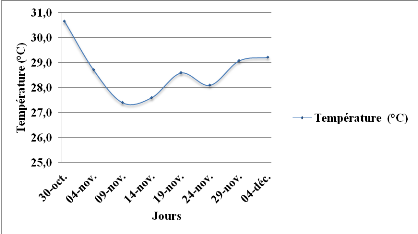
Figure 3: Evolution de la
température journalière durant la période de remplissage
des gousses à la récolte au champ. Echelle (100/112).
3.1.1.1 Effet de la conservation sur le taux de
germination
Le taux moyen de germination était très
élevé pour les accessions d'arachide conservées
«ex situ» durant une courte durée que celles
conservées pendant une durée plus ou moins longue (Tableau IV).
Tableau IV: Effet de la
conservation sur le taux germination
|
Durée de conservation (ans)
|
3
|
4
|
5
|
6
|
8
|
14
|
|
Taux de germination (%)
|
75
|
45,83
|
80
|
53,33
|
60
|
70
|
3.1.1.2 Caractères quantitatifs
Parmi les quinze caractères quantitatifs, cinq
caractères quantitatifs pré-récolte montrent des
différences significatives entre les accessions évaluées.
Par ailleurs, l'analyse de variance n'a pas porté sur les
caractères tels que le début de floraison (DFL), l'indice de
surface foliaire (LAI), le nombre de réticulations par gousse (NRG), le
nombre maximum des graines par gousse (Nmg/G) et le poids moyen d'une graine
(PMg).
Quant aux caractères quantitatifs
post-récolte, le nombre de gousses par plante (NGP), le rendement en
gousses par plante (RGP), le rendement en graines par plante (RgP) ne montrent
aucune différence significative. En outre, le taux de germination (TAG)
varie entre 22,67 % (A15) et 100 % (A13, A19 et A20).
L'ANOVA montre une différence très hautement
significative (F=6,04***) entre les accessions étudiées pour ce
caractère, avec un coefficient de détermination (R2
=74,2 %) et un coefficient de corrélation (R=0,86). Le test de
Tukey regroupe les accessions en quatre sous groupes. Le taux de levée
varie entre 26,67 (A15) et 100 % (A20); ce pourcentage est inférieur
à celui de la germination pour toutes les accessions, à
l'exception des accessions A8, A9, A7, A11, A14, A15, A17, A18 et A20 où
leurs taux de levée sont voisins de leurs taux de germination.
Quant à la date de floraison, les accessions A13 et
A5 fleurissent entre 19 et 25 JAS respectivement (Tableau VI). Alors que la
date de cinquante pour cent de floraison (50 % F) varie entre 23 JAS (A13) et
35 JAS (A15) et une différence très hautement significative
(F=7,07***) a été détectée. Les coefficients de
détermination et de corrélation sont respectivement
R2=66,2 % et R=0,87. Pour le test de ppds, sept sous groupes ont
été distingués.
La surface foliaire des accessions d'arachide varie entre
13, 951 cm2 (A14) et 34,73 cm2 (A17) avec une moyenne de
22,30 #177; 8,01 cm2. La différence est significative
(F=2,00*) entre les accessions. Les coefficients de corrélation et de
détermination sont R=0,69 et R2=48,8 %. Trois sous groupes
ont été identifiés par le test de Duncan.
La hauteur de la plante (HPL) varie entre 7 cm (A15) et
24,16 cm (A13) avec une moyenne de 13,56 #177; 6,5 cm. L'ANOVA montre une
différence très significative (F=2,66**) entre les accessions
d'arachide dont les coefficients de corrélation et de
détermination sont R=0,74 et R2=55,8 % respectivement. Selon
le test de Duncan, les accessions ont été classées en
quatre sous groupes.
La durée du cycle s'étale de 80 jours (A8)
à 98,33 jours (A1) avec une moyenne de 90,75 #177; 6,70 jours. L'analyse
de variance montre une différence hautement significative (F=2,95**).
Les coefficients de corrélation et de détermination sont
respectivement R=0,76 et R2=58,4 %. Le test de ppds regroupe les
accessions en six sous groupes.
Le nombre de gousses par plante (NGP) se fluctue entre
18,60 (A20) et 71,33 (A7) avec une moyenne de 36, 65 #177; 21,17 gousses par
plante. L'ANOVA ne montre aucune différence significative
(F=1,20ns). Le test de ppds a permis de classer les accessions en
trois sous entités.
Quant au rendement en gousses par plante (RGP), il varie
entre 9,20 g/plante (A3) et 57,74 g/plante (A7) avec une moyenne de 21,61 #177;
21,70 g/plante. L'ANOVA ne montre aucune différence significative entre
les moyennes (F=0,11ns). Le rendement en graines par plante (RgP)
varie entre 1,11 g/plante (A19) et 30,83 g/plante (A7) avec une moyenne de 9,03
#177; 13,43 g/plante. Le test de ppds reclasse les accessions en trois groupes.
Pour la teneur en huile des accessions d'arachide, elle
varie entre 36,30 % (A8) et 52,80 % (A20, A2) avec une moyenne de 44,92#177;
12,34 %.
L'analyse de variance ne montre aucune différence
significative (F=0,29ns) entre les accessions d'arachide au risque
alpha égale à 5 %. L'analyse de variance des caractères
au sein des répétitions ne montre aucune différence
significative pour les taux de germination, date de 50 % de floraison,
durée du cycle, nombre de gousses par plante, rendement en gousses par
plante, rendement en graines par plante, la teneur en huile tandis que des
différences significatives sont enregistrées pour la surface
foliaire (F=4,22*) et la hauteur de la plante (F=3,04*) (Tableau V).
Tableau V: Performance de l'ensemble des accessions en
conditions naturelles.
|
|
caractères
|
TAG (J)
|
50 %F(J)
|
SF (cm2)
|
HPL (cm)
|
DC (J)
|
NGP
|
RGP (g/plante)
|
RgP
(g/plante)
|
TEH (%)
|
|
Accessions
|
Fisher
|
6,041***
|
7,079***
|
2,005*
|
2,657**
|
2,950**
|
1,206ns
|
0,119ns
|
1,496ns
|
0,296ns
|
|
Répétitions
|
Fisher
|
0,0003ns
|
0,131ns
|
4,221*
|
3,034*
|
1,475ns
|
1,988ns
|
1,437ns
|
0,801ns
|
0,135ns
|
|
|
Moyennes
|
64,833
|
26,7
|
22,302
|
13,555
|
90,75
|
36,658
|
21,61
|
9,034
|
44,92
|
|
CV (%)
|
44,2
|
3,7
|
35,9
|
44,7
|
7,4
|
57,8
|
10,05
|
14,87
|
27,5
|
|
Ecart type
|
28,671
|
3,233
|
8,011
|
6,057
|
6,691
|
21,171
|
21,61
|
13,433
|
12,343
|
|
Répétition 1
|
64
|
26,4
|
18,474
|
11,885
|
89,25
|
30,835
|
15,61
|
5,921
|
43,28
|
|
Répétition 2
|
65,5
|
26,8
|
22,254
|
12,625
|
90,25
|
35,365
|
22,728
|
10,509
|
46,56
|
|
Répétition 3
|
64,900
|
26,900
|
25,374
|
16,057
|
92,75
|
43,775
|
26,744
|
10,672
|
|
***, très hautement
significative,**, hautement
significative;*,significative; ns,
non significative; 50 % F, cinquante pourcent
floraison; DC, durée du cycle; CV,
coefficient de variation; HPL, hauteur de la plante;
NGP, nombre de gousses par plante; RGP,
rendement en gousses par plante; RgP, rendement en graine par
plante; rep, répétition; SF,
surface foliaire; TAG, taux de germination;
TEH, teneur en huile.
Tableau VI: Performance des accessions d'arachide pour
les 15 caractères quantitatifs
|
Accessions
|
50 % F(j)
|
TAG (%)
|
SF (cm2)
|
HPL (cm)
|
DC(j)
|
NGP
|
RGP (g/plante)
|
RgP (g/plante)
|
TEH (%)
|
DFL (J)
|
T (%)
|
LAI
|
NRG
|
Nmg/G
|
PMg
|
|
A13
|
22,666a
|
100,000 d
|
27,692 c
|
24,167 d
|
85,000 c
|
33,267 b
|
18,429a
|
5,470 a
|
51,300a
|
19
|
80
|
2,51
|
10
|
4
|
0,7
|
|
A19
|
23,000ab
|
100,000 d
|
21,746 c
|
17,267 d
|
93,333 f
|
19,867a
|
10,076a
|
1,117a
|
36,500a
|
20
|
96,67
|
1,41
|
0
|
4
|
0,6
|
|
A20
|
23,000 b
|
100,000 d
|
24,511 c
|
15,833 d
|
90,000 ef
|
18,600a
|
10,852a
|
1,119a
|
52,800a
|
20
|
100
|
1,26
|
0
|
4
|
0,55
|
|
A6
|
24,000 bc
|
96,667 d
|
30,181 c
|
15,333 cd
|
86,667 cd
|
42,667 c
|
26,508 c
|
18,572 c
|
36,800a
|
23
|
76,67
|
1,58
|
12
|
2
|
0,44
|
|
A12
|
25,000 cd
|
63,333 d
|
24,487 c
|
16,700 d
|
88,333 de
|
27,433 b
|
11,970a
|
1,877a
|
43,800a
|
21
|
56,67
|
1,29
|
14
|
4
|
0,43
|
|
A18
|
25,000 d
|
70,000 d
|
19,037 b
|
13,200 c
|
88,333 e
|
39,067 c
|
18,510a
|
7,870a
|
49,300a
|
22
|
70
|
1,08
|
10
|
3
|
0,44
|
|
A9
|
26,000 de
|
70,000 d
|
26,850 c
|
13,767 c
|
85,000 bc
|
56,633 c
|
53,265 c
|
29,888 c
|
41,500a
|
23
|
70
|
2,22
|
12
|
2
|
0,65
|
|
A16
|
26,000 e
|
73,333 d
|
17,867 b
|
10,500 b
|
91,667 f
|
55,100 c
|
30,943 c
|
12,977 bc
|
41,400a
|
24
|
73,33
|
2,8
|
12
|
3
|
0,52
|
|
A11
|
26,000 e
|
46,667 cd
|
22,808 c
|
21,300 d
|
91,667 f
|
35,367 b
|
15,592a
|
9,612ab
|
49,200a
|
22
|
46,67
|
1,07
|
0
|
3
|
0,56
|
|
A10
|
26,000 e
|
76,333 d
|
29,181 c
|
13,267 c
|
81,667ab
|
33,967 b
|
22,265ab
|
14,114 c
|
50,900a
|
23
|
76,67
|
1,2
|
12
|
2
|
0,42
|
|
A8
|
26,000 e
|
96,667 d
|
20,984 c
|
12,560 bc
|
80,000a
|
30,567 b
|
16,549a
|
5,970a
|
36,300a
|
24
|
96,67
|
1,68
|
11
|
2
|
0,47
|
|
A1
|
27,000 ef
|
30,000 b
|
26,080 c
|
16,967 d
|
98,333 f
|
32,167 b
|
14,883a
|
1,295a
|
42,300a
|
33,333
|
24
|
1,06
|
10
|
3
|
0,4
|
|
A7
|
27,000 f
|
63,333 d
|
14,303 b
|
11,330 b
|
96,667 f
|
71,333 c
|
57,749 c
|
30,834 c
|
51,600a
|
24
|
46,67
|
4,33
|
11
|
3
|
0,62
|
|
A4
|
27,000 f
|
36,667 b
|
18,349 b
|
10,330 b
|
95,000 f
|
43,100 c
|
38,783 c
|
14,234 c
|
51,100a
|
24
|
33,33
|
2,26
|
12
|
3
|
0,91
|
|
A3
|
28,000 f
|
50,000 d
|
20,086 bc
|
8,500 b
|
93,333 f
|
30,200 b
|
9,209a
|
1,774a
|
39,500a
|
24
|
46,67
|
2,23
|
12
|
2
|
0,55
|
|
A17
|
29,000 f
|
60,000 d
|
34,731 c
|
17,567 d
|
88,333 e
|
23,833ab
|
15,252a
|
4,233a
|
44,300a
|
24
|
60
|
1,87
|
12
|
4
|
0,71
|
|
A2
|
29,000 f
|
30,000 ab
|
17,938 b
|
9,400 b
|
95,000 f
|
43,333 c
|
25,078 bc
|
5,742a
|
52,800a
|
23
|
30
|
1,95
|
12
|
2
|
0,55
|
|
A14
|
29,333 f
|
43,333 bc
|
13,951a
|
7,867 ab
|
95,000 f
|
38,967 bc
|
14,210a
|
4,364a
|
37,600a
|
23
|
43,33
|
1,08
|
12
|
2
|
0,48
|
|
A5
|
30,000 f
|
56,667 d
|
20,948 c
|
8,733 b
|
95,000 f
|
24,600 b
|
11,970a
|
5,257a
|
46,1
|
25
|
56,67
|
1,34
|
12
|
2
|
0,53
|
|
A15
|
35,000 g
|
26,667 a
|
14,119ab
|
7,000 a
|
96,667 f
|
33,100 b
|
10,089a
|
4,353a
|
43,3
|
20
|
26,67
|
0,59
|
10
|
2
|
0,31
|
|
Moyenne
|
26,7
|
64,833
|
22,302
|
13,555
|
90,75
|
36,658
|
21,61
|
9,034
|
44,92
|
23,067
|
60,53
|
1,7405
|
9,8
|
2,8
|
0,542
|
|
CV(%)
|
3,7
|
44,2
|
35,9
|
44,7
|
7,4
|
57,8
|
10,05
|
14,87
|
27,5
|
1,128
|
3,88
|
4,82
|
4?442
|
2,98
|
2,48
|
|
ppds /HSD
|
2,021
|
3,728
|
2,021
|
2,453
|
2,021
|
2,021
|
2,021
|
2,021
|
2,086
|
-
|
-
|
-
|
--
|
-
|
-
|
|
Ecart type
|
3,233
|
28,671
|
8,011
|
6,057
|
6,691
|
21,171
|
21,61
|
13,433
|
12,343
|
2,953
|
23,47
|
0,8384
|
4,3359
|
0,834
|
0,134188
|
|
Ficher
|
7,079***
|
6,041***
|
2,005*
|
2,657**
|
2,950**
|
1,206ns
|
0,119ns
|
1,496ns
|
0,296ns
|
-
|
-
|
-
|
-
|
-
|
-
|
***, très hautement
significative;**, très significative;*
significative; 50 % F, cinquante pourcent de floraison;
DC, durée du cycle; DFL, Début
de floraison; HPL, hauteur de la plante; ICG,
Icrisat Centre Groundunt; LAI, indice de surface
foliair ; NGP, nombre de gousses par plante;
PMg, Nmg/G, nombre maximum de graines par gousse;
NRG, nombre de réticulations par gousse; poids moyenne
d'une graine; RGP, rendement en gousses par plante;
SF, surface foliaire; TAG, taux de
germination. Les moyennes suivies par les mêmes lettres ne sont pas
significativement différentes à 95 % de niveau de
confiance.
3.1.1.2a Corrélaton entre les
paramètres
La matrice de corrélation a été
établie afin de savoir si la teneur en huile est corrélée
à d'autres caractères. Cette matrice montre qu'il existe une
corrélation entre la teneur en huile et le poids moyen d'une graine.
Cette corrélation est très faible ( r=0,31 ) (Tableau VII).
Tableau VII: Matrice de
corrélation
|
TAG
|
T
|
DFL
|
50%F
|
SF
|
HPL
|
LAI
|
DC
|
NGP
|
NRG
|
Nmg/G
|
RGP
|
RgP
|
PMg
|
TEH
|
|
TAG
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
T
|
0,961***
|
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
DFL
|
-0,430
|
-0,409
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
|
50 %F
|
-0,776**
|
-0,715
|
**0,324
|
1
|
|
|
|
|
|
|
|
|
|
|
|
|
SF
|
0,424
|
0,398
|
-0,040
|
-0,427
|
1
|
|
|
|
|
|
|
|
|
|
|
|
HPL
|
0,481
|
0,405
|
-0,471
|
-0,674*
|
0,668*
|
1
|
|
|
|
|
|
|
|
|
|
|
LAI
|
0,140
|
0,007
|
0,310
|
-0,183
|
-0,150
|
-0,049
|
1
|
|
|
|
|
|
|
|
|
|
DC
|
-0,680*
|
-0,704
|
*0,262
|
0,489
|
-0,572*
|
-0,393
|
0,054
|
1
|
|
|
|
|
|
|
|
|
NGP
|
-0,172
|
-0,272
|
0,390
|
0,060
|
-0,354
|
-0,272
|
0,701*
|
0,117
|
1
|
|
|
|
|
|
|
|
NRG
|
-0,299
|
-0,362
|
0,513*
|
0,371
|
-0,016
|
-0,446
|
0,257
|
-0,102
|
0,378
|
1
|
|
|
|
|
|
|
Nmg/G
|
0,346
|
0,310
|
-0,445
|
-0,521*
|
0,310
|
0,677*
|
0,107
|
0,107
|
-0,312
|
-0,434
|
1
|
|
|
|
|
|
RGP
|
-0,050
|
-0,138
|
0,339
|
-0,031
|
-0,151
|
-0,171
|
0,7204
|
0,004
|
0,901
|
0,301
|
-0,188
|
1
|
|
|
|
|
RgP
|
-0,050
|
-0,009
|
0,341
|
-0,090
|
-0,022
|
-0,105
|
0,619*
|
0,619*
|
0,856**
|
0,254
|
-0,287
|
0,939***
|
1
|
|
|
|
PMg
|
0,057
|
0,017
|
0,156
|
-0,244
|
0,124
|
0,196
|
0,537*
|
0,025
|
0,163
|
-0,054
|
0,356
|
0,396
|
0,261
|
1
|
|
|
TEH
|
-0,158
|
-0,183
|
-0,073
|
-0,062
|
-0,026
|
0,148
|
0,207
|
0,084
|
0,138
|
-0,120
|
0,253
|
0,226
|
0,135
|
0,312
|
1
|
*, corrélation
faible;** corrélation forte, ***,
corrélation très forte; (+) et
(-) corrélation significativement :positivement et
négativement corrélés respectivement; chiffre en gras
montre une corrélation significative; 50 % F, cinquante
pour cent de floraison; DC, durée du cycle;
DFL, Début de floraison; HPL,
hauteur de la plante; NGP, nombre de gousses par plante;
Nmg/G, nombre maximum de graines par gousse;
NRA, nombre des réticulation par gousse;
PMg, poids moyenne d'une graine; RGP ,
rendement en gousses par plante; RgP, rendement en graine par
plante; SF, surface foliaire; T, taux de
levée; TAG, taux de germination; TEH,
teneur en huile.
3.1.1.2b Description des groupes taxonomiques
Afin de décrire les types taxonomiques des
accessions, une analyse factorielle discriminante (AFD) a été
faite sur la base des quinze caractères retenus lors des observations.
En effet, excepté le taux de germination, 14 caractères
permettant de discriminer les différents types taxonomiques ont
été retenus. Ces types taxonomiques sont statistiquement
différents (Lambda= 0,0000001462, F critique (2,645) < observé
(10,02) au risque 0,05, taux d'erreur d'apparence= 0,0005). Le type taxonomique
fastigiata fastigiata Valencia (FST) est le meilleur taxonomique
riche en huile que les autres types taxonomiques (PRU, VUL, HYR et HYR). Les
carrés de distance de Mahalanobis issus de l'AFD montrent que le type
taxonomique VUL (2129,25) est plus proche de celui de FST que les types HYR,
HYB et PRU.
Les types HYR (2270,58) et HYB (7618,31) sont plus proches de
FST et VUL que PRU (2270,58). Chaque caractère est spécifique au
type taxonomique. Les 14 caractères permettant de discriminer les types
taxonomiques sont présentés dans le tableau VIII.
Tableau VIII: Performance
moyennes des groupes taxonomiques
|
Groupes
|
FST
|
HYB
|
HYR
|
PRU
|
VUL
|
Moyennes
|
Ecart type
|
CV(%)
|
Fisher
|
|
Fréquence
|
4
|
4
|
4
|
4
|
4
|
|
T ( %)
|
64,167
|
38,333
|
50,833
|
70,833
|
80,000
|
60,833
|
16,468
|
27,1
|
2,551ns
|
|
DFL (J)
|
21,750
|
23,500
|
23,250
|
21,250
|
22,750
|
22,5
|
0,968
|
4,3
|
2,167ns
|
|
50% F
|
25,667
|
28,333
|
29,500
|
24,500
|
25,500
|
26,7
|
2,113
|
7,9
|
426,449*
|
|
SF cm2)
|
26,067
|
17,581
|
16,809
|
24,206
|
26,849
|
22,302
|
4,465
|
21,4
|
98,803*
|
|
HPL (cm)
|
19,059
|
8,950
|
9,267
|
16,692
|
13,734
|
13,54
|
4,465
|
33
|
1,543ns
|
|
LAI
|
1,633
|
1,880
|
2,265
|
1,255
|
1,670
|
1,741
|
0,37
|
21,2
|
265,942*
|
|
DC (J)
|
88,333
|
94,583
|
95,000
|
92,500
|
83,334
|
90,7
|
4,916
|
5,4
|
2,245ns
|
|
NGP
|
32,884
|
38,900
|
46,033
|
24,517
|
40,959
|
36,659
|
8,262
|
22,5
|
2,458ns
|
|
NRG(g)
|
8,000
|
12,000
|
11,250
|
6,000
|
11,750
|
9,8
|
2,666
|
27,2
|
1,014ns
|
|
Nmg/G
|
3,500
|
2,250
|
2,500
|
3,750
|
2,000
|
2,8
|
0,779
|
27,8
|
0,385ns
|
|
RGP (g)
|
16,946
|
21,820
|
29,188
|
11,945
|
29,647
|
21,909
|
7,694
|
35,1
|
0,645ns
|
|
RgP (g)
|
6,797
|
6,529
|
13,355
|
1,352
|
17,136
|
9,034
|
6,217
|
68,8
|
0,631ns
|
|
PMg (g)
|
0,603
|
0,623
|
0,495
|
0,495
|
0,495
|
0,542
|
0,065
|
12
|
0,633ns
|
|
TEH (%)
|
48,525
|
45,250
|
45,600
|
43,850
|
41,375
|
44,92
|
2,611
|
5,8
|
0,022ns
|
*, significative;
ns, non significative; DC, durée du
cycle; DFL, début de floraison; F,
fréquence; FST, fastigiata fastigiata Valancia;
HPL, hauteur de la plante; HYB, hypogaea
bunch Virginia bunch; HYR, hypogaea runner Virginia runner
LAI, indice de surface foliaire; NGP, nombre
des gousses par plante; Nmg/G, nombre maximum de graines par
gousse ;PMg, poids moyen d'une graine; PRU,
fastigiata peruviana Valencia peruviana; SF, surface foliaire;
RGP, rendement en gousses par plante; RgP,
rendement en graines par plante; TEH, teneur en huile
VUL, fastigiata vulgaris Spanish.
