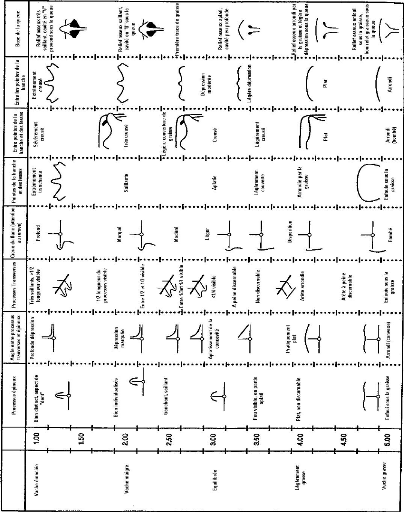
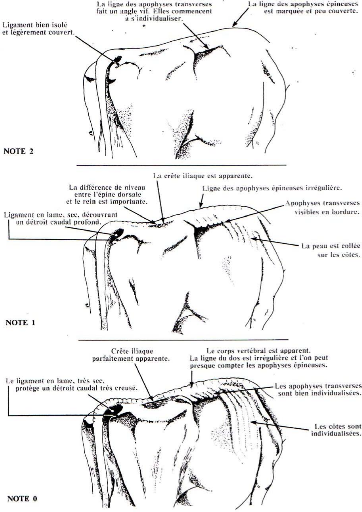
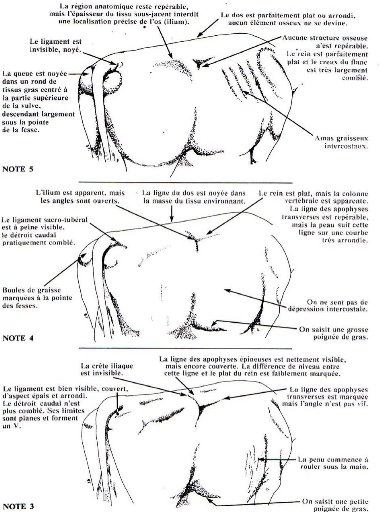
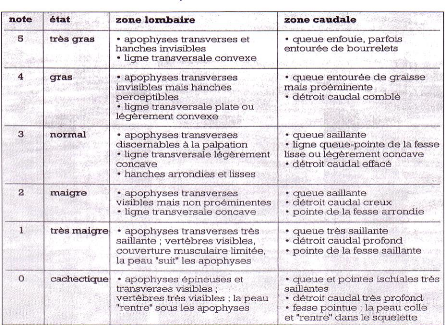
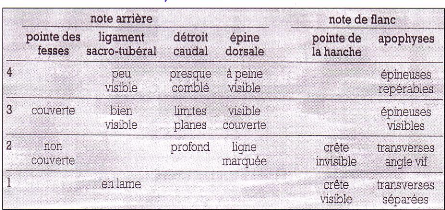
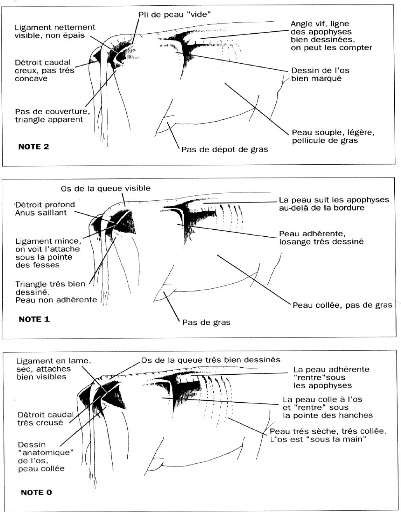
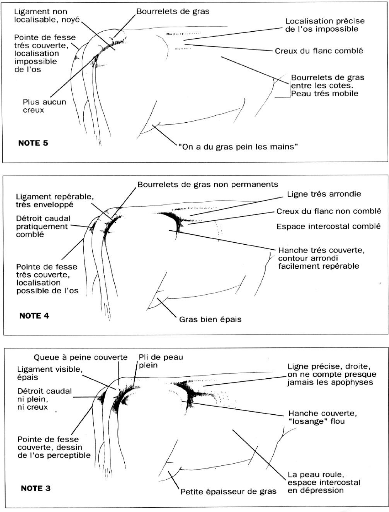
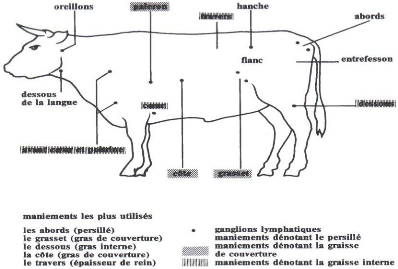
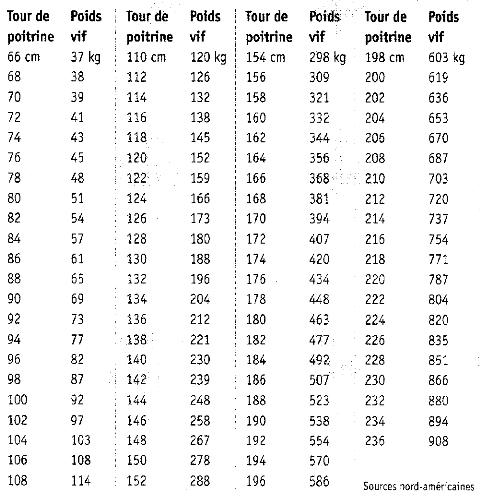
Note d'état corporel et reproduction chez la vache laitière( Télécharger le fichier original )par Pierre Froment Ecole nationale vétérinaire d'Alfort - docteur vétérinaire 2012 |

Mamelle Triglycérides 2) Appréciation par l'évolution de la note d'état corporel
D'une manière générale, l'état corporel des animaux est un des indicateurs (avec les performance de production, les résultats de reproduction et la composition du lait) de l'efficacité et de la sécurité d'une ration. Les recommandations dans ce domaine préconisent moins de 10% des vaches ayant un état supérieur à 4 ou inférieur à 2,5. Il faudra cependant tenir compte du stade physiologique des animaux, cette norme doit être ajustée lorsque beaucoup d'animaux sont en début de lactation [30]. Au cours de la seconde partie de lactation, le retour à un bilan énergétique positif s'accompagnera d'une reprise d'état, traduisant la reconstitution des réserves corporelles [25]. III. VARIATIONS NORMALES DE LA NOTE
D'ETAT
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Corrélation perte |
NEC au vêlage |
NEC post-partum |
||
|
1ère lactation |
2ième lactation |
1ère lactation |
2ième lactation |
|
|
Génotypique |
- 0,15 à - 0,26 |
-0,11 à -0,48 |
-0,70 à -0,99 |
-0,56 à -0,91 |
|
Phénotypique |
0,54 |
-0,65 |
||
4) Saison de vêlage
Dans l'étude de Drame et al. [25] la saison de vêlage s'est révélée être un important facteur de variation de l'état corporel. Les vaches vêlant en période de stabulation ont gardé un état corporel significativement inférieur à celles vêlant en pâture.
5) Cas particulier de la génisse
Si les critères de notation et les objectifs sont globalement les mêmes, il convient d'ajouter une nuance dans la conduite alimentaire des génisses de renouvellement car les besoins ne sont pas les mêmes que ceux des vaches adultes du fait de la croissance. Dans les premiers mois de vie, la croissance peut être très importante. Néanmoins, celle-ci doit être contrôlée par la suite, afin de ne pas trop engraisser la mamelle et de ne pas porter préjudice au potentiel de production laitière.
a) Jusqu'à la puberté
La puberté des génisses survient à un niveau de poids vif constant pour une race donnée. Ainsi, la puberté des races Holstein et Normande a lieu à 40% du poids vif adulte soit environ 9 à 12 mois alors que celle de la race Montbéliarde ou encore Tarine survient à 50% de ce poids ce qui en fait des races plus tardives : 13 à 16 mois [12].
Une croissance prépubère trop élevée engendre au niveau de la mamelle, le développement d'un tissu adipeux au détriment du tissu parenchymateux. Des génisses à haut potentiel ayant eu une croissance excessive (Gain moyen Quotidien, GMQ = 950 g/j) auront une production amoindrie de 5% par rapport à des homologues ayant eu une croissance raisonnable de 750 g/j [12].
De six à douze mois, il est raisonnable de conseiller une croissance de 700 à 750 g/j [2, 12].
b) Après la puberté
La conduite dépendra du schéma d'élevage (vêlage à deux ou trois ans) et de la qualité des pâturages.
- Pour des génisses vêlant à deux ans et qui vont bénéficier d'un pâturage de bonne qualité permettant un GMQ de 1000 à 1100 g/j, une croissance hivernale de 600 g/j est suffisante,
- Si elles bénéficient d'un pâturage de moins bonne qualité, on poussera le GMQ à 750
g/j,
- Dans le cas d'un vêlage à trois ans, le bon compromis économique est celui d'un GMQ de 400 à 500 g/j en bénéficiant d'une bonne croissance compensatrice au pâturage [12].
c) Conséquences sur la note d'état corporel
Les génisses laitières doivent rester « crues » ; c'est-à-dire avoir des reliefs osseux visibles et légèrement recouverts de gras autour de la puberté pour ne pas risquer l'engraissement préjudiciable vu précédemment. La note d'état doit être de trois autour de la puberté (tableau 3).
Tableau 3 : Objectifs de notes d'état corporel au cours de la croissance de la génisse laitière [12].
|
Objectif vêlage Age |
Etat d'engraissement |
Etat d'engraissement génisses Normandes |
|
|
24-27 mois |
24-30 mois |
33 mois |
|
|
6 mois |
2,8 |
2,8 |
2,7 |
|
12 mois |
3,0 |
3,0 |
2,9 |
|
15 mois |
3,1 |
3,1 |
3,0 |
|
18 mois |
3,3 |
3,2 |
3,1 |
d) Pendant la première année
Il convient donc de maintenir la note d'état autour de 3 de la puberté jusqu'à la mise à la reproduction.
Les études conduites dans les élevages français pendant l'hiver ont montré que pour maintenir une note d'état corporel constante chez des animaux de plus d'un an le GMQ doit être de 570 g/j en race Prim Holstein, et de 650 g/j en race Normande [12].
e) Jusqu'au vêlage
Après la puberté, une vitesse de croissance élevée n'a plus de répercussion négative sur le développement du tissu mammaire. L'objectif de croissance est alors de 850 à 900 g/j. Une vitesse de croissance trop élevée peut conduire à un état d'engraissement excessif au vêlage, l'optimum de situant vers 3 à 3,5 et les conséquences des excès étant identiques à celles que l'on observe chez les adultes : moindre appétit post-partum entraînant un risque accru de cétose et syndrome de la vache grasse. Une vitesse de croissance trop lente peut limiter le développement
corporel de la génisse sans affecter celui du veau. Il en résulte alors un risque élevé de vêlage difficile, mais aussi une moindre production laitière par défaut de réserves énergétiques [2].
L'état corporel varie significativement en fonction du moment dans le cycle de production [25]. 1) Note d'état corporel au tarissement
Il est intéressant de commencer par le tarissement dans la mesure où la note d'état corporel devrait rester stable pendant cette période. Le tarissement est une période stratégique et déterminante quant à l'avenir nutritionnel de l'animal et du troupeau.
La note d'état corporel au tarissement est donc celle attendue au vêlage [43]. L'objectif retenu de note d'état au tarissement est situé entre 3 et 3,5 sur une échelle de 0 à 5 (tableau 4). Les variations d'état corporel au tarissement, que ce soit amaigrissement ou reprise d'état, supérieures à un point sont sources de problèmes [14]. Il est d'ailleurs intéressant, lors d'une visite d'élevage, d'analyser deux lots, celui des vaches taries récemment et celui des vaches en fin de tarissement, prêtes à vêler, pour évaluer l'efficacité de la gestion alimentaire au tarissement [48].
Tableau 4 : Recommandations pour la note d'état au tarissement.
|
Référence |
Recommandation |
Echelle utilisée |
Remarque |
|
[44] |
Entre 3,0 et 3,5 |
Notes entre 0 et 5 |
|
|
[3] |
Entre 3,0 et 3,5 |
Notes entre 0 et 5 |
|
|
[67] |
Entre 3,0 et 3,5 |
Notes entre 0 et 5 |
>3,5=tarissement retardé à 5 semaines antepartum |
a) Recommandations usuelles
Les recommandations quant à la note d'état au vêlage sont généralement comprises entre 3 et 4 sur une échelle de allant de 0 à 5 [31, 34, 44, 67, 86], l'idéal étant une note de 3,5 [67] (tableau 5). Pourtant les observations de terrain, en race Prim Holstein, ne confirment pas ces recommandations. Quarante pour cent des bovins seraient en dessous de la note 3 au vêlage et 33 % au dessus de 3,5 ; il ne resterait alors que 27 % des animaux entre 3 et 3,5 [41].
Tableau 5 : Recommandations pour la note d'état corporel au vêlage
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|||
|
|
|
|||
|
|
|
|||
|
|
||||
b) Application à différents systèmes d'élevage
L'objectif de note d'état corporel au vêlage est variable en fonction du type de système de production. De nombreux éléments sont à concilier : une pleine expression du potentiel laitier, des conditions de vêlage faciles, des résultats de reproduction corrects (un retour de cyclicité normal, fertilité, fécondité), éviter des maladies métaboliques...mais il est difficile de trouver le juste équilibre ; équilibre qui n'est d'ailleurs pas le même d'un élevage à l'autre.
Trois types de système peuvent être décrits : le premier favorise la productivité laitière. Les élevages où elle dépasse les 8000 kg/vache laitière/an peuvent choisir d'augmenter l'intervalle vêlage/vêlage (IVV) à 14 mois dans le but d'exprimer pleinement le potentiel laitier. La conduite de reproduction n'est alors plus la même, on accepte une fertilité moindre et la stratégie de réforme devient moins stricte.
Dans le second système, l'objectif est de réduire les coûts de production, notamment alimentaires. Un IVV élevé n'est alors pas souhaitable et peut être raisonnablement contraignant et rentable autour de 13 mois. La conduite de reproduction est basée sur une mise à la reproduction précoce, dès 50 jours, avec une surveillance attentive des différents évènements (détection des chaleurs, interventions sur des vaches non détectées).
Dans le dernier type d'élevage, l'objectif est le regroupement des vêlages. La difficulté est, dans ce système, de garder un IVV de 12 mois avec un taux de réforme faible. La maîtrise de la reproduction devient ici prioritaire, les vaches doivent être cyclées rapidement, et fécondées dans un court intervalle, celles qui ne répondent pas aux contraintes étant alors réformées.
Les objectifs de note d'état en fonction du système de production désiré sont présentés dans le tableau 6.
Tableau 6 : Objectifs d'état corporel au vêlage en fonction des systèmes d'élevage [21].
|
Critères/Systèmes |
Productivité élevée |
Coûts réduits |
Vêlages groupés |
|
NEC multipares |
3,5 à 4 |
3 à 3,5 |
Automne 3 à 3,5 |
|
Printemps 2,5 à 3 |
|||
|
NEC primipares |
3 à 3,5 |
3 |
3 |
La perte d'état corporel en début de lactation est significativement proportionnelle à l'état d'engraissement au vêlage [86].
a) Appétit des vaches
La quantité de matière sèche ingérée en début de lactation diminue en fonction de l'état corporel au vêlage [11]. La relation est proportionnelle pour un état corporel situé entre 1,6 et 3,8 au vêlage (échelle 1 à 5). La différence de matière sèche ingérée par jour est de 1,3 kg entre deux groupes de vache ayant un écart de note d'état d'un point : une vache ayant une note de 3,5 au vêlage consomme 1,3 kg de moins par jour qu'une vache ayant une note de 2,5.
Une vache avec des réserves peut mobiliser 40 à 50 kg de réserves adipeuses ce qui représente 400 à 500 litres de lait. En revanche, une vache maigre mobilise trois à quatre fois moins mais son appétit est supérieur [34].
La mobilisation des réserves doit être raisonnable. Les excès de mobilisation sont néfastes. Plusieurs origines peuvent être répertoriées :
- soit c'est la vache elle-même qui est en cause : les vaches à haut potentiel n'ont pas un appétit plus élevé, ce qui conduit à un déficit énergétique plus élevé et à un excès de mobilisation.
- soit ce sont les apports qui sont insuffisants. C'est alors soit la ration qui est en cause, soit l'appétit des vaches qui est déprécié. L'appétit des vaches peut être déprécié par une maladie concomitante (mammite, métrite, maladie métabolique), par un état d'engraissement exagéré (la mobilisation est d'autant plus importante que cet état a été acquis précocement au tarissement) ou par une transition alimentaire mal conduite et qui ne laisse pas aux papilles ruminales le temps de se développer ni à la flore le temps de s'adapter à la nouvelle ration avec comme conséquence une ration mal valorisée voire une évolution vers l'acidose ruminale [34].
Notons que l'appétit des vaches et leur capacité d'absorption digestive sont liées au développement des papilles du rumen. Leur dimension entre le tarissement et le 3ième mois de lactation (en moyenne) double. Leur capacité d'absorption des acides gras volatils (AGV) triple et la vitesse d'absorption ruminale quintuple alors [67].
