|
Blida, Avril 2011
1
UNIVERSITÉ SAAD DAHLAB DE BLIDA
Faculté des Sciences Agronomiques et
Vétérinaires
Département des Sciences
Agronomiques
MÉMOIRE DE MAGISTER
Spécialité : Amélioration des productions
végétales
ETUDE DE L'INTERACTION GENOTYPE X MILIEU ET DE
SON IMPACT
SUR LA SELECTION DES VARIETES
DE BLE DUR (Triticum durum Desf.)
CULTIVEES EN
ALGERIE
Par
SAFI Rekia
Devant le jury composé de
A. BOUTEKRABT Professeur, U. de Blida Président
A. ACHOUCH Professeur, U. de Blida Examinateur
A. BENBELKACEM Maître de recherche, INRA. Constantine
Examinateur
M. BENMOUSSA Professeur, U. de Blida Rapporteur
2
Mots clés : blé dur, caractères
phéno-morphologiques, génotype x milieu,
adaptation, Sélection, qualité
technologique.
RESUME
L'analyse de l'interaction génotype x environnement et
de la stabilité du rendement pour sélectionner un matériel
végétal à haut potentiel de rendement et adapté aux
différentes zones de production céréalières est
l`objectif de cette étude. L'expérimentation a été
conduite au cours de la campagne 2006/2007 sur trente génotypes de
blé dur (Triticum durum Desf.), évalués sur
quatre sites différents. Les résultats montrent des
différences d'expression des caractères mesurés et leur
relation avec le rendement en grains d'un site à un autre, suite
à la présence de l'interaction génotype x environnement
GxE. Le rendement grain varie significativement par site et entre sites,
suggérant la prépondérance de l'effet milieu sur
l'expression de cette variable. L'analyse de la variance combinée montre
que les effets milieu, génotype et l'interaction génotype x
environnement expliquent, respectivement, 91.78, 2.06 et 6.15% de la variation
totale. L'utilisation de la régression conjointe n'explique que 10.13%,
alors que le modèle AMMI explique 92.65% de la somme des carrées
des écarts de l'interaction. Les résultats indiquent que les
sites-tests peuvent être regroupés en deux domaines de
recommandation, l'un représenté par Khroub, Sétif et
Tiaret et l'autre par Oued Smar. Le génotype Bichena /
Ariza_2//Solga_8 (N°3) se distingue par un haut rendement en grains et une
bonne stabilité. Il présente une large adaptation à tous
les sites d'évaluation. Par contre Gcn/4/D68-1-93A-1A//Ruff / Fg /3/
Mtl-5 (N°12) montre une adaptation spécifique au site Oued Smar.
Ces résultats indiquent que le sélectionneur a le choix entre la
sélection de Bichena / Ariza_2//Solga_8 pour l'ensemble des sites ou
bien recommander ce génotype aux régions
représentées par les sites Khroub, Sétif, et Tiarte et de
sélectionner Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 spécialement
pour la région représentée par le site Oued Smar. Les
résultats montrent aussi que la qualité technologique est
d'autant plus acceptable que le milieu est favorable au rendement grain.
3
jetiIo
|
ÇáÊÎã æ
ÇíßÊÊ
|
,
|
ÉÇíáÁá
ÏæÏÑÇã ÑÏÇÞ
ÊÇ ÉÇíÊÁÇä
ØÁÇãä
ÁÇÞÊäÇ áÌÇ
äã ÉáÛáÇ
ÑÇÑÞÊÓÇ æ
ØÓæ
|
õ äÕ
áÎÇÏÊáÇ
áíáÍÊ
|
ÁØãä 00 ìáá
6002/6002 íÍáÇáÇ
ãÓæãáÇ ááÇÎ
ÊíÑÌ
ÉÇÑÌÊáÇ
.ÉÓÇÑÏáÇ å å
Ïå æå ÈæÇÍáá
ÉÌÊäãáÇ
ÞØÁäãáÇ
ãíÇÞ íÇí
ÊÁÇÞæÑí
ÊÇäíÇ
ÌÁÇÊäáÇ .
ÉÇáÊÎã
ãÇÞÇæã
ÉÇÚÇÑ íÇí
ÁÇåãííÞÊ ãÊ
(Triticum durum Desf.) ÈáÕáÇ
ÍãÞáÇ äã
ÑÇíÛÊÊ
ÉÇíÇÍáÇ
ÉÇáÛáÇ .
ØÇÓæ õ äÇÕ
áÎÇÏÇÊáÇ
ÏæÇÌæá
ÁÇÚÇÊ ÑÇÎÊ
ìÇá ãÇÞæã
äÇã ÉÇíÇÍáÇ
ÉáÛáÁÇ
ÁåÊÞáÇá æ
ÉÓÁÞãáÇ
ÑííÁÚãáÇ
ÊÇÑíËÇÊ
äÁÇÇ ÊÇËí
ÑíÛÊãáÇ
áíáÍÊ .
ÏæÏÑãáÇ ìáá
ØÓæáá
áãÁÔáÇ
ÑíËÊáÇ ßáÇ Ç
ÑÓã o ÁåäãÖ æ
ãÞÇæãáÇ äíÇ
ÑíÇß áßÔÇ
|
áÁãÚÊÇÓÇ
|
. ÉÇíáßáÇ
ÊÇÑÇíÛÊáÇ
äÇã
|
2.76 % æ 6.02 o 87.29
ÁÇÚÇÁÊÊ
ÑÇÓí
|
ØÇÓæ
|
õ
|
äÕ
áÎÇÏÊáÇ æ
äÕáÇ
|
o ØÓæáÇ
|
ÊÁÇíÇÑÍäÍÇ
ÇæÇãÌã äÇã 86.26 %
ÍÇÖæí AMMI Ì
æÇãäáÇ ä
äíÇÍ íÇí 70.70
%ØÇÞí ÑÓí
ßÑÊÔãáÇ
ãÌÇÑÊáÇ
|
ÞØÁÇäã
ãÇÖí áæáÇ
|
o
|
äíáÁÌã ìá
ÁåÚíãÌÊ
äßãí
ÈÑÁÌÊáÇ
ãÞÇæã ä ìá
Áåíáá
áÕÍãáÇ
ÌÁÊäáÇ ÑíÔÊ
|
.ááÁÊáá
ÉíÚíÇÑÊáÇ
|
|
.
ÉíÌæáæäßÊ
Éíáæä
|
,
|
ÈÁÎÊäÇ
|
,
|
íßÊ
|
,
|
ØÓæ
|
õ äÕ
|
,
|
ÉíÌæáæíÑæãæäíáÇ
ÕÌÁÕÎáÇ
|
, ÈáÕáÇ
ÍãÞáÇ
|
Ó 411.111
ûLoIÇA
|
0
ãÇÇÇÞÑ
íËÇÑæÇÇÇáÇ
ØãäÇÇÇáÇ
. ÑÁãÇÇÇÓ
ÏÇæ
ÉÇÇÇÞØäã
íÇÇí
áÇÇÇËãÊí
íäÁÇÇÇËáÇ
ÁÇÇÇã
ÊÑÁÇÇÇíÊ
æ íØÇÇÇÓ o
ÈæÑÇÇÇÎáÇ
íÊáÇ
ãÞÇæãáÇ
ãíãÌæ íßÊí o
ÇÑÇÑÞÊÓÇ
äÓÍáÇ æå æ
íáÁá ÏæÏÑãÇ
ÒíãÊí ) Bichena / Ariza_2//Solga_8)
) Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5) 76
ãÇÞÑ
íËÇÑæÇáÇ
ØãäÇáÇ ä
ÑÇíÛ o ÁÇåÇ
ãíÇíÞÊáÇ
ãÇÊ
æ ãÞÇæãáÇ
ãíãÌá 0 ãÞÑ
íËÇÑæáÇ
ØãäáÇ ÁÞÊäÇ
äíÇ Áã ÑÁÊÎí
ÈÎÁäáÇ áÚÌÊ
ÌÁÊäáÇ å å .
ÑÁãÓ ÏÇæ íí
ÁÕÁÎ ÁíßÊ
ÑåÙÇ
ÏÇæ íí
ÉáËãÊãáÇ
ÉÞØäãáá
ÁÕíÕÎ 76 ãÞÑ
íËÇÑæáÇ
ØãäáÇ
ÈÁÎÊäÇ æ
ÊÑÁíÊ æ íØÓ o
龄뇂
ãÞÇæã íí åÇ
ìÕæí
.ÑËß
ÏæÏÑãá
ÁÇÓÁäã
ØÓæáÇ äÁß
Áãáß äÓÍ
äæßÊ
ÉíÌæáæäßÊáÇ
ÉíáæäáÇ ä
ÁÖí äíÇÊ
ÌÁÊäáÇ
.ÑÁãÓ
4
ABSTRACT
Analysis of genotype x environment interaction and yield
stability to select plant material with high yield potential and adapted to
different areas of cereal production is the main objective of this study. The
experiment was conducted during the 2006/2007 season on thirty genotypes of
durum wheat (Triticum durum Desf.), under four different sites. The
results show differences in the expression of the measured characteristics and
their relationship with grain yield from one site to another, due to the
presence of genotype x environment (GxE) interaction. Grain yield varied
significantly within and among sites, suggesting the preponderance of the site
effect for this variable. The combined analysis of variance showed that
environment, genotype and genotype x environment effects explain respectively
91.78, 2.06 and 6.15% of the total variation. The joint regression explains
only 10.13%, while the AMMI model explained 92.65% of the sum of squared
deviations of the interaction.
Result indicate that the test sites can be grouped into two
areas
of recommendation, one represented by Khroub, Setif Tiaret and one by
Oued Smar. The genotype Bichena/Ariza_2//Solga_8 (N°3) is characterized by
a high and stable grain yield with a wide adaptation to all test sites.
However, Gcn/4/D68-1-93A-1A//Ruff/Fg/3/Mtl-5 (N°12) shows a site-specific
adaptation to Oued Smar. These results indicate that the breeder has a choice
between selecting Bichena / Ariza_2//Solga_8 for all sites or areas to
recommend this genotype represented by sites Khroub, Setif, and Tiaret and
select Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 specifically for the area
represented by the site Oued Smar. It has been also found that the
technological quality is more acceptable when the environment is favorable to
grain yield.
Key words: durum wheat, pheno-morphologic
traits, genotype x environment, Adaptation, selection, technological
quality.
5
REMERCIEMENTS
Tout d'abord, Louanges à ALLAH qui m'a guidé sur
le droit chemin et m'a inspiré les bons pas, sans sa miséricorde,
ce travail n'aura pas abouti.
Mes vifs remerciements s'adressent d'abord à ma
famille, mes parents, mes frères et soeurs pour m'avoir toujours
comprise et soutenue dans mes choix et pour m'avoir donnée la force de
réaliser ce travail.
Mes plus sincères remerciements s'adressent au
professeur Benmoussa qui a accepté de m'encadrer et aussi pour m'avoir
fait confiance dans la conduite de ce sujet. Merci pour votre patience, calme
et surtout pour votre gentillesse, corrections et conseils.
Je remercie Mr Boutekrabt de m'avoir fait l'honneur de
présider la soutenance, qu'il trouve ici ma profonde reconnaissance.
J'adresse toute ma gratitude au Professeur Achouch, pour
l'intérêt qu'il a porté à mon travail et pour avoir
accepté d'évaluer ce travail.
Un grand merci à Mr Benbelkacem Maître de
recherche à l'INRAA de Constantine pour m'avoir proposé de
travailler sur ce sujet, pour l'aide qu'il m'a apporté sur terrain et au
laboratoire, et aussi pour sa gentillesse et entière
disponibilité.
Lors de ce travail, j'ai eu la chance d'être accueillie
dans quatre stations expérimentales ainsi qu' au niveau de la direction
générale de l'ITGC et au laboratoire de qualité
technologique dirigé par Mm Madani, je remercie donc vivement les
différents responsables qui m'ont accueilli. Je transmets un immense
merci à tous les personnels des différents sites qui m'ont
beaucoup aidé au champ et au laboratoire.
Je voudrais aussi remercier tout particulièrement, Mr
Bouzerzour, Professeur à l'université de Sétif pour
m'avoir aidé dans les analyses statistiques. Mes remerciements vont
à Mr Sémiani, chargé de recherches à l'INRAA d'
Alger, merci pour votre aide et vos remarques.
J'ai pleinement profité de la présence et le
soutien de mes amis. Merci à vous tous. Par précaution, Merci
à toutes les personnes qui ont finalement interagit avec moi.
6
TABLE DES MATIERES
RESUME
REMERCIEMENTS
TABLE DES MATIERES
LISTE DES ILLUSTRATIONS, GRAPHIQUES ET TABLEAUX
INTRODUCTION 12
1. LE BLE DUR COMME CEREALE MONDIALE ET CEREALE EN
ALGERIE 14
1.1 Introduction 14
1.2 Le blé dur : situation mondiale en chiffre 14
1.3 Les recherches sur les blés en Algérie 16
1.4 Aire de production 18
1.5 Qualité et différentes formes d'utilisation de
blé dur en Algérie 20
2. LES FACTEURS GENETIQUES ET ENVIRONNEMENTAUX ASSOCIEES 21
AUX PROCESSUS DE CROISSANCE ET DE DEVELOPPEMENT
2.1 Introduction 21
2.2 Les facteurs climatiques 21
2.3 Les facteurs agro-techniques 24
2.4 Les facteurs liés à la variété
d'origine génétique 25
3. L'APPRECIATION DE LA VALEUR PHENOTYPIQUE EN PRESENCE
D'ITERACION GENOTYPE x MILIEU 27
3.1 Introduction 27
3.2 Le génotype et le phénotype 27
3.3 Concept de l'interaction génotype x milieu 28
3.4 L'élaboration du rendement et corrélation
entre caractères 30
4. L'ADAPTATION AU MILIEU COMME OBJECTIF DE SELECTION ET
PRISE EN COMPTE DES INTERACTIONS GENOTYPE x MILIEU 32
4.1 Introduction 32
4.2 Concept de l'adaptation au milieu 32
4.3 Origine génétique de l'adaptation au milieu
34
4.4 Prendre en compte l'adaptation en sélection 36
4.5 La stabilité du rendement . 38
5. 7
MATERIELS ET METHODES 40
41 46 48 51 54 57 62
65
5.1 Caractéristiques pédoclimatiques des sites
expérimentaux
5.2 Conduite des essais
5.3 Analyse des conditions climatiques
5.4 Matériel végétal
5.5 Protocole expérimental
5.6 Suivi et notations
5.7 Analyse des données
6. ANALYSE ET INTERPRETATION DES RESULTATS
6.1 Etude du comportement génotypique intra et inter
sites 65
6.2 Etude des liaisons intra et inter sites entre variables
88
6.3 Etude de l'interaction génotype x environnement
du
rendement 94
6.4 L'adoption des génotypes sur la base du rendement
nominal... 108
6.5 Répétitivité (Reliability) 110
6.6 Appréciation de la qualité technologique par
l'étude de certains
paramètres physico-chimiques 111
6.7 Discussion générale 119
CONCLUSION 126
REFERENCES APENDICES
A. Liste des symboles et des abréviations.
B. Présentation des données des
différents sites d'étude.
C. Données relatives aux variables mesurées.
D. Analyses de la variance.

LISTE DES ILLUSTRATIONS, GRAPHIQUES ET TABLEAUX
Figure 1.2 Le rendement du blé dur en Algérie.
Figure 1.2 Zones céréalières aptes à
la céréaliculture.
Figure 5.1 L'essai répété de rendement de
blé dur 1ère année...
2006/2007 (ITGC Oued Smar).
Figure 5.2 Localisation géographique des quatre sites
d'étude.
Figure 5.3 Variation du cumul pluviométrique par...
site.d'expérimentation durant la campagne 06/07.
Figure 5.4 Variation de la pluviométrie moyenne mensuelle
des...
sites expérimentaux durant la campagne 2006/2007.
Figure 5.5 Variation de la température moyenne mensuelle
des
sites expérimentaux de la campagne 2006/2007.
Figure 5.6 Expérimentation conduite dans les quatre sites
d'étude.
Figure 5.7 Protocole expérimental adopté au niveau
des 4 sites.
Figure 6.1 Variation de la durée de la phase
végétative (en % de la
valeur maximale) Sur les différents sites.
Figure 6.2 Variation de la hauteur du chaume (en % de la
valeur
maximale) sur les différents sites.
Figure 6.3 Variation du nombre de talles herbacées (en %
de la
valeur maximale) sur les différents sites.
Figure 6.4 Variation du nombre d'épis/m2 (en %
de la valeur
maximale) sur les différents sites.
Figure 6.5 Variation de la longueur de l'épi (en % de la
valeur
maximale) sur les différents sites.
Figure 6.6 Variation du nombre de grains par épi (en % de
la valeur
maximale) sur les différents sites.
50
51
70
74
76
78
81
8
16
19
41
45
48
55
56
68
Figure 6.7 Variation du poids de mille grains (en % de la
valeur
maximale) sur les différents sites.
85
88
Figure 6.8 Variation du rendement en grains (en % de la
valeur
maximale) sur les différents sites.
96
Figure 6.9 Performances des génotypes ayant le
meilleur
classement sur l'ensemble des sites en comparaison avec les
performances du témoin Waha et de la moyenne des sites.
102
Figure 6.10 Variation du rendement grain des génotypes
qui
contribuent le plus à l'interaction G x L selon la
fertilité des sites.
105
Figure 6.11 Biplot AMMI1 des effets principaux
génotypes et sites et
de l'interaction de la première composante IPCA1 du
rendement grain.
107
Figure 6.12 Biplot AMMI2 de l'interaction du rendement grain
des 30
génotypes évalués dans 4
localités.
119
Figure 6.13 Rendement nominal des meilleurs génotypes
en fonction
des scores LIPCA1 des 4 sites expérimentaux.
52
Tableau 5.1 Code et pedigree de 30 génotypes
testés dans quatre
sites pédoclimatiques différents en
Algérie durant la campagne 2006-2007 (Benbelkacem, 2007).
53
Tableau 5.2 Caractéristiques des variétés
témoins utilisées dans
l'étude (Boufanar -Z et O. Zaghouane, 2006).
65
Tableau 6.1 Carrés moyens des écarts de
l'analyse de la variance de
la durée de la phase végétative (DVP) et
de la hauteur (HT) de la végétation des 4 sites.
66
Tableau 6.2 Valeurs moyennes caractéristiques de la
durée de la
phase végétative et de la hauteur du chaume
notées sur les différents sites.
71
Tableau 6.3 Carrés moyens des écarts de
l'analyse de la variance de
la capacité de tallage herbacé et du nombre
d'épis/m2 des 4 sites.
72
Tableau 6.4 Valeurs moyennes caractéristiques de la
capacité de
tallage herbacé (TLL) et du nombre
d'épis/m2 (NE) des différents sites.
10
77
77
82
83 90 93 95
97
98
101
Tableau 6.5 Carrés moyens des écarts de l'analyse
de la variance
de la longueur de l'épi et le nombre de grains par
épi des 4 sites.
Tableau 6.6 Valeurs moyennes caractéristiques de la
longueur de
l'épi (LE) et du nombre de grains par épi (NGE) des
différents sites.
Tableau 6.7 Carrés moyens des écarts de l'analyse
de la variance
du rendement en grains et du poids des épis des 4
sites.
Tableau 6.8 Valeurs moyennes caractéristiques du poids de
1000
grains (PMG) et du rendement en grains (RDT) des
différents sites.
Tableau 6.9 Coefficients de corrélation
phénotypiques entre les
variables mesurées par site et pour les 4 sites (en gras
coefficients statistiquement significatifs).
Tableau 6.10 Coefficients de corrélation des rangs de
Sperman de la
même variable mesurée sur les sites
différents (en gras coefficients statistiquement significatifs).
Tableau 6.11 Classement des rendements grainiers des
génotypes
par site, somme des classements par génotype et moyenne
par génotype des rendements des 4 sites.
Tableau 6.12. Analyse de la variance de la régression
conjointe et de l'AMMI (effet moyen additif et interaction multiplicative du
rendement grain.
Tableau 6.13 Rendement moyen (t/ha) et numéro des
génotypes performants par localité.
Tableau 6.14 Moyennes du rendement grain (RDT, t/ha),
coefficient
de régression (b), contribution au carré moyen de
l'interaction (MSGxL), carré moyen de la déviation de
la régression (MSDEV.), ordonnée à l'origine
(a, t/ha) et répétitivité (I) du rendement des
génotypes de blé dur évalués dans 4
localités.
Tableau 6.15 Carrés moyens des écarts de l'analyse
de la variance du 111
taux de mitadinage (MTD) et du taux d'extraction (EXT)
mesurés sur les quatre sites.
Tableau 6.16 Valeurs moyennes caractéristiques du taux
de
mitadinage (MTD) et du taux d'extraction (EXT) notées sur
les différents sites.
11
Tableau 6.17 Carrés moyens des écarts de l'analyse
de la variance du 116
taux de gluten humide (GH) et du taux de gluten sec (GS)
mesurés sur les trois sites (Oued Smar, El Khroub et Sétif).
117
Tableau 6.18 Valeurs moyennes caractéristiques de la
teneur en
gluten humide (GH) et de la teneur en gluten sec (GS)
notées sur les différents sites.
12
INTRODUCTION
Avec 230 kg/hab/an l'algérien se présente comme
l'un des gros consommateurs de céréales, ce qui engendre une
demande qui dépasse les 7.5 millions de tonnes par an [1]. La production
des céréales est, par contre, faible et fluctuante, comme le sont
ses rendements. En effet la faiblesse des rendements est liée à
la présence de contraintes biotiques et abiotiques de nature
intermittente, dont le déficit hydrique est le principal facteur
limitant [2]. Ainsi le rendement en grains peut varier de 0.5 à 3.6
t/ha, sur le même site, au cours des années successives, comme
rapporté par BAHLOULI et al., (2005) [3].
En fait, les faibles rendements sont dus aux effets
combinés des basses températures hivernales, des gelées
printanières, des hautes températures et au déficit
hydrique de fin de cycle. Les variations climatiques influent sur les
conditions de croissance et développement de la plante, ce qui a pour
effet d'affecter le processus de sélection. En effet tous les milieux
n'ont pas la même aptitude à révéler les
différences génotypiques, suite à la présence de
l'interaction.
Cette interaction génotypes x environnement, selon son
amplitude, entraîne des modifications du classement variétal et de
ce fait complique la sélection, vu que chaque environnement aboutit un
classement différent des génotypes. Le choix basé sur la
moyenne générale des milieux n'est plus dans ce cas valable, et
celui par environnement conduit à la sélection d'une multitude de
génotypes, dont la gestion devient onéreuse [4]. L'interaction
génotype x environnement est un problème majeur qui complique
l'interprétation des essais de rendement et rend les recommandations
difficiles, notamment lorsque les génotypes sont
sélectionnés dans un seul environnement et ciblés pour
d'autres environnements [5]; [6]; [7].
L'interaction peut être causée par les
différences de phénologie, de rythme de développement, de
besoins de vernalisation, de réponse à la photopériode, et
aux différences de la capacité des génotypes à
répondre aux conditions de
13
croissance [8]. Selon LECOMPTE et al., (2003) [9], la
connaissance de la nature des réactions ( sensibilité /
tolérance) des variétés aux changements des milieux de
production est la première étape dans la sélection d'un
matériel stable et performant. EBDON et GAUCH (2002) [10] rapportent que
les composantes environnementales sont corrélées avec les
précipitations, les températures moyennes journalières
maximales et minimales, l'altitude, la latitude, la fertilisation azotée
et la teneur en argile.
Dans les programmes de sélection, la signification de
l'interaction génotype x environnement doit être prise en
considération car elle peut être, convenablement, exploitée
d'une manière avantageuse à travers différentes approches.
Son effet par rapport à l'effet du génotype et environnement des
caractères mesurés, doit être estimé. Pour exploiter
positivement l'interaction génotype x environnement, la
régression conjointe et le modèle AMMI (additive models and
multivariate interaction) sont des outils analytiques utiles [11]; [12]; [13];
[14]; [15]. Les résultats de telles analyses peuvent être
utilisés pour grouper des environnements en sous régions sur la
base de la similarité des performances des cultivars [16]; [7].
La présente contribution se propose d'analyser
l'interaction génotype x environnement ainsi que l'identification de la
stabilité du rendement grain en utilisant les données provenant
d'une expérimentation multi- site, de 30 génotypes de blé
dur (Triticum durum Desf.).
14
CHAPITRE 1
LE BLE DUR COMME CEREALE MONDIALE ET CEREALE EN
ALGERIE
1.1 Introduction
Le blé dur (Triticum durum Desf.) occupe,
mondialement, la cinquième place après le blé tendre
(Triticum aestivum.L), le riz (Oryza sativa.L), le maïs
(Zea naydis.L) et l'Orge (Hortodeum vulgare.L) avec une
production de 35.7 millions de tonnes pour la campagne (2006-2007) [17].
Il a été domestiqué au Proche Orient
à partir d'une graminée sauvage (Triticum turgidum ssp.
Dicoccoïdes) qui a ensuite, progressivement, évolué
vers Triticum Turgidum ssp. Dicoccum puis vers Triticum durum
(blé dur cultivé). Les premières cultures apparaissent au
VIII millénaire avant J-C en Mésopotamie et dans les
vallées du tigre et de l'Euphrate (aujourd'hui l'Irak) dans la
région du croissant fertile [18].
1.2 Le blé dur : situation mondiale en chiffre
Sur la scène mondiale, la superficie moyenne
consacrée annuellement à la culture du blé dur,
s'étend sur environ 18 millions d'hectares, avec une production annuelle
moyenne approximative de 30 millions de tonnes [19].
L'union européenne est dans le monde, la
1ère région productrice de blé dur. La
production est concentrée en Italie, l'Espagne, la France et en
Grèce. La production de blé dur en 2007 a été
estimée à 8,3 millions de tonnes quant aux superficies, elles
étaient estimées à 3 millions d'hectares [17]. Cependant,
par rapport à la moyenne des 5 dernières années, celles-ci
ont diminué de 18,9%. Cette baisse est due à une nette diminution
des superficies Italienne et Espagnole au cours de cette période
[17].
15
Le Canada est le second producteur mondial de blé dur
en 2007 avec 1,95 millions d'hectares (Mha) consacré aux producteurs
canadiens de l'Ouest, soit 27% de plus qu'en 2006. Les rendements en 2007
étant estimés par statistique Canada à 1,87 t/ha [20].
La Turquie classée en troisième position
après l'Union Européenne (UE) et le Canada, a produit en moyenne
3.1 millions de tonnes (Mt) au cours des cinq dernières années
[17].
La production céréalière de l'Afrique du
Nord dépend largement des pluies hivernales, qui sont souvent
imprévisibles, par conséquent, la production de blé dur a
été des plus variables au cours de la dernière
décennie. Celle-ci a oscillé, entre 5,6 Mt, en 1996-1997 et 1,7
Mt, en 2000-2001. La production en 2006-2007 atteint 5,5 Mt [21].
Concernant l'Algérie, certaines superficies en
système intensif et soutenues dans le cadre du PNDA (Plan National de
Développement Agricole) dépassent souvent les 20 quintaux, alors
que dans d'autres qui se trouve dans la même zone ou dans des zones non
répertoriées comme zones céréalières, la
production est moins importante.
Pour le blé dur, la production nationale est
passée de 12.389.000 quintaux en 2001 à 15.521.000 quintaux en
2006, soit un accroissement de niveau de productivité de 45.38%. Comme
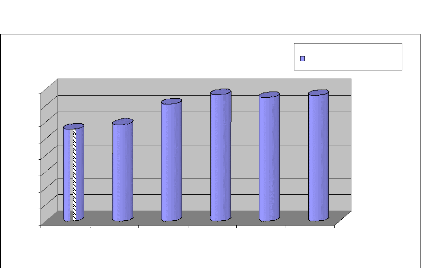
nous pouvons le voir sur la figure (1.1). Les rendements moyens
enregistrés, au cours de la période 2000-2006, ont connu une
progression dans le temps, passant de 11.14 q/ha en 2000-2001 à 15.2
q/ha en 2005-2006 (DSASI, 2007) [22].
16
16
14
12
10
8
6
4
2
0
Années
Rendement (q/ha)
2000-2001 2001-2002 2002-2003 2003-2004 2004-2005 2005-2006
Rendement (q/ha)
Figure 1.1 : Rendement de blé dur en Algérie (2001-
2006) [22] 1.3 Historique de recherches sur les blés en
Algérie
Au temps de l'occupation romaine, l'Afrique du Nord
fournissait à la capitale de l'empire une certaine quantité de
grains, représentant les impôts en nature versés par les
particuliers, ou les redevances des sociétés foncières, a
tel point que l'expression de « Grenier de Rome » s'est
imposée pour désigner l'Afrique du Nord [23].
Avec l'arrivée des arabes et plus
particulièrement pendant les règnes des dynasties
algériennes des Beni Ziri, des Beni Hammad et Beni Ziyane, un certain
élan fut donnée à l'agriculture en général.
Après l'effondrement du royaume Zianide , sous l'administration turque,
la culture du blé devient tellement importante que le gouvernement
algérien a entretenu des relations commerciales fructueuses avec les
différentes républiques italiennes et françaises. En
effet, à cette époque le blé Algérien sauva le
peuple français de la famine pendant les années terribles du
blocus, que fit subit l'Angleterre à la France, durant la guerre
Napoléon [24].
Jusqu'à 1830, l'Algérie n'avait pas
importé de blé pour sa subsistance. DUCELLIER, 1930 [25] a
décrit l'ensemble des espèces de blés cultivées en
Algérie, pour le blé dur barbu, vingt neuf variétés
ou populations ont été
17
mentionnées. Plus de 30 années après les
travaux de Ducellier, les mêmes variétés cultivées
de blé dur ont été mentionnées [23].
Il apparaît que les ressources génétiques
des blés étaient fortement diversifiées non seulement
à travers le nombre de variétés ou population
cultivées mais aussi et surtout à travers la très grande
diversité génétique au niveau de chaque population. Ces
populations de terroirs souvent très bien adaptées aux conditions
du milieu permettaient certainement de répondre aux
préoccupations et aux besoins locaux. Les pressions de sélections
appliquées localement à ce matériel ont permis de
maintenir un certain progrès génétique régulier
mais assez lent [26].
L'introduction de variétés
étrangères de blé dur et des autres céréales
en général, n'a été entreprise qu'au cours de la
campagne 1969-1970 par l'INRAA au niveau de CNRA (Centre National de Recherches
Agronomiques) d'El-Harrach et dans les stations régionales (Sidi
Bel-Abbès, El khroub, Sétif et Guelma) [27].
Au cours de cette période l'amélioration
génétique était à ses débuts et se confinait
dans la sélection massale ou dans le développement de cultivars
issus de croisements à l'intérieur de ces populations dont
l'objectif était l'obtention des plantes mieux adaptées aux
différentes conditions de culture [27]. La collaboration du CIMMYT
(centre international pour l'amélioration du blé et du maïs)
a été total et a rendu possible l'atteinte de nos objectifs.
Au courant des années 1980 l'ICARDA (centre
international pour la recherche agronomique en zones sèches) a
participé au développement de la recherche des grandes cultures
avec l'ITGC [27].
En 1995, l'ITGC a dressé un catalogue des principales
variétés de céréales (blés, orge, avoine,
triticale) cultivées en Algérie. Il est important de mentionner
que plusieurs variétés introduites et sélectionnées
localement ont été développées et parfois les noms
ont été algérianisés [26].
18
Pour le blé dur, 12 variétés sont
actuellement en productions, dont 5 dites améliorées, ont
été introduites depuis 1980. Sur l'ensemble de ce matériel
végétal en production, les variétés Hoggar (Vitron)
et Waha sont les plus demandées sur le marché. En effet, 65 % de
la superficie semencière totale était occupée que par les
deux variétés [28].
Depuis 2004, de nouvelles variétés performantes
ont été homologuées et introduites dans le programme de
multiplication de semences, il s'agit des variétés Cirta,
Gta/Dur... et Boussellem. En 2006, 3 autres variétés ont
été homologuées (ciccio, cannizio et colosseo) [28].
1.4 Aire de production
Le blé dur est cultivé à travers
l'ensemble des zones agro-écologiques de notre pays, mais il est
essentiellement localisé dans les régions semi-arides et
même arides et donc soumises aux aléas climatiques qui
pénalisent fortement les niveaux de productivité [29]. La
superficie globale emblavée durant la période 2000-2006
était de 3.177.500 hectares. Sur ce volume, le blé dur occupait
une superficie moyenne de 1.355.667 hectares [22].
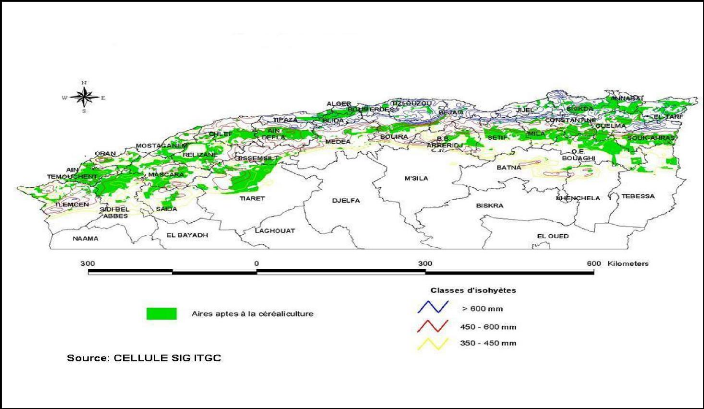
Dans l'ensemble, la sole réservée à la
production céréalière est concentrée dans trois
grandes zones agro climatiques [30] (Figure 1.2) :
? La zone humide à sub-humide incluant les
régions littorales et sub-littorales du centre du pays vers l'Est
caractérisée par une pluviométrie annuelle de 600 mm
permettant une culture intensive;
? La zone semi aride incluant les plaines telliennes
orientales et occidentales
avec une pluviométrie annuelle moyenne variant entre 350
et 500 mm;
? La zone sub-aride qui se confine aux hauts plateaux se
caractérise par une faible pluviométrie (200-350 mm) avec un
système de culture à prédominance agropastorale avec des
altitudes dépassant les 1000 m associés à des stress
biotiques sévères (30 à 40 jours de gel et plus de 40
jours de sirocco par année en moyenne) [31].
19
Figure 1.2 : Zones céréalières aptes
à la céréaliculture
1
20
1.5 Qualité et différentes formes
d'utilisation de blé dur en Algérie
Des grains de céréales et leurs
dérivés représentent l'apport principal en cadres de
l'alimentation humaine [32]. Le blé dur est l'espèce la plus
importante qui se distingue du blé tendre par un ensemble de
caractéristiques, son albumen est vitreux et contient plus de
protéines, donc moins d'amidon. Moulu, il garde une texture granuleuse
(la semoule) qui fait que les pâtes sont fermes, peu collantes et
résistent à la cuisson [33].
De plus le blé dur est riche en pigments
caroténoïdes qui donnent au produit fini, cette couleur
ombrée qui répond à l'aspect esthétique que
recherche le consommateur. Et grâce à ses caractéristiques
nutritionnelles et technologiques, le blé dur se prête à la
fabrication d'une gamme très variée de produits alimentaires dont
les pâtes en constituent la base pour une grande partie du monde, le
couscous et le Frik en Afrique du Nord, El-burghul au Moyen Orient.
En Algérie, le blé dur est consommé sous
plusieurs formes dont essentiellement le couscous, la galette, les pâtes
alimentaires et le Frik [33].
21
CHAPITRE 2
LES FACTEURS GENETIQUES ET ENVIRONNEMENTAUX ASSOCIEES
AUX
PROCESSUS DE CROISSANCE ET DE DEVELOPPEMENT
2.1 Introduction
Les variations du rendement et de ses composantes ne
s'expliquent pas par le niveau des facteurs ou conditions qui sont à un
niveau suffisant, mais seulement par les facteurs limitants [34].
Les caractéristiques climatiques des zones
céréalières sont telles que la culture du blé dur
est exposée à divers stress qui réduisent des rendements
escomptables et engendrent la variabilité de la production d'une
année à l'autre et d'une région à l'autre [35];
[36].
2.2 Les facteurs climatiques
2.2.1 Les précipitations
Dans les régions semi-aride du sud de la
méditerranée y compris l'Algérie, la faiblesse des
précipitations et leur distribution aléatoire se traduisent
souvent par une situation de contrainte hydrique présente pratiquement
tout au long du cycle de développement du blé [37], [38]. En
raison de son importance économique, la production de blé dur
dépendra beaucoup des études consacrées pour une meilleure
compréhension de la sécheresse qui touche environ 40% des terres
cultivées au monde [39]. Ces situations de stress hydriques ou de
sécheresse, présente des intensités et des
fréquences variables selon les régions et les années.
En faite, la sécheresse est définie comme une
absence de pluie durant une période de temps donnée, affectant
par conséquent l'alimentation hydrique des plantes. La sécheresse
permanente ou temporaire limite le développement, la distribution de
végétation naturelle et le rendement des plantes cultivées
plus que tout autre facteur [40]. Selon son intensité et ses dates
d'apparition au cours
22
du cycle de développement des céréales,
le stress hydrique peut causer des dégâts à des
degrés variables en fonction des variétés cultivées
et de la date de semis [41]; [42]; [43]; [44].
La sécheresse au début de cycle affecte
l'installation de la culture, quand le déficit hydrique apparaît
pendant la période de tallage herbacé, la vitesse
d'émission des talles diminue et si le déficit s'accroît
sévèrement, il y a arrêt du tallage [45]. Une
sécheresse importante survenant pendant la période
d'émission des épillets soit approximativement de la mi tallage
au stade 1 à 2 noeuds limite leurs nombre final surtout dans les zones
semi aride et plus tard affecte la formation des fleurs, donc pénalise
le nombre de grains potentiels de l'épi [46].
Un rupture de l'alimentation en eau pendant la phase de
remplissage des grains engendre un ralentissement de la vitesse de migration
des réserves vers le grain : il y a échaudage et par
conséquent des diminutions de poids de 1000 grains [46].
La résistance à la sécheresse d'un
génotype s'évalue par sa capacité à maintenir un
rendement acceptable en conditions de déficit hydrique. Il est,
cependant, difficile de sélectionner directement pour la
résistance à la sécheresse car le contrôle
génétique de ce caractère quantitatif est très
complexe [47].
La sécheresse de fin de cycle est pratiquement quasi
présente au cours de ces dernières années et les efforts
en matière d'amélioration génétique sont
orientés vers la création de variétés
précoces afin d'échapper à ce type de stress.
2.2.2 La température
La température est un principal moteur de
développement, une température de niveau identique n'aura pas la
même efficacité sur la vitesse du développement en hiver ou
au printemps, compte tenu de la variation de la durée du jour sous
l'effet des saisons [46]. Selon la sensibilité variétale, le
seuil thermique de mortalité varie entre -12 et -16 [48]. De plus le
blé est moins sensible à la température durant sa phase
végétative par rapport à sa phase reproductive [49].
23
En effet, plusieurs études ont signalé
l'existence de différences génotypiques en réponses au
stress thermique [50]; [51].
La culture du blé dur, sur les hauts plateaux, est
essentiellement sujette aux effets des basses températures de la
levée au stade épiaison [52]. En effet, le froid peut gêner
la sortie des épis ; cette déformation est temporaire et sans
effet sur le rendement [46]. Cependant, les basses températures au stade
floraison induisent une stérilité des gamètes mâles.
Contrairement aux basses températures survenant au même stade, le
stress thermique induit surtout une stérilité femelle. Lorsque le
grain commence à ce développer, les fortes températures
excessives, supérieures à 25°C accentuent le processus
d'avortement des fleurs [45].
Dans les situations de fin de cycle où les fortes
températures sont fréquentes, il existe un grand risque
d'échaudage des grains. Ce risque est d'autant plus grand, lorsque les
hautes températures persistent [29]. En effet, l'augmentation des
températures accélère la vitesse de croissance des grains
et réduit leur durée de remplissage [53].
2.2.3 Le gel
Des dégâts dus au gel d'hiver dépendent
bien évidemment de la sensibilité variétale, mais aussi du
stade de développement. Aussitôt après la germination, le
blé est particulièrement sensible au gel [48].
Si le gel survient brutalement (forte amplitude thermique),
les tiges qui ont amorcé leur montée sont détruites. Ces
tiges correspondent à celles qui sont aux stades les plus
avancés, les maîtres brins et les premières talles
primaires. Elles correspondent aussi aux talles les plus productives comportant
un nombre supérieur d'épillets et de fleurs par épillets.
Détruites par le gel, ce sont les talles plus jeunes et donc moins
productives qui constitueront le peuplement épis final [45].
24
Le gel marque son effet principalement sur la composante
nombre de grains par épi [45]. Les zones arides et semi arides qui sont
très exposées aux risques précoces de sirocco ainsi qu'au
gel printanier [54]. Dans ces zones des gelées tardives
accompagnées de brouillard, des carences nutritionnelles peuvent
entraîner la non fécondation des épillets terminaux, ainsi
que des 2ème et 3ème fleurs de chaque épillet.
Les étamines ne s'ouvrent pas, une température constamment
inférieure à 15°C pendant la fécondation produit ce
phénomène appelé « coulure » [48].
Il est possible d'y remédier en évitant le choix
de variétés trop précoces pour la région.
2.3 Les facteurs agro-techniques
Les espèces blé et orge, prospèrent sur
une gammes assez variées de sols et l'optimum semble être des
terres neutres, profondes et de texture équilibrée. En sol peu
profond, le rendement en grains des céréales est
pénalisé [55]. La structure, grumeleuse stable ; assure une
pilosité suffisante. Le complexe absorbant important permet à la
plante de se nourrir à partir des réserves chimiques du sol. Ces
critères ne sont pas toujours respectés [48].
La réussite de l'installation de la culture
nécessite la maîtrise du travail du sol et du semis. En effet, les
rotations des cultures permettent l'amélioration de la fertilité,
la structure du sol et le contrôle des maladies, des insectes et des
adventices. La bonne préparation de lit de semences s'avère
fondamentale pour assurer les meilleures conditions de températures,
d'humidité et d'aération aux semences en germination [29].
La date de semis est un facteur important dans la formation du
rendement. Les semis précoces augmentent le rendement, mais notamment en
zones semi arides et arides, peuvent aussi entraîner un risque de
sécheresse qui peut causer la mort des plantules [56]. Les semis tardifs
exposent la phase de formation des grains aux hautes températures qui
accélèrent la maturité des graines avant leurs
remplissage, donnant des graines de petites tailles et moins remplies, et
ainsi
25
un rendement assez bas [57]. Dans ce cas, il est
impératif d'augmenter la densité de plantes pour pallier le
manque de tallage herbacé, et maintenir ainsi le rendement [45].
Dans les zones sub-humides inférieures à 300
mètres d'altitude, avec plus de 600 mm de pluie par an, la culture du
blé souffre encore de plusieurs carences techniques qui peuvent
s'expliquer par le non respect de l'itinéraire technique par les
agriculteurs de la zone. Ainsi le semis se fait encore et le plus souvent
à la volée, le désherbage chimique est faiblement
pratiqué ou appliqué dans de mauvaises conditions (choix de
l'herbicide et stade des adventices au moment de l'application), alors que la
fertilisation azotée de couverture est conditionnée par les
pluies de fin d'hiver [58].