3.1.1.3 Caractères qualitatifs
Les observations faites sur les quatorze caractères
qualitatifs montrent que douze d'entre eux présentent plus d'une
modalité. En effet, les accessions A1, A2, A3, A5, A10, A11, A16, A18,
A19 et A20 sont des accessions dont la forme de leur foliole est
lancéolée. Les accessions A8, A12, A13 sont des accessions de
forme oblongue, et les accessions A7, A9 et A4, A6, A14, A15, A17 sont de forme
ovale et obovale respectivement.
Quatre types de port ont été distingués.
Les accessions A2, A3 et A4 ont un port buissonnant. Les accessions A16, A15,
A14, A5, A7, A1 et A9 sont des accessions à port rampant 1 et rampant 3
respectivement.
Les A11, A13 et A17 ont un port semi-érigé et
les accessions A6, A8, A10, A18, A20, A19, A12 sont des accessions à
port érigé.
Pour la présence ou l'absence de fleurs sur la tige
principale, la fleur est absente chez toutes les accessions à port
rampant 1 et buissonnant. Elle est variable chez les ports érigés
(absence ou présence) et présente chez les semi
érigés.
Les tiges des accessions rampantes et buissonnantes ne sont
pas pigmentées. La pigmentation peut être absente ou
présente chez les accessions à port érigé et
complètement présente chez les accessions à port semi
érigé. Toutes les accessions ont des gynophores
pigmentées, hormis l'accession A1.
Le niveau de ramification varie entre n+1 (A13, A10) et n+3
(A7, A9). Du point de vue pathologique c'est-à-dire du point de vue de
la sensibilité aux maladies, les accessions A6, A8, A2, A9, A10, A16
sont sensibles aux cercosporioses alors que A3 est sensible à la
pourriture du collet respectivement, mais avec une faible incidence.
Pour les caractères qualitatifs pris en période
post-récolte, quatre sous groupes sont identifiés au sein de ces
20 accessions pour les types de bec de la gousse (TBG) et la compacité
de la gousse (COG).
Quatre types de bec (léger: L, modéré:
M, profond: pr, sans bec: Sb) et quatre type de compacité
(légère: L, moyenne: Mo, dure: D et très dure: TD) ont
été notés. Cinq modalités ont été
notées pour les types de réticulation de la gousse (TRG),
d'étranglement de la gousse (TEG). La couleur de la fleur et le type
d'inflorescence sont communs pour toutes les accessions étudiées.
Quant à la couleur de la graine, huit couleurs ont été
identifiées au cours de cette étude. Les graines ont les
mêmes couleurs que celles qui ont été semées.
L'analyse factorielle des correspondances (AFC) montre, qu'il
y a des caractères qui sont propres à certaines accessions d'une
part et d'autre part des caractères permettant de différencier
les accessions qui sont plus ou moins semblable. En effet, les types de
réticulation et d'étranglement de la gousse sont
spécifiques à A11 alors que la couleur de la graine (tanné
très pâle) est liée à A8, la non pigmentation du
gynophore est propre à l'accession A1 (figure 4).
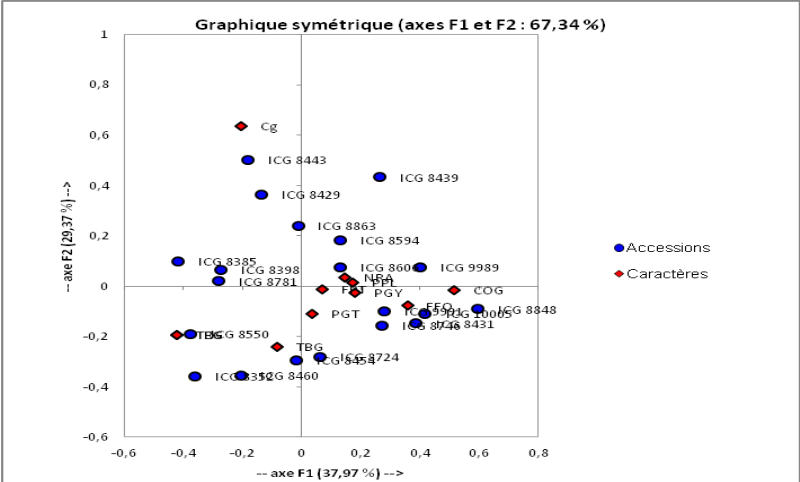
Figure 4:
AFC et classification des individus. Echelle:(28/82)
Lorsque les caractères sont pris ensemble, certaines
accessions sont proches les unes des autres. Ainsi, les accessions A15 et A16
sont proches; les A19 et A18 forment un sous groupe et enfin A3 et A16 sont
intimement liées. La figure 5 du dendrogramme confirme la
proximité des accessions. Ce dendrogramme montre, que les accessions A9,
A7, A17 et A1 forme chacune un sous groupe à part tandis que, les
accessions A15, A14, A5, A4, A16, A3, A2 et A19, A18, A12, A8, A13, A20, A11,
A10, A1 forment les sous groupes 1 et 2 respectivement. Les accessions de ces
deux sous groupes, excepté les accessions A9 et A6 appartiennent aux
sous espèces hyppogaea et fastigiata
respectivement. 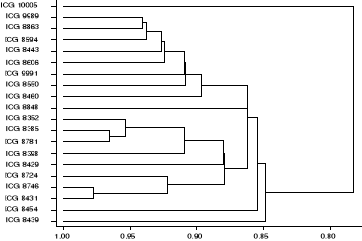
Figure
5: Dendrogramme issu de la classification ascendante
hiérarchique des accessions d'arachide sur la base de la distribution de
leurs caractères qualitatifs. Echelle (146/199)
Tableau IX: Caractères qualitatifs des 20
accessions d'arachide (Arachis hypogaea L.).
|
Accessions
|
CFL
|
FFO
|
FPT
|
TIF
|
SAM
|
PPL
|
PGT
|
PGY
|
NRA
|
TBG
|
TEG
|
TRG
|
COG
|
Cg
|
|
A1
|
J.Org
|
Lancéolée
|
0
|
C
|
0
|
Rampant 1
|
0
|
0
|
n+2
|
Mi
|
L
|
TPr
|
TD
|
Rose
|
|
A2
|
J.Org
|
Lancéolée
|
0
|
C
|
1
|
Buissonnant
|
0
|
+
|
n+2
|
P
|
pr
|
Pr
|
D
|
Rose
|
|
A3
|
J.Org
|
Lancéolée
|
0
|
C
|
1
|
Buissonnant
|
0
|
+
|
n+2
|
P
|
pr
|
M
|
D
|
Tannée claire
|
|
A4
|
J.Org
|
Obovale
|
0
|
C
|
0
|
Buissonnant
|
0
|
+
|
n+2
|
P
|
pr
|
Pr
|
D
|
Tannée claire
|
|
A5
|
J.Org
|
Lancéolée
|
0
|
C
|
0
|
Rampant 1
|
0
|
+
|
n+2
|
P
|
M
|
L
|
D
|
Tannée foncée
|
|
A6
|
J.Org
|
Obovale
|
0
|
C
|
2
|
Erigé
|
0
|
+
|
n+2
|
Mi
|
L
|
M
|
L
|
Rose
|
|
A7
|
J.Org
|
Ovale
|
0
|
C
|
0
|
Rampant 1
|
0
|
+
|
n+3
|
M
|
L
|
Pr
|
TD
|
Tannée foncée
|
|
A8
|
J.Org
|
Oblongue
|
+
|
C
|
2
|
Erigé
|
+
|
+
|
n+2
|
M
|
pr
|
M
|
L
|
Tannée très pâle
|
|
A9
|
J.Org
|
Ovale
|
+
|
C
|
1
|
Rampant 3
|
0
|
+
|
n+3
|
Sb
|
Se
|
L
|
L
|
Rose
|
|
A10
|
J.Org
|
Lancéolée
|
+
|
C
|
2
|
Erigé
|
0
|
+
|
n+1
|
Sb
|
Se
|
L
|
L
|
Rose
|
|
A11
|
J.Org
|
Lancéolée
|
+
|
C
|
0
|
Semi érigé
|
++
|
+
|
n+2
|
Mi
|
Se
|
Sr
|
D
|
Rouge
|
|
A12
|
J.Org
|
Oblongue
|
+
|
C
|
0
|
Erigé
|
+
|
+
|
n+2
|
Mi
|
L
|
L
|
D
|
Rouge claire
|
|
A13
|
J.Org
|
Oblongue
|
+
|
C
|
0
|
Semi érigé
|
++
|
+
|
n+1
|
M
|
L
|
L
|
D
|
Rouge
|
|
A14
|
J.Org
|
Obovale
|
0
|
C
|
0
|
Rampant 1
|
0
|
+
|
n+2
|
P
|
Se
|
M
|
Mo
|
Rose
|
|
A15
|
J.Org
|
Obovale
|
0
|
C
|
0
|
Rampant 1
|
0
|
+
|
n+2
|
M
|
M
|
L
|
L
|
Rose
|
|
A16
|
J.Org
|
Lancéolée
|
0
|
C
|
1
|
Rampant 1
|
0
|
+
|
n+2
|
P
|
pr
|
M
|
L
|
Rougeâtre
|
|
A17
|
J.Org
|
Obovale
|
+
|
C
|
0
|
Semi érigé
|
+
|
+
|
n+2
|
M
|
L
|
M
|
TD
|
Rose
|
|
A18
|
J.Org
|
Lancéolée
|
+
|
C
|
0
|
Erigé
|
++
|
+
|
n+2
|
M
|
M
|
Pr
|
L
|
Rouge foncée
|
|
A19
|
J.Org
|
Lancéolée
|
+
|
C
|
0
|
Erigé
|
++
|
+
|
n+2
|
M
|
L
|
Sr
|
TD
|
Rouge
|
|
A20
|
J.Org
|
Lancéolée
|
+
|
C
|
0
|
Erigé
|
++
|
+
|
n+2
|
TP
|
L
|
Sr
|
TD
|
Rouge
|
0, absence;++,
2 taches/foliole;+, présence ou tache/foliole ;
n+1, premier rameau dérivé de la tige
principale; n+2,
n+3, rameau dérivé du premier
et du deuxième respectivement; C, composé;
CFL, couleur de la fleur; Cg, couleur de la
graine; COG, compacité de la gousse;
D, dure; FFO, forme du foliole;
ICG, Icrisat Centre Groundunt; J.Org, jaune
orangé; L, léger; M,
modéré; Mi, mince; Mo, moyen;
P, prononcé, Pr, proéminent;
pr, profond; FPT, fleur présente sur
la tige principale; PGY, pigmentation du gynophore;
PGT, pigmentation de la tige; PPL, port de la
plante; pr, profond; Se, sans bec;
Sr, sans réticulation; Tr,
très; TBG, Type du bec de la gousse;
TEG, type d'étranglement de la gousse;
TRG, type de réticulation de la gousse;
TIF, type d'inflorescence.
3.1.2 En conditions contrôlées
L'accession A17 n'a pas fait l'objet d'observations ou de
mesures excepté son taux de germination à cause de son faible
nombre des plantes (3 plantes/12) durant l'expérimentation. Cette
expérimentation s'est effectuée dans les conditions de
température et humidité relative moyennes journalières
(figure 6). Quant aux figures 7, 8 et 9, elles montrent l'évolution de
ces deux paramètres pendant les premières phases de restriction
hydrique appliquées aux stades 50 % de floraison, de fructification et
de remplissage des gousses respectivement. Durant la période de ces
restrictions hydriques, les moyennes de température et d'humidité
relative sont: T=32,01° C; HR= 41,78 %; T=29,57° C; HR=29,53 % et
T=29,22° C; HR=28,52 % pour les stades 50 % de floraison, de
fructification et de remplissage des gousses respectivement. Les autres
restrictions hydriques intermittentes n'ont pas fait l'objet
d'établissement des figures montrant l'évolution de ces
paramètres.
Figure
6: Evolution des moyennes de température et
d'humidité relative durant l'expérimentation.
Echelle (100/100).
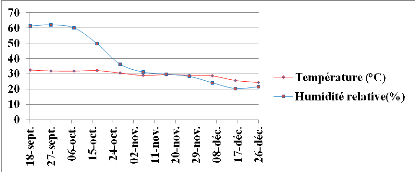
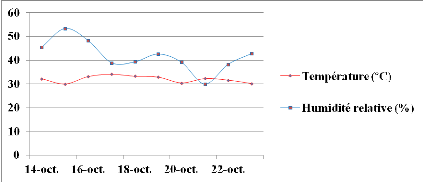
Figure 7:
Evolution des moyennes de température (T)
et d'humidité relative (HR) durant le
1èe stress hydrique appliqué au stade 50 % de
floraison. Echelle (100/111)
Figure 8:
Evolution des moyennes de température (T) et
d'humidité relative (HR) durant le
1èrstress hydrique appliqué au stade fructification.
Echelle (107/128).
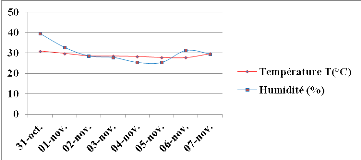
Figure
9: Evolution des moyennes de température (T) et
d'humidité relative (HR) durant le 1èe stress hydrique
appliqué au stade de remplissage de gousses. Echelle
(100/100).
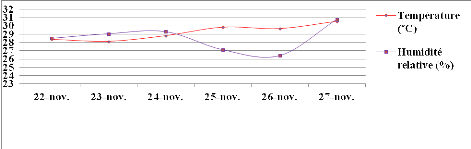
3.1.2.1 Classement des accessions selon le
degré de flétrissement
Les observations faites à 13 heures pendant les
périodes de stress hydriques ont montré les résultats
ci-après.
Pour le traitement T1, les accessions A19, A12,
A20, A18, A13 avaient un degré de flétrissement tel que leurs
feuilles apparaissaient cassantes au toucher; alors que les accessions A14, A2,
A3, A1, A6, A8, A7, A9, A10, A11 et A4 avaient des feuilles qui apparaissaient
sèches au toucher. Le flétrissement est peu perceptible chez les
accessions A16, A15 et A5.
Pour le traitement T2, les accessions A18 et A20
avaient un degré de flétrissement dont les feuilles
apparaissaient sèches au toucher pour la contrainte hydrique
appliquée au stade fructification; contrairement aux autres accessions
qui gardaient les mêmes scores lors de leur classement du traitement
T1.
Pour le traitement T3, deux sous groupes ont
été distingués: le sous groupe dont le
flétrissement est perceptible et celui dont le flétrissement est
peu perceptible: A7, A1, A15, A5, A4, A14, A2, A6.
3.1.2.2 Caractères quantitatifs
Le début de floraison (DF) des 20 accessions
d'arachide en conditions contrôlées varie entre 19 JAS (A13) et 25
JAS (A14, A15, A7) avec une moyenne de 23,42 #177; 1,89 JAS. La date de 50 % F
se fluctue entre 21,00 JAS (A19) et 28,667 JAS (A5) avec une moyenne de 25,03
#177; 2,84 JAS. Pour ce caractère, la différence entre les
accessions est très hautement significative (F=7,53***) avec des
coefficients de corrélation R=0,88 et de détermination R2
=78,1 %. Sept sous groupes ont été distingués selon
le test de ppds (Tableau X).
Dans cette condition d'expérimentation en pots, la
couleur de la fleur (CFL), la forme de la foliole (FFO), la présence de
la fleur sur la tige principale (FPT), la pigmentation de la tige (PGT) et du
gynophore (PGY) sont constantes. Le début floraison, la date de 50 % F
et la surface foliaire avaient des nouvelles valeurs (Tableau X).
3.1.2.2a La surface foliaire (SF)
Le stress hydrique a réduit la surface foliaire de
l'ensemble des accessions d'arachide. En effet, la surface foliaire des plantes
«témoin» varie de 21,31 cm2 (A15) à 43,63
cm2 (A9) avec une moyenne de 32,90 #177; 8,74 cm2 tandis
que celle des plantes «stressées» varie entre 17,17
cm2 (A4) et 27,04 cm2 (A1) avec une moyenne de 21,85
#177; 4,72 cm2. L'ANOVA montre une différence très
hautement significative (F=4,17***) entre les moyennes des plantes
«témoin». Les coefficients de corrélation et de
détermination sont respectivement R= 0,815 et R2= 66,4 %.
Contrairement aux plantes «stressées», l'ANOVA ne montre
aucune différence significative (F=1,28ns). Le test de Duncan
a permis de classer les plantes «témoins» en six sous groupes
et les plantes «stressées» en trois sous groupes. Les
accessions A16, A15, A2, A3 avaient maintenu relativement leur surface foliaire
durant la période de stress hydrique (Tableau X).
Tableau X: Regroupement des accessions en fonction
date de 50 % F et SF
|
Accessions
|
50 % F (jours)
|
SFt (cm2)
|
SFs (cm2)
|
|
A9
|
24,333 de
|
43,638 f
|
22,131 c
|
|
A6
|
22,333 bc
|
39,432 f
|
24,727 c
|
|
A11
|
21,667 b
|
43,515 f
|
19,388 c
|
|
A4
|
25,667 ef
|
24,057ab
|
17,174a
|
|
A19
|
21,000a
|
34,863 f
|
25,745 c
|
|
A13
|
21,667ab
|
36,396 f
|
23,795 c
|
|
A12
|
24,333 e
|
32,318 f
|
19,102 c
|
|
A3
|
27,000 g
|
29,942 de
|
23,228 c
|
|
A18
|
24,667 e
|
30,475 e
|
19,701 c
|
|
A20
|
22,667 c
|
35,582 f
|
21,845 c
|
|
A7
|
27,667 g
|
34,384 f
|
26,785 c
|
|
A14
|
28,333 g
|
25,057 bc
|
18,967 bc
|
|
A16
|
24,000 cd
|
27,466 cd
|
22,789 c
|
|
A10
|
26,000 fg
|
43,615 f
|
23,638 c
|
|
A5
|
28,667 g
|
32,221 ef
|
22,357 c
|
|
A8
|
24,000 d
|
28,189 d
|
19,011 c
|
|
A1
|
24,333 e
|
41,124 f
|
27,044 c
|
|
A15
|
28,667 g
|
21,318a
|
17,912ab
|
|
A2
|
28,667 g
|
21,318a
|
19,920 c
|
|
Moyenne
|
25,035
|
32,905
|
21,856
|
|
CV (%)
|
11,4
|
26,6
|
21,8
|
|
Ppds/ autres
|
2,024
|
2,449
|
2,449
|
|
Ecart type
|
2,847
|
8,741
|
4,772
|
50 % F, cinquante pour cent de
floraison; S, «stressé»; SF,
surface foliaire; t, «témoin». Les
moyennes suivies de mêmes lettres ne sont pas significativement
différentes à un risque alpha égale 5 %.
3.1.2.2b Biomasse sèche (BS)
L'analyse de variance montre une différence
significative entre la biomasse sèche des plantes
«témoins» (F=2,22*) avec des coefficients de
corrélation R=0,71 et de détermination R2=51,3 %. Le
stress hydrique a réduit significativement la biomasse des plantes
«stressées» aux stades de fructification (F=6,30***) et de
remplissage de gousses (3,29**); sans pourtant affecter de manière
significative les plantes «stressées» au stade 50 % F
(F=3,29ns). Cette réduction de la biomasse sèche est
plus importante chez les plantes «stressées» au stade 50 % F
(13,52 #177; 3,13 g, R=0,63, R2=40,9 %) qu'aux stades de
fructification (14,32 #177; 3,84 g, R=0,86, R2=74,9 %) et de
remplissage des gousses (20,05 #177; 3,90 g, R=0,78, R2=62,2 %).
3.1.2.2c Rendement et ses composantes
Dans cette étude, le stress hydrique a réduit
de manière significative le nombre de gousses par plante et le rendement
en graines par plante (RgP) au fur et à mesure que les apports d'eau
diminuent.
