0.4. Les espèces dans la structure verticale :
la problématique des strates et des ensembles structuraux
La structure verticale des écosystèmes
forestiers tropicaux a fait l'objet de plusieurs travaux et débats qui
n'ont pas encore jusqu'à ce jour été résolus. Au
stade mature, la forêt tropicale présente une structure verticale
stratifiée dans laquelle il est parfois difficile de définir les
compartiments. Richards (1952), distingue cinq strates indépendantes
dont trois arborescentes, une arbustive et une autre herbacée.
Senterre propose une structure verticale simplifiée en
quatre strates, que nous avons adoptée dans le cadre de ce travail, mais
en focalisant uniquement notre attention sur les deux strates
supérieures (nommées ici strate A qui renferme les arbres
dominants et les émergents et la strate Ad composée des arbres
dominés).
Deux types de configuration spatiale ressortent de l'analyse
de la structure verticale de cette forêt. Si l'on envisage la structure
spatiale en terme de strates, nous distinguons deux strates arborescentes : la
strate arborescente supérieure ou strate des dominants qui inclut les
Emergents (E) et les Dominants (A) ainsi que la strate arborescente
dominée (Ad) qui inclut l'élément propre de cette strate
et la régénération de la strate supérieure.
Mais lorsqu'on tient compte des compartiments structuraux, on
a une perception différente. L'ensemble des individus matures de
diverses espèces forment « l'ensemble du présent ». Cet
ensemble inclut tous les arbres adultes de deux strates. L'ensemble des
individus immatures constitue « l'ensemble du futur » et inclut toute
la régénération de la strate supérieure
présente dans la strate dominée.
Cette configuration introduit la notion des « ensembles
structuraux » définit par Oldeman, (1974) et Senterre (2005).
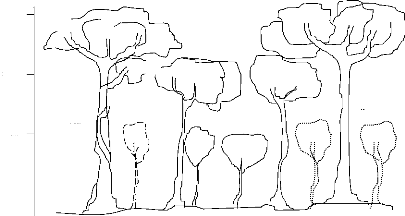
La figure 1 ci-dessous illustre la structure verticale des
espèces de deux strates supérieures étudiées dans
la forêt de notre zone d'étude.
E
A
L
a
b
c
Ad
40 m
25 m
6m
Figure 1 : Physionomie et structuration des deux strates
arborescentes dans les forêts des environs de Kisangani. E+A forme la
strate dominante, Ad : strate dominée ; tandis que a :
régénération de E+A présente dans Ad,
b : régénération de E+A absente dans E+A
c : élément propre de Ad et L : liane
0.5. Approche contingente et conception
déterministe : deux paradigmes d'explication de la diversité et
de la composition floristique
De nombreuses hypothèses ont été
proposées afin d'expliquer la coexistence des espèces et le
maintien des communautés (Palmer, 1994 ; Wright, 2001 et Ricklefs,
2004). En écologie, ces mécanismes dépendent de
l'échelle d'investigation. A une échelle biogéographique
les communautés sont reliées à des causes
géologiques, historiques et climatiques (Ricklefs, 2004 ; Gaston, 2000).
Tandis qu'à une échelle régionale ou locale, ce sont les
facteurs du relief, du substrat et les conditions abiotiques imposées
par le milieu qui définissent les communautés (Tuomisto &
al. 2003 ; Condit & al. 2002).
A l'échelle locale, deux théories majeures sont
généralement mises en avant pour expliquer la diversité
floristique et les variations dans la composition floristique.
La première est basée sur une approche contingente
(facteurs historiques) et la deuxième sur une conception
déterministe (paramètres environnementaux).
Le paradigme de la contingence historique repose sur le fait
que les plantes ont des capacités
de dispersion limitées.
L'hypothèse qui explique cette approche est celle de
l'équivalence
fonctionnelle des espèces connue aussi sous le
terme de la théorie neutre développée par
Hubbell (2005). Elle prône l'équivalence
écologique des espèces et que seuls, la capacité de
dispersion, l'extinction, la spéciation ainsi que des
événements aléatoires déterminent la composition
d'une communauté (Hardy & Sonké, 2004). C'est la
théorie de la dérive écologique (Hubbell, 2001) qui met
l'accent sur les aspects historiques et stochastiques de regroupements.
Le paradigme déterministe se base sur l'idée
selon laquelle la variabilité spécifique dans une
communauté est dictée par les interactions
interspécifiques. Ainsi, il découle de cette explication des
patrons de diversité aux différentes échelles
considérés ce qui suit : la diversité alpha (au niveau
local) dans une communauté est déterminée par des facteurs
biotiques telles que la compétition, et abiotiques (par exemple les
perturbations). La variabilité à l'échelle du paysage de
cette diversité (diversité bêta) est corrélée
à l'hétérogénéité environnementale de
ce paysage (variations topographiques, hydromorphiques, altitudinales, etc.)
(Tuomisto et al. 2003, Chesson & Warner, 1981, Kelly & Bowler,
2002, Dauby, 2007). C'est la théorie de niche écologique selon
laquelle les regroupements d'espèces sont régis par les
interactions entre elles et avec leur milieu. (MacArthur & Levins, 1967 ;
Hutchinson, 1961)
Cependant, certaines caractéristiques des
communautés laissent envisager que des paramètres autres que les
facteurs environnementaux (hétérogénéité
spatiale) participent à l'explication de patrons de diversité
(Flores, 2005).
Il est évident, à en croire Dauby (2007) qu'il
n'y a aucun modèle explicatif unique, chaque modèle apporte sa
part d'explication dans les patrons de diversité et de
variabilité au sein des forêts denses tropicales.
Toutefois, nous estimons que l'approche déterministe
conduit à une définition plus ou moins claire de la typologie des
forêts tropicales.
| 