En parallèle, les travaux conventionnels de travail du
sol ne sont plus adaptés dans les zones où la conservation de
l'eau et des sols est prioritaire [29]. La valorisation des eaux pluviales est,
donc, un facteur déterminant de la production. Les innovations
technologiques, visant à améliorer la production, doivent
réduire des risques climatiques, en maximisant l'infiltration et
l'utilisation des eaux de pluies, améliorer la fertilité des sols
et réduire les pertes du patrimoine sol. Dans la mesure où, suite
à la nature méditerranéenne du climat, 70% des pluies sont
enregistrées en hiver, les techniques culturales doivent assurer aussi
le stockage du maximum de cette eau dans le profil sous climat semi-aride
[59].
De même, il est possible de produire sans labourer dans
le contexte de l'aridité méditerranéenne tout en assurant
la préservation des ressources naturelles et de l'environnement [60];
[61]; [62]; [63]; [64].
2.4 Les facteurs liés à la
variété d'origine génétique
Elle consiste en une période de basses
températures dont ont besoin les céréales d'automne pour
l'initiation florale. Sachant que la plupart des variétés
de
2.4.1 La vernalisation
26
céréales (blé et orge) cultivées
au Maghreb sont de type printemps, leurs besoins en vernalisation sont nuls
[29].
2.4.2 Exigence en durée du jour
Le blé dur est une espèce dite de jours longs
préférentiels ce qui signifie que les jours longs favorisent la
vitesse d'apparition des stades liés au développement floral. Il
faut que la durée d'éclairement soit d'environ 12 heures pour que
l'épi commence à monter dans la tige [48].
Le rendement final est souvent déterminé par les
conditions prévalentes dans le milieu de production et surtout celles de
la période de remplissage du grain. Ainsi, sous conditions de stress
hydrique et thermique sévères en fin de cycle, les
génotypes photo insensibles se comportent relativement mieux que ceux
qui sont photo périodes dépendants. Alors que sous les conditions
de fin de cycle plus clémentes, les génotypes sensibles à
la photopériode produisent plus à cause d'une durée de
cycle plus longue [65].
La manipulation de la sensibilité à la
photopériode est un objet très important en amélioration
des céréales afin d'optimiser la date d'épiaison et de
maturité à la variabilité climatique du milieu de
production [66].
27
CHAPITRE 3
L'APPRECIATION DE LA VALEUR PHENOTYPIQUE EN
PRESENCE
D'INTERACTION GENOTYPE x MILIEU
3.1 Introduction
La meilleure façon de caractériser un milieu,
c'est d'y décrire les performances phénotypiques d'un ensemble de
génotypes et la meilleure façon de décrire un
génotype c'est de décrire la variabilité de son
comportement au travers des différents milieux. Pour comprendre les
relations génotypes/milieu, il faut déployer le génotype
dans différents milieux, mais aussi le milieu sur différents
génotypes [67].
3.2. Le génotype et le phénotype
Le phénotype d'une plante correspond à ses
performances dans un milieu et pour un niveau d'observations donné. Il
est le résultat du « dialogue » entre la plante et son milieu.
L'étude de ce dialogue représente le domaine de la physiologie
végétale, au sens large, même lorsque celui-ci se situe au
niveau moléculaire. Une plante reçoit des signaux du milieu
où elle vit, elle les interprète en fonction de son
génotype et il en résulte une réaction qui est ce que nous
voyons ou mesurons selon le niveau d'observation : c'est le phénotype.
Un génotype peut être défini comme un arrangement de
gènes (dans un cytoplasme particulier) et un phénotype sera la
valeur prise par cet arrangement pour un caractère donné (et donc
un type de mesure) et dans un milieu donné [67].
Dans le cas d'un caractère contrôlé par un
nombre réduit de gènes et pour lequel les expressions dans un
milieu peuvent être réparties sans problème en classe, il
est possible d'identifier les unités de ségrégations en
cause et d'en attribuer certains arrangements à des plantes. Ces
arrangements forment le génotype. Le caractère est alors dit
qualitatif, et l'on parle de gènes à effets majeurs. De tels
gènes sont connus pour de nombreux caractères
28
(résistance aux maladies, nanisme,
stérilité mâle...). Les conclusions peuvent dépendre
du niveau d'étude et les expressions phénotypiques de
différents génotypes peuvent être variables d'un milieu
à l'autre (cas d'interaction génotype x milieu pour des
caractères qualitatifs). Dans ce cas un génotype pourra
être identifié par l'ensemble des expressions d'un même
arrangement de gènes [67].
S'il y a risques d'erreur de répartition des
phénotypes en classes, le caractère est dit quantitatif. Ce sera
le cas avec un fort effet du micro-milieu quel que soit le nombre de
gènes en cause, ou avec un grand nombre de gènes en cause quels
que soit les effets du micro-milieu. Avec l'intervention d'un grand nombre de
gènes, un génotype correspond alors à un arrangement
inconnu de gènes, à la limite c'est l'individu [67].
La plupart des caractères agronomiques que l'on cherche
à sélectionner ne dépendent pas de l'expression d'un ou
deux gènes, mais de plusieurs gènes dont les effets individuels
sont « mineurs » ont dit qu'ils sont gouvernés par des
ensembles polygéniques ou bien que leur hérédité
est quantitative. Ces caractères sont par exemple : la
précocité, le rendement en grains [18].
Le phénotype d'une plante est donc bien
résultante visible de l'expression des gènes de cette plante en
interaction avec le milieu extérieur dans lequel elle se trouve. Selon
une image de JACQUARD (1978) [68], le génotype c'est la partition, le
phénotype le résultat de l'interprétation par le
milieu.
3.3 Concept de l'interaction génotype x milieu
Lorsqu'un caractère est mesuré sur un individu,
la valeur observée représente la valeur phénotypique de
cet individu et toutes les observations doivent être fondées sur
la mesure des valeurs phénotypiques. Dans ce cas, les caractères
quantitatifs, dans leur expression phénotypique sont la
résultante en fait de plusieurs facteurs génétiques et non
génétiques. De ce fait, la variabilité phénotypique
(Vp) est égale à la somme suivante : [67].
(Vp) = VG + V (G ×E) + VE
29
Où (VG) : la variance génétique, (VE) :
la variance environnementale et V (G ×E) : l'interaction génotype
× environnement. Inclure tous les facteurs d'ordre non
génétique dans le terme « environnement » signifie que
le génotype et l'environnement sont par définition les seules
causes qui déterminent la valeur phénotypique [67].
La variance génétique (VG) peut être
elle-même décomposée en variance additive (VA), due
à l'effet moyen des gènes, en variance de dominance (VD) due aux
interactions entre allèles aux même loci et en variance
d'épistasie (VEP), due aux interactions entre allèles à
des loci différents. L'expression de la variance phénotypique
devient :
(VP) = (VA) + (VD) + (VEP) + (VG x E) + (VE)
La variance due au milieu qui par définitions, comprend
toute la variation d'origine non génétique, peut avoir beaucoup
de causes différentes et sa nature dépendra beaucoup du
caractère et de l'espèce étudiée. D'une
façon générale, la variance due au milieu est une cause
d'erreur qui réduit la précision des études
génétiques [67].
L'interaction génotype x environnement
représentent, donc, la variation quantitative et qualitative, entre
divers environnements, des différences entre génotypes.
L'interaction perturbe en fait l'association que l'on peut faire entre valeur
phénotypique et valeur génotypique d'un caractère [69].
C'est l'interaction génotype x environnement qui permet d'expliquer que
certains génotypes soient plus performants que d'autres pour un type
d'environnement donné.
L'interaction GxE peut prendre plusieurs aspects [70] :
- soit qu'une quelconque différence spécifique
du milieu exerce sur certains génotypes un effet plus important que sur
d'autres ;
- soit qu'il y ait un changement dans le classement d'une
série de génotypes que l'on mesure sous différents
milieux, c'est-à-dire que le génotype A qui est supérieur
au génotype B sous le milieu X peut lui être inférieur sous
le milieu Y.
30
3.4 Elaboration du rendement et corrélation
entre caractères
L'amélioration des plantes est pratiquement toujours
multicaractère. Elle l'est d'un point de vue économique, mais
aussi d'un point de vue biologique. En effet, il s'agit de transformer la
plante dans son ensemble. Compte tenu du caractère intégré
de la plante, la transformation sur un caractère entraîne souvent
des modifications d'autres caractères [67]. La connaissance des
relations existantes entre caractères permet donc de mieux transformer
les plantes et comprendre son fonctionnement dans un milieu donné [71];
[72]; [73].
Le rendement est la résultante des effets des
caractères impliqués directement et indirectement dans sa
formation tels que le poids de 1000 grains, le nombre d'épis, le nombre
de grains par épi, la précocité d'épiaison... [74].
Chaque composante s'élabore pendant une phase bien définie et
porte l'empreinte positive ou négative des effets des milieux [75];
[76].
Dans certains cas, l'occurrence de stress hydrique à
différents stades de développement et la présence de
hautes températures de fin de cycle nécessitent la participation
de toutes les composantes pour l'élaboration du rendement car dans ces
conditions la compensation entre les composantes devient faible [41].
En Outre, les composantes du rendement sont
considérées comme des unités à partir desquelles un
bon rendement peut être obtenu [77]. Plusieurs chercheurs rapportent
qu'il est plus facile d'améliorer le rendement grain par le biais de ses
composantes [78] ou de certains caractères morphologiques, surtout dans
les milieux difficiles, qui peuvent servir de critère de
sélection [79]; [80].
D'après FISHER, 1978 [81], le nombre de
grains/m2 est un caractère étroitement lié avec
le rendement en grains, sous différents lieux et pour différents
génotypes. Ce caractère est corrélé souvent
positivement au rendement en grains des céréales d'automne dans
les zones semi arides connues par une sécheresse de fin de cycle [82];
[83]; [84].
31
Dans les régions connues par le gel printanier, en
l'absence de sécheresse de fin de cycle, le poids de mille grains
constitue la principale composante du rendement [85]; [86]. Pour une
variété donnée à partir d'un certain nombre de
grain seuil, le PMG est corrélé négativement au nombre de
grains/m2 [87]; [88]. Ce dernier résultat est la
conséquence d'une compétition qui s'établit entre les
grains pour les assimilats, ce qui engendre souvent la chute du PMG.
En Algérie, la période de remplissage des grains
est souvent confrontée à des contraintes environnementales en fin
de cycle et plus précisément au déficit hydrique et aux
hautes températures. Ceci a pour conséquence de provoquer des
pertes de poids des grains et plus particulièrement chez les
variétés à cycle long et dans le cas de semis tardif.
En conditions de sécheresse, la stabilité
d'expression de la hauteur est importante. Dans ces conditions, une paille
haute est plus apte à stocker plus de réserves glucidiques, qui
sont susceptibles d'être transférées vers le grain au cours
de la phase de remplissage [89].
DOUMEZ et al., (2001) [90], rapportent des gains
génétiques significatifs dans le temps suite aux effets de la
sélection pour le rendement en grains et la durée du cycle. Ils
suggèrent que les composantes du rendement qui se forment au cours de la
phase végétative sont celles qu'il faut améliorer.
LAFITTE et COURTOIS (2002) [91], mentionnent parmi d'autres
caractères, la durée de la phase végétative comme
une source de l'interaction génotype x environnement du rendement en
grains du blé. Les génotypes précoces sont plus productifs
sous conditions de contraintes hydrique et thermique de fin de cycle.
L'idéal pour atteindre les rendements proches du
potentiel est l'élaboration de chacune des composantes de rendement dans
les meilleures conditions. Cependant, la grande variabilité
édapho-climatique de nos régions et celle du matériel
génétique justifie la fluctuation des rendements et de leurs
composantes [29].
32
CHAPITRE 4
L'ADAPTATION AU MILIEU COMME OBJECTIF DE SELECTION ET
PRISE EN COMPTE DES INTERACTIONS GENOTYPE x MILIEU
4.1 Introduction
L'adaptation au milieu est un objectif de sélection
recherché pour un grand nombre d'espèces végétales
et elle fait le plus souvent appel à l'analyse du rendement.
L'améliorateur peut rechercher des génotypes présentant
une "adaptation spécifique", c'est-à-dire une adaptation à
des milieux spécifiques, ou au contraire une "adaptation
générale" à des conditions de milieux variées
[92].
4.2 Concept de l'adaptation au milieu
L'adaptabilité au milieu est un phénomène
essentiel pour que la culture puisse s'exprimer et produire tant soit peu ou
à la limite survivre et se reproduire. L'adaptation est donc synonyme de
survie, mais elle est définie aussi comme la capacité d'une
plante à croître et à donner des rendements satisfaisants
dans les zones à des stress de périodicités connues [93].
Cependant, l'adaptation à des milieux fortement variables ne peut
être qu'un compromis entre les conditions climatiques qui
prévalent au cours d'une année donnée et la
réactivité de la variété (ou l'espèce)
à ces conditions de croissance [94].
Pour une durée d'adoption donnée, une
variété (ou espèce) est dite adaptée si elle
réussit à donner de meilleurs rendements que la
variété (ou l'espèce) qu'elle remplace et ce le plus
fréquemment possible. La comparaison des performances, dans le temps,
des espèces et variétés peut donner des indications sur
les capacités d'adaptation et de performances de ces dernières.
Cette adaptation est à lier à la variabilité climatique,
notamment la distribution des pluies, la demande climatique et la
température du lieu de production qui conditionnent le
développement et la production du végétal [94].
33
La notion d'adaptation se confond parfois avec celle de la
résistance et de tolérance au stress. En fait l'adaptation n'est
que la résultante de la tolérance aux contraintes. Une plante
adaptée est donc celle qui tolère ou résiste à un
stress donné et réussit à produire à un niveau
satisfaisant par rapport à une autre plante qui sera dite non
adaptée [74]; [95].
L'adaptation regroupe, en fait, deux notions distinctes :
L'adaptation générale qui représente la
capacité que possède un génotype à valoriser des
conditions de milieux variées, et l'adaptation spécifique qui
représente la capacité d'un génotype à valoriser
des conditions particulières [92];[96].
L'adaptation générale, parfois appelée
adaptabilité, est conférée par une adaptation
simultanée à un ensemble de contraintes du milieu, telles que le
froid, la sécheresse, le manque d'eau..., etc. C'est en quelque sorte
une somme d'adaptations spécifiques. Ainsi, l'adaptation
générale s'observe le plus souvent en conditions naturelles dans
des réseaux d'expérimentation regroupant un ensemble de milieux
sur plusieurs années, les "réseaux multilocaux et pluriannuels".
La notion d'adaptation est à replacer dans le contexte des interactions
génotype × milieu car des variations d'adaptation se traduisent par
des interactions génotype × milieu [97].
Les génotypes adaptés à une large gamme
de milieux sont particulièrement intéressants, mais ils
présentent généralement des performances plus faibles que
celles des génotypes adaptés à des environnements plus
spécifiques. A l'opposé, si l'adaptation spécifique peut
apparaître comme un frein au développement d'une
variété/lignée, car son créneau de
développement est limité à un ensemble d'environnements
réduits, ces génotypes ont toutes les chances d'être
préférables aux autres dans ce type de conditions, du fait de
leurs niveaux de performances plus élevés [98].
Les causes de l'adaptation plus au moins bonnes d'une
variété à un type d'environnement se trouvent dans le
patrimoine génétique de chaque génotype. Pour le
rendement, les facteurs de régularité tels que la
résistance aux maladies,
34
à la verse, ou au froid ont permis l'émergence
de génotype dont les niveaux de rendement sont stables que ceux des
variétés ne présentent pas ces caractéristiques
[99].
4.3 Origine génétique de l'adaptation au
milieu
L'adaptation d'un génotype au milieu peut avoir deux
origines : la présence de "gènes d'adaptation", ou plus
exactement de gènes qui gouvernent des caractères jouant un
rôle dans l'adaptation, et le pouvoir tampon lié à la
structure génétique de certains génotypes [96]. L'effet de
la structure génétique peut se manifester soit par
l'hétérozygotie conférée par l'état hybride,
soit par l'hétérogénéité du peuplement
cultivé (par exemple, une population, un mélange d'hybrides, une
variété multilignée, une variété multiclone,
etc.) [97].
Les gènes d'adaptation spécifique sont nombreux
: ils peuvent concerner l'évitement de la contrainte (gènes de
précocité) ou bien l'adaptation à la contrainte
elle-même (résistance au froid hivernal, etc.) ou à l'agent
pathogène [97]. Cette résistance au stress se manifeste sous la
forme d'un ensemble de mécanismes qui induisent chez la plante une
capacité à accepter la contrainte mais sans subir les effets.
C'est la résistance génétique. Elle permet à la
plante de résister à la déformation mécanique,
à la dégradation membranaire et de maintenir ses activités
métaboliques sous des niveaux de contrainte assez élevés.
Les génotypes performants en conditions favorables perdent cette
capacité sous conditions de contraintes sévères. Il y a
alors séparation de la capacité d'adaptation et celle de
productivité [100]; [95].
La supériorité de la structure
hétérozygote par rapport à la structure homozygote a
été étudiée depuis longtemps chez beaucoup
d'espèces. L'effet de l'hétérozygotie sur la
stabilité du rendement varie entre les espèces et dépend
de leur système de reproduction [101]. L'effet est fort pour des
espèces comme le maïs, le seigle ou le tournesol. Il est plus
faible pour les espèces partiellement allogames comme le colza, la
féverole ou le sorgho. Enfin, il n'a pas été mis en
évidence de supériorité de la structure
hétérozygote chez des espèces autogames comme le
blé [101] ou le pois [102].
35
La supériorité de l'hybride est due au fait
qu'il permet de réunir dans un même génotype un plus grand
nombre de gènes, allèles ou non, codominants ou dominants
favorables contrôlant l'adaptation à différents milieux
[96].
Les avancées de la biologie moléculaire,
notamment la recherche de QTL (Quantitative Trait Loci ou
régions du génome associées à l'expression «
quantitative » de certains caractères agronomiques) et la
possibilité nouvelle de séquencer le génome d'une
espèce afin d'en faire l'inventaire des gènes, devraient
permettre d'identifier des gènes d'adaptation.
Dans une revue bibliographique consacrée à
l'identification de QTL influencés par le milieu [103], plusieurs
travaux mettent en évidence une interaction QTL/milieu. La
détection de ces interactions QTL/milieu devrait représenter les
effets génétiques responsables de l'interaction génotype x
milieu.
Concernant le blé dur, des derniers travaux de
coopération entre l'INRA Maroc, les institutions françaises
(Inra, Montpellier Sup Agro) et international (ICARDA) ont permis l'isolement
et la caractérisation de nouveaux gènes de blé dur
associés à la tolérance à la sécheresse, il
s'agit de la famille de gènes (LEA).Les protéines Late
Embryogenesis Abondant (ou LEA) sont classées par familles. Parmi ces
familles se trouve le groupe des (déhydrines) pour lequel une
corrélation avec le caractère de tolérance à la
sécheresse a été mise en évidence. Plusieurs clones
codant pour ces déhydrines ayant été isolés au
cours de ces travaux, ils ont servi d'amorces pour la prospection de nouveaux
clones présentant des identités des séquences avec ces
déhydrines [104].
Il s'agissait ainsi de caractériser les gènes
exprimés lors d'un déficit hydrique, de préparer les
banques d'ADNc à partir de quelques génotypes
sélectionnés en conséquence puis de les séquencer
afin d'en déduire les protéines correspondantes. Le programme
prévoyait ensuite la cartographie des gènes reconnus,
l'identification des loci (QTLs) responsable de la variabilité de la
tolérance à la sécheresse et enfin le développement
des marqueurs génétiquement associés au caractère
étudié [104].
36
Ce projet de coopération a permis un transfert de
biotechnologie adaptée à l'amélioration
génétique du caractère "Tolérance au stress
hydrique" chez le blé dur.
4.4 Prendre en compte l'adaptation en sélection
Quelle que soit l'adaptation recherchée,
spécifique ou générale, le sélectionneur peut
être amené à optimiser le dispositif expérimental
surtout pour les expérimentations en conditions non
contrôlées. D'une part, cette optimisation devra considérer
des aspects liés au milieu. Le sélectionneur peut en effet devoir
choisir un nombre de milieux représentatifs d'une région, d'un
pays, etc. Pour établir ce choix, il faut tenir compte de la
répétitivité des mesures de l'adaptation, de l'importance
des effets dus à l'année et des effets dus aux lieux et de la
possibilité de réduire le nombre de répétitions.
D'autre part, l'optimisation devra tenir compte des aspects liés
à l'hérédité du caractère lui-même
(héritabilité). Par exemple, il est nécessaire parfois
d'augmenter le pouvoir discriminant des milieux lorsque
l'héritabilité des paramètres est faible [97].
Il est couramment établi que l'interaction
génotype x année x lieu est plus importante que l'interaction
génotype x année, elle-même plus importante que
l'interaction génotype x lieu [105]. La présence de l'interaction
génotype x année peut être à l'origine du manque de
répétitivité des paramètres mesurant la
stabilité constatée pour bien des espèces [106]. À
volume expérimental constant, une variation du nombre d'années a
plus d'influence sur la précision des performances d'un génotype
entre milieux qu'un changement du nombre de lieux [105]. Par exemple, trois
années d'expérimentation dans trois lieux donnent une meilleure
précision qu'une seule année dans neuf lieux. Un équilibre
est donc nécessaire entre le nombre d'années et le nombre de
lieux. Toutefois, pour raccourcir la durée du processus de
sélection, le sélectionneur préfère augmenter de
façon importante le nombre de milieux pour compenser l'effet
année.
En présence d'interaction génotype x milieu, il
est économiquement plus intéressant de réduire les essais
à une seule répétition, ce qui permet d'augmenter le
nombre de milieux [107]. Ceci rejoint la constatation que la
37
réponse à la sélection n'est pas trop
affectée lorsque le nombre de répétitions n'est pas
optimal [108].
Lorsque le sélectionneur cherche à
réduire le nombre de milieux, la question de la
représentativité devient primordiale. Une stratégie
consiste à échantillonner les milieux de façon à
représenter un maximum de diversité pour les interactions
observées sur le caractère considéré. Le principe
consiste donc à choisir des lieux qui se ressemblent le moins possible
du point de vue de l'interaction [97].
Un milieu peut être dit favorable s'il apporte à
la plante tout ce dont elle a besoin pour avoir la production maximum. Un
milieu favorable doit donc tendre à supprimer les interactions. Par
contre un milieu pauvre avec des facteurs limitants, doit permettre
l'extériorisation d'interaction, les différents génotypes
n'ayant pas les mêmes exigences [67].
Pour un caractère comme le rendement d'une production,
les variances génétiques et environnementales sont en
général plus fortes en conditions favorables. En conditions
défavorables, la sélection portera plus sur « l'adaptation
» et en conditions favorables, elle portera sur le potentiel. Les deux
aspects doivent être considérés par le sélectionneur
[67].
La comparaison des performances génotypiques dans des
environnements favorables et non favorables à l'expression de haut
rendement en grains est souvent utilisée pour identifier les
génotypes tolérants et productifs. Cette approche vise à
minimiser les besoins de rendement en grains dans les environnements non
favorables relativement aux rendements obtenus en milieux favorables [100].
En outre, La variation des performances d'un cultivar
donné en fonction de la variabilité des milieux est la
résultante des variations cumulées par les composantes du
rendement et les caractères morphologiques de la plante, qui influent
directement ou indirectement sur l'élaboration du rendement. Dans les
régions méditerranéennes, les variations morphologiques
dues à l'interaction de
Selon le concept statistique et biologique, un génotype
stable est celui qui présente une faible variance due aux milieux :
stabilité de type I [115]. Le concept
38
l'environnement mettent le sélectionneur dans une
situation complexe pour choisir et retenir à partir des composantes du
rendement [109].
En effet, L'amélioration de la tolérance aux
stress reste un objectif de sélection prioritaire dans les zones
soumises à une forte variabilité climatique [110]; [74]. FISHER
(1985) [111], mentionne que la tolérance aux stress doit être
quantifiée sur la base du niveau de productivité obtenue sous
contrainte.
De même, l'évaluation peut être
alternée entre les deux types de milieux, favorables et
défavorables dans les premières générations de
sélection et suivie d'une évaluation multilocale la plus large
possible dans les générations suivantes
[112].
4.5 La stabilité du rendement
La stabilité du rendement est un critère important
pour le choix d'une variété
[113]. La capacité génotypique à
maintenir un haut rendement en grains quelque soit les conditions de croissance
prévalente est un concept connu aussi sous le vocable stabilité
du rendement [114]. Pour avoir une stabilité du rendement
élevée, le génotype idéal doit contenir le maximum,
sinon la totalité des gènes contrôlant la tolérance
au stress [109].
Parmi la multitude de termes employés, les termes de
stabilité phénotypique, de stabilité de rendement et
d'adaptation ou d'adaptabilité sont le plus souvent utilisés avec
des sens parfois différents. Il n' y a donc pas de définition
unique de stabilité [113].
La stabilité phénotypique caractérise
l'importance des fluctuations du rendement observées pour un même
génotype cultivé dans différents milieux. Ces fluctuations
dépendent des effets des milieux et des interactions génotype X
milieu [113].
39
agronomique fait intervenir des paramètres de
stabilité de type II. Un génotype stable réagit
parallèlement à la réponse moyenne des génotypes
avec lesquels il est testé [116].
La stabilité de type III est reliée à
l'adaptation ou l'adaptabilité [116], le sélectionneur
recherchera une adaptation spécifique lorsque le milieu est bien
contrôlé, comme en conditions artificielles par exemple, et
plutôt une adaptation générale, associée à
une adaptation spécifique, lorsque le milieu est plus ou moins
contrôlé comme en conditions naturelles [92].
Un dernier type de stabilité, le type IV défini
par LIN et al., 1988 [117], fait intervenir le facteur temps. La
variation du milieu est séparée en une composante
prévisible due à l'interaction génotype × lieu et une
composante imprévisible due à l'interaction génotype
× année. Cette dernière permet une mesure de la
stabilité ou de la capacité d'un cultivar à
résister à des variations imprévisibles qui sont en effet
causées par les effets années.
La stabilité est donc une notion plus large que
l'interaction et elles sont complémentaires l'une de l'autre. En outre,
un génotype stable selon un des critères ne l'est pas
forcément selon les autres. Ceci provient du fait que les
réponses des génotypes aux conditions d'environnement sont
multivariables [116].
Etudier l'adaptation et la stabilité d'un
génotype revient donc à analyser l'interaction génotype
× milieu : le sélectionneur recherche des génotypes
présentant une bonne stabilité, tandis que le
généticien est souvent confronté de façon
involontaire à des problèmes d'analyse de l'interaction. Ceci
explique le développement d'un nombre important de modèles pour
l'analyse de l'interaction génotype × milieu. Il nous a paru plus
intéressant donc la caractérisation des génotypes en
présence de différentes méthodes pour donner à
l'améliorateur des points de repère dans la sélection.
40
CHAPITRE 5
MATERIEL ET METHODES
Dans le nouveau processus d'obtention variétale
établi dans le cadre du programme national de l'amélioration du
blé (PNAB) mis en oeuvre par l'institut national de la recherche
agronomique d'Algérie (INRAA) et l'institut technique des grandes
cultures (ITGC), plusieurs étapes relatives aux essais de rendement
été réalisées à travers les
différentes stations de l'ITGC.
En effet, tous les meilleurs produits
générés soit par croisements et sélections locales
à partir de pépinières en ségrégation ou
d'observation, soit du matériel génétique provenant
d'introductions nouvelles à partir de différents centres
internationaux (CIMMYT, ICARDA, ACSAD etc...) sont testés d'abord en
essai préliminaire de rendement au niveau du site principal (Elkhroub)
en une seule répétition. Les lignées avancées ou
génotypes les plus performants découlant de cet essai
préliminaire sont ensuite testées en essai
répété de 1ère année au niveau des grandes
zones agro-écologiques d'Algérie (Littoral, sub littoral, hautes
plaines, hauts plateaux de l'Est et de l'Ouest) et ainsi de suite
jusqu'à la quatrième année. Durant la campagne 2006/2007,
nous avons travaillé sur l'essai répété de
rendement 1ère année de blé dur qui
était aussi considéré comme un essai multi sites (Figure
5.1). Il a été conduit dans quatre lieux représentant
quatre stations expérimentales de l'institut technique des grandes
cultures à savoir : Oued Smar, ELKhroub (Constantine), Sétif et
Tiaret (Figure 5.2).
Ce site se caractérise par des sols de texture
argilo-limoneuse, un pH de l'ordre de 7 à 7.8. Un taux de calcaire nul,
une faible teneur en matière organique
41

Figure 5.1 : Essai répété
de rendement de blé dur 1ère année 2006/2007
(ITGC Oued Smar).
5.1 Caractéristiques pédoclimatiques des
sites expérimentaux
5.1.1 Le site d'oued Smar (Alger)
La station expérimentale de l'ITGC d'Oued Smar
appartient, selon la classification d'emberger [118], à l'étage
bioclimatique sub-humide à hiver doux et pluvieux et à
été chaud et sec. Les cordonnées géographiques de
la station étant les suivantes :
? Altitude moyenne de 24 m ;
? Latitude : 36.7 N ;
? Longitude : 3.2 E.
42
et riche en azote. Concernant le Phosphore et la potasse,
leurs teneurs sont très faibles ce qui nécessite une correction
[119].
La pluviométrie annuelle moyenne est de 672 mm, avec le
mois le plus froid qui est Janvier et le mois le plus chaud qui est Août.
La saison se divise en deux périodes, une période humide et
froide qui va d'Octobre à Avril et une période sèche et
chaude allant de Mai à Septembre [118].
La zone connaît peu d'accidents climatiques. Elle est
directement exposée à l'influence maritime qui se traduit par un
taux d'humidité de l'air ambiant élevé et un risque nul
pour le gel et protégée des vents chauds, venant du sud, par
l'Atlas blidéen.
5.1.2 Le site d' El Khroub (Constantine)
Le site expérimental de l'ITGC d'El Khroub (Constantine)
est situé à une
quinzaine de kilomètres au Sud Est de Constantine avec une
:
? Altitude moyenne de 640 m ;
? Latitude : 36.6 N ;
? Longitude : 6.6 E.
La zone du Khroub est caractérisée par un sol
brun à la surface et brun ocre en profondeur, de texture argileuse
à argilo-limoneuse, le taux d'argile varie de 30 à 39%. Ces sols
ont une caractéristique spécifique; forte rétention en
eau, apparition en période sèche des fissures de retrait.
Les caractéristiques chimiques indiquent que le sol est
basique avec un pH alcalin (8.30-8.40 %), et moyennement riche en calcaire dont
la teneur est de 12%. La teneur en matière organique est faible
(0.95-1.25 %).
En ce qui concerne les profondeurs du sol analysées, la
teneur en azote (N%) dépasse la valeur 0.05%. De ce fait, notre sol est
riche en azote, la teneur en Phosphore et Potasse est très faible ce qui
nécessite une correction [119].
43
La zone d'El Khroub est caractérisée par un
climat continental semi-aride à hiver froid et à
été chaud et sec avec une pluviométrie annuelle qui est de
450 mm. Ce site est caractérisé par des terres vallonnées,
assez fertiles avec quelques risques de gel tardif et de sécheresse en
fin de cycle.
5.1.3 Le site de Sétif
La station expérimentale de l' ITGC de Sétif
située à 4 Km au sud Est de la
ville de Sétif. Le site se trouve en partie dans la petite
vallée de l'oued
Boussellem avec une :
? Altitude moyenne de 1023 m ;
? latitude : 36.12 N ;
? Longitude : 05.24 E ;
? pente moyenne : 2%.
Les sols des hauts plateaux Sétifiens sont
carbonatés dont leur majorité et présentent des
individualisations calcaires continues, dures. Celles ci ne laissent qu'une
mince couche de terre, limitant ainsi l'extension des racines [120].
Les caractéristiques pédologiques du site
expérimental indiquent que le sol est de type argilo-limoneuse selon les
horizons, un pH alcalin (8.2). La teneur moyenne en calcaire actif est de
18.4%, la matière organique est acceptable avec une valeur de 2.6% et le
phosphore assimilable a atteint les 36.0 ppm, il est pauvre en azote avec une
valeur moyenne de 0.07%.
La zone appartient au domaine semi-aride
caractérisé par un climat continental typiquement
méditerranéen de type (d) à hivers froids, et sous
l'influence de l'atlas tellien (qui limite les précipitations
hivernales), de l'atlas saharien et du Sahara (qui sont à l'origine de
vents secs et desséchants qui peuvent intervenir dés le
printemps), et de l'altitude (qui entraîne des gelées tardives)
[121]. La saison sèche s'étale sur 4 à 5 mois, de Mars
à Septembre. Elle est très exposée aux risques
précoces de sirocco ainsi qu'au gel printanier [54]. La
pluviométrie annuelle est de 396 mm et présente une grande
variabilité interannuelle. La willaya de Sétif occupe, à
l'échelle nationale, le troisième rang
44
sur le plan de la superficie des céréales,
après Tiaret et Oum EL-Bouaghi, avec prés de 180.000 hectares
emblavés chaque année [122].
5.1.4 Le site de Tiaret
La station expérimentale de Tiaret située à
45 Km de la ville de Tiaret. Le
site se trouve en partie dans le village de Sabaine avec une :
? Altitude moyenne de 910 m ;
? Latitude : 36.5 N ;
? Longitude : 1.6 E.
La zone de SABAIN est caractérisée par des sols
légèrement dégradés en raison de la topographie
légèrement ondulée .ce sont des sols de profondeur
moyenne.
Le site est représentatif les hauts plateaux de
l'Ouest, le climat de cette région est rigoureux avec une
pluviométrie limitée (348 mm) et très
irrégulière. La sécheresse printanière, les
siroccos précoces sont à craindre à partir de la mi-Avril
et suivent de très près les gelées tardives. Ces stress
sont les principaux facteurs limitant la production
céréalière [54].
Figure 5.2 : Localisation géographique des quatre sites
d'étude.
45
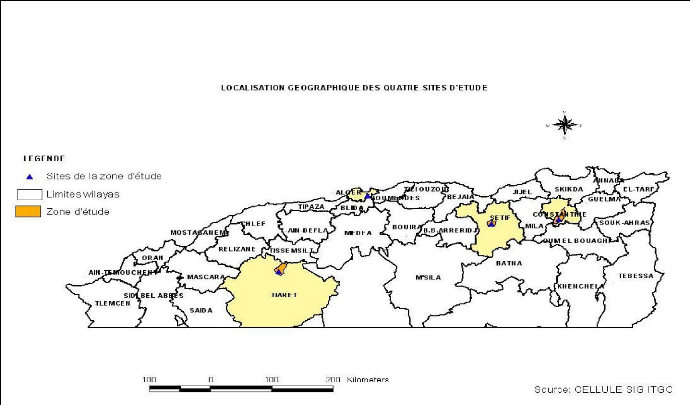
46
5.2 Conduite des essais
5.2.1 Station Oued smar
· Précédant cultural : le
précédant était une jachère travaillée.
· Préparation du sol :
V' Labour : effectué à l'aide d'une
charrue bisoc;
V' Façons superficielles : passage du
cultivateur et de la herse avant le semis.
· Engrais de fond : Epandage d'engrais de type TSP 46 %
avec une dose de 2 q/ha.
· Semis : le semis est effectué le 9 Janvier 2007
à une profondeur de 3 cm.
· Désherbage et fertilisation : Le traitement
phytosanitaire a été effectué à l'aide d'un anti
dicotylédones le 2.4 D (acide 2.4
dichloro-phénoxyacétique) à raison d'un litre/ha et d'un
anti mono cotylédones le Topic à raison de 1l/ha en date du 26
Mars 2007 suivi de l'épandage d'engrais azotée sous forme
d'urée à raison de 1.5 q/ha.
· Récolte : Elle a été
réalisée le 19 Juin 2007 à l'aide de la moissonneuse
batteuse expérimentale Hege125.
5.2.2 Station El khroub
· Précédant cultural : le
précédant était une jachère travaillée.
· Préparation du sol :
V' Labour : effectuée à l'aide d'une
charrue à disque;
V' Les recroisages : le premier effectué
à l'aide d'un cover croop au mois d'Avril 2006 alors que le
deuxième recroisage a été réalisé au mois de
Septembre avec une Herse rotative ;
V' Façons superficielles :
réalisées avant le semis à l'aide d'un roto herse.
· Engrais de fond : Epandage d'engrais de type TSP 46 %
(fertilisation phosphatée) effectuée à l'aide d'un
épandeur centrifuge avec une dose de 1 q/ha au mois d'Octobre 2006.
· Semis : Le semis est réalisé au 6
Janvier 2007 avec un semoir expérimental HIGI.
47
· Désherbage et fertilisation : L'entretien
consiste en un désherbage aux deux désherbants qui sont Topik
(0.9 l/ha) et Zoom (120 gr/ha) au stade tallage et un épandage d'engrais
azotée sous forme d'urée 46 % à raison de 1 q /ha au
même stade.
· Récolte : la récolte a été
faite le 07 Juillet 2007.
5.2.3 Station de Sétif
· Précédant cultural : le
précédant était une jachère travaillée.
· Préparation du sol :
V' Labour : un labour profond suivi de passage du cover croop
pour l'enfouissement des engrais ;
V' Façons superficielles : passage de la herse avant
l'opération semis.
· Engrais de fond : L'épandage de 46
unités/ha de superphosphate à 46 % est effectuée juste
avant le semis.
· Semis : le semis est réalisé le 09
Décembre 2006.
· Désherbage et fertilisation : L'apport de 120
gr/ha d'un désherbant de type Zoom est réalisé au stade
plein tallage suivi de l'épandage d'azote sous forme d'urée
à raison de 1 q/ha au stade début et fin tallage.
· Récolte : la récolte a été
faite le 11 Juillet 2007.
5.2.4 Station de Tiaret
· Précédant cultural : le
précédant était des légumes sec.
· Préparation du sol :
V' Labour : à l'aide d'une charrue à disque,
l'état du sol était semi-humide ;
V' Les recroisages : le premier effectué à
l'aide d'un cover croop au début de novembre, mais le deuxième
recriosage a été réalisé avant le semis (fin
Décembre 2006) avec toujours le cover croop;
V' Façons superficielles : réalisées
avant le semis à l'aide d'une roto herse.
· Semis : le semis réalisé le 04 Janvier
2007 avec un semoir expérimental (Ojord).
· Désherbage et fertilisation : L'entretien
consiste en un désherbage aux deux désherbants Topik et Zoom
à raison de 0.9 l/ha et 120 gr/ha
Figure 5.3 : Variation du cumul pluviométrique par site
d'expérimentation durant la campagne 2006/2007 [123].
48
successivement au mois de Mars, suivi par l'épandage
d'engrais azotée sous forme d'urée (1 q/ha) au stade fin tallage.
? La récolte : la récolte a été
faite le 29 Juin 2007.
5.3 Analyse des conditions climatiques
La quantité moyenne de pluie enregistrée durant
la campagne agricole 2006/2007, a varié de 628.3 mm pour le site d'Oued
Smar à 341.8 mm pour le site de Tiaret (Figure 5.3), soit un
écart de 286.5 mm. Pour ce dernier site. Les précipitations de la
compagne ont été de 10.76 mm inférieurs à celle de
la moyenne des cinq dernières compagnes (Appendice B ; Tableau 1).
600
500
400
P
700
300
200
100
0
Oued S Khroub Sétif Tiaret
Sites
La quantité de pluie était moyenne dans la
région du Khroub (440.9 mm), Le déficit de 46 mm par rapport
à la moyenne des 25 ans ne s'est par fait trop sentir. Nous pouvons
remarquer aussi que les précipitations de cette campagne (2006/2007) ont
été de 78 mm et étant supérieures à celle de
la compagne précédente (Appendice B, Tableau 4). Le site de
Sétif a reçu 392.4 mm en moyenne (Figure 5.3). Cette
quantité été légèrement inférieure
à la moyenne des cinq dernières campagnes (Appendice B, Tableau
5).
49
En effet la variation inter site est très forte.
Puisque le site d'Oued Smar est de loin le plus pluvieux, alors que le site de
Tiaret est plus stressant pour la végétation.
Une comparaison des pluies mensuelle dans les quatre sites
nous révèle des fluctuations de la quantité de pluie selon
les mois (Figure 5.4).
Pour le site d'Oued Smar, l'automne a été
marqué par un déficit en précipitation, ce qui a
retardé en quelque sorte la préparation du lit de semence. Par
ailleurs, la quantité de pluie enregistrée pendant la
période de reproduction (Mars-Avril) a été très
bénéfique pour un bon développement des
céréales (Figure 5.4). Cependant, les dernières pluies ont
été favorables au développement de maladies
cryptogamiques, notamment la septoriose et la rouille brune ce qui a
affecté certaines variétés. Cette situation a
été également bénéfique au
développement des mauvaises herbes dites thermophiles.
Globalement la pluviométrie totale a été
convenable durant cette campagne au Khroub, les 440 mm cumulées de
Septembre à Juin ont été suffisants pour un bon
développement des cultures. La réalisation du semis a
été perturbée par les précipitations abondantes du
mois de Décembre (118.8 mm) où nous avons enregistré un
écart positif de 45.6 mm de plus que la moyenne. Durant les mois de
Janvier et Février tout l'essai a montré une bonne installation
malgré l'occurrence de plusieurs journées gélives.
La période de Mars à Mai a enregistré un
cumul de 210.1 mm (Figure 5.4), cette quantité assez bien
répartie sur ce trimestre est théoriquement suffisante à
une bonne production des céréales si ce n'est l'occurrence des
siroccos du mois de Juin surtout qui ont beaucoup soufflés provoquant
des températures atteignant parfois les 38°C qui ont affecté les
variétés tardives.
50
250 P
200
150
100
50
0
Mois
S O N D J F M A M J
Oued S Khroub Sétif
Tiaret
Figure 5.4 : Variation de la pluviométrie moyenne
mensuelle des sites expérimentaux durant la campagne 2006/2007 [123].
Les mois d'Octobre et de Novembre étaient peu pluvieux,
sur le site de Sétif, est au total de 10.1 mm. L'absence de pluie durant
ces deux mois a permis aux agriculteurs de travailler leurs sols plus ou moins
bien par contre le semis a été retardé par crainte des
pluies parasites.
Au cours des mois de Février et Mars nous avons
enregistré un cumul de 128.6 mm (Figure 5.4), ce qui a permis de dire
que le mois de Mars a été le mois le plus froid et le plus
pluvieux. Cette quantité de pluie a eu un effet très positif sur
le coefficient de tallage mais le manque de précipitation durant les
phases de remplissage a eu un effet très néfaste sur le PMG.
L'absence de pluies durant l'automne a retardé en
quelque sorte la préparation de lit de semence et la réalisation
de l'opération semis au niveau de la zone de Tiaret. Par ailleurs, la
quantité de pluie enregistrée durant les mois de Février
et Mars a été très favorable et a un effet positif sur le
coefficient de tallage, cette précipitation s'élève
à 100.00 mm durant le mois d'Avril (Figure 5.4). La fin de cycle dans
cette zone a été marquée par un déficit en
précipitation associée aux températures
élevées.
51
En plus de ces variations pluviométriques, des
fluctuations thermiques ont été enregistrées dans les
quatre lieux testés (Figure 5.5), marquant d'une part l'effet des basses
températures, comme c'est le cas du site de Tiaret où la
température moyenne atteint 5°C au mois de Mars (Figure 5.5). Ceux qui a
influé sur la fertilité des épis et d'autre part, l'effet
des hautes températures en fin de cycle qui a coïncidé avec
la phase de remplissage des grains.
Oued S
Khroub
20
Sétif
Tiaret
15
10
5
0
S
O N
D J
F
M A
M
J
mois
Figure 5.5 : Variation de la température moyenne mensuelle
des sites
expérimentaux de la campagne 2006/2007 [123].