Au stade 50 % de floraison, le nombre de gousses par plante
(NGP) des plantes «stressées» est de 7,42 #177; 4,20 gousses
avec un coefficient de détermination R2=54,9 % et un
coefficient de corrélation R= 0,44 et celui des plantes
«témoins» est de 8,36 #177; 7,95.
Aux stades de fructification et de remplissage de gousses, le
nombre de gousses par plante pour les plantes «stressées» sont
respectivement: 8,26 #177;4,76 gousses avec R2= 54,9 % et R=0,74; et
14,22 #177; 5,69 gousses avec R2=54,1 % et R=0,73. La
différence de NGP est hautement significative pour les plantes
«stressées» aux stades 50 % de floraison (F=2,57**) et de
fructification (F=2,48**). Alors qu'elle est seulement significative au stade
de remplissage de gousses (F=1,70*). Aucune différence significative
n'est observée chez les plantes «témoin»
(F=1,20ns).
Les accessions A1, A20, A14, A19, A13, A5, A8, A15, A18,
A11, A4, A9, A16, A10, A3 et A6 avaient le plus grand nombre de gousses au
stade 50 % F. Quant au stade de fructification, certaines accessions (A14, A19,
A13, A15, A18, A11, A10, A6) avaient le nombre de gousses plus
élevé. Au stade de remplissage de gousses, les accessions A14,
A15, A9, A16, A10, A6 avaient le nombre de gousses plus élevé. En
outre, les accessions A1, A6, A16, A11, A5 et A19 avaient maintenu relativement
leur nombre de gousses aux différents stress appliqués aux stades
50 % de floraison, de fructification et de remplissage de gousses.
Le rendement en graines par plante varie suivant les stades
de développement de la plante à partir desquels la
sécheresse intervient. En effet les moyennes sont 8,37 #177; 3,88 g avec
R= 0,65, R2=42,4 % pour les plantes «témoin»; 1,77
#177; 1,43 g avec R= 0,66 et R2= 44,7 % pour les plantes
«stressées» au stade 50 % F; 2,43 #177; 1,96 g avec R=0,72,
R2= 52,6 % pour les plantes «stressées» au stade de
fructification et 4,48 #177; 3, 02 g, avec R=0,69, R2 =48 % pour les
plantes «stressées» au stade de remplissage des gousses. La
différence du RgP est significative chez ces plantes
«stressées»
. En effet, il existe une différence significative aux
stades de 50 % de floraison (F=1,70*), de fructification (F=2,34*) et de
remplissage de gousses (F=1,84*); tandis qu'aucune différence
significative n'est observée chez les plantes
«témoins». En effet, les accessions A1, A19 et A6 avaient le
meilleur rendement en graines parmi les plantes «stressées» au
stade 50 %. Les accessions A20, A19, A13, A15, A18, A11, A16, A10 et A6 avaient
le meilleur rendement en graines parmi les plantes «stressées»
au stade de fructification. Alors que les accessions A20, A12, ICG 9998, A13,
A18, A11, A16, A10, A3, A6 avaient le meilleur rendement en graines parmi les
plantes «stressées».
Le stress a réduit le poids moyen d'une graine au
fur et à mesure que la quantité d'eau apportée diminue. En
effet, la moyenne de poids moyen en graines de l'ensemble des plantes
«stressées» au stade 50 % F est de 1,11 #177; 0,23 g; celle
des plantes «stressées» au stade de fructification est de 1,25
#177; 0,29 g.
La moyenne de poids moyen en graines des plantes
«stressées» au stade de remplissage des gousses est de 1,34
#177; 0,26 g. La figure 10 montre que certaines accessions maintiennent
relativement leur PMg par exemple A6 encerclé (figure 10).
Figure 10:
Variation de poids moyen d'une graine des plantes
témoins (T0), stressées aux stades
50% de floraison (T1), de fructification
(T2) et de remplissage des gousses
(T3). Echelle (100/100).
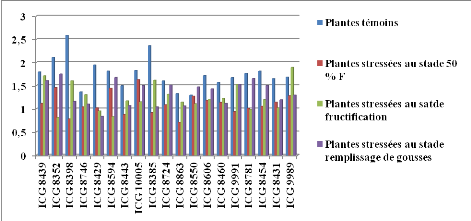
3.1.2.2d Indice de récolte
L'indice de récolte varie selon les conditions de
culture. Cependant, le stress hydrique a affecté de manière
significative les plantes «stressées». Au stade de
fructification, la différence est significative (F=2,21*) avec une
moyenne de 0,19 #177; 0,17 et des coefficients de corrélation R= 0,71 et
de détermination R2=51,2 %. Au stade de remplissage des
gousses la différence est significative (F=2,47*) avec une moyenne de
0,247 #177; 0,19 et des coefficients de corrélation R= 0,74 et de
détermination R2=55,3%. Aucune différence
significative n'est observée entre les plantes
«stressées» au stade 50 % de floraison (F=1,496ns).
Il existe une différence significative entre les plantes
«témoins» (F=2,445*). Les accessions A18, A11, A13, A10, A20,
A6, A19 et celles A18, A13, A10, A19 avaient le meilleur indice de
récolte en conditions stressantes aux stades de fructification et de
remplissage de gousses respectivement. De manière significative, l'effet
de stress hydrique est plus important au stade 50 % de floraison qu'aux stades
de fructification et de remplissage des gousses (Tableau XIII).
3.1.2.2e Indice de résistance à la
sécheresse (IRS)
Il ressort de cette étude que l'indice de
résistance de WILSON et al., (1978) varie suivant les stades de
développement de la plante auxquels intervient la contrainte hydrique.
En effet le stress hydrique a réduit de manière
significative (F=2,05*) l'indice de résistance à la
sécheresse des plantes au stade de fructification. Les coefficients de
détermination et de corrélation sont: R= 0,70, R2=
49,3 %.
Les valeurs moyennes d'IRS sont: 0,49 #177; 0,12; 0,55 #177;
0,14 et 0,72 #177; 0,15 des plantes «stressées» aux stades 50
% F, fructification et remplissage de gousses respectivement (Tableau XIII).
Hormis les accessions A11 et A13, toutes les accessions se sont montrées
performantes vis-à-vis d'un stress intervenu au stade de
fructification.
3.1.2.2f Indice de tolérance à la
sécheresse (ITS)
Pour l'indice de tolérance à la
sécheresse, l'analyse de variance montre une différence
significative uniquement entre les plantes «stressées» au
stade remplissage de gousses. L'indice varie énormément suivant
les accessions et les différents stades de développement auxquels
la sécheresse intervient (Tableau XI).
Tableau XI: Analyse de variance des caractères
quantitatifs
|
Caractères
|
50 % F
|
SF
|
BS
|
NGP
|
RgP
|
Ir
|
IRS
|
ITS
|
|
Plantes témoins
|
|
4,174***
|
2,227*
|
1,207ns
|
1,566ns
|
2,445*
|
0,453ns
|
0,600ns
|
|
Plantes stressées au stade 50 % F
|
7,536***
|
1,282ns
|
1,460ns
|
2,570**
|
1,703*
|
1,496ns
|
2,050*
|
0,600ns
|
|
Plantes stressées au stade fructification
|
|
|
6,301***
|
2,573**
|
2,708*
|
2,212*
|
1,214ns
|
1,221*
|
|
Plantes stressées au remplissage de
goussess
|
|
|
3,293**
|
2,486**
|
1,847*
|
2,471*
|
|
|
***, très hautement
significative; **, hautement significative;*,
significative; 50 % F, cinquante pour cent de floraison;
BS, biomasse sèche; IRS, indice de
résistance à la sécheresse; Ir, indice de
récolte; ITS, indice de tolérance à la
sécheresse; NGP, nombre de gousses par plante;
ns, non significative; RgP,
rendement en graines par plante.
|
Accessions
|
BS/T0
|
BS/T1
|
BS/T2
|
BS/T3
|
NGP/T0
|
NGP/T1
|
NGP/T2
|
NGP/T3
|
RgP/T0
|
RgP/T1
|
RgP/T2
|
RgP/T3
|
|
A1
|
24,700 bc
|
13,033 cd
|
16,167 g
|
16,833 c
|
9,667a
|
6,000 d
|
6,333 d
|
7,000a
|
6,685 cd
|
2,689 b
|
2,071 e
|
3,017 d
|
|
A7
|
36,933 e
|
17,767 d
|
17,600 g
|
26,100 g
|
12,000ab
|
1,667a
|
6,000 bc
|
8,333ab
|
3,421a
|
0,743a
|
2,652 ef
|
1,156ab
|
|
A20
|
21,300a
|
10,567a
|
13,633 f
|
16,700 bc
|
13,333 bc
|
5,333 cd
|
8,000 de
|
11,000 c
|
8,790 e
|
2,204a
|
2,983 f
|
5,150 e
|
|
A12
|
32,600 e
|
11,833ab
|
13,633 ef
|
15,833 b
|
13,667 c
|
5,000 bc
|
5,333 b
|
11,667 cd
|
6,350 c
|
1,525a
|
1,351 d
|
5,520 e
|
|
A14
|
29,533 e
|
13,533 d
|
18,833 g
|
22,267 g
|
14,333 c
|
5,333 d
|
12,667 f
|
19,000 f
|
8,883 e
|
1,157a
|
2,531 e
|
5,029 de
|
|
A19
|
23,000ab
|
13,200 d
|
12,567 de
|
15,800ab
|
15,000 c
|
8,667 d
|
10,000 f
|
13,333 de
|
13,819 e
|
2,651 b
|
5,270 f
|
9,400 e
|
|
A13
|
26,033 c
|
12,700 c
|
9,133 ab
|
14,367a
|
15,667 cd
|
8,333 d
|
9,333 f
|
15,333 ef
|
8,537 e
|
2,612ab
|
3,093 f
|
6,863 e
|
|
A5
|
29,267 de
|
15,967 d
|
18,367 g
|
20,433 ef
|
15,667 d
|
9,000 d
|
6,333 cd
|
8,667 b
|
5,946 ab
|
1,202a
|
0,950 cd
|
0,936a
|
|
A8
|
32,0 33 e
|
14,900 d
|
17,167 g
|
22,367 g
|
18,667 d
|
5,667 d
|
5,333 b
|
15,000 e
|
7,926 de
|
0,935a
|
0,332ab
|
2,273 bc
|
|
A15
|
31,400 e
|
13,500 d
|
15,533 fg
|
18,200 cd
|
19,000 d
|
11,000 d
|
9,333 f
|
20,000 f
|
6,200 bc
|
1,687a
|
2,954 f
|
4,297 d
|
|
A18
|
22,300a
|
10,267a
|
12,167 d
|
16,100 b
|
19,333 d
|
5,667 d
|
12,000 f
|
15,000 e
|
7,239 d
|
0,809a
|
3,429 f
|
5,520 e
|
|
A11
|
25,867 c
|
14,467 d
|
10,733 bc
|
19,333 e
|
19,333 d
|
10,000 d
|
10,000 f
|
13,667 e
|
8,942 e
|
2,063a
|
2,670 f
|
5,591 e
|
|
A4
|
33,267 e
|
17,033 d
|
18,333 g
|
27,100 g
|
20,333 d
|
6,000 d
|
6,000 c
|
13,333 e
|
6,271 c
|
0,689a
|
1,165 d
|
2,678 d
|
|
A9
|
28,000 cd
|
13,533 d
|
7,100a
|
23,500 g
|
21,000 d
|
9,667 d
|
1,667a
|
17,000 f
|
11,945 e
|
1,630a
|
0,662 bc
|
3,788 d
|
|
A16
|
28,000 d
|
14,900 d
|
17,067 g
|
22,267 fg
|
21,333 d
|
11,000 d
|
14,333 ef
|
16,667 f
|
11,536 e
|
2,506a
|
2,662 f
|
5,197 e
|
|
A10
|
26,267 c
|
10,967a
|
12,233 d
|
18,933 de
|
22,667 d
|
6,667 d
|
15,333 f
|
23,000 f
|
9,343 e
|
1,324a
|
4,704 f
|
6,593 e
|
|
A3
|
26,733 c
|
12,267 bc
|
13,700 f
|
25,700 g
|
24,000 d
|
6,333 d
|
6,333 d
|
12,000 d
|
8,064 e
|
1,186a
|
2,030 de
|
2,458 e
|
|
A2
|
35,433 e
|
14,333 d
|
17,700 g
|
24,600 g
|
26,000 d
|
4,000ab
|
3,667ab
|
11,000 c
|
6,973 d
|
1,474a
|
0,172a
|
2,821 d
|
|
A6
|
23,000 b
|
12,133 b
|
11,700 cd
|
18,333 d
|
28,000 d
|
15,667 d
|
14,333 f
|
19,333 f
|
12,305 e
|
4,711 b
|
4,619 f
|
5,636 e
|
|
Moyenne
|
28,202
|
13,521
|
14,328
|
20,053
|
18,368
|
7,421
|
8,544
|
14,228
|
8,378
|
1,779
|
2,437
|
4,489
|
|
Ecart type
|
3,848
|
3,133
|
3,848
|
3,900
|
7,950
|
4,209
|
4,760
|
5,695
|
3,886
|
1,430
|
1,969
|
3,021
|
|
CV (%)
|
26,8
|
23,2
|
26,8
|
23,2
|
43,3
|
56,7
|
57,6
|
40,0
|
46,4
|
80,4
|
80,8
|
67,3
|
|
Ppds
|
2,024
|
2,024
|
2,024
|
2,028
|
2,024
|
2,024
|
2,024
|
2,024
|
2,024
|
2,024
|
2,024
|
2,028
|
Tableau XII: Regroupement des accessions d'arachide en
fonction de leurs BS, RGP et RgP
CV, coefficient de variation;
BS, biomasse sèche; RGP, rendement en
gousses/plante; RgP, rendement en graines/plante;
T0 et T1, T2,
T3, plante «témoin» et
«stressées» aux stades 50 % F, de fructification et de
remplissage des gousses respectivement. Les moyennes suivies de même
lettres ne sont pas significativement différentes au seuil alpha=5
%.
Tableau XIII: Regroupement des accessions d'arachide
en fonction des indices Ir, IRS, ITR
|
Accessions
|
Ir0
|
Ir1
|
Ir2
|
Ir3
|
IRS1
|
IRS2
|
IRS3
|
ITR1
|
ITR2
|
ITR3
|
|
A7
|
0,093a
|
0,041a
|
0,161 e
|
0,043a
|
0,481a
|
0,481 e
|
0,712 d
|
0,232a
|
0,232a
|
0,374 d
|
|
A2
|
0,175ab
|
0,106a
|
0,010a
|
0,110 c
|
0,441a
|
0,544 e
|
0,744 d
|
0,426a
|
0,426a
|
0,306 bc
|
|
A4
|
0,189 bc
|
0,044a
|
0,064 b
|
0,095ab
|
0,513a
|
0,552 e
|
0,815 d
|
0,121a
|
0,121a
|
0,447 d
|
|
A15
|
0,196 cd
|
0,122a
|
0,196 ef
|
0,253 c
|
0,436a
|
0,510 e
|
0,601 bc
|
0,299a
|
0,299a
|
0,688 d
|
|
A5
|
0,206 d
|
0,086a
|
0,051ab
|
0,044a
|
0,549a
|
0,632 e
|
0,693 cd
|
0,230a
|
0,230a
|
0,177a
|
|
A12
|
0,211 d
|
0,151a
|
0,113 c
|
0,340 cd
|
0,366a
|
0,431 e
|
0,489a
|
0,353a
|
0,353a
|
0,813 d
|
|
A8
|
0,267 d
|
0,060a
|
0,019a
|
0,106 c
|
0,498a
|
0,557 e
|
0,710 d
|
0,165a
|
0,165a
|
0,292ab
|
|
A1
|
0,270 d
|
0,202ab
|
0,129 c
|
0,186 c
|
0,528a
|
0,655 e
|
0,682 c
|
0,416a
|
0,416a
|
0,494 d
|
|
A3
|
0,304 d
|
0,096a
|
0,161 e
|
0,096 bc
|
0,460a
|
0,512 e
|
1,036 d
|
0,275a
|
0,275a
|
0,449 d
|
|
A14
|
0,321 de
|
0,068a
|
0,140 cd
|
0,243 c
|
0,463a
|
0,651 e
|
0,766 d
|
0,194a
|
0,194a
|
0,554 d
|
|
A18
|
0,344 e
|
0,080a
|
0,286 f
|
0,362 d
|
0,483a
|
0,555 e
|
0,719 d
|
0,103a
|
0,103a
|
0,749 d
|
|
A11
|
0,349 e
|
0,146a
|
0,278 f
|
0,328 c
|
0,565a
|
0,418 bc
|
0,745 d
|
0,231a
|
0,231a
|
0,618 d
|
|
A13
|
0,370 ef
|
0,215 b
|
0,320 f
|
0,465 d
|
0,525a
|
0,377ab
|
0,594ab
|
0,385a
|
0,385a
|
0,728 d
|
|
A10
|
0,379 f
|
0,101a
|
0,386 f
|
0,363 d
|
0,428a
|
0,478 de
|
0,735 d
|
0,135a
|
0,135a
|
0,671 d
|
|
A20
|
0,406 f
|
0,239 b
|
0,290 f
|
0,314 c
|
0,505a
|
0,623 e
|
0,793 d
|
0,352a
|
0,352a
|
0,614 d
|
|
A16
|
0,420 f
|
0,167a
|
0,156 de
|
0,238 c
|
0,538a
|
0,615 e
|
0,797 d
|
0,230a
|
0,230a
|
0,477 d
|
|
A9
|
0,432 f
|
0,113a
|
0,087 bc
|
0,185 c
|
0,490a
|
0,256 e
|
0,846 d
|
0,140a
|
0,140a
|
0,359 cd
|
|
A6
|
0,549 f
|
0,416 b
|
0,395 f
|
0,318 c
|
0,524a
|
0,530 e
|
0,796 d
|
0,379a
|
0,379a
|
0,460 d
|
|
A19
|
0,612 f
|
0,421 b
|
0,064 f
|
0,597 d
|
0,567a
|
0,549 e
|
0,697 d
|
0,183a
|
0,183a
|
0,687 d
|
|
Moyenne
|
0,321
|
0,142
|
0,193
|
0,247
|
0,493
|
0,522
|
0,724
|
0,255
|
0,285
|
0,527
|
|
Ecart type
|
0,176
|
0,140
|
0,177
|
0,252
|
0,121
|
0,142
|
0,156
|
0,218
|
0,218
|
0,285
|
|
CV (%)
|
54,9
|
9,82
|
9,19
|
7,76
|
2,45
|
2,71
|
2,15
|
8,55
|
7,95
|
5,41
|
|
Ppds
|
2,024
|
2,024
|
2,024
|
2,028
|
2,024
|
2,024
|
2,028
|
2,024
|
2,028
|
2,028
|
50 % F, cinquante pour cent de
floraison; S, «stressé »;
SF, surface foliaire; 0 et
(1, 2, 3) sont des plantes
«témoin» et «stressées» aux stades 50 % de
floraison, de fructification et de remplissage de gousses respectivement;
CV, coefficient de variation; Ir, indice de
récolte; IRS, indice de résistance à la
sécheresse; ITS, indice de tolérance à la
sécheresse; ppds, plus petite différence
significative protégée. Les moyennes suivies de mêmes
lettres ne sont pas significativement différentes au risque alpha
égale 5 %.
3.1.2.3 Caractères Physiologiques
3.1.2.3aTeneur en Chlorophylle (T.Ch)
Dans cette étude, le stress hydrique a réduit
la teneur en chlorophylle des plantes «stressées» uniquement
au stade de fructification. Les coefficients de corrélation R=0,72 et de
détermination R2=52,5% avec une moyenne de 44,32 #177;
6,36mg/g de matière fraîche. Par contre ce stress hydrique n'a pas
réduit la teneur en chlorophylle des plantes
«stressées» aux stades 50 % F et de remplissage des gousses.
L'ANOVA montre une différence significative pour les plantes
«stressées» au stade 50 % de floraison (F=2,33*). Alors que la
différence n'est pas significative chez les plantes
«stressées» aux stades de fructification (F=1,41ns)
et de remplissage des gousses (F=1,57ns). Il en est de même
pour les plantes «témoins» aux différents stades: 50 %
de floraison (F=1,36ns), stade de fructification
(F=0,77ns) et stade de remplissage des gousses
(F=1,47ns).
Le test de plus petite différence significative
protégée (ppds) a permis de classer les accessions en plusieurs
sous groupes. En effet, pour les plantes «témoins» les
accessions A1, A12, A15 avaient une faible teneur en chlorophylle au stade 50 %
F; au stade de fructification, ce sont les accessions A13, A18, A19, A6 qui
avaient une faible teneur en chlorophylle. Quant au stade de remplissage des
gousses, ce sont les accessions A13, A8, A1, A12 avaient une faible teneur en
chlorophylle. Pour les plantes «stressées» aux stades 50 % de
floraison l'accession (A11), de fructification les accessions (A18, A9) et de
remplissage de gousses les accessions (A15, A13, A11) avaient de faible teneur
en chlorophylle. Le test de ppds regroupe les plantes
«stressées» en plusieurs sous groupes. Pour les plantes
«stressées» au stade 50 % F, cinq sous groupes ont
été distingués; pour les plantes
«stressées» au stade de fructification, deux sous groupes ont
été distingués et pour les plantes
«stressées» au stade remplissage de gousses quatre sous
groupes ont été identifiés (Tableau XIV).