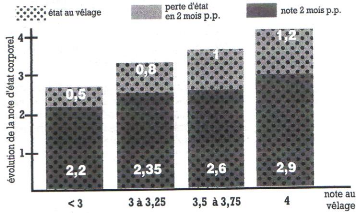
Finalement, l'évolution de l'état corporel, certes cyclique, n'est pas aléatoire ; chaque étape conditionne la suivante (figure 11).
Figure 11 : Relation entre état corporel au vêlage et perte d'état en début de lactation. D'après [67].
L'échelle utilisée compte 5 points.
b) Evolution de la proportion de vaches grasses/vaches maigres
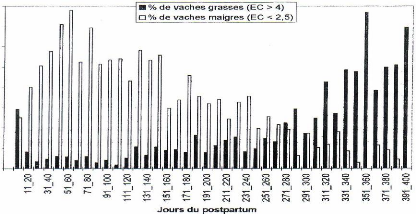
En accord avec ce que nous avons vu concernant l'appétit, la proportion de vaches grasses diminue au cours du post-partum. La distribution du pourcentage des vaches grasses a une répartition inverse à celle des vaches maigres. Dans l'étude de Drame et al. [25], la proportion de vaches maigres passe de 12 à 39% entre le vêlage et le 60ième jour de lactation. Cette proportion diminue voire devient nulle entre le 300ième et le 400ième jour post-partum (figure 12).
Figure 12 : Fréquence des vaches maigres/vaches grasses au cours du post-partum [25].
c) Objectifs d'évolution de l'état corporel au cours du postpartum.
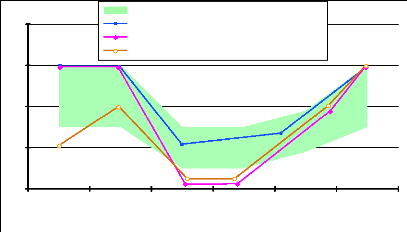
L'état corporel des vaches laitières subit donc une chute au cours des deux voire trois premiers mois de lactation. Elle est inévitable mais doit être maîtrisée et compensée lors de la deuxième période de lactation. Cette perte a été observée sur le terrain. Les observations confirment également qu'elle est d'autant plus élevée que les vaches sont grasses au vêlage. Les pertes sont mesurées à 0,6 unité par point d'état corporel au vêlage [11]. Sur une échelle de 1 à 5, cette perte s'élève à 1,4 point pour les vaches grasses (note d'état au vêlage =4), à 0,5 point pour les vaches normales (note d'état au vêlage comprise entre 2,5 et 3,5) et à 0,05 point pour les vaches maigres (note au vêlage = 2) [25]. Dans l'étude de Fréret et al. [41], 35 % des Prim Holstein ont une perte d'état entre 0 et 60 jours post-partum inférieure à 1 point, 35% perdent entre 1 et 1,5 point et enfin, 30 % d'entre elles perdent plus qu'1,5 point. Les recommandations quant à la perte d'état en post-partum sont présentées dans le tableau 7 et la figure 13. Si les observations de terrain ne sont pas toujours en accord avec celles-ci, il semble néanmoins admis qu'une perte d'état corporel supérieure à un point sur une échelle de 0 à 5 semble préjudiciable et inquiétante.
4,5
2,5
3,5
4
3
Période
sèche
Vêlage Pic Fécond. Fin de lactation
Objectif de moyenne de troupeau Objectif individuel classique
haute production
état médiocre
Tableau 7 : Recommandations pour les pertes d'état corporel en postpartum
|
Référence |
Recommandation |
Echelle utilisée |
||
|
[30, |
33, 34, |
44] |
Moins d'un point |
Echelle de 0 à 5 |
|
[67] |
0,5 à 0,7 point |
Echelle de 0 à 5 |
||
Figure 13 : Evolution de l'état corporel à l'échelle individuelle ou de troupeau et objectifs [28].
L'étude et l'utilisation de l'état corporel pour la conduite d'élevage a plusieurs intérêts. S'il permet de juger évidemment de la conduite nutritionnelle du troupeau, il est alors intéressant de relier l'évolution de ce facteur à deux composantes de l'élevage : la production laitière d'abord aussi bien qualitativement que quantitativement et les résultats de reproduction ensuite.
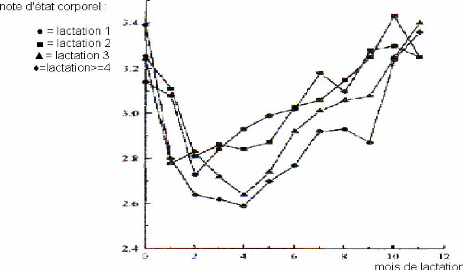
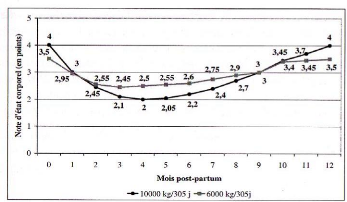
Note d'état corporel et production laitière élevée en début de lactation sont corrélées négativement (figure 14). Nous avons déjà évoqué la raison principale qui est l'appétit des vaches. Rappelons qu'il est indépendant du niveau de production et que le déficit énergétique est supérieur chez les hautes productrices. En conséquence, les vaches les plus hautes productrices ont des notes d'état plus basses [80], ou perdent plus d'état corporel [51, 57], tandis que les vaches moins bonnes productrices peuvent même gagner de l'état en début de lactation [43].
Pourtant, une étude met en évidence une relation favorable entre la production laitière (surtout le taux de matières grasses) et l'augmentation de la note d'état corporel, cet effet se faisant ressentir jusqu'à cent jours de lactation [44]. Il s'agit là bien plus d'un effet de la lipomobilisation permise par un état d'engraissement important induit expérimentalement dans cette étude. En effet, cette donnée est extraite d'un groupe de vaches suralimentées au tarissement de façon à accroître leur taux d'engraissement et leur poids. La mobilisation de ces réserves est accompagnée d'un taux d'AGNE (acides gras non estérifiés) sanguins élevés, AGNE captés par la mamelle, matières premières du taux butyreux du lait (TB).
Il existe donc une relation significative entre le profil d'état et la production au pic. La probabilité d'observer des profils de perte d'état élevée ou d'état insuffisant est diminuée pour une production au pic moyenne par rapport à une production élevée associée à un pic tardif [76].
Figure 14 : Evolution de la note d'état corporel en fonction du niveau de production [78].
Les profils d'état insuffisant sont fréquemment associés à des taux protéiques (TP) au pic inférieurs à 28 g/kg [76] en race Prim Holstein. La moitié des vaches présentant un TP au pic inférieur à 28 g/kg sont dans la classe "état insuffisant" alors que pour celles ayant un TP supérieur à 28 g/kg sont à 32,3% dans la classe "reprise d'état précoce".
Il existe une liaison hautement significative entre le TP et la note d'état [34]. Le principal facteur conditionnant le TP est l'énergie : les apports énergétiques permettent la synthèse de matières protéiques par la mamelle.
L'évaluation du déficit énergétique en début de lactation passe traditionnellement par la notation de l'état corporel mais la notion de TP mini (TP le plus bas enregistré au cours des trois premiers contrôles) est de plus en plus retenue dans ce but [65].
L'évolution de la note d'état corporel au cours du post-partum doit amener à adapter la production laitière en fonction de la perte d'état de l'animal. En effet, si production laitière et note d'état sont corrélés négativement, il en est de même pour la perte d'état en post-partum et les résultats de reproduction. Il apparaît donc nécessaire de faire varier la production laitière pour limiter la perte d'état en post-partum et pour atteindre les objectifs de note d'état au tarissement.
La diminution de la production laitière peut être menée de plusieurs façons, mais les conséquences sont variables. Il est plus avantageux de faire varier le niveau azoté, car si la réduction des apports énergétiques entraîne une perte d'état corporel autant qu'un système restrictif en apport azoté, le second n'entraîne pas de dégradation des performances de reproduction [21]. Là encore plusieurs possibilités quant au niveau azoté de la ration sont à choisir selon le type d'élevage adopté, ils sont présentés dans le tableau 8.
Dans le cas d'un système privilégiant la production laitière, le niveau azoté ne devra pas être trop élevé afin de faire durer la production laitière. L'état d'engraissement doit être suffisant pour exprimer pleinement le potentiel laitier mais l'amaigrissement ne doit pas être trop rapide ni trop intense.
Dans le cas d'un système visant à réduire les coûts, le niveau azoté et l'état d'engraissement doivent être encore plus modérés afin d'éviter des démarrages de lactation trop rapides. Le système privilégie la durée : capacité d'ingestion élevée, sélection sur index fonctionnels de longévité.
Dans le cas du système à groupement de vêlages, les recommandations sont sensiblement les mêmes : il faut éviter des démarrages de lactation trop rapides et permettre une courbe de lactation sans pic, tout doit être mis en oeuvre pour obtenir une fertilité optimisée dans le temps imparti. Il faudra aussi veiller à la croissance des génisses pour assurer d'atteindre 60% du poids adulte à la première IA pour un vêlage à vingt-quatre mois.
Tableau 8 : Recommandations sur le niveau azoté en fonction des objectifs d'élevage [21]
Le niveau azoté est exprimé en gramme de PDI (protéines digestibles dans l'intestin) par kilogramme de matière sèche ; NEC=note d'état corporel
|
Critères/Systèmes |
Productivité élevée |
Coûts réduits |
Vêlages groupés |
|
Niveau azoté |
100 à 105 |
90 à 95 |
90 à 95 |
La notation de l'état corporel s'est développée ces trente dernières années, s'avérant un outil nécessaire pour juger la gestion alimentaire et nutritionnelle du troupeau. Les objectifs d'état corporel pour différents moments du cycle de production de la vache sont validés dans le but de réduire l'incidence des maladies métaboliques et pour assurer la quantité et la qualité de la
production laitière mais aussi pour le rôle capital des variations de note d'état corporel sur les performances de reproduction.
DEUXIEME PARTIE : RELATION ENTRE NOTE
|
La dégradation des performances de reproduction en élevage laitier est devenue la source de développement de nouvelles techniques permettant d'en appréhender les facteurs de risque. Outre la baisse de la fertilité et l'allongement de la fécondité, les anomalies de cyclicité sont aujourd'hui très fréquentes puisqu'elles peuvent concerner de 30 à 50 % des vaches [76]. L'implication de la note d'état corporel et surtout de ses variations en post-partum est largement étudiée. Dans un premier temps, il est important de traiter de cet impact sur la reprise de cyclicité et sur les différents profils de progestérone associés puis, dans un second temps sur les paramètres de fertilité et de fécondité. Enfin, l'influence de l'état corporel sur la réussite du péripartum nécessite une attention particulière. La fertilité des vaches étant d'abord conditionnée par le retour à l'intégrité anatomo-histologique de l'utérus, la maîtrise des évènements accompagnant les six premières semaines post-partum (mise bas, délivrance, métrite etc) est déterminante.
La période immédiate après le vêlage est suivie d'une inactivité ovarienne chez la vache laitière comme chez la vache allaitante [46], au sens d'une absence d'ovulation.
Les mécanismes qui conduisent au rétablissement de l'activité sexuelle chez la vache sont relativement bien connus. Avant le vêlage, les taux élevés d'oestrogènes foetaux et de progestérone maternelle inhibent la sécrétion de LH et de FSH par l'axe hypothalamohypophysaire réduisant l'activité ovarienne [46].
Les concentrations en FSH augmentent en cinq à dix jours après le vêlage, celles de LH commencent à augmenter dix à vingt jours après le part, avec une sensibilité accrue à l'hormone hypothalamique GnRH ou gonadolibérine (Gonadotrophin Releasing Hormone). Mais la faible fréquence des pulses de LH provoque une faible production d'androgènes dans les cellules thécales du follicule. Ces androgènes étant les précurseurs de la synthèse d'oestrogènes par les cellules de la granulosa, l'insuffisance en oestrogènes conduit ces follicules à l'atrésie.
Le facteur crucial déterminant le moment où l'ovulation peut avoir lieu est l'obtention de pulses de LH suffisants : une fréquence de décharge toutes les trois ou quatre heures aboutit à la perte de dominance (cf infra) du follicule et donc à l'absence d'ovulation ; si la fréquence de décharge est d'un pic par heure, l'ovulation peut avoir lieu [35]. Ces pulses sont conditionnés d'une part par la progestérone, principal agent inhibiteur de ces décharges durant la phase lutéinique et d'autre part par les oestrogènes qui, en quantité suffisante, exercent un rétrocontrôle positif sur l'axe hypothalamo-hypophysaire.
L'intervalle vêlage/première ovulation est assez court chez la vache laitière. Il est compris entre quinze et trente jours mais sa variabilité est élevée (écart type de quinze à vingt-cinq jours) [46, 54].
La croissance folliculaire qui conduit un follicule primordial en follicule pré ovulatoire dure environ cinq mois. Les dix derniers jours de cette croissance s'effectuent sous forme de vague. Chaque vague comprend trois phases : recrutement, sélection, dominance. Lors de la première phase, une dizaine de follicules émerge d'un groupe de follicules tertiaires, passent de 2 à 3 mm à 5 à 6 mm et deviennent dépendant à l'hormone folliculostimulante (FSH). Deux ou trois follicules sont alors sélectionnés de cette cohorte initiale et commencent à sécréter des oestrogènes et de l'inhibine. Cette sécrétion exerce un rétrocontrôle négatif sur l'hypophyse, abaissant la sécrétion de FSH à un niveau inférieur aux besoins folliculaires. Seul un follicule, le follicule dominant, va pouvoir se développer en présence de petites quantités de FSH. Il acquiert alors des récepteurs à l'hormone lutéinisante : la LH. Sa taille est alors de 10 mm et sa croissance devient LH dépendante pour atteindre 15 à 20 mm. La suite de sa croissance conduit ou non à l'ovulation selon la présence d'un corps jaune. Si tel est le cas, le rétro-contrôle négatif exercé par la progestérone empêche le pic pré ovulatoire de LH. Le follicule s'atrésie après quelques jours d'attente. S'il devient dominant après la lutéolyse, il ira jusqu'à l'ovulation (figure 15 et 16).
On peut détecter un follicule de plus de 10 mm autour de dix à quatorze jours post-partum chez la vache laitière. Cet intervalle semble plus long au printemps (20 jours) qu'en hiver (7 jours) [35]. Ce premier follicule ovule dans 75% des cas, devient kystique dans 20% des cas ou s'atrésie pour les 5% restant [46]. La première ovulation post-partum a lieu vers 14 à 25 jours en moyenne mais dans deux cas sur trois, elle ne s'accompagne pas de manifestations visibles de chaleurs [35].
Figure 15 : Reprise du développement folliculaire chez la vache laitière en post-partum [35].
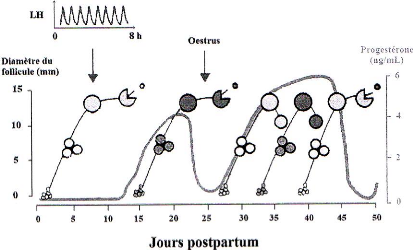
Les femelles bovines présentent de deux à quatre vagues, exceptionnellement jusqu'à six. En majorité ont lieu des cycles à deux ou trois vagues. Lors de cycle à trois vagues, une nouvelle vague émerge à J1, J6 et J16 ; lors de cycle à deux vagues, à J1 et J9 ou J10 [16].
Figure 16 : Les vagues folliculaires chez la vache [16]
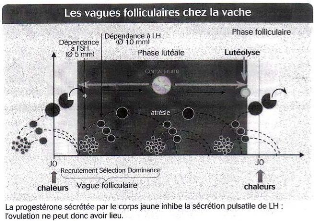
La phase folliculaire dure en moyenne de six à sept jours, la phase lutéale de quatorze à quinze jours.
Tableau 9 : Bilan de la reprise d'activité sexuelle en post-partum (pp). D'après [35].
|
Apparition du 1er follicule dominant |
Première ovulation |
Durée premier cycle |
Devenir 1er follicule |
Chaleurs |
|
10 à 12 jours pp |
10 à 20 jours pp |
8 à 12 jours |
- 70 à 80% ovulation et formation d'un corps jaune - 20 % kyste - 5% atrésie |
Pour 1/3 des premières ovulations |
La succession de vagues aboutit à l'ovulation et à la mise en place d'un corps jaune. La poursuite de la reprise de cyclicité nécessite l'élimination de celui-ci lorsqu'il n'y a pas fécondation.
Le premier corps jaune cyclique apparaît en moyenne à 28 jours pp et 85% des vaches en présentent un avant 50 jours post-partum [20].
L'évolution du corps jaune de la vache se réalise en trois phases :
- une période de croissance de 4 à 5 jours au cours de laquelle il est insensible à l'action des prostaglandines,
- un temps de maintien d'activité pendant 8 à 10 jours,
- enfin, s'il n'y a pas eu de fécondation, une période de lutéolyse d'abord brutale puis plus progressive en 24 à 48 heures.
Du point de vue histologique, le corps jaune est formé de deux types cellulaires. Les cellules de la thèque vont donner une lignée de petites cellules lutéales et celle de la granulosa de grandes cellules lutéales. Pendant la phase lutéale, les cellules formant le corps jaune sécrètent essentiellement de la progestérone.