5.4 Matériel végétal
Trente variétés et lignées de blé
dur (Triticum durum Desf.) Ont été utilisées
dans notre étude dont cinq sont utilisées comme
témoins. Il s'agit des variétés
Waha, Hoggar, Gta/Dur, Boussellem et Cirta. Les
variétés concernées par l'étude
et les caractéristiques de celles des témoins sont
illustrées respectivement
dans les tableaux 5.1 et 5.2.
52
Tableau 5.1 : Code et pedigree des génotypes testés
dans quatre sites durant la campagne 2006-2007 [124].
|
N°
|
Variétés ou Lignées
|
Pedigree
|
Origine
|
|
1
|
Minimus/ Rascon_19
|
CDSS93Y415-2Y-0B-0Y-2B-0Y
|
MULTI1-02
|
|
2
|
Mck 2/Aco89
|
CDSS93Y7-27Y-0M-0Y-0B-1Y-0B
|
MULTI1-03
|
|
3
|
Bichena / Ariza_2//Solga_8
|
CDSS94Y00805T-D-2M-0Y-0B-3Y-0B
|
MULTI1-04
|
|
4
|
Silver_26 / Toska_26
|
CDSS94Y00412S-8Y-0M-0Y-0B-2Y-0B
|
MULTI1-07
|
|
5
|
Waha
|
Témoin
|
Témoin
|
|
6
|
Minimus_7// Auk /Oste /3/ Shag_26
|
CDSS93B00594T-A-3Y-0M-0Y-0B-0Y-4B
|
MULTI1-08
|
|
7
|
Sula /RBCE 2/3/Hui //CIT 71/Cii/4/Ryps27 3/...
|
CDSS95B00359S-0M-1Y-0B-2Y-0B-0Y-0B
|
MULTI1-09
|
|
8
|
Rascon/Sla_3/3/Plata_1/Snm//Plata_9
|
CDSS96Y00558S-11Y-0M-0Y-1B-0Y-0B-0B
|
MULTI1-11
|
|
9
|
Sn Turk Mi 83-84 375/Nigris_5// Tantlo_1/3/...
|
CDSS96Y00442S-7Y-0M-0Y-1B-0Y-0B-0B
|
MULTI1-12
|
|
10
|
Hoggar
|
Témoin
|
Témoin
|
|
11
|
Srn 3 / Nigris 4 // Porto 5
|
CDSS95B00195S-7Y-0M-0Y-1B-0Y-0B-0B
|
MULTI1-13
|
|
12
|
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5
|
ICD95-1302-C-3AP-0AP-1AP-0AP-5AP-AP-5AP-0AP
|
MULTI1-14
|
|
13
|
D68-1-93A-1A//Ruff /Fg /3/ Mtl-5/4/Lahn
|
ICD95-0169-C-0AP-3AP-0AP
|
MULTI1-16
|
|
14
|
Plata_1/Snm // Plata_9/3/Green
|
CDSS95B00284S-0M-9Y-0B-2Y-0B-0Y-0B
|
MULTI1-17
|
|
15
|
Gta/Dur...
|
Témoin
|
Témoin
|
|
16
|
Inter 9/Poho 1
|
CDSS95Y00286S-3Y-0M-0Y-0M-0Y-0B-1Y-0B
|
MULTI1-21
|
|
17
|
Kucük
|
CD91B2620-G-8M-030Y-030M-2Y-0M-2Y-0B
|
MULTI1-22
|
|
18
|
Aaz 77_2/Porron_11//Busca_3
|
CDSS95B00637T-A-0M-2Y-0B-3Y-0B-0Y-0B
|
MULTI1-23
|
|
19
|
Altar84
|
|
|
|
20
|
Boussellem
|
Témoin
|
Témoin
|
|
21
|
D68-1-93A-1A //Ruff/Fg/3/Mtl-5/4/Lahn
|
ICD95-0169-C-0AP-3AP-0AP-5AP-0AP
|
LOC1-01
|
|
22
|
Aram-5/Cali//Rascon-37/3/Plata-8
|
CDSS92B1478-A-10M-0Y-0B
|
LO-07
|
|
23
|
FI 05
|
-
|
LO-09
|
|
24
|
FI 02
|
-
|
LO-08
|
|
25
|
Cirta
|
Témoin
|
Témoin
|
|
26
|
Ting7/2*Green-3
|
CDSS92B1037-G-2M-0Y-0M-0Y-1B-0Y
|
LO-01
|
|
27
|
Boussellem/Ofonto//waha
|
16S-1S-1S-1S-0S
|
Sétif
|
|
28
|
Ofonto/Waha//MBB
|
32S-2S-2S-1S-0S
|
Sétif
|
|
29
|
Tr 32225/Gediz//Ofonto
|
|
Sétif
|
|
30
|
MBB/Ofonto//Radiosso/Waha
|
2S-1S-3S-0S
|
Sétif
|
53
Tableau 5.2 : Caractéristiques des variétés
témoins [28].
|
Caractéristiq
|
|
Pedigree
|
Origine
|
Zone d'adaptation
|
Cycle végétatif
|
Résistance
|
Productivité
|
|
Témoins
|
Obtenteur
|
|
Waha
|
ITGC
El khroub
|
waha `S' plc `S'
/Ruff/ Gta «S» /
3/Rolette
|
CIMMYT (Mexique)
|
Hauts -plateaux et Plaines intérieures.
|
précoce
|
Tolérante au froid, résistante à la verse,
sensible à la sécheresse et aux gelées
printanières.
|
45 q/ha
|
|
Hoggar
|
CIMMYT (Mexique)
|
JO»S» /AA»S» // FG»S»-CM 9799;
|
Espagne
|
Hauts-Plateaux, Plaines
intérieures et zones sahariennes (sous pivot)
|
semi- précoce
|
Tolérante au froid, résistante à la verse,
sensible à la sécheresse et aux gelées
printanières.
|
60 q/ha. Pour les Hauts-Plateaux et Plaines intérieures
|
|
Gta/Dur
|
ITGC Guelma
|
Gaviota /durum
|
CIMMYT (Mexique)
|
Littoral et sub littoral, Plaines intérieures et
Hauts-Plateaux.
|
précoce
|
Tolérante au froid,
moyennement résistante à la verse, et sensible
à la sécheresse.
|
50 q/ha
|
|
Boussellem
|
ITGC Sétif
|
Heider/Marte//Hu evo de oro ICD86-0414-Abl- Otr-4AP-
Otr14AP-Otr.
|
ICARDA (Syrie) - CIMMYT (Mexique)
|
Nord Hauts- Plateaux, plaines intérieures.
|
mi-tardif
|
Tolérante au froid et à la sécheresse,
résistante à la verse.
|
38 q/ha
|
|
Cirta
|
ITGC
EL Khroub
|
Hedba3/GDO VZ 619
|
Algérie
|
Sub littoral et Plaines intérieures.
|
semi- précoce
|
Tolérante au froid et à la sécheresse, assez
résistante à la verse.
|
35 à 40 q/ha
|
54
5.5 Protocole expérimental
Les essais ont été conduits en plein champ, au
cours de la campagne 2006\2007 pour les quatre sites d'étude (Figure
5.6).
Le protocole expérimental adopté au niveau des
04 sites étant celui des blocs aléatoires complets avec 4
répétitions. La distribution des variétés au niveau
des différents blocs est aléatoire (Figure 5.7).
L'essai occupe une superficie de 1127.5 m2 avec 120 parcelles
élémentaires de 6 lignes de 5 m de long avec un espace inter
lignes de 20 cm (soit une superficie parcellaire de 6 m2). Ceci
correspond à une densité de 300 plants / m2.
55

|
Figure 5.6.A : vue de l'essai installé à Oued
Smar.
|
Figure 5.6.B : vue de l'essai installé au Khroub.
|

Figure 5.6.C : vue de l'essai installé à Figure
5.6.D : vue de l'essai installé à
Tiaret. Sétif.
Figure 5.6 : L'expérimentation conduite dans les quatre
sites d'étude.

5m
B I
1.2m 0.5m
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
1m
6 8 7 9 10 12 11 14 5 15 13 3 1 2 4
B II
0.5m
21 23 22 24 25 27 26 29 20 30 28 18 16 17 19
9 10 12 11 14 3 2 1 13 4 7 8 15 6 5
B III
24 25 27 26 29 18 17 16 28 19 22 23 30 21 20
11 5 1 6 13 8 4 15 12 2 14 10 9 7
3
B IV
26 20 16 21 28 23 19 30 27 17 29 25 24 22 18
56
Figure 5.7 : Protocole expérimental adopté au
niveau des 4 sites.
> Nombre de grains par épi (NGE) : Le nombre de grains
par épi a été déterminé sur un
échantillon aléatoire de 10 épis par
variété;
57
5.6 Suivi et notations
Pour caractériser les génotypes
étudiés, des mesures ont été faites sur un ensemble
de paramètres liés à la phénologie et à la
morphologie de la plante tout le long du cycle de développement à
travers les quatre sites expérimentaux. Les paramètres
observés se résument comme suite :
5.6.1 Paramètres morpho phénologiques
> Notation du stade épiaison : le stade
épiaison est défini lorsque 50% des épis sont
complètement dégagés de la gaine de la feuille
étendard de la parcelle élémentaire notée. Cette
date sert à la détermination de la durée de la phase
végétative (DPV) ;
> Nombre de talles par m2 (TLL/m2) a
été réalisé au niveau de trois rangées par
parcelle élémentaire;
> Hauteur des chaumes (HT) : A maturité la hauteur
des plantes a été mesurée sur chaque parcelle
élémentaire à partir de 10 plants choisis au hasard. (la
hauteur est définie comme étant la longueur séparant la
base de la plante à l'extrémité supérieure du
pédoncule);
> Longueur de l'épi (LE) : Pour cet
paramètre, nos mesures ont porté sur un échantillon de 10
plants au hasard. La longueur de l'épi est définie comme
étant la distance séparant la base de l'épi et son sommet
(barbes non incluses);
> Nombre d'épis par m2 (NE/m2)
: Il a été déterminé au niveau de trois
rangées par parcelle élémentaire;
La détermination du taux de mitadinage est faite sur un
échantillon de 100 g par variété et pour les quatre sites,
après avoir procédé au nettoyage des grains.
58
? Poids de mille grains (PMG) : Le comptage et le poids des
grains a été réalisé par un compteur
électronique et une balance de précision de 00.1gr;
? Rendement en grains réel (RDT) est
déterminé suite à la récolte des essais. La
quantité de grains récupérés de chaque unité
est battue, nettoyée, triée et ensuite pesée pour
déterminer le rendement en kg par unité puis reconvertie en
q/ha.
5.6.2 Analyses technologiques
Pour apprécier l'aptitude technologique de chaque
génotype, nous avons jugé nécessaire de procéder
à la détermination de certains paramètres indispensables
qui portent sur la :
? Détermination du taux de mitadinage (MTD); ?
Détermination du taux d'extraction (EXT); ? Détermination du taux
de gluten humide (GH); ? Détermination du taux de gluten sec (GS).
Ces analyses ont été effectuées au
laboratoire de l'institut technique des grandes cultures (ITGC siège
à Alger) sur les échantillons des essais de quatre sites.
? Détermination du taux de mitadinage
Le taux de mitadinage (MTD) est défini comme
étant le pourcentage en nombre de grains de blé dur reconnus non
entièrement vitreux. Le grain de blé dur est normalement
entièrement vitreux. S'il présente la moindre trace d'amande
farineuse, il entre dans la catégorie des grains mitadinés.
1000
59
La méthode de détermination consiste en :
? L' Epandage de l'échantillon dans un bac ;
? L'examen d'une quantité de grains, ces grains sont
coupés transversalement à l'aide du Farinotome de Pohl pour
compter les grains mitadinés, même partiellement. La
méthode est répétée six fois pour chaque
échantillon.
? Détermination du taux d'extraction
Après nettoyage de 500 g de grains de chaque
variété pour les quatre essais, une quantité de 300 g a
été conditionnée. Son objectif est de modifier
l'état physique des grains de manière à faciliter la
séparation du son et de l'amande et le broyage de celle-ci. Un
blé humide sera difficile à travailler, notamment à bluter
; un blé trop sec se prêtera mal à la séparation des
enveloppes du coeur de l'albumen. Le conditionnement a été
conduit en deux étapes :
? Premier mouillage + premier repos;
? Deuxième mouillage + deuxième repos.
Le conditionnement a été réalisé
dans des bocaux, après avoir ajouté au préalable la
quantité d'eau nécessaire. Les bocaux sont ensuite fermés
hermétiquement puis soumis à des mouvements afin que l'eau soit
bien répartie entre les grains, nous laissons enfin les bocaux reposer
horizontalement le temps nécessaire.
La quantité d'eau ajoutée est
déterminée selon l'humidité qui est calculée
après broyage d'une petite quantité de grains, 10 g de produit
pour variété a été pesé et versé dans
une vase métallique. Deux vases par échantillon ont
été introduits dans une étuve pendant 10 mn,
l'humidité et la quantité d'eau ajoutée sont
déterminées selon les formules suivantes :
T= [14 - H] x 12
T 1000g
X 300g
T x 300
X =
60
H : la moyenne de l'humidité pour chaque
échantillon; T : la quantité d'eau ajoutée pour 1000 g de
grains; X : la quantité d'eau ajoutée pour 300 g de grains.
Après l'hydratation de 4 à 5 échantillons
par jour ; un premier mouillage a été effectué pendant 30
mn pour imbiber bien les grains; une durée de repos de 12 heures a
été proposée avant d'effectuer la deuxième
hydratation suivie par le deuxième mouillage.
Après un repos de deux heures, l'opération de
transformation de blé dur en semoule a été
effectuée au laboratoire dans un moulin expérimental de type
CHOPIN. Le produit fini (semoule) a été pesé et le taux
d'extraction est obtenu en suivant les étapes suivantes :
M 300 g
M' 1000 g
Où M : est la quantité de semoule dans 300 g de
grains de blé dur ;
M' : est la quantité de semoule dans 1000 g de grains de
blé dur.
1000g 100 %
M' X %
M'
Le taux d'extraction X =
10
? Détermination du taux du gluten
humide
Décrit pour la première fois en 1745 par JACOPO
BECCARI [125], Professeur à l'université de Bologne, le gluten
est un matériel viscoélastique obtenu par lixiviation (lavage par
l'eau) d'une pâte de blé tendre ou de blé dur.
Principalement constitué de protéines (75 à 85 % selon les
conditions de fabrication) [126].
61
? Mode opératoire
-I'humidité a été déterminé
en pesant 10 g de semoule qui a été préparé avant
et versé dans une vase métallique, deux vase ont
été introduit dans l'étuve pendant 10 mn et
l'humidité calculé selon la même méthode
citée dans le conditionnement ;
-La préparation de NACL doit être
réalisée avec l'eau destillée (20 g de NACL dans un litre
d'eau destillée), nous avons préparé la solution
fraîche chaque jour et l'utilisée à une température
comprise entre 18 et 22°C ;
-La pâte a été préparée en
pesant 10 g de semoule qui ont été introduits dans un mortier et
mélangés avec 10 ml de la solution de chlorure de sodium. La
semoule a été agitée avec la spatule pour former une boule
avec la pâte ;
-La masse obtenue est malaxée durant 10 mn à
l'aide d'une presse à gluten (GLUTOMATIC) au moyen de la solution de
chlorure de sodium jusqu'à l'obtention d'un pâton doux au toucher,
ce réseau élastique a été pesé à
l'état humide après rosage pour donner le gluten humide .
L'opération a été
répétée deux fois pour confirmer les résultats.
? Expression des résultats
Le gluten humide exprimé en pourcentage de masse du
produit rapporté à la matière sèche (GH %) est
égal :

Mh ×10 × 100
GH (%) =
100 - H
Où M h : est la masse, en gramme, de gluten humide.
H : la moyenne de l'humidité.

Y = b I + a Où I = Y.j - Y..
62
? Détermination du taux du gluten sec
Le gluten humide obtenu précédemment est
placé dans un appareil (Gluark) pendant 4 mn pour sa dessiccation, la
masse obtenue a été pesée à l'état sec pour
donner le gluten sec en gramme.
Le gluten sec exprimé en pourcentage de masse du produit
rapporté à la matière sèche (GS%) est égal
:

Ms ×10 × 100
GS (%) =
100 - H
Où Ms : est la masse, en gramme, de gluten sec.
H : la moyenne de l'humidité
5.7 Analyse des données
Les données mesurées ont fait l'objet d'une
analyse de variance par site et combinée sur tous les différents
sites pour analyser l'interaction génotype x environnement (G x L). Ces
analyses ont été réalisées avec le logiciel
CropStat 7.2 (2008) [127]. La plus petite différence significative
(Ppds) au seuil de 5% de probabilité a été utilisée
pour comparer les moyennes variétales et environnementales.
Suite à la multitude des réponses des
génotypes à la variation du milieu, les données ont
été traitées par la technique de la régression
conjointe selon le modèle de FINLY et WILKINSON (1963) [11] qui estime
la régression du rendement de chaque variété par rapport
à l'indice du milieu (I) qui est l'écart de la moyenne des
génotypes dans un lieu (Y.j) déduite de la grande moyenne (Y..).
L'équation de la régression est :
63
Elle décrit une relation linéaire entre I,
variable indépendante (indice du milieu) et Yij variable
dépendante représentant le rendement moyen du génotype i
dans l'environnement j. a est l'ordonnée à l'origine et b la
pente de la droite, ou coefficient de régression.
Une valeur de b significativement inférieure à 1
est indicatrice d'une adaptation spécifique aux environnements
défavorables tandis que des valeurs supérieures à 1
caractérisent une adaptation spécifique aux environnements
favorables. Une valeur de b égale à l'unité est indicative
d'une large adaptation. Cette méthode permet de calculer la droite de
régression du rendement moyen du génotype en fonction du
rendement moyen de tous les environnements [11].
La méthode des effets principaux additifs et de
l'interaction multiplicative (AMMI) a été aussi utilisée,
pour étudier l'interaction génotype x environnement. Cette
technique associe l'analyse de variance de modèle additif à
l'analyse en composantes principales des interactions. Elle sépare les
effets dus au comportement génotypique (pattern) des effets dus à
l'erreur dit bruit de fond (bruit =noise) [13].
Une étude des corrélations a été
réalisée, pour déceler les liens éventuels entre
les caractères mesurés. La connaissance des relations entre
caractères permet d'identifier ceux qui contribuent directement ou
indirectement au rendement, pour leurs utilisations comme critères
directes ou indirectes de sélection pour le rendement.
Le degré de similitudes des différents sites est
mesuré par le coefficient de corrélation de rang de Spearman
(rs) qui a été calculé selon la
procédure décrite par STEEL et TORRIE (1980) [128]. La
signification statistique du coefficient de rang de Spearman rs est
testée par comparaison du t de Student calculé avec le t de table
au seuil de 5% de probabilité.
64
Le t de Student est calculé selon STEEL et TORRIE, (1980)
[128] par :
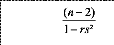
tobs = rs
Où n= nombre de génotype et rs est le
coefficient de rang de Spearman. Le t de table a n-1 degrés de
libertés.
Le rendement nominal de chaque génotype a
été estimé par la somme de l'effet principal du
génotype plus l'interaction génotype x environnement. Cette
dernière est égale au produit des scores du génotype
(GIPCA1) et du site considérés (LIPCA1). Le rendement nominal
permet d'identifier les génotypes qui se partagent le même domaine
de recommandations.
L'indice I (reliability index) est calculé selon la
procédure décrite par ANNICHIARICO (2002) [129]. Cet indice donne
le plus faible rendement du génotype considéré dans les
plus mauvaises conditions (risque). Il a la forme :

Ii = Yi. - Zpó
Où Yi. L'effet moyen génotype,
ó est l'écart type de la variance inter sites et Zp est
égale à 1.645, suite à la forte fréquence des
mauvaises campagnes qui caractérisent la céréaliculture
algérienne.
65
ns, *, **, et *** = effet non significatif et
significatif au seuil de 5 et 1% respectivement.
CHAPITRE 6
ANALYSE ET INTERPRETATION DES RESULTATS
6.1 Etude du comportement génotypique intra
et inter sites
6.1.1 Durée de la phase végétative et
hauteur de la végétation
6.1.1.1 Durée de la phase
végétative
L'analyse de la variance de la durée de la phase
végétative et de la hauteur du chaume, effectuée par site
et pour les quatre sites d'étude, indique un effet génotype
significatif à très hautement significatif pour les deux
variables (Tableau 6.1). L'effet significatif indique que des
différences génotypiques, pour la durée de la phase
végétative et la hauteur du chaume, existent entre les
différentes variétés évaluées. Ceci offre
l'opportunité de sélectionner pour ces deux variables,
individuellement ou en combinaison, si les différences existantes entre
génotypes affectent la capacité de rendement en grains [130] ou
si elles sont désirables parce qu'elles sont actives dans l'esquive des
stress et le rendement économique [131].
Tableau 6.1 : Carrés moyens des écarts de
l'analyse de la variance de la durée de la phase
végétative (DVP) et de la hauteur (HT) de la
végétation des 4 sites.
Variable Source ddl OSM KHR SET TRT
DVP Génotype 29 28.94*** 35.6
*** 21.6 *** 16.84***
(j) Blocs 3 0.81 7.43 3.26 3.74
Résiduelle 87 0.5 3.9 1.0 1.63
HT Génotype 29 70.41*** 68.15***
33.95*** 27.91*
(cm) Blocs 3 98.51 31.03 164.03 21.97
Résiduelle 87 9.11 15.22 10.9 17.76
66
La durée de la phase végétative (semis-
épiaison), sur le site d'oued Smar (OSM), est en moyenne de 100.8 jours
et varie de 97.0 à 108.3 jours, soit une amplitude de 11.3 jours, qui
peut être exploiter en sélection vu que la plus petite
différence significative n'est que de 2.5 jours (Tableau 6.2). Les
génotypes Fl 02 (n° 24) et Cirta sont les plus tardifs, alors que
le reste des variétés testées présente une
durée de la phase végétative inférieure à
100 jours (Appendice C, Tableau 1). Ce résultat montre le sens de la
sélection pratiquée qui privilège le raccourcissement du
cycle pour esquiver les stress de fin de cycle. Le stade épiaison est
une importante étape dans le cycle de vie de la plante. C'est à
ce stade que l'architecture de la plante devient apparente et atteint son
maximum, elle donne le plus souvent une indication sur les capacités
différentielles de production des variétés [132].
CECCARELLI et al., (1992) [74] mentionnent qu'une
moindre sensibilité à la photopériode permet à la
variété d'épier plus tôt, et donc d'être moins
exposée aux stress abiotiques de fin de cycle.
Tableau 6.2 : Valeurs moyennes caractéristiques de la
durée de la phase végétative et de la hauteur du chaume
notées sur les différents sites.
|
Sites
|
Valeurs
|
DPV (jours)
|
HT (cm)
|
|
OSM
|
Max
|
108,3
|
90,1
|
|
Min
|
97,0
|
71,3
|
|
Amplitude
|
11,3
|
18,9
|
|
Moyenne
|
100,8
|
80,1
|
|
Ppds5%
|
2,5
|
4,2
|
|
KHR
|
Max
|
121,0
|
97,8
|
|
Min
|
104,8
|
80,8
|
|
Amplitude
|
16,3
|
17,0
|
|
Moyenne
|
117,5
|
88,6
|
|
Ppds5%
|
2,7
|
5,5
|
|
SET
|
Max
|
146,0
|
97,3
|
|
Min
|
138,0
|
83,5
|
|
Amplitude
|
8,0
|
13,8
|
|
Moyenne
|
141,7
|
90,2
|
|
Ppds5%
|
1,41
|
4,6
|
|
TRT
|
Max
|
130,0
|
87,1
|
|
Min
|
121,0
|
76,9
|
|
Amplitude
|
9,0
|
10,3
|
|
Moyenne
|
124,5
|
82,0
|
|
Ppds5%
|
1,8
|
5,9
|
67
Sur le site du Khroub (KHR), la durée de la phase
végétative varie de 104.8 j, valeur observée pour le
génotype Waha, à 121.0 j pour le génotype Srn_3 / Nigris_4
// Porto_5 (n° 11). La moyenne est de 117.5 jours, associée
à la plus forte amplitude comparativement à celle notée
sur le site OSM (Tableau 6.2). Ceci indique que les conditions de croissance du
site du khroub étaient plus favorables à une meilleure expression
de la durée de la phase végétative. Le manque d'eau et
l'élévation de la température comptent parmi les facteurs
qui ont pour effet de raccourcir la durée du cycle de la phase
végétative. Des températures plus douces allongent la
durée de la phase végétative [133].
Avec une moyenne de 141.7 jours, la durée de la phase
végétative est encore plus longue sur le site de Sétif
(SET). La valeur minimale de 138.0 jours est plus supérieure à la
durée la plus longue sur les sites du KHR et OSM (Tableau 6.2). Il
semble que l'altitude et la continentalité du climat ont fortement
affecté le développement de la plante sur ce site, qui montre
cependant une plus faible amplitude, liée au fait que les stress de fin
de cycle sont plus intenses et surviennent rapidement, une fois
l'épiaison réalisée [134]. Le site de Tiaret (TRT)
ressemble plus à celui de Sétif puisque situé sur le
même étage (hauts plateaux) mais à l'ouest du pays. En
effet la durée de la phase végétative sur ce site est plus
proche de celle exprimée sur le site de Sétif. Les valeurs
extrêmes sont de 121.0 et 130.0 jours (Tableau 6.2).
Les génotypes les plus précoces, à
Sétif, sont Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12),
Inter_9/Poho_1 (n°16) et Aaz 77_2/Porron_11//Busca_3 (n°18), et le
plus tardif est le génotype Minimus_7// Auk /Oste /3/ Shag_26 (n°
6) (Appendice C, Tableau 3). Par contre sur le site de Tiaret les
génotypes précoces sont Sn Turk Mi 83-84 375/Nigris_5//
Tantl_1/3/... (n° 9) et Hoggar (n°10) et le plus tardif est le
génotype Altar84 (n°19) (Appendice C, Tableau 4). Ces
résultats montrent que l'expression de la durée de la phase
végétative est variable selon le site expérimental et
aussi selon le génotype. Ils montrent aussi que le génotype
précoce sur un site n'est pas forcement le plus précoce sur les
autres sites. Ceci s'explique par la présence de l'interaction
génotype x milieu pour la durée de la phase
végétative.
68
Exprimées en pourcent de la valeur maximale, la
durée de la phase végétative a logiquement
été la plus longue sur le site de SET, suivi de TRT. Elle a
été la plus courte sur le site de OSM, et intermédiaire
sur le site du KHR (Figure 6.1). L'effet de l'altitude semble le plus
prépondérant pour expliquer ces différences inter sites,
d'expression de la durée de la phase végétative. Les
différences entre génotypes et la sensibilité
génotypique au changement de site est plus liée aux facteurs
génétiques (gènes de précocité,
sensibilité à la photopériode, sensibilité à
la vernalisation) [8].
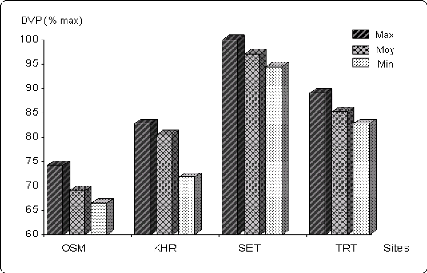
Figure 6.1 : Variation de la durée de la phase
végétative (en % de la valeur maximale) sur les différents
sites.
6.1.1.2 Hauteur de la végétation
L'analyse de la hauteur du chaume montre que Cirta est la
variété la plus haute sur le site OSM, elle présente une
hauteur de 90.1cm, valeur significativement différente de la moyenne de
l'essai qui est de 80.1 cm (Tableau 6.2). Aram-5/Cali//Rascon-37/3/Plata-8
(n° 22) et Gcn/4/D68-1-93A-1A//Ruff/Fg/3/Mtl-5 (n°12) se
caractérisent par une hauteur de végétation la plus courte
de l'essai, avec respectivement des moyennes de 74.8 et 71.2 cm (Appendice C,
Tableau 1). Ceci représente une différence de sélection
pour la
69
hauteur du chaume de 19 cm par rapport à la hauteur
moyenne de Cirta (Tableau 6.2). Ce résultat montre l'importance des
variétés locales du point de vue longueur de la paille (Cirta
ayant comme parent femelle la variété Hedba3). D'après
DJEBRANI (2000) [135], les variétés locales algériennes
sont généralement dites à paille haute, elles ont
également une surface foliaire élevée et une masse
végétative importante.
Sur le site du khroub, les génotypes Minimus_7// Auk
/Oste /3/ Shag_26 (n°6), Tr 32225/Gediz//Ofonto (n°29) et Sula
/RBCE_2/3/Hui //CIT 71/Cii/4/Ryps27_3/... (n°7) ont des tiges plus hautes
avec respectivement 97.7, 94.5 et 94.0 cm. Le génotype
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12) est le plus court avec 80.7
cm. L'amplitude de ce site est de 17cm autour d'une moyenne
générale de 88.5 cm (Tableau 6.2).
D'après BEN ABDELLAH et BEN SALEM (1993) [136], les
variétés à paille haute ont une meilleure adaptation au
déficit hydrique. La préférence des
agriculteurs-éleveurs Algériens va vers les
variétés à paille longue, qui généralement
produisent plus de paille. La paille du blé dur est fortement
demandée comme aliment du bétail. Le bénéfice
apporté par la paille, en année sèche, est souvent plus
important que celui du grain dont le prix est fixé par l'état
[137]. Selon ANNICHIARICO et al., (2005) [131], le rendement
économique
(paille + grain) est souvent supérieur chez les
variétés à paille haute
comparativement aux
génotypes plus nains.
Sur le site de Sétif, la hauteur de la paille est plus
proche de celle exprimée sur le site du khroub, mais les
génotypes les plus hauts sur le site du khroub ne sont pas les
mêmes sur le site de Sétif. En effet, dans cette zone
d'étude, la hauteur de la paille est à l'avantage des
génotypes Rascon/Sla_3/3/Plata_1/Snm//Plata_9 (n°8) et FI 05
(n°23) avec respectivement 97.2 et 95.2cm. Le témoin Boussellem a
une hauteur qui atteint 94.5 cm. La valeur la plus faible de 83.5 cm est
affichée par le génotype Plata_1/Snm // Plata_9/3/Green
(n°14). Sur ce site, plus de la moitie des génotypes testés
ont des hauteurs supérieures à 90.0 cm (Appendice C, Tableau 3),
ce qui indique que
70
les conditions de croissance de cette campagne ont permis une
meilleure expression de ce caractère.
Comparé au site de Sétif, les résultats
indiquent que le site de Tiaret a été moins favorable à
l'expression de la hauteur du chaume. En effet le génotype
Inter_9/Poho_1 (n°16), le plus haut sur ce site, présente une
hauteur proche de celle du génotype le plus court sur les sites du
khroub et Sétif soit 87.1cm (Tableau 6.2).
Exprimées en pourcent de la valeur maximale, la hauteur
de la tige la plus longue a été enregistrée sur les sites
du khroub et de Sétif, suivi par le site d'Oued Smar et enfin le site de
Tiaret a été le dernier pour la valeur maximale de ce
caractère (Figure 6.2).
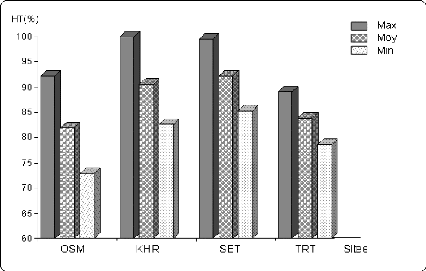
Figure 6. 2 : Variation de la hauteur du chaume (en % de la
valeur maximale) sur les différents sites.
Ces résultats montrent des différences
significatives entre les sites pour l'expression d'un même
caractère, différences de classement et différences de
niveau atteint par les valeurs moyennes du caractère
considéré (Tableau 6.2).
71
Les différences d'expression génotypique, entre
les quatre sites testés, s'expliquent par la variation de la gamme des
conditions entre les quatre milieux, variation qui est une source d'interaction
génotype x milieu. Les variétés de céréales
à pailles hautes sont réputées plus tolérantes
à la sécheresse, suite à la relation qui existe entre la
hauteur de la plante et le développement d'un système racinaire
plus profond. Ceci donne un avantage marqué aux variétés
à pailles hautes, du point de vue extraction d'eau en conditions
sèches, comparativement aux variétés à pailles
courtes [138].
6.1.2 Tallage herbacé et nombre
d'épis/m2
6.1.2.1 Tallage herbacé
L'analyse de la variance indique un effet génotypique
significatif uniquement sur les sites Oued Smar et El Khroub (Tableau 6.3).
L'effet n'est pas significatif sur les sites de Sétif et Tiaret, suite
à une forte erreur résiduelle.
Tableau 6.3 : Carrés moyens des écarts de
l'analyse de la variance de la capacité de tallage herbacé (TLL)
au m2 et du nombre d'épis/m2 (NE) des 4 sites.
|
Variable
|
Source
|
ddl
|
OSM
|
KHR
|
SET
|
TRT
|
|
TLL
|
Génotype
|
29
|
25958.4*
|
43995.5*
|
13832.5ns
|
20549.6ns
|
|
(m-2)
|
Blocs
|
3
|
10072.5
|
56586.3
|
131869
|
148690
|
|
Résiduelle
|
87
|
16342.8
|
24906.6
|
12893.1
|
18429
|
|
NE
|
Génotype
|
29
|
10722.9***
|
8885.32**
|
5088.4ns
|
7733.25ns
|
|
(m-2)
|
Blocs
|
3
|
771.32
|
2481.11
|
12898.5
|
31113
|
|
Résiduelle
|
87
|
203.07
|
4461.71
|
3881.7
|
5486.26
|
ns, * et *** = effet non significatif et significatif au seuil de
5 et 1% respectivement.
Sur le site Oued Smar, le nombre de talles herbacées,
produites par unité de surface semée, varie de 478.7 à
810.0 talles/m2 autour d'une moyenne de 655.7 talles/m2
et une Ppds de 179.6 (Tableau 6.4). La capacité de tallage
herbacé est indicatrice de celle des épis par unité de
surface, et donc du rendement en grains attendu. Plus une variété
talle, plus son potentiel de
Sur le site du Khroub, la capacité de tallage de
l'ensemble des génotypes testés est élevée, elle
varie de 640.0 à 1136.3 talles/m2. L'amplitude est
très
72
production est élevé, pour peu que les
conditions de croissance qui viennent après la période de tallage
soient favorables à la montée du potentiel
épis/m2.
A Oued Smar, Cirta et Boussellem/Ofonto//Waha (n° 27)
présentent les moyennes les plus élevées avec
respectivement 810.0 et 802.5 talles/m2. La valeur minimale est
observée chez Boussellem avec 478.8 talles/m2 (Appendice C,
Tableau 1), en effet le croisement Boussellem/Ofonto//Waha a permis
d'améliorer le nombre de talles par unité de surface de leur
parent Boussellem. Selon GATE (1995) [45], en l'absence de facteurs limitants,
le tallage herbacé dépend essentiellement du nombre de plants
installés et de la durée effective du tallage, exprimée en
phyllotherme. Cette durée s'accroît au fur et à mesure que
le semis est précoce et la variété est tardive au stade
épi1cm.
Tableau 6.4 : Valeurs moyennes caractéristiques de la
capacité de tallage herbacé (TLL) et du nombre
d'épis/m2 (NE) des différents sites.
|
Sites
|
Valeurs
|
TLL/m2
|
NE/m2
|
|
OSM
|
Max
|
810.0
|
497.5
|
|
Min
|
478.8
|
267.5
|
|
Amplitude
|
331.3
|
230.0
|
|
Moyenne
|
655.8
|
386.0
|
|
Ppds5%
|
179.6
|
123.1
|
|
KHR
|
Max
|
1136.3
|
536.3
|
|
Min
|
640.0
|
353.8
|
|
Amplitude
|
496.3
|
182.5
|
|
Moyenne
|
860.5
|
451.3
|
|
Ppds5%
|
221.8
|
93.8
|
|
SET
|
Max
|
890.0
|
512.5
|
|
Min
|
626.3
|
353.8
|
|
Amplitude
|
263.8
|
158.8
|
|
Moyenne
|
770.5
|
413.5
|
|
Ppds5%
|
159.6
|
87.6
|
|
TRT
|
Max
|
1012.5
|
553.8
|
|
Min
|
766.3
|
332.5
|
|
Amplitude
|
246.3
|
221.3
|
|
Moyenne
|
904.1
|
423.4
|
|
Ppds5%
|
190.8
|
104
|
73
élevée, prenant une valeur de 496.3
talles/m2, soit l'équivalent du nombre d'épis produit
par m2. La Ppds est de 221.8 talles/m2 (Tableau 6.4). Les
génotypes qui se classent en tête pour cette variable sont
Boussellem/Ofonto//Waha (n°27), Plata_1/Snm// Plata_9/3/Green (n°14)
et FI 02 (n°24), avec respectivement des moyennes de 1136.3, 1037.5 et
1005.0 talles/m2 (Appendice C, Tableau 2).
Ces valeurs sont assez élevées au niveau de
cette zone malgré leur semis tardif, ceci ne peut s'expliquer que par la
pluviométrie enregistrée à ce stade. Les basses
températures durant le mois de Janvier à Mars qui ont ralenti la
durée de tallage, ceux qui favorisent la formation d'un plus grand
nombre de talles. Cette idée peut être complétée par
HION et al., (2002) [139] qui trouvent que le nombre moyen de talles
herbacées régresse avec la tardivité du semis,
contrairement, l'apport en eau l'augmente.
Les génotypes qui produisent le moins de talles
herbacées sont Mck_2/Aco89 (n°2) et Ting7/2*Green-3 (n°26),
avec des moyennes respectives de 640.0 et 667.0 talles/m2 (Appendice
C, Tableau 2).
Sur les sites de Sétif et de Tiaret, suite à la
valeur élevée de la résiduelle, les différences
entre génotypes ne sont pas significatives, d'où la conclusion
que tous les génotypes testés ont la même capacité
de tallage herbacé.
En comparaison des quatre sites d'étude, la
capacité de tallage herbacé s'est mieux exprimée sur le
site du Khroub, elle est moindre sur le site d'Oued Smar (Figure 6.3).
74
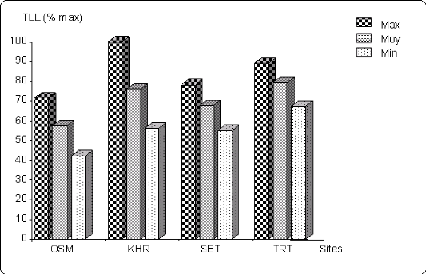
Figure 6.3 : Variation du nombre de talles herbacées
(en % de la valeur maximale) sur les différents sites.
La capacité de tallage herbacé est une variable
désirable uniquement lorsque les conditions de croissance conduisent
à une liaison assez étroite entre cette variable et le nombre
d'épis/m2 qui conditionne le rendement en grains. Sous le
stress de nature intermittente, une production élevée de talles
herbacées est vue comme une surconsommation du stock d'eau du sol,
laissant peu pour la montée des épis et pour un bon remplissage
du grain [140], [141].
6.1.2.2 Nombre d'épis par m2
Comme pour la capacité de tallage herbacé, le
nombre d'épis/m2, montre un effet génotypique
significatif sur les sites Oued Smar et Khroub uniquement et non significatif
sur les sites de Sétif et Tiaret (Tableau 6.3). Cette variable varie de
267.5 à 497.5 épis/m2 sur le site OSM, avec une
moyenne de 386.0. Sur le site KHR, la moyenne est de 451.3
épis/m2, variant de 353.8 et 536.3 épis/m2
(Tableau 6.4). Les génotypes qui affichent les meilleures moyennes sont
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12) et Altar84 (n°19) sur
le site OSM, et Bichena/Ariza_2//Solga_8 (n°3) et Gcn/4/D68-1-93A-1A//Ruff
/ Fg /3/ Mtl-5 (n°12) sur le site KHR (Appendice C, Tableau 1 et 2). Ces
résultats indiquent que
75
Gcn/4/D68-1-93A-1A//Ruff/Fg/3/Mtl-5 montre une adaptation de
la capacité de production des épis sur les deux sites OSM et KHR.
Par contre les génotypes Altar84 (n°19) et Bichena/Ariza_2//Solga_8
(n°3) montrent une adaptation spécifique.
Les génotypes qui extériorisent un faible
potentiel de production d'épis/m2, sont Ofonto/Waha//MBB
(n°28) et MBB/Ofonto//Radiosso/Waha (n°30) sur les deux sites. Sur le
site OSM, les témoins se classent en dernière position avec un
nombre moyen d'épis/m2 de 341.5 (Appendice C, Tableau 1 et
2). Ceci suggère que les nouvelles sélections apportent un plus,
désirable pour cette composante.
Le nombre d'épis/m2 présente des
moyennes de 413.5 et 423.4 épis/m2 sur les sites Sétif
et Tiaret (Tableau 6.4). L'analyse de la variance ne montre pas d'effet
génotypique significatif, de ce fait, tous les génotypes
extériorisent des capacités similaires.
La comparaison des sites pour l'expression du nombre
d'épis/m2, montre que c'est le site de OSM qui
présente la plus grande différence entre les génotypes, ce
site est donc plus discriminant. Les meilleures valeurs moyennes sont
notées sur le site de TRT, pour la valeur maximale, la meilleure moyenne
de l'essai est notée sur le site KHR et la plus faible moyenne est
notée sur le site OSM (Figure 6.4). Ceci indique que des
différences entre génotypes ne sont pas constantes d'un lieu-
test à un autre.
76
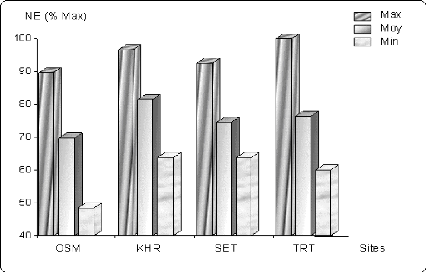
Figure 6.4 : Variation du nombre d'épis/m2
(en % de la valeur maximale) sur les différents sites.
6.1.3 Longueur de l'épi et nombre de grains par
épi
6.1.3.1 Longueur de l'épi
L'analyse de la variance de la longueur de l'épi
effectué par site indique un effet génotypique très
hautement significatif sur le site d' Oued Smar, significatif sur le site du
khroub et un effet génotypique non significatif sur les sites de
Sétif et de Tiaret. (Tableau 6.5).
La comparaison des différents génotypes du point
de vue longueur de l'épi sur le site OSM montre que Cirta et Minimus_7//
Auk /Oste /3/ Shag_26 (n°6) présentent les valeurs les plus
élevées avec respectivement 8.9 et 8.4 cm. La moyenne
générale de l'essai est de 7.7cm. Le génotype Silver_26 /
Toska_26 (n°4) présente la plus faible longueur, soit 6.6 cm
(Tableau 6.6).
77
Tableau 6.5 : Carrés moyens des écarts de
l'analyse de la variance de la longueur de l'épi (LE) et le nombre de
grains par épi (NGE) des 4 sites.
|
Variable
|
Source
|
ddl
|
OSM
|
KHR
|
SET
|
TRT
|
|
LE
|
Génotype
|
29
|
0.93***
|
1.9*
|
0.7ns
|
0.44ns
|
|
(cm)
|
Blocs
|
3
|
0.95
|
2.9
|
0.32
|
4.73
|
|
Résiduelle
|
87
|
0.25
|
1.0
|
0.27
|
0.68
|
|
NGE
|
Génotype
|
29
|
71.14***
|
97.7***
|
62.4***
|
44.83***
|
|
Blocs
|
3
|
28.44
|
39.25
|
0.85
|
73.09
|
|
Résiduelle
|
87
|
20.82
|
24.3
|
1.53
|
14.44
|
ns, * et *** = effet non significatif et significatif au seuil de
5 et 1% respectivement.