Tableau XIV: Regroupement des accessions
d'arachide en fonction de la T.Ch
|
Accession
|
T.ChT0/TI
|
T.ChT1
|
T.ChT0/T2
|
T.Ch T2
|
T.ChT0/T3
|
T.Ch T3
|
|
A1
|
37,033a
|
35,067a
|
52,700 c
|
45,300 b
|
40,000 b
|
49,833 d
|
|
A12
|
37,967ab
|
46,100 ef
|
47,400 c
|
43,100 a
|
40,900 bc
|
51,567 d
|
|
A15
|
39,067 bc
|
44,233 de
|
52,533 c
|
46,767 b
|
41,367 c
|
42,400a
|
|
A13
|
39,900 c
|
45,333 e
|
49,600 c
|
42,800a
|
36,533a
|
43,333ab
|
|
A6
|
40,633 c
|
52,500 f
|
51,300 c
|
44,900 b
|
44,100 d
|
50,267 d
|
|
A3
|
41,000 c
|
47,700 f
|
44,300 bc
|
49,350 b
|
46,400 d
|
51,600 d
|
|
A10
|
41,133 c
|
42,967 d
|
50,200 c
|
48,600 b
|
44,367 d
|
53,033 d
|
|
A16
|
41,200 c
|
42,467 d
|
50,300 c
|
46,633 b
|
48,667 d
|
48,800 d
|
|
A4
|
41,500 cd
|
48,867 f
|
48,767 c
|
46,300 b
|
45,400 d
|
53,033 d
|
|
A19
|
41,500 d
|
41,200 d
|
41,167 b
|
43,667a
|
44,167 d
|
46,133 c
|
|
A20
|
41,633 d
|
40,767 cd
|
41,167 b
|
45,667 b
|
41,067 c
|
49,467 d
|
|
A8
|
41,967 d
|
43,333 d
|
49,100 c
|
46,533 b
|
38,900ab
|
48,767 d
|
|
A5
|
41,967 d
|
47,733 f
|
52,567 c
|
47,700 b
|
47,033 d
|
54,733 d
|
|
A18
|
42,533 d
|
41,467 d
|
40,800ab
|
43,567ab
|
41,233 c
|
47,500 cd
|
|
A2
|
42,600 d
|
39,833 bc
|
45,433 c
|
42,800a
|
44,633 d
|
51,533 d
|
|
A7
|
42,833 d
|
46,800 f
|
48,900 c
|
48,233 b
|
45,800 d
|
55,600 d
|
|
A14
|
42,933 d
|
54,367 f
|
58,950 c
|
49,067 b
|
46,667 d
|
55,400 d
|
|
A11
|
43,933 d
|
38,700ab
|
49,433 c
|
43,833 b
|
41,533 cd
|
45,933 bc
|
|
A9
|
46,467 d
|
42,700 3d
|
39,667a
|
46,167 b
|
50,033 d
|
48,833 d
|
|
Moyenne
|
41,463
|
44,323
|
48,436
|
45,852
|
43,621
|
49,820
|
|
Ecart type
|
3,291
|
6,370
|
8,918
|
3,303
|
5,355
|
5,624
|
|
CV(%)
|
7,9
|
14,5
|
18,4
|
7,2
|
12,3
|
11,3
|
|
ppds
|
2,024
|
2,024
|
2,024
|
2,024
|
2,024
|
2,028
|
CV, coefficient de variation;
ppds, plus petite différence significative
protégée; T.Ch, teneur en chlorophylle;
T.Ch T0/, teneur en chlorophylle des plantes
«témoin» aux différents stades ;
T0, plantes «témoins»;
T1, plantes «stressées» au stade
50 % F;T2, plantes «stressées» au
stade de fructification; T3, plantes
«stressées» au stade de remplissage de gousses. Les moyennes
suivies de mêmes lettres ne sont pas significativement différentes
au risque alpha=0,05 %.
3.1.2.3bTeneur relative en eau (TRE)
Le stress hydrique a réduit la teneur relative en eau
des feuilles pour les plantes «stressées» aux stades 50 % de
floraison, de fructification et de remplissage de gousses. Par ailleurs, la
teneur relative en eau des feuilles des plantes «stressées» au
stade 50 % de floraison varie entre 23,28 % (A9) et 63,34 % (A15) avec une
moyenne de 43,80 #177; 16,73 %. Celle des plantes «stressées»
au stade de fructification varie entre 14,70 % (A9) et 51,59 % (A2) avec une
moyenne de 35,94 #177; 15,41 %. Quant aux plantes «stressées»
au stade de remplissage des gousses, elle varie de 23,48 % (A16) à 65,64
% (A7) avec une moyenne de 34,05 #177; 15,33 %.
L'analyse de variance montre que la différence entre
les moyennes des accessions est hautement significative pour les plantes
«stressées» aux stades 50 % de floraison (F=2,58**) et
fructification (F=2,54**). Aucune différence significative n'est
enregistrée chez les plantes «stressées» au stade de
remplissage de gousses (F=1,01ns).
Pour la teneur relative en eau, l'ANOVA ne montre aucune
différence significative pour les plantes «témoins» aux
stades 50 % de floraison (F=1,15ns), de fructification
(F=1,08ns) et de remplissage de gousses (F=1,12ns). Le
coefficient de corrélation est très forte (R=0,74) pour les
plantes «stressées» aux stades 50 % F et de fructification (R=
0,73). Les coefficients de détermination sont R2 =55 % et
R2= 54,4 % aux stades 50 % de floraison et de fructification
respectivement. Ces coefficients montrent que 55 % et 54,4 % des variations
sont dues à la variabilité des accessions aux stades 50 % F et
fructification respectivement.
Le test de ppds distingue trois sous groupes au stade 50 % F,
sept sous groupes au stade de fructification et deux sous groupes au stade de
remplissage des gousses. Les accessions A1 et A15 avaient maintenu presque leur
teneur relative en eau lors d'un stress intervenu au stade 50 % de floraison.
Par contre, les accessions A15, A2 et A7 avaient maintenu leur teneur relative
en eau aux stades de fructification et de remplissage de gousses
respectivement. Par contre, les accessions A9, A6 avaient une faible teneur
relative en eau au stade 50 % F. Quant au stade de fructification,
c'étaient les accessions A9, A6 et A11 qui avaient une faible teneur
relative en eau.
3.1.2.3c Déficit hydrique (DH)
Le déficit hydrique (DH) varie suivant les
génotypes de la plante, mais aussi avec la période à
laquelle intervient la sécheresse. En effet, pour les plantes
«stressées» au stade 50 % F, le DH varie entre 30,7
(A15) et 64,7 % (A9) avec une moyenne de 46,2 #177; 13,8 %. Alors que celui des
plantes «stressées» au stade de fructification varie entre
41,4 (A2) et 73,5 % (A9) avec une moyenne de 55,6 #177; 13,2 %. Quant à
celui des plantes «stressées» au stade de remplissage, il
varie entre 27,49 et 62,29 avec une moyenne de 50,06 #177; 10,71 %. L'ANOVA
montre que la différence est significative pour les plantes
«stressées» au stade 50 % F (F=2,37*) avec des coefficients de
corrélation R=0,72 et de détermination R2=53 %. La
différence est hautement significative (F=2,65**) pour les plantes
«stressées» au stade de fructification avec des coefficients
de corrélation R=0,74 et de détermination R2=55,7 %.
En outre, aucune différence significative n'est enregistrée pour
les plantes «stressées» au stade de remplissage de gousses
(F=1,49ns). Le test de Duncan a permis de regrouper les accessions
en 6 sous groupes pour les plantes «stressées» aux stades 50 %
F et de fructification. Pour les plantes «stressées» au stade
de remplissage de gousses, deux sous groupes ont été
distingués.
Les accessions A9, A6, A4, A19, A13, A12, A20, A16, A1 et A2
avaient un déficit hydrique le plus important au stress intervenu au
stade 50 % F. Quant au stress intervenu au stade de fructification,
c'était les accessions A9, A6, A11, A4, A19, A13, A12, A3, A18 et A20
avaient le déficit hydrique le plus important au stress intervenu au
stade de fructification.
3.1.2.3d Teneur en eau par rapport à la masse
sèche (TE/MS)
La teneur en eau par rapport à la masse sèche
(TE/MS) varie selon les accessions et les conditions de culture. La TE/MS des
plantes «témoins» varie de 240,51 % (A5) à 370,10 %
(A13) avec une moyenne de 291,62 #177; 69,04 %. Alors que celle des plantes
«stressées» varie de 220,74 % (A15) à 325,72 % (8550)
avec une moyenne de 261,63 #177; 92,23 %.
L'ANOVA ne montre aucune différence significative chez
les plantes «stressées» (F=0,54ns) et
«témoin» (F= 0,34ns) (Tableau XV). Le ppds regroupe
les accessions en plusieurs sous groupes. La teneur en eau par rapport à
la masse des accessions A6, A11, A19, A12, A5, A1 est plus élevée
en conditions stressantes qu'en conditions normales (Tableau XVI).
Tableau XV: Analyse de
variance des caractères physiologiques
|
Caractères
|
T.Ch t
|
T.Ch s
|
TRE t
|
TRE s
|
DH
|
TE/MS t
|
TE/MS s
|
|
Stade 50 % F
|
1,361ns
|
2,334*
|
1,152ns
|
2,581**
|
2,379*
|
0,542ns
|
0,347ns
|
|
Stade fructification
|
0,772ns
|
1,416ns
|
1,084ns
|
2,542**
|
2,651**
|
|
|
|
Stade remplissage de gousses
|
1,472ns
|
1,577ns
|
1,127ns
|
1,014ns
|
1,495ns
|
|
|
**; hautement
significative;*, significative; DH,
déficit hydrique; s, «stressé»;
t, «témoin»; T.Ch, teneur en
chlorophylle; TE/MS, teneur en eau par rapport à la
masse sèche; ns, non significative;
TRE, teneur relative en eau des feuilles
.
Tableau XVI: Regroupement des accessions d'arachide en
fonction de TRE, TE/MS et DH aux différents traitements
|
Accessions
|
TRE/T0
|
TRE/T1
|
TRE/T2
|
TRE/ T0
|
TRE/T0
|
TRE/T3
|
TE/MSt
|
TE/MSs
|
DH1
|
DH2
|
DH3
|
|
A9
|
47,985ab
|
23,277a
|
14,701a
|
92,803 c
|
72,415a
|
32,440a
|
313,813b
|
277,507a
|
64,7 f
|
73,5 f
|
55,259 b
|
|
A6
|
73,305 c
|
26,597 bc
|
15,794ab
|
88,505 c
|
78,229 c
|
33,878a
|
296,320b
|
317,834a
|
62,6 f
|
72,2 f
|
49,552 b
|
|
A11
|
76,137 c
|
53,956 f
|
23,504 bc
|
87,460 c
|
76,040 c
|
32,088a
|
303,766b
|
325,722a
|
37,9 bc
|
66,4 f
|
48,928 b
|
|
A4
|
70,634 c
|
28,750 c
|
23,983 cd
|
88,783 c
|
79,429 bc
|
26,462a
|
268,123b
|
235,843a
|
59,8 f
|
66 f
|
54,344 b
|
|
A19
|
77,805 c
|
24,301a
|
26,922 de
|
87,339 c
|
75,785 b
|
30,930a
|
282,196b
|
318,232a
|
60,8 f
|
63,4 f
|
54,948 b
|
|
A13
|
80,230 c
|
31,131 cd
|
27,718 e
|
86,128 c
|
84,328 c
|
36,003a
|
370,109b
|
261,633a
|
56,4 f
|
62,9 f
|
45,936 b
|
|
A12
|
76,254 c
|
46,108 f
|
29,893 ef
|
89,291 c
|
85,556 c
|
34,522a
|
273,697b
|
321,387a
|
44,9 f
|
61,3 f
|
48,838 b
|
|
A3
|
68,821 c
|
48,714 f
|
33,225 fg
|
79,661a
|
81,293 c
|
28,214a
|
273,461b
|
224,282a
|
42,7 ef
|
57,4 f
|
56,209 b
|
|
A18
|
73,104 c
|
54,440 f
|
36,374 g
|
88,426 c
|
75,062 b
|
24,532a
|
311,486b
|
275,650a
|
40,2 de
|
55,7 f
|
62,296 b
|
|
A20
|
74,202 c
|
42,290 f
|
36,926 g
|
88,944 c
|
75,550 b
|
36,547a
|
290,321b
|
263,601a
|
48,7 f
|
55,2 ef
|
52,359 b
|
|
A7
|
75,362 c
|
54,095 f
|
39,270 g
|
88,839 c
|
84,744 c
|
65,649b
|
323,483b
|
267,376a
|
38,1 c
|
52,3 de
|
27,492a
|
|
A14
|
70,307 c
|
45,467 f
|
40,079 g
|
85,863 c
|
87,737c
|
27,998a
|
250,441ab
|
227,906a
|
41,1 e
|
52,6 e
|
47,865 b
|
|
A16
|
47,050a
|
38,224 de
|
42,832 g
|
88,684 c
|
75,041ab
|
23,485a
|
270,174b
|
269,656a
|
50,4 f
|
49,3 d
|
52,819 b
|
|
A10
|
75,181 c
|
53,050 f
|
44,650 g
|
90,813 c
|
82,001 c
|
35,065a
|
323,825b
|
247,169a
|
39,3 cd
|
47,9 d
|
50,714 b
|
|
A5
|
67,729 c
|
58,948 f
|
47,132 g
|
86,930 c
|
87,160c
|
37,401a
|
240,517a
|
247,028a
|
34,1ab
|
46,4 d
|
43,081ab
|
|
A8
|
80,361 c
|
38,442 e
|
47,659 g
|
85,171 bc
|
68,404a
|
29,122a
|
322,727b
|
303,239a
|
35,8 b
|
45,6 cd
|
56,040 b
|
|
A1
|
72,128 c
|
60,173 f
|
49,358 g
|
83,807 b
|
59,030a
|
33,863a
|
302,291b
|
306,182a
|
46,2 f
|
44,0 bc
|
50,718 b
|
|
A15
|
66,712 bc
|
63,339 f
|
51,275 g
|
83,242ab
|
81,528 c
|
40,007a
|
260,258b
|
220,748a
|
30,7a
|
42,0ab
|
45,982 b
|
|
A2
|
75,181 c
|
41,022 ef
|
51,597 g
|
85,643 c
|
82,161 c
|
36,892a
|
263,258b
|
239,410a
|
43,1 f
|
41,4a
|
51,882 b
|
|
Moyenne
|
70,995
|
43,807
|
35,942
|
87,175
|
78,398
|
34,052
|
291,621
|
271,074
|
46,200
|
55,600
|
50,061
|
|
CV (%)
|
21,2
|
38,2
|
42,9
|
5,1
|
14,9
|
45
|
23,7
|
34,0
|
30,0
|
23,7
|
21,4
|
|
ppds/autres
|
2,024
|
2,024
|
2,024
|
2,024
|
2,028
|
2,028
|
2,024
|
2,024
|
2,449
|
2,449
|
2,449
|
|
Ecart type
|
15,036
|
16,730
|
15,415
|
4,965
|
11,681
|
15,331
|
69,045
|
92,231
|
0,138
|
0,132
|
10,715
|
CV, coefficient de variation;
DH, déficit hydrique; ppds, plus
petite différence significative protégée;
s, plantes «stressées»; SF,
surface foliaire; T1,
T2, T3 stress hydriques
appliqués respectivement aux stades 50 % de floraison, de fructification
et de remplissage de gousses; t, plantes
«témoin»; TE/MS, teneur en eau par rapport la
masse sèche; TRE, Teneur relative en eau de la
troisième feuille de la tige principale des plantes. Les moyennes
suivies par les mêmes lettres ne sont pas significativement
différentes au risque alpha=0,05 %.
3.1.2.3e Relation entre les paramètres
étudiés
La matrice de corrélation établie avec les
plantes «stressées» aux différents stades 50 % F, de
fructification et de remplissage des gousses montre que la corrélation
entre les paramètres étudiés varie suivant les stades de
développement de la plante. En effet, la biomasse sèche (BS) est
positivement et significativement corrélée (r=0,70) à la
teneur relative en eau (TRE) au stade de fructification. Alors qu'aucune
corrélation significative n'est enregistrée aux stades 50 % de
floraison et de remplissage des gousses entre la biomasse sèche (BS) et
les autres paramètres. Ce paramètre (BS) est négativement
et significativement corrélé (r=-0,66) au déficit hydrique
(DH), à l'indice de récolte (r=-0,52) au stade de fructification.
Il est cependant positivement et significativement corrélé
(r=0,56) à la teneur en chlorophylle (T.Ch), à l'indice de
résistance à la sécheresse (r=0,60). Il est aussi
négativement et significativement corrélé (r=-0,51) au
rendement en graines par plante (RgP), à l'indice de récolte
(r=-0,79), à l'indice de tolérance à la sécheresse
(r=-69) au stade de remplissage de gousses. Enfin aucune corrélation
significative n'est enregistrée entre la biomasse sèche et les
autres paramètres au stade de 50 % de floraison.
L'indice de résistance à la sécheresse
(IRS) est négativement et significativement corrélé au DH
(r=-0,57) au stade de fructification. Quant à l'ITR, il est positivement
et significativement corrélé au rendement en graines par plante
(r=0,54), au poids moyen d'une graine (r=0,64) au stade 50 % F et à
l'indice de récolte (r= 0,60) au stade de remplissage de gousses
respectivement. Cet indice de tolérance est négativement et
significativement corrélé à la biomasse sèche
(r=-0,69) au stade de remplissage des gousses.
La teneur en eau par rapport à la masse sèche
(TE/MS) est positivement et significativement corrélée au poids
moyen d'une graine (r=0,52) et à l'indice de récolte (r=0,53)
(Tableau XVII).
Tableau XVII: Matrice de
corrélation en conditions contrôlées.
XVII-a-Plantes stressées au stade 50 %
floraison
|
T.Ch
|
TRE
|
DH
|
TE/MS
|
SF
|
BS
|
NGP
|
RgP
|
PMg
|
Ir
|
IRS
|
ITR
|
|
T.Ch
|
1,000
|
|
|
|
|
|
|
|
|
|
|
|
|
TRE
|
-0,218
|
1,000
|
|
|
|
|
|
|
|
|
|
|
|
DH
|
0,098
|
0,879**
|
1,000
|
|
|
|
|
|
|
|
|
|
|
TE/MS
|
-0,408
|
-0,253
|
0,273
|
1,000
|
|
|
|
|
|
|
|
|
|
SF
|
-0,174
|
-0,057
|
0,279
|
0,248
|
1,000
|
|
|
|
|
|
|
|
|
BS
|
0,181
|
-0,025
|
-0,090
|
-0,118
|
-0,043
|
1,000
|
|
|
|
|
|
|
|
NGP
|
0,125
|
-0,135
|
0,202
|
0,059
|
-0,039
|
-0,122
|
1,000
|
|
|
|
|
|
|
RgP
|
-0,038
|
-0,362
|
0,525*
|
0,488
|
0,460
|
-0,308
|
0,628*
|
1,000
|
|
|
|
|
|
PMg
|
-0,370
|
0,105
|
0,049
|
0,532
|
0,372
|
-0,042
|
-0,055
|
0,392
|
1,000
|
|
|
|
|
Ir
|
-0,083
|
-0,457
|
0,578*
|
0,537*
|
0,480
|
-0,376
|
0,483
|
0,888**
|
0,343
|
1,000
|
|
|
|
IRS
|
-0,189
|
-0,260
|
0,325
|
0,273
|
0,345
|
0,296
|
0,281
|
0,394
|
-0,095
|
0,401
|
1,000
|
|
|
ITR
|
-0,148
|
0,090
|
0,039
|
0,226
|
0,261
|
-0,177
|
0,096
|
0,543*
|
0,645*
|
0,393
|
-0,109
|
1,000
|
XVII-b Plantes stressées au stade
fructification
|
T.Ch
|
TRE
|
DH
|
BS
|
NGP
|
RgP
|
PMg
|
Ir
|
IRS
|
ITR
|
|
T.Ch
|
1,000
|
|
|
|
|
|
|
|
|
|
|
TRE
|
0,236
|
1,000
|
|
|
|
|
|
|
|
|
|
DH
|
-0,284
|
-0,441
|
1,000
|
|
|
|
|
|
|
|
|
BS
|
0,404
|
0,709*
|
-0,660*
|
1,000
|
|
|
|
|
|
|
|
NGP
|
0,085
|
-0,174
|
0,054
|
-0,139
|
1,000
|
|
|
|
|
|
|
RgP
|
-0,045
|
-0,346
|
0,221
|
-0,351
|
0,831**
|
1,000
|
|
|
|
|
|
PMg
|
0,330
|
0,130
|
0,198
|
0,012
|
0,050
|
0,373
|
1,000
|
|
|
|
|
Ir
|
-0,054
|
-0,388
|
0,268
|
-0,523*
|
0,752**
|
0,705*
|
-0,029
|
1,000
|
|
|
|
IRS
|
0,253
|
0,458
|
-0,572*
|
-0,52*3
|
0,238
|
0,068
|
0,047
|
0,166
|
1,000
|
|
|
ITR
|
-0,378
|
-0,179
|
-0,169
|
0,012
|
-0,141
|
-0,064
|
-0,335
|
0,108
|
0,104
|
1,000
|
XVII-c Plantes stressées au stade
remplissage des gousses
|
T.Ch
|
TRE
|
DH
|
BS
|
NGP
|
RgP
|
PMg
|
Ir
|
IRS
|
ITR
|
|
T.Ch
|
1,000
|
|
|
|
|
|
|
|
|
|
|
TRE
|
0,235
|
1,000
|
|
|
|
|
|
|
|
|
|
DH
|
-0,308
|
-0,891**
|
1,000
|
|
|
|
|
|
|
|
|
BS
|
0,560*
|
0,123
|
0,145
|
1,000
|
|
|
|
|
|
|
|
NGP
|
-0,251
|
-0,334
|
0,250
|
-0,111
|
1,000
|
|
|
|
|
|
|
RgP
|
-0,512*
|
-0,330
|
0,250
|
-0,682*
|
0,483
|
1,000
|
|
|
|
|
|
PMg
|
0,037
|
0,216
|
0,232
|
0,021
|
-0,214
|
0,142
|
1,000
|
|
|
|
|
Ir
|
-0,575*
|
-0,250
|
0,326
|
-0,794**
|
0,376
|
0,977***
|
0,095
|
1,000
|
|
|
|
IRS
|
0,279
|
-0,282
|
0,326
|
0,609*
|
0,047
|
0,250
|
0,185
|
0,356
|
1,000
|
|
|
ITR
|
-0,489
|
-0,155
|
0,177
|
-0,699*
|
0,328
|
0,775**
|
0,137
|
0,808**
|
-0,434
|
1,000
|
*, corrélation
faible;** corrélation forte, ***,
corrélation très forte; (-), corrélation
négative; Valeur en gras, montre le degré de
significativité avec coefficient de corrélation de Pearson (0,05
%); BS, biomasse sèche; DH,
déficit hydrique; Ir, indice de récolte;
IRS, indice de résistance à la
sècheresse; ITR, indice de tolérance à la
sécheresse; NGP, nombre de gousses par plante;
PMg, poids moyen d'une graine, T.Ch, teneur
en chlorophylle; TE/MS, teneur en eau par rapport à la
masse sèche.
