Impact du réchauffement climatique sur la distribution spatiale des ressources halieutiques le long du littoral français: observations et scénarios( Télécharger le fichier original )par Sylvain Lenoir Université Lille 1 Science - Doctorat 2011 |

2.2. IntroductionClimate warming has become unequivocal (Intergovernmental Panel on Climate Change 2007b) and over the last 40 years the ocean has absorbed 84% of the heat added to the climate system (Levitus et al. 2005). Several significant correlations between changes in climate and in the abundance of marine species, ranging from plankton to fish and seabirds (Aebischer et al. 1990; Beaugrand & Reid 2003; Frederiksen et al. 2006), suggest that marine ecosystems have responded quickly to a warming of temperature (Richardson & Shoeman 2004; Beaugrand et al. 2009; Drinkwater et al. 2010). For example, a sustained climate-induced change occurred in the trophodynamics of the North Sea ecosystem during the 1980s (Kirby et al. 2009) that altered the phenology (Edwards & Richardson 2004) and the biogeography of many species (Stebbing et al. 2002; Genner et al. 2004; Perry et al. 2005) across several trophic levels (Frederiksen et al. 2004; Kirby & Beaugrand 2009), and including the breeding success of seabirds such as black-legged kittiwakes and common guillemots (Wanless et al. 2005 ; Harris et al. 2007; Harris et al. 2008). North Sea seabird breeding success appears to be controlled partially by the abundance (Poloczanska et al. 2004; Frederiksen et al. 2005), distribution (Shealer 2002; Grémillet & Boulinier 2009) composition and nutritional quality of the prey the parent birds feed to their chicks on the nest (Wanless et al. 2005; Harris et al. 2008). As a consequence, an observed reduction in the breeding success of some North Sea seabird colonies has been attributed to dietary changes (Wanless et al. 2005; Harris et al. 2008). In particular, three species of fish in the North Sea, the lesser sandeel (Ammodytes marinus), the European sprat (Sprattus sprattus) and the snake pipefish (Entelurus aequoreus) seem to be important determinants of North Sea seabird breeding success (Harris et al. 2007, Harris et al. 2008); While the lesser sandeel and the European sprat, which are high energy fish, are the preferred prey, the snake pipefish, although frequently recorded in the diet of seabirds since 2004, and found discarded at nest sites, is a poor food for young chicks (Harris et al. 2008). Changes in the abundance and nutritional value of lesser sandeels and sprats, and in the abundance of snake pipefish, due to a combination of overfishing and the effects of hydroclimatic change (Frederiksen et al. 2004) seem to have acted synergistically to compromise North Sea seabird breeding success recently (Wanless et al. 2005; Wanless 2007). To further explore the possible effect of hydroclimatic change on the North Sea ecosystem with regard to the main fish prey of North Sea seabirds we applied a recently developed ecological niche model to forecast past and future spatial changes in the probability of occurrence of North Sea sandeels, sprats, and snake pipefish(Beaugrand et al in press; Lenoir et al in press). We took into account sea surface temperature (SST), bathymetry, sea surface salinity (SSS) (see electronic supplementary material, figure IV.S1), and, in the case of sandeels, bottom-sediment type (figure IV.S2). We then applied our model to determine whether future changes in the hydroclimatic regime of the North Sea may lead to a future increase in the abundance of two warm-water southern fish species, the European sardine (Sardina pilchardus) and the European anchovy (Engraulis encrasicolus). In a similar way to which climate altered the dietary composition of common guillemots (Uria aalge) in the California Current (Veit et al. 1996; Oedekoven et al. 2001), an increase in the abundance of sardine and anchovy in the North Sea could potentially compensate any decrease in the abundance of North Sea sprats and sandeels. ? Biological data Sandeel, European sprat, snake pipefish, European anchovies and European sardine data (post-larval stages) were obtained from the Ocean Biogeographic Information System (OBIS; available at: http://www.iobis.org/), the Global Information Biodiversity Service (GBIF; available at: http://data.gbif.org/), and the ICES-FishMap online atlas (available at: http://www.ices.dk/marineworld/fishmap/ices/). Both temporal and spatial data are available for the majority of occurrences with the exception of the sardine where, due to reduced spatial coverage, we determined the current spatial range based upon expert knowledge and examination of the OBIS data. First, we determined the minimum and maximum latitudinal occurrence of the sardine by examining the OBIS data. Second, we completed the occurrence point between 20°N and 54°N in each node of a spatial grid of 0.1° longitude x 0.1° latitude where bathymetry was between 0 and 3000m (North Atlantic, including the Mediterranean Sea and the North Sea). ? Environmental data Mean annual SST for the period 