Tableau 6.6 : Valeurs moyennes caractéristiques de la
longueur de l'épi (LE) et du nombre de grains par épi (NGE) des
différents sites.
|
Sites
|
Valeurs
|
LE (cm)
|
NGE
|
|
OSM
|
Max
|
8.9
|
54.0
|
|
Min
|
6.6
|
33.3
|
|
Amplitude
|
2.3
|
20.8
|
|
Moyenne
|
7.7
|
46.1
|
|
Ppds5%
|
0.7
|
6.4
|
|
KHR
|
Max
|
8.8
|
51.8
|
|
Min
|
6.0
|
30.8
|
|
Amplitude
|
2.8
|
21.0
|
|
Moyenne
|
7.1
|
42.5
|
|
Ppds5%
|
1.4
|
6.9
|
|
SET
|
Max
|
6.8
|
44.0
|
|
Min
|
5.4
|
27.8
|
|
Amplitude
|
1.4
|
16.3
|
|
Moyenne
|
5.9
|
35.5
|
|
Ppds5%
|
0.73
|
5.61
|
|
TRT
|
Max
|
7.3
|
39.5
|
|
Min
|
5.8
|
25.5
|
|
Amplitude
|
1.5
|
14.0
|
|
Moyenne
|
6.4
|
32.5
|
|
Ppds5%
|
1.16
|
5.3
|
Sur le site du khroub, dont la moyenne générale
de la longueur de l'épi est de 7.1 cm, Ting7/2*Green-3 (n°26) et
Aram-5/Cali//Rascon-37/3/Plata-8 (n°22) ont les épis les plus
longs, avec respectivement 8.8 et 8.0 cm. La plus
78
faible longueur de l'épi est observée chez
Silver_26 / Toska_26 (n°4) et D68-1-93A-1A //Ruff/Fg/3/Mtl-5/4/Lahn
(n°21) (Appendice C, Tableau 2).
Sur les sites de Sétif et Tiaret, les
différences entre les génotypes testés, pour cette
variable, ne sont pas significatives.
La variation des valeurs extrêmes de la longueur de
l'épi (exprimée en % de la valeur maximale observée), sur
les quatre sites d'étude, indique que la meilleure valeur maximale a
été enregistrée sur le site de OSM, suivi par le site du
KHR. La plus faible valeur maximale a été notée sur le
site de SET qui a donné aussi la plus faible valeur moyenne et la plus
faible valeur minimale. Les meilleures valeurs moyennes sont notées sur
le site de OSM (Figure 6.5).
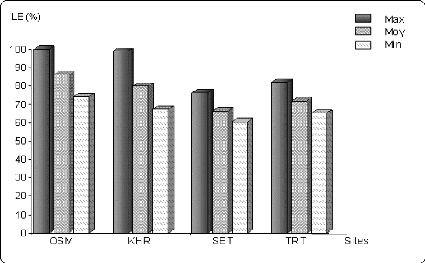
Figure 6.5 : Variation de la longueur de l'épi (en % de
la valeur maximale) sur les différents sites.
6.1.3.2 Nombre de grains par épi
Bien que la fertilité de l'épi soit un
caractère variétal, elle dépend des conditions climatiques
de la pré et post-anthèse [142]. L'analyse de la variance du
nombre de grains par épi effectué par site et pour les quatre
sites testés indique
79
un effet génotypique très hautement significatif
sur les quatre sites d'étude (Tableau 6.5). Les résultats du site
Oued Smar montrent que le nombre moyen de grains par épi est de 46.1
grains. Le nombre de grains par épi varie de 33.3 pour FI 05 (n°23)
à 54.0 grains par épi pour Ofonto/Waha//MBB (n°28), soit une
amplitude de 20.7 grains par épi avec une Ppds de 6.4 (Tableau 6.6).
En outre, les génotypes Aaz 77_2/Porron_11//Busca_3
(n°18), Minimus/ Rascon_19 (n°1), Silver_26/Toska_26 (n°4) et
MBB/Ofonto//Radiosso/Waha (n°30) sont intéressants, car ils ont des
valeurs comprises entre 53.0 et 49.0 grains par épi, proches de la
valeur de la meilleur variété (Appendice C, Tableau 1).
La réduction du nombre d'épis par m2
est compensée par l'augmentation du nombre de grains par épi chez
les génotypes Minimus/ Rascon_19 (n°1), Hoggar et Ofonto/Waha//MBB
(n°28), ce dernier génotype donne le nombre de grains par
épi le plus élevé. BENBELKACEM et al., (1984)
[143] rapportent que les lignées à faible nombre de talles
compensent le désavantage du faible nombre d'épis par la
production d'un nombre élevé de grains par épi.
Sur le site du khroub, le nombre de grains par épi
varie de 49.5 à 51.8 grains par épi pour les génotypes
Kucük (n°17), Aram-5/Cali//Rascon-37/3/Plata-8 (n°22),
Inter_9/Poho_1 (n°16), Srn_3 / Nigris_4 // Porto_5 (n°11) et Altar84
(n°19). Parmi les témoins Boussellem présente le meilleur
nombre de grains par épi avec une moyenne de 44.7 grains par épi
(Appendice C, Tableau 2).
Le génotype Plata_1/Snm // Plata_9/3/Green (n°14)
a le nombre de grains par épi, le plus faible de 30.8 grains par
épi. Ce génotype présente le nombre de talles par
m2 le plus élevé. Ceci conforte l'idée de GATE
(1995) [45] qui mentionne qu'un excès de talles herbacées suivi
de conditions climatiques particulièrement favorables affecte le nombre
de grains par épi.
Sur le site expérimental de Sétif, la mauvaise
répartition de la pluie durant les phases critiques d'installation de la
culture et de remplissage du grain, a eu un effet négatif sur le
tallage-épi, la fertilité et le poids de 1000 grains. Le
génotype
80
Aram-5/Cali//Rascon-37/3/Plata-8 (n°22) est classé
en tête avec 44.0 grains par épi et Kucük (n°17) a le
nombre de grains par épi le plus faible, 27.8 grains par épi.
L'amplitude est de 16.3 grains et la Ppds est de 5.61 (Tableau 6.6). Le
témoin Cirta a une moyenne de 41 grains par épi, ce qui le classe
en deuxième position parmi les génotypes testés (Appendice
C, Tableau 3).
Sur le site de Tiaret, le nombre de grains par épi
varie de 25.5 pour le génotype Rascon/Sla_3/3/Plata_1/Snm//Plata_9
(n°8) à 39.5 grains par épi pour Cirta, soit une amplitude
de 14.0 grains. La Ppds est de 5.3, alors que la moyenne générale
de l'essai est de 32.5 grains par épi (Tableau 6.6). D'après
GRIGNAC (1981) [144], les basses températures pendant la montaison
causent la régression des talles montantes et peuvent engendrer des
pertes de rendement en grains considérables. La méiose pollinique
et la phase qui l'encadre de quelques jours apparaissent comme
extrêmement sensibles à des stress climatiques : des basses
températures, un défaut de rayonnement, un excès thermique
ou un stress hydrique peuvent produire les mêmes effets : un
déficit de grains par épi consécutif à une absence
de fécondation sans limiter pour autant la croissance [45].
Le froid et les faibles rayonnements induisent la
stérilité des gamètes mâles et leurs effets sont
variables selon la variété, pouvant atteindre 50% de perte de
grains par épi chez les variétés sensibles [45]. Les
génotypes Rascon/Sla_3/3/Plata_1/Snm//Plata_9 (n°8), Sn Turk Mi
83-84 375/Nigris_5// Tantlo_1/3/... (n°9), Kucük (n°17), Sula
/RBCE_2/3/Hui //CIT 71/Cii/4/Ryps27_3/... (n°7) et Ofonto/Waha//MBB
(n°28), présentent une faible fertilité, semblent sensible
au froid caractéristique de cette zone d'altitude (Appendice C, Tableau
4).
La variation du nombre de grains par épi d'un site
à l'autre, exprimé en pourcent de la valeur maximale, montre que
le site d'Oued Smar a été la plus favorable à l'expression
du nombre de grains par épi. Le site du khroub qui a été
aussi classé le deuxième pour les trois valeurs qui ont
été un peu plus proche de celles d'Oued Smar (Figure 6.6). Ces
valeurs sont intermédiaires sur le site de Sétif. Contrairement,
le site de Tiaret a donné le plus faible pourcentage pour les trois
valeurs maximale, moyenne et minimale (Figure 6.6).
81
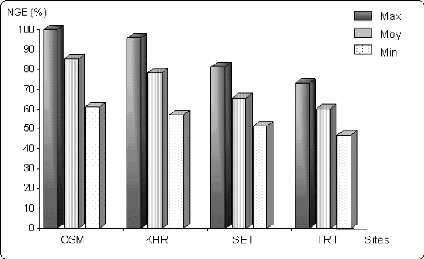
Figure 6.6 : Variation du nombre de grains par épi (en %
de la valeur maximale) sur les différents sites.
6.1.4 Poids de 1000 grains et le rendement en
grains
6.14.1 Poids de 1000 grains
Parmi les composantes du rendement, le poids de mille grains
est une caractéristique purement variétale et qui est très
fluctuant en fonction des conditions de la fertilité du sol, des
techniques culturales appliquées et surtout des facteurs climatiques
[145]. L'analyse de la variance pour le poids de mille grains a
révélé un effet génotypique très hautement
significatif concernant les quatre sites d'étude (Tableau 6.7).
Sur le site d'Oued Smar, la valeur du poids de mille grains la
plus élevée est celle du génotype Sula /RBCE_2/3/Hui //CIT
71/Cii/4/Ryps27_3/... (n°7) avec 47.1 g. Les génotypes qui donnent
des poids de mille grains les plus faibles, sont Inter_9/Poho_1 (n°16) et
Altar84 (n°19) avec respectivement 25.57 et 24.82 g (Appendice C, Tableau
1). D'après HAMADACHE et al., (1998) [146], la rouille brune
cause souvent, au niveau de la zone littorale sub-humide et en cas d'attaque
précoce, une chute de la fertilité de l'épi et
l'échaudage du grain.
82
La moyenne générale de l'essai est de 33.35 g
avec une amplitude de 22.24 g et la Ppds est de 5.4 (Tableau 6.8). La
réduction de la fertilité est compensée par l'augmentation
du poids de mille grains chez les génotypes Sula/ RBCE_2/3/Hui //CIT
71/Cii/4/Ryps27_3/... (n°7), D68-1-93A-1A//Ruff /Fg /3/ Mtl-5/4/Lahn
(n°13) et Mck_2/Aco89 (n°2).
Tableau 6.7 : Carrés moyens des écarts de
l'analyse de la variance du poids de mille grains (PMG) et du rendement en
grains (q/ha) des 4 sites.
|
Variable
|
Source
|
ddl
|
OSM
|
KHR
|
SET
|
TRT
|
|
PMG
|
Génotype
|
29
|
115.15***
|
59.8***
|
53.5***
|
18.4***
|
|
(g)
|
Blocs
|
3
|
27.04
|
31.5
|
4.2
|
2.6
|
|
Résiduelle
|
87
|
15.29
|
14.1
|
1.4
|
1.86
|
|
RDT
|
Génotype
|
29
|
316.88***
|
74.9***
|
73.72***
|
9.5***
|
|
(q/ha)
|
Blocs
|
3
|
222.57
|
41.82
|
13.94
|
3.54
|
|
Résiduelle
|
87
|
41.20
|
10.12
|
1.62
|
2.0
|
ns, * et *** = effet non significatif et significatif au seuil de
5 et 1% respectivement.
Les valeurs du poids de mille grains varient entre 34.6 g pour
Inter_9/Poho_1 (n°16) et 48.0 g pour Mck_2/Aco89 (n°2), sur le site
du khroub (Tableau 6.8). Plus des 2/3 des génotypes ont un poids de
mille grains supérieur à 40.0 g. Ceci indique que, pour cette
zone d'étude, le poids de mille grains est élevé,
malgré l'occurrence des siroccos du mois de Juin avec des
températures de 38°C, qui ont plus touché les
génotypes tardifs, Srn_3 / Nigris_4 // Porto_5 (n°11) et Kucük
(n°17) (Appendice C, Tableau 2).
WARDLAW (2002) [147] mentionne que certains génotypes
minimisent l'effet des hautes températures sur le poids de mille grains
en réduisant le nombre de grains par épi. Parmi les
témoins, Waha a le plus faible poids de 1000 grains et Boussellem a un
poids de 1000 grains élevé, 47.99 g (Appendice C, Tableau 2).
D'autre part, le site de Sétif ressemble plus à celui du khroub,
en effet, le poids de mille grains sur ce site est plus proche de celui
exprimé sur le site du khroub. Les moyennes générales sont
de 43.00 g et 42.6 g pour les deux sites (Tableau 6.8).
83
Tableau 6.8 : Valeurs moyennes caractéristiques du
poids de 1000 grains (PMG) et du rendement en grains (RDT) des
différents sites.
|
Sites
|
Valeurs
|
PMG (g)
|
RDT (q/ha)
|
|
OSM
|
Max
|
47.1
|
51.2
|
|
Min
|
24.8
|
17.0
|
|
Amplitude
|
22.2
|
34.2
|
|
Moyenne
|
33.3
|
30.6
|
|
Ppds5%
|
5.4
|
9.0
|
|
KHR
|
Max
|
48.0
|
65.6
|
|
Min
|
34.6
|
44.8
|
|
Amplitude
|
13.4
|
20.8
|
|
Moyenne
|
43.0
|
54.5
|
|
Ppds5%
|
5.3
|
11.3
|
|
SET
|
Max
|
49.7
|
57.9
|
|
Min
|
35.4
|
38.9
|
|
Amplitude
|
14.3
|
19.0
|
|
Moyenne
|
42.6
|
49.4
|
|
Ppds5%
|
3.31
|
4.64
|
|
TRT
|
Max
|
25.7
|
13.1
|
|
Min
|
17.0
|
7.2
|
|
Amplitude
|
8.7
|
5.8
|
|
Moyenne
|
20.4
|
9.9
|
|
Ppds5%
|
5.5
|
3.4
|
Le génotype FI 05 (n°23) a la valeur moyenne la
plus élevée, sur le site de Sétif avec 49.75 g, suivi de
Boussellem (48.78 g). Tr 32225/Gediz//Ofonto (n°29), Inter_9/Poho_1
(n°16), Hoggar, Sn Turk Mi 83-84 375/Nigris_5// Tantlo_1/3/... (n°9),
Ofonto/Waha//MBB (n°28) et Bichena/ Ariza_2//Solga_8 (n°3) qui
donnent des poids de mille grains assez proches allant de 45.0 à 47.0 g.
Cependant, les génotypes Mck_2/Aco89 (n°2) et D68-1-93A-1A//Ruff
/Fg /3/ Mtl-5/4/Lahn (n°13) semblent remplir moins bien ses grains que les
autres génotypes testés (Appendice C, Tableau 3).
Le poids de mille grains est réduit par le
déficit hydrique de post-floraison, suite à l'altération
de la durée de remplissage par l'accélération de la
sénescence foliaire. BENBELKACEM et KELLOU, (2000) [148] montrent qu'un
manque d'eau après floraison combiné aux températures
élevées entraîne une diminution du poids de mille grains
par altération de la vitesse et/ou de la durée de remplissage ce
qui se traduit par l'échaudage des grains.
84
Selon TRIBOI et al., (1985) [149], le flux des
assimilats vers les grains dépend d'une part de la quantité
d'assimilats stockée dans les tiges et d'autre part de l'assimilation
post-anthèse. Cette quantité est fonction du nombre de tiges
produites par m2, du type de variété, de la hauteur du
chaume et des conditions climatiques spécifiques de l'année. En
outre, en conditions de stress hydrique, une paille haute est plus apte
à stocker plus de réserves glucidiques, les quels sont
susceptibles d'être transférer vers le grain au cours du
remplissage [136]. Dans ce cas de notre essai, les deux premières
variétés qui donnent le poids de mille grains le plus
élevé celles qui caractérisent parmi les plus hautes mais
ce ci ne suffit pas de donner un meilleur rendement chez ces deux
variétés à cause du faible nombre d'épis par
m2 et leurs nombre moyen de grains par épi.
Les valeurs de poids de mille grains notées sur le site
de Tiaret indiquent que les génotypes Silver_26 / Toska_26 (n°4),
Ting7/2*Green-3 (n°26), Rascon/Sla_3/3/Plata_1/ Snm//Plata_9 (n°8) et
Inter_9/Poho_1 (n°16) viennent en tête de classement avec des
valeurs qui varient de 22.0 à 25.7 g (Appendice C, Tableau 4). Ces
moyennes sont inférieures aux valeurs minimales enregistrées sur
les sites du khroub et Sétif et proches de celles d'Oued Smar. Le
génotype Boussellem/Ofonto//waha (n°27) présente un poids de
1000 grains de 17.00 g et qui est le plus faible sur ce site (Tableau 6.8).
Les conditions climatiques de ce site, associées
à un semis tardif qui décala la phase de remplissage du grain et
par conséquence elle coïncide avec le déficit hydrique de la
fin du cycle (Mai) qui affecte fortement le poids de mille grains.
MEKHLOUF et al., (2001) [150] montrent que la
période pluvieuse est associée à une faible demande
climatique mais aussi de basses températures hivernales qui
décalent la croissance active de la plante vers le printemps, moment
où la demande climatique augmente et la pluie devient plus rare. La
migration de substances de réserve accumulées
antérieurement dans les feuilles et la tige (amidon, protéines) a
lieu essentiellement au cours de cette période.
85
La comparaison des quatre sites d'étude pour
l'expression du poids de mille grains exprimé en pourcent de la valeur
maximale montre que le site de Sétif a été le plus
favorable à l'expression du poids de mille grains pour la valeur
maximale suivi par le site du khroub et Oued Smar qui ont donné des
valeurs maximales plus proches à celle de Sétif (Figure 6.7). La
valeur maximale sur le site de Tiaret a été la plus faible. Les
meilleures valeurs moyennes ainsi que les valeurs minimales sont notées
sur le site du khroub et Sétif. La plus faible valeur minimale est
notée sur le site de Tiaret (Figure 6.7).
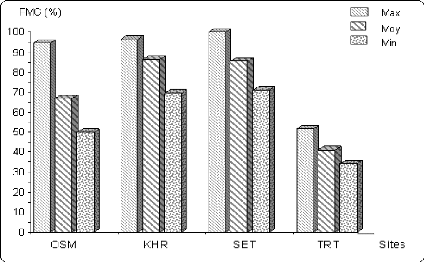
Figure 6.7 : Variation du poids de mille grains (en % de la
valeur maximale) sur les différents sites.
6.1.4.2 Rendement en grains
L'analyse de la variance par site ou localité montre un
effet génotypique très hautement significatif, suggérant
de la variabilité génétique à exploiter en
sélection (Tableau 6.7). L'effet significatif indique que, dans chaque
site, des différences génotypiques existent entre les
différentes lignées testées. Ceci montre que la
capacité de production est différente d'un génotype
à l'autre et d'un milieu à l'autre.
86
En effet, pour Oued Smar, Gcn/4/D68-1-93A-1A//Ruff / Fg /3/
Mtl-5 (n°12) présente le rendement le plus élevé avec
51.2 q/ha. La moyenne de l'essai sur ce site est de 30.6 q/ha avec une
amplitude de 34.2 q/ha et la Ppds est de 9.0 (Tableau 6.6). Les rendements les
plus faibles sont observés chez MBB/Ofonto// Radiosso/Waha (n°30),
Ofonto/Waha//MBB (n°28), Inter_9/Poho_1 (n°16) et
Aram-5/Cali//Rascon-37/3/ Plata-8 (n°22) avec respectivement 19.9, 19.1,
19.1 et 17.0 q/ha (Appendice C, Tableau 1).
Avec une moyenne générale de l'essai de 54.5
q/ha, le rendement le plus élevé sur le site du khroub est celui
de Bichena / Ariza_2//Solga_8 (n°3), avec 65.6 q/ha. Il faut noter que, le
rendement dépasse les 50.0 q/ha chez 25 génotypes parmi les 30
testés sur le site du khroub. Le plus faible rendement est celui du
témoin Boussellem avec 44.8 q/ha. L'amplitude du site est de 20.8 q/ha,
avec une Ppds de 11.3 (Tableau 6.8).
Ces résultats s'expliquent par les conditions
climatiques qui étaient favorables. La période de Mars à
Mai a enregistré un cumul pluviométrique de 210.1 mm. Le
désherbage chimique ainsi que les potentialités
élevées du milieu et l'absence des gelées
printanières ont favorisé l'expression du potentiel de rendement.
D'après BOUZERZOUR et al., (2000) [100], l'amélioration
de la production est possible en adoptant un itinéraire technique plus
intensif dans les plaines intérieures où les conditions de
production sont nettement plus favorables et par l'adoption de cultivars plus
flexibles vis-à-vis des contraintes climatiques.
L'étude des valeurs moyennes du rendement, sur le site
de Sétif, montre que le génotype Bichena/Ariza_2//Solga_8
(n°3) se distingue par le rendement en grains le plus élevé,
57.9 q/ha. Minimus/Rascon_19 (n°1) et Minimus_7// Auk /Oste /3/ Shag_26
(n°6) produisent le moins (Appendice C, Tableau 3).
La moyenne générale de l'essai est de 49.4 q/ha
(Tableau 6.8) qui reste une valeur importante malgré la mauvaise
répartition de pluviométrie durant les phases critiques, ceci
peut être expliqué par la compensation entre les caractères
relatives aux rendement d'une part et la capacité de certains
génotypes
87
à maintenir l'activité photosynthétique
sous stress hydrique, le plus long temps possible, par l'utilisation des
assimilats stockées dans les tiges d'autre part. Selon BAHLOULI et
al., (2008) [151] le rendement est lui-même la résultante des
sites, du nombre de grains par m2 (épis par m2 et
grains par épi) et de leur remplissage.
L'impact des conditions climatiques (les basses
températures et le déficit hydrique de fin du cycle) du site
expérimental de Tiaret s'est répercuté sur les composantes
du rendement ce qui a affecté énormément le rendement
final. En effet, les rendements obtenus ont été tellement faibles
qu'ils ne permettent pas de se prononcer sur le niveau de productivité
des génotypes (Appendice C, Tableau 4). Les niveaux de rendement varient
de 7.2 q/ha pour Rascon/Sla_3/3/Plata_1/Snm//Plata_9 (n°8) à 13.1
q/ha pour Inter_9/Poho_1 (n°16). La moyenne générale de
l'essai est de 9.9 q/ha (Tableau 6.8).
Selon MECKLOUF et al., (2006) [134] si
l'épiaison coïncide avec l'avènement de températures
minimales trop basses, au cours du mois d'Avril, le potentiel de production est
réduit. Ceci semble être le cas pour ce site. COVENTRY et al.,
(1993) [152] montrent aussi que le rendement diminue avec les semis
tardifs.
Exprimé en pourcent de la valeur maximale. Le rendement
a été le plus élevé sur le site du khroub, suivi de
Sétif, il est moyen sur le site d'Oued Smar et le plus faible a
été noté sur le site de Tiaret (Figure 6.8).
88
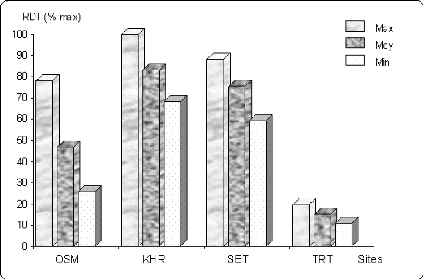
Figure 6.8 : Variation du rendement en grains (en % de la
valeur maximale) sur les différents sites.
6.2 Etude des liaisons intra et inter sites
entre variables
6.2.1 Liaisons intra site
L'objectif de l'étude des liaisons entre
caractères aide à identifier les traits qui varient ensemble et
ceux qui sont indépendants. La sélection sur la base d'un
caractère engendre forcément des changements dans les moyennes
des caractères qui lui sont liés, dans le sens positif ou
négatif selon le signe de la corrélation. Par contre les
changements chez les caractères non liées peuvent être
positifs, négatifs ou neutres ; selon le génotype et les
conditions.
Dans la présente étude, sur le site Oued Smar,
le nombre d'épis/m2 est positivement corrélé
avec le nombre de talles herbacées, ce qui paraît logique. La
hauteur du chaume l'est positivement avec la durée de la phase
végétative, la longueur de l'épi avec le poids de 1000
grains ainsi que le poids de 1000 grains avec le rendement en grains (Tableau
6.9).
89
Ces résultats suggèrent que sur ce site, les
génotypes qui produisent beaucoup de talles herbacées, produisent
aussi plus d`épis/m2. La sélection précoce sur
la capacité de tallage herbacé engendre celle de
l'épis/m2, avec le jeu de variétés
étudiées. Les génotypes tardifs au stade épiaison,
présentent une hauteur de végétation plus importante.
D'après BERGER (1985) [153] La précocité au stade
épiaison est associée à la réduction de la hauteur
du chaume et à l'augmentation de la capacité de tallage
épi.
Les génotypes dont l'épi est long, se
caractérisent aussi par un poids de 1000 grains élevé. Les
génotypes dont le poids de 1000 grains est élevé, donnent
un haut rendement en grains, qui est le plus souvent noté chez les
génotypes dont la durée de la phase végétative est
courte (génotypes précoces), suite à la corrélation
négative entre le rendement et la durée de la phase
végétative (Tableau 6.9). Selon GOSS et al., (1986) in
RIOU, (1993) [154], le rendement dépend de la période de
croissance de la plante et la vitesse à la quelle le feuillage se met en
place.
Sur le site du Khroub, nous notons une corrélation
positive entre la capacité de tallage herbacé et le nombre
d'épis/m2. Ceci traduirait une influence déterminante
du tallage dans les conditions non limitantes. La hauteur du chaume est
positivement corrélée avec le poids de 1000 grains et
négativement avec le nombre de grains par épi. Ces deux
dernières variables sont négativement corrélées
entre elles (Tableau 6.9).
SLAFER et SAVIN, (1994) [155] interprètent la liaison
négative entre le nombre de grains par unité de surface et le
poids de mille grains par, le fait que, l'augmentation du nombre de grains est
induite par une biomasse aérienne élevée au stade
épiaison. Le nombre de grains, en plus, se situe en des endroits
éloignés du tiers médian de l'épi. Ces grains
acquièrent un poids individuel plus faible que celui des grains du tiers
médian. Le poids moyen du grain est globalement réduit, suite
à la répartition de la même quantité d'assimilats
entre un nombre de grains plus élevé.
90
D'autre part, le rendement est corrélé
positivement seulement avec le nombre d'épis/m2 (Tableau
6.9), ceci est en parfait accord avec les conclusions de l'équipe de
cette station qui sélectionnent indirectement pour le rendement sur la
base du nombre d'épis. MONNEVEUX et al., 1993 [156], sont
arrivées aussi à la même conclusion car ils ont
trouvé que les hauts rendements sont associés à un
tallage-épis élevé ou au maintient d'une bonne
fertilité de l'épi.
Il est important de noter que sur ces deux sites d' Oued Smar
et du khroub, il y a peu de ressemblance pour les corrélations inter
caractères, hormis le tallage herbacé et le nombre
d'épis/m2.
Tableau 6.9 : Coefficient de corrélation
phénotypique entre les variables mesurées par site et pour les
quatre sites (en gras coefficients statistiquement significatifs)
|
Trait
Ligne
|
Tall
|
NE
|
PHT
|
LE
|
NGE
|
PMG
|
RDT
|
DVP
|
|
Tall L1
|
|
0.481
|
0.199
|
0.041
|
-0.084
|
-0.179
|
0.081
|
0.358
|
|
L2
|
|
0.441
|
0.318
|
-0.206
|
-0.397
|
0.048
|
0.149
|
-0.051
|
|
NE L1
|
0.023
|
|
-0.085
|
-0.139
|
-0.205
|
-0.156
|
0.227
|
0.125
|
|
L2
|
0.348
|
|
0.163
|
-0.344
|
-0.260
|
-0.097
|
0.333
|
-0.310
|
|
PHT
|
-0.223
|
-0.209
|
|
0.215
|
0.170
|
-0.084
|
-0.091
|
0.362
|
|
-0.200
|
0.108
|
|
-0.210
|
0.440
|
0.381
|
0.279
|
0.107
|
|
LE
|
0.285
|
-0.157
|
0.024
|
|
-0.109
|
0.472
|
0.207
|
0.027
|
|
-0.152
|
-0.240
|
0.027
|
|
0.123
|
-0.215
|
-0.200
|
0.127
|
|
NGE
|
0.157
|
-0.334
|
-0.262
|
0.033
|
|
-0.208
|
0.014
|
-0.092
|
|
-0.207
|
-0.199
|
-0.036
|
0.361
|
|
-0.439
|
-0.115
|
0.239
|
|
PMG
|
-0.058
|
-0.273
|
0.283
|
0.078
|
-0.537
|
|
0.749
|
-0.225
|
|
-0.046
|
-0.065
|
0.170
|
0.055
|
-0.435
|
|
-0.071
|
0.179
|
|
RDT
|
0.318
|
0.030
|
-0.272
|
-0.036
|
0.380
|
0.129
|
|
-0.419
|
|
-0.365
|
-0.030
|
0.323
|
0.229
|
0.437
|
0.062
|
|
-0.165
|
|
DVP
|
0.239
|
-0.011
|
-0.144
|
0.218
|
0.110
|
-0.208
|
-0.027
|
|
|
0.353
|
-0.132
|
-0.378
|
-0.172
|
0.102
|
0.122
|
-0.257
|
|
Au dessus L1 = OSM, L2= KHR, en dessous, L1 = SET, L2 = TRT,
n-2 = 28, r5%= 0.361, r1% =0.463.
Sur le site de Sétif, les seules liaisons
significatives et notables sont entre le poids de 1000 grains et le nombre de
grains par épi et entre le rendement et le nombre de grains par
épi, confirmant ainsi les résultats de GRIGNAC (1973) [157] qui a
trouvé que le nombre de grains par épi est la composante la plus
étroitement liée au rendement. Celle-ci augmente rapidement avec
l'amélioration de l'environnement de la culture.
Le peu de liaisons entre les variables mesurées par
site est suggestif de la variabilité des itinéraires ou
alternatives qui conduisent à l'expression du
91
Les mêmes liaisons sont aussi notées sur le site
de Tiaret où la hauteur du chaume est négativement liée
avec la durée de la phase végétative et le tallage
herbacé est négativement corrélé avec le rendement
(Tableau 6.9).
Il ressort de l'analyse des liaisons inter caractères,
que le nombre d'épis/m2 est dépendant de la
capacité de tallage herbacé sur les deux sites les plus
favorables Oued Smar et Khroub. Sur les sites du Khroub, Sétif et
Tiaret, le poids de 1000 grains se matérialise au détriment du
nombre de grains par épi, probablement par l'effet de concurrence pour
les assimilats. Les conditions climatiques de ces sites ne permettent pas la
réalisation concomitante d'un nombre de grains par épi et d'un
poids élevé de 1000 grains. Sur les sites de Sétif et
Tiaret, les différences de rendement en grains entre génotypes
émergent le plus souvent suite à des différences du nombre
de grains par épi (Tableau 6.9).
En matière de sélection de nouvelles
variétés, le caractère le plus intéressant est
surtout le rendement en grains. L'étude des corrélations indique
que sur le site du Khroub, suite à l'existence d'une liaison entre le
rendement et le nombre d'épi/m2, une sélection
précoce sur la capacité de ce dernier peut contribuer au
classement des génotypes performants. HSU et WALTAN (1971) [158] ont
noté que le nombre d'épi est la composante la plus importante
dans la détermination du rendement.
Sur le site d'Oued Smar, une courte durée de la phase
végétative et un poids élevé de 1000 grains peuvent
aider à l'identification de génotypes à haut rendement en
grains. La variation du rendement grain est, donc, étroitement
liée à l'expression du poids de mille grains pour cette zone
d'étude. MC NEAL et al., (1978) [159] indiquent que le poids du
grain est un bon critère de sélection indirecte pour
améliorer le rendement. Par contre, sur les sites de Sétif et
Tiaret, seul le nombre de grains par épi semble faire la
différence entre génotypes à haut rendement et ceux dont
le potentiel est faible. La fertilité de l'épi peut être
définie comme objectif de sélection [160].
92
rendement en grains chez les génotypes
étudiés. Ceci a pour effet de compliquer la sélection
indirecte, dans un site pour les autres sites et induit la sélection
spécifique à chaque site, sur la seule base du rendement en
grains.
6.2.2 Liaisons inter sites
L'évaluation multi sites des génotypes vise
l'indentification des sites qui sont similaires, nécessitant une
sélection à large adaptation et ceux qui sont distinctifs
nécessitant une sélection spécifique. Dans ce contexte le
degré de ressemblance entre sites pour une même variable
mesurée est important à déterminer. Il permet de grouper
les sites qui se ressemblent et de ne garder pour l'évaluation future
que le site le plus discriminant, et d'identifier les sites spécifiques
[129].
L'analyse des coefficients de corrélation de rang de
Sperman est utilisée pour étudier la ressemblance entre sites
pour une variable donnée. FALCONER (1982) [161] mentionne, à ce
sujet, qu'un caractère mesuré sur deux sites peut être
considéré comme deux caractères différents si la
corrélation entre sites pour ce caractère est non significative.
Dans ce cas la sélection sur la base de ce caractère sur un site
n'engendre pas de réponse corrélative significative sur l'autre
site. Si la corrélation est significative, il y a une réponse
corrélative positive.
L'étude des coefficients de corrélation de rang
de Sperman indique que la durée de la phase végétative, la
hauteur de plante et le poids de mille grains montrent un degré de
ressemblance entre les sites OSM et KHR significatif, un degré de
ressemblance significatif est aussi noté entre les sites de OSM et TRT
concernant la variable nombre d'épi/m2 (Tableau 6.10). Les
deux sites SET et TRT sont similaires pour les deux variables, nombre de grains
par épi et poids de mille grains (Tableau 6.10).
Le reste des variables montrent peu de ressembles entre sites.
Ceci indique que le classement des génotypes pour un caractère
donné sur un site ne suit pas fidèlement le classement du
même caractère sur les autres sites. Ces résultats mettent
en évidence la spécificité des sites d'évaluation
étudiés et suggèrent la présence d'interaction
génotype x milieu pour l'ensemble des variables mesurées.
93
Cette interaction génotype x milieu représente
un des problèmes les plus importants que l'améliorateur doit
résoudre. Si cette interaction est très accentuée il sera
recommandé de créer des variétés
spécifiquement adaptées à chaque type de milieu [11];
[12]; [162].
Tableau 6.10 : Coefficient de corrélation des rangs de
Sperman de la même variable mesurée sur les différents
sites (en gras coefficients statistiquement significatifs)
|
Variables
|
Sites
|
KHR
|
SET
|
TRT
|
|
DVP
|
OSM
|
0.470
|
0.303
|
0.127
|
|
KHR
|
|
0.027
|
0.252
|
|
SET
|
|
|
0.006
|
|
HT
|
OSM
|
0.717
|
0.038
|
0.145
|
|
KHR
|
|
0.098
|
-0.010
|
|
SET
|
|
|
0.336
|
|
TLL
|
OSM
|
0.316
|
-0.206
|
-0.006
|
|
KHR
|
|
-0.064
|
-0.338
|
|
SET
|
|
|
-0.059
|
|
NE
|
OSM
|
0.306
|
0.213
|
0.472
|
|
KHR
|
|
0.316
|
0.103
|
|
SET
|
|
|
0.252
|
|
LE
|
OSM
|
0.222
|
-0.286
|
-0.291
|
|
KHR
|
|
0.127
|
-0.497
|
|
SET
|
|
|
0.118
|
|
NGE
|
OSM
|
0.093
|
-0.134
|
-0.010
|
|
KHR
|
|
-0.071
|
-0.227
|
|
SET
|
|
|
0.580
|
|
PMG
|
OSM
|
0.549
|
-0.394
|
-0.377
|
|
KHR
|
|
-0.155
|
-0.344
|
|
SET
|
|
|
0.487
|
|
RDT
|
OSM
|
0.119
|
-0.129
|
-0.034
|
|
KHR
|
|
0.161
|
-0.035
|
|
SET
|
|
|
0.175
|
Diverses méthodes, paramétriques et non
paramétriques, sont proposées dans la littérature pour
approcher de telles situations. Parmi les méthodes non
paramétriques figurent la méthode de la somme des classements du
caractère d'intérêt dans les différents sites [163].
Parmi les méthodes paramétriques figure la technique de la
régression conjointe et l'approche AMMI (Additive Main effect and
Multiplicative Interaction) [13].
94
Dans ce qui suit, nous concentrons sur l'analyse du rendement
en grains qui est l'objectif principal de la sélection. Nous appliquons
la méthode non paramétrique pour identifier les génotypes
les plus stables et les plus performants. Puis nous utilisons l'analyse AMMI
dans le même sens, tout en décrivant l'architecture des meilleurs
génotypes identifiés (Winner).
6.3 Etude de l'interaction génotype x
environnement du rendement en grains
6.3.1 Identification des génotypes performants
L'utilisation de la méthode des classements des
rendements des différents génotypes par site indique que le
génotype Bichena / Ariza_2//Solga_8 (n°3) se classe
5ième sur le site OSM, 1ier sur les sites KHR et
SET et enfin 7ième sur le site TRT, finissant avec la plus
petite somme de classement de 14. Il se classe premier une moyenne de rendement
inter sites de 43.85 q/ha (Tableau 6.11).
Le second génotype, du point de vue somme des
classements des rendements, est la variété
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12). Cette
variété se classe première sur le site OSM,
5ième sur le site KHR, 3ième sur SET et
6ième sur le site TRT. Elle fait une somme des classements de
15, avec une moyenne des rendements inter sites de 43.6 q/ha (Tableau 6.11).
Ces deux génotypes sont suivis par le témoin Waha qui fait un
score de classements de 40 pour une moyenne de rendement de 37.32 q/ha. Les
performances de ces deux génotypes en comparaison avec celles du
témoin Waha et de la moyenne de tous les génotypes
évalués par site sont données en figure (6.9).
95
Tableau 6.11 : Classement des rendements en grains des
génotypes par site, somme des classements par génotype et moyenne
par génotype des rendements des 4 sites.
|
N° Géno
|
OSM
|
KHR
|
SET
|
TRT
|
? classements
|
RDT moyen
|
|
1
|
4
|
12
|
29
|
8
|
53
|
36.9
|
|
2
|
7
|
21
|
14
|
24
|
66
|
36.9
|
|
3
|
5
|
1
|
1
|
7
|
14
|
43.85
|
|
4
|
8
|
24
|
21
|
11
|
64
|
36.6
|
|
Waha
|
15
|
14
|
2
|
9
|
40
|
37.3
|
|
6
|
11
|
2
|
30
|
19
|
62
|
36.1
|
|
7
|
2
|
20
|
28
|
27
|
77
|
38.6
|
|
8
|
18
|
3
|
11
|
30
|
62
|
36.7
|
|
9
|
9
|
26
|
19
|
18
|
72
|
35.7
|
|
10
|
10
|
19
|
16
|
28
|
73
|
36.3
|
|
11
|
14
|
16
|
8
|
5
|
43
|
37.1
|
|
12
|
1
|
5
|
3
|
6
|
15
|
43.6
|
|
13
|
3
|
23
|
24
|
29
|
79
|
38.4
|
|
14
|
20
|
15
|
7
|
16
|
58
|
35.8
|
|
15
|
6
|
27
|
6
|
14
|
53
|
37.3
|
|
16
|
29
|
8
|
18
|
1
|
56
|
34.5
|
|
17
|
25
|
9
|
27
|
26
|
87
|
32.4
|
|
18
|
16
|
22
|
5
|
4
|
47
|
37.0
|
|
19
|
19
|
10
|
15
|
23
|
67
|
35.6
|
|
20
|
21
|
30
|
23
|
3
|
77
|
32.7
|
|
21
|
12
|
25
|
26
|
12
|
75
|
35.1
|
|
22
|
30
|
17
|
4
|
17
|
68
|
33.6
|
|
23
|
23
|
13
|
25
|
13
|
74
|
34.0
|
|
24
|
17
|
11
|
22
|
10
|
60
|
36.1
|
|
25
|
22
|
6
|
10
|
15
|
53
|
36.0
|
|
26
|
26
|
28
|
17
|
20
|
91
|
32.3
|
|
27
|
13
|
7
|
9
|
22
|
51
|
37.4
|
|
28
|
28
|
29
|
12
|
21
|
90
|
31.8
|
|
29
|
24
|
4
|
13
|
25
|
66
|
35.1
|
|
30
|
27
|
18
|
20
|
2
|
67
|
33.5
|
96
Les performances de ces deux génotypes restent
supérieures ou égales à celles du témoin Waha sur
l'ensemble des sites (Figure 6.9). Ce résultat indique leur large
adaptation relative à l'ensemble des environnements testés.
L'alternative est de faire une sélection plus spécifique, avec le
choix du génotype Bichena / Ariza_2//Solga_8 (n°3) pour les sites
KHR et SET où il se classe 1ier, avec des rendements de 65.6
et 57.9 q/ha. Puis le génotype Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5
(n°12), pour le site OSM avec une moyenne de rendement de 51.25 q/ha, et
enfin le génotype Inter_9/Poho_1 (n°16) pour le site TRT avec un
rendement moyen de 13.1 q/ha, soit une moyenne sur l'ensemble des sites de
46.96 q/ha, ce qui représente un avantage relativement à la
première alternative de 3.26 q/ha = (46.96-43.7).
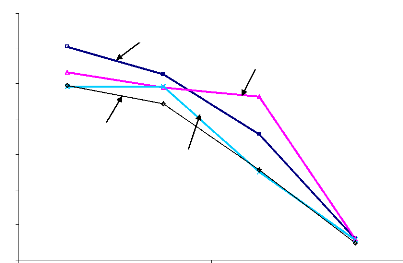
35
Waha
25
15
5
RDT (q ha-1)
Bichena / Ariza_2//Solga_8 (n°3)
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12)
55
45
Moyenne essai
75
65
KHR SET OSM TRT Sites
Figure 6.9 : Performances des génotypes ayant le
meilleur classement sur l'ensemble des sites en comparaison avec les
performances du témoin Waha et de la moyenne des sites.
97
6.3.2 Performances de rendement et importance de
l'interaction G x E
L'analyse de la variance combinée, intégrant
tous les sites, confirme la présence d'une l'interaction significative
pour le rendement en grains et un effet site hautement significatif (Tableau
6.12). Ces résultats indiquent que sous ces conditions pédo-
climatiques, la culture de blé dur est doublement affectée par un
effet localité et par une interaction génotype x localité.
L'effet localité explique 91.78% de la somme des carrés des
écarts, alors que l'effet génotype et l'interaction n'expliquent
que 2.06 et 6.15% de la somme des carrées des écarts des
traitements du rendement en grains (Tableau 6.12).