A l'issue de cette analyse de corrélation, les
caractères rendement en graines par plante (RgP), poids moyen d'une
graine (PMg) et l'indice de récolte (Ir) sont corrélés
à l'indice de tolérance à la sécheresse (ITS). Le
dendrogramme de la figure 11 (établi sur la base de ces
caractères) regroupe les accessions en trois sous groupes suivant leur
tolérance ou sensibilité à la sécheresse lorsque le
niveau de troncature n=0,95 (flèche). Ce dendrogramme confirme aussi les
performances des accessions A6, A1 et A2 vis-à-vis de la
sécheresse. Ces accessions sont considérées comme des
accessions tolérantes à la sécheresse dont A6 est la
meilleure accession tolérante à la sécheresse (figure
12).
Figure 11:
Dendrogramme d'identification des accessions
tolérantes à la sécheresse sur la base des
caractères corrélés à l'indice de tolérance
à la sécheresse. Echelle (157/194).
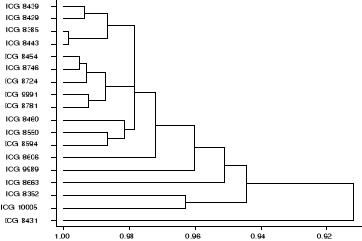

Figure 12:
Photo de Graines d'A6 (accessions tolérante) aux
différents traitements(T1, stress appliqué au stade de 50 % F;T2,
stress appliqué au stade de fructification; T3,stress appliqué au
stade de remplissage de gousses
3.1.3 Quelques caractères influencés par
l'environnement
Dans cette étude, la germination, les stades de
début de floraison, de cinquante pour cent de floraison et la surface
foliaire varient suivant les accessions et les conditions de culture. La
température journalière durant la période
«semis-cinquante pour cent de floraison» varie de 22,75° C
à 32° C (28,39 #177; 2,25°C) et de 29,55° C à
30,1° C (30,42 #177; 0,99°C) en conditions naturelles et
contrôlées respectivement. Quant à la radiation solaire,
elle varie de 5,1 à 18,6 (21,22 #177; 4,29 MJ/m2) en
conditions naturelles et de 23,5 à 24,5 (21, 49 #177; 2,95
MJ/m2) en conditions contrôlées (figure 13). Cela veut
dire que ces caractères (germination, début de floraison et 50 %
F) sont très influencés par les facteurs environnementaux
(facteurs climatiques et édaphiques). En effet, 50 % F est plus
précoce en conditions contrôlées qu'en conditions
naturelles. Quant à la surface foliaire de la troisième feuille
de la tige principale des accessions d'arachide, elle est plus large en
conditions contrôlés qu'en conditions naturelles (Tableau
XVIII).
Nombre de jours après semis (JAS). Echelle
(114/103)
Figure 13:
Evolution des températures minimale (T
Min) et maximale (T Max) en conditions naturelles (N)
et contrôlées (C) du semis au stade 50% de
floraison.
.
Température (°C)
Radiation solaire (MJ/m2)
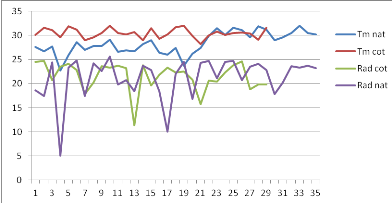
Tableau XVIII: Comparaison de DF et 50 % F dans les
deux conditions de culture
|
Accessions
|
DF(Jours)
|
50 % F(Jours)
|
SF (cm2)
|
|
Conditions
Naturelles
|
Conditions
Contrôlées
|
Conditions
Naturelles
|
conditions
contrôlées
|
Conditions naturelles
|
Conditions contrôlées
|
|
A13
|
19
|
21
|
22,666
|
21,667a
|
27,692
|
36,396
|
|
A19
|
20
|
22
|
23,000
|
21,000a
|
21,746
|
34,863
|
|
A20
|
20
|
23
|
23,000
|
22,667 c
|
24,511
|
35,582
|
|
A6
|
23
|
24
|
24,000
|
22,333bc
|
30,181
|
39,432
|
|
A12
|
21
|
23
|
25,000
|
24,333 d
|
24,487
|
32,318
|
|
A18
|
22
|
23
|
25,000
|
24,667 d
|
19,037
|
30,475
|
|
A9
|
23
|
24
|
26,000
|
24,333 d
|
26,850
|
43,638
|
|
A16
|
24
|
23
|
26,000
|
24,000 cd
|
17,867
|
27,466
|
|
A11
|
22
|
23
|
26,000
|
21,667 b
|
22,808
|
43,515
|
|
A10
|
23
|
27
|
26,000
|
26,000 d
|
29,181
|
43,615
|
|
A8
|
22
|
23
|
26,000
|
24,000 d
|
20,984
|
28,189
|
|
A1
|
24
|
24
|
27,000
|
24,333 d
|
26,080
|
41,124
|
|
A7
|
24
|
25
|
27,000
|
27,667 d
|
14,303
|
34,384
|
|
A4
|
24
|
26
|
27,000
|
25,667 d
|
18,349
|
24,057
|
|
A3
|
24
|
26
|
28,000
|
27,000 d
|
20,086
|
29,942
|
|
A171
|
24
|
x
|
29,000
|
x
|
34,731
|
x
|
|
A2
|
23
|
27
|
29,000
|
28,667 d
|
17,938
|
21,318
|
|
A14
|
23
|
27
|
29,333
|
28,333 d
|
13,951
|
25,057
|
|
A5
|
25
|
26
|
30,000
|
28,667 d
|
20,948
|
32,221
|
|
A15
|
20
|
27
|
35,000
|
28,667 d
|
14,119
|
21,318
|
|
Moyenne
|
22,5
|
23,42
|
26,700
|
25,035
|
22,302
|
32,89
|
|
CV( %)
|
7,56
|
8,09
|
3,7
|
11,4
|
35,9
|
26,6
|
|
Ecart type
|
1,701
|
1,894
|
3,233
|
2,847
|
8,0110
|
7,27
|
1accession non
évaluée; 50 % F, Cinquante pour cent de
floraison; CV, coefficient de variation; DF,
début de floraison; x, non évalué. Les
moyennes subies de mêmes lettres ne sont pas significativement
différentes au seuil alpha=0,05 %.
III.2 Discussion
3.2.1 Conditions
naturelles (Champ)
L'analyse de variance montre que
la différence entre les moyennes des accessions est significative pour
le taux de germination, la date de 50 % F, la surface foliaire, la hauteur de
la tige principale et la durée du cycle. Alors qu'aucune
différence significative n'est observée pour le nombre de gousses
par plante, le rendement en gousses par plante, rendement en graines par plante
et la teneur en huile. Quatre caractères quantitatifs montrent une
hétérogénéité au sein des 20 accessions
évaluées. Le taux de germination (CV=44,20 %), la surface
foliaire (CV=35,90 %), la hauteur de la tige principale (CV=44,70 %) et le
nombre de gousses par plante (CV=36,65 %) montrent que les 20 accessions sont
hétérogènes. Alors que la date de 50 % de floraison
(CV=3,700 %), la durée du cycle (CV=7,40 %), le rendement en gousses par
plante (CV=10,05 %), le rendement en graines par plante (CV=14,87 %) et la
teneur en huile (27,50 %) montrent que ces 20 accessions sont homogènes.
L'homogénéité de l'échantillon peut être due
à l'influence des conditions environnementales (facteurs climatiques et
édaphiques) sur ces cinq caractères.
L'hétérogénéité de l'échantillon pour
les quatre caractères montre que ces caractères seraient moins
influencés par les conditions environnementales. La variabilité
existante au sein de cet échantillon pour ces quatre caractères
peut être expliquée à 74,2 % pour le taux de germination
(TAG), 48,8 % pour la surface foliaire (SF), 55,8 % pour la hauteur de la tige
principale (HPL) et 36,4 % pour le nombre de gousses par plante (NGP).
L'analyse factorielle discriminant (AFD) montre que parmi les 15
caractères fixés, 14 permettent de discriminer les types
taxonomiques des accessions.
En effet, le taux de germination (TAG) varie entre 22, 66 %
et 100 % selon les accessions. Ce taux de germination est l'un des
critères de la certification des semences de l'arachide. Il renvoie
à la capacité d'une semence de réaliser toutes les
fonctions essentielles pour la croissance normale d'une plante dans des
conditions idéales (température, humidité, aération
convenable à sa germination). Le tableau IV, montre que le taux
de germination devient de plus en plus faible au fur et à mesure que la
durée de conservation devient importante, excepté les accessions
conservées pendant 4 ans et 5 ans. Le faible taux de germination
pourrait être dû soit à la longue durée de
conservation, soit à la dormance favorisant la pourriture des graines.
Selon SINGH et al., (1991), les graines de nombreuses espèces
d'Acacia utilisées en Inde ne germent pas lorsque le potentiel
hydrique atteint -9 ou -12 bars. Selon NEFFATI et al., (1997) et WAHBI
et al., (2010), les légumineuses des zones arides sont capables
de germer sous une large gamme de températures (15 ° C et 40°
C). Ceci montre que la température n'est pas le facteur affectant du
taux de germination des accessions conservées pendant 4 ans. Puisque, la
température durant l'expérimentation est comprise entre
27,3° C et 30,7° C. Il en est de même pour l'eau car selon
NDOUR et al., (1998), l'eau n'est pas un facteur limitant de la
germination chez les légumineuses de zones arides.
Au regard de tout ce qui précède, ce faible taux
de germination des accessions conservées pendant 4 ans pourrait
être dû aux facteurs internes de la graine, le mauvais
conditionnement des semences ou aux intempéries de coupures
d'électricité affectant le pouvoir germinatif de ces semences. Ce
taux de germination (TAG) est positivement et significativement
corrélé (r=0,961) au taux de levée. Ceci montre que plus
le taux de germination est important, plus le taux de levée est
élevé.
Le taux de levée des accessions varie entre 26,66 %
(A15) et 100 % (A20). Pour les types taxonomiques, ce taux de levée des
VUL (80 %), PRU (70,83 %) et FST (64,16 %) est plus élevé que
celui des types HYB (38,33 %) et HUR (50,83 %). Ce faible taux de levée
des HYR et HYB pourrait être expliqué par la dormance de leurs
graines, dormance qui occasionne la pourriture de la graine. Ce taux de
levée est négativement et significativement corrélé
(r=-0,70) à la durée du cycle (DC). Ceci montre que, plus les
graines sont dormantes plus la durée du cycle est prolongée. Il
est aussi négativement et significativement corrélé
à la date de 50 % F (r=- 0,71). Ceci montre que plus le taux de
levée est énorme, plus la date de 50 % de floraison est
prolongée. La date de 50 % F varie selon les taxons; elle varie entre
23 JAS et 35 JAS pour les accessions. La date de 50 % F des types taxonomiques
HYB (29 JAS) et HYR (30 JAS) est plus tardive que celles des types FST (26
JAS), VUL (25 JAS) et PRU (25 JAS). Quant au début de floraison, il
varie entre 19 JAS (A13) et 25 JAS (A5). Les types taxonomiques HYR (23 JAS) et
HYB (24 JAS) fleurissent tardivement que les types taxonomiques PRU (21 JAS),
FST (22 JAS) et VUL (23 JAS). Les résultats de début de floraison
et la date de 50 % F sont conformes à ceux de la
«classification botanique» de SINGH et al., (1998)
et HERSELMAN (2003). Selon cette classification, les accessions de la sous
espèce hypogaea sont des accessions tardives. Chez l'arachide,
la date de floraison pourrait être un paramètre de grande
importance dans la détermination du rendement comme cela a
été démontré chez Vigna subterrenea
par KUMAGA et al., (2003), MAKANDA et al.,
(2009) et ONWUBIKO et al., (2011). Une sélection se basant sur
ce caractère permet de regrouper les accessions. En effet, les
accessions A13, A19, A20, A6 et A12 peuvent être
considérées comme des accessions précoces (cycle court);
les accessions A1, A7, A4, A3, A17, A2, A14, A15 et A5 peuvent être
considérées comme des accessions tardives (cycle long). Alors que
les autres accessions peuvent être considérées des
accessions intermédiaires (cycle semi-précoce).
Le type d'inflorescence (fleur composée) et l'absence
des fleurs sur la tige principale des accessions A1 et A6 ne corroborent pas
ceux de la «classification botanique» de SINGH et
al., (1998) et HERSELMAN (2003); selon laquelle la sous espèce
fastigiata porte des fleurs sur leur tige principale. Pour les types
taxonomiques, la durée de cycle est plus précoce chez les types
VUL (83,33 JAS) et FST (88,33 JAS) que chez les types PRU (92,50 JAS), HYB
(94,58 JAS) et HYR (95,50 JAS). Ceci montre la précocité de la
sous espèce Arachis hypogaea fastigita par rapport à la
sous espèce Arachis hypogaea hypogaea.
Cette durée du cycle est significative et positivement
corrélée (r=0,61) au rendement en graines par plante. Ceci montre
que plus la durée du cycle est prolongée, plus le nombre de
gousses augmente et plus le rendement en graines augmente. Cette durée
de cycle est négativement et significative corrélée
(r=-0,68) au taux de germination et à la surface foliaire (r= -0,57).
La surface foliaire des accessions varie entre 13,95
cm2 (A14) et 34,73cm2 (A17). Cette surface foliaire des
types taxonomiques HYB (17,58 cm2) et HYR (16,80 cm2) est
plus étroite que celle des FST (26,06 cm2), PRU (24,20
cm2) et VUL (26,84 cm2). Cette étroite surface
foliaire observée chez les types HYR et HYB pourrait être une
stratégie d'augmentation de leur nombre des feuilles afin de recouvrir
toute la surface du sol, donc une stratégie d'avoir un LAI.
L'indice de surface foliaire (LAI) varie entre 0,59 (A15) et
4,33 (A7). Ceci montre que le caractère rampant ne conditionne pas le
recouvrement du sol par la plante. Les types taxonomiques PRU (1,25), FST
(1,63) et VUL (1,67) ont un LAI plus grand que les types taxonomiques HYB
(1,88) et HYR (2,26). Cette importante valeur de LAI chez les HYB et HYR
confirme le caractère rampant de la sous espèce Arachis
hypogaea hypogaea.
La hauteur de la tige des plantes varie entre 7 cm (A15) et
24,16 cm (A13). Elle est inferieure à celle de RABECHAULT (1960). Elle
varie selon les types taxonomiques. Elle est plus petite chez les types
taxonomiques HYB (8,95) et HYR (9,26 cm) que chez les types VUL (13,73 cm), PRU
(16,69 cm) et FST (19,05 cm). Ceci confirme aussi le caractère rampant
de la sous espèce Arachis hypogaea hypogaea et le
caractère érigé et semi érigé de la sous
espèce Arachis hypogaea fastigiata. Elle est positivement et
significativement corrélée à la surface foliaire (r=0,66).
Le nombre de gousses par plante (NGP) varie entre 19,86
(A19) et 71,33 (A7). Il est très élevé chez les types
taxonomiques HYB (38,90), HYR (46,033) que chez les types taxonomiques FST
(32,88) et PRU (24,51 excepté VUL (40,95). Ce nombre élevé
de gousses par plante chez les HYR et HYB confirme leur caractère
rampant; puisque, plus le nombre de ramifications (envergure) augmente, plus le
nombre de gynophores augmente et plus le nombre de gousses augmente. Quant au
nombre élevé de gousses chez le type taxonomique VUL, il pourrait
être dû à la fertilité des gynophores tardives.
Les types taxonomiques FST (3,5 graines), PRU (3,7 graines)
contiennent plus des graines dans leurs gousses que les types taxonomiques VUL
(2,0 graines), HYB (2,2 graines) et HYR (2,5 graines). Ces résultats
sont conformes à ceux de la «classification
botanique» selon laquelle, les accessions de la sous espèce
hypogaea possèdent 2 graines dans leurs gousses. Mais ces
résultats ne sont pas conformes à ceux de la même
«classification botanique» de l'arachide cultivée,
selon la quelle, le type taxonomique HYB possède 3 graines dans leur
gousse. Ce Nmg/G est positivement et significativement corrélée
(r=0,67) à la hauteur de la plante (HPL). Il existe des
corrélations positives entre: NGP et RGP (r=0,90); NGP et RgP (r=0,85);
RGP et RgP (r=0,93).
Le rendement en gousses par plante (RGP) varie entre 18,42
(A13) et 57,74 (A7). Il est plus grand chez les types taxonomiques HYR (29,18),
HYB (21,82) et VUL (29,64) que chez le types FST (16,94) et PRU (11,94). Ceci
montre que le rendement en gousses n'est pas lié au caractère
rampant.
La teneur en huile (TEH) entre les accessions varie entre
36, 30 % (A8) et 52, 8 % (A20). Elle est plus importante chez le type
taxonomique FST (48,52 %) que chez les types taxonomiques VUL (41,37 %), PRU
(43,85 %), HYB (45,25 %) et HYR (45,60 %). Ces résultats sont en accord
avec ceux de CELIA. et al., (2005); selon lesquels, les accessions de
type taxonomique FST sont plus riches en huile que les autres types
taxonomiques. En particulier, les accessions A11 (49,20 %), A18 (49,30 %), A10
(50,90 %), A4 (51,0 %), A13 (51,30 %), A7 (51,60 %), A2 (52,80 %), A20 (52,80
%) sont les meilleures accessions de bonne teneur en huile. Elles peuvent
être considérées comme des accessions d'industrie (bonne
teneur en huile). Alors que, les autres accessions seront
considérées comme des accessions de «bouche». Cette
importante teneur en huile du type taxonomique FST montre que, tout programme
de création des variétés d'arachide de bonne teneur en
huile doit tenir compte de la sous espèce fastigiata comme
témoin. En plus de la teneur en huile, la couleur des huiles
diffère suivant les accessions. C'est ainsi que l'accession A17 avait
une couleur en huile totalement différente de la couleur habituelle
d'arachide. La matrice de corrélation montre une corrélation
significative entre la teneur en huile et le poids moyen d'une graine. Cette
corrélation est très faible. Elle pourrait être forte
lorsque toutes les accessions appartenant au type taxonomique fastigiata
fastigiata Valencia seront évaluées.
3.2.2 Conditions
contrôlées (pots)
L'identification des variétés productives en
zones aride et semi-aride caractérisées par des épisodes
de déficit hydrique et de hautes températures, qui peuvent
apparaître de façon progressive, brutale, au début, au
milieu ou en fin de saison s'avère nécessaire dans le monde en
particulier en Afrique Subsaharienne. Ces variétés productives
possèdent de mécanismes d'adaptation leur permettant de produire
dans des conditions stressantes. Selon HAYEK et al., (2000), la
résistance à la sécheresse est liée à la
capacité d'une variété de développer un nombre
élevé de mécanismes d'adaptation et non pas à la
présence d'un seul mécanisme. Ces variétés
résistantes développent une stratégie d'adaptation
regroupant en même temps un ensemble des mécanismes d'adaptation
selon REJEB et al., (1993). Ce terme résistance diffère
de la tolérance par le fait que la résistance à la
sécheresse est associée à plusieurs
caractéristiques d'ordre phénologique, morphologique,
physiologique, biochimique reflétant différents types
d'adaptation (Esquive, Evitement et Tolérance) selon TUNER, en 1979.
La teneur en chlorophylle, le déficit hydrique, la
teneur relative en eau et la teneur en eau par rapport à la masse
sèche sont des paramètres d'identification des
variétés résistantes à la sécheresse. En
effet, la teneur en chlorophylle (T.Ch), la teneur relative en eau (TRE), le
déficit hydrique (DH) et la teneur en eau par rapport à la masse
sèche (TE/MS) des accessions d'arachide étudiées varient
suivant les stades de développement de la plante et les conditions de
culture. Pour la teneur en chlorophylle, l'analyse de variance montre une
différence significative au stade 50 % de floraison (F=2,33*). Alors que
pour la teneur relative en eau des feuilles, l'analyse de variance montre une
différence hautement significative au stade 50 % de floraison (F=2,58**)
et au stade de fructification (F=2,54**). Quant au déficit hydrique,
l'analyse montre une différence significative au stade 50 % de floraison
(F=2,37*) et une différence hautement significative au stade
fructification (F=2,65**). Aucune différence significative n'est
observée pour la teneur en eau par rapport à la masse
sèche.
