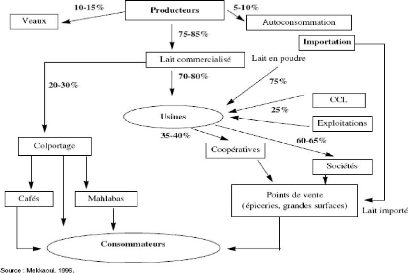
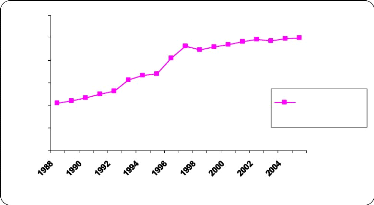
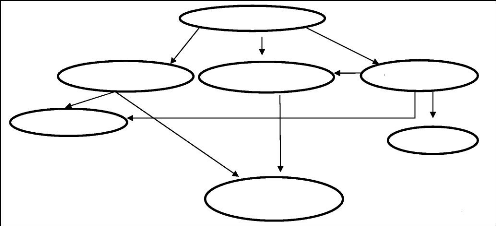
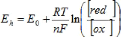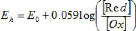Contribution à l'évaluation des pratiques frauduleuses dans le lait à la réception( Télécharger le fichier original )par El Houssain BOUICHOU - Ingénieur zootechnicien 2009 |

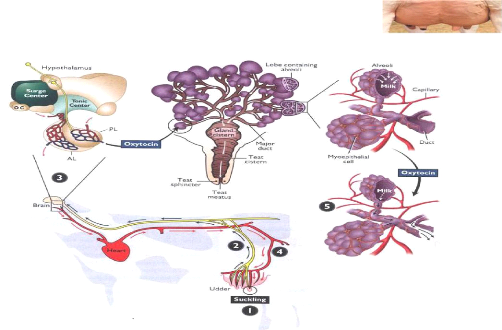
B/ Libération d'ocytocine dans le sangSous la commande de l'ocytocine, les muscles entourant l'alvéole se contractent, obligeant ainsi le lait contenu dans les alvéoles à sortir par le canal. Cette hormone (ocytocine), transportée par voie sanguine, provoque la contraction des cellules myoépithéliales des acini mammaires et l'éjection du lait alvéolaire dans les canaux galactophores puis dans la citerne du pis.
Contribution à l'évaluation des pratiques frauduleuses dans le lait à la réception Figure11 : libération d'ocytocine dans le sang 3- Physiologie de la descente du lait Sans le concours de l'ocytocine, il n'est pas possible de récolter le lait alvéolaire qui représente 70% à 80 % de la qualité totale. Il n'est pas possible non plus de recueillir tout le lait alvéolaire, ce qui reste est le lait résiduel qui devrait être une fraction d'au plus 6% du lait total. Chez la truie, les stimuli sont nombreux puisque les porcelets tètent 10 à 20 fois par jour. Mais l'injection ne dure que 20 secondes environ. Chaque porcelet consommera tout de même de 10 à 50 g par porcelet par tétée, ce qui classe la truie proportionnelle à la pression à l'intérieur de la glande. Ainsi, après une tétée ou après une traite la pression est très faible dans la glande, alors la sécrétion lactée recommence et elle diminue graduellement à mesure que la pression augmente à cause du lait qui s'accumule. C'est pourquoi les traites ou les tétées fréquentes augmentent la production de lait. L'effet peut être inverse si la glande est vidangée moins souvent ou incomplètement. Chez la vache Toute stimulation tactile des trayons déclenche immédiatement un influx nerveux en direction du système nerveux central. Une fois stimulée, la posthypophyse libère l'hormone ocytocine.
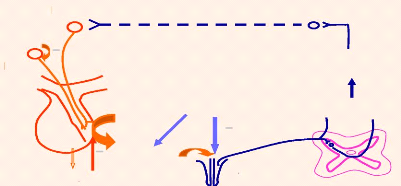
Figure13. Reflexe neuroendocrinien d'éjection du lait Réflexe neuroendocrinien
Stimulation Endorphine Système Réseau Système adrénergique NPV ocytocine - NSO Faisceau Tronc cérébral - Ocytocine - d'éjection du lait L'ocytocine met environ 50 secondes pour arriver au pis après transmission du réflexe nerveux et son action dure de 2 à 8 minutes (durée de demi-vie dans le sang de 4min) (optimum d'activité jusqu'à 5 min après la stimulation). Ce premier réflexe neuro-endocrinien est secondé par un réflexe nerveux autonome local qui a pour effet une dilatation des canaux galactophores et du sphincter des trayons. Le débit sanguin du pis est également augmenté pendant la traite, ce qui se traduit notamment par une légère érection du trayon. L'éjection des premiers jets de lait représente la meilleure stimulation tactile des trayons avant la traite. La présence des corpuscules thermo-récepteurs montre l'importance de la température sur la traite. Il est toujours conseillé de travailler à une température voisine de la bouche du veau. La descente du lait peut également être déclenchée par des stimuli visuels, auditifs, ou autres (heure de traite, entrée en salle d'attente ou en salle de traite, vue du veau,...). Les pertes de lait parfois observées avant la traite n'ont par contre aucun lien avec une augmentation des taux circulants d'ocytocine.B. Boudry. 2005. Si la méthode de stimulation n'a pas d'influence sur la quantité d'ocytocine libérée pendant la traite, l'intervalle de temps entre cette stimulation et le début de la traite a lui une grande importance sur la traite, la production laitière et la santé mammaire. Comme précédemment décrit, le lait avant la traite est répartie dans le quartier en une portion alvéolaire (80%) et une portion citernale (20%). Si cette dernière est directement disponible pour la traite, la partie alvéolaire nécessite l'action de l'ocytocine avant d'être éjectée. Si la pose de la griffe ne coïncide pas avec la descente du lait, la portion citernale sera traite avant l'arrivée du lait alvéolaire dans la citerne du pis. Il va s'ensuivre une chute temporaire, partielle ou complète, du débit déjection
du lait lors du passage entre lait citernal et alvéolaire, phénomène appelé traite bimodale. L'objectif premier de la routine de traite est de préparer les vaches dans un ordre permettant de synchroniser la descente du lait et la pose de la griffe. L'intervalle de temps optimum entre la stimulation et la descente du lait varie respectivement de 50 à 90 secondes pour les vaches en début et en fin de lactation. Idéalement, l'intervalle de temps entre la première stimulation des trayons et la pose de la griffe varie entre 60 et 90 sec afin de profiter au maximum du réflexe de l'ocytocine. Avec une stimulation adaptée, la traite est rapide, ininterrompue et complète ; la surtraite et la durée de traite à faible débit (< 1kg/min) sont réduites au minimum. Le gain moyen lié à une stimulation adéquate serait de 0,6 min par vache sur le temps de traite (gain de 10%) et de 0,3 Kg de lait par traite (gain de 1%). De plus, la réduction du temps de traite préserve l'intégrité des trayons et donc les défenses aspécifiques de la mamelle. Le temps de préparation des trayons permet d'évaluer l'importance des mesures d'hygiènes apportées aux trayons et le niveau de stimulation des trayons avant la traite. Le temps de pose du faisceau mesure la synchronisation entre la descente du lait et le début de la traite. La durée de traite sera quant à elle dépendante de la correcte stimulation de la vache mais également d'autres facteurs liés à la machine à traire (niveau de vide, rapport de pulsation, débit seuil et temporisation du décrochage automatique,...) ou à l'animal (rapidité de traite, niveau de production, nombre de lactation, ...). Mein (98) conclut que la durée de traite d'une vache produisant 15 kg de lait par traite s'approche des 6 minutes (plus ou moins une minute) et que cette durée peut être augmentée d'une minute par tranche de 5kg de lait supplémentaire par traite. Cette mesure de la durée de traite peut être complémentée par un calcul du débit moyen d'éjection du lait si les niveaux de productions individuelles sont connus. Pour le troupeau, un débit d'éjection moyen du lait par traite ne devrait jamais être inférieur à 2,2 L /min, l'objectif étant d'atteindre des valeurs de 2,5 L/min.
Partie II. Compositions & Comportement du lait face à différents traitements. EL.BOUICHOU
1. Lait Le lait est un milieu réactionnel complexe dont le rôle premier est de satisfaire tous les besoins nutritionnels du nourrisson. Il contient principalement des glucides, de la matière grasse, des protéines et des sels minéraux. D'autres constituants se retrouvent dans le lait sous forme de traces tels que les vitamines, les enzymes et les gaz dissous. La composition détaillée du lait est donnée à la section 1.1 . Le pH naturel du lait se situe entre 6,6 et 6,8 et il est grandement influencé par sa composition. Le lait se présente sous la forme d'une émulsion de particules de gras dans un environnement aqueux. Les principales protéines laitières sont les caséines qui se présentent sous la forme d'une dispersion de petites particules colloïdales (<300 nm) nommées micelles. 1.1 Composition du lait Les constituants majeurs du lait de vache sont présentés au Tableau 4 . De ceux-ci, les protéines présentent un intérêt tout particulier car ce sont les responsables majeures des propriétés technologiques du lait. Les protéines représentent 95 % de la matière azotée du lait et peuvent être départagées en deux groupes : les caséines et les protéines du lactosérum. L'azote non-protéique est composé de substances diverses telles que l'urée, l'ammoniac, l'acide urique, des acides aminés libres, des peptides, etc. Tableau 4. Composition générale du lait de vache (Amiot et al ., 2002).
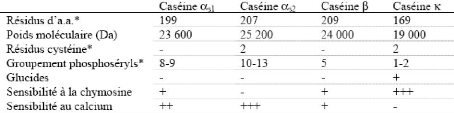
1.1.1 Caséines Les quatre principales caséines qui existent naturellement dans le lait sont les caséines ás1 , ás2 , â et ê. Les caséines Y sont, pour leur part, des fragments peptidiques issus de la dégradation de la â-caséine par la plasmine. Les caséines se distinguent par leur faible solubilité à pH 4,6 et elles sont différenciées sur la base de la distribution des charges et de la sensibilité à la précipitation par le calcium. Leurs caractéristiques physicochimiques sont présentées au Tableau 5 . Elles partagent un facteur de composition commun puisque ce sont des protéines conjuguées, la plupart avec des groupements phosphate estérifiés à des résidus sérine. Les sites phosphoséryls sont souvent regroupés, créant des zones hydrophiles dans leur chaîne (Brulé et al ., 1997). La capacité des caséines à fixer le calcium diminue dans l'ordre B1;s2>
B1;s1> â> ê, correspondant à leur contenu décroissant en phosphosérine. Le groupe phosphate des résidus phosphosérine constitue le principal site de liaison du calcium (Rollema, 1992), contribuant à la structure de la micelle. Tableau 5. Caractéristiques physico-chimiques des caséines (Brulé et al. ,1997).