Tableau 6.12 : Analyse de la variance de la régression
conjointe et de l'AMMI (effet moyen additif et interaction multiplicative) du
rendement en grains.
|
Source de variation
|
ddl
|
SCE
|
CME
|
F-obs
|
F-test
|
|
Traitements
|
119
|
1651.46
|
13.88
|
74.41
|
**
|
|
Localité (L)
|
03
|
1515.78
|
505.26
|
315.95
|
**
|
|
Bloc (localité)
|
12
|
19.19
|
1.60
|
8.57
|
**
|
|
Génotype (G)
|
29
|
34.05
|
1.17
|
1.01
|
ns
|
|
G x L
|
87
|
101.63
|
1.17
|
6.26
|
**
|
|
Régression
|
29
|
10.30
|
0.36
|
0.23
|
ns
|
|
Déviation de la régression
|
58
|
91.33
|
1.57
|
8.44
|
**
|
|
IPCA1
|
31
|
77.25
|
2.49
|
9.01
|
**
|
|
IPCA2
|
29
|
16.91
|
0.58
|
2.11
|
**
|
|
Résiduelle
|
27
|
7.47
|
0.28
|
1.48
|
ns
|
|
Erreur pondérée
|
348
|
64.9
|
0.19
|
----
|
---
|
|
Totale
|
479
|
1735.55
|
----
|
|
---
|
ns,*,**= effets non significatifs et significatifs au seuil de 5
et 1%, respectivement.
Sur les quatre milieux testés, Les moyennes des
rendements varient de 0.72 à 6.55 t/ha, moyennes extrêmes
mesurées, respectivement, chez le génotype
Rascon/Sla_3/3/Plata_1/Snm//Plata_9 (n°8) sur le site de Tiaret et le
génotype Bichena / Ariza_2//Solga_8 (n°3) sur le site du Khroub
(Appendice C, Tableau 5). La moyenne des rendements des localités
98
(effet site) varie de 1.00 à 5.45 t/ha, valeurs
mesurées respectivement à Tiaret et au Khroub (Tableau 6.13).
Lorsque l'interaction génotype x environnement est
significative, le sélectionneur ne peut se baser sur les performances
moyennes inter-sites pour choisir le meilleur génotype. En effet une
interaction génotype x environnement significative indique que chaque
environnement a son propre classement des performances génotypiques et
donc son meilleur génotype. Ceci est d'autant plus vrai que les
environnements expérimentés dans l'évaluation des
génotypes divergent fortement du point de vue capacité
d'expression de potentiel de production.
Cette procédure rend plus aisé le choix d'un
génotype donné dans une série ou groupe d'environnements
qui classent de manière uniforme les génotypes
évalués. Les génotypes Bichena / Ariza_2//Solga_8,
(n°3) Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12), Aaz
77_2/Porron_11//Busca_3 (n°18) et le témoin Waha figurent parmi les
meilleurs génotypes dans au moins deux sites (Tableau 6.13).
Tableau 6.13 : Rendement moyen (t/ha) et numéro des
génotypes performants Par localité.
|
Sites
|
OSM
|
KHR
|
SET
|
TRT
|
|
RDT (t/ha)
|
3.06
|
5.45
|
4.93
|
1.00
|
|
Ppds5%
|
0.9
|
1.13
|
0.45
|
0.34
|
|
Numéro des a
Génotypes Performants
|
12
|
3
|
3
|
16
|
|
7
|
6
|
5
|
30
|
|
13
|
8
|
12
|
20
|
|
29
|
22
|
18
|
|
12
|
18
|
11
|
a = génotypes dont le RDT se situe dans la marge de RDTmax
- 1Ppds5%
L'identification du génotype le plus performant sur des
sites spécifiques est désirable pour les sélectionneurs et
les producteurs. Dans le cas où ces résultats se
répètent dans le temps et l'espace, un gain de rendement de 1.05
t/ha = (4.69-3.64), sera réalisé, si la sélection retient
le génotype Bichena / Ariza_2//Solga_8 (n°3) pour KHR et SET, le
génotype
99
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12) pour OSM
et le génotype Inter_9/Poho_1 (n°16) pour TRT (Tableau 6.13).
D'après FINLY et WINLKINSON, (1963) [11] le
modèle de l'analyse de la variance n'est pas approprié pour
décrire ces changements dans les environnements peu favorables qui
induisent des changements des réponses des génotypes. Il est
souvent fait recours aux modèles linéaires et multiplicatifs pour
caractériser le comportement de chaque génotype lorsque le milieu
varie [13].
6.3.3 L'analyse de la régression
Quoique plusieurs chercheurs (WRIGHT, 1971 [164]; ZOBEL et
al., 1988 [13]; ANNICCHIARICO, 1997 [165]; EBDON et GAUCH, 2002 [10])
montrent que la technique de la régression conjointe ne
différencie pas entre l'effet principal et l'interaction, cette
technique a été utilisé pour analyser les résultats
de la présente étude. La composante linéaire,
testée par rapport à la déviation de la régression,
n'était pas significative, suggérant
l'homogénéité des coefficients de régression. Par
contre, la déviation de la régression était hautement
significative, retenant une part appréciable, non expliquée, de
la somme des carrés des écarts (89.86%) de l'interaction G x L
(Tableau 6.12).
Les résultats du test t des coefficients de
régression montrent que Rascon/Sla_3/3/Plata_1/Snm//Plata_9 (n°8)
et Boussellem/Ofonto//waha (n°27) présentent un coefficient de
régression significativement supérieur à l'unité,
alors que les autres génotypes se caractérisent par des
coefficients de régression non significativement différents de
l'unité (Tableau 6.14).
Les génotypes Minimus_7// Auk /Oste /3/ Shag_26
(n°6), Sula /RBCE_2/3/Hui //CIT 71/Cii/4/Ryps27_3/... (n°7),
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12), D68-1-93A-1A//Ruff /Fg /3/
Mtl-5/4/Lahn (n°13) et Aram-5/Cali//Rascon-37/3/Plata-8 (n°22)
contribuent significativement à l'interaction génotype x milieu.
Alors que les génotypes Minimus_7// Auk /Oste /3/ Shag_26 (n°6),
Sula /RBCE_2/3/Hui //CIT 71/Cii/4/Ryps27_3/... (n°7),
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12), D68-1-93A-1A//Ruff /Fg /3/
Mtl-5/4/Lahn (n°13), Inter_9/Poho_1(n°16),
Aram-5/Cali//Rascon-37/3/Plata-8 (n°22)
100
et MBB/Ofonto//Radiosso/Waha (n°30) présentent un
écart significatif du carré moyen de la régression
(Tableau 6.14).
Les génotypes Minimus/ Rascon_19 (n°1), Silver_26
/ Toska_26 (n°4), Sula /RBCE_2/3/Hui //CIT 71/Cii/4/Ryps27_3/...
(n°7), Sn Turk Mi 83-84 375/Nigris_5// Tantlo_1/3/... (n°9),
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12), D68-1-93A-1A//Ruff /Fg /3/
Mtl-5/4/Lahn (n°13), et D68-1-93A-1A //Ruff/Fg/3/Mtl-5/4/Lahn (n°21)
présentent une ordonnée à l'origine positive, égale
ou supérieure à 0.3 t/ha, suggérant que ces
génotypes minimisent la baisse de rendement sous conditions de
production défavorables (Tableau 6.14).
La représentation linéaire des génotypes
qui contribuent le plus à l'interaction selon la fertilité des
sites montre que le génotype
101
Tableau 6.14 : Moyennes du rendement en grains (RDT, t/ha),
coefficient de régression (b), contribution au carré moyen de
l'interaction (MSGxL), carré moyen de la déviation de
la régression (MSDEV.), ordonnée à l'origine
(a, t/ha) et répétitivité (I) du rendement des
génotypes de blé dur évalués dans 4
localités.
|
Genotype
|
RDT
|
b
|
MSGxL
|
MSDEV
|
a
|
I
|
|
1
|
3.69
|
0.871
|
0.32ns
|
0.38ns
|
0.33
|
0.445
|
|
2
|
3.69
|
0.983
|
0.12ns
|
0.19ns
|
0.11
|
0.321
|
|
3
|
4.38
|
1.183
|
0.16ns
|
0.03ns
|
0.01
|
0.320
|
|
4
|
3.66
|
0.932
|
0.09ns
|
0.11ns
|
0.33
|
0.540
|
|
5
|
3.73
|
1.016
|
0.07ns
|
0.11ns
|
0.03
|
0.270
|
|
6
|
3.61
|
0.984
|
0.64**
|
0.96**
|
0.12
|
0.127
|
|
7
|
3.86
|
0.842
|
1.48**
|
2.06**
|
0.80
|
0.425
|
|
8
|
3.67
|
1.131*
|
0.08ns
|
0.01ns
|
-0.38
|
0.000
|
|
9
|
3.57
|
0.884
|
0.29ns
|
0.35ns
|
0.42
|
0.549
|
|
10
|
3.63
|
0.955
|
0.13ns
|
0.18ns
|
0.25
|
0.457
|
|
11
|
3.71
|
1.015
|
0.01ns
|
0.02ns
|
0.09
|
0.350
|
|
12
|
4.36
|
0.972
|
0.81**
|
1.21**
|
0.83
|
0.772
|
|
13
|
3.84
|
0.936
|
0.80**
|
1.17**
|
0.49
|
0.416
|
|
14
|
3.58
|
1.075
|
0.08ns
|
0.08ns
|
-0.26
|
0.000
|
|
15
|
3.73
|
0.986
|
0.10ns
|
0.15ns
|
0.27
|
0.489
|
|
16
|
3.45
|
1.008
|
0.44ns
|
0.65*
|
-0.22
|
0.000
|
|
17
|
3.24
|
0.979
|
0.23ns
|
0.34ns
|
-0.42
|
0.000
|
|
18
|
3.70
|
0.985
|
0.13ns
|
0.20ns
|
0.05
|
0.264
|
|
19
|
3.56
|
1.068
|
0.04ns
|
0.03ns
|
-0.33
|
0.000
|
|
20
|
3.27
|
0.941
|
0.09ns
|
0.11ns
|
0.07
|
0.293
|
|
21
|
3.51
|
0.875
|
0.10ns
|
0.06ns
|
0.33
|
0.538
|
|
22
|
3.36
|
1.097
|
0.59**
|
0.83**
|
-0.63
|
0.000
|
|
23
|
3.39
|
0.981
|
0.10ns
|
0.15ns
|
-0.18
|
0.038
|
|
24
|
3.61
|
0.956
|
0.02ns
|
0.02ns
|
0.16
|
0.408
|
|
25
|
3.60
|
1.087
|
0.17ns
|
0.21ns
|
-0.36
|
0.000
|
|
26
|
3.24
|
0.968
|
0.24ns
|
0.35ns
|
-0.28
|
0.000
|
|
27
|
3.74
|
1.120*
|
0.06ns
|
0.00ns
|
-0.28
|
0.017
|
|
28
|
3.18
|
0.981
|
0.35ns
|
0.53ns
|
-0.39
|
0.000
|
|
29
|
3.51
|
1.132
|
0.26ns
|
0.28ns
|
-0.59
|
0.000
|
|
30
|
3.35
|
1.056
|
0.45ns
|
0.66*
|
-0.32
|
0.000
|
* pente significativement > 1; ns, *, ** = composantes de
la variance non significative et significative au seuil de 5% et de 1%,
respectivement; = valeur négative de I est représentée
par zéro.
102
Bichena / Ariza_2//Solga_8 (n°3) présente une
adaptation spécifique aux régions représentées par
les sites KHR et SET. Le génotype Gcn/4/D68-1-93A-1A//Ruff / Fg /3/
Mtl-5 (n°12) est à recommander aux régions
représentées par les sites OSM et TRT (Figure 6.10). Un gain de
rendement en grains de 0.67 t/ha, pour l'ensemble des régions couvertes
par les 4 sites, peut être obtenu par l'adoption des génotypes
Bichena / Ariza_2//Solga_8 (n°3) pour KHR et SET, et
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12) pour OSM et TRT (Figure
6.10).
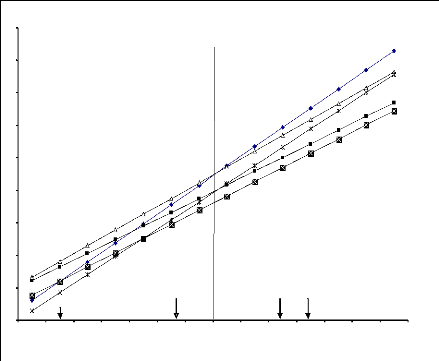
9
8
7
6
5
4
3
2
1
0
0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6 6.5 7
Genotype yield (t/ha)
Gran mean =Y.. = 3.64 t/ha
3
12
27
7
21
12
7
SET KHR
OSM
Yield index (t/ha)
TRT
27
Figure 6.10 : Variation du rendement en grains des
génotypes qui contribuent le plus à l'interaction selon la
fertilité des sites.
103
6.3.4 L'analyse AMMI
Les résultats de l'analyse AMMI montrent la
signification des deux premières composantes de l'interaction (IPCA),
qui sont utilisées pour construire le biplot AMMI2. Les deux composantes
expliquent 92.65% de la somme des carrés des écarts de
l'interaction G x L (76.01% pour le IPCA1 et 16.64% pour le IPCA2). La
résiduelle du modèle AMMI2 n'est statistiquement pas
significative (Tableau 6.12). Comparativement aux 10.13% de l'interaction G x L
non significatifs, indiqués par le modèle de l'analyse de la
régression conjointe, le modèle AMMI apparaît plus
efficace, vu qu'il arrive à expliquer une importante proportion de
l'interaction G x L (Tableau 6.12).
Le biplot AMMI1, est une représentation graphique de
l'effet principal génotype, l'effet localité et de l'interaction
G x L expliquée par la première composante principale IPCA1 du
rendement en grains. Le graphe explique 98.52% de la somme des carrés
des écarts des traitements, qui se répartissent entre 91.78,
somme des carrés des écarts des localités ; 2.06, somme
des carrés des écarts des génotypes et 4.67%, pour la
somme des carrés des écarts de l'interaction G x L (Figure 6.11,
Tableau 6.12). La somme des carrés des écarts de la
première composante IPCA1, qui représente 76.01% de la somme des
carrés des écarts de l'interaction G x L, est plus
élevée en valeur que la somme des carrés des écarts
génotype, suggérant l'importance de tenir compte de l'interaction
dans l'estimation des effets génotypes évalués dans
différentes localités.
Selon GAUCH et ZOBEL, (1996) [166] dans des essais multi site,
on trouve souvent que la variation due à l'interaction est plus
importante que celle due à l'effet génotype. Le score d'un
génotype sur de la première composante IPCA1, sur le plan
représenté par le biplot AMMI1 est un indicateur de la
stabilité des performances de ce génotype dans les
différentes localités. Plus élevée est la valeur,
négative ou positive, du score, plus le génotype en question
montre une adaptation spécifique à un site ou de groupe de sites
donnés. Un score de l'IPCA proche de zéro (près de
l'origine) est indicateur d'une faible interaction et donc d'une
stabilité élevée [13].
104
Les génotypes Bichena/Ariza_2//Solga_8 (n°3),
Rascom/Sla_3/3/Plata_1/ Snm// Plata_9 (n°8), Boussellem/Ofonto//waha
(n°27) et FI 02 (n°24) présentent un faible score sur l'IPCA1;
ils contribuent, donc, relativement moins à l'interaction G x L. Ils
sont considérés comme des variétés à large
adaptation pour l'ensemble des sites d'évaluations. Ces génotypes
diffèrent cependant du point de vu effet principal du rendement en
grains (Figure 6.11).
Les génotypes Bichena / Ariza_2//Solga_8 (n°3) et
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12) comme les génotypes
Sula /RBCE_2/3/Hui //CIT 71/Cii/4/Ryps27_3/... (n°7), D68-1-93A-1A//Ruff
/Fg /3/ Mtl-5/4/Lahn (n°13), Gta/Dur... (n°15) et
Rascon/Sla_3/3/Plata_1/Snm//Plata_9 (n°8) ont le même effet
principal mais diffèrent pour la contribution à l'interaction.
Par contre les génotypes Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5
(n°12), Gta/Dur... (n°15), Mck_2/Aco89 (n°2), Silver_26 /
Toska_26 (n°4), Waha (n°5), Boussellem (n°20), FI 05
(n°23), Kucük (n°17) et Ofonto/Waha//MBB (n°28)
diffèrent pour l'effet principal et l'interaction (Figure 6.11).
Vu leurs scores sur l'IPCA1, toutes les localités se
caractérisent par une interaction G x L élevée. KH et ST
se regroupent dans le quadrant inférieur, montrant un effet principal
élevé et partageant les mêmes meilleurs génotypes
(winners). De ce fait ces sites représentent un même domaine de
recommandations (Figure 6.11). Le site OS s'oppose aux sites KH et ST; il
classe différemment les génotypes comparativement aux autres
sites. OS montre une performance de rendement moyenne alors que le site de TR
présente la plus faible performance de rendement en grains (Figure
6.11).
Une relation de type linéaire apparaît entre les
scores sur l'IPCA1 et l'effet moyen du biplot AMMI1, suggérant que les
génotypes qui se caractérisent par des effets principaux
extrêmes : Les génotypes Sula /RBCE_2/3/Hui //CIT
71/Cii/4/Ryps27_3/... (n°7), Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5
(n°12) et D68-1-93A-1A//Ruff /Fg /3/ Mtl-5/4/Lahn (n°13) avec
Aram-5/Cali//Rascon-37/3/Plata-8 (n°22), MBB/Ofonto//Radiosso/Waha
(n°30), et Inter_9/Poho_1 (n°16) présentent des scores de
l'interaction G x L opposés. Le premier groupe présente
Au contraire, les génotypes Waha (n°5), Srn_3 /
Nigris_4 // Porto_5 (n°11), Plata_1/Snm // Plata_9/3/Green (n°14),
Inter_9/Poho_1 (n° 16), Kucük (n°17),
105
des scores positifs et le second groupe présente des
scores négatifs et par conséquence ces deux groupes de
génotypes sont adaptés à différents milieux.
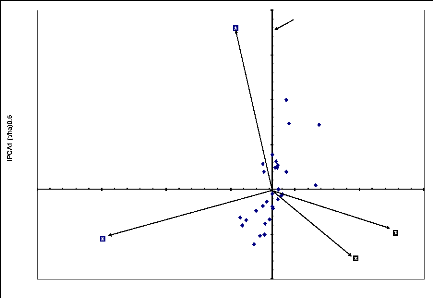
-0.500
-1.000
2.000
0.500
0.000
1.500
1.000
0.00 1.00 2.00 3.00 Grain yield (t/ha) 4.00 5.00
6.00
TR
OS
28
17
23 20
24
26
22
16
21
1
29 25
30
9
14
5
7
8
15
13 12
27
Y.. =3.64
3
ST
KH
Figure 6.11 : Biplot AMMI1 des effets principaux
génotypes et sites et de l'interaction de la première composante
IPCA1 du rendement en grains.
Analysant ensemble les scores de l'interaction et les effets
principaux,
les génotypes : Minimus/ Rascon_19 (n°1),
Mck_2/Aco89 (n°2),
Bichena / Ariza_2//Solga_8 (n°3), Silver_26 /
Toska_26 (n°4), Minimus_7// Auk /Oste /3/ Shag_26 (n°6), Sula
/RBCE_2/3/Hui //CIT 71/Cii/4/Ryps27_3/... (n°7),
Rascon/Sla_3/3/Plata_1/Snm//Plata_9 (n°8), Sn Turk Mi 83-84 375/Nigris_5//
Tantl_1/3/... (n°9), Hoggar (n°10), Gcn/4/D68-1-93A-1A//Ruff / Fg /3/
Mtl-5 (n° 12), D68-1-93A-1A//Ruff /Fg /3/ Mtl-5/4/Lahn (n°13),
Gta/Dur... (n°15), et D68-1-93A-1A //Ruff/Fg/3/Mtl-5/4/Lahn (n°21)
ont des scores IPCA1 positifs comme celui du site OS. Ces génotypes
montrent donc une adaptation spécifique à ce site.
106
Aaz 77_2/Porron_11//Busca_3 (n°18), Altar84 (n°19),
Boussallem (n°20), Aram-5/Cali//Rascon-37/3/Plata-8 (n°22), FI 05
(n°23), FI 02 (n°24), Cirta (n°25), Ting7/2*Green-3 (n°26),
Boussellem/Ofonto//waha (n°27), Ofonto/Waha//MBB (n°28), Tr
32225/Gediz//Ofonto (n°29) et MBB/Ofonto//Radiosso/Waha (n°30)
montrent une adaptation aux sites KH, ST et TR qui se distinguent par des
scores IPCA1 négatifs (Figure 6.11).
Le génotype Bichena / Ariza_2//Solga_8 (n°3) se
distingue par un haut rendement grain et une bonne stabilité, il
présente une large adaptation à trous les sites
d'évaluation. Par contre les génotypes Gcn/4/D68-1-93A-1A//Ruff /
Fg /3/ Mtl-5 (n°12), D68-1-93A-1A//Ruff /Fg /3/ Mtl-5/4/Lahn (n°13)
et Sula/Rbce2/3/Hui//Cit71/CII/4/ Ryps27_3/... (n°7) montrent une
adaptation spécifique au site OS (Figure 6.11). Aucun génotype
à haut rendement ne montre une adaptation spécifique aux sites
KH, ST et TR (Figure 6.11).
Ces résultats indiquent que le sélectionneur a
le choix entre la sélection du génotype Bichena /
Ariza_2//Solga_8 (n°3) qui montre une adaptation générale en
plus de son potentiel de rendement élevé, pour tous les sites ou
bien recommander ce génotype aux régions
représentées par les sites KH, ST et TR et de sélectionner
le génotype Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12)
spécialement pour la région représentée par le site
OS.
Le gain de rendement attendu est estimé selon la
méthode décrite par ZOBEL et al., (1988) [13] à
partir de l'effet principal génotype + l'effet principal site - la
moyenne générale de l'ensemble des sites + le produit des scores
IPCA1 du génotype x celui du site. L'adoption du génotype Bichena
/ Ariza_2//Solga_8 (n°3) pour toutes les localités donne un gain de
rendement de 0.74 t/ha = (4.38 -3.64), par contre le gain de rendement produit
par l'alternative est égale à 1.0 t/ha = (4.64-3.64).
La stratégie associant l'adaptation
générale à une large région
représentée par les trois sites KH, ST et TR avec l'adaptation
spécifique pour OS, induit un gain de rendement de 5.94% =
(4.64-4.38/4.38), et cela sur la base de l'information fournie par le biplot du
modèle AMMI1 (Figure 6.11).
107
L'étude du biplot du modèle AMMI2, construit en
utilisant les scores des génotypes et des sites
généré par les deux premières composantes, indique
que les sites OS et TR ont des scores opposés pour l'IPCA1, alors que
les sites KH et ST ont des scores opposés pour l'IPCA2. Les sites OS, KH
et ST sont une source d'interaction plus forte que celle du site de TR, comme
indiqué par les valeurs prises par leurs scores (Figure 6.12).
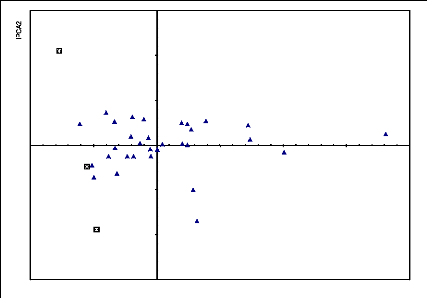
-1.000 -0.500 0.000 0.500 1.000 1.500 IPCA1 2.000
8
13
4
7
1
6
9
10
12
3
15 2
21
OS
ST
18
28
5
26
22
25
19
11
14
27
16
24
29
TR
30 17
23 20
KH
1.500
1.000
0.500
0.000
-0.500
-1.000
-1.500
Figure 6.12 : Biplot AMMI2 de l'interaction du rendement en
grains des 30 génotypes évalués dans 4
localités.
Les génotypes (Bichena / Ariza_2//Solga_8),
(Rascon/Sla_3/3/Plata_ 1/Snm//Plata_9), (Srn_3 / Nigris_4 // Porto_5),
(Plata_1/Snm // Plata_9/3/Green), (Altar84), (D68-1-93A-1A
//Ruff/Fg/3/Mtl-5/4/Lahn), (FI 02) et (Boussellem/Ofonto// waha) qui
représentent respectivement les numéros: 3, 8, 11, 14, 19, 21, 24
et 27, positionnés prés de l'origine, contribuent peu à
l'interaction G × L. Par contre les génotypes
(Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5), (D68-1-93A-1A//Ruff /Fg /3/
Mtl-5/4/Lahn), (Minimus_7// Auk /Oste /3/ Shag_26), (MBB/Ofonto//Radiosso/
Waha), (Inter_9/Poho_1) et (Aram-5/Cali//Rascon-37/3/Plata-8) qui sont
représentés respectivement par les numéros: 12, 13, 6, 30,
16, et 22 contribuent
108
fortement à l'interaction (Figure 6.12). Les sites KH
et TR présentent le même type d'interaction, alors que les sites
de ST et OS présentent des types différents, vu qu'ils se
positionnent dans différents quadrants du biplot (Figure 6.12).
Le génotype Sula /RBCE_2/3/Hui //CIT
71/Cii/4/Ryps27_3/... (n°7) montre une adaptation spécifique au
site OS. Le génotype MBB/Ofonto//Radiosso/Waha (n°30) est
spécifiquement adapté au site du KH. Le génotype
Inter_9/Poho_1 (n°16) au site TR et le génotype
Aram-5/Cali//Rascon-37/3/Plata-8 (n°22) montre une adaptation
spécifique au site ST (Figure 6.12).
Sur la base des informations fournies par le biplot AMMI2,
l'adoption des génotypes à adaptation spécifique permet
d'exploiter l'interaction G x L et génère, par conséquent,
un gain de rendement égale à 0.64 t/ha = (4.28-3.64).
6.4. L'adoption des génotypes sur la base du
rendement nominal
Le calcul du rendement nominal est utile pour
déterminer l'adaptabilité à des régions
spécifiques identifiées par leurs scores sur la première
composante de l'interaction (LIPCA1). La figure (6.13) indique que le
génotype Bichena / Ariza_2//Solga_8 (n°3) présente le
rendement nominal le plus élevé pour des sites dont le score de
la première composante LIPCA1 est inférieur à -0.700. Le
site de ST se trouve dans cette plage de scores LIPCA1. De ce fait, et sur la
base du rendement nominal, ce génotype Bichena / Ariza_2//Solga_8
(n°3) est spécifiquement adapté à la région
représentée par le site de Sétif.
Le génotype Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5
(n°12) présente un rendement nominal élevé pour des
sites dont le score LIPCA1 est supérieur à -0.700. Les sites TR,
KH et OS se situent dans cette plage des valeurs des scores LIPCA1. De ce fait
ce génotype est à recommander aux régions
représentées par ces trois sites, où il montre une
adaptation spécifique, donc un haut rendement suite à une
interaction positive (Figure 6.13).
109
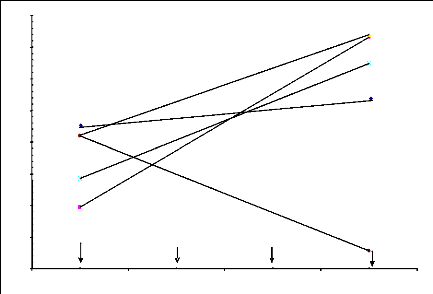
6.00
5.50
5.00
4.50
4.00
3.50
3.00
2.50
2.00
-0.771 -0.554 -0.481 L IPCA1 scores 1.806
Nominal yield (t/ha)
12
7
13
3
TR KH
22
ST
OS
Figure 6.13 : Rendement nominal des meilleurs génotypes
en fonction des scores LIPCA1 des 4 sites expérimentaux.
La variation du rendement nominal entre les différents
sites peut être utilisée comme un indicateur de la
stabilité génotypique. Le génotype Bichena /
Ariza_2//Solga_8 (n°3) montre une stabilité de type biologique vu
que son rendement nominal varie de 4.18, rendement nominal du site ST, à
4.55 t/ha à OS (Figure 6.13). Le rendement nominal du génotype
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12) varie, quant à lui
de 4.10 à 5.70 t/ha. Comparativement le génotype Sula
/RBCE_2/3/Hui //CIT 71/Cii/4/Ryps27_3/... (n°7) qui se classe en
tête pour le rendement nominal sur le site OS présente une faible
stabilité (instable), vu que son rendement nominal varie largement de
2.97 à 5.66 t/ha (Figure 6.13).
Suite à leur haut rendement nominal relativement
stable, les génotypes Bichena / Ariza_2//Solga_8 (n°3) et
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12) ressortent comme les plus
performants parmi les 30 génotypes évalués et de ce fait
sont à recommander sur l'ensemble des régions
représentées par
110
les 4 sites d'évaluation. Ces génotypes peuvent
être aussi utilisés comme témoins pour tester l'adaptation
générale et l'adaptation spécifique des futures
lignées avancées entrant les essais multi- locaux, tel que
mentionné par SAMONTE et al., (2005) [167].
6.5 Répétitivité
(Reliability)
Vu que le niveau technologique de la
céréaliculture algérienne est faible, faisant de cette
activité, une activité de subsistance, le génotype le
moins risqué est le génotype Gcn/4/D68-1-93A-1A//Ruff / Fg /3/
Mtl-5 (n°12), qui présente une valeur de l'indice I de
répétitivité égale à 0.772 t/ha. Ce
génotype est recommandé, sur la base de son rendement nominal,
aux régions représentées par les sites OS, KH et TR. Le
génotype Bichena / Ariza_2//Solga_8 (n°3), recommandé pour
la région représentée par le site ST, se classe
13ième pour la valeur de l'indice I (Tableau 6.14). Les
valeurs de l'indice I sont significativement corrélées avec
l'effet principal génotype (rs = Spearman coefficient de
corrélation = 0.581**), et avec les scores IPCA1 des
génotypes (rs= 0.858**).
111
6.6 Appréciation de la qualité
technologique par l'étude de certains paramètres
physico-chimiques
6.6.1 Taux de mitadinage et taux
d'extraction
6.6.1.1 Taux de mitadinage
La structure vitreuse du grain de blé dur est un
paramètre déterminant pour sa valeur d'utilisation. L'existence
des parties farineuses dans l'albumen des grains mitadinés, affecte la
valeur semoulière en diminuant le rendement en semoule [168].
L'analyse de la variance montre un effet génotype,
milieu et une interaction génotypes x milieu très hautement
significatifs pour les deux variables taux de mitadinage et taux d'extraction
(Tableau 6.15).
Tableau 6.15 : Carrés moyens des écarts de
l'analyse de la variance du taux de mitadinage (MTD) et du taux d'extraction
(EXT) mesurés sur les quatre sites.
|
Source
|
ddl
|
MTD (%)
|
EXT (%)
|
|
Totale
|
479
|
196.98
|
54.46
|
|
Génotype (G)
|
29
|
230.17***
|
105.64***
|
|
Environnement (E)
|
3
|
22209.54***
|
3535.32***
|
|
(G) × (E)
|
87
|
238.36***
|
137.19***
|
|
Résiduelle
|
360
|
0.47
|
1.64
|
ns,*,*** = effets non significatifs et significatifs au seuil de
5 et 1%, respectivement.
Un effet génotypique très hautement significatif
indique que les différences entre les génotypes
évalués sont donc plus importantes, le génotype
Boussellem/Ofonto//waha (n°27) se distingue par le taux de mitadinage le
plus élevé (15.29%) que celui des autres variétés,
alors que le génotype Altar84 (n°19) donne le taux le plus faible
(3.5%), soit une amplitude de 11.79% (Tableau 6.16).
La variété est un élément de
gestion du risque de mitadinage, mais l'influence du climat et de l'azote
étant très forte aussi [170].
112
Le génotype Bichena / Ariza_2//Solga_8 (n°3) et le
génotype Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12) qui ont
été sélectionnés comme des génotypes
à large adaptation donnent des taux de mitadinage inférieurs
à la moyenne générale avec respectivement 5.03 et 7.2 %
(Tableau 6.16).
Les taux de mitadinage élevés influencent sur la
qualité des semoules, en effet, d'après ANNOU, (2002) [169] Les
piqûres blanches issues de blé mitadinés
déprécient la couleur des pâtes alimentaires car elles
s'hydratent difficilement quant elles sont suffisamment volumineuses et
résistent à la désagrégation lors du
pétrissage.
Tableau 6.16 : Valeurs moyennes caractéristiques du
taux de mitadinage (MTD) et du taux d'extraction (EXT) notées sur les
différents sites.
|
Les Valeurs
|
MTD (%)
|
EXT (%)
|
|
Effet moyen génotype (G)
|
|
Max
|
15.29
|
58.72
|
|
Min
|
3.5
|
47.10
|
|
Amplitude
|
11.79
|
11.62
|
|
Le génotype n° 3
|
5.03
|
58.72
|
|
Le génotype n° 12
|
7.2
|
50.50
|
|
Effet moyen Environnement (E)
|
|
E1 Oued Smar
|
2.92
|
48.71
|
|
E2 El Khroub
|
4.34
|
55.75
|
|
E3 Sétif
|
29.42
|
61.69
|
|
E4 Tiaret
|
0.08
|
53.08
|
|
Interaction (G) × (E)
|
|
Max
|
56.13
|
66.23
|
|
Min
|
0
|
31.68
|
|
Amplitude
|
56.13
|
34.55
|
|
|
La moyenne générale
|
9.19
|
54.80
|
|
C V (%)
|
7.5
|
2.1
|
|
E.T
|
0.69
|
1.16
|
Nous constatons, donc, que tous les génotypes
évalués font apparaître une augmentation significative des
grains mitadinés dans le milieu de Sétif par rapport
113
En effet, sous la dépendance du facteur milieu, sur le
site de Sétif, le taux de mitadinage atteint 29.42%, sur le site du
Khroub et Oued Smar, le taux de mitadinage apparaît faible à
très faible avec 4.34 et 2.92% respectivement, alors que le milieu de
Tiaret caractérisé par une absence totale des grains
mitadinés sur la majorité des génotypes
évalués, le taux de mitadinage est presque nul (0.08%) (Tableau
6.16). Ceci peut être expliqué par la richesse du sol en azote
minéral, étant donné que le précédent
cultural était une légumineuse alimentaire.
Selon BRAUN, (1994) [171] Le mitadinage est fortement
limité par fractionnement des apports d'azote, un apport au stade
gonflement de la culture est mieux recommandé que les apports tardifs.
Ceci est lié au coefficient apparent d'utilisation de l'azote qui est
plus élevé à ce stade.
La qualité d'un blé dur est essentiellement
régie par les effets conjugués du génotype et des facteurs
agro-climatiques. La variété est un facteur important dans la
détermination de la qualité, toutefois, l'expression du potentiel
génétique est étroitement liée au mode de conduite
de la culture. Le blé exprime son plus haut potentiel de quantité
et de qualité seulement sous une optimisation des facteurs de production
[168]. En effet, les valeurs du taux de mitadinage pour l'interaction
génotype X environnement sont comprises entre zéro pourcent (pour
l'ensemble des génotypes à Tiaret) et à 56.13% pour le
génotype Boussellem/Ofonto//waha (n°27) à Sétif
(Tableau 6.16).
L'effet de l'interaction entre les deux facteurs sur le taux
de mitadinage est marquant, Les génotypes évalués
acquièrent une vitrosité d'autant plus grande que le milieu
devient plus favorable. Chez le génotype Boussellem/Ofonto//waha
(n°27) qui s'exprime par la valeur la plus élevée du taux de
mitadinage à Sétif (56.13%), par contre à Tiaret son taux
de mitadinage est nul. Sur les deux sites Khroub et Oued Smar, ses valeurs sont
faible à très faible (3.38 et 1.67% respectivement). Par
conséquent, dans l'ensemble, le taux de mitadinage varie beaucoup d'un
site à l'autre pour le même génotype (Appendice C, Tableau
7).
De même, les résultats de l'interaction
génotype x environnement montrent que le taux d'extraction chez le
génotype Mck_2/Aco89 (n°2) à Sétif est le plus
114
aux sites d'El Khroub et Oued Smar, tandis qu'à Tiaret
le taux de mitadinage ne dépasse pas 0.5% (Appendice C, Tableau 7 ).
De même, nous pouvons considérer que le
génotype à plus faible taux de mitadinage, dans le milieu de
Sétif, est celui qui est le plus résistant au mitadinage, dans ce
cas c'est le génotype Altar84 (n°19) avec 10.13%.
Sachant que le mitadinage est directement lié à
la teneur en protéines, donc à la nutrition azotée de la
culture, si au cours du remplissage du grain la matière protéique
se trouve en quantité suffisante, l'albumen prendra un aspect vitreux.
En revanche, la carence protéique conduit à la formation de
nombreuses vacuoles d'air au sein de l'albumen, lui conférant une
apparence opaque ou farineuse [172].
6.6.1.2 Taux d'extraction
La comparaison des moyennes, pour l'effet génotype,
indique que la valeur maximale est obtenue chez le génotype Bichena /
Ariza_2//Solga_8 (n°3) avec un taux d'extraction de 58.72%, alors que le
génotype Altar84 (n°19) présente la valeur minimale (47.10%)
soit une amplitude de 11.62%. Le génotype Gcn/4/D68-1-93A-1A//Ruff / Fg
/3/ Mtl-5 (n°12) donne un taux d'extraction proche à la moyenne
générale de l'essai (54.80%) (Tableau 6.16).
Il faut souligner que plus des 2/3 des génotypes ont
des taux d'extraction supérieur à 55% (Appendice C ; Tableau 6).
Pour la réalisation des testes technologiques tels que
l'alvéographe Chopin, BOURDET (1976) [172] préconise un taux
d'extraction de 60 à 70%. En se référant au (Tableau
6.16), nous constatons que les moyennes des taux d'extraction oscillent entre
61.69% pour le site de Sétif et 48.71% pour le site d'Oued Smar, les
valeurs du Khroub et de Tiaret sont respectivement de 55.75 et 53.08%.
115
élevé avec 66.23% (Tableau 6.16). Ce
génotype donne des taux d'extraction respectivement de 56.81, 52.53 et
46.41% à El Khroub, Tiaret et Oued Smar.
Le taux d'extraction le plus faible a été obtenu
par le génotype MBB/Ofonto//Radiosso/Waha (n°30) qui a une valeur
de 31.68% enregistré à Oued Smar, alors qu'il a des taux
d'extraction de 61.74% à Sétif, de 56.08% à El Khroub et
de 52.33% à Tiaret (Appendice C, Tableau 8). D'après ces
résultats, nous remarquons que l'amplitude est de 34.55% qui reste
supérieur à la valeur minimale. Ceux peut s'expliquer par la
forte variabilité inter sites entre génotypes et pour le
même génotype et abouti à une forte interaction
génotype × environnement.
DEXTER et MATSUO, (1978) [174] ont montré que la teneur
en cendre des semoules augmentent quand le taux d'extraction passe de 57.7%
à 75.9%. Dans ce cas, nous pouvons constater que les génotypes
qui ont des taux d'extractions les plus élevés sont ceux qui
donnent des teneurs en matière minérale élevées.
Parmi eux, les génotypes Bichena / Ariza_2//Solga_8 (n°3), Sn Turk
Mi 83-84 375/Nigris_5// Tantl_1/3/... (n°9) et les deux témoins
Cirta et Waha et ce grâce à leurs moyennes obtenues sur les quatre
sites (Appendice C, Tableau 6). L'augmentation du taux de cendre en fonction du
taux d'extraction est due à la contamination de la semoule par les
couches périphériques des grains qui sont beaucoup plus
minéralisées que l'albumen amylacé [174], [175].
6.6.2 Teneur en gluten humide et en gluten sec
Le gluten est un complexe protéique constitue
essentiellement de gluadines et de gluténines, il confère
à la pâte ses propriétés plastiques [176].
L'analyse de la variance des deux paramètres
mesurés ( teneur en gluten humide et celui en gluten sec) sur l'ensemble
des génotypes évalués sur les trois sites (Oued Smar, El
Khroub et Sétif) indique des effets génotype, milieu et
interaction génotype X milieu très hautement significatifs
(Tableau 6.17).
116
Tableau 6.17 : Carrés moyens des écarts de
l'analyse de la variance du taux de gluten humide (GH) et du taux de gluten sec
(GS) mesurés sur les trois sites (Oued Smar, Khroub et Sétif).
|
Source
|
ddl
|
GH (%)
|
GS (%)
|
|
Totale
|
459
|
81.22
|
9.77
|
|
Génotype (G)
|
29
|
150.30***
|
19.54***
|
|
Environnement (E)
|
2
|
8219.08***
|
684.03***
|
|
(G) × (E)
|
58
|
134.93***
|
22.62***
|
|
Résiduelle
|
270
|
1.98
|
0.96
|
ns, * et *** = effet non significatif et significatif au seuil de
5 et 1% respectivement.
Le génotype Plata_1/Snm // Plata_9/3/Green (n°14)
présente les teneurs en gluten humide et en gluten sec les plus
élevées avec respectivement 34.13 et 12.74%, alors que le
témoin Hoggar donne les valeurs les plus faibles (20.81% en gluten
humide et 5.93% en gluten sec) avec une amplitude de 13.32% pour le gluten
humide et une amplitude de 6.54% pour le gluten sec (Tableau 6.18). Celui peut
s'expliquer par la variation significative du gluten humide et sec et ce selon
la variété.
Les génotypes Bichena / Ariza_2//Solga_8 (n°3) et
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12) présentent des
teneurs en gluten humide et en gluten sec proches de la moyenne
générale de l'essai de chaque paramètre qui est
respectivement de 29% et 8.76% (Tableau 6.18).
Les propriétés fonctionnelles
(élasticité, extensibilité et viscosité) de la
plupart des produits de la filière blé, sont largement
déterminées par le comportement viscoélastique des
protéines du blé [177]. Selon DJEMA, (2000) [178] les teneurs en
gluten sec et gluten humide sont généralement plus
élevées chez les semoules obtenues aux taux d'extraction de 70%
que celles obtenues aux taux d'extraction de 60 à 63%.
117
Tableau 6.18 : Valeurs moyennes caractéristiques de la
teneur en gluten humide (GH) et de la teneur en gluten sec (GS) notées
sur les différents sites.
|
Les Valeurs
|
GH (%)
|
GS (%)
|
|
Effet moyen génotype (G)
|
|
Max
|
34.13
|
12.47
|
|
Min
|
20.81
|
5.93
|
|
Amplitude
|
13.32
|
6.54
|
|
Le génotype n° 3
|
29.82
|
9.23
|
|
Le génotype n° 12
|
29.10
|
9.07
|
|
Effet moyen Environnement (E)
|
|
E1 Oued Smar
|
36.18
|
10.65
|
|
E2 El Khroub
|
30.88
|
9.57
|
|
E3 Sétif
|
19.95
|
6.08
|
|
Interaction (G) X (E)
|
|
Max
|
44.06
|
19.47
|
|
Min
|
10.62
|
2.89
|
|
Amplitude
|
33.44
|
16.58
|
|
|
La moyenne générale
|
29.00
|
8.76
|
|
C V (%)
|
4.90
|
11.20
|
|
E.T
|
1.41
|
0.98
|
Pour l'effet milieu et d'après nos résultats
(Tableau 6.18), nous constatons que le classement des sites concernant les deux
paramètres reste le même, en effet, le site d'Oued Smar marque les
valeurs les plus élevées en gluten humide et en gluten sec avec
respectivement 36.18 et 10.65% et le site de Sétif indique les valeurs
les plus basses de 19.95% pour la teneur en gluten humide et de 6.08% pour la
teneur en gluten sec. Cependant, .au Khroub, la teneur en gluten humide
présente une valeur intermédiaire (30.88%) et celle du gluten sec
marque une valeur proche de celle enregistrée à Oued Smar
(9.57%).
Etant sous la dépendance du facteur milieu, le site
d'Oued Smar est plus favorable à l'expression des deux teneurs en gluten
sec et humide que le site de Sétif qui est montré comme
étant plus défavorable. Ceci peut s'expliquer par les taux
élevés de mitadinage sur ce site. CORBELLINI et al.,
(1998) [179] et LUO et al., (2000) [180] ont noté que les
propriétés rhéologiques du gluten sont affectées de
même que la quantité et la qualité des protéines,
par le niveau de fertilité du sol notamment les disponibilités de
l'azote et de soufre.