Au stade de fructification, la teneur en chlorophylle (48,43
mg/g Mf) des plantes «témoins» est supérieure à
celles des plantes «témoins» aux stades 50 % de floraison
(41,46 mg/g Mf) et de remplissage des gousses (43,62 mg/g Mf). Ceci pourrait
montrer que l'arachide a besoin de l'azote aux stades 50 % de floraison et de
remplissage des gousses en conditions normales (non stressantes). La forte
teneur en T.Ch au stade de fructification chez les plantes
«témoins» montre que cette phase serait la phase la plus
sensible à la sécheresse. Puisque, plus la teneur en chlorophylle
est importante, plus l'absorption de l'énergie lumineuse est importante
et plus la demande en eau pour les réactions biochimiques
(photosynthèse, respiration et autres réactions biochimiques) est
importante. Par contre celles des plantes «stressées» varient
des 44,32 mg/g Mf (stade 50 % F), 45,85 mg/g Mf (stade fructification) et 49,82
mg/g Mf (stade remplissage des gousses). La faible teneur en chlorophylle
pourrait être expliquée par la diminution de la durée
d'ensoleillement, la sénescence foliaire, la diminution de sa
synthèse ou encore la diminution de l'absorption de l'énergie
lumineuse nécessaire à cette photosynthèse (BLUM
al., 2005).
La différence non significative des accessions
d'arachide aux stades de fructification et de remplissage des gousses, pourrait
être due au fait que l'arachide est tolérante à la
sécheresse, comme cela a été montré par NOMBRE,
(2012) chez le voandzou. La différence significative chez les plantes
«stressées» au stade 50 % de floraison serait due à la
haute température (32,01° C) et à la faible humidité
relative (41,78 %). Par contre, aux stades de fructification et de remplissage
des gousses les valeurs de ces paramètres sont T=29,57° C, HR=29,53
% et T=29,22° C, HR=28,52 % respectivement. Ces résultats
corroborent ceux de WOOD et al., (1968) et BOLHUIS et al.,
(1959). Le stress hydrique a réduit la teneur en chlorophylle uniquement
chez les plantes «stressées» au stade de fructification (94,66
%). Aucune réduction de T.Ch n'est observée chez les plantes
«stressées» aux stades 50 % de floraison et de remplissage des
gousses.
Au contraire, les T.Ch des plantes
«stressées» aux stades 50 % de floraison et de remplissage des
gousses ont été augmentées. Ces résultats
corroborent ceux de ATTI et al., (2004) chez le soja et NOMBRE, (2012)
chez le voandzou. La diminution de la teneur en chlorophylle pourrait
être due à la synthèse et à l'accumulation de la
proline (synthétisée à partir d'un même
précurseur que les pigments chlorophylliens: acide glutamique). En
effet, plus la concentration en proline augmente plus celle des pigments
chlorophylliens diminue. Pour le stress appliqué au stade 50 % de
floraison, les accessions A9, A11, A18, A20 et A1 avaient une teneur en
chlorophylle plus faible que les autres accessions. Alors que chez les plantes
«stressées» au stade de fructification c'étaient les
accessions A3, A19, A20, A13 et A9 qui avaient une teneur en chlorophylle
élevée. Quant au stress appliqué au stade de remplissage
des gousses, les accessions A16 et A9 présentaient une teneur
relativement plus faible. En se basant sur la fréquence d'apparition des
accessions d'arachide présentaient de faible teneur en chlorophylle aux
différents traitements, l'accession A9 devrait être la meilleure
accession tolérante à la sécheresse.
Le déficit hydrique (DH) est plus important au stade
de fructification (55,60 %) qu'aux stades 50 % de floraison (46,200 %) et de
remplissage des gousses (50,061%). Par ailleurs, la comparaison des plantes
«stressées» aux différents stades (50 % de floraison,
de fructification et de remplissage des gousses) montre que le DH d'une
même accession est très variable d'un stade à un autre.
Cette différence s'expliquerait non seulement par la croissance des
feuilles mais aussi par la variation des conditions climatiques. C'est ainsi
qu'au: (i) stade 50 % de floraison, la température moyenne et
l'humidité relative moyennes sont 32,01° C et 41,78 %
respectivement, (ii) stade de fructification, les températures et
l'humidité relative moyennes sont: 28,86° C et 30,03 %, (iii) stade
remplissage des gousses, elles sont d'ordre suivant: 29,22° C et 28,52 %.
Ces résultats confirment que, le stade de fructification est la phase la
plus sensible à la sécheresse. Ces résultats corroborent
ceux de ROY-MACAULEY, (1999) chez le raisin et ceux de HAMIDOU, (2006) chez le
niébé et ceux de AMANI et al., (2007) chez l'arachide.
Cette importante valeur de DH pendant la phase de fructification pourrait
être expliquée par l'utilisation de l'eau à la fois par la
croissance des gynophores, leur fixation au sol et la croissance des parties
végétatives. Alors que l'utilisation de l'eau pendant le stade
(50 % F) serait faite uniquement au cours de la croissance
végétative et des réactions biochimiques. Selon GABRIEL,
(2008) une diminution de 10 % de la teneur relative en eau (TRE) d'un tissu
indique que son volume décroit d'environ 10 % qui correspondent à
une augmentation de 10 % du DH. Une sélection s'opérant aux
stades 50 % de floraison et de fructification, montre que les accessions A9
(73,5 %) et A6 (72,2 %) paraissaient des accessions tolérantes à
la sécheresse. Ces accessions A6 et A9 avaient un DH plus important aux
différents stades de développement. Malgré leurs DH
élevés, ces deux accessions persistent à la dessiccation
des feuilles.
Ces accessions posséderaient un nombre
élevé des vacuoles d'accumulation des composés osmotiques.
Ces vacuoles maintiennent la turgescence des cellules à résister
à la sécheresse.
La teneur relative en eau des feuilles (TRE) varie suivant
les stades de développement de la plante et des conditions de culture.
Cependant, la TRE des feuilles est plus importante au stade fructification
(87,17 %) qu'aux stades 50 % F (70,99 %) et de remplissage des gousses (78,39
%). Ceci montre que le stade le plus exigeant en eau est celui de
fructification, donc le stade le plus sensible à la sécheresse.
Le stress hydrique a réduit de manière significative la teneur
relative en eau des feuilles (TRE) des plantes «stressées» aux
stades 50 % de floraison et de fructification. En revanche, le stress n'a pas
affecté de manière significative les plantes
«stressées» au stade de remplissage des gousses. Cette
réduction de TRE au stade 50 % de floraison corrobore le résultat
obtenu par NOMBRE, (2012) chez le voandzou. De manière significative les
valeurs de la teneur relative en eau des plantes «stressées»
sont très inférieures à celles obtenues par NOMBRE, (2012)
chez le voandzou. Ceci pourrait être expliqué par le fait que la
teneur relative en eau des feuilles varie entre les espèces (inter
espèces), au sein d'une même espèce (intra espèce),
par la texture du sol, les conditions climatiques et les différents
stades de développement de la plante. Selon COLLINSON et al.,
(1997) la teneur relative en eau correspond à une signification
physiologique de l'état hydrique de la plante. C'est un indicateur de la
résistance des espèces vis-à-vis d'un stress hydrique.
Cependant, la réduction de la teneur relative des feuilles aux
différents stades de développement varie des 61,70 % (50 % de
floraison), 41,22 % (fructification) et 43,43 % (remplissage des gousses). Ce
paramètre confirme la sensibilité de la phase fructification
à la sécheresse. Au stade 50 % de floraison, les accessions A9
(23,277) et A19 (24,535) avaient de teneur relative des feuilles très
basse. Alors qu'aux stades de fructification et de remplissage des gousses, les
accessions A9 (14,70 %), A6 (15,794 %) et A16 (23,48 %), A18 (24,30 %)
présentaient de TRE très basse respectivement. La faible teneur
relative en eau des accessions pourrait être expliquée par la
réduction de la transpiration d'eau par les feuilles. En effet, plus la
teneur en eau est faible dans les feuilles, moins de stomates sont fonctionnels
donc moins de transpiration. Pour ce paramètre physiologique, peu
d'accessions maintenaient leur teneur relative en eau de feuilles en conditions
stressantes. Ces résultats ne corroborent pas ceux obtenus par COLLINSON
et al., (1997) et VURAYAI et al., (2012) chez le voandzou.
Cela pourrait expliquer que le maintien de la teneur relative en eau des
feuilles lors d'un stress hydrique permet certes, aux plantes de continuer
à réaliser leurs activités métaboliques, mais ne
conditionnerait par la circulation de la sève élaborée
dans le phloème afin d'assurer le remplissage des gousses.
Une sélection basée sur ce paramètre en
confrontant les différentes valeurs des différents traitements,
permet d'identifier les accessions potentiellement tolérantes à
la sécheresse par leur maintien en TRE. En effet, aux stades 50 % de
floraison et de fructification, les accessions A1 et A15 maintenaient plus ou
moins leur TRE des feuilles. Alors qu'au niveau de stade remplissage des
gousses, l'accession A15 a été remplacée par A7. Par
conséquent l'accession A1 semble être la plus tolérante
à la sécheresse.
La teneur en eau par rapport à la masse sèche
(TE/MS) a été affectée par le stress hydrique, mais de
manière non significative. En effet, cette teneur en eau par rapport
à la masse sèche varie entre 291,62 % et 271,07 % chez les
plantes «témoins» et «stressées»
respectivement.
Cette énorme perte de TE/MS a été
estimée à 92,95 %. Perte qui pourrait être due à la
réduction de la surface foliaire des nouvelles feuilles apparaissaient
après la levée du première contrainte hydrique. Chez les
plantes «stressées», les accessions A1 (306,18 %), A8 (303,23
%), A6 (317,83%), A19 (318,23 %), A12 (321,38%) et A11 (325,72 %),)
présentaient de TE/MS plus élevée. Ces valeurs de TE/MS
permettraient à ces accessions de produire mieux, donc d'augmenter leurs
composantes du rendement. Par ailleurs, le stress hydrique a réduit les
composantes du rendement. Cette réduction des composantes du rendement
devient de plus en plus énorme au fur et à mesure que la
quantité d'eau apportée pour les plantes diminue.
La biomasse sèche (BS) des plantes
«stressées» varie des 47,94 % (50 % de floraison), 50,80 %
(fructification) et 71,10 % (remplissage de gousses). Ces résultats sont
conformes à ceux de COLLINSON et al., (2000) et VURAYAI et
al., (2000) chez le voandzou. Au stade 50 % de floraison, les
accessions A20 (10,56 g), A10 (10,96 g), A18 (10,26 g) avaient la biomasse la
plus faible. Quant au stade de fructification, les accessions A13 (9,13 g), A11
(10,73 g), A6 (12,13 g) qui avaient une faible biomasse. Au stade remplissage
des gousses, les accessions A13 (14,36 g), A19 (15,80 g), A12 (15,83 g) et A20
(16,70 g) étaient les plus affectées par le stress hydrique. Ces
résultats corroborent ceux de MERRIEN et al., (9981a) chez le
tournesol. Cette réduction pourrait être due soit à la
sénescence prématurée des feuilles (induit par ce stress
hydrique), soit à la diminution des composés organiques dans les
organes de réserve et le faible nombre des feuilles spécifiques
au génotype de la plante. La réduction de cette biomasse
sèche serait non seulement un mécanisme de résistance
à la sécheresse, en réduisant la surface
évaporatrice de la plante, mais aussi en limitant des réactions
biochimiques. Cette réduction de biomasse sèche affecte peu les
accessions rampantes en particulier celles de type 1. Ceci pourrait être
expliqué que le recouvrement du sol par les feuillages de la plante.
C'est aussi un moyen pour les accessions rampantes de maintenir une certaine
humidité du sol. Cette biomasse sèche (BS), est positivement et
significativement corrélée à la teneur relative en eau
(r=0,70). Ceci montre que plus la quantité d'eau apportée
à la plante est importante, plus la biomasse sèche augmente.
Cette biomasse sèche est aussi positivement et
significativement corrélée à la teneur en chlorophylle
(r=0,56) et à l'indice de résistance (r=0,60). Ceci montre que
plus les pigments chlorophylliens sont importants, plus la biomasse
sèche est importante; plus l'indice de résistance augmente. Cette
biomasse sèche (BS) est négativement et significativement
corrélée au déficit hydrique (DH), au rendement en graines
par plante (RgP), à l'indice de récolte. Cette corrélation
significative, pourrait être expliquée respectivement: (i) plus le
déficit hydrique est important, plus il y a moins de réaction
chimique. Plus les matières organiques diminuent, (ii) plus la BS
diminue (induit par la sénescence prématuré), plus la
photosynthèse est perturbée, plus le remplissage des graines est
perturbé. Cette importante richesse en biomasse sèche augmente
l'indice de récolte. Ce dernier a été beaucoup
affecté par les contraintes hydriques lors de cette étude. Il est
fonction de la durée des épisodes de déficit hydrique, des
conditions climatiques et du moment où la contrainte hydrique
intervient.
Au regard de tout ce qui précède, la teneur en
chlorophylle et la biomasse sèche ne sont pas des critères de
sélection des accessions tolérantes. Par contre, la teneur
relative en eau des feuilles (TRE), le déficit hydrique (DH) et la
teneur en eau par rapport à la masse sèche (TE/MS) sont des
critères de sélections des accessions tolérantes à
la sèche. Cependant, les accessions tolérantes ont une TRE
très basse, un DH plus important et une TE/MS plus élevée
lors d'un stress hydrique. Ceci se confirme par l'indice de tolérance
à la sécheresse.
Cet indice est positivement et significativement
corrélé au rendement en graines par plante (RgP), au poids moyen
d'une graine (PMg), à l'indice de récolte (Ir). Cela montre que
ces caractères sont des paramètres pertinents pour
sélectionner des variétés tolérantes à la
sécheresse en conditions stressantes. Ces résultats sont
conformes à ceux de HAMIDOU et al., (2006) sur Vigna
unguiculata. Cette corrélation positive pourrait être
expliquée: (i) plus le poids moyen d'une graine augmente, plus le
rendement en graines augmente et plus l'indice de récolte est
élevé, (ii) plus la biomasse sèche est moins importante,
plus la perte est moindre et plus l'indice de récolte est
élevé. Par contre, cet indice de tolérance à la
sécheresse est négativement et significativement
corrélé à la biomasse sèche. Cet indice de
récolte varie entre 44,23 % (50 % de floraison) et 76,94 % (remplissage
des gousses). Les accessions A13 (0,21), A20 (0,23) et A6 (0,41), A19 (0,42)
avaient un indice de récolte très élevé lors du
stress hydrique appliqué au stade 50 % de floraison.
Alors que, pour le stress hydrique appliqué au stade de
fructification, c'étaient les accessions A16 (0,15), A15 (0,19), A13
(0,32), A11 (0,27), A18 (0,28), A20 (0,29), A6 (0,39) qui avaient un indice de
récolte plus élevé. Quant aux plantes
«stressées» au stade de remplissage des gousses, les
accessions A13 (0,46 g), A19 (0,59) possédaient un indice de
récolte plus important.
En outre, les accessions A1 (Ir0=0,27,
Ir1=0,20, Ir2=0,12 et Ir3=0,18, A13
(Ir0=0,37, Ir1=0,21, Ir2=0,32,
Ir3=0,46), A15 (Ir0=0,19, Ir1=0,12,
Ir2=0,19 et Ir3=0,25) et A6 (Ir0=0,54,
Ir1=0,41, Ir2=0,39 et Ir3=0,31), maintenaient
plus ou moins leurs indices de récolte aux différents
traitements. Le maintien de cet indice de récolte assure le maintien du
rendement en graines de la plante dans les conditions stressantes et la faible
biomasse sèche. Il pourrait être expliqué par le faible
rendement en graines lié au génotype de chaque accession dans des
conditions normales (conditions non stressantes). Puisque, plus le rendement
d'une accession serait faible plus l'effet du stress sur le rendement serait
peu perceptible. Ce rendement en graines est beaucoup plus affecté par
le stress hydrique de manière générale. Il devient de plus
en plus faible au fur et à mesure que la sécheresse persiste. Par
ailleurs, le rendement en graines par plante se fluctue des 21,23 % (50 % de
floraison), 29,08 % (fructification) et 53,58 % (remplissage des gousses).
Cette réduction de rendement en graines est significative pour les
plantes «stressées» aux stades 50 % de floraison, de
fructification et de remplissage des gousses. Ce rendement en graines est
très élevé chez les plantes «stressées»
au stade 50 % de floraison. En effet, A13 (4,70), A19 (2,65), A1 (2,68), A6
(4,71) avaient un meilleur rendement en graines au stade 50 % de floraison. Au
stade de fructification, c'étaient les accessions A7 (2,65), A11 (2,67),
A15 (2,954), A20 (2,98), A13 (3,09), A6 (4,61), A10 (4,70), ICG (5,27)
présentaient un rendement en graines très élevé.
Quant au stress hydrique appliqué au stade de remplissage des gousses,
la majorité des accessions avait un bon rendement. En plus de cela, les
accessions A1 (RgP0=6,68, RgP1=2,68,
RgP2=2,07, RgP3=3,01), A6 (RgP0=12,30,
RgP1=4,71, RgP2=4,61, RgP3=5,63) avaient
maintenu relativement leur rendement en graines par plante (RgP) aux
différents stress hydriques. Ces accessions sont
considérées comme des accessions tolérantes à la
sécheresse (rendement en graine satisfaisant).
Toutes les accessions qui avaient presque un fort indice de
résistance à la sécheresse n'avaient pas le même
rendement en graines. Cela pourrait être expliqué que cet indice
(IRS) ne devrait pas être utilisé pour sélectionner des
accessions tolérantes à la sécheresse: c'est-à-dire
des accessions qui produisent mieux dans des conditions stressantes. Cet indice
IRS pourrait être utilisé pour identifier des accessions à
caractère fourrager et non à caractère alimentaire. Cet
IRS devient de plus en plus faible au fur et à mesure que la
quantité d'eau apportée diminue. Les moyennes enregistrées
au cours de cette étude étaient de 0,49 #177; 0,12 (50 % de
floraison); 0,52 #177; 0,14 (fructification) et 0,72 #177; 0,15 (remplissage de
gousses). Le rendement en graines par plante (RgP) est significativement et
positivement corrélé au nombre de gousses par plante (NGP).
Le nombre de gousses par plante diminue au fur et mesure que
la quantité d'eau apportée pour les plantes diminue. En effet, le
nombre de gousses par plante varie entre 40,40 % (50 % de floraison), et 53,58
% (remplissage des gousses). Chez les plantes «stressées» au
stade 50 % de floraison, l'accession A6(15,66) avait le plus grand nombre de
gousses par plante.
Alors que chez les plantes «stressées» au
stade de fructification, c'étaient les accessions A13 (15,33), A16
(16,66), A9 (17,00), A15 (20,00), A14 (19,00), A6 (19,33) et A10 (23,00) qui
avaient le plus grand nombre de gousses par plante (NGP). Quant aux plantes
«stressées» au stade de remplissage des gousses, les
accessions A13 (15,33), A16 (16,66), A9 (17,00), A14 (19,00), A6 (19,33), A15
(20,00) avaient donné un nombre de gousses par plante plus
élevé. Ce NGP est très faible chez les
variétés rampantes, en particulier les accessions A7 et A14.
Cette diminution de nombre de gousses a affecté négativement le
rendement en graines par plante (RgP) et le poids moyen d'une graine (PMg).
Elle pourrait être due à: (i) la limitation de la fixation du
nombre maximum des gynophores sur la surface du substrat de pot, (ii) la
réduction du nombre des gynophores induit par la limitation de la
croissance végétative (réduction du nombre des rameaux).
En outre, l'accession A6 a maintenu relativement le NGP (NGP0=28,00,
NGP1=15,66, NGP2=14,33, NGP3=19,33) aux
différents traitements (T1, T2 et T3). Ceci montre que l'accession A6
est une accession tolérante à la sécheresse. Le
remplissage des gousses assure le poids moyen d'une graine.
Chez les plantes «témoins» le poids moyen
d'une graine varie entre 1,29 g (A11) et 2,58 g (A4). Ces résultats ne
corroborent pas ceux décrits par RABECHAULT (1960) dont leur poids moyen
varie entre 0,3 g et 0,5 g. Ce PMg élevé montre que ces
accessions étudiées sont des accessions à très
grand poids moyen de graine. Ce PMg est non seulement lié au
génotype de la plante, mais aussi à l'environnement. Le poids
moyen a été affecté par le stress hydrique. Il devient de
plus en plus faible au fur et à mesure que la durée de la
contrainte hydrique devient très importante. Les accessions A1, A6, A2,
A12 et A19 ont donné un PMg satisfaisant lors du stress intervenu au
stade 50 % F. Ce résultat confirme que les accessions tolérantes
ont un PMg élevé BLUM et al., (1998). Quant aux plantes
«stressées» au stade de fructification, les accessions A9, A7,
A15, A13, A4, A19, A6 avaient un PMg plus élevé. Contrairement
aux plantes «stressées» au stade de remplissage des gousses,
trois accessions (A8, A18, A3) présentaient un faible PMg. Le faible
poids moyen d'une graine pourrait être dû à la fois à
la réduction de la photosynthèse et à la réduction
du nombre de feuilles. A l'issu de la matrice de corrélation, l'indice
de récolte, le rendement en graines par (RgP) et le poids d'une graine
(PMg) sont positivement et significative corrélés à
l'indice de tolérance (ITS). Le dendrogramme établi sur la base
des caractères (RgP, PMg et Ir) associées à l'indice de
tolérance à la sécheresse confirme, que les accessions A6,
A1 et A2 sont des accessions tolérantes à la sécheresse
donc des variétés productives en Afrique Subsaharienne. La
tolérance des accessions A6 et A1 pourrait être due à
l'absence des fleurs sur leurs tiges principales malgré qu'elles
appartiennent à la sous espèce fastigiata.