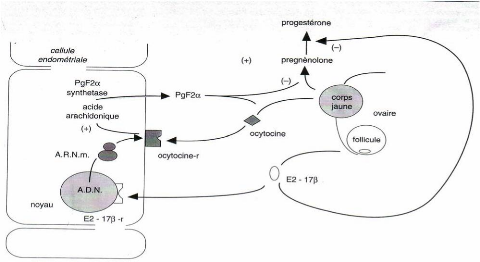
D'un point de vue biologique, la lutéolyse se réalise selon plusieurs modalités (figure 17) :
- indirectement par l'intermédiaire de l'ocytocine, la PGF2á d'origine utérine entraîne une vasoconstriction provoquant une ischémie du corps jaune.
- directement, d'une part la PGF2á se fixe sur des récepteurs spécifiques présents en surface du corps jaune et diminue l'action lutéotrope de la LH par blocage de l'activité de l'adényl-cyclase et inhibition de l'AMP (adénosyl mono-phosphate), d'autre part, la PGF2á exercerait ses effets sur la cellule lutéale par l'intermédiaire d'une augmentation du taux de calcium intracellulaire et par l'activation d'une protéine kinase C [39].
La lutéolyse résulte d'une boucle d'interaction entre l'utérus et l'ovaire. Au moment de la lutéolyse, l'étude des variations hormonales montre une augmentation du taux d'oestrogènes suivie par une libération d'ocytocine puis de PGF2á. En fin de phase lutéale, les oestrogènes folliculaires induisent la synthèse de récepteurs à l'ocytocine au niveau utérin [35, 39]. A cette période, les grandes cellules lutéales ne synthétisent plus de la progestérone mais de l'ocytocine. Celle-ci va se fixer sur les récepteurs utérins initiant un pulse de sécrétion de prostaglandines qui exercent alors une double action sur le corps jaune : d'une part, l'inhibition de la synthèse de progestérone, d'autre part, le déclenchement d'un pulse important d'ocytocine d'où une série d'interactions entre utérus et corps jaune qui aboutissent à la lutéolyse [39].
Figure 17 : Déterminisme de la lutéolyse chez les ruminants. D'après [39]
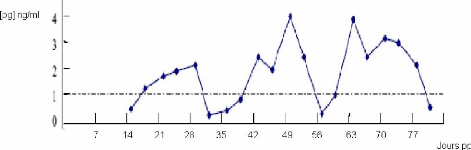
La réalisation de dosages fréquents (deux ou trois fois par semaine dans le lait) de progestérone permet de suivre le profil de rétablissement de l'activité sexuelle cyclique après vêlage (figure 18) [46].
Figure 18 : Profil de progestérone normal, avec chaleurs régulières [91].
Cinquante à soixante-dix pour cent des vaches laitières présentent une reprise de cyclicité postpartum (tableau 10). Chez la vache laitière, cinq grands types d'anomalies de reprise de cyclicité ont été identifiés [46].
Tableau 10 : Fréquence des anomalies de reprise d'activité sexuelle après vêlage chez la vache laitière.
|
Référence |
[41] |
[97] |
[90] |
[54] |
[71] |
[95] |
[70] |
|
|
N |
275 |
91 |
110 |
102 |
448 |
37 p |
125 |
97 |
|
Nombre de |
10 |
1 |
2 |
6 |
1 |
2 |
4 |
|
|
Niveau de |
>7500 |
8350 |
6941 |
7417 |
9300/ |
6600/ |
||
|
Profil |
73,45 |
70,3 |
49 |
59 |
53,5 |
45 |
63,4 |
63,9 |
|
Inactivité |
7,3 |
15,4 |
14 |
11 |
20,5 |
24 |
25,2 |
22,7 |
|
Phase |
NC |
NC |
5 |
7 |
0,5 |
|||
|
Cessation |
4,0 |
1,1 |
5 |
7 |
3,0 |
5 |
10,6 |
10,3 |
|
Phase |
9,45 |
13,2 |
37 |
15 |
20,0 |
26,3 |
0,8 |
0,3 |
|
Profil |
5,8 |
NC |
NC |
1 |
2,5 |
|||
|
Pays |
France |
France |
Japon |
France |
Belgique |
RU |
Japon |
|
n : nombre de profils ; p : primipares ; RU : Royaume-Uni ; VL : Vache laitière ; NC : non communiqué
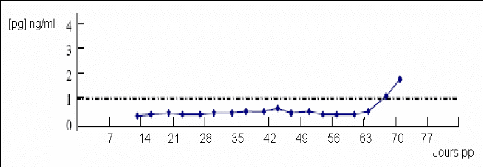
La reprise d'activité différée se caractérise par un premier signe d'activité lutéale postérieur à 45 à 50 jours après le vêlage (figure 19). Une vache est considérée en activité lutéale dès lors qu'elle présente au moins deux dosages de progestérone (pris deux fois par semaine) dans le lait supérieur ou égal à 1 ng/ml [91] ou un dosage supérieur à 5ng/ml [54]. Cette inactivité représenterait 10 à 20% des animaux. L'inactivité ovarienne prolongée touche essentiellement les primipares, notamment celles qui vêlent avec un état corporel inférieur à 2,5. Les difficultés de vêlage, les non délivrances, les mauvaises involutions utérines et les métrites perturbent la reprise de la cyclicité post-partum et sont autant de facteurs de risque connus d'inactivité ovarienne [21].
Figure 19 : Profil de progestérone lors d'absence d'activité ovarienne entre 30 et 50 jours [91].
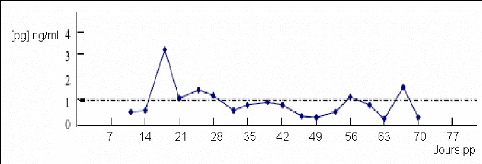
On remarque une interruption de la sécrétion de progestérone pendant 12 à 14 jours (figure 20). L'interruption de cyclicité est plus rare et touche 1 à 13% des animaux.
Figure 20 : Profils de progestérone correspondant à une interruption de cyclicité [41].
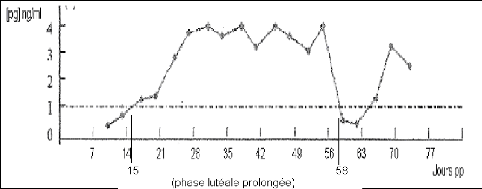
On parle également de corps jaune persistant ; la sécrétion de progestérone a lieu pendant plus de 19 à 28 jours au lieu de 16 à 17 jours physiologiquement (figure 21). Elle représente 12 à 35% des profils post-partum. Le corps jaune qui persiste suit le plus souvent une première ovulation précoce et peut sécréter de la progestérone très au-delà de cinquante jours de lactation [21]. En plus des ovulations précoces, les pertes de poids excessives et une haute production sont des facteurs de risque de cette anomalie de cyclicité.
Figure 21 : Profils de progestérone correspondant à une phase lutéale prolongée [91].
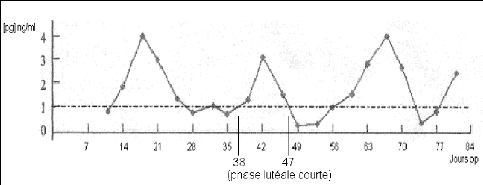
La sécrétion de progestérone a lieu pendant moins de 10 jours (figure 22). C'est un cas plus rare (moins de 5% des cas) et qui est jugé normal quand il intervient après la première ovulation [46].
Figure 22 : Profil de progestérone correspondant à une phase lutéale courte [91].
Cette catégorie comprend des profils de progestérone non classables dans les catégories précédentes.
Les kystes ovariens sont considérés comme une cause majeure d'infertilité. Cependant 50% des kystes diagnostiqués disparaissent spontanément et ne perturbent pas la cyclicité. De plus, les kystes ne concernent que 10 à 15% des vaches laitières qui présentent des troubles de la reproduction.
On définit le kyste ovarien comme une structure de type
folliculaire dont la taille est supérieure à
25 mm et qui
persiste plus de dix jours. Il est également possible que des structures
de plus petite
taille, mais qui persistent dans le temps en soient également, mais leur diagnostic en pratique est difficile.
Deux types de kystes sont distingués : le kyste folliculaire dont les parois sont fines et qui sécrète rarement de la progestérone ; et le kyste lutéal qui possède des parois épaisses et qui est associé à une production variable de progestérone. Les kystes folliculaires s'accompagnent de quatre dominantes comportementales : comportement normal, irrégularité et al.longement des cycles, anoestrus, nymphomanie [94]. Les kystes lutéaux s'accompagnent exclusivement d'anoestrus.
Le suivi de kystes marqués montre que dans 13% des cas seulement, le kyste persiste réellement mais dans 87% des cas, il est remplacé par un follicule qui ovule (sept cas sur vingt) ou qui est suivi du développement d'un autre kyste sur l'un ou l'autre des ovaires. Le kyste n'ovule en aucun cas [68].
Notons que la présence de kyste peut être responsable de l'apparition d'anomalie de cyclicité. En effet, des hypothèses mettent en cause le rôle de kystes folliculaires dans le cas de reprise d'activité différée ou de kystes lutéaux lors de phase lutéale prolongée (mais le corps jaune persistant en reste le principal responsable).
b) Effet de l'alimentation
L'alimentation fait là encore partie des facteurs de risque contribuant à l'apparition de kystes sans qu'en soit précisé le type. Si l'insuffisance d'apport énergétique ou l'excès d'azote non protéique en période post-partum sont souvent évoqués, aucun effet de l'état corporel au tarissement, au vêlage, à trente jours post-partum, ni de la perte d'état entre le vêlage et trente jours post-partum n'a été mis en évidence [59, 68]. En revanche, la note d'état au tarissement [43], et bien plus encore, le gain d'un point de NEC pendant le tarissement est associé à un risque de kyste 8,4 fois plus élevé [68]. D'une manière générale, l'état d'embonpoint est un facteur de risque [43].
c) Physiopathogénie
L'apparition de kyste est due à un défaut d'ovulation ou d'atrésie du follicule dominant. La fréquence des pulses de LH peut être insuffisante pour provoquer l'ovulation, mais suffisante pour permettre le maintien de la croissance du follicule dominant et de la production d'oestrogènes. Le déficit énergétique est alors mis en cause car il est à l'origine d'une diminution
de la synthèse de LH. De la même manière, le gain d'état corporel durant la période sèche révèle une "sur-alimentation" et une accumulation de gras qui sera mobilisé en post-partum. L'augmentation de la concentration sanguine en acides gras non estérifiés est alors dans ce cas fortement suspectée comme facteur de risque de kystes ovariens [59].
1) Chez les génisses
Une reprise d'activité ovarienne retardée est associée à des états corporels insuffisants au moment du vêlage. Cette situation est rencontrée lorsque les apports alimentaires dans le dernier tiers de gestation sont insuffisants (génisses au pré sans complémentation avec vêlage d'automne). A cette période, le GMQ doit être au moins de 500 g/j pour assurer les besoins de la gestation [36].
2) Chez les multipares
Il semble difficile d'établir une relation entre le profil de cyclicité et la note d'état au vêlage. A l'inverse, la perte d'état en post partum est un facteur de risque mis en évidence dans plusieurs études. Fréret et al. [41] trouvent une proportion de vaches ayant une perte supérieure à 1,5 entre 0 et 60 jours significativement différente selon les profils. On les retrouve en majorité dans les profils d'absence de cyclicité ou de phase lutéale prolongée. Cinquante pour cent des femelles ayant eu une note minimale inférieure ou égale à 1,5 présentent ce type de cycle (47,4% de celles-ci sont dans la catégorie "absence de cyclicité" et 41,7% dans la catégorie "phase lutéale prolongée"). Même si Touze et al. [97] trouvent un poids vif au vêlage significativement plus faible chez les vaches en inactivité ovarienne prolongée que chez les vaches à profil normal, ils ne mettent pour autant pas en évidence un rôle déterminant de la note d'état.
Opsomer et al. [72], dans une étude sur les facteurs de risque des dysfonctionnements ovariens en post-partum, mettent en évidence la perte d'état corporel à trente jours, soixante jours, et cent vingt jours après vêlage comme facteurs de risque significativement plus important de l'inactivité ovarienne prolongée comparativement aux vaches à profil normal et à phases lutéales prolongées. La perte d'état à trente et soixante jours multiplie par 18,7 et 10,9 fois (odds ratio) le risque de manifester une inactivité ovarienne prolongée plutôt qu'un profil normal. La perte s'élève à 0,26 et 0,29 points au premier et au second mois respectivement pour les vaches
normales et à 0,39 et 0,49 points respectivement pour les autres. Les auteurs rapportent que l'importance de la note d'état au vêlage et d'une bien moindre importance comparativement à celle de la perte d'état pendant les premiers mois de lactation.
Shrestha et al. [90] placent l'état corporel comme le paramètre le plus impliqué dans la reprise d'activité ovarienne. Ils trouvent une note d'état corporel à cinq, sept, neuf et onze semaines post-partum significativement plus faible pour les vaches présentant une inactivité ovarienne prolongée. La perte d'état supérieure ou égale à un point apparaît aussi comme un facteur de risque de retard d'activité ovarienne en général. Mais la note d'état dès cinq semaines postpartum est aussi significativement plus faible pour l'inactivité ovarienne prolongée que pour les cycles normaux ou les phases lutéales prolongées. Il s'en suit donc un retard d'ovulation plus important pour les vaches présentant une perte d'état corporel modérée (0,5 à 1 unité) ou sévère (>1 point) comparativement à celle perdant peu (<0,5 point) [14, 44].
Dans l'étude de Benaich et al. [8], il existe une corrélation positive entre la durée de l'intervalle vêlage reprise d'activité ovarienne et le degré de mobilisation des réserves corporelles. Il s'agit d'une étude ayant pour objectif d'évaluer l'effet de la mobilisation des réserves sur les paramètres de reproduction dans huit fermes (216 animaux) au Maroc. La ferme où les vaches ont le moins mobilisé leurs réserves corporelles a les paramètres de reproduction les meilleurs et inversement dans la ferme où les vaches ont le plus maigri.
L'étude de Disenhaus et al. [22] traite de la cyclicité en fonction de la production laitière. Les profils de cyclicité post-partum sont classés différemment. Les vaches sont classées en 3 catégories : cyclicité normale, cyclicité anormale et anoestrus selon leurs profils de progestérone (vu précédemment). La probabilité (le seuil de significativité ayant été fixé à 10 % par les auteurs) de présenter une anomalie de cyclicité est augmentée par l'augmentation de la production laitière corrigée pendant les trois premières semaines (p=0,06), par la matière sèche volontairement ingérée au cours des trois premières semaines post-partum (p=0,03), par l'état d'engraissement au vêlage (p=0,06) et par la perte d'état entre le vêlage et soixante jours postpartum (p=0 ,06). C'est d'ailleurs l'interaction entre ces deux derniers paramètres qui a eu un effet négatif (p=0,06). En ce qui concerne l'anoestrus, le bilan énergétique des semaines sept à dix en est un facteur de risque (p=0,07), tout comme les AGNE plasmatiques moyens des semaines quatre à six et sept à dix (p=0,07 et p=0,04 respectivement) et la glycémie minimale aux mêmes périodes (p=0,08). En dehors de ces paramètres métaboliques, on retrouve la
production laitière corrigée des semaines sept à dix (p=0,02), la perte d'état entre le vêlage et 60 jours post-partum et le poids moyen des vaches entre la semaine quatre et six (p=0,05) (tableau 11).
Tableau 11 : Comparaison des caractéristiques zootechniques et métaboliques de vaches laitières selon leur profil de cyclicité post-partum [22].
PL : production laitière ; AGNE : acides gras non estérifiés ; MSI : matière sèche ingérée ; pp : post-partum
|
Etude 1 : anormale vs normale |
Etude 2 : anoestrus vs normale |
|||
|
Cyclicité |
Anormale n=15 |
Normale n=35 |
Anoestrus n=14 |
Normale n=35 |
|
Bilan énergétique (UFL/j) |
1,90#177;0,49 |
0,78#177;52 |
||
|
PL 4% moyenne (kg/j) |
33,3#177;0,9 |
29,8#177;0,9 |
||
|
PL 4% moyenne (kg/j) |
27,0#177;1,1 |
23,1#177;1,1 |
||
|
MSI (kg/j) |
16,1#177;0,7 |
14,2#177;0,6 |
||
|
Poids moyen (kg) |
559#177;14 |
579#177;13 |
||
|
Etat d'engraissement au |
3,27#177;0,11 |
2,78#177;0,13 |
2,9#177;0,1 |
2,9#177;0,1 |
|
Perte d'état |
0,84#177;0,13 |
0,55#177;0,12 |
0,79#177;0,11 |
0,41#177;0,11 |
|
AGNE max (microM) 1 à |
775#177;85 |
629#177;74 |
||
|
AGNE moyen |
314#177;40 |
229#177;37 |
||
|
Glycémie moyenne (g/L) |
0,664#177;0,018 |
0,695#177;0,015 |
||
|
Glycémie minimale (g/L) |
0,583#177;0,017 |
0,653#177;0,016 |
||
Finalement, l'évolution de l'état corporel
influence significativement la reprise et la régularité de
la
cyclicité entre 30 et 80 jours post-partum. Il convient de
considérer la perte d'état d'un côté et
la note
en elle-même de l'autre. Le risque de présenter une phase
lutéale prolongée ou une
inactivité ovarienne prolongée est plus élevée chez les vaches perdant plus de 1 à 1,5 point entre 0 et 60 jours ainsi que chez celles présentant une note insuffisante à 30 jours de lactation (note<2). Une note intermédiaire (entre 2 et 2,5) diminue le risque d'activité ovarienne désordonnée. Le risque de reprise de cyclicité anormale est également élevé pour les vaches en bon état, voire grasses.