118
D'autre part, l'interaction entre les deux facteurs
génotype et milieu, pour le gluten humide, montre que la valeur maximale
est obtenue chez le génotype Aaz 77_2/Porron_11//Busca_3 (n°18)
à Oued Smar (44.06%). Ce génotype donne une teneur de 30.05
à El Khroub et de 27.81% à sétif. Alors que la valeur
minimale est marquée chez le génotype Sn Turk Mi 83-84
375/Nigris_5// Tantl_1/3/... (n°9) à Sétif soit de 10.62%.
Ce génotype présente une teneur de 35.9% à Oued smar et
31.26% au Khroub. Ce dernier donne la valeur maximale en gluten sec (19.74%) au
Khroub ainsi que la valeur minimale (2.89%) à Sétif (Appendice C,
Tableau 9 et 10).
L'examen des valeurs maximales et minimales indiquent que
l'amplitude, pour le gluten humide, est de 33.44% et 16.58% c'est l'amplitude
concernant le gluten sec, représenté par des valeurs
supérieures aux moyennes générales des deux
paramètres.
En se basant sur l'échelle de classement proposé
par MATVEEF, (1966) [181] concernant le gluten sec, on peut noter que, si le
gluten sec est inférieur à 11% c'est un blé insuffisant et
si le gluten sec est compris entre 11% et 15% c'est un blé de bonne
valeur pastière et s'il est supérieur à 15%, il est
considéré comme un blé de force. Il est constaté,
donc, que le génotype Sn Turk Mi 83-84 375/Nigris_5// Tantl_1/3/...
(n°9) au Khroub possède une qualité pastière
supérieure (c'est un blé de force). Le génotype
Plata_1/Snm // Plata_9/3/Green (n°14) présente des valeurs plus
proches d'un environnement à l'autre pour le gluten sec qui sont
respectivement de 13.76, 12.57 et 11.09% dans les sites de Sétif, Oued
Smar et Khroub .Ce génotype peut donner un blé de bonne
qualité que les autres génotypes testés (Appendice C,
Tableau 10).
Nos résultats montrent également qu'à
Sétif et El Khroub, tous les génotypes se distinguent par leurs
teneurs en gluten sec inférieures à 11% à l'exception des
génotypes Plata_1/Snm // Plata_9/3/Green (n°14) et Altar84
(n°19) (Appendice C, Tableau 10). Ceux-ci peuvent être
expliqués par les taux élevés de mitadinage suite à
un manque de fertilisation azotée durant les stades critiques ou
à un lessivage d'azote suite à une forte pluie. Ces
résultats pourraient influencer la teneur en protéines. HOULIARO
et al., (1981) [182] montrent que la teneur en protéines est
corrélée significativement et positivement au gluten sec.
119
6.7 Discussion générale
De nombreux chercheurs de la filière blé
réalisent une évaluation variétale, pour différents
objectifs :
? Au niveau de la sélection, pour trier les
génotypes qui seront gardés à l'étape suivante du
processus de sélection;
? Au niveau de la multiplication, pour identifier, parmi les
variétés inscrites au catalogue, celles qui seront
intéressantes à développer;
? Au niveau de la production pour choisir les
variétés qui seront mises au culture dans chaque parcelle de
l'exploitation;
? Au niveau de l'utilisation pour repérer, à
travers la variété, les lots de blé qui, à priori,
répondront le mieux aux exigences du marché.
Cette évaluation variétale s'appuie sur
l'expérimentation pour acquérir une meilleure connaissance des
variétés, donc sur des essais variétaux (regroupant
plusieurs génotypes ou variétés), multi locaux,
généralement pluriannuels. En effet, l'analyse du comportement
des génotypes en fonction des caractéristiques du milieu est
depuis longtemps un sujet important.
De nombreux traits comme le rendement et les caractères
technologiques, sont variables d'un environnement à un autre pour un
génotype donné. Ces variations sont la résultante de trois
facteurs : le génotype, l'environnement au sens large, et l'interaction
génotype x environnement. Dans notre cas, l'étude du comportement
génotypique à travers les quatre milieux d'étude souligne
la diversité de comportement intra et inter sites chez les trente
génotypes évalués.
Ces génotypes manifestent différentes
réponses, traduisant les variations du niveau d'expression des
paramètres mesurés et de l'ordre de classement des
génotypes selon les environnements. En se basant sur les
caractéristiques des milieux expérimentaux qui permettent de
noter les conditions propices au développement et à la
constitution du rendement d'une part et la sensibilité
génotypique au changement de site d'autre part.
120
Les résultats montrent que les sites du Khroub et
Sétif sont les plus favorables à l'expression de haut rendement
en grains grâce aux potentialités naturelles offertes par le
milieu physique pour cette année d'étude, le
phénomène de compensation entre variables et la maîtrise
des facteurs agro-techniques qui jouent un rôle important dans
l'amélioration de la production et celle qui est la cause principale de
la faiblesse du rendement en zone sub-humide. Notamment les adventices et les
maladies fongiques, la rouille brune (Puccinia Triticina) en
particulier. D'après HAMADACHE et al., (2002) [58],
l'amélioration et la stabilité de la productivité du
blé dur au niveau de la zone sub-humide de l'Algérie du nord,
passe par l'introduction des légumineuses annuelles dans l'assolement,
la maîtrise des adventices annuelles et une protection
intégrée et efficace de la culture contre les maladies foliaires
(Tache auréolée, et septoriose surtout ainsi que la rouille
brune).
Contrairement au site semi aride de l'ouest où les
conditions de croissance ont été nettement défavorables,
notamment les basses températures pendant la phase
méiose-épiaison, affectent la fertilité de l'épi,
les conditions d'alimentation hydrique limitantes en Mai ainsi que la
tardivité du semis qui induit une maturité plus tardive exposant
certains génotypes aux effets de stress de fin de cycle.
Dans de telles conditions environnementales, la réponse
de la céréale risque d'être variable selon les conditions
de croissance de la plante, le choix de l'espèce et de la
variété n'est pas suffisant à lui seul pour
déterminer la production de blé dur, il faut aussi tenir compte
du cumul pluviométrique et de la demande climatique qui accompagnent le
choix variétal en question. C'est ainsi qu'une année pluvieuse ou
sèche mais douce n'autorise pas le même rendement qu'une
année pluvieuse ou sèche mais froide [183].
De même, la connaissance des changements qui ont
accompagné l'amélioration de la productivité est
essentielle pour pouvoir faire plus de progrès. Elle permet de
comprendre les facteurs limitants l'obtention de hauts rendements en grains et
de développer des stratégies appropriées. Dans ce
contexte, Nous nous sommes intéressés, en premier lieu, aux
variations du rendement associées aux variations des principaux
caractères identifiés comme ayant une influence
121
non négligeable sur cette variable. D'après
celui-ci nous pouvons constater l'avantage relatif des génotypes
précoces évalués au site sub-humide, cette
précocité a contribué à l'augmentation du rendement
et seul le PMG s'est manifesté de manière positive sur ce
dernier. Ceci suggère que la sélection précoce sur le PMG
engendre celle du rendement en grains. La stabilité du rendement est
donc dépendante, en partie, de celle du PMG. D'autre part, le nombre
d'épi et le nombre de grains par épi sont peu utiles dans la
prévision du rendement en grains.
Dans les plaines intérieures, le rendement est
tributaire du nombre d'épis par unité de surface, et ce dernier
dépend du nombre de talles par m2. Il ressort, donc, que la
sélection des épis produits par unité de surface est
capable d'identifier des génotypes performants. Pour les hauts plateaux
de l'est et de l'ouest, le caractère qui contribue à faire les
différences de rendement entre les génotypes est le nombre de
grains par épi. En effet, la faible valeur de ce caractère est
indicatrice de faible rendement.
Ces changements de liaisons des variables avec le rendement,
d'un lieu test à un autre, indiquent que ces dernières sont
irrégulières dans leur relation les uns avec les autres, à
cause des phénomènes de compensation et de sensibilité aux
changements de milieux. Le peu de liaison entre caractères, intra et
inter sites, et la variation de ces liaisons selon le milieu ont pour effet de
compliquer la sélection indirecte sur la base des variables autres que
le rendement en grains.
C'est pour cette raison, que nous avons,dans un
deuxième temps, envisagés les différentes méthodes
d'étude de l'interaction génotype x environnement
du rendement en grains qui est l'objectif principal de la
sélection.
L'identification des génotypes performants indique
que les changements dans les conditions environnementales entraînent des
fluctuations du rendement pour chaque génotype.
La sélection faite sur la base des plus faibles somme
de classement qui sont indicatrices de l'adaptabilité de leurs
génotypes aux différents environnements. De même, nous
pouvons choisir l'adaptation spécifique sur la même base
122
(classement) où le gain de rendement est
supérieur à celui obtenu par l'adaptation générale,
comme proposé par MALHOTRA et al., (1991) [184] qui trouve que
la classification améliore la prévision des performances
génotypiques à l'intérieure des différents groupes
d'environnements, et conduit à une sélection pour l'adaptation
spécifique. Cependant, l'objectif de sélectionneur est, en effet,
de développer des génotypes stables qui répondent
favorablement à la variation des environnements.
L'analyse de la variance combinée, intégrant
tous les sites, indique que la culture de blé dur est doublement
affectée, par l'effet milieu et l'effet interaction génotype x
milieu. En effet, l'existence de l'interaction génotype x milieu
significative rend difficile la recommandation, d'un génotype
particulier dans un environnement donné, vu que chaque site à ses
meilleurs génotypes particuliers, et qu'il est rare de trouver un
génotype qui reste supérieur sur l'ensemble des lieux
testés. Dans de telles conditions, l'analyse de la variation des
rendements est approchée, par la technique de la régression
conjointe, qui permet de décrire les réponses génotypiques
selon la variation des environnements testés.
La variation des coefficients de régression obtenus,
met en évidence la sensibilité du rendement aux effets de
l'interaction génotype x environnement. Des valeurs de coefficients de
régression significativement supérieures à l'unité,
caractérisent deux génotypes qui répondent favorablement
à l'amélioration de l'environnement. Des valeurs de
l'ordonnée à l'origine positive, égale ou
supérieure à 0.3 t/ha caractérisent les génotypes
qui minimisent la baisse de rendement sous conditions de production
défavorables.
Les génotypes sélectionnés, sur la base
de la représentation linéaire des génotypes qui
contribuent plus à l'interaction, présentent une adaptation
spécifique aux deux types d'environnements, et se distinguent par des
coefficients de régression non significativement différents de
l'unité. Cependant, le modèle de l'analyse de la
régression conjointe n'arrive à expliquer que 10.1 pourcent de
l'interaction par rapport au modèle de l'analyse AMMI qui montre la
signification des deux premières composantes de l'interaction (IPCA), en
expliquant une importante proportion de l'interaction génotype x
environnement.
123
Nous avons, donc, choisi les techniques multivariées
comme celle des effets principaux additifs et l'interaction multiplicatives
(additive main effects and multiplicative interaction = AMMI) pour la partition
et l'interprétation de l'interaction génotype x environnement.
Selon GAUCH, (1992) [185] l'analyse AMMI est plus efficiente parce qu'elle
explique une grande proportion de la somme des carrés des écarts
de l'interaction. De plus cette analyse sépare les effets principaux de
l'interaction, permettant ainsi une meilleure interprétation des
données issues d'une expérimentation multi-site.
La représentation graphique de l'effet principal
génotype et de l'interaction génotype x environnement,
indiquée par le biplot AMMI1, montre les différences des effets
additifs, c'est-à-dire les différences de rendement entre
génotypes et entre environnements ainsi que les différences des
effets multiplicatifs, liés à l'IPCA1. Ce graphe permet de
prédire le rendement pour toute combinaison génotype x
environnement en utilisant le modèle. Le graphe bidimensionnel du
modèle AMMI1 montre que le score d'un génotype sur de la
1ère composante IPCA1 est un indicateur de la
stabilité des performances de ce génotypes dans les
différents sites.
Les génotypes à large adaptation,
présentent un faible score sur l'IPCA1 et contribuent, moins à
l'interaction. De même les sites qui présentent le même
domaine de recommandation, se caractérisent par un effet principal
élevé, se partageant les mêmes meilleurs génotypes.
D'autre part, quand les scores sur l'IPCA1 et l'effet moyen du biplot AMMI1
sont de même signe, leurs interactions sont positives, alors que si elles
sont des signes contraires, leurs interactions sont négatives. Il en
résulte que la similarité des signes des scores de l'interaction
et les effets principaux d'un couple génotype x environnement
dénote l'adaptation du génotype aux conditions agro-climatiques
de l'environnement en question. Par contre, si les signes sont opposés,
le génotype n'est pas adapté à l'environnement en
question.
Le biplot AMMI2 permet de ranger les génotypes selon
leurs zones d'adaptation, ses zones sont classées selon leur type
d'interaction. Le graphe bidimensionnel procure ainsi un moyen simple pour
déterminer les zones
124
d'adaptation des différents génotypes. Par
conséquent, les analyses du modèle AMMI sont des techniques
statistiques utiles pour l'interprétation des données d'essais
multilocaux, pour le groupement des génotypes et d'environnements selon
des caractéristiques appropriées, et pour l'identification des
zones d'adaptation des génotypes [186].
Une autre méthode a été pratiquée
dans notre étude pour déterminer l'adaptation à des
régions spécifiques identifiées par leurs scores sur la
1ère composante de l'interaction (LIPCA1). Elle est basée sur le
calcul du rendement nominal, Les résultats montrent que les
environnements sont classés selon les valeurs du score de la
1ère composante LIPCA1.De ce fait, chaque génotype a
un rendement nominal le plus élevé dans la plage des scores
LIPCA1 où se trouve l'un des quatre sites ou plus, ce génotype
est donc spécifiquement adapté à la région
représentée par ces sites.
De plus, la variation du rendement nominal entre les
différents sites peut être utilisée comme un indicateur de
la stabilité génotypique, de sorte que, plus la variation du
rendement nominal d'un génotype est faible, plus sa stabilité est
augmente. Bien qu'en générale, une bonne productivité soit
liée à une faible stabilité, PELTONEN-SAINIO et
al., (1993) [187] estiment qu'il est possible de sélectionner des
génotypes d'avoine à la fois productifs et stables, tandis que,
chez le blé tendre d'hivers, OZGEN, (1991) [188] mentionne que de
nouveaux génotypes à la fois productifs et adaptés
à une large gamme de milieux peuvent être créés.
Chez le blé dur, pour notre étude, nos
résultats montrent qu'il est, en effet, possible de cumuler ces
différentes caractéristiques en sélection pour deux
génotypes parmi les trente génotypes évalués. Cette
association entre productivité et adaptation est plus difficile à
réaliser dans les environnements variables, lorsque la sélection
est faite sur la seule base du rendement grain [189], [190].
D'autre part, nous nous sommes intéressés
à des caractères qualitatifs pour l'appréciation de la
qualité technologique. De ce fait, nous avons montré que le taux
de mitadinage est fortement influencé par le site de culture et
l'interaction
125
génotype x milieu que par le facteur
génétique, la diminution des valeurs moyennes de ce
paramètre est proportionnelle à la richesse du sol en azote, et
donc, une forte contribution du facteur environnement qui modifié
l'expression de ce paramètre. Cette idée peut compléter
celle de ABDELLAOUI et MARICHE, (2002) [168] qui notent que la qualité
du blé dur dépend essentiellement de l'accumulation des
protéines. Ces dernières sont largement tributaires de la
nutrition azotée durant le développement de la culture.
Le taux d'extraction, qui varie d'un génotype à
un autre pour le même site et d'un site à un autre pour le
même génotype qui abouti à une forte interaction
génotype × environnement, peut être expliqué par la
variation des conditions de culture d'une part, et la capacité
d'expression de ce paramètre liée à chaque génotype
d'autre part. Il est constaté que les variétés qui
présentent les meilleurs taux d'extraction sont celles qui sont
évaluées dans les milieux les plus favorables à
l'expression des hauts rendements en grains.
D'autre part, l'analyse du comportement viscoélastique
des protéines dans notre essai, montre que la teneur en gluten sec, est
d'autant plus basse que la teneur en gluten humide est plus faible. Ceci
explique les valeurs maximales et minimales qui sont données par les
mêmes génotypes. De même, les résultats obtenus
soulignent la différence d'expression des deux paramètres chez
les 30 génotypes évalués sur les trois sites, ces
génotypes manifestent différentes réponses, traduisant
leur capacité d'expression des deux teneurs en gluten sec et humide
d'une part et l'influence du milieu de culture pour le même
génotype d'autre part. ceux qui abouti à une forte interaction
génotype x milieu.
ABDELLAOUI et MARICHE, (2002) [168] à travers une
étude sur deux variétés de blé dur (waha et
vitron), indiquent que les apports d'azote exercent une action croissante sur
les teneurs en gluten sec des deux variétés. La maîtrise de
la nutrition azotée et la fertilisation du sol, en
générale, des variétés locales et introduites
pourraient contribuer à l'amélioration de la qualité du
blé dur en Algérie.
126
CONCLUSION
La fluctuation des conditions de croissance d'un environnement
à un autre et l'instabilité climatique qui caractérise les
zones céréalières en Algérie sont vues comme des
contraintes à l'origine de la variabilité des rendements.
En effet, à travers l'étude du comportement
génotypique intra et inter sites, nous pouvons constater que, le
classement des génotypes pour un caractère donné, sur un
site ne suit pas fidèlement le classement du même caractère
sur les autres sites. Ainsi que, les changement des caractères qui
conditionnent le rendement d'un lieu test à un autre. Ceux qui
illustrent, clairement, l'influence de la diversité géographique
et climatique sur les potentialités génétiques des
génotypes évalués.
Ces résultats incitent à la sélection de
génotypes adaptés aux exigences environnementales locales, ce qui
requiert l'analyse des interactions génotypes x milieu du rendement en
grains à l'aide des différentes méthodes. Ces
dernières sont fondées sur des analyses statistiques
dérivées de la régression ainsi que le modèle AMMI.
Ce dernier à prouver une efficacité que l'analyse de la
régression conjointe qui n'arrive à expliquer qu'une faible
proportion de l'interaction. Cette étude indique que les conditions
pédo-climatiques des lieux testés, affectent significativement le
comportement des génotypes et induisent l'interaction génotype x
environnement.
L'analyse AMMI regroupe les différents sites en deux
domaines de recommandation Khroub, Sétif et Tiaret vs Oued Smar. Le
génotype Bichena / Ariza_2//Solga_8 (n°3) présente un
rendement en grains élevé associé à une bonne
stabilité, et une large adaptation à tous les sites
d'évaluation, par contre le génotype Gcn/4/D68-1-93A-1A//Ruff /
Fg /3/ Mtl-5 (n°12) montre une adaptation spécifique au site Oued
Smar.
.
127
Ces résultats indiquent que le sélectionneur a
le choix entre la sélection du génotype Bichena /
Ariza_2//Solga_8 (n°3) pour tous les sites ou bien recommander ce
génotype aux régions représentées par les sites
Khroub, Sétif et Tiaret et de sélectionner le génotype
Gcn/4/D68-1-93A-1A//Ruff / Fg /3/ Mtl-5 (n°12) spécialement pour la
région représentée par le site Oued Smar. La
stratégie associant l'adaptation générale à une
large région représentée par les trois sites Khroub,
Sétif et Tiaret avec l'adaptation spécifique pour Oued Smar
induit à un gain de rendement estimé à 5.94%.
Ces deux génotypes ressortent aussi comme les plus
performants parmi les 30 génotypes évalués sur la base de
leur haut rendement nominal relativement stable et de ce fait sont à
recommander sur l'ensemble des régions représentées par
les 4 sites.
Les variables relatives à l'appréciation de la
qualité technologique, quant à elles ont été
affectées par l'effet génotype, environnement et l'interaction
génotype x environnement. De ce fait, les génotypes
évalués acquièrent une qualité technologique
d'autant plus acceptable que le milieu devient plus favorable, notamment la
fertilité du sol.
La recherche des génotypes performants,
caractérisés par une large adaptation ou par une adaptation
spécifique à une zone déterminée fait appel
à l'étude et la modélisation de l'interaction. Elle
nécessite une adaptation des méthodes, tant au niveau statistique
qu'au niveau agronomique, pour structurer l'information et adapter les
modèles de fonctionnement des cultures à la
caractérisation d'un réseau multi local et
pluriannuel.
Il est suggéré d'augmenter le nombre de lieux
tests pour mieux appréhender la réponse des génotypes aux
variations des lieux et d'identifier les contraintes qui ont une occurrence
régulière pour sélectionner un matériel
tolérant et stable. Ces premiers résultats obtenus peuvent
constituer une base de travail dans ce domaine de recherche sur les blés
en Algérie.
128
REFERENCES
1. Hervieu, B., Capone, R., S. Abis, S., «The challenge
posed by the cereals
sector in the Mediterranean», Ciheam analytical
note, n°9, (2006), 14 p.
2. Chenafi, H., Bouzerzour, H., Aidaoui, A. and Saci, A.,
«Yield response of
durum wheat (Triticum durum, Desf) cultivar Waha to
deficit irrigation under semi arid growth conditions», Asian Journal plant
Sci., n°5, (2006), 854-860.
3. Bahlouli, F., Bouzerzour, H., Benmahammed, A. and Hassous,
K.L.,
«Selection of high yielding and risk efficient durum wheat
(Triticum durum Desf.) Cultivars under semi arid conditions», J. Agro,
n°4, (2005), 360-365.
4. Yan, W., Hunt, L.A., Sheng, Q.and Szlavnics,Z.,
«Cultivar evaluation and
mega-environment investigation based on the
GGE biplot», Crop Sci, n°40, (2000), 597-605.
5. De la Vega, A.J., Chapman, S.C. et Hall, A.J.,
«Genotype by environment
interaction and indirect selection for yield
in sunflower. I. Two-mode pattern analysis of oil and biomass yield across
environments in Argentina», Field Crops Res, n° 72, (2001), 17-38.
6. Farshadfar, E. et Sutka, J., «Locating QTLs
controlling adaptation in wheat
using AMMI model», Cereal Res. Commun,
n°31, (2003), 249-255.
7. Annicchiarico, P., Bellah, F. et Chiari, T.,
«Repeatable genotype x location
interaction and its exploitation by
conventional and GIS-based cultivar recommendation for durum wheat in
Algeria», Eur, J. Agronomy, n°24, (2006), 70-81.
8. Oosterom, V.E.J., Klein, D., Ceccarelli, S. et Nachit,
M.M., «Genotype x
Environment Interactions of barley in the
Mediterranean region», Crop Science, n°33, (1993), 669-674
9. Lecomte, C., Brancourt-Hulmel, M., Doussinault, G.,
Berard, P., Le Buanec,
V. et Trotte, M., «Genetic improvement of
agronomic traits of winter wheat cultivars released in France from 1946 to
1992», Crop Sci, n° 43, (2003), 37-45.
10. Ebdon, J.S. et Gauch H.G., «Additive main effect and
multiplicative
interaction analysis of national turf grass performance
trials: I. Interpretation of genotype x environment interaction», Crop
Sci, n°42, (2002), 489-496.
11.
129
Finlay, K.W. et Wilkinson, G.N., «The analysis of
adaptation in a plant
breeding programme», Aust. J. Agric, Res,
n°14, (1963), 742-754.
12. Eberhart, S.A. et Russell, W.A., «Stability
parameters for comparing
varieties», Crop Sci, n°6, (1966),
36-40.
13. Zobel, R.W., Wright, M.J. et H.G. Gauch, H.G.,
«Statistical analysis of a
yield trial», Agron. J, n°80,
(1988), 388-393.
14. Yan, W. et Rajcan, I., «Biplot evaluation of test
sites and trait relations of
soybean in Ontario», Crop Sci, n°42,
(2002), 11-20.
15. Fan, X.M., Kang, M.S., Chen, H.M., Zhang, Y., Tan, J. et
Xu. C., «Yield
Stability of Maize Hybrids Evaluated in
Multi-Environment Trials in Yunnan», China. Agron. J., n°99, (2007),
220-228.
16. Cooper, M. et Delacy, I.H., «Relationships among
analytical methods used
to study genotypic variation and
genotype-by-environment interaction in plant breeding multi-environment
experiments», Theor, Appl. Genet, n°88, (1994), 561-572.
17. C.I.C., «Perspectives agricoles», Anonyme,
Conseil international des céréales, (2007).
18. Bonjean, A., Picard, E., «Les céréales
à paille. Origine, histoire, économie, sélection»,
(1990), 205 p.
19. C.I.C., «Perspectives agricoles», Anonyme,
Conseil international des céréales, (2002-2003).
20. Statistique Canada., «Perspectives agricoles Canada
», Anonyme, (2007).
21. C.I.C., «Perspectives agricoles», Anonyme,
Conseil international des céréales, (2006-2007).
22. DSASI., «Statistiques agricoles, superficies et
productions, serie B. 2000 à 2007», Ed. Direction des statistiques
agricoles et des systèmes d'information. Ministère de
l'agriculture et Développement rural, Algérie, (2007).
23. Laumont, p., Erroux, J., «Inventaire des blés
durs rencontrés et cultivés en Algérie»,
Mémoire soc, Hist. Afri Nord, n°5, (1961), 1-96.
24. Anonyme., «La céréaliculture en
Algérie », Ed. Centre d'études économiques °77,
(1970), 66p
25. Ducellier, L., «Espèces et
variétés de céréales cultivées en
Algérie» Direction de l'agriculture et de la colonisation, (1930),
1-62.
130
26. Abdelguerfi, A. et Laouar, M., «Les ressources
génétiques des blés en Algérie. Passé,
présent et avenir», In Actes du premier symposium international sur
la filière blé : Enjeux et stratégie, Alger, (7 au 9
février 2000), 133-148.
27. Benbelkacem, A., «La recherche variétale sur
les blés en Algérie»,
Céréaliculture,
(1993), n°26, 3-8.
28. Boufanar -Z, F. et Zaghouane, O., «Guide des
principales variétés de
céréales à paille
en Algérie : blé dur, blé tendre, orge et avoine»,
Première édition 2006. ITGC/ICARDA/INRA/CNCC.MADR, ITGC, (2006),
154p.
29. Boulal, H., Zaghouane, O., El Mourid, M. et Rezgui, S.,
«Guide pratique de
la conduite des céréales d'automne
(blés et orge) dans le Maghreb (Algérie, Maroc, Tunisie) »,
ITGC/INRAA/ICARDA. MADR, Ed. ITGC, Algérie, (2007), 180p.
30. Benbelkacem, A., «Etude de l'adaptation
variétale des céréales cultivées
en
Algérie Sous différentes conditions
agro-écologiques», Céréaliculture, ITGC /MADR,
n°31, (1997), 17-22.
31. Hakimi, M., «l'orge une culture de survie en
Algérie», in Séminaire sur
:
Agrométéorologique des systèmes d'agriculture
pluviale fondés sur la culture de l'orge. ONM-ICARDA, (1989), 1-7.
32. Cheftel, J.C., Cheftel, H. et Besançon, P.,
«introduction à la biochimie et à
la technologie des
aliments», Ed, Lavoisier, V.1, (1977).
33. Zaghouane-B, F., Merabti, A., Zaghouane, O., Bouabdelli,
F., Ait Abdellah,
J. et Amrani, M., «Le blé dur :
qualité, importance et utilisation dans la région des hauts-
plateaux (Tiaret et Tissemsilt) », Projet IRDEN, DOC. ITGC, (2003),
44p.
34. Nolot, J.M., «L'analyse des interactions
génotype x environnement :
l'approche agrophysiologiques», In :
Séminaire sur la Tolérance à la sécheresse des
céréales en zones méditerranéennes.
Diversité génétiques et amélioration
variétale, Monpellier, France, Ed. Ben Salem, M et Monneveux, P., INRA,
Les colloques, n°64, (15-17 décembre 1992), 101-116.
35. Baldy, G., «Contribution à l'étude
fréquentielle des conditions climatiques et
de leurs influences sur
la production des principales zones céréalières», Doc
du projet céréale, (1974a), 170p.
36. Abbassenne, F., Bouzerzour, H. et Hachemi, L.,
«Phénologie et production du blé dur (Triticum durum
Desf.) en zone semi-aride d'altitude», Annales Agronomiques de l'INA,
n°8, (1997a), 16-22.
37. Benchohra, B. et Khelloufi, B., «Impact du risque
climatique sur le
rendement du blé dur (Triticum durum
Desf.) en zone semi-aride : Approche
131
éco-physiologique», Science et changements
planétaires/Sécheresse, V. 11, n° 1, (Mars 2000), 45-51
38. Nachit, M.M., Picard, E., Monneuveux, P., Labhilili, M. et
Rivool, R.
«Présentation d'un programme international
d'amélioration du blé dur pour le bassin
méditerranéen», Cahier d'étude et de recherche
Francophones / Agriculture, Réseau SEOUANA, V. 7, n° 6,
(novembre-décembre 1998), 510p.
39. Hsissou, D. et Bouharmont, J., «in vitro selection
and characterization of
drought tolerant plants of durum wheat (Triticum
durum Desf.) », Agronomie, n° 2, (1994), 65-70.
40. Hurd, E.A., Townly, S., Patterson, L. et Owen, Ch.,
«Technics used in
producing wascana wheat», Can J paint Sc,
n° 52, (1972), 689-691.
41. El Mourid, M., «Performance of wheat and barley
cultivars under different
soil moisture regimes in a semi-arid region»,
Ph. D thesis, Iowa State University, (1988), 229p.
42. El Falah, M., Chalbi, N. et El Guazzah, M., «Analyse
de l'adaptation à
l'aridité de quelques ressources
génétiques locales d'orge (Hordeum Vulgare.L)
comparativement à des variétés
améliorées», In: L'amélioration des plantes pour
l'adaptation aux milieu arides, AUPELF-UREF, Ed. John Libbey, Paris, (1991),
197-209.
43. Samarah, N.H., «Effect of drought stress on growth
and yield of barley»,
Agronomie, Sustain, Dev, n° 25, (2005),
145-149.
44. Tambussi E.A., Nogués, S., Ferrio, P., Voltas, J.
et Araus, J.L., «Does
higher yield potential improve barley performance
in Mediterranean conditions? », Field crop research, n°91, (2005),
149-160.
45. Gate, P., «Ecophysiologie du blé»,
Technique et documentation Lavoisier, Paris, (1995), 351p.
46. Gate, P., «Stades du blé », Ed. ITCF,
France, (2003), 68p.
47. Sarrafi, A., Mentewab, A. et Monneuveux, P.,
«Variabilité génétique de la fluorescence
chlorophyllienne chez des haploïdes doubles d'orge et son utilisation dans
la sélection pour la résistance au stress hydrique », In :
Séminaire sur la Tolérance à la sécheresse des
céréales en zones méditerranéennes.
Diversité génétiques et amélioration
variétale, Montpellier, France, Ed. Ben Salem, M et Monneveux, P., INRA,
Les colloques, n°64, (15-17 décembre 1992), 397-404.
48. Simon, H., Codaccioni, P. et Lecoeur, X., «Produire
des céréales à paille», Technique et documentation-
Lavoisier, Paris, (1989), 333p.
49.
132
Entez, M.H. et Fowler, D.B., «Critical stress periods
affecting productivity of
no- till winter wheat in western Canada »,
Agron. Journal, V. 80, n° 6 (1988), 987-992.
50. Al Khatib, K. et Paulsen, G.M., «Photosynthsis and
productivity during high
temperature stress of wheat genotype from major
world regions. Crop Sci. n°30, (1990), 1127-1132
51. Ouabbou, H., «Physiological aspects of Recovery and
evaluation of wheat
during high temperature stress», Phd. Kansas State
University, USA, (1995).
52. Gazeau, C., «Les végétaux et les
basses températures», Marina Botard ep. Raffaud tuteur : Claude
Gazeau, Université Pierre et Marie curie (Paris 6), DEUG science et vie,
(2002).
53. Gate, P., Brain, P., Colnenne, J. et Briffeaux, J.,
«Pour les céréales à paille
à chaque
variété son époque de semis», Perspective Agricole,
n°148, (1990), 20-27.
54. Baldy, C.H., «Progrès récents
concernant l'étude du système racinaire du
blé»,
Annal Agronomie, V. 24, n° 2, (1974b), 241-276.
55. El Mourid, M., Lamine, M., Boutfiass, M. et Farihane, H.,
«Simulation de
l'effet de l'irrigation d'appoint sur la
productivité du blé tendre dans les régions semi- arides
du Maroc», In: Supplementary irrigation and drought water management,
International conference , Valenzano, Bari, V. 1, (Septembre 27- octobre 2
1992), 1-14.
56. Harris, H.C., Goebbel, W. et Cooper, P.G.M., «Crop
genotype environnement interaction», ICARDA, (1987), 37-47.
57. EL-Khachin, A., «Les grandes cultures du
blé», Ed. EL maarf, (1996),
32-42.
58. Hamadache, A., Abdellaoui, Z. et Aknine, M.,
«Facteurs agrotechniques
d'amélioration de la
productivité du blé dur en Algérie. Cas de la zone
sub-humide», Revue de la Recherches Agronomiques de l'INRA,
Algérie, n°10, (Juin 2002), 5-18.
59. Mecklouf, A., « Etude de la variabilité
génétique du blé dur (Triticum durum,
Desf), pour la
tolérance au froid », Thèse de doctorat, INA, EL Harrach,
Algérie, (2008), 130p.
60. Wilhelm, W.W., Mielke, L.N. et Fenster, C.R., «Root
development of winter
wheat as related to tillage practice in Western
Nebraska», Agron. J, n° 74, (1982), 85-88.
61.
133
Kribaa, M., Hallaire, S. et Curmi, J., «Effects of
tillage methods on soil
hydraulic conductivity and durum wheat grain yield
in semi-arid area», Soil and Tillage, n° 37, (2001), 17-28.
62. Klein, J.D., Cohen I., Hebbe, S., Asido, Y., Dolgin, B.
et Bonil, D. J.,
«Establishment of wheat seedlings after early soming
and germination in an arid Mediterranean environment», Agron. J,
n°94, (2002), 585-893.
63. Saber, N. et Mrabet, R., «Impact of no tillage and
crop sequence on
selected soil quality attributes of a vertic calcixeroll
soil in Morocco», Agronomy, n°22, (2002), 451-459.
64. Yalcin, H., Cakir, E. et Aykas, E., «Tillage
parameters and economic
analysis of direct seeding, minimum and conventional
tillage in wheat», Journal of agronomy, n°4, (2005), 329-332.
65. Worland, A.J., Apendina, M.L. et Sayer, E.J., «The
distribution in European
winter wheat of genes that influence ecoclimatic
adaptability while determining photoperiod intensively and plant height»,
Euphytica, n°80, (1994), 219-228.
66. Lin, Z., Pinson, S.R.M., Stansel, J.W. et Park, W.D.,
«Identification of
quantitative trait loci (QTLs) for heading date and
plant height in cultivated rice (Oryza sativa L.) », Theor. Appl. Genet,
n°91, 374-381.
67. Gallais, A., «Théorie de la sélection
en amélioration des plantes»,
Collection sciences agronomiques,
Ed. Masson, (1990), 588p.
68. Jacquard, P., Rotili, P. et Zannone, L., «Les
interaction génotype x milieu
biologique : analyse diallèle
des aptitudes à l'association entre populations de trèfle
violet», Ann. Amélior. Plantes, 26, (1978), 309-325.
69. Kempton, R.A. et Fox, P.N., «Statistical methods for
plant variety evaluation
Chapman et Hall, London, (1997).
70. Haldane, J.B.S., «The interaction of nature and
nurture», Ann. Eugen.
Lond, (1946), 197-205.
71. Grafius, J.E., «Multiple characters and correlated
responses», Crop sci,
n°18, (1978), 931-934.
72. Benmahammed, A., «Association et
héritabilités de quelques caractères à
variation
continue chez l'orge ( Hordeum Vulgare L.) », Thèse de
Magister , INA, EL Harrach, (1996), 80p.
73. Abbassenne, F., «Etude génétique de la
durée des phases de développement et influence sur le rendement
et ces composantes chez le blé dur (Triticum durum, Desf)»,
Thèse de Magister. INA, El harrach, Algérie, (1997),62p.
74.
134
Ceccarelli, S., Grando, S. et Hamblin, J., «Relationship
between barley
grain yield measured in low and high yielding
environments», Euphytica, n°64, (1992), 49-58.
75. Gate, P., Brain, P., Colnenne, J. et Briffeaux, J.,
«Pour les céréales à paille
à chaque
variété son époque de semis», Perspective Agricole,
n°148, (1990), 20-27.
76. Oosterom, VE. et Acevedo, E., «Adaptation of barley
to harsh Mediterranean environments. II Apical development leaf and tiller
appearance», Euphytica, n°62, (1992), 15-27.
77. Le Guis, J., «Grain filling and shoot growth of
2-row and 6-row winter barley
varieties», Agronomie, n°13, (1993),
545-552.
78. Martine, O.G., «Structure de la variabilité
du rendement du blé d'hiver dans
quelques département
céréalier français, I. variabilité entre
années et entre départements du peuplement épis et du
poids du grain par épi», Agronomie 3, n°9, (1993), 917-924.
79. Acevedo, E., Graufurd, P.Q., Austin, R.B. et Perez-Marco,
P., «Traits
associated with high yield in barley in low rainfall
environments», J. Agr, Sci. Camb, n°116, (1991), 23-36.
80. Ali Dib, T. et Monneveux, P., «Adaptation à
la sécheresse et notion d'idéiotype chez le blé dur. I.
Caractères morphologiques d'enracinement», Agronomie, n°12,
(1992), 371-379.
81. Fisher, R.A. et Maurer, R., «Drought resistance in
spring wheat cultivars.I.
Grain yields responses», Aust. J. Agr, Res
n°29, (1978), 897-912.
82. Ouhajou, L., «Efficience d'utilisation de l'eau et
stabilité des rendements
des céréales»,
Mémoire de 3ème cycle IAV, Institut Agronomique et
Vétérinaire, Hassan II, Département production
végétale, Rabat, (1991).
83. Hamadi, I., «Effet du régime hydrique et de
la variété sur la production du
blé tendre en zone
semi-aride», Mémoire de 3ème cycle, ENS, Meknes, (1992),
136p.
84. Bouzerzour, H. et Monneveux, P., «Analyse des
facteurs de stabilité du
rendement de l'orge dans les conditions des
Hauts-plateaux de l'Est algérien», In : Tolérance à
la sécheresse des céréales en zones
méditerranéennes. Diversité génétiques et
amélioration variétale, Montpellier, France, Ed. Ben Salem, M et
Monneveux, P., INRA ed, Les colloques, n°64, (15-17 décembre 1992),
139-158.
85. Mouret, J.C., Conesa, A.P., Bouchier, A., Gaid, M. et
Ouled Said, O.,
«Identification des facteurs de variabilité du
rendement du blé dur en conditions hydriques limitantes dans la
région de Sidi Bel Abbès (Algérie).
135
(1991)», In: Acevedo, E., Conesa, A.P., Monneveux, P. and
Srivastava, J.P., Edts. Physiology breeding of winter cereals for stressed
Mediterranean Environments, INRA ed, Montpollier, France, (3-6 July 1989),
35-47.
86. Meziani, L., Bamoun, A., Hamou, M., Brinis, L. et
Monneveux, P., «Essai de
définition des caractères
d'adaptatin du blé dur dans différentes zones agro-climatiques de
l'Algérie», In : Séminaire sur la Tolérance à
la sécheresse des céréales en zones
méditerranéennes. Diversité génétiques et
amélioration variétale, Montpellier, France, Ed. Ben Salem, M et
Monneveux, P., INRA ed, Les colloques, n°64, (15-17 décembre 1992),
191-203.
87. Triboi, E., «Modèle d'élaboration du
poids du grain chez le blé tender
(Triticum aestivum. En
thell) », Agronomie, n°10, (1990), 191-200.
88. Auriau, P., Doussinault, G., Jahier, J., Lecomte, C.,
Pierre, J., Pluchard, P.,
Rousset, M., Saur, L. et Trottet, M., «Le
blé tendre », In : Gallais, A. et Bannerot, H., Eds;
Amélioration de espèces végétales Cultivées,
INRA ed, (1992), 22-38.
89. Nachit, M.M. et Jarrah, M., «Association of some
morphological characters
to grain yield in durum wheat under Mediterranean
dry land conditions», Rachis, n°5, (1986), 33-34.
90. Donmez, E., Sears, R.G., Shroyer, J.P. et Paulsen, G.M.,
«Genetic grain in
yield attributes of winter wheat in the great plains
», Crop Science, n°41, (2001), 1412-1419.
91. Lafitte, HR. et Courtois, B., «Interpreting cultivar
x environment interaction
for yield in upland rice assigning value to
drought-adaptive trait », Crop Science, n°42, (2002), 1409-1420.
92. Gallais, A., «Adaptation et adaptabilité en
amélioration des plantes», Sel Fr,
n°42, (1992a), 55-57.
93. Papadakis, J.S., «Ecologie agricole », Eds;
Jules Duculot, Gembloux,
(1932), 303p.
94. Bouzerzour, H., Adjabi, A., Benmahammed, A., Hadj
Sahraoui, A. et
Harkati, N., «Production et adaptation comparées
des variétés de céréales en zone semi-aride
d'altitude », Céréaliculture, n°37, (2002), 4-13.
95. Fellah, A., Benmahammed, A., Djekoun, A. et Bouzerzour,
H., «selection
pour améliorer la tolérance aux stress
abiotique chez le blé dur (Triticum durum Desf.) », Actes
de l'IAV, Hassen II, Maroc, n°22, (2002), 161-170.
96. Gallais., A., «Bases génétiques et
stratégie de sélection de
l'adaptation
générale», Sel Fr, n°42, (1992b),
59-78.
136
97. Brancourt-Hulmel, M., «Sélection
variétale et milieu Sélection pour l'adaptation au milieu et
prise en compte des interactions génotype x milieu», Dossier :
Agriculture, recherche et territoire, V 7, n° 6, (Novembre -
Décembre 2000), 504-511.
98. Barbottin, A., «Utilisation d'un modèle de
culture pour évaluer le comportement des genotypes : pertinence de
l'utilisation d'Azodyn pour analyser la variabilité du rendement et de
la teneur en protéines du blé tendre», Thèse de
Doctorat, INA, Paris-Grignon, (2004), 167p.
99. Trottet, M. et Doussinault, G., «Analyse du
progrès génétique chez le blé
tendre au cours du
XXème siècle », le Sélectionneur Français,
(2002), n°53, 3-13.
100. Bouzerzour, H., Benmahammed, A., Benbelkacem, A.,
Hazmoune, T., Mimoun, H., Bourmel, S. et Mekhlouf, A., «stabilité
des performances phéno-morphologiques de quelques variétés
de blé dur (Triticum durum, desf.), issues d'une
sélection multilocale», In: Actes du premier symposium
International sur la filière blé . Enjeux et Stratégie,
Alger, (7-9 février 2000), 187-195.
101. Léon, J., «Mating system and the effect of
heterogeneity and heterozygoty on phenotypic stability», In: VAN OOIJEN
FW, JANSEN J, eds. Biometrics in plant breeding. Proc. Meeting of the eucarpia
section biometrics in plant breeding 9th, (6-8 July 1994), 19-31.
