I.1.2.2. Immunité adaptative ou spécifique
Intervient secondairement après une phase de
reconnaissance de l'antigène, de prolifération lymphoïde et
de différenciation en cellules productrices d'anticorps (lymphocytes B /
plasmocytes) et de cytotoxicité (lymphocytes TCD3+). Cette
immunité adaptative est douée de mémoire et un
deuxième contact avec l'antigène permet une réponse rapide
et puissante de type secondaire (Olivier, 2005). L'immunité adaptative,
antérieurement qualifiée d'immunité spécifique, est
nécessaire pour le contrôle des infections de longue durée
et pour la mise en place d'une mémoire immunologique sur laquelle
s'appuie la vaccination (Cavaillon, 2010).
Kouassi et al., (2003) intitule que, l'immunité
spécifique est apparue lors de la divergence entre
vertébrés et invertébrés. Elle est
caractérisée par un ensemble de molécules de structure
extrêmement diversifiée appartement toutes à la
superfamille des immunoglobulines (Ig) : les anticorps, les récepteurs
d'antigène des lymphocytes T (TCR) et les molécules CMH. Les
molécules d'Ig existent sous forme soluble (les anticorps
répartis en cinq classes de fonctions biologiques différentes
chez l'homme : IgM, IgG, IgA, IgD et IgE) et sous forme de récepteurs
membranaires des lymphocytes B (BCR). La molécule d'anticorps,
formée en général de 2 chaînes
légères, interagit par son site de liaison ou paratope avec une
zone de l'antigène appelée épitope. Chaque épitope
correspond à une zone de 2 à 3 nm de diamètre (soit
environ 15 acides aminés).
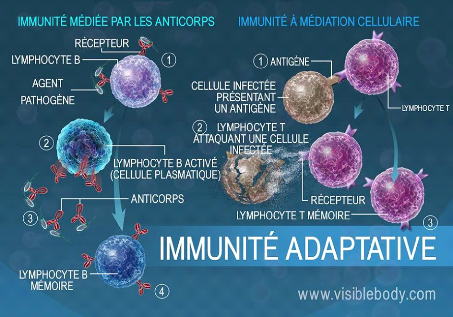
Figure 11 : organisation de l'immunité
adaptative.
I.1.2.2.1. Les acteurs de l'immunité adaptative
Il s'agit principalement des lymphocytes B et T, les
lymphocytes B étant responsables de la réponse immunitaire
humorale (production d'anticorps) et les lymphocytes T des réponses
cellulaires (auxiliaire, cytotoxique ou régulatrice). Les lymphocytes B
et les lymphocytes T ont une morphologie similaire, avec un rapport
nucléo-cytoplasmique élevé sans granulation. Ils sont
capables de reconnaître spécifiquement des antigènes via
leurs immunorécepteurs de type BCR ou TCR. Le BCR se lie à
l'antigène natif alors que le TCR se lie à des antigènes
apprêtés et présentés sous forme de peptide
associé aux molécules du CMH. Il existe des sous-populations
fonctionnelles de lymphocytes T et B définies par leur phénotype,
c'est-à-dire un ensemble de caractéristiques moléculaires
membranaires, et des propriétés fonctionnelles
différentes. Par exemple, parmi les lymphocytes T, on distingue deux
sous-populations majeures : les lymphocytes T auxiliaires ou helpers (Th) et
les lymphocytes T cytotoxiques. Les lymphocytes T auxiliaires
sécrètent des cytokines et sont responsables de l'organisation
des réponses immunitaires innées et adaptatives. Les lymphocytes
T cytotoxiques provoquent la mort des cellules présentant des
antigènes étrangers (dans le cas d'une infection virale ou
d'autres pathogènes intracellulaires) ou des
antigènes du soi anormal en termes qualitatif et/ou quantitatif (dans le
cas d'une cellule tumorale) (Jonathan et al., 2018).
Il existe également des lymphocytes T
régulateurs exerçant des fonctions de régulation et
d'inhibition des réponses immunitaires. Au-delà de leur
rôle de précurseur des plasmocytes, cellules principalement
présentes dans la moelle osseuse ayant pour fonction la production des
anticorps en grande quantité et pendant une longue durée, les
lymphocytes B ont également un rôle de CPA aux lymphocytes T.
Cette propriété est à la base de la coopération
cellulaire entre les lymphocytes T et B afin de réguler l'activation de
ces derniers et ainsi la production des anticorps. Au décours des
réponses immunitaires, les lymphocytes B comme les lymphocytes T donnent
naissance à des cellules mémoires à durée de vie
longue dont le rôle est de répondre plus efficacement à une
nouvelle exposition à un antigène donné (réponse
secondaire) (Jonathan et al., 2018).
| 