3) Aspect physiopathologique
La régulation endocrine de la reprise d'activité sexuelle fait intervenir la leptine. Cette hormone est sécrétée par le tissu adipeux et par le placenta [36] mais aussi, dans une moindre mesure, par la mamelle, le rumen, l'abomasum et/ou le duodénum et l'hypophyse [17]. Sa concentration sérique est associée à la quantité d'adipocytes et à l'importance des réserves corporelles en graisse. Elle agit sur l'hypothalamus en favorisant la sécrétion de GnRH ou plus exactement en limitant son inhibition. L'hypophyse possède également des récepteurs à la leptine laissant supposer une action directe. La leptine pourrait ainsi renseigner l'hypothalamus sur les réserves énergétiques à long terme et sur les capacités de l'animal à mener à bien la croissance folliculaire, l'ovulation, et enfin la gestation. Notons que la leptinémie des génisses est physiologiquement inférieure à celle des multipares et peut être en rapport avec une reprise d'activité ovarienne plus tardive chez celles-ci (figure 23) [36].
La leptinémie des vaches commence à diminuer de quatre à une semaine avant le vêlage, elle atteint un nadir autour de 3 à 6 ng/ml au cours de la première semaine post-partum (figure 23) pour réaugmenter ensuite. Cette diminution dès la fin de gestation/début de lactation résulterait de la diminution de la prise alimentaire, de la négativation du bilan énergétique, d'une insulinorésistance, et d'une diminution de l'état corporel durant cette période [17]. Elle aurait également comme rôle de diminuer la sensibilité à l'insuline et la synthèse protéique, de diminuer l'activité thyroidienne et d'augmenter l'efficacité alimentaire. Cette activité participe au partage homéorhétique des nutriments notamment pour favoriser l'approvisionnement de la mamelle (insulino-résistante, voir plus bas) mais a pour contrepartie un blocage de la reproduction [17].
Figure 23 : Influence de l'état corporel au vêlage sur la leptinémie des vaches avant et après vêlage [36].
Note (V) : Note d'état corporel au vêlage.
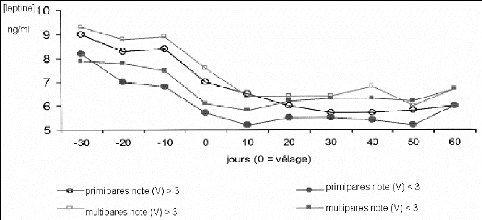
La leptinémie est donc corrélée positivement à la note d'état corporel au pic de lactation, les vaches et les génisses ayant une note d'état corporel supérieure à trois ont des taux d'Insulin Growth Factor 1 (IGF1) et de leptine plasmatique plus élevés [17]. De même, la chute du taux de leptine est plus prononcée chez les génisses et se met en place plus tardivement chez les vaches grasses. Enfin, les multipares et les génisses grasses présentent une leptinémie post-partum supérieure à 5,5 ng/ml et retrouvent une activité ovarienne cyclique plus tôt contrairement aux génisses maigres. Un pic de leptine supérieur à 5 ng/ml est d'ailleurs généralement observé (parallèlement a une diminution du taux d'AGNE sanguins) une semaine avant le premier pic de progestérone en post-partum [17].
L'utilisation du glucose est également déterminante pour la reprise d'activité ovarienne. Il semble être la principale source d'énergie utilisée par l'ovaire. Or, certains tissus non insulinodépendants [17]ont alors prioritaires. C'est le cas du cerveau, du coeur ou de la mamelle. Il est probable que l'entrée du glucose dans l'ovaire soit insulino-dépendante. Ceci pourrait également expliquer pourquoi les primipares, plus insulino-résistante que les multipares présentent des
délais de reprise de cyclicité post-partum plus longs que les multipares malgré un bilan énergétique plus favorable, Grimard citée par Ennuyer [36].
Le rôle du déficit énergétique est également prépondérant. D'un point de vue biochimique, les vaches en bilan énergétique négatif ont des concentrations sanguines en IGF1, en glucose et en insuline insuffisantes et des concentrations d'hormone de croissance et d'acides gras libres élevées. D'un point de vue physiopathologique, les effets de ce déficit énergétique sur la pulsatilité de LH expliquent le retard de la première ovulation après le vêlage.
La concentration d'IGF1 a été démontré comme variant dans le même sens que la balance énergétique [92] ; et en inversement à la production laitière. Cet accroissement d'IGF1 est également lié à l'augmentation de la sécrétion de progestérone au cours du cycle oestral. La réduction du taux d'IGF1 semble accompagnée d'une réduction d'activité ovarienne et compromet la croissance et la qualité des follicules [90].
Notons que le déficit est d'autant plus élevé que les vaches produisent beaucoup (forte expression du pic), que leur capacité d'ingestion est limitée (cas des génisses) ou quelles ingèrent moins (pathologie) [36].
Un bilan énergétique négatif altère donc la sécrétion d'IGF1. Il diminue la taille, altère la qualité et la croissance du follicule. En outre, ce follicule est le principal sécréteur d'oestrogènes, hormones responsables du comportement d'oestrus.
Les premières chaleurs post-partum sont l'évènement le plus concret que l'éleveur va détecter et qui témoigne d'une cyclicité normale. C'est également une date prise en compte pour détecter les chaleurs suivantes. Pourtant, la détection des premières chaleurs est de plus en plus difficile car elles durent de moins en moins longtemps [20] : à l'heure actuelle en moyenne entre quatre et quatorze heures toutes les trois semaines [88]. Or la détection des chaleurs a une influence majeure sur les paramètres de reproduction, notamment sur l'intervalle vêlage/première insémination artificielle. Les premières chaleurs sont détectées en moyenne à 59 jours postpartum chez les vaches laitières mais l'intervalle vêlage/premières chaleurs peut parfois s'étendre à plus de 70 jours post-partum (tableau 12) [41]. Pourtant, les chaleurs utilisables pour une mise à la reproduction normales sont définies comme celles détectées entre 50 et 70 jours post-partum [54].
Tableau 12 : Intervalle vêlage-première chaleur. D'après [41]. L'intervalle moyen est de 59 jours.
|
Intervalle |
<50 jours post-partum |
[50-70] jours post-partum |
>70 jours post-partum |
|
% |
45 |
25 |
30 |
Environ 50% des chaleurs ne sont pas détectées en élevage actuellement et 5 à 20% des vaches sont inséminées en phase lutéale ou en début de gestation du fait d'une mauvaise détection de celles-ci. Une partie de ces mauvais résultats vient souvent de facteurs humains : le temps consacré par l'éleveur à cette tâche primordiale diminue avec l'augmentation de la taille des troupeaux [88].
L'acceptation du chevauchement reste le principal facteur caractérisant l'oestrus : la vache s'immobilise lors d'un chevauchement par l'arrière ou avance un peu sous le poids de sa congénère [20]. Mais certaines, plus actives que d'autres vont vers les autres et acceptent difficilement voire refusent d'être chevauchées. Il faudra aussi prendre en considération d'autres facteurs notamment environnementaux : un sol glissant inhibe les manifestations d'oestrus, le mode de stabulation en logettes y est aussi moins favorable comparativement à la stabulation libre. D'autres vaches en chaleurs peuvent également inhiber le comportement de certaines. L'acceptation du chevauchement n'est jamais exprimée en phase lutéale et est donc spécifique de l'oestrus. D'autres critères peuvent être repérés mais peuvent aussi être observés en phase lutéale. C'est l'augmentation de la fréquence d'un ou de plusieurs de ces critères chez un même animal qui doit alerter : agitation, mictions, beuglement, chevauchements initiés par la vache en chaleur, pose ou frottement du menton sur la croupe ou le dos d'une congénère, flairage ou léchage de la vulve avec ou sans signe du flehmen. Ces derniers comportements peuvent être initiés ou acceptés par la vache en chaleur. Une vache en chaleur se déplace significativement plus et reste moins longtemps couchée [20].
Associés à ces signes comportementaux, interviennent des modifications physiques : vulve congestionnée, présence d'un mucus vulvaire transparent et filant d'origine cervicale, poils ébouriffés voire des érosions cutanées au niveau de la croupe, de la base de la queue et des tubérosités ischiatiques, témoins de chevauchements répétés.
Les signes secondaires sont donc assez subjectifs, ce qui rend difficile la détection des chaleurs. La tâche est encore compliquée par deux phénomènes : d'une part, nombre d'ovulations sont silencieuses. La première ovulation n'est accompagnée de signes d'oestrus que dans un cas sur trois [35]. Mais en dehors de celle-ci, des études rapportent que 14% des vaches n'expriment aucun comportement de chaleurs et par conséquent sont indétectables [20]. D'autre part, les vaches peuvent manifester également des fausses chaleurs. On parle de fausses chaleurs lorsque l'animal présente des signes d'oestrus alors que la concentration en progestérone excède 5 ng/ml. Dans l'étude de Kerbrat et Disenhaus [54], six vaches sur cent deux observées ont présenté des fausses chaleurs. Cinq d'entre elles avaient un profil de reprise de cyclicité anormal et une avait un profil normal. Cette tendance est confirmée par d'autres études : le pourcentage de fausses chaleurs est particulièrement élevé pour les vaches ayant présenté une cyclicité post-partum
anormale ou retardée (respectivement 30% et 14%) alors qu'il n'est que de 8% pour les vaches à cyclicité normale [23].
Il est donc important de ne pas négliger cet évènement et de recommander aux éleveurs une observation matin et soir, pendant trente minutes au total, et en ne se consacrant qu'à l'observation des animaux : le temps de traite, de distribution de la ration ou le paillage, ne doivent pas être considérés dans le temps recommandé. Il existe également de nombreux dispositifs d'aide à la détection des chaleurs. Pour ceux qui prennent en compte le critère d'acceptation du chevauchement pour leur fonctionnement, il faudra donc rester méfiant quant aux vaches n'exprimant pas ce comportement d'autant qu'il en existe d'autres, tels que le podomètre, qui ne sont pas basés sur ce principe. La prise en compte de la seule acceptation du chevauchement permet de détecter seulement 37 à 53 % des vaches en chaleurs, même avec deux observations d'au moins trente minutes par jour [88]. Afin de prendre en compte les autres modifications associées à l'oestrus, une grille attribuant un certain nombre de points selon les différents signes observés pour un animal donné a été proposée (tableau 13). Pour une vache donnée, les points s'additionnent sur une période de vingt-quatre heures. Avec deux ou trois observations de trente minutes par jour, une vache est considérée en chaleurs pour un score qui atteint au moins cinquante points [88].
Tableau 13 : Grille de pointage pour la détection visuelle des chaleurs [88].
|
Signes d'oestrus |
Points |
Nombre |
Total |
|
Présence de mucus à la vulve |
3 |
||
|
Interactions avec d'autres vaches |
3 |
||
|
Agitation |
5 |
||
|
Flairage de la vulve d'une autre vache |
10 |
||
|
Appui du menton sur une autre vache |
15 |
||
|
Chevauchée sans immobilisation |
10 |
||
|
Chevauchement (ou essai) d'autres vaches |
35 |
||
|
Chevauchement d'autres vaches par l'avant |
45 |
||
|
Acceptation du chevauchement |
100 |
||
|
TOTAL |
|||
10 à 15% des vaches laitières présentent des dysfonctionnements ovariens qui perturbent l'expression des chaleurs au moment de l'insémination artificielle (IA) première, c'est-à-dire audelà de cinquante jours post-partum [88] ; c'est 5 à 10% de plus que dans les années 80 [20].
Quatre vaches sur cinq normalement cyclées depuis le vêlage sont détectées correctement. Cette proportion devient significativement inférieure pour les autres : en effet, malgré l'attention particulière portée à la détection, lorsque l'activité cyclique post-partum est irrégulière ou retardée, la détection des chaleurs de mise à la reproduction n'est réalisée que pour une vache sur deux (figure 24). Cette moindre détectabilité des chaleurs des vaches dont la cyclicité est rétablie va dans le sens d'une expression de l'oestrus plus faible au cours de la première ovulation à la fois en terme de nombre de signes (acceptation du chevauchement) que de l'intensité et de la durée de ceux-ci. Une difficulté supplémentaire s'ajoute alors à l'éleveur pour les animaux présentant des anomalies de cyclicité, il n'a alors aucun repère fiable et prévisible pour les chaleurs suivantes (de mise à la reproduction, après cinquante jours) [21].
Mais au-delà des vaches présentant des dysfonctionnements ovariens, celles dont la cyclicité est régulière n'ont pas toujours une activité oestrale aussi bien orchestrée. En effet, plus d'un tiers (35%) des vaches dont la cyclicité a été vérifiée par des profils de progestérone n'ont pas de chaleurs détectées entre 50 et 70 jours. Dans cette étude, 73 % des vaches ont eu un profil de cyclicité normal mais seulement 32 % ont présenté des chaleurs régulières. 16% ont eu des chaleurs irrégulièrement et 26 % n'en ont pas eu. Enfin, 63 % des vaches à profil normal ont eu au moins une chaleur non détectée entre 30 et 80 jours post-partum [41].
Les anomalies de cyclicité sont un facteur significatif influençant la détection des chaleurs. Le pourcentage de détection de celles-ci varie considérablement selon les profils (tableau 14).
Figure 24 : Efficacité de la détection des chaleurs entre 50 et 90 jours post-partum en fonction des profils d'activité cyclique [21].
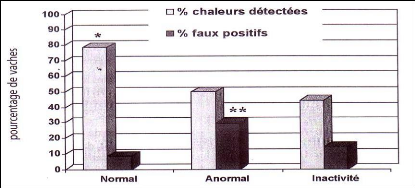
% significativement différents entre colonnes de la même couleur à *p<0,05 et **p<0,01.
Tableau 14 : Pourcentage de chaleurs détectées selon le profil de cyclicité
Dans ces deux études, les chaleurs ont été détectées selon l'habitude des éleveurs mais ont été néanmoins confirmées à posteriori par les dosages de progestérone sur le lait congelé et analysé en fin de collecte.
|
Profils |
Normal |
Inactivité |
Interruption |
Phase |
Activité |
|
% chaleurs régulières p<0,0001 |
43,1 |
5 |
27,3 |
3,85 |
37,5 |
|
% chaleurs détectées |
76 |
46 |
|||
Il semble difficile d'établir une relation directe entre note d'état et expression des chaleurs. Dans une étude visant à étudier les relations entre la note d'état de 0 à 120 jours post-partum et les paramètres de reproduction [76], les éleveurs devaient noter les signes d'oestrus. Soit ils observaient un seul signe (acceptation du chevauchement ou signe non spécifique), soit ils observaient plusieurs types de signes (dont acceptation du chevauchement ou non). La proportion de chaleurs comportant un seul signe d'oestrus noté a eu tendance à diminuer lorsque la perte d'état entre zéro et trente jours a été inférieure à un point. Ces résultats peuvent être à la fois mis en relation avec une moindre expression des chaleurs chez les femelles présentant une perte d'état élevée au cours du premier mois de lactation, ainsi qu'avec des facteurs liés à la détection des chaleurs, illustrant le rôle prépondérant de l'éleveur.
En ce qui concerne l'intervalle vêlage-première chaleur, la première chaleur a été vue en moyenne 57#177;31 jours après le vêlage et les femelles inséminées 82#177;26 jours après le vêlage. La perte d'état corporel supérieure à un point entre 0 et 30 jours, autant qu'un état corporel insuffisant au vêlage ou encore une affection du post-partum ont significativement allongé le délai moyen d'apparition des premières chaleurs après vêlage. Il existe d'ailleurs dans cette étude, des différences significatives au sein même des multipares : les femelles présentant un bon état corporel ont été vues en chaleur, puis inséminées dans des délais plus courts après vêlage, alors que les femelles en état corporel insuffisant ont présenté les délais les plus longs [76].
Le bilan énergétique a une influence majeure à cette période, l'expression des chaleurs diminue à chaque cycle tant qu'il reste négatif. En effet, au cours du deuxième cycle, seulement 66,7% des vaches en bilan énergétique négatif manifestent leurs chaleurs avant l'ovulation, contre 80% des vaches en bilan énergétique positif [92].