102. Sarawat, P., Stoddard, FL, Marshall,
DR. et ALI, SM., «Hererosis for yield
and related characters in pea», Euphytica, n°80, (1994), 39-48
103. Beavis,
WD. et Keim, P., «Identification of
quantitative trait loci that are affected by environment», In: KANG MS,
GAUCH HG, eds. Genotype-by-environment interaction, Boca Raton: CRC-Press,
(1996), 123-49.
104. Labhilili, M. et Bentata, F.,
«caractérisation de gènes de tolérance à la
sécheresse chez le blé dur», Dossier d'Agropolis
International : La plate-forme de recherches avancées Agropolis,
n°5, (2004), 30-31.
105. Talbot., «Resource allocation for selection systems
», In : KEMPTON RA, FOX PN, eds. Statistical methods for plant variety
evaluation. London : Chapman & Hall, (1997), 162-174.
106. Weber, WE., Wricke, G. et Westermann, T.,
«Selection of genotypes and prediction of performance by analyzing
genotype-by-environment interactions», In : KANG MS, GAUCH HG, eds.
Genotype-by-environment interaction. Boca Raton: CRC-Press, (1996), 353-71.
107. Moreau, L., Lemarle, S., Charsset, A. et Gallais, A.,
«Economic efficiency of one cycle of marker-assisted selection», Crop
Sci, n°40, (2000), 329-37.
137
108. Wricke, G. et Wener, WE., «Selection between clones
and homozygous lines. In : WRICKE G, WEBER WE, eds. Quantitative genetics and
selection in plant breeding»,Berlin, New York : De Gruyter, (1986),
171-94.
109. Hachemi, L., «Le blé dur en Algérie. 5th
cereal workshop on the gap between present farm yield and the potential»,
(1979), 19-75.
110. Blum, A., «Plant breeding for stress
environments», CRC press, Inc, Florida, USA, (1988), 223p.
111. Fisher, R.A., «Number of kernels in wheat crop and the
influence of solar radiation and temeratures», J Agric.Sci, Camb,
n°105, (1985), 447-461.
112. Braun, HJ., Rajram, S. et Van Ginkel, M., «CIMMYT's
approach to breeding for wide adaptation», In : TIGERSTEDT PMA, ed.
Adaptation in plant breeding. The Netherlands: Kluwer Academic Publishers,
(1997), 197-205.
113. Brancourt-Hulmel, M., Biarnès-Dumoulin, V. et
Denis, JB., «Points de repère dans l'analyse de la stabilité
et de l'interaction génotype/milieu en amélioration des
plantes», Agronomie, n°17, (1997), 219-246
114. Cattivelli, L., Baldi, P., Crosatti, N., Di Fonza, N. et
Stanca, M., «Chromosome regions and stress related sequences involved in
resistance to abiotic stress in Triticeae », Plant Molecular Biology,
n°48, (2002), 649-665.
115. Lin, C.S., Binns, M.R. and Lefkovitch, L.P.,
«Stability analysis: where do we stand? », Crop Sci, n°26,
(1986), 894-900.
116. Brancourt-Hulmel, M. et Lecomte, C., «Slection et
stabilité du rendement chez le blé tendre d'hiver »,
Agronmie, n°14, (1994), 611-625.
117. Lin,
CS. et Binns, MR., «A method of
analyzing cultivar x location x year experiment: a new stability
parameter», Theor Appl Genet, n°76, (1988), 425-430.
118. Hamadache, A., Abdellaoui, Z. et Aknine, M.,
«Facteurs agrotechniques d'amélioration de la productivité
du blé dur en Algérie. Cas de la zone sub-humide», Revue de
la Recherches Agronomiques de l'INRA, Algérie, n°10, (Juin 2002),
5-18.
119. ITGC., «Recueil des fiches techniques des cereals
d'hiver, fourrages, légumineuses alimentaires et
oléagineuses», Institut technique des grandes cultures, MADR,
(2006).
120. Meslem, M., «Le phosphore assimilable dans les sols
des hautes plaines Sétifienne : Analyses chimiques et isotopiques et
incidence sur la fertilisation phosphatée», Thèse mag, INES
Biol, Univ. Sétif, (1992), 106p.
138
121. Baldy, C., Ruelle, P. et Fernandes, A.,
«Résistance à la sécheresse du sorgho-grain en climat
méditerranéen», Sécheresse, n°4, (1993),
85-93.
122. Makhlouf, M., Hamadache, A. et Harkati, N., «Effets
de la date et de l'outils de travail du sol sur le comportement des bromes
(Bromus sp.) et le rendement du blé dur (Triticum durum Desf.)
dans la région de Sétif », Céréaliculture,
n°37, (2002), 24-29.
123. O.N.M., «Données
météorologiques, Série A», Anonyme, Office Nationale
de Météorologie, (2008).
124. Benbelkacem, A. et Zeltni, A., «Présentation
des résultats des activités amélioration des
céréales 2006/2007», Bilan Amélioration 2006/2007,
ITGC El Khroub, (2007).
125. Beccari, J., «De Bobniensi Scientarum et Artium
Instituto atque Academia», n°2, (1745), 122-127.
126. Pierre, F., «Le grain de blé composition et
utilisation», INRA, Paris, France, (2000), 308p.
127. Cropstat 7.2.3., «Software package for windows
», International Rice Research Institute, IRRI, Manila, (2008).
128. Steel, R.G.D. et Torrie, J.H., «Principles and
procedures of statistics, a biometrical approach», Mc Graw Hill Inc,
(1980), 633 pp.
129. Annicchiarico, P., «Genotype by environment
interactions: Challenges and opportunities for plant breeding and cultivar
recommendations», FAO, Rome, (2002), 159 p.
130. Ceccarelli, S., Grando, S. et impigha, A., «Choice
of selection strategy inbreeding barley for stress environment»,
Euphytica, n° 103, (1998), 307-313.
131. Annichiarico, P., Abdellaoui, Z., Kelkouli , M. et
Zerargui, H., «Grain yield, straw yield and economic value of tall and
semi-dwarf durum wheat cultivars in Algeria », Journal of Agricultural
Science, n°143, (2005), 57-64.
132. Kirby, E.J.M., Spink, J.h., Frost, D.l et Evans, E.J.,
«A study of wheat development in the field: Analysis by phases»,
European journal of Agronomy, n°11, (1999), 63-82.
133. Simane, B., Peacock, JM. et Struik, PC.,
«Differences in development plasticity growth rate among drought.
Resistant and susceptiblecultivars of durum wheat (Triticum turgidum L.
var.durum) », Plant and soil, 157, (1993), 155-166.
139
134. Mekhlouf, A., Bouzerzour, H., Benmahammed, A., Hadj
Sahraoui, A. et Harkati, N., «Adaptation des variétés de
blé dur (Triticum durum Desf.) au climat semi-aride»,
Sécheresse, article scientifique, 17(4), (2006), 507-513.
135. Djebrani, M., «Adaptation au dificit hydrique de
quatre variétés de blé dur», In: Actes du premier
symposium International sur la filière blé . Enjeux et
Stratégie, Alger, (7-9 février 2000), 161-169.
136. Ben Abdellah, N. et Ben Salem., «Paramètres
morphophysiologiques de sélection pour la résistances à la
sécheresse des céréales », In : Séminaire sur
la Tolérance à la sécheresse des céréales en
zones méditerranéennes. Diversité génétiques
et amélioration variétale, Montpellier, France, Ed. Ben Salem, M
et Monneveux, P., INRA ed, Les colloques, n°64, (15-17 décembre
1992), 173-190.
137. Bouzerzour, H. et Oudina, M., «Variabilité
du rendement de l'orge sous l'influence du climat des hauts plateaux», In
: Séminaire W.O.M, Eds. ICARDA, (1989), 110-119.
138. Subbiah, B.V., Katyal, J.C., Narasimham, R.L. et
Dakshinamurti, C., «Preliminary investigation on root distribution of
yielding wheat varieties », Inst, J, Appl.Radiat, Isot, n°19, (1968),
385-390.
139. Hiou, S., dekak, A. et Khamar, L., «Effets des
dates de semis et le traitement hydrique sur 12 variétés de
blé dur en zone semi-aride », Céréaliculture, ITGC,
n°37, (2002), 17-23.
140. Abbassene, F., Bouzerzour, H. et Hachemi, L., «Taux
de remplissage et durée de la phase de remplissage chez 14
génotypes de blé dur (Triticum durum Desf.) en zone
semi-aride», annale agronomique, n°43, (1998), 75-81.
141. Benmahammed, A., Nouar, H., Haddad, L., Laala, Z.,
Oulmi, A. et Bouzerzour, H., «Analyse de la stabilité des
performances de rendement du blé dur (Triticum durum Desf.)
sous conditions semi-arides », Biotechnol. Agron. Soc. Environ, n°
14, (2010), 1-12,
142. Melki, M., Dahman, A.B.K. et Garoui, A., «effet de
la variation saisonnières des facteurs climatiques sur les composantes
du rendement de céréales (blé dur et orge) », Revue
de l'INAT, vol.10, n°1, (Juin 1995), 105-114.
143. Benbelkacem, A., Mekni, M.S. et Rasmusson, D.C.,
«Breeding for high tiller number and yield in barley», Crop Sci,
n°24, (1984), 968-972.
144 Grignac, P., «Rendement et composantes du rendement
du blé d'hiver dans l'environnement méditerranéen
français CEE, division de la coordination de la recherche »,
Séminaire de Bari (Italie), 1-2, (30/10/1981), 185-195.
140
145. M'hedbi, K., «Effets de la dose de semis sur le
rendement et les composantes du rendement de l'orge cultivée en sec
», In : Actes des 2ème journées nationales sur les acquis de
la recherche Agronomique Halieutique et Vétérinaire, Kef
(Tunisie), V.1, (8-10 décembre 1995), 94-99.
146. Hamadache, A., Ait Abdellah, F. et Ladada, M.,
«Synthèse des travaux de recherche réalisés par les
fermes expérimentales sur la protection des grandes cultures »,
Bilan de la recherche sur les grandes cultures : 1977-1997, ITGC, (1998),
13p.
147. Wardlaw, I.F., «Interaction between drought and
chronic high temperature during kernel filling in wheat in a controlled
environment», Ann. Bot, n°90, (2002), 469-476.
148. Benbelkacem, A. et Kellou., «Evaluation du
progrès génétique chez quelques variétés de
blé dur (Triticum durum Desf.) cultivées en
Algérie», In: Actes du premier symposium International sur la
filière blé . Enjeux et Stratégie, Alger, (7-9
février 2000), 123-131.
149. Triboi, E., Blancon, J. et Magne, J.,
«Déterminisme du poids moyen du grain chez le blé »,
C.R. Acad, Agri, France, n°71, (1985), 871-876.
150. Mekhlouf, A., Bouzerzour, H., Dehbi, F. et Hannachi, A.,
«Rythme de développement et variabilité de réponse du
blé dur aux basses températures. Tentatives de sélection
pour la tolérance au gel », In : séminaire sur la
valorisation des milieux semi-arides, centre Universitaire OEB, (mai 2001).
151. Bahloul, F., Bouzerzour, H. et Benmahammed, A.,
«Effet de la vitesse et de la durée du remplissage du grain ainsi
que de l'accumulation des assimilas de la tige dans l'élaboration du
rendement du blé dur (Triticum durum Desf.), dans les
conditions de culture des hautes plaines orientales d'Algérie»,
Biotechnol. Agron. Soc. Environ, 12(1), (2008), 31-39.
152. Coventry, D.R., Brouke, H.D., Conn, D.R. et Reevest,
T.G., «Influence of genotype, sowing date and seeding rate on wheat
development and yield», Australian of Agronomy, (1993), 751-757.
153. Berger, M., «Etude des caractères
morpho-physiologiques, des composantes du rendement et de leurs
corrélations génétiques et environnementales chez le
blé tendre», Thèse Docteur Ingenieur, ENSA, Toulouse,
(1985), 182 p.
154. Riou, C., «La sécheresse, diagnostics et
conséquences peut-on en limiter les effets ? », Centre INRA,
Bordeaux, (1993), 1-16.
155. Slafer, G.A. et Savin, R., «Post-an thesis green
area duration in a semi-dwarf and standard height wheat cultivars as affected
by sink strength», Aust. J. Agro, Res : 45, (1994), 1337-1346.
141
156. Monneveux, P., Méziani, F., Bamoun, A., Hamou, M.
et Brinis, L., «Essai de définition des caractères
d'adaptation du blé dur dans différentes zones agroclimatiques de
l'Algérie», In : Séminaire sur la Tolérance à
la sécheresse des céréales en zones
méditerranéennes. Diversité génétiques et
amélioration variétale, Montpellier, France, Ed. Ben Salem, M et
Monneveux, P., INRA, Les colloques, n°64, (15-17 décembre 1992),
299-303.
157. Grignac, P., «Relation between yield, yield
components and some morphological characters », Genetics and breeding of
durum wheat sump, Bari, (1973), 275-284.
158. Hsu, P. et Malton, P.D., «Relation ships between
yield and its components and structures above the flag rode in spring wheat
», Crop sci, 11, (1971), 190-193.
159. MC Neal, F.H., Qualset, G.O., Baldridje, D.E. et
Stewart, V.R., «selection for yield and yield components in wheat »,
Crop sci, n°18, (1978), 145-152.
160. Bouzerzour, H. et Monneveux, P., 1992. «Analyses
des facteurs de stabilité du rendement de l'orge dans les conditions des
hauts plateaux algériens», Les Colloques, n°64, (1992),
205-215.
161. Falconer, D., «Introduction to quantitative
genetics », Eds Longman group limited, New York, (1982), 318-355.
162. Roseille, A. A. et Hamblin, J., «Theoritical aspect
of selection for yield in stress and non stress environments », Crop Sci.,
n°21, (1981), 943-946.
163. Kang, MS. et Magari, R., «Stable: a basic program
for calculating stability and yield stability statistics», Agron. J,
n°87, (1995), 276-277.
164. Wright, A.J., «The analysis and prediction of some
two factor interactions in grass breeding », Journal Agri, Sci Camb 76,
(1971), 301-306.
165. Annicchiarico, P., «Joint regression vs. AMMI
analysis of genotype x environment interactions for cereals in Italy»,
Euphytica, n°94, (1997), 53-62.
166. Gauch, H.G. et Zobel, R.W., «Identifying
mega-environments and targeting genotypes», Crop Sci., n°37, (1996),
311-326.
167. Samonte, S.O.P.B., Wilson, L.T., McClung, A.M. and
Medley, J.C., «Targeting cultivars onto rice growing environments using
AMMI and SREG GGE biplot analysis», Crop Sci, .n°45, (2005),
2414-2424.
168. Abdellaoui, Z. et Mariche, O., «Effet de la
fertilisation azotée sur l'expression de la qualité technologique
du blé dur : propriétés physico-chimiques »,
Céréaliculture, ITGC, n°50, (2002), 18-28.
142
169. Annou, B.S., «Aptitude technologiques de quelques
variétés de blé dur local: Interaction
amidon*protéine », Mémoire de magister de l'INA,
Algérie, (2002), 96p.
170. Gate, P., Jezequel, S., Castillon, P. et Laconde, J.P.,
«Bien remplir le grain et limiter le mitadinage », Brochure de
blé dur. ITCF, France, (1996), 36-43.
171. Braun, P.H., «Bons résultats pour l'apport
tardif d'azote », Perspective agricole, Ed. ITCF, France, n°188,
(1994), 30-34.
172. Matveef, M., «Le mitadinage des blés durs,
son évaluation et son influence sur le rendement et la valeur
semoulière », Bull de l'EFM, n°213, (1963), 133-138.
173. Bourdet, A., «L'identification des
variétés de blé par électrophorèse »,
In : symposium INRA, Electrophorèse, Paris, (24-25 février
1976).
174. Dexter, J.E. et Matsuo, R.R., «Effect of starch on
pasta dough and rheology and spaghetti cooking quality », Cereal chem.,
n°59, (1979), 190-195.
175. Abecassis, et Feillet, P., «Pureté des semoules
de blé dur, taux de cendre et règlementation », Ind,
céréales, n°31, (1985), 13-18.
176. Sadli, F., «La qualité technologique et
biochimique des blés cultivés en Algérie»,
Céréaliculture, n°26, (Mai 1993), 9-15.
177. Michel., «Comportement rhéologique du gluten,
influence de la composition en prolamines et de leur état
d'association», Thèse de doctorat d'université UFR de
science, Nantes, (1994).
178. Djema, I., «Influence du taux d'extraction et de
granulométrie de la semoule sur la qualité technologique du
couscous du blé dur», Thèse de magister, INA,
(Algérie), (2000).
179. Corbellini, M., Mazza, L., Ciaffi, M., Lafianda, D. et
Borghi, B., «Effet of heat shock during grain fillingon protein
composition and technological quality for wheats», In: wheat, prospects
for global improvement, proc. Of the International wheat conference Ankara,
Turkey, (10-14 June 1996), kluwer Academic publisher, Dordrechtt, Netherlands,
(1998), 213-220.
180. Luo, C., Branland, W.B., Griffin, W.B. et MC Neil, D.L.,
«The effect of nitrogen and sulfur fertilization and their interaction
with genotype on wheat glutenins and quality parameters», J. cereal sci,
n°31, (2000), 185-194.
181. Matveef, M., «Influence du gluten des blés
durs sur la valeur des pâtes alimentaires», Bull. Anc, Elèves
Ed. Fr meunerie, 213, (1966), 133-138.
182. Houliaro Poulos, S., Abecassis, J. et Autran, J.C.,
«Produits de mouture de blé dur : coloration et
caractéristiques culinaires», Ind. Céréales,
n°12, (1981), 3-13.
143
183. Bouzerzour, H., Benmahammed, A., Benkharbache, N. et
Hassous, K.L., «Contribution des nouvelles obtention à
l'amélioration et à la stabilité du rendement d'orge en
zone semi-aride d'altitude», Revue de la Recherche Agronomique de l'INRA,
Algérie, (2001b).
184. Malhotra, J.M., Everson, EH. et Hamblin, J.,
«Classification of environnements and genotypes in chickpeas»,
Euphtica, n°50, (1991), 161-168.
185. Gauch, H. G. «Statistical analysis of regional
yield trials: AMMI analysis of factorial designs», Elsevier Health
Sciences, Amsterdam, Netherlands, (1992).
186. Nachit, M.M., Nachit, G. et Ketata, H., «Analyse
des effets additifs et multiplicatifs des interactions
génotype*environnement chez le blé dur dans le bassin
méditerranéen», », In : Séminaire sur la
Tolérance à la sécheresse des céréales en
zones méditerranéennes. Diversité génétiques
et amélioration variétale, Montpellier, France, Ed. Ben Salem, M
et Monneveux, P., INRA, Les colloques, n°64, (15-17 décembre 1992),
159-169.
187. Peltonen-Sanino, P., Moore, K. et Pehu, E.,
«Phenotypic stability of oats measured with different stability
analyses», J Agri Sci 121, (1993), 13-19.
188. Ozgen, M., «Yield stability of winter wheat
(Triticum sp) cultivars and lines», J Agri Crop Sci 166, (1991),
318-325.
189. Pfeiffer, WH., «Drought tolerance in bread wheat.
Analysis of yield improvement over years in Cimmyt germplasm», In : Proc
of the Ind. Conf. on wheat production constraints in Tropical Environments.
Klatt eds, Mexico, D.F, Cimmyt, (1993), 274-284.
190. Fellah, A., Benmahammed, A., Djekoun, A. et Bouzerzour,
H., «Sélection pour améliorer la tolérance aux stress
abiotiques chez le blé dur (Triticum durum Desf.) », Actes
de l'IAV, Hassen II, Maroc, (2001).
144
APPENDICE A
LISTE DES SYMBOLES ET DES ABREVIATIONS
Kg/hab/an : Kilogramme par Habitant et par An.
t/ha : Tonnes par Hectare.
GxE : Interaction génotypes x environnement GxE.
AMMI : Effet principal additif et interaction multiplicative.
CIC : Conseil International des céréales.
Mha : millions d'hectares.
UE : Union Européen.
Mt : millions de tonnes.
PNDA : Plan National de Développement Agricole.
INRAA : Institut National de la Recherche Agronomique
d'Algérie.
ITGC : Institut Technique des Grandes Cultures.
CIMMYT : Centre International pour l'amélioration du
blé et du maïs.
ICARDA : Centre International pour la recherche agronomique en
zones
sèches.
QTL : Quantitative Traits Loci.
LEA : Les protéines Late Embryogenesis Abondant.
ONM : Office National de Météorologie.
T : Température.
P : Précipitation.
2.4D : Acide 2.4 dichloro-phénoxyacétique.
H : Humidité.
PVG : Durée de la Phase Végétative.
TLL : Capacité de Tallage herbacé.
HT : Hauteur des chaumes.
LE : Longueur de l'épi.
NE : Nombre d'épis.
NGE : Nombre de grains par épi.
PMG : Poids de Mille Grains.
145
RDT : Rendement en grains.
MTD : Taux de Mitadinage.
EXT : Taux d'Extraction.
GH : Gluten Humide.
GS : Gluten Sec.
Mn : Minute.
Ddl : degré de liberté.
OSM : Oued Smar.
KHR : Khroub
SET : Sétif.
TRT : Tiaret.
Max : Valeur Maximale.
Min : Valeur Minimale.
Ppds : Plus petite différence significative.
C V : Coefficient de variation.
E.T : Ecart type.
SCE : Somme Carré des Ecarts.
CME : Carré Moyen des Ecarts.
L : Localité.
IPCA : Première Composante Principale de
l'Interaction.
MG : Moyenne Générale.
B : Coefficient de régression.
MSGxL : Contribution au carré moyen de
l'interaction.
MSDEV. : Carré moyen de la déviation de la
régression.
a : Ordonnée à l'origine ( t/ha).
I : Répétitivité du rendement.
146
APPENDICE B
PRESENTATION DES DONNEES DES DIFFERENTES ZONES
D'ETUDE
Tableau 1 : Pluviométrie moyenne mensuelle
enregistrée durant la compagne 2006/2007 des quatre sites
d'étude.
|
Mois
|
Oued S
|
Khroub
|
Sétif
|
Tiaret
|
|
S
|
26
|
26,2
|
52
|
6
|
|
O
|
14,5
|
10
|
1
|
6
|
|
N
|
21
|
19,1
|
9,1
|
3,2
|
|
D
|
218,5
|
118,8
|
45
|
58,2
|
|
J
|
13,6
|
14,2
|
10,2
|
19,5
|
|
F
|
67,7
|
28,9
|
48,5
|
87
|
|
M
|
175,4
|
117,8
|
101,8
|
62
|
|
A
|
70,6
|
66,2
|
88,6
|
100
|
|
M
|
16
|
26,1
|
28,2
|
14,5
|
|
J
|
5
|
13,6
|
8
|
0,6
|
|
Total
|
628.3
|
440.9
|
392.4
|
341.8
|
Source : ONM (2008).
Tableau 2 : Température moyenne mensuelle
enregistrée durant la compagne 2006/2007 des quatre sites
d'étude.
|
Mois
|
Oued S
|
Khroub
|
Sétif
|
Tiaret
|
|
S
|
23,30
|
21,50
|
22,13
|
19,96
|
|
O
|
21,40
|
19,70
|
19,70
|
20,12
|
|
N
|
17,50
|
12,90
|
12,14
|
13,53
|
|
D
|
12,30
|
8,70
|
7,85
|
8,06
|
|
J
|
11,00
|
8,70
|
8,35
|
8,10
|
|
F
|
13,50
|
9,70
|
8,15
|
9,40
|
|
M
|
12,60
|
9,30
|
8,05
|
5,52
|
|
A
|
15,80
|
13,60
|
12,40
|
12,35
|
|
M
|
19,30
|
16,90
|
16,80
|
16,66
|
|
J
|
22,70
|
23,40
|
24,10
|
22,40
|
Source : ONM (2008).
147
Tableau 3 : Pluviométrie mensuelle enregistrée
à Oued Smar durant la compagne 2006/2007 en comparaison avec celles
enregistrées durant la période 89/04.
|
Mois
|
Pluviométrie (mm)
|
Total
mensuel
06/07
|
Moyennes
mensuelles
89/04
|
Ecarts de
précipitation
|
|
décade1
|
Décade2
|
Décade3
|
|
S
|
00
|
26
|
00
|
26
|
30.6
|
-4.6
|
|
O
|
00
|
14.5
|
00
|
14.5
|
57.7
|
-43.2
|
|
N
|
17
|
00
|
04
|
21
|
81.4
|
-60.4
|
|
D
|
33
|
71.5
|
113.8
|
218.5
|
74.1
|
+144.2
|
|
J
|
00
|
00
|
13.6
|
13.6
|
96.9
|
-83.3
|
|
F
|
14.2
|
6.5
|
47
|
67.7
|
74.5
|
-6.8
|
|
M
|
59.2
|
59.2
|
57
|
175.4
|
50.3
|
+125.1
|
|
A
|
07
|
50.9
|
12.7
|
70.6
|
65.8
|
+4.8
|
|
M
|
06
|
00
|
10
|
16
|
49.4
|
-33.4
|
|
J
|
05
|
00
|
00
|
05
|
8.3
|
-3.3
|
|
Total
|
141.4
|
228.6
|
254.1
|
623.1
|
589
|
+39.1
|
Source : ONM (2008).
Tableau 4 : Données climatiques de la compagne
2006/2007 enregistrées à la station expérimentale agricole
du Khroub.
|
Mois
|
Pluvio
05/06
(mm)
|
Pluvio
06/07
(mm)
|
Pluvio
(25 ans)
|
Nbre J
gelées
|
Nbre J
sirocco
|
Autres
|
|
S
|
14.4
|
26.2
|
37.5
|
|
|
|
|
O
|
2.1
|
10.0
|
38.6
|
|
|
|
|
N
|
18.6
|
19.1
|
44.6
|
4
|
|
|
|
D
|
63.1
|
118.8
|
73.2
|
3
|
|
|
|
J
|
78.9
|
14.2
|
62.8
|
7
|
|
|
|
F
|
50.3
|
28.9
|
53.8
|
5
|
|
|
|
M
|
33.4
|
117.8
|
56.2
|
2
|
|
|
|
A
|
14.7
|
66.2
|
59.0
|
1
|
|
|
|
M
|
83.8
|
26.1
|
42.3
|
|
5
|
|
|
J
|
3.6
|
13.6
|
19.3
|
|
15
|
|
|
Total
|
362.9
|
440.9
|
486.5
|
22
|
20
|
|
Source : ITGC El Khroub (2007).
148
Tableau 5 : Données climatiques de la compagne
2006/2007 enregistrées à la station expérimentale agricole
de Sétif.
|
Mois
|
Pluvio 06/07
|
My 25 camp
|
Ecarts de P
|
Gelées
|
Sirocco
|
|
S
|
52
|
39.84
|
+12.1
|
|
|
|
O
|
1.0
|
23.66
|
-22.6
|
|
|
|
N
|
9.1
|
36.77
|
-27.67
|
|
|
|
D
|
45
|
48.64
|
-3.64
|
10
|
|
|
J
|
10.2
|
44.3
|
-34.17
|
17
|
|
|
F
|
26.8
|
34.63
|
-7.85
|
4
|
|
|
M
|
101.8
|
32.68
|
+69.1
|
6
|
|
|
A
|
88.6
|
38.04
|
+50.5
|
1
|
|
|
M
|
28.20
|
48.31
|
-20.11
|
|
|
|
J
|
30.0
|
21.71
|
+8.29
|
|
9
|
|
Total
|
392.72
|
398.65
|
-5.93
|
|
|
Source : ITGC Sétif (2007).
Tableau 6 : Données climatiques de la compagne
2006/2007 enregistrées à la station expérimentale agricole
de Sabaine (Tiaret).
|
Mois
|
Pluvio 06/07
|
My 5 camp
|
Ecarts de P
|
Gelées
|
Sirocco
|
|
S
|
6.0
|
21.62
|
-16.6
|
|
|
|
O
|
6.0
|
36.88
|
-30.88
|
|
|
|
N
|
3.2
|
49.4
|
-46.2
|
1
|
|
|
D
|
58.2
|
46.42
|
+11.78
|
3
|
|
|
J
|
19.5
|
41.96
|
-22.46
|
18
|
|
|
F
|
87
|
46.6
|
+40.4
|
|
|
|
M
|
62
|
26.72
|
+35.28
|
5
|
|
|
A
|
100
|
32.04
|
+67.96
|
|
|
|
M
|
14.5
|
37.54
|
-23.04
|
|
1
|
|
J
|
0.6
|
14.5
|
-13.9
|
|
3
|
|
Total
|
351
|
353.68
|
-2.68
|
|
|
Source : ITGC Tiaret (2007).
149
APPENDICE C
DONNEES RELATIVES AUX VARIABLES MESUREES
150
Tableau 1 : Valeurs moyennes des variables mesurées (site
d'Oued Smar)
|
Géno
|
EPS
|
TALL
|
PHT
|
LE
|
NE
|
NGE
|
PMG
|
RDT
|
|
1
|
100,00
|
567,50
|
82,750
|
8,125
|
368,75
|
50,25
|
39,0750
|
41,2450
|
|
2
|
100,50
|
627,50
|
75,625
|
7,875
|
410,00
|
43,25
|
37,8875
|
35,8300
|
|
3
|
99,00
|
682,50
|
81,875
|
8,125
|
395,00
|
48,50
|
32,4775
|
40,8300
|
|
4
|
99,75
|
710,00
|
82,375
|
6,625
|
397,50
|
50,00
|
26,6550
|
35,8300
|
|
5
|
100,00
|
673,75
|
76,375
|
7,875
|
355,00
|
45,75
|
32,8425
|
29,9975
|
|
6
|
101,00
|
710,00
|
86,125
|
8,375
|
428,75
|
43,00
|
34,5850
|
33,3325
|
|
7
|
99,50
|
663,75
|
81,375
|
8,125
|
425,00
|
43,50
|
47,0625
|
50,8300
|
|
8
|
98,00
|
648,75
|
82,250
|
7,250
|
381,25
|
48,75
|
30,6850
|
29,1625
|
|
9
|
99,50
|
678,75
|
79,625
|
7,125
|
405,00
|
46,25
|
31,3050
|
35,4125
|
|
10
|
98,00
|
572,50
|
76,500
|
7,750
|
311,25
|
48,25
|
37,5325
|
34,1625
|
|
11
|
99,50
|
642,50
|
79,875
|
7,875
|
363,75
|
47,25
|
30,5825
|
30,8300
|
|
12
|
98,00
|
682,50
|
71,250
|
8,125
|
497,50
|
48,25
|
40,7300
|
51,2475
|
|
13
|
98,00
|
576,25
|
76,500
|
8,000
|
318,75
|
43,50
|
46,1150
|
46,6625
|
|
14
|
97,00
|
730,00
|
78,875
|
8,000
|
373,75
|
40,00
|
35,1700
|
27,0800
|
|
15
|
98,00
|
641,25
|
81,500
|
8,125
|
328,75
|
44,25
|
34,8075
|
36,6650
|
|
16
|
99,00
|
535,00
|
76,500
|
7,500
|
372,50
|
48,00
|
25,5675
|
19,1625
|
|
17
|
102,00
|
648,75
|
79,500
|
7,575
|
432,25
|
48,00
|
26,7900
|
22,0775
|
|
18
|
101,00
|
742,50
|
85,500
|
7,250
|
446,25
|
53,00
|
29,2350
|
29,9975
|
|
19
|
102,00
|
735,00
|
77,875
|
7,125
|
455,00
|
48,75
|
24,8150
|
27,4950
|
|
20
|
102,75
|
478,75
|
84,000
|
7,500
|
373,75
|
46,25
|
34,0000
|
26,2475
|
|
21
|
103,00
|
740,00
|
75,375
|
6,750
|
377,50
|
44,50
|
35,2625
|
32,9125
|
|
22
|
104,00
|
712,50
|
74,875
|
7,250
|
390,00
|
45,75
|
26,3050
|
17,0800
|
|
23
|
100,75
|
665,00
|
79,125
|
7,625
|
421,25
|
33,25
|
31,0525
|
23,2475
|
|
24
|
107,00
|
652,50
|
86,000
|
7,375
|
386,25
|
41,50
|
37,0025
|
29,5800
|
|
25
|
108,25
|
810,00
|
90,125
|
8,875
|
338,75
|
48,00
|
33,9300
|
24,2475
|
|
26
|
104,25
|
666,25
|
75,125
|
7,875
|
432,50
|
38,50
|
30,0600
|
20,8300
|
|
27
|
102,75
|
802,50
|
85,625
|
8,000
|
428,75
|
48,25
|
32,6450
|
31,2450
|
|
28
|
102,00
|
580,00
|
78,375
|
8,000
|
267,50
|
54,00
|
32,2750
|
19,1650
|
|
29
|
102,00
|
610,00
|
81,000
|
7,750
|
421,25
|
45,50
|
30,4725
|
22,4975
|
|
30
|
98,00
|
486,25
|
81,875
|
7,500
|
277,50
|
49,00
|
33,5325
|
19,9950
|
|
M.g
|
100,82
|
655,75
|
80,125
|
7,711
|
386,03
|
46,10
|
33,3486
|
30,6633
|
|
M.t
|
101,40
|
635,25
|
81,700
|
8,025
|
341,50
|
46,50
|
34,6225
|
27,7805
|
|
C.V
|
0.7
|
19.5
|
3.8
|
6.5
|
22.7
|
9.9
|
11.7
|
20.9
|
|
E.T
|
0.7117
|
127.84
|
3.0183
|
0.5
|
87.626
|
4.5639
|
3.92
|
6.42
|
|
Ppds
|
1.00
|
179.653
|
4.2416
|
0.7034
|
123.142
|
6.41375
|
5.4960
|
9.0213
|
151
Tableau 2 : Valeurs moyennes des variables mesurées (site
ELKhroub)
|
Géno
|
EPS
|
Tall
|
PHT
|
LE
|
NE
|
NGE
|
PMG
|
RDT
|
|
1
|
116,75
|
857,50
|
90,750
|
7,500
|
505,00
|
37,00
|
41,7775
|
55,8225
|
|
2
|
119,00
|
640,00
|
88,000
|
7,750
|
425,00
|
43,75
|
48,0100
|
52,8850
|
|
3
|
114,75
|
968,75
|
87,000
|
7,000
|
516,25
|
40,50
|
43,1675
|
65,5850
|
|
4
|
119,00
|
905,00
|
92,000
|
6,000
|
485,00
|
41,50
|
37,3000
|
51,9500
|
|
5
|
104,75
|
921,25
|
86,000
|
6,250
|
513,75
|
39,75
|
39,0425
|
54,3000
|
|
6
|
116,00
|
805,00
|
97,750
|
7,500
|
431,25
|
40,00
|
44,5550
|
63,1075
|
|
7
|
117,75
|
947,50
|
94,000
|
7,500
|
488,75
|
41,25
|
45,8850
|
52,9325
|
|
8
|
116,50
|
852,50
|
91,250
|
6,500
|
420,00
|
46,00
|
42,1950
|
59,4650
|
|
9
|
114,75
|
902,50
|
86,750
|
7,750
|
432,50
|
37,50
|
41,2575
|
49,7500
|
|
10
|
114,75
|
862,50
|
87,500
|
7,250
|
417,50
|
44,00
|
45,0750
|
53,2925
|
|
11
|
121,00
|
973,75
|
86,500
|
7,750
|
456,25
|
49,50
|
35,0625
|
54,1250
|
|
12
|
116,75
|
850,00
|
83,000
|
6,500
|
536,25
|
37,50
|
44,7575
|
58,3350
|
|
13
|
116,75
|
716,25
|
83,250
|
6,250
|
442,50
|
43,00
|
46,1625
|
51,9850
|
|
14
|
118,50
|
1037,50
|
90,750
|
7,500
|
510,00
|
30,75
|
42,1125
|
54,1650
|
|
15
|
117,25
|
870,00
|
86,750
|
7,500
|
393,75
|
39,25
|
45,5025
|
49,4000
|
|
16
|
115,00
|
797,50
|
85,250
|
7,000
|
497,50
|
50,50
|
34,5925
|
57,3425
|
|
17
|
120,00
|
772,50
|
86,750
|
6,750
|
430,00
|
51,75
|
37,3875
|
56,5400
|
|
18
|
116,75
|
867,50
|
91,500
|
6,750
|
491,25
|
38,75
|
46,2975
|
52,7000
|
|
19
|
118,00
|
797,50
|
85,250
|
7,750
|
465,00
|
49,50
|
40,6825
|
56,1675
|
|
20
|
119,00
|
892,50
|
93,000
|
7,250
|
458,75
|
44,75
|
47,0250
|
44,7775
|
|
21
|
120,00
|
877,50
|
88,250
|
6,000
|
457,50
|
43,75
|
46,5500
|
51,2500
|
|
22
|
119,00
|
863,75
|
82,500
|
8,000
|
463,75
|
51,25
|
38,2600
|
53,9175
|
|
23
|
118,25
|
728,75
|
91,250
|
6,750
|
436,25
|
42,25
|
44,9000
|
54,5075
|
|
24
|
118,50
|
1005,00
|
93,500
|
7,000
|
440,00
|
38,00
|
46,8550
|
56,0400
|
|
25
|
118,00
|
852,50
|
90,750
|
7,750
|
371,25
|
37,00
|
43,0025
|
57,9600
|
|
26
|
117,75
|
667,50
|
83,250
|
8,750
|
377,50
|
45,00
|
38,7125
|
49,3250
|
|
27
|
119,50
|
1136,25
|
92,500
|
6,250
|
488,75
|
39,50
|
47,9850
|
57,5000
|
|
28
|
120,75
|
815,00
|
80,750
|
7,500
|
353,75
|
48,00
|
45,1225
|
48,1550
|
|
29
|
120,50
|
800,00
|
94,500
|
6,250
|
457,50
|
41,25
|
47,4350
|
59,0850
|
|
30
|
118,25
|
830,00
|
86,500
|
6,500
|
377,50
|
43,00
|
41,8125
|
53,3400
|
|
M.g
|
117,45
|
860,46
|
88,558
|
7,092
|
451,33
|
42,52
|
42,9494
|
54,5236
|
|
M.t
|
114,75
|
879,75
|
88,800
|
7,200
|
431,00
|
40,95
|
43,9295
|
51,9460
|
|
C.V
|
1.7
|
18.3
|
4.4
|
14.1
|
14.8
|
11.6
|
8.7
|
14.7
|
|
E. T
|
1.97
|
157.82
|
3.90
|
1.00
|
66.80
|
4.93
|
3.76
|
8.03
|
|
Ppds
|
2.7666
|
221.78
|
5.4825
|
1.4098
|
93.8693
|
6.9260
|
5.2790
|
11.2811
|
152
Tableau 3 : Valeurs moyennes des variables mesurées (site
de Sétif).
|
Géno
|
EPS
|
Tall
|
PHT
|
LE
|
NE
|
NGE
|
PMG
|
RDT
|
|
1
|
140,00
|
762,50
|
92,750
|
5,800
|
433,75
|
28,00
|
40,1250
|
39,6700
|
|
2
|
144,00
|
807,50
|
89,750
|
5,700
|
452,50
|
38,25
|
36,6350
|
50,5975
|
|
3
|
140,00
|
777,50
|
88,500
|
5,700
|
486,25
|
34,75
|
45,3600
|
57,8550
|
|
4
|
140,25
|
785,00
|
89,250
|
6,525
|
436,25
|
31,50
|
43,9925
|
47,9525
|
|
5
|
145,00
|
857,50
|
90,250
|
5,925
|
421,25
|
38,00
|
40,9525
|
54,1525
|
|
6
|
140,00
|
673,75
|
91,000
|
5,850
|
411,25
|
34,75
|
42,8750
|
38,8675
|
|
7
|
144,00
|
757,50
|
91,500
|
5,450
|
407,50
|
35,50
|
38,3650
|
42,6625
|
|
8
|
138,00
|
800,00
|
97,250
|
6,075
|
377,50
|
36,75
|
44,2475
|
51,1100
|
|
9
|
144,00
|
735,00
|
92,250
|
6,650
|
418,75
|
32,25
|
45,8375
|
48,4725
|
|
10
|
140,00
|
826,25
|
88,250
|
5,725
|
375,00
|
36,75
|
46,5475
|
49,9275
|
|
11
|
144,00
|
718,75
|
91,750
|
5,575
|
387,50
|
39,25
|
42,8200
|
52,1700
|
|
12
|
138,00
|
746,25
|
91,000
|
5,400
|
437,50
|
35,25
|
41,9850
|
54,0050
|
|
13
|
142,00
|
800,00
|
91,250
|
6,450
|
417,50
|
39,75
|
35,4375
|
47,2025
|
|
14
|
140,00
|
823,75
|
83,500
|
6,000
|
425,00
|
34,50
|
44,4900
|
52,5075
|
|
15
|
142,00
|
751,25
|
88,500
|
5,725
|
391,25
|
34,25
|
44,6775
|
52,7600
|
|
16
|
138,00
|
750,00
|
89,500
|
5,700
|
442,50
|
32,50
|
46,5850
|
48,5675
|
|
17
|
143,75
|
731,25
|
91,250
|
5,575
|
512,50
|
27,75
|
41,5800
|
42,9325
|
|
18
|
138,00
|
780,00
|
89,500
|
5,500
|
420,00
|
35,50
|
42,5375
|
53,7525
|
|
19
|
144,00
|
775,00
|
86,250
|
6,750
|
430,00
|
33,75
|
41,2425
|
50,3775
|
|
20
|
144,00
|
890,00
|
94,500
|
6,800
|
353,75
|
34,25
|
48,7775
|
47,3100
|
|
21
|
140,00
|
685,00
|
89,000
|
5,475
|
408,75
|
37,50
|
37,5575
|
45,6925
|
|
22
|
142,00
|
740,00
|
90,000
|
6,150
|
391,25
|
44,00
|
40,1625
|
53,9725
|
|
23
|
140,00
|
626,25
|
95,250
|
5,525
|
357,50
|
32,25
|
49,7475
|
46,9525
|
|
24
|
140,00
|
766,25
|
91,000
|
6,000
|
381,25
|
40,75
|
39,8750
|
47,9125
|
|
25
|
144,00
|
881,25
|
87,250
|
5,725
|
452,50
|
41,00
|
37,3575
|
51,6500
|
|
26
|
146,00
|
748,75
|
89,500
|
5,525
|
398,75
|
33,25
|
44,6250
|
49,9175
|
|
27
|
142,00
|
732,50
|
88,250
|
5,750
|
397,50
|
41,00
|
38,7100
|
52,1025
|
|
28
|
142,00
|
732,50
|
93,500
|
6,150
|
423,75
|
31,00
|
45,7150
|
50,9850
|
|
29
|
142,00
|
855,00
|
91,000
|
5,450
|
396,25
|
30,50
|
47,1200
|
50,6475
|
|
30
|
144,00
|
797,50
|
84,250
|
6,125
|
361,25
|
39,75
|
40,6475
|
48,2925
|
|
M.g
|
141,70
|
770,46
|
90,225
|
5,892
|
413,54
|
35,48
|
42,5529
|
49,3659
|
|
M.t
|
143,00
|
841,25
|
89,750
|
5,980
|
398,75
|
36,85
|
43,6625
|
51,1600
|
|
C.V%
|
0.7
|
14.7
|
3.7
|
8.8
|
15.1
|
11.3
|
5.5
|
6.4
|
|
E.T
|
1.00
|
113.55
|
3.30
|
0.52
|
62.304
|
3.1
|
2.35
|
3.17
|
|
Ppds
|
1.41
|
159.57
|
4.64
|
0.73
|
87.56
|
5.61
|
3.31
|
4.46
|
153
Tableau 4 : Valeurs moyennes des variables mesurées (site
de Tiaret) :
|
Géno
|
EPS
|
Tall
|
PHT
|
LE
|
NE
|
NGE
|
PMG
|
RDT
|
|
1
|
125,25
|
787,50
|
84,500
|
5,875
|
413,75
|
32,50
|
21,5000
|
10,9575
|
|
2
|
124,50
|
928,75
|
78,250
|
6,375
|
417,50
|
33,25
|
18,9000
|
8,3725
|
|
3
|
124,25
|
766,25
|
80,625
|
6,500
|
395,00
|
36,75
|
20,1000
|
11,1550
|
|
4
|
124,50
|
907,50
|
81,125
|
6,875
|
452,00
|
32,50
|
25,6500
|
10,6650
|
|
5
|
124,75
|
876,25
|
83,250
|
7,250
|
346,25
|
37,25
|
19,2000
|
10,8300
|
|
6
|
126,00
|
836,25
|
77,000
|
6,300
|
427,50
|
32,25
|
20,8000
|
9,2450
|
|
7
|
125,25
|
998,75
|
83,875
|
5,750
|
553,75
|
28,50
|
19,2500
|
7,9975
|
|
8
|
126,00
|
853,75
|
83,375
|
6,750
|
365,00
|
25,50
|
24,3000
|
7,2450
|
|
9
|
121,00
|
877,50
|
82,000
|
6,375
|
415,00
|
26,00
|
20,6500
|
9,3275
|
|
10
|
121,50
|
845,00
|
82,375
|
6,375
|
411,25
|
32,00
|
18,2000
|
7,9975
|
|
11
|
122,50
|
777,50
|
86,125
|
6,125
|
388,75
|
33,25
|
20,3000
|
11,4550
|
|
12
|
123,00
|
962,50
|
82,375
|
6,500
|
516,25
|
30,50
|
20,5000
|
11,1650
|
|
13
|
126,25
|
970,50
|
80,125
|
6,625
|
431,25
|
37,00
|
18,4500
|
7,7475
|
|
14
|
126,00
|
867,50
|
77,000
|
6,750
|
402,50
|
34,50
|
19,9000
|
9,5775
|
|
15
|
125,75
|
1012,50
|
76,875
|
6,175
|
375,00
|
30,00
|
18,9000
|
10,3300
|
|
16
|
121,25
|
873,75
|
87,125
|
6,625
|
431,25
|
32,50
|
22,2500
|
13,0775
|
|
17
|
122,50
|
927,50
|
84,500
|
6,625
|
441,25
|
26,25
|
21,9000
|
8,1625
|
|
18
|
125,50
|
883,75
|
84,250
|
6,625
|
436,25
|
33,75
|
20,1000
|
11,5375
|
|
19
|
130,00
|
997,50
|
79,125
|
6,000
|
463,75
|
31,50
|
19,9000
|
8,4125
|
|
20
|
124,25
|
778,75
|
84,125
|
6,375
|
407,50
|
35,25
|
22,0500
|
12,3725
|
|
21
|
123,75
|
885,00
|
81,625
|
6,875
|
470,00
|
33,50
|
17,5000
|
10,5800
|
|
22
|
122,00
|
902,50
|
82,625
|
6,375
|
455,00
|
33,25
|
18,3000
|
9,4125
|
|
23
|
123,25
|
890,00
|
81,125
|
6,000
|
431,25
|
31,50
|
18,6500
|
10,4975
|
|
24
|
126,00
|
978,75
|
84,750
|
6,375
|
420,00
|
33,25
|
20,6000
|
10,8300
|
|
25
|
125,75
|
956,25
|
85,250
|
6,625
|
412,50
|
39,50
|
17,1000
|
10,1225
|
|
26
|
124,25
|
1011,25
|
82,375
|
6,375
|
446,25
|
31,00
|
25,0000
|
9,1625
|
|
27
|
121,75
|
905,00
|
81,375
|
6,375
|
405,00
|
34,50
|
17,0000
|
8,7475
|
|
28
|
126,00
|
1001,25
|
82,375
|
6,000
|
425,00
|
28,75
|
21,9000
|
8,8725
|
|
29
|
128,25
|
940,00
|
80,625
|
6,125
|
332,50
|
31,25
|
21,9500
|
8,2050
|
|
30
|
124,50
|
925,00
|
80,250
|
6,750
|
412,50
|
36,75
|
20,1000
|
12,4975
|
|
M.g
|
124,52
|
904,14
|
82,013
|
6,424
|
423,36
|
32,48
|
20,3633
|
9,8819
|
|
M.t
|
124,40
|
893,75
|
82,375
|
6,560
|
390,50
|
34,80
|
19,0900
|
10,3305
|
|
C.V
|
1
|
15.00
|
5.1
|
12.9
|
17.5
|
11.7
|
19.1
|
24.7
|
|
E .T
|
1.28
|
135.75
|
4.21
|
0.83
|
74.07
|
3.8
|
3.89
|
2.44
|
|
Ppds
|
1.8
|
190.776
|
5.92
|
1.16
|
104.09
|
5.34
|
5.46
|
3.43
|
154
Tableau 5 : Valeurs moyennes de l'effet interaction
génotype x milieu du rendement en grains RDT (q/ha) par ordre
décroissant.