*nombre par mole La forte présence de résidus proline induit des coudes dans la chaîne qui empêchent la formation d'une structure secondaire serrée et ordonnée. De plus, l'absence de structure tertiaire est responsable de la grande résistance à la dénaturation thermique de ces protéines. 1.1.1.1 Caséine ê Certes, la caséine la plus étudiée est la caséine ê (ê-CN), probablement à cause de son importance dans la stabilité de la micelle et de son rôle en transformation laitière. La caséine ê se différencie des autres caséines par sa grande sensibilité à la chymosine, son peu d'affinité pour le calcium et la présence de résidus glucidiques. La ê-CN favorise la stabilité de la micelle de deux façons. Tout d'abord, prises individuellement et en présence de calcium, les caséines B1;s1 , B1;s2 sont insolubles et la caséine â n'est soluble qu'entre 0-4 o C. L'association de ces caséines avec la caséine ê est une condition essentielle à leur dispersion en présence de calcium (Holt et Horne, 1996). Ensuite, la ê-CN possède deux régions qui sont séparées lors de l'hydrolyse du lien Phe105 -Met106 par la chymosine. La partie N-terminale se nomme para-ê-CN et est de nature hydrophobe. Elle demeure attachée à la micelle. La partie C-terminale de la ê-CN est le caséinomacropeptide (CMP), de nature hydrophile et chargée négativement. La partie CMP de la ê-CN s'étend dans le sérum et forme le chevelu micellaire d'une épaisseur de 5 à 10 nm (Dalgleish, 1998). Ce dernier participe à la stabilisation des micelles par répulsion électrostatiques et stériques (Brulé et al ., 1997; Holt et Horne, 1996; Lucey, 1995; Fox, 1993). La protéine qui perd le chevelu micellaire par hydrolyse n'a plus la capacité de stabiliser les autres caséines (Amiot et al. , 2002). La ê-CN est également la seule caséine
à avoir des résidus glucidiques dans sa
n'a pas de rôle spécifique mais renforce le caractère hydrophile de la partie Cterminale en plus d'augmenter l'encombrement stérique à la surface de la micelle. 1.1.2 Protéines du lactosérum Les autres protéines du lait sont présente dans le lactosérum. Les protéines sériques sont définies comme étant les protéines solubles présentes dans le lactosérum suivant la précipitation des caséines à pH 4,6 à 20 °C. Les protéines sériques englobent une première fraction protéique (80 %) composée de f3-lactoglobuline (f3-LG), d' a- lactalbumine (a-LA), d'albumine de sérum bovine (BSA) et d'immunoglobulines, et une deuxième fraction non protéique (20 %) composée de protéoses peptones et de composés azotés (De Wit, 1981). 1.1.2.1 f3-lactoglobuline Les deux principales protéines sériques sont la f3-LG et l'a-LA. À elle seule, la f3-LG représente 50 à 60 % des protéines du lactosérum (Amiot et al. , 2002). Cette protéine est hautement nutritive puisqu'elle contient tous les acides aminés essentiels. Certains auteurs ont démontré la capacité de la f3-LG à lier l'acide rétinoïque ou certains ligands hydrophobes tels que les vitamines liposolubles et les acides gras. Cependant, la fonction biologique réelle de la protéine demeure inconnue (Kontopidis et al., 2002). La structure primaire de la f3-LG, qui possède un poids moléculaire de 18 362 Da, est composée de 162 résidus d'acides aminés dont la séquence est donnée à la Figure 14. Il existe plusieurs variants génétiques de cette protéine, les plus communs étant le A et le B qui se différencient par les résidus d'acides aminés en position 64 (aspartyle et glycyle respectivement) et 118 (valyle et alanyle respectivement) (Morr et Ha, 1993). Figure 14. Structure primaire de la â-lactoglobuline A (Adapté de Walstra et Jenness, 1984).
La structure secondaire de cette protéine contient
environ 50 % de feuillets f3, 15 %
66-160 et 106-119, pour stabiliser la structure. La Cys121 exhibe donc un thiol libre libre pouvant induire des réactions d'échange SH/S-S intra et intermoléculaires et qui est responsable de la formation d'agrégats lors du chauffage (Sawyer et al. , 2002). Le premier pont disulfure, impliqué dans le maintien du coeur hydrophobe, est faiblement accessible et donc peu réactif comparativement au second (Cayot et Lorient, 1998). La â-lactoglobuline, très compacte de par sa structure, est très résistante à l'hydrolyse, propriété conférée par l'action stabilisante des deux ponts disulfures (Papiz et al., 1986), ce qui en fait une protéine qui résiste à l'hydrolyse gastrique. Selon les conditions du milieu, la structure quaternaire de cette protéine prend différentes formes. Figure . 1 5 Structure tridimensionnelle d'un monomère de la â-lactoglobuline A. Source : Protein Data Bank (Qin et al. , 1998).
Le point isoélectrique de la â-LG du lait des ruminants est 5,2 (Kinsella et al. , 1989) et la conformation de cette protéine est affectée par les variations de pH. Au pH naturel du lait (pH 6,8), la â-LG se retrouve principalement sous forme de dimère. À mesure que le pH diminue, les dimères tendent à se polymériser en octomères de 147 kDa (Verheul et al. , 1999), mais à pH<3,5, les dimères se dissocient en monomères par répulsions électrostatiques. À pH 7, un dépliement partiel de la structure de la protéine entraîne un changement de conformation. L'exposition du groupement carbonyle du résidu Glu89, favorisé à pH élevé, serait responsable de ce changement de conformation, appelé la transition de Tanford (Qin et al. , 1998). À pH>8, les molécules partiellement dénaturées par la transition de Tanford s'associent de manière irréversible par l'établissement de ponts disulfures intermoléculaires qui peuvent mener à l'agrégation des protéines (Verheul et al. , 1999). La température influence également la structure tridimensionnelle de la â-LG. Bien que la â-LG se retrouve principalement sous forme de dimères au pH naturel du lait, les dimères se dissocient en monomères lorsque la température atteint environ 65 °C. Au-delà de cette température survient un dépliement de la structure qui provoque la dénaturation irréversible de la protéine (Sawyer et al. , 2002).
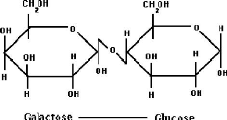
En plus des changements structuraux causés par la température et/ou le pH, des réactions d'échange SH/S-S surviennent également selon l'exposition du résidu thiol libre en position 121 (Cys121). Ces réactions sont impliquées dans le mécanisme de dénaturation de la â-LG, de même que dans sa gélification (Shimada et Cheftel, 1989) et sont favorisées à pH neutre ou basique. À pH plus faible, l'augmentation de la température est nécessaire afin d'initier le dépliement et le processus de dénaturation. La formation de ponts disulfures intermoléculaires est à la base de la polymérisation irréversible ainsi que de la gélification de la â-LG (Monahan et al. , 1995). 1.1.3 Lactose Le principal glucide du lait est le lactose, un disaccharide composé de B1;-Dglucose et de â-D-galactose, tel qu'illustré à la Figure 16. Puisque le carbone anomérique du glucose n'est pas engagé dans le lien glycosidique, le lactose est un sucre réducteur pouvant réagir avec une protéine lors de la réaction de Maillard, détaillée à la section 1.4.3.3.2. Quoique le lait contienne près de 4,6 % de lactose, il n'a pas de goût sucré, le pouvoir sucrant du lactose n'étant que de 22 par rapport au saccharose à qui une valeur de 100 est attribuée (Amiot et al ., 2002). Le lactose est utilisé comme substrat lors de la fermentation du lait par les bactéries lactiques, phénomène à la base de produits fermentés tels que le fromage et le yogourt. Figure 16. Structure du lactose.
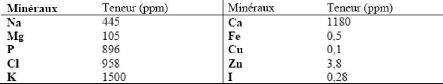
1.1.4 Minéraux Les minéraux jouent un rôle important dans l'organisation structurale des micelles de caséine. Les principaux minéraux présents dans le lait sont donnés au Tableau 17. et de nombreux autres sont présents à l'état de traces. Les minéraux sont répartis entre l'état soluble, sous la forme d'ions ou de sels, et l'état colloïdal, associés à la micelle de caséine. De plus, les minéraux sont en équilibre entre les deux états. Tout
particulièrement, le phosphore et le calcium jouent un rôle prépondérant dans le maintien de l'intégrité de la micelle de caséine. Cet aspect sera approfondi à la section 1.2.3.3 . Tableau 17. Teneur de différents minéraux dans le lait (Amiot et al ., 2002).
1.1.5 Constituants mineurs En plus des protéines, glucides, lipides, et minéraux, le lait contient des vitamines et des enzymes. Les vitamines A, D, E et K sont liposolubles, ainsi se retrouvent-elle dans la matière grasse et peuvent être perdues lors de l'écrémage. D'autres vitamines sont hydrosolubles et se retrouvent dans le sérum. C'est le cas de l'acide ascorbique (C) qui s'y retrouve en plus grande concentration à 2 mg/100 ml. La thiamine (B1), la riboflavine (B2), la pyroxydine (B6), la cyanocobalamine (B12), la niacine, l'acidepantothénique, l'acide folique et la biotine (H) (Amiot et al. , 2002). De ces vitamines, l'acide ascorbique joue un rôle important car non seulement est-ce la vitamine présente en plus grande concentration mais c'est un agent anti-oxydant et un stabilisateur du potentiel d'oxydoréduction du lait, tel qu'il sera démontré à la section 1.3.4.1. Des vitamines peuvent être dégradées sous conditions oxydatives, c'est le cas des vitamines C, B2, A et acide folique (Rada-Mendoza et al. , 2002). Pour leur part, les pertes en vitamines B12, A et E dépendent de la quantité d'oxygène dans le lait (Schrodes, 1982). Le lait contient un certain nombre d'enzymes, dont certaines sont oxydoréductrices. C'est le cas des enzymes lactate dehydrogénase, malate déhydrogénase, xanthine oxidase, lactoperoxidase et sulphydryl oxydase (Walstra et Jenness, 1984). Leur rôle en relation avec les réactions d'oxydoréductions du lait sera approfondi à la section 1.3.4.4 . Comme autre constituant mineur, l e lait contient des gaz dissous. Le lait cru contient environ 6 mg/L d'oxygène; cette présence peut avoir un effet négatif sur les qualités nutritionnelles du produit (Walstra et Jenness, 1984). 1.1.6 Variabilité de composition La composition du lait de vache varie selon différents facteurs que sont l'individualité, la race, le stade de lactation, l'alimentation, la saison et l'âge (Amiot et al ., 2002). Notamment, la saison possède une grande influence sur la composition générale du lait, ce qui peut avoir des conséquences directes en technologie laitière.
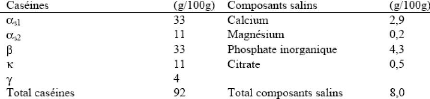
2. Micelle de caséine 2.1 Composition La micelle de caséine renferme, en plus des différentes caséines et composants salins présentés au Tableau 18, du lactosérum enchâssé et les enzymes lipase et plasmine. La micelle de caséine est constituée à 92 % des caséines B1;s1 , B1;s2 , â et ê et le 8 % restant correspond aux composants salins dont les deux plus importants sont le calcium et le phosphore (Amiot et al. , 2002). Les proportions exactes des constituants sont variables selon l'influence sur la composition du lait des facteurs énumérés à la section 1.1.6 . Quoique la micelle de caséine soit passablement stable, il y a quatre façons principales d'induire l'agrégation. Ce sont l'utilisation d'enzymes protéolytiques, les conditions acides, les traitements de chaleur et la gélification causée par le vieillissement. Tableau 18. Composition générale de la micelle de caséine (Brulé et al ., 1997).