1960-2008 were obtained from the International Comprehensive Ocean-Atmosphere Data Set (ICOADS, http://icoads.noaa.gov; Woodruff et al. 1987) and from the ECHAM 4 model (EC for European Centre and HAM for Hamburg; for modelled SST data based upon climate change scenarios A2 (scenario of a divided world) and B2 (scenario of a divided world, but more ecologically friendly; Intergovernmental Panel on Climate Change 2007b). The SST data are therefore modelled data based on scenario A2 (a concentration of carbon dioxide of 856 ppm by 2100, which represents an increase of CO2 similar to that currently observed) and B2 (a concentration of carbon dioxide of 621 ppm by 2100) and they reflect world populations of 15.1 and 10.4 billion people in 2100, respectively (Intergovernmental Panel on Climate Change 2007b). For comparison purposes SST were also averaged for the decades, 1960-1969, 2050-2059 and 2090-2099, and for the period 2000-2008. In addition to hydroclimatic change, other physical habitat variables that influence the distribution of fish species are sea surface salinity (SSS) (electronic supplementary material, figure IV.S1) (Schmidt-Nielsen 1990) and bathymetry (Louisy 2002), and, for species such as the sandeel that has a benthic phase, the bottom-sediment type. We obtained data for mean annual SSS from both Levitus' climatology (Levitus 1982) and ICES ( http://www.ices.dk; Lenoir et al. 2010); we did not include temporal changes in SSS for two reasons, firstly, spatial variance in salinity is much more pronounced than the temporal variance and secondly, temporal variance is poorly assessed in the Atmosphere-Ocean General Circulation (AO-GCM) models (Martin Visbeck, Personal Communication). We obtained bathymetry data from the global ocean bathymetry map (Smith & Sandwell 1997) and for the sandeel we obtained data on bottom-sediment type from the "Service Hydrogéographique et Océanographique de la Marine" (SHOM). (electronic supplementary material, figure IV.S2). ? Calculation of thermal niches We used occurrence data to assess the thermal niche (sensu Hutchinson, Hutchinson 1957) of each species by calculating the number of individuals recorded per degree of annual SST (figure IV.1). ? Mapping of species occurrence We used the non-parametric probabilistic ecological niche model (NPPEN; Beaugrand et al. in press; Lenoir et al. 2010, and electronic supplementary material 1), to assess the probability of species occurrence and to map their potential spatial distribution. Briefly, the technique tests whether environmental conditions within a cell in a geographical space belong to the estimated ecological niche of a species. The model tests whether or not an observation (i.e. the j geographical cells of our environmental grids), characterised by an environmental stratum (p variables), belongs to a reference matrix, here matrix Zm,p. Mathematically, the technique is nonparametric using a modified version of the multiple response permutation procedure (MRPP; Mielke et al. 1981). The technique uses the Generalised Mahalanobis distance (Mahalanobis, 1936), which enables the correlation between variables to be considered (Farber & Kadmon 2003). The distance was calculated as follows:
With x the vector of length p, representing
the values of the three/four environmental stratums to be tested
(j=2,530,206), Rp,p the correlation matrix of
reference matrix Zm,p and ? Long-term changes in the probability of fish occurrence Long-term spatial changes in the probability of fish occurrence were calculated for the five species on an annual basis from 1960 to 2100. Maps of species occurrence were built for the period 1960-1969, 2000-2008, 2050-2059 and 2090-2099 within the North Sea spatial domain ranging from 4°W to 9°E and from 51°N to 62°N (figures IV.2?IV.3; electronic supplementary material, figures IV.S3?IV.S7). ? Validation of the model We used correlation analysis (Spearman's rank correlation) to validate the modelled spatial distributions of each fish species by comparing the probabilities of occurrence modelled by NPPEN with the percentage of the number of actual presence data points in the North Sea for 2000-2008 (figure IV.4). Only half of the data, not incorporated to the model, were used for validation. We also use correlation analysis to compare the long-term changes in spawning stock biomass (SSB) of the lesser sandeel (two-year old fish) to the annual average probabilities of occurrence modelled by NPPEN in the ICES subarea IV for the period 1983-2003 (figure IV.5). No SSB data were available for the four other species in this area. |
|


 (1)
(1)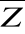 the average environmental condition inferred from
Zm,p. The significance of the Generalised
Mahalanobis distance was then assessed using a simplified version of the MRPP
(Beaugrand et al. in press; Lenoir et al. 2010).
the average environmental condition inferred from
Zm,p. The significance of the Generalised
Mahalanobis distance was then assessed using a simplified version of the MRPP
(Beaugrand et al. in press; Lenoir et al. 2010).