|
N° (GxE)
|
RDT
|
N° (GxE)
|
RDT
|
N° (GxE)
|
RDT
|
N° (GxE)
|
RDT
|
|
V3M2
|
65,5850
|
V11M3
|
52,1700
|
V7M3
|
42,6625
|
V16M4
|
13,0775
|
|
V6M2
|
63,1075
|
V27M3
|
52,1025
|
V1M1
|
41,2450
|
V30M4
|
12,4975
|
|
V8M2
|
59,4650
|
V13M2
|
51,9850
|
V3M1
|
40.8300
|
V20M4
|
12,3725
|
|
V29M2
|
59,0850
|
V4M2
|
51,9500
|
V1M3
|
39,6700
|
V18M4
|
11,5375
|
|
V12M2
|
58,3350
|
V25M3
|
51,6500
|
V6M3
|
38,8675
|
V11M4
|
11,4550
|
|
V25M2
|
57,9600
|
V21M2
|
51,2500
|
V15M1
|
36,6650
|
V12M4
|
11,1650
|
|
V3M3
|
57,8550
|
V12M1
|
51,2475
|
V2M1
|
35,8300
|
V3M4
|
11,1550
|
|
V27M2
|
57,5000
|
V8M3
|
51,1100
|
V4M1
|
35,8300
|
V1M4
|
10,9575
|
|
V16M2
|
57,3425
|
V28M3
|
50,9850
|
V9M1
|
35,4125
|
V5M4
|
10,8300
|
|
V17M2
|
56,5400
|
V7M1
|
50,8300
|
V10M1
|
34,1625
|
V24M4
|
10,8300
|
|
V19M2
|
56,1675
|
V29M3
|
50,6475
|
V6M1
|
33,3325
|
V4M4
|
10,6650
|
|
V24M2
|
56,0400
|
V2M3
|
50,5975
|
V21M1
|
32,9125
|
V21M4
|
10,5800
|
|
V1M2
|
55,8225
|
V19M3
|
50,3775
|
V27M1
|
31,2450
|
V23M4
|
10,4975
|
|
V23M2
|
54,5075
|
V10M3
|
49,9275
|
V11M1
|
30,8300
|
V15M4
|
10,3300
|
|
V5M2
|
54,3000
|
V26M3
|
49,9175
|
V5M1
|
29,9975
|
V25M4
|
10,1225
|
|
V14M2
|
54,1650
|
V9M2
|
49,7500
|
V18M1
|
29,9975
|
V14M4
|
9,5775
|
|
V5M3
|
54,1525
|
V15M2
|
49,4000
|
V24M1
|
29,5800
|
V22M4
|
9,4125
|
|
V11M2
|
54,1250
|
V26M2
|
49,3250
|
V8M1
|
29,1625
|
V9M4
|
9,3275
|
|
V12M3
|
54,0050
|
V16M3
|
48,5675
|
V19M1
|
27,4950
|
V6M4
|
9,2450
|
|
V22M3
|
53,9725
|
V9M3
|
48,4725
|
V14M1
|
27,0800
|
V26M4
|
9,1625
|
|
V22M2
|
53,9175
|
V30M3
|
48,2925
|
V20M1
|
26,2475
|
V28M4
|
8,8725
|
|
V18M3
|
53,7525
|
V28M2
|
48,1550
|
V25M1
|
24,2475
|
V27M4
|
8,7475
|
|
V30M2
|
53,3400
|
V4M3
|
47,9525
|
V23M1
|
23,2475
|
V19M4
|
8,4125
|
|
V10M2
|
53,2925
|
V24M3
|
47,9125
|
V29M1
|
22,4975
|
V2M4
|
8,3725
|
|
V7M2
|
52,9325
|
V20M3
|
47,3100
|
V17M1
|
22,0775
|
V29M4
|
8,2050
|
|
V2M2
|
52,8850
|
V13M3
|
47,2025
|
V26M1
|
20,8300
|
V17M4
|
8,1625
|
|
V15M3
|
52,7600
|
V23M3
|
46,9525
|
V30M1
|
19,9950
|
V7M4
|
7,9975
|
|
V18M2
|
52,7000
|
V13M1
|
46,6625
|
V28M1
|
19,1650
|
V10M4
|
7,9975
|
|
V14M3
|
52,5075
|
V21M3
|
45,6925
|
V16M1
|
19,1625
|
V13M4
|
7,7475
|
|
V11M3
|
52,1700
|
V20M2
|
44,7775
|
V22M1
|
17,0800
|
V8M4
|
7,2450
|
M1: Oued Smar, M2: El Khroub, M3: Sétif, M4: Tiaret
155
Tableau 6 : Valeurs moyennes de l'effet génotype
des variables relatives à la qualité (MTD et EXT sur les
quatre sites et GH et GS sur les trois sites, OSM, KHB et SET).
|
N° Géno
|
MTD (%)
|
EXT (%)
|
GH (%)
|
GS (%)
|
|
1
|
3,949
|
50,083
|
32,158
|
9,7425
|
|
2
|
5,678
|
55,495
|
28,688
|
9,9229
|
|
3
|
5,027
|
58,721
|
29,816
|
9,2325
|
|
4
|
4,427
|
53,387
|
31,464
|
9,595
|
|
5
|
6,944
|
55,697
|
29,307
|
8,7375
|
|
6
|
5,237
|
55,186
|
31,152
|
9,5242
|
|
7
|
6,532
|
55,798
|
31,754
|
9,4325
|
|
8
|
11,174
|
56,286
|
30,193
|
8,92
|
|
9
|
7,529
|
57,222
|
25,929
|
11,0283
|
|
10
|
12,809
|
57,012
|
60,909
|
5,9283
|
|
11
|
5,399
|
55,351
|
32,044
|
9,715
|
|
12
|
7,199
|
50,497
|
29,075
|
9,0708
|
|
13
|
4,156
|
55,592
|
32,063
|
9,7233
|
|
14
|
11,597
|
56,161
|
34,126
|
12,4708
|
|
15
|
9,016
|
55,886
|
30,162
|
8,8992
|
|
16
|
7,957
|
51,082
|
27,879
|
8,5525
|
|
17
|
7,102
|
53,421
|
32,442
|
9,8283
|
|
18
|
10,845
|
54,429
|
33,97
|
8,2342
|
|
19
|
3,496
|
47,104
|
28,488
|
8,5058
|
|
20
|
10,157
|
55,216
|
29,999
|
8,6
|
|
21
|
14,38
|
56,884
|
28,257
|
8,3625
|
|
22
|
15,012
|
54,953
|
29,785
|
8,8267
|
|
23
|
13,267
|
56,114
|
21,232
|
6,4058
|
|
24
|
12,499
|
55,882
|
28,16
|
8,4308
|
|
25
|
6,488
|
57,139
|
28,34
|
8,285
|
|
26
|
13,072
|
54,079
|
31,499
|
9,2108
|
|
27
|
15,295
|
56,735
|
29,116
|
8,4075
|
|
28
|
13,047
|
55,829
|
21,118
|
6,4242
|
|
29
|
11,165
|
55,322
|
30,265
|
8,7258
|
|
30
|
14,809
|
50,457
|
22,739
|
6,7292
|
156
Tableau 7 : Valeurs moyennes de l'effet interaction
génotype x milieu du taux de mitadinage MTD (%) par ordre
décroissant.
|
N° (G*E)
|
MTD
|
N° (G*E)
|
MTD
|
N° (G*E)
|
MTD
|
N° (G*E)
|
MTD
|
|
V27M3
|
56,132
|
V23M2
|
10,000
|
V7M2
|
2,915
|
V14M2
|
0,365
|
|
V22M3
|
52,242
|
V20M2
|
8,715
|
V1M1
|
2,837
|
V19M4
|
0,340
|
|
V30M3
|
51,622
|
V29M2
|
8,170
|
V13M2
|
2,692
|
V29M4
|
0,273
|
|
V26M3
|
49,262
|
V11M2
|
7,720
|
V18M1
|
2,587
|
V28M4
|
0,220
|
|
V21M3
|
48,537
|
V8M2
|
7,257
|
V26M1
|
2,522
|
V16M4
|
0,213
|
|
V28M3
|
45,292
|
V17M2
|
7,200
|
V16M2
|
2,327
|
V7M4
|
0,168
|
|
V14M3
|
43,845
|
V12M1
|
6,982
|
V22M1
|
2,302
|
V8M4
|
0,163
|
|
V24M3
|
43,832
|
V17M1
|
6,705
|
V25M1
|
2,195
|
V15M4
|
0,163
|
|
V10M3
|
38,825
|
V30M2
|
6,572
|
V14M1
|
2,180
|
V10M4
|
0,110
|
|
V23M3
|
38,355
|
V10M2
|
6,510
|
V19M1
|
2,180
|
V11M4
|
0,108
|
|
V18M3
|
37,085
|
V12M2
|
6,300
|
V4M2
|
2,170
|
V13M4
|
0,050
|
|
V29M3
|
35,512
|
V10M1
|
5,780
|
V24M1
|
1,975
|
V1M4
|
-
|
|
V8M3
|
33,147
|
V21M1
|
5,665
|
V6M1
|
1,827
|
V2M4
|
-
|
|
V15M3
|
32,570
|
V9M2
|
5,505
|
V15M2
|
1,817
|
V3M4
|
-
|
|
V20M3
|
30,500
|
V22M2
|
5,505
|
V1M2
|
1,685
|
V4M4
|
-
|
|
V16M3
|
28,385
|
V28M2
|
5,307
|
V27M1
|
1,670
|
V5M4
|
-
|
|
V9M3
|
23,792
|
V4M3
|
5,222
|
V7M1
|
1,530
|
V6M4
|
-
|
|
V7M3
|
21,517
|
V3M1
|
4,412
|
V15M1
|
1,512
|
V9M4
|
-
|
|
V5M3
|
20,225
|
V23M1
|
4,275
|
V20M1
|
1,412
|
V12M4
|
-
|
|
V25M3
|
20,057
|
V24M2
|
4,187
|
V28M1
|
1,367
|
V14M4
|
-
|
|
V6M3
|
18,710
|
V8M1
|
4,127
|
V19M2
|
1,337
|
V17M4
|
-
|
|
V2M3
|
16,520
|
V5M2
|
4,012
|
V30M1
|
1,042
|
V18M4
|
-
|
|
V12M3
|
15,512
|
V18M2
|
3,707
|
V16M1
|
0,902
|
V20M4
|
-
|
|
V17M3
|
14,502
|
V25M2
|
3,700
|
V13M1
|
0,875
|
V21M4
|
-
|
|
V11M3
|
13,012
|
V5M1
|
3,540
|
V9M1
|
0,820
|
V22M4
|
-
|
|
V13M3
|
13,005
|
V27M2
|
3,377
|
V11M1
|
0,757
|
V24M4
|
-
|
|
V3M3
|
12,517
|
V21M2
|
3,317
|
V29M1
|
0,705
|
V25M4
|
-
|
|
V1M3
|
11,275
|
V3M2
|
3,180
|
V26M2
|
0,502
|
V26M4
|
-
|
|
V4M1
|
10,315
|
V2M2
|
3,172
|
V23M4
|
0,438
|
V27M4
|
-
|
|
V19M3
|
10,127
|
V2M1
|
3,020
|
V6M2
|
0,412
|
V30M4
|
-
|
M1: Oued Smar, M2: El Khroub, M3: Sétif, M4: Tiaret
157
Tableau 8 : Valeurs moyennes de l'effet interaction
génotype x milieu du taux d'extraction EXT (%) par ordre
décroissant.
|
N° (G*E)
|
EXT
|
N° (G*E)
|
EXT
|
N° (G*E)
|
EXT
|
N° (G*E)
|
EXT
|
|
V2M3
|
66,230
|
V3M2
|
59,667
|
V20M2
|
55,515
|
V28M4
|
52,250
|
|
V27M3
|
64,645
|
V17M2
|
59,430
|
V19M2
|
55,392
|
V23M1
|
52,225
|
|
V3M4
|
64,490
|
V23M2
|
59,395
|
V28M2
|
55,370
|
V7M4
|
52,170
|
|
V20M3
|
64,480
|
V14M2
|
58,450
|
V21M4
|
55,360
|
V4M4
|
52,055
|
|
V28M3
|
64,250
|
V10M2
|
57,827
|
V24M4
|
55,320
|
V28M1
|
51,445
|
|
V17M3
|
63,740
|
V29M2
|
57,797
|
V27M4
|
55,250
|
V26M4
|
51,000
|
|
V25M3
|
63,515
|
V8M4
|
57,760
|
V5M1
|
55,145
|
V6M1
|
50,670
|
|
V9M3
|
63,100
|
V7M2
|
57,605
|
V9M2
|
55,130
|
V23M4
|
50,330
|
|
V24M3
|
63,063
|
V14M1
|
57,503
|
V11M2
|
55,112
|
V27M1
|
50,050
|
|
V18M3
|
62,895
|
V9M4
|
57,330
|
V12M4
|
55,038
|
V4M2
|
49,860
|
|
V16M3
|
62,740
|
V17M4
|
57,300
|
V25M1
|
55,033
|
V3M1
|
49,813
|
|
V8M3
|
62,620
|
V5M2
|
57,117
|
V13M4
|
54,660
|
V5M4
|
49,740
|
|
V23M3
|
62,505
|
V27M2
|
56,995
|
V22M2
|
54,172
|
V4M1
|
49,610
|
|
V26M3
|
62,445
|
V19M4
|
56,840
|
V20M4
|
54,047
|
V13M1
|
49,593
|
|
V29M3
|
62,220
|
V2M2
|
56,810
|
V15M1
|
53,885
|
V1M2
|
49,418
|
|
V10M3
|
62,210
|
V15M2
|
56,662
|
V21M1
|
53,818
|
V11M1
|
49,333
|
|
V6M3
|
62,153
|
V1M4
|
56,660
|
V1M1
|
53,795
|
V24M1
|
49,015
|
|
V4M3
|
62,025
|
V21M2
|
56,500
|
V25M4
|
53,570
|
V18M1
|
48,780
|
|
V13M3
|
62,000
|
V25M2
|
56,440
|
V9M1
|
53,328
|
V22M1
|
48,630
|
|
V21M3
|
61,860
|
V26M2
|
56,362
|
V29M4
|
53,230
|
V8M1
|
48,608
|
|
V30M3
|
61,740
|
V19M1
|
56,300
|
V18M4
|
53,190
|
V29M1
|
48,043
|
|
V22M3
|
61,010
|
V8M2
|
56,157
|
V7M1
|
53,075
|
V14M4
|
47,942
|
|
V3M3
|
60,915
|
V24M2
|
56,133
|
V18M2
|
52,850
|
V20M1
|
46,820
|
|
V11M3
|
60,905
|
V13M2
|
56,117
|
V12M2
|
52,825
|
V26M1
|
46,510
|
|
V12M3
|
60,795
|
V30M2
|
56,080
|
V15M4
|
52,680
|
V2M1
|
46,410
|
|
V5M3
|
60,785
|
V11M4
|
56,055
|
V2M4
|
52,530
|
V1M3
|
40,460
|
|
V14M3
|
60,750
|
V22M4
|
56,000
|
V10M1
|
52,430
|
V12M1
|
33,330
|
|
V7M3
|
60,343
|
V16M4
|
55,943
|
V16M2
|
52,410
|
V16M1
|
33,238
|
|
V15M3
|
60,315
|
V6M4
|
55,660
|
V30M4
|
52,330
|
V17M1
|
33,213
|
|
V19M3
|
59,885
|
V10M4
|
55,580
|
V6M2
|
52,260
|
V30M1
|
31,680
|
M1: Oued Smar, M2: El Khroub, M3: Sétif, M4: Tiaret
158
Tableau 9 : Valeurs moyennes de l'effet interaction
génotype x milieu du teneur en gluten humide GH (%) par ordre
décroissant.
|
N° (GxE)
|
GH
|
N° (GxE)
|
GH
|
N° (GxE)
|
GH
|
|
V18M1
|
44,055
|
V19M1
|
33,735
|
V30M2
|
26,758
|
|
V14M1
|
42,860
|
V3M1
|
33,430
|
V29M2
|
26,425
|
|
V29M1
|
42,560
|
V4M1
|
33,295
|
V1M3
|
25,863
|
|
V20M1
|
42,380
|
V17M3
|
33,205
|
V3M3
|
25,618
|
|
V30M1
|
41,460
|
V15M2
|
33,200
|
V2M1
|
25,432
|
|
V26M1
|
40,307
|
V26M2
|
33,100
|
V13M3
|
24,913
|
|
V27M1
|
39,920
|
V23M1
|
33,060
|
V10M1
|
23,935
|
|
V15M1
|
39,075
|
V2M2
|
31,935
|
V6M3
|
23,493
|
|
V11M1
|
38,765
|
V8M2
|
31,760
|
V19M2
|
23,068
23,068
|
|
V8M1
|
38,335
|
V21M1
|
31,307
|
V22M3
|
|
V16M1
|
37,718
|
V9M2
|
31,265
|
V21M3
|
22,950
|
|
V22M1
|
37,438
|
V5M2
|
31,140
|
V14M3
|
22,940
|
|
V14M2
|
36,578
|
V23M2
|
30,638
|
V12M3
|
22,915
|
|
V7M1
|
36,280
|
V21M2
|
30,513
|
V7M3
|
22,865
|
|
V24M1
|
36,220
|
V3M2
|
30,400
|
V29M3
|
21,810
|
|
V5M1
|
36,150
|
V18M2
|
30,048
|
V26M3
|
21,090
|
|
V7M2
|
36,118
|
V12M2
|
30,035
|
V5M3
|
20,630
|
|
V25M1
|
36,035
|
V25M2
|
29,740
|
V8M3
|
20,485
|
|
V9M1
|
35,905
|
V17M1
|
29,615
|
V24M3
|
19,743
|
|
V28M1
|
35,752
|
V22M2
|
28,850
|
V20M3
|
19,543
|
|
V13M1
|
35,640
|
V11M2
|
28,800
|
V25M3
|
19,245
|
|
V13M2
|
35,638
|
V2M3
|
28,698
|
V27M3
|
18,810
|
|
V1M1
|
35,612
|
V19M3
|
28,663
|
V15M3
|
18,213
|
|
V6M2
|
35,140
|
V27M2
|
28,618
|
V10M3
|
12,740
|
|
V1M2
|
35,000
|
V11M3
|
28,568
|
V16M3
|
11,513
|
|
V6M1
|
34,825
|
V24M2
|
28,518
|
V10M2
|
10,750
|
|
V17M2
|
34,505
|
V20M2
|
28,075
|
V9M3
|
10,618
|
|
V16M2
|
34,408
|
V18M3
|
27,808
|
V23M3
|
-
|
|
V12M1
|
34,275
|
V4M3
|
27,718
|
V28M3
|
-
|
|
V4M2
|
33,830
|
V28M2
|
27,603
|
V30M3
|
-
|
M1: Oued Smar, M2: El Khroub, M3: Sétif,
159
Tableau 10 : Valeurs moyennes de l'effet interaction
génotype x milieu du teneur en gluten sec GS (%) par ordre
décroissant.
|
N° (GxE)
|
GS
|
N° (GxE)
|
GS
|
N° (GxE)
|
GS
|
|
V9M2
|
19,4700
|
V3M1
|
10,2000
|
V3M3
|
8,2650
|
|
V14M3
|
13,7600
|
V4M2
|
10,2000
|
V30M2
|
8,2650
|
|
V14M1
|
12,5675
|
V17M3
|
10,0525
|
V18M3
|
8,1550
|
|
V26M1
|
12,3425
|
V23M1
|
9,9425
|
V1M3
|
8,0625
|
|
V29M1
|
12,1970
|
V4M1
|
9,9125
|
V29M2
|
8,0425
|
|
V20M1
|
11,9625
|
V2M2
|
9,8700
|
V12M3
|
7,6750
|
|
V16M1
|
11,9550
|
V19M1
|
9,8000
|
V13M3
|
7,6425
|
|
V30M1
|
11,9225
|
V8M2
|
9,6425
|
V10M2
|
7,5175
|
|
V8M1
|
11,6575
|
V5M2
|
9,4425
|
V18M1
|
7,4625
|
|
V22M1
|
11,4450
|
V15M2
|
9,4425
|
V19M2
|
7,2000
|
|
V11M1
|
11,3575
|
V26M2
|
9,4150
|
V6M3
|
7,1350
|
|
V27M1
|
11,3050
|
V2M1
|
9,3925
|
V21M3
|
7,0525
|
|
V13M1
|
11,2700
|
V23M2
|
9,2750
|
V10M1
|
6,8225
|
|
V24M1
|
11,2575
|
V21M1
|
9,2725
|
V7M3
|
6,4875
|
|
V15M1
|
11,2225
|
V3M2
|
9,2325
|
V22M3
|
6,3950
|
|
V14M2
|
11,0850
|
V12M2
|
9,0925
|
V15M3
|
6,0325
|
|
V6M1
|
11,0325
|
V18M2
|
9,0850
|
V5M3
|
6,0050
|
|
V7M1
|
10,9825
|
V17M1
|
9,0675
|
V29M3
|
5,9375
|
|
V1M1
|
10,8450
|
V11M2
|
9,0375
|
V26M3
|
5,8750
|
|
V7M2
|
10,8275
|
V25M2
|
8,9425
|
V24M3
|
5,6300
|
|
V5M1
|
10,7650
|
V27M2
|
8,9125
|
V8M3
|
5,4600
|
|
V9M1
|
10,7300
|
V21M2
|
8,7625
|
V25M3
|
5,3625
|
|
V28M1
|
10,6225
|
V11M3
|
8,7500
|
V20M3
|
5,2075
|
|
V16M2
|
10,5575
|
V4M3
|
8,6725
|
V27M3
|
5,0050
|
|
V25M1
|
10,5500
|
V28M2
|
8,6500
|
V10M3
|
3,4450
|
|
V12M1
|
10,4450
|
V22M2
|
8,6400
|
V16M3
|
3,1450
|
|
V6M2
|
10,4050
|
V2M3
|
8,6350
|
V9M3
|
2,8850
|
|
V17M2
|
10,3650
|
V20M2
|
8,6300
|
V23M3
|
-
|
|
V1M2
|
10,3200
|
V19M3
|
8,5175
|
V28M3
|
-
|
|
V13M2
|
10,2575
|
V24M2
|
8,4050
|
V30M3
|
-
|
M1: Oued Smar, M2: El Khroub, M3: Sétif,
160
APPENDICE D
ANALYSES DE LA VARIANCE
1/ Site d'Oued Smar
Tableau 1.1 : Analyse de la variance de la durée de la
phase végétative (DVP).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
839.467
|
28.9471
|
57.15
|
0.000
|
|
2 Répétition
|
3
|
2.43334
|
0.81111
|
1.60
|
0.193
|
|
Résiduelle
|
87
|
44.0666
|
0.50651
|
|
|
|
Totale
|
119
|
885.967
|
7.44510
|
|
|
Tableau 1.2 : Analyse de la variance de la hauteur de la
végétation (HT).
==========================================================
Source de Variation DDL SCE CM Test F Proba
==========================================================
|
1 Génotype
|
29
|
2042.00
|
70.4138
|
7.73
|
0.000
|
|
2 Répétition
|
3
|
295.542
|
98.5139
|
10.81
|
0.000
|
|
Résiduelle
|
87
|
792.583
|
9.11015
|
|
|
|
Totale
|
119
|
3130.12
|
26.3036
|
|
|
Tableau 1.3 : Analyse de la variance de la capacité de
tallage herbacé (TLL)
==========================================================
Source de Variation DDL SCE CM Test F Proba
==========================================================
|
1 Génotype
|
29
|
752795
|
25958.4
|
1.59
|
0.045
|
|
2 Répétition
|
3
|
30217.5
|
10072.5
|
0.62
|
0.610
|
|
Résiduelle
|
87
|
0.14218
|
16342.8
|
|
|
|
Totale
|
119
|
0.22048
|
18528.0
|
|
|
Tableau 1.4 : Analyse de la variance du nombre d'épis par
m2 (NE).
===========================================================
Source de Variation DDL SCE CM Test F Proba
===========================================================
|
1 Génotype
|
29
|
310964
|
10722.9
|
52.80
|
0.000
|
|
2 Répétition
|
3
|
2313.96
|
771.320
|
3.80
|
0.013
|
|
Résiduelle
|
87
|
17667.3
|
203.072
|
|
|
|
Totale
|
119
|
330945
|
2781.05
|
|
|
161
Tableau 1.5 : Analyse de la variance de la longueur de
l'épi (LE).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
1 Génotype 29 27.1884 0.93753 3.74 0.000
2 Répétition 3 2.86892 0.95630 3.82 0.013
Résiduelle 87 21.7986 0.25055
Totale 119 51.8559 0.43576
Tableau 1.6 : Analyse de la variance du nombre de grains par
épi (NGE).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
1 Génotype 29 2063.30 71.1483 3.42 0.000
2 Répétition 3 85.3333 28.4444 1.37 0.258
Résiduelle 87 1812.17 20.8295
Totale 119 3960.80 33.2840
Tableau 1.7 : Analyse de la variance du poids de 1000 grains
(PMG).
Source de Variation DDL SCE CM Test F Proba
1 Génotype 29 3339.54 115.157 7.53 0.000
2 Répétition 3 81.1407 27.0469 1.77 0.158
Résiduelle 87 1330.70 15.2955
Totale 119 4751.39 39.9276
Tableau 1.8 : Analyse de la variance du rendement en grains
(RDT)
=============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
1 Génotype 29 9189.59 316.883 7.69 0.000
2 Répétition 3 667.721 222.574 5.40 0.002
Résiduelle 87 3585.19 41.2091
Totale 119 13442.5 112.962
162
2/Site du Khroub
Tableau 2.1 : Analyse de la variance de la durée de la
phase végétative (DVP).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
1032.20
|
35.5931
|
9.18
|
0.000
|
|
2 Répétition
|
3
|
22.3000
|
7.43333
|
1.92
|
0.131
|
|
Résiduelle
|
87
|
337.200
|
3.87586
|
|
|
|
Totale
|
119
|
1391.70
|
11.6950
|
|
|
Tableau 2.2 : Analyse de la variance de la hauteur de la
végétation (HT).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
1976.34
|
68.1497
|
4.48
|
0.000
|
|
2 Répétition
|
3
|
93.0917
|
31.0306
|
2.04
|
0.113
|
|
Résiduelle
|
87
|
1324.16
|
15.2202
|
|
|
|
Totale
|
119
|
3393.59
|
28.5176
|
|
|
Tableau 2.3 : Analyse de la variance de la capacité de
tallage herbacé (TLL)
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
0.12758
|
43995.5
|
1.77
|
0.023
|
|
2 Répétition
|
3
|
169759
|
56586.3
|
2.27
|
0.085
|
|
Résiduelle
|
87
|
0.21668
|
24906.6
|
|
|
|
Totale
|
119
|
0.36125
|
30357.1
|
|
|
Tableau 2.4 : Analyse de la variance du nombre d'épis
par m2 (NE).
===========================================================
Source de Variation DDL SCE CM Test F Proba
===========================================================
|
1 Génotype
|
29
|
257674
|
8885.32
|
1.99
|
0.008
|
|
2 Répétition
|
3
|
7443.34
|
2481.11
|
0.56
|
0.649
|
|
Résiduelle
|
87
|
388169
|
4461.71
|
|
|
|
Totale
|
119
|
653287
|
5489.80
|
|
|
163
Tableau 2.5 : Analyse de la variance de la longueur de
l'épi (LE).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
53.7417
|
1.85316
|
1.84
|
0.016
|
|
2 Répétition
|
3
|
8.69168
|
2.89723
|
2.88
|
0.040
|
|
Résiduelle
|
87
|
87.5583
|
1.00642
|
|
|
|
Totale
|
119
|
149.992
|
1.26043
|
|
|
Tableau 2.6 : Analyse de la variance du nombre de grains par
épi (NGE).
============================================================
Source de Variation DDL SCE CM Test F Proba
===========================================================
|
1 Génotype
|
29
|
2832.97
|
97.6885
|
4.02
|
0.000
|
|
2 Répétition
|
3
|
117.767
|
39.2556
|
1.62
|
0.190
|
|
Résiduelle
|
87
|
2113.23
|
24.2900
|
|
|
|
Totale
|
119
|
5063.97
|
42.5543
|
|
|
Tableau 2.7 : Analyse de la variance du poids de 1000 grains
(PMG).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
1734.25
|
59.8018
|
4.24
|
0.000
|
|
2 Répétition
|
3
|
94.3972
|
31.4657
|
2.23
|
0.089
|
|
Résiduelle
|
87
|
1227.67
|
14.1111
|
|
|
|
Totale
|
119
|
3056.32
|
25.6833
|
|
|
Tableau 2.8 : Analyse de la variance du rendement en grains
(RDT)
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
2170.86
|
74.8572
|
7.40
|
0.000
|
|
2 Répétition
|
3
|
125.480
|
41.8266
|
4.13
|
0.009
|
|
Résiduelle
|
87
|
880.554
|
10.1213
|
|
|
|
Totale
|
119
|
3176.89
|
26.6966
|
|
|
164
3/Site de Sétif
Tableau 3.1 : Analyse de la variance de la durée de la
phase végétative (DVP).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
625.700
|
21.5759
|
21.40
|
0.000
|
|
2 Répétition
|
3
|
9.80000
|
3.26667
|
3.24
|
0.026
|
|
Résiduelle
|
87
|
87.7000
|
1.00805
|
|
|
|
Totale
|
119
|
723.200
|
6.07731
|
|
|
Tableau 3.2 : Analyse de la variance de la hauteur de la
végétation (HT).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
984.675
|
33.9543
|
3.12
|
0.000
|
|
2 Répétition
|
3
|
492.092
|
164.031
|
15.05
|
0.000
|
|
Résiduelle
|
87
|
948.158
|
10.8984
|
|
|
|
Totale
|
119
|
2424.93
|
20.3775
|
|
|
Tableau 3.3 : Analyse de la variance de la capacité de
tallage herbacé (TLL)
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
401144
|
13832.5
|
1.07
|
0.389
|
|
2 Répétition
|
3
|
395607
|
131869
|
10.2
|
0.000
|
|
Résiduelle
|
87
|
0.11217
|
12893.1
|
|
|
|
Totale
|
119
|
0.19184
|
16121.4
|
|
|
Tableau 3.4 : Analyse de la variance du nombre d'épis
par m2 (NE).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
147564
|
5088.40
|
1.31
|
0.169
|
|
2 Répétition
|
3
|
38695.6
|
12898.5
|
3.32
|
0.023
|
|
Résiduelle
|
87
|
337711
|
3881.73
|
|
|
|
Totale
|
119
|
523970
|
4403.11
|
|
|
165
Tableau 3.5 : Analyse de la variance de la longueur de
l'épi (LE).
============================================================
Source de Variation DDL SCE CM Test F Proba
=============================================================
|
1 Génotype
|
29
|
18.9917
|
0 .65488
|
2.45
|
0.125
|
|
2 Répétition
|
3
|
0.97966
|
0.32655
|
1.22
|
0.306
|
|
Résiduelle
|
87
|
23.2403
|
0.26713
|
|
|
|
Totale
|
119
|
43.2117
|
0.36312
|
|
|
Tableau 3.6 : Analyse de la variance du nombre de grains par
épi (NGE).
=============================================================
Source de Variation DDL SCE CM Test F Proba
=============================================================
|
1 Génotype
|
29
|
1810.18
|
62.4198
|
40.77
|
0.000
|
|
2 Répétition
|
3
|
2.55833
|
0.85277
|
0.56
|
0.649
|
|
Résiduelle
|
87
|
133.192
|
1.53094
|
|
|
|
Totale
|
119
|
1945.92
|
16.3523
|
|
|
Tableau 3.7 : Analyse de la variance du poids de 1000 grains
(PMG).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
1551.15
|
53.4881
|
38.13
|
0.000
|
|
2 Répétition
|
3
|
12.6227
|
4.20758
|
3.00
|
0.034
|
|
Résiduelle
|
87
|
122.057
|
1.40296
|
|
|
|
Totale
|
119
|
1685.83
|
14.1667
|
|
|
Tableau 3.8 : Analyse de la variance du rendement en grains
(RDT)
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
2137.90
|
73.7207
|
45.40
|
0.000
|
|
2 Répétition
|
3
|
41.8336
|
13.9445
|
8.59
|
0.000
|
|
Résiduelle
|
87
|
141.273
|
1.62383
|
|
|
|
Totale
|
119
|
2321.01
|
19.5043
|
|
|
166
4/Site de Tiaret
Tableau 4.1 : Analyse de la variance de la durée de la
phase végétative (DVP).
|
Source de Variation
|
DDL
|
SCE
|
CM
|
Test F
|
Proba
|
|
1 Génotype
|
29
|
488.467
|
16.8437
|
10.30
|
0.000
|
|
2 Répétition
|
3
|
11.2333
|
3.74444
|
2.29
|
0.083
|
|
Résiduelle
|
87
|
142.267
|
1.63525
|
|
|
|
Totale
|
119
|
641.967
|
5.39468
|
|
|
Tableau 4.2 : Analyse de la variance de la hauteur de la
végétation (HT).
|
Source de Variation
|
DDL
|
SCE
|
CM
|
Test F
|
Proba
|
|
1 Génotype
|
29
|
809.669
|
27.9196
|
1.57
|
0.046
|
|
2 Répétition
|
3
|
65.9229
|
21.9743
|
1.24
|
0.301
|
|
Résiduelle
|
87
|
1545.14
|
17.7602
|
|
|
|
Totale
|
119
|
2420.73
|
20.3423
|
|
|
Tableau 4.3 : Analyse de la variance de la capacité de
tallage herbacé (TLL)
=============================================================
Source de Variation DDL SCE CM Test F Proba
=============================================================
|
1 Génotype
|
29
|
595939
|
20549.6
|
1.12
|
0.340
|
|
2 Répétition
|
3
|
446071
|
148690
|
8.07
|
0.000
|
|
Résiduelle
|
87
|
0.16033
|
18429
|
|
|
|
Totale
|
119
|
0.26453
|
22229.7
|
|
|
Tableau 4.4 : Analyse de la variance du nombre d'épis
par m2 (NE).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
224264
|
7733.25
|
1.41
|
0.113
|
|
2 Répétition
|
3
|
93339
|
31113.0
|
5.67
|
0.001
|
|
Résiduelle
|
87
|
477305
|
5486.26
|
|
|
|
Totale
|
119
|
794908
|
6679.90
|
|
|
167
Tableau 4.5 : Analyse de la variance de la longueur de
l'épi (LE).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
12.7674
|
0.44025
|
0.65
|
0.909
|
|
2 Répétition
|
3
|
14.1909
|
4.73031
|
6.93
|
0.000
|
|
Résiduelle
|
87
|
59.3816
|
0.68254
|
|
|
|
Totale
|
119
|
86.3399
|
0.72554
|
|
|
Tableau 4.6 : Analyse de la variance du nombre de grains par
épi (NGE).
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
1300.18
|
44.8336
|
3.10
|
0.000
|
|
2 Répétition
|
3
|
219.292
|
73.0972
|
5.06
|
0.003
|
|
Résiduelle
|
87
|
1256.46
|
14.4420
|
|
|
|
Totale
|
119
|
2775.93
|
23.3271
|
|
|
Tableau 4.7 : Analyse de la variance du poids de 1000 grains
(PMG).
=============================================================
Source de Variation DDL SCE CM Test F Proba
=============================================================
|
1 Génotype
|
29
|
533.759
|
18.4055
|
9.89
|
0.000
|
|
2 Répétition
|
3
|
7.78800
|
2.59600
|
1.39
|
0.249
|
|
Résiduelle
|
87
|
161.972
|
1.86175
|
|
|
|
Totale
|
119
|
703.519
|
5.91192
|
|
|
Tableau 4.8 : Analyse de la variance du rendement grainier
(RDT)
|
Source de Variation
|
DDL
|
SCE
|
CM
|
Test F
|
Proba
|
|
1 Génotype
|
29
|
275.492
|
9.49973
|
4.66
|
0.000
|
|
2 Répétition
|
3
|
10.6366
|
3.54554
|
1.74
|
0.163
|
|
Résiduelle
|
87
|
177.246
|
2.03731
|
|
|
|
Totale
|
119
|
463.375
|
3.89390
|
|
|
168
Tableau 5 : Analyse de la variance du taux de mitadinage (MTD)
des 4 sites.
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
6674.83
|
230.17
|
488.38
|
0.000
|
|
2 Environnement
|
3
|
66628.61
|
22209.54
|
47125.09
|
0.000
|
|
Interaction (G)x(E)
|
87
|
20737.02
|
238.36
|
505.75
|
0.000
|
|
Résiduelle
|
320
|
128.22
|
0.47
|
|
|
|
Totale
|
478
|
84610.13
|
196.68
|
|
|
Tableau 6 : Analyse de la variance du taux d'extraction (EXT) des
4 sites.
|
Source de Variation
|
DDL
|
SCE
|
CM
|
Test F
|
Proba
|
|
1 Génotype
|
29
|
3063.61
|
105.64
|
79.83
|
0.000
|
|
2 Environnement
|
3
|
10605.97
|
3535.32
|
4641.51
|
0.000
|
|
Interaction (G)x(E)
|
87
|
11935.41
|
137.19
|
102.50
|
0.000
|
|
Résiduelle
|
320
|
181.81
|
1.34
|
|
|
|
Totale
|
478
|
26086.81
|
54.46
|
|
|
Tableau 7 : Analyse de la variance du taux de gluten humide
(GH) des 3 sites.
============================================================
Source de Variation DDL SCE CM Test F Proba
============================================================
|
1 Génotype
|
29
|
4358.82
|
150.30
|
75.87
|
0.000
|
|
2 Environnement
|
2
|
16438.16
|
8219.08
|
4148.82
|
0.000
|
|
Interaction (G)x(E)
|
58
|
7825.93
|
134.93
|
68.11
|
0.000
|
|
Résiduelle
|
270
|
534.89
|
1.98
|
|
|
|
Totale
|
368
|
29157.80
|
81.22
|
|
|
Tableau 8 : Analyse de la variance du taux de gluten sec (GS)
des 3 sites.
===========================================================
Source de Variation DDL SCE CM Test F Proba
===========================================================
|
1 Génotype
|
29
|
622.20
|
19.54
|
20.32
|
0.000
|
|
2 Environnement
|
6
|
1368.06
|
684.03
|
711.42
|
0.000
|
|
Interaction (G)x(E)
|
69
|
1312.15
|
22.62
|
23.53
|
0.000
|
|
Résiduelle
|
270
|
259.60
|
0.69
|
|
|
|
Totale
|
359
|
3602.41
|
9.77
|
|
|
169
|
|