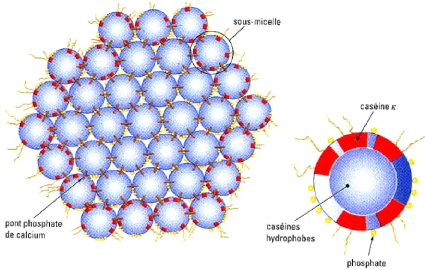
2.2 Structure La grande majorité des caséines sont présentes sous la forme de particules colloïdales nommées micelles de caséine qui sont passablement stables. Les quatre principales façons d'induire l'agrégation sont l'utilisation d'enzymes protéolytiques, les conditions acides, les traitements de chaleur et la gélification causée par le vieillissement. Toutefois, la structure exacte de la micelle de caséine fait encore l'objet de spéculations. Il existe plusieurs modèles de micelles de caséines dont le plus répandu est le modèle avec sous-unités de Schmidt (1982). Cependant, le modèle « à dualité des liens » de Horne (1998) tend à vouloir s'imposer chez certains. 1.2.2.1 Modèle avec sous-unités Le modèle avec sous-unités, présenté à la Figure 19 , suggère que les sous-unités n'étaient pas toutes de même composition mais que certaines étaient plus riches en êCN et étaient situées en surface de la micelle. Finalement, Schmidt (1982) précisa que les sous-micelles étaient reliées par des agrégats de Ca9 (PO4 )6 . Dans le modèle actuel, les micelles sont en fait composées de sous-unités comprenant de 10 à 100 molécules de caséines et nommées sous-micelles. Les sous-micelles sont reliées entre elles par des ponts phosphate de calcium. La composition des sous-micelles au centre et en périphérie est différente. En effet, les caséines â et ás1 sont
plus présentes au centre de la micelle et forment le coeur hydrophobe alors que la partie externe, davantage hydrophile, est formée de caséine ás1 , ás2 et ê (Amiot et al. , 2002). Figure 19. Modèle de micelle de caséine avec sous-unités (Amiot et al., 2002).
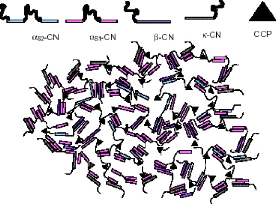
2.2.2 Modèle à dualité des liens Un nouveau modèle, nommé « à dualité de liens », a récemment été proposé par Horne (1998). Ce modèle est proposé pour remédier au manque d'un mécanisme plausible pour expliquer l'assemblage, la croissance et la terminaison de la croissance des autres modèles, tout en étant conforme à la réaction des micelles suite à un changement de pH, de température, d'addition d'urée ou de retrait de phosphate de calcium avec l'EDTA (Horne, 1998). Le modèle propose deux voies de polymérisation pour l'assemblage des micelles. Ces voies sont la polymérisation de caséines individuelles via les régions hydrophobes des caséines et le pontage grâce au phosphate de calcium colloïdal (CCP). La formation des micelles est contrôlée par l'équilibre entre les forces d'attraction et de répulsion dans les micelles, notamment les excès localisés
d'attraction hydrophobe et les répulsions électrostatiques (Horne, 1998). La structure de la micelle selon le modèle à dualité des liens est présentée à la Figure 20 . Figure 20. Structure de la micelle selon le modèle à dualité des liens (Horne, 1998).
2.3 Caractéristiques physicochimiques de la micelle de caséine 2.3.1 Diamètre micellaire Le diamètre de la micelle varie entre 50 à 600 nm et le diamètre moyen est de 120 nm (Fox, 1993). Le rayon de la micelle de caséine est inversement proportionnel au contenu en caséine ê (Horne et al ., 1997). En effet, les plus petites micelles renferment de plus fortes proportions de caséine ê en raison du ratio surface-volume élevé des petites micelles (van den Berg et al ., 1992). Quoique le contenu en calcium et en phosphore micellaire augmente avec l'augmentation du diamètre micellaire, il ne contrôle pas la taille des micelles. La quantité de caséine ê est donc le facteur dominant (Umeda et Aoki, 2002). 2.3.2 Hydratation La quantité d'eau qui se lie à la micelle dépend de sa composition mais aussi de d'autres facteurs. Ces facteurs sont le nombre de groupes polaires exposés, la conformation des protéines superficielles, la polarité en surface, le pH, les sels présents et leur concentration (Kinsella, 1984). L'eau fixée au chevelu micellaire, soit
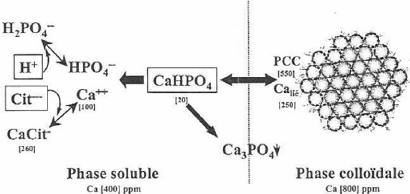
la couche d'hydratation en surface, est faiblement liée et contribue à la stabilisation de la micelle. La micelle fixe environ 3 g d'eau/g de protéine pour un volume hydrodynamique de 3,9 ml/g de protéines lorsque le chevelu micellaire est inclus (Walstra et van Vliet, 1986; Dalgleish, 1981). Cependant, l'hydratation interne de la micelle est d'environ 2 g d'eau/g protéine (Fox, 1993). Il y a une forte corrélation positive entre la quantité de caséine ê présente et le degré d'hydratation. La capacité d'hydratation et le diamètre micellaire sont donc étroitement reliés. De ce fait, les petites micelles contiennent davantage de caséine ê et sont plus hydratées (Anema et Creamer, 1993). 2.3.3 Minéraux La composition minérale du lait de vache a été abordée à la section 1.1.4 et le rôle de certains minéraux dans le maintien de l'intégrité de la micelle a été mentionné. Les deux principaux minéraux d'importance dans la structure de la micelle sont le calcium et le phosphore dont la teneur dans le lait excède leur solubilité. C'est grâce à l'interaction avec les micelles que la précipitation est évitée. L'équilibre entre les formes solubles et associées de ces minéraux est représenté à la Figure 21 . Figure 21. Les principaux équilibres salins du lait (Britten et Pouliot, 2002).
Environ le tiers du calcium se retrouve dans la phase soluble qui se divise en 20 % de sel citrate ou phosphate et 10 % d'ions calcium libres. Le deux-tiers restant est soit directement liés à la micelle ou fait partie des complexes de phosphate de calcium à l'intérieur de celle-ci (FAO, 1998). Dans le cas du phosphore, la majeure partie (60 %) est sous forme de phosphate organique, dont la moitié est liée à la caséine et l'autre en solution, 20 % est lié aux groupements hydroxyles de certains acides aminés et le reste se partage entre les phospholipides et les esters hydrosolubles (FAO, 1998). La Figure 22. résume la distribution du calcium et du phosphore dans le lait bovin.
Figure 22. Répartition du calcium et du phosphore dans le lait (adapté de FAO, 1998).
Les équilibres salins sont influencés par le pH, la température, la nature et la concentration des sels. Une diminution de pH entraîne la dissociation du phosphate de calcium (Amiot et al. , 2002); l'acidification cause donc une décalcification de la micelle (St-Gelais et TirardCollet, 2002). Au contraire, une augmentation de pH et l'augmentation de la concentration en Ca2+ déplacent les équilibres pour entraîner la formation de phosphate de calcium tricalcique qui tend à précipiter (Amiot et al. , 2002). L'effet de la température sur la solubilité du phosphate de calcium est contraire aux principes généraux de solubilité puisque sa solubilité diminue à mesure que la température augmente (Amiot et al. , 2002). Au chauffage, le déplacement de l'équilibre a lieu vers la forme colloïdale, ce qui augmente la minéralisation de la micelle. Le déplacement vers la forme colloïdale fait également en sorte que le H2 PO4 - se dissocie et libère des ions H+ , abaissant le pH. Finalement, la variation de la force ionique du lait causée par l'ajout de sels tels que le chlorure de sodium a pour effet de solubiliser le phosphate de calcium colloïdal qui peut mener jusqu'à la dissociation de la micelle (Britten et Pouliot, 2002; Horne, 1998). Lorsqu'il s'agit d'un sel calcique, tel que le chlorure de calcium, il y a augmentation du calcium soluble. Celui-ci, en plus de causer une légère baisse de pH par dissociation du H2 PO4 - , tend à se fixer à la micelle. Sous l'addition de calcium, ces protéines demeurent solubles jusqu'à ce qu'une quantité critique soit liée après laquelle la solubilité diminue (Rollema, 1992).
3 Propriétés électrochimiques 3.1 Réactions d'oxydoréduction Les réactions d'oxydoréduction impliquent le transfert d'électrons entre atomes et molécules. Le transfert d'oxygène, d'hydrogène ou des deux à la fois peut se produire. L'oxydation et la réduction sont, respectivement, la perte et le gain d'électrons. S'il n'y a pas de flux d'électrons qui entrent ou sortent du système à l'étude, les réactions redox sont nécessairement couplées, une réaction d'oxydation (perte d'électrons) ayant lieu simultanément à une réaction de réduction (gain d'électrons) (Walstra et Jenness, 1984). Une réaction de réduction réversible d'un composé en un autre s'écrit comme suit : Ox + n e- ?Red Comme par exemple : Fe3+ + e- ? Fe2+ Les réactions d'oxydoréduction dans les systèmes laitiers sont influencés par les traitements thermiques, la concentration en oxygène dissous, la concentration en ions métalliques tels que le Cu2+ , l'exposition à la lumière et par les oxydoréductases, à la fois du lait et des microorganismes (Singh et al., 1997; Walstra et Jenness, 1984). 3.2 Mesure du potentiel redox Le potentiel redox est l'aptitude d'un système à capter et céder des électrons. Le système de mesure du potentiel d'un système à transférer des électrons comprend une électrode inerte en or ou platine et une électrode de référence de potentiel connu (Walstra et Jenness, 1984; Sherbon, 1999). Tous les potentiels sont exprimés envers un standard commun, généralement l'électrode à hydrogène (SHE) (Walstra et Jenness, 1984). L'électrode à hydrogène possède une valeur assignée de zéro quand une électrode inerte trempe dans une solution d'activité unitaire en regard des protons (pH=0) en équilibre avec l'hydrogène gazeux à une pression de 1 atmosphère. Le potentiel redox (Eh) est relié aux concentrations des formes oxydées et réduites du composé suivant l'équation de Nernst :
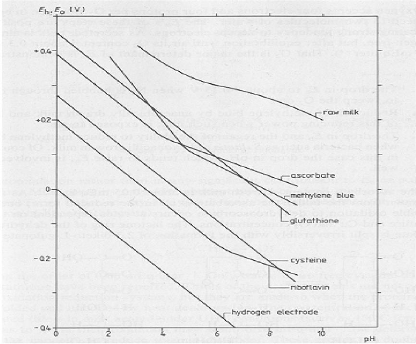
En accord avec la convention utilisée par les biochimistes, le potentiel redox tend vers des valeurs plus positives lorsque le ratio [Ox]/[Red] augmente. Le potentiel redox standard du système, caractéristique propre à chaque système redox, est une mesure de l'habilité relative du système à donner ou accepter des électrons. En pratique pour les solutions diluées, les concentrations molaires des composés sont utilisées dans les calculs plutôt que leur activité(Walstra et Jenness, 1984). De plus, le potentiel standard du système dépend aussi du pH. La Figure 23 présente le potentiel redox du lait et le potentiel standard de plusieurs systèmes en fonction du pH (Walstra et Jenness, 1984) Figure 23. Potentiel redox du lait et le potentiel standard de plusieurs systèmes en fonction du pH (Walstra et Jenness, 1984).