Tableau 15 : Note d'état corporel et reprise d'activité sexuelle d'après [76]. Echelle utilisée allant de 0 à 5.
|
PROFILS DE CYCLICITE (%) |
|||||||
|
Variable |
Classe |
Première chaleur vue par l'éleveur |
p |
Cyclicité |
Cyclicité chaleurs |
Phase |
Cyclicité |
|
Affection |
non |
Réf |
0,001 |
66,7 |
66,7 |
39,5 |
33,3 |
|
oui |
+ 11 j |
33,3 |
33,3 |
60,5 |
60,7 |
||
|
Perte d'état |
< 1 |
Réf |
0,004 |
||||
|
=1 |
+ 8 j |
||||||
|
Perte d'état |
<1,5 |
74,7 |
70,5 |
51,2 |
87,0 |
||
|
=1,5 |
25,3 |
29,5 |
48,8 |
13,0 |
|||
|
Note |
=2,5 |
Réf |
0,02 |
||||
|
]1,5-2,5[ |
+ 5 j |
||||||
|
=1,5 |
+ 16 j |
||||||
|
Note |
<2 |
30,7 |
40,0 |
58,1 |
34,8 |
||
|
[2-2,5] |
33,3 |
29,5 |
23,2 |
8,7 |
|||
|
>2,5 |
36,0 |
30,5 |
18,6 |
56,5 |
|||
|
Rang de |
Primipares |
Réf |
>0,05 |
||||
|
Multipares |
+ 2 j |
||||||
L'étude de Villa-Godoy et al. [99] s'intéresse aux génisses. Différentes rations leur sont proposées. Une première division selon l'état corporel (maigre ou non) et la deuxième selon le bilan énergétique (négatif ou non) forment 4 lots. Les auteurs se sont ensuite intéressés au comportement d'oestrus décrits précédemment. On retrouve 98,7% des comportements d'acceptation du chevauchement et 91,7% des chevauchements initiés qui correspondent à des taux de progestérone basaux (<1ng/ml) c'est-à-dire à l'oestrus. Or, dans cette étude, la
manifestation des chaleurs par l'acceptation du chevauchement, tant en durée qu'en intensité, n'a pas été significativement liée au bilan énergétique ni à la note d'état d'engraissement. Mais dans la catégorie des génisses en mauvais état corporel et en déficit énergétique, la durée de manifestation de ce signe diminue au cours des cycles successifs. C'est donc la persistance d'un bilan énergétique négatif, corroboré par une perte d'état persistante, qui a un impact négatif sur le signe majeur d'oestrus : l'acceptation du chevauchement. La manifestation des chaleurs par l'activité d'acceptation du chevauchement est par ailleurs significativement supérieure à celle de chevauchement initié chez les vaches en bilan énergétique négatif. Les relations entre note d'état, bilan énergétique indépendamment ou de façon liée et les principaux critères de manifestation d'oestrus ne sont donc pas toujours évidentes.
Seule une étude datant de 1992 ne note réellement pas d'effet ni de la note d'état au vêlage, ni de la perte d'état pendant les 80 premiers jours de lactation, ni de la production laitière cumulée sur cette même période sur l'intervalle vêlage/premier signe d'oestrus [87].
Un ou deux follicules pré ovulatoires sont présents au moment de l'oestrus. D'un diamètre de 18 à 20 mm, ils sécrètent des quantités élevées d'oestradiol 17â responsable à la fois du comportement d'oestrus, du déclenchement de l'ovulation et de la préparation du tractus génital à l'accouplement et au transport des spermatozoïdes. L'action des oestrogènes provoque en particulier l'épaississement de la muqueuse vaginale et l'augmentation de la teneur en eau du mucus cervical qui s'écoule alors par la vulve [88].
Chez des vaches recevant une ration à concentration énergétique modérée et lors de déficit énergétique, on observe une moindre réceptivité des ovaires à la sécrétion de LH. Le déficit énergétique affecterait la croissance folliculaire, surtout par altération des petits follicules, soixante à quatre-vingt jours avant l'ovulation, lorsque les cellules de la thèque interne, sensibles à la LH, se multiplient ainsi que celles de la granulosa, sensibles à la FSH. Ces altérations pourraient gêner le développement ultérieur du follicule, conduisant à une atrésie du follicule secondaire ou tertiaire, ou à un retard d'ovulation. Un tel phénomène explique le décalage observé entre le déficit énergétique et ses conséquences sur l'activité ovarienne [32].
De même, la concentration en oestradiol du liquide folliculaire chez des vaches qui reçoivent une ration à concentration énergétique modérée est plus faible, ce qui pourrait être une conséquence d'une altération de la granulosa. Une telle diminution de sécrétion d'oestradiol pourrait être à l'origine d'un retard d'ovulation en retardant le pic de sécrétion de LH. [32]. Elle pourrait aussi être à l'origine de la moindre expression des chaleurs observée [92].
60 % des éleveurs classent l'infécondité comme pathologie majeure de l'élevage [35]. 1) Definitions
On peut définir la fertilité comme la capacité à se reproduire, c'est-à-dire la capacité à produire des ovocytes fécondables.
Pour ce qui est de la fécondité, elle caractérise l'aptitude pour une femelle à mener à terme sa gestation, dans les délais requis. La fécondité englobe alors la fertilité, le développement embryonnaire et foetal, la mise bas et la survie du nouveau-né (figure 25).
Il est important de noter que cette distinction entre fertilité et fécondité ne se retrouve pas dans la littérature anglo-saxonne, le terme ?fertility? englobant les deux notions retrouvées dans la littérature française.
Figure 25 : Notions de fertilité et de fécondité en élevage bovin laitier [89].
fécondité (intervalle entre deux vêlages)

vêlage n IA1 IA2 IAf vêlage
n+1

délai de mise fertilité durée de gestation
à la reproduction (temps perdu par échec des IA)
En pratique courante, plusieurs paramètres sont employés pour caractériser la fertilité (tableau
16). D'une importance majeure, on retrouve le pourcentage de réussite en première IA (TRIA1 = taux de réussite à l'IA première) ainsi que le nombre de vaches à trois IA et plus (ou plus de deux IA) [35]. L'évaluation du nombre d'IA pour obtenir l'insémination fécondante est également très importante (IA/IF).
En ce qui concerne la fertilité, on s'intéresse surtout à l'intervalle vêlage-vêlage (IVV), à l'intervalle vêlage-première insémination (IVIA1) et à l'intervalle vêlage-insémination fécondante (IVIF). Les paramètres calculés pour caractériser la fertilité et la fécondité sont présentés dans le tableau 16.
Tableau 16 : Définition des variables intéressant la fertilité et la fécondité chez la vache laitière.
IA : Insémination Artificielle ; IF : Insémination artificielle Fécondante ; IA1 : Insémination Artificielle première ; V : Vêlage.
|
Paramètres de fertilité |
|
Taux de mise bas = Nb de mise bas à terme suite à IA1 X 100 |
|
Nb d'IA1 Taux de non-retour = Nb d'IA1 sur mois (n) sans retour jusqu'à la fin du mois (n+2) X 100 |
|
Nb d'IA1 sur mois (n) Taux de réussite = Nb d'IA1 suivi de gestation à 90 jours (ou de fécondation) X 100 |
|
en première IA Nb d'IA1 IA / IF = Nb d'IA pour toutes les vaches pour obtenir une IF X 100 |
|
Somme des IF Proportion des vaches > 2 IA = Nb de vaches > 2 IA (fécondantes ou non) X 100 |
|
Nb d'IA1 |
|
Paramètres de fécondité |
|
- intervalle Vn-Vn+1 = nombre de jours entre le vêlage (n) et la date estimée du vêlage (n+1) - intervalle Vn-IFn+1 = nombre de jours entre Vn et l'insémination fécondante suivante -proportion des intervalles Vn-IA1n+1>60jours = nb d'intervalle Vn-IA1n+1>60 jours |
|
nb d'intervalles Vn-IA1n étudiés - proportion des intervalles Vn-IFn+1>60 jours = nb d'intervalles Vn-IFn+1>60 jours |
|
nb d'intervalles Vn-IFn+1 étudiés |
L'optimisation du bilan de reproduction nécessite de se fixer des objectifs. La comparaison des valeurs de l'élevage avec celles des objectifs doit se faire avec précaution en nuançant la critique selon la stratégie d'élevage choisie (voir plus loin).
Tableau 17 : Objectifs standards pour la reproduction des vaches laitières d'après [98].
|
FERTILITE |
OBJECTIFS |
|
IA/IF |
< 1,6 |
|
% vaches à 3 IA et plus |
< 15 % |
|
TRIA1 |
> 60 % |
|
FECONDITE |
|
|
IVIA1 |
70 jours |
|
% vaches ayant IVIA1 > 80 jours |
< 15 % |
|
IVIF |
90 jours |
|
% vaches ayant IVIF >110 jours |
< 15 % |
|
IVV |
365 jours |
Le kit fécondité est un outil proposé par la Société Nationale des Groupements Techniques Vétérinaires. C'est un outil qui permet d'effectuer l'analyse de la fécondité d'un troupeau laitier lors d'un audit de reproduction ou lors d'un suivi d'élevage. Il est basé sur le calcul de sept paramètres définissant quatre problèmes :
Trois paramètres sont utilisés pour définir les chaleurs non vues : le pourcentage de vaches non vues en chaleur 60 jours post-partum (IVCH1>60 jours), le pourcentage de vaches non inséminées 90 jours post-partum, le pourcentage de vaches à IVIF supérieur à 120 jours et moins de trois inséminations.
Deux paramètres définissent la fertilité : le pourcentage de réussite en première IA et le pourcentage de vaches à 3 inséminations et plus.
La régularité des cycles est également prise en compte par le pourcentage de retours tardifs et, enfin, le pourcentage de métrites évalue un problème sanitaire. Les objectifs prévus pour ces différents paramètres sont indiqués dans le tableau 18.
Tableau 18 : Objectif pour la reproduction des vaches laitières d'après le kit fécondité de la SNGTV [35].
IVCH1 : Intervalle Vêlage/Chaleurs n°1 ; IA1 : Insémination Artificielle n°1 ; IF : Insémination artificielle Fécondante.
|
PARAMETRES |
OBJECTIFS |
||
|
% de vaches à IVCH1 > 60 jours |
< |
25 |
% |
|
% de vaches à IVIA1 > 90 jours |
< |
20 |
% |
|
% de réussite en IA1 |
> |
50 |
% |
|
% de vaches en 3ième IA et plus |
< |
20 |
% |
|
% de vaches à retours tardifs |
< |
15 |
% |
|
% de vaches à IVIF > 120 jours |
< |
20 |
% |
|
% de métrites |
< |
15 |
% |
|
% de vaches réformées pour infécondité |
< |
15 |
% |
L'évolution de la conduite des élevages oblige à nuancer les repères usuellement utilisés en fonction de la conduite d'élevage choisie par l'éleveur (tableau 19). Nous l'avons déjà largement évoqué plus haut. Trois stratégies semblent se détacher, vraisemblablement pour des raisons économiques. La stratégie visant à solliciter le potentiel génétique laitier se voit allonger les durées de lactation et par là même, les intervalles de reproduction. La stratégie ?coûts réduits? cherche à réduire les coûts de production, essentiellement alimentaires, en valorisant surtout les fourrages. Le respect des recommandations en terme de paramètres de reproduction doit alors être plus strict. S'il accepte une production laitière limitée par le potentiel des fourrages, ce système ne se veut économique que si les résultats de reproduction n'aggravent pas ce manque. Le système à vêlages groupés, tout comme le précédent, a une exigence double : les vaches doivent être fécondées et, de plus, fécondées dans l'intervalle souhaité et nécessitent donc une fertilité élevée.
Tableau 19 : Objectifs de reproduction adaptés aux trois principaux systèmes d'élevage [21].
|
Critères / Systèmes |
Productivité laitière |
Coûts réduits |
Vêlages groupés |
|
Intervalle VV |
12 à 14 mois |
13 mois |
12 mois |
|
Intervalle VIF |
85-140 jours |
85-100 jours |
80-90 jours |
|
Intervalle VIA1 |
50-100 jours |
50-80 jours |
Variable selon période d'IA |
|
TRIA1 |
45% |
> 50% |
> 55% |
|
Taux de gestation |
> 80% |
> 85% |
> 90% |
La valeur absolue de la note d'état corporel ainsi que sa variation influencent la fertilité et/ou la fécondité. Un mécanisme pathogénique possible est une diminution de la progestéronémie. En effet, la sécrétion de progestérone par le corps jaune est limitée chez des vaches qui ont subi un déficit énergétique, au moins jusqu'au cinquième cycle post-partum, et le taux de réussite à l'insémination s'en trouve très affecté [32].
Dans une étude comparative des résultats de différentes études, on trouve un lien de dépendance significatif entre la NEC au vêlage et le TRIA1 seulement pour les vaches ayant une NEC faible au vêlage par rapport à celles ayant une note intermédiaire (OR=0,91 contre OR=1,04). Les vaches vêlant en état insuffisant se voient diminuer de dix point leur TRIA1. Cette relation de dépendance n'est pas retrouvée pour les vaches ayant une note élevée au vêlage, mais, pour celles-ci, l'IVIF est supérieur [51, 61] (figure 26).
A l'inverse, une différence significative sur le taux de
réussite en première IA existe dans l'étude
de Heuer
et al. [51] entre un groupe de vaches ayant une note d'état
excessive (>4) au vêlage et
un groupe ayant une note normale
(comprise entre 2 et 4). Cette différence n'est pas retrouvée
entre le groupe de vaches ayant une note faible (<2) et le groupe ayant une note normale. Les différences sur les autres paramètres de fertilité ne sont pas significatives [51, 100].
Figure 26 : Relation entre note d'état corporel au vêlage et principaux paramètres de reproduction [29].

Les résultats concernant la NEC à la première insémination sont homogènes. Que ce soit pour une note élevée ou faible, la relation n'est pas significative. Le tableau 19 présente les résultats de quelques études ayant cherché les relations entre la note à un instant donné et les résultats de reproduction.
Tableau 20 : Bilan de la relation note d'état corporel/paramètres de reproduction NS : non significatif ; NEC : note d'état corporel.
|
Etudes |
NEC |
TRIA1 |
IVIF |
IVIA1 |
IA/IAF |
|
[42] |
3 classes selon la note à un |
p < 0,05 |
|||
|
[64] |
NEC vêlage |
OR=0,8 |
|||
|
NEC tarissement |
OR=0,8 |
IVIF >120 j |
|||
|
[100] |
NEC tarissement, vêlage, 30 et |
NS |
NS |
||
|
[51] |
3 classes selon la note au |
p < 0,05 pour NS pour les autres |
NS |
NS |
NS |
|
[24] |
Effet de la note au tarissement, |
OR=0,8 mais NS |
La note d'état a donc une influence sur les résultats de reproduction peu évidente. La significativité des relations est très peu constatée à l'exception de celle entre le taux de réussite en première IA et la note d'état au vêlage. Néanmoins, on observe malgré tout des tendances à la dégradation des résultats pour des notes extrêmes (figure 25 et tableau 19). Bon nombre d'auteurs soulignent d'ailleurs la limite d'interprétation de leurs résultats en évoquant qu'une étude forçant la note d'état corporel vers des notes extrêmes pourrait être judicieuse.
Les conséquences d'une perte d'état sur la reproduction sont plus évidentes que celles de la valeur absolue de la note d'état (tableau 21). Dans l'étude de Lopez-Gatius et al. [61], répertoriant les résultats de nombreuses études, le lien entre cette perte et le TRIA1 est faible pour la catégorie de vaches perdant peu. La relation devient plus évidente quand la perte dépasse un point. Dans cette même étude, la perte d'état corporel a un impact surtout sur l'IVIF et surtout pour les vaches connaissant une perte sévère supérieure à un point. L'IVIF de ces animaux augmente de 10,6 jours.
Dans l'étude de Mayne et al. [66], de telles observations sont également faites ; et les auteurs arrivent à la conclusion que les vaches qui ont un meilleur intervalle vêlage/vêlage sont celles qui, outre une meilleure détection des chaleurs, un meilleur intervalle vêlage/première insémination, un meilleur taux de réussite en première insémination, ont une note d'état inférieure au tarissement (3,0 contre 3,3, p<0,05) et surtout une perte d'état en début de lactation inférieure (0,3 point contre 0,6 point ; p<0,05) comparativement à celle ayant un intervalle vêlage/vêlage plus long.
De nombreuses études trouvent des résultats similaires, une synthèse de celles-ci est présentée dans le tableau 21.
Tableau 21 : Significativité de la relation perte de note d'état corporel/paramètres de reproduction selon les études.
NEC : note d'état corporel ; TRIA1 : taux de réussite en première insémination ; IA/IF nombre d'insémination par insémination fécondante ; IVIA1 : intervalle vêlage première insémination ; IVIF : intervalle vêlage insémination fécondante ; pp : post-partum ; NS : non significatif ; OR : odd ratio.
|
Etudes |
NEC |
TRIA1 |
IA/IF |
IVIA1 |
IVIF |
IVIF > 120 j |
|
|
[69] échelle de 1 à 5 |
Perte ou gain > 1 point pp ou < |
Relation significative sur la fertilité (p < 0,05) : taux de gestation, IA/IF, TRIA1. |
|||||
|
[64] échelle de 1 à 5 |
Perte de NEC tarissement |
NS |
OR = 1,8 p < 0,05 |
||||
|
[8] échelle de 1 à 9 |
8 fermes ayant profils de perte d'état entre 5 et 45 jours pp différents |
p < 0,05 |
p < 0,01 |
||||
|
[57] échelle de 1 à 5 |
Notation tous les mois. Perte sur 100 jours |
p<0,05 |
|||||
|
[47] échelle de 0 à 5 |
3 classes : perte d'état <0,5 ou >1,0 ou 0,5<...<1,0 en début de lactation |
p<0,05 pour perte >1,0 vs les deux autres |
NS |
p<0,05 pour perte >1,0 vs les deux autres |
|||
|
[76] |
Perte entre 0,5 et 1 point entre 0 et 30 jours ou >1,5 point entre 0 et 60 jours pp |
p<0,05 |
p<0,05 |
||||
|
[45] échelle de |
3 lots, perte sévère, |
p < 0,01 pour |
p < 0,05 |
NS |
p < 0,05 |
||
L'analyse des résultats d'enquêtes montre une tendance générale vers une détérioration des résultats de reproduction lorsque cette perte après vêlage s'accroît. Tant que cette perte reste inférieure à un point, l'influence de l'amaigrissement sur la reproduction reste modeste. A l'inverse, quand la perte d'état dépasse 1,5 point, la dégradation concerne tous les paramètres de reproduction calculés ou observés (figure 27 et 28).