CONCLUSION & PERSPECTIVES
Conclusion &
perspectives
La caractérisation a relevé une certaine
hétérogénéité pour les caractères
tels que la surface foliaire, la hauteur de la plante, le nombre de gousses par
plante et le taux de germination. La germination est beaucoup plus
influencée par le conditionnement des semences (durée de
conservation etc.). Tous les caractères quantitatifs mesurés
à l'exception du taux germination, permettent de discriminer les types
taxonomiques. Le type taxonomique fastigiata fastigiata Valencia est
le meilleur taxon riche en huile. La richesse en huile d'arachide est
liée au poids moyen d'une graine. Les accessions de poids moyen d'une
graine pourraient être recommandées aux industries d'huilerie.
. Cette étude a permis aussi de montrer que les
accessions d'arachide n'ont pas les mêmes comportements vis-à-vis
de la sécheresse. Les accessions tolérantes ont une teneur
relative en eau (TRE), une teneur en eau par rapport à la masse
sèche (TE/MS) et un déficit hydrique (DH) très importants
que les accessions sensibles lors d'une sécheresse. En plus, elles ont
des indices de récolte et de tolérance très
élevés. Ces caractères pourraient être des
indicateurs pour une éventuelle sélection. L'accession ICG 8431
est la meilleure accession tolérante à la sécheresse. Elle
peut être mise à la disposition des paysans. Afin d'apporter des
éclaircissements sur les «zones d'ombre», il est
nécessaire de:
Ø Identifier des variétés de bonne teneur
en huile uniquement chez la sous espèce Arachis fastigiata
hypogaea, en particulier celles qui ne possèdent pas de
fleur sur leur tige principale.
Ø Analyser la composition chimique de la teneur en
huile, c'est-à-dire analyser la composition en tocophérol et
phytostérol de l'huile d'arachide.
Ø Evaluez les variétés rampantes au champ
afin de déterminer leur performance agronomique en conditions
stressantes, c'est à dire leur potentiel génétique en
conditions stressantes afin de mettre au point un idéotype issu entre
ICG 8431 et la variété rampante potentiellement tolérante
à la sécheresse et résistante aux maladies.
Ø Evaluer l'accession ICG 8431 vis-à-vis des
maladies d'une part et d'autre part tester sa stabilité du rendement
vis-à-vis de la tolérance à la sécheresse dans des
environnements pédoclimatiques différents.
Ø Doser la proline et les pigments chlorophylliens afin
de confirmer si la faible teneur en chlorophylle de l'arachide est liée
à l'augmentation de la proline.
Ø Identifier des QTL associés à la teneur
en huile, à la teneur relative en eau, au rendement en graines par
plante, au poids moyen d'une graine, indice de récolte en conditions
stressantes.
Ø Evaluer les autres accessions de NARS (National
Agriculture Research System) afin de déterminer leur potentiel
génétique qui pourrait être avantagé dans zones
aride et semi aride.
Références
bibliographiques
ABE H., YAMAGUSHI-SHINOZAKI K., URAO T., IWASAKI T.,
HOSOKAWA D. and SHINOZAKI K., 1997. Role of Arabidopsis MYC and MYB
homologs in drought-and abscissic acid-regulated gene expression. Plant
Cell., 9, pp. 1859-1868.
ABEBE T., GUENZI A. C., MARTIN B. and CUSHMAN J. C.,
2003. Tolerance of mannitol-accumulating transgenic wheat to water
stress and salinity. AGPM-Info Technique, 353, 2006p.
AMANI L. K. et BERARD T., 2007.
Prévisions de la production nationale d'arachide au
Sénégal à partir du modèle agro
météorologique AMS et NDVI. Des interuniversitaires en
gestion des risques naturels, 54p.
AMIGUES J. P., DEBAEKE P., ITIER B., LEMAIRE G.,
SEGUIN B., TARDIEU F. et THOMAS A., 2006. Sécheresse et
Agriculture: Réduire la vulnérabilité de l'Agriculture
à un risque accru de manque d'eau. Exp. Sci. Collec. Syn. Rap.
INRA (France), 72p.
ATTI F., 2007. Effet de stress hydrique
sur le comportement hydrique et la maturité phénologique de la
vigne (Vitis vinifera L.): Etude de cinq Cépages autochtones de
Midi-Pyrénées. Thèse Doct., Univ. Tolouse, 185p.
BAINA D., MOUTARI A., MAHAMANE S., MAHAMADOU M-A.,
ABDOU M. et ABOUBACAR M. K., 2007. Deuxième rapport sur
l'état des Ressources Phyto-génétiques pour l'alimentation
et l'Agriculture, 71p.
BARAKAT F. et HANDOUFE A. 1997. La
sécheresse Agricole au Maroc in sustainability of water resources under
increasing uncertainty (proceeding of the Rabat symposium), Sér.1,
240, 11p.
BEN CEUR M., GHARBI M. et PAUL R., 1999.
L'Amélioration variétale et les autres actions
contribuant à la sécurité alimentaire en Tunisie en
matière des céréales. Sécheresse. 10, pp.
27-33.
BEN SALEM M. et VIEIRA D., A SILVA J. P.,
1991. Polymorphisme variétal de résistance à la
sécheresse chez les céréales à paille. Cas du
blé. L'amélioration de l'adaptation des plantes aux milieux
arides. John Libbey.AUPELF-UREF (Eds.), pp. 25-34.
BLANCHON D., 2009. Atlas mondial de
l'eau. Eds., autrement, 209p.
BLUM A. and EBERCON A., 1981. Cell membrane
stability as measure of drought and heat tolerance in wheat. Crops
Sci., 21, pp. 43-47.
BLUM A., 2005. Drought
resistance water-use efficiency and yield potential-are they
compatible dissonant or mutually exclusive. Aust., J. Agric. Res. 56,
pp. 1159-1168.
BOIS J. F., COUCHAT Ph. et MOUTONNET P.,
1983. Etude de la réponse à un stress hydrique de
quelques variétés de riz pluvial et de riz irrigué. In
plant soil, 11p.
BOLHUIS G., 1959. Nether. J. Agric.
Sc., 7, 4, pp. 317-326.
BOUFFIL F., 1951. Biologie évolution
et sélection de l'arachide au Sénégal. Cell Tech.
Agric. Trop., 111p.
BOYER J. S., 1968. Plant productivity and
environment. Sci., 218, pp. 443-448.
BOYLE M. G., BOYER J. S. and MORGAN P.W.,
1991. Stem infusion of liquid culture medium prevents reproductive
failure of maize at low water potential. Crop Sci., 31, pp.
1246-1252.
BRAY E. A., 1997. Plant responses to water
deficit. Trends Plant Sci., 2, pp. 48-54.
BRAY E. A., BAILEY-SERRES J. and WERETILNYK E., 2000.
Responses to abiotic stress. In Biochemistry and Molecular Biology
of Plant. Buchnau B. B., Guissem W.et Jones R.L. (Eds.), American society
of plant physiologists, Rockville, MD, pp. 1158-1203.
CARPENTER J. F., CROWE L. M. and ARAKAWA,
1990. Comparaison of solute-induced protein stabilization in aqueous
solution and in the frozen and dried states. J. Dairy. Sci., 73, pp.
3627-3636.
CECCARELLI S. and GRANDO S., 1996. Drough as
a challenge for the plant breeder. Plant growth regul., 2, pp.
55-149.
CELIA N. et SYNTHIA S., 2005. L'allergie
alimentaire à l'arachide, Univ. Paris XII-vol. de Marne 61. Avenue
du General de Gaulle 94010 Creteil Cedex, 40p.
CHEVALIER A., 1934. Etude sur les prairies de
l'Afrique de l'Ouest: Les graminées. Rev. Bot. Appl. Agric.
Trop., IV, pp. 17-48.
CHIMENTI C. A., PEARSON J. and HAL A. J., 2002.
Omotic adjustmet and yield maintenance under drought in sunflower.
Field Crops Res., 75, pp. 235-246.
CHIMENTI C. A., MARCANTONIO M. and HAL A. J., 2006.
Divergent selection for osmotic adjustment results in improved drought
tolerance in maize in both early growth and flowering phase. Field Crops
Res., 95, pp. 305-315.
CHOWDRHY M. A., RASOOL I., KHALIQ I. MEMOOD T. and
GILLIANI, 1999. Genetics of some metric traits in spring wheat under
normal and drought environments. RAHIS, 18(1), pp. 34-39.
CLARKE J. M. and CAG T. N., 1982. Evaluation
of techniques for screening for drough resistance in wezt. Crops Sci.,
22, pp. 503-506.
COLLINSON S., BRINK M. and WIGGLESWORTH D. J.,
1997.Characteristic of bambara groundnut, cultivation in
Botsxan. In Proceeding of int., bambara groundnut symposium. Univ. of
Nottingham, United Kingdom, pp. 133-142.
COLLINSON S. T., SIBUGA K. K. P., TARIMO A. J. P. and
AZAM ALI S. N., 2000. Influence of sowing date on the growth and yield
of bambara groundnut landraces in Tanzani. Exp., Agric., (36), pp.
1-13.
CONKERTONE E. E., J. et ORY L., 1989. Les
effets du stress hydrique sur la composition d'arachide, vol.1, 144 (12), pp.
593-602.
COSGROVE D. J., 1993. Water uptake by growing
cells: An assessment of the controlling roles of wall relaxation, solute
uptake, and hydraulic conductance. In J.Plant Sci., 154, pp. 10-20.
CRUZIAT P., 1974. Détermination des
plantes en eau subies par les différents organes d'une plante soumise au
desséchement. Ann. Agri., pp. 254-539.
DAALOUL A., BCHINI H. et SAYAR R., 2013.
Variabilité génétique de quelques
paramètres du système racinaire du blé (Triticum
duram Desp.) sous deux régimes hydrique. Bull. Res. Genet.
FAO-Biodiversity (Eds.), 129, pp. 25-31.
DAVIS W. J., TRADIEU F. and TREJO C. L.,
1994. How do chemical signals work in plants that grow in drying soil.
Plant Physiol., 104, pp. 309-314.
DEBAEKE P., CASADEBAIG P., HAQUIN B., MESTTRIES E.,
PALEAU J. P. et SALVI F., 2004. Simulation de la réponse
variétale du tournesol à l'environnement à laide du
modèle. SUNFL., 17, pp. 51-143.
DURE L., CROUCH M., HARAD J., HO T-HD., MUNDY J.,
QUATRRANO R., THOMAS T. and SUNG Z. R., 1989. Common amino acid
sequence domains among the LEA proteins of higher plants. Plant Mol.
Bio., 12, pp. 475-486.
FAOSTAT, 2013. Production agricole, cas
d'arachide. (http // faostat.fao.org.)
FUKAI S. and COOPER M., 1995. Development of
drought-resistant Landers using physiological traits in rice yield. Crop
Res., 40, pp. 67-86.
FELICITHY V., ANNE-MARIE T., JEREMIE L., PATRICK V. et
PHILIPPE D., 2010. Les possibilités de sélection pour le
rendement en grain du tournesol par l'étude de caractères
morphologiques et architectures. Agr. Enviro., Sér.3,
vol.17, pp. 139-142.
GABRIEL C. and MASSACCI A., 1996. Leaf
photosynthesis under drought stress in photosynthesis and the environment.
Academic publisher Dordreth. Baker N. R. (Eds. Kluwer). Advences in
photosynthesis govindjee, vol.5, ISBN 07923-14316.
GABRIEL CORNIC, 2008. Effet de la contrainte
hydrique sur la photosynthèse foliaire. Foxit corporation for
évaluation only, foxit reader, 36p.
GILLIER P. et SIVESTRE P., 1969. L'arachide
collection: Techniques agricoles et productions tropicales, pp.
16-22.
GIMENEZ C., MITCHELL VJ. and LAWLOR DW., 1992.
Regulation of photosynthetic rate of two sunflower hybrids under water
stress. Plant Physiol, 98, pp. 516-524.
GREGORY W. C. and GREGORY M. P., 1967. Indice
mutation and species hybridization and depreciation of arachis. Ciencia
culture; 19, pp. 166-174.
GRENIER C. H., INZE D. and TARDIEU F., 2000.
Spatial distribution cell division rate can be deduced from that of P34 kinas
activity in maize leaves grown in contrasting conditions of temperature and
water status. Plant Physiology, 124, pp. 1393-1402.
GUPTA S. A. and BERKOWITZ G. A., 1987.
Osmotic adjustment, symplast volume, and none stoma tally mediated
water stress inhibition of photosynthesis in wheat. Plant Physiol, 87,
pp. 1040-1047.
HAMIDOU F., 2006. Paramètres
physiologiques, biochimiques et agronomiques pertinents pour les programmes
d'amélioration de l'adaptation du niébé (Vigna
unguiculata L. Wap). Thèse doct. Univ., Ouaga, 154p.
HAMMONS S., 1973. The origin and history of
the groundnut Landers Royaume. Crop, J., Smartt, Uni., chapman and
Hall, pp. 24-42.
HARBI A., KRISHMAN A., AMBAVARAM M. M. and PEREIRA A.,
2010. Molecular and physiological analysis of drought stress in
Arabidopsis reveals early responses leading to acclimation in plant growth.
Plant Physiology, 54, pp. 1254-1271.
HASSAAN R. K., 2003. Effect of drought stress
and yield components of some Triticale genotypes. Ann. Agric. Sci.
(Cairo). 48(1), pp. 117-129.
HERSELMAN L., 2003. Genetic variation among
Southern African cultivated peanut (Arachis hypogaea L.) genotypes as
revealed by AFLP analysis. Euphytica; 133, pp. 27-319.
HOPKINS W. G., RAMBOUR S. et EVARD C. M.,
2003. Physiologie végétale, ISBN 27445-0089-5,
532p.
IBPGR (Now Biodiversity international) and ICRISAT,
1992. Descriptors for groundnut, 125p.
IJAZ RASOOL N., ELBRAMAWY M. A. S., SABA T. and
AHMAD-UR-R. S., 2012. Consumptive use of water and genetically
assessment of wheat genotypes to defy worrisome prevalence of water stress.
Sarhad .J. Agri. Sér.4, Vol. 28, 6p.
JONES M. M. and TURNER N. C., 1980. Turgot
maintenance by osmotic adjustment, a review and evaluation. Adaptation of
plant to water and High Temperature stress. Wiley, New York, USA. pp.
87-107.
JONES H. G., 1992. Plant and
Microclimates: A quantitative approach to environmental plant physiology.
Cambridge Univ. Press London, Sp.
JONGDEE B., FUKI S. and COOPER M., 2002. Leaf
water potential and osmotic adjustment as physiological traits to improve
drought tolerance in rice. Field Crop Res., 76, pp. 153-163.
KHALFAOUI J. L., 1988. Approche de
l'amélioration génétique de l'adaptation à la
sécheresse des espèces cultivées en zone semi- arides.
Application au cas de l'arachide (Arachis hypogaea L.) destinée à
la région sèche du Sénégal. Thes. Doct.Univ.
du Paris-Sud., pp. 67-68.
KIM J. Y. MAHE A., BRANGEON J. and PRIOUL J. L.,
2000. A maize vacuolar vertase, IVR2, is induced by water stress.
Organ/tissue specificity and diurnal modulation of expression. Plant
physiol., 124, pp. 71-84.
KRAVICKAS A, 1969. The origin variability and
spread of the groundnut (Arachis hypgaea L.) In :The Domestication
and exploitation of plant and animals Ucko R.J. Dimbledy W. C. (Eds.).
Londres GB : Duckworth, pp. 40-427.
KUMAGA F. K., ADIKU S .G. K. and OFORI K., 2003.
Effect of post-floering Water stress on dry matter and yield of three
Tropical Grain legumes. Int. J. Agric. Biol., 4 pp. 405-407.
LACAZE X., 2006. Développer des
plantes résistantes à la sécheresse. Exemple des
recherches sur les céréales menées à l'institut de
l'évolution. Sci. Tech., 16p.
LANDI P., SANGUINETI M. C., CONTI S. and TUBEROSA R.,
2001. Direct and correlate responses to divergent selection for leaf
abscissic acid concentration in two maize populations. Crop Sci., 41,
pp. 334-335.
LARBI A., MEKLICHE A., ABDELLLAOUI K., BENNOUR A.,
BETAHAR N., KADI S. et ABDELGUERFI A., 1997. Effet du stress hydrique
sur le comportement physiologique des dix populations de
Médicago intertxrta. In: CIHEAM, pp.167-171.
LEVITT J., 1980. Reponses of plants to
environmental stress. water radiation salt and other stress, vol.1 New
York, USA, pp. 395-434.
LUDLOW M. M., CHU A. C. P., CLEMENTS R. J. and
KERSLAKE R. G., 1983. Adaptation of species of Centrosema to water
stress; Australian. J. Plant Physiol, 10, pp. 119-130.
LUU D.T. and MAUREL C., 2005. Aquaporines in
challenging environment: molecular gear for adjusting plant water status.
Plant Cell and Environment, 28, pp. 85-96.
MAKANDA I. TONGOONA P., MAAMBA R., ICISHAHAYO D. and
DERERA J., 2009. Path coefficient analysis of bambara groundnut pod
yield components at for planting dates. Res. J. Agric. Biol. Sc., 5
(3), pp. 289-292.
MAUREL C. and CHRISPEELS M. J., 2001.
Aquaporines: entry into plant water relations. Plant Physiol,
125, pp. 135-138.
MATTHEWS M. A. and BOYER J. S., 1984.
Acclimation of photosynthesis to low leaf water potentials. Plant
Physiol, 74, pp. 161-166.
MAYEUX A. H., 1999. Etude sur le taux
d'allogamie chez l'arachide. Oléagineux, 18, pp. 571-574.
MERAH O., 1999. Utilisation de la
discrimination isotopique du carbone pour l'amélioration de la
tolérance à la sécheresse chez le blé dur
(Triticum durum Dest.) dans les régions
méditerranéennes. Thés. Doct. Ensa-Rennes, 172p.
MERRIEN A., BLANCHET R. and GELFI N., 1981a.
Relationship between water supply, leaf area development and survival,
and production in sunflower (Helianthus annuus L.). Agri.1,
pp. 917-922.
MERREIN A. and GRANDIN L., 1990. Comportement
hydrique du tournesol: Synthèse des essais d'irrigation. Le
tournesol et l'eau, Banchet R. et Merrien Cetiom (Eds.) Pub., Paris, ISBN
9993-88, pp. 75-90.
MOHARRAM J. and HABIB M., 2011. Evaluation of
10 wheat cultivar under water stress at Moghan (Iran) conditions. Afric.J.
Biotech. 10 (53), pp. 10900-10905.
MOJAYAD F., 1993. Adaptation à la
sécheresse, photosynthèse et photoinhibition chez le
tournesol (Helianthus annuus L.). PhD Thesis. Institut National
Polytechnique, Toulouse, France, 79p.
MOJAYAD F. and PLANCHON C., 1994. Stomatal
and photosyntgetic adjustment to water deficit as the expression of heterosis
in sunflower. Crop Sci., 34, pp. 103-107.
MORGAN J. M., 1983. Osmoregulation as a
selection criterion for drought tolerance in wheat Aust. J. Agric.
Res., 34, pp. 607-614.
MORGAN J., 1984. Osmoregulation and water
stress in higher plants, Ann. Rev. Plant Physiol, 35, pp.
299-319.
MORGAN J. M., 2005. Osmoregulation and water
stress in higher. Ann. Rev. Plant physiol., 35p.
MORIZET J. et MERRIEN A., 1990. Principaux
traits du comportement hydrique du tournesol. Le tournesol et l'eau.
Blanchet R. et Merrien A., Cetiom (Eds.) Pub., Paris, pp. 7-21.
NDOUR P. et DANTHU P., 1998. Effet des
contraintes hydriques et amines sur la germination de quelques Acacia
africana. Campa C., Grignon C., Gueye M. et Hamon S., eds., colloque
et séminaires: l'acacia au Sénégal. Paris:
ORSTOM, pp. 105-122.
NEFFATI M., 1997. Caractérisation
morpho-biologique de certaines espèces végétales nord
africaines: implication pour l'amélioration pastorale. Thès.
Doc. Univ. Grande Bretagne (Belgique), Sp.
NIGAM S. N. RAO V. RAMANATHA and GIBBONS R. W., 1983.
Utilization of natural hybrids in the improvement of groundnut
(Arachis hypogaea L.). Exp. Agric., 19, pp.
355-359.
NOMBRE ASSET, 2012. Sélection des
variétés de voandzou [Vigna subterranea
(L.) Verdic. Comb. Nov.] pour la tolérance au
stress hydrique. Master Univ. Ouaga (Burkina Faso), 66p.