Finalement, la cinétique des réactions d'oxydoréduction est importante à considérer dans la mesure du potentiel redox. Certaines réactions présentent un haut niveau d'énergie d'activation ce qui implique qu'un délai considérable peut s'écouler avant que l'équilibre ne soit atteint. Des mesures exactes de potentiel redox sont donc longues dans le lait qui possède plusieurs systèmes qui se superposent et dont certains ne sont que très lentement ou incomplètement réversibles. De plus, la diffusion
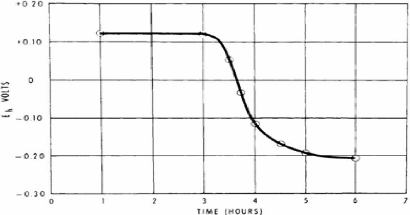
d'oxygène atmosphérique dans le lait peut compliquer les mesures (Walstra et Jenness, 1984). Le rôle de l'oxygène sera approfondi à la section 3.4 . 3.3 Potentiel redox du lait Le potentiel redox d'échantillons individuels de lait en équilibre avec l'air, puisque lorsque sécrété le lait est presque entièrement dépourvu d'oxygène, est généralement compris entre +250 à +350 mV (électrode de référence SHE, 25 °C, pH 6,6-6,7). (Walstra et Jenness, 1984). L'alimentation des vaches pourrait y jouer un rôle, les vaches en pâturage produisant du lait présentant un potentiel redox de 20 mV inférieur à celui des vaches nourries à la fois de pâturage et s'ensilage ( Sherbon, 1999). Les principaux systèmes du lait qui déterminent son potentiel redox sont l'oxygène dissous, l'acide ascorbique et la riboflavine. Le système thiol-ponts disulfures, lorsqu'activé par traitement thermique, contribue également (Walstra et Jenness, 1984). D'autres systèmes pourraient influencer le potentiel redox du lait mais ne sont pas réversibles. C'est le cas du système lactate-pyruvate qui n'est pas réversible à moins d'être activé par des enzymes et qui est présent en quantité négligeable dans le lait frais. Le groupement aldéhyde du lactose est oxydable en carboxyl à pH élevé, mais ce n'est pas un système réversible qui contribue au potentiel redox à pH 6,6 (Walstra et Jenness, 1984). En dernier lieu, certaines enzymes oxydo-réductrices pourraient jouer un rôle mineur. 3.4 Oxygène dissous L'importance de l'oxygène dissous dans la valeur du potentiel redox du lait se démontre de plusieurs façons. Tout d'abord, la diminution du potentiel redox du lait à environ -0,15 mV suite au barbotage d'azote pour remplacer l'oxygène est observée (Walstra et Jenness, 1984). Ensuite, le bleu de méthylène est réduit par du lait tiré de façon anaérobe mais cette capacité réductrice est perdue lorsqu'un tel lait est exposé à l'air. Finalement, la baisse du potentiel redox et le retour de la capacité réductrice sont observés quand des bactéries telles S. lactis ou des lactobacilles croissent dans le lait. Cependant, dans ce dernier cas, la croissance bactérienne cause une diminution de pH qui contribue à l'augmentation du potentiel redox (Walstra et Jenness, 1984). La Figure 24 présente la chute de potentiel redox suite à la consommation d'oxygène et la production de réducteurs par une bactérie lactique, conférant au fromage et laits fermentés des potentiels négatifs (Fox et McSweeney, 1998; Sherbon, 1999). Selon l'espèce bactérienne, le potentiel redox peut chuter jusqu'à -100 ou -200 mV (Walstra et al. , 1999)
Figure 24. Diminution du potentiel redox du lait pendant l'incubation avec une souche de S. Lactis à 25 °C (Fox et McSweeney, 1998; Sherbon, 1999).
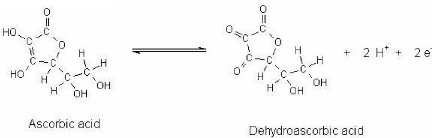
3.4.1 Acide ascorbique Le contenu en acide ascorbique (Figure 25 ) du lait frais est d'environ 11,2 à 17,2 mg/L et se présente entièrement sous la forme réduite lorsque tiré du pis. L'oxydation réversible en acide déhydroascorbique a lieu à un taux qui dépend de la température et de la concentration en cuivre et oxygène (Walstra et Jenness, 1984; Singh et al. ,1997). L'acide ascorbique stablilise le potentiel redox du lait désaéré à 0 mV et celui du lait oxygéné à 200-300 mV (Fox et McSweeney, 1998; Sherbon, 1999). L'acide ascorbique est préservé dans le lait par la prévention de la contamination au cuivre et au fer et par désaération. Des études ont démontré une corrélation inverse significative entre le potentiel redox et le contenu en acide ascorbique réduit dans le lait frais préalablement équilibré avec l'air (Walstra et Jenness, 1984).
Figure 25. Acide ascorbique et déhydroascorbique.
3.4.2 Riboflavine La concentration de riboflavine libre dans le lait est d'à peine 4 uM et sa structure est donnée à la Figure 26 . Son importance dans le phénomène redox réside dans son rôle dans la photo-oxydation plutôt que par un apport significatif au potentiel redox (Walstra et Jenness, 1984). Figure 26. Formes oxydées et réduites de la riboflavine.
3.4.3 Système thiol-ponts disulfures Le lait ne contient que de faibles concentrations de thiols de faible poids moléculaire, telle la cystéine libre, et n'ont pas d'influence significative sur le potentiel redox du lait. Les thiols de protéines natives, soit ceux de la â-LG, ne sont pas actifs dans les systèmes redox mais sont activés quand les protéines sont ouvertes et déplissées par dénaturation thermique. Le potentiel standard des thiols du lait liés à la protéine n'est pas connu et le système pourrait ne pas être facilement réversible (Walstra et Jenness, 1984). 3.4.4 Enzymes oxydoréductrices Les enzymes oxydoréductrices du lait sont en temps normal très peu impliquées dans les réactions redox du lait. La lactate dehydrogénase et la malate déhydrogénase sont des enzymes dépendantes au NAD mais il n'y a pas de preuve de la participation ces enzymes dans les réactions redox. (Walstra et Jenness, 1984).
La xanthine oxydase (XO) est une métalloprotéine présente dans le lait bovin à concentration de 35 mg/L. Elle est principalement associée à la membrane des globules de gras et produit H2 O2 ou O2 ?- si ses substrats, que sont la xanthine, l'hypoxanthine et des aldéhydes, lui sont founis. Telle qu'isolée du lait, la XO utilise l'oxygène moléculaire comme un accepteur d'électrons, mais si une partie de ses disulfures sont réduits par le dithiothretol elle devient une déhydrogénase dépendante du NAD+. La réoxydation des thiols en disulfures avec l'enzyme laitière sulphydryl oxydase rétablit la spécificité de l'enzyme pour l'oxygène comme accepteur d'électrons. L'activité de la XO dans le lait frais est accrue d'environ 4 fois par le rangement à 4 °C, le chauffage à 70 °C pendant 5 minutes, l'homogénéisation ou l'incubation avec des préparations commerciales de protéinase ou lipase parce que de tels traitements transfèrent une grande partie de l'enzyme des globules de gras vers le sérum ( Walstra et Jenness, 1984). Les études sur la possibilité que la XO soit impliquée dans le développement des saveurs oxydées du lait n'ont pas encore donné de résultats probants (Walstra et Jenness, 1984). La catalase qui s'accumule dans la couche de crème catalyse la décomposition du H2 O2 en eau et oxygène (Walstra et Jenness, 1984). La lactoperoxydase (LP) peut composer jusqu'à 1 % des protéines sériques du lait. Son activité dans le lait augmente au cours de la lactation pour atteindre un maximum à 40 jours post-partum et diminuer ensuite. Elle catalyse l'oxydation par l'H2 O2 d'une longue liste de composés donneurs d'électron, incluant des amines aromatiques, des phénols, des acides aromatiques, la tyrosine et tryptophane, l'acide ascorbique, des nitrites, des thiocyanates et d'autres encore. Le thiocyanate est un constituant naturel du lait. Son oxydation mène à un produit inhibiteur de certaines bactéries (Walstra et Jenness, 1984). La superoxyde dismutase (SOD) catalyse la dismutation de l'ion superoxyde O2 ?- en H2 O2 et O2 . La structure primaire est connue, elle possède un thiol libre et un lien disulfure. Ce pourrait être un important antioxydant protégeant les constituants du lait de l'oxydation par l'ion superoxyde généré par les oxydations catalysées par la xanthine oxydase et la lactoperoxydase ainsi que des photo-réactions de la riboflavine (Walstra et Jenness, 1984). La sulphydryl oxydase (SO), présente dans le lait bovin à raison de 3 mg/L, catalyse l'oxydation des thiols à la fois dans les petits composés et les protéines en disulfures en utilisant l'oxygène moléculaire comme accepteur d'électron. C'est une oxydase anaérobe, réduisant l'oxygène en H2 O2 plutôt qu'en eau selon l'équation suivante : 2 RSH + O2 ? RSSR + H2O2 De la SO immobilisée sur billes de verre a été utilisée dans le lait UHT dans le but d'atténuer la saveur de « cuit » puisqu'elle pourrait catalyser l'oxydation des thiols en disulfures dans le lait (Walstra et Jenness, 1984). Finalement, quelques unes de ces enzymes sont des
métalloprotéines (xanthine
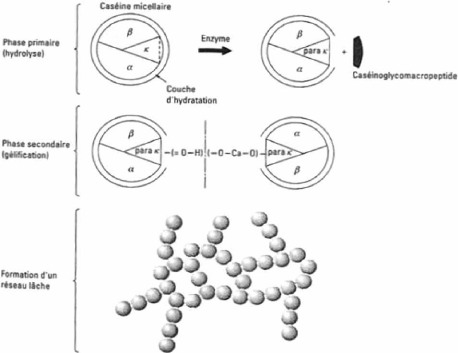
causée par le chauffage mais ceci n'est pas une activité enzymatique (Walstra et Jenness, 1984). 4 Comportement du lait face à différents traitements Les différents procédés industriels appliqués au lait visent à assurer la qualité et la stabilité des produits. De ces procédés, les traitements de chaleur sont très utilisés en technologie laitière. L'aptitude du lait à supporter les hautes températures est une caractéristique technologique importante, tout comme l'aptitude à la fabrication fromagère (Robitaille, 1995). 4.1 Coagulation par la présure Diverses enzymes protéolytiques ont la capacité de coaguler le lait mais la présure est la plus utilisée. La présure est constituée de deux enzymes, soit la chymosine, qui permet l'hydrolyse de la caséine K, et la pepsine (Ruettiman et Ladisch, 1987). La coagulation du lait par la présure est divisée en trois étapes : la phase d'hydrolyse enzymatique, la phase d'agrégation et la phase de formation du gel (Brown et Ernstrom, 1988). Elles sont décrites plus en détail ci-après et illustrées à la Figure 27 . 4.1.1 Phase d'hydrolyse enzymatique Lors de cette première étape, l'enzyme vient couper le lien peptidique Phe105 -Met106 de la caséine K et la protéine est scindée en deux peptides, le CMP et la para-k-CN, dont les propriétés ont été énoncées à la section 1.1.1.1 . Le mode d'action de l'enzyme n'est pas encore bien défini. Il est possible que l'enzyme s'attaque à une micelle, en suive la surface et hydrolyse toutes les caséines K sur son passage ou bien qu'après l'attaque d'une molécule de caséine K elle retourne en solution pour diffuser jusqu'à la prochaine micelle (Ruettimann et Ladisch, 1987). L'hydrolyse progressive de la caséine K durant la phase primaire altère les propriétés des micelles à un point où elles deviennent susceptibles à l'agrégation, qui représente la seconde phase de la réaction (Lucey, 1995). 4.1.2 Phase d'agrégation Lors de la seconde phase, les micelles déstabilisées peuvent se rapprocher et former des liens hydrophobes. Ceci est possible parce que les charges en surface des micelles diminuent lorsque le CMP est libéré. La perte de ce segment réduit le potentiel zeta de la micelle d'environ 30 à 50 % et diminue les interactions de nature entropique (stérique), ce qui permet un rapprochement des micelles attaquées par l'enzyme et facilite l'agrégation (Brulé et al ., 1997; Lucey, 1995; Singh et Fox, 1989; Dalgleish, 1981). L'agrégation devient possible lorsqu'un certain degré d'hydrolyse est atteint puisqu'il existe un niveau minimum de caséine K nécessaire à la stabilisation de la micelle (Horne et al ., 1993). Selon les auteurs, l'agrégation commence lorsque de 60 à 90 % de la caséines K est hydrolysée. Le temps écoulé pour l'atteinte de cette phase se nomme temps de coagulation par la présure ou RCT (pour «Rennet Coagulation Time ») (Dalgleish, 1981). En pratique, le RCT est défini comme le temps écoulé jusqu'à l'observation visuelle de floculation.