Figure 27 : Relation entre la perte d'état corporel 2 mois après vêlage et les performances de reproduction [28].
IV-Ovulation IV-Chaleurs 1 IV-IA1
% réuss. IA1 IA/Fécondation
Intervalles (j) et % de reussite
40
20
90
80
70
60
50
30
10
0
2,5
2
0,5
0
1,5
1
IA par fecondation
< 0,5 0,5 à 1 > 1
Baisse de note d'état après vêlage
Figure 28 : Relation entre la perte d'état corporel 2 mois après vêlage et les performances de reproduction [32].
|
40 |
IV-IF > 110 j |
|||
|
% de vaches |
30 25 20 15 10 5 0 |
0 0,5 1 1,5
Baisse de note d'état après vêlage
Chez la vache laitière, les taux de vêlage après insémination sont proches voire inférieurs à 50%. Pourtant, plusieurs expériences ont démontré que les taux de fécondation étaient supérieurs à 80% (jusqu'à 90%) [21, 52, 77]. Hormis les cas d'avortement d'origine pathologique, les cas de mortalité foetale chez les bovins sont peu nombreux (5%). En revanche, 30 à 40 % des embryons meurent après fécondation [79].
Il existe deux types de mortalité embryonnaire : la mortalité embryonnaire précoce (MEP) survient avant le 16ième jour de gestation les retours en chaleurs sont alors réguliers (entre 18 et 25 jours); et la mortalité embryonnaire tardive (MET) qui intervient après le 16ième jour post IA et occasionne des retours en chaleurs décalés, en effet, l'embryon a alors eu le temps d'émettre un signal de maintien du corps jaune, dû à l'action antilutéolytique de l'interféron ô ce qui entraîne un allongement du cycle sexuel [56].
On parle de mortalité foetale après 43 jours de gestation car l'embryon devient alors un foetus. Ce type de cas, mises à part les origines pathologiques, représente 5% des cas.
La détermination des différentes situations après insémination, présentée dans le tableau 22, peut être faite par un dosage de progestérone à 21-24 jours après l'insémination et un constat ultérieur de gestation soit par dosage de la PSPB (Pregnancy Specific Protein B) soit par échographie à 30-35 jours. Cette combinaison permet de différencier les cas de MEP, MET et de mortalité foetale [77]. Il n'est cependant pas possible de faire la distinction entre non fécondation (NF) et MEP, car, dans les deux cas, le taux de progestérone à J21-24 est faible et le constat de gestation à J30-35 est négatif. Les femelles pour lesquelles le taux de progestérone est élevé à J21-24 mais qui sont trouvées non gravides à J30-35 sont considérées comme des cas de MET [75], même si certains de ces cas peuvent correspondre à des cas de phases lutéales prolongées.
Enfin, les vaches qui présentent un taux élevé de progestérone à J21-24, un constat de gestation à J30-35 positif mais une palpation transrectale (PTR) négative à J60-100, sont considérées comme des cas de mortalité foetale.
Tableau 22 : Protocole de suivi de gestation à partir des résultats de dosages hormonaux et de diagnostics de gestation [77].
P4 : progestérone ; PTR : palpation trans rectale ; Echo : échographie ; PSPB: pregnancy specific protein B ; NF : non fécondation ; MEP : mortalité embryonnaires précoce ; MET : mortalité embryonnaire tardive.
|
P4 à J0 |
P4 à J21-24 |
PSPB ou écho J30- |
PTR à J60-100 |
|
|
NF ou MEP |
- |
- |
- |
- |
|
MET |
- |
+ |
- |
- |
|
Mortalité foetale |
- |
+ |
+ |
- |
Les situations qui compromettent le synchronisme observé entre les gamètes mâles et femelle ou entre la vésicule embryonnaire et l'ensemble utéro-ovarien de la mère sont susceptibles d'entraîner l'absence de fécondation ou l'arrêt de la gestation [56].
i) Non-fécondation, mortalité embryonnaire précoce
Le déficit énergétique de la vache en début de lactation retentit à la fois sur la qualité de l'embryon et sur l'environnement hormonal qui accompagne le développement de l'embryon. La mise à la reproduction trop précoce d'animaux dont l'état corporel est trop dégradé ou pas encore stabilisé augmente donc le risque de mortalité embryonnaire.
Dans l'étude de Fréret et al. [41], la perte d'état entre 0 et 60 jours post-partum a eu un effet sur le taux de NF-MEP : ce taux est de 41,7% pour une perte supérieure à un point, contre 29,8% lorsque la perte est inférieure à un point. Remarquons qu'aucune relation n'a été observé dans cette étude entre la note d'état au vêlage et les performances de reproduction après insémination artificielle.
Pinto et al. [75] mettent en évidence un taux de gestation plus élevé dans la classe de vaches présentant un TP supérieur (TP>30g/kg) par rapport aux autres femelles (47,1 et 41,3% respectivement). Ceci est lié à une diminution des taux de NF-MEP pour les animaux de cette classe (28,6% et 32,8% pour la classe TP bas).
ii) Mortalité embryonnaire tardive
Les études montrent là encore un effet néfaste d'un mauvais état corporel (excessif ou insuffisant) sur la mortalité embryonnaire. Les taux de MET sont plus faibles chez les vaches maigres ou en état correct que chez les vaches grasses au moment de l'insémination, avec respectivement 13,5%, 15,1% et 24,5% de MET soit un odd ratio de 2,0 pour le dernier groupe (p<0,01) [42].
De nombreux auteurs mettent alors en évidence le rôle prépondérant du déficit énergétique sur le taux de MET. Le suivi de la note d'état en post-partum est alors important : le risque de mortalité embryonnaire tardive est multiplié par 2,4 pour chaque unité d'état corporel perdu durant le premier mois de lactation [60]. De même, Tillard et al. [96] mettent en évidence la
relation entre le profil d'état corporel et le taux de MET. 8 profils sont tracés selon la note de départ et les pertes ; les profils extrêmes correspondant soit à une perte trop élevée soit à une note constamment inférieure d'1,5 point à la normale ont des taux de MET de 29 et 27 % alors que chez les animaux à profil optimal, ce taux est de 19%.
Un effet défavorable d'une production laitière élevée sur le taux de MET est également observée. Une augmentation des MET est notée chez les femelles hautes productrices (18,7% pour les vaches produisant plus de 39 kg de lait par jour contre 13,5% pour les classes de production moyenne ou faible, p<0,03). Il existe alors une interaction forte avec l'état d'engraissement, cet effet défavorable de la production laitière élevée étant essentiellement observé chez les femelles en bon état au moment de l'insémination artificielle [75].
La relation entre la note d'état corporel et les paramètres de reproduction fait intervenir le déficit énergétique. Notons qu'il existe une association entre le TP minimum enregistré sur les trois premiers mois de lactation et le taux de NF-MEP [75]. Il est également noté un écart de dix point de fertilité entre les lots de vaches TP mini faible (TP<28g/kg) et le lot de vaches TP mini élevés (TP>32g/kg). Une corrélation forte et positive existe également entre le TP mini et le TRIA1. Un objectif de TP mini>28g/kg est donc à retenir pour que l'énergie ne soit pas un facteur limitant pour la fertilité, [65].
Les hypothèses les plus couramment admises font appel au rôle de l'insuline et des IGF sur la croissance folliculaire et sur le contrôle hormonal de l'activité ovarienne. Les IGFs sont synthétisées sous stimulation de l'hormone de croissance GH (Growth Hormone ou somatotropine). Lors de déficit énergétique, la synthèse des IGFs diminue ; en début de lactation la forte sécrétion d'hormone de croissance ne s'accompagne pas d'une forte sécrétion d'IGF (phénomène de découplage) [32].
Des épisodes de mortalité embryonnaire peuvent survenir en raison d'une faible sécrétion de progestérone par le corps jaune au moment de la reconnaissance maternelle de la gestation. En effet, la progestérone est indispensable à l'établissement et au maintien de la gestation. En début de gestation, l'augmentation de sa concentration circulante suffisamment tôt permet, grâce aux
modifications de l'environnement utérin, un développement optimal de l'embryon et ainsi une production d'interféron ô satisfaisante pour inhiber le mécanisme de lutéolyse dû à la PGF2á [79]. La sécrétion de progestérone est également essentielle durant la phase lutéale pour assurer la nutrition de l'embryon [79]. Une moindre sécrétion de progestérone par le corps jaune après les premières ovulations, lié à une moindre sensibilité du corps jaune à la LH voire à une lutéolyse précoce, pourrait entraîner des risques de mortalité embryonnaire [29]. En période de déficit énergétique, la sécrétion pulsatile de LH est compromise mais les réserves pituitaires en LH augmentent ce qui semble indiquer que c'est la décharge de GnRH qui est diminuée. La moindre sécrétion de LH diminue son effet lutéotrope sur le corps jaune et pourrait donc diminuer la sécrétion de progestérone. L'apport énergétique en début de lactation conditionne donc la qualité du corps jaune et sa sécrétion de progestérone nécessaire à l'établissement d'un environnement utérin compatible avec le développement de l'embryon. Mais cet environnement est fragile, un apport excessif en matières azotées a des conséquences catastrophiques. En plus d'aggraver le bilan énergétique en raison de l'énergie nécessaire pour détoxifier l'ammoniaque, les vaches alimentées avec une ration à matières azotées totales élevées ont des concentrations de progestérone diminuée (diminution des concentrations sanguines de cholestérol, précurseurs des hormones ovariennes stéroïdiennes.) ainsi qu'un pH utérin diminué par la diffusion d'urée et d'ammoniac dans le fluide utérin auquel l'embryon est très sensible [79]. Notons enfin que les apports lipidiques ont également une influence sur la mortalité embryonnaire en favorisant la synthèse de progestérone voire en en diminuant la clairance par augmentation de la cholestérolémie et de la proportion de lipides dans le corps jaunes. Une moindre sécrétion d'oestrogènes ou de prostaglandines lors d'ajout de matières lipidiques dans la ration pourrait également être l'un des éléments d'explication d'un meilleur fonctionnement du corps jaune [79].
Les vaches en inactivité ovarienne prolongée ont tendance à être mise à la reproduction plus tardivement. L'intervalle vêlage-première ovulation est plus court chez les vaches à corps jaune persistant que chez les vaches normales, [97]. L'intervalle vêlage-reprise d'activité ovarienne est corrélé positivement avec les intervalles IVA1 (p<0,05) et IVIF (p<0,01), [8], eux-mêmes liés au bilan énergétique et à la vitesse de mobilisation des réserves corporelles, [15].
L'étude de Shrestha et al. [91] étudie
l'effet des anomalies de cyclicité au cours des premiers
cycles sur
les suivants et les résultats de reproduction conséquent. Dans
cette étude, 42,9% des
vaches ont présenté des cycles normaux, 35,2% ont eu une phase lutéale prolongée (profil A), 13,2% n'ont pas ovulé ou ont eu une ovulation retardée (profil B), et 8,8% ont eu des cycles irréguliers. Les résultats sont présentés dans le tableau 23, le degré de significativité donné correspond à la comparaison avec les vaches à profil de cyclicité normal.
Tableau 23 : Cyclicité anormale au cours du premier cycle et cycles suivants [91].
|
Paramètre Profil |
%Insémination à 100 jours |
Taux de conception |
Taux de gestation |
IVIA1 |
IVIf |
|
Profil A |
P<0,05 |
p<0,05 |
p<0,01 |
p<0,01 |
p<0,01 |
|
Profil B |
p<0,05 |
p<0,01 |
p<0,05 |
p<0,01 |
Le diagnostic étiologique de l'avortement est assez difficile en élevage. La mise en évidence d'un rôle direct de la note d'état corporel l'est tout autant. Une étude de Lopez-Gatius et al. [60] étudie les facteurs de risque d'avortement entre trente jours et quatre-vingt dix jours suivant l'insémination. Une perte d'état corporel élevée entre le vêlage et trente jours post-partum en est un : une perte autour d'une unité de NEC est associé à un risque 2,4 fois plus élevé d'arrêt de gestation pendant la période étudiée.
La mise bas est un phénomène physiologique continu que l'on a pour habitude de diviser en quatre phases.
Pendant la première phase, dite préparatoire, s'initie le développement mammaire. La vulve devient oedémateuse et relâchée. Les ligaments sacro-sciatiques sont de plus en plus lâches. La parturition est imminente lorsque l'on peut les déplacer crânialement de plus de trois centimètres. Le bouchon muqueux du col utérin se liquéfie entre trois et quelques heures avant la mise bas.
Chez les ruminants, les surrénales foetales jouent un rôle clé dans le déclenchement de la parturition. L'augmentation exponentielle du cortisol foetal pendant les dernières semaines provoque une séquence d'évènements endocriniens qui aboutira à la parturition. L'augmentation marquée de l'ACTH (Adreno Cortico Tropic Hormone) foetal entraîne une hausse importante du cortisol, ce qui présuppose un axe hypothalamo-hypophyso-surrénalien foetal fonctionnel. Le point de départ de cette activation surrénalienne résulte d'un processus d'auto-amplification : le cortisol foetal agit par rétrocontrôle positif au niveau hypophysaire, en amplifiant l'effet stimulant du CRF (Cortisol Releasing Factor) et de l'AVP (vasopressine) sur la sécrétion d'ACTH, [4]. Cette hypercortisolémie est essentiel à la mise en place du surfactant pulmonaire sans lequel la vie extra-utérine est impossible. L'augmentation du cortisol est aussi responsable de l'importante synthèse d'oestrogènes, surtout de l'oestrone par les cotylédons, [49]. La production d'oestrogènes (oestradiol et oestrone) se fait au dépend de celle de progestérone. La diminution du rapport progestérone/oestrogènes agit sur les tissus concernés (myomètre et col utérin) directement ou par le biais d'autres hormones (prostaglandines, ocytocine).
La production de prostaglandines (PGF2á et PGE2) se fait
essentiellement par l'endomètre mais
également par le placenta
à partir de 100 jours de gestation. Leurs taux circulants
s'élèvent
brutalement au moment du terme sous l'effet
stimulant des oestrogènes sur la phospholipase A2
et la cyclo-oxygénase. Elles accélèrent alors la lutéolyse (ce qui accélère la chute du rapport progestérone/oestrogène donc l'auto-amplification), provoquent des contractions utérines et la dilatation cervicale et augmentent la sensibilité du myomètre à l'ocytocine.
La relaxine est synthétisée par le corps jaune essentiellement. Sa concentration augmente progressivement en fin de gestation pour s'effondrer au moment de la mise-bas. C'est une hormone favorisant le relâchement des ligaments sacro-sciatiques, du col utérin et des fibres lisses myométriales. La formation de ses récepteurs est stimulée par les oestrogènes.
Le contrôle hormonal de la mise-bas agit donc en boucle d'auto-amplification ; chaque hormone a son propre rôle, le déterminisme de leur sécrétion se fait en cascade aboutissant aux modifications des tissus mous décrits précédemment ainsi qu'à la dégradation enzymatique du collagène du col utérin aboutissant à sa maturation et relaxation. Le cortisol foetal agirait également pour stimuler la décharge lutéolytique de prostaglandines par l'endomètre directement ou par le biais des oestrogènes [4] (figure 29).
La phase suivante est la première phase de la mise bas proprement dite. Elle commence avec le début de dilatation du col utérin. Les contractions utérines longitudinales dilatent surtout l'ostium externe du col et lui confèrent un diamètre interne de 8 à 12 cm. Puis commence la dilatation active réalisée par l'allanto-chorion poussé à travers le col par des contractions utérines plus fortes et plus fréquentes. L'utérus est innervé par des fibres du système nerveux sympathique qui provient du nerf hypogastrique. Les cellules du myomètre possèdent des récepteurs adrénergiques á à effet contractile et des récepteurs adrénergiques â à effet relaxant. Pendant la parturition, la stimulation des récepteurs adrénergiques participerait au contrôle de la qualité et de la longueur des contractions utérines. Les prostaglandines sont aussi associées à la commande des contractions utérines. L'effacement complet du col signe la fin de la phase 1 et prend 2 à 6 heures.
Figure 29 : Déterminisme de la mise-bas chez les ruminants [4].