NOORKA I. R. KHALIQ R. I., AKRAM Z. and IQBAL S.,
2009a. Inheritance studies of physio-genetic traits in spring wheat
under normal and moisture stress environment. J. Agric. Sci 1(1), pp.
29-34.
NOORKA I. R. and AFZAL., 2009b. Global
climatic and environmental change impact on agricultural research challenges
and wheat productivity in Pakistan. Earth Sci. O
Frontier, 16, 100p.
OBER E. S., LE-BLOA M., CHARK C. J. A., ROYAL A.,
JAGGARD K.W. and PIDGEON J.D., 2005. Evaluation of physiological trait
as selection criteria for drought tolerance in sugar beet, Field Crops
Res., 91, pp. 231-249.
ONWUBIKO N. I.C., ODUM O. B. UTTAZI C.O. and POLY-
MBAH P. C., 2011. Studies on the adaptation of bambara groundnut
(Vigna subterranea L. Verdc): In owerri Southeastern Nigeria. New York Sc.
J., 4(2), pp. 60-67.
OUVRARD O., CELLIER F., FERRARE K., LAMAZE T., DUPUIS
J-M. and PASSIOURA J. B., 1996 The yield of crops in relation to
drought. In: Physiology and determination of Crop yield. Crop sci.
Boote k. J. et al.(Eds.). Society of America.Madison
WI, pp. 343-359.
PASSIOURA J. B. 1996. Drough and drough
tolerance. plant growth regul. SORRELLS M. E., DIAB A., NACCHIT
M.(Eds.).Comparative genetic of drought tolerance. Options
méditerranéenne. Séminaire méditerranéen,
20, pp. 79-83.
PERRIER A., 1975. Etude physique de
l'évapotranspiration dans les conditions naturelles. An.,
Agron. 26(2), pp. 105-123.
RABECHAULT H., 1960. Recueil iconographique
des espèces comestibles de légumineuses africaines. «
Riz et riziculture » Paris, 61p.
RAMANJULU S. and BARTELS D., 2002. Drought
and dessication-induced modulation of gene expression in plants. Plant Cell
Environ, 25, pp. 141-151.
REDDY A.R., CHAITANYA K.V. and VIVEKANADAN M., 2004.
Drought-induced response of photosynthesis and antioxidant metabolism
in higher plants. J. Plant physiol, 161, pp. 1189-1202.
REJEB, M. N. et BEN SALEM M., 1993. Les
divers mécanismes d'adaptation à la sécheresse chez les
végétaux supérieurs, cas du blé et du caroubier.
Bul., Soc. Sc. Nat., Tunisie, 22, pp. 49-52.
REKIKA D., 1997. Identification et
analyse génétique des caractères physiologiques
liés au rendement en conditions de sécheresse chez le blé
dur. Intérêt potentiel des espèces sauvages
apparentées pour l'amélioration de ces caractères.
Thèse Doct. Ensa-Montpollier, 158p.
REYMOND M., MULLER B., LEONARDI A., CHARCOSSET A., and
TARDIEU F., 2003. Combining to quantitative trait loci analysis and an
ecophysilogical model to analyze the genetic variability of the respones of
leaf growth to temperature and water deficit. Plant Physiology, 131,
pp. 664-675.
REYNIER F.N. et JACQUOT M., 1978. Demande
pour l'obtention de la résistance variétale à la
sécheresse cas de riz pluvial. Agro. Trop., XXXII-4,4P.
ROY-MACAULEY H., PIMENTEL, ABDOU A. C.S. DIOUF O., and
SARR B., 1999. Effect of irrigation regimes on the water status of
cowpea cultivated in the field. Physiol. Mol. Biol Plants. (5), pp.
153- 159.
SAUGER L., 1949. Hybridation de l'arachide
à Bambey. Paris. Agr. Trop., vol. 4,12, pp. 618-624.
SCHONFELD M. A. JOHNSON B. F. CRAVER B. F. and
MORHINWEG D. W., 1988. Water relations in winter wheat as drought
resistance indicators. Crop Sci., 22, pp. 526-531.
SCHWACKE R., GRALLATH S., BREITKREUZ K. E., STRANSKY
H., FROMMER W. B. and RENTSCH D., 1999. Le ProT1 a transporter for
proline glycine betaine and ?-amino butric acid in tomato pollen. Plant
Cell, 11, pp. 377-391.
SEIFOLLAH POORMOHAMMAD K., 2007. Analyse
génétique des réponses physiologiques de tournesol
(Hélianthus annuus L.) soumis à la
sécheresse. Thèse doct. Univ., de Toulouse, 213p.
SEKI M., NARUSAKA M., ISHILA J., NANJO T., FUJITA M.,
OONO Y., KAMIYA A., NAKAJIMA M., ENJU A., SAKURAI T., SATOU M., AKIYAMA K.,
TAJI T., YAMAGUCHI-SHINOZKI K., CARNINCI P., KAWAI J., HAYASHIZAKI Y. and
SHINOZAKI K., 2002. Monitoring the expression profiles of 7000
Arabidopsis genes under drought, cold and high-salinity stress using a
full-length cDNA microarray. Plant Cell, 13, pp. 61-72.
SHIMOZAKI K., YAMAGUSHI-SHINOZAKI K. and SEKI M.,
2003. Regulatory network of gene expression in the drought and cold
stress responses. Current Opinion. Plant Biology, 6, pp. 410-417.
SHIMOZAKI K., YAMAGUSHI-SHINOZAKI K., 2007.
Gene networks involved in drought stress response and tolerance. J. of
Experimental Bot., 58, pp. 221-227.
SINGH A., SMAARTT J., SIMPSON C. E. and RAINA S. N.,
1998. Genetic variation molecular polymorphism in groundnut
(Arachis hypogaea L.). Gen. Res.Crop Eval., 45, pp.
26-119.
SINGH C., KHAJURIA H. N. SINGH A. and SHARMA F., 1991.
Acacias forarid regions. Acta. Bot. indica, 19, pp. 29-32.
SLAMA A., BEN SALEM M., BEN NACEUR M. et ZID E. D.,
2005. Les céréales en Tunisie: production,
effet de la sécheresse et mécanisme de résistance.
Nat. Res. Agr. Tunisie (Inrat). Univ. Elmanar. (
http://www.john libbeyeurotext.).
SMART L. B. MOSKAL W. A., CAMERON K. D. and BENNETT A.
B., 2001. MIP Genes are down-regulated under drought. In: induced
mutation against plant disease, pp. 385-392.
SORRELLS M. E, DIAB A. and NACHIT M., 2000.
Comparative genetics of drought tolerance. Options
mediterraneennes, Ser. A, 40, pp. 191-201.
SUBBARAO G. V., JOHANSEN C., SLIKARD A. E., NAGESWARA
R. C., SAXENA N. P. and CHAUHAN Y. S., 1995. Strategy for improving
drought resistance in grain legume. Plant Sci., 14, pp. 469-523.
TARDIEU F., ZHANG J., and GOWING D. J. G., 1993
a. A model of stomata control by both ABA concentration in the xylem
sap and leaf water status. Test of the model and of alternative mechanisms for
droughted and ABA-fed field-grown maize. Plant Cell Environ.,
16, pp. 413-420.
TARDIEU F. and DAVIES W. J., 1993 b.
Integration of hydraulic and chemical signalling in the control of stomata
conductance and water status of droughted plants. Plant Cell and
Environment, 16, pp. 341-349.
TEULAT B., 1997 a. Recherche de marqueurs
moléculaires liés à la tolérance au déficit
hydrique chez l'orge (Hordeum vulgare L.). Thés. Doct.
Ensa-Montpollier, 191p.
TEULAT B., REKIKA D. NACHIT M. and MONNEVEUX P., 1997
b. Comparative osmotic adjustments in barley and tetraploid wheats.
Plant Breeding, 116, pp. 519-523.
TEULAT-MERAH B., ZOUMAROU W., DOUIMI R., BEN SALEM M.,
BAHRI H. et THIS D., 2000. Etude de la tolérance à la
sécheresse chez les céréales: L'orge comme modèle
biologique approche QTL. Modèles biologiques à
l'amélioration des plantes, 24p.
TEULAT B. BORRISE and THIS D., 2001. News QTL
identified for plat water statue, water soluble carbohydrate and osmotic
adjustment in a barbey population growth chamber under two water regimes.
Theor. Appl. Genet, 103, pp. 161-170.
TEYRMAN A. W. MITCHALL V., DRISCOLL S.P. and LAWLOR
D.W., 2002. Effect of water deficit and its interaction with
CO2 supply on the biochemistry and physiology of
photosynthesis in sunflower. J. Exp. Bot., 375, pp. 1781-1791.
TRIBOI E. and TRIBOI-BOLNDEL AM., 2002.
Productivity and grain or seed
compostion: a new approach to an old problem. Eur J.
Agr.,16, pp. 86-163.
TRIBOI BLONDEL A. M., MESSOUD J., DEBAEKE P., LECOEUR
J. and VEAR F., 2004. Heredity of Sunflower leaf characters useable as
yield predictors. Proc.16 thint Sunflower Conf. Fargo (USA),
2, pp. 24-517.
TUBERO R., SANGUINETI M. C., LANDI P., SALVIE S.,
CASARINI E. and CONTI S., 1998. RFLP mapping of quantitative trait
loci controlling abscissic acid concentration in drought-stressed maize
(Zea mays L.). Theor App Genet, 97, pp. 744-755.
TUNER N. C., 1979. Drought resistance and
adaptation to water deficit in crops plants. Stress physiol. In Crop
Plants, Mussell H. et Staples R. C. (éds). Wiley interscience,
New York, pp. 303-372.
TURNER N. and JONES M., 1980. Turgot
maintenance by adjustment: a review and evaluation. In Adaptation
of plants to water and high temperature stress, N Turner and kramer P.
(eds.), wiley, New York, pp. 87-107.
TURNER N., 1986. Adaptation to water deficit:
A changing perspective. Aust, J. plant physiol., 13, pp. 90-175.
TURNER N. C., 1997. Further progress in crop
water relation. Adv. Agr. 58, pp. 293-338.
TURNER N. C., WRIHT G. C., SIDDIQUE K. H., 2001.
Adaptation of grain legume to water limited environment, Adv.
Agri., 76, pp. 193-231.
TEYRMAN S. D., NIEMIETZ C.M. and BRAMLEY H.,
2002. Plant aquaporins: multifunctions water and solute channels with
expanding roles. Plant Cell Environ, 25, pp. 173-194.
UNO Y., FURIHATA T., ABE H., YOSHIDA R., SHINOZAKI K.,
and YAMAGUSHISHINOZAKI K., 2000. Arabidopsis basic leucine zipper
transcription factors involved in abscssic acid-dependant signal transduction
paf way under drought and high-salinity conditions. Proceedings of the
Nat. Acad. Of Sci. Of the USA, 97, pp. 11632-11637.
YAMADA S., KOMORI. T, MEYERS P. N., KUWATA S., KUBO T.
and IMASEKI H., 1997. Expression of plasma membrane water channel
genes under water stress in Nicotiana excelsior. Plant Cell Physiol,
38, pp. 1226-1231.
YAMAGUCHI-SHINOZAKI K., KOIZUMI M., URAO S. and
SHINOZAKI K., 1992. Molecular cloning of 9 cDNA that are responsives
to dessication in Arabidopsis thaliana Sequence analysis of one cDNA
that encodes a putative transmembrane channel protein. Plant Cell
Physiol., 33, pp. 217-224.
YAMAGUCHI-SHINOZAKI K. and SHINOZAKI K., 2005.
Organization of cis-acting regulatory elements in osmotic and
cold-stress-responsive promoters. Trends Plant Sci., 10, pp. 88-94.
VURAYAI R., EMONGOR V. and MOSEKI B., 2011b.
Effect of water stress imposed at different ground and development
stage on morphological traits and yield of Bambara groundnut (Vigna
subterranean L. Verdc). An., J.,of Plant physiol., ISSN 1557-4539
6(1): pp. 17-27.
WAHBI JAOUAD, LAMIA H., NAOUFEL S. et MOHAMED L. K.,
2010. Etude de la germination des graines d'Acacia tortilis
sous différentes contraintes abiotiques. Biotechnol. Agron. Soc.
Environ.14(4), pp. 643-652.
WILSON J., FISHER M., SCULTZE E., DOLBY G. and LYDLLOW
M., 1978. Comparison between pressure volume and dew point-hygrometry
technique for determining water relation characteristics of grass and legume
leaves: Oecologia, 41, pp. 77-88.
WIKIPEDIA, 2014.
http://fr.wikipedia.org/wiki/Huile_d%27arachide
WOOD L. M. W., 1968. Oléagineux J.
Agric. Res., 19, 2 pp. 241-251.
WU Y. and COSGROVE D. J., 2000. Adaptation of
roots to low water potentials by changes in cell wall extensibility and cell
wall proteins. J. of Exp. Bot., 51, pp. 1543-1553.
YOSHIDA S., 1981. Fundamentals of rice crop
Sc., International. Rice Res Int., P.O. Box 933, Manilia Philipines,
269p.
ZAGRE B. et BALMAD
D., 1998. Point sur la recherche des
variétés adaptées à la sécheresse.
Rapport final 17p.
ZAGRE M'BI BERTIN, 2004.
Hérédité de la précocité et de
quelques caractères associés au rendement chez l'arachide
(Arachis hypogaea L.). Thèse, Doct., Univ. Cocody (Cote
d'Ivoire), 110p.
ZHANG J. and DAVIES WJ., 1989. Abscisic acid
produced in dehydrating roots maize noble the plant to measure the water stats
of soil. Plant Cell Environ., 12, pp. 73-82.
ZHANG J., NGUYEN H. T. et BLUM A., 1999.
Genetic analysis of osmotic adjustment in crop plants. J. of
Exp.Bot., 50, pp. 291-302.
ZHU C., SCHRAUT D., HARTUNG W. and SCHAFFINER A. R.,
2006. Differential responses of maize MIP genes to salt stress and
ABA. J. of Exp. Bot., 56, pp. 2971-2981.
ZINSELMEIR C., LAUER M. J. and BOYER J. S., 1995.
Reversing drought-induced losses in grain yield: sucrose maintains
embryo growth in maize potential disrupts carbohydrate metabolism in maize
(Zea mays L.) ovaries. Plant Physiol., 107, pp. 385-391.
ZINSELMEYER C., JEONG B.R., and BOYER J.S.,
1999. Starch and the control of kernel number in maize at low
potentials. Plant physiol., 121, pp. 25-26.
Annexe 1: Extraction d'huile
Les étapes de cette extraction d'huile sont les
suivantes:
- Oter la pellicule des graines (tégument
séminal).
- Broyer les graines.
- Peser 5 g de broyat par accessions.
- Emballer avec papier filtre et le mettre dans le soxhlet.
- Mettre en place la plaque chauffante à 70°C, le
système de refroidissement et la pierre ponce dans le ballon à
col rodé (29/32).
- Peser le ballon contenant la pierre ponce (P0).
- Faire couler l'hexane (200 ml) dans le soxhlet contenant
l'échantillon.
- Mettre en marche la plaque chauffante et le système
de refroidissement pendant une durée de 3 heures dont le nombre de
siphonage varie de 12 à 14 siphonages.
- Evaporer l'hexane au rotavapor à la
température d'ébullition du solvant (61° C).
- Mettre le ballon contenant l'huile à l'étuve
à 100° C.
- Effectuer une série de pesé (3 fois) à
chaque 40 mn afin d'obtenir un poids constant de l'échantillon.
- Calculer la teneur en huile selon la formule suivante:
TEH(%)= 100*(P1-P0)/E, avec P0= poids du ballon vide contenant la pierre, P1=
poids du ballon contenant l'huile plus la pierre ponce, E=prise d'essai (5
g).
Photo 5: Huiles des accessions ICG8385, A17, A8
respectivement de gauche à droite
 
Photo 3:Aperçu des huiles de certaines
Accessions
|
Dates d'arrosage
|
Quantité d'eau
apportée/arrosage
|
Traitements arrosés
|
|
16/11.13
|
1000 ml
|
T0 ;T2 ;T3
|
|
18/11/13
|
800 ml
|
T0;T2;T3
|
|
19/10/13
|
1000 ml
|
T0;T2;T3
|
|
20/10/13
|
1000 ml
|
T0;T1;T2;T3
|
|
21/11/13
|
1000 ml
|
T0;T1;T3
|
|
22/11/13
|
1000 ml
|
T0;T1;T3
|
|
24/11/13
|
1000 ml
|
T0;T1
|
|
26/11/13
|
1000 ml
|
T0; T1; T2
|
|
27/11/13
|
1000 ml
|
T0;T1;T2;T3
|
|
28/11/13
|
1000 ml
|
T0;T2;T3
|
|
29/11/13
|
1000 ml
|
T0;T2;T3
|
|
01/12/13
|
1000 ml
|
T0;T2;T3
|
|
02/12/13
|
1000 ml
|
T0;T1;T2;T3
|
|
03/12/13
|
1000 ml
|
T0;T1
|
|
04/12/13
|
1000 ml
|
T0
|
|
05/12/13
|
1000 ml
|
T0
|
|
07/12/13
|
1000 ml
|
T0
|
|
09/12/13
|
1000 ml
|
T0;T1;T2;T3
|
|
10/12/13
|
1000 ml
|
T0;T1;T2;T3
|
|
14/12/13
|
1000 ml
|
T0;T1;T2;T3
|
|
16/12/13
|
500 ml
|
T0;T1;T2;T3
|
|
17/12/13
|
1000 ml
|
T0
|
|
19/12/13
|
1000 ml
|
T0
|
|
21/12/13
|
1000 ml
|
T0
|
|
22/12/13
|
1000 ml
|
T0
|
|
26/12/13
|
100 ml
|
T0;T1;T2;T3
|
T0 : Plantes « témoin».
T1 : Plantes « stressées » au stade 50%
floraison fructification suivis des stress intermittents.
Durée totale stress : 10+ 7+7+6+7+10=47jours de stress.
T2 : Plantes « stressées » au stade de
fructification suivi s des stress intermittents.
Durée totale de stress : 7+6+8+10= 31 jours de stress.
T3 Plantes « stressées » au stade remplissage
des gousses suivis des stress intermittents.
Durée totale de stress : 6+8+10=24 jours de stress.
Annexe 2: Calendrier d'arrosage
durant toute l'expérimentation en pots
|
Dates d'arrosage
|
Quantité d'eau
apportée/arrosage
|
Traitements arrosés
|
|
18/09/13 au 14/10/13
|
1000 ml chaque 2jrs
|
T0 ;T1 ;T2 ;T3
|
|
17/10/13
|
800 ml
|
T0; T2;T3
|
|
18/10/13
|
1000 ml
|
T0;T2;T3
|
|
19/10/13
|
1000 ml
|
T0;T2;T3
|
|
21/10/13
|
1000 ml
|
T0;T2;T3
|
|
22/10/13
|
1000 ml
|
T0;T2;T3
|
|
23/10/13
|
1000 ml
|
T0; T1 ;T2;T3
|
|
24/10/13
|
1000 ml
|
T0;T1;T2;T3
|
|
25/10/13
|
1000 ml
|
T0;T1;T2;T3
|
|
26/10/13
|
1000 ml
|
T0;T1;T2;T3
|
|
28/10/13
|
1000 ml
|
T0;T1;T2;T3
|
|
29/10/13
|
1000 ml
|
T0;T1;T2;T3
|
|
30/10/13
|
1000 ml
|
T0;T2;T3
|
|
31/10/13
|
1000 ml
|
T0;T2;T3
|
|
01/11/13
|
1000 ml
|
T0;T3
|
|
03/11/13
|
1000 ml
|
T0; T3
|
|
05/11/13
|
1000 ml
|
T0;T3
|
|
06/1113
|
1000 ml
|
T0;T1;T3
|
|
0711/13
|
1000 ml
|
T0;T1;T2;T3
|
|
10/11/13
|
1000 ml
|
T0;T1;T2;T3
|
|
11/11/13
|
1000 ml
|
T0;T1;T2;T3
|
|
12/11/13
|
1000 ml
|
T0;T1;T2;T3
|
|
14/11/13
|
1000 ml
|
T0;T1;T2;T3
|
|
15/11/13
|
1000 ml
|
T0;T2;T3
|
Annexe 3: Vue d'ensemble des deux
expérimentations
Vue d'ensemble de l'essai en conditions naturelles (Au
champ)

L'essai en conditions contrôlées, après
applications des deux premiers traitements (T1 et T2)

Annexe 4: Dispositif
expérimental en conditions naturelles
Répétition I
ICG8550
ICG8431
ICG8352
ICG8594
ICG8781
ICG8460
ICG8454
ICG8443
ICG9991
ICG9989
ICG8746
ICG8385
ICG8863
ICG8724
ICG8848
ICG8606
ICG8439
ICG10005
ICG8429
ICG8398
1m
Répétition II
ICG8431
ICG8460
ICG10005
ICG8385
ICG8746
ICG8439
ICG8398
ICG8848
ICG8863
ICG8724
ICG8606
ICG8454
ICG8443
ICG8594
ICG9991
ICG8550
ICG9989
ICG8352
ICG8781
ICG8429
1m
ICG8724
ICG8443
ICG8781
ICG9991
ICG8863
ICG8429
ICG8594
ICG9989
ICG8385
ICG8431
ICG8352
ICG8439
ICG10005
ICG8460
ICG8606
ICG8550
ICG8746
ICG8398
ICG8848
ICG8454
Répétition II
Annexe 5: Dispositif
expérimental en conditions contrôlées

|
|