4.1.3 Phase de formation du gel La troisième phase mène à la formation d'un réseau tridimensionnel continu nommé gel. Les agrégats augmentent d'abord de taille. Par la suite, la réticulation entre les chaînes et la fusion des particules transforment le lait en gel (Ruettiman et Ladisch, 1987). Le caillé se raffermit et la synérèse débute. Figure 27. Phases du processus de coagulation enzymatique du lait (St-Gelais et Tirard-Collet, 2002).
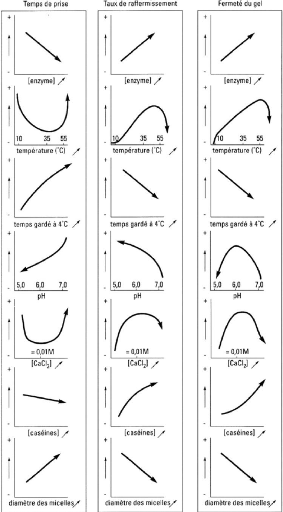
4.2 Facteurs influençant la coagulation La production de coagulum satisfaisant est importante dans la fabrication fromagère car la nature du caillé formé détermine la qualité du produit final. De nombreux facteurs influencent la coagulation du lait. Ce sont la composition, la température, le pH, l'ajout de sels, la concentration en présure et le diamètre micellaire. Les effets des facteurs principaux qui influencent les paramètres de coagulation sont résumés à la Figure 28 . Figure 28. Facteurs influençant les paramètres de coagulation enzymatique du lait (St-Gelais et TirardCollet, 2002).
4.2.1 Composition du lait La composition du lait influence la coagulation du lait de différentes façons. Une augmentation du niveau de caséines dans le lait résulte en un temps de coagulation plus court, un taux de raffermissement plus élevé et un caillé plus ferme au temps de coupage (Hill, 1995). Une augmentation du degré de glycosylation de la caséine ê augmente la vitesse de raffermissement du gel sans toutefois influencer le RCT (Robitaille et al ., 1995). Du point de vue bactériologique, un compte en cellules somatiques supérieur à 500 000 cellules/ml est caractérisé par une augmentation du RCT et une pertes de fines plus importante menant à une baisse de rendement (Roupas, 2001). 4.2.2 Température d'emprésurage et de coagulation L'activité enzymatique est influencée par la température. L'activité de la présure est donc reliée à la température à laquelle les étapes d'emprésurage et de coagulation sont conduites (St-Gelais et Tirard-Collet, 2002). La présure montre une activité maximale entre 30 et 42 °C et est inhibée à des températures supérieures à 55 °C (St-Gelais et Tirard-Collet, 2002). Le Tableau 29 résume l'influence de la température au moment où l'enzyme est ajoutée sur la coagulation. Tableau 29. Impact de la température sur la coagulation du lait par la présure (St-Gelais et Tirard-Collet, 2002).
4.2.3 Traitement thermique du lait Les traitements thermiques appliqués au lait altèrent ses propriétés coagulantes, ce qui rend le lait chauffé inapte à la fabrication fromagère. La fabrication fromagère est réalisée de préférence sur des laits non chauffés ou ayant subi une thermisation (Vasbinder et al. , 2003). Ensuite, les traitements thermiques imposés au lait modifient les cinétiques de coagulation. D'une part, l'entreposage du lait au froid conduit à une augmentation du temps de prise puisque les minéraux de la phase colloïdale sont solubilisés (Raynal et Remeuf, 2000). D'autre part, il est connu que le lait surchauffé prend plus de temps à coaguler (Moir et al. , 1930, 1931). La littérature explique la baisse de l'aptitude à
la coagulation des laits chauffés par une
à la précipitation du phosphate de calcium et la stabilisation de la micelle de caséine grâce au recouvrement avec des protéines sériques dénaturées chargées positivement (Vasbinder et al. , 2003). Ensuite, les études démontrent qu'une augmentation de la température de chauffage se traduit par une augmentation du RCT qui augmente de 10 à 15 fois pour les laits chauffés (Vasbinder et al ., 2003; Dalgleish, 1990; Singh et al. , 1988). Un traitement thermique plus sévère que la pasteurisation se traduit par une augmentation du temps de coagulation et à des gels plus faibles. Finalement, lors d'un traitement thermique très sévère (>90 °C, 10 min), le lait perd l'aptitude à coaguler sous l'action de la présure (Singh et al. , 1988). Un des effets du chauffage est le déplacement des équilibres ioniques vers les formes insolubles. Les modifications de composition de la surface des micelles et la précipitation des phosphates diminuent la stabilité de la phase colloïdale. La diminution de calcium soluble rend plus difficile la création de liaisons entre les micelles lors de la formation du gel. Les temps de coagulation allongent alors que la fermeté du caillé et la vitesse d'égouttage diminuent (St-Gelais et Tirard-Collet, 2002). En ce qui a trait à l'augmentation du temps de prise, la formation du complexe f3-LGic-CN serait responsable de la baisse de sensibilité à la chymosine (Morgan et al. , 2000; Lucey, 1995; Singh et al ., 1988). Il a été démontré que le temps de coagulation augmente avec l'augmentation du degré de dénaturation de la f3-lactoglobuline et avec le pourcentage de cette protéine associé aux micelles (Singh et Waungana, 2001). La complexation des protéines du lactosérum dénaturées avec la caséine-ic modifie la conformation de la ic-CN et rendrait le site d'hydrolyse moins accessible pour l'enzyme. De plus, une bonne partie de la f3-lactoglobuline ne s'associe pas et se présente sous forme d'agrégats dans le sérum qui pourraient interférer dans les processus d'agrégation et de formation du réseau, d'où les gels plus faibles (Waungana et al ., 1996; Lucey, 1995; Lucey et al ., 1993; McMahon et al ., 1993; Dalgleish, 1990). Toutefois, l'équipe de Vasbinder et coll. (2003) soutient que le début de la coagulation, soit le relâchement du CMP par hydrolyse enzymatique, n'est pas affecté par le chauffage. En effet, l'application d'un traitement thermique à 90 °C pendant 10 minutes au lait s'est traduit par une diminution de 6 % du CMP relâché, diminution insuffisante pour inhiber l'agrégation des micelles (Vasbinder et al ., 2003). L'agrégation des micelles a lieu indépendamment du traitement thermique appliqué cependant les micelles perdent la capacité de former un gel. Cette observation correspond à un temps de coagulation plus élevés et une diminution du module d'élasticité (G') tel que rapportés dans la littérature et qui sont attribués à une diminution de l'activité enzymatique, la précipitation du calcium et la dénaturation des protéines sériques (Vasbinder et al ., 2003). Puisque les travaux ne démontrent aucun effet du traitement thermique sur l'hydrolyse enzymatique, la précipitation du phosphate de calcium par la chaleur comme facteur influençant l'hydrolyse enzymatique est exclue (Vasbinder et al ., 2003). L'altération des propriétés coagulantes du lait chauffé est entièrement attribuée aux protéines sériques dénaturées
qui enrobent les micelles de caséine puisqu'en absence de protéines sériques, les micelles de caséines du lait chauffé s'agrègent (Vasbinder et al ., 2003). 4.2.4 pH L'influence du pH est double. Tout d'abord, le pH optimal de la chymosine se situe entre 5,3 et 5,5 (Lenoir et al. , 1997; Fox, 1993). L'acidification du lait à pH de 6,3 améliore les propriétés de coagulation à la présure (Renault et al. , 2000). Il y a augmentation de la vitesse d'hydrolyse enzymatique, réduction du RCT et formation d'un gel plus ferme (Roupas, 2001; Lenoir et al. , 1997; Horne et Muir, 1994). Ensuite, l'acidification mène à la neutralisation des charges de répulsion puisque le nombre de charges positives augmente, ce qui déstabilise la micelle et permet davantage d'attraction électrostatique entre les micelles (Renault et al , 2000). La diminution du pH mène également à la solubilisation du phosphate de calcium micellaire et fait augmenter le calcium soluble. Le calcium qui se lie à la surface des micelles contribue à modifier leur charge nette en plus d'accroître l'efficacité des collisions entre l'enzyme et le substrat (Roupas, 2001). 4.2.5 Concentrations en sels La composition minérale du lait va affecter la coagulation et tout particulièrement la quantité de calcium ajouté au lait. En effet, le temps de prise diminue tandis que le taux de raffermissement et la fermeté du gel vont augmenter jusqu'à des concentrations de CaCl2 d'environ 0,01 M. On attribue cet effet à l'augmentation d'ions calcium. Cependant, à des concentrations supérieures, on observe un phénomène inverse pour chacun des paramètres de coagulation (St-Gelais et TirardCollet, 2002). Le calcium n'est pas requis dans la phase initiale d'hydrolyse enzymatique mais est essentiel à l'agrégation des micelles. L'ajout de CaCl2 en technologie fromagère stimule la coagulation, le raffermissement du caillé et permet de réduire les pertes dans le lactosérum, tout en diminuant le pH et réduisant la charge de la micelle (Roupas, 2001). L'addition de NaCl, pour sa part, augmente la force ionique et cause une déminéralisation de la micelle. L'augmentation de la force ionique entraîne une modification des charges de la chymosine et de la ê-caséine, ce qui entrave l'attraction entre l'enzyme et son substrat (Roupas, 2001). 4.2.6 Concentration présure L'activité de l'enzyme dépend du pH, de la température et la concentration en calcium, sujet qui ont été traités précédemment. Il existe une corrélation linéaire entre la concentration en présure et le temps de coagulation, ce dernier devenant plus court à mesure que la concentration en présure augmente (Horne et Muir, 1994). Le taux de raffermissement et la fermeté du gel augmentent quant à eux avec la concentration en présure (St-Gelais et Tirard-Collet, 2002).
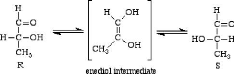
4.2.7 Diamètre micellaire La coagulation enzymatique est un phénomène de surface et elle est influencée par le diamètre des micelles de caséine. En général, plus le diamètre est grand, plus le temps de prise est long et plus le taux de raffermissement et la fermeté finale sont faibles (StGelais et Tirard-Collet, 2002). En fromagerie, de petites micelles seront donc recherchées puisqu'elles sont associées à un temps de prise plus court et un taux de raffermissement plus rapide (Park et al. , 1999). De plus, les micelles de petites tailles fusionnent plus étroitement entre elles et sont donc caractérisées par la formation d'un réseau protéique plus dense, plus cohérent et plus ferme que celles de grande taille (Roupas, 2001; Lenoir et al. , 1997; Delacroix-Buchet et al ., 1993). 4.3 Traitement thermique Les divers traitements thermiques appliqués au lait en industrie, tels que la pasteurisation et la stérilisation, peuvent avoir des effets dénaturants sur les protéines du lait. Des réactions de dégradation des protéines, de dégradation du lactose et des modifications de l'équilibre minéral sont observées. En effet, lors du chauffage, des réactions de dénaturation et d'agrégation de protéines se produisent et le phosphate de calcium précipite. 4.3.1 Potentiel redox Les impacts des traitements thermiques sur le potentiel redox du lait ont été considérablement étudiés jusqu'au milieu du siècle dernier. Une baisse marquée du potentiel coïncide avec la libération des groupements thiols par dénaturation des protéines, principalement la â-lactoglobuline. ( Sherbon, 1999). Tout d'abord, le chauffage du lait en présence d'oxygène accélère l'oxydation des thiols et la destruction du système acide ascorbique-dehydroascorbique. Ensuite, le chauffage favorise la réaction de Maillard entre le lactose et les protéines et la production de réducteurs de type énediol (Figure 1.14 ) (Walstra et Jenness, 1984 ). Figure 30. Formation d'un intermédiaire énediol.