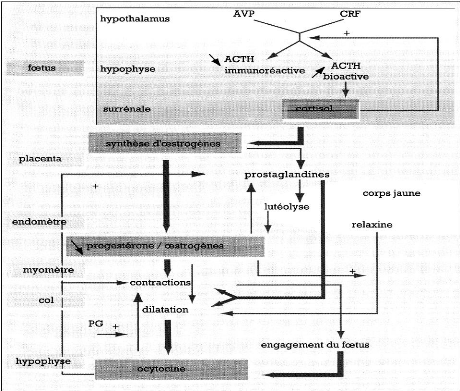
Les fibres innervant le col et le vagin antérieur régularisent la mise en circulation de l'ocytocine par la neurohypophyse. Le taux plasmatique de celle-ci augmente alors après la dilatation du col et surtout du vagin antérieur. C'est le réflexe de Fergusson [4]. A ce moment là, les contractions utérines deviennent beaucoup plus puissantes et régulières, engageant le foetus dans la filière pelvienne [4, 49]. Les contractions sont responsables de 90% du travail de dilatation et d'expulsion. Une fois engagé, le rôle des contractions abdominales devient aussi essentiel à l'expulsion du veau. La durée de l'expulsion (stade 2) est très variable d'une race à l'autre et est toujours plus longue chez les primipares. Bien que le foetus puisse vivre 8 à 12 heures après le début du stade 2, il est normalement expulsé en moins de 4 heures. Souvent, il y a arrêt momentané des efforts d'expulsion au moment de la rupture de l'allanto-chorion ; les contractions reprenant quand l'amnios apparaît à la vulve. Après la rupture de l'amnios, la
fréquence de la presse abdominale est de une à deux toutes les deux à trois minutes. En présentation antérieure, il y a presque toujours une pause au moment où le front arrive à la vulve qui peut alors se dilater. Après l'extériorisation de la tête, l'expulsion totale du veau est très rapide.
Les membranes foetales sont alors évacuées dans les heures qui suivent, ce qui correspond, avec l'involution utérine, au troisième et dernier stade de la parturition [49].
Il y a dystocie quand le déroulement de la parturition est prolongée ou arrêté à la suite d'un problème physique ou physiologique. La moindre entrave aux contractions abdominales ou utérines peut entraîner une dystocie. Elle concerne moins de 5% des gestations, la prévalence est plus élevée chez les vaches allaitantes et chez les primipares [49].
Les dystocies sont souvent classées selon leur origine, foetale ou maternelle. Il est toutefois fréquent de diagnostiquer plusieurs problèmes. Une dizaine de diagnostics peuvent expliquer ces dystocies. 75% des dystocies chez les vaches adultes et 90% des cas chez les génisses résultent d'un défaut de présentation, de position ou de posture du veau ou d'une disproportion foetopelvienne. Le poids du veau à la naissance et le diamètre pelvien de la parturiente sont les deux éléments les plus critiques dans l'échec de mise-bas naturelle. Le sexe du veau est également particulièrement déterminant dans l'incidence des dystocies : les veaux mâles ont une gestation plus longue de deux jours en moyenne et pèsent plus lourd. La présentation est aussi déterminante : les présentations postérieures comptent 65% de veaux mâles et sept fois plus de dystocie qu'en présentation antérieures. Mais elles ne représentent que 5% des veaux uniques [4].
D'un point de vue endocrinien, une concentration sérique plus faible en oestrogènes et plus élevée en progestérone chez la parturiente après le 260ième jour de gestation a été associée à une augmentation des taux de dystocies. L'insuffisance oestrogénique a un impact sur la préparation des tissus mous vue précédemment ; la constriction de la vulve et du vagin de même que la dilatation insuffisante du col utérin sont souvent diagnostiquées chez les génisses vêlant avant terme. Ces problèmes s'expliquent par un défaut de synchronisme des changements endocriniens en peri-partum [4].
Le niveau énergétique de la ration dans la deuxième moitié de la gestation est l'élément le plus déterminant pour le poids du veau à la naissance. Le développement du diamètre pelvien est lui aussi dépendant, entre autres, de l'apport énergétique pendant la gestation. Le diamètre pelvien des génisses nourries avec une ration plus énergétique est en moyenne plus grand que celui des génisses nourries avec une ration moins énergétique. Cependant, quand la ration devient trop énergétique, le pourcentage de dystocie augmente sensiblement, dans ce cas, l'augmentation des dépôts de gras dans la filière pelvienne en réduit le diamètre. La note d'état corporel présente là encore un intérêt, les animaux en état d'engraissement excessif (note d'état > 4 sur une échelle de 0 à 5) sont plus à risques d'un excès de gras dans la filière pelvienne, d'où un diamètre pelvien plus faible et un risque de dystocie plus élevé, surtout pour les primipares [43].
Au-delà de 15 % de vaches atteintes de rétention placentaire dans un troupeau, un diagnostic de troupeau devient nécessaire. Notons que l'estimation de ce chiffre est difficile car cette affection fait de moins en moins l'objet de visite vétérinaires. La démarche diagnostique (figure 28) prend en compte l'évaluation du statut énergétique en fin de gestation.
L'état d'engraissement a là encore son rôle à jouer qu'il soit insuffisant ou excessif. L'état d'engraissement au vêlage inférieur à 2, révélateur d'un état de sous-nutrition, peut entraîner une fréquence plus élevée de rétentions placentaires [40]. A l'inverse, un état d'engraissement excessif au vêlage (>4) est également un facteur de risque de rétention placentaire [64, 74]. Il favorise un part languissant, élevant le risque de non-délivrance et de métrite. Il favorise aussi les hypocalcémies qui retardent elles-mêmes l'involution utérine, le risque de métrite s'en trouvant ensuite accru. Une vache trop grasse développe souvent une stéatose hépatique. Les capacités de détoxification du foie diminuent, ce qui favorise les non-délivrances et les métrites. Notons que cette stéatose favorise également d'autres affections métaboliques (coma vitulaire), elles-mêmes facteurs de risque des non-délivrances et métrites [3].
L'évolution de l'état corporel pendant le tarissement ne semble pas jouer de rôle significatif sur le taux de rétention placentaire [55].
Figure 30 : Démarche diagnostique lors de non-délivrances et/ou métrites en élevage [3].
Etape 1 : Eliminer les causes non alimentaires (hérédité, liées au part, infectieuses)
Etape 2 : Etudier le statut énergétique en fin de gestation : état d'engraissement insuffisant ou excessif
Etape 3 : Etudier la nutrition azotée : excès ou déficit pendant le tarissement, excès en début de lactation
Etape 4 : Contrôle du déficit énergétique en début de lactation
Etape 5 : Mise en évidence d'une acidose ruminale
Etape 6 : Contrôle de l'équilibre minéral de la ration : calcium, magnésium
Etape 7 : Détection des carences en oligo-éléments et en vitamines : sélénium, iode, vitamines E et A
On distingue la métrite puerpérale aiguë qui apparaît entre cinq et quinze jours suivant le vêlage, de la métrite chronique qui se manifeste plus tardivement. Le caractère aigu ne fait pas référence à la gravité de l'affection mais s'oppose au terme chronique, qui caractérise les infections utérines tardives, détectées plus de trente jours post-partum.
La durée d'évolution d'une métrite puerpérale aiguë est variable, de deux à quinze jours. Les signes cliniques sont dominés par la perte d'appétit, une chute de la production laitière et un état fébrile transitoire (T°>39,5°C).
Les causes et facteurs favorisants sont obstétricaux. Toute anomalie dans le déroulement de la mise-bas constitue une situation à risque de métrite. La rétention placentaire en demeure une cause majeure : 96% des rétentions placentaires évoluent vers une infection utérine avec une hyperthermie (> 39,5°C) [1]. Les causes et facteurs favorisants peuvent également être métaboliques. L'atonie utérine fréquemment associée à l'hypocalcémie ou aux déséquilibres nutritionnels en est un. L'immunodépression physiologique en post-partum est également un facteur de risque, elle entraîne une diminution des capacités inflammatoires et immunitaires. L'accroissement du taux circulant de corps cétoniques concourt à la diminution de l'activité bactéricide : la dépression des défenses naturelles est donc aggravée lors d'hypoglycémie associée à la cétose. Notons que l'utérus est naturellement contaminé par les bactéries d'origine périnéale au moment du vêlage mais qui sont éliminées normalement vers la sixième semaine après vêlage. Toute perturbation des mécanismes d'élimination peut conduire à une métrite puerpérale aiguë.
La prévention de ces situations à risque passe donc par deux volets : d'une part, il convient de sécuriser le post-partum et de redoubler d'attention lors d'incidents obstétricaux (avortement, gémellité, veau mort-né, induction de la parturition, dystocies aggravées par des manipulations gynécologiques, extractions forcées, vaginite avec ou sans déchirure [1]) , lors de prolapsus utérin, lors de rétention placentaire, de part languissant laissant suspecter une hypocalcémie. D'autre part, il convient de maîtriser les facteurs en amont. De nombreuses situations peuvent être prévenues avec une bonne maîtrise de l'alimentation. Les déséquilibres minéraux (Ca,P), les carences fréquentes en vitamines A et E, en sélénium (et iode), les acidoses latentes en font partie.
Les situations d'état d'engraissement extrêmes, notamment au tarissement, soit en excès (cétose) soit en défaut (déficit énergétique) sont des situations à risque [1]. Les vaches en état corporel insuffisant sont plus à risque de développer une métrite dans les 20 jours post-partum que celles ayant une note d'état corporel normale [51].
Elles se caractérisent par une infertilité plus ou moins persistante, sans répercussion sur l'état général mais avec des conséquences économiques marquées. C'est une affection courante : 10% des vaches au cours des trois premiers mois suivant le part, 40 à 60% des causes d'infertilité, 10% des causes de réformes [9].
Parmi les causes prédisposantes, figure l'état corporel et le déficit énergétique. En effet, la mobilisation des réserves s'effectue au détriment de l'involution utérine et de la résistance de l'endomètre aux infections. Outre le retard à la relance du cycle ovarien nécessaire à une involution utérine normale, la dépression de l'axe hypothalamo-hypophysaire du fait de la sous-alimentation diminue la résistance des épithéliums en entravant la multiplication cellulaire et en entraînant leur kératinisation. On retrouve cet effet négatif de la note d'état corporel au vêlage sur le taux de métrites mais aussi l'effet de la perte d'état en post-partum, avec un odd ratio de 2,2 [64]. Dans cette étude, cet effet est noté sur les multipares. Aucun effet n'est noté de la note d'état corporel au vêlage sur les affections utérines en post-partum chez les primipares.
A l'inverse, un état d'engraissement excessif au vêlage (syndrome de la vache grasse) favorise les mise bas difficiles et le retard de l'involution utérine [9]. Notons que l'effet négatif d'une note d'état corporel excessive à trente jours post-partum est encore significatif une fois exclus les effets d'une mise bas dystocique ou d'une rétention placentaire. Ce risque est malgré tout accru après de tel évènements post-partum [43]. L'impact sur les résultats de reproduction est alors considérable : augmentation de l'IVIA1, diminution du TRIA1, augmentation d'IA/IF, augmentation de l'IVV, augmentation du taux de kystes ovariens et enfin des réformes pour infécondité [43].
La perte d'état durant le tarissement a une influence significative sur le taux de métrite, les vaches ayant subi une perte de note d'état corporel supérieure à un point présentent un taux de métrites (quelles qu'elles soient) significativement supérieur à celles dont la perte a été plus limitée [55].
La note d'état corporel a son importance en suivi d'élevage en tant que révélateur du déficit énergétique. Ce sont alors les variations de note d'état corporel surtout en post-partum qui en témoignent. Les variations excessives, caractérisées par une variation de note d'état supérieur à un point sont souvent associées à des résultats de reproduction dégradés. Une perte d'état trop élevée a un impact sur la reprise de cyclicité et l'expression de l'oestrus. L'effet sur les différents paramètres de fertilité et/ou de fécondité est direct [47], ou indirect : une détection de l'oestrus médiocre augmente l'intervalle vêlage-premier oestrus et par la même l'intervalle vêlagepremière insémination puis le taux de réussite à l'insémination, la vache étant bien souvent inséminée au mauvais moment [22]. Il en est de même pour les différents évènements postpartum (expulsion des membranes foetales, métrites etc) dont la normalité conditionne les évènements suivants. La réussite de la mise à la reproduction de la vache laitière est une succession d'évènements conditionnés les uns par les autres. Tout écart est préjudiciable sur les performances de reproduction, notamment celle de la variation de l'état corporel.
L'élevage de la vache laitière se complique. La productivité croissante désirée ces dernières décennies a posé de nouvelles problématiques ; la sélection selon certains critères se fait toujours en élevage au détriment d'autres : la reproduction semble dès lors le facteur limitant de la croissance génétique de l'élevage laitier.
L'amélioration des résultats de reproduction demande un suivi plus technique tout au long du cycle physiologique de l'animal, et pas seulement du post-partum : le tarissement reste, nous l'avons vu, une période à ne pas négliger pour réussir l'entrée en lactation de la vache.
Le déficit énergétique a incontestablement sa part de responsabilité dans la dégradation de ces performances. Ne négligeons pas non plus les causes infectieuses, qui pour certaines peuvent avoir des conséquences désastreuses sur ces mêmes critères de reproduction.
Toute la reproduction est construite sur des équilibres hormonaux qui demeurent instables. Toute perturbation vient rompre ces équilibres, les conséquences se faisant ressentir aussi bien au niveau sanitaire, économique que zootechnique. La maîtrise de ces équilibres nécessitait de mettre en oeuvre des moyens concrets d'appréciation des facteurs de risques. De nombreux outils de suivi se sont développés, la notation de l'état corporel dont nous avons vu l'utilité fait ou doit faire parti de ceux-ci.
Mais la productivité laitière augmente vraisemblablement plus vite que les capacités d'ingestion, le déficit énergétique semble alors devoir se creuser. Des interrogations quant à la politique de sélection et de conduite d'élevage semblent alors légitimes ; d'autant plus qu'elles doivent tenir compte de la mondialisation des échanges, de la fragilité des quotas et de la nécessaire compétitivité mondiale...
1. ALZIEU J.P., AUBADIE-LADRIX M., BOURDENX L., ROMAIN-BENYOUSSEF D., SCHMITT E.J., CHASTANT-MAILLARD S., Les infections utérines précoces. Point Vét, 2005, 36 (N° spécial reproduction des ruminants): p. 66-70.
2. ARZUL P.,ENJALBERT F. Conduite de la génisse laitière du sevrage au vêlage. Conséquences des déséquilibres alimentaires. Régimes alimentaires pratiqués en France. In: Journées nationales des GTV. Dijon 17 18 19 mai 2006, 175-181
3. AUBADIE-LADRIX M., Non-délivrance et métrites chez la vache laitière. Point Vét, 2005, 259: p. 42-45.
4. BATTUT I., BRUYAS J.F., FIENI F., TAINTURIER D., La mise bas : déterminisne et maîtrise pharmacologique. Point Vet, 1996, 28 (N° spécial reproduction des ruminants): p. 67-72.
5. BAUMAN D.E.,CURRIE W.B., Partitioning of nutrients during pregnancy and lactation : a review of mechanisms involving homeostasis and homeorhesis. J Dairy Sci, 1980, 63: p. 1514-1529.
6. BAZIN S., Grille de notation de l'état d'engraissement des vaches pies-noires. ITEBRNED. 1984, Paris (France). 31 p.
7. BAZIN S., Grille de notation de l'état d'engraissement des vaches montbéliardes. ITEBRNED. 1989, Paris (France). 27 p.
8. BENAICH S., GUEROUALI A., BELAHSEN R., MOKHTAR N., AGUENAOU H., Effet du degré de mobilisation des réserves corporelles après le vêlage sur la fonction reproductive de la vache laitière en post-partum. Revue de Méd. Vét, 1999, 150 (5): p. 441-446.
9. BENCHARIF D.,TAINTURIER D., Les métrites chroniques chez les bovins. Point Vét, 2005, 36 (N° spécial reproduction des ruminants): p. 72-77.
10. BERTICS S.J., GRUMMER R.R., CADORGINA-VALINO C., STODDARD E.E., Effect of prepartum dry matter intake on liver triglyceride concentration and early lactation. J. Dairy Sci., 1992, 75 (1914-1922).
11. BROSTER W.H.,BROSTER V.J., Body score of dairy cows. J Dairy Res, 1998, 65: p. 155-173.
12. BRUNSCHWING P. La puberté de la génisse laitière : les conséquences d'une croissance excessive. In: Journées nationales des GTV. Dijon, 17 18 19 mai 2006, 169-174
13. BUTLER W.R., Nutritional interactions with reproductive performance in dairy cattle. Animal Reproduction Science, 2000, 60-61: p. 449-457.
14. BUTLER W.R., Nutrition, negative energy balance and fertility in the post partum dairy cows. Cattle practice, 2005, 13 (1): p. 13-17.
15. BUTLER W.R.,SMITH R.D., Interrelationships between energy balance and post-partum reproductive function in dairy cattle. J. Dairy Sci., 1989, 72: p. 767-783.
16. CHASTANT-MAILLARD S., FOURNIER R., REMMY D., Actualité sur le cycle de la vache. Point Vét, 2005, 36 (N° spécial reproduction des ruminants): p. 10-15.
17. CHILLIARD Y., DELAVAUD C., BONNET M., Leptin expression in ruminants : Nutritionnal and physiological regulations in relation with energy metabolism. Domestic Animal Endocrinology, 2005, 29: p. 3-22.
18. CHILLIARD Y., REMOND B., AGABRIEL J., ROBELIN J., VERITE R., Variations du contenu digestif et des réserves corporelles au cours du cycle gestation-lactation. Bull Tech CRZV Theix INRA, 1987, 70: p. 117-131.
19. DECHOW C.D., ROGERS G.W., CLAY J.S., Heritability and correlations among body condition score loss, body condition score, production and reproduction performance. J Dairy Sci, 2002, 85: p. 3062-3070.