Ces diverses réactions diffèrent d'énergie d'activation. En conséquence, la balance globale du système redox après traitement thermique dépend grandement du temps et de la température utilisés. De plus, le taux de production de réducteurs par réaction de Maillard augmente avec la concentration en réactifs. Il y a donc davantage de ces composés réducteurs résultant du traitement thermique appliqué dans la concentration et le séchage du lait que de ceux utilisés dans les laits non-concentrés (Walstra et Jenness, 1984 ). Cependant, la désaération et traitements thermiques HTST résultent en un potentiel redox minimal et produisent des poudres de lait de qualité supérieure contre la dégradation produisant une saveur oxydée ( Sherbon, 1999).
Des études, rapportées par Walstra et Jenness (1984), sur la capacité réductrice et le potentiel redox du lait sous différentes conditions montrent que le potentiel redox du lait témoin se situe à +315 mV. Lorsque ce lait est chauffé à 85 °C pendant 30 minutes, le potentiel redox diminue à +200 mV. Cependant, lorsque l'atmosphère n'est plus de l'air ambiant mais de l'azote, le potentiel redox chute à 0 mV, mettant en évidence l'importance des gaz dissous sur le potentiel redox des laits chauffés. 4.3.2 Dégradation des protéines Alors que les caséines sont très résistantes à la chaleur, les protéines sériques le sont moins. Malgré cette résistance, les caséines peuvent tout de même être affectées et déstabilisées par la chaleur. (O'Connell et Fox, 2000). 4.3.2.1 Dénaturation des protéines sériques Les différentes protéines sériques n'ont pas la même résistance face aux traitements thermiques. L'ordre de sensibilité à la chaleur est immunoglobulines > sérumalbumines > f3-lactoglobuline > B1;-lactalbumine (Corredig et Dalgleish, 1996). Le chauffage des protéines sériques entraîne l'ouverture des structures secondaire et tertiaire qui permet l'agrégation de ces protéines entre elles ou avec les micelles de caséine. Le chauffage mène à une perte de la structure native de la protéine. La dénaturation des protéines sériques résulte en le déplissement de leurs polypeptides, exposant ainsi des résidus habituellement cachés à l'intérieur de la structure globulaire (Singh et Waungana, 2001; Calvo, 1995). Dans le cas de la f3-lactoglobuline, ce processus intramoléculaire qui expose le coeur hydrophobe expose en même temps le groupement thiol libre ainsi que des ponts disulfures très réactifs (Fryer et al ., 1995). Les protéines déplissées présentent donc une configuration instable qui est ensuite stabilisée par la phase d'agrégation. La dénaturation de la f3-lactoglobuline est réversible jusqu'à ce que l'agrégation prenne place tandis que la dénaturation de l'B1;-lactalbumine est réversible jusqu'à 85 °C (Kinsella, 1984). La disponibilité de nouveaux sites exposés par le déplissement des protéines permet la formation d'agrégats constitués exclusivement de protéines sériques ou d'un mélange avec les caséines. Le fait que la f3-lactoglobuline s'associe avec la micelle ou bien forme des agrégats exclusivement composés de protéines sériques dépend de plusieurs facteurs tels que la durée et l'intensité du chauffage, le pH, la force ionique, la concentration en calcium et en phosphate solubles ainsi que le degré de dissociation de la caséine K de la micelle (Devold et al ., 2000). L'B1;-lactalbumine, bien qu'elle participe aussi aux interactions, ne se lie pas directement aux caséines, elle le fait plutôt par l'intermédiaire de la f3-lactoglobuline (Dzurec et Zall, 1985). En effet, il a été démontré que l'B1;-lactalbumine ne polymérise pas en l'absence des autres protéines sériques, même si des caséines sont présentes (Calvo, 1995). De plus, la présence de f3-lactoglobuline est nécessaire à l'interaction de l'B1;-lactalbumine avec les micelles. En effet, en dessous de 90 °C, l'B1;-lactalbumine et la f3- lactoglobuline s'associent pour former des complexes intermédiaires qui eux s'associent ensuite aux micelles (Corredig et Dalgleish, 1999). L'attachement de
protéines sériques dénaturées à la surface de la micelle, fait l'objet de la prochaine section. 4.3.2.2 Formation d'un complexe entre la â- LG et la caséine ê Les groupements thiols libres présents dans la structure native de la â-lactoglobuline sont à l'origine des complexes formés entre les protéines sériques et les micelles de caséine sous l'effet de la chaleur. Durant le traitement thermique, il y a interaction entre la â-lactoglobuline dénaturée et la caséine ê située à la surface de la micelle et formation d'un complexe stabilisé par des interactions mettant principalement en cause les groupements thiols mais aussi des groupements hydrophobes et ioniques (Devold et al ., 2000; Singh, 1995; Hill, 1989). Le chauffage à pH inférieur à 6,7 mène à une plus grande quantité de complexes formés avec les micelles alors qu'à pH supérieur à 6,7, les complexes se dissocient de la micelle à cause de la dissociation de la caséine ê elle-même (Singh et Waungana, 2001). L'étendue des interactions augmente avec la durée et l'intensité du traitement thermique appliqué (entre 75 et 90 °C) et l'interaction entre la â-lactoglobuline et la caséine ê atteint un plateau laissant croire qu'une quantité limitée de sites sont disponibles pour l'interaction (Corredig et Dalgleish, 1999). Il est bien connu aussi que la formation du complexe entre la â-LG et la ê-CN est responsable de la baisse de sensibilité à la chymosine (Morgan et al. , 2000). Quand la â-LG et la ê-CN sont chauffées ensemble entre 80 et 90 °C, elles interagissent par la formation des liens S-S. Morrissey suggérait dès 1969 que les interactions entre la âLG et les caséines au chauffage affectent de façon marquée la coagulation à la présure du lait (Morrissey et al ., 1969; Ustumol et Brown, 1985). 4.3.3 Dégradation du lactose Le lactose est impliqué dans deux types de réactions de dégradation dans le lait, soit l'isomérisation et la réaction de Maillard.Alors que la première est plus importante d'un point de vue quantitatif à des températures supérieures à 100 °C, c'est la seconde qui prédomine à des températures inférieures (van Boekel, 1998 ; Berg et van Boekel, 1994). 4.3.3.1 Isomérisation du lactose Le lactose est sujet à des réactions d'isomérisation dans le lait (Berg et van Boekel, 1994, van Boekel, 1996). La principale dégradation est celle de Lobry-de-BruinAlberda-van-Ekenstein où les protéines agissent comme catalyseur. Les produits de la réaction sont le lactulose et l'épilactose, des disaccharides formés de galactose et de, respectivement, fructose et mannose. L'énolisation du lactulose mène ensuite à la formation de produits de dégradation tels le galactose, le tagatose, l'acide formique ainsi que différents composés carbonés. L'acide formique formé en grande quantité est largement responsable de la baisse de pH induite par la chaleur, de concert avec l'altération l'équilibre salin.
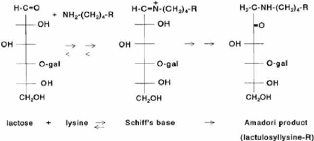
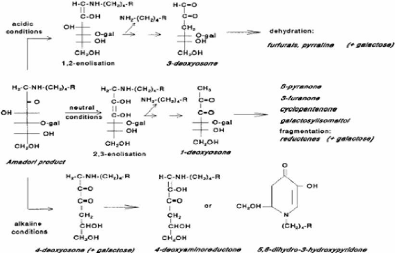
4.3.3.2 Réaction de Maillard La réaction de Maillard est une réaction chimique des groupements aminés et des sucres réducteurs qui mène à la formation de composés bruns. Ce brunissement non enzymatique est influencé par la température, le pH, l'activité de l'eau et la présence de certains sels et vitamines. Quoique cette réaction soit désirable dans certains produits, elle est le plus souvent indésirable en transformation laitière (Walstra et Jenness, 1984). Dans le cas du lait, les groupements aminés en cause sont principalement des résidus lysine dans les protéines laitières puisque le contenu en acide aminé est très faible (Walstra et Jenness, 1984). Les résidus lysine des caséines semblent être plus réactifs que les protéines sériques et la ê-CN semble être la caséine la plus réactive (van Boekel, 1998 ). Le sucre réducteur du lait est le lactose (van Boekel, 1998 ). Dans le but de systématiser l'ensemble des réactions impliquées, une classification en trois stades est généralement utilisée. Récemment toutefois, une classification conceptuelle a été proposée par Yaylayan (1997). La classifications usuelle en trois stades est utilisée ici. Le stade initial, illustré à la Figure 31 , comprend la condensation du sucre réducteur avec le groupement aminé et donne, via la base de Schiff et le réarrangement d'Amadori (transformation d'un aldose en cétose), le composé d'Amadori. Dans le lait, le composé d'Amadori est la lactulosyllysine, attachée aux protéines. Au stade avancé, illustré à la Figure 32 , il y a brisure du composé d'Amadori qui, par les voies que sont l'énolisation, la cyclisation et la déshydratation, mènent à des produits de réaction différents. Le stade final, moins bien caractérisé d'un point de vue chimique, comprend la condensation de composés aminés et de fragments de sucre en protéines polymérisées et pigments bruns, appelées mélanoïdines (Van Boekel, 1998). Les principaux produits de dégradation de la lactulosyllysine liée à des protéines dans le lait sont le galactose et l'acide formique (Berg et van Boeckel, 1994). Figure 31 . Aperçu du stade initial de la réaction de Maillard (gal = galactose, R = chaîne protéique) (van Boekel, 1998).
Figure 32. Dégradation du composé d'Amadori au stade avancé de la réaction de Maillard sous conditions acides, neutres et alcalines (gal = galactose, R = chaîne protéique) (van Boekel, 1998) .
L'un des facteurs, nommés ci-avant, intervenant dans le développement de la réaction de Maillard est le pH. L'augmentation du pH de 6,6 à 7,0 se traduit par une augmentation de la formation de lactulose, composée intervenant dans la réaction de Maillard puisque l'isomérisation des sucres réducteurs est favorisée à pH élevé (Berg, 1993). Les conséquences de la réaction de Maillard dans le lait sont importantes et comprennent la perte de valeur nutritive car les résidus lysine sont bloqués et non assimilés, une digestibilité réduite et l'inhibition d'enzymes, la production de saveur, la formation de composés anti-oxydants mais aussi de composés mutagènes, antimutagènes et anticancérigènes, la formation possible de composés antibactériens, la diminution de l'allergénicité, la polymérisation des protéines du lait ainsi que le développement de coloration brune à cause des mélanoïdines (van Boekel, 1998). 4.3.3.3 Acides organiques L'isomérisation du lactose ainsi que la réaction de Maillard mènent tous deux à la formation d'acide formique, un produit de la dégradation du galactose. Un chauffage intense peut aussi mener à la formation d'acide acétique, lactique, propionique, butyrique et laevulinique (Berg et van Boekel, 1994 ; Walstra et Jenness, 1984).
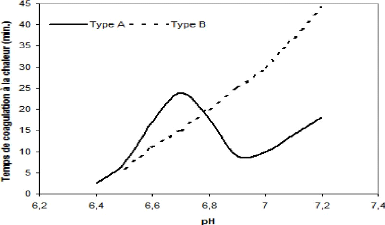
4.3.4 Modification de l'équilibre minéral Les traitements thermiques appliqués au lait entraînent la réduction du calcium total soluble et du calcium ionique dans le lait. Le phosphore soluble subit des changements similaires (Ustunol et Brown, 1985). L'équilibre entre le phosphate de calcium colloïdal et soluble affecte l'équilibre des constituants de caséine et leur degré d'association (Ustunol et Brown, 1985). 4.3.5 Modification du pH L'application d'un traitement de chaleur cause l'abaissement du pH suite à trois réactions. Ce sont l'oxydation thermique du lactose en acides organiques (50 % de l'abaissement de pH), l'hydrolyse du phosphate organique (phosphosérines) (30 %) et enfin la précipitation du phosphate de calcium tricalcique et le relâchement concomitant d'ion H+ (20 %) (Singh, 2004). 4.3.6 Stabilité thermique L'augmentation de la stabilité thermique du lait est désirable dans plusieurs applications alimentaires dans le but, par exemple, d'assurer la vie de tablette des laits stérilisés en évitant la gélification à l'entreposage (Singh, 2004). La stabilité du lait à chaleur réfère à sa résistance à la coagulation à des températures de stérilisation (Singh et Creamer, 1992) alors que Singh définissait la stabilité thermique comme étant la capacité du lait de résister aux hautes températures des procédés industriels sans coagulation visible ou gélification (Singh, 2004). Le facteur le plus important vis-à vis de la stabilité est le pH. Les autres facteurs qui influencent la stabilité thermique sont les sels, les protéines du lait (Morgan et al. , 2000; Singh et Creamer, 1992) ainsi que les traitements tels le préchauffage, la concentration et l'homogénéisation, qui peuvent causer la coagulation partielle ou complète pendant la transformation ou l'entreposage (Singh et Creamer, 1992, Singh, 2004). La façon la plus commune de mesurer la stabilité thermique du lait est d'établir le profil du temps de coagulation à la chaleur en fonction du pH (HCT-pH) tel que présenté à la Figure 33. Le lait est classé en deux types selon son profil de stabilité. Le lait de type A, prédominant au Québec, possède un minimum de stabilité à pH 6,9 et un maximum à pH 6,7 alors que la stabilité thermique du lait de type B augmente continuellement en fonction du pH (Singh, 2004; O'Connell et Fox, 2000; Singh et Creamer, 1992).
Figure 33. Profil de stabilité à la chaleur en fonction du pH (Adapté de Singh, 2004).
Le minimum de stabilité peut être supprimé en modifiant la composition du lait. Une légère diminution de la concentration totale en ions calcium et magnésium, de l'ordre de 11 à 13 mM, élimine le minimum alors qu'une augmentation diminue la stabilité pour tous les pH. L'addition de phosphates augmente le HCT, et la diminution de phosphate soluble déplace le profil vers les valeurs alcalines. L'addition de citrate a pour effet de déplacer le maximum vers des valeurs plus acides. Le retrait de 40 % du phosphate de calcium colloïdal augmente le HCT entre pH 6,4 et 7,4 alors que le retrait plus prononcé, de l'ordre de 60 à 100 %, n'augmente le HCT qu'entre pH 6,4 et 7,0 et présente un effet déstabilisant à pH plus élevé (Singh, 2004). L'urée est le seul constituant indigène du lait dont la présence corrèle fortement avec les variations normales de stabilité à la chaleur. L'addition à de faibles concentrations n'affecte pas le HCT dans la région du maximum mais à forte concentration elle augmente le HCT (Singh, 2004). De plus, il est bien établi que l'urée est apte à stabiliser à la chaleur le lait de type A dans la région du pH de stabilité maximale et du lait de type B à travers les pH de 6,4 à 7,4 (Holt et al., 1978). La relation entre les protéines du lactosérum et la stabilité à la chaleur est bien établie pour le lait de vache : la â-LG et la ê-CN sont les protéines qui ont le plus d'influence sur le profil du temps de coagulation à la chaleur (HCT). De plus, la â-LG est nécessaire au développement d'un profil de type A. En effet, la stabilité des micelles de caséines en absence de protéines sériques augmente continuellement avec le pH alors que l'addition de â-LG à une dispersion de micelles de caséines introduit un minimum et un maximum dans le profil de stabilité. D'autre part, l'enrichissement du lait en ê-CN accroît la stabilité dans la région du minimum et convertit un lait de type A en lait de type B (Singh, 2004).
De nombreux additifs peuvent modifier le profil de stabilité à la chaleur en fonction du pH. Leimyms agents bloqueurs de thiols tel le N-ethylmaleimide réduisent le HCT dans la région du max. L'addition d'agents réducteurs, tel le 2-mercaptoéthanol, déstabilise le lait sur toute la gamme de pH. L'addition d'agents oxydants, tel le KBrO4 , éliminent le minimum dans la courbe HCT-pH. Le KIO3 , pour sa part, a un effet stabilisant marqué à travers la gamme de pH 6,5 à 7,3. Le formaldéhyde augmente la stabilité, particulièrement dans la région du minimum qui est éliminé. Les détergents anioniques, comme le SDS, augmentent la stabilité dans la région du minimum mais déplacent aussi la courbe vers des pH plus élevés. Les détergents cationiques, tel le cetylmethylammonium, déplacent aussi la courbe dans le même sens mais augmentent au contraire la stabilité dans la région du maximum. Finalement, les extraits riches en polyphénols, comme l'acide caféique, contribue à augmenter le HCT, surtout dans la région du minimum (Singh, 2004). 4.3.7 Coagulation à la chaleur La coagulation du lait suite à un chauffage prononcé à des températures entre 120 et 140 °C est une conséquence de la perte de stabilité de la micelle résultant de nombreux changements physiques et chimiques de ses composants. Les traitements de chaleur modifient à la fois les micelles de caséines et l'environnement de la phase sérique autour d'elles. Les changements qui ont lieu dans les micelles de caséines elles-mêmes sont l'association des protéines sériques, les changements des équilibres minéraux, la déphosphorylation et la dissociation des caséines. Dans la phase sérique, des changements de pH et la concentration des sels solubles sont observés ainsi que l'hydrolyse du lactose. (Singh, 2004). 4.3.7.1 Changement dans les micelles de caséines Le pH de chauffage a une grande influence sur l'étendue de l'association des protéines sériques aux micelles de caséine. À des valeurs de pH inférieures à 6,8, une vaste majorité des complexes de protéines sériques demeure associée à la surface de la micelle de caséine. À des valeurs de pH plus élevées, les agrégats de protéines sériques demeurent dans le sérum et la ê-CN se dissocie de la micelle. Singh mentionne que le mécanisme exact n'est pas encore élucidé. Il serait possible que les interactions entre la â-LG et la ê-CN aient lieu dans la micelle et que le complexe se dissocie dans la phase sérique ou encore que le complexe soit formé dans le sérum. Néanmoins, la présence de protéines du lactosérum accroît de façon marquée la dissociation de la êCN micellaire à des valeurs de pH>6,9. D'autres caséines (ás1 , ás2 et â) se dissocient de la micelle lors du chauffage mais de façon beaucoup moins marquée (Singh, 2004). Selon le pH au chauffage, deux types de particules de caséine sont produites avec une structure et une stabilité différente. L'association des protéines du lactosérum aux micelles à des pH<6,8 modifie la surface des particules de caséine. Il a été démontré récemment que l'association de protéines sériques dénaturées avec les micelles de caséine augmente leur taille et que le potentiel zeta des micelles enveloppées de protéines sériques est plus grand que celui des micelles natives, ce qui contribue à la stabilité des ces particules (Anema et Li, 2003). En conséquence, les micelles
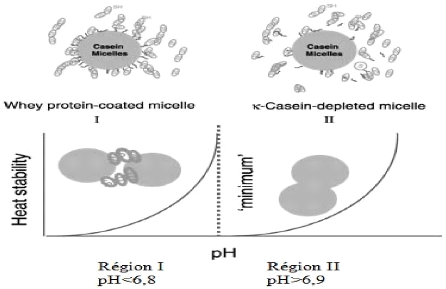
enveloppées de protéines sériques sont plus stables à la chaleur, aux ions calcium, à l'éthanol et à la présure que les micelles de caséines natives (Singh et Fox, 1986). Les micelles de caséine appauvries en ê-CN formées par chauffage du lait à pH>6,8 ont un potentiel zeta réduit et présentent une sensibilité accrue aux ions calcium, à l'éthanol et à la chaleur comparativement aux micelles natives. Cependant, la détermination de l'état moléculaire de la ê-CN qui se dissocie de la micelle est compliquée par le fait que, lors du chauffage, les protéines sériques interagissent avec celle-ci via des réactions d'échange thiols-disulphide. L'hypothèse avancée par Singh (2004) pour expliquer la dissociation de la ê-CN à pH légèrement alcalin repose sur les interactions hydrophobes qui seraient insuffisantes pour retenir la ê-CN une fois une valeur critique de charge de surface atteinte. Les répulsions électrostatiques entre la êCN et les autres composantes de la micelle causeraient alors la dissociation. L'autre hypothèse implique la conversion du phosphate de calcium colloïdal en une forme alternative moins apte à lier les molécules de caséine et de maintenir la structure de la micelle. Cependant, puisque l'attachement de la ê-CN à la micelle n'implique pas de phosphate de calcium colloïdal, un tel changement influencerait la dissociation des autres caséines mais très peu la dissociation de la ê-CN (Singh, 2004). 4.3.7.2 Changements dans la phase sérique Le chauffage affecte aussi la phase sérique qui entoure les micelles. En effet, le pH du lait décroît graduellement avec l'augmentation du temps de chauffage à 140 °C. De plus, les traitements thermiques diminuent les concentrations de phosphate soluble ainsi que du calcium soluble et ionique. Toutefois, l'activité de l'ion calcium, qui dépend du pH initial du lait, décroît pendant le chauffage malgré que le pH diminue (Singh, 2004). 4.3.7.3 Mécanismes de coagulation du lait à la chaleur Un mécanisme pour expliquer les profils HCT-pH des laits a été développé. Tout d'abord, la courbe HCT-pH est divisée en deux régions en fonction du pH (Figure 34 ). Figure 34. Interactions induites par la chaleur selon le pH (Singh, 2004).