20. DISENHAUS C. Mise à la reproduction chez la vache laitière : actualités sur la cyclicité post-partum et l'oestrus. In: Journées nationales des GTV. Tours, 26 27 28 mai 2004, 859-865
21. DISENHAUS C., GRIMARD B., TROU G., DELABY L., De la vache au système : s'adapter aux différents objectifs de reproduction en élevage laitier? Renc. Rech.Ruminants, 2005, 12: p. 125-135.
22. DISENHAUS C., KERBRAT S., PHILIPOT J.M., La production laitière des 3 premières semaines est négativement associée avec la normalité de la cyclité chez la vache laitière. Renc. Rech.Ruminants, 2002, 9: p. 147-150.
23. DISENHAUS C., KERBRAT S., PHILIPOT J.M., Entre fureur et pudeur : actualités sur l'expression de l'oestrus chez la vache laitière, in journée bovine nantaise. 2003: Nantes.
24. DOMECQ J.J., SKIDMORE A.L., LLOYD J.W., KANEENE J.B., Relationship between body condition scores and conception at first artificial insemination in a large dairy herd of hight yielding holstein cows. J Dairy Sci, 1997, 80: p. 113-120.
25. DRAME E.D., HANZEN C., HOUTAIN J.Y., LAURENT Y., FALL A., Profil de l'état corporel au cours du post-partum chez la vache laitière. Ann. Med. Vét., 1999, 143: p. 265-270.
26. EARLE D.F., A guide to scoring dairy cows. J. Agric., 1976, 74: p. 228-231.
27. EDMONSON A.J., LEAN I.J., WEAVER L.D., FARVER T., WEBSTER G., A body condition scoring chart for holstein dairy cows. J Dairy Sci, 1989, 72: p. 68-78.
28. ENJALBERT F. Alimentation et reproduction chez la vache laitière. [En-ligne]. Créé en
1998. [ http://luzernes.org/docs/Fertilit%E9%20ENJALBERT.doc], (consulté le
20/05/2007)29. ENJALBERT F., Relations alimentation-reproduction chez la vache laitière. Point Vét, 1994, 158: p. 77-83.
30. ENJALBERT F., Conseil alimentaire et maladies métaboliques en élevage. Point Vét, 1995, 27 (N° spécial maladies métaboliques): p. 33-38.
31. ENJALBERT F., Rationnement en peripartum et maladies métaboliques. Point Vét, 1995, 27 (N° spécial maladies métaboliques): p. 39-45.
32. ENJALBERT F., Relations entre alimentations et fertilité : actualités. Point Vét, 2002, 227: p. 46-50.
33. ENJALBERT F., Reproduction et fertilité des vaches laitières. Activéto, 2002, 14: p. 16- 17.
34. ENJALBERT F., Les contraintes nutritionnelles autour du vêlage. Point Vét, 2003, 236: p. 40-44.
35. ENNUYER M., Les vagues folliculaires chez la vache, applications pratiques à la maîtrise de la reproduction. Point Vét, 2000, 31 (209): p. 9-15.
36. ENNUYER M. Post-partum réussi=gestation suivante. In: Journées nationales des GTV. Dijon, 17 18 19 mai 2006, 229-238
37. FERGUSON J.D., AZZARO G., LICITRA G., Body condition assessment using digital images. J Dairy Sci, 2006, 89: p. 3833-3841.
38. FERGUSON J.D., GALLIGAN D.T., THOMSEN N., Principal descriptor of body condition score in holstein cow. J Dairy Sci, 1994, 77: p. 2695-2703.
39. FIENI F., TAINTURIER D., BRUYAS J.F., BATTUT I., Physiologie de l'activité ovarienne cyclique chez la vache. Bull. GTV, 1995, 4: p. 35-49.
40. FOURICHON C., SEEGERS H., BAREILLE N., Bilan de santé : évaluer les fréquences et les conséquences des maladies de production dans un troupeau bovin laitier. Bull. GTV, 2004, 25: p. 29-36.
41. FRERET S., CHARBONNIER G., CONGNARD V., JEANGUYOT N., DUBOIS P., LEVERT J., et al., Expression et détection des chaleurs, reprise de la cyclicité et perte d'état corporel après vêlage en élevage laitier. Renc. Rech.Ruminants, 2005, 12: p. 149- 152.
42. FRERET S., GRIMARD B., PINTO A., BOUCA P., CHEVALLIER A., HUMBLOT P., Facteurs de variation du taux de réussite en première insémination dans les troupeaux laitiers à mauvaise fertilité. Journée bovine nantaise, 2001: p. 51-54.
43. GEARHART M.A., CURTIS R., ERB H.N., SMITH R.D., SNIFFEN C.J., CHASE L.E., et al., Relationship of changes in condition score to cow health in holsteins. J Dairy Sci, 1990, 73: p. 3132-3140.
44. GERLOFF B.J., Body condition scoring in dairy cattle. Agri-practice, 1987, 8 (7): p. 31- 36.
45. GILLUND P., REKSEN O., GROHN Y.T., KARLBERG K., Body condition related to ketosis and reproductive performance in norvegian dairy cows. J Dairy Sci, 2001, 84: p. 1390-1396.
46. GRIMARD B.,DISENHAUS C., Les anomalies de reprise de la cyclicité après vêlage. Point Vét, 2005, 36 (N° spécial reproduction des ruminants): p. 16-21.
47. GRUMMER R.R. Optimization of transition period energy status for improved health and reproduction. In: World Buiatrics Congress. Nice, France, 15-19 octobre 2006, 191- 202
48. HADY P.J., DOMECQ J.J., KANEENE J.B., Frequency and precision of body condtion scoring. J Dairy Sci, 1994, 77: p. 1543-1547.
49. HARVEY D., Parturition normale et dystocie chez la vache. Méd. Vét. Québec, 1988, 18 (2): p. 89-92.
50. HEINONEN K., ETTALA E., ALANKO M., Effect of postpartum live weight loss on reproductive functions in dairy cows. Acta vet. Scand., 1988, 29 (2): p. 249-254.
51. HEUER C., SCHUKKEN Y.H., DOBBELAAR P., Post-partum body condition score and results from the first test day milk as predictors of disease, fertility, yields, and culling in commercial dairy herds. J. Dairy Sci., 1999, 82: p. 295-304.
52. HUMBLOT P., Use of pregnancy specific proteins and progesterone assays to monitor pregnancy and determine the timing, frequencies and sources of embryonic mortality in ruminants. Theriogenology, 2001, 56: p. 1417-1433.
53. JEFFERIES B.C., Body condition scoring and its use in management. Tasmanian J. Agric., Min. Agric., 1961, 32: p. 1-9.
54. KERBRAT S.,DISENHAUS C., Profils d'activité lutéale et performances de reproduction du vêlage à la prémière insémination. Renc. Rech.Ruminants, 2000, 7: p. 227-230.
55. KIM I.H.,SUH G.H., Effect of the amount of body condition score loss from the dry to the near calving periods on the subsequent body condition change, occurence of postpartum diseases, metabolic parameters and reproductive performance in holstein dairy cows. Theriogenology, 2003, 60: p. 1445-1456.
56. LEDOUX D., HUMBLOT P., CONSTANT F., PONTER A., GRIMARD B., Echecs précoces de gestation chez la vache laitière. Point Vet, 2006, 37 (numéro spécial reproduction des ruminants): p. 50-55.
57. LOEFFLER S.H., DEVRIES M.J., SCHUKKEN Y.H.;, The effects of time of disease occurence, milk yield, and body condition on fertility of dairy cows. J. Dairy Sci., 1999, 82: p. 2589-2604.
58. LOPEZ-GATIUS F., Is fertility declining in dairy cattle ? a retrospective study in northeastern Spain. Theriogenology, 2003, 60: p. 89-99.
59. LOPEZ-GATIUS F., SANTOLARIA P., YANIZ J., FENECH M., LOPEZ-BEJAR M., Risk factors for postpartum ovarian cysts and their spontaneous recovery or persistence in lactating dairy cows. Theriogenology, 2002, 58: p. 1623-1632.
60. LOPEZ-GATIUS F., SANTOLARIA P., YANIZ J., RUTLAND J., LOPEZ-BEJAR M., Factors affecting pregnancy loss from gestation day 38 to 90 in lactating dairy cows from a single herd. Theriogenology, 2002, 57: p. 1251-1261.
61. LOPEZ-GATIUS F., YANIZ J., MADRILES-HELM D., Effects of body condition score and score change on the reproductive performance of dairy cows : a meta-analysis. Theriogenology, 2003, 59: p. 801-812.
62. LOUBETTE R.,MOINEL J.M. Conduite et profil génétique des génisses pour le renouvellement du troupeau. In: Journées nationales des GTV. Dijon, 17 18 19 mai 2006, 819-823
63. LUCY M.C., Reproductive loss in hight producing dairy cattle : where will it end? J. Dairy Sci., 2001, 84 (6): p. 1277-1293.
64. MARKUSFELD O., GALON N., EZRA E., Body condition score, health, yield and fertility in dairy cows. Vet Rec, 1997, 141: p. 67-72.
65. MARTINOT Y. TP mini : un outil de mesure du déficit énegétique. In: Journées nationales des gtv. Dijon, 17 18 19 mai 2006, 709-714
66. MAYNE C.S., McCOY M.A., LENNOX S.D., MACKEY D.R., VERNER M., D.C. CATNEY, et al., Fertility of dairy cows in northern Ireland. Vet Rec, 2002, 150: p. 707- 713.
67. MEISSONNIER E., Tarissement modulé, conséquence sur la production, la reproduction et la santé des vaches laitières. Point Vét, 1994, 26: p. 69-75.
68. MIALOT J.P., HOUARD J., CONSTANT F., CHASTANT-MAILLARD S., Les kystes ovariens chez la vache. Point Vet, 2005, 36 (N° spécial reproduction des ruminants): p. 90-93.
69. MONTIEL F.,AHUJA C., Body condition and suckling as factors influencing the duration of post partum anestrus in cattle : a review. Animal Reproduction Science, 2005, 85: p. 1-26.
70. NAKAO T., MORISYOSHI M., KAWATA K., The effect of postpartum ovarian dysfunction and endometritis on subsequent reproductive performance in hight and medium producing dairy cows. Theriogenology, 1991, 37: p. 341-349.
71. OPSOMER G., CORYN M., DELUYKER H., An analysis of ovarian dysfunction in high yielding dairy cows after calving based on progesterone profiles. Reproduction in Domestic Animals, 1998, 33: p. 193-204.
72. OPSOMER G., GROHN Y.T., HERTL J., CORYN M., DELUYKER H., DE KRUIF A., Risk factors for post-partum ovarian dysfunction in high producing dairy cows in belgium : a field study. Theriogenology, 1999, 53: p. 841-857.
73. OTTO K.L., FERGUSON J.D., FOX D.G., Relationship between condition score and composition of ninth to eleventh rib tissue in holstien dairy cows. J Dairy Sci, 1991, 74: p. 852-859.
74. PEDRON O., CHELI F., SENATORE D.B., RIZZI R., Effect of body condition score at calving on performance, some blood parameters, and milk fatty acid composition in dairy cows. J Dairy Sci, 1992, 76: p. 2528-2535.
75. PINTO A., BOUCA P., CHEVALLIER A., FRERET S., GRIMARD B., HUMBLOT P., Sources de variation de la fertilité et des fréquences de mortalité embryonnaire chez la vache laitière. Renc. Rech.Ruminants, 2000, 7: p. 213-215.
76. PONSART C., DUBOIS P., CHARBONNIER G., LEGER T., FRERET S., HUMBLOT P. Evolution de l'état corporel entre 0 et 120 jours de lactation et reproduction des vaches laitières hautes productrices. In: Journées nationales des GTV. Nantes, 23 24 25 mai 2007, 347-356
77. PONSART C.,HUMBLOT P., Mortalité embryonnaire et retours décalés : que peut apporter le suivi de gestation? Bulletin Technique de l'Insémination Animale, 2003, 110: p. 35-40.
78. PONTER A., Pourquoi parler d'alimentation? Bulletin Technique de l'Insémination Animale, 2003, 43 (110).
79. PONTER A., GUELOU K., DUVAUX-PONTER C., Influence de l'alimentation sur la mortalité embryonnaire. Point Vet., 2005, 36: p. 100-105.
80. PRYCE J.E., COFFEY M.P., SIMM G., The relationship between body condition score and reproductive performance. J Dairy Sci, 2001, 84: p. 1508-1515.
81. PRYCE J.E.,HARRIS B.L., Genetics of body condition score in New Zealand dairy cows. J Dairy Sci, 2006, 89: p. 4424-4432.
82. PUSHPAKUMARA P.G.A., GARDNER N.H., REYNOLDSC.K., BEEVER D.E., WATHES D.C., Relationships between transition period diet, metabolic parameters and fertility in lactating dairy cows. Theriogenology, 2003, 60: p. 1165-1185.
83. REIST M., ERDIN D.K., VON EUW D., TSCHUMPERLIN K.M., LEUENBERGER H., HAMMON H.M., et al., Postpartum reproductive function : association with energy, metabolic and endocrine statusin hight yielding dairy cows. Theriogenology, 2003, 59: p. 1707-1723.
84. ROCHE J.R., DILLON P.G., STOCKDALE C.R., BAUMGARD L.H., VANBAALE M.J., Relationships among international body condition scoring systems. J Dairy Sci, 2004, 87: p. 3076-3079.
85. ROCHE J.R., MACKEY D.R., DISKIN M.G., Reproductive management of post-partum cows. Animal Reproduction Science, 2000, 60-61: p. 703-712.
86. RUEGG P.L., Body condition scoring in dairy cows : Relationships with production, reproduction, nutrition and health. The Compendium North America Edition, 1991, 13 (8): p. 1309-1313.
87. RUEGG P.L., GOODGER W.J., HOLMBERG C.A., WEAVER L.D., HUFFMAN E.M., Relation among body condition score, serum urea nitrogen and cholesterol concentrations, and reproductive performance in hight-producing holsteins dairy cows in early lactation. American Journal of Veterinary Research, 1992, 53: p. 10-14.
88. SAINT-DIZIER M., La détection des chaleurs chez la vache. Point Vét, 2005, 36 (N° spécial reproduction des ruminants): p. 22-27.
89. SEEGERS H.,MALHER X., Analyse des résultats de reproduction d'un troupeau laitier. Point Vet, 1996, 36: p. 127-135.
90. SHRESTHA H.K., NAKAO T., HIGAKI T., SUZUKI T., AKITA M., Relationships between body condition score, body weight, and some nutritional parameters in plasma and resumption of ovarian cyclicity postpartum during pre-service period in high-producing dairy cows in a subtropical region in Japan. Theriogenology, 2005, 64: p. 855- 866.
91. SHRESTHA H.K., NAKAO T., SUZUKI T., HIGAKI T., AKITA M., Effects of abnormal ovarian cycles during pre-service period postpartum on subsequent
reproductive performance of high-producing Holstein cows. Theriogenology, 2004, 61: p. 1559-1571.
92. SPICER L.J., TUCKER W.B., ADAMS G.D., Insulin-like growth factor-1 in dairy cows : relationships among energy balance, body condition, ovarian activity, and estrous behavior. J Dairy Sci, 1990, 73: p. 929-937.
93. STAPLES C.R.,THATCHER W.W., Relationships between ovarian activity and energy status during the early postpartum period of high producing dairy cows. J Dairy Sci, 1990, 73: p. 938-947.
94. TAINTURIER D., Pathologie de la reproduction de la vache. La Dépêche vétérinaire, supplément technique, 1999, 64: p. 47p.
95. TAYLOR V.J., BEEVER D.E., BRYANT M.J., WATHES D.C., Metabolic profiles and progesterone cycles in first lactation dairy cows. Theriogenology, 2003, 59: p. 1661- 1677.
96. TILLARD E., HUMBLOT P., FAYE B., Impact des déséquilibres énergétiques postpartum sur la fécondité des vaches laitières à la réunion. Renc. Rech. Ruminants, 2003, 10: p. 127-129.
97. TOUZE J.L., LAIGRE P., THOMERET F., BOSC M., GRIMARD B., Anomalies des profils de rétablissement de la cyclicité post-partum chez la vache laitière Prim Hostein : relations avec les caractéristiques zootechniques. Renc. Rech.Ruminants, 2004, 11: p. 400.
98. VALLET A.,PACCARD P., Définition et mesures des paramètres de l'infécondité et de l'infertilité. B.T.I.A., 1984, 32: p. 2-3.
99. VILLA-GODOY A., HUGHES T.L., EMERY R.S., STANISIEWSKI E.P., FOGWELL R.L., Influence of energy balance and body condition on estrus and estrous cycle in Holstein heifers. J Dairy Sci, 1990, 73: p. 2759-2765.
100. WALTNER S.S., McNAMARA J.P., HILLERS J.K., Relationships of body condition score to production variables in high producing Holstein dairy cattle. J Dairy Sci, 1993, 76: p. 3410-3419.
101. WILDMAN E.E., JONES G.M., WAGNER P.E., BOMAN R.L., TROUTT H.F., LESCH T.N., A dairy cow body condition scoring system and its relationship to selected production characteristics. J Dairy Sci, 1982, 65: p. 495-501.