Dans la région I (pH<6,8), la stabilité des micelles est assurée par le recouvrement en protéines sériques, A pH bien en-dessous du maximum de stabilité, le lait coagule rapidement à cause du faible pH et de la diminution des répulsions électrostatiques. De plus, les grandes quantités de protéines sériques associées aux micelles peuvent favoriser l'agrégation des particules de caséine par réticulation des protéines sériques liées à des micelles différentes. La présence d'un maximum (pH=6,7-6,8) dans les profils de stabilité à la chaleur est attribuable à la plus grande stabilité des micelles recouvertes de protéines sériques. La formation du complexe â-LG-ê-CN à la surface des micelles de caséine altère les interactions stériques et électrostatiques et prévient la dissociation de la ê-CN micellaire (Singh, 2004; O'Connell et Fox, 2003). Dans la région II, la stabilité thermique décroît suite à la dissociation de la ê-CN micellaire qui avait un effet stabilisant. Le minimum de stabilité dans le profil HCTpH est le résultat de la coagulation induite par les sels des micelles appauvries en êCN, plus sensibles à la présence d'ions calcium. A pH supérieur au minimum de stabilité, et ce même si la dissociation de la ê-CN micellaire augmente, il y a accroissement de la stabilité thermique grâce à l'augmentation de la charge protéique et de la faible activité de l'ion calcium (Singh, 2004; O'Connell et Fox, 2003). Cependant, O'Connell et Fox (2003) ont émis une hypothèse alternative pour expliquer la dépendance au pH de la stabilité thermique du lait qui amplifie le rôle joué par le calcium. Toutefois, cette hypothèse n'explique pas la dissociation de la kCN à pH>6,9, telle qu'observée par de nombreux chercheurs (Singh, 2004). 5 Electrolyse à membrane Les électrotechnologies, basées sur la
l'utilisation d'électrodes qui peuvent ajouter ou
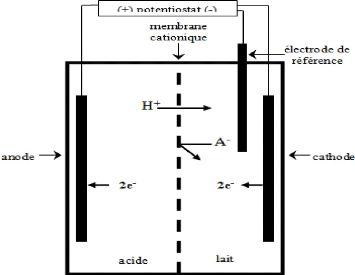
métaux lourds des eaux usées, la dépollution d'air contaminé et le recyclage d'effluents (Pletcher et Weinberg, 1992b). Certaines sont des technologies « vertes » puisqu'elles ne produisent pas d'effluents et qu'elles n'utilisent ni ne produisent de matériel toxique. La technologie est basée sur l'utilisation de cellules électrolytiques, combinées ou non à des membranes perméables aux ions, qui utilisent l'énergie électrique pour forcer des réactions à la surface d'une électrode immergée dans le liquide à traiter. Ces cellules fonctionnent en mode continu, semi-continu et discontinu (Pletcher et Weinberg, 1992a,b). 5.1 Principe Les modifications chimiques qui ont lieu dans une cellule d'électrolyse sont déterminées par trois facteurs à savoir les réactions d'oxydation à l'anode, les réactions de réduction à la cathode et la nature des espèces chimiques circulant à travers des séparateurs entre les électrodes. Les séparateurs sont le plus souvent des membranes qui peuvent être perméable aux anions ou au cations (Pletcher et Weinberg, 1992a,b). Les membranes laissent passer le courant et sont utilisées pour permettre l'équilibre des charges du système. La neutralité électrique évite une éventuelle oxydation, à l'anode, de l'espèce réduite à la cathode et vice-versa (Bazinet et al ., 1998). Dans le cas de l'électroréduction, les électrons fournis par une source de courant sont transférés de la cathode aux espèces en solution et les cations circulent de l'anode vers la cathode à travers une membrane de telle sorte qu'une charge équivalente à celle transférée aux électrodes soit transportée à travers la membrane (Bazinet et al ., 1998 ; Pletcher et Weinberg, 1992a,b). De ce fait, un composé n'est oxydé que si un autre est réduit (Bazinet et al ., 1998). 5.2 Applications connues De nombreuses applications des traitements d'électrolyse dans le monde alimentaire ont été développées. Les traitements électrolytiques sont utilisés pour coaguler les protéiques sériques (Janson et Lewis, 1994), prévenir le brunissement du jus de pomme (Tronc et al ., 1998), produire du tofu (Hara et al ., 2003) et obtenir de l'huile de canola hydrogénée comportant moins d'acides gras trans (Mondal et Lalvani, 2003). De le domaine laitier, un brevet a été obtenu pour la production de poudre de lait de meilleure saveur et odeur (Inoue et al ., 2003). La seule application connue de l'électroréduction dans le secteur laitier est la réduction, jusqu'à 39 % sous certaines conditions, des ponts disulfures des protéines de lactosérum (Bazinet et al ., 1997). Ces travaux ouvrent la voie à l'empêchement possible de l'oxydation des groupements thiols de la â-lactoglobuline menant à la fixation de celle-ci sur la ê-caséine par l'application d'un traitement d'électroréduction au lait. 5.3 Cellule d'électrolyse Une cellule d'électrolyse à membrane pour l'électroréduction en mode statique est généralement composée de deux électrodes métalliques immergées dans des électrolytes séparés par une membrane cationique (Bazinet et al ., 1998). La Figure 35 illustre une cellule de verre typiquement utilisée en laboratoire. La solution à traiter est placée dans le compartiment cathodique alors qu'un acide dilué est placé dans le compartiment anodique. Une membrane cationique de grade alimentaire assure la
séparation des liquides tout en laissant passer le courant et les cations pour assurer l'électroneutralité (Bazinet et al ., 1998). Les électrodes d'acier inoxydable sont reliées à un générateur de courant et divers appareils de mesure peuvent être introduits dans le système. Une électrode de référence est insérée tout près de la cathode afin de contrôler le potentiel appliquée à celle-ci. Finalement, les compartiments sont munis de barreaux magnétiques pour permettre une agitation constante dans le but de minimiser les zones de polarisation, le colmatage à la surface de la membrane et la formation de dépôts à la surface des électrodes. Figure 35. Schéma d'une cellule d'électrolyse à membrane, de type « statique ».
6 Approche proposée 6.1 Réduire la sensibilité à l'oxydation A notre connaissance, aucune étude n'a porté sur la modulation du potentiel d'oxydoréduction (redox) pour contrôler les réactions d'oxydation dans le lait. La valeur du potentiel redox (ORP) est une mesure de l'affinité pour les électrons ou du potentiel à donner des électrons d'un système redox. Plus le potentiel redox est négatif, plus la tendance à donner des électrons est grande. Le potentiel redox augmente donc avec l'augmentation d'énergie des électrons (Patushenko et al. , 2000). Certaines études ont étudié l'effet de l'ajout de substances chimiques anti-oxydantes dans le lait avec des résultats mitigés (Halliwell et al. , 1995). De plus, ces substances ne sont pas toutes permises dans le lait et le consommateur est réticent à l'ajout d'additifs chimiques. Pour contourner ce problème, nous proposons la modulation du potentiel
redox par électrolyse à membrane pour augmenter stabilité lait à la chaleur. Le procédé d'électrolyse à membrane utilise l'énergie électrique pour forcer des réactions à la surface d'une cathode inerte en contact avec le produit à traiter (Wang et al. , 1991). Les réactions qui surviennent à la cathode sont responsables de la réduction des espèces actives et de l'abaissement du ORP. Les protons utilisés sont remplacés par ceux de l'anolyte qui traversent la membrane cationique pour maintenir l'électroneutralité de la solution. Une telle cellule d'électrolyse a été utilisée par Bazinet et collaborateurs (Bazinet et al ., 1997) pour réduire les ponts disulfures de la â-LG. Par cette technologie, il sera possible de contrôler la concentration d'oxygène dissous dans le lait, le potentiel redox et l'état d'oxydation des espèces chimiques actives pour augmenter la stabilité du lait à la chaleur et ainsi prévenir les pertes de lysine, l'apparition de la coloration brune et la coagulation à l'entreposage des laits stérilisés, tout en favorisant l'attaque du lait à la présure en fabrication fromagère(Marie-Michèle Filion, 2006).
Partie III : Organisation de la filière laitière au Maroc
Introduction L'élaboration du plan laitier en 1975 a eu pour conséquence une dynamique manifeste de la filière lait durant les trois dernières décennies. Ceci, pour faire face à une demande croissante due à une explosion démographique. Au Maroc, la production bovine laitière jouit d'un statut très particulier dans les plans de développement agricole (Hamama et al, 2005). En effet, elle remplit des rôles sociaux et économiques non négligeables pour créer des emplois et des richesses dans nombreuses exploitations agricoles. Pour cela, nous allons présenter l'évolution de la production laitière au niveau national dans ce chapitre, les circuits de commercialisation, les infrastructures de collecte, ainsi que les facteurs limitant la qualité du lait.
IX. Présentation de la filière L'industrie laitière au Maroc peut être considérée comme un champ d'activité stratégique dans lequel des opérateurs économiques développent des approches différentes (souvent complémentaires parfois même contradictoires), dont les objectifs en reflètent les logiques d'action (régulation ou rentabilité). Les principaux acteurs du processus d'industrialisation de cette production sont notamment l'Etat, les Centres de Collecte privés (CCP), les coopératives de collecte de lait et les grandes sociétés de transformation du lait. L'évolution du secteur laitier est tout d'abord orientée par :
Avec l'arrivée des entreprises espagnoles, la production est faite sous contrat avec les producteurs. En effet, les agriculteurs individuels reçoivent les plants en contrepartie, ils livrent leur production à l'entreprise qui la conditionne et l'exporte. Aucun contrat n'est signé entre le producteur et l'entreprise qui fixe le prix en fonction du marché, par conséquent, les agriculteurs supportent seuls la variation des prix de vente. Des groupes d'agriculteurs se sont décidés de se réunir pour créer des coopératives (Faysse et al, 2007). 2.2 La concurrence entre les coopératives de collecte de lait et le secteur informel Quoique l'objectif de mon travail se limite d'analyser l'état de la concurrence dans lequel s'insèrent les coopératives et les CCP qui opèrent dans le circuit formel, il s'est avéré nécessaire de montrer que le secteur informel est aussi en concurrence avec les coopératives. Le bureau international du travail (BIT) distingue sept critères pour définir le secteur informel : la facilité d'entrée à l'activité, un marché de concurrence non réglementé, la propriété familiale des entreprises, l'échelle restreinte des activités, le recours aux réserves locales, le recours à une technologie adaptée et à une forte intensité de travail, une formation acquise hors du système scolaire. La dernière définition et critères de l'informalité proposés par cet organisme est « le faible lien avec les institutions officielles, un faible niveau d'organisation, des unités qui opèrent à petite échelle et de manière spécifique, avec peu ou pas de division entre le travail et le capital en tant que facteurs de production. Les relations de travail, lorsqu'elles existent, sont surtout fondées sur l'emploi occasionnel, les relations de parenté ou les relations personnelles et sociales plutôt que sur des accords contractuels comportant des garanties en bonne et due forme » (BIT, 1993a et b). Dans le monde, le secteur informel laitier contribue pour une part significative, à l'activité économique totale de ce secteur. Son expansion a été fortement accentuée par l'explosion démographique, l'urbanisation, l'exode rural et la libéralisation de l'économie. Le secteur informel échappe à tout contrôle des pouvoirs publics qu'il soit fiscal, social ou sanitaire. Il constitue, selon certains dires, non seulement une concurrence déloyale pour le sous-secteur organisé mais aussi un danger de santé publique. Il est pourtant fortement présent dans les pays en voie de développement. Le secteur informel est dynamique dans le secteur laitier. Selon les pays, l`informel draine entre 30 et 98 % de la production commercialisée dont une partie très faible drainée par les coopératives et représente entre 25 et 75 % du chiffre d`affaires du secteur laitier. Ainsi, la part des coopératives dans le lait usiné dans quelques pays est donnée par le tableau ci-dessous :
* Source : DE, 2007 Source : Staal, 2000 Quelques exemples de la concurrence entre les coopératives et le secteur informel : É Au Kenya, le marché informel domine le commerce du lait, 86% du lait est vendu cru par des petits vendeurs ambulants. Plus de 42% du lait est vendu directement du producteur au consommateur. Un quart du lait passe par les coopératives laitières, et une partie est vendue sous forme cru. De plus, plusieurs sociétés étrangères sont rentrées en compétition avec les coopératives laitières locales en vendant du lait pasteurisé. La libéralisation du marché laitier pose des défis importants pour les coopératives laitières (ILRI, Kenya, 2005). É En Inde, dans le cas de la coopérative laitière de Baroda, une coopérative avait des problèmes parce que pendant l'été, les petits vendeurs informels proposaient un prix plus haut qu'elle, et comme les villages étaient prêts de la ville de vente, ils pouvaient sans problème écouler leur marchandise. Ceci affaiblissait grandement la coopérative, (Shah, 1995).
Maitrise de la qualité du lait cru de la ferme ii l'usine de transformation EL.BOUICHOU
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||